Embed Size (px)
Citation preview

Beitrag zur Wirkungsweise der dehydrierenden Enzyme von Milchsaurebakterien ;
von Alfred Bertho und Bruno von Zychlinski. [Aus dem Chem. Laboratorium der Bayer. Akad. der Wissenschaften
5u Munchen.] (Eiogelaufen am 30. Mai 1934.)
Vor einiger Zeit wurde von A. B e r t h o und H. Gluckl ) gezeigt, daB beim AtmungsprozeS katalasefreier, fakultativ anaerober Milch- saurebakterien der aufgenommene Sauerstoff zu praktisch 100 Proa. als Hydroperoxyd wiedergefunden wird und in seiner Acceptorwirkung durch Metbylenblau und Chinon ersetzt werden kann. I n diesem Falle waren nach unserer Ansicht die bekannten Forderungen der Dehy- drierungstheorie voll erfullt. Unsere Befunde wurden mittlerweile von verschiedener Seite bestatigt 2) 3). Bald nach dem Erscheineu der oben erwahnteu Arbeiten berichteteu 0. W a r burg und W. C h r i s t i a n 0 uber ein zuerst aus Hefe isoliertes gelbrotes Oxydationsferment vom Typus eines Ubertragerkatalysators, das imstande war, im Verein mit einem sogenanuten Zwischenferment und einem dazugehSrigen Co- ferment Hexosemonophosphat (Robisonester) in geringem Ma6 zu dehy- drieren.
Kuree Zeit danach wurde von W a r b u r g und Chr is t ian6) das gelbrote Ferment in dem von uns untersuchten B. Delbriickii auf- gefnuden und ohne Angaben von VersuchEdaten berichtet, daB die isolierten Fermente des obigen Schemas Robisonester in vitro bei Gegenwart von Sauerstoff unter Hpdroperoxydbildung zu oxydieren vermogen, und daB bei dieser Oxydation Sauerstoff durch MB ersetat werden kann. Die Bildung von Hydroperoxyd kommt nach diesen Autoren durch Reoxydation der Leukoform des gelbroten Ferments eustande. Ausfuhrlich mit Versuchen belegt wird dieses Atmuugs- schema in einer spateren Arbeit 6).
I)a nach W a r b u r g und C h r i s t i a n das gelbrote Fer- ment in vitro und in vivo gleich wirkt, ware nach diesen Autoren unsere Auffassung vom Atmungsvorgang der Milch-
Der chemische Verlauf der Reaktion blieb ungeklart.
l) A. 494, 159 (1932); Naturw. 19, 88 (1931). 8, I. G. D a v i s , Bio. Z. 265, 94; 267, 355 (1933). *) C1. F r o m a g e o t u. I. R O U X , Bio. Z. 267, 202 (1933). 4, Bio. Z. 264, 438 (1932).
Bio. 2. 266, 377 (1933). 5, Bio. 8. 260, 499 (1933).
Annalen der Chemie. 612. Band. 6

82 B e r l h o und von Zyychlinski,
saurebakterien abznlehnen. Wir miissen uns aus ver- schiedenen Grunden gegen diese Behauptung wenden. Zu- nachst ist in keiner der Arbeiten W a r b u r g s eine Angabe vorlianden, wonach es ihm gelungen ware, aus Milchsaure- bakterien Fermente vom Typus des Zwischenferments und Coferments zu isolieren, urn sie bei seinen Versuchen in vitro zu benutzen. Die Versuche wurden vielmehr mi t Zwischenferment aus Hefe und Coferment am Pferdeblut- korperchen durchgefiihrt. Schon aus diesem Grund ist die obige Behauptung nicht stichhaltig. Reiterhin ist fest- zustellen, daW die Versuche in vitro nur zu einer sehr un- vollstandigen Oxydation des Hexosephosphats fuhren und niernals einen respiratorischen Quotienten von 0,5 ergeben, wie ihn eine vollstandige Veratmung von Glucose bei gleichzeitiger Hydroperoxydbildung erfordert l).
TVir haben nach den Angaben W a r b n r g s 2 ) sein Zwischenferment aus Hefe, das zugehorige Coferment aus Pferdeblutkorperchen und das gelbrote Ferment ebenfalls aus Hefe hergestellt und die Oxydation von Hexosemono- phosphat mit Hilfe dieser 3 Fermente bei Gegenwart von Sauerstoff untersucht. Dabei entsprach der Sauerstoff- verbrauch etwa dem der Warburgschen Versuche, doch ist es uns dabei niemals gegluckt, in blausaurefreien An- satzen die Bildung von Hydroperoxyd nachzuweisen. duch
T a b e l l e I. Je 2 ccm ,,Zwischenfermcnt"-Ltisung, j e 1 ccm ,,Coferment"-Ltisung und LSsung des ,,gelben Ferments". J e 0,5 ccm m/lo-Hexosephosphat- 18snng und 2 ccm "/,,-Phosphatpuffer pH = 6,8, Gesamtvol. jeweils 10 ccm. Temp. 39,o ". Iieaktionsgas Liift. Zwei Teroperaturkontrollen.
m/l,-Rlausauiure
30 ., 11 45 ,, 18
ccm e/,o-Thiosulfat . . l) Siehe S. 81, Anm. 1. a ) Siehe S. 81, Anm. 4 u. 6.

Wirkungsweise der dehydrierenden Enzyme usw. 83
bei Gegenwart von Blausaure, die wegen des Katalase- gehaltes des Zwischenferments einer Hydroperoxydzersetzung wahrend des Versuchs entgegenwirkt, blieben die auf- gefundenen Hydroperoxydmengen weit hinter den zu er- wartenden. Die experimentellen Belege ergeben sich aus Tab. I.
In GefSiB 5 und 6 mit einem Zusatz von je 1 ccm n/lo-Hydro- peroxyd ist bei Versurhsende das Hydroperoxyd vollstandig verscb wun- den, und die Sauerstoffaufnahme wird durch den durch Hydroperoxyd- zersetzung entwickelten Sanerstoff weitgebend kompensiert. m/luo-Blau- silure bewirkt, wie ein Versuch zeigte, eine starke Hemmung dieser Hy droperoxydzersetzung.
Weitere Einwkinde, die wir gegen die oben angefuhrte Behauptung W a r b u r g s machen miissen, ergeben sich aus den Versuchsergebnissen, die wir m i t dem Walburgschen Fermentsystem bei Verwendung anderer Atmungssubstrate erzielten. Wir haben friiher gezeigt l), daB die Milchsanre- bildner Glucose, Methylglyoxal, Dioxyaceton und Glycerin- aldehyd zu veratmen vermiigen. Il’eiterhin ist durch Unter- suchungen von anderer Seite bekannt geworden, dad auch Milchsaure 2, nnd Brenztraubensaure 2, durch Milchsaure- bakterien oxydiert werden konnen. Falls die W a r b u r g - sche Behauptung zutrifft, da13 das System in vitro und in vivo gleich wirkt, ist zu erwarten, daB eben diese Sub- stanzen - vielleicht mit Ausnahme der Glucose, deren Ver- atmung erst nach vorausgehender Phosph’orylierung erfolgen wird - durch die isolierten Fermentkomponenten bei Gegen- wart von Sauerstoff angegriffen werden. Es miiBte dies urn so mehr der Fall sein, als auch lebende Unterhefe diese Substanzen des glycolytischen Zuckerabbaues ebenso wie mi t Sauerstoff mit MB oder Chinon zu oxydieren vermag3). Es hat sich jedoch gezeigt, dal3 ebensowenig wie Glucose die C,- und C,-Korper 1 (+)-Milchsaure, Athylalkohol, Brenz- t raubenshre und Methylglyoxal durch das isolierte W a r - burgsche System oxydiert werden. Aus den in Tabb. 11,
l) A. B e r t h o u. H. Gli ick, a. a .0 . 3 I. G. D a v i s , a. a. 0. 8, H. W i e l a u d u. B. C l a r e n , A. 492, 126 (1932); 609 ,182 (1934).
6*
84 B e r t h o und von Z y c h l i n s k i ,
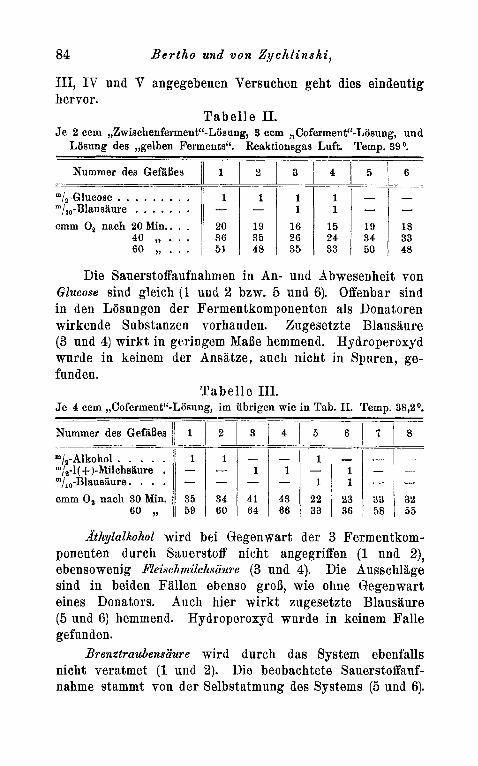
111, I V und V angegebenen Versuchen geht dies eindeutig hervor.
T a b e l l e 11. Je 2 ccm ,,Zwischenferment"-Losung, 3 ccm ,,Coferment"-LSsung, und
Losung des ,,gelben FermentdL. Reaktionsgas Luft. Temp. 39 O.
15 24 33
1 - 1 -
19 34 50
2
1
19 35 48
__ __
-
60 ,, . . .
3
1 1
16 26 35
_ _ _
I 51
Die Sauerstoffaufnahmen in An- und Glucose sind gleich (1 und 2 bzw. 5 und
18 33 48
Abwesenheit von ;). Offenbar sind
in den Losungen der Fermentkomponenteh als Donatoren wirkende Substanzen vorhanden. Zugesetzte Blausaure (3 und 4) wirkt in geringem MaBe hemmend. Hydroperoxyd wurde in keinem der Ansatze, auch nicht in Spuren, ge- funden.
T a b e l l e 111. Je 4 ccm ,,Coferrnent"-Lijsung, im ubrigen wie in Tab. 11. Temp. 38,2O.
Nummer des GefaEes . -
"/,-Alkohol . . . . . m/s-l( +)-Milchsaure . m/,,-Blausaure. . . . cmm 0, nach 30 Min.
60 1,
1 - 1
1 1 22 23 33 36
-
AthyZaZkohoZ w i d bei Gegenwart der 3 Fermentkom- ponenten durch Sauerstoff nicht angegriffen (1 und 2), ebensowenig Reischniilchsaure (3 nnd 4). Die Ausschlage sind in beiden Fallen ebenso grog, wie ohne Gegenwart eines Donators. Auch hier wirkt zugesetzte Blausaure (5 und 6) hemmend. Hydroperoxyd wurde in keinem Falle gefunden.
Brenztraubensiiure wird durch das System ebenfalls nicht veratmet (1 uiid 2). Die beobachtete Sauerstoffauf- nahme stammt von der Selbstatmung des Systems (5 und 6).
Wirkungsweise der dehydrierenden Bnzyme usw. 85
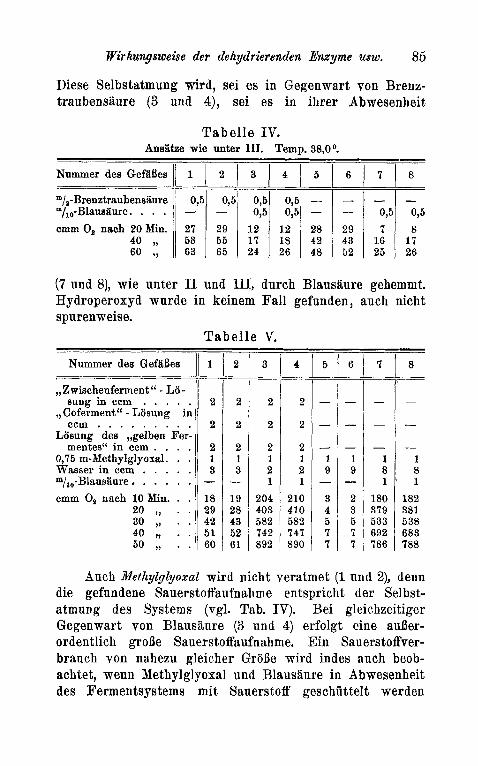
Diese Selbstatmung wird, sei es in Gegenwart von Brenz- traubensaure (3 nnd 4), sei es in ihrer Abwesenheit
1 ' 180 379 533 692 786
T a b e l l e IV. Ansatze wie unter 111. Temp. 38.04
_ _ - - - - 1 1 8 8
1
182 381 538 683 788
Nummer des GefiBes
m/,-Brenztraubensaure "/,,-Blaus&ure. . . .
,, Coferment '' - Losung in ccm . . . . . . . . .
Losung des ,,gelben Fer- mentes" in ccrn . . . .
0,75 m-Methylglyoxal. . . Wasser in ccm . . . . .
cmm 0, nach 10Min. . . 20 ,, . . 30 ,, . . 40 ,, . . 50 ,, . .
m/lo-Blausaure. . . . . ,
(7 und 8), wie unter I1 und 111, durch Blausaure gehemmt. Hydroperoxyd wurde in keinem Fall gefunden, auch nicht spurenweise.
T a b e l l e V.
2
2 1 3
18 29 42 51 60
-
- - 2 - -
2
2
2 1 3
19 28 43 52 61
-
____
2 2
2 2
2 2 1 1 2 2 1 1
204 210 403 410 582 582 142 141 892 890
Auch Methylglyoxal wird nicht veratmet (1 und 2), denn die gefundene Sauerstoffaufnahme entspricht der Selbst- atmung des Systems (vgl. Tab. IV). Bei gleichzeitiger Gegenwart von Blausiiure (3 und 4) erfolgt eine auber- ordentlich grobe Sauerstoffaufnahme. Ein Sauerstoffver- brauch von nahezu gleicher GroBe wird indes auch beob- achtet, wenn Methylglyoxal und Blausiiure in Abwesenheit des Fermentsystems mit Sauerstoff geschuttelt werden

86 B e r t h o und v o n Z y c h l i n s k i ,
(7 und 8). Diese Peststellung deckt sich mit den von C. V. S m y t h e l ) gemachten Beobachtungen. Methylglyoxal selbst ist , wie wir auch schon friiher beobachtet haben, nicht autoxydabelz). Der geringe Mehrverbrauch an Sauer- stoff von 3 und 4 gegeniiber 7 und 8 durfte in der Haupt- sache durch Donatorwirkung des Fermentsystems zustande kom men.
Nach diesen Befunden bleibt das Warburgsche gelb- rote Oxydationsferment in seiner zu dem noch unvoll- standigen IVirkung als Oxydationskatalysator auf das Substrat Hexosemonophosphat (Ro bisonester) beschrankt und kann niemals Aiispruch erheben, generell als Uber- tragerkatalysator fur den oxydativen Abbau anderer Atmungssubstrate zn gelten. hbgesehen davon, daB die Zwischenschaltung des gelbroten Fermentes die Tatsache einer Uehydrierung nicht aus der Welt schafft, bleibt zu mindesten f u r die erwahnten anderen Substrate nur die Dehydrierung niit Hilfe des 1)ehydrasesystems der Milcli- sgurebakterien im Sinne Wie la nd s diskutierbar.
Wir liaben weiterhin versucht, das Dehydrasesystem von B. DeZbrucRzi durch Plasmolyse und Autolyse der Bakterienleiber abzutrennen, wiewohl fruhere Versuche in einem anderen Fall ohne Ergebnis geblieben waren3). Zu diesem Zweck wurde einerseits das Bakteriensediment ab- wechselnd i n flussiger Luft gefroren und wieder aufgetaut und andererseits Autolyse unter Zusatz von Toluol ver- sucht. I n keinem Fall gelang es, mit Hilfe der erhaltenen zellfreien Autolysate irgendwelche Sauerstoffaufnahmen bei Gegenwart von Glucose zu erzielen. Stets blieb die Wirk- samkeit im abgeschleuderten Bakterienrest erhalten.
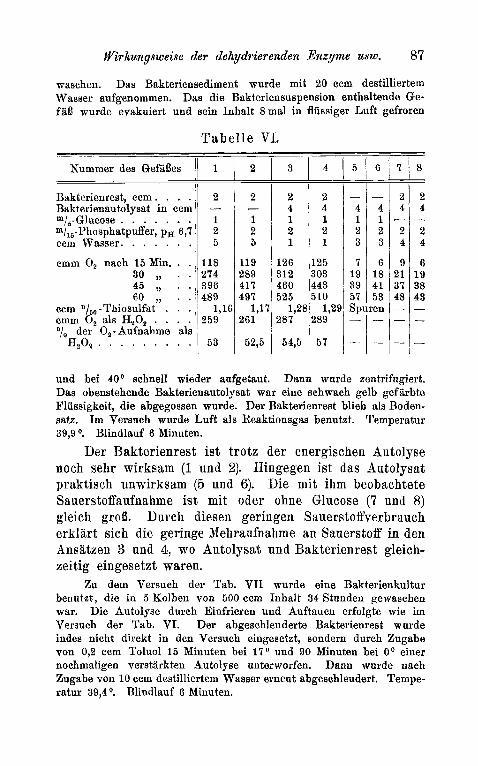
In Tab. VI ist ein Versuch niedergelegt zu dem Rakterien (B. DeZbriickii), die 24 Stunden in vier 500 ccm fassenden Kolben auf Bierwurze gewachsen waren, benutzt wurden. Die Zellen wurden abgeschleudert und 2 ma1 mit pbysiologischer Kochsalzlosung ge-
l) B. 65, 819, 1268 (1932).
3, H. W i e l a n d u. A. B e r t h o , A. 467, 95 (1928). A. B e r t h o u. H. G l u c k , a. a. 0.
Wirkungsweise der dehydrierenden Bnzyme usw. 07
wasehen. Das Bakteriensediment wurde mit 20 ccm destilliertem Wasser aufgenommen. Das die Bakteriensuspension enthaltende Ge- faB wurde evakuiert und sein Inhalt 8mal in flussiger Luft gefroren
T a b e l l e VI,
Bakterienautolysat in ccm
ml15-Phosphatpuffer, pH 6,7 ccm Wasser. . . . . . . cmm 0, nach 15 Min. . . 1
30 ,, . . 45 ,, . . 60 ,, . .
o/o der 0,-Aufnahme als
m/5-Glucose . . . . . . .
ccm n/,o-Thiosulftlt . . . cmm 0, als H,O, . . . .
R,O, . . . . . . . . .
- 1 2 5
118 1274 I396 1489
259
53
1,16
- __ 2
2
1 2 5
119 289 417 497
261
- -
-
1,1:
52,5
2 2 4 4 1 1 , 2 2 1 1
126 125 312 303 460 443 525 510
287 289 1,28 1,2!
54,5 57
___
5 i 6 - -
4 4 1 1 2 2 3 3
7 6 19 18 39 41 57 53 Spuren
und bei 40° schnell wieder aufgetaut. Dann wurde zentrifugiert. Das obenstehende Bakterienautolysat war eine schwach gelb gef iirbte Flussigkeit, die abgegossen wurde. Der Bakterienrest blieb als Boden- satz. Im Versuch wurde Luft als Reaktionsgas benutzt. Temperatur 39,9 O. Blindlauf 6 Minuten.
Der Bakterienrest ist trotz der energischen Autolyse noch sehr wirksam (1 und 2). Hingegen ist das Autolysat praktisch unwirksam (5 und 6). Die m i t ihm beobachtete Sauerstoffaufnahme ist mit oder ohne Glucose (7 und 8) gleich grob. Durch diesen geringen Sauerstoffverbrauch erklart sich die geringe Mehraufnahme an Sauerstoff in den Ansatzen 3 und 4, wo Autolysat und Bakterienrest gleich- zeitig eingesetzt waren.
Zu dem Versuch der Tab. VII wurde eine Bakterienkultur benutzt, die in 5 Kolben von 500 ccm Inhalt 34 Stunden gewaschen war. Die Autolyse durch Eiufrieren und Auftauen erfolgte wie im Versuch der Tab. VI. Der abgeschleuderte Bakterienrest wurde indes nicht direkt in den Versuch eingesetzt, sondern durch Zugabe von 0,2 ccm Toluol 15 Minuten bei 17" und 90 Minuten bei O o einer nochmaligen verstarkten Autolyse unterworfen. Dann wurde nach Zugabe von 10 ccm destilliertem Wasser erneut abgeschleudert. Tempe- ratur 39,4O. Blindlauf 6 Minuten.

88 B e r t h o u. v o n Z y c h l i n s k i , Behydyierende Enzyme usw.
- - 3 1 2 6
8 1 132 198 246
234 1,04
95,5
Tabel le VII.
- - 3
3 - 1 1 2 2 6 6
80 0 133 0 209 0 258 0
1 , l O - 246 -
-
95,5 I-
__ -
Nummer des GefidSes - -__ -
Bakterienautolysat in ccm 3 Bakterienautolysat nach
Zugabe v. Toluol in ccm - Bakterienrest in ccm. . . - m/,-Glucose . . . . . . . 1 m/,5-Phosphatpuffer, pH 6,7 2 Wasser in ccm . . . . . 6
"j0 d. 0,-Aufnahme als H,O, 1 -
3
3 3 1 2
88 138 201 259
246 95
-
1 , l O
3
3 3 1 2
95 142 210 265
235
-
1,05
88,5
Das Bakterienautolysat (1 und 2) ist wie im Versnch der Tab. V I kaum wirksam. Das gleiche gilt fur das durch Zugabe von Toluol erhaltene Autolysat (5 und 6). Hingegen ist der energisch durch Einfrieren und Auftauen und durch Zugabe von Toluol autolysierte Bakterienrest sehr stark wirksam (3 und 4). Durch Zusammengeben der beiden Antolysate mit dem Bakterienrest wird die Sauer- stoflanfnahme nicht wesentlich verandert (7 und 8).
Die vorliegende Arbeit wurde durch ein dem einen von UDB (v. Z.) gewahrtes 1. ff.-Stipelzdium ermaglicht, wofur wir auch an dieser Stelle bestens dsnken. Die Konigsstiftung xum A. v. Baeyer- Jzcbilaum unterstutzte die Arbeit in dankenswerter Weise durch Ge- wiihrung von Barmitteln.





























