Embed Size (px)
Citation preview

5.1 Einführung
Die warm-gemäßigte Klimazone liegt ungefähr zwi-schen den Wendekreisen (23°30') und dem 35. Brei-tengrad am Rande der Tropen („Subtropen“). Sie isteine Übergangszone zwischen den durch Strahlungs-überschuss charakterisierten Tropen und der durchmarkante solarklimatische und thermische Jahreszei-ten gekennzeichneten kühl-gemäßigten (nemoralen)Zone (Lauer & Rafiqpoor 2002). Das Klima der Sub-tropen zeichnet sich also (auf Meeresniveau) durchwarme bis heiße Sommer aus, wobei mindestenssechs Monate Mitteltemperaturen von über 18 °Cerreichen. Im Winter können Fröste bis zu –10 °Cauftreten (s. Abschn. 1.3.1, Tab. 1-8). Die Grenze zwi-schen Tropen und Subtropen ziehen Köppen-Tre-wartha bei einer Jahresisotherme von +18 °C. Diezonale Vegetation besteht überwiegend aus immer-grünen Laubwäldern, die sich von den tropischenWäldern durch geringere Wuchshöhe, weniger Artenund kompaktere, kleinere Blätter unterscheiden.Frostempfindliche Baumarten fehlen. Charakteris-tisch für die Subtropen ist das verstärkte Auftretenvon Nadelhölzern, die in den Tropen nur in Gebirgeneine gewisse Rolle spielen.
In die warm-gemäßigte Zone fallen auch zahlrei-che markante Hochgebirge (Burga et al. 2004). Nebeneinem relativ kleinen Anteil der Anden zwischenetwa 25 und 35° S gehören in Mittel- und Nordame-rika die westliche Kette der Sierra Madre und derSüdteil der Sierra Nevada in Kalifornien dazu. Dasgrößte subtropische Hochgebirge ist aber zweifel-sohne der Himalaya mit dem Hochland von Tibetsowie Karakorum und Hindukusch. In Afrika wer-den der Atlas (Marokko) und die Drakensberge(Südafrika), in Europa die Sierra Nevada (Spanien),der Apennin mit den Abruzzen (Italien), der Taurus(Türkei) und das Dinarische Gebirge (Dinariden) aufdem Balkan sowie einige kleinere, isolierte Berge wie
der Olymp zu den Subtropen gestellt. Den meistendieser Gebirge ist ein höherer Anteil an Nadelhölzerngemeinsam, der besonders auf der Nordhemisphäreausgeprägt ist und dort in den hochmontanen Nadel-wäldern borealen Charakters („Taiga“) zum Aus-druck kommt; die Vegetation der alpinen Stufe trägtmit ihren Gras- und Zwergstrauchtundren häufigschon polare Züge. Großrosettenpflanzen fehlen. Inden trockensten, oft wüstenartigen Gebirgen (vorallem Zentralasiens) können auch Dornpolster auf-treten. Hartpolster sind vor allem in den Anden häu-fig.
Die Klimazone zerfällt in drei ökologisch unter-schiedliche Gebiete, nämlich die winterfeuchten Sub-tropen an der Westseite der Kontinente (Cs-Klimanach Köppen-Trewartha; Leitvegetation Hartlaub-wälder), die immerfeuchten Subtropen an deren Ost-seite (Cf-Klima; Leitvegetation Lorbeerwälder) unddie subtropischen Trockengebiete dazwischen (s. Kap.4). Die Westseiten stehen im Sommer unter dem Ein-fluss des Subtropenhochs, während im Winter dieWestwindtrift mit ihren Regenfällen das Wetter be -stimmt. Das Klima der Ostseiten ist monsunalgeprägt: Im Sommer fließen aus Südosten (Nordhe-misphäre) bzw. Nordosten (Südhemisphäre) feucht-warme Luftmassen landeinwärts, die Niederschlägebringen; im Winter weht ein trockener und kalterNordwest- bzw. Südwestwind. Damit sind die Ge -biete im Landesinnern im Winter vergleichsweiseniederschlagsarm, sodass man hier von einem som-merfeuchten Klima sprechen kann; nur die Küsten -regionen sind ganzjährig humid. Aber auch hier fälltder meiste Regen im Sommer (s. Abb. 1-23). Dem -entsprechend unterscheidet sich auch die Vegeta-tion: Für die Teilzone der winterfeuchten Subtropenmit einem sommertrockenen Klima sind xerophyti-sche Hartlaubwälder und -gebüsche charakteristisch,während in den immerfeuchten Subtropen immer-grüne Laubwälder dominieren. Die physiognomi-schen, ökophysiologischen und floristischen Unter-
5Die warm-gemäßigte (subtropische) Zone
J. S. Pfadenhauer et al., Vegetation der Erde© Springer-Verlag Berlin Heidelberg 2014

schiede zwischen beiden rechtfertigen, dass wir siegetrennt behandeln.
5.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen)
5.2.1 Grundlagen
Wie oben bereits angesprochen, liegen die Gebietemit einem subtropisch-warmen und humiden Klimazwischen dem 30. und 40. Breitengrad auf der Ost-seite der Kontinente (s. Abb. 1-30). Dort gehen sieäquatorwärts kontinuierlich in die immerfeuchtenTropen über (Brasilien, Ostasien, Südafrika), währendsie polwärts an die feuchte nemorale Zone (Nord-amerika, Japan, Korea, Neuseeland) oder an dieHartlaubvegetation der winterfeuchten Subtropen(Afrika, Australien) angrenzen. Im Westen stoßen siemehr oder minder abrupt an die tropisch-subtropi-schen Trockengebiete im Innern der Kontinente.
Die immerfeuchten Subtropen nehmen rund 6 Mio. km2 ein (= 4 % der Festlandsfläche; Schultz2000). Sie umfassen nach Schultz (2000) • in Nordamerika den Südosten der USA mit den
Bundesstaaten South Carolina, Georgia, Alabama,Mississippi, Louisiana und Florida (etwa bis 35° Nund bis 95° W),
• in Südamerika die südlichen Bundesstaaten Brasi-liens (Paraná, Santa Catarina und Rio Grande doSul), ganz Uruguay und den Nordosten von Ar -gentinien einschließlich des größten Teiles derPampa,
• in Ostasien einen großen Teil von China (etwazwischen 24 und 32° N, westlich bis zu den nachSüden streichenden Ausläufern des Himalaya inYunnan und Sichuan, z. B. das Hengduan-Ge bir -ge) sowie Südjapan (bis etwa 37° N) und die Süd-spitze von Südkorea,
• in Ostaustralien die Küstenregion und ihre Rand-stufe (Great Dividing Range) zwischen 23 und 37° S,
• im pazifischen Raum Neukaledonien und dieNordinsel von Neuseeland,
• in Afrika den Südosten von Südafrika (östlichesKapland, Teile von KwaZulu-Natal) und
• in Europa kleine, im Vergleich zu den übrigenVorkommen floristisch verarmte Gebiete der Flo-
renregion Makaronesien (Teneriffa, La Gomeraund Gran Canaria, Madeira und die Azoren).
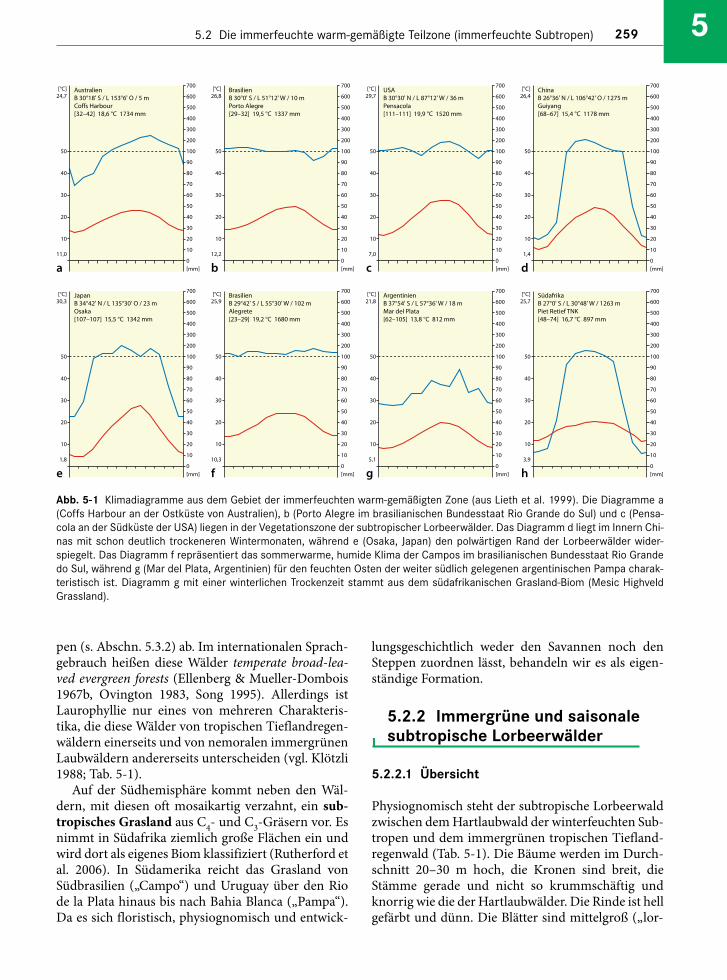
Das Klima ist im Allgemeinen ganzjährig humid,mit einem Sommermaximum und Jahresnieder-schlägen zwischen 800 mm (Abb. 5-1a) und mehr als 1.500 mm (Abb. 5-1b, c, d). Im Übergang zu deninnerkontinentalen Trockengebieten werden die Win-ter nie der schlagsärmer (Abb. 5-1e). Polwärts, gegendie nemoralen Laubwälder zu, verschärfen sich diether mischen Jahreszeiten (Abb. 5-1f). Der Jahres-gang der Temperatur (auf Meeresniveau) übertrifftmit Unterschieden von mindestens 12–15 °C zwi-schen Sommer und Winter den Tagesgang derimmerfeuchten Tropen um rund das Doppelte.Fröste sind auf Meeresniveau in Küstennähe selten,kommen aber in höheren Berglagen oberhalb 700–800 m NN und im Landesinnern bei Kaltlufteinbrü-chen aus den polaren Gebieten regelmäßig vor. Zwarbleibt die Mitteltemperatur des kältesten Monatsimmer über 5 °C; dennoch gibt es für die Vegetationeine Winterruhe, die Teil des jahreszeitlichen Aspekt-wechsels der Vegetation ist.
Die Böden nehmen eine Mittelstellung zwischenden tropischen Ferralsolen und den nemoralen Cam-bisolen ein. Es handelt sich um basenarme, vielfachAl-reiche Acrisole (mit geringer Kationenaustausch-kapazität, KAK) und Alisole (mit hoher KAK; Zechet al. 2014). Sie zeigen eine ausgeprägte Tonverlage-rung (Lessivierung). Der Ah-Horizont ist humus-und tonarm. Der Bt-Horizont ist oft intensiv gelb-braun oder rot gefärbt. Azonal treten auch Vertisoleauf. Im Übergang zu den Trockengebieten gibt esauch Solonetze.
Die zonale Vegetation der immerfeuchten Subtro-pen ist ein immergrüner Laubwald, der wegen desCharakters seiner Blätter (s. Abschn. 1.3.4.1) undeiner gewissen Häufung von Vertretern der Laura-ceae in der deutschsprachigen Literatur als subtropi-scher Lorbeerwald bezeichnet wird (Klötzli 1988).Der Begriff stammt in seiner latinisierten Form Lau-risilva von den Schweizer Botanikern Brockmann-Jerosch & Rübel (1912); als laurophyll werden Bäumebezeichnet, deren Blätter zwischen dem eher weichen(ledrigen) und dem eher harten (sklerophyllen) Typstehen (s. ausführlich in Kasten 1-7). Auch wir ver-wenden für die zonalen Wälder der immerfeuchtenSubtropen den Begriff Lorbeerwald und grenzen ihnbegrifflich als „subtropischen Lorbeerwald“ vom„nemoralen Lorbeerwald“ der feuchten kühl-gemä-ßigten Zone (s. Abschn. 6.2.4.2), dem oreotropischenLorbeerwald der Feuchttropen (s. Abschn. 2.3.2) so -wie vom „Hartlaubwald“ der winterfeuchten Subtro-
258 5 Die warm-gemäßigte (subtropische) Zone5
pen (s. Abschn. 5.3.2) ab. Im internationalen Sprach-gebrauch heißen diese Wälder temperate broad-lea-ved evergreen forests (Ellenberg & Mueller-Dombois1967b, Ovington 1983, Song 1995). Allerdings istLaurophyllie nur eines von mehreren Charakteris-tika, die diese Wälder von tropischen Tieflandregen-wäldern einerseits und von nemoralen immergrünenLaubwäldern andererseits unterscheiden (vgl. Klötzli1988; Tab. 5-1).
Auf der Südhemisphäre kommt neben den Wäl-dern, mit diesen oft mosaikartig verzahnt, ein sub-tropisches Grasland aus C4- und C3-Gräsern vor. Esnimmt in Südafrika ziemlich große Flächen ein undwird dort als eigenes Biom klassifiziert (Rutherford etal. 2006). In Südamerika reicht das Grasland vonSüdbrasilien („Campo“) und Uruguay über den Riode la Plata hinaus bis nach Bahia Blanca („Pampa“).Da es sich floristisch, physiognomisch und entwick-
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 259
lungsgeschichtlich weder den Savannen noch denSteppen zuordnen lässt, behandeln wir es als eigen-ständige Formation.
5.2.2 Immergrüne und saisonale subtropische Lorbeerwälder
5.2.2.1 Übersicht
Physiognomisch steht der subtropische Lorbeerwaldzwischen dem Hartlaubwald der winterfeuchten Sub-tropen und dem immergrünen tropischen Tiefland-regenwald (Tab. 5-1). Die Bäume werden im Durch-schnitt 20–30 m hoch, die Kronen sind breit, dieStämme gerade und nicht so krummschäftig undknorrig wie die der Hartlaubwälder. Die Rinde ist hellgefärbt und dünn. Die Blätter sind mittelgroß („lor-
AustralienB 30°18' S / L 153°6' O / 5 mCoffs Harbour[32–42] 18,6 °C 1734 mm
700
600
500
400
300
200
10050
[°C]24,7
11,0
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
BrasilienB 30°0' S / L 51°12' W / 10 mPorto Alegre[29–32] 19,5 °C 1337 mm
700
600
500
400
300
200
10050
[°C]26,8
12,2
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
USAB 30°30' N / L 87°12' W / 36 mPensacola[111–111] 19,9 °C 1520 mm
700
600
500
400
300
200
10050
[°C]29,7
7,0
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
ChinaB 26°36' N / L 106°42' O / 1275 mGuiyang[68–67] 15,4 °C 1178 mm
700
600
500
400
300
200
10050
[°C]26,4
1,4
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
JapanB 34°42' N / L 135°30' O / 23 mOsaka[107–107] 15,5 °C 1342 mm
700
600
500
400
300
200
10050
[°C]30,3
1,8
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
BrasilienB 29°42' S / L 55°30' W / 102 mAlegrete[23–29] 19,2 °C 1680 mm
700
600
500
400
300
200
10050
[°C]25,9
10,3
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
ArgentinienB 37°54' S / L 57°36' W / 18 mMar del Plata[62–105] 13,8 °C 812 mm
700
600
500
400
300
200
10050
[°C]21,8
5,1
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
a b c d
e f g h
SüdafrikaB 27°0' S / L 30°48' W / 1263 mPiet Retief TNK[48–74] 16,7 °C 897 mm
700
600
500
400
300
200
10050
[°C]25,7
3,9
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
Abb. 5-1 Klimadiagramme aus dem Gebiet der immerfeuchten warm-gemäßigten Zone (aus Lieth et al. 1999). Die Diagramme a(Coffs Harbour an der Ostküste von Australien), b (Porto Alegre im brasilianischen Bundesstaat Rio Grande do Sul) und c (Pensa-cola an der Südküste der USA) liegen in der Vegetationszone der subtropischer Lorbeerwälder. Das Diagramm d liegt im Innern Chi-nas mit schon deutlich trockeneren Wintermonaten, während e (Osaka, Japan) den polwärtigen Rand der Lorbeerwälder wider-spiegelt. Das Diagramm f repräsentiert das sommerwarme, humide Klima der Campos im brasilianischen Bundesstaat Rio Grandedo Sul, während g (Mar del Plata, Argentinien) für den feuchten Osten der weiter südlich gelegenen argentinischen Pampa charak-teristisch ist. Diagramm g mit einer winterlichen Trockenzeit stammt aus dem südafrikanischen Grasland-Biom (Mesic HighveldGrassland).
beerartig“), mehr oder minder eiförmig, ganzrandigoder schwach gezähnt und haben häufig eine glän-zende Oberseite (Abb. 5-2). Die Strauchschicht ist inder Regel gut entwickelt und ebenfalls meist immer-grün. Wie bei den tropischen Tieflandregenwäldernfehlt auch hier eine Feldschicht weitgehend, sofern essich um ein Reifestadium handelt. Denn die Baum-kronen lassen kaum Licht auf den Boden. LediglichFarne sind gut vertreten. Artenzahl und Abundanzepiphytischer Kryptogamen (Moose, Flechten) sindgrößer als in tropischen Wäldern. Bäume mit Brett-wurzeln sind seltener; auch Palmen treten deutlichzurück. Baumwürger der Gattung Ficus kommendagegen noch recht häufig vor. In besonders regen-oder nebelreichen Lagen wie auf der Nordinsel vonNeuseeland, in Südostaustralien und an den Hängender Serra Geral in Südbrasilien treten Baumfarne auf.Dort ist auch der Bewuchs mit Kormo-Epiphytenbesonders reichlich.
Der Anteil laubabwerfender Bäume steigt imÜbergang zu den sommergrünen nemoralen Laub-wäldern (Nordamerika, China, Japan) und zu deninnerkontinentalen Trockengebieten (Brasilien). DieWälder erhalten damit einen saisonalen Charakter.
Besonders in den höheren Lagen (untere und mitt-lere montane Stufe der Gebirge) treten vermehrtaltertümliche Koniferen der Familien Podocarpa-ceae, Cupressaceae, Araucariaceae und Taxaceae auf.Sie bilden als Emergenten gelegentlich Dominanz-bestände („subtropische Reliktkoniferenwälder“, s.Kasten 5-2 zu den nemoralen Reliktkoniferenwäl-dern s. Abschn. 6.2.3); die Laurophyllen sind aberimmer in der zweiten Baumschicht reichlich vertre-ten. Ihre Assimilationsorgane erinnern in einigenFällen eher an Laubblätter als an die Nadeln der Pina-ceae. Neben Ginkgo biloba gehören hierzu einige Ver-treter der Podocarpaceae mit blattartigen Phyllokla-dien, die aus Kurztrieben hervorgegangen sind (wiedie Gattung Phyllocladus) und die Araucariaceen-Gattung Agathis (Abb. 5-2, 5-8).
Die Artenzahl der Bäume und Sträucher der sub-tropischen Lorbeerwälder ist geringer (Einwirkungvon Frost) als in den tropischen Tieflandregenwäl-dern, aber deutlich höher als in den Hartlaubwäldern(Einwirkung von Frost und Trockenheit). Sie liegt jenach Region zwischen zehn und über 60 Arten proHektar. Am artenreichsten sind die Wälder derimmerfeuchten Subtropen in China (Wang 1961).
260 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 5-1 Unterschiede zwischen den immergrünen (laurophyllen) subtropischen Laubwäldern und den tropischen Tieflandregen-wäldern sowie den subtropischen Hartlaubwäldern.
Merkmale immergrüne immergrüne immergrüne tropische subtropische subtropischeTieflandregenwälder Lorbeerwälder1 Hartlaubwälder
Wuchshöhe der Bäume 30–50 m 20–30 m 10–20 m 2
(Emergenten bis 60 m) (selten bis 40 m)
Kronenform der Bäume häufig schirmförmig rundkronig ausladend-breitkronig
Stamm schlank, hochwüchsig, schlank, eher gedrungen, gedrungen, oft krumm-Brettwurzeln häufig Brettwurzeln selten schäftig, Borke, Brettwurzeln
fehlen
Blätter mittel bis groß (noto- bis mittel (bis klein) mittel bis klein (mikro- makrophyll, überwiegend ledrig, (überwiegend notophyll), bis notophyll), häufig hartTräufelspitzen häufig glänzend, Träufelspitzen und leicht brechend,
meist fehlend keine Träufelspitzen
Baumartenzahl (ha–1) 100–300 10–60 5–10
Gefäß-Epiphyten mittel bis häufig gering bis mittel sehr gering bis fehlend(in Nebellagen) häufig in Nebellagen)
Reliktkoniferen fehlend vorhanden bis häufig fehlend
Vegetationszeit (Monate)3, 12, keine jahreszeitliche 12, jahreszeitliche 6–9, jahreszeitlichePhänologie Rhythmik Rhythmik winterbedingt Rhythmik kälte- und
trockenheitsbedingt
1 Ohne die nemoralen Lorbeerwälder; 2 außer Australien (bis über 60 m); 3 Optimalausbildung ohne trockene Monate.

Die charakteristischen Pflanzenfamilien der subtro-pischen Lorbeerwälder sind neben den pantropi-schen Lauraceae (Beilschmidia, Cinnamomum, Cryp-tocarya, Laurus, Lindera, Litsea, Ocotea, Persea) aufder Nordhemisphäre die Fagaceae mit den Gattun-gen Castanopsis, Lithocarpus und Quercus, die Thea-ceae mit Camellia und Schima, die Magnoliaceae(Magnolia, Michelia) sowie die Hamamelidaceae(Distylium), auf der Südhemisphäre die Cunonia-ceae mit den Gattungen Caldcluvia, Cunonia, Eucry-phia und Weinmannia, die Monimiaceae (Laurelia,Laureliopsis) und die Proteaceae (Faurea, Knightia;Tab. 5-2). Aus vorwiegend tropisch verbreitetenFamilien stammen Vertreter der Moraceae (mit derGattung Ficus), der Myrsinaceae (Myrsine), der Myr-taceae (in den Lorbeerwäldern vor allem auf der Süd-hemisphäre, z. B. Eucalyptus in Australien) und derSymplocaceae. Die einzige Gattung der Aquifolia-ceae, Ilex, ist mit etwa 400 Arten zwar weltweit ver-breitet (Heywood et al. 2007), hat aber einen merk-lichen Schwerpunkt in den Lorbeerwäldern vonSüdamerika und Südostasien.
Lorbeerwälder gelten als Relikte der spättertiärenimmergrünen Laubwälder, die in Räumen mit einem
kontinuierlich gemäßigt warmen und feuchtenKlima die pleistozänen Kaltzeiten überdauern konn-ten. Diese über Millionen von Jahren andauerndeKontinuität macht sich in den vielen altertümlichenTaxa bemerkbar, die in diesen Wäldern leben. So sindzahlreiche ursprüngliche Blütenpflanzen (wie dieChloranthaceae, Lauraceae und Magnoliaceae) Be -standteile des Lorbeerwaldes, ebenso wie die vermut-liche Ausgangssippe der Angiospermen, Amborellatrichopoda, die in den laurophyllen BergwäldernNeukaledoniens vorkommt (s. Abschn. 1.2.1). Dergroße Anteil altertümlicher Gymnospermen (s. oben)ist ebenfalls ein Hinweis auf das Alter dieser Wälder.Offensichtlich waren die Bedingungen für ihr Über-leben hier besonders günstig: Das kühlere, subtropi-sche Klima begrenzte einerseits die Konkurrenz deraggressiven jüngeren Angiospermensippen der feuch-ten Tropen und war andererseits frei vom Kältestressder nemoralen Zone (Schroeder 1998).
Im Folgenden werden wir die zonalen subtropi-schen Lorbeerwälder in den einzelnen Kontinentenseparat beschreiben, weil sie sich floristisch undstrukturell so deutlich voneinander unterscheiden,dass sie einen eigenständigen Charakter haben. Wir
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 261
b c
d e
a
f
Abb. 5-2 Assimilationsorgane von Bäumen der subtropischen Lorbeerwälder. a = Laurusnovocanariensis, Lauraceae (Teneriffa); b = Cunonia capensis, Cunoniaceae (Südafrika); c =Castanopsis cuspidata, Fagaceae (Japan); d = Pittosporum undulatum, Pittosporaceae (Brasi-lien); e = Phyllokladien von Phyllocladus trichomanoides (links) und P. glauca (rechts), Podo-carpaceae (Neuseeland); f = Afrocarpus falcatus, Podocarpaceae (Südafrika).
beginnen mit den südostasiatischen LorbeerwäldernChinas und Japans, die von allen am artenreichstensind und den Typus des subtropischen Lorbeerwal-des am besten repräsentieren, gefolgt von Makarone-sien, Nordamerika, Südamerika, Afrika, Australienund Neuseeland. Immergrüne laurophylle Wäldergibt es aber auch außerhalb ihres zonalen Vorkom-mens in den immerfeuchten Subtropen, nämlichextrazonal in feuchten Tälern der sonst von Hart-laubwäldern eingenommenen winterfeuchten Sub-tropen und als oreotropische immergrüne Laubwäl-der in der montanen Stufe feuchttropischer Gebirge(Klötzli 1988). Die zuletzt genannten Wälder haben
wir schon in Abschn. 2.3.2 besprochen. Schließlichkommen Lorbeerwälder auch in der feuchten nemo-ralen Zone vor, allerdings nur unter hochozeani-schen Bedingungen wie in Südchile (nordpatagoni-scher und valdivianischer Lorbeerwald). DieseWälder werden in Abschn. 6.2.4 behandelt.
5.2.2.2 Südostasien
Weltweit die größte Fläche, die potenziell von subtro-pischen Lorbeerwäldern eingenommen wird, liegtzwischen 24 und 32° N sowie 99 und 123° O in China
262 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 5-2 Charakteristische Baumfamilien der Lorbeerwaldgebiete (nach Schroeder 1998, modifiziert). Angegeben ist die Zahl derGattungen mit immergrünen Gehölzen. Im gleichen Gebiet vorkommende Sommergrüne sind nicht berücksichtigt. Nord- und süd-hemisphärische Lorbeerwälder unterscheiden sich deutlich; lediglich Mexiko nimmt eine Zwischenstellung ein.
Familie China, Nord- Makaro- Mexiko Austra- Neu- Süd Süd-Japan amerika nesien lien seeland brasilien afrika
nord- und südhemispärisch
Lauraceae 9* 1* 4* 6* 6* 2* 4* 2*
Oleaceae 1 1 2* 1 1 1 1 3*
Cupressaceae 4 1 1 2 2 1 1
Myrsinaceae 4 3 3 2 1 1 2*
Aquifoliaceae 1 1 1* 1 1* 1*
Celastraceae 2 1 3 1 4*
Araliaceae 4 1 2 3 2 2 2
Arecaceae 2 2 1 3 1 2
vorwiegend nordhemisphärisch
Fagaceae 3* 1* 1*
Theaceae 9* 1* 1 2 1
Pinaceae 2 1* 1 2*
Magnoliaceae 4* 1* 1*
Taxaceae 3 2 1 1
vorwiegend südhemisphärisch
Cunoniaceae 1 10* 2* 2* 2*
Proteaceae 1 1 8* 1* 2 2
Araucariaceae 2 1* 1*
Myrtaceae 2 1 2 9* 3* 8* 2
Monimiaceae 2 5* 2 1 1
Podocarpaceae 2 1 5 3 1* 1
Winteraceae 1 1 2 1
* Maßgeblich an der oberen Baumschicht beteiligt, zumindest in bestimmten Ausbildungen.

(Wang 1961, Song 1988). Rund ein Viertel der Flächedes Landes, nämlich etwa 2 Mio. km2, wäre von die-sem Waldtyp bedeckt. Heute sind nur noch wenigeReste vorwiegend auf heiligen Bergen und in Schutz-gebieten erhalten geblieben. Die Wälder sind floris-tisch sehr reichhaltig; sie enthalten rund zwei Drittelder 4.000 in China vorkommenden Gattungen unddie Hälfte (15.000) aller chinesischen Gefäßpflanzen-arten.
Diese Zahlen dürfen nicht darüber hinwegtäu-schen, dass die dominanten Baumarten nur einigenwenigen Gattungen angehören, nämlich Beilschmie-dia (39 Arten), Camellia (97 Arten, davon 76 ende-misch), Castanopsis (58 Arten), Lithocarpus (123 Ar -ten in China, davon 69 endemisch), Michelia (37Arten) und Quercus (35 Arten in China; Artenzahlennach eFloras 2008). Die großen Artenzahlen dieserGattungen sind auf eine beachtliche allopatrischeDifferenzierung in dem durch Gebirge stark geglie-derten Gebiet zurückzuführen; auf Teilflächen, wiesie für Vegetationsaufnahmen verwendet werden,kommen aber oft nur wenige Arten vor. So liegt dieBaumartenzahl in Plots von 20 × 20 m Seitenlängezwischen 15 und 25; die Gesamtzahl der Gefäßpflan-zenarten variiert zwischen 45 (im Norden) und 100pro 400 m2 (im Süden).
Die Wälder werden in eine große Zahl physiogno-misch und floristisch verschiedener Vegetations -typen gegliedert (Wang 1961). Vereinfacht kann mandie eher im Westen verbreiteten Castanopsis dela-vayi- und Quercus delavayi-Wälder von den weiterim Osten vorkommenden Waldtypen unterschieden,in denen verschiedene Castanopsis- und Quercus-Arten zur Dominanz gelangen (wie z. B. C. eyrei, Q.glauca, Machilus (Persea) thunbergii). Die Lorbeerwäl -der setzten sich über Südwestchina bis nach Nepal ineiner Höhenlage zwischen 1.000 und 2.000 m NNbandartig als oreotropischer (laurophyller) Laubwaldfort; im Bereich Nordindien-Assam ist dieser Wald-typ die Heimat der Teepflanze (Camellia sinensis).
Im Reifestadium sind die Wälder überwiegendimmergrün (Abb. 5-3a). Sommergrüne Bäume wieverschiedene Quercus- (und Fagus-)Arten treten inden höheren Gebirgslagen mit nemoralem Klimaoder im Übergang zu den sommergrünen Laubwäl-dern auf (wie in dem ca. 81 km2 umfassenden Schutz-gebiet Gutianshan National Nature Reserve zwischenGangzhou und Nanchang). Sommergrüne (wie dieAlnus-Arten) und immergrüne Pioniere (hier vorallem Pinus spp. wie P. yunnanensis im SüdwestenChinas und P. massoniana im Südosten) stellen sichnach Aufgabe landwirtschaftlicher Nutzflächen raschein und kommen dann vorübergehend zur Domi-
nanz. Im Lauf der Sukzession werden sie von Lauro-phyllen abgelöst. Viele dieser Laurophylle wie dieCastanopsis-Arten sind resprouter; ihr Stockaus-schlagsvermögen ist beachtlich und beschleunigt dieRegeneration der Wälder nach intensiver Holznut-zung (Wang et al. 2007). Vermutlich ist diese Fähig-keit ein Vorteil in Gebieten, die regelmäßig von tro-pischen Wirbelstürmen heimgesucht werden (wiebesonders in Japan; Fuiji et al. 2009; Kasten 5-1). DerWald regeneriert sich schneller durch Stockausschlagals durch Wiederbesiedlung einer freigeräumten Flä-che mittels Samen. Die heute vielerorts dominieren-den Lorbeerwälder zeigen deshalb einen niederwald-artigen Charakter.
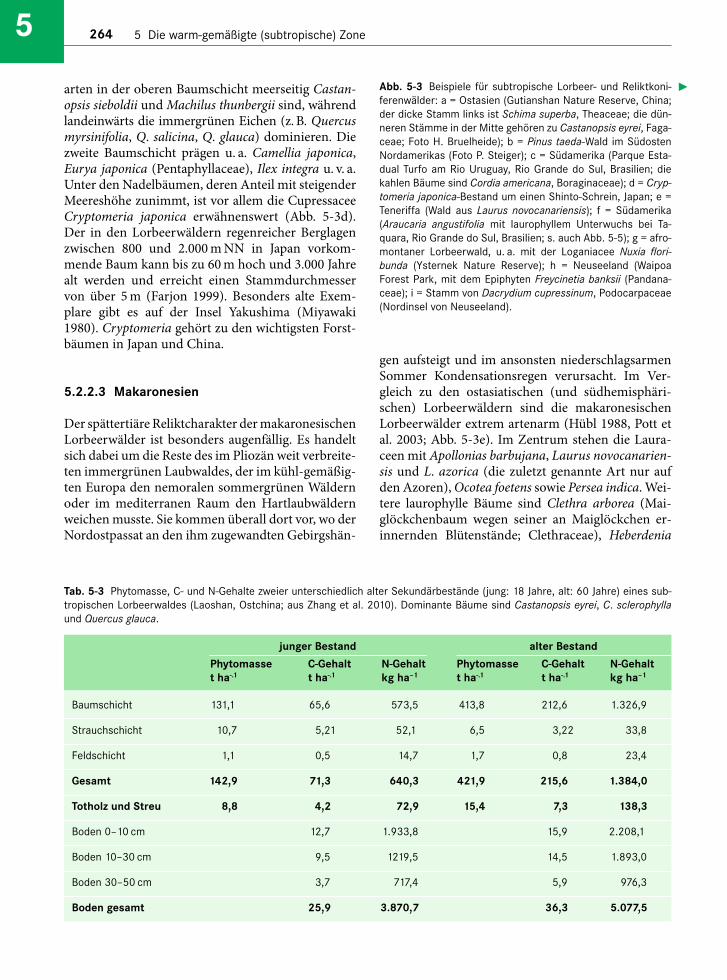
Die Phytomasse ist bemerkenswert hoch (Tab. 5-3). Sie erreicht in 60 Jahre alten Waldbeständen beieiner Baumhöhe von 13 m immerhin 420 t ha–1 (ober-und unterirdisch), von denen nur etwa 2 % auf dieStrauch- und Feldschicht entfallen (Zhang et al.2010). Damit unterscheiden sich diese Wälder nichtwesentlich von tropischen Tieflandregenwäldern (s.Abschn. 2.1.6). Der C- bzw. N-Gehalt im Boden be -trägt 14 bzw. 77 % des gesamten Pools, sodass bezüg-lich des Stickstoffs eher der Boden, bezüglich desKohlenstoffs eher die (lebende) Phytomasse als Spei-chermedium dient. Während der Entwicklung vomJung- zum Altbestand akkumulieren subtropischeLorbeerwälder in China in relativ kurzer Zeit be -trächtliche Mengen an C und N (Tab. 5-3); so hat sichder Phytomassespeicher innerhalb der kurzen Zeit-spanne von 42 Jahren verdreifacht (im Fall von C)bzw. mehr als verdoppelt (im Fall von N).
Nicht ganz so artenreich wie die chinesischen sinddie subtropischen Lorbeerwälder in Japan. So stehenden 159 Gattungen in der Provinz Yunnan (Südwest-china) 79 Gattungen in Südwestjapan gegenüber;44 Gattungen wie Castanopsis, Camellia, Quercus(Untergattung Cyclobalanopsis) kommen in beidenRegionen vor (Tang & Ohsawa 2009). Ähnlich wie inChina sind die Wälder auch hier weitgehend ver-schwunden und nur um Tempelanlagen, in schwerzugänglichen Gebirgsregionen oder in für Ackerbauzu steilem Gelände erhalten geblieben. Die Artenzahlder Bäume (> 4,5 cm Brusthöhendurchmesser, BHD)variiert nach einer Zusammenstellung von Itô (1997)zwischen durchschnittlich 13 (Wälder auf Kyushuund den Ryukyu-Inseln) und 24 (Okinawa) auf je -weils 400 m2 großen Probeflächen. Die japanischenLorbeerwälder, deren Hauptverbreitung auf Süd-Honshu, Shikoku und Kyushu liegt (s. Abb. 5-7), sindpflanzensoziologisch detailliert erfasst und klassifi-ziert (Miyawaki 1979): Sie werden in der KlasseCamellietea japonicae zusammengefasst, deren Kenn-
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 263
arten in der oberen Baumschicht meerseitig Castan-opsis sieboldii und Machilus thunbergii sind, währendlandeinwärts die immergrünen Eichen (z. B. Quercusmyrsinifolia, Q. salicina, Q. glauca) dominieren. Diezweite Baumschicht prägen u. a. Camellia ja ponica,Eurya japonica (Pentaphyllaceae), Ilex integra u. v. a.Unter den Nadelbäumen, deren Anteil mit steigenderMeereshöhe zunimmt, ist vor allem die CupressaceeCryptomeria japonica erwähnenswert (Abb. 5-3d).Der in den Lorbeerwäldern regenreicher Berglagenzwischen 800 und 2.000 m NN in Japan vorkom-mende Baum kann bis zu 60 m hoch und 3.000 Jahrealt werden und erreicht einen Stammdurchmesservon über 5 m (Farjon 1999). Besonders alte Exem-plare gibt es auf der Insel Yakushima (Mi yawaki1980). Cryptomeria gehört zu den wichtigsten Forst-bäumen in Japan und China.
5.2.2.3 Makaronesien
Der spättertiäre Reliktcharakter der makaronesischenLorbeerwälder ist besonders augenfällig. Es handeltsich dabei um die Reste des im Pliozän weit verbreite-ten immergrünen Laubwaldes, der im kühl-gemäßig-ten Europa den nemoralen sommergrünen Wäldernoder im mediterranen Raum den Hartlaubwäldernweichen musste. Sie kommen überall dort vor, wo derNordostpassat an den ihm zugewandten Gebirgshän-
gen aufsteigt und im ansonsten niederschlagsarmenSommer Kondensationsregen verursacht. Im Ver-gleich zu den ostasiatischen (und südhemisphäri-schen) Lorbeerwäldern sind die makaronesischenLorbeerwälder extrem artenarm (Hübl 1988, Pott etal. 2003; Abb. 5-3e). Im Zentrum stehen die Laura-ceen mit Apollonias barbujana, Laurus novocanarien-sis und L. azorica (die zuletzt genannte Art nur aufden Azoren), Ocotea foetens sowie Persea indica. Wei-tere laurophylle Bäume sind Clethra arborea (Mai-glöckchenbaum wegen seiner an Maiglöckchen er -innernden Blütenstände; Clethraceae), Heberdenia
264 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 5-3 Phytomasse, C- und N-Gehalte zweier unterschiedlich alter Sekundärbestände (jung: 18 Jahre, alt: 60 Jahre) eines sub-tropischen Lorbeerwaldes (Laoshan, Ostchina; aus Zhang et al. 2010). Dominante Bäume sind Castanopsis eyrei, C. sclerophyllaund Quercus glauca.
junger Bestand alter Bestand
Phytomasse C-Gehalt N-Gehalt Phytomasse C-Gehalt N-Gehaltt ha-.1 t ha-.1 kg ha–1 t ha-.1 t ha-.1 kg ha–1
Baumschicht 131,1 65,6 573,5 413,8 212,6 1.326,9
Strauchschicht 10,7 5,21 52,1 6,5 3,22 33,8
Feldschicht 1,1 0,5 14,7 1,7 0,8 23,4
Gesamt 142,9 71,3 640,3 421,9 215,6 1.384,0
Totholz und Streu 8,8 4,2 72,9 15,4 7,3 138,3
Boden 0–10 cm 12,7 1.933,8 15,9 2.208,1
Boden 10–30 cm 9,5 1219,5 14,5 1.893,0
Boden 30–50 cm 3,7 717,4 5,9 976,3
Boden gesamt 25,9 3.870,7 36,3 5.077,5
Abb. 5-3 Beispiele für subtropische Lorbeer- und Reliktkoni -ferenwälder: a = Ostasien (Gutianshan Nature Reserve, China;der dicke Stamm links ist Schima superba, Theaceae; die dün-neren Stämme in der Mitte gehören zu Castanopsis eyrei, Faga-ceae; Foto H. Bruelheide); b = Pinus taeda-Wald im SüdostenNordamerikas (Foto P. Steiger); c = Südamerika (Parque Esta-dual Turfo am Rio Uruguay, Rio Grande do Sul, Brasilien; diekahlen Bäume sind Cordia americana, Boraginaceae); d = Cryp-tomeria japonica-Bestand um einen Shinto-Schrein, Japan; e =Teneriffa (Wald aus Laurus novocanariensis); f = Südamerika(Araucaria angustifolia mit laurophyllem Unterwuchs bei Ta -quara, Rio Grande do Sul, Brasilien; s. auch Abb. 5-5); g = afro-montaner Lorbeerwald, u. a. mit der Loganiacee Nuxia flori-bunda (Ysternek Nature Reserve); h = Neuseeland (WaipoaForest Park, mit dem Epiphyten Freycinetia banksii (Pandana-ceae); i = Stamm von Dacrydium cupressinum, Podocarpaceae(Nordinsel von Neuseeland).
�

55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 265
g
f
c
d e
h i
a b

bahamensis (Primulaceae), Picconia excelsa (Olea-ceae) undVisnea mocanera (Theaceae). Die Aquifoli-aceae sind mit den Ilex-Arten I. canariensis und I.perado, die vorwiegend subtropisch verbreitetenMyricaceen mit Myrica faya vertreten; Myrica bildet
266 5 Die warm-gemäßigte (subtropische) Zone5
zusammen mit Erica arborea und E. scoparia einGebüsch oder einen niedrigen Wald als Ersatzvegeta-tion der Lorbeerwälder nach deren Beseitigung oderin windausgesetzten Berglagen (wie auf der Hochflä-che im Westen von Madeira).
Taifun und WalddynamikBestandsdynamik und Nährstoffumsatz werden in den Lor-beerwäldern des ostasiatischen Raumes, vor allem in denküstennahen Gebieten Chinas, Japans und auf Taiwan, vontropischen Wirbelstürmen (Zyklonen) geprägt, die im Nord-westpazifik als Taifun bezeichnet werden. Im Nordatlantikheißen sie hurricane bzw. huracán (span.), ein Wort, von demsich die deutsche Bezeichnung Orkan ableitet (Häckel2005). Sie treten mit hoher Frequenz auf (zwischen 0,5- und1,1-mal pro Jahr), und zwar meist in den Monaten Juli, Augustund September. In der Saffir-Simpson-Skala (NWS 2011)werden nach der Windgeschwindigkeit fünf Stufen unter-schieden (Stufe 1: 118–154 km h–1, Stufe 5: > 249 km h–1).Von den in Nordosttaiwan zwischen 1951 und 2005 aufge-tretenen 41 Taifunen gehörten 19 zu den Stufen 4 und 5, 22zu 1 bis 3 (Lin et al. 2012).
Die starken Taifune (Stufe 4 und 5) sind zwar seltener alsdiejenigen der Stufe 1 bis 3, können aber eine verheerendeWirkung entfalten. So verwüstete der Taifun Larry die Regen-wälder in Nordostaustralien in einem Umkreis von 30 km umdas „Auge“ des Wirbelsturmes und knickte bzw. entwurzelteüber 90 % aller Bäume (Turton 2008). Wirbelstürme habendort, wo sie regelmäßig auftreten, einen erheblichen Einflussauf die Baumartenzusammensetzung: Sie verändern den fürOrganismen verfügbaren Raum, sie erhöhen die landschaft-liche Heterogenität und die Variabilität von Stoffkreisläufen,sie setzen Sukzessionsprozesse in Gang und sie formen dieStruktur der Wälder; sie bringen Organismen in Bewegungund sie initiieren evolutionäre Veränderungen durch natürli-che Selektion (Lugo 2008). Dieser Einfluss ist nicht nur derkinetischen Energie der Windbewegung geschuldet, die intropischen Wirbelstürmen 3.500- bis 15.000-mal höher istals im globalen Mittel (ebd.), sondern auch den außeror-dentlich hohen Niederschlägen. Sie führen in der Folge einesWirbelsturmes zu Erosion von Bodenmaterial in den durchWindwurf geöffneten Wäldern und zum Austrag von Nähr-stoffen. Solche Nährstoffverluste können in den feuchtenTropen und Subtropen für N und P das 1.300- bis 2.900-Fache des Austrags aus einem ungestörten, bewaldeten Ein-zugsgebiet betragen (Lugo 2008).
Die Wirkung von Wirbelstürmen auf tropische und sub-tropische Wälder hängt nicht nur von der Stärke der Wind-bewegung und Art der Windbewegung ab, sondern auch vonder Resistenz der Bäume gegenüber Druck und Zug (s.Hubrig 2004). Durchwurzelungstiefe, Holzeigenschaften,
Baumhöhe, Kronendichte und viele weitere Eigenschaftenentscheiden darüber, ob ein Baum geknickt oder entwurzeltwird oder lediglich Druckschäden erleidet (permanent schiefstehende oder umgebogene Bäume). In der Realität einestropischen Wirbelsturmes zeigt sich jedoch, dass einzelneBäume und einzelne Baumarten abhängig vom jeweiligenEreignis in unterschiedlichem Ausmaß betroffen sind, so -dass sich eine Vorhersage der zu erwartenden Schädenkaum treffen lässt (Duryea et al. 2007). In Gebieten mithochfrequenten Taifunen wie im westpazifischen Raum sinddie Wälder niedriger, die Kronen ihrer Bäume kleiner und derAnteil an Pionierbäumen und lichtbedürftigen Lianen ist grö-ßer als dort, wo Wirbelstürme selten sind oder fehlen (Webb1958). In solchen Wäldern ist zudem der Anteil von Bäumenmit Stockausschlag (wie bei den Castanopsis-Arten; s.Abschn. 5.2.2.2) und weiteren stressbedingten Merkmalen(vorgezogene Blühperiode, kurze Lebensspanne) größer(Lugo & Zimmermann 2002).
Bezogen auf diese Kennzeichen ist die Vegetation desostasiatischen Raumes offensichtlich besser an hochfre-quente Taifune angepasst als die Pflanzendecke in Gebieten,in denen tropische Wirbelstürme eher selten auftreten.Selbst heftige Ereignisse der Stufen 3 und 4 schaffen in denan Wirbelstürme „gewöhnten“ subtropischen Lorbeerwäl-dern von Nordosttaiwan nur Lücken mit einem Durchmesservon weniger als 100 m (Lin et al. 2011). Stattdessen reißensie oft nur die Blätter von den Bäumen, sodass eine großeMenge an grüner Streu auf die Bodenoberfläche gelangt. Dadie frischen Blätter höhere Nährstoffkonzentrationen auf-weisen als die im normalen Alterungsprozess des Laubesentstehende Blattstreu, wirken sich entblätternde Taifunebeschleunigend auf den Nährstoffumsatz aus. Denn imfeuchtwarmen Klima tropisch-subtropischer Wälder wird dieproteinreiche Streu rasch abgebaut; die frei werdendenNährstoffe stehen den Bäumen also innerhalb von wenigenMonaten nach dem Taifun zur Erneuerung ihrer Blattmassezur Verfügung (Xu et al. 2004, Ostertag et al. 2005). Gleich-zeitig ist die Entblätterung der Bäume auch ein Schutz gegenWindwurf, weil die kahlen Kronen dem Sturm weniger Wider-stand entgegensetzen. Die Nettoprimärproduktion solcherhäufig gestörter Waldbestände wird dagegen eher gebremst,weil ein Teil des Nährstoffkapitals für die ständige Erneue-rung der Blattmasse verbraucht wird.
Kasten 5-1

5.2.2.4 Nord- und Mittelamerika
Im Südosten von Nordamerika sind die sonst so cha-rakteristischen immergrünen laurophyllen Laubwäl-der nur kleinflächig ausgebildet (Knapp 1965, Fuji-wara & Box 1994, Christensen 2000). Es handelt sichentweder um lichte, immergrüne Bestände von Quer-cus virginiana und Q. hemispherica mit einigen wei-teren laurophyllen Arten (wie Morella cerifera, Perseaborbonia) und der Palme Sabal palmetto auf Sandbö-den (mit Schwerpunkt in Florida; Greller 2004) oderum Mischwälder aus immergrünen (wie Magnoliagrandiflora) und sommergrünen Bäumen (wie Fagusgrandifolia) auf frischen, basenreichen Standorten,die zu den nemoralen Wäldern überleiten (s. Abschn.6.2.2.6.4). Beigemischt ist Sabal palmetto. Die Zahllaurophyller Gehölze ist gegenüber den südostasiati-schen Lorbeerwäldern erheblich eingeschränkt: DieLauraceen sind lediglich durch zwei Persea-Artenvertreten; von den Fagaceen kommen nur acht Quer-cus-, aber weder Castanopsis- noch Lithocarpus-Arten vor; die für Ostasien so typischen GattungenSchima und Camellia (Theaceae) fehlen. Lediglichdie Gattung Ilex bildet mit sechs Arten ein wichtigesElement der südostamerikanischen Lorbeerwälder.
Meist dominieren auf den großflächig verbreitetenSandböden dieser erdgeschichtlich jungen Küsten-ebene jedoch feuerstabilisierte Kiefernwälder ausPinus palustris, P. taeda (Abb. 5-3b), P. elliottii oder P. caribaea mit sommer- und immergrünem Unter-wuchs (verschiedene Quercus-Arten), denen erhebli-che wirtschaftliche Bedeutung für die Papierindus-trie zukommt (s. Abschn. 5.2.4). Nach Fujiwara &Box (1994) dürfte es sich bei den meisten dieserNadelwälder um eine anthropogene Ersatzvegetationder potenziell natürlichen laurophyllen Laubwäldermit ihrer Dominanz aus immergrünen Eichen (mitBrettwurzeln) handeln. Die dazwischen liegendengrundwasserbeeinflussten Gebiete werden von som-mergrünen Sumpfzypressenwäldern oder immergrü-nen Moorwäldern eingenommen (Pocosins; s.Abschn. 5.4.1). Auf der Halbinsel von Florida be -schränkt sich das potenzielle Lorbeerwaldgebiet aufdie karibische Küstenebene; ansonsten kommt einsklerophyller Buschwald vor, dessen Hartlaubigkeitauf die nährstoffarmen Sandböden zurückzuführensein dürfte (Haeupler 1994).
In Mexiko gedeiht der Lorbeerwald in der SierraMadre Oriental in 800–2.000 m NN unter der Be -zeichnung „Bosque mesóphilo de montaña“ (Rze-dowski 2006) und ist auf Standorte mit hoher Luft-feuchte (Nebellagen, Schluchten) beschränkt. Erer reicht 15–25 m Höhe und besteht aus immergrü-
nen Vertretern der Fagaceae, Clethraceae, Fabaceae s. l., Lauraceae, Melastomataceae und Rubiaceae.Regelmäßig kommen der sommergrüne Baum Liqui-dambar styraciflua (Hamamelidaceae) und verschie-dene immergrüne Quercus-Arten vor (v. a. Q. xa la-pensis). Hinzu treten Podocarpus-Arten wie P.matudae. Die Wälder sind reich an Farnarten undMoosen sowie an Epiphyten, unter denen vor allemdie Orchidaceae in der Artenzahl dominieren.
5.2.2.5 Südamerika
Hueck (1966) und Hueck & Seibert (1981) beschrei-ben „wechselgrüne, mesophytische, subtropischeWälder Ost- und Südbrasiliens, z. T. mit starkemAnteil immergrüner Arten“, die sich ab etwa 15° Svom Rio Doce aus nach Süden bzw. Südwesten ent-lang des Rio Paraná erstrecken, am Wendekreis biszum Rio Paraguay bei Asunción und im Süden biszum Rio Jacuí bei Porto Alegre reichen (Abb. 5-4).Nach der Klimaklassifikation von Troll & Paffen(1964) bzw. Köppen (1931) ist allerdings nur derSüden (etwa südlich einer Linie vom Wendekreis ander Atlantikküste bei São Paulo über die Iguaçu-Was-serfälle bis etwa zur Einmündung des Rio Paraguayin den Rio Paraná bei Corrientes) subtropisch (sub-tropisches sommerfeuchtes bzw. Cfa-Klima). DieWälder nördlich dieser Linie sind tropisch, wobei wires mit einem Ökoton zu tun haben, das die Festle-gung einer Grenze zwischen tropischen und subtro-pischen Wäldern erschwert.
So nehmen von Norden nach Süden die für Lor-beerwälder charakteristischen Taxa Nectandra undOcotea (Lauraceae), Myrsine (Myrsinaceae) und Ilex(Aquifoliaceae) zu (Oliveira-Filho & Fontes 2000);die meisten Bäume kommen jedoch aus eher tropi-schen Familien wie Annonaceae, Melastomataceae,Moraceae, Myrtaceae (z. B. mit der Gattung Euge-nia) u. a. In den Wäldern südlich der o. g. Linie (inden brasilianischen Bundesstaaten Paraná, SantaCatarina und Rio Grande do Sul sowie in der Pro-vinz Missiones in Argentinien) sind Lauraceen mitNectandra megapotamica, Ocotea und Persea sowieFabaceae (wie Dalbergia) und Caesalpiniaceen (Apu-leia) besonders häufig (wie im Waldreservat Turvoam Rio Uruguay; Ruschel et al. 2005; Abb. 5-3c).Dalbergia- und Apuleia-Arten treten als Emergentenauf, die bis zu 30 m hoch werden können und etwadie Hälfte der Baumarten stellen; sie sind laubab-werfend, obwohl in dieser Höhenlage (200–300 mNN) weder Fröste noch Trockenzeiten vorkommen.Möglicherweise ist dieser saisonale Charakter auf
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 267
268 5 Die warm-gemäßigte (subtropische) Zone5
30°
b
a
c
MVBA
BB
PA
Cu
d
e
123456789a9b9c9d9e
R. Uruguay
R. Parana
R. P
aran
a
R. P
arag
uay
R. Iguacu
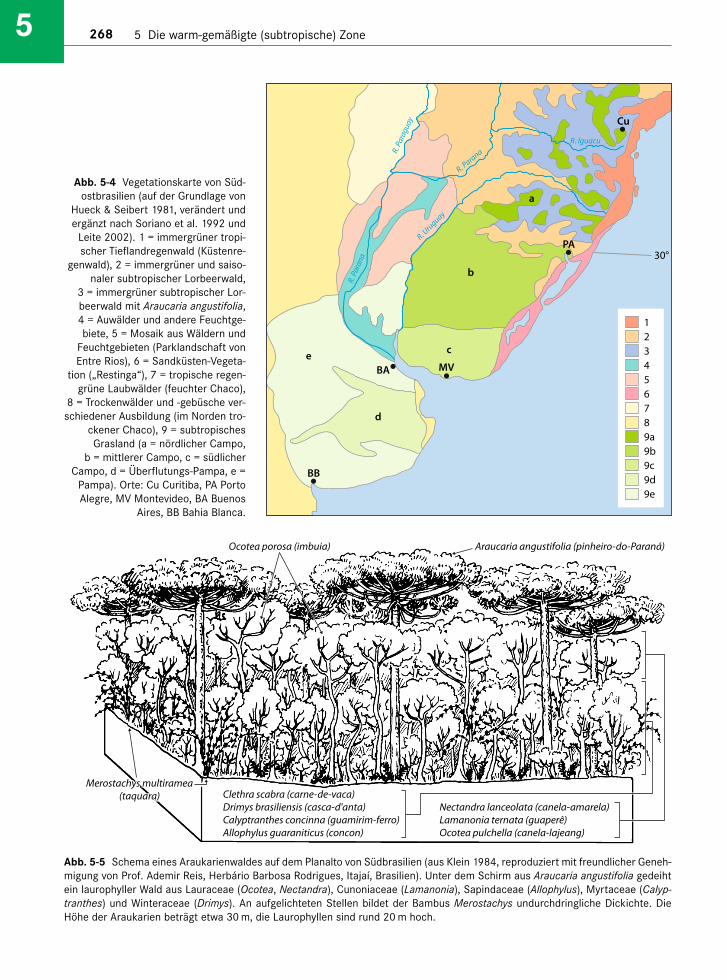
Abb. 5-4 Vegetationskarte von Süd-ostbrasilien (auf der Grundlage von
Hueck & Seibert 1981, verändert undergänzt nach Soriano et al. 1992 und
Leite 2002). 1 = immergrüner tropi-scher Tieflandregenwald (Küstenre-
genwald), 2 = immergrüner und saiso-naler subtropischer Lorbeerwald,
3 = immergrüner subtropischer Lor-beerwald mit Araucaria angustifolia, 4 = Auwälder und andere Feuchtge-biete, 5 = Mosaik aus Wäldern und
Feuchtgebieten (Parklandschaft vonEntre Rios), 6 = Sandküsten-Vegeta-
tion („Restinga“), 7 = tropische regen-grüne Laubwälder (feuchter Chaco),
8 = Trockenwälder und -gebüsche ver-schiedener Ausbildung (im Norden tro-
ckener Chaco), 9 = subtropischesGrasland (a = nördlicher Campo,
b = mittlerer Campo, c = südlicherCampo, d = Überflutungs-Pampa, e =
Pampa). Orte: Cu Curitiba, PA PortoAlegre, MV Montevideo, BA Buenos
Aires, BB Bahia Blanca.
Ocotea porosa (imbuia) Araucaria angustifolia (pinheiro-do-Paraná)
Merostachys multirameaMerostachys multiramea(taquara)(taquara)
Merostachys multiramea(taquara) Clethra scabra (carne-de-vaca)
Drimys brasiliensis (casca-d'anta)Calyptranthes concinna (guamirim-ferro)Allophylus guaraniticus (concon)
Nectandra lanceolata (canela-amarela)Lamanonia ternata (guaperê)Ocotea pulchella (canela-lajeang)
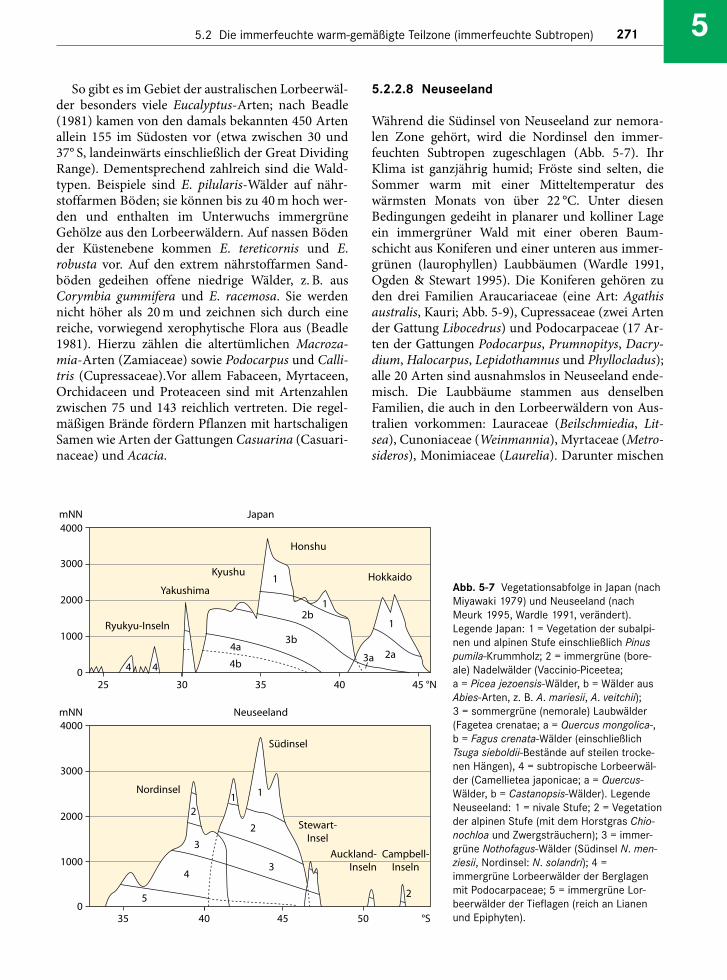
Abb. 5-5 Schema eines Araukarienwaldes auf dem Planalto von Südbrasilien (aus Klein 1984, reproduziert mit freundlicher Geneh-migung von Prof. Ademir Reis, Herbário Barbosa Rodrigues, Itajaí, Brasilien). Unter dem Schirm aus Araucaria angustifolia gedeihtein laurophyller Wald aus Lauraceae (Ocotea, Nectandra), Cunoniaceae (Lamanonia), Sapindaceae (Allophylus), Myrtaceae (Calyp-tranthes) und Winteraceae (Drimys). An aufgelichteten Stellen bildet der Bambus Merostachys undurchdringliche Dickichte. DieHöhe der Araukarien beträgt etwa 30 m, die Laurophyllen sind rund 20 m hoch.

trockenere Klimaphasen im Pleistozän zurückzufüh-ren und hat nichts mit den gegenwärtigen Klimabe-dingungen zu tun (Behling et al. 2004). Einige derEmergenten haben deshalb keine längere kahlePhase; sie treiben innerhalb weniger Tage nach demLaubabwurf wieder aus (wie Cordia americana,Boraginaceae), ein Rhythmus, den man auch bei densaisonalen Bäumen der Araukarienwälder beobach-tet (Laubfall in der regenärmsten Periode von Aprilbis Juni; Marques et al. 2004). In den regenreichenGipfellagen der Serra do Mar, mit Niederschlägenüber 2000 mm, sind die Wälder be sonders reich anEpiphyten und Baumfarnen (Dicksonia sellowiana).Hier, wie auch in den tieferen Lagen, ist Ilex para-guayensis beheimatet; aus seinen fermentierten undgerösteten Blättern wird Mate-Tee zubereitet (Abb.5-6).
Auf dem nach Westen abfallenden Hochplateaudes Planalto gedeiht der südbrasilianische Arauka-rienwald mit Araucaria angustifolia. Er ist mosaikar-tig von subtropischem Grasland durchsetzt, das alsRelikt einer trockenen Klimaperiode gedeutet wird(s. Abschn. 5.2.3). Das Gebiet mit einer Höhenlagezwischen 600 und 1.600 m NN wird regelmäßig vonFrösten heimgesucht; selbst Schneefall ist durchaushäufig, wenngleich die Schneedecke kaum einmalmehrere Zentimeter überschreitet und auch nurwenige Tage Bestand hat. Unter dem Schirm ausAraukarien findet sich eine zweite Baumschicht ausLauraceen (Ocotea, Nectandra), den südhemisphäri-schen Cunoniaceen (Lamanonia, Weinmannia), den
Winteraceen (Drimys), den Myrtaceen (Calyptran-thes) und den Aquifoliaceen (Ilex; Klein 1984; Abb. 5-3f, 5-5). Hinzu tritt als weitere altertümliche Gym-nosperme Podocarpus lambertii.
Die südbrasilianischen Lorbeerwälder einschließ-lich ihrer von Araukarien dominierten Ausbildungauf dem Planalto haben zusammen mit den tropi-schen Tieflandregenwäldern der Küstenebene (Oli-veira-Filho & Fontes 2000) als „Mata Atlantica“ we -gen ihrer hohen Artenzahlen unter Wissenschaftlernund in der (nicht nur brasilianischen) Öffentlichkeitgroße Aufmerksamkeit gefunden (Galindo-Leal &Camara 2003). Sie bilden ein Beispiel für denerschreckenden Rückgang tropischer und subtropi-scher Vegetation innerhalb weniger Jahrzehnte (s.Abschn. 5.2.4).
5.2.2.6 Südafrika
Unter den südhemisphärischen subtropischen Lor-beerwäldern ist der südafrikanische Anteil der klein-ste; er nimmt lediglich 0,25 % der Fläche der Repu-blik Südafrika ein (Eeley et al. 2001). Die Wälder sindauf Western Cape, Eastern Cape und KwaZulu-Natalbeschränkt, wo sie zwischen 25 und 35° S in der Küs-tenebene und dem westlich anschließenden Berglanddes „Great Escarpment“ (Drakensberge) in Formstark fragmentierter, von subtropischem Graslandumgebener Flecken vorkommen (Mucina & Gelden-huys 2006). Das größte und unter Schutz gestellte
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 269
a b
Abb. 5-6 Mate (Ilex paraguayensis,Aquifoliaceae): a = weiblicher Baum, b = Mate-trinkender Gaucho (Rio Grande do Sul, Brasilien).

Waldgebiet, das den Charakter der südafrikanischenLorbeerwälder wohl am besten zeigt, ist der KnysnaForest in der Umgebung der gleichnamigen Stadt(Abb. 5-3g). Er gehört zu einem Waldtyp, der in Süd-afrika als „Afromontane Forest“ bezeichnet wirdund außer Lauraceae (wie Ocotea bullata), Aquifolia-ceae (Ilex mitis) und Oleaceae (Olea capensis) eineReihe alter Gondwana-Elemente temperierter Her-kunft aufweist (Midgley et al. 1997). Hierzu gehörenu. a. die beiden Podocarpaceen Podocarpus latifoliusund Afrocarpus falcatus, ferner Faurea macnaughto-nii (Proteaceae) sowie Cunonia capensis. In der Pro-vinz Western Cape finden sich diese Wälder auf Mee-resniveau; nach Norden steigen sie weiter nach obenund gedeihen in KwaZulu-Natal in Höhenlagen zwi-schen 800 und 1.500 m NN. Hier, im Tiefland derKüstenebene des Indischen Ozeans, ist der zweiteWaldtyp vertreten, der als „Coastal Forest“ bezeich-net wird und in dem die oben genannten temperier-ten Taxa weitgehend fehlen. Seine Artenzusammen-setzung hat somit eher tropischen Charakter. Derdritte Waldtyp nimmt floristisch eine Zwischenstel-lung zwischen beiden ein; er gedeiht im Hügellandder Küstenebene („Scarp Forest“).
Während die Wälder der Tieflagen wohl erst vorrund 8.000 Jahren von Norden her in das Gebiet ein-gewandert sind, dürften die afromontanen Wälderschon vor dem Pleistozän existiert haben, und zwarschon immer in einer stark zersplitterten Form mitzahlreichen kleineren und größeren Waldinseln ineiner Matrix aus Grasland (Eeley & al. 1999; s.Abschn. 5.2.3). Deshalb findet man – im Gegensatzzu den erst in jüngster Zeit anthropogen fragmen-tierten Wäldern aller Klimazonen – keine Hinweiseauf einen Randeffekt (wie verringerte Artenzahlenmit abnehmender Inselgröße und zunehmender Dis-tanz zwischen den Inseln; Kotze & Lawes 2007).
5.2.2.7 Australien
Entlang der Ostküste von Australien verläuft einnach Osten abfallender Gebirgszug geringer Höhe(Great Dividing Range), der die humide Küstenebenevon dem trockeneren Hinterland trennt. Hier wie anden Gebirgshängen kommen immergrüne Laubwäl-der von der Cape-York-Halbinsel bis an die Südspitzevor (Beadle 1981, Webb & Tracy 1994, Busby &Brown 1994). Wie sonst nur in Südostasien gehen dietropischen Tieflandregenwälder (s. Abschn. 2.1.8) imNorden, die subtropischen Lorbeerwälder in derMitte und die nemoralen Lorbeer- und Nothofagus-Wälder im Süden kontinuierlich ineinander über.
Entlang dieses Gradienten nehmen die Artenzahl,der Reichtum an Kormo-Epiphyten und die Regen-waldmerkmale (Brettwurzeln, Träufelspitzen, Kauli-florie) von Nord nach Süd ab. Auf der SüdspitzeAustraliens südlich der Australischen Alpen und aufTasmanien bestehen die Wälder nur mehr aus weni-gen immergrünen Baumarten mit Eucalyptus-Domi-nanz; das Vorkommen der Gattungen Nothofagusund Eucryphia verweist auf die Verwandtschaft mitden südchilenischen immergrünen Laubwäldern(nemorale Lorbeerwälder, immergrüne nemoraleNothofagus-Wälder; s. Abschn. 6.2.4).
Im Vergleich zu Südamerika und Ostasien sind diebis zu 40 m hohen subtropischen LorbeerwälderAustraliens artenarm. Sie werden von zwei Arten dersüdhemisphärischen Gondwana-Familie Cunoniceaedominiert, nämlich Ceratopetalum apetalum undSchizomeria ovata, denen sich einige weitere Artendieser Familie sowie einige Lauraceae beimischen(wie verschiedene Cryptocarya- und Endiandra-Ar -ten). Hinzu kommen Vertreter der Monimiaceae(Doryphora), der Myrtaceae (wie Syzygium) und derAraucariaceae (Araucaria cunninghamii). Die zuletztgenannte Baumart bildet, ähnlich wie A. angustifoliain Südbrasilien, Dominanzbestände mit einer lauro-phyllen zweiten Baum- und Strauchschicht. Faba-ceen sowie typisch tropische Baumfamilien (Melia-ceae) treten zurück. Der krautige Unterwuchs bestehthauptsächlich aus Farnen (Adiantum-Arten sowieBaumfarne der Gattungen Dicksonia und Cyathea).Landeinwärts folgen mit zunehmender Trockenheitoffene sklerophylle Eucalyptus-Wälder (z. B. mit E.albens und E. melliodora).
Die subtropischen Lorbeerwälder beginnen in derHöhe des Wendekreises (bei Rockhampton) undenden ungefähr bei 37° S. Sie bilden kein zusammen-hängendes Band, sondern sind in Teilgebiete frag-mentiert, wenn auch nicht in so extremer Weise wiein Südostafrika. Diese Fragmentation hängt damitzusammen, dass die Unregelmäßigkeit des Nieder-schlagsgeschehens (wie überall in Australien) unddie stellenweise extrem nährstoffarmen oder nassenBöden nicht überall das Gedeihen eines Lorbeerwal-des zulassen. Solche extremen Standorte werden vonEucalyptus-Wäldern eingenommen, da es unter denüber 600 Arten praktisch immer eine oder mehreregibt, die mit den jeweiligen Stressfaktoren gut zu -rechtkommen (Beadle 1981). Eucalyptus-Arten tre-ten außerdem als Pioniere auf, wenn Lorbeerwälderdurch Rodung oder Windwurf zerstört wurden. Diestresstoleranten Eukalypten übernehmen also inAustralien die Rolle der Föhren auf der Nordhemi-sphäre.
270 5 Die warm-gemäßigte (subtropische) Zone5
So gibt es im Gebiet der australischen Lorbeerwäl-der besonders viele Eucalyptus-Arten; nach Beadle(1981) kamen von den damals bekannten 450 Artenallein 155 im Südosten vor (etwa zwischen 30 und37° S, landeinwärts einschließlich der Great DividingRange). Dementsprechend zahlreich sind die Wald-typen. Beispiele sind E. pilularis-Wälder auf nähr-stoffarmen Böden; sie können bis zu 40 m hoch wer-den und enthalten im Unterwuchs immergrüneGehölze aus den Lorbeerwäldern. Auf nassen Bödender Küstenebene kommen E. tereticornis und E.robusta vor. Auf den extrem nährstoffarmen Sand -böden gedeihen offene niedrige Wälder, z. B. ausCorymbia gummifera und E. racemosa. Sie werdennicht höher als 20 m und zeichnen sich durch einereiche, vorwiegend xerophytische Flora aus (Beadle1981). Hierzu zählen die altertümlichen Macroza-mia-Arten (Zamiaceae) sowie Podocarpus und Calli-tris (Cupressaceae).Vor allem Fabaceen, Myrtaceen,Orchidaceen und Proteaceen sind mit Artenzahlenzwischen 75 und 143 reichlich vertreten. Die regel-mäßigen Brände fördern Pflanzen mit hartschaligenSamen wie Arten der Gattungen Casuarina (Casuari-naceae) und Acacia.
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 271
5.2.2.8 Neuseeland
Während die Südinsel von Neuseeland zur nemora-len Zone gehört, wird die Nordinsel den immer-feuchten Subtropen zugeschlagen (Abb. 5-7). IhrKlima ist ganzjährig humid; Fröste sind selten, dieSommer warm mit einer Mitteltemperatur deswärmsten Monats von über 22 °C. Unter diesenBedingungen gedeiht in planarer und kolliner Lageein immergrüner Wald mit einer oberen Baum-schicht aus Koniferen und einer unteren aus immer-grünen (laurophyllen) Laubbäumen (Wardle 1991,Ogden & Stewart 1995). Die Koniferen gehören zuden drei Familien Araucariaceae (eine Art: Agathisaustralis, Kauri; Abb. 5-9), Cupressaceae (zwei Artender Gattung Libocedrus) und Podocarpaceae (17 Ar -ten der Gattungen Podocarpus, Prumnopitys, Dacry-dium, Halocarpus, Lepidothamnus und Phyllocladus);alle 20 Arten sind ausnahmslos in Neuseeland ende-misch. Die Laubbäume stammen aus denselbenFamilien, die auch in den Lorbeerwäldern von Aus -tra lien vorkommen: Lauraceae (Beilschmiedia, Lit- sea), Cunoniaceae (Weinmannia), Myrtaceae (Metro-sideros), Monimiaceae (Laurelia). Darunter mischen
25
4 4 4b
3b
3a
2b
2a
1
1
1
4a
Ryukyu-Inseln
Yakushima
Kyushu
Honshu
Hokkaido
30 35
Japan
40 45 °N0
1000
2000
3000
4000mNN
Nordinsel
Südinsel
Stewart-Insel
Auckland-Inseln
Campbell-Inseln
35
5
4
3
3
2
2
21 1
40 45
Neuseeland
50 °S0
1000
2000
3000
4000mNN
Abb. 5-7 Vegetationsabfolge in Japan (nachMiyawaki 1979) und Neuseeland (nachMeurk 1995, Wardle 1991, verändert).Legende Japan: 1 = Vegetation der subalpi-nen und alpinen Stufe einschließlich Pinuspumila-Krummholz; 2 = immergrüne (bore-ale) Nadelwälder (Vaccinio-Piceetea; a = Picea jezoensis-Wälder, b = Wälder ausAbies-Arten, z. B. A. mariesii, A. veitchii); 3 = sommergrüne (nemorale) Laubwälder(Fagetea crenatae; a = Quercus mongolica-,b = Fagus crenata-Wälder (einschließlichTsuga sieboldii-Bestände auf steilen trocke-nen Hängen), 4 = subtropische Lorbeerwäl-der (Camellietea japonicae; a = Quercus-Wälder, b = Castanopsis-Wälder). LegendeNeuseeland: 1 = nivale Stufe; 2 = Vegetationder alpinen Stufe (mit dem Horstgras Chio-nochloa und Zwergsträuchern); 3 = immer-grüne Nothofagus-Wälder (Südinsel N. men-ziesii, Nordinsel: N. solandri); 4 =immergrüne Lorbeerwälder der Berglagenmit Podocarpaceae; 5 = immergrüne Lor-beerwälder der Tieflagen (reich an Lianenund Epiphyten).

Standort und Breiten- bzw. Höhenlage, wird aberauch von Störungen durch Vulkanausbrüche beein-flusst. So kann man mehrere Entwicklungsstadiender Sukzession auf Lava oder vulkanischer Asche inräumlicher Nachbarschaft studieren. Sie unterschei-den sich u. a. durch ihren Anteil an Koniferen, die inder Jugend lichtbedürftig sind, eher Pioniercharakterhaben und daher die ersten Sukzessionsstadien auf-bauen. Da einige, wie Podocarpus totara, langlebig(bis zu 1000 Jahre) sind, findet man sie auch noch imEndstadium der Vegetationsentwicklung, bei demdas vulkanische Ereignis schon Jahrhunderte zurück-liegt. Welche Baumarten am Waldaufbau beteiligtsind, hängt aber auch von der jeweiligen Phase imRegenerationszyklus ab. Nach einem altersbedingten(kleinflächigen) Bestandszusammenbruch sind Laub-bäume (wie die Cunoniacee Weinmannia und dieMyrtacee Leptospermum) die Pioniere, in derenSchatten die Koniferen keimen und als „Oskars“
272 5 Die warm-gemäßigte (subtropische) Zone5
sich regelmäßig Vertreter der Proteaceae (z. B. Knigh-tia excelsa) und der Elaeocarpaceae. Der Unterwuchsbesteht aus Farnen (z. B. Blechnum) und Baumfarnen(Cyathea, Dicksonia). Die für Australien prägendeGattung Eucalyptus fehlt vollständig. Die hohe Luft-feuchte in dem hochozeanischen Klima fördert dieEntwicklung austrocknungsempfindlicher Kryptoga-men, darunter viele Vertreter der Hymenophyllaceae(Hautfarne), die Stämme und Äste der Bäume dichtüberziehen. Ihre Blätter bestehen aus einer ein-schichtigen Lamina ohne Spaltöffnungen mit gro-ßen, manchmal mit dem bloßen Auge erkennbarenZellen. Die Familie enthält mehrere Gattungen mitinsgesamt über 600 Arten, von denen 25 auf Neusee-land vorkommen.
Diese Koniferen-Laubbaum-Mischwälder stellendie natürliche Vegetation auf der Nordinsel sowie imNorden und in der Mitte der Südinsel dar (Abb. 5-3h, i). Ihre Artenzusammensetzung variiert je nach
a
c
b d
Abb. 5-8 Agathis australis, Waipoa Forest Park, NordinselNeu seeland: a = Alter Baum mit geradem Stamm und hoch an -gesetzter Krone; b = Jungbäume; c = männliche Zapfen; d = weib-licher Zapfen.

(zum Begriff s. Abschn. 3.1.2.4) auf günstige Lichtbe-dingungen für ihre weitere Entwicklung zu adultenIndividuen warten. In der Regel zeichnet sich die Rei-fephase deshalb durch eine physiognomische Domi-nanz verschiedener Podocarpaceen wie des häufigen,bis zu 50 m hohen Emergenten Dacrydium cupressi-num aus (Abb. 5-3i).
Einen derartigen Waldentwicklungszyklus zeigenz. B. die Kauri-Wälder auf dem „Northland“ derNordinsel. Die von den Ureinwohnern Neuseelands,den Maori, Kauri genannte Araucariacee Agathisaustralis beeindruckt als bis zu 60 m hoher Emergentunter ungestörten Bedingungen mit ihrem zylinder-förmigen, unverzweigten Stamm (Durchmesser 3–4 m) und ihrer mächtigen, ausladenden Krone (Abb. 5-8a). Der Waldentwicklungszyklus (Abb. 5-9) be -ginnt mit der Erstbesiedlung einer Bestandslückedurch die Pioniersträucher Leptospermum scopariumund Kunzea ericoides (Proteaceae). Unter derenSchirm entwickeln sich reich beastete Jungbäumevon Agathis mit langgestreckter Kronenform (Abb. 5-8b). Nach dem Absterben der Pioniere wächst Aga-this hoch und bildet den für erwachsene Individuencharakteristischen astlosen Stamm mit breiter Krone.Unter diesem Schirm kommt dann im nächstenSchritt die zweite Baumschicht aus immergrünenLaubbäumen hoch. Die Reifephase ist erreicht. Sie istreich an Lianen, Baumfarnen und Epiphyten underinnert strukturell an oreotropische Nebelwälder.Unter den Lianen fällt die zu den indo-pazifischenPandanaceae gehörende Gattung Freycinetia mitihren langen, schmalen Blättern auf.
Kauri wurde von den europäischen Einwanderernwegen des widerstandsfähigen (harzreichen) Holzesfür den Bau von Schiffen und die Produktion vonMöbeln intensiv ausgebeutet. Das bernsteinartigeHarz fand (und findet) Verwendung bei der Herstel-lung von Schmuck. Mehr als 94 % des ursprünglichenBestands (13.000 km2) der Wälder sind auf dieseWeise innerhalb der letzten 150 Jahre verschwunden.Deshalb steht Kauri heute unter strengem Schutz.
5.2.3 Subtropisches Grasland
5.2.3.1 Merkmale
Nach Walter & Breckle (1999) wird die zonale Vege-tation außerhalb der Gebirge in ebener Lage und auf„mittleren“ Böden, die weder zu nass noch zu tro-cken sind, vom Allgemeinklima bestimmt. In denimmerfeuchten Subtropen besteht die zonale Vegeta-tion aus immergrünen bzw. saisonalen Laubwäldern,deren Physiognomie und Artenzusammensetzungmit den in den Klimadiagrammen zum Ausdruckkommenden thermischen und hygrischen Merkma-len gut übereinstimmt. Umso mehr verwundert es,dass es in Südamerika und in Südafrika neben diesenWäldern ein baumfreies Grasland gibt, dessen Matrix
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 273
weitere Arten
Leptospermum
Kauri-Sämling
Kauri
Kunzea
a
c
e
b
d
Abb. 5-9 Waldentwicklungszyklus von Agathis australis-(Kauri-)Wäldern auf der Nordinsel von Neuseeland (aus Ogden & Stewart 1995, geringfügig verändert). a = Bestandslücke durchStörung (in der Realität meist größer als hier dargestellt), b (nach 50 Jahren) = Leptospermum scoparium (Myrtaceae)und Kunzea ericoides (Proteaceae) siedeln sich als Pioniere an,dazwischen keimen Kauri-Samen; c (nach 150 Jahren) = Kunzeastirbt altersbedingt ab, die Agathis-Sämlinge sind zu Jungbäu-men mit schmaler Krone herangewachsen; d (nach 250 Jahren)= Bildung der für alte Bäume typischen Agathis-Krone; e (nach350 Jahren) = Reifephase des Waldbestands mit ausgewachse-nen Agathis-Emergenten und einer zweiten Baumschicht auslaurophyllen Laubbäumen.

überwiegend aus C4-Gräsern gebildet wird (Abb. 5-10a, b, c). Der Anteil der C3-Gräser nimmt mit stei-gender Meereshöhe und geographischer Breite zu.Das Klima unterscheidet sich nicht wesentlich vondemjenigen der Waldgebiete: Die Jahresniederschlägeliegen zwischen 600 (an der Grenze zu den westlichanschließenden Trockengebieten) und 2.000 mm inden Gebirgen; im kühlen Winter mit gelegentlichemFrost kann eine kurze (maximal dreimonatige) Tro-ckenzeit auftreten (Juni bis August; s. Abb. 5-1f, g, h).Das Klima ist subtropisch-sommerfeucht nach Troll& Paffen (1964) und dem Klimatyp Cfa (warm-feuchtgemäßigt) bzw. Cfb (kühl-feuchtgemäßigt; inGebirgslagen) nach Köppen-Trewartha zuzuordnen(Trewartha & Horn 1980).
Die Vegetation wird überwiegend von C4-Horst-gräsern der Poaceen-Unterfamilien Chloridoideaeund Panicoideae geprägt. In Südamerika sind die C3-Arten der Pooideae regelmäßig beigemischt; sie wer-den nach Süden immer häufiger, bis sie südlich desRio de la Plata die Vorherrschaft übernehmen. InAfrika dominieren die C3-Gräser in Lagen oberhalb2.000 m NN. Die Pflanzendecke wird unter günstigenBedingungen (tiefgründige Böden, warmes Klima)bis über 1 m hoch. Die jährlich erzeugte oberirdischePhytomasse beträgt 5 t ha–1 oder mehr. Die C4-Gräservertrocknen in der kühlen Jahreszeit, sodass es zuEngpässen bei der Futterversorgung der Weidetierekommen kann. Verbunden mit dem langsamenAbbau der nährstoffarmen Streu gehören Grasfeuer
274 5 Die warm-gemäßigte (subtropische) Zone5
Die altertümlichen KoniferenDie Ausbreitung der Angiospermen und der Rückgang derGymnospermen sind zwei der wichtigsten phytogeographi-schen Prozesse in der Erdgeschichte (Enright et al. 1995).Die Gymnospermen, deren Entstehungszeit in das mittlereDevon datiert (etwa 365 Mio. Jahre BP), dürften aber selbstzur Zeit ihrer größten Vielfalt nicht mehr als einige TausendArten erreicht haben. Heute sind noch rund 900 Artenübrig. Sie gehören zu den Ordnungen Cycadales, Ginkgo -ales, Gnetales (mit Gnetaceae, Ephedraceae und Welwit-schiaceae) und Pinales (= Coniferales; Koniferen). DerenName („Zapfenträger“) ist nicht ganz korrekt, weil bei vie-len Taxa die weiblichen Zapfen auf eine Blüte reduziert sindund die Samenschuppe den Samen als fleischiges Organ(Epimatium) umhüllt (Bresinsky et al. 2008), sodass dieFrüchte wie eine Beere aussehen (z. B. bei Taxus). Dierezenten Pinales bestehen aus sieben Familien (mit 615Arten; Farjon 2010a), von denen einige Vertreter der aus-schließlich auf der Nordhalbkugel vorkommenden Pinaceae(zwölf Gattungen, 232 Arten) Reinbestände vorzugsweise inden kalten Klimaten aufbauen. Die übrigen Familien und eingroßer Teil der Pinaceae haben ihren Schwerpunkt in dernemoralen und subtropischen Zone; nordhemisphärischsind die Taxaceae (fünf Gattungen, ca. 25 Arten), dieCephalotaxaceae (eine Gattung, ca. sechs Arten, Ostasien)sowie die monotypische Familie Sciadopityaceae, die mitnur einer Art in Japan vorkommt (Sciadopitys verticillata).Auf der Nord- und auf der Südhemisphäre gleichermaßengedeihen als einzige kosmopolitische Familie der Koniferendie Cupressaceae, die (neuerdings einschließlich der Taxo-diaceae) 29 Gattungen mit 135 Arten umfassen, währendAraucariaceae (drei Gattungen, ca. 23 Arten) und Podocar-paceae (18 Gattungen, 174 Arten) fast ausnahmslos südlichdes Äquators vorkommen (Farjon 2010a, Turner & Cernusak2011).
Von den sieben Familien der Pinales sind Araucariaceenund Podocarpaceen mit ersten fossilen Nachweisen in derälteren Trias die ältesten Koniferen (Hill 1995). Auslöser fürihre Entwicklung dürfte die zunehmende Austrocknung amEnde des Paläozoikums gewesen sein. Die rezente GattungAraucaria gibt es seit der frühen Kreidezeit, während Fossi-lien von Agathis in Ostaustralien aus dem Jura stammen. DieAraucariaceen erreichten ihre maximale Diversität in Juraund Kreide (Kershaw & Wagstaff 2001); sie waren in dieserZeit nahezu auf allen Kontinenten, auch in Europa, verbreitet(Kunzmann 2007). Der letzte Nachweis in Europa stammtaus der oberen Kreidezeit (Maastricht); die dort gefundenenZapfenschuppen entsprechen ziemlich genau der heutenoch existierenden Araucaria heterophylla (Van der Ham etal. 2010), heute endemisch auf den zwischen Neukaledonienund Neuseeland liegenden Norfolkinseln. Im Tertiär unterla-gen sie der Konkurrenz der Angiospermen und überlebten inden wenigen Refugien auf der Südhemisphäre. Ihre heutigenVorkommen sind also reliktisch. Auch die Pinaceen entwi-ckelten sich bereits während der frühen Jurazeit, und zwarauf Laurasia; Pinus entstand vermutlich vor rund 135 Mio.Jahren (Unterkreide), Abies vor 110 und Larix vor 115 Mio.Jahren (Lin et al. 2010). Die südhemisphärischen Gattungender Cupressaceae wie Libocedrus, Callitris und Austrocedrussind fossil erst aus dem frühen Tertiär nachgewiesen.
Koniferen sind, bezogen auf das einzelne Individuum, imVergleich zu vielen Angiospermen-Bäumen meist ziemlichlanglebig. Bei einem Vergleich von 44 Koniferen und 76Angiospermen in Nordamerika erreichten Erstere ein durch-schnittliches maximales Lebensalter von 400 Jahren, wäh-rend Letztere nur 250 Jahre alt wurden (Loehle 1988). Aufder Südhemisphäre werden Fitzroya cupressoides (3.600Jahre), gefolgt von Lagarostrobos franklinii (2.260), Agathisaustralis (1.680) und Araucaria araucana am ältesten (1.300;
Kasten 5-2

vor allem am Ende des eher trockenen Winters zumErscheinungsbild der Vegetation. Der größte Teil derBrände ist anthropogen; das Grasland der großenViehfarmen wird abgebrannt, um den Weiderest zubeseitigen und den Wiederaustrieb im Frühjahr zubeschleunigen (Abb. 5-10e). Dass Feuer aber schonvor der Einwanderung der Europäer aufgetreten sindund wohl ein intrinsisches Merkmal jeder hoch-wüchsigen Graslandvegetation darstellen, zeigt sichan den Merkmalen der beteiligten Arten (Overbeck& Pfadenhauer 2007, Fidelis et al. 2010a, b): DieHorstgräser schützen ihr Meristem vor Hitze durchdicht gepackte Sprosse (wie im Fall von Andropogonlateralis; Abb. 5-10f); die Zwergsträucher besitzenHypokotylknollen (Lignotuber), aus denen sie wie-
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 275
der austreiben, wenn die oberirdischen Organe ver-brannt sind; der Austrieb der zahlreichen monokoty-len Geophyten des südafrikanischen Graslandes wirddurch die Beseitigung der Grasstreu stimuliert (Abb.5-10g).
In Abschn. 5.2.2.1 haben wir darauf hingewiesen,dass die subtropischen Wälder mosaikartig mit demGrasland verzahnt sind. In Südamerika findet manein derartiges Mosaik in den sog. nördlichen Campos(s. unten) als ein Ökoton zwischen Araukarienwaldund Grasland in Südbrasilien; in Südafrika sind dieLorbeerwälder inselförmig im Grasland verteilt. Siegedeihen entweder als Galeriewälder entlang vonFließgewässern, in Tälern, auf den feuchteren undkühleren Nordhängen oder auf felsigen Kuppen. Da
Enright & Ogden 1995). Auf der Nordhemisphäre können dieCupressaceen Sequoiadendron giganteum (Kalifornien), Cu -pressus nootkatensis (Nordwestküste Nordamerika), Cha-maecyparis obtusa (Japan) und Cryptomeria japonica (derNationalbaum von Japan) sowie Pinus longaeva (White Moun-tains, Kalifornien; über 4.000 Jahre) ein Alter von 3.000 Jah-ren erreichen; zwischen 1.000 und 2.000 Jahren liegen u. a.Pseudotsuga menziesii (1.200), Sequoia sempervirens (2.200),Taxodium distichum und Thuja plicata (beide 1.200 Jahre;Suzuki & Tsukahara 1986, Loehle 1988, Schütt et al. 2004,Eckenwalder 2009). Der älteste Baum der Erde ist ein Exem-plar von Pinus longaeva mit 4.806 Jahren (Lanner 2007).Diese Langlebigkeit ist das Ergebnis von langsamem Wachs-tum und chemischer Verteidigung gegenüber Schadorganis-men durch Baumharz. Die Mehrzahl der Koniferen ist in derJugend lichtbedürftig und zeigt somit Pioniercharakter; dieSamen sind häufig leicht und anemochor. Zoochorie (durchVögel) tritt vor allem bei Podocarpaceen und manchen Arau-caria-Arten auf. Die Blätter sind nadel- oder schuppenförmigmit nur einer zentralen Blattader ohne Verzweigung.
Vermutlich als Reaktion auf die Entstehung des Laub-blattes haben manche Koniferen ihre Sprosse zu blattartigenOrganen umgebildet. So zeigen einige der südhemisphäri-schen Taxa mehr oder minder ganzrandige, aus Kurzspros-sen entstandene Phyllokladien (wie die Gattung Phyllocla-dus) oder sie haben die ursprünglich radial angeordnetenNadeln in eine Ebene gedreht, sodass der Eindruck einesgefiederten Blattes entsteht (Dacrycarpus; Hill 1995). Eineähnliche, wenn auch nicht so perfekte Annäherung an einAngiospermenblatt zeigt sich auch bei den Taxaceen und dereuropäischen Weißtanne (Abies alba). Solche komplexenAssimilationsorgane sind in den lichtintensiven Tropen undSubtropen photosynthetisch effizienter als die skleromor-phen, äquifazialen Nadeln. Taxa mit dieser Ausstattungkonnten deshalb mit der Angiospermenentwicklung im Alt-tertiär besser mithalten (Coomes & Bellingham 2011; vgl.
hierzu auch das „buchenähnliche“ Verhalten von Abies albaund Taxus baccata in montanen mitteleuropäischen Wäl-dern; Ellenberg & Leuschner 2010; s. Abschn. 6.2.2.6.2).Hier zeigt sich, dass laubbaumähnliche Entwicklungen beiden Koniferen Koexistenz zwischen beiden Wuchsformenermöglichen, wie im Fall der Mischwälder in Neuseeland,Tasmanien und Südchile zu sehen ist (s. auch Kap. 6).
Koniferen mit Nadeln oder Schuppen sind den Blüten-pflanzen dagegen nur unter extremeren Umweltbedingun-gen überlegen. Sofern sie nicht im Lauf des Neogen frost -hart wurden, überlebten sie die Angiospermenausbreitungunter subtropischen oder nemoralen Klimabedingungen vorallem als Pioniere (z. B. nach massiven Störungen wieErdrutsch, Vulkanausbruch oder Feuer wie im Fall der nemo-ralen Nadelwälder des nordöstlichen Nordamerika mit Pseu-dotsuga menziesii), aber auch auf trockenen oder feuchtenbis nassen Standorten (wie Austrocedrus chilensis und Fitz-roya cupressoides in Chile) und in Gebirgslagen (wie Arauca-ria angustifolia in Südbrasilien und Araucaria araucana inChile). Ihre Langlebigkeit erlaubt ihnen eine jahrhunderte-lange Präsenz in ansonsten von Laubbäumen dominiertenWäldern (Enright et al. 1999). Tiefe Temperaturen ertragensie nur begrenzt (Bannister 2007). Ganz anders ist die Situ-ation im Fall der frostharten Pinaceen auf der Nordhemi-sphäre. Die Entwicklung dieser „modernen“ Bäume geht aufden Abkühlungsprozess am Ende des Tertiärs zurück. VieleArten der Gattungen Pinus, Picea, Abies und Larix könnensomit in der borealen Zone zur Dominanz gelangen, auchwenn immer sommergrüne Laubbäume (wie Populus undBetula) beigemischt sind. Die Nadelbäume wachsen im kalt-gemäßigten Klima zwar schneller als Laubbäume, weil siefrüher im Jahr mit der Assimilation beginnen und späterdamit aufhören, verlieren diesen Vorteil aber durch diehöhere Assimilationsleistung der Laubbäume, die dafür ihreBlattmasse jährlich erneuern müssen (Schulze et al. 2002).

die beteiligten Baumarten nicht feuerresistent sind,liegt der Schluss nahe, dass dieses Mosaik das Ergeb-nis regelmäßiger Brände ist (Pillar & Quadros 1997),stellenweise kombiniert mit dem Verbiss durch Wei-detiere. Denn alle derzeit waldbestandenen Flächenwerden weniger leicht von Feuer erfasst als die freienHänge und Plateaus, sei es wegen der höheren Bo -den- und Luftfeuchte, sei es wegen des Mangels anbrennbarem Material (Felskuppen).
Im Gegensatz zu den Savannen ist das subtropi-sche Grasland völlig baumfrei. Zwar gibt es am Randmanchmal savannenartige Strukturen (wie die „Espi-nales“ aus Acacia- und Prosopis-Arten am Westranddes Graslandes in Südamerika); innerhalb des Gras-landes treten Bäume aber nur auf Standorten auf, dievor Feuer geschützt sind, oder es handelt sich umSukzessionsflächen, die von Pioniergehölzen besie-delt werden. Auch gibt es keine feuerresistentenBäume oder Sträucher mit dicker Borke und resprou-ter-Eigenschaften. Von dort sind in der Vergangen-heit auch keine Holzpflanzen eingewandert, auch
276 5 Die warm-gemäßigte (subtropische) Zone5
wenn Savannen an das Grasland angrenzen wie inSüdafrika. Die Ursache liegt vermutlich darin, dassdie Savannenbäume tropischen Ursprungs mit denkühlen Temperaturen im Winter nicht zurechtkom-men (Bredenkamp et al. 2002).
In der Literatur wird das subtropische Graslandunter verschiedenen Bezeichnungen geführt. Ver-breitet ist die Bezeichnung Steppe, wegen der physio-gnomischen und floristischen Ähnlichkeit (Vorkom-men von C3-Gräsern) mit der baumfreien Vegetationder trockenen nemoralen Zone Nordamerikas undAsiens (IBGE 2004). Ökologisch unterscheidet sichdas subtropische Grasland aber fundamental von dennemoralen Steppen, worauf wir unten zu sprechenkommen (s. Henning 1988). Hueck (1966) wendetden regional auf das Grasland südlich des Rio de laPlata beschränkten Begriff Pampa auch für (auf denersten Blick) ähnliche Vegetationstypen in Uruguayund Südbrasilien an, obwohl das eine mit demandern kaum Gemeinsamkeiten aufweist (s. Abschn.5.2.3.3). Auch die Bezeichnung „Grassavanne“ ist
a b
c d
Abb. 5-10 Beispiele für subtropische Grasländer. a = nördlicher Campo mit Galeriewald aus Araucaria angustifolia bei São Fran-cisco de Paula, Rio Grande do Sul, Brasilien; b = mittlerer Campo bei Dom Pedrito, Rio Grande do Sul, Brasilien, c = Pampa ausNasella neesiana, Bromus catharticus und Paspalum dilatatum, im Hintergrund Celtis iguanaea, Cannabaceae, auf einem Dünenrü-cken (Foto F. Klötzli); d = Grasland im Golden Gate National Park, Südafrika, mit Themeda triandra als dominanter Art.

nicht angebracht; denn die Savannen sind in densommerfeuchten Tropen nur auf Vertisolen und sehrflachgründigen Standorten baumfrei (s. das „Savan-nengrasland“ in Abschn. 3.3.3). Solche Standortbe-dingungen treffen aber für das subtropische Graslandnicht zu. Wir bleiben deshalb bei der Bezeichnung„subtropisches Grasland“ und verwenden zusätzlichdie regionalen Begriffe Campo (Pl. Campos) für Süd-brasilien und Uruguay, Pampa für Argentinien undGrassland für Südafrika (s. unten).
5.2.3.2 Vorkommen und Verbreitung
5.2.3.2.1 SüdamerikaDas subtropische Grasland reicht in Südamerika vonetwa 25 (etwa auf der Höhe von Curitiba) bis 38° S
(Bahia Blanca in Argentinien; Soriano et al. 1992,Leite 2002, Royo Pallarés et al. 2005; Abb. 5-4). Esbesteht aus zwei Teilen, nämlich einem nördlich desRio de la Plata gelegenen Gebiet (die „Campos“ vonSüdbrasilien und Uruguay) und der südlich davongelegenen Pampa Argentiniens. „Pampa“ bedeutet inder Sprache der einheimischen Indianerstämme(Quechua) „baumlose Ebene“ und entspricht damitdem spanisch-portugiesischen Wort campo. DieCampos werden in einen nördlichen, mittleren undsüdlichen Abschnitt untereilt, die sich floristisch undstrukturell voneinander unterscheiden: • Der nördliche Campo liegt nördlich des 30. Brei-
tengrads auf einem Hochland („Planalto“), dasvon 1.500 m der Küstenrandstufe („Serra Geral“)im Osten nach Westen bis zum Rio Paraná aufunter 600 m NN abfällt (Abb. 5-10a). Er bildet ein
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 277
f g
e
Abb. 5-10 (Fortsetzung) e = anthropo-gen abgebranntes Grasland bei Bad-plaas, Südafrika; f = Wiederaustriebeines Horstes von Andropogon lateralisnach Feuer; g = Hypoxis sp. (Hypoxida-ceae) als Beispiel eines nach Feuerrasch austreibenden Geophyten ineinem südafrikanischen Grasland.

Mosaik mit Araukarien- und subtropischen Lor-beerwäldern. Biogeographisch gehört er zusam-men mit diesen zur Region Mata Atlântica und istfloristisch mit den zentralbrasilianischen Savan-nen verwandt. Die jährlichen Niederschläge be -tragen zwischen 1.500 und 2.000 mm; die Jahres-mitteltemperatur variiert je nach Höhenlagezwischen 16 und 22 °C. Die sauren, basenarmenBöden sind überwiegend Alisole mit einem zuStaunässe neigenden Bt-Horizont; in höherenLagen fallen Böden mit bis zu 40 cm mächtigemA-Horizont auf, die zu den Umbrisolen gestelltwerden (Dümig et al. 2008a). Die Vegetation wirdvon hochwüchsigen C4-Gräsern dominiert (wieAndropogon lateralis, Schizachyrium tenerum, S.spicatum); darunter kommen in einer zweitenGrasschicht verschiedene Axonopus- und Paspa-lum-Arten vor. Asteraceen stellen mit 24 % diemeisten Arten; dazu gehören einige Zwergsträu-cher aus der Gattung Baccharis (wie B. trimera mitgeflügelten Ästen). Physiognomisch fallen dieroten Blüten von Petunia-Arten (Solanaceae) unddie hochwüchsigen Rosettenpflanzen von Eryn-gium horridum (Apiaceae) mit scharfen Stachelnan den Blatträndern auf. Das Grasland bedeckt dieHänge und Kuppen des Planalto überall dort, wotiefgründige Böden vorherrschen. Die Wälderwachsen auf felsigen Kuppen oder galerieartig ent-lang von Fließgewässern, können aber auch großezusammenhängende Flächen einnehmen. DieWald-Grasland-Grenze ist scharf.
• Nach Süden, jenseits des 30. Breitengrads, folgtder mittlere, ebenfalls von Feuer geprägte Ab -schnitt der Campos im Zentrum und im Südenvon Rio Grande do Sul und in Uruguay (mittlererCampo; Abb. 5-10b). Hier treten zu den domi-nanten C4-Gräsern wie Andropogon lateralis undmehreren Aristida-Arten (wie Aristida jubata) C3-Gräser der mit Stipa verwandten Gattung Nasella(z. B. N. neesiana), Melica und Briza hinzu. Sie bil-den die bis zu 1 m hohe obere Grasschicht. Dieuntere Grasschicht besteht aus Paspalum- undAxonopus-Arten. Neben den zahlreich vertrete-nen Asteraceen sind Le gumi nosen häufig, die vorallem durch die Gattung Adesmia repräsentiertwerden. Klassische Nutzungsweise ist auch hierdie Rinderbeweidung. Florengeographisch gehörtdas Gebiet zur Pampa-Provinz (Cabrera & Willink1980); die jährlichen Niederschläge sind mit1.200–1.600 mm geringer und die Jahresmittel-temperaturen mit 20–16 °C etwas niedriger als imNorden. Die Zahl der Gefäßpflanzenarten wirdfür den Teil der nördlichen und mittleren Cam-
pos, die zu Brasilien gehören („Campos Sulinos“),mit mehr als 3.000 angegeben, wovon die Gräserrund 25 % ausmachen, gefolgt von den Asteraceenmit 20 % und den Fabaceen mit 7 % (Boldrini2009). Auf skelettreichen Böden kommen Gebü-sche vor, die vorwiegend aus Asteraceen der Gat-tungen Baccharis, Eupatorium und Heterothala-mus sowie der weltweit verbreiteten SapindaceeDodonaea viscosa bestehen.
• Der südliche Campo am Rio de la Plata ist floris-tisch und physiognomisch der Hügelpampa (s.unten) sehr ähnlich; hier wie dort dominierenNasella-Arten (wie N. charruana), und die Pflan-zendecke ist nur einschichtig. Er erstreckt sich vonder Atlantikküste bis zum Rio Uruguay und um -fasst westlich dieses Flusses auch das Grasland imsüdlichen Entre Rios.
• Die Pampa südlich des Rio de la Plata grenzt imSüden und Westen an die Trockenwälder und -gebüsche des Monte und des Chaco (Abb. 5-10c).Diese Trockenwälder tragen wegen ihres dornigenCharakters die Bezeichnung „Espiñal“; sie beste-hen aus regengrünen Mimosaceen wie Acaciacaven und verschiedenen Prosopis-Arten (P. calde-nia, P. nigra) sowie der Fabacee Geoffroea decorti-cans. Abgesehen von der flachwelligen Landschaftim Nordwesten („Hügelpampa“ bei Buenos Aires)ist das Gebiet der Pampa eine Ebene in einerHöhenlage zwischen 0 und 200 m NN, unterbro-chen von zwei niedrigen Bergländern im Zentrumund im Süden (Abb. 5-4). Die jährliche Mitteltem-peratur liegt zwischen 16 °C im Norden und 13 °Cim Süden. Die Niederschläge nehmen von Nordennach Süden und von Osten nach Westen ab; sie lie-gen im Nordosten noch bei 1.000 mm, erreichenim Südwesten aber lediglich rund 400 mm. Abge-sehen von den Bergländern ist das Gebiet von Lössbedeckt. Die Böden werden teilweise den Phaeo-zemen zugeordnet, ein Bodentyp, der besondersgroßflächig in den Waldsteppen und Steppen dernemoralen Zone der Nordhemisphäre auftritt(Zech et al. 2014): Er ist basenreich und zeichnetsich durch einen humusreichen, mächtigen Ober-boden aus. Im Süden können auch Kastanozemeund Chernozeme auftreten. Sonst gibt es großflä-chig Vertisole. In der Pflanzendecke dominierenmit den Gattungen Nasella (23 Arten), Poa,Melica, Piptochaetium und Bothriochloa die C3-Gräser, wenn auch C4-Gräser nicht fehlen (Aris-tida, Panicum, Paspalum u. a.). Etwa ein Dritteldes Gebiets wird regelmäßig überflutet („Überflu-tungs-Pampa“). Hier sind die Bö den stellenweisesalzbeeinflusst (Solonetz) und in dem niedrigen
278 5 Die warm-gemäßigte (subtropische) Zone5

Grasland ist das halophytis che Gras Distichlis sco-paria häufig. Abgesehen von solchen Standortenist die Pampa heute intensiv agrarisch genutzt;naturnahes Grasland ist auf wenige kleine Rest-bestände beschränkt.
5.2.3.2.2 SüdafrikaIn Südafrika gehört das subtropische Grasland zum„Grassland Biome“ (Mucina et al. 2006), das mitknapp 355.000 km2 nach dem „Savanna Biome“(413.000 km2) das zweitgrößte Biom der RepublikSüdafrika ist und 28 % der Landesfläche einnimmt(Rutherford et al. 2006). Das Biom reicht von 25 bis33° S und besteht aus vier ökologisch und biogeogra-phisch verschiedenen Graslandregionen:• „Drakensberg Grassland“ (im Süden des Hochlan-
des mit den höchsten Erhebungen; ganzjährighumid mit Jahresniederschlägen zwischen 700und 1.200 mm mit kaltem Winter; in höherenLagen Vorkommen von C3-Gräsern),
• „Dry Highveld Grassland“ (im Landesinnern imÜbergang zu den zentralen Trockengebieten undbis zu viermonatiger Trockenzeit im Winter undJahresniederschlägen zwischen 400 und 600 mm),
• „Mesic Highveld Grassland“ (im Osten des Hoch-landes mit maximal dreimonatiger Trockenzeitund Jahresniederschlägen zwischen 600 und 1.400 mm; Abb. 5-10d) und
• „Sub-Escarpment Grassland“ (am Fuß der Dra-kensberge und des Escarpments mit Niederschlä-gen zwischen 500 und 1.000 mm und einer bis zuzwei Monate dauernden Trockenzeit).
Das südafrikanische Grasland grenzt im Norden undOsten an die Savannen der sommerfeuchten Tropenund im Süden und Südwesten an die Zwergstrauch-halbwüsten der Karoo. Die Böden sind in den höhe-ren Gebirgslagen saure, basenarme, intensive ge -färbte Cambi- und Luvisole oder Leptosole. DieVegetation besteht aus C4-Gräsern. Überall verbreitetund häufig ist Themeda triandra, ein bis zu 150 cmhoch werdendes Horstgras, das meist in Savannenund somit auch in Südostasien und Australien vor-kommt; hinzu treten verschiedene Eragrostis-, Cym-bopogon-, Setaria-, Aristida-Arten und Vertreterzahlreicher weiterer Gattungen (O’Connor & Bre-denkamp 1997). In der Vegetationskarte von Süd-afrika werden zahlreiche lokale Ausbildungen be -schrieben, die sich floristisch, geographisch undstandörtlich voneinander unterscheiden. So nehmeninnerhalb des „Mesic Highveld Grassland“ (mit ins-gesamt rund 4.000 Kormophyten; Cowling et al.1989) zwischen etwa 28 und 29° S das „Eastern Free
State Clay Grassland“ (Gm 3 in Mucina et al. 2006)und das „Eastern Free State Sandy Grassland“ (Gm 4)die größten Flächen ein. Beide Graslandtypen be -finden sich auf einer Höhe zwischen 1.400 und 1.800 m NN und liegen in einer hügeligen Landschaftmit mittleren Jahresniederschlägen um 700 mm.Beweidung mit Rindern ist weit verbreitet. In Gm 4liegt der Golden Gate National Park, einer der be -kanntesten Nationalparks von Südafrika, mit einerbeeindruckenden Gebirgslandschaft und einer rei-chen Fauna.
Erwähnenswert sind einige der unter dem Über-begriff „Drakensberg Grasslands“ zusammengefas-sten Graslandtypen oberhalb 1.900 m NN, weil sie,wie das „Lesotho Highland Basalt Grassland“, be -sonders reich an Endemiten sind. C3-Gräser wie Fes -tuca- und Koeleria-Arten sind regelmäßig vorhan-den. Dazwischen mischen sich Hemikryptophytenund Chamaephyten, darunter viele Asteraceae (z. B.zahlreiche Arten von Helichrysum). Auffallende Blü-tenteppiche bildet Kniphofia caulescens, die zu denXanthorrhoeaceae gehört, einer auch in Australienverbreiteten, südhemisphärischen Familie (Gras-baumgewächse). Einige Flächen dieser Formationliegen ebenfalls im Golden Gate National Park.
Allerdings sind keineswegs alle Gebiete reineGrasländer, wie der Name „Grassland Biome“ sugge-rieren mag (Mucina et al. 2006). So gibt es außer denverstreuten Inseln afromontaner Wälder, die wirbereits in Abschn. 5.2.2.6 behandelt haben, flächen-haft ausgedehnte Gebüsche auf steinigen, flachgrün-digen Böden, an felsigen Hängen oder lokalklima-tisch trockenen Leegebieten mit je nach Lageunterschiedlicher Artenzusammensetzung. Ein Bei-spiel ist das „Basotho Montane Shrubland“ mit Budd-leja salviifolia, Euclea crispa und einer afrikanischenUnterart von Olea europaea (Free State Province,Lesotho). Auch savannenartige Strukturen kommenvor, z. B. mit Protea rubripilosa auf Quarzit im nörd-lichen Abschnitt des Drakensberg-Hochlandes.
5.2.3.3 Grasland gegen Wald und Wald gegen Grasland?
5.2.3.3.1 Grasland gegen Wald – der Vergleich mit Savannen und Steppen Die Gründe für das Vorkommen von Grasländern inden immerfeuchten Subtropen der Südhemisphärewaren lange Zeit ein Gegenstand kontroverser Dis-kussionen. Unter deutschen Ökologen und Pflanzen-geographen hat man sich vor allem mit der PampaArgentiniens und ihrer Natürlichkeit beschäftigt (das
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 279

„Pampa-Problem“; s. Ellenberg 1962, Walter 1967,Troll 1968, Henning 1988). Die nördlich anschlie-ßenden Campos blieben dagegen weitgehend außerAcht, obwohl schon der deutschstämmige Jesuiten-pater B. Rambo in seiner Fisionimia do Rio Grande doSul (1956) vermutet hat, dass sie ein Relikt einer frü-heren Trockenperiode sein müssten (vgl. Overbeck etal. 2007). Eine ähnliche Diskussion wie über diePampa gab es auch im Fall des südafrikanischenGraslandes (Bredenkamp 2002).
Immer dann, wenn in einem Gebiet Pflanzenge-meinschaften vorkommen, die nicht mit der aus demAllgemeinklima zu erwartenden zonalen Vegetationübereinstimmen, ist die Ursache für diese Abwei-chung entweder auf die Tätigkeit des Menschen(anthropogene Vegetation) zurückzuführen oderbestimmte Bodeneigenschaften verhindern die Aus-bildung der zonalen Vegetation (azonale Vegetation).Dass die Ursachen in den verwendeten Klimadatenselbst liegen können, wird oft zu wenig bedacht. Sogeben die Daten, wie sie in den Klimadiagrammennach Walter und Lieth zum Ausdruck kommen, nurdas Allgemeinklima eines beschränkten Zeitraumeswieder (zwischen zehn und 60 Jahre, selten mehr; s.Abschn. 1.3.1); sie müssen also nicht repräsentativfür eine zonale Vegetation sein, die sich vor mehrerenJahrhunderten oder gar Jahrtausenden entwickelthat. Außerdem spiegeln die Diagramme kurzfristigeUnregelmäßigkeiten im Niederschlagsgeschehen nichtwider, obwohl diese für die Vegetation von Bedeu-tung sein können.
Um die Frage nach der Natürlichkeit der subtropi-schen Grasländer Südamerikas und Afrikas zu klä-ren, sind ein Rückblick auf die Savannen der som-merfeuchten Tropen (s. Abschn. 3.1.2.2) und eineVorschau auf die Steppen der nemoralen Zone hilf-reich (s. Abschn. 6.3). Das baumfreie Savannengras-land ist azonal; es gedeiht auf Sonderstandorten,nämlich auf Böden, deren Eigenschaften das Vor-kommen von Bäumen verhindern (extreme Flach-gründigkeit durch oberflächennahe Petroplinthit-oder andere Krusten, Vertisole). Auf den normaldrainierten Nitisolen bzw. Lixisolen erschwert dieregelmäßige Störung durch Feuer die Ausbildungeines dichten Waldes; die Gehölze, die dennoch unterdiesen Bedingungen wachsen, sind einigermaßenfeuerresistent. Trotz eines Klimas, das mit bis zu sie-ben ariden Monaten trockener ist als das Klima derimmerfeuchten Subtropen, wären die sommerfeuch-ten Tropen ein Waldland.
Das wirklich einzige und eindeutig zonale Gras-land (außerhalb der alpinen Stufe der Hochgebirgeund der polaren Tundra) sind die nemoralen Steppen
der Nordhemisphäre. Sie sind völlig baum-, wennauch nicht völlig gehölzfrei. Der Grund hierfür liegtnicht primär in einer negativen klimatischen Wasser-bilanz; denn das Klima ist selbst im kontinentalenZentralasien noch humid, wenngleich sich im Früh-jahr und Herbst aride Phasen andeuten (s. ausführ-lich in Abschn. 6.3.1). Den Baumwuchs verhindertvielmehr die lange thermische Winterruhe mit denkaum von Schnee geschützten, tief gefrorenen Bö -den; im zwar heißen, aber kurzen Sommer reichendann die Niederschläge nicht für den erfolgreichenAbschluss eines Vegetationszyklus aus.
Dagegen gibt es im subtropischen Grasland(außerhalb der Pampa) weder Baumwuchs verhin-dernde Krusten und Tonböden wie im Savannen-grasland noch kalte Winter mit kurzer Vegetations-periode wie in den nemoralen Steppen. Klima- undBodeneigenschaften sprechen also für Wald, jeden-falls unter den gegenwärtigen Klimabedingungen.
5.2.3.3.2 Menschlicher EinflussBliebe also noch die Frage, ob das Grasland anthro-pogen ist. Diese Frage ist naheliegend, denn auch ver-meintlich (d. h. nach unserer heutigen Vorstellungeiner technisierten Landnutzung) „extensiv“ wirt-schaftende indigene Kulturen können die Vegetationeiner Landschaft im Lauf der Jahrtausende nachhal-tig und tiefgreifend verändern. Jede Bevölkerungs-gruppe strebt danach, im Rahmen ihrer technischenMöglichkeiten so viel Ertrag wie möglich zu erzielen.Zu den besonders auffälligen Erscheinungen gehörtdie anthropogene Veränderung der Lage der Wald-und Baumgrenzen, selbst bei einer geringen Bevölke-rungsdichte, wie sich am Beispiel der tropischenHochgebirge eindrucksvoll zeigt (s. Kasten 4-5 inAbschn. 4.5.2). Deshalb liegt die Vorstellung nahe, imFall der subtropischen Grasländer handele es sichebenfalls um das Ergebnis einer ausgedehnten Wald-zerstörung, vorbereitet von der indigenen Bevölke-rung durch die Anwendung von Feuer für die Jagd,und zu Ende gebracht von den europäischen Ein-wanderern, die mit ihren Rindern, Schafen und Zie-gen großflächige Weidesysteme aufbauten. Unter-stützt wird diese Ansicht durch den Erfolg, denLandeigner mit Aufforstungen aus nicht-heimischenBaumarten haben, vor allem mit Pinus-Arten nord-amerikanischer Provenienz und verschiedenen Eu -kalypten (s. Abschn. 5.2.4). Auch ist die anthropo-gene Beseitigung von Wäldern in manchen Gebieten,in denen heute Grasland dominiert, nachweisbar, wiebeispielsweise in den Küstenregionen von KwaZulu-Natal und Eastern Cape in Südafrika (Mucina & Gel-denhuys 2006).
280 5 Die warm-gemäßigte (subtropische) Zone5

Gegen diese Hypothese sprechen allerdings so -wohl die Aufzeichnungen früher Forschungsreisen-der (wie des französischen Botanikers Auguste deSaint-Hilaire aus den Jahren 1820 und 1821; s. Gau-treau 2010), die übereinstimmend von ausgedehntenGrasländern in Uruguay berichteten, als auch die Er -gebnisse pollenanalytischer (palynologischer) Unter-suchungen (Meadows & Linder 1993 für Südafrika,Prieto 1996 für die Pampa und Behling 2002 für dienördlichen Campos in Brasilien). Nach den Pollen-funden in Sedimenten, die mehr als 10.000 Jahre altsind, hat das subtropische Grasland in beiden Konti-nenten schon im Spätpleistozän existiert, zu einerZeit also, in der das Klima trockener und kühler warals heute. Nimmt man an, dass die ersten Indios umdiese Zeit in den Südosten von Südamerika einge-wandert sind (z. B. Flegenheimer & Zárate 1993),muss das Grasland also schon vor der menschlichenBesiedlung bestanden haben. Auch die erfolgreichenAufforstungen mit exotischen Baumarten sprechennicht für den anthropogenen Charakter der Campos.Sie belegen lediglich deren potenzielle Baumfähigkeitunter den heutigen Klimabedingungen.
5.2.3.3.3 Wald gegen Grasland – die historischePerspektiveSeit dem Beginn der Besiedlung des südamerikani-schen Raumes durch die Indios um 10.000 Jahre BPsind hochfrequente Feuer (im Schnitt alle ein bis fünfJahre) durch den deutlichen Anstieg von Kohlen-stoffpartikeln in den Pollendiagrammen nachgewie-sen (Behling et al. 2004). Die Grasländer müssen alsoschon von jeher feuergeprägt gewesen sein. Anhandder Pollendiagramme der nördlichen Campos vonSüdbrasilien lässt sich zeigen, dass vor etwa 4.000Jahren die bis heute andauernde feuchte Phase aufdem Planalto begann (Abb. 5-11); ab diesem Zeit-punkt konnten sich die Araukarienwälder von ihrentrockenzeitlichen Refugien an der Atlantikküste vor-wiegend entlang von Flusstälern auf das südbrasilia-nische Hochland hinauf ausbreiten und erobertenschließlich ab etwa 1.300 Jahre BP größere Gebieteam Ostrand des Plateaus (Behling et al. 2004). An -hand der 13C-Signatur in der organischen Substanzdes Bodens (δ13C von –18,7 bis –14,3 ‰ bei C4-Pflanzen) konnten Dümig et al. (2008b) nachweisen,dass sich die Araukarienwälder auf dem Planalto vonRio Grande do Sul ab etwa 1.300 Jahre BP auf ehe-maligem Graslandboden entwickelt haben. Damithat sich die schon von Rambo (1956) ausgesprocheneHypothese von der Ursprünglichkeit der Campos be -stätigt. Im 17. Jahrhundert kamen mit den jesuiti-schen Missionen Pferde und Rinder in das Gebiet;
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 281
die Inbesitznahme des Graslandes durch Viehfarmen(„Façendas“) drängte die Wälder wieder zurück undschärfte die Wald-Grasland-Grenze (Pillar & Qua-dros 1997). Offensichtlich haben sich Wald-Gras-land-Grenzen aber seit dem Beginn des 19. Jahrhun-derts nicht mehr verändert, wie Gautreau (2010)durch Auswertung alter Karten und Aufzeichnungenfür Uruguay nachweisen konnte.
Auch das Grasland-Biom von Südafrika dürfte aufein trockenes Klima während der Glazialzeitenzurückzuführen sein (Mucina & Geldenhuys 2006).Wie in Südbrasilien ist die Ausbreitung von Bäumen(wenigstens in tieferen Lagen) während einer humi-den Phase zwischen 4.000 und 2.500 Jahren BP polle-nanalytisch nachweisbar. In den höheren Lagen(oberhalb 1.500 m NN) könnte das Fehlen von Lor-beerwäldern nach Bredenkamp et al. (2002) daranliegen, dass sich im Baumartenpool des gesamtensüdafrikanischen Raumes keine kältetoleranten Baum- arten finden, die potenziell in der Lage wären, dasGrasland zu besiedeln. Die feuerresistenten Holzge-wächse der Savannen sind thermisch zu anspruchs-voll, um in das kühle subtropische Grasland vorzu-dringen. Wie in den Campos Südamerikas ist aberauch im Grasland-Biom Südafrikas Feuer der ent-scheidende Faktor für die Persistenz des Graslandes.
5.2.3.3.4 Das Pampa-Problem: immer noch ein Problem?Eine Ausnahme bilden die Pampa und Teile des süd-lichen Campo: Das Klima ist ganzjährig durch unre-gelmäßig auftretende Trockenzeiten von mehrerenWochen Dauer geprägt (Soriano et al. 1992). Siekommen im Klimadiagramm nicht zum Ausdruck,weil sie im langjährigen Mittel von den reichlich fal-lenden Niederschlägen der übrigen Jahre überdecktwerden. Ohne diese Trockenzeiten wäre die Salzan-reicherung um die zahlreichen abflusslosen Senkennicht zu erklären. Auf die Diskrepanz zwischen Kli-madiagramm und tatsächlichem Klimageschehenhat übrigens schon Walter (1967) bei der Diskussionum das „Pampa-Problem“ hingewiesen.
Hinzu kommen für Bäume ungünstige Boden -eigenschaften. Da die grobporenarmen, tonreichenLössböden in ebener Lage auch noch regelmäßigüberflutet werden, wirken sich periodische Wasserbi-lanzdefizite ähnlich auf die Vegetation aus wie imSavannengrasland (Troll 1968). In der Tat gibt esWälder in Gebieten ohne Lössdecke auf grobkörni-gem Substrat, wie beispielsweise die „Espiñales“westlich der Pampa (sogar bei geringeren Nieder-schlägen), sowie die Celtis spinosa-Bestände (Canna-baceae) auf fossilen Sanddünen und Strandwällen
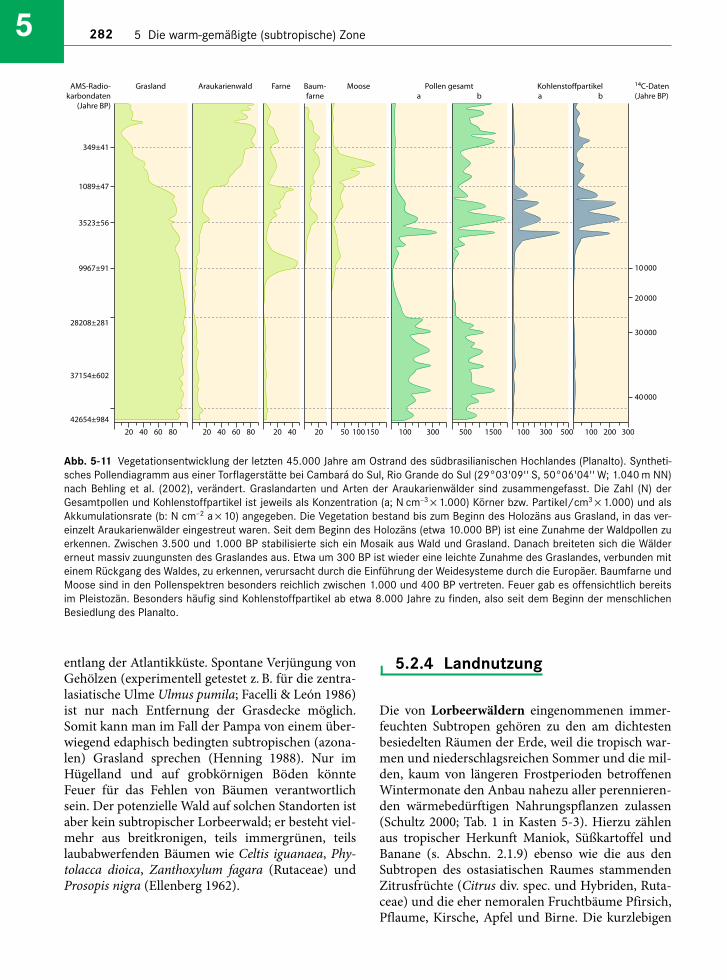
entlang der Atlantikküste. Spontane Verjüngung vonGehölzen (experimentell getestet z. B. für die zentra-lasiatische Ulme Ulmus pumila; Facelli & León 1986)ist nur nach Entfernung der Grasdecke möglich.Somit kann man im Fall der Pampa von einem über-wiegend edaphisch bedingten subtropischen (azona-len) Grasland sprechen (Henning 1988). Nur imHügelland und auf grobkörnigen Böden könnteFeuer für das Fehlen von Bäumen verantwortlichsein. Der potenzielle Wald auf solchen Standorten istaber kein subtropischer Lorbeerwald; er besteht viel -mehr aus breitkronigen, teils immergrünen, teilslaubabwerfenden Bäumen wie Celtis iguanaea, Phy-tolacca dioica, Zanthoxylum fagara (Rutaceae) undProsopis nigra (Ellenberg 1962).
5.2.4 Landnutzung
Die von Lorbeerwäldern eingenommenen immer-feuchten Subtropen gehören zu den am dichtestenbesiedelten Räumen der Erde, weil die tropisch war-men und niederschlagsreichen Sommer und die mil-den, kaum von längeren Frostperioden betroffenenWintermonate den Anbau nahezu aller perennieren-den wärmebedürftigen Nahrungspflanzen zulassen(Schultz 2000; Tab. 1 in Kasten 5-3). Hierzu zählenaus tropischer Herkunft Maniok, Süßkartoffel undBanane (s. Abschn. 2.1.9) ebenso wie die aus denSubtropen des ostasiatischen Raumes stammendenZitrusfrüchte (Citrus div. spec. und Hybriden, Ruta-ceae) und die eher nemoralen Fruchtbäume Pfirsich,Pflaume, Kirsche, Apfel und Birne. Die kurzlebigen
282 5 Die warm-gemäßigte (subtropische) Zone5
40 000
30 000
20 000
10 000
14C-Daten(Jahre BP)
42654±984
37154±602
28208±281
9967±91
3523±56
1089±47
349±41
AMS-Radio-karbondaten
(Jahre BP)
20 40 60 80
Grasland
20 40 60 80
Araukarienwald
20 40
Farne
20
Baum-farne
50 100 150
Moose Pollen gesamt Kohlenstoffpartikel
100 300
a
500 1500
b
100 300 500
a
100 300200
b
Abb. 5-11 Vegetationsentwicklung der letzten 45.000 Jahre am Ostrand des südbrasilianischen Hochlandes (Planalto). Syntheti-sches Pollendiagramm aus einer Torflagerstätte bei Cambará do Sul, Rio Grande do Sul (29°03'09'' S, 50°06'04'' W; 1.040 m NN)nach Behling et al. (2002), verändert. Graslandarten und Arten der Araukarienwälder sind zusammengefasst. Die Zahl (N) derGesamtpollen und Kohlenstoffpartikel ist jeweils als Konzentration (a; N cm–3 × 1.000) Körner bzw. Partikel/cm3 × 1.000) und alsAkkumulationsrate (b: N cm–2 a × 10) angegeben. Die Vegetation bestand bis zum Beginn des Holozäns aus Grasland, in das ver-einzelt Araukarienwälder eingestreut waren. Seit dem Beginn des Holozäns (etwa 10.000 BP) ist eine Zunahme der Waldpollen zuerkennen. Zwischen 3.500 und 1.000 BP stabilisierte sich ein Mosaik aus Wald und Grasland. Danach breiteten sich die Wäldererneut massiv zuungunsten des Graslandes aus. Etwa um 300 BP ist wieder eine leichte Zunahme des Graslandes, verbunden miteinem Rückgang des Waldes, zu erkennen, verursacht durch die Einführung der Weidesysteme durch die Europäer. Baumfarne undMoose sind in den Pollenspektren besonders reichlich zwischen 1.000 und 400 BP vertreten. Feuer gab es offensichtlich bereitsim Pleistozän. Besonders häufig sind Kohlenstoffpartikel ab etwa 8.000 Jahre zu finden, also seit dem Beginn der menschlichenBesiedlung des Planalto.

55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 283
(therophytischen), stärkeliefernden Getreidepflan-zen Mais, Reis und Weizen können in günstigen Jah-ren sogar mehrmals im Jahr angebaut werden. Dastrifft auch für die unzähligen Bohnensorten der Gat-tungen Vigna (altweltlich) und Phaseolus (neuwelt-lich) sowie für die Sojabohne (Glycine max) zu, dievon der Wildform Glycine max ssp. soja aus dem sub-tropischen Ostasien abstammt und erstmals um dasJahr 1.000 v. Chr. aus China beschrieben wurde (Lie-berei & Reisdorff 2007). Die Sojabohne ist die welt-weit bedeutendste unter den Bohnen und ein wichti-ges Exportgut vieler (vor allem südamerikanischer)Länder. Charakteristisch sind auch Tee und Kaffee,deren Kultivierung wir schon in der Lorbeerwald-stufe der feuchttropischen Gebirge in Abschn. 2.1.9beschrieben haben.
Die Anbauweisen in den Lorbeerwaldgebietensind unterschiedlich. Neben der in bereits Abschn.2.1.9 beschriebenen Wald-Feld-Wechselwirtschaftmit jahrzehntelangen Brachestadien (Entwicklungvon Sekundärwäldern unterschiedlichen Alters, Abb.5-12a) ist eine kleinbäuerliche Subsistenzwirtschaftauf wenigen Hektaren weit verbreitet, vor allem inBrasilien und China (hier besonders Nassreisanbau).Es gibt aber auch eine spezialisierte Ackerbauwirt-schaft auf größeren Betriebsflächen (Farmen), dieMarktfrüchte erzeugen (Weizen und Reis; Abb. 5-12b).
Außer Ackerbau wird Weidewirtschaft nur klein-flächig für den Eigenbedarf betrieben; lediglich inhochozeanischen Gebieten wie auf der Nordinsel vonNeuseeland haben sich ausgedehnte Weidesystememit Rindern und/oder Schafen etabliert (Abb. 5-12c).Die Vegetation besteht z. T. aus einheimischen, rege-nerationsfreudigen C4-Gräsern, wenn das Arten -potenzial im natürlichem Grasland desselben Natur-raumes zur Verfügung steht (wie Paspalum notatumin den Campos von Südbrasilien und Zoysia japonicain den Weidegebieten Japans), z. T. aus Ansaaten tro-pischer Futtergräser wie Axonopus fissifolius (Her-kunft Mittel- und Südamerika) und Cynodon dacty-lon (Herkunft Afrika). In den LorbeerwaldgebietenJa pans dient das bis zu 2,5 m hohe Gras Miscanthussinensis (Panicoideae) zur Gewinnung von Winter-futter (Ito 1990). In Regionen ohne natürliches Gras-land rekrutiert sich die Pflanzendecke überwiegendaus europäischen Grünlandpflanzen, die eingeschlepptoder angesät wurden. Hier findet man z. B. Loliumperenne, Festuca arundinacea, Cynosurus cristatusund Trifolium repens. Einige der eingeführten Pflan-zen haben sich zwischenzeitlich als aggressive Inva-soren entpuppt. So bilden die beiden aus dem hoch-ozeanischen Westeuropa stammenden Leguminosen
Stechginster (Ulex europaeus) und Besengins ter (Cy -tisus scoparius) in allen Gebieten der immerfeuchtenSubtropen Zwergstrauchheiden auf regelmäßig ge -störten Flächen (Straßenränder, Brachen und andereruderale Standorte). Beispielsweise bedeckt Ulex inNeuseeland inzwischen 3,5 % der Landesfläche (Rees& Hill 2001, McQueen et al. 2006). Der Stechginsterwurde im 19. Jahrhundert von den ersten europäi-schen Einwanderern für das Anlegen von Schutz-pflanzungen eingeführt.
Die forstwirtschaftliche Nutzung der subtropi-schen Lorbeerwälder beschränkte sich bis in die 60erJahre des 20. Jahrhunderts auf die selektive Heraus-nahme von älteren Individuen sogenannter Werthöl-zer. Das waren besonders die altertümlichen Konife-ren wie z. B. Araucaria angustifolia in Südbrasilienund Agathis australis sowie Dacrydium cupressinumin Neuseeland, deren Holz wegen ihrer Resistenz undder geraden Stämme für Schiff- und Möbelbaubesonders geschätzt war. Zwischen 1915 und 1960exportierte Brasilien mehr als 18 Mio. m3 Holz, über-wiegend aus dem Gebiet des „Mata Atlântica“ (Koch& Corrêa 2002). Heute werden aufgegebene land-wirtschaftliche Nutzflächen teils mit einheimischenGehölzen wie im Fall von Araucaria in Südbrasilien,überwiegend aber mit exotischen Baumarten, vorallem mit nordamerikanischen Pinus-Arten aufge-forstet. Auf der Nordinsel von Neuseeland sind diePinus radiata-Plantagen (Umtriebszeit 28 Jahre) mitt- lerweile auf rund 1,2 Mio. ha angewachsen (Reif1997; Abb. 5-12d). In China verwendet man alsschnellwachsende Bäume die einheimischen Pionier-Kiefern wie Pinus yunnanensis und P. massonianasowie Cryptomeria japonica; die letztgenannte Art istder bedeutendste Forstbaum in den feuchten Subtro-pen von Japan und China (5-12e).
Als Konsequenz dieser agrarischen und forst-lichen Nutzung sind subtropische Lorbeerwälder aufwenige Restbestände zusammengeschrumpft. Dasgilt für ostasiatische Länder mit ihren jahrtausende-alten Kulturen ebenso wie für solche, in denen derintensive Ackerbau erst mit der Kolonisierung durchdie Europäer begann. Bei vielen dieser Restbeständehandelt es sich – im Vergleich zu den primären Vor-kommen – um artenärmere Sekundärwälder. DiePrimärwaldreste sind vielerorts ihrer älteren Bäumeberaubt. Wie rasch der Rückgang der Wälder voran-geschritten ist, zeigt sich u. a. in Neuseeland: Dortschrumpften die ursprünglich über 1 Mio. ha umfas-senden Kauri-Wälder nach der Einwanderung euro-päischer Siedler Mitte des 19. Jahrhunderts auf 7.500 ha (Steward & Beveridge 2010). Von den zu„Mata Atlântica“ zusammengefassten subtropischen
284 5 Die warm-gemäßigte (subtropische) Zone5
Tropisch-subtropische KulturpflanzenVon den rund 352.000 bisher beschriebenen Gefäßpflan-zenarten (Paton et al. 2008) werden etwa 20.000 (5,6 %)vom Menschen genutzt (WBGU 2000). Sie liefern Stärke,Fett und/oder Eiweiß in Samen, Wurzeln oder Knollen sowieOst und Gemüse (Nahrungspflanzen). Pflanzen mit sekun-dären Pflanzenstoffen (Sekundärmetaboliten) dienen als sti-mulierende Drogen (Genussmittel- und Drogenpflanzen)oder werden für die Herstellung von Arzneimitteln verendet(Arzneipflanzen). Samen, Blätter und weitere Organe mitätherischen Ölen werden zu Gewürzen verarbeitet (Gewürz-pflanzen). Futterpflanzen sichern die Ernährung der Haus-tiere. Technische Nutzpflanzen stellen Grundstoffe für dieProduktion von Kosmetika, Schmiermittel im Maschinenbau(Rhizinus) und Gebrauchsgegenstände zur Verfügung (z. B.Faserpflanzen wie Baumwolle und Sisalagave, Holz oderkautschukliefernde Pflanzen). Die Produkte mancher, früherausschließlich der menschlichen Ernährung dienenden Fut-terpflanzen werden heute als Energiepflanzen angebaut. Inden Tropen und Subtropen gehören dazu in erster LinieZuckerrohr (für Bioäthanol) und Ölpalmen (für Biodiesel).
Die Anzahl der für einen oder mehrere dieser Zweckelandwirtschaftlich oder gärtnerisch angebauten Nutzpflan-zen dürfte (einschließlich der niederen Pflanzen) knapp5.000 be tragen. Rund 660 davon werden angebaut („kulti-viert“), davon 160 in großem Maßstab. Etwa 90 % der welt-weiten Ernährung des Menschen werden von nicht mehr als20 Pflanzen sichergestellt. Sechs davon decken über 60 %der globalen Kalorienversorgung ab, nämlich Weizen, Reis,Mais, Kartoffeln, Süßkartoffeln und Maniok (WBGU 2000).Bezogen auf die An baufläche stehen Weizen, Mais und Reisan erster Stelle, gefolgt von Sojabohnen, Kartoffeln und
Maniok (FAO 2012; Tab. 1). Die auf das Frischgewicht bezo-genen Hektarerträge sind mit über 20 t im Jahr besondershoch bei Bananen, Papayas, Wassermelonen und Zucker-rohr. Als in zweifacher Hinsicht problematisch erweist sichder großflächige Anbau von Energie- und Futterpflanzen wiez. B. Soja für die Tiermast in entwickelten Ländern. Dennerstens werden hierdurch tropisch-subtropische Primär- und(artenreiche) Sekundärwälder in großem Maßstab irreversi-bel zerstört und zweitens gehen die Anbaugebiete für diemenschliche Ernährung verloren.
Die meisten der Kulturpflanzen stammen aus den gebir-gigen Tropen und Subtropen. Ihre wild wachsenden Ur -sprungssippen kamen, soweit sie überhaupt bekannt sind,bis auf wenige Ausnahmen (wie die Kakao-Pflanze Theo-broma cacao und die kautschukliefernde EuphorbiaceeHevea brasiliensis) außerhalb der geschlossenen tropischenTiefland- und Gebirgsregenwälder vor; sie waren Besiedlervon Sukzessionsflächen (nach Störung) und Bestandteileoffener regengrüner Wälder, Trockengebüsche oder Halb-wüsten. Ihre Herkunftsgebiete werden nach ihrer Entde-ckung durch den russischen Pflanzengenetiker Nikolai Vavi-lov (1887–1943) als Vavilov-Zentren bezeichnet. Siestimmen teilweise mit den Entstehungszentren des Acker-baus überein, der auf allen Kontinenten (außer Nordamerika)fast simultan vor etwa 10.000 bis 11.000 Jahren begann(Martin & Sauerborn 2006) und in einigen Fällen mit der Ent-wicklung früher Hochkulturen zusammenfällt (Tab. 2). Ver-mutlich haben sich Hochkulturen dort gebildet, wo nebenanderen günstigen Umweltbedingungen auch Wildpflanzenvorhanden waren, die sich als Nahrungsquelle eigneten undzüchterisch bearbeitet werden konnten.
Kasten 5-3
Tab. 1 Botanische Zuordnung, Herkunft, Produktion, Ertrag und Anbaufläche einiger in den Tropen und Subtropen angebau-ter Nahrungspflanzen (zusammengestellt nach Lieberei & Reisdorff 2007 sowie FAO 2010). Bezogen auf die Anbaufläche unddie weltweite Produktion stehen Weizen, Mais und Reis an der Spitze, gefolgt von Sojabohnen, Kartoffeln und Maniok.
botanischer Name Familie und Herkunft Produktion Ertrag FlächeUnterfamilie weltweit (t ha–1)1 (¥ 1.000
(Mio t)1 km2)
Ananas Ananas comosus Bromeliaceae tropisches 19 21,4 91Südamerika
Äpfel Malus domestica Rosaceae Gebirge 70 14,7 47,3Mittelasiens2
Bananen Musa ¥ paradisiaca Musaceae Südostasien 102 21,4 47,7
Bohnen Phaseolus Fabaceae Mittel- und 23 0,8 63,1(trocken) Südamerika
Vigna Fabaceae Südasien
Dattelpalme Phoenix dactylifera Arecaceae Persischer Golf 8 7,2 10,9
Erdnüsse Arachis hypogaea Fabaceae Anden 383 1,63 240,7
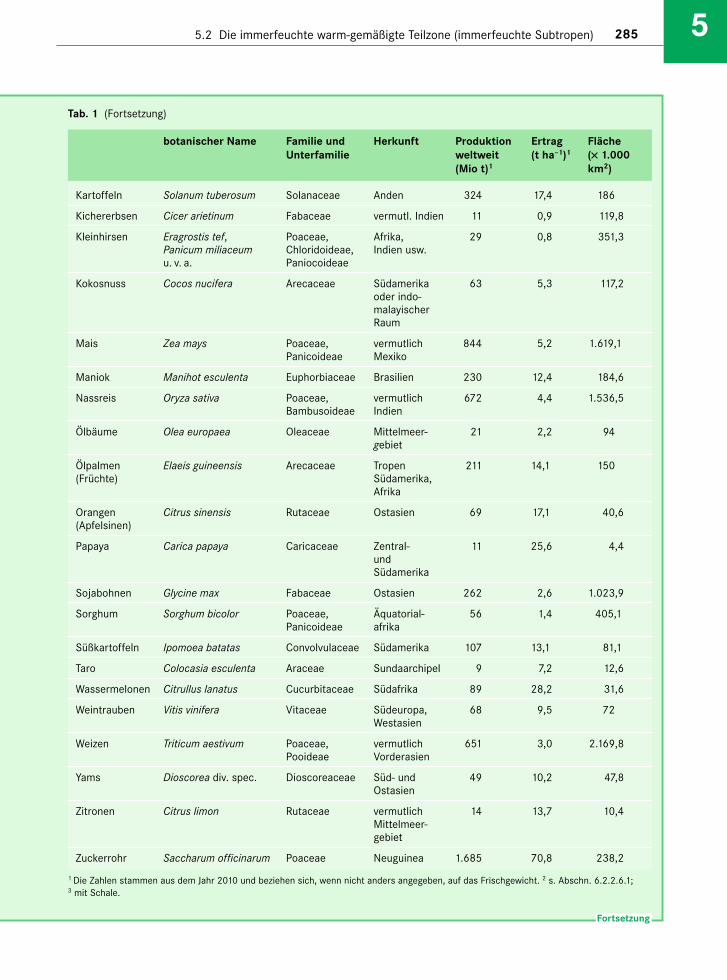
55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 285
Tab. 1 (Fortsetzung)
botanischer Name Familie und Herkunft Produktion Ertrag FlächeUnterfamilie weltweit (t ha–1)1 (¥ 1.000
(Mio t)1 km2)
Kartoffeln Solanum tuberosum Solanaceae Anden 324 17,4 186
Kichererbsen Cicer arietinum Fabaceae vermutl. Indien 11 0,9 119,8
Kleinhirsen Eragrostis tef, Poaceae, Afrika, 29 0,8 351,3Panicum miliaceum Chloridoideae, Indien usw.u. v. a. Paniocoideae
Kokosnuss Cocos nucifera Arecaceae Südamerika 63 5,3 117,2oder indo-malayischer Raum
Mais Zea mays Poaceae, vermutlich 844 5,2 1.619,1Panicoideae Mexiko
Maniok Manihot esculenta Euphorbiaceae Brasilien 230 12,4 184,6
Nassreis Oryza sativa Poaceae, vermutlich 672 4,4 1.536,5Bambusoideae Indien
Ölbäume Olea europaea Oleaceae Mittelmeer- 21 2,2 94gebiet
Ölpalmen Elaeis guineensis Arecaceae Tropen 211 14,1 150(Früchte) Südamerika,
Afrika
Orangen Citrus sinensis Rutaceae Ostasien 69 17,1 40,6(Apfelsinen)
Papaya Carica papaya Caricaceae Zentral- 11 25,6 4,4und Südamerika
Sojabohnen Glycine max Fabaceae Ostasien 262 2,6 1.023,9
Sorghum Sorghum bicolor Poaceae, Äquatorial- 56 1,4 405,1Panicoideae afrika
Süßkartoffeln Ipomoea batatas Convolvulaceae Südamerika 107 13,1 81,1
Taro Colocasia esculenta Araceae Sundaarchipel 9 7,2 12,6
Wassermelonen Citrullus lanatus Cucurbitaceae Südafrika 89 28,2 31,6
Weintrauben Vitis vinifera Vitaceae Südeuropa, 68 9,5 72Westasien
Weizen Triticum aestivum Poaceae, vermutlich 651 3,0 2.169,8Pooideae Vorderasien
Yams Dioscorea div. spec. Dioscoreaceae Süd- und 49 10,2 47,8Ostasien
Zitronen Citrus limon Rutaceae vermutlich 14 13,7 10,4Mittelmeer-gebiet
Zuckerrohr Saccharum officinarum Poaceae Neuguinea 1.685 70,8 238,2
1 Die Zahlen stammen aus dem Jahr 2010 und beziehen sich, wenn nicht anders angegeben, auf das Frischgewicht. 2 s. Abschn. 6.2.2.6.1; 3 mit Schale.
Fortsetzung
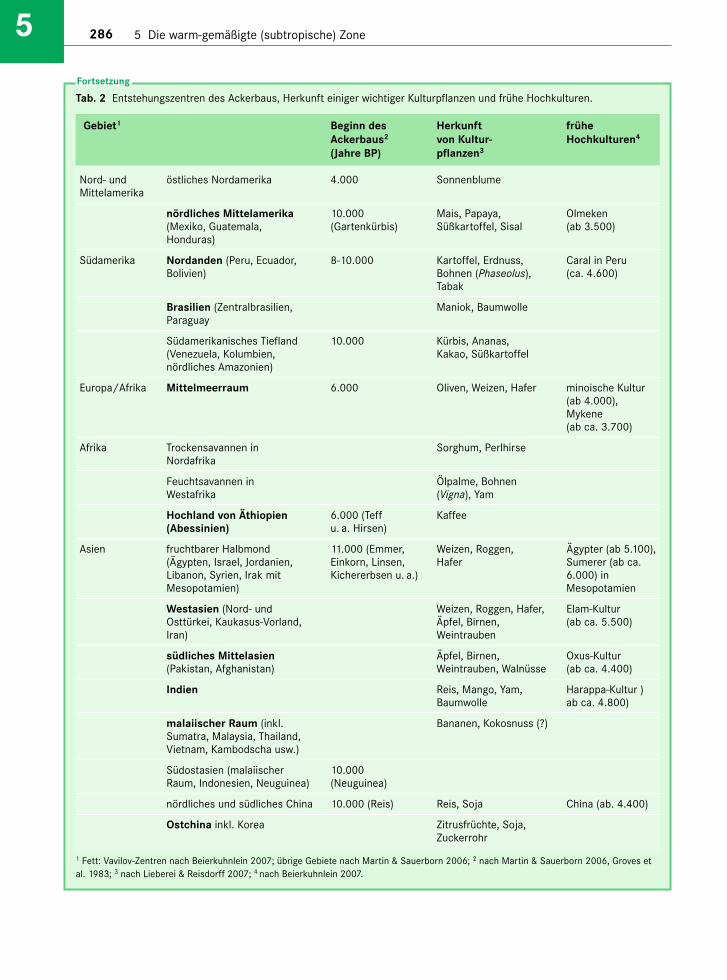
286 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 2 Entstehungszentren des Ackerbaus, Herkunft einiger wichtiger Kulturpflanzen und frühe Hochkulturen.
Gebiet1 Beginn des Herkunft frühe Ackerbaus2 von Kultur- Hochkulturen4
(Jahre BP) pflanzen3
Nord- und östliches Nordamerika 4.000 SonnenblumeMittelamerika
nördliches Mittelamerika 10.000 Mais, Papaya, Olmeken(Mexiko, Guatemala, (Gartenkürbis) Süßkartoffel, Sisal (ab 3.500)Honduras)
Südamerika Nordanden (Peru, Ecuador, 8-10.000 Kartoffel, Erdnuss, Caral in Peru Bolivien) Bohnen (Phaseolus), (ca. 4.600)
Tabak
Brasilien (Zentralbrasilien, Maniok, BaumwolleParaguay
Südamerikanisches Tiefland 10.000 Kürbis, Ananas, (Venezuela, Kolumbien, Kakao, Süßkartoffelnördliches Amazonien)
Europa/Afrika Mittelmeerraum 6.000 Oliven, Weizen, Hafer minoische Kultur (ab 4.000), Mykene (ab ca. 3.700)
Afrika Trockensavannen in Sorghum, PerlhirseNordafrika
Feuchtsavannen in Ölpalme, Bohnen Westafrika (Vigna), Yam
Hochland von Äthiopien 6.000 (Teff Kaffee(Abessinien) u. a. Hirsen)
Asien fruchtbarer Halbmond 11.000 (Emmer, Weizen, Roggen, Ägypter (ab 5.100),(Ägypten, Israel, Jordanien, Einkorn, Linsen, Hafer Sumerer (ab ca. Libanon, Syrien, Irak mit Kichererbsen u. a.) 6.000) in Mesopotamien) Mesopotamien
Westasien (Nord- und Weizen, Roggen, Hafer, Elam-Kultur Osttürkei, Kaukasus-Vorland, Äpfel, Birnen, (ab ca. 5.500)Iran) Weintrauben
südliches Mittelasien Äpfel, Birnen, Oxus-Kultur(Pakistan, Afghanistan) Weintrauben, Walnüsse (ab ca. 4.400)
Indien Reis, Mango, Yam, Harappa-Kultur )Baumwolle ab ca. 4.800)
malaiischer Raum (inkl. Bananen, Kokosnuss (?)Sumatra, Malaysia, Thailand, Vietnam, Kambodscha usw.)
Südostasien (malaiischer 10.000 Raum, Indonesien, Neuguinea) (Neuguinea)
nördliches und südliches China 10.000 (Reis) Reis, Soja China (ab. 4.400)
Ostchina inkl. Korea Zitrusfrüchte, Soja, Zuckerrohr
1 Fett: Vavilov-Zentren nach Beierkuhnlein 2007; übrige Gebiete nach Martin & Sauerborn 2006; 2 nach Martin & Sauerborn 2006, Groves etal. 1983; 3 nach Lieberei & Reisdorff 2007; 4 nach Beierkuhnlein 2007.
Fortsetzung

55.2 Die immerfeuchte warm-gemäßigte Teilzone (immerfeuchte Subtropen) 287
a
e
b
c d
Abb. 5-12 Beispiele für Kulturlandschaften aus dem Gebiet der subtropischen Lor-beerwälder. a = Wald-Feld-Wechselwirtschaft in der Serra do Mar (Santa Catarina,Brasilien) mit verschieden alten Brachstadien und Sekundärwäldern; b = Weizenan-bau anstelle von Lorbeerwäldern mit Araucaria angustifolia (Rio Grande do Sul, Brasi-lien); c = Weidelandschaft auf der Nordinsel von Neuseeland mit Podocarpus totaraals Schattenbaum; d = Aufforstungen mit Pinus radiata auf der Nordinsel von Neu-seeland; e = Cryptomeria japonica-Forst, Honshu, Japan, mit Hydrangea sp. als Klet-terpflanze an den Stämmen.
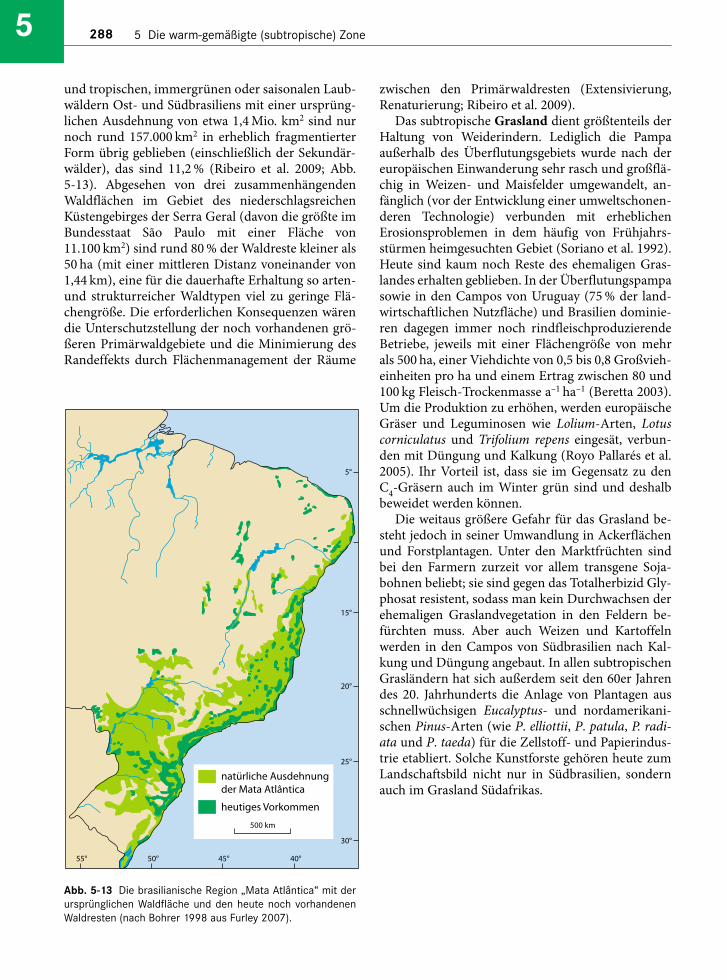
und tropischen, immergrünen oder saisonalen Laub-wäldern Ost- und Südbrasiliens mit einer ursprüng-lichen Ausdehnung von etwa 1,4 Mio. km2 sind nurnoch rund 157.000 km2 in erheblich fragmentierterForm übrig geblieben (einschließlich der Sekundär-wälder), das sind 11,2 % (Ribeiro et al. 2009; Abb. 5-13). Abgesehen von drei zusammenhängendenWaldflächen im Gebiet des niederschlagsreichenKüstengebirges der Serra Geral (davon die größte imBundesstaat São Paulo mit einer Fläche von 11.100 km2) sind rund 80 % der Waldreste kleiner als50 ha (mit einer mittleren Distanz voneinander von1,44 km), eine für die dauerhafte Erhaltung so arten-und strukturreicher Waldtypen viel zu geringe Flä-chengröße. Die erforderlichen Konsequenzen wärendie Unterschutzstellung der noch vorhandenen grö-ßeren Primärwaldgebiete und die Minimierung desRandeffekts durch Flächenmanagement der Räume
zwischen den Primärwaldresten (Extensivierung,Renaturierung; Ribeiro et al. 2009).
Das subtropische Grasland dient größtenteils derHaltung von Weiderindern. Lediglich die Pampaaußerhalb des Überflutungsgebiets wurde nach dereuropäischen Einwanderung sehr rasch und großflä-chig in Weizen- und Maisfelder umgewandelt, an -fänglich (vor der Entwicklung einer umweltschonen-deren Technologie) verbunden mit erheblichenErosionsproblemen in dem häufig von Frühjahrs-stürmen heimgesuchten Gebiet (Soriano et al. 1992).Heute sind kaum noch Reste des ehemaligen Gras-landes erhalten geblieben. In der Überflutungspampasowie in den Campos von Uruguay (75 % der land-wirtschaftlichen Nutzfläche) und Brasilien dominie-ren dagegen immer noch rindfleischproduzierendeBetriebe, jeweils mit einer Flächengröße von mehrals 500 ha, einer Viehdichte von 0,5 bis 0,8 Großvieh -einheiten pro ha und einem Ertrag zwischen 80 und100 kg Fleisch-Trockenmasse a–1 ha–1 (Beretta 2003).Um die Produktion zu erhöhen, werden europäischeGräser und Leguminosen wie Lolium-Arten, Lotuscorniculatus und Trifolium repens eingesät, verbun-den mit Düngung und Kalkung (Royo Pallarés et al.2005). Ihr Vorteil ist, dass sie im Gegensatz zu denC4-Gräsern auch im Winter grün sind und deshalbbeweidet werden können.
Die weitaus größere Gefahr für das Grasland be -steht jedoch in seiner Umwandlung in Ackerflächenund Forstplantagen. Unter den Marktfrüchten sindbei den Farmern zurzeit vor allem transgene Soja-bohnen beliebt; sie sind gegen das Totalherbizid Gly-phosat resistent, sodass man kein Durchwachsen derehemaligen Graslandvegetation in den Feldern be -fürchten muss. Aber auch Weizen und Kartoffelnwerden in den Campos von Südbrasilien nach Kal-kung und Düngung angebaut. In allen subtropischenGrasländern hat sich außerdem seit den 60er Jahrendes 20. Jahrhunderts die Anlage von Plantagen ausschnellwüchsigen Eucalyptus- und nordamerikani-schen Pinus-Arten (wie P. elliottii, P. patula, P. radi-ata und P. taeda) für die Zellstoff- und Papierindus-trie etabliert. Solche Kunstforste gehören heute zumLandschaftsbild nicht nur in Südbrasilien, sondernauch im Grasland Südafrikas.
288 5 Die warm-gemäßigte (subtropische) Zone5
5°
15°
20°
25°
30°
40°45°50°55°
natürliche Ausdehnungder Mata Atlântica
heutiges Vorkommen
500 km
Abb. 5-13 Die brasilianische Region „Mata Atlântica“ mit derursprünglichen Waldfläche und den heute noch vorhandenenWaldresten (nach Bohrer 1998 aus Furley 2007).

5.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen)
5.3.1 Grundlagen
5.3.1.1 Vorkommen und Verbreitung
Mit rund 3,16 Mio. km2 (= 2,1 % der Landoberflächeder Erde) nehmen die winterfeuchten Subtropen(subtropische Winterregengebiete) die kleinste Flä-che von allen Klimazonen ein. Sie kommen aus-schließlich an der Südwest- und Westseite der Konti-nente zwischen 30 und 40° südlicher bzw. nördlicherBreite vor und grenzen äquatorwärts an die tropisch-subtropischen Trockengebiete, polwärts an die ne -mo rale Zone (s. Abb. 1-30). Von den insgesamt fünfTeilgebieten sind zwei auf der Nordhemisphäre(mediterranes und kalifornisches Winterregenge-biet) und drei auf der Südhemisphäre vertreten (chi-lenisches, südafrikanisches und australisches Winter-regengebiet; Tab. 5-4): 1. Das mediterrane Winterregengebiet ist mit etwa
1,8 Mio. km2 das größte der fünf Teilgebiete. SeineWest-Ost-Ausdehnung reicht von Portugal überden Irak bis an den Fuß des Hindukusch über eineDistanz von rund 4.000 km; entlang dieses Gra-dienten kommt es zu einem ausgeprägten Ozea-nitäts-Kontinentalitäts-Gefälle, das den übrigenTeilgebieten dieser Klimazone fehlt. In der Vegeta-tion kann man deshalb zwischen einem eher oze-anisch geprägten westmediterranen und einemeher kontinental geprägten ostmediterranenRaum unterscheiden. Die maximale Nord-Süd-Ausdehnung zwischen dem nördlichsten Vorkom-men am Golf von Genua bei etwa 44° N und demsüdlichsten in Südwestmarokko bei etwa 29° Nbeträgt etwa 1.700 km. Mit Ausnahme von Slowe-nien und Kroatien, die zur nemoralen Zone gehö-ren, und Ägypten, das gänzlich von tropisch-sub-tropischen Trockengebieten eingenommen wird,haben alle Anrainerstaaten des Mittelmeers einenmehr oder minder großen Anteil an dieser Öko-zone.
2. Das kalifornische Winterregengebiet besteht auseinem bis zu 200 km breiten und rund 1.800 kmlangen Streifen entlang der Pazifikküste, unterbro-chen vom Kalifornischen Längstal (Sacramentound San Joaquin Valley) mit seiner Halbwüsten -vegetation. Es reicht vom südlichen Oregon bei
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 289
42° N bis nach Mexiko (Baja California), wo esrund 250 km südlich der Grenze zu den USA mitzunehmenden Sommer- und abnehmenden Win-terregen in tropische Trockenwälder übergeht.Seine Ausdehnung nach Osten wird durch die Kli-mascheide der Hochgebirge der Sierra Nevadaund ihrer südlichen Ausläufer verhindert, in derenRegenschatten die Wüstengebiete von Nevada undKalifornien (Mojave, Sonora) vorkommen.
3. Das chilenische Winterregengebiet liegt inMittelchile und reicht von 31° bis 38° S. Es geht imNorden in die Atacama-Wüste über. Im Südenschließen sich mit abnehmender sommerlicherTrockenzeit überwiegend immergrüne nemoraleWälder an. Im Osten bilden die Anden die Grenze.
4. Das kleinste der subtropischen Winterregenge-biete ist mit knapp 90.000 km2 das südafrikani-sche Winterregengebiet (Kapregion). Es umfasstden äußersten Süden und Südwesten Afrikas süd-lich der Trockengebiete von Karoo und Namib,liegt innerhalb der kapensischen Florenregion (s.Abschn. 1.2.2) und ist nahezu identisch mit demFynbos-Biom.
5. Das australische Winterregengebiet besteht auszwei Teilgebieten, die durch die Halophyten-Halb-wüste der Nullarbor-Plains voneinander getrenntsind (s. Abb. 4-23), nämlich ein stark durchGebirgszüge gegliedertes Teilgebiet im Süden umAdelaide, das nach Osten in die immerfeuchtenSubtropen übergeht, und ein zweites, weitgehendebenes Teilgebiet im Südwesten des Kontinents(um Perth). Beide grenzen gegen das Landesin-nere an Trockengebüsche und Halbwüsten.
5.3.1.2 Klima und Boden
Das subtropische Winterregenklima ist eine erdge-schichtlich junge Erscheinung. Die Klimaänderungzu einem trockenen Sommer und einem feuchten,thermisch milden Winter vollzog sich synchron inallen fünf Teilgebieten erst am Ende des Pliozän vorrund vier Mio. Jahren und stabilisierte sich endgültigvor ca. 2,5 Mio. Jahren, also kurz vor dem Beginn desPleistozäns. Das Klima ist also durch ein Wasserdefi-zit während der für den Pflanzenwuchs thermischgünstigen Jahreszeit gekennzeichnet. Im Winter fälltder überwiegende Teil des Niederschlags, wobeiregelmäßig Winterstürme auftreten und auch Schneebis auf Meeresniveau fallen kann. In allen fünf Gebie-ten nimmt die Dauer der sommerlichen Trockenzeitäquatorwärts zu, bis bei etwa 300 mm Jahresnieder-schlag und mehr als sieben Monaten Trockenzeit die
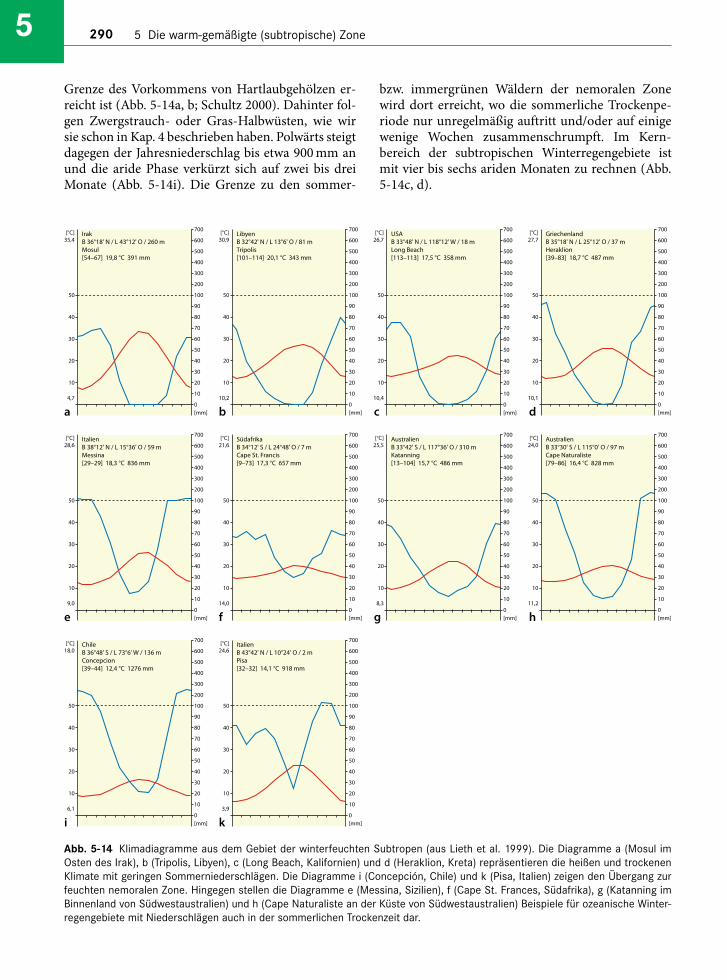
Grenze des Vorkommens von Hartlaubgehölzen er -reicht ist (Abb. 5-14a, b; Schultz 2000). Dahinter fol-gen Zwergstrauch- oder Gras-Halbwüsten, wie wirsie schon in Kap. 4 beschrieben haben. Polwärts steigtdagegen der Jahresniederschlag bis etwa 900 mm anund die aride Phase verkürzt sich auf zwei bis dreiMonate (Abb. 5-14i). Die Grenze zu den sommer-
290 5 Die warm-gemäßigte (subtropische) Zone5
bzw. immergrünen Wäldern der nemoralen Zonewird dort erreicht, wo die sommerliche Trockenpe-riode nur unregelmäßig auftritt und/oder auf einigewenige Wochen zusammenschrumpft. Im Kern-bereich der subtropischen Winterregengebiete ist mit vier bis sechs ariden Monaten zu rechnen (Abb.5-14c, d).
IrakB 36°18' N / L 43°12' O / 260 mMosul[54–67] 19,8 °C 391 mm
700
600
500
400
300
200
10050
[°C]35,4
4,7
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
LibyenB 32°42' N / L 13°6' O / 81 mTripolis[101–114] 20,1 °C 343 mm
700
600
500
400
300
200
10050
[°C]30,9
10,2
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
USAB 33°48' N / L 118°12' W / 18 mLong Beach[113–113] 17,5 °C 358 mm
700
600
500
400
300
200
10050
[°C]26,7
10,4
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
GriechenlandB 35°18' N / L 25°12' O / 37 mHeraklion[39–83] 18,7 °C 487 mm
700
600
500
400
300
200
10050
[°C]27,7
10,1
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
ItalienB 38°12' N / L 15°36' O / 59 mMessina[29–29] 18,3 °C 836 mm
700
600
500
400
300
200
10050
[°C]28,6
9,0
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
SüdafrikaB 34°12' S / L 24°48' O / 7 mCape St. Francis[9–73] 17,3 °C 657 mm
700
600
500
400
300
200
10050
[°C]21,6
14,0
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
AustralienB 33°42' S / L 117°36' O / 310 mKatanning[13–104] 15,7 °C 486 mm
700
600
500
400
300
200
10050
[°C]25,5
8,3
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
a b
ChileB 36°48' S / L 73°6' W / 136 mConcepcion[39–44] 12,4 °C 1276 mm
700
600
500
400
300
200
10050
[°C]18,0
6,1
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
ItalienB 43°42' N / L 10°24' O / 2 mPisa[32–32] 14,1 °C 918 mm
700
600
500
400
300
200
10050
[°C]24,6
3,9
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]i k
c d
e f g h
AustralienB 33°30' S / L 115°0' O / 97 mCape Naturaliste[79–86] 16,4 °C 828 mm
700
600
500
400
300
200
10050
[°C]24,0
11,2
40
30
20
10
90
80
70
60
50
40
30
20
10
0[mm]
Abb. 5-14 Klimadiagramme aus dem Gebiet der winterfeuchten Subtropen (aus Lieth et al. 1999). Die Diagramme a (Mosul imOsten des Irak), b (Tripolis, Libyen), c (Long Beach, Kalifornien) und d (Heraklion, Kreta) repräsentieren die heißen und trockenenKlimate mit geringen Sommerniederschlägen. Die Diagramme i (Concepción, Chile) und k (Pisa, Italien) zeigen den Übergang zurfeuchten nemoralen Zone. Hingegen stellen die Diagramme e (Messina, Sizilien), f (Cape St. Frances, Südafrika), g (Katanning imBinnenland von Südwestaustralien) und h (Cape Naturaliste an der Küste von Südwestaustralien) Beispiele für ozeanische Winter-regengebiete mit Niederschlägen auch in der sommerlichen Trockenzeit dar.
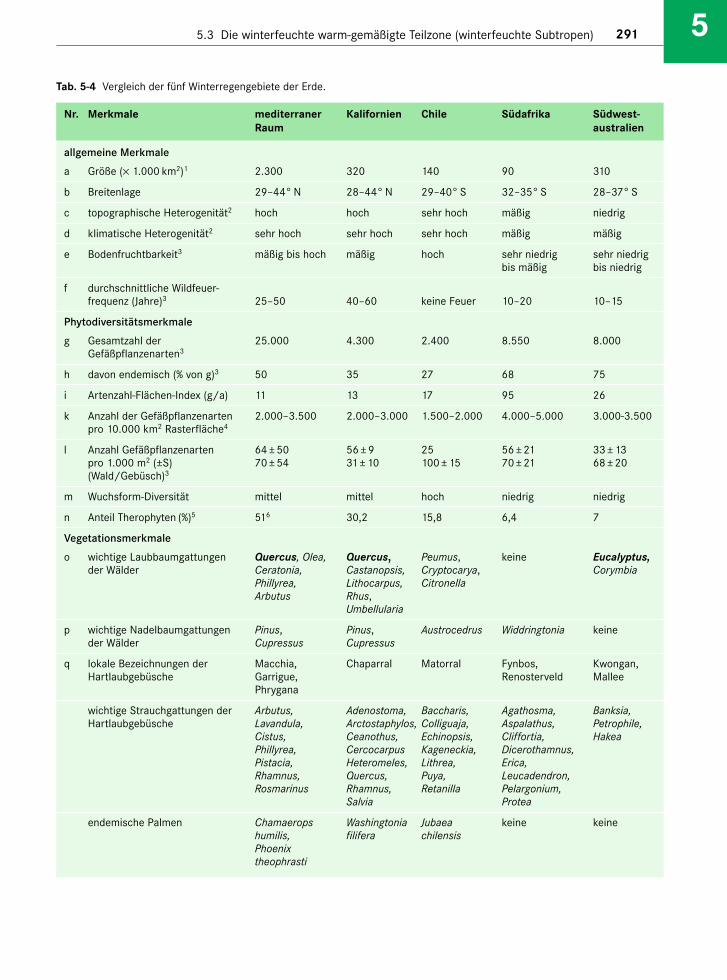
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 291
Tab. 5-4 Vergleich der fünf Winterregengebiete der Erde.
Nr. Merkmale mediterraner Kalifornien Chile Südafrika Südwest-Raum australien
allgemeine Merkmale
a Größe (¥ 1.000 km2)1 2.300 320 140 90 310
b Breitenlage 29–44° N 28–44° N 29–40° S 32–35° S 28–37° S
c topographische Heterogenität2 hoch hoch sehr hoch mäßig niedrig
d klimatische Heterogenität2 sehr hoch sehr hoch sehr hoch mäßig mäßig
e Bodenfruchtbarkeit3 mäßig bis hoch mäßig hoch sehr niedrig sehr niedrig bis mäßig bis niedrig
f durchschnittliche Wildfeuer-frequenz (Jahre)3 25–50 40–60 keine Feuer 10–20 10–15
Phytodiversitätsmerkmale
g Gesamtzahl der 25.000 4.300 2.400 8.550 8.000Gefäßpflanzenarten3
h davon endemisch (% von g)3 50 35 27 68 75
i Artenzahl-Flächen-Index (g/a) 11 13 17 95 26
k Anzahl der Gefäßpflanzenarten 2.000–3.500 2.000–3.000 1.500–2.000 4.000–5.000 3.000-3.500pro 10.000 km2 Rasterfläche4
l Anzahl Gefäßpflanzenarten 64 ± 50 56 ± 9 25 56 ± 21 33 ± 13pro 1.000 m2 (±S) 70 ± 54 31 ± 10 100 ± 15 70 ± 21 68 ± 20(Wald/Gebüsch)3
m Wuchsform-Diversität mittel mittel hoch niedrig niedrig
n Anteil Therophyten (%)5 516 30,2 15,8 6,4 7
Vegetationsmerkmale
o wichtige Laubbaumgattungen Quercus, Olea, Quercus, Peumus, keine Eucalyptus,der Wälder Ceratonia, Castanopsis, Cryptocarya, Corymbia
Phillyrea, Lithocarpus, CitronellaArbutus Rhus,
Umbellularia
p wichtige Nadelbaumgattungen Pinus, Pinus, Austrocedrus Widdringtonia keineder Wälder Cupressus Cupressus
q lokale Bezeichnungen der Macchia, Chaparral Matorral Fynbos, Kwongan, Hartlaubgebüsche Garrigue, Renosterveld Mallee
Phrygana
wichtige Strauchgattungen der Arbutus, Adenostoma, Baccharis, Agathosma, Banksia, Hartlaubgebüsche Lavandula, Arctostaphylos, Colliguaja, Aspalathus, Petrophile,
Cistus, Ceanothus, Echinopsis, Cliffortia, HakeaPhillyrea, Cercocarpus Kageneckia, Dicerothamnus, Pistacia, Heteromeles, Lithrea, Erica,Rhamnus, Quercus, Puya, Leucadendron,Rosmarinus Rhamnus, Retanilla Pelargonium,
Salvia Protea
endemische Palmen Chamaerops Washingtonia Jubaea keine keinehumilis, filifera chilensisPhoenix theophrasti
Abgesehen von der ostmediterranen Teilzone,dem nordafrikanischen Raum und Kalifornien, wodie Sommer sehr heiß werden und kaum Regen fällt(Abb. 5-14a–d), zeigt der Temperatur- und Nieder-schlagsverlauf eher ozeanische Merkmale: Die Mit -teltemperatur des wärmsten Monats übersteigt selten20–25 °C, diejenige des kältesten Monats liegt bei 7–13 °C. Längere (mehrwöchige) Frostperioden feh-len. Auch sind die Sommermonate nicht gänzlichniederschlagsfrei; im Übergang zur feuchten nemo-ralen Zone (Concepción an der Südgrenze des chile-nischen Winterregengebiets; Abb. 5-14i), vor allemaber in der Kapregion Südafrikas (Abb. 5-14f) und inSüdwestaustralien (Abb. 5-14g, h) regnet es auch imSommer regelmäßig. Nach einer Analyse von Cow-ling et al. (2005) sind hier die Regenereignisse gleich-mäßiger über das Jahr verteilt und die Variabilitätzwischen den einzelnen Jahren ist geringer als in denanderen Winterregengebieten. Im mediterranenRaum und in Kalifornien regnet es dagegen seltener,dafür aber heftiger, und die Unterschiede zwischenden Jahren sind größer. Wir werden später sehen,dass ein verlässliches Niederschlagsregime (rainfallreliability) eine (von mehreren) Ursachen für diehohe Phytodiversität der australischen und südafri-kanischen Winterregengebiete ist.
Nach Zech et al. (2014) sind die maßgeblichenProzesse der Bodengenese in den winterfeuchtenSubtropen vom Wechsel zwischen winterlicherRegen- und sommerlicher Trockenzeit bestimmt.Entkalkung und Anreicherung von Residualton inden feuchten Wintermonaten sind in Landschaftenmit vorherrschenden Karbonatgesteinen wie immediterranen Raum weit verbreitet. Hinzu kommt,dass im Winter entlang von Grobporen (z. B. som-
merlichen Trockenrissen) Ton mechanisch nachunten verlagert wird, sodass Tonanreicherungs- (Bt-)Horizonte entstehen. Während der trocken-heißenWitterungsbedingungen im Sommer entwickelt sichaus den durch Verwitterung des Gesteins gebildetenhydratisierten Eisenoxiden der rot gefärbte Hämatit(„Rubefizierung“). Ältere Landoberflächen, derenBodenbildung ins Spättertiär zurückreicht, zeigenhohe Anteile von Kaolinit, die sonst eher für dieBöden der immerfeuchten Tropen und Subtropencharakteristisch sind.
Als Ergebnis von Tonbildung, Tonverlagerungund Rubefizierung dominieren deshalb intensiv rotgefärbte, mehr oder minder tiefgründige, gut durch-wurzelbare Böden mit reichem Bodenleben währenddes Winters und pH-Werten von 5–7. Sie werden imMittelmeergebiet traditionell als Terra rossa bzw.Terra fusca bezeichnet und sind heute als ChromicCambisol (ohne Bt-Horizont) und Chromic Luvisol(mit Bt-Horizont) klassifiziert. Beide Böden sind inallen fünf Winterregengebieten zonal verbreitet und(bei guter Nährstoffversorgung) ackerbaulich nutz-bar; in Südafrika und Australien, wo die Böden auspräkambrischen und früh-paläozoischen P-armenGe steinen alter Landoberflächen entstanden, sind sieallerdings sehr nährstoffarm und sauer. Daneben gibtes eine Vielzahl azonaler Böden, unter denen beispiels-weise Arenosole aus küstennahen Sandablagerungensowie Salzböden (wie in Süd- und Südwestaustralien)zu nennen sind. Vor allem im Mittelmeergebiet sindaußerdem Leptosole in Hang- und Kuppenlagen weitverbreitet; sie sind vielfach das Ergebnis mensch-licher Übernutzung, die schon in der Antike zu groß-räumiger Bodenerosion geführt hat.
292 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 5-4 (Fortsetzung)
Nr. Merkmale mediterraner Kalifornien Chile Südafrika Südwest-Raum australien
Landnutzung
r Besiedlung durch Europäer >10.000 200 450 350 120(Jahre BP)2
s Beginn des Getreideanbaus ca. 8.000 140 450 250 100und der Viehhaltung2
1 Wegen der unterschiedlich vorgenommenen Abgrenzung der Winterregengebiete variieren die Flächenangaben in der Literatur beträchtlich. Schlägtman beispielsweise dem mediterranen Raum auch die Übergänge zur nemoralen Zone („submediterran“ bzw. „supramediterran“ in den Gebirgen) undzu den tropisch-subtropischen Trockengebieten (Gras-Halbwüsten = „mediterrane Steppen“) zu, dann erhöht sich dessen Flächengröße auf etwa 3,5 Mio. km2 und entspricht damit der Größe der mediterranen Florenregion. Wir beschränken uns hier auf die thermo- und mesomediterrane Zone(s. Abschn. 5.3.2.2), das sind dann etwa 2,3 Mio. km2. Daten nach Rebelo et al. 2006 (Südafrika), übrige Gebiete nach Cowling et al. (1996). 2 nachGrove & Rackham (2001) sowie Fox & Fox (1986), 3 nach Cowling et al. (1996), Hopper (1992) sowie Keeley & Swift (1995); 4 nach Barthlott et al.(2007), 5 nach Arroyo et al. (1995), 6 Daten aus Israel; bezogen auf den ganzen mediterranen Raum dürfte der Anteil bei etwa 30 % liegen; 7 nach Fox& Fox (1986).

5.3.1.3 Flora und Phytodiversität
Die Anzahl der Gefäßpflanzenarten ist trotz dergeringen Flächengröße der winterfeuchten Subtro-pen beachtlich hoch (Zeilen g–l in Tab. 5-4). Die fünfWinterregengebiete beherbergen auf nur 2 % derLandoberfläche rund 20 % aller bisher bekanntenTracheophyten (Cowling et al. 1996). Aufgrund sei-ner Größe von 2,3 Mio. km2 (= 73 % der gesamtenKlimazone) und des von Hochgebirgen bestimmtenReliefs enthält der mediterrane Raum mit rund25.000 Gefäßpflanzenarten die meisten Spezies, ge -folgt von der Kapregion (mit lediglich 0,09 Mio. km2
das kleinste Gebiet) und Südwestaustralien mit je -weils 8.000 und mehr (davon über 60 % endemisch).Bezogen auf die gleiche Flächeneinheit (100 × 100km2 Rasterfläche, Barthlott et al. 2007) liegen dieArtenzahlen allerdings mindestens um die Hälfteunter denjenigen der feuchttropischen Hotspots derPhytodiversität (s. Tab. 1-7), nämlich zwischen 2.000(Chile) und 5.000 (Kapland) pro Rasterfläche.Immerhin übersteigen sie diejenigen der benachbar-ten Formationen in der nemoralen Zone und dentropisch-subtropischen Trockengebieten um mehrals die Hälfte, sodass man entlang des Phytodiversi-tätsgradienten vom Äquator zu den Polen neben denfeuchten Tropen von einem zweiten Maximum desArtenreichtums sprechen kann. Auch makroskaligvariiert die Phytodiversität beträchtlich, wie sich beieinem Vergleich der Artenzahlen auf 1.000 m2 zeigt(Tab. 5-4). Am artenreichsten sind die offenen, regel-mäßig von Feuer heimgesuchten Gebüsche inAustralien und Südafrika sowie die beweideten Ge -hölzformationen des ostmediterranen Raumes, amartenärmsten der dichte Chaparral Kaliforniens unddie Wälder (z. B. die Eucalyptus-Wälder Südwestaus-traliens).
Diese vergleichsweise hohen Artenzahlen in denwinterfeuchten Subtropen sind darauf zurückzufüh-ren, dass mehrwöchige Frostperioden wie in dernemoralen Zone und lange Trockenzeiten wie in dentropisch-subtropischen Trockengebieten fehlen. Ther-mischer und hygrischer Stress sind also gegenüberden benachbarten Klimazonen reduziert. Darüberhinaus beeinflussen aber auch Geodiversität, Feuer,Klimageschichte und Isolation die Phytodiversitätder fünf Winterregengebiete auf unterschiedliche,noch immer nicht gänzlich geklärte Weise (Cowlinget al. 1996, Rundel 1998, Médail 2009; Tab. 5-4): 1. Das mediterrane, das kalifornische und das chile-
nische Winterregengebiet sind von jungen Falten-gebirgen und Vulkanismus geprägt. Dadurch er -höhen sich die räumliche (topographische) und
klimatische Vielfalt (Geodiversität). In allen dreiRegionen folgt die Vegetation einem Höhengra-dienten; extrazonal findet man sommergrüneWälder in den Hochlagen und laurophylle Wälderin feuchten Taleinschnitten. Es wäre also zu erwar-ten, dass auf mittlerer Maßstabsebene (mesoska-lig) besonders viele Arten vorkommen (Zeilen kund l in Tab. 5-4). Dies ist der Fall im Mittelmeer-gebiet und in Kalifornien, nicht aber in Chile (mitlediglich 2.500 Gefäßpflanzenarten pro Rasterflä-che). Stattdessen sind die Kapregion und Südwest-australien am artenreichsten. Dort konzentrierensich die winterfeuchten Subtropen auf mäßig relie-fierte Landschaften ohne ausgeprägte Höhengra-dienten; Hochgebirge fehlen. Die hohe Phytodi-versität mit dem größten Endemitenanteil mussalso in diesen beiden Gebieten andere Ursachenhaben.
2. Beide Gebiete zeichnen sich durch alte Landober-flächen mit extrem nährstoffarmen Böden aus.Nährstoffmangel kann vor allem in Offenland-schaften, also außerhalb von Wäldern, Koexistenzvon Pflanzen fördern, sodass unter solchen Bedin-gungen mehr Arten pro Flächeneinheit gedeihenkönnen als unter günstigeren Nährstoffbedingun-gen (Huston 1994). Das gilt besonders dort, wo ein einigermaßen kontinuierliches Klimaregimeüber Jahrmillionen hinweg die Bildung speziellerPflanzenfunktionstypen ermöglichte. Auf derSüdhalbkugel und in Kalifornien haben sich diepleistozänen Klimaschwankungen weniger bemerk-bar ge macht als im mediterranen Raum, der wäh-rend der Kaltzeiten kühler und trockener war alsdie übrigen Winterregengebiete. In Südafrika undin Australien ist ein solches verlässliches Nieder-schlagsregime (rainfall reliability) in gleicherWeise wie in den tropischen Tieflandregenwäl-dern vermutlich mitverantwortlich für die Bil-dung zahlreicher Sippen ähnlicher ökologischerAnsprüche (Cowling et al. 2005). Es beschleunigtdie Entwicklung neuer Arten und senkt die Aus-sterberaten. Außerdem erleichtern schwache, aberregelmäßig auftretende Regen vor allem im Spät-sommer und Herbst, der Zeit der meisten Brände,die Keimung der Samen und Etablierung derJungpflanzen auf den frisch abgebrannten Flä-chen.
3. Hier zeigt sich, dass auch Feuer eine treibendeKraft hoher Artendiversität in den winterfeuchtenSubtropen sein kann. Sowohl Kalifornien als auchdas südafrikanische und australische Winter -regengebiet werden regelmäßig von Brändenheimgesucht, deren Abstand in den artenreichen
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 293

Hartlaubgebüschen Afrikas (Fynbos und Renos-terveld) und Australiens (Kwongan) mit zehn bis20 Jahren geringer ist als in Kalifornien und imMittelmeergebiet (40 bis 50 Jahre; s. Tab. 5-4), abergrößer als z. B. in den meisten Savannen. Bei einersolchen „mittleren“ Brandfrequenz in Fynbos undKwongan können mehr Pflanzen mit unterschied-lichen Regenerations- und Etablierungsmechanis-men gedeihen als dort, wo die Feuerereignissehäufiger oder seltener sind. Außerdem verhinderndie nährstoffarmen Böden ein schnelles Zuwach-sen der feuerbedingten Lücken und ermöglichendie Koexistenz einer Fülle von Pflanzenarten mitähnlichen Wuchsformen, wie man beispielsweiseim Fynbos Südafrikas gut beobachten kann (Cow-ling et al. 1997a): Solche Lücken werden vonEphemeren und kurzlebigen Gehölzen genutzt,die sich nach Feuer aus unterirdischen Organenoder Samen vorübergehend etablieren. In derKapregion hat die überproportionale Radiationverschiedener Gehölzgattungen zu einer enormenArtenfülle geführt, wie im Fall der Gattung Ericamit 657 Arten, von denen 97 % endemisch sind(Goldblatt & Manning 2002).
4. Die Unterschiede in der Artenausstattung der fünfWinterregengebiete sind ein Ergebnis der Floren-und Vegetationsgeschichte (Axelrod 1973, 1975).Paläoökologisch lässt sich nachweisen, dass dieheute dominierenden sklerophyllen Gehölze größ-tenteils mitteltertiären Ursprungs sind, also keinerezente phylogenetische Antwort auf die zuneh-mende Sommertrockenheit am Ende des Tertiärsund im Pleistozän darstellen. Sie waren Bestand-teil der überwiegend immergrünen (größtenteilslaurophyllen) Wälder im Miozän, die physiogno-misch den heutigen Lorbeerwäldern der immer-feuchten Subtropen ähnelten. Der Heterogenitätder Standorte entsprechend enthielten diese Wäl-der bereits immergrüne hartlaubige und sommer-grüne (weichlaubige) Bäume und Sträucher, vondenen sich Erstere mit zunehmender Sommertro-ckenheit im ausgehenden Pliozän und im Pleisto-zän zuungunsten der laurophyllen Taxa durch-setzten. Die Laurophyllen überlebten in feuchtenBachschluchten und stellenweise (wie in Chile) imÜbergang zur hochozeanischen nemoralen Zone,die Sommergrünen in den höheren Gebirgslagen. Zu dieser „vor-pliozänen“ Artengruppe gehörenGehölze, denen außer der Sklerophyllie eine Reiheweiterer Eigenschaften (phylogenetisch „alte“Merkmale) gemeinsam ist (Herrera 1992, Verdú etal. 2003; „Konvergenz“; s. Abschn. 5.3.1.4): Siesind in der Lage, nach Beschädigung durch Tier-
fraß, Windwurf oder Feuer rasch aus dem Stockauszuschlagen (resprouter), tragen fleischige,großsamige und zoochore Früchte und verhaltensich wie Klimaxbaumarten. Beispiele aus demMittelmeergebiet sind die Gattungen Quercus,Arbutus und Olea. Daneben gibt es eine physio-gnomisch weniger einheitliche „nach-pliozäneArtengruppe“, die sich während und nach der Eta-blierung des Winterregenklimas entwickelt hat.Ihre Vertreter sind entweder aus benachbartenVegetationszonen eingewandert (wie Heteromelesin Kalifornien aus der kühl-gemäßigten Zone)oder hatten genetisch flexible Vorfahren in denspättertiären Wäldern (Ackerly 2009). Es handeltsich um niedrigwüchsige, malakophylle Sträucher,die mit ihren kleinen, anemochoren Samen in derLage sind, offene Flächen nach Feuer oder ande-ren Störungen rasch zu besiedeln (reseeder). Bei-spiele sind die Gattungen Calicotome (Fabaceae),Cistus und Lavandula im Mittelmeergebiet (Blon-del & Aronson 1995).
5. Bei einem Vergleich der Flora der fünf Winterre-gengebiete fällt auf, dass die floristische Ähn-lichkeit untereinander begrenzt ist. Denn ihreFlorenausstattung ist vom jeweiligen regionalenArtenpool geprägt, aus dem sich die heutige Vege-tation zusammensetzt (Raven 1973). DieserArtenpool ist im mediterranen Raum überwie-gend holarktisch mit einigen wenigen tropischenElementen (wie Ceratonia, Chamaerops, Laurus,Myrtus und Olea), ebenso in Kalifornien (mit alt-tertiären Pflanzen semiarider Gebiete wie Arcto -staphylos und Ceanothus). Holarktische Florenele-mente wie Quercus und Pinus verbinden diebeiden nordhemisphärischen Winterregengebiete;diese Übereinstimmung ist der kreidezeitlichenund tertiären Nähe der beiden Kontinente Nord-amerika und Eurasien geschuldet. Die Flora vonMittelchile ist dagegen fast ausschließlich neotro-pisch. In Afrika und Australien prägen Vertreterder alten Gondwana-Flora (vor allem Proteaceaeund Restionaceae) das Bild der Vegetation, diesich unter den Bedingungen eines kontinuier-lichen Klimas und nährstoffarmer Böden be son -ders reich entfalten konnte. Floristische Ähnlich-keiten mit Chile oder der mediterranen Flora sindnicht zu erkennen.
5.3.1.4 Sklerophyllie und Konvergenz
Ein allen Winterregengebieten gemeinsames Merk-mal ist die Sklerophyllie (Hartlaubigkeit) vieler Ge -
294 5 Die warm-gemäßigte (subtropische) Zone5

hölze. Allerdings ist das Spektrum morphologischer,anatomischer und funktionaler Blatteigenschaftensehr groß. Die Assimilationsorgane können kleinund schmal (wie bei Olea europaea) oder breit undlorbeerartig sein (wie bei Arbutus unedo). Es gibtSträucher und Bäume mit nadel- (erikoiden) undschuppenförmigen (cupressoiden) Blättern. Ersterekommen nicht nur bei der Gattung Erica, sondernauch unter Proteaceae, Asteraceae und Thymelaea-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 295
ceae vor, Letztere trägt z. B. der Renosterbos Dicero-thamnus rhinocerotis (Asteraceae) in der Kapregion.Darüber hinaus gibt es immergrüne malakophylleGehölze wie die Cistus-Arten, die Winter- und Som-merblätter erzeugen und diese abwerfen können,wenn die Wasserversorgung nicht mehr ausreicht(Abb. 5-15).
In Abschn. 5.3.1.3 wurde darauf hingewiesen, dasssklerophylle Gehölze bereits im Mitteltertiär als
a c d
e g h
b
f
i
Abb. 5-15 Beispiele für Wuchsformen der Winterregengebiete. a = Quercus ilex, Fagaceae(sklerophyll; Mallorca, Spanien); b = Arbutus unedo, Ericaceae (sklerophyll-großblättrig; Mal-lorca, Spanien); c = Banksia coccinea, Proteaceae (proteoider Typ des sklerophyllen Blattes mitSpaltöffnungen auf beiden Blattseiten; Stirling Range, Australien); d = Andersonia echinoce-phala, Ericaceae (sklerophyll-kleinblättrig; Stirling Range, Australien); e = Erica sp., Ericaceae(sklerophyll-erikoid; Kapregion, Südafrika); f = Cistus albidus, Cistaceae (malakophyll; Mallorca,Spanien); g = Retanilla ephedra, Rhamnaceae (Rutenstrauch; La Campana, Chile); h = Asphode-lus ramosus, Asphodelaceae (Geophyt; Kreta); i = Lagurus ovatus, Poaceae (Therophyt; Italien).
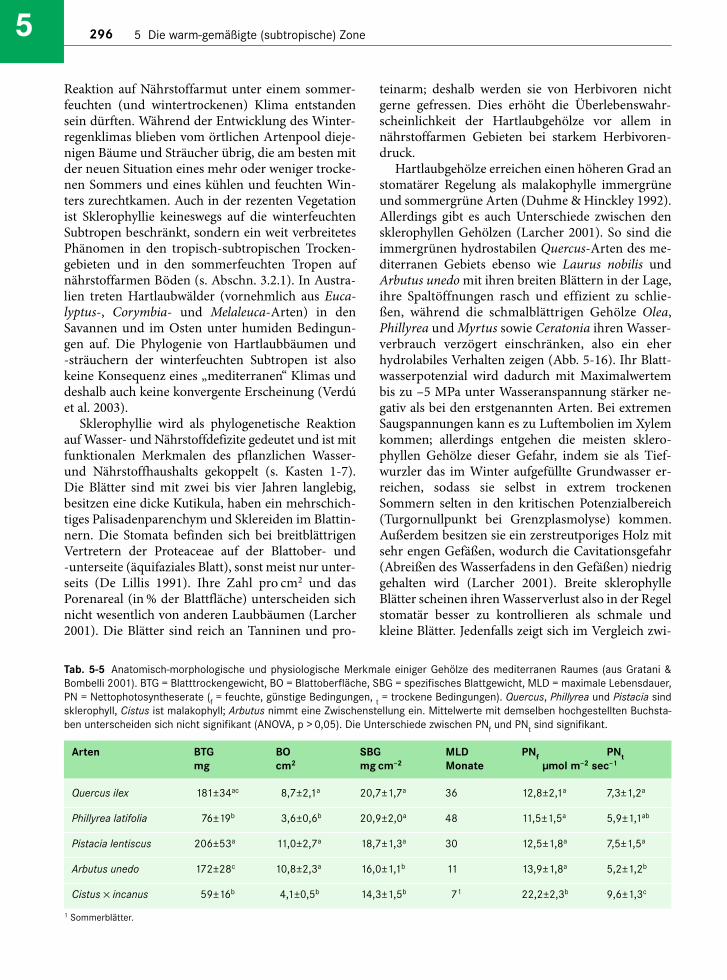
Reaktion auf Nährstoffarmut unter einem sommer-feuchten (und wintertrockenen) Klima entstandensein dürften. Während der Entwicklung des Winter-regenklimas blieben vom örtlichen Artenpool dieje-nigen Bäume und Sträucher übrig, die am besten mitder neuen Situation eines mehr oder weniger trocke-nen Sommers und eines kühlen und feuchten Win-ters zurechtkamen. Auch in der rezenten Vegetationist Sklerophyllie keineswegs auf die winterfeuchtenSubtropen beschränkt, sondern ein weit verbreitetesPhänomen in den tropisch-subtropischen Trocken-gebieten und in den sommerfeuchten Tropen aufnährstoffarmen Böden (s. Abschn. 3.2.1). In Austra-lien treten Hartlaubwälder (vornehmlich aus Euca-lyptus-, Corymbia- und Melaleuca-Arten) in denSavannen und im Osten unter humiden Bedingun-gen auf. Die Phylogenie von Hartlaubbäumen und -sträuchern der winterfeuchten Subtropen ist alsokeine Konsequenz eines „mediterranen“ Klimas unddeshalb auch keine konvergente Erscheinung (Verdúet al. 2003).
Sklerophyllie wird als phylogenetische Reaktionauf Wasser- und Nährstoffdefizite gedeutet und ist mitfunktionalen Merkmalen des pflanzlichen Wasser-und Nährstoffhaushalts gekoppelt (s. Kasten 1-7).Die Blätter sind mit zwei bis vier Jahren langlebig,besitzen eine dicke Kutikula, haben ein mehrschich-tiges Palisadenparenchym und Sklereiden im Blattin-nern. Die Stomata befinden sich bei breitblättrigenVertretern der Proteaceae auf der Blattober- und -unterseite (äquifaziales Blatt), sonst meist nur unter-seits (De Lillis 1991). Ihre Zahl pro cm2 und dasPoren areal (in % der Blattfläche) unterscheiden sichnicht wesentlich von anderen Laubbäumen (Larcher2001). Die Blätter sind reich an Tanninen und pro-
teinarm; deshalb werden sie von Herbivoren nichtgerne gefressen. Dies erhöht die Überlebenswahr-scheinlichkeit der Hartlaubgehölze vor allem innährstoffarmen Gebieten bei starkem Herbivoren-druck.
Hartlaubgehölze erreichen einen höheren Grad anstomatärer Regelung als malakophylle immergrüneund sommergrüne Arten (Duhme & Hinckley 1992).Allerdings gibt es auch Unterschiede zwischen densklerophyllen Gehölzen (Larcher 2001). So sind dieimmergrünen hydrostabilen Quercus-Arten des me -di terranen Gebiets ebenso wie Laurus nobilis undArbutus unedo mit ihren breiten Blättern in der Lage,ihre Spaltöffnungen rasch und effizient zu schlie-ßen, während die schmalblättrigen Gehölze Olea,Phillyrea und Myrtus sowie Ceratonia ihren Wasser-verbrauch verzögert einschränken, also ein eherhydrolabiles Verhalten zeigen (Abb. 5-16). Ihr Blatt-wasserpotenzial wird dadurch mit Maximalwertembis zu –5 MPa unter Wasseranspannung stärker ne -gativ als bei den erstgenannten Arten. Bei extremenSaugspannungen kann es zu Luftembolien im Xylemkommen; allerdings entgehen die meisten sklero-phyllen Gehölze dieser Gefahr, indem sie als Tief-wurzler das im Winter aufgefüllte Grundwasser er -reichen, sodass sie selbst in extrem trockenenSommern selten in den kritischen Potenzialbereich(Turgornullpunkt bei Grenzplasmolyse) kommen.Außerdem besitzen sie ein zerstreutporiges Holz mitsehr engen Gefäßen, wodurch die Cavitationsgefahr(Abreißen des Wasserfadens in den Gefäßen) niedriggehalten wird (Larcher 2001). Breite sklerophylleBlätter scheinen ihren Wasserverlust also in der Regelstomatär besser zu kontrollieren als schmale undkleine Blätter. Jedenfalls zeigt sich im Vergleich zwi-
296 5 Die warm-gemäßigte (subtropische) Zone5
Tab. 5-5 Anatomisch-morphologische und physiologische Merkmale einiger Gehölze des mediterranen Raumes (aus Gratani &Bombelli 2001). BTG = Blatttrockengewicht, BO = Blattoberfläche, SBG = spezifisches Blattgewicht, MLD = maximale Lebensdauer,PN = Nettophotosyntheserate (f = feuchte, günstige Bedingungen, t = trockene Bedingungen). Quercus, Phillyrea und Pistacia sindsklerophyll, Cistus ist malakophyll; Arbutus nimmt eine Zwischenstellung ein. Mittelwerte mit demselben hochgestellten Buchsta-ben unterscheiden sich nicht signifikant (ANOVA, p > 0,05). Die Unterschiede zwischen PNf und PNt sind signifikant.
Arten BTG BO SBG MLD PNf PNtmg cm2 mg cm–2 Monate μmol m–2 sec–1
Quercus ilex 181±34ac 8,7±2,1a 20,7±1,7a 36 12,8±2,1a 7,3±1,2a
Phillyrea latifolia 76±19b 3,6±0,6b 20,9±2,0a 48 11,5±1,5a 5,9±1,1ab
Pistacia lentiscus 206±53a 11,0±2,7a 18,7±1,3a 30 12,5±1,8a 7,5±1,5a
Arbutus unedo 172±28c 10,8±2,3a 16,0±1,1b 11 13,9±1,8a 5,2±1,2b
Cistus × incanus 59±16b 4,1±0,5b 14,3±1,5b 71 22,2±2,3b 9,6±1,3c
1 Sommerblätter.
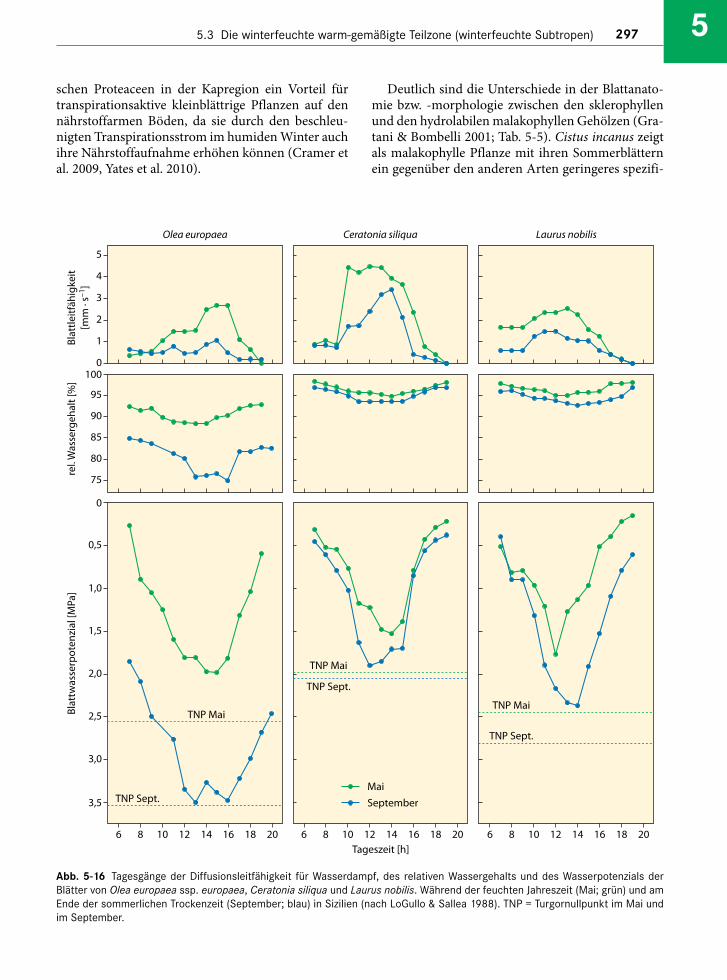
schen Proteaceen in der Kapregion ein Vorteil fürtranspirationsaktive kleinblättrige Pflanzen auf dennährstoffarmen Böden, da sie durch den beschleu-nigten Transpirationsstrom im humiden Winter auchihre Nährstoffaufnahme erhöhen können (Cramer etal. 2009, Yates et al. 2010).
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 297
Deutlich sind die Unterschiede in der Blattanato-mie bzw. -morphologie zwischen den sklerophyllenund den hydrolabilen malakophyllen Gehölzen (Gra- tani & Bombelli 2001; Tab. 5-5). Cistus incanus zeigtals malakophylle Pflanze mit ihren Sommerblätternein gegenüber den anderen Arten geringeres spezifi-
5
4
3
2
1
0
Blat
tleitf
ähig
keit
[mm
· s–1
]
Olea europaea Ceratonia siliqua Laurus nobilis
100
95
90
85
80
75
rel.
Was
serg
ehal
t [%
]
0
1,0
0,5
1,5
2,0
2,5
3,0
3,5
6 8 10 12 14 16 18 20
Blat
twas
serp
oten
zial
[MPa
]
TNP Mai
TNP Sept.
6 8 10 12Tageszeit [h]
14 16 18 20
TNP Mai
TNP Sept.
6 8 10 12 14 16 18 20
TNP Mai
TNP Sept.
MaiSeptember
Abb. 5-16 Tagesgänge der Diffusionsleitfähigkeit für Wasserdampf, des relativen Wassergehalts und des Wasserpotenzials derBlätter von Olea europaea ssp. europaea, Ceratonia siliqua und Laurus nobilis. Während der feuchten Jahreszeit (Mai; grün) und amEnde der sommerlichen Trockenzeit (September; blau) in Sizilien (nach LoGullo & Sallea 1988). TNP = Turgornullpunkt im Mai undim September.

sches Blattgewicht, übertrifft in der Photosynthese-leistung bei günstigen Feuchtebedingungen die Skle-rophyllen um fast das Doppelte und ist selbst amEnde des trockenen Sommers noch leistungsfähigerals diese. Denn wegen des teilweisen oder gänzlichenSpaltenschlusses verringert sich die C-Akquisitionder Hartlaubgehölze ausgerechnet im Sommer, alsoin der thermisch eigentlich günstigen Jahreszeit. IhreWachstumsraten sind deshalb niedrig, außer wäh-rend der kurzen Perioden im Frühling und imHerbst. Arbutus nimmt eine Zwischenstellung ein.Im Vergleich zu den laurophyllen Bäumen (wie imFall von Persea indica auf Teneriffa; Gonzáles-Rodrí-guez et al. 2002) tendieren die Sklerophyllen desmediterranen Raumes zu einer eher konservativenStrategie; sie transpirieren deutlich weniger, habenmit 200–400 mmol m–2 s–1 eine niedrigere stomatäreLeitfähigkeit (P. indica: 800 mmol m–2 s–1) und mit3–5 μmol CO2 mmol–1 H2O einen weitaus höherenWassernutzungskoeffizienten der Photosynthese alsdie hydrolabileren laurophyllen Gehölze (P. indica:mittleres Maximum 1,5 μmol mmol–1; alle Da tenunter feuchten Bedingungen erhoben).
5.3.2 Vegetation
5.3.2.1 Übersicht
Trotz der geringen Flächengröße gehören die winter-feuchten Subtropen, was Vegetation und Fauna be -trifft, zu den am besten untersuchten Zonen derErde. Außer in den Vegetationsmonographien ein-zelner Kontinente und Länder (z. B. Klötzli et al. 2010für Europa; Hueck 1966 und Veblen et al. 2007 fürSüdamerika; Knapp 1965 und Barbour & Billings2000 für Nordamerika; Knapp 1973, Cowling et al.1997b und Mucina et al. 2006 für Südafrika; Beadle1981und Groves 1994 für Australien) und Lehrbü-chern (z. B. Walter & Breckle 1991, Archibold 1995,Schroeder 1998, Schultz 2000, Pugnaire & Valladares2007, Jørgensen 2009) wird die Ökologie der fünfWinterregengebiete in umfangreichen Sammelbän-den und Monographien dargestellt (Di Castri &Mooney 1973, Di Castri et al. 1981, Kruger et al.1983, Cowling 1992, Arianoutsou & Groves 1994,Arroyo et al. 1995, Davis & Richardson 1995, Moreno& Oechel 1995, Dallman 1998, Rundel et al. 1998,Barbour et al. 2007). Speziell für den mediterranenRaum sei auf Blondel & Aronson (1999), Rodá et al.(1999), Grove & Rackham (2001), Quézel & Médail(2003), Mazzoleni et al. (2004) sowie Thompson(2005) verwiesen.
Die zonale Vegetation der fünf Winterregenge-biete besteht aus Hartlaubwäldern und -gebüschen,in denen sklerophylle Gehölze dominieren. DieUnterschiede bezüglich Artenzusammensetzung undStruktur sind allerdings so groß, dass Zweifel an derEinheitlichkeit und Eigenständigkeit der Formationangebracht sind (Walter & Breckle 1991). Ein unbe-fangener Betrachter dürfte kaum eine physiognomi-sche Ähnlichkeit zwischen den Steineichenwäldernund Macchien des mediterranen Raumes, dem süd-afrikanischen Fynbos und den hochwüchsigen Karri-(Eucalyptus diversicolor-)Wäldern Australiens erken-nen können. Letztere gleichen den Eukalyptusbe -ständen der immerfeuchten Subtropen Südostaus-traliens mehr als den unter demselben Klimavorkommenden Cryptocarya alba-Wäldern in Mit -tel chile.
Außerhalb von Südwestaustralien erreichen dieWälder im mediterranen Raum, in Kalifornien undin Chile im Optimalzustand lediglich 10–20 m Höhe.Wegen ihres immergrünen Charakters haben sie eindichtes Kronendach, das nur wenig Licht in denBestand eindringen lässt. Der Unterwuchs ist deshalbmeist spärlich ausgebildet und besteht aus immer-grünen Kleinsträuchern und Chamaephyten. Nur dieEukalyptuswälder sind wegen der senkrechten Stel-lung der Blätter hell genug für eine höhere und üppi-gere Strauchschicht. Das Wurzelsystem der Bäume istdimorph: Es setzt sich aus einer (10 bis über 20 m)tief reichenden Pfahlwurzel und einem oberflächen-nahen, ausgreifenden lateralen Wurzelgeflecht zu -sam men, sodass die Pflanzen sowohl Grund- alsauch Oberflächenwasser aus spärlichen Niederschlä-gen und abtropfendem Küstennebel aufnehmen kön-nen (Kummerow 1981). Die Pfahlwurzel entstehtsofort nach der Keimung der Samen und wird, wiebei dem chilenischen Hartlaubbaum Quillaja sapo-naria, schon im ersten Jahr der Jungpflanzenent-wicklung 1 m lang (Canadell & Zedler 1995); hat diePflanze den trockenen Sommer überlebt, entstehendie Seitenwurzeln.
Unter hygrisch suboptimalen Bedingungen wer-den die Wälder niedriger und lichter. Dann bildetsich eine bis zu 6 m hohe Strauchschicht, bevor dieBäume schließlich ganz verschwinden und ein Hart-laubgebüsch entsteht (Tab. 5-4). An flachgründigen,sonnseitigen Hängen oder (anthropogen) nach de -gradierender Bodennutzung werden diese Gebüschezu lockeren, rund 1 m hohen heideartigen Bestän-den, in denen malakophylle Kleinsträucher und sol-che mit erikoiden oder cupressoiden Blättern zurDominanz gelangen können. Dann kommen, wie imchilenischen Matorral, sogar Wuchsformen vor, die
298 5 Die warm-gemäßigte (subtropische) Zone5

sonst eher für tropisch-subtropische Trockengebietecharakteristisch sind, nämlich Sukkulente (wie Echi-nopsis chiloensis), Rutensträucher (wie die Rhamna-cee Retanilla ephedra) mit reduzierten Blättern undgrünen Sprossen sowie Pflanzen mit Dornen bzw.Stacheln. Solche Organe zur mechanischen Verteidi-gung gegen Herbivore sind ansonsten in den Hart-laubgebüschen weniger häufig; in Südafrika undAustralien fehlen sie gänzlich. Stattdessen spielt diechemische Verteidigung mittels Tanninen und äthe-rischen Ölen in den Blättern eine größere Rolle(Blondel & Aronson 1995). Vor allem bei den Sklero-phyllen verlängert sich dadurch die Blattlebensdauer.
Das Wurzelsystem der Hartlaubgebüsche ist imGegensatz zu dem der Wälder eher flach und weitausstreichend (Canadell & Zedler 1995). In dennährstoffarmen Böden der Kapregion und Südwest-australiens haben die Proteaceae eine besondere Stra-tegie der Nährstoff-Akquisition entwickelt: Nachdem ersten Regen bilden sich unter der Streuauflage
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 299
unzählige Feinwurzeln, welche die Hauptwurzeln wieein Kokon umgeben und sehr effizient die im Ober-boden freigesetzten Nährstoffe aufnehmen (Lamberset al. 2006; Kasten 5-4).
In diesen Hartlaubgebüschen sind ephemerePflanzen, vor allem Zwiebel- und Knollengeophytenverschiedener monokotyler Familien (wie Iridaceae,Amaryllidaceae) häufig. Viele von ihnen blühen imHerbst und entwickeln anschließend die Blätter imWinter; den Sommer überdauern sie unterirdisch.Therophyten, deren Anteil an der Flora in der Medi-terraneis besonders hoch ist (Zeile n in Tab. 5-4), kei-men überwiegend im Herbst und blühen im Spät-winter oder Frühjahr. Vor allem das Frühjahr stelltpflanzenökologisch eine besonders günstige Periodedar: Die winterliche Feuchtigkeit und die rasch an -steigenden Temperaturen ermöglichen einen Höhe-punkt der Blütenbildung in dieser Jahreszeit nichtnur bei den Ephemeren, sondern auch bei Sträuchernund Bäumen. Eine zweite, wenn auch weniger ausge-
a b
dc
Abb. 5-17 Beispiele für Anpassungen von Pflanzen südhemisphärischer Hartlaubwälder und -gebüsche an regelmäßig wiederkeh-rende Brände (s. auch Abb. 1 in Kasten 3-4). a = resprouter: Berzelia intermedia, Bruniaceae (Fynbos, Südafrika); b = Keimlinge (spa-telförmige Blätter) des reseeders Leucadendron eucalyptifolium, Proteaceae (montaner Fynbos, Südafrika); c = nach Feuer geöff-nete Kapseln von Corymbia calophylla, Myrtaceae, Südwestaustralien (viele Eukalypten und Proteaceen bilden eineKronensamenbank); d = büschelförmiger Laubaustrieb der Kronen von Eucalyptus diversicolor ca. 50 Jahre nach Feuer (Südwest-australien).

prägte Blütezeit ist der Herbst, wenn die ersten Regennach dem trockenen Sommer gefallen sind.
Die Hartlaubgebüsche haben in den Winterregen-gebieten verschiedene Bezeichnungen (Tab. 5-4). Immediterranen Raum sind Macchie (für hohe Gebü-sche) und Garrigue (für niedrige Gebüsche) ge -bräuchlich; Letztere heißen im ostmediterranen RaumPhrygana. In Kalifornien spricht man von Chaparralund in küstennahen Gebieten von Coastal Scrub. DieHartlaubgebüsche in Chile werden Matorral ge -nannt. In der Kapregion unterscheidet man zwischenFynbos (auf nährstoffarmen, sandigen oder kiesigenBöden) und Renosterveld (auf lehmigen Böden), inAustralien zwischen Kwongan (einer von Proteaceen
dominierten Gebüschvegetation) und Mallee (einemEucalyptus-Trockengebüsch, bereits in Abschn.4.3.3.3 besprochen). Außer in Chile, wo natürlicheFeuer nicht vorkommen (s. Abschn. 5.3.2.3), handeltes sich bei den Gebüschen häufig um eine Feuerkli-max. Das gilt besonders für Südafrika und Austra-lien, größtenteils auch für Kalifornien. Im mediterra-nen Raum ist die heutige Verbreitung hingegenüberwiegend auf die jahrtausendelange Tätigkeit desMenschen zurückzuführen (s. aber Abschn. 5.3.2.2).Seit dem Altertum haben Waldrodung, Brände undlandwirtschaftliche Nutzung in erheblichem Maß zurAusbreitung von Macchie und Garrigue beigetragen.Wir haben es also hier entweder mit Degradations-
300 5 Die warm-gemäßigte (subtropische) Zone5
Phosphoraufnahme von Pflanzen auf armen Standorten
sert die Aufnahmeeffizienz der Pflanzen und erlaubt Wachs-tum und Reproduktion derart spezialisierter Arten auf P-limi-tierten Böden.
Wurzelexsudate sind organische Säuren wie Citrat undMalat sowie das Enzym Phosphatase, welche schwer ver-fügbaren (in der organischen Substanz oder in Schwerme-tallkomplexen gebundenen) Phosphor in eine lösliche Formüberführen, sodass die Pflanzen ihn über die Wurzeln auf-nehmen können. Diese Wurzelexsudate sind nur dann wirk-sam, wenn genug Feuchtigkeit im Boden vorhanden ist. InTrockengebieten geben deshalb die oberflächennahen Wur-
Kasten 5-4
In den Winterregengebieten Südafrikas und Australiens sindder gesteinsgebundene Vorrat und die pflanzenverfügbareMenge von Phosphor so gering, dass die Pflanzen spezielleMechanismen entwickelt haben, um ihre P-Versorgungsicherzustellen (Lambers et al. 2006). Abgesehen von denrecht häufigen karnivoren Drosera-Arten tun sie dies aufzwei verschiedenen Wegen, indem sie erstens die geringeMenge an P im Boden durch Wurzelexsudate in eine fürPflanzen leicht verfügbare Form überführen, und zweitensdie Wurzeloberfläche durch Mykorrhizierung und/oder ver-stärkte Bildung von Feinwurzeln vergrößern. Beides verbes-
Abb. 1 Beispiele für Feinwurzel-büschel (root clusters) bei Protea-
ceae und Restionaceae (FotosMichael Shane und Hans Lam-bers). Die Pflanzen wurden im
Gewächshaus in einer Nährlösungunter Phosphormangelbedingun-gen (P-Gehalten ≤ 1 μg) kultiviert(Lambers et al. 2006). a = Bank-
sia prionites (Skalenstrich 13 mm,proteoid roots); b = Hakea pro-
strata (Skalenstrich 4 mm, prote-oid roots). a b
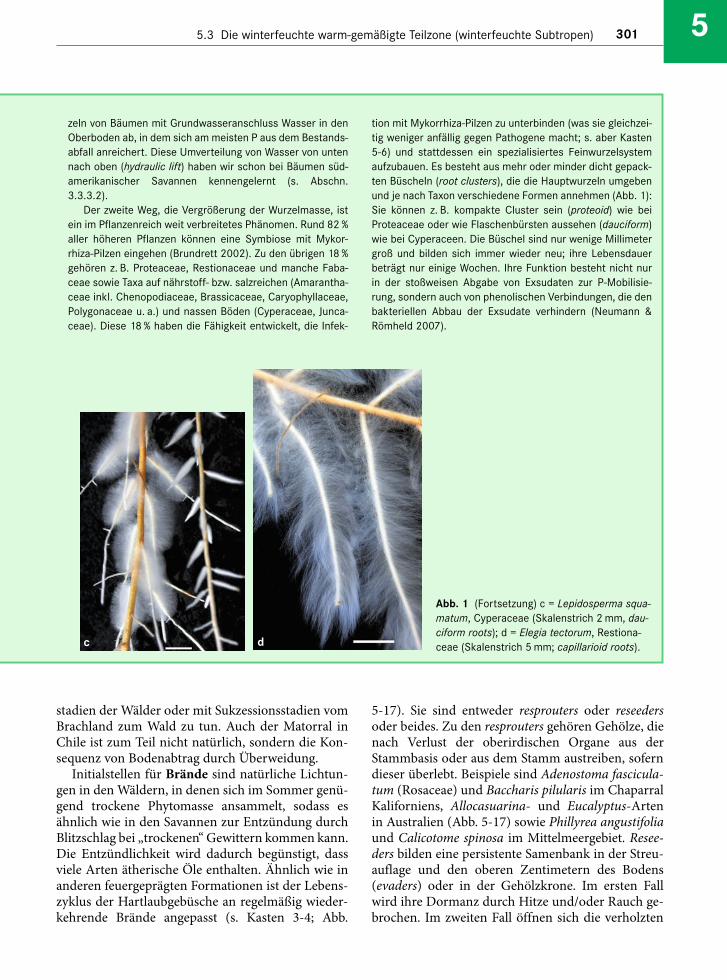
stadien der Wälder oder mit Sukzessionsstadien vomBrachland zum Wald zu tun. Auch der Matorral inChile ist zum Teil nicht natürlich, sondern die Kon-sequenz von Bodenabtrag durch Überweidung.
Initialstellen für Brände sind natürliche Lichtun-gen in den Wäldern, in denen sich im Sommer genü-gend trockene Phytomasse ansammelt, sodass esähnlich wie in den Savannen zur Entzündung durchBlitzschlag bei „trockenen“ Gewittern kommen kann.Die Entzündlichkeit wird dadurch begünstigt, dassviele Arten ätherische Öle enthalten. Ähnlich wie inanderen feuergeprägten Formationen ist der Lebens-zyklus der Hartlaubgebüsche an regelmäßig wieder-kehrende Brände angepasst (s. Kasten 3-4; Abb.
5-17). Sie sind entweder resprouters oder reseedersoder beides. Zu den resprouters gehören Gehölze, dienach Verlust der oberirdischen Organe aus derStammbasis oder aus dem Stamm austreiben, soferndieser überlebt. Beispiele sind Adenostoma fascicula-tum (Rosaceae) und Baccharis pilularis im ChaparralKaliforniens, Allocasuarina- und Eucalyptus-Artenin Australien (Abb. 5-17) sowie Phillyrea angustifoliaund Calicotome spinosa im Mittelmeergebiet. Resee-ders bilden eine persistente Samenbank in der Streu-auflage und den oberen Zentimetern des Bodens(evaders) oder in der Gehölzkrone. Im ersten Fallwird ihre Dormanz durch Hitze und/oder Rauch ge -brochen. Im zweiten Fall öffnen sich die verholzten
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 301
zeln von Bäumen mit Grundwasseranschluss Wasser in denOberboden ab, in dem sich am meisten P aus dem Bestands-abfall anreichert. Diese Umverteilung von Wasser von untennach oben (hydraulic lift) haben wir schon bei Bäumen süd-amerikanischer Savannen kennengelernt (s. Abschn.3.3.3.2).
Der zweite Weg, die Vergrößerung der Wurzelmasse, istein im Pflanzenreich weit verbreitetes Phänomen. Rund 82 %aller höheren Pflanzen können eine Symbiose mit Mykor-rhiza-Pilzen eingehen (Brundrett 2002). Zu den übrigen 18 %gehören z. B. Proteaceae, Restionaceae und manche Faba-ceae sowie Taxa auf nährstoff- bzw. salzreichen (Amarantha-ceae inkl. Chenopodiaceae, Brassicaceae, Caryophyllaceae,Polygonaceae u. a.) und nassen Böden (Cyperaceae, Junca-ceae). Diese 18 % haben die Fähigkeit entwickelt, die Infek-
tion mit Mykorrhiza-Pilzen zu unterbinden (was sie gleichzei-tig weniger anfällig gegen Pathogene macht; s. aber Kasten5-6) und stattdessen ein spezialisiertes Feinwurzelsystemaufzubauen. Es besteht aus mehr oder minder dicht gepack-ten Büscheln (root clusters), die die Hauptwurzeln umgebenund je nach Taxon verschiedene Formen annehmen (Abb. 1):Sie können z. B. kompakte Cluster sein (proteoid) wie beiProteaceae oder wie Flaschenbürsten aussehen (dauciform)wie bei Cyperaceen. Die Büschel sind nur wenige Millimetergroß und bilden sich immer wieder neu; ihre Lebensdauerbeträgt nur einige Wochen. Ihre Funktion besteht nicht nurin der stoßweisen Abgabe von Exsudaten zur P-Mobilisie-rung, sondern auch von phenolischen Verbindungen, die denbakteriellen Abbau der Exsudate verhindern (Neumann &Römheld 2007).
Abb. 1 (Fortsetzung) c = Lepidosperma squa-matum, Cyperaceae (Skalenstrich 2 mm, dau-ciform roots); d = Elegia tectorum, Restiona-ceae (Skalenstrich 5 mm; capillarioid roots).dc

Kapseln des Fruchtstandes erst während des nächst-folgenden Feuers, um die Samen freizusetzen (sero-tiny). Gehölze mit einer Samenbank in der Kronesind besonders häufig in den südafrikanischen undaustralischen Hartlaubgebüschen (wie beispielsweisedie Vertreter der Gattungen Leucadendron und Bank-sia). Das weitgehende Fehlen vieler Merkmale derreseeder-Strategie im mediterranen Raum, vor allembei den Therophyten, ist ein Hinweis auf die – imVergleich zu Kalifornien – geringere Bedeutung vonWildfeuern in der Vergangenheit (Pausas et al. 2006).
Hartlaubwälder und -gebüsche sind nicht die ein-zigen Formationen der winterfeuchten Subtropen.Auf der Nordhemisphäre sind Nadelwälder ausPinus-, Cupressus- und Juniperus-Arten (außerhalbder Gebirge) auf trockenen und flachgründigenStandorten und als Pioniervegetation regional ver-breitet. Sie werden zusammen mit den Hartlaubwäl-dern in den Abschn. 5.3.2.2 und 5.3.2.3 behandelt. Inder montanen Stufe der Gebirge kommen extrazonalsommergrüne (nemorale) Laubwälder (Quercus inKalifornien und im Mittelmeergebiet, Nothofagus inChile) und nemorale Gebirgsnadelwälder vor (z. B.aus Cedrus atlantica im Atlas-Gebirge, aus verschie-den Pinus-, Abies- und Tsuga-Arten sowie dem Mam-mutbaum Sequoiadendron giganteum in der SierraNevada von Kalifornien), die zusammen mit derVegetation oberhalb der Baumgrenze in Abschn. 5.5besprochen werden. Ein integraler Bestandteil derVegetation der winterfeuchten Subtropen sind fernerdie Flussauen und Sümpfe sowie die Küstenvegeta-tion der Marschen und Dünen (s. Abschn. 5.4).
Die winterfeuchten Subtropen sind wegen der lan-gen Sonnenscheindauer, der Lage am Meer (Fische-rei, heute Tourismus) und der klimatischen Eignungfür den Anbau tropischer und temperater Nutzpflan-zen ein bevorzugter Siedlungsraum des Menschen(Schultz 2000). Dies gilt besonders für den mediter-ranen Raum mit seiner Jahrtausende zurückreichen-den Kulturgeschichte. Aber auch in den übrigenWinterregengebieten bestimmen heute Weide- undAckerflächen sowie Aufforstungen weiträumig denLandschaftscharakter. Die anthropogene Vegetationprägt deshalb mehr als in den bisher behandeltenÖkozonen den Landschaftscharakter. Ihre Physio-gnomie und Struktur differiert zwischen den fünfWinterregengebieten erheblich; Gemeinsamkeitensind kaum erkennbar, sieht man vom Auftreten euro-päischer Gräser in Amerika und Australien ab. Des-halb besprechen wir diese synanthropen Formatio-nen nicht wie sonst in einem separaten Abschnitt,sondern zusammen mit den Hartlaubwäldern und -gebüschen der jeweiligen Regionen.
5.3.2.2 Das mediterrane Winterregengebiet
5.3.2.2.1 Vegetations- und Landschafts -geschichteVegetation und Landschaft des mediterranen Rau-mes sind so, wie sie sich heute darstellen, das Ergeb-nis prähistorischer und historischer Prozesse, derenRolle nicht immer leicht einzuschätzen ist (Grove &Rackham 2001). Palynologisch lässt sich belegen,dass das Klima während des letzten Hochglazialskühler, aber auch trockener war als heute. Währendder Glazialzeiten dominierte weiträumig eine step-penartige Offenlandvegetation, die sich in den ein-schlägigen Pollendiagrammen niederschlägt: DerAnteil windbestäubter Artemisia-Pollen in den Pro-ben aus dieser Zeit beträgt bis zu einem Drittel des gesamten Pollenspektrums. In den etwas feuchte-ren Interglazialen und im frühen Holozän breitetensich Bäume aus. Ob dabei Wälder mit einem ge -schlossenen Kronendach entstanden oder nur eineArt offener Waldbestand (woodland), ist umstritten.Jedenfalls hat dieses Hin und Her zwischen Gehölz-beständen und Steppe zum Aussterben vieler Ele-mente der alten pliozänen Baumflora geführt; dieperiodische pleistozäne Trockenheit des mediterra-nen Raumes, der ja auch die glazialen Refugien fürdie Waldarten der nemoralen Zone stellte, ist wohldie Hauptursache für die Artenarmut der europä -ischen Wälder, verglichen mit Nordamerika und Ost-asien (Lang 1994). Dennoch konnten zahlreichePflanzen aus dem Pliozän in Nischen überleben, wiesich u. a. auf der Iberischen Halbinsel belegen lässt(Gonzáles-Samperiz et al. 2010). Beispiele sind diefrost empfindliche Palme Phoenix theophrasti, dieman heute an wenigen küstennahen Stellen in Kretaund in der Ägäis als Tertiärrelikt findet, und Rhodo-dendron ponticum, ein immergrüner Strauch, der infeuchten Tälern im Südwesten der Iberischen Halb-insel überlebte. Sein Hauptvorkommen liegt aller-dings in den nemoralen Wäldern des südöstlichenSchwarzmeergebiets.
Ab etwa 12.000 Jahren BP konnten sich immer-grüne und sommergrüne Bäume erneut ausbreiten;nach 7.000 BP sind alle heute bekannten Arten derHartlaubwälder und -gebüsche, aber auch die ther-mophilen sommergrünen Eichenarten wie Quercuspubescens, Q. faginea, Q. cerris u. a. sowie die som-mergrünen, rezent vorwiegend nördlich der Alpenverbreiteten Gattungen Acer, Alnus, Betula, Carpinus,Fagus und Populus pollenanalytisch nachweisbar.Zwischen 7.000 und 5.000 BP verschwanden diesommergrünen zugunsten der immergrünen Ge höl -ze (Quercus ilex im Fall von Marokko und Korsika;
302 5 Die warm-gemäßigte (subtropische) Zone5

et al. 2003) als zonale Formation für den mediterra-nen Raum angesehen; danach würden sie heutebeträchtliche Flächen in der Südhälfte der IberischenHalbinsel, in Marokko und auf den Inseln (Kreta,Zypern, Mallorca, Korsika, Sardinien) sowie aufeinem bis zu 70 km breiten Küstenstreifen von Ita-lien, der Balkanhalbinsel und von Kleinasien einneh-men, während das Binnenland größtenteils bereitsnemoral (sub- bzw. supramediterran; s. unten) ist(Abb. 5-18). Die Vegetation ist von immergrünensklerophyllen Bäumen und Sträuchern besonders derGattungen Quercus, Pinus, Juniperus, Olea und Pista-cia geprägt (Raus & Bergmeier 2003). Pflanzensozio-logisch werden alle Hartlaubwälder einschließlichder thermo- und mesomediterranen Nadelwälder inder Klasse Quercetea ilicis zusammengefasst (z. B.Rivas-Martínez et al. 2001 für die Iberische Halbin-sel; Mucina 1997; Tab. 5-6). Die Hartlaubgebüscheauf Karbonatgestein unterscheiden sich dagegen flo-ristisch so deutlich von denjenigen auf silikatischenBöden, dass sie in zwei verschiedene Klassen gestelltwerden (Cisto-Lavenduletea auf Silikat- bzw. Cisto-Micromerietea auf Karbonatgestein). Natürliche, vonperennen Gräsern dominierte Pflanzengemeinschaf-ten fehlen außerhalb der Hochgebirge; es gibt sie nuranthropogen bei permanenter Beweidung und/odernach Feuer (Lygea sparti-Stipetea tenacissimae).
Den mediterranen Raum kann man von untennach oben (Höhengradient) bzw. von Süden nachNorden (Breitengrad-Gradient) in acht thermischeKlimazonen einteilen, denen jeweils eigene Vegeta-tionskomplexe zugewiesen sind (Tab. 5-7; Abb. 5-19;s. auch Tab. 5-11). Die Hartlaubwälder und -gebü-sche konzentrieren sich auf die thermo- und meso-mediterrane Zone. Die supra- bzw. submediteraneund die montan-mediterrane Stufe sind überall dortvon extrazonalen sommergrünen Laubwäldern ge -prägt, wo die Sommertrockenheit die Dauer voneinem Monat nicht überschreitet oder ganz fehlt. Wo
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 303
Reille 1992, Reille et al. 1996). Die Ursachen sindumstritten und werden entweder einer Klimaände-rung hin zu längeren, trockeneren Sommern (LeHouérou 1981) oder dem Menschen zugeschrieben(Rodung der sommergrünen Wälder auf den besse-ren, tiefgründigen Standorten für den Anbau vonFeldfrüchten; Lang 1994). Für die anthropogene Ur -sache spricht die neuerdings beobachtete Zunahmevon Quercus pubescens etwa im Süden Frankreichsoder auf der Iberischen Halbinsel (Neff & Franken-berg 1995, Quézel 2004); selbst im kalifornischenWinterregengebiet mit seinen besonders trockenenund heißen Sommermonaten sind sommergrüneEichen weit verbreitet (s. Abschn. 5.3.2.4). Somitscheint der immergrüne Charakter der Gehölze nurdann von Vorteil zu sein, wenn die Böden gleichzei-tig nährstoffarm sind wie in der Kapregion Südafri-kas und in Südwestaustralien, wo sommergrüneBäume und Sträucher komplett fehlen.
Vermutlich war die Pflanzendecke schon um 5.000 BP (Beginn der Bronzezeit und Entwicklungder ersten Hochkulturen wie der Minoer in Kreta) einMosaik aus parkartigem Offenland, Gebüsch undWäldern, das durch die vorhellenistischen Kulturenauf Kreta, in Sardinien, auf dem Peloponnes, in Süd-frankreich und in Süditalien geprägt war. Denn nachGrove & Rackham (2001) lassen sich pollenanalytischweder geschlossene Wälder in dieser Zeit nachweisennoch Belege für eine Degradation von Wäldern wäh-rend der folgenden hellenistischen und römischenPeriode finden. Deshalb bleiben Zweifel an der groß-flächigen Existenz prähistorischer Hartlaubwälder.Auch spricht der hohe Anteil von Neoendemiten, diekeine Waldarten sind, für die pleisto- und holozäneKontinuität des Offenlandes (Thompson 2005).
5.3.2.2.2 HartlaubwälderTrotzdem werden Hartlaubwälder und -gebüsche inder Karte der natürlichen Vegetation Europas (Bohn
Ar
Abb. 5-18 Potenzielles Verbreitungs-gebiet der mesomediterranen Stein -eichen- (Quercus ilex-)Wälder (schrägschraffiert), der thermomediterranenÖlbaum- (Olea europaea-Pistacia len-tiscus-)Wälder (punktiert) und derinframediterranen Argania-Gehölze inWestmarokko (Ar; nach Braun-Blan-quet 1964 aus Walter 1968).

dies nicht der Fall ist, wie im Süden und Osten (z. B.Kreta, Südtürkei), gibt es endemische trockenheitsre-sistente Nadelwälder aus Pinus-, Cedrus- und Junipe-rus-Arten.
304 5 Die warm-gemäßigte (subtropische) Zone5
Die inframediterrane Zone nimmt die wärmsten,absolut frostfreien Gebiete im äußersten Südwestendes mediterranen Raumes ein (Mauretanien, Ma -rokko). In Südwestmarokko sind die beiden charak-
Tab. 5-6 Übersicht über die thermo- und mesomediterranen Syntaxa des Mittelmeergebiets (nach Mucina 1997). Die thermome-diterranen Nadelwälder aus Pinus halepensis, P. brutia und Cupressus sempervirens werden wegen ihres sklerophyllen Unter-wuchses in die Ordnung Pistacio-Rhamnetalia gestellt (s. auch Bergmeier 2003).
Klasse Erläuterung
Wälder und Gebüsche
Quercetea ilicis Br.-Bl. et O. de Bolós 1958 mediterrane, immergrüne Eichenwälder und Macchien:Quercetalia ilicis (mesomediterrane Steineichenwälder)Pistacio lentisci-Rhamnetalia alaterni (thermomediterrane Ölbaum- und Nadelwälder)
Cisto-Lavenduleta Br.-Bl. In Br.-Bl. et al. 1940 niedrigwüchsiges mediterranes Hartlaubgebüsch (Garrigue, Phrygana) auf Silikat (eher westmediterran)
Cisto-Micromerietea julianae Oberd. 1954 niedrigwüchsiges mediterranes Hartlaubgebüsch (Garrigue, Phrygana) auf Karbonat (eher ostmediterran)
Nero-Tamaricetea Br.-Bl. et O. de Bolós 1958 mediterrane Galeriewälder und ähnliche Gebüsche
Grasland
Daphno-Festucetea Quézel 1964 griechisches und ägäisches, oromediterranes Grasland und Phrygana auf Karbonatgestein
Lygea sparti-Stipetea tenacissimae Rivas-Martínez 1978 halbnatürliches mediterranes Grasland
Annuellenfluren
Thero-Brachypodietea Br.-Bl. ex A. de Bolós y mediterrane terrestrische niedrige Annuellenfluren aus Gräsern Vayreda 1950 und Kräutern
Gebüsch ausAlnus alnobetula ssp. suaveolusundJuniperus communis var. saxatilis(schattseitig)
Gebüsch ausBerberis aetnensisundGenista lobelii(sonnseitig)
Fagus sylvatica(schattseitig)
Fagus sylvatica-Abies alba-Wald
Quercus pubescens-Wald
Quercus ilex-Wald Quercus suber-Wald
Aleria
Mt. Cinto (2710 m)
Calvi
Pinus nigrassp. laricio(sonnseitig)
0
1000
2000
3000mNN NW SO
a
Pachenes, Lefka Ori (2453 m)
Dornpolster-gebüsch
offener Cupressussempervirens-Wald
Olea europaea-Ceratonia siliqua-Wald
Quercus coccifera-Wald
Pinus brutia-Wald
Chora SfakionChania0
1000
2000
3000mNN NNW SSO
b
Abb. 5-19 Schema der Vegetationsabfolgen in Korsika (a, westmediterran) und Kreta (b, ostmediterran), nach Vorlagen von H.Albrecht. Quercus suber- (Korkeichen-), Olea europaea-Ceratonia siliqua- (Ölbaum-Johannisbrotbaum-) und Pinus brutia-Wälder sindthermomediterran, Quercus ilex- (Steineichen-) und Q. coccifera- (Kermeseichen-)Wälder sind mesomediterran, Quercus pubes-cens- (Flaumeichen-)Wälder sind supramediterran, Fagus sylvatica- (Rotbuchen-), Pinus nigra ssp. larico- und Cupressus sempervi-rens var. horizontalis-Wälder sind montan-mediterran.
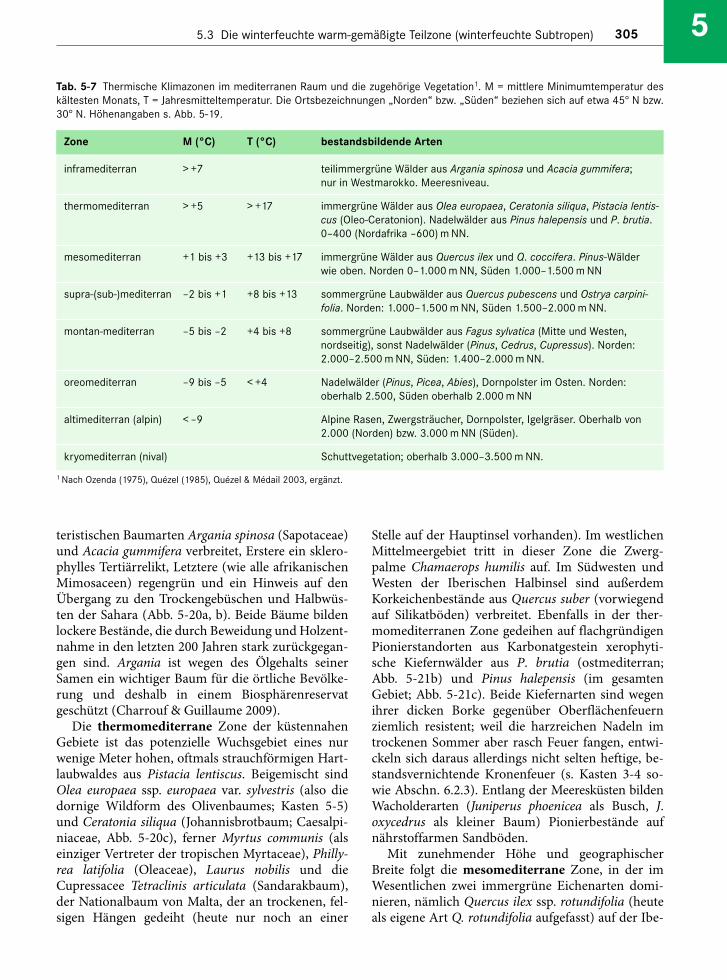
teristischen Baumarten Argania spinosa (Sapotaceae)und Acacia gummifera verbreitet, Erstere ein sklero-phylles Tertiärrelikt, Letztere (wie alle afrikanischenMimosaceen) regengrün und ein Hinweis auf denÜbergang zu den Trockengebüschen und Halbwüs-ten der Sahara (Abb. 5-20a, b). Beide Bäume bildenlockere Be stän de, die durch Beweidung und Holzent-nahme in den letzten 200 Jahren stark zurückgegan-gen sind. Argania ist wegen des Ölgehalts seinerSamen ein wichtiger Baum für die örtliche Bevölke-rung und deshalb in einem Biosphärenreservatgeschützt (Charrouf & Guillaume 2009).
Die thermomediterrane Zone der küstennahenGebiete ist das potenzielle Wuchsgebiet eines nurwenige Meter hohen, oftmals strauchförmigen Hart-laubwaldes aus Pistacia lentiscus. Beigemischt sindOlea europaea ssp. europaea var. sylvestris (also diedornige Wildform des Olivenbaumes; Kasten 5-5)und Ceratonia siliqua (Johannisbrotbaum; Caesalpi-niaceae, Abb. 5-20c), ferner Myrtus communis (alseinziger Vertreter der tropischen Myrtaceae), Philly-rea latifolia (Oleaceae), Laurus nobilis und dieCupressacee Tetraclinis articulata (Sandarakbaum),der Nationalbaum von Malta, der an trockenen, fel-sigen Hängen gedeiht (heute nur noch an einer
Stelle auf der Hauptinsel vorhanden). Im westlichenMittelmeergebiet tritt in dieser Zone die Zwerg-palme Chamaerops humilis auf. Im Südwesten undWesten der Iberischen Halbinsel sind außerdemKorkeichenbestände aus Quercus suber (vorwiegendauf Silikatböden) verbreitet. Ebenfalls in der ther-momediterranen Zone gedeihen auf flachgründigenPionierstandorten aus Karbonatgestein xerophyti-sche Kiefernwälder aus P. brutia (ostmediterran;Abb. 5-21b) und Pinus halepensis (im gesamtenGebiet; Abb. 5-21c). Beide Kiefernarten sind wegenihrer dicken Borke gegen über Oberflächenfeuernziemlich resistent; weil die harzreichen Nadeln imtrockenen Sommer aber rasch Feuer fangen, entwi-ckeln sich daraus allerdings nicht selten heftige, be -standsvernichtende Kronenfeuer (s. Kasten 3-4 so -wie Abschn. 6.2.3). Entlang der Meeresküsten bildenWacholderarten (Juniperus phoe nicea als Busch, J.oxycedrus als kleiner Baum) Pionierbestände aufnährstoffarmen Sandböden.
Mit zunehmender Höhe und geographischerBreite folgt die mesomediterrane Zone, in der imWesentlichen zwei immergrüne Eichenarten domi-nieren, nämlich Quercus ilex ssp. rotundifolia (heuteals eigene Art Q. rotundifolia aufgefasst) auf der Ibe-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 305
Tab. 5-7 Thermische Klimazonen im mediterranen Raum und die zugehörige Vegetation1. M = mittlere Minimumtemperatur deskältesten Monats, T = Jahresmitteltemperatur. Die Ortsbezeichnungen „Norden“ bzw. „Süden“ beziehen sich auf etwa 45° N bzw.30° N. Höhenangaben s. Abb. 5-19.
Zone M (°C) T (°C) bestandsbildende Arten
inframediterran > +7 teilimmergrüne Wälder aus Argania spinosa und Acacia gummifera; nur in Westmarokko. Meeresniveau.
thermomediterran > +5 > +17 immergrüne Wälder aus Olea europaea, Ceratonia siliqua, Pistacia lentis-cus (Oleo-Ceratonion). Nadelwälder aus Pinus halepensis und P. brutia.0–400 (Nordafrika –600) m NN.
mesomediterran +1 bis +3 +13 bis +17 immergrüne Wälder aus Quercus ilex und Q. coccifera. Pinus-Wälder wie oben. Norden 0–1.000 m NN, Süden 1.000–1.500 m NN
supra-(sub-)mediterran –2 bis +1 +8 bis +13 sommergrüne Laubwälder aus Quercus pubescens und Ostrya carpini-folia. Norden: 1.000–1.500 m NN, Süden 1.500–2.000 m NN.
montan-mediterran –5 bis –2 +4 bis +8 sommergrüne Laubwälder aus Fagus sylvatica (Mitte und Westen, nordseitig), sonst Nadelwälder (Pinus, Cedrus, Cupressus). Norden:2.000–2.500 m NN, Süden: 1.400–2.000 m NN.
oreomediterran –9 bis –5 < +4 Nadelwälder (Pinus, Picea, Abies), Dornpolster im Osten. Norden: oberhalb 2.500, Süden oberhalb 2.000 m NN
altimediterran (alpin) < –9 Alpine Rasen, Zwergsträucher, Dornpolster, Igelgräser. Oberhalb von2.000 (Norden) bzw. 3.000 m NN (Süden).
kryomediterran (nival) Schuttvegetation; oberhalb 3.000–3.500 m NN.
1 Nach Ozenda (1975), Quézel (1985), Quézel & Médail 2003, ergänzt.

rischen Halbinsel (Abb. 5-20d) und in Südfrankreich,ansonsten Q. ilex ssp. ilex (Q. ilex), und Q. coccifera(Kermeseiche) im Osten (Abb. 5-19, Abb. 5-20e). ImUnterwuchs dieser Wälder kommen eine Reihe vonGeophyten vor wie verschiedene Arten der GattungCyclamen, und, falls der Bestand nicht zu dicht ist,hartlaubige Sträucher, die zu den Gattungen Philly-rea, Arbutus, Rhamnus und Viburnum gehören. Alsholzige Liane wächst Smilax aspera. Anstelle der Wäl-der sind heute Hartlaubgebüsche (Macchie, Garriguebzw. Phrygana, s. unten) häufig anzutreffen. Auchhier kommen auf flachgründigen Böden die obengenannten Pinus-Wälder vor. Sowohl die thermo- alsauch die mesomediterrane Zone sind überwiegend inlandwirtschaftliche Nutzflächen überführt.
Der supramediterranen Höhenstufe in den medi-terranen Gebirgen entspricht der submediterranesüdliche Rand der nemoralen Zone in Oberitalien,auf der Balkanhalbinsel, in Südfrankreich (soweitnicht thermo- bzw. mesomediterran) und im Nordender Iberischen Halbinsel (s. Abschn. 6.2.2.6.2). In
beiden Fällen besteht die Baumschicht aus sommer-grünen Bäumen, zu denen vor allem die FlaumeicheQuercus pubescens (mit regionalen Kleinarten) unddie Hopfenbuche Ostrya carpinifolia gehören, auf derIberischen Halbinsel und im Atlas-Gebirge auch Q.faginea. Nur in der Strauchschicht macht sich nochder sklerophylle Charakter bemerkbar, z. B. durchdas Vorkommen von Ruscus aculeatus, dem Mäuse-dorn. Ansonsten fehlen die sklerophyllen Arten derthermo- und mesomediterranen Zone. Stattdessengedeihen hier zahlreiche nemoral verbreitete, (ausmitteleuropäischer Sicht) thermophile, laubabwer-fende Sträucher (wie der Perückenstrauch Cotinuscoggygria) und Kräuter. Im Westen treten kleinblät-trige Ahornarten wie Acer monspessulanum und A.opalus auf, im Osten Q. cerris und eine Reihe weite-rer sommergrüner Eichen. Eine Besonderheit sinddie beiden weltweit einzigen immergrünen Acer-Arten (von über 100), nämlich A. sempervirens inKreta und A. obtusifolium im Taurus-Gebirge (Tür-kei), im Libanon und in Israel (s. Abb. 5-23c).
306 5 Die warm-gemäßigte (subtropische) Zone5
Herkunft, Bedeutung und Kultivierung des OlivenbaumesEine der ältesten Kulturpflanzen und ein Symbol des Mittel-meerraumes ist der Olivenbaum (Olea europaea ssp. euro-paea; Abb. 1). Sein Areal umfasst das gesamte mediterraneGebiet mit Schwerpunkt in der thermomediterranen Zone(Abb. 2). Exklaven gibt es in der Umgebung der oberitalieni-schen Seen, entlang der Südküste des Schwarzen Meeresund auf der nördlichen Iberischen Halbinsel mit frostarmemKlima.
Die paläotropische Gattung Olea umfasst 33 Arten, dievon Afrika bis nach Südchina verbreitet sind. Der europäi-
sche Olivenbaum Olea europaea spaltet sich in sechs Unter-arten mit disjunktem Vorkommen auf: ssp. europaea immediterranen Raum, ssp. cuspidata von Südafrika über Süd-ägypten und die Arabische Halbinsel bis Südindien, ssp.laperrinei in den zentralsaharischen Gebirgen, ssp. maroc -cana in Südwestmarokko, ssp. cerasiformis (Madeira) undssp. guanchica auf den Kanarischen Inseln (Green 2002,Mabberley 2008). Die heute überall angebauten Kulturfor-men (var. europaea) sind das Ergebnis einer jahrtausende-langen Selektion großfrüchtiger Sorten aus verschiedenenWildarten (zu denen wohl auch ssp. cuspidata gehört) mitkleineren Früchten, deren Ölgehalt geringer ist als derjenigeder Kultursorten. Deren große genetische und morphologi-sche Vielfalt ist das Ergebnis von häufiger Hybridisierungzwischen kultivierten und wilden Populationen (Thompson2005). Die Wildform (var. sylvestris), die man überall imUmfeld von Olivenkulturen finden kann, ist genetisch unein-heitlich; sie besteht aus ursprünglichen Populationen undverwilderten Kultursorten (Zohary & Spiegel-Roy 1975).
Die Kultivierung des Olivenbaumes begann vermutlich im6. Jahrtausend BP im Nahen Osten nördlich des Toten Mee-res und breitete sich von dort über den gesamten Mittel-meerraum aus. Dabei entstanden Kultursorten auch auslokalen Populationen der Wildart, wie beispielsweise in Spa-nien schon zu Beginn der Bronzezeit, also vor der Einführungkultivierter Pflanzen aus dem Nahen Osten (Terral 2004).Olivenbaumkultivare werden durch Propfen von Edelreisern
Kasten 5-5
Abb. 1 Olivenbaumkultur (Mallorca, Spanien).

Im Apennin und auf Korsika kommen oberhalbvon etwa 2.000 m NN Wälder aus Fagus sylvatica vor.Sie gehören zur montan-mediterranen Stufe, die indiesem Teil des mediterranen Raumes der Zone dermitteleuropäischen Buchenwälder entspricht (s.Abschn. 6.2.2.6.2). Die Wälder sind also extrazonal.Nemoral-kühle, aber sommertrockene Gebirgslagenwerden dagegen von Nadelhölzern eingenommen,im Westen und Norden südseitig (wie Pinus nigrassp. laricio auf Korsika), im kontinentalen Osten un -abhängig von der Exposition. So dominiert auf Kretaoberhalb von 1.000– 1.500 m NN ein lichter Wald ausder breitkronigen (indigenen) Varietät Cupressussempervirens var. horizontalis (Schütt et al. 2004; Abb.5-21a). Die schmalkronige Zierform (var. stricta)wurde bereits von den Römern gepflanzt und prägtheute die „klassische“ mediterrane Kulturlandschaft(s. Abb. 5-23a).
Auf die nach oben anschließenden Zonen (oreo-mediterran, altimediterran, kryomediterran) werdenwir in Abschn. 5.5 eingehen. Neben der Gebirgstaiga,
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 307
die mit Pinus-, Picea- und Abies-Arten bereits anboreale Nadelwälder erinnert (oreomediterran), gibtes in der altimediterranen Stufe (anthropogen auchin tieferen Lagen) eine für sommertrockene Gebirgs-lagen charakteristische Dornpolstervegetation, inden humiden nord- und westmediterranen GebirgenZwergstrauchheiden und alpine Rasen, wie man siein ähnlicher Form auch in den nemoralen Hochge-birgen findet.
5.3.2.2.3 Hartlaubgebüsche Hartlaubgebüsche aus sklerophyllen und malako-phyllen Sträuchern und einer großen Anzahl anEphemeren waren wahrscheinlich schon in prähisto-rischer Zeit vorhanden. Vermutlich bildeten sie einMosaik mit Hartlaubwäldern und waren mit diesendurch Sukzessionsprozesse verbunden (z. B. in Formvon Regenerationszyklen nach Feuer; Abb. 5-22).Heute ist ihre landschaftsbestimmende Dominanzeine Folge der anthropogenen Degradation von Wäl-dern (Braun-Blanquet 1964). So entsteht aus einem
auf Wildlinge vermehrt, um genetisch einheitliche Klone zuerhalten. Die Vermehrung über Samen geschieht lediglichzum Zweck der Züchtung neuer Sorten. Für den Anbau eig-nen sich nur Gebiete ohne längere Frostperioden; allerdingsbenötigen die Bäume eine winterliche Kältezeit zwischen 0und 10 °C, um zum Blühen zu gelangen (Lieberei & Reisdorff2007). Die Kultivare werden bis zu 20 m hoch (im Gegensatzzur Wildform mit kaum mehr als 10 m) und erreichen einAlter von nahezu 2.000 Jahren (Zohary & Spiegel-Roy 1975).Die oft bizarren Stammformen alter Olivenbäume gehen aufdie Infektion durch den holzbewohnenden BasidiomycetenFormitiporia punctata (Stammfäule) zurück. Die befallenenStellen werden zwar immer wieder ausgeschnitten, infizieren
sich aber jedesmal von neuem, sodass schließlich durchbro-chene und zerteilte Stämme entstehen. Die Vitalität derBäume und ihre Reproduktion werden dadurch aber nichtbeeinträchtigt.
Die Früchte werden kurz vor der Vollreife im Dezemberund Januar geerntet, wenn der Ölgehalt im Mesokarp beietwa 50 % liegt. Das wertvollste Öl gewinnt man aus den mitHand gepflückten Oliven, weil die Früchte dabei nichtbeschädigt werden. Druckschäden, wie sie auftreten kön-nen, wenn die Früchte mit einem Rechen abgestreift oderabgeschüttelt werden, mindern die Qualität, weil durch dieVerletzung freie Fettsäuren entstehen (Lieberei & Reisdorff2007).
Abb. 2 Verbreitung der Kultursorte(var. europaea, rot schraffiert) und derWildform (var. sylvestris, grün) von Oleaeuropaea ssp. europaea im mediterra-nen Raum (nach Walter & Straka 1970,Zohary & Spiegel-Roy 1975, verändert).

308 5 Die warm-gemäßigte (subtropische) Zone5
dc
a b
Abb. 5-20 Beispiele für Hartlaubwälder und Hartlaubgebüsche im Mittelmeergebiet. a = Argania spinosa (im Vordergrund) und Aca-cia gummifera (dahinter, links), Marokko; b = Früchte von Argania, Marokko (Fotos a, b W. Zielonkowski); c = Ceratonia siliqua-Bestand, Kreta; d = Quercus ilex-Wald, Mallorca.
a b
Abb. 5-21 Beispiele für Nadelwälder im Mittelmeergebiet. a = Reste von Cupressus sempervirens-Wäldern; b = offener Pi nus bru-tia-Wald. Beide Aufnahmen stammen aus Kreta.

55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 309
Quercus ilex-Wald durch Niederwaldbetrieb zu -nächst ein bis zu 6 m hohes Gebüsch, in dem derWaldunterwuchs z. B. aus Phillyrea latifolia, Arbutusunedo, Rhamnus alaternus, Pistacia lentiscus, Ericaarborea und anderen Hartlaubsträuchern zur Domi-nanz gelangt. Dieses Gebüsch ist eine Macchie (Abb.5-20g). Bei fortschreitender Nutzung, insbesondereBeweidung verbunden mit Feuer, entsteht darauseine Vegetation aus Klein- und Zwergsträuchern wieRosmarinus officinalis, Lavandula latifolia,Thymusvulgaris u. v. a. (Garrigue), die wohl die aromatisch-ste aller Pflanzengemeinschaften der Erde ist (Klötzliet al. 2010; Abb. 5-20f). Der hohe Gehalt an verschie-denen Terpenen bedeutet aber auch eine er höhteBrandgefahr. So ist die Garrigue also eine Vegetation,
g h
e f
Abb. 5-20 (Fortsetzung) e = Quercus coccifera-Bestand, Kreta; f = Garrigue aus Cistus salviifolius und Euphorbia dendroides, Mal-lorca; g = Macchie aus Myrtus communis und Arbutus unedo, Mallorca; h = Phrygana mit Sarcopoterium spinosum (Rosaceae) undPhlomis lanata (Lamiaceae), Kreta.
c
Abb. 5-21 (Fortsetzung) c = Pinus halepensis-Wald auf Mal-lorca.
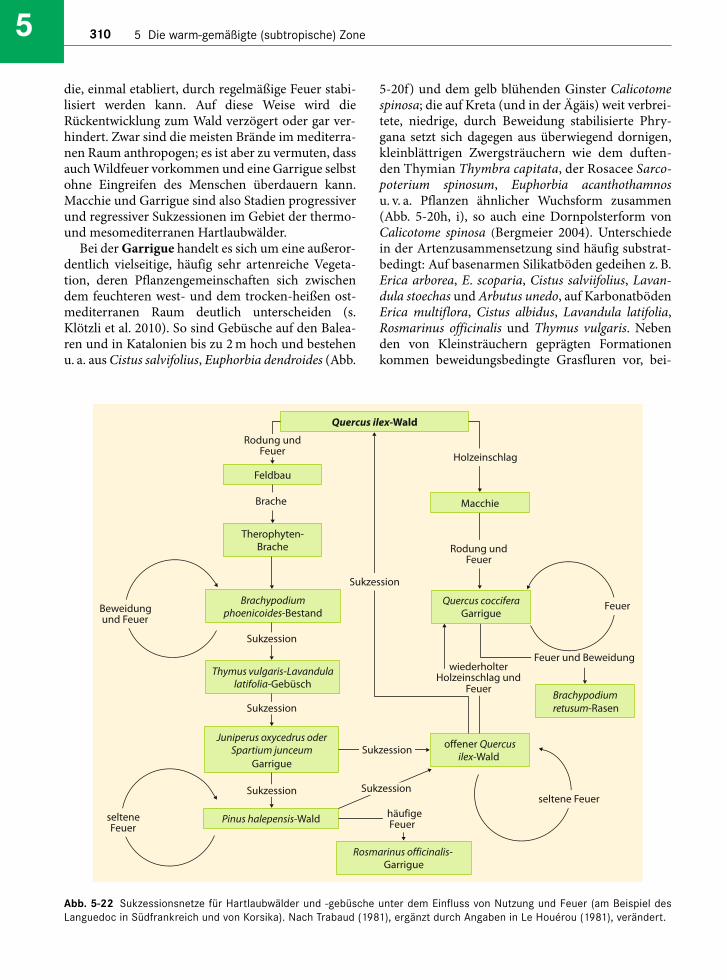
die, einmal etabliert, durch regelmäßige Feuer stabi-lisiert werden kann. Auf diese Weise wird dieRückentwicklung zum Wald verzögert oder gar ver-hindert. Zwar sind die meisten Brände im mediterra-nen Raum anthropogen; es ist aber zu vermuten, dassauch Wildfeuer vorkommen und eine Garrigue selbstohne Eingreifen des Menschen überdauern kann.Macchie und Garrigue sind also Stadien progressiverund regressiver Sukzessionen im Gebiet der thermo-und mesomediterranen Hartlaubwälder.
Bei der Garrigue handelt es sich um eine außeror-dentlich vielseitige, häufig sehr artenreiche Vegeta-tion, deren Pflanzengemeinschaften sich zwischendem feuchteren west- und dem trocken-heißen ost-mediterranen Raum deutlich unterscheiden (s.Klötzli et al. 2010). So sind Gebüsche auf den Balea-ren und in Katalonien bis zu 2 m hoch und bestehenu. a. aus Cistus salvifolius, Euphorbia dendroides (Abb.
5-20f) und dem gelb blühenden Ginster Calicotomespinosa; die auf Kreta (und in der Ägäis) weit verbrei-tete, niedrige, durch Beweidung stabilisierte Phry -gana setzt sich dagegen aus überwiegend dornigen,kleinblättrigen Zwergsträuchern wie dem duften-den Thymian Thymbra capitata, der Rosacee Sarco-poterium spinosum, Euphorbia acanthothamnosu. v. a. Pflanzen ähnlicher Wuchsform zusammen(Abb. 5-20h, i), so auch eine Dornpolsterform vonCalicotome spinosa (Bergmeier 2004). Unterschiedein der Artenzusammensetzung sind häufig substrat-bedingt: Auf basenarmen Silikatböden gedeihen z. B.Erica arborea, E. scoparia, Cistus salviifolius, Lavan-dula stoechas und Arbutus unedo, auf KarbonatbödenErica multiflora, Cistus albidus, Lavandula latifolia,Rosmarinus officinalis und Thymus vulgaris. Nebenden von Kleinsträuchern geprägten Formationenkommen beweidungsbedingte Grasfluren vor, bei-
310 5 Die warm-gemäßigte (subtropische) Zone5
Quercus ilex-Wald
Feldbau
Therophyten-Brache
Brachypodium phoenicoides-Bestand
Thymus vulgaris-Lavandula latifolia-Gebüsch
Brachypodiumretusum-Rasen
Quercus coccifera Garrigue
offener Quercus ilex-Wald
Macchie
Pinus halepensis-Wald
Juniperus oxycedrus oder Spartium junceum
Garrigue
Rodung und Feuer
Rodung und Feuer
Feuer und Beweidung
Brache
Holzeinschlag
wiederholter Holzeinschlag und
Feuer
Sukzession
Sukzession
Sukzession
Sukzession
Rosmarinus officinalis- Garrigue
häufigeFeuer
Sukzession
Sukzession
Beweidung und Feuer
seltene Feuer
Feuer
seltene Feuer
Abb. 5-22 Sukzessionsnetze für Hartlaubwälder und -gebüsche unter dem Einfluss von Nutzung und Feuer (am Beispiel desLanguedoc in Südfrankreich und von Korsika). Nach Trabaud (1981), ergänzt durch Angaben in Le Houérou (1981), verändert.

spielsweise aus dem Tiefwurzler Brachypodium retu-sum und dem bis zu 3 m hohen Horstgras Ampelo-desmos mauritanicus. Geophyten wie Asphodelus-und Narcissus-Arten sind weit verbreitet.
5.3.2.2.4 Anthropogener EinflussDie mediterrane Kulturlandschaft ist vielerorts nochaußerordentlich divers (Abb. 5-23). Diese Vielfaltberuht erstens auf den unterschiedlichen landwirt-schaftlichen Betriebsgrößen, die von großen markto-rientierten Latifundien bis zu kleinen, stellenweisesubsistenzorientierten (d. h. für den Eigenbedarf pro-duzierenden) Gartenbaubetrieben reichen, zweitensauf den verschiedenen Nutzungssystemen wie Re -gen feldbau im Winter, Bewässerungslandbau imSommer sowie Dauerkulturen aus Fruchtbäumenund drittens auf der großen Zahl von Nutzpflanzen,die aus unterschiedlichen Klimazonen stammen undim mediterranen Raum kultiviert werden können(Schultz 2000). So erlauben die kühlen Temperaturenim Winter den Anbau von Feldgemüse (Tomaten,Kartoffeln, Salat) und Getreide (Weizen, Gerste) ausder kühl-gemäßigten (bzw. oreotropischen) Zone inForm des Regenfeldbaus. Viele Gemüsepflanzen wiePorree, Gemüsefenchel, Knollensellerie, Rote Bete(nach der Wildform Beta vulgaris), Artischocke so -wie alle Kopf- und Blattkohlsorten (Brassica oleracea)sind mediterranen Ursprungs. Im Sommer ist dieKultivierung von Reis möglich, sofern genug Wasserwie in Küstennähe und in Flusstälern zur Verfügungsteht. Während die einheimischen Fruchtbäume Öl -baum, Feige und Mandel sowie die Weinrebe diesommerliche Trockenphase ohne Probleme vertra-gen, müssen Obstbäume wie Zitrusfrüchte, derenHeimat die immerfeuchten Subtropen Ostasiens sind,gelegentlich bewässert werden.
Abgesehen von den besten (tiefgründigen) Böden,die ackerbaulich genutzt werden oder mit Dauerkul-turen bestanden sind, wird nahezu der gesamtemediterrane Raum mit Schafen und Ziegen beweidet(Le Houérou 1990). Beweidung gehört zur Kulturge-schichte des Mittelmeergebiets und prägt vor allemdie Gebirge. In Kombination mit Feuer stabilisiert siedie Garrigue und schafft vielerorts eine savannenar-tige Landschaft aus einzeln stehenden Bäumen undGehölzgruppen (Abb. 5-23). Abgekoppelt von derrauen Wirklichkeit des Hirtenlebens und schon inhellenischer Zeit mythisch verklärt, wurde sie im 18.und 19. Jahrhundert als „Arkadien“ (eigentlich eineWeidelandschaft auf dem Peloponnes und heute einVerwaltungsbezirk) zum Sehnsuchtsort der nachSüden reisenden Bildungsbürger und damit zum Ide-albild einer ländlichen Idylle, in der Mensch und Tier
glücklich miteinander leben (Frizell 2009). Ein Bei-spiel für eine derartige Landschaft ist die spanischeDehesa, eine Parklandschaft aus Quercus rotundifoliaund Q. suber im Südwesten von Spanien (Extrema-dura, Andalusien) und Portugal, die heute überwie-gend in Privatbesitz ist (Joffre et al. 1999). Ihre Flächebeträgt rund 50.000 km2 und wird hauptsächlich vonSchafen, Ziegen und Schweinen verschiedener loka-ler Rassen beweidet. Die Dehesa besteht aus einem
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 311
a
b
c
Abb. 5-23 Beispiele für mediterrane Kulturlandschaften. a =traditionelle mediterrane Kulturlandschaft mit Cupressus sem-pervirens var. stricta, Mallorca; b = Äcker und Olivenhaine inder fruchtbaren Ebene bei Festos, Kreta; c = arkadische Wei-delandschaft mit Acer sempervirens, Kreta.

Mo saik aus Weideflächen, Brachland (Garrigue) undAckerflächen und ist ein Brut- und Nahrungsgebietfür europa- und weltweit gefährdete Tierarten wieden Mönchsgeier.
Aufforstungen der Garrigue wurden bis vor 30 Jah-ren vielerorts mit Pinus-Arten (besonders P. halepen-sis und P. pinea) durchgeführt. Diese unterschiedensich kaum von den natürlichen Beständen mit sklero-phyllem Unterwuchs. Wegen der harzreichen Streusind sie stark feuergefährdet. Die großen verheeren-den Waldbrände der letzten Jahrzehnte im mediterra-nen Raum gehen auf (anthropogene) Feuer in den
Pinus-Forsten zurück, z. T. mit verheerenden Folgen inBezug auf Bodenabtrag und Hochwasser (Pausas et al.2008). Heute werden immer mehr Exoten angepflanzt,insbesondere Eucalyptus-Arten wie E. globulus.
5.3.2.3 Das kalifornische Winterregengebiet
5.3.2.3.1 Hartlaubgebüsche (Chaparral und Coastal Scrub)Die Vegetation des kalifornischen Winterregenge-biets wird (außerhalb des zentralen Längstales mit
312 5 Die warm-gemäßigte (subtropische) Zone5
e f
ba
dc
Abb. 5-24 Beispiele für Wälder und Gebüsche des kaliforni-schen Winterregengebiets. a = montaner Chaparral östlich vonLos Angeles; b = „Chamissal“ aus Adenostoma fasciculatum; c = Arizona-Chaparral mit Quercus emoryi (im Mittelgrund) undCercocarpus montanus im Vordergrund; d = Quercus agrifolia-Hartlaubwald, Monterey; e = Cupressus macrocarpa-Bestand,Monterey-Halbinsel; f = Pinus radiata-Bestand mit Equisetumtelmateia ssp. braunii im Vordergrund. Fotos: d H. Albrecht, dieübrigen F. Klötzli.

55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 313
große Menge an brennbarem Material aus abgestor-benen Ästen und Blättern in der Strauchschicht ange-häuft hat. Wegen des Holzanteils sind die Brände mitdurchschnittlich 500 °C heißer als die Grasfeuer derSavannen und Prärien, sodass die oberirdische Phy-tomasse komplett verbrennt. Bei starkem Wind (wiedie Santa-Ana-Stürme im Spätherbst und Winter)können mehrere Tausend Hektar betroffen sein. Dievegetationsfreien Brandflächen werden fast aus-schließlich von Pflanzen besiedelt, die sich entwederaus den überlebenden unterirdischen Organen bzw.einem Lignotuber (resprouter) oder aus der Samen-bank im Boden (reseeder) zu regenerieren vermögen.Zuwanderung über Fernausbreitung ist selten. Manunterscheidet a) obligate reseeders ohne resprouter-Eigenschaften, b) fakultative reseeders mit resprouter-
Gras- und Zwergstrauchhalbwüsten) großräumigvon einem niedrigen (1,5–4 m hohen) Hartlaubge-büsch (Chaparral) bestimmt (Knapp 1965, Keeley2000, Keeley & Davis 2007; Abb. 5-24a, c). Physio-gnomisch entspricht der Chaparral der Macchie immediterranen Raum, ist aber im Gegensatz zu diesermeist kein vorübergehendes Degradationsstadiumder Hartlaubwälder, sondern entweder eine azonaleVegetation auf extrem flachgründigen Steilhängenoder, häufiger, eine Feuerklimax, stabilisiert durchwiederholte natürliche Feuerereignisse. Er kommt inHöhenlagen zwischen 0 und 1.500 m NN bei mittle-ren Jahresniederschlägen zwischen 300 und 650 mmvor und überzieht im südlichen Kalifornien dieLandschaft wie eine Decke von der Küste bis zumFuß der Sierra Nevada. Die heutige Verbreitung desChaparral ist auf eine frühholozäne Klimaänderunghin zu trockeneren und längeren Sommern zurück-zuführen; dadurch konnte sich das Hartlaubgebüschum ca. 500 km nach Norden ausbreiten.
Die Flora des Chaparral enthält 1.177 Tracheo-phytenarten (davon 42 % endemisch), die sich u. a.aus über 219 Straucharten, 409 Therophyten und 389mehrjährigen Gräsern und Kräutern zusammenset-zen (Keeley & Davis 2007). In ungestörtem Zustand(d. h. mehr als 20 Jahre nach dem letzten Feuerereig-nis) dominieren sklerophylle Sträucher und Halb-sträucher; sie bestehen u. a. aus den Gattungen Ade-nostoma (Rosaceae), Arctostaphylos (Ericaceae, über80 auf das kalifornische Winterregengebiet be -schränkte Arten), Ceanothus (Rhamnaceae, über 50Arten), Cercocarpus und Heteromeles (beide Rosa-ceae) sowie Quercus mit fünf strauchförmig wach-senden Arten, von denen Quercus berberidifolia amhäufigsten ist . Die wenigen malakophyllen Sträucherstammen aus den Gattungen Artemisia und Salviaund sind besonders im küstennahen Sage Brush (s.unten) verbreitet. Wegen des ausgeprägten Reliefsgibt es eine Vielzahl von Pflanzengemeinschaften. Sowerden die Südhänge bevorzugt von Adenostoma fas-ciculatum (Chamissal) besiedelt, einem bis zu 3 mhohen erikoiden Kleinstrauch mit nadelförmigenBlättern (Abb. 5-24b). Auch kleinblättrige Ceano-thus- und Arctostaphylos-Arten kommen hier vor.Dagegen wachsen auf den kühleren und feuchterenNordhängen breitblättrige Sträucher (wie Quercusberberidifolia, Frangula californica, Prunus ilicifoliau. a.). Einige Arten sind in Abb. 5-25 zeichnerischdargestellt (aus Walter 1968).
Wildfeuer prägen Artenzusammensetzung undStruktur des Chaparral entscheidend. Sie entwickelnsich als Kronenfeuer im Mittel alle 40 bis 60 Jahre amEnde des langen trockenen Sommers, wenn sich eine
1
3
4
5
6
2
Abb. 5-25 Einige Arten aus dem kalifornischen Chaparral. 1 =Arctostaphylos tomentosa, Ericaceae; 2 = Adenostoma fascicu-latum, Rosaceae; 3 = Ceanothus cuneatus, Rhamnaceae; 4 = C.papillosus; 5 = Pickeringia montana, Fabaceae; 6 = Quercus cf.berberidifolia, Fagaceae. Zeichnungen von R. Anheisser, ausWalter (1968).

Eigenschaften und c) obligate resprouters. Zum Typ agehören neben den zahlreichen annuellen Kräuternauch viele Sträucher aus den Gattungen Ceanothusund Arctostaphylos. Ihre Samen keimen bei ausrei-chender Bodenfeuchte und günstigen Temperaturenunmittelbar nach dem Feuer. So entsteht schon imersten Jahr eine geschlossene, blütenreiche Pflanzen-decke. Die Quercus-, Prunus- und Rhamnus-Artensind obligate resprouters, die sich ausschließlich vege-tativ regenerieren und dafür mehrere Jahre brauchen.Sie bilden keine persistente Samenbank, da ihreSamen kurzlebig sind und meist unmittelbar nachder Reife keimen. Die übrigen Gehölze wie Adeno -stoma fasciculatum sowie die Mehrzahl der Ceano-thus- und Arctostaphylos-Arten sind fakultative resee-ders, die sich sowohl aus Samen als auch ausStockausschlägen regenerieren.
Die obligaten wie auch die fakultativen reseedersbilden eine persistente (1–5 Jahre) oder gar eine per-manente Samenbank (> 5 Jahre) im Boden. DieSamen bleiben bis zum nächsten Feuerereignis dor-mant. Dann wird die Keimruhe entweder durchHitze oder durch die chemische Wirkung des Rau-ches gebrochen. So keimen die Ceanothus-Arten erstnach einem Hitzeschock. Die Samen der meisten Sip-pen werden aber durch organische Verbindungen imRauch zur Keimung angeregt (chemische Stratifika-tion). Hierzu gehören die zahlreichen annuellen undperennen Gräser und Kräuter, die zusammen über 60 % der Chaparral-Flora stellen. Die meisten vonihnen blühen und fruchten in den ersten Jahren nachFeuer und überdauern den Rest des Feuerintervallsin der Samenbank. Einige persistieren aber auch inden gut entwickelten älteren Gebüschen überall dort,wo Lücken entstehen. Die am häufigsten vertretenenFamilien sind Asteraceae (Gnaphalium), Boragina-ceae (wie einige der in Mitteleuropa als Bienenweideund zur Bodenverbesserung angepflanzten Phacelia-Arten), Fabaceae (Lupinus), Portulaccaceae undScrophulariaceae.
Da die Samen der meisten Reseeders unter günsti-gen Umständen mehrere 100 Jahre keimfähig blei-ben, verändert sich die Artenzusammensetzung desChaparral nicht wesentlich, auch wenn die Feuerin-tervalle über 100 Jahre dauern. Umgekehrt benötigenvor allem die Gehölze nach einem Feuerereignis aus-reichend Zeit (zehn bis 20 Jahre), um den Samenvor-rat im Boden wieder aufzufüllen. Da heute die meis-ten Feuer anthropogen entstehen und deshalb dieFeuerintervalle kürzer sind als bei Wildfeuern, isteine erfolgreiche Reproduktion der Gehölze nichtmehr möglich. So entsteht aus einem Chaparral all-mählich ein Grasland, und zwar vorwiegend aus
314 5 Die warm-gemäßigte (subtropische) Zone5
europäischen C3-Gräsern (Bromus, Lolium, Vulpia)und Kräutern (Centaurea, Lactuca). Vor allem die ausdem Mittelmeergebiet bzw. aus den mittelasiatischenTrockengebieten stammenden Therophyten Bromusmadritensis und B. tectorum bilden überall dichteBestände.
Unter extrem trockenen Bedingungen, wozu nichtnur südexponierte Steilhänge aus Karbonatgestein,sondern auch die niederschlagsarmen südkaliforni-schen Küstengebiete mit Jahresniederschlägen unter250 mm gehören, gedeiht eine Vegetation aus mala-kophyllen Halbsträuchern wie Artemisia californica(sage brush), Eriogonum fasciculatum (buck wheat,Polgonaceae) und Salvia mellifera (black sage), denenzahlreiche weitere trockenresistente Zwergsträucherder Halbwüsten (wie Encelia farinosa) und Sukku-lente (Opuntia) beigemischt sind. Dieser CoastalScrub kennzeichnet den Übergang zu den Zwerg-strauch- und Sukkulenten-Halbwüsten der So no ra-und Mojave-Wüste, findet sich aber auch extra- bzw.azonal inmitten des kalifornischen Winterregenge-biets in einem Vegetationsmosaik aus Chaparral undEichenwäldern (Abb. 5-26).
Ein dem kalifornischen Chaparral strukturell undfloristisch vergleichbares Hartlaubgebüsch gibt esöstlich der Rocky Mountains in Arizona (Arizona-Chaparral; Abb. 5-24c). Es ist von teils subtropi-schen, im Norden schon nemoralen Halbwüsten undWüsten umgeben und enthält Sommerniederschläge,ist also eientlich kein Winterregengebiet mehr. Häu-fige Arten sind Quercus turbinella, Juniperus deppi-ana, Cercocarpus montanus und Rhus ovatus.
1S N
1
2
2
4
3
65 3
Abb. 5-26 Vegetationsprofil aus den tieferen Lagen der SanGabriel Mountains nördlich von Los Angeles, 550 m NN (ausKnapp 1965). 1 = Sage Brush mit Artemisia californica, 2 = Cha-parral, 3 = Quercus chrysolepis-Wald, 4 = Pseudotsuga macro-carpa-Bestand, 5 = laurophyller Umbellularia californica-Bestand, 6 = sommergrüner Laubwald aus Acer macrophyllum.Wie in den anderen Winterregengebieten gedeihen in kühlen,feuchten Tälern extrazonal Lorbeer- oder sogar sommergrüneWälder (s. auch Abb. 5-27b).

5.3.2.3.2 HartlaubwälderAußerhalb des von Chaparral eingenommen Gebietskommen Hartlaubwälder aus Quercus- und Nadel-wälder aus Pinus-, Cupressus- und Juniperus-Artenvor (Knapp 1965, Barbour & Minnich 2000, Allen-Diaz et al. 2007, Barbour 2007, Minnich 2007; Abb.5-24). Die Baumschicht der Hartlaubwälder des kali-fornischen Winterregengebiets wird einzeln oder inKombination aus acht Eichenarten gebildet, vondenen drei immergrün (Quercus agrifolia, Q. wislize-nii, Q. chrysolepis) und fünf sommergrün sind (Q.douglasii, Q. lobata, Q. engelmannii, Q. kelloggii, Q.garryana). Die Wälder werden in der Regel nur 10–15 m hoch, unter besonders günstigen Bedingungenauch über 20 m (Abb. 5-24d). Das Kronendach istnicht immer geschlossen; vielmehr bilden die Eichenauch lockere, offene (savannenartige) Bestände(Woodlands), die schon vor der europäischen Ein-wanderung existierten und deren Unterwuchs ver-mutlich aus Hemikryptophyten und Geophyten so -wie vereinzelten Hartlaubsträuchern bestand. DieVegetation wurde wohl schon in voreuropäischerZeit von den Haustieren der indigenen Bevölkerungbeweidet. In den dichteren Waldbe ständen gedei-hen auch einige hartlaubige Sträucher aus dem Cha-parral wie Arctostaphylos- und Ceanothus-Arten,decken aber nur wenige Prozent. Vielleicht handelt essich bei den Woodlands also um vom Menschendurch Feuer und Beweidung stabilisierte Relikte vonWaldentwicklungsstadien aus dem Mit telholozän.Jedenfalls sind wie andernorts ge schlossene Wälderauf waldfähigen Standorten wahrscheinlich. Heutedominieren im Unterwuchs eingeschleppte annuelleGräser und Kräuter.
Die Eichenwälder gedeihen bevorzugt auf eda-phisch oder klimatisch feuchteren Standorten, alsoentweder an Unterhängen, in Tälern und auf tief-gründigen Böden oder in schattseitigen Lagen bzw.im Einflussbereich des Küstennebels. Allen-Diaz etal. (2007) unterscheiden „Valley Oak Woodland“ ausQuercus lobata (valley oak) auf tiefgründigen Bödenin Tallagen, „Blue Oak Woodland“ aus Q. douglasii(blue oak) auf flachgründigen Böden, z. T. bereits mitPinus sabiniana, am Fuß der Sierra Nevada und„Coastal Oak Woodlands“ aus Q. agrifolia (coast liveoak) in Küstennähe. Dieser überwiegend immer-grüne Wald ist am ehesten mit den Q. ilex-Wälderndes mediterranen Raumes vergleichbar; er enthält auffeuchteren Böden einige weitere immergrüne Bäumeund Sträucher wie Q. chrysolepis, Arbutus menziesiiund Umbellularia californica (Lauraceae).
In Küstennähe schließlich gedeihen vereinzeltartenarme Nadelwälder auf sehr nährstoffarmen,
durchlässigen Böden aus Sand, Kies oder Serpentin(Abb. 5-24e, f). Die Wälder sind häufig von Chapar-ral umgeben und unterliegen wie dieser regelmäßi-gen Feuern im Abstand von etwa 50 Jahren. Bei allenNadelbäumen bleiben die Zapfen auch nach derSamenreife geschlossen (serotiny); erst bei Feuer öff-nen sie sich, sodass die Samen auf die abgebranntenFlächen fallen und dort ein ideales Keimbett vorfin-den. Von den zahlreichen Cupressus- und Pinus-Ar -ten sind C. macrocarpa und P. radiata deshalb be son -ders erwähnenswert, weil sie als Exoten in vielenGebieten der Subtropen für Aufforstungen oder alsornamentale Bäume verwendet werden (C. macro-carpa in Südamerika, P. radiata in Südafrika undNeuseeland).
5.3.2.4 Das chilenische Winterregengebiet
Die Hartlaubwälder und -gebüsche des chilenischenWinterregengebiets werden pflanzensoziologisch seitOberdorfer (1960) durch die Klasse Lithraeo-Cryp-tocaryetea und ihre einzige Ordnung Cryptocaryeta-lia repräsentiert. Sie reichen im Norden an der Küstewegen der noch ausreichenden Wasserversorgungdurch den Küstennebel etwa bis 31° S, im Landesin-neren dagegen nur etwa bis 33° S. Dort gehen sie ineine sukkulente Dornstrauchvegetation über, diebereits zu den tropisch-subtropischen Trockengebie-ten gezählt wird (Schmithüsen 1956; Abb. 5-27).Anders im Süden: Dort kommen sie gerade noch biszu einer Linie zwischen Concepción und Temuco beietwa 38° S vor, während am Westhang der feuchterenKüstenkordillere und in den Anden bereits lauro-phyll geprägte Wälder dominieren (Abb. 5-27).
Die Bäume der Hartlaubwälder sind gedrungenund sklerophyll und zählen zu den Tiefwurzlern(Abb. 5-28a). Charakteristisch ist eine reiche Ent-wicklung von Geophyten wie Solenomelus peduncu-latus (Iridaceae) im Frühling, wenn genügend Feuch-tigkeit vorhanden ist. Charakteristische Bäume sindCryptocarya alba (Lauraceae), Quillaja saponaria(eine Rosacee, aus deren Rinde früher Saponin fürdie Seifenherstellung gewonnen wurde), Peumus bol-dus (Monimiaceae), Myrceugenia obtusa (Myrtaceae)und Maytenus boaria (Celastraceae). Lianen sinddurch Proustia pyrifolia ssp. pyrifolia (Asteraceae)und Cissus striata (Ampelidaceae) vertreten. Bemer-kenswert sind die wenigen noch vorhandenenBestände von Jubaea chilensis, der einzigen einheimi-schen Palme Chiles, die ein Bestandteil der Hartlaub-wälder war und heute nur noch auf wenigen, strenggeschützten Flächen vorkommt (Abb. 5-28c). Inner-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 315

halb der Hartlaubvegetation gibt es in feuchten Bach-tälern einen extrazonalen Lorbeerwald mit Persealingue, Drimys winteri und Dasyphyllum excelsum.Hier kommt eine der (als „Monumento Natural“)besonders streng geschützten Baumarten Chiles vor,nämlich Beilschmiedia miersii, eine bis 20 m hochwerdende Lauracee.
Charakteristische Matorral-Bildner sind die Eu -phorbiacee Colliguaja odorifera, die Asteracee Bac-cha ris linearis sowie Kageneckia oblonga (Rosaceae)und Lithrea caustica (Anacardiaceae). Die letztge-nannte Art kann nach Blattkontakt bei disponiertenPersonen eine schwere Allergie auslösen; das Aller-gen ist Katechol (Kalergis & al. 1997). An be sondersflachgründigen nordexponierten Hängen kommenSäulenkakteen (Echinopsis chiloensis), die Puya-Arten P. chilensis, P. berteroana und P. coerulea sowieDornsträucher wie Retanilla trinervia, ein malako-phyller Xerophyt mit Blattabwurf in der sommer-
316 5 Die warm-gemäßigte (subtropische) Zone5
1N
a
S2 3 4 5 6 7 8
b
1 2 3 4 5 6
Abb. 5-27 a: Schematische Abfolge der Klima- und Vegetationszonen in Mittelchile vom Nordrand der Atacama (ca. 27° S; arid;1) bis zum Valdivianischen Lorbeerwald (ca. 39° S; perhumid; 8; aus Di Castri 1973, verändert; reproduziert mit Genehmigung vonSpringer Science+Business Media). 2 = Sukkulenten-Halbwüste, 3 = Trockengebüsch mit Sukkulenten, 4 = niedriges Hartlaubge-büsch (Matorral), 5 = hohes Hartlaubgebüsch (Matorral), 6 = Hartlaubwald, 7 = Sommer-Lorbeerwald. 1–3 gehören zu den tropisch-subtropischen Trockengebieten mit Winterregen, bei 7 und 8 handelt es sich um saisonale und immergrüne Lorbeerwälder dernemoralen Zone. b: Vegetationsprofil durch ein Tal in Mittelchile (aus Di Castri 1973, verändert; reproduziert mit Genehmigung von Springer Science+Business Media). Links Nord-, rechts Südhang. 1 = erodierter Oberhang mit Halbwüsten-Vegetation, 2 = Trockengebüsch mit Echi-nopsis chiloensis und Puya-Arten, 3 = extrazonaler Lorbeerwald mit Beilschmiedia miersii und Persea lingue, 4 = Hartlaubwald mitCryptocarya alba u. a., 5 = hohes Hartlaubgebüsch (Matorral), 6 = niedriges Hartlaubgebüsch (Matorral) mit dem regengrünen Dorn-strauch Retanilla trinervia (Rhamnaceae).
lichen Trockenzeit, und Rutensträucher wie Re -tanilla ephedra (Rhamnaceae) und Ephedra chilensisvor (Abb. 5-28b).
Mittelchile ist das einzige der fünf Winterregenge-biete, aus dem keine Wildfeuer bekannt sind. Dennin der gefährdeten Jahreszeit, also im trockenenSommer, können sich im Gegensatz zu den anderenmediterran geprägten Räumen kaum Gewitter bil-den, weil die Anden das Eindringen feuchter Luft-massen von Osten her verhindern (Armesto et al.2007). Deshalb fehlt in den chilenischen Hartlaubge-bieten die für winterfeuchte Subtropen sonst so typi-sche und häufige Erscheinung, nämlich eine durchHitze oder Rauch aktivierbare Kronen- bzw. Boden-samenbank (Montenegro & al. 2004). Während sichalso in Kalifornien die Mehrzahl der Chaparral-Sträucher nach dem Brand aus der Samenbank rege-neriert, sind viele Matorral-Sträucher resprouters. Dasich die Fähigkeit, nach Störung aus Wurzelsystem,

Stammbasis oder Stamm wieder auszutreiben, durchBeweidung oder Massenbewegung (etwa im Zu sam -menhang mit Vulkanausbrüchen) entwickelt habenkann, ist die große Zahl von resprouters im chileni-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 317
schen Matorral kein Beleg für Wildfeuer. Deren Feh-len drückt sich auch dadurch aus, dass vorwiegendnicht-heimische Arten der Feldschicht von denanthropogenen Bränden profitieren, nicht dagegendie einheimischen Kräuter und Gräser (Sax 2002).
Die Wälder sind zum größten Teil durch land-und forstwirtschaftliche Nutzung seit der Kolonisa-tion des Landes durch die Europäer verschwunden.Das Klima und die (verglichen mit dem trockenenNorden) günstigeren Böden ließen im Winterregen-gebiet von Chile eine intensive Bodennutzung zu,während der Süden (südlich des Rio Biobio) kaumzugänglich und von Indianerstämmen besiedelt war.Somit konzentrierten sich Besiedlung und Nutzungauf Mittelchile. Während auf den tiefgründigen allu-vialen Böden permanenter Ackerbau betrieben wird,entwickelte sich sonst anstelle der Hartlaubwälderund des Matorral durch Beweidung eine savannenar-tige Vegetation, die von Acacia caven und einer vor-wiegend aus Europa stammenden Feldschicht gebil-det wird (Abb. 5-29). Hier dominieren einjährigeGräser und Kräuter wie Bromus mollis, Avena bar-bata, Lolium multiflorum, Carduus pycnocephalusu. v. a. (Ovalle & al. 1996), alle aus der mediterranenFlorenregion. Diese als Espinales bezeichneten an -thropogenen „Savannen“ (die mit einer echten Sa -vanne der sommerfeuchten Tropen nichts zu tunhaben) nehmen in Mittelchile rund 2 Mio. ha ein.Acacia caven ist eine regengrüne Art, die in Chileunter dem Einfluss des Winterregenklimas in derkühleren Jahreszeit Blätter trägt und im trockenenSommer je nach Dauer der Trockenzeit das Laubbehält oder abwirft (fakultativer Laubabwurf). Siestammt vermutlich aus dem Chaco östlich derAnden, also aus einem tropisch-subtropischen Tro-ckengebiet mit Sommerregen, und gilt in Chile als
a
b
c
Abb. 5-28 Beispiele für Hartlaubwälder und -gebüsche ausMittelchile. a = Die namengebende Baumart der Klasse Lith-raeo-Cryptocaryetea, Cryptocarya alba (Lauraceae); b = Mator-ral aus Retanilla trinervia, Rhamnaceae, mit dem SäulenkaktusEchinopsis chiloensis und verschiedenen Puya-Arten (Bromeli-aceae); c = Palmenhain aus Jubaea chilensis, im VordergrundRetanilla trinervia. Alle Aufnahmen stammen aus dem National-park La Campana.
Abb. 5-29 Weidelandschaft mit Acacia caven in Mittelchile(Foto F. Klötzli).

nicht-heimisch (Aronson 1992). Die N2-fixierendeMimosacee erlaubt je nach Standort ein recht nach-haltiges Landnutzungssystem, nämlich permanenteBeweidung mit Rindern und Schafen in ebener Lageoder ein Rotationssystem aus Ackerbau (Weizen)und Beweidung in besser drainiertem hügeligemGelände (Ovalle & al. 2006). Acacia caven wird tradi-tionell für die Gewinnung von Feuerholz und zurHerstellung von Holzkohle regelmäßig abgeschlagen,treibt aber rasch wieder aus.
5.3.2.5 Das südafrikanische Winterregengebiet
Das kleinste der fünf Winterregengebiete liegt an derSüdspitze Afrikas; es umfasst rund 90.000 km2 undgehört zur kapensischen Florenregion (s. Abschn.1.2.2; Abb. 5-30). Seine natürliche Vegetation ist einimmergrünes Hartlaubgebüsch mit hohem Anteil anerikoiden Sträuchern und Kleinsträuchern (Cowling1992, Cowling et al. 1997a, Rebelo et al. 2006), beste-hend zu zwei Dritteln aus Fynbos auf den im Gebietvorherrschenden nährstoffarmen Sandböden und zuknapp einem Drittel aus Renosterveld auf tonreichenBöden; dazu kommen rund 3.000 km2 Buschland aufkarbonatreichen Sandböden in der Küstenebene ent-lang des Indischen Ozeans (Western Strandveld).Wälder fehlen fast völlig, sieht man von flächenhaftunbedeutenden Vorkommen einzelner, bis zu 10 mhoher Bestände aus der Proteacee Leucadendronargenteum (silvertree) ab. Nordöstlich von Kapstadtwächst vereinzelt die Reliktkonifere Widdringtoniawallichii (Cupressaceae). Das gesamte Gebiet ist vonbis zu 2.000 m hohen Gebirgen durchzogen und des-halb klimatisch ziemlich heterogen. So kommt es zuextrazonalen Vorkommen von Zwergstrauch-Halb-wüsten (wie im Fall der Little Karoo) und von lauro-phyllen afromontanen Wäldern um Knysna und Port
318 5 Die warm-gemäßigte (subtropische) Zone5
Elizabeth (im Einflussbereich der Steigungsregen; s.Abschn. 5.2.2.6) sowie am Grund tief eingeschnitte-ner Täler (Abb. 5-30).
Der Fynbos hat seinen Namen von den zahlrei-chen erikoiden (feinblättrigen) Gehölzen, die nebenden breitblättrigen Sträuchern der Proteaceae, vorallem aus den Gattungen Leucadendron und Protea,die Physiognomie dieses immergrünen, feuergepräg-ten Hartlaubgebüschs bestimmen. Erikoide Blätterbesitzen nicht nur die Vertreter der Ericaceen, son-dern auch zahlreicher anderer Familien wie Astera-ceae, Rhamnaceae, Thymelaeaceae und Rutaceaesowie die endemischen und subendemischen Bruni-aceae, Pennaeaceae und Grubbiaceae (Abb. 5-31).Bedornte Kleinsträucher aus den Gattungen Cliffor-tia (Rosaceae) und Aspalathus (Fabaceae) sowiehalbsukkulente Halbsträucher wie Pelargonium(Geraniaceae) sind häufig. In diese Matrix aus 1–4 mhohen Gebüschen, gelegentlich überragt von Proteanitida, einem bis zu 5 m hohen Kleinbaum, mischensich die an Juncus-Arten erinnernden Horste derGondwana-Familie Restionaceae sowie Gräser undCyperaceen. Im Frühjahr und nach Feuer entwickelnsich zahlreiche Geophyten aus den Familien Irida-ceae (wie z. B. Freesia, Gladiolus, Ixia), Amaryllida-ceae, Asphodelaceae, Orchidaceae und Asparagaceae(Ornithogalum). Artenzusammensetzung und Struk-tur des Fynbos unterscheiden sich nach Standort undgeographischer Lage (Campbell & Werger 1988); sowächst z. B. die von Poaceen („Grassy fynbos“) oderAsteraceen dominierte Ausbildung („Asteraceousfynbos“) auf flachgründigen Böden in eher trocke-nen Gebieten, während man den von Proteaceenbeherrschten Typus („Proteoid fynbos“) auf Kollu-vien am Hangfuß der Gebirgszüge findet (5-32a, b).
Das Renosterveld ist nach der dominierendenAsteracee Dicerothamnus rhinocerotis benannt, demRenosterbos, einem 1–2 m hohen, graugrünen Strauch
CapeTown
Cape Agulhas
Breede River
26°O
18°O
34°S
22°O35°S
32°S
Fynbos
Renosterveld
Karroid shrubland
Afromontane forest
Thicket
Fynbos/thicket-Mosaik
Abb. 5-30 Vegetationsgliederunginnerhalb der kapensischen Florenre-
gion (aus Richardson et al. 1995; repro-duziert mit Genehmigung von SpringerScience+Business Media). Außer Fyn-
bos, Renosterveld und Western Strand-veld (= „Fynbos/thicket“-Mosaik in der
Karte) kommen Zwergstrauch- undSukkulenten-Halbwüsten (= Karroid
shrubland) in intramontanen Beckenla-gen („Little Karoo“) sowie im Nordenund Nordwesten des Gebiets vor; diebereits immerfeuchten Küstengebieteum Knysna werden von afromontanen(laurophyllen) Wäldern eingenommen.

mit cupressoiden (schuppenförmigen) Blättern (Abb.5-32c). Die Vegetation ist immergrün und von fre-quenten Feuerereignissen geprägt (im Durchschnittalle drei bis fünf Jahre). Proteaceen und Restionaceentreten zurück. Wegen der vergleichsweise fruchtba-ren Böden in der ansonsten nährstoffarmen Kapre-gion wurde das Renosterveld in den vergangenen 100Jahren vor allem in den Küstenebenen nahezu voll-ständig in Ackerflächen umgewandelt. Kaum vonFeuer geprägt ist die als Western Strandveld be zeich-nete Vegetation aus niedrigen hartlaubigen Gehöl-zen. Sukkulente wie Aloe, Ruschia, Mesembryanthe-mum sind häufig, Proteaceen fehlen.
In Abschn. 5.3.1.3 haben wir schon auf die hohenArtenzahlen und den außerordentlichen Endemiten-reichtum vieler Familien und Gattungen hingewiesen(Goldblatt & Manning 2002). Die in Tab. 5-8 aufge-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 319
führten 15 Familien decken rund 70 % der Kapfloraab. Die artenreichsten Gattungen sind neben Erica(657 Arten) Aspalathus (Fabaceae; 272 Arten; unterihnen der als Tee verwendete Rooibos A. linearis),Pelargonium (Geraniaceae; 148 Arten) und Agathos -ma (Rutaceae; 143 Arten). Insgesamt umfassen 13Gattungen mehr als 100 Arten. Die meisten dieserartenreichen Gattungen einschließlich Protea undLeucadendron (außer Pelargonium) haben denSchwerpunkt ihrer Verbreitung auf Sandböden undim Bergland. Obwohl der Beginn der Radiation beivielen Abstammungslinien in das mittlere Tertiärzurückreicht, ist die Mehrzahl der Arten, wie im Fallder Restionaceae, erst im Pliozän und im Pleistozänentstanden, möglicherweise als Reaktion auf das
Tab. 5-8 Artenzahl und Endemitenanteil der 15 größten Fami-lien in der kapensischen Florenregion (überwiegend, aber nichtausschließlich Fynbos, Renosterveld und Coastal Strandveld),aus Goldblatt & Manning (2002).
Familie Artenzahl Zahl der (Anzahl Gattungen endemisch) (Anzahl endemisch)
Asteraceae 1.036 (655) 121 (33)
Fabaceae1 761 (629) 37 (6)
Iridaceae 677 (540) 28 (6)
Aizoaceae 659 (524) 76 (18)
Ericaceae 657 (637) 1 (0)
Scrophulariaceae 414 (297) 33 (7)
Proteaceae 329 (319) 14 (9)
Restionaceae 318 (294) 19 (10)
Rutaceae 273 (257) 15 (6)
Orchidaceae 227 (138) 25 (2)
Poaceae 207 (80) 61 (3)
Cyperaceae 206 (101) 29 (4)
Hyacinthaceae2 191 (83) 14 (0)
Campanulaceae 183 (140) 13 (6)
Asphodelaceae 157 (81) 8 (0)
1 Einschließlich Caesalpiniaceae und Mimosaceae; 2 heute zu Asparaga-ceae.1 Nach Ozenda (1975), Quézel (1985), Quézel & Médail 2003,ergänzt.
1
45 6
7
23
Abb. 5-31 Einige Arten mit schmalen oder erikoiden Blätternaus dem südafrikanischen Fynbos. 1 = Gnidia pinifolia, Thyme-laeaceae; 2 = Leucadendron levisanus, Proteaceae; 3 = Berze-lia abrotanoides, Bruniaceae; 4 = Diosma oppositifolia, Ruta-ceae; 5 = Phylica sp. (Rhamnaceae); 6 = Coleonema album,Rutaceae; 7 = Agathosma capitata, Rutaceae. Zeichnungen vonR. Anheisser, aus Walter (1968).

Winterregenklima in einer isolationsfördernden ge -birgigen Umwelt (Linder & Hardy 2004, Linder2005).
Die Diversität physiognomischer Pflanzenfunk-tionstypen (Anzahl und Anteil am Gesamtspektrum)ist im Vergleich mit den übrigen Winterregengebie-
320 5 Die warm-gemäßigte (subtropische) Zone5
ten gering. Klein- und Zwergsträucher dominierenmit 53,3 % (Chile: 17,8 %, Kalifornien: 11,0 %;Arroyo et al. 1995, Goldblatt & Manning 2002). Vondiesen regenerieren sich fast alle nach Feuer ausSamen (reseeders). Nur wenige sind resprouters, wieder Kleinbaum Protea nitida, der nach Feuer amStamm wieder austreibt. Viele Vertreter der Protea-ceae öffnen ihre Fruchtstände (Zapfen) erst bei Feuer(serotiny) und entlassen endospermreiche Samen,deren Proteinvorräte die Keimung auf den nährstoff-armen Böden erleichtert. Die Samen der erikoidenSträucher, aber auch einiger Proteaceen sind häufigmyrmekochor; sie tragen ein zucker- und ölhaltigesAnhängsel, also ein Elaiosom, und werden deshalbvon Ameisen in ihre Nester verschleppt. Dort bildensie eine persistente Samenbank, die durch Rauchaktiviert wird. Möglicherweise ist Myrmekochorieein Selektionsvorteil für Pflanzen armer Standorte(Cowling et al. 1994): Da viele Arten der Proteaceennur wenige potenziell keimfähige Samen erzeugen,müssen diese vor Fressfeinden geschützt werden. Dasgelingt am besten, indem sie rasch von Ameisen fort-geschafft und vergraben werden.
Alle Fynbos-Gehölze sind immergrün; Malako-phylle fehlen vollständig. Lediglich auf den besserenBöden (Renosterveld, Strandveld) gibt es vereinzeltregengrüne Arten. Wie in anderen saisonal trocke-nen Gebieten mit nährstoffarmen Böden (Savannenund Wälder der sommerfeuchten Tropen) sind somitauch in der Kapregion (und in Australien) laubab-werfende Gehölze nährstoffökologisch benachteiligt.Beide Winterregengebiete stellen deshalb gute Bei-spiele für die in Kasten 1-7 diskutierte Hypothese dar,dass Sklerophyllie eher auf Nährstoffarmut als aufAridität zurückzuführen ist.
Der Anteil der Therophyten an der Flora der Kap-region ist mit 6,7 % (= 609 Arten) sehr gering, imGegensatz zu Chile (16 %) und Kalifornien (30 %),wo Annuelle im ersten Jahr nach einem Feuerereignisreichlich blühen und fruchten. Im Fynbos wird dieseRolle von den Geophyten (den „fire lilies“, wie z. B.der Amaryllidacee Cyrtanthus, der Iridaceaee Watso-nia und der Asphodelacee Kniphofia) übernommen.Diese sind mit über 1.300 Arten in der Flora der Kap-region vertreten (= mehr als 17 %); davon entfällt dieHälfte (662 Arten) auf die Iridaceae.
Landnutzung: Die südafrikanischen Hartlaubgebü-sche wurden in der Vergangenheit ähnlich wie inanderen Winterregengebieten durch Umwandlung inAckerflächen (vorwiegend Renosterveld, aber auchFynbos in tieferen Lagen auf besseren Böden), durchAufforstung mit nicht-heimischen Sippen (Pinus
a
b
c
Abb. 5-32 Beispiele für Hartlaubgebüsche der Kapregion inSüdafrika. a = montaner Fynbos mit Leucadendron eucalyptifo-lium, Proteaceae, mit Resten afromontaner (laurophyller) Wäl-der im feuchten, feuerfreien Talgrund; b = Leucadendron euca-lyptifolium mit gelben Hochblättern; c = Renosterveld mitDicerothamnus rhinocerotis, Asteraceae (dominant) und begin-nender Einwanderung von Pinus radiata. Alle Bilder aus demOuteniqua-Gebirge.

radiata, diverse Eucalyptus-Arten) sowie durch spon-tane Invasion und Ausbreitung einst als Forst- oderZierpflanzen eingeführter oder eingeschleppter Taxa(wie verschiedene Hakea- und Acacia-Arten ausAustralien) dezimiert (Rebelo et al. 2006). So durch-dringen und überwachsen Pinus radiata und Hakeasericea (Proteaceae) den Fynbos auf Sandböden inner-halb von zwei oder drei Feuerzyklen (< 50 Jahre), weildie Invasoren als hochwüchsige reseeders mit ihremkurzen Jugendstadium und den endo spermreichen,anemochoren Samen einen Konkurrenzvorteil ge gen-über den einheimischen Proteaceen haben (Richard-son et al. 1997). Schließlich bilden beide Bäume einhohes, undurchdringliches Ge strüpp, das sich durchdie heißen Kronenfeuer selbst stabilisiert und dasAufkommen von Fynbos-Arten verhindert. Der Auf-wand, den man betreiben muss, um diesen Prozessrückgängig zu machen, ist groß: Die Pinus-Hakea-Bestände werden gefällt; Stämme und Zweige bleibenzwölf bis 18 Monate liegen, bis die Samen ausgefallensind und gekeimt haben. Dann wird das gesamteorganische Material abgebrannt, um alle Sämlinge zuvernichten (Richardson et al. 1997).
5.3.2.6 Die australischen Winterregengebiete
Trotz mancher Ähnlichkeit mit dem Winterregenge-biet Südafrikas (arme Böden, hohe Feuerfrequenz,großer Endemitenanteil, Dominanz der Proteaceae inder Strauchschicht u. a.; s. Tab. 5-4) unterscheidensich beide Gebiete klimatisch und pflanzengeogra-phisch beträchtlich (Beard 1990, Hobbs et al. 1995):So hebt sich das CSb-Klima nach Köppen-Trewarthain Süd- und Südwestaustralien durch Jahresnieder-schläge bis über 1.200 mm von den übrigen winter-feuchten Subtropen deutlich ab, zumal auch die Som-mer nicht niederschlagsfrei sind (Abb. 5-33). Da dasWinterregenklima in Australien erst seit rund 2,5Mio. Jahren existiert und zwischenzeitlich immerwieder feucht-warme Perioden aufgetreten sind(Lamont et al. 1984), ist von den pliozänen, Eucalyp-tus-dominerten Wäldern (die wegen der Nährstoff -armut der Böden ohnehin schon hartlaubig waren)mehr übrig geblieben als andernorts. Das gilt auch füralle skleromorphen Arten und Gattungen (Tab. 5-9).
In Südwestaustralien kann man nach der Höhedes Jahresniederschlags und in Abhängigkeit von denBodenbedingungen (flachgründig oder tiefgründig,nur mäßig oder extrem nährstoffarm) folgende For-mationen unterscheiden (Beadle 1981; Abb. 5-33): 1. Bei Jahresniederschlägen von mehr als 1.000 mm
wachsen auf tiefgründigen Sand- und jungen Ver-
witterungsböden Wälder aus Eucalyptus diversico-lor, einem bis zu 80 m hoch werdenden Baum, dendie Aborigines „Karri“ nennen (Abb. 5-34a). Im„Valley of the Giants“ bei Pemberton wurde dasKronendach mittels Fußgängerbrücken für Tou-risten zugänglich gemacht, sodass man bequemden von Kronenfeuern verursachten büschelför-migen Wiederaustrieb sehen kann (Abb. 5-17d).Die Feuerereignisse treten im Abstand von 50 bis100 Jahren selbst in den niederschlagsreichstenGebieten Südwestaustraliens auf. E. diversicolor istein Beispiel für Eukalypten mit dicker, einigerma-ßen brandgeschützter Borke; er regeneriert sichaus Adventivknospen im Stamm- und Kronenbe-reich, ist also ein resprouter. Karri-Wälder gehörenneben den nemoralen Eucalyptus regnans- (Tas-manien) und Sequoiadendron giganteum-Bestän-den (Sierra Nevada, Kalifornien) zu den höchst-wüchsigen Wäldern der Erde (s. Kasten 6-4). Derreichliche Unterwuchs besteht aus Grasbäumenwie Xanthorrhoea preissii (Xanthorrhoeaceae) undeiner Reihe von endemischen immergrünenSträuchern, unter ihnen Acacia pulchella, eine derwenigen australischen Acacia-Arten, die keinePhyllodien, sondern gefiederte Blätter aufweist,Spyridium globulosum (Rhamnaceae) und dieintensiv blau blühende Fabacee Hovea elliptica(Abb. 5-34b).
2. Innerhalb dieser regenreichen Zone Südwestaus-traliens mit mehr als 800 mm Jahresniederschlagfolgen landeinwärts sowie auf trockenen nähr-stoffarmen Böden mehr oder weniger offene, mitbis zu 40 m deutlich niedrigere Wälder, in denenbesonders häufig Eucalyptus marginata („Jar-rah“) auftritt (Abb. 5-34c; Dell et al. 1989). IhrUnterwuchs besteht aus Proteaceen mit harten,dornigen Blättern wie z. B. den Gattungen Petro-phile und Dryandra. Hier kommt auch derimmergrüne, kleinblättrige Strauch Gastrolobiumparvifolium (Fabaceae: Abb. 5-34g) vor, dessenBlätter Natrium-Monofluorazetat enthalten. DieSubstanz ist giftig für nicht-heimische Säugetiereeinschließlich des Menschen (3 mg ge nügen alsletale Dosis) und verursachte während der Kolo-nisationszeit im 19. Jahrhundert er hebliche Aus-fälle unter den eingeführten Haustierherden(Chandler et al. 2002). Heute wird das Mittelunter dem Namen „Teneighty“ („1080“) synthe-tisch hergestellt und zur Bekämpfung von invasi-ven Säugern wie Wildschweinen eingesetzt.
3. Zwischen 300 und 800 mm Niederschlag folgenoffene Wälder aus zahlreichen Eucalyptus-Arten,unter anderem aus E. wandoo, sowie ein Mosaik
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 321
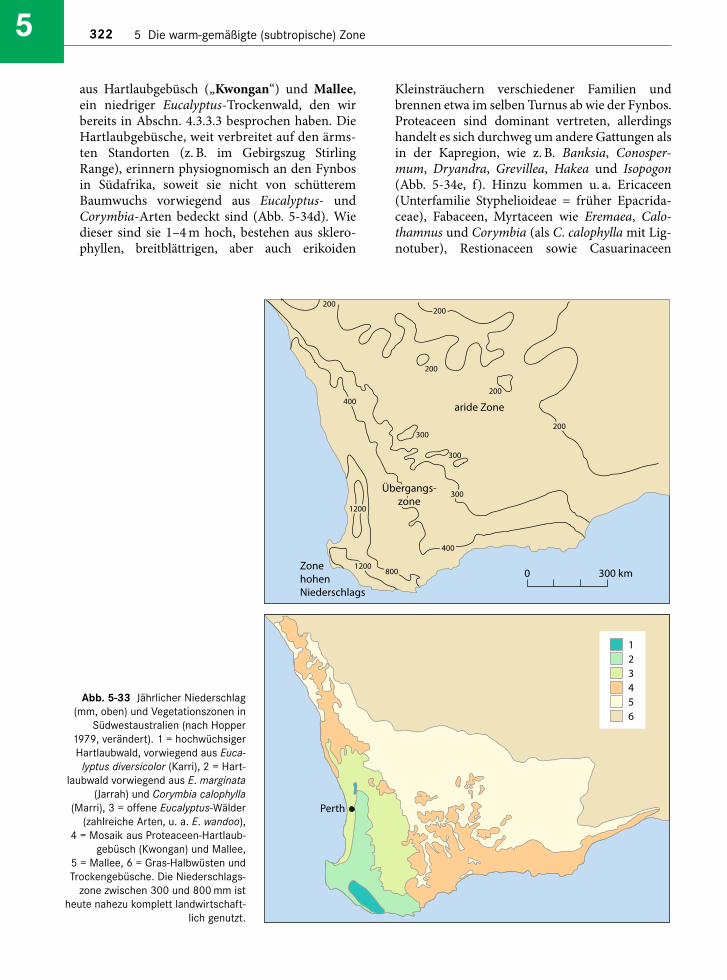
aus Hartlaubgebüsch („Kwongan“) und Mallee,ein niedriger Eucalyptus-Trockenwald, den wirbereits in Abschn. 4.3.3.3 besprochen haben. DieHartlaubgebüsche, weit verbreitet auf den ärms -ten Standorten (z. B. im Gebirgszug StirlingRange), erinnern physiognomisch an den Fynbosin Südafrika, soweit sie nicht von schütteremBaumwuchs vorwiegend aus Eucalyptus- undCorymbia-Arten bedeckt sind (Abb. 5-34d). Wiedieser sind sie 1–4 m hoch, bestehen aus sklero-phyllen, breitblättrigen, aber auch erikoiden
Kleinsträuchern verschiedener Familien undbrennen etwa im selben Turnus ab wie der Fynbos.Proteaceen sind dominant vertreten, allerdingshandelt es sich durchweg um andere Gattungen alsin der Kapregion, wie z. B. Banksia, Conosper-mum, Dryandra, Grevillea, Hakea und Isopogon(Abb. 5-34e, f). Hinzu kommen u. a. Ericaceen(Unterfamilie Styphelioideae = früher Epacrida-ceae), Fabaceen, Myrtaceen wie Eremaea, Calo-thamnus und Corymbia (als C. calophylla mit Lig-notuber), Restionaceen sowie Casuarinaceen
322 5 Die warm-gemäßigte (subtropische) Zone5
200
200
200
300
300
300
400
400
800
1200
1200
200
200
300
400
400
800
1200
1200
200
200
300 km0
aride Zone
Übergangs-zone
Übergangs-zone
ZonehohenNiederschlags
Perth
123456
Abb. 5-33 Jährlicher Niederschlag(mm, oben) und Vegetationszonen in
Südwestaustralien (nach Hopper1979, verändert). 1 = hochwüchsigerHartlaubwald, vorwiegend aus Euca-
lyptus diversicolor (Karri), 2 = Hart-laubwald vorwiegend aus E. marginata
(Jarrah) und Corymbia calophylla(Marri), 3 = offene Eucalyptus-Wälder
(zahlreiche Arten, u. a. E. wandoo), 4 = Mosaik aus Proteaceen-Hartlaub-
gebüsch (Kwongan) und Mallee, 5 = Mallee, 6 = Gras-Halbwüsten undTrockengebüsche. Die Niederschlags-
zone zwischen 300 und 800 mm istheute nahezu komplett landwirtschaft-
lich genutzt.
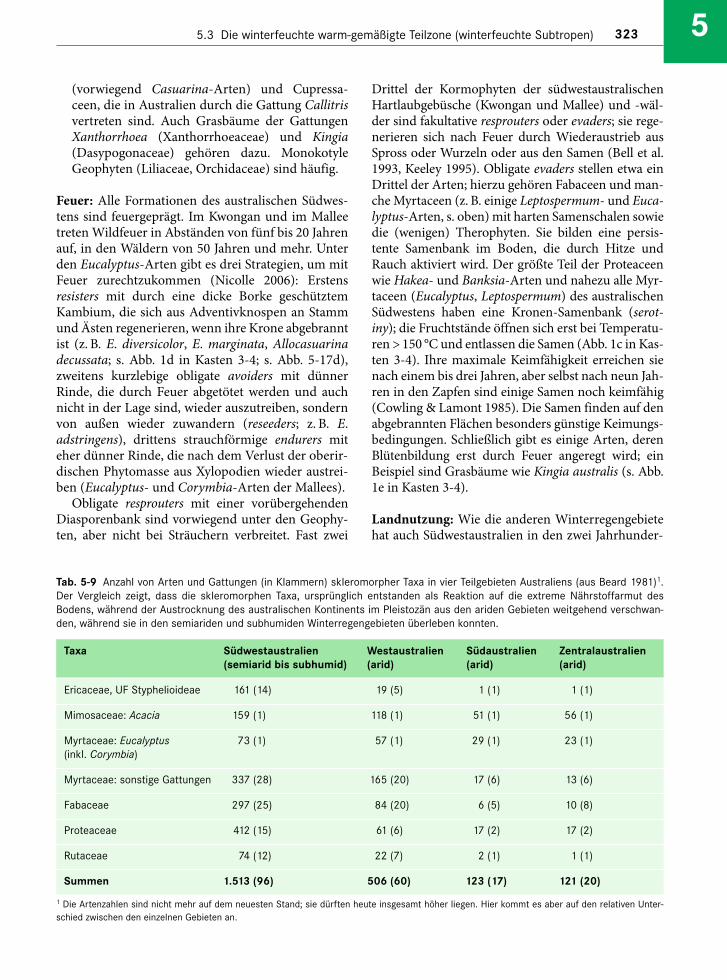
(vorwiegend Casuarina-Arten) und Cupressa-ceen, die in Australien durch die Gattung Callitrisvertreten sind. Auch Grasbäume der GattungenXanthorrhoea (Xanthorrhoeaceae) und Kingia(Dasypogonaceae) gehören dazu. MonokotyleGeophyten (Liliaceae, Orchidaceae) sind häufig.
Feuer: Alle Formationen des australischen Südwes-tens sind feuergeprägt. Im Kwongan und im Malleetreten Wildfeuer in Abständen von fünf bis 20 Jahrenauf, in den Wäldern von 50 Jahren und mehr. Unterden Eucalyptus-Arten gibt es drei Strategien, um mitFeuer zurechtzukommen (Nicolle 2006): Erstensresisters mit durch eine dicke Borke geschütztemKambium, die sich aus Adventivknospen an Stammund Ästen regenerieren, wenn ihre Krone abgebranntist (z. B. E. diversicolor, E. marginata, Allocasuarinadecussata; s. Abb. 1d in Kasten 3-4; s. Abb. 5-17d),zweitens kurzlebige obligate avoiders mit dünnerRinde, die durch Feuer abgetötet werden und auchnicht in der Lage sind, wieder auszutreiben, sondernvon außen wieder zuwandern (reseeders; z. B. E.adstringens), drittens strauchförmige endurers miteher dünner Rinde, die nach dem Verlust der oberir-dischen Phytomasse aus Xylopodien wieder austrei-ben (Eucalyptus- und Corymbia-Arten der Mallees).
Obligate resprouters mit einer vorübergehendenDiasporenbank sind vorwiegend unter den Geophy-ten, aber nicht bei Sträuchern verbreitet. Fast zwei
Drittel der Kormophyten der südwestaustralischenHartlaubgebüsche (Kwongan und Mallee) und -wäl-der sind fakultative resprouters oder evaders; sie rege-nerieren sich nach Feuer durch Wiederaustrieb ausSpross oder Wurzeln oder aus den Samen (Bell et al.1993, Keeley 1995). Obligate evaders stellen etwa einDrittel der Arten; hierzu gehören Fabaceen und man-che Myrtaceen (z. B. einige Leptospermum- und Eu ca-lyptus-Arten, s. oben) mit harten Samenschalen so wiedie (wenigen) Therophyten. Sie bilden eine per sis-tente Samenbank im Boden, die durch Hitze undRauch aktiviert wird. Der größte Teil der Proteaceenwie Hakea- und Banksia-Arten und nahezu alle Myr-taceen (Eucalyptus, Leptospermum) des australischenSüdwestens haben eine Kronen-Samenbank (sero t-iny); die Fruchtstände öffnen sich erst bei Temperatu-ren > 150 °C und entlassen die Samen (Abb. 1c in Kas-ten 3-4). Ihre maximale Keimfähigkeit erreichen sienach einem bis drei Jahren, aber selbst nach neun Jah-ren in den Zapfen sind einige Samen noch keimfähig(Cowling & La mont 1985). Die Samen finden auf denabgebrannten Flächen besonders günstige Keimungs-bedingungen. Schließlich gibt es einige Arten, derenBlütenbildung erst durch Feuer angeregt wird; einBeispiel sind Grasbäume wie Kingia australis (s. Abb.1e in Kasten 3-4).
Landnutzung: Wie die anderen Winterregengebietehat auch Südwestaustralien in den zwei Jahrhunder-
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 323
Tab. 5-9 Anzahl von Arten und Gattungen (in Klammern) skleromorpher Taxa in vier Teilgebieten Australiens (aus Beard 1981)1.Der Vergleich zeigt, dass die skleromorphen Taxa, ursprünglich entstanden als Reaktion auf die extreme Nährstoffarmut desBodens, während der Austrocknung des australischen Kontinents im Pleistozän aus den ariden Gebieten weitgehend verschwan-den, während sie in den semiariden und subhumiden Winterregengebieten überleben konnten.
Taxa Südwestaustralien Westaustralien Südaustralien Zentralaustralien(semiarid bis subhumid) (arid) (arid) (arid)
Ericaceae, UF Styphelioideae 161 (14) 19 (5) 1 (1) 1 (1)
Mimosaceae: Acacia 159 (1) 118 (1) 51 (1) 56 (1)
Myrtaceae: Eucalyptus 73 (1) 57 (1) 29 (1) 23 (1)(inkl. Corymbia)
Myrtaceae: sonstige Gattungen 337 (28) 165 (20) 17 (6) 13 (6)
Fabaceae 297 (25) 84 (20) 6 (5) 10 (8)
Proteaceae 412 (15) 61 (6) 17 (2) 17 (2)
Rutaceae 74 (12) 22 (7) 2 (1) 1 (1)
Summen 1.513 (96) 506 (60) 123 (17) 121 (20)
1 Die Artenzahlen sind nicht mehr auf dem neuesten Stand; sie dürften heute insgesamt höher liegen. Hier kommt es aber auf den relativen Unter-schied zwischen den einzelnen Gebieten an.

ten seit Einwanderung der Europäer einen tiefgrei-fenden Landschaftswandel erlebt (Abb. 5-35). EinTeil des Gebiets, das zwischen der Mallee-Region imOsten und der Southern-Forest-Region im Westenliegt und jährliche Niederschläge zwischen 300 und
800 mm erhält, wurde als „Wheatbelt“ zur Kornkam-mer Australiens (Beard 1990). Die Erschließung die-ser rund 155.000 km2 umfassenden Region begann inder ersten Hälfte des 19. Jahrhunderts und beschleu-nigte sich zu Beginn des 20. Jahrhunderts und noch-
324 5 Die warm-gemäßigte (subtropische) Zone5
gfe
a b
c d
Abb. 5-34 Beispiele für Hartlaubwälder und -gebüsche in Südwestaustralien und einige typische Sippen. a = Karri-Wald aus Euca-lyptus diversicolor (Karri), Valley of the Giants bei Walpole; b = Unterwuchs in einem Karri-Wald mit Xanthorrhoea preissii, Xanthor-rhoeaceae und Hovea elliptica (blau blühend, Fabaceae, bei Walpole); c = Jarrah-Wald aus Eucalyptus marginata mit dem bizarrenStrauch Dryandra nobilis, Proteaceae, und den weißen Stämmen von E. wandoo im Hintergrund (Reservat Dryandra Woodland); d= Kwongan aus verschiedenen Proteaceen (u. a. Hakea conchifolia, rechts) und lockerem Überstand aus Corymbia calophylla, Myr-taceae (Nationalpark Stirling Range). Besonders auffallende Blüten haben u. a. die Proteaceen Isopogon cuneatus (e) und Grevilleaarmigera (f); in Jarrah-Wäldern wächst die Fabacee Gastrolobium parvifolium (g), deren Blätter das für Säugetiere giftige Natrium-Monofluorazetat („Teneighty“) enthalten.

mals nach dem 2. Weltkrieg (Hobbs 1998). Heutewird die Region zu 93 % landwirtschaftlich genutzt,wobei auf etwa vier Fünftel der Nutzfläche Weizen,Gerste und Raps angebaut werden; ein Fünftel dientder Produktion von Lammfleisch und Schafwolle(Wheatbelt Development Commission 2011). Ledig-lich auf 7 % sind Reste der ursprünglichen Vegetationaus Jarrah, Mallee und Kwongan erhalten, von deneneinige in kleineren Schutzgebieten einem Verände-rungsverbot unterliegen.
Mit der Besiedlung durch europäische Einwande-rer und der nahezu vollständigen Umwandlung der
Region in landwirtschaftliche Nutzflächen im Ver-lauf der vergangenen 80 Jahre hat sich eine Reihe vonUmweltproblemen eingestellt. Dazu gehört z. B. dasAuftreten des in Südostasien heimischen OomycetenPhytophthora cinnamomi, der die einheimische Ge -hölzvegetation schädigt (Kasten 5-6). Neben derBoden- und Winderosion auf den großen Acker-schlägen schränkt vor allem die zunehmende Versal-zung der Böden die Nutzbarkeit der Äcker ein undbedroht inzwischen auch die Vegetation der Schutz-gebiete (dry salinity; Clarke et al. 2002): Währendunter transpirationsaktiven Wäldern ein großer Teildes Niederschlagswassers von der Vegetation ver-braucht wird, kommt es unter Ackernutzung zu einerZunahme der Grundwasserneubildung. In Gebietenmit Salzeinwehung aus benachbarten Ozeanen odermit salzhaltigen marinen Sedimenten im Untergrundwerden mit dem verstärkten GrundwasserstromSalze gelöst und reichern sich in Geländemulden an.Während trockenerer Perioden gelangen sie mit demaufsteigenden Wasser an die Bodenoberfläche. Mitt-lerweile dürften rund 10 % der Region von Versal-zung bedroht sein, mit weiter zunehmender Tendenz(Bari & Smettem 2006). Eine Maßnahme, den An -stieg des Grundwassers aufzuhalten, besteht in derKombination von streifen- oder blockförmigen Auf-forstungen tiefwurzelnder, transpirationsintensiverEucalyptus-Arten (Farrington & Salama 1996).Außerdem versucht man, die isolierten Reste der
55.3 Die winterfeuchte warm-gemäßigte Teilzone (winterfeuchte Subtropen) 325
Abb. 5-35 Im „Wheatbelt“ von Südwestaustralien bei Katan-ning (Rapsfelder).
Phytophthora cinnamomiDer Oomycet Phytophthora cinnamomi gehört taxonomischin die Ordnung Peronosporales, die als „Falscher Mehltau“vorwiegend höhere Landpflanzen befallen (Bresinsky et al.2008). Der vermutlich aus Papua-Neuguinea stammendepathogene Pilz kann jahrelang saprophytisch im Bodenleben und bildet unter günstigen Bedingungen (nach Nieder-schlägen, wenn die Bodenporen wassergefüllt sind) mit zweiGeißeln versehene bewegliche Zoosporen, die in der Boden-lösung an die Wurzeln eines potenziellen Wirtes gelangen.Dort bilden sie Cysten, die in die Interzellularen keimen undzu einem Mycel im Pflanzenkörper heranwachsen.
In den 20er Jahren des 20. Jahrhunderts wurde Phytoph-thora cinnamomi unabsichtlich mit infizierter Baumschul-ware nach Perth eingeschleppt (Hardham 2005). Von dortbreitete sich der Pilz zunächst in die Eucalyptus marginata-Wälder aus und verursachte dort ein als „Jarrah dieback“bekannt gewordenes Waldsterben. Inzwischen ist Phytoph-thora in den Wäldern und Gebüschen Südwestaustraliens
ebenso angekommen wie im Süden (Victoria) und auf Tas-manien. Er befällt bevorzugt immergrüne Gehölze und rich-tet nicht nur in Forstplantagen (z. B. Pinus-Aufforstungen),sondern auch in natürlichen Ökosystemen beträchtlicheSchäden an. So sind 85 % aller in Australien vorkommendenProteaceen anfällig für die Pilzinfektion, ebenso wie vieleEucalyptus-Arten. Im Stirling-Range-Nationalpark, einem dergroßen Kwongan- und Mallee-Schutzgebiete Südwestaustra-liens, dürften nach vorsichtiger Schätzung etwa 2.000 Arteninfektionsgefährdet sein. Der Pilz lässt sich zwar durch spe-zielle Fungizide bekämpfen; da deren Anwendung in Schutz-gebieten allerdings wegen der unkontrollierbaren Auswir-kungen auf andere Mikroorganismen problematisch ist,bleiben nur die Isolation der befallenen Flächen (Betretungs-verbot) sowie die Desinfektion von Fahrzeugen und Wander-stiefeln vor und nach dem Besuch gefährdeter Gebiete alsvorbeugende Maßnahmen übrig (Hardham 2005).
Kasten 5-6

ursprünglichen Vegetation mittels eines Korridorsaus renaturierten Wäldern und Heiden zu verknüp-fen (Projekt Gondwana Link; Fischer et al. 2008).
5.4 Azonale Vegetation
5.4.1 Feuchtgebiete
5.4.1.1 Begriffe
Feuchtgebiete sind Ökosysteme, die regelmäßig ent-weder von Fremdwasser überflutet oder von Grund-wasser überstaut werden, und zwar mit einer Fre-quenz und Dauer, die zur Dominanz einer anwassergesättigte Bodenbedingungen angepasstenPflanzendecke führt (vgl. Joosten & Clarke 2002; s.Kasten 6-12 in Abschn. 6.4.2). Die Untergliederungder Feuchtgebiete und die Terminologie der Subty-pen sind international uneinheitlich und umstritten(dto.). Der Einfachheit halber unterscheiden wirSüß- und Salzwasser-Feuchtgebiete; zu den Süßwas-ser-Feuchtgebieten gehören Moore (engl. mires) mitbiogener Sedimentation (Torf, Kalk), Sümpfe (engl.swamps; ohne biogene Sedimentation) und Fluss auen(engl. floodplains; mit Sedimentation von Fremdma-terial); die Vegetation kann aus Wäldern (Auwälderbei Überflutung, Moor- bzw. Sumpfwälder bei Über-stau) oder aus Cyperaceen- bzw. Poaceen-Beständenmit je nach Trophie (verfügbare Nährstoffmenge)unterschiedlicher Bestandshöhe bestehen. Salzwas-ser-Feuchtgebiete (Sedimentation von Fremdmate-rial unter Salzeinfluss) erreichen ihre größte Flächen-ausdehnung an Flachküsten, in Ästuaren undMündungsdeltas; sie sind in den Tropen häufig be -waldet (Mangroven), in den übrigen Klimazonenbaumfrei (Salzmarschen).
Flussauen sind in den Subtropen nicht nur einschmales Band periodisch überfluteter Wälder undSümpfe; vor ihrer Kultivierung begleiteten sie diegroßen Ströme (wie den Mississippi und den Paraná)beidseitig entlang des gefällearmen Unterlaufs undim Mündungsgebiet mit einem bis zu 100 km breitenStreifen. Nordhemisphärisch dominieren sommer-grüne Gattungen wie Salix, Populus, Platanus undQuercus, aber auch hoch spezialisierte Nadelbäumewie Taxodium und Chamaecyparis; südhemisphä-risch sind vorwiegend immergrüne Gehölze aus demtropisch-subtropischen Verwandtschaftskreis cha-rakteristisch.
Verlandungs- und Versumpfungsniedermooresind vor allem in den Küstenebenen und küstenna-hen gefällearmen Niederungen, aber auch inniederschlags- und nebelreichen Gebirgslagen weitverbreitet. So liegen die größten Moore in denimmerfeuchten Subtropen von Nord- und Südame-rika, wie die Everglades in Florida und die Esteros delIberá in Nordargentinien. Sie nahmen vor ihrer Kul-tivierung im 20. Jahrhundert Tausende von Quadrat-kilometern ein und waren weitgehend unzugänglich.Die im natürlichen Zustand verbliebenen Reste,dominiert von Röhrichten aus hochwüchsigen Grä-sern und Cyperaceen wie den Gattungen Cyperus,Scirpus, Cladium, Panicum und Phragmites sowieSumpfwäldern aus Hypoxie-toleranten Bäumen,unterliegen meist strengen Schutzauflagen und sindheute beliebte und bekannte Ziele für an Naturbeob-achtung interessierte Touristen. In Gebirgslagen undbei extremer Nährstoffarmut kommen Moore mitombrotrophem (vom Regenwasser gespeisten) Cha -rakter mit einer Moosdecke aus Vertretern der Gat-tung Sphagnum als Torfbildner vor (s. auch Kasten 6-12 in Abschn. 6.4.2).
In den winterfeuchten Subtropen treten dagegenSüßwasser- gegenüber Salzwasser-Feuchtgebieten zu -rück. Salzverbrackung ist wegen des trockenen Som-mers vor allem in Flussdeltas und entlang der Küsteneine häufige Erscheinung. So findet man Salzmar-schen mit dominanten Chenopodiaceen wie denGattungen Salicornia, Arthrocnemum und salzver-träglichen Gräsern wie Spartina in den Mündungsge-bieten der Flüsse (wie z. B. in der „Camargue“ ge -nannten Landschaft im Rhone-Delta).
5.4.1.2 Süßwasser-Feuchtgebiete
5.4.1.2.1 FlussauenZu den größten Feuchtgebieten der Erde gehört dasMississippi-Becken, das südlich von St. Louis (Mis-souri) beginnt und bis zum Mündungsdelta im Golfvon Mexiko reicht (Fraser & Keddy 2005, Mitsch &Gosselink 2007). Es umfasst eine Fläche von rund108.000 km2 und besteht aus gefällearmen Überflu-tungsebenen, die von den Mäandern des Mississippiund seiner Nebenflüsse geprägt sind und zahlreicheAltwasserarme und Umlaufseen enthalten. Das Ge -biet war vor Beginn der europäischen Besiedlung fastgänzlich von Auen- und Moorwäldern bedeckt (bot-tomland hardwood forests). Diese wurden beschleu-nigt zwischen 1900 und 1970 n. Chr. abgeholzt und inlandwirtschaftliche Nutzflächen überführt. Die ver-bliebenen Waldreste, etwa 20 % ihres ursprünglichen
326 5 Die warm-gemäßigte (subtropische) Zone5
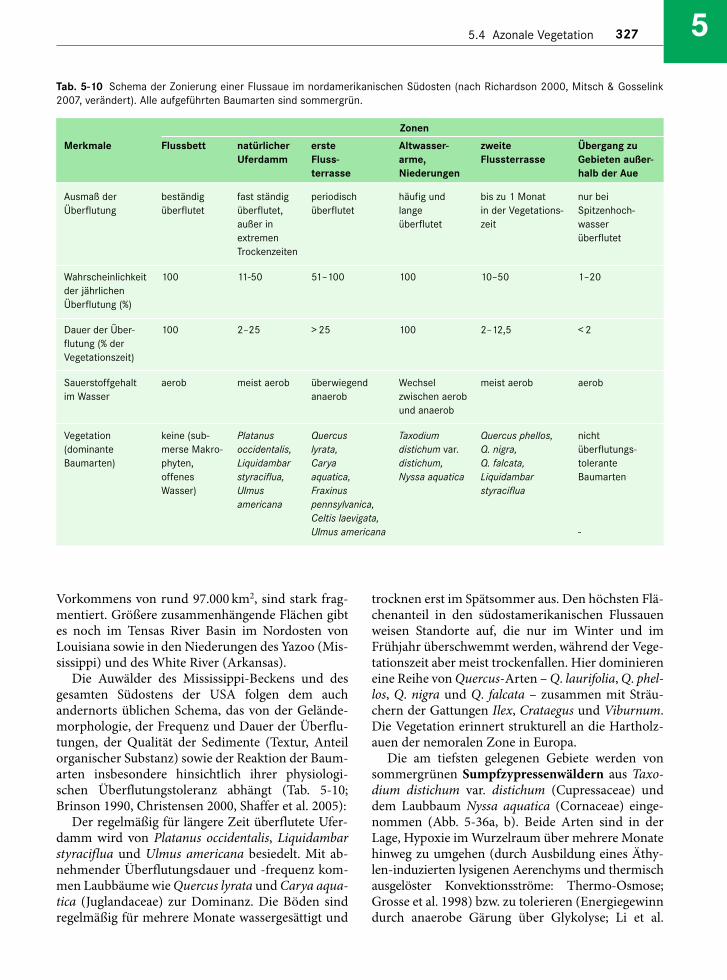
Vorkommens von rund 97.000 km2, sind stark frag-mentiert. Größere zusammenhängende Flächen gibtes noch im Tensas River Basin im Nordosten vonLouisiana sowie in den Niederungen des Yazoo (Mis-sissippi) und des White River (Arkansas).
Die Auwälder des Mississippi-Beckens und desgesamten Südostens der USA folgen dem auchandernorts üblichen Schema, das von der Gelände-morphologie, der Frequenz und Dauer der Überflu-tungen, der Qualität der Sedimente (Textur, Anteilorganischer Substanz) sowie der Reaktion der Baum-arten insbesondere hinsichtlich ihrer physiologi-schen Überflutungstoleranz abhängt (Tab. 5-10;Brinson 1990, Christensen 2000, Shaffer et al. 2005):
Der regelmäßig für längere Zeit überflutete Ufer-damm wird von Platanus occidentalis, Liquidambarstyraciflua und Ulmus americana besiedelt. Mit ab -nehmender Überflutungsdauer und -frequenz kom-men Laubbäume wie Quercus lyrata und Carya aqua-tica (Juglandaceae) zur Dominanz. Die Böden sindregelmäßig für mehrere Monate wassergesättigt und
55.4 Azonale Vegetation 327
trocknen erst im Spätsommer aus. Den höchsten Flä-chenanteil in den südostamerikanischen Flussauenweisen Standorte auf, die nur im Winter und imFrühjahr überschwemmt werden, während der Vege-tationszeit aber meist trockenfallen. Hier dominiereneine Reihe von Quercus-Arten – Q. laurifolia, Q. phel-los, Q. nigra und Q. falcata – zusammen mit Sträu-chern der Gattungen Ilex, Crataegus und Viburnum.Die Vegetation erinnert strukturell an die Hartholz -auen der nemoralen Zone in Europa.
Die am tiefsten gelegenen Gebiete werden vonsommergrünen Sumpfzypressenwäldern aus Taxo-dium distichum var. distichum (Cupressaceae) unddem Laubbaum Nyssa aquatica (Cornaceae) einge-nommen (Abb. 5-36a, b). Beide Arten sind in derLage, Hypoxie im Wurzelraum über mehrere Monatehinweg zu umgehen (durch Ausbildung eines Äthy-len-induzierten lysigenen Aerenchyms und thermischausgelöster Konvektionsströme: Thermo-Os mo se;Grosse et al. 1998) bzw. zu tolerieren (Energiegewinndurch anaerobe Gärung über Glykolyse; Li et al.
Tab. 5-10 Schema der Zonierung einer Flussaue im nordamerikanischen Südosten (nach Richardson 2000, Mitsch & Gosselink2007, verändert). Alle aufgeführten Baumarten sind sommergrün.
Zonen
Merkmale Flussbett natürlicher erste Altwasser- zweite Übergang zu Uferdamm Fluss- arme, Flussterrasse Gebieten außer-
terrasse Niederungen halb der Aue
Ausmaß der beständig fast ständig periodisch häufig und bis zu 1 Monat nur bei Überflutung überflutet überflutet, überflutet lange in der Vegetations- Spitzenhoch-
außer in überflutet zeit wasser extremen überflutetTrockenzeiten
Wahrscheinlichkeit 100 11-50 51–100 100 10–50 1–20der jährlichen Überflutung (%)
Dauer der Über- 100 2–25 > 25 100 2–12,5 < 2 flutung (% der Vegetationszeit)
Sauerstoffgehalt aerob meist aerob überwiegend Wechsel meist aerob aerobim Wasser anaerob zwischen aerob
und anaerob
Vegetation keine (sub- Platanus Quercus Taxodium Quercus phellos, nicht (dominante merse Makro- occidentalis, lyrata, distichum var. Q. nigra, überflutungs-Baumarten) phyten, Liquidambar Carya distichum, Q. falcata, tolerante
offenes styraciflua, aquatica, Nyssa aquatica Liquidambar BaumartenWasser) Ulmus Fraxinus styraciflua
americana pennsylvanica, Celtis laevigata, Ulmus americana -

2010). Ob die meist etwa 1 m hohen Kniewurzeln vonTaxodium zum Gasaustausch beitragen, also ähnlichwie im Fall der Mangroveart Bruguiera gymnorhiza(s. Abb. 2-26) Pneumatophoren sind, ist umstritten(Mitsch & Gosselink 2007, Mitsch et al. 2009); sieerhöhen jedenfalls die Standfestigkeit der Bäume,indem sie das lateral ausgedehnte Wurzelwerk ver-stärken. Sumpfzypressenwälder sind deshalb ziem-lich unempfindlich gegenüber den tropischen Wir-
328 5 Die warm-gemäßigte (subtropische) Zone5
belstürmen der Karibik, wie sich im Fall des Hurri-kans Katrina im Jahr 2005 erneut gezeigt hat (Midd-leton 2009). Taxodium wird bis zu 40 m hoch undkann ein Alter von mehr als 2000 Jahren erreichen.Die Keimung der nautochoren Samen und die Ent-wicklung der Jungpflanzen sind auf die kurzen Zeit-fenster beschränkt, in denen die Wälder nicht über-flutet sind. Das Holz ist weich, aber resistent gegenTermiten und Pilzbefall und war deshalb in der Ver-
b c
d e
a
Abb. 5-36 Beispiele für subtropische Süßwasser-Feuchtgebiete: a, b =Taxodium dis-tichum-Sumpf im Winter (Mississippi-Delta, USA; Foto A. Fidelis) und im Sommer(Everglades; Foto ©Rudy Umans, www.123RF.com); die Kniewurzeln sind in a deut-lich zu sehen; c = Auwald aus Platanus orientalis (Kreta); d = Cladium mariscus ssp.jamaicense-Grundwassermoor, Everglades, Florida (Foto F. Klötzli); e = Sphagnum-Moor auf dem „Planalto“ in Südbrasilien (Rio Grande do Sul) mit Araucaria angusti-folia im Hintergrund.

gangenheit als Bauholz für den Außenbereich, für dieHerstellung von Holzschindeln sowie für die Innen-ausstattung sehr begehrt (Schütt et al. 2004).
Es fällt auf, dass die Bäume der südostamerikani-schen Auwälder im Herbst ihr Laub verlieren, wäh-rend die Wälder außerhalb der Feuchtgebiete über-wiegend immergrün sind (s. Abschn. 5.2.2.4). ImGegensatz zu den tropischen Varzea-Wäldern desAmazonasbeckens, in denen ebenfalls laubabwer-fende Bäume häufig auftreten (s. Abschn. 2.2.3.2),sind sie aber nicht zum Höchststand der Überflutungkahl (was unter Hypoxie-Stress sinnvoll wäre), son-dern unabhängig vom Überflutungszeitraum in derkühlen Jahreszeit. In anderen Gebieten der immer-feuchten Subtropen enthalten die Auwälder nebenlaubabwerfenden auch immergrüne laurophylle Ar -ten, z. B. in Südbrasilien (Budke et al. 2008).
Dieser vor allem nordhemisphärisch verbreitetesaisonale Charakter der Auwälder wird besondersdeutlich in den subtropischen Winterregengebieten:So kommen in Kalifornien verbreitet Salix-, Populus-und Platanus-Arten vor (wie Populus fremontii undPlatanus racemosa), vergesellschaftet mit weiterenLaubbäumen (z. B. Acer negundo). Populus und vorallem die thermophile Gattung Platanus gelten alsRelikte der pliozänen Tertiärflora (Holstein 1984).Wegen ihrer Pioniereigenschaften (wie Anemocho-rie, vegetative Ausbreitung über Wurzelausläufer,Schnellwüchsigkeit) konnten sie die von Sediment-transport und Überflutung geprägten Flussauenbevorzugt besiedeln. Ganz ähnlich ist die Situationim mediterranen Raum, dessen Auwälder auf derIberischen Halbinsel und im Süden Frankreichs ausSalix-Arten, Populus alba, P. nigra und Fraxinusangustifolia, im Osten (Griechenland, Türkei) ausPlatanus orientalis, oft vergesellschaftet mit demimmergrünen Oleander (Nerium oleander) und derlaubabwerfenden Mönchspfeffer Vitex agnus-castus,einer Lamiacee, bestehen (Abb. 5-36c).
5.4.1.2.2 Sümpfe und MooreIm Südosten von Nordamerika sind Feuchtgebietemit Torfbildung in den Küstenebenen der Bundes-staaten North und South Carolina, Georgia und Flo-rida erhalten geblieben. Unter ihnen sind die Ever -glades mit heute noch 3.500 km2 Fläche (im süd-lichen Florida) und die Okefenokee Swamps mit1.770 km2 (an der Grenze zwischen Georgia und Flo-rida) am bekanntesten (Richardson 2000, 2008; Abb.5-36d). In beiden Fällen handelt es sich überwiegendum Versumpfungsniedermoore (zur Terminologie s.Suc cow & Joosten 2001; Kasten 6-12), die vor etwa5.000 bis 6.000 Jahren entstanden, als der ansteigende
Meeresspiegel das Grundwasser in den tief gelegenenflachen Becken anhob. Die Vegetation der Evergladesbestand ursprünglich zu 60 % aus einem Cladiummariscus ssp. jamaicense-(Schneidried-)Röhricht,das den europäischen Einwanderern als unendlicheWeite erschien („never glades“) und von den einhei-mischen Indianerstämmen Gras-See („Pa-hay-okee“)genannt wurde (Richardson & Huvane 2008). Cla-dium ist eine torfbildende Cyperacee karbonatreicherNassböden, die Dominanzbestände bildet und demeklatanten P-Mangel (P-Festlegung als Calcium-Phosphat = Apatit) durch eine höchst effiziente P-Nutzungseffizienz begegnet. Diese Röhrichte, sta-bilisiert durch regelmäßige leichte Wildfeuer, unter-brochen von Bauminseln und zahlreichen Schlenken,Stand- und Fließgewässern (mit einer reichen Fauna),wurden seit den 30er Jahren des 20. Jahrhundertssukzessive entwässert und kultiviert. Mit steigendemP-Eintrag wurden Schneidried- durch Rohrkolben-Röhrichte aus Typha latifolia und T. domingensisersetzt. Heute steht der Südteil als Everglades-Natio-nalpark unter strengem Schutz; der Mittelteil ist alsWasserschutzgebiet ausgewiesen und der Norden istüberwiegend landwirtschaftlich genutzt.
Außer diesen von Grasartigen dominierten Moo-ren kommen in den Okefenokee Swamps, den Ever -glades und in weiteren kleineren Feuchtgebieten desnordamerikanischen Südostens gehölzbestandeneMoore vor, die teilweise sogar ombrotrophen Cha -rakter annehmen können (Hofstetter 1983, Christen-sen 2000). So entstanden aus Verlandungsnieder-mooren in abgeschnittenen Flussarmen ab etwa6.000 Jahren BP schwach aufgewölbte, vorwiegendvon Regenwasser gespeiste Moore, die an Hoch-moore erinnern und wegen dieser Wölbung von denIndianern „Pocosin“ (= „Moor auf einem Hügel“)genannt wurden. Die Pocosins sind von einem un -durchdringlichen Gestrüpp aus überwiegend immer-grünen, mykorrhizierten Sträuchern bewachsen, zudenen Ilex glabra, Cyrilla racemiflora (Cyrillaceae,mit den Ericaceae verwandt), Lyonia lucida (Erica-ceae) und der auch in borealen Mooren Nordameri-kas und Eurasiens verbreitete Kleinstrauch Chamae-daphne calyculata (Ericaceae) gehören. Im Zentrumdieser Moore leben torfbildende Sphagnum-Arten(wie S. magellanicum und S. cuspidatum). Das Ge -büsch wird von einzeln stehenden Pinus serotina-Bäumen überragt.
Zwei weitere bewaldete Moortypen möchten wirnoch erwähnen, nämlich erstens die „White CedarSwamp Forests“ aus Chamaecyparis thyoides (whitecedar) in feuchten Senken der Küstenebene auf sau-ren Histosolen, und zweitens die „Cypress Domes“
55.4 Azonale Vegetation 329

genannten Kleinmoore, die regelmäßig in denCyperaceen-Mooren auf lokalen Torfakkumulatio-nen sowie in der Küstenebene in isolierten Senkenmit stagnierendem Wasser vorkommen. Sie sind vonTaxodium distichum var. imbricatum (früher T. as -cendens) bewachsen. Die Bezeichnung „Dome“ gehtauf die kuppelförmige Gestalt der Wäldchen zurück,die dadurch zustande kommt, dass die Bäume imInnern größer werden als am Rand.
In den immerfeuchten Subtropen von Südame-rika, Afrika und Ostasien sind Moore flächenhaftvon geringerer Bedeutung als in Nordamerika, weilgroße, gefällearme Ebenen fehlen oder, wie inChina, nahezu alle Feuchtgebiete kultiviert wurden.In der Küstenebene von Südbrasilien gibt es Moor-wälder auf mehrere Meter mächtigen Histosolen,deren Baumschicht, verglichen mit den oben be -schriebenen nordamerikanischen Beständen, über-raschend artenreich ist. In St. Catarina und im Nor-den von Rio Grande do Sul enthalten sie noch bis zu50 Baumarten pro ha, unter denen Palmen (wie Sya-grus romanzoffiana), Myrtaceen (wie Myrcia multi-flora) und andere tropische Familien dominierenund die Verwandtschaft zu den tropisch-subtropi-schen Wäldern des „Mata Atlântica“ (s. Abschn.5.2.2.5) erkennen lassen (Dorneles & Waechter2004). Diese Moorwälder mussten häufig Zucker-rohrplantagen zur Produktion von Bioethanol wei-chen und sind bis auf wenige Restbestände ver-schwunden. Nach Süden zu werden sie artenärmer,bilden eher Dominanzbestände aus (mit Erythrinacrista-galli und Ficus cestrifolia) und machenschließlich Beständen aus graminoiden Helophyten(= Pflanzen, die im Wurzelraum an Wasserüber-schuss adaptiert sind) Platz, unter denen Scirpusgiganteus und Cyperus giganteus (ökologisch ver-gleichbar mit Cyperus papyrus in Af ri ka) besondersweit verbreitet sind. Solche Cyperaceen-Sümpfebestimmen, zusammen mit Palmenhainen (in denenneben der o. g. Syagrus-Art auch Butia yatay großeFlächen einnimmt), schwimmenden Wiesen ausRhynchospora asperula (Cyperaceae) und Gehölzin-seln das Bild der Esteros del Iberá im nördlichenEntre Rios (Zweistromland) zwischen den FlüssenParaná und Uruguay (Neiff 2001). Im Araukarienge-biet Südbrasiliens gibt es schließlich kleine Moorge-biete in den Senken des Basaltplateaus mit 2–3 mmächtigen Torfschichten und einer Vegetation ausSphagnum-Arten sowie verschiedenen tropischenund andinen Florenelementen (wie der EricaceeGaylussacia; Pfadenhauer & Castro Boechat 1981;Abb. 5-36e); als vegetationsgeschichtliches Ar chivlieferten sie den pollenanalytischer Beweis für den
Reliktcharakter des südbrasilianischen Graslandes(Behling et al. 2004; s. Abschn. 5.2.3.3.3).
5.4.1.3 Subtropische und nemorale Salzwasser-Feuchtgebiete
Außerhalb der Tropen wird der Übergang zwischenLand und Meer an Sand- und Schlickküsten in derRegel nicht mehr von Mangroven, sondern von Salz-marschen eingenommen. Unter einer Salzmarschversteht man eine natürliche oder halbnatürliche (d. h. partiell durch Landgewinnungsmaßnahmenentstandene) Vegetation, die aus niedrigen halophy-tischen (d. h. salzverträglichen) Gefäßpflanzen (Gras-land, Annuellenfluren, Klein- und Zwergstrauch-Vegetation) besteht und auf regelmäßig von Salzwas-ser überfluteten alluvialen Fluss-, See- und Meeres -sedimenten am Rand von salzhaltigen Gewässernwächst (Beeftink 1977, ergänzt; s. auch Adam 1990).Marschen sind von den Subtropen bis zur borealenKlimazone weit verbreitet. Wir finden sie im Binnen-land in der Umgebung von Salzquellen sowie in Tro-ckengebieten am Rande von Salzseen. Flächenhaftvon Bedeutung sind sie aber an Meeresküsten, dievor Wellenschlag durch vorgelagerte Inseln geschütztsind (ähnlich den Mangroven im Schutz von Koral-lenriffen), an Flussmündungen (Ästurare und Del-tas) und entlang von flachen Küstengewässern (La - gunen). An den Meeresküsten mit Tidehub, alsoeinem Wechsel zwischen Flut (submerse Phase) undEbbe (emerse Phase) in sechsstündigem Rhythmus,unterscheidet man die niedrige (zwischen mittleremNiedrig- und mittlerem Hochwasser; Eulitoral) vonder hohen Marsch (zwischen mittlerem Hochwasserund Springtide; Supralitoral). Die niedrige Marsch istwegen der extremen Lebensbedingungen jedenfallsim meernahen Bereich vegetationsfrei.
Die regelmäßige, durch den Tidehub hervorgeru-fene Überflutung, verbunden mit reduzierendenBedingungen im Substrat, und der hohe Salzgehaltbedingen die Artenarmut der Vegetation. Weit ver-breitet sind die beiden C4-Gras-Gattungen Spartina(Amerika, Europa, Nordafrika; heute auch in Ost-asien) und Distichlis (Amerika, Australien) sowie alsnur mäßig salztolerante (salzindifferente) SippenPhragmites australis und Typha (Kosmopoliten),unter den Juncaceen die Gattung Juncus (Kosmopo-lit), unter den Chenopodiaceen Arthrocnemum (vor-wiegend mediterran), Atriplex (weltweit außerhalbder borealen und polaren Zone), Salicornia, Sarco -cornia und Suaeda (kosmopolitisch; Verbreitungs -angaben aus Mabberley 2008). Hinzu treten lokal
330 5 Die warm-gemäßigte (subtropische) Zone5

eine Reihe weiterer Sippen aus verschiedenen Fami-lien, u. a. der Plumbaginaceae (mit der kosmopoliti-schen Gattung Limonium; Abb. 5-37e).
Die Vegetation folgt einem Gradienten aus Sali-nität und Bodentextur. Der Salzgehalt im Bodenkann im Überflutungsbereich landeinwärts zuneh-men, wenn die Niederschläge nicht ausreichen, dasSalz im Boden auszuspülen, und die Böden tonreichsind. Er nimmt ab, wenn die Böden im nieder-
55.4 Azonale Vegetation 331
schlagsreichen Sommer regelmäßig durchspült wer-den, vorausgesetzt, das Substrat ist ausreichendporös. Daraus leiten sich die wesentlichen Unter-schiede zwischen den Salzmarschen der immer-feuchten warm- und kühlgemäßigte Zone einerseits(eher Grasartige und annuelle Chenopodiaceen) undder winterfeuchten Subtropen andererseits ab (eherperennierende, häufig sukkulente Chenopodiaceen-Kleinsträucher). So werden z. B. im mediterranen
a b
c d
e
Abb. 5-37 Beispiele für die Vegetation von subtropischen Salzwasser-Feuchtgebie-ten. a = Arthrocnemum macrostachyum, Chenopodiaceae, im Rhone-Delta; b = Sali-cornia dolichostachya- (Vordergrund) und Spartina anglica-Bestände (Mittelgrundrechts) in der niederen Marsch der Nordseeküste (bei Spiekeroog); c = Meerbälle ausPosidonia oceanica bei Choggia, Italien; d = Strandnelkenrasen auf Norderney mitblühendem Limonium vulgare (Foto K. Kiehl); e = Limonium vulgare, ein typischerBestandteil von Salzwiesen an der Nordseeküste.

Raum regelmäßig überflutete, im Sommer aber tro-ckenfallende Bereiche innerhalb der Salzmarschenvon sukkulenten Arthrocnemum- und Sarcocornia-Arten besiedelt (wie A. macrostachyum und S. fruti-cosa auf Tonböden im Rhone-Delta; Britton & Pod-lejski 1981, Klötzli et al. 2010; Abb. 5-37a), die alsHolzpflanzen ähnlich wie Sarcocaulon und Anabasisin Halophyten-Halbwüsten der tropisch-subtropi-schen Trockengebiete (s. Abschn. 4.2.8 bzw. 4.3.4.3)mit Salzgehalten bis zu 0,6 % gut zurechtkommen.Hierbei handelt es sich um Xerohalophyten, derenrelative Wachstumsrate durch eine erhöhte Salzkon-zentration im Substrat gefördert wird (wie im Fallvon A. macrostachyum mit optimalem Wachstum zwi-schen 200 und 500 mmol = 0,6–1,4 % NaCl; Re -dondo-Gómez et al. 2010).
In den Marschen der immerfeuchten Subtropenund der kühl-gemäßigten (nemoralen) Zone domi-nieren dagegen die weniger salztoleranten Gräser,Grasartigen und Binsen. An der Atlantikküste Nord-amerikas gedeiht Spartina alterniflora von der bore-alen Zone bis in die Subtropen am meerwärtigenRand der niedrigen Marsch; dahinter, in der hohenMarsch, folgen Wiesen aus S. patens und Distichlisspicata. Am landeinwärtigen Rand der Salzwiesenbilden Juncus gerardii (New England), weiter südlichJ. roemerianus ausgedehnte eintönige Bestände(Mendelssohn & McKee 2000, Mitsch et al. 2009).Chenopodiaceen wie Salicornia spp. findet mankleinflächig an Stellen mit lokaler Salzanreicherung(Montague & Wiegert 1990).
In der niederen Marsch Europas, vor allem imWatt der Nordseeküste, gehört der Queller (Salicor-nia) zum charakteristischen Erscheinungsbild (Abb.5-37b). Von den zwölf Arten in West-, Mittel- undNordeuropa dringen die tetraploiden Spezies (wie S.dolichostachya) am weitesten meerwärts vor, wäh-rend die diploiden Sippen (wie S. europaea) ehermeerfern oder entlang der Ostseeküste vorkommen(Kadereit et al. 2007). Sonst wird die niedere Marschvon Spartina-Arten besiedelt, in Europa von S. angli -ca (Schlickgras). Das Schlickgras entwickelte sich ausder Hybridisierung der ursprünglich auf den Briti-schen Inseln heimischen Sippe S. maritima mit demim frühen 19. Jahrhundert nach England einge-schleppten C4-Gras S. alterniflora (Thompson 1991).Durch Verdoppelung des Chromosomensatzes ent-stand aus der diploiden, sterilen Hybridform (= S. ×townsendii) das tetraploide, kräftige, hochwüchsige(50–150 cm), phänotypisch plastische C4-Rhizom-gras S. anglica, das konkurrenzstärker ist als die Aus-gangssippen und diese heute weitgehend verdrängthat. Die Pflanze wurde in der ersten Hälfte des
19. Jahrhunderts an der deutschen und niederländi-schen Küste zur Neulandgewinnung angepflanzt undbreitet sich zulasten von Salicornia in der unterenMarsch weiter aus, in den letzten Jahrzehnten be -schleunigt durch die Klimaerwärmung (Nehring &Hesse 2008).
Auch in Kalifornien treten auf Böden mit denhöchsten Salzkonzentrationen Arthrocnemum-Artenauf, nämlich A. subterminale und A. macrostachyum(Knapp 1965). Im australischen Südwesten zeigtsich die sommerliche Salzanreicherung am Vorkom-men von Gebüschen aus Halosarcia- und Scleroste-gia-Arten (subendemisch bzw. endemisch), währendan der humiden Ostküste Sarcocornia quinqueflora inder niedrigen Marsch und Grasartige in der oberenMarsch mit geringeren Salzgehalten vorkommen(Adam 1994).
Eine Besonderheit sind die Seegraswiesen immediterranen Raum und an der Süd- und Südwest-küste von Australien (Larkum et al. 2006). Als See-gras bezeichnet man die Vertreter vor allem zweierFamilien der Monokotylen, nämlich der Zosteraceae(mit drei Gattungen, darunter Zostera) und derPosidoniaceae (eine Gattung, Posidonia, mit neunArten). Während die Gattung Zostera auch einigeVertreter in den Meeren der kühl-gemäßigten Zonestellt (z. B. an der europäischen Nord- und Ostsee-küste mit Zostera marina und Z. noltii), ist Posidoniaeine Pflanze der Subtropen (P. oceanica im Mittel-meer und weitere acht Arten an der Südküste vonAustralien). Die Pflanzen bilden ausgedehnte Be stän -de im Sublitoral der Küstengewässer bis etwa 40 mTiefe. Am Boden verankern sie sich mithilfe ihreskriechenden Rhizoms. Ihre Blätter sind linear oderfadenförmig, die Blüten reduziert (keine Blütenblät-ter), die Bestäubung erfolgt unter Wasser. Seegras-wiesen sind be deutende Nahrungsquellen für Was-servögel und Fische. An der Mittelmeerküste findetman manchmal runde Gebilde („Meerbälle“), dieaus abgerissenen Rhizomen und Blättern von Posi-donia oceanica bestehen und durch die Wellenbewe-gung bei Frühjahrs- und Herbststürmen entstehen(Abb. 5-37c).
In der feuchten nemoralen Zone entwickeln sichoberhalb des mittleren Tidehochwassers Salzwiesen(Abb. 5-37d). Besonders großflächig sind sie entlangder Nordseeküste von Belgien bis Dänemark ausge-bildet, kommen aber auch an der Ostsee vor (Ellen-berg & Leuschner 2010, Klötzli et al. 2010). Sie beste-hen (u. a.) aus salztoleranten Gräsern wie Puccinelliamaritima (Andel) und Festuca rubra ssp. litoralis,Binsen (z. B. Triglochin maritima, Juncaginaceae)sowie fakultativen Halophyten aus den Familien
332 5 Die warm-gemäßigte (subtropische) Zone5
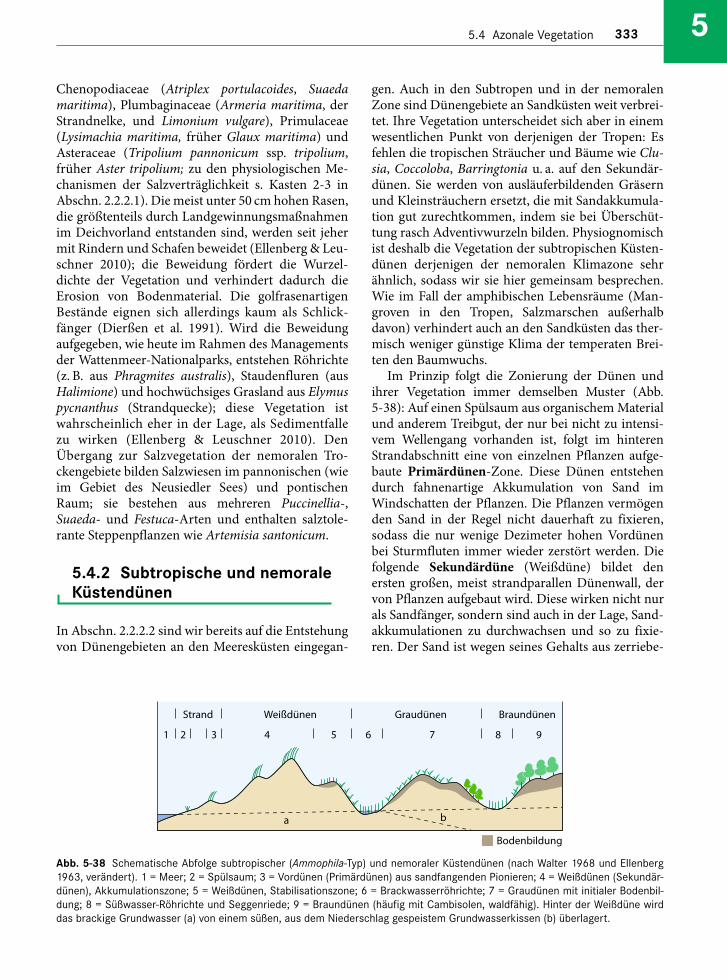
gen. Auch in den Subtropen und in der nemoralenZone sind Dünengebiete an Sandküsten weit verbrei-tet. Ihre Vegetation unterscheidet sich aber in einemwesentlichen Punkt von derjenigen der Tropen: Esfehlen die tropischen Sträucher und Bäume wie Clu-sia, Coccoloba, Barringtonia u. a. auf den Sekundär-dünen. Sie werden von ausläuferbildenden Gräsernund Kleinsträuchern ersetzt, die mit Sandakkumula-tion gut zurechtkommen, indem sie bei Überschüt-tung rasch Adventivwurzeln bilden. Physiognomischist deshalb die Vegetation der subtropischen Küsten-dünen derjenigen der nemoralen Klimazone sehrähnlich, sodass wir sie hier gemeinsam besprechen.Wie im Fall der amphibischen Lebensräume (Man-groven in den Tropen, Salzmarschen außerhalbdavon) verhindert auch an den Sandküsten das ther-misch weniger günstige Klima der temperaten Brei-ten den Baumwuchs.
Im Prinzip folgt die Zonierung der Dünen undihrer Vegetation immer demselben Muster (Abb. 5-38): Auf einen Spülsaum aus organischem Materialund anderem Treibgut, der nur bei nicht zu intensi-vem Wellengang vorhanden ist, folgt im hinterenStrandabschnitt eine von einzelnen Pflanzen aufge-baute Primärdünen-Zone. Diese Dünen entstehendurch fahnenartige Akkumulation von Sand imWindschatten der Pflanzen. Die Pflanzen vermögenden Sand in der Regel nicht dauerhaft zu fixieren,sodass die nur wenige Dezimeter hohen Vordünenbei Sturmfluten immer wieder zerstört werden. Diefolgende Sekundärdüne (Weißdüne) bildet denersten großen, meist strandparallen Dünenwall, dervon Pflanzen aufgebaut wird. Diese wirken nicht nurals Sandfänger, sondern sind auch in der Lage, Sand -akkumulationen zu durchwachsen und so zu fixie-ren. Der Sand ist wegen seines Gehalts aus zerriebe-
55.4 Azonale Vegetation 333
Chenopodiaceae (Atriplex portulacoides, Suaedamaritima), Plumbaginaceae (Armeria maritima, derStrandnelke, und Limonium vulgare), Primulaceae(Lysimachia maritima, früher Glaux maritima) undAsteraceae (Tripolium pannonicum ssp. tripolium,früher Aster tripolium; zu den physiologischen Me -chanismen der Salzverträglichkeit s. Kasten 2-3 inAbschn. 2.2.2.1). Die meist unter 50 cm hohen Rasen,die größtenteils durch Landgewinnungsmaßnahmenim Deichvorland entstanden sind, werden seit jehermit Rindern und Schafen beweidet (Ellenberg & Leu-schner 2010); die Beweidung fördert die Wurzel-dichte der Vegetation und verhindert dadurch dieErosion von Bodenmaterial. Die golfrasenartigenBestände eignen sich allerdings kaum als Schlick-fänger (Dierßen et al. 1991). Wird die Beweidungaufgegeben, wie heute im Rahmen des Managementsder Wattenmeer-Nationalparks, entstehen Röhrichte(z. B. aus Phragmites australis), Staudenfluren (ausHalimione) und hochwüchsiges Grasland aus Elymuspycnanthus (Strandquecke); diese Vegetation istwahrscheinlich eher in der Lage, als Sedimentfallezu wirken (Ellenberg & Leuschner 2010). DenÜbergang zur Salzvegetation der nemoralen Tro-ckengebiete bilden Salzwiesen im pannonischen (wieim Gebiet des Neusiedler Sees) und pontischenRaum; sie bestehen aus mehreren Puccinellia-,Suaeda- und Festuca-Arten und enthalten salztole-rante Steppenpflanzen wie Artemisia santonicum.
5.4.2 Subtropische und nemorale Küstendünen
In Abschn. 2.2.2.2 sind wir bereits auf die Entstehungvon Dünengebieten an den Meeresküsten eingegan-
1 2 3 4
a
Bodenbildung
b
5 6 7 8 9
Strand Weißdünen Graudünen Braundünen
Abb. 5-38 Schematische Abfolge subtropischer (Ammophila-Typ) und nemoraler Küstendünen (nach Walter 1968 und Ellenberg1963, verändert). 1 = Meer; 2 = Spülsaum; 3 = Vordünen (Primärdünen) aus sandfangenden Pionieren; 4 = Weißdünen (Sekundär-dünen), Akkumulationszone; 5 = Weißdünen, Stabilisationszone; 6 = Brackwasserröhrichte; 7 = Graudünen mit initialer Bodenbil-dung; 8 = Süßwasser-Röhrichte und Seggenriede; 9 = Braundünen (häufig mit Cambisolen, waldfähig). Hinter der Weißdüne wirddas brackige Grundwasser (a) von einem süßen, aus dem Niederschlag gespeistem Grundwasserkissen (b) überlagert.

nen marinen Muschel- und Schneckenschalen kar-bonatreich. Er wird so lange akkumuliert, bis dieDüne instabil wird; bei Stürmen entstehen dannWindanrisse, aus denen der Sand in Form von Para-beldünen landeinwärts verfrachtet wird. Im An -schluss daran baut sich die Sekundärdüne erneut auf.Das auf die Sekundärdünen folgende ältere Dünen-gebiet (in Europa Grau- und Braundünen genannt)ist in humiden Klimaten durch mehr oder wenigerfixierten Sand (abgesehen von den wanderndenParabeldünen) gekennzeichnet. Die Karbonate sindausgewaschen, die Böden sind umso tiefgründigerentwickelt, je weiter man sich von der Küste entfernt.
Im Folgenden geben wir einen vergleichendenÜberblick über die Vegetationsabfolge der subtropi-schen und nemoralen Küstendünen (Abb. 5-39) undbeziehen uns weitgehend auf die zahlreichen Beiträgezu einzelnen Dünengebieten in Van der Maarel(1993a, b, 1997a) und Martínez & Psuty (2004) sowieauf Kohler (1970, Chile), Eskuche (1973, Uruguayund Argentinien), Pfadenhauer (1979, Südbrasilien),Lubke et al. (1997, Südafrika), Clarke (1994, Austra-lien) und Haacks (2003, Neuseeland).
Spülsaum: Auf dem vom Meer angespülten und imvorderen Drittel des Sandstrandes linienförmig ange-häuften Treibgut aus organischem Material (Pflan-zen- und Tierkadavern) und zerriebenen Schnecken-und Muschelschalen siedelt eine Vegetation, die ausnitrophytischen, salztoleranten (sukkulenten), ephe-meren Pflanzen besteht. Kennzeichnend sind auf derNordhemisphäre die überwiegend annuellen Artender Gattung Cakile (z. B. C. maritima in Europa, C.edentula in Nordamerika; Abb. 5-39a) sowie Salsolakali agg. (C4-Chenopodiacee), in Japan und Neusee-land auch Calystegia soldanella (Convolvulaceae), dieMeerwinde, ein ursprünglich altweltliches, heuteweltweit verbreitetes Taxon, das sonst (z. B. im medi-terranen Raum) eher in den Dünen vorkommt undlange, kriechende Ausläufer bildet (Abb. 5-39b).Beide Cakile-Arten sind weltweit verschleppt.
Primärdünen (Vordünen; hinterer Strandab-schnitt): Die Vegetation besteht entweder aus peren-nierenden salztoleranten Gräsern oder aus niedrigen,häufig blattsukkulenten Geophyten mit Ausläufern(oder aus beidem). Die Pflanzen sind Sandfänger,besitzen aber nicht die Fähigkeit, Sandakkumulatio-nen dauerhaft zu durchwachsen und auf diese Weisehohe Dünen aufzubauen. Im Mittelmeerraum ist hierElymus farctus (Poaceae) weit verbreitet, ebenso ander Nordsee in den Dünengebieten der West-, Ost-und Nordfriesischen Inseln (hier vergesellschaftet
mit der Salzmiere Honckenya peploides, Caryophylla-ceae, mit dichten Wurzelwerk), in Kalifornien Ley-mus mollis sowie der aus Europa eingeführte Strand-hafer Ammophila arenaria. Die Primärdünen imWinterregengebiet von Chile werden von Nolanaparadoxa besiedelt, einer Solanacee, deren große,trichterförmige Blüten an diejenigen von Calystegiasoldanella erinnern (Nolana vielleicht von lat. nola =„kleine Glocke“; Genaust 1996). In Südbrasilien undUruguay kommen Spartina ciliata, Blutaparon portu-lacoides (eine blattsukkulente, kriechende Amaran-thacee) und Paspalum vaginatum vor. Paspalum wirdauf Golfplätzen in den Tropen und Subtropen alsErsatz für das Bermudagras Cynodon dactylon ver-wendet, weil es im Gegensatz zu diesem mit Meer-wasser gegossen werden kann (Duncan & Carrow1999). In Südafrika ist die Asteracee Arctotheca popu-lifolia heimisch (in Australien eingebürgert; Abb. 5-39c). Im australischen Südwesten (winterfeuchteSubtropen) schließlich wächst Spinifex hirsutus aufden Primärdünen, im Südosten (immerfeuchte Sub-tropen) S. sericeus. Die Spinifex-Arten (nicht zu verwechseln mit den umgangssprachlich Spinifexgenannten Igelgräsern der Gattung Triodia der inner-australischen Trockengebiete; s. Abschn. 4.3.4.4) sindsandfixierende, diözische Gräser mit bis zu 5 m lan-gen Ausläufern und stachelspitzigen Blättern.
Sekundärdünen (erster Dünenwall): Sekundärdü-nen sind das Ergebnis kräftig wachsender, nährstoff-bedürftiger Pflanzen, die nicht nur in der Lage sind,Sand vorübergehend zu akkumulieren, sondern ihnauch dauerhaft zu fixieren. Dies erreichen siedadurch, dass sie den angewehten Sand mit Rhizo-men, Sprossen und Wurzeln oder einer kräftigenVerzweigung mittels aktivierbarer Adventivknospendurchwachsen. Im Wesentlichen kann man zweiphysiognomische Pflanzenfunktionstypen unter-scheiden, nämlich (a) steife, hochwüchsige C3-Gräseroder Cyperaceen ähnlicher Physiognomie mit weitverzweigten Rhizomen („Ammophila-Typ“) oder (b)Sträucher mit Wurzelausläufern bzw. Sprossbewurze-lung („Ambrosia-Typ“; Abb. 5-40):a. Ammophila-Typ: Ammophila arenaria (Strandha-
fer) und der Bastard mit Calamagostis epigejos,Ammocalamagrostis baltica, sind in den Weißdü-nen Europas heimisch (Abb. 5-39d) und wurdenvon dort aus wegen ihrer hervorragenden Eigen-schaften für die Fixierung von Küstendünen inviele Gebiete der Welt eingeführt und erfolgreichangepflanzt. So findet man Ammophila-bewach-sene Sekundärdünen heute von Australien überKalifornien bis nach China (z. B. in Abb. 5-39f).
334 5 Die warm-gemäßigte (subtropische) Zone5

Dieser Export ist dort sinnvoll, wo es in Küstendü-nen keine einheimischen Gräser ähnlichen Ver-haltens gibt und durch Beweidung oder Tou-rismus Vegetationsschäden entstehen. Dieselassen sich mit dem wachstumsstarken Strandha-fer rasch heilen. An der warm-gemäßigten Atlan-tikküste Südamerikas gibt es allerdings mit demC4-Gras Panicum racemosum einen Dünenfixie-rer, der sich wie Ammophila verhält (Abb. 5-39e,
55.4 Azonale Vegetation 335
5-40b), ebenso im Südosten von Nordamerika mitUniola paniculata und Panicum amarum sowie inJapan mit Carex kobomugi. Alle genannten Artenzeichnen sich durch rasigen Wuchs aus und bildenein dichtes Rhizomgeflecht, das die Sekundärdü-nen bis zum süßen Grundwasserspiegel durchzieht(Abb. 5-40b). Die mehr oder minder dicht gepack-ten Sprosse sind skleromorph und tragen einedicke Kutikula (Schutz gegen Sandschliff und
hg
a b c
d
f
e
Abb. 5-39 Beispiele für die Vegetation subtropischer und nemoraler Küstendünen: a = Cakile maritima im Spülsaum (Nordsee,Spiekeroog); b = Calystegia soldanella bei Chioggia, Italien; c = Primärdünen mit Arctotheca populifolia (Asteraceae) bei Walpole,Australien; d = Sekundärdünen mit Ammophila arenaria (Nordsee, Amrum); e = Sekundärdünen an der Atlantikküste südlich vonPorto Alegre mit Panicum racemosum mit (links) und ohne ständige Sandzufuhr als Kümmerform (rechts); f = Sekundärdünen beiWalpole, Südwestaustralien mit Scaevola crassifolia, im Hintergrund Ammophila arenaria, eingebürgert; g = Scaevola crassifolia; h= Dünenwäldchen aus Juniperus phoenicea bei Chioggia, Italien.

Salzspray; Malloch 1997). In Australien über-nimmt neben den blattsukkulenten Sträuchernder Gattung Scaevola (s. unten) auch Spinifex wiein den Primärdünen die Rolle des Sandfixierers.Alle Arten wachsen am besten bei ständiger Sand -akkumulation, weil nur dann die Nährstoffversor-gung gesichert ist; hört die Sandnachlieferung auf,beginnen die Pflanzen zu kümmern.
b. Ambrosia-Typ: In Kalifornien und Mittelchile istAmbrosia chamissonis (Asteraceae) der Hauptdü-nenbildner: Der Kleinstrauch durchwächst denakkumulierten Sand und bildet an den verschütte-ten Sprossen Adventivwurzeln (Abb. 5-40a). Am -brosia ist in Nordamerika heimisch, wurde nachChile eingeschleppt und ist heute dort naturali-siert. In Südafrika dominieren im Bereich der Pri-mär- und Sekundärdünen blattsukkulente Sträu-cher der tropischen Gattung Scaevola (z. B. S.plumieri; Goodeniaceae; 5-39f, g) und Tetragoniadecumbens (Aizoaceae). Beide bilden ausgedehnteKlone über Wurzelausläufer. Wie Arctotheca
336 5 Die warm-gemäßigte (subtropische) Zone5
stammt Tetragonia aus Südafrika und ist inAustralien naturalisiert. Auch in Australien sindScaevola-Arten für die Winterregengebiete (S.crassifolia) und die immerfeuchten Subtropen (S.calendulacea) charakteristisch. Nach Genaust(1996) wurde der lateinische Name Scaevola vonLinné unter Anlehnung an den Beinamen desrömischen Patriziers C. Mucius Scaevola gewählt,der sich als Zeichen seiner Furchtlosigkeit dierechte Hand abbrannte (lat. scaevus = links,schief), und zwar in Anspielung auf die zygomor-phe Blütenkrone, die aussieht, als fehle eine Hälfte(Abb. 5-39g).
In den älteren Dünengebieten mit festgelegtemSand, aber ohne Bodenentwicklung (Graudünen)gedeihen niedrige Gebüsche oder Zwergstrauchhei-den; die beteiligten Arten sind nur mäßig tolerantgegenüber Sandakkumulation. Viele Arten sindimmergrün und besitzen erikoide oder schmale skle-rophylle Blätter, vermutlich als Reaktion auf die
0
20
40
cm
Luv Lee
PrimärwurzelAdventivwurzel
verholzteRhizome
a
c
b
Abb. 5-40 Sandakkumulierende und -fixierende Pflanzen: a = Ambrosia
chamissonis, chilenische Küstendünen(aus Kohler 1970); b = Panicum race-
mosum, südbrasilianische Küsten-dünen (aus Pfadenhauer 1979);
c = Sekundärdüne bei Maulin, Chile,mit Ambrosia und Ammophila arenaria.
Reproduziert mit Genehmigung vonwww.schweizerbart.de.

Nährstoffarmut in dieser Zone. Salze und Karbonatesind in den humiden Subtropen ausgewaschen. DieArtenzusammensetzung ist je nach Floreninventarsehr unterschiedlich: Die Gehölzbestände werden immediterranen Raum von Crucianella maritima(Rubiaceae), im Südosten von Nordamerika von Ilexvomitoria und Morella cerifera (Myricaceae), in Kali-fornien von Ericameria ericoides (Asteraceae) undLupinus arboreus, in Mittelchile von Margyricarpuspinnatus und Baccharis concava, in Südbrasilien undUruguay von Baccharis leucopappa und B. gnaphalo-ides, in Südafrika von Passerina rigida (Thymelaea-ceae), in Australien von Proteaceen-Gebüschen (imSüdwesten) und Eucalyptus-Beständen (im Südos-ten) und in Japan von Vitex trifolia ssp. litoralis (Ver-benaceae; kriechende Liane) bestimmt. In kühlerenGebieten wie im Nord- und Ostseeraum dominierenGrasartige wie Carex arenaria mi langen Ausläufernund der Therophyt Corynephorus canescens (Silber-gras).
Die älteren Dünengebiete mit ausgeprägterBodenentwicklung (Braundünen) sind von niedri-gen Wäldern entweder als vorübergehendes Wald-entwicklungsstadium oder bereits als Endstadiumder Vegetationsentwicklung bewachsen. Beispielesind die 2–3 m hohen Gebüsche aus Juniperus-Arten(J. oxycedrus, J. phoenicea; Abb. 5-39h) im mediterra-nen Raum, die mit Hartlaubgehölzen wie Phillyreaangustifolia vergesellschaftet sind. Im Südosten Nord-amerikas gibt es Quercus virginiana-Gebüsche undeine Reihe eher azonaler Kiefernwälder. In Kalifor-nien ist das Endstadium der Vegetationsentwicklungentweder ein Artemisia-reicher Küsten-Chaparral(Coastal Scrub) oder ein Wald aus Pinus radiata undanderen Kiefernarten. In Chile wächst auf den altenDünen ein Hartlaubwald aus windverformten Bäu-men der Flacourtiacee Azara celastrina. Aus Südbra-silien werden epiphytenreiche Dünenwälder lauro-phyllen Charakters aus Myrsine- (Myrsinaceae),Cupania- (Sapindaceae), Nectandra-Arten (Laura-ceae) beschrieben. In den Winterregengebieten Süd-afrikas dominiert der Küsten-Fynbos. In Ja pan kom-men im Bereich der subtropischen LorbeerwälderGebüsche aus Quercus phillyraeoides vor. In dernemoralen Zone Europas sind in eher ozeanischenLagen (Niederlande) Quercus robur und Q. petraeaals Endstadium der Vegetationsentwicklung zu er -warten (wenn die hohe Dynamik aus Sandumlage-rung eine ungestörte Sukzession überhaupt zu lässt),während unter kontinentalen Bedingungen (Polen,Baltikum) Pinus sylvestris zur Dominanz gelangt, inbeiden Fällen mit Empetrum nigrum als arktisch-borealem Zwergstrauch im Unterwuchs.
55.5 Subtropische Hochgebirge 337
5.5 Subtropische Hochgebirge
Wie wir in Abschn. 1.3.4.3 erläutert haben, ähnelt dievertikale Abfolge von Klima und Vegetation deraußertropischen Hochgebirge der horizontalen, brei-tengradparallelen Zonierung (s. Abb. 1-32). Die Ur -sache hierfür liegt im Wechsel vom tropischen Tages-zum außertropischen Jahreszeitenklima jenseits des20. Breitengrads. Dieser Wechsel vollzieht sich gra-duell und nicht abrupt; er macht sich zu allererst inden oberen Lagen der Gebirge bemerkbar. Währendnämlich die Wintermonate in der planaren, kollinenund submontanen Stufe der Subtropen mild sind undvielen frostempfindlichen Arten einschließlich einerReihe tropischer Kulturpflanzen das Gedeihen er -möglichen, können wegen der kalten Winter in dermontanen und subalpinen Stufe subtropischer Ge -birge (montan bis alpin) nur noch solche Bäumenwachsen, die mit einer dormanten Phase längereFrostperioden zu überstehen vermögen. Dabei han-delt es sich um Laub- und Nadelbäume, die denSchwerpunkt ihrer Verbreitung in der kühl- (nemo-ralen) bzw. kalt-gemäßigten (borealen) Klimazonehaben. Nemoral-nordhemisphärisch sind die som-mergrünen Arten u. a. der Gattungen Quercus, Fagusund Acer, nemoral-südhemisphärisch die immer-und sommergrünen Vertreter der Gattung Nothofa-gus. Nemorale Nadelbäume gehören u. a. zu den phy-logenetisch alten (artenarmen) Gattungen Cedrus(vier Arten), Chamaecyparis (fünf), Pseudotsuga(vier), Sequoia (eine), Sequoiadendron (eine), Tsuga(neun) und Taxus (neun; Mabberley 2008); einigevon ihnen bilden die Nadelwälder des kühl-gemäßig-ten pazifischen Nordwestens der USA (Nordkalifor-nien, Oregon, Washington; s. Abschn. 6.2.3). Über-wiegend borealen Charakter tragen Abies, Larix,Picea und Pinus. Die Arten dieser Gattungen domi-nieren in den borealen Nadelwäldern (s. Kap. 7),kommen aber auch häufig in den außertropischenHochgebirgen der Nordhalbkugel vor. Eher borealverbreitet sind einige sommergrüne Laubbäume mitPioniercharakter wie die Vertreter der GattungenAlnus, Betula, Populus und Sorbus (in Europa z. B. S.aucuparia, die Vogelbeere). In Eurasien kommenaußerdem breitblättrige immergrüne Kleinbäumeund Sträucher der Ericaceen-Gattung Rhododendron(montan bis subalpin) vor.
Wir sehen also in den subtropischen Hochgebir-gen (Auflistung in Kasten 5-7) eine zonale Parallele(Tab. 5-11): Die oreonemorale (Gebirgs-)Stufe ent-

licher Trockenheit betroffenen Hochlagen der nord-hemisphärischen Gebirge. Eine idealisierte Höhen-stufung in den Hochgebirgen der Subtropen würdedemnach folgendermaßen aussehen (Abb. 5-41): a. Planar bis submontan: subtropische Stufe mit
immergrünen laurophyllen oder sklerophyllenWäldern;
b. tief- und mittelmontan: oreonemorale Stufe mitsommer- (nordhemisphärisch) bzw. sommer- undimmergrünen (südhemisphärisch) Laubbäumenund/oder nemoralen Nadelbäumen;
c. hochmontan und subalpin: oreoboreale Stufe mitborealen immergrünen Nadelbäumen („Gebirgs -taiga“) und/oder borealen Pionierbäumen (som-mergrün), subalpin häufig Krummholz ausniederliegenden, Polykormone bildenden Bäu-men (wie Nothofagus-Arten) und Sträuchern (wiez. B. Alnus alnobetula, Rhododendron-Arten undPinus mugo bzw. P. pumila; zum Begriff Krumm-holz s. Abschn. 6.5.3);
d. alpine Stufe: Gras- oder Zwergstrauchtundrenmit Arten, von denen manche auch in den polarenTundren gedeihen.
Hochgebirge, die dieser Zonierung weitgehend fol-gen, sind u. a. die Pyrenäen (mit Pinus uncinata inder borealen Stufe), die östlichen Ausläufer des Hi -
338 5 Die warm-gemäßigte (subtropische) Zone5
spricht der nemoralen (kühl-gemäßigten) Zone, dieoreoboreale (Gebirgs-)Stufe der borealen (kalt-gemäßigten) Zone. „Subalpin“ als Ökoton zwischenWald- und Baumgrenze ist mit „subpolar“ vergleich-bar. Die Vegetation der alpinen Stufe weist physio-gnomisch und teilweise auch floristisch eine gewisseÄhnlichkeit mit derjenigen der polaren Tundren auf(„oreopolar“), jedenfalls in den nicht von sommer-
S N S N
S N
0
1
2
3
4
5
445
5
6
109
116
33
2 2
1 11
2
3
45
Mittel-Nepal28–29° N / 84° O
Hoher Atlas31° N / 8° O
Hindukusch37° N / 73° O
70 km 6,7 km
6
¥ 10
00 m
NN
7
7
7
8
8
Abb. 5-41 Höhenstufen einiger subtropischer Gebirge (s. auchAbb. 5-7, 5-19). Mittlerer Himalaya, Nepal (randtropisch, nacheiner Vorlage aus Schroeder 1998, verändert und ergänzt nachMiehe 2004a): 1 = saisonierter Regenwald (warm-tropischeStufe), 2 = oreotropischer Lorbeerwald (subtropische Stufe), 3 = oreotropischer Nebelwald (unterer Nebelwald; subtropi-sche Stufe), 4 = Tsuga dumosa-Quercus semecarpifolia-Wald(mittlerer Nebelwald; nemorale Stufe), 5 = Abies spectabilis-Abies densa-Wald (oberer Nebelwald; boreale Stufe), 6 = Krumm-holz aus Rhododendron-Arten, 7 = Vegetation der alpinen Stufeaus Rhododendron-Zwergstrauchheiden bzw. Kobresia nepa-lensis-Hochgebirgssteppen; leeseitig: 9 = Trockengebüsche, 10 = Nadelwald aus Pinus wallichiana und Cupressus torulosa,11 = Pinus wallichiana-Wald.Hoher Atlas, Marokko (südmediterran, nach Walter 1968 undKlötzli & Burga 2004): 1 = potenziell Stipa tenacissima-Gras-Halbwüsten und hartlaubige Trockengebüsche aus Argania spi-nosa (inframediterran), 2 = Tetraclinis articulata-Bestände mitJuniperus phoenicea (thermomediterran); 3 = Quercus ilex-Wald, im Osten des Gebirges mit Cedrus atlantica (mesomedi-terran); 4 = Juniperus thurifera-Gebüsch; 5 = Dornpolster-Vege-tation.Hindukusch, Afghanistan (nach Breckle & Frey 1974). 1 = sub-tropisches Trockengebüsch, 2 = Hartlaubwälder aus Quercusbaloot, in den obersten Lagen Q. semecarpifolia ; 3 = Nadel-wälder aus Pinus gerardiana, in oberen Lagen Cedrus deodara;4 = Krummholz aus Juniperus- und Betula-Arten; 5 = alpineStufe, 6 = nivale Stufe; 7 = sommergrüne Offenwälder (Pistaciavera und Amygdalus bucharica); 8 = Juniperus-Offenwälder mitsommergrünen Gehölzen.
Tab. 5-11 Vergleich der gebräuchlichen Höhenstufen-Modellein subtropischen Hochgebirgen
allgemeine thermische Höhenstufen klimatisch-geo- Höhenstufen in subtropischenmorphologische im mediter- HochgebirgenHöhenstufen1 ranen Raum2 als zonale
Parallele3
planar thermomediterran subtropisch
kollin mesomediterran
submontan
tiefmontan supramediterran oreonemoral
mittelmontan montan-mediterran
hochmontan oreomediterran oreoboreal
subalpin
alpin altimediterran alpin
nival kryomediterran nival
1 Aus Abschn. 1.3.4.3; 2 aus Tab. 5-7; 3 in Anlehnung an Schroeder(1998), verändert.

ma laya-Systems in Westchina (Wang 1961), die japa-nischen Hochgebirge (Yoshino 1978; s. Abb. 5-7),soweit sie in den Subtropen liegen, einschließlich desFuji-san (Miyawaki et al. 1994), und die Westseite derSierra Nevada in Kalifornien (Barbour & Minnich2000, Billings 2000, Fites-Kaufman et al. 2007). Inden japanischen Alpen (Hida-Gebirge) besteht dieVegetationsabfolge aus Lorbeerwald bis etwa 600 mNN, gefolgt von einem sommergrünen Wald ausFagus crenata (bis 1.600 m NN) und einer Gebirg-staiga aus Abies mariesii und A. veitchii (borealeStufe). Subalpin herrscht ein Krummholz aus Pinuspumila vor. Ganz ähnlich ist die Abfolge am Fuji-san;nur sind dort in der subalpinen Stufe auf nährstof-freichen und feuchten Böden Betula ermanii und auftrockenen Böden die niederliegende Larix-Art L.kaempferi verbreitet; Letztere ist zudem ein Pionierauf vulkanischer Asche.
In der dem Pazifik zugeneigten Seite der SierraNevada wächst (über einer planar-submontanensubtropischen Zone mit Quercus chrysolepis) ab etwa800 m NN ein vierschichtiger nemoraler Nadelwaldaus Pseudotsuga menziesii (Douglasie), Abies concolorund Pinus ponderosa. Diese Bäume bilden eine obere,30–60 m hohe Baumschicht; nur vereinzelte Grup-pen aus Sequoiadendron giganteum, dem Mammut-baum, können bis zu 90 m hoch werden (Kasten 5-8).Darunter, in einer zweiten Baumschicht (8–15 m),wachsen sommergrüne Acer-, Cornus-, Corylus- undQuercus-Arten (wie Q. kelloggii). Die dritte Schichtbesteht aus einem niedrigen Gebüsch, u. a. aus Arc-tostaphylos, Ceanothus, Cercocarpus, und die vierteSchicht aus nemoralen Hemikryptophyten und Geo-phyten (wie Adenocaulon, Galium, Iris, Viola u. a.).Diesen „Midmontane Forest“ werden wir in ähn-licher Artenzusammensetzung weiter nördlich als
55.5 Subtropische Hochgebirge 339
Subtropische HochgebirgeZu den subtropischen Hochgebirgen gehört das hinsichtlichMeereshöhe und Fläche bedeutendste Hochgebirge derErde, das zentralasiatische Himalaya-Gebirgssystem. Es er -streckt sich von etwa 40 bis 25° S sowie von 75 bis 105° Ound umfasst den Himalaya (im eigentlichen Sinn) zwischender Indus-Schlucht im Westen, der Brahmaputra-Schlucht imOsten, dem nordindischen Vorgebirge im Süden und demTsangpo-Tal im Norden mit dem Mt. Everest (8.846 m NN),den Transhimalaya und das nördlich anschließende Hochandvon Tibet sowie dessen Randketten zum Tarimbecken (Kun-lun Shan, Altun Shan, Nan Shan, 6.346 m NN, bereits nemo-ral), ferner das Karakorum-Gebirge in Nordwestindien (Kash-mir) und Nordpakistan mit dem Chogori (= K2; 8.611 m NN)und die östlich des Hochlands von Tibet liegenden Gebirgs-ketten im Quellgebiet des Mekong und des Chang Jiang(Jangtsekiang) in Westchina (Heng Duan Shan, Bayan HarShan, Gongga Shan 7.590 m NN u. a.). Während der Hima-laya ein randtropisches Gebirge ist, das zwischen den Tro-pen und den Subtropen vermittelt, sind die westchinesi-schen Gebirge in der Provinz Yunnan und Sichuan reinsubtropisch (z. B. Emei Shan; Tang & Ohsawa 1997).
Außerhalb des Himalaya-Systems gehören in Ostasiender T'ai-wan Shan in Taiwan (Yü Shan 3.997 m NN; randtro-pisch), in Japan das Hida-Gebirge (Hotaka-dake 3.190 m NN)und der Fuji-san (3.776 m NN), im südpazifischen Raum dieAustralischen Alpen mit dem Mt. Kosciusko (2.230 m NN, im Südosten des Kontinents) und die Gebirge auf der Nord-insel von Neuseeland (Tongariro National Park mit demMt. Ruapehu, 2.797 m NN und dem Mt. Taranaki = Mt. Eg -mont 2.518 m NN) zu den immerfeuchten Subtropen.
Den winterfeuchten Subtropen sind im Mittelmeergebietdie Abruzzen (Gran Sasso 2.914 m NN) und der Ätna (3.340 m NN) in Italien, die korsischen Gebirge (Monte Cinto2.710) in Frankreich, die Sierra Nevada (Mulhacén, 3.478 mNN) und die Pyrenäen (Pico de Aneto, 3404 m NN) in Spa-nien, der Olymp (Mitikos, 2918 m NN) in Griechenland, dasTaurus-Gebirge (Akdag, 3.086 m NN) in der Türkei und derHohe Atlas (Jabal Tubga–l, 4.165 m NN) in Marokko zuzurech-nen. Östlich an die Mediterraneis schließen sich die Hoch-gebirge im Iran mit dem Zagros im Südwesten des Landes(Oshtora–n Ku–h, 4.331 m NN) und dem Alborz (Takt-e Soley -ma–n, 4.820 m NN) an. Von Winterregen ist auch die Vege-tation von Afghanistan geprägt, wenngleich es sich überwie-gend um Wüsten, Halbwüsten und Trockengebüsche handelt; dennoch treten oberhalb des Trockengürtels im Hin-dukusch (Tirich Mı–r 7.699 m NN in Pakistan) Hartlaubwäldermit mediterranen Florenelementen auf. Die amerikanischenWinterregengebiete sind durch die Sierra Ne vada in Kalifor-nien (Mt. Whitney 4.418 m NN) und die Anden in Chile (mitt-lere Anden; Aconcagua 6.959 m NN) vertreten. Die Drakens-berge in Südafrika (Thabana Ntlenyana am Ostrand vonLesotho, 3.482 m NN) nehmen eine Sonderstellung ein; siesind, soweit sie zu den immerfeuchten Subtropen gehören,in den höheren Lagen, vermutlich bis zur potenziellen Wald-grenze, von einem ausgedehnten Themeda-Grasland ge -prägt. In der winterfeuchten Kapregion fehlen Hochgebirge,ebenso in Südwestaustralien.
Kasten 5-8

zonale Vegetation des amerikanischen Nordwestens(s. Abschn. 6.2.3) wieder antreffen. In der borealenStufe (oberhalb von etwa 2.300 m NN) dominierenPinus contorta, Abies magnifica und Populus tremulo-ides, gefolgt von einem subalpinen Offenwald ausPinus albicaulis, P. longaeva, P. flexilis und Tsugamertensiana (mountain hemlock). P. albicaulis (Abb.5-42a) ist in schneegeschützten Lagen ein mehrstäm-miger 10–12 m hoher Baum, der in der subalpinen
Stufe auch als Krummholz vorkommt. Er erinnertphysiognomisch an Pinus sibirica und die nah ver-wandte Zirbelkiefer P. cembra. Wie diese erzeugt ergroße essbare Samen, die vom Kiefernhäher Nuci-fraga columbiana (dem amerikanischen Verwandtendes Tannenhähers) ausgebreitet werden. Der Vogelversteckt mehr Samen im Boden, als er als Winter-vorrat benötigt; die nicht verbrauchten oder verges-senen Nüsse wachsen zu Baumgruppen heran (Hut-
340 5 Die warm-gemäßigte (subtropische) Zone5
Sequoiadendron giganteum (Riesenmammutbaum)
Kasten 5-9
Mit bis zu 90 m Höhe, einem Stammdurchmesser oberhalbder Stammbasis von bis zu 11 m und einer Borke, die eineDicke von 60 cm erreichen kann, ist Sequoiadendron gigan-teum neben Sequoia sempervirens, dem Küstenmammut-baum, der 120 m Höhe erreichen kann, eine der eindrucks-vollsten Baumgestalten der Erde (Watson & Eckenwalder1993, Farjon 2010b). Sein natürliches Vorkommen ist aufetwa 67 Einzelbestände („Groves“) zwischen 1 und 1.600 haGröße am Westabfall der Sierra Nevada beschränkt, diezusammen rund 146 km2 bedecken und zwischen 1.400 und2.400 m NN liegen (Fites-Kaufman et al. 2007). BevorzugterStandort sind tief eingeschnittene Täler und nordseitigeHänge mit ausreichend Bodenfeuchte auch während der tro-ckenen Sommermonate. Die „Groves“ sind keine Reinbes-tände von Sequoiadendron; die Art ist vielmehr den nemora-len Nadelwäldern aus Abies concolor und Abies magnifica (inhöheren Lagen) beigemischt. Das derzeit mächtigste Rie-senmammutbaum-Individuum mit einem Holzvolumen von1.486 m3 („General Sherman Tree“ im Sequoia & KingsCanyon National Park) hat ein Alter von etwa 1.650 Jahren(Stephenson 2000); vermutlich kann die Art aber mehr als2.000 Jahre alt werden (Farjon 2010b). Das rotbraune Kern-holz, von dessen Farbe der amerikanische Name redwoodkommt, ist termitenresistent, zäh und leicht zu bearbeiten.Es diente früher als Bau- und Möbelholz. Heute sind die Rest-bestände des Riesenmammutbaumes (66 % des ursprüng-lichen Vorkommens) streng geschützt.
Sequoiadendron produziert pro Zapfen rund 200 kleine,geflügelte, anemochore Samen, die am besten auf Mineral-boden in Bestandslücken keimen, vorzugsweise nachdemdie Streu- und Humusauflage durch Feuer zerstört wurde.Die Jungpflanzen sind schnellwüchsig; unter günstigenUmständen (ausreichend Licht und Bodenfeuchte) kann dasHöhenwachstum bis zu 0,7 m pro Jahr betragen (Schütt et al.2004). Sequoiadendron ist also ähnlich wie Fitzroya cupres-soides in den chilenischen Anden (s. Abschn. 6.2.4.3) einelanglebige Pionierart (York et al. 2011), deren Regenerationvon feuerbedingten Bestandslücken mit allerdings großemzeitlichen Abstand (viele 100 Jahre) abhängt. Mit häufigem,leichtem Oberflächenfeuer kommt sie gut zurecht.
Abb. 1 Mariposa Grove mit Sequoiadendron giganteum,Sierra Nevada, Kalifornien (Foto H. Albrecht).

chins & Lanner 1982). Pinus longaeva (Abb. 5-42b)gehört weltweit zu jenen Baumarten, die das höchsteAlter erreichen; einzelne Individuen können mehrereTausend Jahre alt werden. Die Baumgrenze liegt beietwa 3.500 m NN. Darüber gedeiht ein außerordent-lich artenreicher alpiner Rasen, dessen Matrix ausGrasartigen (besonders Carex mit 29 Arten und ver-schiedenen Gräsern der Gattungen Agrostis, Bromusund Poa) gebildet wird (Rundel 2011).
Einige der subtropischen Hochgebirge weichenvon diesem allgemeinen Schema ab. Hierzu gehörendie randtropischen Hochgebirge mit ihrer intermedi-ären Stellung zwischen Tropen und Subtropen, dienordmediterranen und südhemisphärischen Hoch-gebirge, denen die boreale Stufe fehlt, die süd- undostmediterranen Hochgebirge, bei denen die som-merliche Trockenzeit bis in die alpine Stufe reicht,und schließlich die iranisch-afghanischen Hochge-birge, die aufgrund ihrer Lage einen eigenständigenCharakter haben. 1. Randtropische Hochgebirge sind in den unteren
Lagen durch eine warmtropische (mit tropischenTieflandregenwäldern) und eine oreotropischeStufe (mit laurophyllen Wäldern) gekennzeichnet,tragen anstelle der oreotropischen Heidewälderaber bereits Nadelwälder mit borealem Charakter.Ein Beispiel ist der T‘ai-wan Shan auf Taiwan:Hier kommen ab etwa 2.900 m NN Nadelwälderaus Abies kawanakii vor, denen bis zur Baum-grenze ein Gebüsch aus Rhododendron-Arten undJuniperus squamata folgt (boreale Stufe; Schroeder1998). Randtropisch ist auch der Himalaya(Schweinfurth 1957, Miehe in Walter et al. 1991,Miehe 2004a); sein vom Monsun geprägter Süd-ostteil (Mittelnepal; 83°30' O; Abb. 5-40) ist aufder Südseite bis in eine Höhe von 900–1.200 m NNvon einem tropischen, saisonalen Tieflandregen-wald aus Dipterocarpaceen (vor allem aus Shorearobusta) bedeckt (s. Abschn. 2.1.8). Darüber folgtals westliche Fortsetzung der südostasiatischenLorbeerwald-Region ein oreotropischer Lorbeer-wald, in dem immergrüne Quercus-Arten undzahlreiche weitere Laurophylle (wie Schima, Cast-anopsis, Beilschmiedia) dominieren und der ab2.000 m NN nebelwaldartigen Charakter mit rei-chem Epiphytenbewuchs vorwiegend aus Moosenund Flechten annimmt. Die unteren Lagen desLorbeerwaldes sind häufig anthropogen oder inkonvexen Hanglagen durch eine feuerstabilisierteKlimax aus Pinus roxburghii ersetzt. In der mittle-ren Nebelwaldstufe (nemorale Stufe) wachsen mitTsuga dumosa und Quercus semecarpifolia diegrößten Waldbäume des Himalaya (Abb. 5-42c);
55.5 Subtropische Hochgebirge 341
beide Arten werden bis zu 40 m hoch. Zahlreichegroßblättrige Rhododendron-Arten, mannshoheFarne und einige sommergrüne Gehölze (wie dieGattung Schefflera, Araliaceae) sind charakteris-tisch. Ab etwa 3.000 m NN dominieren Koniferen,insbesondere Abies spectabilis und A. densa, ver-mischt mit Betula utilis (boreale Stufe, bis 3.800 mNN; Abb. 5-42e). Subalpin folgen dann ein Rho-dodendron-Krummholz (Abb. 5-42f) und, schatt-seitig, oberhalb von etwa 4.500 m NN eine Rho-dodendron-Zwergstrauchheide (vorwiegend R.an thopogon), südseitig ein (teilweise anthropoge-ner) von Kobresia-, Carex- und Festuca-Artendominierter Rasen (Abb. 5-43a).
2. In den Gebirgen des nördlichen mediterranenRaumes (kontinuierliche Zunahme der Humiditätmit steigender Meereshöhe und entsprechendeAbnahme der Dauer der sommerlichen Trocken-periode) fehlt die boreale Stufe (s. auch unterPunkt 3) und die derzeitige Waldgrenze (s. Abb. 5-19, Korsika-Nordseite) befindet sich viel niedriger,als sie eigentlich sein müsste, verglichen mit ande-ren nordhemisphärischen Gebirgen gleicher Brei-tenlage und ähnlicher klimatischer Bedingungen(Sierra Nevada in Kalifornien; Körner 2012). Soliegen die Bodentemperaturen im Waldgrenzbe-reich beispielsweise des Apennin (1.820 m NN,Fagus sylvatica), des Olymp (2.320 m NN, Pinusheldreichii) oder südgriechischer Gebirge (Pelo-ponnes, 2.100 m NN, Abies cephalonica) bei 10,5bzw. 8,0 bzw. 11,1 °C und damit deutlich höher alsder von Körner (2012) angenommene thermischeGrenzwert des Baumwachstums von +6,7 °C in 10 cm Bodentiefe (gemessen in Hochgebirgenebenso wie an der boreal-polaren Waldgrenzewährend der Vegetationszeit; Körner & Paulsen2004; s. Abschn. 1.3.4.3). Die heutige Waldgrenze dürfte somit anthropogensein; ohne menschlichen Einfluss läge sie wohl umeiniges höher. Belege dafür liefern palynologischeDaten, beispielsweise aus dem Apennin: Dort gingAbies alba seit Beginn der Bronzezeit (vor etwa5.000 Jahren BP) kontinuierlich zurück, währendFagus zunahm und die Feuerfrequenz anstieg(Vescovi et al. 2010). Anstelle einer vermutlichvon Abies- und Picea-Arten gebildeten Gebirg-staiga haben sich heute Gebüsche aus Juniperuscommunis, Zwergstrauchheiden aus Vaccinium-Arten und Weiderasen aus Festuca-Arten undKobresia myosuroides angesiedelt.
3. Auch in den südhemisphärisch-subtropischenHochgebirgen fehlt die boreale Stufe. Die nemo-ralen Wälder aus Nothofagus-Arten in Chile

(Oberdorfer 1960) und in Neuseeland (Wardle1991) bzw. aus Eucalyptus pauciflora ssp. nipho-phila in den Australischen Alpen (Williams &Costin 1994) reichen deshalb bis zur Waldgrenze(s. Abb. 5-7), wo sie alle eine niedrige, krüppeligeForm annehmen und ein (subalpines) Krumm-holz bilden, das gegen die alpine Stufe scharf abge-grenzt ist. Ähnlich wie im Fall der nordmediterra-nen Gebirge stimmt auch hier der thermischeGrenzwert mit den vor Ort gemessenen Tempera-turen nicht überein (Termas de Chillán, Chile,Nothofagus pumilio, 8,7 °C; Körner 2012). DieUrsache für die zu tief liegende Waldgrenze habenwir schon in Abschn. 1.3.4.3 angesprochen: Aufder Südhemisphäre fehlen die an boreale Hochla-genklimate adaptierten Baumarten. Es handeltsich hier also im Sinn von Körner & Paulsen(2004) um eine Baumartengrenze (tree speciesline) und nicht um eine Baumgrenze (tree line).Im Einzelnen sieht die Höhenstufung wie folgtaus: Im chilenischen Winterregengebiet folgt aufden Hartlaubwald ab etwa 1.200 m NN einenemorale Stufe mit Nothofagus macrocarpa undeinem N. antarctica- (beide Arten sommergün)bzw. Kageneckia angustifolia-Krummholz bis zurWaldgrenze bei etwa 2.000 m NN. Darüber wächstein Gebüsch aus Chuquiraga oppositifolia (Astera-ceae) und Berberis empetrifolia, das oberhalb von2.600 m NN von einer Vegetation aus Polster-pflanzen abgelöst wird (Cavieres et al. 2000). Aufder Nordinsel von Neuseeland wiederum werdendie Wälder aus N. fusca (Untergrenze bei etwa 600m NN) ab etwa 1.000 m NN von N. solandri ssp.cliffortioides (immergrünes Krummholz) abgelöst;die Waldgrenze liegt tiefer als hinsichtlich der geo-graphischen Breite zu erwarten wäre, nämlich nurbei etwa 1.300 m NN. Darüber sind Zwergstrauch-heiden aus Ericaceae, stellenweise mit mehr oderminder dominanten, hochwüchsigen Horstgrä-sern der Gattungen Chionophila (Neuseeland undAustralien) bzw. Festuca (Chile; überwiegend F.thermarum) ausgebildet.
4. Hochgebirge der winterfeuchten Subtropen, indenen auch in höheren Lagen noch eine ausge-prägte sommerliche Trockenzeit auftritt oder gardie Niederschläge oberhalb der Wolkenkondensa-tionszone wieder abnehmen, zeichnen sich da -durch aus, dass ihnen eine nemorale Stufe fehltund die boreale Stufe, wenn vorhanden, durchxerophytische Gehölze (auf der Nordhalbkugel v. a.mit der Gattung Juniperus) geprägt ist. Das ist derFall im Norden des chilenischen Winterregenge-biets, wo über dem Hartlaubgürtel ein sukkulen -
tenreicher Trockenbusch mit vielen Säulenkak-teen wächst. Vor allem aber gehören viele süd-und ostmediterrane Hochgebirge zu diesemTypus. Beispiele sind die Sierra Nevada in Süd -spanien (Pauli & Gottfried 2004), das Atlas-Ge -birge (Walter 1968; Abb. 5-40), das Taurus-Ge -birge und Kreta (s. Abb. 5-19) sowie der Olymp,wo in schattigen Nordlagen auch Fagus mit relativvereinzelten Pinus halepensis erscheint. In all die-sen Gebirgen gibt es in mittleren Höhenlagen(über thermomediterranen Wäldern, s. Abschn.5.3.2.2.2) Hartlaubwälder aus Quercus ilex (west-mediterran) bzw. Q. coccifera (ostmediterran), anSüdhängen auch Pinus-Arten (meist P. halepensis,im Osten P. brutia; Abb. 5-21b), im Atlas bzw. imTaurus angereichert mit Cedrus atlantica bzw. C.libani (Abb. 5-42g). Dieses Waldgebiet reicht imAtlas fast bis 3.000 m NN (Quercus ilex sogar dar-über), sonst aber kaum über 2.000 m NN. Darüberfolgen Offenwälder bzw. -gebüsche aus Cupressus-oder Juniperus-Arten (z. B. Genista versicolor-Juni-perus communis-Zwergstrauchheiden in der SierraNe vada, Juniperus thurifera-Offenwald im Atlas,Cupressus sempervirens-Offenwald im ostmediter-ranen Raum wie auf Kreta, Abb. 5-21a, aber nichtauf Zypern) und schließlich die alpinen Dorn-pols terfluren (Abb. 5-43b). Sie bestehen aus halb-kugelförmigen, stark bedornten Sträuchern mitkaum mehr als 50 cm Höhe und unterschiedlichertaxonomischer Zugehörigkeit, die in der Literaturals Beispiel einer morphologischen Konvergenzangesehen werden (Walter et al.1991; Kasten 5-9).
5. Eine Sonderstellung nehmen die iranisch-afgha-nisch-himalayischen Hochgebirge am kontinen-talen Ostrand der eurasiatischen winterfeuchtenSubtropen ein (iranische und afghanische Ge-birge, Hindukusch, westlicher Karakorum). Erstensliegen sie größtenteils in tropisch-subtropischenTrockengebieten, sodass die untere (subtropische)Stufe überwiegend aus Trockengebüschen besteht(für Afghanistan s. Breckle 2007). Zweitens sindihre floristischen Beziehungen größer zur irano-turanischen als zur mediterranen Florenregion (s.Abschn. 1.2.1; Meusel & Schubert 1971). Drittensmacht sich im Osten, vor allem in den oberenGebirgslagen, bereits der Einfluss des Sommer-monsuns bemerkbar, sodass die Niederschlags-kurve zweigipfelig wird. Die Vegetationsabfolgeweicht deshalb von derjenigen der bisher bespro-chenen subtropischen Hochgebirge ab. Im Hindukusch findet sich folgende Zonierung(Breckle & Frey 1974; Abb. 5-41): Auf der Südost-abdachung dieses Gebirges reicht ein Trockenge-
342 5 Die warm-gemäßigte (subtropische) Zone5

büsch aus Acacia modesta, Calotropis procera undZiziphus nummularia (s. Abschn. 4.3.3.2) bis etwa1.300 m NN. Darüber gedeiht ein Hartlaubwald,dessen Artenzusammensetzung mediterrane Ver-wandtschaft erkennen lässt. Denn er besteht aus
55.5 Subtropische Hochgebirge 343
der mit Quercus ilex nahe verwandten EichenartQ. baloot und anderen Vikarianten meditererranerSippen (wie Olea ferruginea, Pistacia khinjuk; Frei-tag 1982). Er ist in den vom Monsun geschütztenTälern bis nach Nordpakistan verbreitet (Schick-
a
d e
f g
b c
Abb. 5-42 Beispiele für oreonemorale und -boreale Wälder subtropischer Gebirge. a = Pinus albicaulis (weiße Stämme) mit P. con-torta (oreonemoral; Yosemite National Park, USA, Foto H. Albrecht); b = Pinus longaeva (oreonemoral, Arizona; Foto F. Klötzli); c = oreonemoraler Tsuga dumosa-Wald (Himalaya; Foto G. Miehe); d = Abies densa-Wald (oreoboreal; Nordbhutan; Foto G. Miehe);e = subalpines Betula utilis-Gebüsch (oreoboreal, Kashmir, 3.650 m NN; Foto F. Klötzli); f = subalpines Rhododendron-Gebüsch (oreo boreal, Nordbhutan; Foto G. Miehe); g = oreonemoraler Cedrus atlantica-Bestand (1.500–2.000 m NN, Hoher Atlas, Marokko;Foto W. Zielonkowski).

hoff 1994). An der Obergrenze dieses Waldeskommt mit Q. semecarpifolia eine Baumart hinzu,deren Hauptvorkommen in der mittleren Nebel-waldstufe der Himalaya-Südabdachung liegt(Shrestha 2003). Oberhalb von etwa 2.300 m NNtreten Nadelwälder auf, die in ihren tieferen Lagensowie im trockeneren Westen und in den inner-montanen Tälern aus der mit Pinus nigra eng ver-wandten Kiefer Pinus gerardiana bestehen. In denoberen Lagen dieser Nadelwälder tritt bis zurWaldgrenze Cedrus deodara auf. Gegen Ostenkommen mit zunehmendem MonsuneinflussAbies spectabilis, Pinus wallichiana und Picea smi-thiana hinzu. Die Krummholzstufe zwischenWald- und Baumgrenze (3.100–3.400 m NN) wird
von Juniperus-Arten (z. B. J. indica) und von Be -tula utilis gebildet. Die Nordwestabdachung des Hindukusch stehtunter dem Einfluss eines winterfeuchten konti-nentalen Trockenklimas. Unterhalb von 1.800 mNN dominieren sommergrüne, offene Gehölzflu-ren, in denen u. a. Pistacia vera wächst, die Aus-gangssippe der essbaren Pistazien. Darüberwächst ein Wacholder-Offenwald u. a. aus Junipe-rus macrocarpa bis zur Baumgrenze bei 3.800 mNN; er ist heute durch Beweidung und Holznut-zung weitgehend zerstört und hat ausgedehntenDornpolsterfluren aus Astragalus-, Acantholimon-(Plumbaginaceae) und Acanthophyllum-Arten(Caryophyllaceae) Platz gemacht (Abb. 5-43c, d).
344 5 Die warm-gemäßigte (subtropische) Zone5
Dornpolster-VegetationDornpolster sind dem Europäer vor allem aus den trockene-ren mediterranen Gebirgen bekannt, so z. B. aus dem Atlas,der Sierra Nevada, dem Ätna und dem südlichen Apennin,dem Olymp, dem Taurus und anderen griechischen und tür-kischen Hochgebirgen; ihre Hauptverbreitung haben sie je -doch in den mittelasiatischen Hochgebirgen Karakorum,Hindukusch und Pamir mit sommerlicher Trockenheit (Kür-schner 1986); von dort reichen sie bis in das Altai-Gebirge (s. Abb. 6-52i). Ihre natürlichen Vorkommen sind auf dieobere montane und untere alpine Stufe beschränkt. Ihrebenachbarten Formationen sind in der Regel mehr oder min-der lockere Wälder bzw. Gebüsche aus Pinus- und Juniperus-Arten. Das Klima zeichnet sich durch eine geringe, aberzuverlässige winterliche Schneedecke, relativ hohe Som-mertemperaturen mit Maxima noch auf 3.000 m NN nahe30°C und starker Sonneinstrahlung aus, gemildert durchgelegentliche Sommergewitter. In extremeren Lagen kom-men Dornpolster vorzugsweise nordexponiert vor und sindmeist stark windausgesetzt. Sie gedeihen eher auf ober-flächlich etwas bewegtem Schutt als auf Gesteinsrohböden,meist im Beweidungsbereich von Wiederkäuern wie Wildzie-gen, Wildschafen und Yaks.
Die Dornpolster stammen aus den Familien Apiaceae(Bupleurum, Mulinum, Platytaenia), Asteraceae (Centaurea),Caryophyllaceae (Acanthophyllum, Arenaria), Brassicaceae(Alyssum, Ptilotrichum, Vella), Euphorbiaceae (Euphorbia),
Fabaceae (Anthyllis, Astragalus, Caragana, Cytisus, Genista,Onobrychis, Oxytropis), Lamiaceae (Stachys, Thymbra),Plumbaginaceae (Acantholimon) und Rosaceae (Sarcopo -terium). Sie besitzen eine ausgeprägte Pfahlwurzel mitextensiver Verzweigung und sind meist immergrün. In derspanischen Sierra Nevada findet man z. B. Vella spinosa(Brassicaceae) und Astragalus sempervirens (Fabaceae), imAtlas Ptilotrichum spinosum (Brassicaceae) und Cytisusbalansae (Fabaceae), auf Kreta Euphorbia acanthothamnos,Thymbra capitata (Lamiaceae) und Sarcopoterium spinosum(Rosaceae). Die ausgedehnte Verbreitung der Dornpolster-Vegetation aus Astragalus-, Acantholimon- und Oxytropis-Arten in den iranischen und mittelasiatischen Gebirgen (wieim Alburz und Hindukusch; Breckle & Frey 1974, Noroozi etal. 2007) sowie ihre Ähnlichkeit mit den Mulinum spinosum-Halbwüsten Patagoniens (s. Abschn. 6.3.4.3; Abb. 6-36d)und den nemoralen Caragana gerardiana-Beständen im tibe-tischen Himalaya (Miehe et al. 2002) lassen vermuten, dasses sich hier um beweidungsresistente Wuchsformen in som-mertrockenen, winterkalten Gebieten handelt (Klötzli2004e). Dass solche Polsterfluren durch Waldvernichtungund anschließende Beweidung mit Haustieren (vornehmlichZiegen) heute auch anstelle von Wäldern stehen (wie aufKorsika an Südhängen bis Meeresniveau), zeigt wieder ein-mal, dass der menschliche Einfluss auch in vermeintlichextremen Lebensräumen nicht zu unterschätzen ist.
Kasten 5-10

55.5 Subtropische Hochgebirge 345
a b
c d
Abb. 5-43 Beispiele für die alpine Vegetation subtropischer Gebirge. a = Mosaik aus Rhododendron setosum-Zwergstrauchheideund Kobresia nepalensis-Gebirgssteppe im Zentralhimalaya (Nordbhutan; Foto G. Miehe); b = Dornpolsterflur (Kreta) mit Astraga-lus angustifolius (im Vordergrund) und Euphorbia acanthothamnos (gelbgrün); c, d = Hochgebirgshalbwüste aus Acantholimon lyco-podioides (Karakorum, Hunzatal,, Pakistan; Fotos F. Klötzli).


![Qualitätsbericht, Klinik in der Zarten [260832552]€¦ · garantiert gemäßigte Temperaturen im Sommer, einen sonnigen und nebelfreien Herbst, klare Winter und einen späten, zarten](https://img.pdfslide.org/doc/110x75/5f7e50b20879590680423071/qualittsbericht-klinik-in-der-zarten-260832552-garantiert-gemigte-temperaturen.jpg)


























