Embed Size (px)
Citation preview

Respiration Physiology (1973) 19, 153-161; North-Holland Publishing Company, Amsterdam
CHANGES IN RED CELL 2,3-DIPHOSPHOGLYCERATE
CONCENTRATION AS CAUSE OF THE POSTNATAL DECREASE OF PIG BLOOD OXYGEN AFFINITY’
R. BAUMANN, F. TEISCHEL, R. ZOCH and H. BARTELS
Physiologisches Institut der Medizinischen Hochschule Hannover, Karl- Wiechert-Allee 9, Hannover 3,
Germany
Abstract. Oxygen affinity (expressed as oxygen half saturation pressure P,,) and red cell 2,3-diphos-
phoglycerate (2.3 DPG) concentration of miniature pig blood was examined in fetuses (955110 days
gestational age) and piglets from birth until 50 days post partum. Fetuses had the highest oxygen
affinity (P,, 23 mm Hg) and lowest 2.3 DPG concentration (2.40 mM 2.3 DPG/l, RBC). Both P,,
and 2.3 DPG concentration increased rapidly during the first week after birth towards adult levels.
Haemoglobin electrophoresis did not show different behaviour of haemoglobin obtained from foetal,
postnatal or adult pig blood. The oxygen equilibrium curves of the respective dialysed haemoglobin
solutions were identical and upon addition of 2,3 DPG the same decrease in oxygen affinity was ob-
served. It is concluded that the postnatal decrease in pig blood oxygen affinity is caused by a con-
comitant increase in red cell 2,3 DPG concentration.
Blood
Foetal
Haemoglobin
Miniature pig
Neonatal
Oxygen affinity
2.3 DPG
In most mammalian species foetal blood has a higher oxygen affinity than adult blood. Transition towards adult oxygen affinity usually takes place during the early postnatal period and is accomplished by different mechanisms. Either like in sheep or goats (Blunt et al., 197 1; Baumann, Bauer and Rathschlag-Schaefer, 1972) foetal haemoglobin is replaced by adult haemoglobin of intrinsically lower oxygen affinity or as for example in man (Bauer et al., 1969; Tyuma and Shimizu, 1969) foetal and adult haemoglobin show the same oxygen affinity but react differently with 2,3- diphosphoglycerate (2,3 DPG) with regulates the oxygen affinity of several species (Benesch and Benesch, 1969; Bunn, 1971). In addition variations in red cell pH can also play a role as cause for postnatal changes in oxygen affinity (Hilpert et al.,
1963).
Accepted for publication 23 June 1973.
’ Financial support by Deutsche Forschungsgemeinschaft is gratefully acknowledged.
153

154 R. BAUMANN. F. TEISCHEL. R. ZOCH AND H. BARTELS
If a haemoglobin reacts with 2.3 DPG like human haemoglobin, changes in the
concentration of 2,3 DPG should also lead to affinity changes. The possible role of
2,3 DPG concentration changes as cause for postnatal oxygen affinity changes was
investigated in miniature pigs, since Novy et ~1. (1967) found identical functional
characteristics of dialysed foetal and adult pig haemoglobin, while on the other
hand whole blood oxygen affinity differed largely. Like Tautz and Kleihauer (1972)
he was unable to detect any difference between the haemoglobin type present in
adult or foetal red cells, which would rule out a change in haemoglobin type as
cause for the different whole blood oxygen affinities. This led Novy et al. (1967)
to the conclusion that differences in red cell environment had to be responsible for
this phenomenon. The results of our study confirm his view and show that 2,3 DPG is
the only factor responsible for postnatal changes in pig blood oxygen affinity in
accordance with the study of Duhm and Kim (1972) which came to our attention
while this investigation was in progress.
Material and methods
Three miniature pig sows of mixed breed (Giittingen) and their offspring (21
piglets) were used for the investigation. In addition blood samples from eight
miniature pig fetuses2 of 95-110 days gestational age of the same breed were
examined.
Blood samplirlg
Blood was taken from the piglets by heart puncture in light halothane anesthesia
and by puncture of the superior vena cava from the sows. Sampling started immedia-
tely after birth and was repeated at daily intervals during the first week using 2-6
animals alternately. Sampling was continued at larger intervals until the 50th day
post purtum. Powdered sodium heparinate was used to prevent clotting.
Blood gus anulyses
Whole blood: Samples of blood were equilibrated for 20 min at 37 “C and a Pco2
of 40 mm Hg with water vapor saturated gas mixtures of known composition.
The pH of the equilibrated samples was measured with a Radiometer microglass
electrode Type G 297/G2 and pH meter 4 calibrated with Radiometer precision
buffers. Micromanometric analyses for 0, and CO, were made on 0.05 ml samples
after Van Slyke and Plazin (1961).
Oxygen capacity and oxygen affinity (expressed as oxygen half saturation
pressure P,, at plasma pH 7.4, 37 “C in whole blood) were calculated after Bartels
and Harms (1959). Haemoglobin was measured as cyanmethaemoglobin (Kleihauer
and Betke. 1957) and methaemoglobin with the method of Evelyn and Malloy (1938).
Haematocrit values were determined in duplicate with the Hawskley micro-method.
For assays of 2.3 DPG one volume of freshly drawn blood was added to nine
volumes of icecold 69/i perchloric acid and centrifuged after stirring. Aliquots of the
’ Kindly provided by Dr. R. Ruefer ( Max-Planck-Institut fiir experimentelle Medizin. G6ttingen)

2.3 DPG AND PIG BLOOD OXYGEN AFFINITY 155
supernatant were neutralized with 2 M K&O,. 2,3 DPG was measured using the
enzymatic test of Krimsky (1962). All enzymes and substrates for the test were
purchased from Boehringer Co., Mannheim.
Prepuration of haemoglobin solutions
Packed erythrocytes were washed three times with 0.9% NaCl and haemolyzed by
addition of one volume of distilled water to one volume of packed cells and centri-
fuged thereafter for 1 hr at 100,000 g. The clear supernatant was placed in Visking
bags and dialysed at 4 “C for three days against several changes of 0.05 M Bis-Tris
buffer in 0.1 M NaCl (pH 7.2 at 20 “C).
After dialysis the haemoglobin solution was diluted with the buffer to a final
concentration of 1 x lo-’ mM/Hb, and divided into three parts: A) no 2,3 DPG
added, B) 30 PM 2,3 DPG/g Hb added, C) 45 PM 2,3 DPG/g Hb added. Oxygen
equilibrium curves were then determined with the method of Benesch, MacDuff and
Benesch (1965) using the Hitachi 1247 Spectrophotometer. Haemoglobin electro-
phoresis was carried out on cellulose acetate strips (Tris-EDTA buffer pH 8.2) with
the Boskamp microphor electrophoresis system.
Results
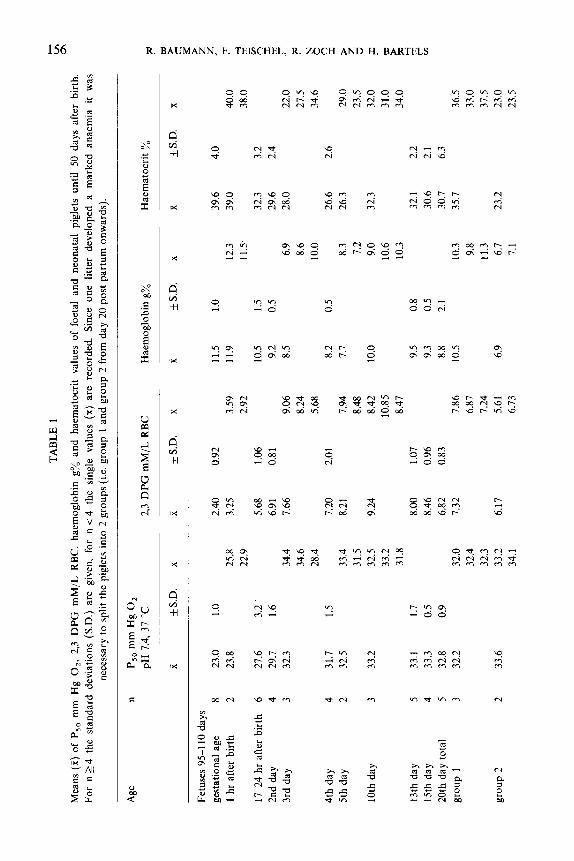
BLOOD 2.3 DPG CONCENTRATION AND P,,(table 1)
Fetuses showed the lowest P,, (23.0 mm Hg) and 2,3 DPG concentration (2.40
mM/L RBC). During the first day after birth both values increased to 27.6 mm Hg
P,, and 5.68 mM 2,3 DPG/L RBC respectively. By the fifth day post partum the
piglets had already established P,, and 2,3 DPG values comparable to adult ones.
Haemoglobin and haematocrit values (table 1) decline during the first five days after
birth and reflect a genuine decrease, since during this period no animal was bled
twice. In the later course however group 2 animals became markedly more anaemic
than group 1 animals since in the former group blood sampling had started at
birth whereas in the latter group it started by the third day posf partum. P,,
and 2,3 DPG values of group 2 animals were markedly elevated at the fortieth
day as compared to group 1 animals. whereas at all other dates no significant
difference was observed.
The P,O and 2,3 DPG of piglets 50 days after birth were slightly elevated in
comparison to the adult values like in all other species so far investigated. No
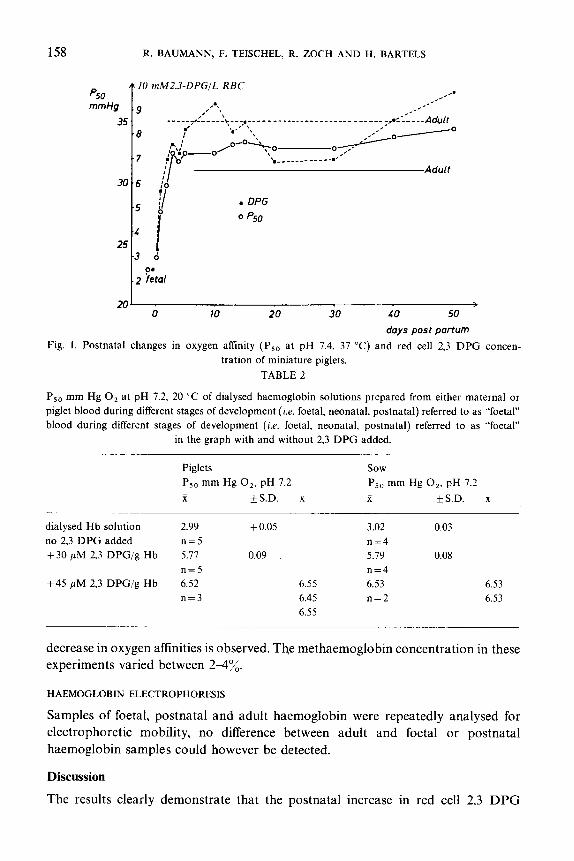
statement can be made about the significance due to the small sample size. Figure 1
shows the time course of the postnatal change in P,, and 2,3 DPG concentration.
DIALYSED HAEMOGLOBIN SOLUTIONS
Table 2 summarizes P,, values of dialysed haemoglobin solutions prepared from
adult blood and from foetal, neonatal and postnatal piglet blood respectively, with
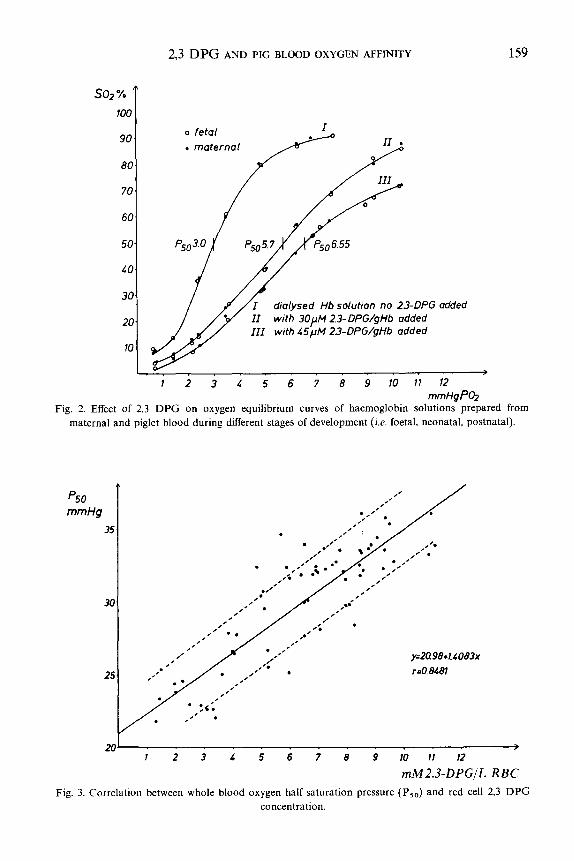
and without 2,3 DPG added. As can be seen from fig. 2 the oxygen equilibrium
curves of both groups are identical and upon addition of 2,3 DPG the same
TA
BL
E
1 5
Mea
ns
(X)
of
P,,
mm
H
g 0,
. 2.
3 D
PG
mM
/L
RB
C,
haem
oglo
bin
8%
and
haem
atoc
rit
valu
es
of
foet
al
and
neon
atal
pi
glet
s un
til
50
days
af
ter
birt
h.
For
n 2
4 th
e st
anda
rd
devi
atio
ns
(SD
.)
are
give
n,
for
n <
4 th
e si
ngle
va
lues
(x
) ar
e re
cord
ed.
Sinc
e on
e lit
ter
deve
lope
d a
mar
ked
anae
mia
it
was
nece
ssar
y to
sp
lit
the
pigl
ets
into
2
grou
ps
(i.e
. gr
oup
1 an
d gr
oup
2 fr
om
day
20 p
ost
part
um
onw
ards
).
Age
n
P,a
mm
H
g O
2 pH
7.
4,
37
‘C
x +S
.D.
x
2,3
DPG
m
M/L
R
BC
x
Hae
mog
lobi
n 8%
H
aem
atoc
rit
y0
&SD
. x
%
_+S.
D.
x x
&SD
. x
Fetu
ses
95-l
10
day
s
gest
atio
nal
age
8
1 hr
af
ter
birt
h 2
17-2
4 hr
af
ter
birt
h 6
2nd
day
4
3rd
day
3
4th
day
4
5th
day
2
10th
da
y 3
13th
da
y 5
15th
da
y 4
20th
da
y to
tal
5
grou
p 1
3
grou
p 2
2
23.0
23.8
27.6
29.7
32.3
31.7
32.5
33.2
33.1
33.3
32.8
32.2
33.6
1.0
25.8
22.9
3.2
1.6
34.4
34.6
28.4
1.5
33.4
31.5
32.5
33.2
31.8
1.7
0.5
0.9
32.0
32.4
32.3
33.2
34.1
2.40
3.25
5.68
6.9
1
7.66
7.20
8.21
9.24
8.00
8.46
6.82
7.32
6.17
0.92
1.06
0.8
1
2.01
1.07
0.96
0.83
3.59
2.92
9.06
8.24
5.68
7.94
8.48
8.42
10.8
5
8.47
7.86
6.87
7.24
5.61
6.73
11.5
11.9
10.5
9.2
8.5
8.2
7.7
10.0
9.5
9.3
8.8
10.5
6.9
1.0
12.3
11.5
.
1.5
0.5
6.9
8.6
10.0
0.5
8.3
1.2
9.0
10.6
10.3
0.8
0.5
2.1
10.3
9.8
11.3
6.7
7.1
39.6
39.0
32.3
29.6
28.0
26.6
26.3
32.3
32.1
30.6
30.7
35.7
23.2
4.0
40.0
7
38.0
;;
I
3.2
K
2.4
!z
22.0
.p
27.5
7
34.6
2.
6 ! z
29.0
>
23.5
3
32.0
?
31.0
34
.0
$
2.2
E
2.1
2
6.3
36.5
33.0
37.5
23.0
23.5

30th
da
y to
tal
grou
p 1
grou
p 2
40th
da
y to
tal
grou
p 1
grou
p 2
50th
da
y to
tal
grou
p 1
grou
p 2
Adu
lt
3 32
.8
6.86
2 32
.7
33.6
6.
88
31.9
1 32
.9
4 33
.7
3.0
8.60
2 32
.5
6.99
30.0
2 36
.0
10.2
0
36.3
3 34
.4
9.72
1 33
.6
2 34
.8
33.9
10
.22
35.7
2 31
.1
30.6
8.
48
31.7
8.6
7.16
10
.3
6.60
6.81
1.99
10
.7
7.47
11
.4
6.51
10.9
5 8.
8
9.46
9.5
8.70
11.1
0 7.
7
9.35
8.42
13
.1
8.53
__
31.0
8.4
32.5
29
.0
12.2
36
.0
5.2
28.0
1.4
32.9
5.
8
11.2
37
.2
37.0
11.6
36
.5
10.0
29
.0
32.0
7.5
26.0
t3
31.3
13.0
39
.5
;s
7.7
27.2
27
.5
8
7.7
27.0
12.9
38
.7
38.0
39.5
w
‘t
158 R. BAUMANN, F. TEISCHEL, R. ZOCH AND H. BARTELS
PSO I 10 mM2.3-DPGJL RBC
_*.
mmHs 9 ,,T _.-’
35 ,’ ‘\ _.*’
-----F----- ~___,._________________________--__,.-_:~___Adu[~
8 7’ “.: \\
8’ o-a+o 0
‘;_________--4’ .’
Adult .7
30 .6
-5
-4 25
.3
. DPG
0 650
d 0.
t 2 ietol
20’ 0 10 20 30
, 40 50
days past partum
Fig. 1. Postnatal changes in oxygen affinity (P,, at pH 7.4. 37 “C) and red cell 2,3 DPG concen-
tration of miniature piglets.
TABLE 2
Ps, mm Hg 0, at pH 7.2, 20 “C of dialysed haemoglobin solutions prepared from either maternal or
piglet blood during different stages of development (i.e. foetal, neonatal, postnatal) referred to as “foetal”
blood during different stages of development (i.e. foetal, neonatal, postnatal) referred to as “foetal”
in the graph with and without 2,3 DPG added.
Piglets sow
P,, mm Hg O,, pH 7.2 P,, mm Hg O,, pH 7.2
x +S.D. x x &SD. x
dialysed Hb solution 2.99 + 0.05 3.02 0.03
no 2.3 DPG added n=5 n=4
+30 PM 2.3 DPG/g Hb 5.77 0.09 5.79 0.08
n=5 n=4
+45 PM 2,3 DPG/g Hb 6.52 6.55 6.53 6.53
n=3 6.45 n=2 6.53
6.55
decrease in oxygen affinities is observed. The methaemoglobin concentration in these
experiments varied between 224%.
HAEMOGLOBIN ELECTROPHORESIS
Samples of foetal, postnatal and adult haemoglobin were repeatedly analysed for
electrophoretic mobility, no difference between adult and foetal or postnatal
haemoglobin samples could however be detected.
Discussion
The results clearly demonstrate that the postnatal increase in red ceil 2,3 DPG
2,3 DPG AND PIG BLOOD OXYGEN AFFINITY 159
SOz% h 100
90.
80.
70,
60.
50.
I dialysed Hb solution no 2.3-DPG added . 20. II
30. ,y
with 2.3- DPG/gHb added III with
30pM 15pM 2%DPG/gHb added
10.
,
1 2 3 I 5 6 7 8 9 10 11 12
mmHg& Fig. 2. Effect of 2.3 DPG on oxygen equilibrium curves of haemoglobin solutions prepared from
maternal and piglet blood during different stages of development (i.e. foetal. neonatal: postnatal).
PSO mmHg
35.
y=209844083x
r--O.8481
20 > 1 2 3 4 5 6 7 8 9 10 11 12
mM2.3-DPGJL RBC Fig. 3. Correlation between whole blood oxygen half saturation pressure (P5,,) and red cell 2.3 DPG
concentration.

160 R. BAUMANN, F. TEISCHEL, R. ZOCH AND H. BARTELS
concentration is responsible for the decrease of pig blood oxygen affinity during this period. as shown by fig. 3 which demonstrates the close correlation between P,, of whole blood and red cell 2,3 DPG concentration, The oxygen equilibrium curves of dialysed foetal, postnatal and maternal blood were identical and upon addition of 2,3 DPG the same shift in oxygen affinity is observed in agreement with Duhm and Kim (1972). Although some authors (Glauser and Glauser, 1971) hold the view that there is a foetal haemoglobin in pigs, this does not affect our conclusion, since as far as oxygen affinity and response to 2,3 DPG are concerned haemoglobin solutions derived from fetuses and adult pigs show the same characteristics.
The reason for the sudden increase in red cell 2,3 DPG concentration during the first few days after birth is unknown. While for human red cells it is established that an increase in red cell pH leads to increased synthesis of 2,3 DPG (Asakura et al., 1966), it is not clear if the large increase in blood pH of piglets during the first 48 hr after birth (,Randall, 1972) might act as a stimulus towards higher 2,3 DPG synthesis since no detailed data are available about glycolysis of neonatal pig red cells. Moreover, the fact that the glycolytic activity of adult pig red cells is by far the lowest of all animals so far investigated (MacManus and Kim, 1968) makes it difficult to apply the same principles that have been established for the regulation of 2,3 DPG metabolism in the human red cell.
References
Asakura. T., Y. Sato, S. Minakami and H. Yoshikawa (1966). pH dependency of 2.3 diphosphoglycerate
in red blood cells. Clin. Chim. Acta 14: 84%841.
Bartels, H. and H. Harms (1959). Sauerstoffdissoziationskurven des Blutes von Saugetieren. Pfliigars
Arch. ges. Phpsiol. 268: 334-365.
Batter, Ch., M. Ludwig, I. Ludwig and II. Bartels (1969). Factors governing the oxygen affinity of
human adult and foetal blood. Respir. P~~siol. 7: 211-277.
Baumann, R., Ch. Bauer and A. M. Rathschiag-Schaefer (1972). Causes of the postnatal decrease of
blood oxygen affinity in lambs. Respir. Physiol. 15: 1.51-158.
Benesch. R.. G. MacDuff and R. E. Benesch (1965). Determination of oxygen equilibrium curves with
a versatile new tonometer. Ancalyt. &o&em. 11: 8 l-87.
Benesch, R. and R. E. Benesch (1969). Intracellular organic phosphates as regulators of oxygen release
by haemoglobin. Nature 221: 618-622.
Blunt, M. H., J. L. Kitchens, S. M. Mayson and T. H. J. Huisman (1971). Red cell 2.3 DPG and
oxygen affinity in newborn goats and sheep. Proc. Sot. Exp. Bid. Med. 138: W-803.
Burnt, H. E. (1971). Differences in the interactions of 2.3 diphosphoglycerate with certain mammalian
hemoglobins. Science 172: 1049.
Duhm, J. and H. D. Kim (1972). Effect of the rapid postnatal increase of 2.3 diphosphoglycerate
concentration in erythrocytes on the oxygen afhnity of pig blood. IInd International Symposium
on Metabolism and Membrane Permeability of Erythrocytes, Thrombocytes and Leucocytes.
Vienna 14-16 June 1972 (in press).
Evelyn, K. A. and H. T. Malloy (1938). Microdetermination of oxyhaemoglobin, methaemoglobin and
sullhaemoglobin in a single sample of blood. J. Biol. Chem. 126: 655-662.
Glauser. C. G. and E. M. Gtauser (1971). A comparison of the hemoglobins occurring in fetal and
adult pigs. Proc. Sot. Exp. Bid. Med. 137: 144991451.
Hilpert, P., R. G. Fleischmann, D. Kempe and H. Bartels ( 1963). The Bohr effect related to blood and
erythrocyte pH. Am. J. Physiol. 205: 337-340.

2,3 DPG AND PIG BLOOD OXYGEN AFFINITY 161
Kleihauer, E. and K. Betke( 1957). Zur HImoglobinbestimmung mittels Cyanhamiglobin nach Betke und
Savelsberg. Arztf. Lab. 3: 202-205.
Krimsky, I. (1962). D-Glycerat 2,3 Diphosphat. In: Methoden der enzymatischen Analyse. Ed. H.-U.
Bergmeyer, Verlag Chemie, Weinheim/Bergstra,Be, pp. 238-240.
McManus, T. J. and H. D. Kim (1968). Energy metabolism in the pig red cell. In: I. International
Symposium on Metabolism and Membrane Permeability of Erythrocytes and Thrombocytes.
Heidelberg, Georg Thieme Verlag, pp. 4348.
Novy, M. J., A. S. Hoversland, R. D. Koler and J. Metcalfe (1967). A comparison of oxygen
affinity and hemoglobin type in the adult and fetal miniature pig. Federarion Proc. 26: 485.
Randall, G. C. B. (1972). pH values and blood gas tensions in the normal piglet during the first
48 hours of life. Biol. Neonate 20: 68-73.
Tautz, C. H. and E. Kleihauer (1972). Gibt es ein fetales Haemoglobin beim Schwein? II. Analysen des
Haemoglobins. Res. exp. Med. 159: 4449.
Tyuma, I. and K. Shimizu (1969). Different response to organic phosphates of human fetal and adult
hemoglobins. Arch. Biochem. Biophys. 129: 404405.
Van Slyke. D. D. and J. Plazin (1961). Micromanometric Analyses. Baltimore, The Williams and Wilkins
Company.





























