Embed Size (px)
Citation preview

Die Angewandte Makromolekulare Chemie 85 (1980) 41-77 (Nr. 1325)
Institut fur Organische Chemie der Freien Universitat Berlin D-1000 Berlin 33, Takustr. 3
An Synthesepulp immobilisierte Enzyme, I.
Georg Manecke und Hans-Gunter Vogt
H e m Prof. Dr. M. Seefelder zum 60. Geburtstag gewidmet
(Eingegangen am 4. Juli 1979)
ZUSAMMENFASSUNG: Synthesepulp - ein Polyethylen-Fibrid, das Polyvinylalkohol enthalt - wurde mit 2-
(3-Aminophenyl)-l,3-dioxolan umgesetzt und das Aminoderivat in die entsprechende Diazoniumverbindung iiberfuhrt. Der Gehalt an reaktionsflhigen Diazoniumgruppen wurde durch Umsetzung mit Tyrosin ermittelt. Die Diazoniumderivate des Synthese- pulps wurden fur die Immobilisierung der Hydrolasen Trypsin, Chymotrypsin, Papain, Aminoacylase, Esterase und Urease, der Oxidoreduktasen Glucose-Oxydase, Katalase, Peroxydase und Glucose-6-phosphat-dehydrogenase und der Transferase Hexokinase eingesetzt. Weiterhin wurde Sojabohnen-Trypsininhibitor an Synthesepulp durch Azo- kupplung immobilisiert. Das Proteinbindungsvermogen der reaktiven Trager und die enzymatischen Eigenschaften der Immobilisierungsprodukte wurden untersucht. Die Thermostabilitat von immobilisiertem Trypsin und immobilisierter Urease sowie die Temperaturabhangigkeit der Aktivitat von immobilisiertem Trypsin wurde untersucht. Die Aktivitaten von immobilisiertem Trypsin wurden gegenuber niedermolekularen und hochmolekularen Substraten ermittelt. Die pH-Optima von Trypsin, Chymotryp- sin, Papain und Urease wurden bestimmt, und der EinfluB von Puffersubstanzen auf die pH-Aktivitatsprofile wurde untersucht. Hexokinase und Glucose-6-phosphat-dehydro- genase wurden gemeinsam als Beispiel fur ein Zwei-Enzymsystem an Synthesepulp im- mobilisiert und die Eigenschaften des immobilisierten Systems bestimmt.
SUMMARY: Synthetic pulp - a polyethylene fibrid which contains poly(viny1 alcohol)- was
reacted with 2-(3-aminophenyl)-l,3-dioxolane. The amino derivative was transferred into the reactive diazonium compound. The content of reactive diazonium groups was determined by reaction with tyrosine. The diazonium derivatives of synthetic pulp were used for the immobilization of the hydrolases trypsin, chymotrypsin, papain, aminoacy- lase, esterase and urease of the oxidoreductases glucose oxidase, catalase, peroxidase, and glucose-6-phosphate-dehydrogenase, and of the transferase hexokinase. Further- more soybean trypsin inhibitor was immobilized on synthetic pulp via azo coupling. The protein binding ability of the reactive carriers and the enzymatic properties of the im- mobilization products were investigated. The thermostability of immobilized trypsin
41

G. Manecke und H.-G. Vogt
and immobilized urease, as well as the dependence of the immobilized tryptic activity on the temperature were studied. The activities of immobilized trypsin were assayed with low molecular weight substrates as well as with high molecular weight substrates. The pH-optima of immobilized chymotrypsin, papain and urease were studied and the influence of buffers on the pH-activity profiles was investigated. Hexokinase and glu- cose-6-phosphatedehydrogenase were co-immobilized on synthetic pulp as an example for a two enzyme system and the properties of the immobilization products were investi- gated.
1. Einleitung
Fur die kovalente Immobilisierung von Enzymen sind synthetische polyme- re Trager unterschiedlicher Morphologie (z. B. Granulate, Perlpolymere, Fil- me, Fasern, Hohlfasern sowie Membranen) eingesetzt worden. Als Tragerma- terialien kommen vor allem mechanisch stabile Polymere in Betracht, die au- Ber den zur Enzymimmobilisierung bestimmten reaktiven Zentren und den hydrophilen Gruppen von der Matrix her chemisch inert sind. Neben der che- mischen Struktur der Trager ist die Tragermorphologie fur die Eigenschaften der immobilisierten Enzyme von Bedeutung.
Da haufig die Diffusion der geschwindigkeitsbestimmende Schritt fur die Aktivitat immobilisierter Enzyme ist, ist es giinstig, Tragermaterialien auszu- wahlen, die es gestatten, das Enzym ausschlieBlich an der Trageroberflache zu immobilisieren. Eine groBe Trageroberflache und ein inerter Kern, der dem Trager eine ausreichende mechanische Festigkeit verleiht, ist hierbei von Vor- teil.
Diese Eigenschaften werden von SWP (synthetic wood pulp), einem synthe- tischen Faserstoff, der urspriinglich von Crown-Zellerbach fur die Papierher- stellung entwickelt wurde', in idealer Weise erfullt. SWP ist eine neue Erschei- nungsform des Polyethylens, die bei der sog. scherinduzierten Kristallisation von Polyethylen aus Polymerlosungen gebildet wird2. Polyethylen kristallisiert hierbei zu sogenannten Shish-Kebabs. Die Herstellung von SWP laBt sich in Gegenwart von Polyvinylalkohol (PVA) als Hydrophilierungsmittel durchfuh- ren, wobei dieser fest an der Oberflache der Fibride fixiert wird2. Derartige Polyethylenfibride sind kommerziell z. B. als Hostapulp@ (Hoechst) erhaltlich. Fur die vorliegende Arbeit wurde Synthesepulp der Firma Hoechst mit den Typenbezeichnungen SWP-E 400 und SWP-R 830 eingesetzt, deren PVA-Ge- halt bei 1,55 bzw. bei 4,2% lag.
Die Derivatisierung des PVA-Anteils im Synthesepulp und die Enzymim- mobilisierungen erfolgten nach Methoden, die teilweise bereits an PVA-Tra-
42

An Synthesepulp immobilisierte Enzyme, I.
gem anderer Morphologie erprobt ~ u r d e n ~ - ~ . Die an Synthesepulp immobili- sierten Enzyme lassen sich verfahrenstechnisch sowohl in Ruhrkesseln als auch in Saulenreaktoren einsetzen und bilden durch ihre spezielle Struktur weiterhin die Moglichkeit zu einem Einsatz als mit Enzymen beschichtete Fil- terpapiere. Zur Darstellung reaktiver Trager auf der Basis von Synthesepulp wurde als Aktivierungsmoglichkeit die Umsetzung mit 2-(3-Aminophenyl)- 173-dioxolan gewahlt, die fur vernetzte PVA-Trager bereits in friiheren Arbei- ten beschrieben wurde3:
2-(3-Aminophenyl)- 1,3-dioxolan > pq=E~q NH2 N , + C i
SWP-E 400 bzw SWP-R 830
SWP-E 400-Ia bzw. SWP-R 830-Ia-c SWP-R 830-IIa-c
SWP-E 400-1Ia bzw
Synthesepulp wurde mit unterschiedlichen Mengen 2-(3-Aminophenyl)-l,3- dioxolan in 0,3 M HC115 h bei 50 "C unter Ruhren umgesetzt. Diese Derivati- sierungen wurden an den Synthesepulp-Typen SWP-E 400 und SWP-R 830 mit den in Tab. 1 aufgefwen Mengen eingesetzten Dioxolans durchgefuhrt. Die so erhaltenen SWP-Aminoderivate Ia-c wurden mit Wasser gewaschen, mit Aceton extrahiert und getrocknet. Die Darstellung reaktiver Trager erfolg- te durch Diazotierung zu SWP-E 400-IIa bzw. SWP-R 830-IIa-c. Wegen des geringen Gehaltes an funktionellen Gruppen lieBen sich die SWP-Derivate weder elementaranalytisch noch IR-spektroskopisch eindeutig charakterisie- ren. Die Bestimmung des Gehalts an reaktiven Diazoniumgruppen wurde da-
Tab. I. Umsetzung von Synthesepulp mit 2-(3-Aminophenyl)-l,3-dioxolan zu den Aminoderivaten SWP-E 400-Ia und SWP-R 830-Ia-c und Kupplung der dia- zotierten Derivate mit Tyrosin.
Diazonium- derivat des SWP
SWP-E 400-IIa SWP-R 830-IIa SWP-R 830-IIb SWP-R 830-IIc
Darstellung der Aminoderivate Bindung von Tyrosin an die eingesetzte Menge Diazoniumderivate I1 des SWP 2-(3-Aminophenyl)-l,3-dioxolan in pl/g in mmol/g in mg/g in pmol/g SWP SWP SWP-Derivat SWP-Derivat
200 1,46 0,3 1,5 200 1,46 1,4-1,5 7,5-8 100 0,13 0,5 3 20 0,15 0,4-0,5 2-3
43

G. Manecke und H.-G. Vogt
her durch Modellumsetzungen mit Tyrosin durchgefuhrt. Die Tyrosinkupp- lung erfolgte in 0,05 M Natriumcarbonatlosung unter 2-stiindigem Riihren bei 0 "C. Das iiberschiissige Tyrosin wurde ausgewaschen und der Tyrosingehalt der Waschlosung mit Folin-Reagenz bestimmt. Die auf diese Weise ermittel- ten Bindungsvermogen der reaktiven Trager fur Tyrosin sind in Tab. 1 aufge- fuhrt, sie entsprechen dem fur die Enzymimmobilisierung effektiv zur Verfu- gung stehenden Gehalt an reaktiven Diazoniumgruppen.
Der unter den gewahlten Bedingungen erzielte hochste Gehalt an reaktiven Diazoniumgruppen betragt mit 8 Fmol/g SWP etwa 2% des theoretischen Ma- ximalwertes fur vollstandige Acetalisierung des PVA-Anteils im Synthesepulp. Dieser niedrige Substitutionsgrad zeigt, daB der iiberwiegende PVA-Anteil im Synthesepulp nicht fur Oberflachenreaktionen zur Verfugung steht.
2. Azokupplung von Enzymen an Diazoniumderivate(II) des Synthesepubs
Der EinfluB des Gehalts der Trager SWP-E 400-IIa und SWP-R 830-1Ia-c an reaktiven Gruppen auf das Enzymbindungsvermogen wurde anhand der Enzyme Trypsin und Chymotrypsin untersucht. Hierzu wurden jeweils 25 mg Enzym in 10 cm3 Phosphatpuffer pH 8,O (Ionenstarke p=O,15) mit 100 mg SWP-Derivat I1 2 h bei 0 "C umgesetzt. Das Enzymbindungsvermogen der un- tersuchten Trager ist in Tab. 2 gezeigt.
Tab. 2. Immobilisierung von Enzymen an Diazoniumderivate I1 des Synthesepulps. Abhangigkeit des Enzymbindungsvermogens vom Gehalt an Diazoniumgrup- pen.
SWP-Derivat Gehalt an Immobilisiertes Enzymb reaktiven Trypsin Chymotrypsin Gruppen" in in mg/g in pmol/g in mg/g in pmol/g wmol/g SWP I1 SWPII SWPII SWPII SWPII
SWP-E 500-IIa 1,s 15 0,6
SWP-R 830-IIb 3 35 1 s 20 03 SWP-R 830-IIc 2-3 12 0 s 16 0,6
a bestimmt aus dem Tyrosinbindungsvermogen Kupplungsbedingungen: 25 mg Enzym; 10 cm3 Phosphatpuffer pH 8,0 (Ionenstarke p=O,lS); 100 mg SWP-Derivat 11; 2 h 0°C.
SWP-R 830-IIa 7,5-8 47 290 35 1,4
44

An Synthesepulp immobilisierte Enzyme, I.
Wie aus Tab. 2 zu ersehen ist, ist das Enzymbindungsvermogen der diazo- tierten SWP-Derivate in Relation zum Gehalt an reaktiven Gruppen erstaun- lich hoch. Fur die untersuchten Trager betragt das molare Verhdtnis von ge- bundenen Enzymmolekulen und reaktiven Gruppen der Matrix l : 2 bis l : 6. Der Anteil der durch eine Einzelbindung an den SWP-Trager gebundenen Enzymmolekiile sollte daher bei den Immobilisierungsprodukten relativ hoch sein. Durch weitere Verdunnung der reaktiven Gruppen am Trager sollte es moglich sein, Mehrfachbindungen eines Enzymmolekuls nahezu ausschlie- Oen zu konnen. Friihere Untersuchungen an Diazoniumgruppen-haltigen ver- netzten PVA-Gelen3 mit 1,4 mmol Diazoniumgruppen pro g zeigten ein weit ungunstigeres molares Verhaltnis von immobilisierten Trypsinmolekulen zu reaktiven Gruppen. In diesem Falle betrug der molare UberschuO an reakti- ven Gruppen der Trager das 80- bis 380fache verglichen mit der molaren im- mobilisierten Trypsinmenge. Die sehr vie1 bessere Ausnutzung der reaktiven Gruppen zur Enzymbindung bei den SWP-Derivaten ist durch die Struktur der SWP-Fasern bedingt. Der inerte Kern der Fibride bewirkt, daO Derivati- sierungen und Immobilisierungen praktisch ausschliefilich an der auOeren Oberflache erfolgen und sterische Behinderungen bei der Enzymimmobilisie- rung weit weniger auftreten, als im Falle von Tragern mit Porenstruktur.
Das Enzymbindungsvermogen der Synthesepulpderivate ist auOer von de- ren Gehalt an reaktiven Gruppen auch von den jeweiligen Kupplungsbedin- gungen abhiingig. Fur die Enzymimmobilisierung durch Azokupplung wird haufig ein schwach allcalischer pH und eine niedrige Kupplungstemperatur gewahlt. In Tab. 3 ist der EinfluB der Kupplungstemperatur an Beispielen fur
Tab. 3. Abhangigkeit der Menge des an SWP-R 830-IIb immobilisierten Enzyms und der relativen Aktivitat (im pH-Optimum) von der Kupplungstemperatur".
Kupplungs- Immobilisiertes Trypsin Immobilisiertes Chymotrypsin temperatur (mg Enzym relative (mg EmYm relative
pro g SWP) Aktivitat pro g SWP) Aktivitat ("C) (% b, (% 7
0 25
35 55
a Kupplungsbedingungen: 25 mg Enzym/lO cm3 Phosphatpuffer pH 8,O (p=0,15) 100 mg SWP-R 803-IIb; 2 h 0,Ol M BAEE, 0,2 M NaC1, 0,l M Tris-Puffer 0,02 M ATEE, 0,2 M KC1, 5 Vo1.-%Ethanol. '
45

G. Manecke und H.-G. Vogt
die Immobilisierung von Trypsin bzw. Chymotrypsin an SWP-R 830-IIb ge- zeigt.
Sowohl Trypsin als auch Chymotrypsin werden infolge der hoheren Reakti- onsgeschwindigkeit bei hoherer Kupplungstemperatur in groBeren Mengen gebunden. Die Zersetzung reaktiver Diazoniumgruppen scheint wiihrend der gewahlten Kupplungszeit von 2 h bei 25 "C unbedeutend zu sein. Bei einer Kupplungstemperatur von 0 "C ist jedoch die relative Aktivitat der immobili- sierten Enzympraparate wesentlich besser, was einerseits auf die bessere Zu- ganglichkeit infolge der geringeren Dichte des Enzyms am Trager, als auch z. T. auf die bessere Stabilitat der Enzymlosung bei 0 "C zuriickzufuhren ist. Alle folgenden Enzymimmobilisierungen an SWP I1 wurden daher bei 0°C durchgefuhrt. Tab. 4 gibt einen Uberblick uber Enzyme, die sich durch Azo-
Tab. 4. Immobilisierung von Enzymen durch Azokupplung an SWP-R 830-IIa".
Immobilisiertes Eingesetzte Eingesetzte Immobilisiertes Bindungs- Protein Protein- Proteinmenge Protein ausbeute
konzentration SWP-R 830-IIa SWP-R 830-IIa (mg/cm3) (mg/g) (mg/g) (%I
Trypsin Trypsin Chymotrypsin Papain Peroxy dase Aminoacylase Urease Esterase Trypsininhibitor Katalase Glucose-Oxydase
250 100 250 240 100 100 90
100 50 25 12,5
47 34 35
147 43 59 50 42 38 22 11
19 34 14 61 43 59 56 43 76 96 87
a Kupplungszeit 2 h, 0°C.
kupplung an SWP-R 830-IIa immobilisieren lieBen. Wie Tab. 4 zeigt, wird das Enzymbindungsvermogen der reaktiven Trager auBer durch das zu bindende Enzym selbst entscheidend durch die Menge und die Konzentration des fur die Immobilisierung eingesetzten gelosten Enzyms bestimmt. Fur einige im- mobilisierte Enzyme konnte gezeigt werden, daB bei genugend geringem En- zymangebot das zu bindende Enzym sich nahezu vollstbdig an SWP-R 830-
46

An Synthesepulp immobilisierte Enzyme, I.
IIa kovalent immobilisieren 1aBt. Daraus laBt sich ersehen, daB der reaktive Trager nicht zu einer unspezifischen Enzymadsorption neigt, die den kovalen- ten Bindungsanteil schmalern wiirde.
Die in Tab. 4 angegebenen Proteinbindungsvermogen von SWP-R 830-IIa wurden durch EiweiBbestimmung mit Folin-Reagenz' ermittelt. Weiterhin wurde die Proteinbindung gravimetrisch aus der Gewichtsdifferenz des ge- friergetrockneten immobilisierten Enzyms zur eingesetzten Tragermenge be- stimmt. Voraussetzung fur relevante Ergebnisse bei dieser Methode ist, daB aul3er der Gewichtszunahme durch die Enzymbindung keine weitere Ge- wichtsanderung durch andere Reaktionen, wie z. B. Hydrolyse der reaktiven Gruppen, Salzbildung, Adsorption etc. auftritt. Diese Voraussetzungen sind beim SWP in idealer Weise erfullt: hoher Gehalt des Tragers an inertem Poly- ethylen, sehr geringer Gehalt an reaktiven Gruppen und vergleichsweise hohe Enzymbindung. Nach beiden Proteinbestimmungsmethoden lieB sich eine Ubereinstimmung von etwa f 1 mg immobilisiertes Protein pro g Trager er- zielen. Die gravimetrische Proteinbestimmungsmethode ist gegeniiber der Fo- lin-Methode dann von Vorteil, wenn das zur Immobilisierung eingesetzte En- zym nicht einheitlich ist. So tritt z. B. im Fall von Papain, das nicht in hochge- reinigter Form eingesetzt wurde, eine deutliche Differenz bei beiden Bestim- mungsmethoden auf, wobei der Folin-Wert mit 147 mg Papain/g offensicht- lich zu hoch liegt, da die Gewichtszunahme nur einer Menge von 90 mg im- mobilisiertem Papain pro g SWP-R 830-IIa entspricht.
2.1 Charakterisierung des an S WP-R 830-IIa immobilisierten Trypsins
An SWP-R 830-IIa immobilisiertes Trypsin wurde hinsichtlich seiner ther- mischen Stabilitat und der Temperaturabhangigkeit der Aktivitat gegeniiber Na-Benzoyl-L-arginin-ethylester (BAEE) untersucht. Neben BAEE wurde als niedermolekulares Substrat auch Nm-Benzoyl-arginin-4-nitroanilid (BAPA) untersucht, das sowohl als L-BAPA als auch in der racemischen Form DL- BAPA eingesetzt wurde. Die enzymatischen Eigenschaften wurden weiterhin gegeniiber Casein als hochmolekularem Substrat untersucht. Weitere Untersu- chungen der Eigenschaften des immobilisierten Trypsins erfolgten gegenuber Sojabohnen-Trypsininhibitor, wodurch Informationen iiber die Zuganglich- keit der aktiven Zentren des immobilisierten Trypsins fur die Affmitatsadsorp- tion dieses hochmolekularen Inhibitors erhalten wurden.
47
G. Manecke und H.-G. Vogt
2 .1 .1 Thermische Stabil i tat und Temperaturabhangigkeit der Ak- tivitat von an SWP-R 830-IIa immobilisiertem Trypsin
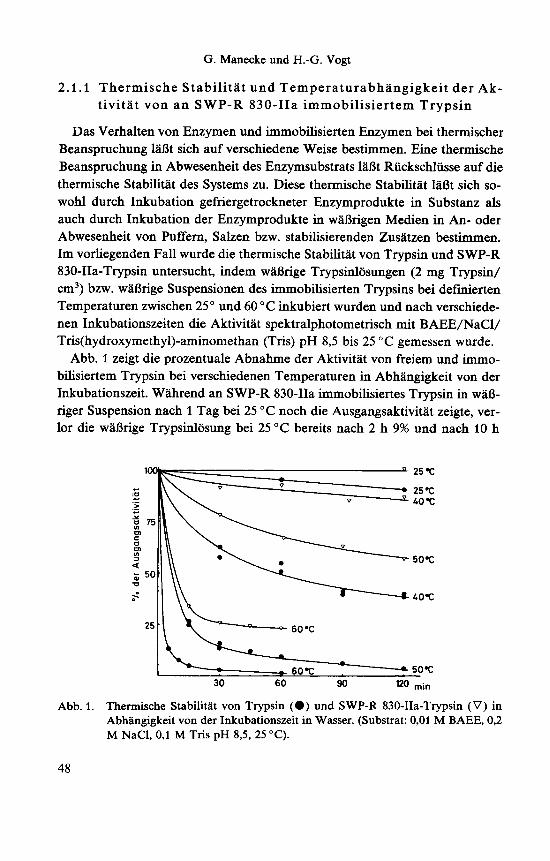
Das Verhalten von Enzymen und immobilisierten Enzymen bei thermischer Beanspruchung laBt sich auf verschiedene Weise bestimmen. Eine thermische Beanspruchung in Abwesenheit des Enzymsubstrats 1aBt Riickschliisse auf die thermische Stabilitat des Systems zu. Diese thermische Stabilitat laBt sich so- wohl durch Inkubation gefriergetrockneter Enzymprodukte in Substanz als auch durch Inkubation der Enzymprodukte in waBrigen Medien in An- oder Abwesenheit von Puffern, Salzen bzw. stabilisierenden Zusatzen bestimmen. Im vorliegenden Fall wurde die thermische Stabilitat von Trypsin und SWP-R 830-IIa-Trypsin untersucht, indem waBrige Trypsinlosungen (2 mg Trypsin/ cm') bzw. waBrige Suspensionen des immobilisierten Trypsins bei definerten Temperaturen zwischen 25 " und 60 "C inkubiert wurden und nach verschiede- nen Inkubationszeiten die Aktivitat spektralphotometrisch mit BAEE/NaCl/ Tris(hydroxymethy1)-aminomethan (Tris) pH 8,5 bis 25 " C gemessen wurde.
Abb. 1 zeigt die prozentuale Abnahme der Aktivitat von freiem und immo- bilisiertem Trypsin bei verschiedenen Temperaturen in Abhangigkeit von der Inkubationszeit. Wahrend an SWP-R 830-IIa immobilisiertes Trypsin in waB- riger Suspension nach 1 Tag bei 25 "C noch die Ausgangsaktivitat zeigte, ver- lor die waBrige Trypsinlosung bei 25 "C bereits nach 2 h 9% und nach 10 h
25.C
25.C V 40.C
50%
40%
a. 601: 50%
30 60 90 120 min
Abb. 1. Thermische Stabilitat von Trypsin (0) und SWP-R 830-IIa-Trypsin (V) in Abhangigkeit von der Inkubationszeit in Wasser. (Substrat: 0,Ol M BAEE, 0,2 M NaCl, 0,l M Tris pH 8,5, 25 "C).
48

An Synthesepulp immobilisierte Enzyme, I.
15% der Ausgangsaktivitat. Bei hoheren Inkubationstemperaturen nimmt das geloste Trypsin rasch an Aktivitat ab, das immobilisierte Trypsin zeigt hier eine deutlich erhohte thermische Stabilitat. Dieser Effekt ist jedoch hauptsach- lich auf die Verhinderung der Autolyse von Trypsin durch die Immobilisie- rung zuriickzufuhren. Wahrend die zeitliche Aktivitatsabnahme des freien Trypsins einem Zeitgesetz zweiter Ordnung folgt, was auf Inaktivierung infol- ge von Autolyse deutet, lieB sich fur die Inaktivierung von immobilisiertem Trypsin keine ganzzahlige Reaktionsordnung bestimmen. Die thermische Inaktivierung sollte einem Geschwindigkeitsgesetz 1. Ordnung folgen. Voraus- setzung hierfur ware allerdings, daB alle Enzymmolekiile in gleicher Weise und in der gleichen Mikroumgebung an die Matrix gebunden waren. Die Inaktivierung des immobilisierten Trypsins zeigt, daB diese Voraussetzungen nicht erfullt sind. Nach einer gewissen Inaktivierung zu Beginn der Inkuba- tionszeit verlauft die weitere Inaktivierung immer langsamer, als es die Kine- tik des Anfangsverlaufes erwarten 1aBt. Offenbar benotigen die verschieden gebundenen Trypsinmolekule unterschiedliche Energien fur ihre Denaturie- rung.
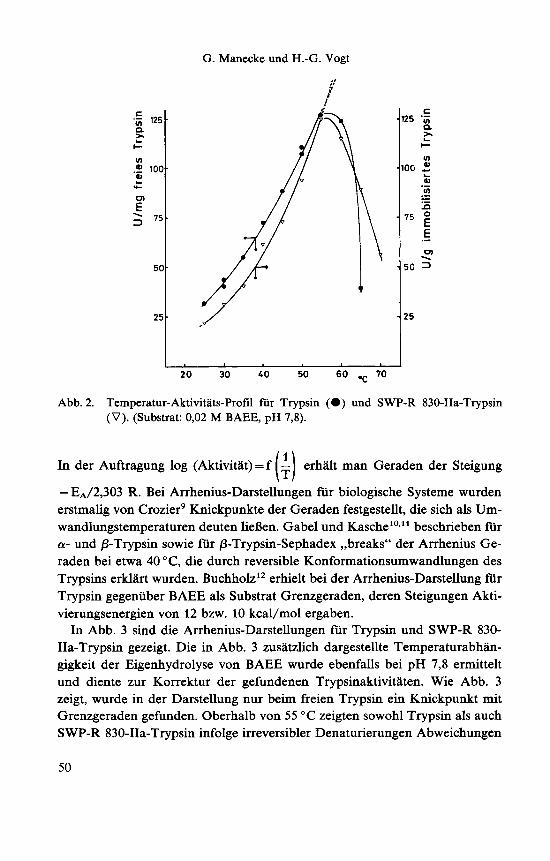
Setzt man ein Enzym oder immobilisiertes Enzym mit Substrat bei verschie- denen Temperaturen um, so laBt sich die Temperaturabhangigkeit der Aktivi- tat bestimmen. Die Aktivitaten von Trypsin und SWP-R 830-IIa-Trypsin wur- den n i t BAEE im Bereich 25-70 "C titrimetrisch bestimmt. Die jeweiligen An- fangsaktivitaten bei den Aktivitatsbestimmungen (abzuglich der jeweiligen Ei- genhydrolyse von BAEE) sind als Funktion der Temperatur in Abb. 2 darge- stellt. Abb. 2 zeigt sowohl fur das freie als auch fur SWP-R 830-IIa-Trypsin maximale Anfangsaktivitaten bei etwa 55 "C. Die Lage der Maxima ist zeitab- haingig*. Mit zunehmenden Reaktionszeiten bei den Aktivitatsbestimmungen erfolgt eine Verschiebung der Maxima zu tieferen Temperaturen. Die Maxima lassen sich dadurch erklaren, daB mit zunehmender Temperatur aui3er der Re- aktionsgeschwindigkeit fur den Substratumsatz auch die Reaktionsgeschwin- digkeit fur die irreversible Denaturierung und bei freiem Trypsin auch die der Autolyse des Enzyms zunehmen, so daB die wahren Anfangsgeschwindigkei- ten bei noch hoheren Temperaturen meBtechnisch nicht mehr faBbar sind. Die theoretischen Anfangsgeschwindigkeiten fur Temperaturen oberhalb von 55 "C sind in Abb. 2 als gestrichelte Kurve dargestellt.
Die Abhangigkeit der Enzymaktivitat von der Temperatur laBt sich durch die Arrhenius-Gleichung beschreiben:
EA Ink= - - + const. RT
49
G. Manecke und H.-G. Vogt
-
C -125 '5 n
F I-
ul -10'2 2
Q ul .- .- - .- n
E - 7 5 g
.- rn
50 3
125 I . I 20 30 40 50 60 .c 70
Abb. 2. Temperatur-Aktivitats-Profil fur Trypsin (0) und SWP-R 830-IIa-Trypsin (V). (Substrat: 0,02 M BAEE, pH 7,8).
In der Auftragung log (Aktivitat) = f - erhalt man Geraden der Steigung
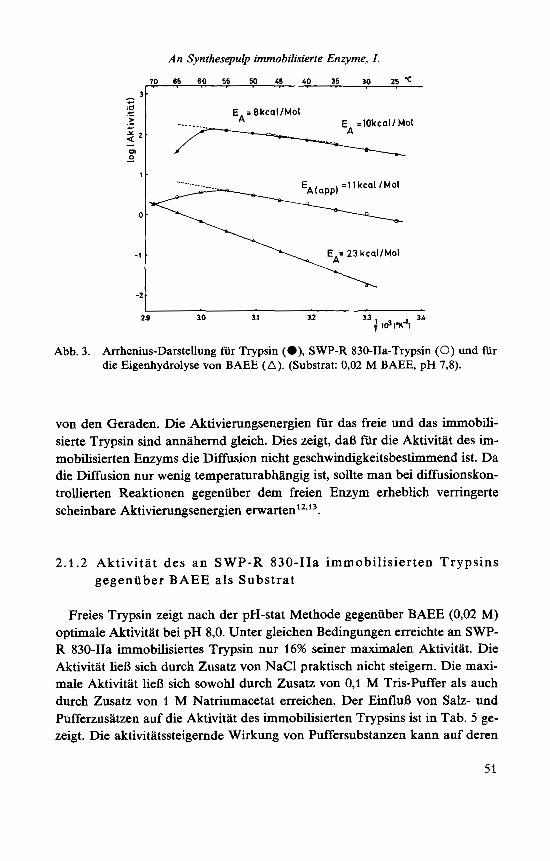
- EA/2,303 R. Bei Arrhenius-Darstellungen fur biologische Systeme wurden erstmalig von Crozier' Knickpunkte der Geraden festgestellt, die sich als Um- wandlungstemperaturen deuten lieBen. Gabel und Kasche'o*" beschrieben fur a- und p-Trypsin sowie fur p-Trypsin-Sephadex ,,breaks" der Arrhenius Ge- raden bei etwa 40 "C, die durch reversible Konformationsumwandlungen des Trypsins erklart wurden. Buchholz'* erhielt bei der Arrhenius-Darstellung fur Trypsin gegeniiber BAEE als Substrat Grenzgeraden, deren Steigungen Akti- vierungsenergien von 12 bzw. 10 kcal/mol ergaben.
In Abb. 3 sind die Arrhenius-Darstellungen fur Trypsin und SWP-R 830- IIa-Trypsin gezeigt. Die in Abb. 3 zusatzlich dargestellte Temperaturabhh- gigkeit der Eigenhydrolyse von BAEE wurde ebenfalls bei pH 7,8 ermittelt und diente zur Korrektur der gefundenen Trypsinaktivitaten. Wie Abb. 3 zeigt, wurde in der Darstellung nur beim freien Trypsin ein Knickpunkt mit Grenzgeraden gefunden. Oberhalb von 55 "C zeigten sowohl Trypsin als auch SWP-R 830-IIa-Trypsin infolge irreversibler Denaturierungen Abweichungen
($1
50
An Synthesepulp immobilisierte Enzyme, I .
70 65 60 55 50 45 40 35 30 25
--__ E I -.._ - -
'u -1 EA= 2 3 kcal/Mol
2 9 30 3.1 32 3 3 3L ; Id 1x41 Abb. 3. Arrhenius-Darstellung fur Trypsin (a), SWP-R 830-IIa-Trypsin (0) und fur
die Eigenhydrolyse von BAEE (A). (Substrat: 0,02 M BAEE, pH 7,8).
von den Geraden. Die Aktivierungsenergien fur das freie und das immobili- sierte Trypsin sind annahernd gleich. Dies zeigt, daR fur die Aktivitat des im- mobilisierten Enzyms die Diffusion nicht geschwindigkeitsbestimmend ist. Da die Diffusion nur wenig temperaturabhangig ist, sollte man bei diffusionskon- trollierten Reaktionen gegenuber dem freien Enzym erheblich verringerte scheinbare Aktivierungsenergien e r~ar ten '~ . '~ .
2.1.2 Aktivitat des an SWP-R 830-IIa immobilisierten Trypsins gegenuber BAEE als Substrat
Freies Trypsin zeigt nach der pH-stat Methode gegenuber BAEE (0,02 M) optimale Aktivitat bei pH 8,O. Unter gleichen Bedingungen erreichte an SWP- R 830-IIa immobilisiertes Trypsin nur 16% seiner maximalen Aktivitat. Die Aktivitat lieR sich durch Zusatz von NaCl praktisch nicht steigern. Die maxi- male Aktivitat lieR sich sowohl durch Zusatz von 0,l M Tris-Puffer als auch durch Zusatz von 1 M Natriumacetat erreichen. Der EinfluR von Salz- und Pufferzusatzen auf die Aktivitat des immobilisierten Trypsins ist in Tab. 5 ge- zeigt. Die aktivitatssteigernde Wirkung von Puffersubstanzen kann auf deren
51

G. Manecke und H.-G. Vogt
Tab. 5. EinfluB von NaC1, Natriumacetat und Tris-Puffer auf die Aktivitat von an SWP-R 830-IIa immobilisiertem Trypsin gegeniiber BAEE als Substrat.
Bestimmungs- Konzentrationen im Test Aktivitat bei methode BAEE NaCl CH3COONa Tris pH 8,O in
(mol/l) (mol/l) (mol/l) (mol/l) U/g SWP- Trypsin
~~ ~~
27 titrimetrisch 0,02 28 95 titrimetrisch 0,Ol - 0,Ol -
156 titrimetrisch 0,Ol -
spektralphoto- 0,Ol 02 - 0,l 149 metrisch
- - - - - titrimetrisch 0,02 0,4
- 1 ,oo
Pufferwirkung in der Mikroumgebung des immobilisierten Enzyms zuriickge- fuhrt werden, wodurch dort der durch Protonenanreicherung absinkende pH dem der AuBenlosung wieder angeglichen wird. Neben dieser statischen Funktion des Puffers postulierten Horvath und EngasserI4 eine dynamische Funktion von Puffersubstanzen. Puffer erleichtern danach den Transport von Protonen als auch von Sauren und Basen, die an der enzymatischen Reaktion beteiligt sind. Engasser und Horvath zeigten in einer theoretischen StudieI5, daB die Erleichterung des Protonentransportes uber den Bereich der Pufferka- pazitat des Saure-Basen-Paares hinaus mit dem pH der AuBenlosung zu- nimmt. Als Beispiel wurde der EinfluB von Acetat auf die Aktivitat von immo- bilisiertem Trypsin gegeniiber BAEE bei pH 7,8 be~chrieben'~. Acetat besitzt bei diesem pH keine nennenswerte Pufferkapazitat. Seine Wirkung wurde von den Autoren folgendermaBen erklart:
Die an der enzymatisch aktiven Oberflache gebildeten Protonen konnen ge- ma6 ihrem Konzentrationsgradienten frei in die Losung diffundieren. Alter- nativ konnen die Protonen in der Mikroumgebung des Enzyms durch Acetat- Ionen gebunden werden und als Essigsaure entlang ihrem eigenen Konzentra- tionsgradienten in die Losung diffundieren. Im Gleichgewichtszustand wird das Proton in der Makroumgebung wieder freigegeben, und Acetat-Ionen keh- ren zuriick an die Oberflache, und der Zyklus wiederholt sich. Insgesamt er- gibt sich somit eine alternative Transportmoglichkeit fur Protonen, so daB der Protonentransport erleichtert wirdI4.
Im vorliegenden Fall bewirkte 0,Ol M Natriumacetat eine Aktivitatssteige- rung auf das 3,5fache und 1 M Natriumacetat auf nahezu das 6fache, womit
52
An Synthesepulp irnmobilisierte Enzyme, I.
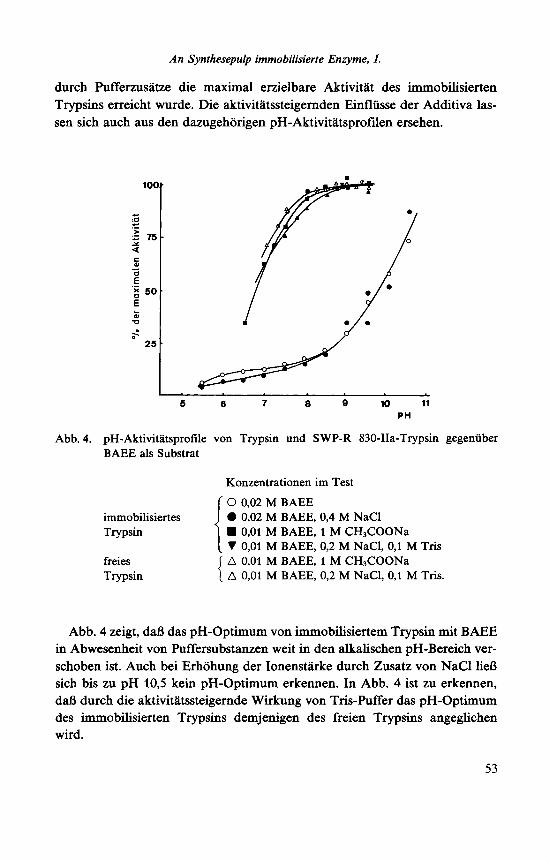
durch Pufferzusatze die maximal erzielbare Aktivitat des immobilisierten Trypsins erreicht wurde. Die aktivitatssteigernden Einfliisse der Additiva las- sen sich auch aus den dazugehorigen pH-Aktivitatsprofilen ersehen.
5 6 7 8 9 10 11 PH
Abb. 4. pH-Aktivitatsprofile von Trypsin und SWP-R 830-IIa-Trypsin gegeniiber BAEE als Substrat
Konzentrationen im Test
0 0,02 M BAEE 0 0,02 M BAEE, 0,4 M NaCl H 0,Ol M BAEE, 1 M CH,COONa V 0,Ol M BAEE, 0,2 M NaCI, 0,l M Tris A 0,Ol M BAEE, 1 M CH3COONa A 0,Ol M BAEE, 0,2 M NaC1, 0,l M Tris.
immobilisiertes Trypsin
freies Trypsin
Abb. 4 zeigt, daB das pH-Optimum von immobilisiertem Trypsin mit BAEE in Abwesenheit von Puffersubstanzen weit in den alkalischen pH-Bereich ver- schoben ist. Auch bei Erhohung der Ionenstarke durch Zusatz von NaCl lie6 sich bis zu pH 10,5 kein pH-Optimum erkennen. In Abb. 4 ist zu erkennen, daf3 durch die aktivitatssteigernde Wirkung von Tris-Puffer das pH-Optimum des immobilisierten Trypsins demjenigen des freien Trypsins angeglichen wird.
53
G. Manecke und H.-G. Vogt
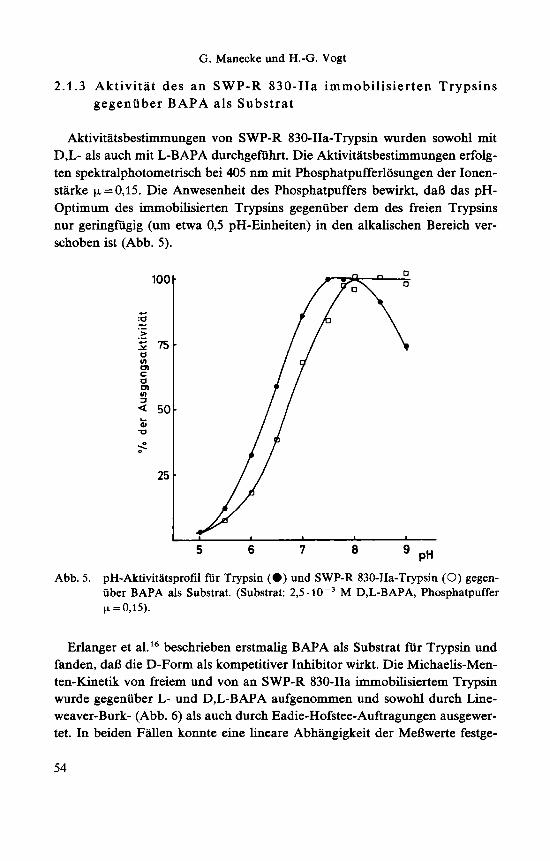
2.1.3 Aktivitat des an SWP-R 830-IIa immobilisierten Trypsins gegenuber BAPA als Substrat
Aktivitatsbestimmungen von SWP-R 830-IIa-Trypsin wurden sowohl mit D,L- als auch mit L-BAPA durchgefuhrt. Die Aktivitatsbestimmungen erfolg- ten spektralphotometrisch bei 405 nm mit Phosphatpufferlosungen der Ionen- starke p,=O,15. Die Anwesenheit des Phosphatpuffers bewirkt, daB das pH- Optimum des immobilisierten Trypsins gegenuber dem des freien Trypsins nur geringfugig (um etwa 0,5 pH-Einheiten) in den alkalischen Bereich ver- schoben ist (Abb. 5) .
5 6 7 8 PH Abb. 5. pH-Aktivitatsprofil fur Trypsin (0) und SWP-R 830-IIa-Trypsin (0) gegen-
uber BAPA als Substrat. (Substrat: 2,s. M D,L-BAPA, Phosphatpuffer p. = 0,15).
Erlanger et a1.I6 beschrieben erstmalig BAPA als Substrat fur Trypsin und fanden, daB die D-Form als kompetitiver Inhibitor wirkt. Die Michaelis-Men- ten-Kinetik von freiem und von an SWP-R 830-IIa immobilisiertem Trypsin wurde gegenuber L- und D,L-BAPA aufgenommen und sowohl durch Line- weaver-Burk- (Abb. 6) als auch durch Eadie-Hofstee-Auftragungen ausgewer- tet. In beiden Fallen konnte eine lineare Abhangigkeit der MeBwerte festge-
54
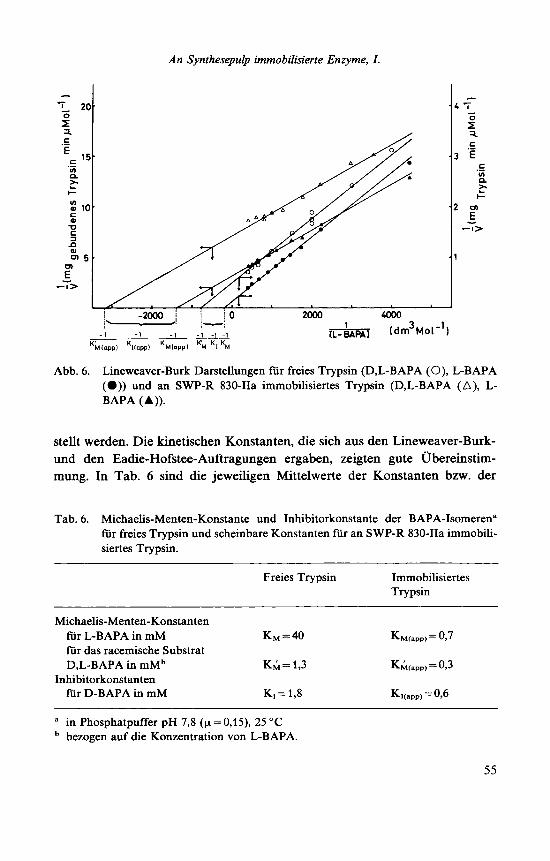
An Synthesepulp immobilisierte Enzyme, I .
a. c .- E c ' 5 .- II) P x I-
ul
c a2 TJ 3
al
L
al 10
n m 5
E" 1
-1 )
- 1 - 1 - 1 - 1 - 1 - 1 __ ~ - _-_ KMlappI Klloppl KMlappl Kl KM
- 4-i -
0 I 7 c
3 'E c II) P x I-
.-
L
2 c n E -
- 1 )
1
Abb. 6. Lineweaver-Burk Darstellungen fur freies Trypsin (D,L-BAPA (0), L-BAPA (a)) und an SWP-R 830-IIa immobilisiertes Trypsin (D,L-BAPA (A), L- BAPA I A )).
stellt werden. Die kinetischen Konstanten, die sich aus den Lineweaver-Burk- und den Eadie-Hofstee-Auftragungen ergaben, zeigten gute Ubereinstim- mung. In Tab. 6 sind die jeweiligen Mittelwerte der Konstanten bzw. der
Tab. 6. Michaelis-Menten-Konstante und Inhibitorkonstante der BAPA-Isomeren" fur freies Trypsin und scheinbare Konstanten fur an SWP-R 830-IIa immobili- siertes Trypsin.
Freies Trypsin Immobilisiertes Trypsin
Michaelis-Menten-Konstanten fur L-BAPA in mM KM = 40 K~i(app)=0,7 fur das racemische Substrat D,L-BAPA in mMb K& = 1,3 KM(,pp, = O J
fur D-BAPA in mM KI = 1,8 Ki(,pp, = 096 Inhibitorkonstanten
a in Phosphatpuffer pH 7,8 (p = 0,15), 25 "C bezogen auf die Konzentration von L-BAPA.
55

G. Manecke und H.-G. Vogt
scheinbaren Konstanten angegeben. Die Werte fur die scheinbare Michaelis- Menten-Konstante (KMM(app)) als auch fur die scheinbare Inhibitorkonstante
sind beim immobilisierten Trypsinderivat niedriger als beim freien Trypsin. Eine mogliche Erklarung hierfur ware, daR sowohl das Substrat als auch der Inhibitor in der Mikroumgebung des immobilisierten Trypsins even- tuell in erhohter Konzentration vorliegen.
2.1.4 Aktivitat des an SWP-R 830-IIa und SWP-E 400-IIa immo- bilisierten Trypsins gegenuber Casein als Substrat
Die Bestimmungen der caseinolytischen Aktivitat von Trypsin und immobi- lisiertem Trypsin wurden in Anlehnung an die fur freies Trypsin angewandte Methode nach Bergmeyerl' durchgefw. Um die Wirksamkeit des immobili- sierten Trypsins mit der des freien Trypsins gegeniiber diesem Substrat ver- gleichen zu konnen, wurde zunachst die Aktivitat des freien Trypsins gegen- uber Casein ermittelt. Hierzu wurden 1 proz. Caseinlosungen in Phosphatpuf- fer pH 8,0 (Ionenstarke p = 0,15) mit 0-100 pg gelostem Trypsin 20 min bei 37 "C inkubiert, mit 5prOZ. CC13COOH gefallt und die Absorption der abzen- trifugierten Losung bei 280 nm gemessen. Aus den Absorptionen bei 280 nm und der eingesetzten Menge Trypsin wurde eine Eichkurve erstellt, die im un- teren Bereich der Trypsinkonzentrationen einen linearen Verlauf zeigte. Pro- ben des immobilisierten Trypsins wurden unter Ruhren in entsprechender Weise mit Casein umgesetzt und die Enzymreaktion nach 20 min durch Ab- saugen der Losung beendet. Nach der Fallung von Casein wurden die Absorp- tionen der Losung bei 280 nm gemessen und aus der Eichkurve die diesen Ab- sorptionen entsprechenden Mengen freien Trypsins entnommen. Die relativen Aktivitaten ergeben sich aus dem Verhaltnis der so bestimmten Enzymmenge zum Trypsingehalt der eingesetzten immobilisierten Trypsinderivate.
In Tab. 7 sind die caseinolytischen Aktivitaten von Trypsin, immobilisiert an SWP-R 830-IIa und SWP-E 400-IIa, gezeigt. Die caseinolytischen Aktivita- ten beider Trypsin-Derivate sind etwa gleich, gleiche Einwaagen beider immo- bilisierter Trypsinderivate bewirkten bei dieser Bestimmungsmethode etwa gleiche Absorptionsanderungen. Die besseren relativen Aktivitaten von SWP- E 400-IIa-Trypsin sind wahrscheinlich auf den geringeren Trypsingehalt und die damit verbundenen geringeren sterischen Behinderungen zuriickzufuhren. Die relativen Aktivitaten bei mehreren aufeinanderfolgenden Bestimmungen nahmen ab, obwohl die verwendeten immobilisierten Trypsinderivate bei Um- setzungen mit niedermolekularen Substraten sich sehr stabil verhielten. Es ist
56

An Synthesepub immobilisierte Enzyme, I.
Tab. 7. Aktivitat von an Synthesepulp SWP-R 830-IIa und SWP-E 400-IIa immobili- siertem Trypsin gegeniiber Casein als Substrat.
SWP-R SWP-E 830-IIa-Trypsin 400-IIa-Trypsin
immobilisiertes Trypsin in
relative Aktivitat gegeniiber
relative Aktivitat bei mehr- fachen Bestimmungen mit
mg/g SWP-Derivat 41
BAEE (96) 19,6
Casein (%) 1. Bestimmung 0,61 2. Bestimmung 0,65 3. Bestimmung 0,43 4. Bestimmung 0,39
15
9,O
2,55 1,97 1,58 1 4
daher anzunehmen, daf3 die Abnahme der relativen Aktivitaten durch Ausfal- lungen von Casein bzw. Caseinabbauprodukten in der Mikroumgebung des immobilisierten Trypsins hervorgerufen wird. Ahnliche Befunde wurden auch von ReimerdesI8 beschrieben.
2.1.5 Das Verhalten von an SWP-R 830-IIa immobilisiertem Trypsin gegeniiber Sojabohnen-Trypsininhibitor
Sojabohen-Trypsininhibitor (STI) ist ein makromolekularer Inhibitor mit einem Molekulargewicht von M = 22OOOI9, der mit Trypsin einen 1 : 1 Kom- plex bildet. 1 mg STI vermochte 1,15 mg freies Trypsin (Merck) vollstandig zu inhibieren. Die Affinitatsadsorption des Inhibitors wurde an SWP-R 830-IIa- Trypsin mit iiberschussigem STI in Phosphatpuffer pH 8,O (Ionenstarke p. = 0,15) unter einstundigem Ruhren bei Raumtemperatur durchgefuhrt. Uberschiissiges STI wurde mit Phosphatpuffer pH 8,O ausgewaschen und die Waschlosungen gesammelt. Die Desorption wurde portionsweise mit 0,001 M HC1 unter Ruhren bei Raumtemperatur durchgefuhrt, wobei die Eluate eben- falls gesammelt wurden. Der STI-Gehalt der Waschlosungen wurde bestimmt. Die affinitatsgebundene STI-Menge wurde aus diesen Werten berechnet, es lieBen sich pro g SWP-R 830-IIa-Trypsin (Trypsingehalt: 32,6 mg/g Immobili- sierungsprodukt) 0,5 mg STI reversibel binden. Dies entspricht einer relativen
57
G. Manecke und H.-G. Vogt
Aktivitat des immobilisierten Trypsins gegeniiber STI von 1,5%. Im Gegensatz zur Aktivitat gegenuber Casein nimmt die Akthitat gegeniiber STI nicht bei wiederholten Bestimmungen ab. Der adsorbierte Inhibitor lieB sich quantitativ mit verdiinnter HC1 wieder desorbieren.
2.2 Immobilisierung von Sojabohnen-Trypsininhibitor (STI) an S WP-R 830- IIa
Es wurde nun umgekehrt der Inhibitor, der selbst ein Protein ist, immobili- siert und dessen Aff~tatsaktivitat untersucht. Fur die Azokupplung von STI an SWP-R 830-IIa wurde eine 0,lproz. Losung von STI in Phosphatpuffer pH 8,0 eingesetzt. Es lieBen sich 37,7 mg STI pro g SWP-R 830-IIa immobilisie- ren. Die EiweiBbestimmung mit Folin-Reagenz und der gravimetrisch aus der Gewichtszunahme ermittelte Wert zeigte Ubereinstimmung. Die Bindungs- ausbeute lag bei 76%.
Zur Untersuchung der Inhibitoreigenschaften des immobilisierten STI wur- de dieser bei pH 8,0 in Phosphatpuffer mit Trypsin unter Riihren bei Raum- temperatur umgesetzt. Abb. 7 zeigt den Verlauf fur die Adsorption von Tryp- sin. Die Adsorptionsreaktion war unter den gegebenen Bedingungen praktisch
0 10 20 30 0 10 20 min
Abb. 7. Adsorption (A) und Desorption (B) von Trypsin an SWP-R 830-IIa-Tryp- sininhibitor in Abhangigkeit von der Reaktionszeit.
58

An Synthesepulp immobilisierte Enzyme, I.
nach 20 min abgeschlossen. Die Halbwertzeit fur die Adsorption sowie auch fur die Desorption - Abb. 7 - betrug jeweils etwa 0,5 min. Die von SWP-R 830-IIa-STI komplexierte Menge Trypsin wurde aus EiweiBbestimmungen mit Folin-Reagenz sowie aus der Aktivitat des desorbierten Trypsins gegen- uber BAEE ermittelt. Es konnte festgestellt werden, da8 pro g eingesetztes SWP-R 830-IIa-STI 6 mg Trypsin reversibel gebunden werden konnten. Die von 1 g SWP-R 830-IIa-STI inhibierte Trypsinaktivitat betrug 240 U (u G pmol Substratumsatz pro min), was bei einer spezifischen Aktivitat des eingesetzten Trypsins von 42,5 U/mg einen gut ubereinstimmenden Wert fur die Trypsinbindung ergibt. 1 mg freies STI vermochte 1,15 mg Trypsin zu in- hibieren. Verglichen mit dem freien STI besitzt das immobilisierte STI eine re- lative Inhibitorwirkung von 14%. In Tab. 8 sind die Ergebnisse dieser Untersu- chungen zusammengefafit.
Tab. 8. Bildung des Trypsin-Sojabohnen-Trypsininhibitor (ST1)-Komplexes an dem SWP-R 830-IIa-Trypsin und an dem SWP-R 830-STI.
SWP-R 830-IIa- SWP-R 330-IIa- Trypsin STI
Molekulargewicht des immobilisierten Pro-
kovalent gebundenes Protein teins
in mg/g Immobilisierungsprodukt in p,mol/g Immobilisierungsprodukt
affinitats-adsorptiv gebundenes STI in mg/g SWP-R 830-IIa-Trypsin
affinitats-adsorptiv gebundenes Trypsin in mg/g SWP-R 830-IIa-STI
Aktivitat von immobilisiertem Trypsin gegen- uber BAEE" in U/g SWP-R 830-IIa-Tryp- sin
Inhibitoraktivitat von immobilisiertem STI in BAEE-Einheiten in U/g SWP-R 830-IIa-STI
relative Aktivitat von SWP-R 830-IIa-Trypsin gegenuber
BAEE in % STI in %
relative Aktivitat von immobilisiertem STI ge- geniiber Trypsin in %
23 400 22 OOO
32,6 36,3 1,39 1,65
6
149
240
12 1.5
14
a 0,Ol M BAEE, 0,2 M NaC1, 0,l M Tris-Puffer pH 8,0, 25 "C.
59
G. Manecke und H.-G. Vogt
Die reversible Bindung von Trypsin an SWP-R 830-IIa-STI wurde auBer im ,,batch-Verfahren" auch im Saulenverfahren untersucht. Die DurchfluBge- schwindigkeit durch die Saule wurde so gewahlt, da8 sich bei der Adsorption eine Verweilzeit des Trypsins in der Saule von ca. 1 min ergab. Wie erwartet, konnte unter diesen experimentellen Bedingungen die volle Bindungskapazi- tat nicht ausgeschopft werden.
2.3 Eigenschaften und pH-Aktivitatsprofile von an S WP-R 830-IIa-c immobili- siertem Chymotrypsin, Papain und Urease
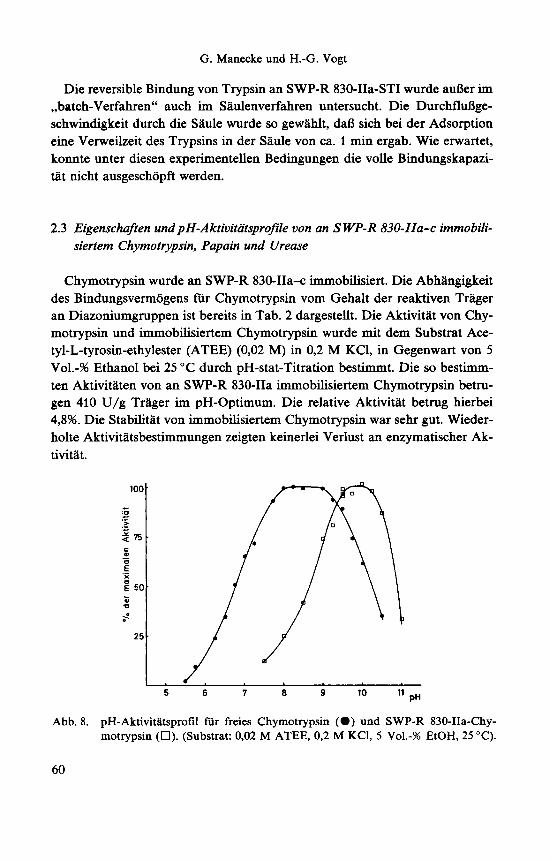
Chymotrypsin wurde an SWP-R 830-IIa+ immobilisiert. Die Abhangigkeit des Bindungsvermogens fur Chymotrypsin vom Gehalt der reaktiven Trager an Diazoniumgruppen ist bereits in Tab. 2 dargestellt. Die Aktivitat von Chy- motrypsin und immobilisiertem Chymotrypsin wurde mit dem Substrat Ace- tyl-L-tyrosin-ethylester (ATEE) (0,02 M) in 0,2 M KC1, in Gegenwart von 5 Vo1.-% Ethanol bei 25 "C durch pH-stat-Titration bestimmt. Die so bestimm- ten Aktivitaten von an SWP-R 830-IIa immobilisiertem Chymotrypsin betru- gen 410 U/g Trager im pH-Optimum. Die relative Aktivitat betrug hierbei 4,8%. Die Stabilitat von immobilisiertem Chymotrypsin war sehr gut. Wieder- holte Aktivitatsbestimmungen zeigten keinerlei Verlust an enzymatischer Ak- tivitlt.
5 6 7 0 9 10 11 pH
Abb. 8. pH-Aktivitatsprofil fur freies Chymotrypsin (0) und SWP-R 830-IIa-Chy- motrypsin (0). (Substrat: 0,02 M ATEE, 0,2 M KCI, 5 Vo1.-% EtOH, 25 "C).
60

An Synthesepulp immobilisierte Enzyme, I.
Abb. 8 zeigt das pH-Aktivitatsprofil von Chymotrypsin und SWP-R 830- IIa-Chymotrypsin. Das pH-Optimum des immobilisierten Chymotrypsins liegt bei pH 10 und ist somit gegeniiber dem pH-Optimum von freiem Chymotryp- sin um etwa 1,5 pH-Einheiten in den alkalischen Bereich verschoben. Da Chy- motrypsin ATEE zu N-Acetyl-L-tyrosin, also zu einer Saure, umsetzt, ist auch hier die pH-Optimumsverschiebung durch Produktakkumulierung zu erkla- ren. Da sowohl das Produkt der enzymatischen Reaktion als auch das Enzym bei pH-Werten oberhalb von 9 negativ geladen sind, ist eine elektrostatische AbstoBung zwischen immobilisiertem Enzym und Produkt denkbar, die einer Produktakkumulierung entgegenwirkt. Dies mag dazu beitragen, daR das pH- Optimum des immobilisierten Chymotrypsins nicht so drastisch in den alkali- schen pH-Bereich verschoben ist, wie das des immobilisierten Trypsins. Beim letzteren wurde die pH-Abhhgigkeit mit BAEE bestimmt, dessen Produkt in diesem pH-Bereich zwitterionisch vorliegt.
Als weitere Hydrolase wurde Papain an Diazoniumderivate des Synthese- pulps SWP-R 830 immobilisiert. Von allen untersuchten Enzymen fuhrte die Immobilisierung durch Azokupplung bei Papain zu den hochsten Bindungs- mengen. Gravimetrisch lien sich eine Papainaufnahme von 90 mg/g SWP-R 830-IIa ermitteln. Die relative Aktivitat gegeniiber BAEE als Substrat betrug bei pH 6 14,7% und im pH-Optimum @H 7 3 ) 18,4%. Die Papainimmobilisie- rung an SWP-R 830-IIc, dem SWP-R 830-11-Trager mit dem niedrigsten Ge- halt an Diazoniumgruppen, ergab gravimetrisch bestimmte Bindungswerte von 28 mg Papain/g (15,5% relative Aktivitat bei pH 6). Die immobilisierten Papainderivate zeigten keinerlei Verlust an enzymatischer Aktivitat nach wie- derholten Umsetzungen mit dem Substrat BAEE.
Abb. 9 zeigt das pH-Aktivitatsprofil von Papain und SWP-R 830-IIa-Pa- pain. Obwohl hier das gleiche Substrat, BAEE, wie bei immobilisiertem Tryp- sin verwendet wurde, traten hier keine extremen Verschiebungen des pH-Op- timums auf. Das pH-Optimum des immobilisierten Papains lag bei pH 7,25- 7 3 . Verglichen mit immobilisiertem Trypsin ist diese geringe Verschiebung des pH-Optimums auf die Anwesenheit von Cystein und EDTA bei den Akti- vitatsbestimmungen von immobilisiertem Papain zuriickzufuhren. Cystein und EDTA konnen nach Engasser und H o r ~ a t h ' ~ den Transport von Protonen und BAEE erleichtern und wirken somit einer Verschiebung der pH-Optima infolge von Produktakkumulation entgegen.
Obwohl Cystein und EDTA in verhaltnismaBig niedriger Konzentration vorliegen, vermogen sie offensichtlich wirkungsvoll eine groJ3ere Verschiebung der pH-Optima in den alkalischen pH-Bereich zu verhindern. Entsprechende
61
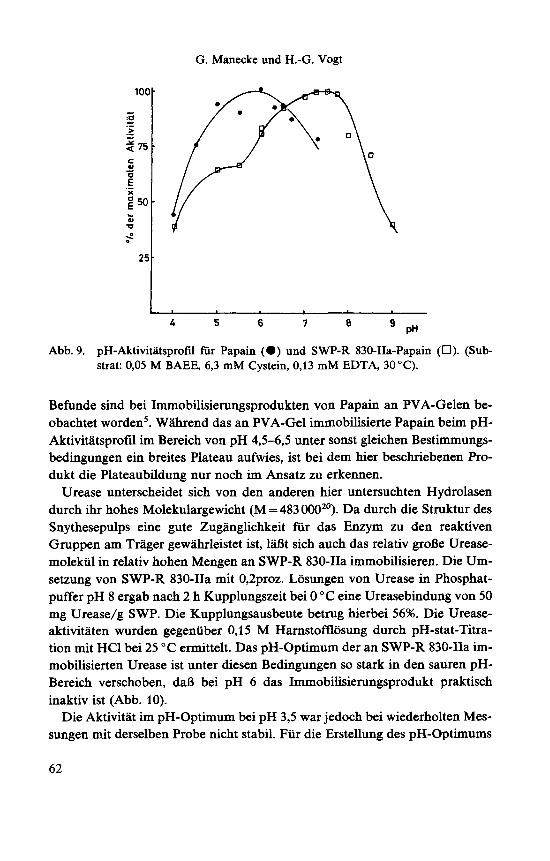
G. Manecke und H.-G. Vogt
9 P H 4 5 6 7 8
Abb. 9. pH-Aktivitatsprofil fur Papain (0) und SWP-R 830-IIa-Papain (0). (Sub- strat: 0,05 M BAEE, 6,3 mM Cystein, 0,13 mM EDTA, 30 "C).
Befmde sind bei Immobilisierungsprodukten von Papain an PVA-Gelen be- obachtet worden5. Wiihrend das an PVA-Gel immobilisierte Papain beim pH- Aktivitatsprofil im Bereich von pH 4 , 5 4 5 unter sonst gleichen Bestimmungs- bedingungen ein breites Plateau aufwies, ist bei dem hier beschriebenen Pro- dukt die Plateaubildung nur noch im Ansatz zu erkennen.
Urease unterscheidet sich von den anderen hier untersuchten Hydrolasen durch ihr hohes Molekulargewicht (M = 483 OOOZo). Da durch die Struktur des Snythesepulps eine gute Zugihglichkeit fur das Enzym zu den reaktiven Gruppen am Trager gewahrleistet ist, l33t sich auch das relativ grol3e Urease- molekul in relativ hohen Mengen an SWP-R 830-IIa immobilisieren. Die Um- setzung von SWP-R 830-IIa mit 0,2proz. Losungen von Urease in Phosphat- puffer pH 8 ergab nach 2 h Kupplungszeit bei 0 "C eine Ureasebindung von 50 mg Urease/g SWP. Die Kupplungsausbeute betrug hierbei 56%. Die Urease- aktivitaten wurden gegenuber 0,15 M Hamstofflosung durch pH-stat-Titra- tion mit HCl bei 25 "C ermittelt. Das pH-Optimum der an SWP-R 830-IIa im- mobilisierten Urease ist unter diesen Bedingungen so stark in den sauren pH- Bereich verschoben, daB bei pH 6 das Immobilisierungsprodukt praktisch inaktiv ist (Abb. 10).
Die Aktivitat im pH-Optimum bei pH 3,5 war jedoch bei wiederholten Mes- sungen mit derselben Probe nicht stabil. Fur die Erstellung des pH-Optimums
62
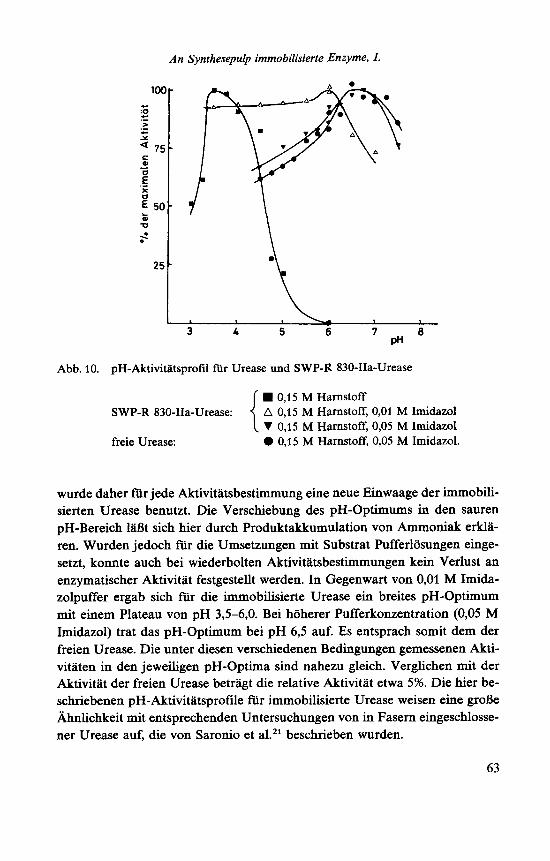
An Synthesepulp immobilisierte Enzyme, I.
Abb. 10. pH-Aktivitatsprofil fur Urease und SWP-R 830-IIa-Urease
0,15 M Harnstoff A 0,15 M Harnstoff, 0,Ol M Imidazol V 0,15 M Harnstoff, 0,05 M Imidazol 0 0,15 M Harnstoff, 0,05 M Imidazol.
SWP-R 830-IIa-Urease:
freie Urease:
wurde daher fur jede Aktivitatsbestimmung eine neue Einwaage der immobili- sierten Urease benutzt. Die Verschiebung des pH-Optimums in den sauren pH-Bereich la& sich hier durch Produktakkumulation von Ammoniak erkla- ren. Wurden jedoch fur die Umsetzungen mit Substrat Pufferlosungen einge- setzt, konnte auch bei wiederholten Aktivitatsbestimmungen kein Verlust an enzymatischer Aktivitat festgestellt werden. In Gegenwart von 0,Ol M Imida- zolpuffer ergab sich fur die immobilisierte Urease ein breites pH-Optimum mit einem Plateau von pH 3,5-6,0. Bei hoherer Pufferkonzentration (0,05 M Imidazol) trat das pH-Optimum bei pH 6,5 auf. Es entsprach somit dem der freien Urease. Die unter diesen verschiedenen Bedingungen gemessenen Akti- vitaten in den jeweiligen pH-Optima sind nahezu gleich. Verglichen mit der Aktivitat der freien Urease betragt die relative Aktivitat etwa 5%. Die hier be- schriebenen pH-Aktivitatsprofde fur immobilisierte Urease weisen eine groBe Ahnlichkeit mit entsprechenden Untersuchungen von in Fasern eingeschlosse- ner Urease auf, die von Saronio et a1.21 beschrieben wurden.
63
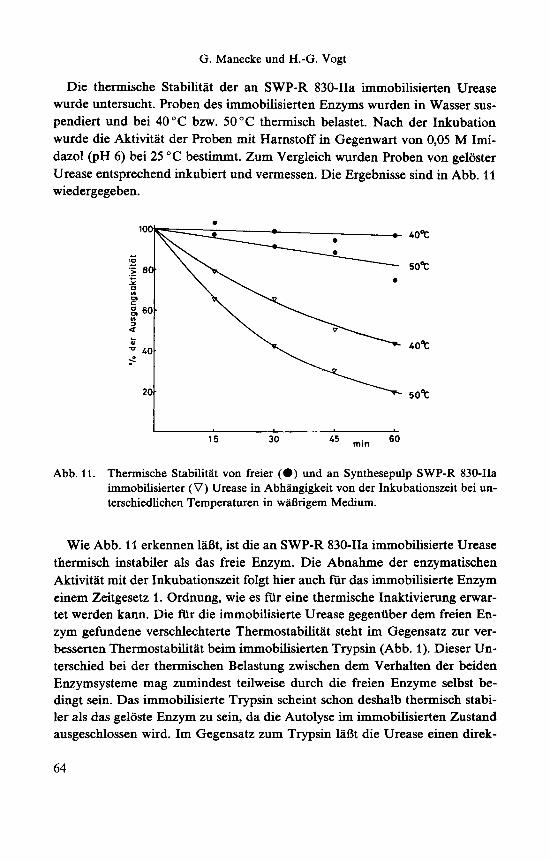
G. Manecke und H.-G. Vogt
Die thermische Stabilitat der an SWP-R 830-IIa immobilisierten Urease wurde untersucht. Proben des immobilisierten Enzyms wurden in Wasser sus- pendiert und bei 40°C bzw. 50°C thermisch belastet. Nach der Inkubation wurde die Aktivitat der Proben mit Harnstoff in Gegenwart von 0,05 M Imi- dazol @H 6) bei 25 "C bestitnmt. Zum Vergleich wurden Proben von geloster Urease entsprechend inkubiert und vermessen. Die Ergebnisse sind in Abb. 11 wiedergegeben.
&Ot
2ot
40%
50%
40%
50%
1 15 30 45 60
Abb. 11. Thermische Stabilitat von freier (0) und an Synthesepulp SWP-R 830-IIa immobilisierter (V) Urease in Abhangigkeit von der Inkubationszeit bei un- terschiedlichen Temperaturen in wiinrigem Medium.
Wie Abb. 11 erkennen lafit, ist die an SWP-R 830-IIa immobilisierte Urease thermisch instabiler als das freie Enzym. Die Abnahme der enzymatischen Aktivitat mit der Inkubationszeit folgt hier auch fur das immobilisierte Enzym einem Zeitgesetz 1. Ordnung, wie es fur eine thermische Inaktivierung erwar- tet werden kann. Die fur die immobilisierte Urease gegeniiber dem freien En- zym gefundene verschlechterte Thermostabilitat steht im Gegensatz zur ver- besserten Thermostabilitat beim immobilisierten Trypsin (Abb. 1). Dieser Un- terschied bei der thermischen Belastung zwischen dem Verhalten der beiden Enzymsysteme mag zumindest teilweise durch die freien Enzyme selbst be- dingt sein. Das immobilisierte Trypsin scheint schon deshalb thermisch stabi- ler als das geloste Enzym zu sein, da die Autolyse im immobilisierten Zustand ausgeschlossen wird. Im Gegensatz zum Trypsin laBt die Urease einen direk-
64

An Synthesepulp immobilisierte Enzyme, I.
ten Vergleich der Thermostabilitat des freien und des immobilisierten Enzyms zu. Die gemessenen Thermostabilitaten gelten nur fur die in waBrigem Medi- um untersuchten Enzyme; in der gefriergetrockneten Form ist immobilisierte Urease weitaus thermostabiler als im waBrigen Medium.
2.4 Eigenschaften weiterer an S WP-R 830-IIa immobilisierten Enzyme
Des weiteren wurden an das Synthesepulpderivat SWP-R 830-IIa die nach- folgenden Enzyme immobilisiert:
Die Hydrolasen: Aminoacylase und Esterase die Oxidoreduktasen:
und die Transferase: Hexokinase. Bis auf Glucose-6-phosphat-dehydrogenase und Hexokinase wurde das Bin-
dungsvermogen des reaktiven Diazoniumderivates SWP-R 830-IIa fur die be- schriebenen Enzyme mit Folin-Reagenz bzw. gravimetrisch bestimmt (Tab. 4). Die Immobilisierung der Aminoacylase wurde in Phosphatpuffer (PH 8,O; p =0,15) in Gegenwart von 1,5. lo-’ M CoClz durchgefuhrt. Von 100 mg ein- gesetzter Aminoacylase pro g SWP-R 830-IIa lieBen sich aus einer 0,2 proz. Losung des Enzyms 59 mg Aminoacylase pro g SWP-R 830-IIa immobilisie- ren. Die Aktivitaten von Aminoacylase und SWP-R 830-IIa-Aminoacylase wurden gegeniiber dem Substrat N-Acetyl-DL-methionin bestimmt. Die Akti- vitat von an SWP-R 830-IIa immobilisierter Aminoacylase betrug etwa 1 U/g SWP (0,196 relative Aktivitat).
Esterase (aus Bacillus subtilis) wurde in 0,l proz. Losung in Phosphatpuffer (pH 8,O) fur die Immobilisierung an SWP-R 830-IIa eingesetzt. Pro g Synthe-
mobilisieren. Die Bestimmung der Esteraseaktivitat wurde gegeniiber DL- Acetylpantolacton nach der pH-stat-Methode vorgenommen. Da fur das pH- Optimum der immobilisierten Esterase eine Verschiebung in den alkalischen Bereich zu erwarten war, wurden die Aktivitatsbestimmungen in Gegenwart von 1,3. lop3 M Boratpuffer pH 8 durchgefuhrt, um das AusmaB der pH-Op- timumsverschiebung zu verringern. Die um die Eigenhydrolyse komgierten Aktivitaten der immobilisierten Esterase betrugen 60-80 U/g Immobilisie- rungsprodukt, die relative Aktivitat betrug somit etwa 2,5%.
12,5 mg Glucose-Oxydase (GOD) wurden als 0,025 proz. Losung des En- zyms in Phosphatpuffer pH 8,0 (p=O,15) pro g SWP-R 820-IIa fur die Kupp-
Glucose-Oxydase, Katalase, Peroxydase, Glucose-6- phosphat-dehydrogenase
sepulp lieBen sich 42 mg Esterase bei einer Kupplungsausbeute von 43Y 0 lm- ’
65

G. Manecke und H.-G. Vogt
lungsreaktion eingesetzt. Bei einer Kupplungszeit von 2 h bei 0°C wurden 87% der eingesetzten GOD an den Trager fmiert. Die Bestimmungen der enzy- matischen Aktivitaten erfolgten nach einer modif~erten Methode3 nach Wer- ner et a1.22. SWP-R 830-IIa-GOD zeigte Aktivitaten von 10-20 U/g Immobili- sierungsprodukt (0,4-0,8% relative Aktivitat).
Peroxydase wurde als 0,23 proz. und Katalase als 0,05 proz. Losung in Phos- phatpuffer pH 8 an SWP-R 830-IIa immobilisiert. Es lienen sich 43 mg Per- oxydase bzw. 22 mg Katalase pro g SWP-R 830-IIa immobilisieren. Die Kupplungsausbeute betrug bei Peroxydase 43%, Katalase wurde infolge des geringen Enzymangebots nahezu quantitativ gebunden (Kupplungsausbeute 95%). Die Aktivitat beider immobilisierten Enzyme wurde gegeniiber H202, bei Peroxydase in Gegenwart von 2,2'-Azino-di-(3-ethylbenzthiazolin-6-sulfo- nat) (ABTS) als Wasserstoff-Donator, bestimmt. Pro g Immobilisierungspro- dukt betrug die Peroxydase-Aktivitat ca. 7 U (relative Aktivitat 0,04%) bzw. die Katalase-Aktivitat 1 300 U (relative Aktivitat 0,1%).
Weiterhin wurden an SWP-R 830-IIa die Enzyme Hexokinase (HK) und Glucose-6-phosphatdehydrogenase (G-6-PDH) coimmobilisiert. Hierzu wur- den pro g des diazotierten Tragers 13 0oO U HK und 1 500 U G-6-PDH einge- setzt, was einem Gesamtproteinangebot von ca. 100 mg entspricht. Die Coim- mobilisierung wurde in 0,5 M Triethanolamin-Puffer pH 8,O bei 25°C 2 h durchgefuhrt. Das Coimmobilisierungsprodukt zeigte eine HK-Aktivitat von 12,6 U/g, was einer Aktivitatsausbeute von 0,1% entspricht, und eine G-6- PDH-Aktivitat von 8,9 U/g, entsprechend einer Aktivitatsausbeute von 0,696. Die gemeinsame Aktivitat des coimmobilisierten Zweienzymsystems betrug
In der vorliegenden Arbeit wurde die kovalente Enzymimmobilisierung an Synthesepulp durch Azokupplung beschrieben. Weitere Methoden zur kova- lenten Bindung von Enzymen an dieses Polymerprodukt sind Gegenstand ei- ner weiteren Veroffentlichung.
494 u/g.
3. Experimenteller Teil
Herkunft der verwendeten Substanzen: Synthesepulp SWP"-R 830 und SWP"-E 400 (Hoechst-Zellerbach); Papain, EC
3.4.22.2 (Merck, Kat. Nr. 7144); Trypsin, EC 3.4.21.4 (Boehringer/Mannheim, Kat. Nr. 109819; Merck, Kat. Nr. 24579), a-Chymotrypsin, EC 3.4.21.1 (Boehringer/Mannheim); Aminoacylase EC 3.5.1.14 (Boehringer/Mannheim); Esterase (aus Bacillus subtilis) EC
66

An Synthesepulp immobilisierte Enzyme, I.
3.1.1.6 (BoehringedMannheim); Urease EC 3.5.1.5 (Boehringer/Mannheim); L-Amino- saure-Oxydase, EC 1.4.3.2 (L-AOD, Boehringer/Mannheim); Glucose-Oxydase, EC 1.1.3.4 (GOD, Grad I, Boehringer/Mannheim); Katalase, EC 1.11.1.6 (Boehringer/ Mannheim, Kat. Nr. 106810); Peroxydase, EC 1.11.1.7 (POD, Boehringer/Mannheim Kat. Nr. 15301 und 15302); Glucose-6-phosphat-dehydrogenase, EC 1.1.1.49 (Boehrin- ger/Mannheim, Kat. Nr. 197726); Hexokinase, EC 2.7.1.1 (Boehringer/Mannheim, Kat. Nr. 208540); Sojabohnen-Trypsininhibitor (Worthington); Casein nach Hammarsten (Merck); N*-Benzoyl-L-arginin-ethylester . HCI (BAEE, Merck); N*-Benzoyl-D,L-argi- nin-4-nitroanilid. HCl (D,L-BAPA, Boehringer/Mannheim); Na-Benzoyl-L-arginin-4- nitroanilid (L-BAPA, Merck); Acetyl-L-tyrosin-ethylester (ATEE, Merck); Acetyl-DL- methionin (Merck); Acetyl-D,L-pantolacton (BASF); Glucose-6-phosphat (Boehringer/ Mannheim); Nicotinamid-adenin-dinucleotidphosphat (NADP, Boehringer/Mann- heim, Kat. Nr. 127353); Adenosin-5'-triphosphat (ATP, Boehringer/Mannheim); 2,2'- Azino-di-(3-ethylbenzthiazolin-6-sulfonat) (ABTS, Boehringer/Mannheim).
Gerate: Absorptionsmessungen: Beckman Spectrophotometer DB-GT mit Schreiber; pH-stat-
Titrationen: automatischer Titrierstand (Metrohm), bestehend aus Dosimat E 535, Im- pulsomat E 473, pH-Meter E 500 sowie W + W-Zweikanalschreiber 312.
3.1 Darstellung von Derivaten des Synthesepulps mit Diazoniumgruppen als reaktive Komponente
3.1.1 Umsetzung von Synthesepulp mit 2-(3-Aminophenyl)-l,3- dioxolan zu Ia-c
10,OO g Synthesepulp (getrocknet) wurden in 200 cm3 Wasser suspendiert, 5 cm3 konz. HCl (D = 1,19) hinzugefugt und unter Riihren mit den in Tab. 9 angeffirten Mengen 2- (3-Aminophenyl)-1,3-dioxolan versetzt. Das Reaktionsgemisch wurde 15 h bei 50 "C ge- riihrt, anschlieRend abfiltriert und unter Riihren portionsweise mit Wasser neutral ge- waschen. Das Produkt wurde mit Aceton gewaschen, 15 h mit Aceton extrahiert und i. Vak. getrocknet.
Tab. 9. Umsetzungen von Synthesepulp mit 2-(3-Aminophenyl)-l,3-dioxolan.
SWP-Derivat Pro 10,OO g Synthesepulp eingesetzte Menge 2- (3-Aminophenyl)-1,3-dioxolan (cm3) (mmol)
SWP-E 400-Ia 2,oo 14,6 SWP-R 830-Ia 2,oo 14,6 SWP-R 830-Ib 1 ,oo 7,3 SWP-R 830-IC 0,20 125
67

G. Manecke und H.-G. Vogt
3 . 1 . 2 Darstellung der diazotierten Derivate IIa-c des Synthese- Pulps
200,O mg Ia-c wurden in 20 cm3 Wasser bei etwa 50 "C suspendiert, abfiltriert, an- schliel3end bei 0°C rnit 20 cm3 0,s M HCl versetzt und nach Zusatz von 1 cm3 1 M NaN02 30 min bei 0 "C geriihrt. Die diazotierten Derivate I I a x wurden abfiltriert, in- tensiv mit Eiswasser gewaschen und direkt fur die jeweilige Kupplungsreaktion einge- setzt.
3 . 1 . 3 Ermittlung des Gehalts an kupplungsfahigen Diazonium- gruppen der Derivate IIa-c (Umsetzung rnit Tyrosin als Mo- de llsu b s t anz)
100-500 mg genau eingewogener Trager Ia-c wurden nach 3.1.2 zu IIa-c diazotiert und mit 10 cm3 4. M Tyrosinlosung in 0,05 M Na2C03 2 h bei 0 "C geriihrt. Die Losung wurde in einen 50 cm3 MeBkolben filtriert und das Kupplungsprodukt unter Ruhren portionsweise rnit insgesamt 40 cm3 Wasser gewaschen und in den MeBkolben iiberfuhrt. Proben von 1,00 cm3 Waschlosung bzw. von Tyrosin-Eichlosungen des Kon- zentrationsbereichs 8 . bis 8 . l o - ' M Tyrosin wurden rnit Folin-Reagenz (Lowry et al.') versetzt und bei 620 nm Absorptionsmessungen durchgefuhrt. Anhand einer Eich- kurve wurde der Tyrosingehalt der Waschlosung ermittelt und daraus die von den Tra- gem IIa-c gebundene Menge Tyrosin berechnet.
3.2 Azokupplung von Enzymen an Synthesepulp-Derivate
Die nach 3.1.2 hergestellten Derivate IIa-c des Synthesepulps SWP-E 400 bzw. SWP- R 830 wurden rnit den in Tab. 10 aufgefuhrten Proteinen unter den angegebenen Kupp- lungsbedingungen umgesetzt. Das Immobilisierungsprodukt wurde rnit dem auch zur Kupplung eingesetzten Phosphatpuffer, dann rnit 0,5 M NaCl und tridest. Wasser gewa- schen. Die Waschlosungen wurden in einem MeBkolben aufgefangen und der EiweiB- gehalt aliquoter Teile der Waschlosung mit Folin-Reagenz (Lowry et al.') bestimmt. Das immobilisierte Enzym wurde gefriergetrocknet, ausgewogen und im Kiihlschrank bei 4 "C gelagert. Abweichend hiervon wurden immobilisierte Hexokinase und Glucose- 6-phosphat-dehydrogenase in tridest. Wasser bei 4 "C im Kuhlschrank aufbewahrt.
3.3 Charakterisierung der an Synthesepulp-Derivate immobilisierten Enzyme
3 . 3 . 1 Bestimmung der an den Trager gebundenen Menge Enzym
Die von dem jeweiligen Derivat des Synthesepulps gebundene Menge Enzym wurde aus dem Proteingehalt der Waschlosung und dem einer entsprechend verdiinnten Enzym- eichlosung durch Bestimmung rnit Folin-Reagenz (Lowry et al.') ermittelt. Zusatzlich wurde die an Synthesepulp-Derivaten immobilisierte Menge Enzym gravimetrisch be- stimmt. Hierzu wurde der jeweilige reaktive Trager vor der Enzymkupplung genau ein-
68
An Synthesepulp immobilisierte Enzyme, I.
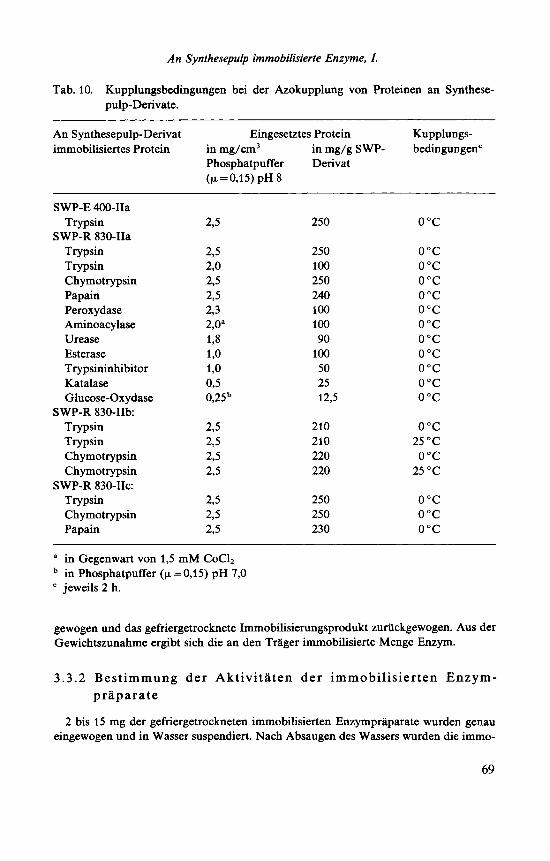
Tab. 10. Kupplungsbedingungen bei der Azokupplung von Proteinen an Synthese- pulp-Derivate.
An Synthesepulp-Derivat Eingesetztes Protein Kupplungs- immobilisiertes Protein in mg/cm3 in mg/g SWP- bedingungcn'
Phosphatpuffer Derivat (+=0,15)pH 8
SWP-E 400-IIa
SWP-R 830-IIa Trypsin
Trypsin Trypsin Chymotrypsin Papain Peroxydase Aminoacylase Urease Esterase Trypsininhibitor Katalase Glucose-Oxydase
Trypsin Trypsin Chymotrypsin Chymotrypsin
Trypsin Chymotrypsin Papain
SWP-R 830-IIb
SWP-R 830-IIc:
250
250 100 250 240 100 100 90
100 50 25 12.5
210 210 220 220
250 250 230
0 "C
0 "C 0 "C 0 "C 0 "C 0 "C 0 "C 0 "C 0 "C 0 "C 0 "C 0 "C
0 "C 25 "C 0 "C
25 "C
0 "C 0 "C 0 "C
~ ~~~~~
a in Gegenwart von 1,5 mM CoC12 in Phosphatpuffer (p=0,15) pH 7,O jeweils 2 h.
gewogen und das gefriergetrocknete Immobilisierungsprodukt zuriickgewogen. Aus dcr Gewichtszunahme ergibt sich die an den Trager immobilisierte Menge Enzym.
3 . 3 . 2 Bestimmung der Aktivitaten der immobilisierten Enzym- praparate
2 bis 15 mg der gefriergetrockneten immobilisierten Enzympraparate wurden genau eingewogen und in Wasser suspendiert. Nach Absaugen des Wassers wurden die immo-
69

G. Manecke und H.-G. Vogt
bilisierten Enzympraparate in einem thermostatisierten G e f a unter Riihren mit den je- weiligen Enzymsubstraten umgesetzt und die Substratlosung mit einer peristaltischen Pumpe in einem Umlaufverfahren iiber eine Fritte durch die gewtihlte MeBzelle (DurchfluBkiivette bzw. pH-MeBzelle) gepumpt.
3.3.2.1 Bestimmung der Aktivitat von immobilisiertem Trypsin
3.3.2.1.1 Bestimmung der Aktivitat von immobilisiertem Trypsin gegeniiber BAEE
Die Aktivitaten von an Synthesepulp-Derivaten immobilisiertem Trypsin wurden ge- geniiber BAEE unter Verwendung der in Tab. 11 angegebenen Konzentrationen im
Tab. 11. Aktivitatsbestimmungen von immobilisiertem Trypsin mit BAEE als Sub- strat.
Methode Konzentrationen im Test BAEE NaCl Tris CH,COONa mM M mM M
- - - titrimetrisch 20
titrimetrisch 10 - 0,Ol 1 ,oo titrimetrisch 10
spektralphotometrisch 10 02 100,o -
- - titrimetrisch 20 0,4 - - -
Test i. a. bei 25 "C bestimmt. Die Temperaturabhagigkeit der Aktivitat wurde im Be- reich 25-70 "C untersucht. Zur Ermittlung der pH-Aktivitatsprofile wurde der jeweilige pH durch Pufferzusatz bzw. durch pH-stat-Titration konstant gehalten. Oberhalb von pH 8,O wurden die gemessenen Aktivitaten durch den Betrag der nach gleicher Bestim- mungsmethode ermittelten Eigenhydrolyse des Substrats korrigiert.
3.3.2.1.2 Bestimmung der Aktivitat von immobilisiertem Trypsin gegenuber BAPA als Substrat
Die Aktivitat von an Synthesepulp immobilisiertem Trypsin wurde gegenuber D,L- BAPA in Anlehnung an die Methode von Erlanger et a1.'6 ermittelt, die in einer fur das immobilisierte Trypsin angepaBten Form bereits in Lit.3 beschrieben wurde. Nach glei- cher Methode wurden auch die Aktivitaten gegenuber L-BAPA ermittelt.
70

An Synthesepub immobilisierte Enzyme, I.
3.3.2.1.3 Bestimmung der Aktivitat von immobilisiertem Trypsin gegeniiber Casein als Substrat (modifizierte Methode nach Bergme~er'~)
1,00 g Casein wurde in einem 100 cm3 MeBkolben im siedenden Wasserbad in Phos- phatpuffer (Ionenstarke p =0,15) pH 8,0 gelost und die Losung bei Raumtemperatur mit Phosphatpuffer auf 100 cm3 aufgefit . 0-100 p1 einer wanrigen Trypsinlosung (ent- sprechend 0-100 pg Trypsin) bzw. ca. 50 mg an SWP-Derivaten immobilisiertes Tryp- sin wurden mit 10 cm3 Caseinlosung bei 37 "C 20 min inkubiert, wobei das immobili- sierte Enzym geriihrt wurde. AnschlieBend wurden beim freien Trypsin 15 cm3 5proz. CCI3COOH bzw. beim immobilisierten Trypsin zu 5 cm3 der abfiltrierten Losung 7,5 cm3 Sproz. CC1,COOH hinzugefugt und der Niederschlag 30 min stehengelassen und danach 20 min zentrifugiert. Die Absorption des Uberstandes wurde bei 280 nm gemes- sen und anhand einer fur das freie Trypsin erstellten Eichkurve die relative Aktivitat des immobilisierten Trypsins ermittelt.
3.3.2.1.4 Bestimmung der Thermostabilitat von an SWP-R 830-IIa immobili- siertem Trypsin
Einzelproben von 10-20 mg SWP-R 830-IIa-Trypsin wurden in Wasser suspendiert und in einem thermostatisierten Wasserbad bei 25-60°C 15-120 min inkubiert. An- schlieaend wurden die Proben in Eiswasser gekuhlt und die jeweilige Aktivitat gegen- uber BAEE ennittelt. Freies Trypsin wurde analog behandelt, indem 0,Zproz. waBrige Trypsinlosungen inkubiert wurden.
3.3.2.1.5 Bestimmung der Eigenschaften von immobilisiertem Trypsin fur die Affmitatsadsorption von Sojabohnen-Trypsininhibitor (STI)
50 mg an SWP-R 830-IIa-Trypsin wurden unter Riihren in 2 cm3 Phosphatpuffer pH 8,O (p=O,15) suspendiert und 2 cm3 einer Losung von 0,50 mg STI in Phosphatpuffer pH 8,0 (p, = 0,15) hinzugefugt. Nach einstundigem Riihren bei Raumtemperatur wurde die Losung abgesaugt und in einen 25 cm3 MeBkolben uberfuhrt. Das SWP-Derivat wurde portionsweise mit Phosphatpuffer pH 8,0 gewaschen und die Waschlosungen in den MeBkolben gegeben und dieser bis zur 25 cm3 Marke aufgef i t (Waschlosung A). Daraufhin wurde das SWP-Derivat zunachst zweimal mit je 5 cm3 Wasser gewaschen und die Waschlosung verworfen. Zur STI-Desorption wurde das SWP-Derivat 5mal mit je 5 cm3 I mM HCI jeweils 10 min unter Ruhren gewaschen und die Waschlosungen in einen 25 cm3 MeBkolben uberfuhrt (Waschlosung B). Der Eiweiagehalt der Waschlo- sungen A und B wurde mit Folin-Reagenz (Lowry et al.') bestimmt. Weiterhin wurde die Inhibitoraktivitat der Waschlosungen A und B sowie die einer STI-Standardlosung bestimmt und daraus die reversibel vom immobilisierten Trypsin gebundene Menge STI ermittelt.
71

G. Manecke und H.-G. Vogt
3.3.2.1.6 Bestimmung der Inhibitoraktivitat von Sojabohnen-Trypsininhibi- tor
Die Trypsinaktivitat gegenuber BAEE wurde spektralphotometrisch, wie in 3.3.2.1.1 beschrieben, bestimmt. Zur Bestimmung der Inhibitoraktivitat wurde eine Losung von 0,l mg Trypsin in 0,l cm3 1 mM HC1 mit 5,O cm3 0,2 M Tris-Puffer pH 8,O versetzt, I cm3 einer waBrigen STI-Standardlosung oder -Waschlosung ( = 0,05 mg STI/cm3) hin- zugefugt und die Losung 5 min bei 25 "C stehengelassen. Darauf wurden 5,O cm3 0,02 M BAEE/0,4 M NaCl hinzugegeben und die Aktivitat durch Messung der Absorptionszu- nahme bei 255 nm (~=0,808 cm2 pmol-1)23 in einer 0,l cm Kiivette ermittelt. Aus den Trypsinaktivitaten in Abwesenheit und in Gegenwart von STI wurden die spezifische Inhibitoraktivitat in U/mg STI und die pro mg STI inhibierte Menge Trypsin berech- net.
3.3.2.2 Bestimmung der Inhibitoreigenschaften von an SWP-R 803-IIa immo- bilisiertem Sojabohnen-Trypsinibitor
3.3.2.2.1 Umsetzung von SWP-R 830-IIa-STI rnit Trypsin
50,O mg SWP-R 830-IIa-STI wurden unter Ruhren rnit einer Fritte rnit 3 cm3 Phos- phatpuffer pH 8,O (p=O,15) suspendiert. 2 cm3 einer frisch zubereiteten Losung von 0,6 mg Trypsin (Merck) in Phosphatpuffer pH 8,O (p = 0,15) wurden in einem Umlaufver- fahren mit dem immobilisierten Trypsininhibitor unter Riihren zur Reaktion gebracht und die Absorptionsabnahme der durch eine 1 cm-DurchfluBkuvette gepumpten Reak- tionslosung bei 280 nm zeitlich verfolgt. Nach einer Reaktionszeit von 1 h wurde die Umsetzung abgebrochen, die Losung in einen 25 cm3 MeBkolben uberfuhrt, der Trager unter Ruhren portionsweise rnit Phosphatpuffer pH 8,O (p = 0,15) gewaschen und die Waschlosung auf 25 cm3 aufgefullt. Der EiweiBgehalt der Waschlosung wurde rnit Fo- lin-Reagenz (Lowry et al.') bestimmt.
3.3.2.2.2 Desorption von an SWP-R 830-IIa-STI gebundenem Trypsin
Das nach 3.3.2.2.1 an SWP-R 830-IIa-STI gebundene Trypsin wurde in einer Fritte mit 5 cm3 Wasser unter Ruhren gewaschen und die Losung in einem Umlaufverfahren durch eine I cm-DurchfluBkuvette gepumpt. Die Losung wurde abgesaugt, unter Ruh- ren 5 cm3 1 mM HC1 hinzugefugt und die Desorption des Trypsins durch Absorptions- messung bei 280 nm verfolgt. Zur Vervollstandigung der Desorption wurden viermal 5 cm3 1 mM KC1 unter Ruhren zum Trager gegeben und die Eluate in einem 25 cm3 MeBkolben gesammelt und die Waschlosung bis zur Marke aufgefullt. Der EiweiBge- halt des Eluats wurde mit Fohn-Reagenz (Lowry et al.') und die Aktivitat des Eluats rnit BAEE als Substrat bestimmt.
72

An Synthesepulp immobilisierte Enzyme, I.
3.3.2.2.3 Adsorption und Desorption von Trypsin an SWP-R 830-IIa-STI in einer durchstromten Saule
40 mg SWP-R 830-IIa-STI wurden unter Riihren in Phosphatpuffer pH 8,O disper- giert und in eine Saule (Durchmesser 0,3 cm) gefut. Auf die Saule wurde eine Losung von 0,25 mg Trypsin in 0,025 cm3 Phosphatpuffer pH 8,O gegeben und rnit demselben Puffer eluiert. Das Eluat wurde mit einer peristaltischen Pumpe bei einer Pumpge- schwindigkeit von 0,25 cm3/min durch eine 0,2 cm DurchfluRkiivette geschickt und der Absorptionsverlauf bei 280 nm registriert. Nach Zugabe von 10 cm3 Elutionspuffer wur- de die Saule mit Wasser gespiilt und anschlieRend die Desorption mit 1 mM HCl bei gleicher Durchfluageschwindigkeit durchgefuhrt.
3.3.2.3 Bestimmung der Aktivitat von immobilisiertem Chymotrypsin gegen- uber ATEE als Substrat
Die Aktivitaten von an Synthesepulp-Derivaten immobilisiertem Chymotrypsin wur- den gegenuber dem Substrat N-Acetyl-L-tyrosinethylester (ATEE) bestimmt. Abgewo- gene Proben des gefriergetrockneten immobilisierten Chymotrypsins wurden hierzu rnit 15 cm3 0,02 M ATEE (0,2 M KCl, 5 Vo1.-% EtOH) durch pH-stat-Titration rnit 0,s M NaOH bei 25 "C untersucht. Als pH-Bereich fur die Untersuchung der pH-Aktivitats- profile wurde pH 7,s-11,O gewahlt. Die Eigenhydrolyse des ATEE wurde fur pH-Werte oberhalb von pH 9,s in analoger Weise bestimmt und die daraus ermittelten Werte bei der Berechnung der Enzymaktivitat beriicksichtigt.
3.3.2.4 Bestimmung der Aktivitat von immobilisiertem Papain gegenuber BAEE als Substrat
Die Aktivitaten von an Synthesepulp immobilisiertem Papain wurden rnit BAEE, wie bereits in einer friiheren Arbeit' beschrieben, bestimmt.
3.3.2.5 Bestimmung der Aktivitat von immobilisierter Urease
3.3.2.5.1 Bestimmung der Aktivitat und der pH-Aktivitatsprofde von immobi- lisierter Urease
Die Aktivitat von immobilisierter Urease wurde rnit 15 cm3 einer 0,15 M Harnstofflo- sung, die zur Untersuchung des Puffereintlusses 0,Ol M bzw. 0,05 M Imidazolpuffer ent- hielt, bei 25 "C durch pH-stat-Titration mit 0,l M HC1 bestimmt. Zur Bestimmung der pH-Aktivitatsprofile wurde der pH-Bereich von pH 3,0-7,s erfaRt. Die Aktivitaten wur- den als pMol umgesetzter Harnstoff pro min und g Immobilisierungsprodukt berech- net.
73

G. Manecke und H.-G. Vogt
3.3.2.5.2 Bestimmung der Thermostabilitat von immobilisierter Urease
Die Bestimmung der Thermostabilitat von an SWP-R 830-IIa immobilisierter Urease erfolgte durch Inkubation von in Wasser suspendierten Proben des immobilisierten En- zyms bei 40 bzw. 50 "C in einem thermostatisierten Wasserbad. Freie Urease wurde als 0,OSproz. warige Enzymlosung in analoger Weise inkubiert. Nach Inkubationszeiten von 15-60 min wurden die Proben mit Eiswasser gekiihlt und die Aktivitat geman 3.3.2.5.1 bestimmt und in Relation zur Ausgangsaktivitat gesetzt.
3.3.2.6 Bestimmung der Aktivitat von immobilisierter Aminoacylase gegen- uber N-Acetyl-D,L-methionin (nach einer Testvorschrift des Kontroll- Labors der Boehringer-Mannheim GmbH, Biochemica Werk Tut- zing24)
Die Aktivitat von immobilisierter Aminoacylase wurde gegeniiber N-Acetyl-D,L- methionin bestimmt. Hierzu wurden zum immobilisierten Enzym 50 cm3 0,l M Tris- Puffer pH 7,4,2 cm3 0,015 M CoC12 und 2 cm3 0,5 M N-Acetyl-D,L-methionin gegeben und bei 25 "C umgesetzt. Nach 5 , l O und 15 min wurden je 5 cm3 der Substratlosung ab- filtriert und der Gehalt an zu L-Methionin umgesetztem Substrat bestimmt. Hienu wurden in eine 1 cm Kiivette 2,70 cm3 der mit O2 gesattigten, filtrierten Substratlosung gegeben, 0,l an3 0,Ol M o-Dianisidin und 0,Ol cm3 (0,l mg) POD hinzugefugt. Die Ab- sorption bei 436 nm wurde gemessen und anschlieRend 0,l cm3 L-AOD hinzugegeben und der sich einstellende Endwert der Absorption gemessen. Aus der Absorptionsdif- ferenz wurde der Methionin-Gehalt in den nach 5, 10 und 15 min gezogenen Proben ermittelt ( E = 8,3 cm2 pmol-') und daraus die Aminoacylaseaktivitat berechnet.
3.3.2.7 Bestimmung der Aktivitat von immobilisierter Esterase gegenuber Acetyl-D,L-pantolacton
Die Aktivitat von an Synthesepulp immobilisierter Esterase wurde mit Acetyl-D,L- pantolacton durch pH-stat-Titration mit 0,05 M NaOH bei pH 8 (25 "C) bestimmt. Zur Aktivitatsbestimmung wurden 0,025 cm3 Acetyl-D,L-pantolacton, 15 cm3 0,l M NaCl und 0,2 cm3 0,l M Boratpuffer pH 8 eingesetzt. Zur Berechnung der Enzymaktivitaten wurde die Eigenhydrolyse des Substrats beriicksichtigt.
3.3.2.8 Bestimmung der Aktivitat von immobilisierter Glucose-Oxydase
Die Aktivitat der immobilisierten Glucose-Oxydase wurde nach der ABTS/POD- Methode" wie bereits beschrieben3 bestimmt.
74

An Synthesepulp immobilisierte Enzyme, I .
3.3.2.9 Bestimmung der Aktivitat von immobilisierter Katalase
Die Katalase-Aktivitaten wurden UV-spektralphotometrisch in Anlehnung an die von Bergmeye?' beschriebene Methode bestimmt'.
3.3.2.10 Bestimmung der Aktivitat von immobilisierter Peroxydase (modifi- zierte Methode nach Werner et a1." mit ABTS als Wasserstoffdona- tor)
15 cm3 einer 0,025 M Losung von ABTS in Phosphatpuffer pH 7,O (p=O,15) wurden mit 0,5 cm3 einer 0,03proz. H202-Losung versetzt, die eingewogene Probe des immobili- sierten Enzyms hinzugegeben und bei 25 "C geriihrt. Die Losung wurde iiber eine Fritte durch eine 0,2 cm DurchfluBkuvette gepumpt und die Absorption bei 436 nm ( E = 29,3 cm2 pmol- ') verfolgt.
3.3.2.1 1 Bestimmung der Aktivitaten von coimmobilisierter Hexokinase/Glu- cose-6-phosphat-dehydrogenase (modifuierte Methode nach Berg- meyer26)
3.3.2.1 1.1 Bestimmung der gemeinsamen Aktivitat des Immobilisierungspro- duktes
6 cm3 der auf pH 7,6 eingestellten HK/G-6-PDH-Substratlosung bestehend aus 3 cm3 0,44 M Glucose (80 mg/cm3), 2 cm3 0,l M Tris-Puffer pH 7,6,0,4 cm3 0,l M MgCI2, 0,2 cm3 81. MM ATP (50 mg/cm3) und 0,4 cm3 11. l o - ' M NADP (10 mg/cm3) wur- den zu dem Immobilisierungsprodukt gegeben und bei 25 "C in einem Umlaufverfahren durch eine 1 cm DurchfluBkiivette gepumpt und die Absorption bei 334 nm ( E = 6,O cm2 pmol- ') registriert.
3.3.2.1 1.2 Bestimmung der Einzelaktivitat von immobilisierter Hexokinase
Die HK-Aktivitat wurde entsprechend wie unter 3.3.2.1 1.1 beschrieben bestimmt, zu- satzlich wurden jedoch zur Substratlosung 0,Ol cm3 (0,5 U) G-6-PDH-Losung hinzuge- Wgt.
75

G. Manecke und H.-G. Vogt
3.3.2.1 1.3 Bestimmung der Einzelaktivitat von immobilisierter Glucosed- phosphat-dehydrogenase
Das Immobilisierungsprodukt wurde mit 6 cm3 G-6-PDH-Substrat pH 7,6, bestehend aus 0,2 cm3 Glucose-6-phosphat (10 mg/cm3), 5,2 cm3 0,l M Tris-Puffer pH 7,6,0,4 cm3 0,l M MgC12 und 0,2 cm3 11. lo-' M NADP (10 mg/cm3), bei 25 "C umgesetzt und die Absorption bei 334 nm der durch eine 1 cm DurchfluBkuvette geleiteten Losung regi- striert ( E = 6,O cm2 pmol-I).
Fur die Bereitstellung der in dieser Arbeit eingesetzten Synthesepulp-Typen sei H e m Prof. Dr. H. Cherdron gedankt. Fur die Uberlassung von Sojaboh- nen-Trypsininhibitor und die Anregung zu dessen Immobilisierung an Synthe- sepulp sowie fur die Zusammenarbeit bei der Charakterisierung des immobili- sierten Inhibitors sei Herrn Prof. Dr. V. Kasche gedankt. Die fur diese Arbeit eingesetzte Esterase wurde uns freundlicherweise von Herrn Dr. D. Jaworek (Boehringer/Mannheim) zur Verfugung gestellt. Unser Dank gilt weiterhin Herrn R. Zander fur die Unterstutzung bei der experimentellen Durchfuhrung der in dieser Arbeit beschriebenen Untersuchungen.
Ger. Offen. 2,249,604 (1973), Crown Zellerbach International, C. A. 79 (1973) 7064 p W. Gordon, H. J. Leugering, H. Cherdron, Angew. Chem. 90 (1978) 833 G. Manecke, H.-G. Vogt, Makromol. Chem. 177 (1976) 725 G. Manecke, H.-G. Vogt, Immobilized Enzymes on Poly(viny1 alcohol) Camers, in Analysis and Control of Immobilized Systems, Hrsg. D. Thomas, J.-P. Kernevez, North Holland Publishing Company, Amsterdam 1976, S. 105 G. Manecke, H.-G. Vogt, Angew. Makromol. Chem. 78 (1979) 21 G. Manecke, H.-G. Vogt, J. Solid-Phase Biochem., im Druck 0. H. Lowry, N. J. Rosebrough, A. L. Farr, R. J. Randall, J. Biol. Chem. 193 (1951) 265 D. J. Lartigue, Basic Enzymology in Immobilized Enzymes for Industrial Reactors, Hrsg. R. A. Messing, Academic Press, New York 1975, S. 11 W. J. Crozier, J. Gen. Physiol. 7 (1925) 123
lo D. Gabel, V. Kasche, Biochem. Biophys. Res. Commun. 48 (1972) 1011 l1 D. Gabel, V. Kasche, Acta Chem. Scand. 27 (1973) 1971 l2 K. Buchholz, Kinetische Messungen mit polymer gebundenen Enzymen, in Deche-
ma-Monographien Nr. 1351-1369, Bd. 71, Hrsg. H. J. Rehm, Verlag Chemie, Wein- heim/Bergstr. 1973, S. 315
' '
76

An Synthesepulp immobilisierte Enzyme, I.
l3 J. M. Engasser, C. Horvath, Diffusion and Kinetics with Immobilized Enzymes, in Applied Biochemistry and Bioengineering, Hrsg. L. B. Wingard, Jr., E. Katchalski- Katzir, L. Goldstein, Academic Press, New York 1976, Bd. I, S. 127
l4 J. M. Engasser, C. Horvath in Analysis and Control of Immobilized Enzyme Sy- stems (Eds. D. Thomas, J. P. Kemevez) North Holland Publishing Company, Am- sterdam 1976, S. 187 J. M. Engasser, C. Horvath, Biochem. Biophys. Acta 358 (1974) 178 B. F. Erlanger, N. Kokowsky, W. Cohen, Arch. Biochem. Biophys. 95 (1961) 271 H. U. Bergmeyer, Methoden der enzymatischen Analyse, 3. Aufl., Verlag Chemie, Weinheim/Bergstr. 1974, Bd. 1, S. 1506 E. H. Reimerdes, First European Congress on Biotechnology, Preprints, Interlaken 1978, Part 1, S. 43 M. Yamamoto, T. Ikenaka, J. Biochem. (Tokyo) 62 (1967) 141 F. J. Reithel, J. E. Robbins, G. Gorin, Arch. Biochem. Biophys. 108 (1964) 409 C. Saronio, M. Salmona, E. Mussini, S. Garattini, Biochemical Studies on Some Fi- ber-Entrapped Enzymes, in Insolubilized Enzymes (Hrsg. M. Salmona, C. Saronio, S. Garattini), Raven Press, New York 1974, S. 143
22 W. Emer, H. G. Rey, H. Wielinger, Fresenius Z. Anal. Chem. 252 (1970) 224 23 J. F. Kezdy, L. Lorand, K. D. Miller, Biochemistry 4 (1965) 2302 24 Boehringer-Mannheim GmbH, unveroffentlicht 25 H. U. Bergmeyer, Methoden der enzymatischen Analyse, 3. Aufl., Verlag Chemie,
Weinheim/Bergstr. 1974, Bd. 1, S. 713 26 H. U. Bergmeyer, Methoden der enzymatischen Analyse, 3. Aufl., Verlag Chemie,
Weinheim/Bergstr. 1974, Bd. 1, S. 487 und S. 502
l.5
l6
l7
l9
2o
21
77





























