Embed Size (px)
Citation preview

Genetica (1969) 40:566-576
D E S Y N A P T I S C H E M U T A N T E N MIT U N T E R S C H I E D E N IM
U N I V A L E N T E N V E R H A L T E N
H. D. KLEIN
Institut ffir Genetik der Universit~t Bonn
(Eingegangen am 2-2. Mai, 1969)
After application of neutrons on dry seeds of Pisum sativum three reces- sive mutants were isolated showing irregularities in the course of meiosis. A cytogenetical analysis showed that at metaphase I, a varying number of univalents are formed, most likely as a result of reduced chiasma frequen- cies. At anaphase I, some univalents divide precociously in mutant 2932 but none do so in mutants 2989 and 2552. As a consequence, most cells of 2989 and 2552 built up more than two spindles at anaphase II, whereas the majority of cells in 2982 form two spindles. This situation is reflected in the frequency distribution of "gones per PMC" at tetrad stage. The discussion deals with the possible causes of univalent formation at meiotic prophase and the variability of univalent behaviour at anaphase I.
Einleitung
Mutan ten mit St6rungen im Stadienablauf der Meiosis s ind h~ufig,
insbesondere an pflanzlichen Objekten, beschrieben worden (zusam-
menfassende Darstel lungen u.a. bei JAIN, 1955, RILEY & LAW, 1965;
RILEY, t966). Eine groBe Gruppe innerha lb dieser Mutan ten nehmen
dabei Formen mit Abweichungen vom normalen Konjugat ionsverhal -
ten der homologen Chromosomen ein. Sie werden gew6hnlich in
asynaptische und desynaptische Mutan ten unter te i l t je nachdem, ob
die Homologenpaarung oder der Prozess der Chiasmenbi ldung gest6rt
ist. Die kausalen Grundlagen dieser St6rungen sind his heute unbe-
kannt . Unabh~ngig davon, ob Asynapsis oder Desynapsis vorliegt,
t re ten als gemeinsames Charakter is t ikum in beiden F~llen Univa len te
auf. Hinsichtl ich ihres Verhaltens im Ablauf der meiotischen Teilungen
lassen sich zwar einige allgemeingfiltige Regeln aufstellen, in Einzel-
heiten ergeben sieh jedoch erhebliche Unterschiede. Beispiele hierffir
liefern drei Mutan ten von P i s u m sa t i vum. Die Ergebnisse ihrer zytoge-
net ischen Bearbei tung werden im folgenden dargestellt.

D E S Y N A P S I S I N P I S U M 567
Material und Methode
Die drei Mutanten mit den Sortimentsbezeichnungen 2552, 2982 und 2989 gehen auf Neutronenbestrahlungen ruhender Samen yon Pisum sativum, Handelssorte "Dippes gelbe Viktoria", zuriick. Die Bestimmung der Spaltungsverh~iltnisse und die Vermehrung der Mu- tanten erfotgte fiber heterozygote M2- und M3-Pflanzen. Die mutierten Pflanzen zeigen in ihrer Morphologie und Anatomie sowie hinsichtlich ihrer Vitalit~tseigenschaften keine Ver~nderungen gegentiber der Nor- malform. Die GenwJrkungen sind ausschlieglich auf die Sporogenese gerichtet. Sie bewirken Meiosis-St6rungen, welche die Fertilit~t der mutierten Pflanzen so weitgehend einschr~nken, dab nur vereinzelt Samenansatz erfolgt.
Die Spaltungsergebnisse legen nach statistischer Priifung jeweils einen monomeren Erbgang mit Rezessivit~tt des mutierten Gens nahe. Zur Untersuchung der Mikrosporogenese wurden die Antheren junger Bltitenknospen in Alkohol-Eisessig 3 : 1 fixiert, mit Karmin- bzw. Orcein-Essigs~ure gef/irbt und zu Quetschpr~paraten verarbeitet.
Ergebnisse
Die Untersuchung des Pachyt~ins, die zur Unterscheidung zwischen a- und desynaptischen Mutanten von Bedeutung ist, ergab keine Ab- weichungen vom Normalverhalten. Die homologen Chromosomen waren in allen ]3ereichen ihres L~ingsverlaufs exakt gepaart. Die an- schliel3enden Stadien Diplot~n und Diakinese sind bei Pisum wegen der untibersichtlichen Kernverh~iltnisse ftir die mikroskopische Untersuchung ungeeignet, so dab eine Weiterverfolgung des meiotJ- schen Stadienablaufs erst wieder v o n d e r Metaphase I an erfolgen kann.
In der Metaphase I werden in allen drei Mutanten Univalente in wechselnden H~iufigkeiten vorgefunden (Abb. 1). Ftir ihr Zustande- kommen nimmt man Vorg~nge w~thrend der Konjugationsphase der homologen Chromosomen an, welche die Austauschfrequenz herab- setzen. Es handelt sich also im vorliegenden Falle definitionsgem~tl3 um desynaptische Mutanten. Einen n~iheren Einblick in das Ausmal3
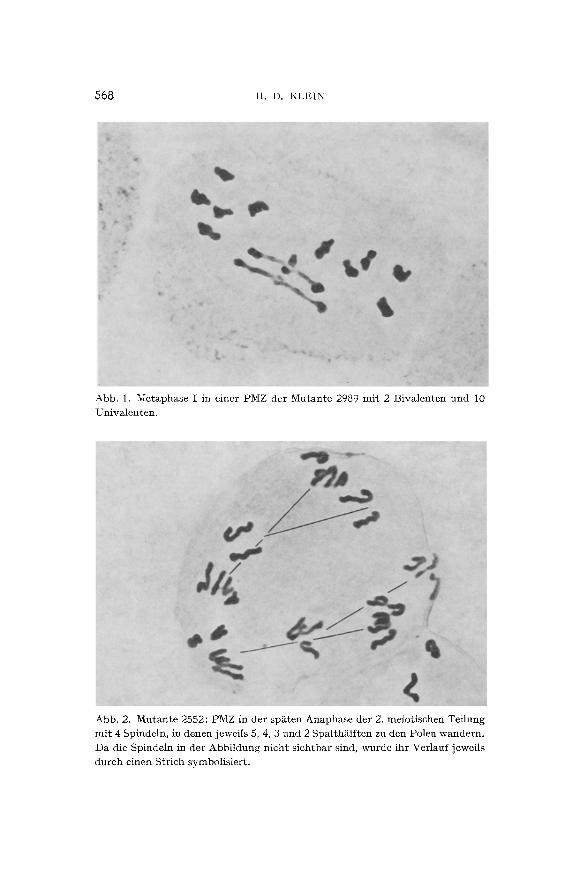
568 H . D . KLEIN
Abb. 1. Me taphase I in einer PMZ der M u t a n t e 2989 m i t 2 B iva len ten u n d 10 Univa len ten .
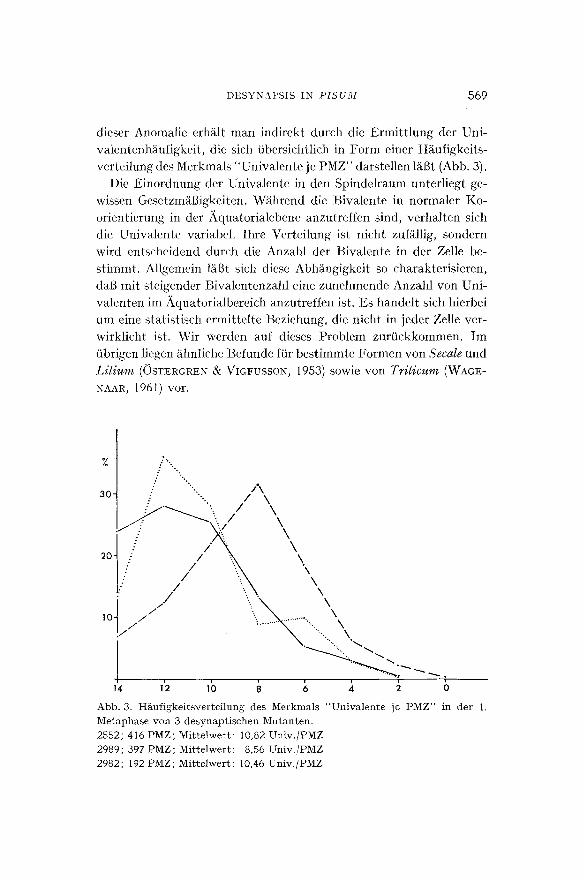
Abb. 2. M u t a n t e 2552: P M Z in der sp~ten A n a p h a s e der 2. meio t i schen Te i lung
mi t 4 Spindeln, in denen jeweils 5, 4, 3 und 2 Spalth~itf~en zu den Polen wande rn . Da die Spindeln in der Abb i l dung n ich t s i ch tbar sind, wurde ihr Verlauf jeweils
du rch e inen St r ich symbol is ier t .
DESYNAPSIS IN P I S U M 569
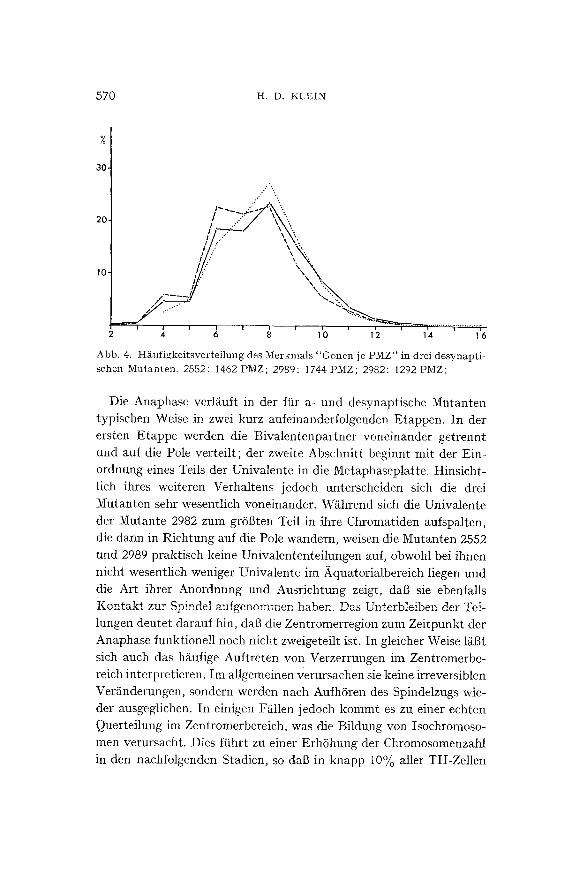
dieser Anomalie erh/ilt man indirekt durch die Ermit t lung der Uni- valentenh~iufigkeit, die sich tibersichtlich in Form einer H~iufigkeits- verteilung des Merkmals "Univalente j e PMZ" darstellen l~iBt (Abb. 3).
Die Einordnung der Univalente in den Spindelraum unterliegt ge- wissen Gesetzm/il3igkeiten. W~ihrend die Bivalente in normaler Ko- orientierung in der Aquatorialebene anzutreffen sind, verhalten sich die Univalente variabel. Ihre Verteilung ist nicht zuf/illig, sondern wird entscheidend durch die Anzahl der Bivalente in der Zelle be- stimmt. Allgemein 1/iI3t sich diese Abh/ingigkeit so charakterisieren, daS mit steigender Bivalentenzahl eine zunehmende Anzahl yon Uni- valenten im Aquatorialbereich anzutreffen ist. Es handelt sich hierbei um eine statistisch ermittelte Beziehung, die nicht in jeder Zelle ver- wirklicht ist. Wir werden auf dieses Problem zurtickkommen. Im tibrigen liegen tthnliche Befunde ftir bestimmte Formen von Secale und Lilium ((~STERGREN 8C VIGFUSSON, 1953) sowie yon Triticum (WAGE-
NAAR, 1 961) vor .
30" :" ""-.. / \ . . ~ . "-, / \
~ . - . . . . / / \ : " ~ \ : i :' \
2 0 - . / ... \ .: / "-.. .\
:" / '.. \ I:" / \ /
/ ' . ~ ............ \ ". . \ 10" / / . . . . . . . . . . . . . /
�9 .... \ \ / �9 ~ . """...... " ~ . ~
)4 f2 )b ; ; ~ ~ o Abb. 3. H&ufigkeitsvertei]ung des Merkmals "Univalente je P M Z " in der I, Metaphase yon 3 desynaptischen Mutanten, 2552; 416 PMZ; Mittelwert: 10,82 Univ./PMZ 2989; 397 PMZ; Mittelwert: 8,56 Univ./PMZ 2982; 192 PMZ; Mittelwert: 10,46 Univ./PMZ
570 ft. D. KLEIN
30-
. . ' \ .s .:
! .." \ ',
/ / . .... -,x;
2 4 6 8 I 0 12 14 16
Abb. 4. t t / iufigkeitsverteilung des Merkmals "Gonen je PMZ" in drei desynapti-
schen Mutanten. 2552:1462 PMZ; 2989:1744 PMZ; 2982:1292 PMZ;
Die Anaphase verl~tuft in der fiir a- und desynaptische Mutanten typischen Weise in zwei kurz aufeinanderfolgenden Etappen. In der ersten Etappe werden die Bivalentenpartner voneinander getrennt und auf die Pole verteilt; der zweite Abschnitt beginnt mit der Ein- ordnung eines Teils der Univalente in die Metaphaseplatte. Hinsicht- lich ihres weiteren Verhaltens jedoch unterscheiden sich die drei Mutanten sehr wesentlich voneinander. W~thrend sich die Univalente der Mutante 2982 zum gr613ten Teil in ihre Chromatiden aufspalten, die dann in Richtung auf die Pole wandern, weisen die Mutanten 2552 und 2989 praktisch keine Univalententeilungen auf, obwohl bei ihnen nicht wesentlich weniger Univalente im Aquatorialbereich liegen und die Art ihrer Anordnung und Ausrichtung zeigt, dab sie ebenfalls Kontakt zur Spindel aufgenommen haben. Das Unterbleiben der Tei- lungen deutet darauf bin, dab die Zentromerregion zum Zeitpunkt der Anaphase funktionell noch nicht zweigeteilt ist. In gleicher Weise l/tl3t sich auch das h~ufige Auftreten yon Verzerrungen im Zentromerbe- reich interpretieren. Im allgemeinen verursachen sie keine irreversiblen Ver~inderungen, sondern werden nach Aufh6ren des Spindelzugs wie- der ausgeglichen. In einigen F~illen jedoch kommt es zu einer echten Querteilung im Zentromerbereich, was die Bildung von Isochromoso- men verursacht. Dies fiihrt zu einer Erh6hung der Chromosomenzahl in den nachfolgenden Stadien, so dab in knapp 10yo aller TII-Zellen

D E S Y N A P S I S I N P I S U M 571
(Mutante 2552) mehr als 28, maximal 31 Chromosomen zu z~ihlen sin& Das Unterbleiben oder die vorzeitige Durchfiihrung der Aquations-
teilung hat ftir die Anordnung der Chromosomen und den weiteren Ablauf der Meiose folgende Konsequenzen:
In der Mutante 2982 wird das Kernmaterial im Zuge der Anaphase im wesentlichen auf die beiden Polbereiche verteilt ; dazwischen liegen einzelne Chromosomen oder Chromatiden, die die Pole nicht erreicht haben. In den Mutanten 2989 und 2552 hingegen erfolgt eine Konzen- trierung in drei oder vier Kernen, von denen 1 bis 2 im Mittelbereich und jeweils einer an den Polen liegen. In der Interkinese lind in der Metaphase I I wird diese Verteilungssituation ohne wesenttiche Ver~tn- derungen wiederholt. In Obereinstinmmng damit werden in der zwei- ten Meiose in der Mutante 2982 vorwiegend zwei Spindeln gebildet, w~ihrend in den beiden anderen Mutanten aberwiegend 3 und 4, verein- zelt sogar 5 Spindeln ausdifferenziert werden (Tabelle 1 und Abb. 2). Durch die Ausbildung yon mehr als 2 Spindeln in 60~o bzw. 86~o bei den Mutanten 2989 und 2552 ist die Mehrzahl der Gonen und Pollen- k6rner in diesen F~illen stark hypohaploid.
TABELLE I
ANAPHAS]E II. ANZAHL DER SPINDELN IN DEN PMZ D]~R MUTANTEN
2982, 2989 UND 2552
Spindeln 2982 2989 2552 Zellen % Zellen % Zellen %
2 94 93,8 124 40,0 9 13,8 3 7 6,2 136 43,6 33 50,8 4 -- 45 14,5 20 30,8 5 -- 6 1,9 3 4,6
101 100,0 311 100,0 65 100,0
Vorzeitig geteilte Univalente wirken sich auf den Ablauf der zweiten Teilung ungiinstig aus, weil die Spalth~itften funktionell ungeteilt sind ; sie k6nnen sich nicht ein zweites Mal teilen. In der Regel bleiben sie im Mittelbereich in wechselnder Entfernung yon den Polen liegen und veranlassen spiiter die Bildung yon Mikrogonen. H~iufig weisen sie Verzerrungen in den Zentromerregion auf, die jedoch nur vereinzelt in vollst~indige Querteilungen iibergehen. Da die Mutanten 2552 und 2989

572 H.D. KLEIN
fast keine vorzeitigen Univalententeilungen aufweisen, sind St6rungen dieser Art hier entsprechend selten.
Die Ausbildung zus~itzlicher Spindeln und das weitgehende Fehlen von "laggards" begiinstigen die Ausbildung von PMZ mit geraden Gonenzahlen. Diese Tatsache spiegelt sich im Kurvenverlauf der Abb. 4 wieder, in der die H~tufigkeitsverteilung des Merkmals "Gonenzahl je PMZ" fiir die Mutanten 2989 und 2552 im Vergleich zu der Mutante 2982 mit vorzeitiger Univalententeilung dargestellt ist. W~hrend die Kurve im letzteren Falle gleichm~il3ig ansteigt, ist fiir 2989 und 2552 ein treppenf6rmiger Anstieg zu verzeichnen: die Anteile 5- und 7- goniger PMZ sind relativ gering, da die H~iufigkeit der Mikronuclei- Bildung zwischen den Polbereichen der TH-Kerne wesentlich ver- mindert ist.
Die geschilderte Situation in den Mutanten 2989 und 2552 bedingt zwar optisch fiir die zweite Teilung ein h6heres NaB an Ordnung, im Hinblick auf die chromosomale Ausstattung der TiI-Kerne hat sie jedoch keine positiven Konsequenzen : Die Anzahl genomatisch ausge- wogener Gonen bleibt auch in diesem Falle eine Funktion der H~tufig- keit und des Verteilungscharakters der Univalente in der Metaphase I.
Diskussion
URSACHEN D E R U N I V A L E N T E N - E N T S T E H U N G
Ffir die Entstehung der Univalente in desynaptischen Mutanten wird gew6hnlich eine Verminderung der Chiasmenbildung verant- wortlich gemacht. Daneben besteht jedoch eine zweite M6glichkeit, die bereits von MOFJ?ET (1932), BEADLE (I933) und sp/iter u.a. yon NAYYAR (1965) diskutiert wurde. Danach sollen Chiasmen in unver~in- derter H~iufigkeit ausgebildet sein, jedoch schneller als gew6hnlich terminalisiert und noch vor Erreichen der Metaphase I aufgel6st wer- den. Eine Mutante dieser Art ist zwar bisher noch nicht bekannt, die Beeinflul3barkeit dieser Vorg~inge wurde jedoch nachgewiesen (STRAUB, 1936; Univalenten-Entstehung durch experimentelle Beein- flussung des Terminalisationsprozesses). Sollte diese letztere Vorstel- lung far unsere Mutanten zutreffen, so w~re zu erwarten, dab beziig- lich der Lage der Chiasmen in den Bivalenten eine kontinuierliche

D E S Y N A P S I S I N P I S U M 573
Reihe von proximal nach distal anzutreffen ist, wobei die Mehrzahl der Chiasmen an den Chromosomenenden liegen sollte. Dies ist jedoch nicht der Fall: Die Chiasmen sind nicht gleichm~13ig iiber die Chromo- somenschenkel verteilt, sondern liegen, wie in der Meiose der Normal- form, in der t~berwiegenden Mehrzahl proximal und interstitiell, w~ihrend terminale Chiasmen extrem selten sind.
Der exakte Nachweis, welcher der beiden Mechanismen wirksam wird, ist nur durch die Ermitt lung der Austauschh~tufigkeit zwischen Markierungsgenen zu erbringen. Eine Ver~nderung der Chiasmafre- quenz muB von einer entsprechenden Senkung der Crossover-Werte begleitet sein, w~ihrend Ver~inderungen im Terminalisationsgeschehen diesbeztiglich keinen EinfluB nehmen kSnnen. Eine solche Kombina- tion zytologischer lind genetische Methoden wurde yon ENNS & LARTER (1942) far eine desynaptische Mutante yon Hordeum vorge- nommen. Sie konnten zeigen, dab mit der Verringerung der Chiasmen- h~iufigkeit eine Senkung der Austauschwerte zwischen zwei Markie- rungsgenen von 40-43% auf 14-16~ verbunden ist. Damit war die Reduktion der Chiasmenfrequenz als Ursache ftir die Univalenten- Entstehung erwiesen.
Sollte das in unserem Falle ebenfalls zutreffen, so bleibt noch die Frage often, welches die Grtinde fiir diese verminderten Austausch- h~iufigkeiten sind.
W~ihrend man his vor kurzem annahm, dab der Nachweis einer lichtmikroskopisch exakten Paarung im Pachyt~n ausreiche, die St6rungen ausschliel31ich im Crossover-Prozel3 zu suchen, wird heute vielfach die Meinung vertreten, dab auch submikroskopische Ver~inde- rungen im Paarungsverhalten in Betracht zu ziehen sind (RILEY, 1966, RILEY 6: LAW, 1965). Danach w~ire die eigentliche Ursache ft~r das Ausbleiben von Chiasmen in Vorg~ingen w~ihrend der Paarung zu suchen, die auf submikroskopischer Ebene ablaufen und die ordnungs- gem~iBe Durchft~hrung des Crossover-Prozesses ausschliegen. Die Paarung w~ire in diesem Falle als "nicht-effektiv" zu bezeichnen.
Wir haben an anderer Stelle (KLEIN, 1969) die M6glichkeit erwogen, dab der gesamte Konjugationsprozess aus mindestens drei Teil- schritten besteht : 1. Exakte und enge Paarung 2. Auftreten von Brtichen an korrespondierenden Stellen yon Nicht-
Geschwister-Chromatiden homologer Chromosomen

574 H.D. KLEIN
3. 13berkretlz-Verheilung der Bruchfl~tchen ist einer dieser Prozesse gest6rt, so kann kein Crossover zustande
kommen. Das, was man als "nicht-effektive Paarung" bezeichnet, k6nnte dann m6glicherweise darin bestehen, dab entweder weniger Brtiche als normal eintreten, oder dab bei gleicher Bruchfrequenz ein Teil der Bruchfl~tchen restituiert und nicht tiberkreuz rekombiniert wird, wie es zur Entstehung eines Chiasmas notwendig w~ire.
U N I V A L E N T E N V E R H A L T E N IN D E R 1. ANAPEIASE D E R MEIOSIS
Treten als Folge yon St6rungen im Konjugationsgeschehen Univa- lente auf, so wird sich ein Teil von ihnen in der Aquatorialebene der Metaphase I einfinden. Wie grog dieser Anteil im Durchschnitt ist, h~ingt einmal vonde r Anzahl der Bivalente (siehe S. 003) alo (KLEIN, 1968) und wird zum anderen auch yon bestimmten strukturellen Eigenschaften der Chromosomen bestimmt. Zeigen die Chromosomen n~imlich eine ausgepfiigte Tendenz, untereinander zu verkleben ("stickiness"), so werden mehr Univalente als sonst tiber Verbindung mit den Bivalenten im Mittelbereich der Zelle fixiert. Sie besitzen somit die lagem~il3ige Voraussetzung ftir eine vorzeitige Aquationstei- lung. In der desynaptischen Mutante 500 unseres Sortimentes (KLEIN, 1968), die einen durchschnittlichen Univalentenanteil yon 5, 17 je PMZ aufweist, werden wegen einer sehr starken Verklebungsbereit- schaft naheza alle Univalente im Aquatorialbereich angetroffen und wir registrieren eine praktisch 100% -ige Vorwegnahme der Aquations- teilung. Das gleiche trifft ftir eine Form yon Scilla zu (REEs, 1952), obwohl hier "stickiness" nicht vorliegt. Das zeigt im tibrigen, dal? unabh~ngig v o n d e r Bivalentenzahl und der "stickiness" zus~itzliche Faktoren ftir die Einordnung der Univalente in die Metaphase-platte der 1. meiotischen Teilung zust~indig sein mtissen.
Die Tatsache, dab Univalente zur Zeit der 1. Teilung im Aquatorial- bereich liegen, besagt jedoch noch nicht, dab sie tats/ichlich die Aquationsteilung durchffihren. Das beweisen die Mutanten 2552 und 2989 mit fehlender Univalententeilung in der Anaphase I, obwohl bei ihnen nicht weniger Univalente als in der Mutante 2982 die lage- m~Bige Voraussetzung hierftir haben. Vergleichbare VerMltnisse sind in der Literatur aus den Arbeiten von LEVAN (1940) an Allium und

DESYNAPSIS IN P I S U M 575
von Ross, SANDERS ~r FRANZKE (1960) an Sorghun~ bekannt geworden. Neben diesen Formen mit 100%-iger Vorwegnahme und fehlender Aquationsteilung der Univalente in der Anaphase I sind am h~iufigsten ~Verte realisiert, die zwischen diesen Extremen liegen. Welche Grfinde fiir dieses unterschiedliche Verhalten der Univalente maBgebend sind, l~Bt sich nicht beurteilen. Line Erkl~irungsm6glichkeit bestiinde darin, dab fiir den Ablauf der Anaphase I eine unterschiedlich lange Zeit ver- fiigbar ist. Gewisse Anzeichen dafiir liegen in unserem Falle vor. Die durchschnittlichen Antherengr6Ben, die man als ZeitmaB verwenden kann, sind bezfiglich eines bestimmten Entwicklungsstadiums des Archespors in den Mutanten 2552 und 2989 etwas kleiner als die Ver- gleichswerte in der Mutante 2982 und in der Normalform. Die Unter- schiede sind jedoch so gering, dab sie statistisch nicht abgesichert werden k6nnen. Neben dem Zeitfaktor mug man auch die M6glichkeit in Betracht ziehen, dab die Ursachen Ifir das unterschiedliche Univa- lenten-Verhalten in bestimmten strukturellen Besonderheiten der Chromosomen selbst, insbesondere ihrer Zentromerregionen zu suchen sind. Hierffir spricht, dab die Spindel in den Mutanten 2552 und 2989 Kontakt mit den Univalenten aufgenommen hat, die Zentromerregion jedoch nicht - wie die ausgedehnten Verzerrungen in diesem Bereich zeigen - in der Lage ist, sich bereits zu diesem Zeitpunkt in zwei voll funktionsfithige Untereinheiten aufzugliedern.
Die Arbei t wurde aus Mit te ln des Bundes ln in i s t e r s fiir Wissenscha f t l i che
F o r s c h u n g der B u n d e s r e p u b l i k un te rs t i i t z t . I ch bedanke reich bei H e r r n Prof . Dr. GOTTSCHALK fiir die kr i t i sche Durch -
s ich t des Manusk r ip t e s .
LITERATUR
BEADLE, G. W. (1933). F u r t h e r s tud ies of a s y n a p t i c maize. Cytolog{a 4: 269-287. ENNS, H. & 2 , N. LARTER (1962). L inkage re la t ions of ds: a gene govern ing
ch romosome behav i ou r in ba r ley and its effect on genet ic r ecombina t ion . Can. J. Genet. Cytol. 4: 263-266.
JAIN, S. 1~. (1955). Male-s ter i l i ty in f lowering p lan ts . Bibliographia Genet. 18:
101-166. KLEIN, H. D. (1968). Die Bee in f lussung der Meiosis du rch m u t i e r t e Gene.
Disse r t a t ion Math . Nat . Fak. Univ . Bonn.
KLEIN, i . D. (1969). Asynaps i s and ex tens ive c h r o m o s o m e b reakage in Pisum. I m Druek.

576 H . D . KLEIN
LEVAN, A. (1940). The cytology of Allium amplectens and the occurrence in na ture of its asynapsis. Hereditas 26 : 353-394.
MOFFETT, A. A. (1932). Chromosome studies in Anemone. I. A new type of chiasma behaviour. Cytologia 4: 26~37.
lXIAYYAR, R. P. (1965). IZaryotype studies in the genus Netopterus (Lacepede).
The occurrence and fate of univalent chromosomes in spermatocytes of N. chitala. Genetica 36: 398-405.
(~STERGREN, G.I• ]~. VIGFUSSON (1953). On position correlation of univalents and quasi-bivalents formed by sticky univalents. Hereditas 39: 33-50.
REES, H. (1952). Asynapsis and spontaneous chromosome breakage in Scilla. Heredity 26: 353-394.
RILEY, R. (1966). Genetics and the regulation of meiotic chromosome behaviour. Sci. Progr. 193-207.
RILEY, R. & C. N. LAW (1965). Genetic variation in chromosome pairing. Advances in Genetics 13:57-114.
l~oss, J. G., M. ]~. SANDERS & C. J. FRANZKE (1960). Asynapsis in Sorghum. Hereditas 46: 470-480.
STRAUB, J. (1936). Untersuchungen znr Physiologie der Meiosis. II. Z. Bot. 30: 1-57.
WAGENAAR, E. B. (1961). Cytological studies of the development of metaphase I in Triticum hybrids. II. The behavioar of univalents in meiotic cell division. Can. J. Genet. Cytol. 3: 204-225.





























