Embed Size (px)
Citation preview

BIOCHIMICA ET BIOPHYSICA ACTA 2 9
BBA 12336
A N R E I C H E R U N G U N D C H A R A K T E R I S I E R U N G E I N E R
1 7 f l - H Y D R O X Y S T E R O I D : N A D ( P ) - O X Y D O R E D U C T A S E
D E R R A T T E N N I E R E
H. BREUER UND K. DAHM
Chemische Abteilung der Chirurgischen Universitiitsklinik, Bonn (Deutschland)
(Eingegangen am 27. August, i963)
SUMMARY
Partial purification and characterisation of a z7fl-hydroxysteroid: N A D ( P )-oxidor eductase of rat kidney
The cytoplasm of rat kidney contains a i7fl-hydroxysteroid dehydrogenase which has been purified I2-fold by precipitation of the 20 ooo × g supernatant with am- monium sulfate. The enzyme reacts with both phenolic and neutral steroids; either NAD or NADP can be utilised as cofactors.
The oxidation of testosterone to androst-4-en-3,I7-dione shows a maximum at p H 8.7, whereas the reduction of androst-4-en-3,i7-dione to testosterone is optimum at pH 6.6. The Michaelis-Menten constants were found to be 18.5"1o -5 M for testosterone and 4O.lO -5 M for NAD. A value of 9.1 . io -gM was obtained for the equilibrium constant. The enzyme shows an activation energy of 13.2 kcal/mole and an inactivation energy of --24.0 kcal/mole within the range of lO-37°; the Q10 values varied between 1.6 and 1. 5 .
The enzymic activity is inhibited by SH blocking agents, but activated by zink ions in low concentrations (lO -5 M). The physiological significance of the 17fl- hydroxysteroid dehydrogenase of rat kidney is discussed briefly.
EINLEITUNG
xTfl-Hydroxysteroid-Dehydrogenasen (17fi-Hydroxysteroid: NAD(P)-Oxydoreducta- sen) spielen bei der Biogenese und im Zwischenstoffwechsel der C-I8 und C-I 9- Steroide eine wichtige Rolle; sie katalysieren die Bildung von Oestradiol-i7fl und Testosteron aus den biologisch weniger aktiven iT-Oxo-Verbindungen. Obgleich I7fl-Hydroxysteroid-Dehydrogenasen in zahlreichen Organen nachgewiesen worden sind (vgl. Ref. i), liegen Angaben fiber ihre Anreicherung und Charakterisierung bis heute nur ffir die Placenta 2-4, die Lebere, ~, die NebenniereS, 9 und die Erythro- zyten 1° vor.
Untersuchungen aus neuerer Zeit lassen erkennen, dass neben der Leber auch die Niere in gr6ssere m Umfange am Stoffwechsel der Steroide teilnimmt (vgl. Ref. 1). Es erschien deshalb wfinschenswert, n/ihere Angaben tiber die Steroidenzyme der Niere zu erhalten. In Fortsetzung unserer Arbeiten tiber die XT/~-Hydroxysteroid-
Biochim. Biophys. Acta, 85 (z964) 29-37

3 ° H. BREUER, K. DAHM
Dehydrogenasen bei S~ugetierenS, "~ und niederen Vertebraten ~ berichten wit im folgenden tiber die Anreicherung und Charakterisierung einer cytoplasmatischen I7/5-Hydroxysteroid:NAD(P)-Oxydoreductase aus der Rattenniere. Durch Behand- lung mit Ammoniumsulfat liess sich eine I2-fache Anreicherung der Enzymaktivit~it erzielen; auf Grund seiner Eigenschaffen kann das Enzym in die Klasse der bisher bekannten I7~-Hydroxysteroid-Dehydrogenasen mit funktionellen SH-Gruppen ein- gereiht werden.
METHODIK
Reagentien und L6sungen
Alle verwendeten Reagentien waren von p.a. Reinheitsgrad (E. Merck, Darm- stadt); Glycerin (E. Merck, Darmstadt) war zweimal destilliert. Die Medien A und B wurden in Anlehnung an JARABAK, AI)AM,% WILLIAMs-ASHMAN UND TALALAY 4 her- gestellt; sie enthielten o.oi M Phosphat-Puffer (pH 7.o), o.0o 5 M EDTA und 2o'/~, Glycerin (Medium A) bzw. 50% Glycerin (Medium B). Alle organischen L6sungs- mittel wurden vor dem Gebraueh zweimal destilliert.
Versuchstiere
MS.nnliche und weibliche Wistar-Ratten im Alter yon 6-8 Wochen wurden nach 24-sttindigem Nahrungsentzug durch Dekapitation get6tet und die Nieren nach dem Ausbluten der Tiere entnommen.
Zellfraktionierung
Alle Schritte wurden bei o ° ausgeftihrt. Das Nierengewebe wurde in einem M6rser zerkleinert und in Medium A mit einem Glashomogenisator homogenisiert. Das I5% ige Homogenat wurde 60 rain bei 5000 x g zentrifugiert; anschliessend wurde der t]berstand 6o rain bei 20 ooo x g zentrifugiert.
Fraktionierge Fdllung mit Ammoniumsulfat
Der 2o ooo x g (}berstand wurde bei o ° der fraktionierten F~tllung mit Am- moniumsulfat unterworfen. Durch Zugabe wm 4%igem Ammoniak wurde der pH- Wert w/ihrend der F~llung zwischen 6.8 und 7.2 gehalten. Der Zusatz yon Ammonium- sulfat bis zur 4o%igen S~ttigung erfolgte innerhalb von 2 h. Die L6sung wurde dann etwa 20 h stehen gelassen, der Niederschlag abzentrifugiert und das Sedi- ment in Medium B aufgenommen. Der ~]berstand der 4o% igen Ammoniumsulfat- Fiillung wurde his zur 8o%igen Siittigung mit Ammoniumsulfat versetzt und die L6sung wie oben beschrieben aufgearbeitet. Die 4 °- und 8o% igen Ammoniumsulfat- F~illungen sowie der i3berstand wurden bei - -6 ° autbewahrt.
Inkubationen Wenn nicht anders vermerkt, hat ten die Versuchsans~ttze folgende Zusammen-
setzung: o. 7/~Mol Testosteron (gel6st in o.05 ml Methanol), 0.5 ml Enzyml6sung, 5/~Mol NAD oder NADP und 0.4 M Tris-HC1 Puffer (pH 8.8) in einem Gesamt- volumen yon 3.5 ml.
Biochim. Biophys. Acta, 85 (1964) 29-37

I7/~-HYDROXYSTEROID-DEHYDROGENASE DER RATTENiNIERE 31
Aufarbeitung der Versuchsansdtze
Nach Beendigung der Inkubationen wurden die L6sungen dreimal mit je 5 ml Aether-Chloroform (3:1) extrahiert, die Extrakte vereinigt und unter vermindertem Druck im Rotationsverdampfer zur Trockne gebracht. Die Rtickst/inde der Inkuba- tionen mit Testosteron wurden der Papierchromatographie im System Petrol- aether (kp. 4o-6o°)-Methanol-Wasser (lOO:85:15) unterworfen; Laufzeit 2 h. Die Lokalisierung yon Testosteron und Androst-4-en-3,I7-dion erfolgte durch Kontakt- photographie im Ultraviolett. Die Rtickst/inde der Inkubationen mit Oestradiol-i7fl wurden auf Propylenglycol impr~gnierten Papier im System Cyclohexan-Benzol (I : I) ftir 7 h chromatographiert. Oestradiol-I7fl und Oestron wurden durch Anf/irben der Randstreifen mit FOLIN-CIOCALTEUS 12 Reagens lokalisiert.
Quantitative Bestimmungen
Neutrale Steroide (Aa-3-Oxosteroide): Die durch ultraviolette Kontaktphoto- graphie ermittelten steroidhaltigen Positionen wurden ausgeschnitten und die Papierstticke nach dem Zerkleinern mit 3 ml Methanol in einem 25 ml-Erlenmeyer- kolben mit Schliff fiir IO min eluiert. Die methanolischen L6sungen wurden vor- sichtig abpipettiert und die Extinktionen in io-mm Quarzktivetten bei 238 m/~ ge- messen. Nach Papierchromatographie und Elution betrug die Wiederfindung von Testosteron und Androst-4-en-3,i7-dion 85 bis 90% (IO Versuche). Die Gesamt- wiederfindung ftir beide Steroide (Bereich 5-4 °/~g) nach Zusatz zu Standardinkuba- tionen, die an Stelle des Enzyms o.5ml einer o.2~oigen Rinderserumalbumin- L6sung enthielten, betrug 7O~o (8 Versuche).
Phenolische Steroide: Die Oestrogen-haltigen Positionen wurden, wie oben be- schrieben, mit Methanol eluiert, die Eluate durch eine Glasfritte (Schott, Mainz) G 4 filtriert und eingedampft. Die quantitative Bestimmung yon Oestradiol-I7fl und Oestron wurde mit der Kober-Reaktion durchgeff~hrt; Einzelheiten siehe BREUER UND NOCKE 13.
Eiweissbestimmung: Diese erfolgte nach LOWRY et al. 14.
ERGEBNISSE
Nach Inkubation yon Testosteron mit verschiedenen Fraktionen des Nierencyto- plasmas entstand Androst-4-en-3.i7-dion in unterschiedlicher Ausbeute. Wie aus Tabelle I hervorgeht, land sich die h6chste spezifische Aktivit~t in der 8o°/0igen Ammoniumsulfat-Fraktion. Diese Fraktion zeigte gegeniiber dem Homogenat eine etwa I2-fach h6here spezifische Aktivit/it und wurde in allen weiteren Versuchen ver- wendet; unter den vorliegenden Bedingungen wurde ausser Androst-4-en-3.i7-dion kein weiterer Metabolit yon Testosteron gefunden.
Die Enzympr/iparation oxydierte Oestradiol-I7fl in etwas geringerem Umfange (etwa 7O~o) als Testosteron, Mit NAD und NADP wurden gleich grosse Ums/itze beobachtet.
Kinetische Untersuchungen
Wie aus Fig. I hervorgeht, zeigt die Oxydation von Testosteron zu Androst- 4-
Biochim. Biophys. Acta, 85 (1964) 29-37
3 2 I-I. B R E U E R , K. DAHM
T A B E L L E 1
G E S A M T A K T I V I T . : ~ T E N U N D S P E Z I F I S C H E A K T I V I T f i , T E N D E R N 2 ~ D - A B H ~ k N G I G E N J 7[J- H Y D R O X Y S T E R O I D - D E H Y D R O G E N A S E N I N V E R S C H I E D E N E N P R A P A R A T I O N E N
D E R R A T T E N N I E R E
0. 7 /dv lo l T e s t o s t e r o n w u r d e n m i t 0. 5 m l de r j c w e i l i g e n E n z y m p r ~ p a r a t i o n , 5 l tMol N A D u n d o. 4 M T r i s - H C 1 Puf fe r ( p t I 8.8) i n e i n e m G e s a m t v o l u m e n v o n 3.5 m l f ~ r 60 ra in be i 37 :~ u n t e r
I m f t i n e i n e m S c h i i t t e l t h e r m o s t a t e n i n k u b i e r t . \Ve i te re E i n z e l h e i t e n s iehe METHODIK.
Gob ihlctc M enge I:'ia;eis sgcha It
F, nzymprgpr~ration .! ndrost 4-on dcr Enzym Gesamt- Spczifischc ¢.z7-dio~ prc~paration Aktivit~t* Aktivitdt** ( M d/ (mg/o.5 rid)
o,5 ml ~, h)
H o l n o g e n a t 5 <) 26.5 355 ° i .0 l oo ooo × g ( ~ b e r s t a n d ,t o 5.5 ~88o 7.-' 40°//o A m m o n i u m s u l f a t - F ~ l l u n g 28 z.3 1ooo i2 .1 8 0 % A m m o n i u m s u l f a t - F ~ l l u n g 1"4 5.5 I8OO 22.() 15be r s t and d e r 8O~o A m m o -
n i u m s u l f a t - F ~ l l u n g o o.3 • o
• G e s a m t a k t i v i t a t - m/~-I°l- j )ndr°st-4-er~-3"~I7-di°n- gel2ilde-t" h
** Spez i f i sche A k t i v i t A t = mfd~'I°l ia~nclr°st-4-eI1--3'I7 -di°l} gebiM_ e t
m g E i w e i s s X h
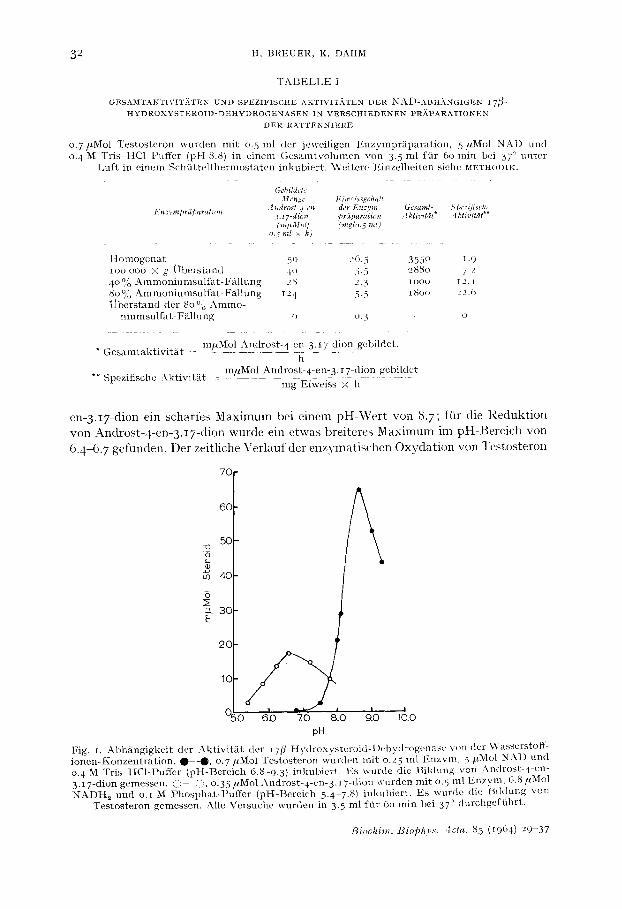
en-3.I7-dion ein scharfes Maximum bei einem pH-Wert von 8. 7 ftir die Reduktion von Androst-4-en-3.IT-dion wurde ein etwas breiteres Maximum im pH-Bereich von 6.4-6. 7 gefunden. Der zeitliche Verlauf der enzymatischen Oxydation von Testosteron
7 0
6 0
5O -o
3 0 E
2 0
q i 6.0 7.0 8.O 9.'0 10.O
pH
Fig . i . A b h / ~ n g i g k e i t dc r A k t i v i t / i t d e r i 7 / ~ - H y d r o x y s t e r o i d - l ) e h y d r o g e n a s ~ yon de r ~Vasserstoff- i o n e n - l , 2 o n z e n t r a t i o n . • - 0 , o . 7 / , M o l T e s t o s t e r o n w u r d e n m i t o.25 m l l : .nzym, 5 / t M o l N A I ) u n d 0. 4 M T r i s - H C 1 - P u f f e r ( p H - B e r e i c h 6.8 9.3) i n k u b i e r t . Es w u r d e d ie B i k h m g yon ; \ n d r o s t - 4 - c n - 3 . I 7 -d ion g e m e s s e n . ()- , : , o .35 /2Mol A n d r o s t - 4 - e n - 3 . I 7 -d ion w u r d e n m i t o.5 m l E n z y m , 6.8 ffMol NADH,z u n d o.~ M P h o s p h a t - P u f f e r ( p H - B e r e i c h 5.4 7.8) i n k u b i e r t . E s w u r d e (lie B i M u n g v o n
T e s t o s t e r o n gemessen . Alle V e r s u c h e w u r d e n in 3.5 m l flhr 6o m i n be i 37 ° d u r c h g e f g h r t .
Bioahim. 1Mophys. ,It/a, 83 ( t9~4) 29-37
I?f l -HYDROXYSTEROID-DEHYDROGENASE DER RATTENNIERE 33
-I() -5
._o c 100 -o I
I e-
l ~t
I
5O o
" o c <
-6 "3
E
C o ! I ,5 2'o 6o 9'o
Zeit (rain)
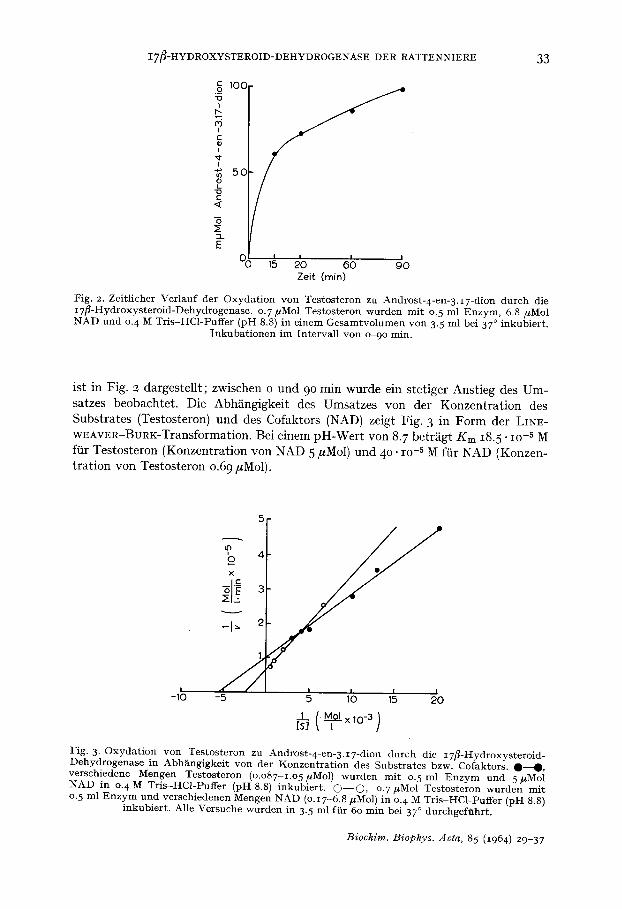
Fig. 2. Zeit l icher Ver lauf der O x y d a t i o n von Tes tos te ron zu Andros t -4-en- 3 . I7-dion du rch die I7 f l -Hydroxys te ro id -Dehydrogenase . o. 7/~Mol Tes tos te ron w u r d e n mi t 0. 5 ml E n z y m , 6.8 /~Mol N A D und o. 4 M Tr i s -HCl -Puf fe r (pH 8.8) in e inem G e s a m t v o l u m e n yon 3.5 ml bei 37 ° inkubier t .
I n k u b a t i o n e n i m In te rva l l yon o~)o min .
ist in Fig. 2 dargestellt; zwischen o und 9 ° min wurde ein stetiger Anstieg des Um- satzes beobachtet. Die AbMngigkeit des Umsatzes yon der Konzentration des Substrates (Testosteron) und des Cofaktors (NAD) zeigt Fig. 3 in Form der LINE- WEAVER-BORK-Transformation. Bei einem pH-Wert yon 8. 7 betr/igt Km 18.5" lO -5 M f~r Testosteron (Konzentration von NAD 5 #Mol) und 4 ° . lO -5 M ftir NAD (Konzen- tration von Testosteron 0.6 9 #Mol).
I ! I I 5 10 15 20
, - -
E - b 4
;K¢.
2 ¸
/
Fig. 3. O x y d a t i o n von Tes tos te ron zu Andros t -4 -en-3 . i7 -d ion du rch die i7 f l -Hydroxys te ro id - D e h y d r o g e n a s e in Abhi~ngigkeit yon tier K o n z e n t r a t i o n des Subs t r a t e s bzw. Cofaktors . S - - Q , verschiedene Mengen Tes tos te ron (o.o87-1.o 5/~Mol) wurden mi t 0. 5 ml E n z y m u n d 5/*Mol N A D in 0. 4 M Tr i s -HCl -Puf f e r (pFI 8.8) inkubier t . O - - O , o.7/~Mol Tes tos te ron w u r d e n m i t o. 5 ml E n z y m u n d versch iedenen Mengen N A D (o.17-6.8/~Mol) in 0. 4 M Tris-HC1-Puffer (pH 8.8)
inkubier t . Alle Versuche wurden in 3.5 ml fQr 60 min bei 37 ° durchgeff ihr t .
Biochim. Biophys. Acta, 85 (1964) 29-37
34 H. BREUER, K. DAHM
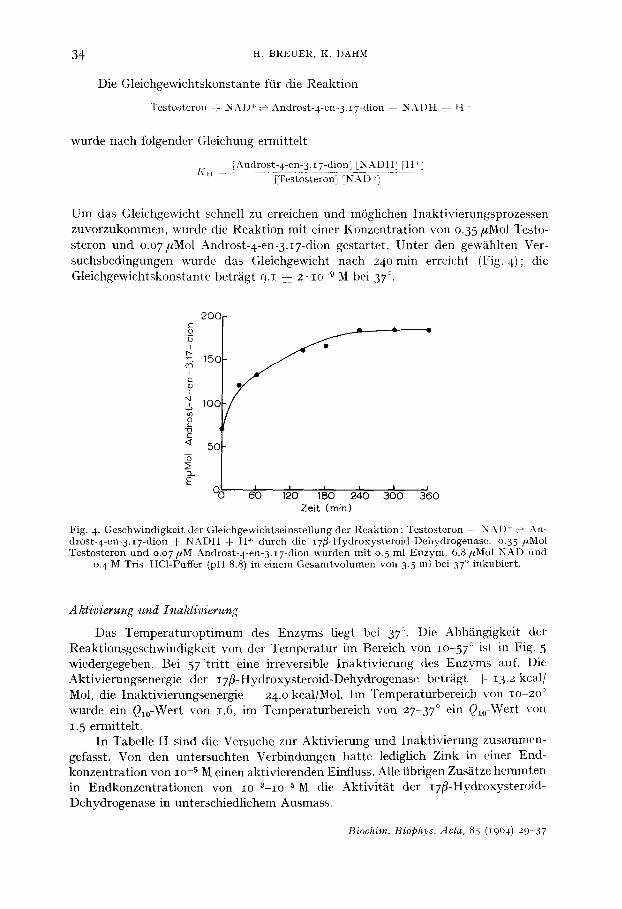
Die Gleichgewichtskonstante fiir die Reakt ion
Testosteron F NAD~-o Androst-4-en-3.i7-dion + N:kI)H T t l ,
wurde nach folgender Gleichung ermit te l t
[Androst-4-en- 3. t 7-dion] IN ADH] [H Ix" u . . . . . . . . . ['l'estosteroIi] [N-.S~I) +~ - -
U m das Gleichgewicht schnell zu erreichen und m6glichen Inakt ivierungsprozessen zuvorzukommen, wurde die Reakt ion mit einer Konzent ra t ion von o.35/~Mol Testo- s teron und o.o 7 #Mol Androst-4-en-3. i7-dion gestartet. Unte r den gew~ihlten Ver- suchsbedingungen wurde das Gleichgewicht nach 2 4 o m i n erreicht (Fig. 4); die Gleichgewichtskonstante betr~igt 9.1 ~_ 2- IO -9 M bei 37 °.
.0 ? L #
2
EL E
200
15C
IOC
50
f I I I
6o 12o 40 300 3;0 Zeit (rain)
Fig. 4. Geschwindigkeit der Gleichgewichtseinstellung der Reaktion: Testosteron -i NAD + ~ An- drost-4-en-3.i7-dion + NADH -}- H + dutch die i7fl-Hydroxysteroid-Dehydrogenase, o.35 /,Mol Testosteron und o.o 7/*M Androst-4-en-3.i7-dion wurden mit o.5 ml Enzym, 6.8/,Mol NAD und
o. 4 M Tris-HC1-Puffer (pH 8.8) in einem Gesamtvolumen von 3.5 ml bei 37 ° inkubiert.
Aktivierung und Inaktivierung
Das Tempera tu rop t imum des Enzyms liegt bei 37 °. Die Abh~ingigkeit der Reaktionsgeschwindigkeit v o n d e r Tempera tur im Bereich yon lO-57 ° ist in Fig. 5 wiedergegeben. Bei 57°tri t t eine irreversible Inak t iv ie rung des Enzyms auf. Die Aktivierungsenergie der i7/5-Hydroxysteroid-Dehydrogenase betr/igt ~ 13.2 kcal/ Mol, die Inakt ivierungsenergie - - 24.0 kcal/Mol. Im Temperaturbereich wm Io-2o ° wurde ein Q10-Wert yon 1.6, im Temperaturbereich yon 27-37 ° ein Q10-Wert wm 1.5 ermittel t .
I n Tabelle I I sind die Versuche zur Akt ivierung und Inak t iv ie rung zusammen- gefasst. Von den untersuchten Verbindungen hat te lediglich Zink in einer End- konzent ra t ion von IO 5 M einen akt ivierenden Einfluss. Alle iibrigen Zus/itze hemmten in Endkonzen t r a t i onen yon Io -S- Io -5 M die Aktivit~it der i7fl-Hydroxysteroid- Dehydrogenase in unterschiedlichem Ausmass.
Biochim. Biophys. Acta, 85 (1964) 29 37

I7fl-HYDROXYSTEROID-DEHYDROGENASE DER RATTENNIERE 35
- 3 . 7
--~'o -4.C
: I_
E
-4 .3 o
I I I J I I ~I 3.2 3.3 3.4 :~5 3.6
l x 1 0 3
Fig. 5. Abhi~ngigkeit der O x y d a t i o n yon Tes tos te ron zu Andros t -4 -en-3 . i7 -d ion du rch die I7fl- H y d r o x y s t e r o i d - D e h y d r o g e n a s e yon der T e m p e r a t u r . 0. 7 #Mol Tes tos te ron w u r d e n m i t 0.25 ml E n z y m , 6 .8/ ,Mol N A D u n d o. 4 M Tris-HC1-Puffer (pH 8.8) in e inem G e s a m t v o l u m e n yon 3-5 ml
ft~r I2o rain inkubier t . Die T e m p e r a t u r wurde im Bereich y o n ro -57 ° vari iert .
DISKUSSION
D i e v o r l i e g e n d e n U n t e r s u c h u n g e n z e i g e n , d a s s s i c h i n d e r 1 6 s l i c h e n F r a k t i o n d e r
R a t t e n n i e r e e i n e i 7 f l - H y d r o x y s t e r o i d - D e h y d r o g e n a s e b e f i n d e t , d i e s o w o h l m i t N A D
a l s a u c h m i t N A D P a l s C o f a k t o r r e a g i e r t . D i e s e s E r g e b n i s b e s t / ~ t i g t d i e k f i r z l i c h
v e r 6 f f e n t l i c h t e n A n g a b e n y o n AOSHIMA UND KOCHAKIAN 15, w o n a c h n u r d i e c y t o -
p l a s m a t i s c h e , n i c h t a b e r d i e m i k r o s o m a l e F r a k t i o n d e r R a t t e n n i e r e f i b e r e i n e N A D /
N A D P - a b h / i n g i g e I 7 f l - H y d r o x y s t e r o i d - D e h y d r o g e n a s e v e r f f i g t . D a s h i e r n a c h -
T A B E L L E I I
Z U S A M M E N S T E L L U N G D E R V E R S U C H E Z U R A K T I V I E R U N G U N D I N A K T I V I E R U N G
D E R I 7 f l - H Y D R O X Y S T E R O I D - D E H Y D R O G E N A S E D E R R A T T E N N I E R E
o. 7 #Mol Tes tos t e ron w u r d e n re.it o. 5 ml E n z y m , 5/~Mol NAD, 0. 4' M Tr is -HC1-Puffer (pH 8.8) u n d den jeweil igen Zus~ tzen (alle in w~ssriger L6sung) in e inem G e s a m t v o l u m e n y o n 3.5 ml ffir 60 rain bei 37 ° inkubier t . Vor Beg inn der I n k u b a t i o n wurde das E n z y m m i t d e m jeweiligen
Zusa t z ffir 5 rain pr '~inkubiert .
Zuscaz
Gebildete Xnderung Menge gegen~bev
Endkonzen#ation Androst-4-en. dem (M) 3.xT-dion Konlrollwert
(ra~Mol) (%)
Keiner - - 132 - - ZnC12 1°-8 3 ° - - 77
xo -5 I52 + 1 5 MgCl~ 10 -3 i 12 -- 15
I O - 5 9 8 - - 26 CRCI2 1o-3 93 - - 3 °
I O - s 96 - -27 Glu t a th ion i o -a 64 - - 5 i
xo -s 89 -- 32 J o d a c e t a m i d Io -8 61 -- 54
I O - 5 1 2 1 - - 8 o-JodosobenzoesAure TO--S 54 - - 59
lO -5 I16 - -12 p -Chlormercur ibenzoesgure lO-4 o -- ioo
Biochim. Biophys. A a a , 85 (z964) 29-37

3 6 H. BREUER, K. DAHM
gewiesene Enzym konntc durch fraktionierte F~illung des 2o ooo x g Uberstandes mit Ammoniumsulfat bei 5o%iger Ausbeute um etwa das I2-fache angereichert werden. Zum Vergleich sei angegeben, dass die bisher bekannt gewordenen An- reicherungen fiir die cytoplasmatische t7fi-Hydroxysteroid-Dehydrogenasen aus der Leber des Meerschweinchens das 5-lathe v, aus der Placenta des Menschen das 5o-fache ~ bzw. 25oo-faehe a, aus der Nebenniere der Rat te das 7(>lathe' und aus der Leber der Ratte das I7o-fache ~6 betragen.
Die I7fl-Hydroxysteroid-Dehydrogenase der Rattenniere oxydiert Oestradiol- I7fl in etwas geringerem Umfange als Testosteron. Demnach ist die Substratspezifitttt dieses Enzyms welt weniger ausgeprggt als diejenige des placentaren Enzyms, das nur phenolische Steroide angreift2, tv. Das pH-Opt imum der enzymatischen Dehy- drierung (Testosteron-~ Audrost-4-en-3.i7-dion ) liegt bei 8,7, wS, hrend die Hydrie- rung (Androst-4-en-3.i7-dion-> Testosteron) ein Optimum bei 6.6 zeigt. Aus dem Verlauf beider pH-Kurven geht hervor, dass der Umsatz im alkalischen Bereich etwa 4 mal gr6sser ist als im sauren. Ftir Testosteron betr/igt K m I8. 5 " IO .5 M,
wtihrend fiir NAD ein Km Wert yon 4 ° . IO '~ M gefunden wurde. Beide \Verte liegen in der fiir Hydroxysteroid-Dehydrogenasen bekannten Gr6ssenordnung (vgl. Ref. I). Ein Vergleich der Michaelis-Menten Konstanten lttsst erkennen, dass das Enzvm zu NAD eine etwas geringere Afflnitttt besitzt als zu Testosteron. Demnach scheint die Bildung des Enzym-NAD Komplexes bei der Dehydrierung yon Testosteron der geschwindigkeitsbestimmende Schritt zu sein.
Wegen ihrer relativ geringen spezifischen AktivitS, ten stellen tierische z7fi- Hydroxysteroid-Dehydrogenasen das Gleichgewicht zwischen oxydierter und re- duzierter Verbindung hmgsamer ein als entsprechende Enzyme bakteriellen Ur- sprungs. Unter den bier gewtthlten Vcrsuchsbedingungen wurde das Gleichgewicht nach 3-4 h erreicht; die Gleichgewichtskoi~stante KH betr~igt bei 3 7 9, I :_ 2 • io-" M und entspricht anntihernd den Werten fiir die,, z7fl-Hydroxyster(fid-Dehydrogenasen der menschlichen Placenta" (18 :q~ 5" IO :) M bei 35 °) und aus Pseudomom~s testo- steroni is (26. io '~ M bei 35°). Dutch eine Steigerung der Temperatur yon io auf 2o :~ nimmt die Reaktionsgesehwindigkeit um das 1.6-fache, durch eine Steigerung yon 27 ° auf 37 ° um das 1.5-fache zu. Die geringe Temperaturabh~ingigkeit der ()t0-\Verte spricht fiir eine geringc Thermohtbilit~tt des Enzvnls im Bereieh \'~m z,~ .}7 und damit fiir eine optimale Anpassung des Enzyms an das Substrat. Die Aktivierungs- energie des Enzyms betrtigt -k I3.2 kcal/Mol, dic lnaktivierungsenergie z4.o kcal/ M ol. Diese fiir eine I7fi-Hydroxysteroid-Dehydrogenase erstmalig mitgeteilten \\:crte liegen in dem fflr Dehydrogenasen zu erwartenden lntervall.
Die I7fi-Hydroxysteroid-Dehydrogenase dvr Rattenniere besitzt, fihnlich wie die NAD/NADP-abhSngigen Dehydrogenasen, fllnktionelle Stt-(;ruppen. l)iese k6nnen durch Schwermetallionen sowie durch typische SH-(;ruppen blockierende Verbindungen, wie 0-,lodosobenz(~e<iure und/5-Ctfloromercuribenzoes~ture, gehemmt werden. Besonders charakteristisch wird die Wirksamkeit der SH-Gruppen durch den aktivierenden Einfluss yon Zinkionen in geringer Konzentration (Io .5 M) her- vorgehoben. Dieser Befund spricht fiir die Beteiligung e, ines Zink-Sulthydryl- Protein Komplexes an der enzymatischen Dehydrierung von Testosteron 2".
I)ie hier nfitgeteilten Ergebnisse unterstiitzen die Annahme, dass die Nitre nicht nur durch die Ausscheidung yon wasserl6slichen Konjugaten, sondern ;ruth durch Oxydoreduktionen regulierend in den Stoffwechsel der Steroide eingreift.
Hiochim. 13iophys. Ac/a, 85 U964) '29-37

I7fl-HYDROXYSTEROID-DEHYDROGENASE DER RATTENNIERE 37
DANK
Die vorliegende Untersuchung wurde mit Untersttitzung der Deutschen Forschungs- gemeinschaft durchgeftihrt.
ZUSAMMENFASSUNG
Die cytoplasmatische Fraktion der Rattenniere enth~ilt eine i7fl-Hydroxysteroid- Dehydrogenase, die durch F~tllung mit Ammoniumsulfat etwa I2-fach angereichert werden konnte. Das Enzym zeigt gegentiber Testosteron und Oestradiol-I7fl an- n~thernd die gleiche spezifische Aktivit~tt und reagiert sowohl mit NAD als auch mit NADP als Cofaktor.
Das Optimum der Oxydation yon Testosteron zu Androst-4-en-3.I7-dion liegt bei einem pH-Wert yon 8.7; ftir die Reduktion von Androst-4-en-3.I7-dion zu Testo- steron wurde ein pH-Optimum yon 6.6 gefunden. Die Michaelis-Menten Konstanten betragen 18.5" lO -5 M ftir Testosteron und 40" lO -5 M ftir NAD. Die Gleichgewichts- konstante der Oxydation yon Testosteron zu Androst-4-en-3.I7-dion hat einen Weft von 9.1 .1o -9 M. Das Temperaturoptimum liegt bei 37°; die Aktivierungs- energie des Enzyms betr~igt + 13.2 kcal/Mol, die Inaktivierungsenergie -- 24.0 kcal/ Mol. Im Bereich von lO°-2o ° sowie im Bereich yon 270-37 ° wurden Q10-Werte von 1.6 bzw. 1.5 ermittelt.
Durch Zusatz yon SH-Gruppen blockierenden Verbindungen wird das Enzym gehemmt; Zinkionen (lO -5 M) i~ben einen aktivierenden Einfluss aus. Die physiolo- gische Bedeutung der i7fl-Hydroxysteroid-Dehydrogenase in der Rattenniere wird kurz besprochen.
L I T E R A T U R
1 H. BREUER, Hoppe-Seyler/Thierfelder Handbuch der Physiologisch- und Palhologisch-chemischen Analyse, Band 6/1, io. Auflage, Springer, Heidelberg, 1964, S. 423 .
2 L. J. LANDER UND L. L. ENGEL, J . Biol. Chem., 233 (1958) 538. 8 m. m. HAGERMAN UND C. A. VILLEE, J . Biol. Chem., 234 (1959) 2o31. 4 j . JARABAK, J. A. ADAMS, H. G. WILLIAMs-ASHMAN UND P. TALALAY, J. Biol. Chem., 237
(1962) 345. 5 M. L. SWEAT, L. T. SAMUELS UND ]~. LUMRY, d r. Biol. Chem., 185 (195 o) 75. 6 C. A. VILLEE UND J. M. SPENCER, J. Biol. Chem., 235 (196o) 3615.
G. L. ENDAHL UND C. D. KOCHAKIAN, Biochim. Biophys. Acta, 62 (1962) 245. 8 /-i. BREUER UND K. DAHM, Acta Endocrinol., 45 (1964) 47. 9 K. DAHM UND H. BREUER, Z. Physiol. Chem., im Druck.
10 H. J. PORTIUS END K. REPKE, Arch. Exptl. Palhol. Pharmahol., 239 (196o) 144. 11/-I. BREUER, R. OZON DND C. MITTERMAYER, Z. Physiol. Chem., 333 (1963) 272- 12 O. FOLIN UND V. CIOCALTEU, J. Biol. Chem., 73 (1927) 627. 13 H. BREUER UND L. NOCKE, Acta Endocrinol,, 29 (1958) 489 . 14 O. H. LowRY, •. J. ROSEBOROUGH, A. L. FARR UND R. J. RANDALL, J. Biol. Chem., 193 (1951)
265. 15 y . AOSHIMA UND C. D. KOCHAKIAN, Endocrinology, 72 (1963) lO6. 16 C. MITTERMAYER UND H. ]3REUER, Naturwissenschaften, 333 (1963) 272. x7 L. J. LANDER, J. A. ALEXANDER UND L. L. ENGEL, d r. Biol. Chem., 234 (1959) 2609. 18 p. "]['ALALAY UND P. I. MARCUS, J. Biol. Chem., 218 (1956) 675. 19 H. NETTER, Theoretische Biochemie, Springer, Heidelberg, 1959, S. 562. 20 K. WALLENFELS, Steric Course of Microbiological Reactions, Ciba Foundation Study Group
No. 2, Churchill, London, 1959, S. 32.
Biochim. Biophys. Acta, 85 (1964) 29-37





























