Embed Size (px)
Citation preview

Aus dem Institut fiir Pflanzenziichtung GroS-Liisewifz der Deutschen Akademie der Landwirtschaftswissenschaften zu Berlin
Ein experimenteller Test der Halbgeschwistercovarianz als Parameter zur Vorhersage des genetischen Gewinns
K. BELLMANN und I. GRIESS
Problem
Die Vorhersage des genetischen Gewinns auf der Grundlage der Ergeb- nisse von Sippenanalysen erfolgt nach
worin S die Selektionsdifferenz, a$ eine Schtitzung der Varianzkomponente Vater, P der fiir die betrachtete Population gultige Inzuchtkoeffizient und a;h die phanotypische Varianz ist. Betrachtet wird im folgenden die Eignung des Experiments I (COMSTOCK und ROBINSON, 1952), das eine hierarchisch klassifizierte Sippenanalyse darstellt. Bei dieser Analyse werden V Zufalls- pflanzen als Vater zufallig ausgewahlt und zur Schatzung von je m, stets anderer, ebenso zufallig ausgewahlter Pflanzen (Mutter) verwendet. Die Nachkommen jeder Mutter bilden eine Vollgeschwister-, die Nachkommen eines Vaters eine Halbgeschwisterfamilie. Aus anlagetechnischen Griinden erfolgt meist eiiie Zusammenfassung mehrerer Vater, d. h. Halbgeschwister- familien, zu einem Satz. Es ergibt sich das hierarchische Model1 (11):
y.. - t j k l - + ‘i -k bij + cijk f eijkl
a, b, Ci jk : Effekte der k-ten Mutter des j-ten Vaters (m = Zahl der Mutter je
eijkl : Effekte des I-ten Nachkommen der k-ten Mutter ( n = Zahl der
Die Varianztabelle enthalt die folgenden M Q : MQ,, MQ,, MQM, MQN
: Effekte des i-ten Satzes (s = Zahl der Satze) : Effekte des j-ten Vaters im i-ten Satz (w = Zahl der Vater je Satz)
Vater)
Nachkommen je Mutter).
mit den Erwartungswerten (orthogonaler Fall) E ( M Q , ) = u’+ nu$ + n s m a , 2
E(M Q N ) =
E ( M Q M ) = CT; + 12 2
19 Biometr. Z. 11,4

266 K. BELLMAEX und I. GRIESS
Hieraus konnen bekanntlich die Schatzungen der Variaiizkomponeiiten fiir Vater, Mutter und Nachkommen erhalten werden.
u$ entspricht der Covarianz zwischen Halbgeschwistern, fur die hinsichb lich der genetischen Varianzkomponenten folgende Zusammensetzung gilt ( F =k 0 ) :
4 4 - - . ~ ~- 6; bzw. G p Cov(H G) ist danacli der Zahler der Schatzung des 1 + F Heritabilitlitskoeffizienten
dessen Sicherheit die Grundlage fur eine brauchbare Vorhersage des gene- tischen Gewinns in (1) ist.
Die Giiltigkeit von (2), d. h. die Giiltigkeit der Interpretation (gelegentlich als ,,genetischer Erwartungswert" bezeichiiet) der phinotypischeii Varianz- komponente $v in Termini genetischer Varianzkomponenteii (ui usw.) in (2) ist nur dann gegeben, wenn die Modellannahmen, die dem Experiment zur Schatzung der additiven genetischen Varianz zugrunde liegen, in der betrachteten Population erfiillt sind. Bei Erfullung dieser Annahmen mufiten Populationen vorliegen, die im Hinblick auf ihre geiietische Struktur Quasi-Ein-Locus-Fklle mit Gleichgewichtsstruktur darstellen. COYSTOCX und ROBINSON (1 952) fordern
I. diploides Verhalten,
2 . Genfrequenzen 0,5 an allen spaltenden Loci,
3. keine multiple Allelie,
4. keine Kopplung bzw. kein Kopplungsphasengleicligewicht , 5. keine Epistasie.
Man muate noch hinzusetzen : 6. unendlich groDe Populationen und 7. Population unterliegt keiner gerichteten Selektion (Gefahr der Heraus- bildung von gametischem Ungleichgewicht, vgl. u. a. NEI 1963, FELSEN-
Mit Ausnahme der Forderung 1 mu0 a priori fur echte Populationen angenommen werden, dal3 keine der genannten Voraussetzungen erfullt ist, und man mu13 fragen, welches Vertrauen h2-Schatzungen auf der Basis von Cov ( H G ) entgegengebracht werden kann.
Eine Entscheidung dariiber, ob die nach (3) erhaltene Schatzung h$,, geeignet ist, 5 G hinreichend genau vorherzusagen, kann getroffen werden,
STEIN 1965).

Experjmenteller Test der Halbgesch~~istercovarianz zur Vorhersage 267
wenn A G(,,, [nach ( l ) ] mit dem tatsachlich erzielten Selektionsgewinn in der betrachteten Population I? verglichen wird, d. h. Vergleich
A
AG(,G, VS. I? bzw.
2 h
( H G ) vs. x = I;(",.) h?
(h,2,., = realisierte Heritabilitat).
Mater ia l und Methode
Es wurden hierzu mehrjahrige experimeiitelle Untersuchungen vorgenom- men, uber die berichtet werden soll. Als Versuchsobjekt diente Zea mays (Sorte ,,Schindelmeister") und Secale cereale (Landsorte aus Bruck a. d. Laita, Osterreich). Beim Mais erfolgte die Selektion in der Ausgangspopula- tion au f Temperaturempfindlichkeit des Langenwachstums (Parameter b, der Regressionefunktion yi = u + b, (ti - t 0 ) 2 + b w (wt - wo)2, worin die yi die Liingenzunahmen in 24 Stunden, die ti die mittlere Tagestemperatur und die wt die mittleren Windgeschwindigkeiten in dieser Zeit sind; vgl. die Phanometrieuntersuchungen am Mais : BELLMANN et al. 1962, 1964, 1967 ; MRAZEK et al. 1965; RAEUBER et a1 1961) und auf Wachstumsleistung bei 13 "C (ylaOc) selektiert ; beim Roggen wurde auf Blattbreite der Rosetten- blitter und auf Zahl der Bestockungstriebe im zeitigen Friihjahr selektiert. Die Selektion nahmen wir in 2 Richtungen vor; d. h. aus den auf hohe Werte selektierten Populationen (Plus-Populationen) wurden stets die V O / ~
besten, aus den auf niedrige Werte selektierten Populationen (Minus- Populationen) stets die v0l0 schlechtesten Pflanzen ausgewiihlt. Es erfolgte eine viermalige Auslese. Die standardisierten Selektionsdifferenzen sind in Abb. i und 2 angegeben. Bei Mais erfolgte eine Auslese von 4 aus durch- schnittlich 180, bei Roggen von durchschnittlich 50 aus durchschnittlich 1200 Pflanzen (der Umfang der Auslesepopulationen und die Zahl der selektierten Pflanzen variierte von Jahr zu Jahr urn etwa 150/,). Bei allen untersuchten Merkmalen erfolgte die Auslese vor Bluhbeginn ; die selek- tierten Pflanzen bliiliten panmiktisch unter sich ab.
Zur Vermeidung von Umweltinteraktionen haben wir alle 9 Populationen ( I+ bis 4+, 1- bis 4-, Grundpopulation) gemeinsam (d. h. unter gleichen Umweltbedingungen) angebaut und die Populationsparameter (Mit- tel : Z T , Varianz : sib) und die Trunkationspunkte sowie die Elternmittel (5,) fur die n-te Generation bestimmt. Es gilt dann
'
wobei und

268 K. BELLMANN und I. GRIESS
Die Sippenanalysen wurden 3jlihrig (Mais) bzw. 2jahrig (Roggen) wieder- holt. In jedem Jahr haben wir 32 Vaterpflanzen mit je 4 Mutterpflanzen gekreuzt und im Folgejahr die Vollgeschwisterfamilien mit je 7 Nachkom- men angebaut. Die 3jahrigen Ergebnisse wurden zu einer Gesamtinfor- mation gepoolt, da dadurch aus stichprobentheoretischer Sicht und wegen moglicher Genotyp-Umwelt-Wechselwirkungen eine relativ sichere Schat- zung fur Cov ( H G) vorliegt
die auf N = 2688 (Mais) bzw. N = 1792 (R.oggen) beruht. Entsprechend wurde 6$h errechnet :
wobei
und dann
Ergebnisse
Die Ergebnisse der Selektionsversuche sind in Abb. 1 und 2 dargestellt und zeigen, da13 bei den 4 betrachteten Merkmalen mit unterschiedlichem Erfolg selektiert worden ist. Wahrend Selektion auf Blattbreite und Be- stockungstriebe erfolgreich war (vgl. h&), trifft dies fur bt und y13 oc nicht zu. (Eine zuchtungstheoretische Betrachtung in ertragsphysiologischer Hinsicht erfolgt zusammen mit weiteren Ergebnissen an anderer Stelle. )
Im Fail der Maisselektionen unterscheiden sich die Phanotypenmittel (und damit auch Genotypenmittel) der Plus- und Minuspopulationen weder untereinander noch von der Ausgangspopulation wesentlich. Im Mittel des Gesamtversuchs wurden durchschnittliche Weritabilitatskoeffizienten fiir b,von 0,07, x;r,fur y130c von 0,09 rnit mittleren Streuungen 3 (errechnet
nach PROUT'S Formel, 1962) von 0,07 (fur b,) undvon 0,06 (fur yisoc) errechnet (vgl. Tabelle 1).
In Tabelle 2 sind die entsprechenden Werte fur die Selektion auf die Roggenmerkmale gegeben. Hier ergibt sich fur die Anfangsphase der Selektion (Selektion von der 0. zur 1. und 1. zur 2. Generation) e k e mittlere realisierte Heritabilitat x;,.) = 0,34 fur Blattbreite und %& = 0,12 fur
Z(t9
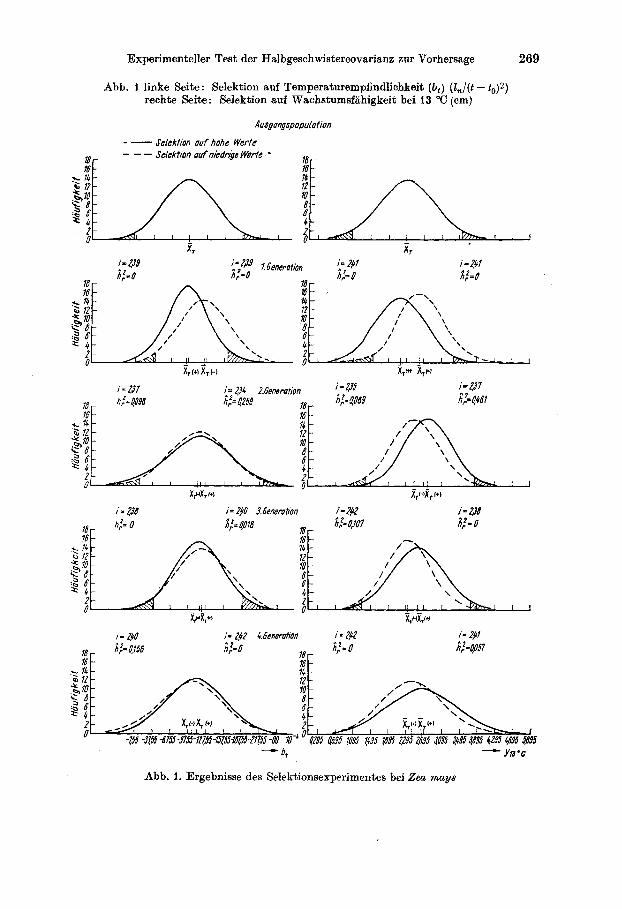
Experimenteller Test der Halbgeschwistercovarianz zur Vorhersage 269
Abb. I linke Seite: Selektion auf Temperaturempfindlichkeit (6,) (Z,/(t - t , )z) rechte Seite: Selektion auf Wachstumsfiihigkeit bei 13 "C (cm)
Ausgongspopuiolion
- - Selektion auf hohe Werte
>;I A . l
12 -3 '0 Q 8 2 6 -k
*: 0 -
%
Abb. 1. Ergebnisse des Selektionsexperimentes bei Zea mays
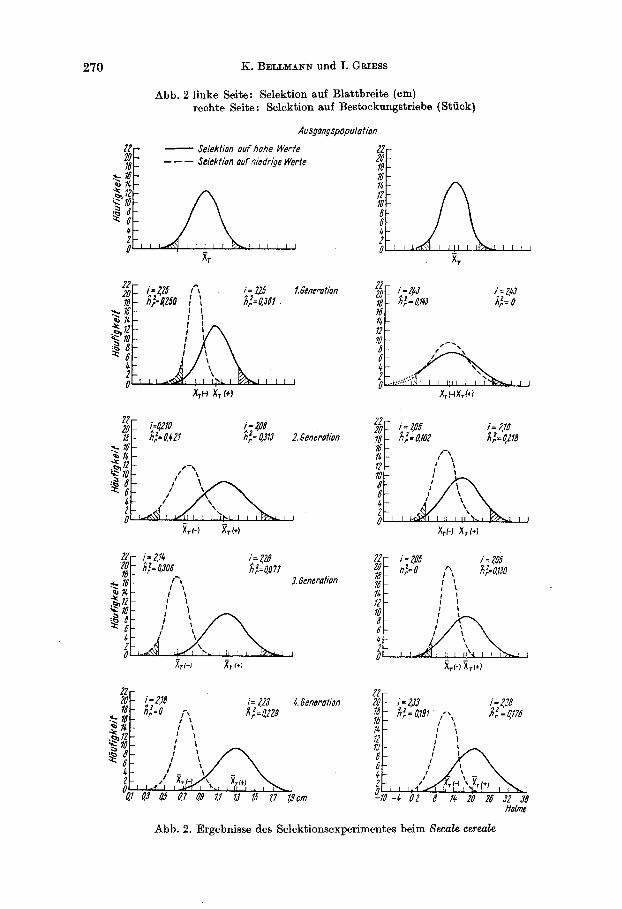
270 K. BELLMANN und I. GRIESS
Abb. 2 linke Seite: Selektion auf Blattbreite (em) rechte Seite : Selektion auf Besbckungstriebe (Stuck)
Ausyanyspopulation
Selektion auf hohe Werte - 22 ;;E _ _ _ Selektion auf niedrige Werle
, Generation
X,l-I X , l + )
Abb. 2. Ergebnisse des Selektionsexperimentes beim Secale cereale
Experimenteller Test der Halbgeschwistercovarianz zur Vorhersage 271
0,81 0,091 0,42 0,03
0,62 0,115 0 0,Ol 0,72 0,114
0,73 0,168 0,31 0,Ol I 1- 2 - 3 4
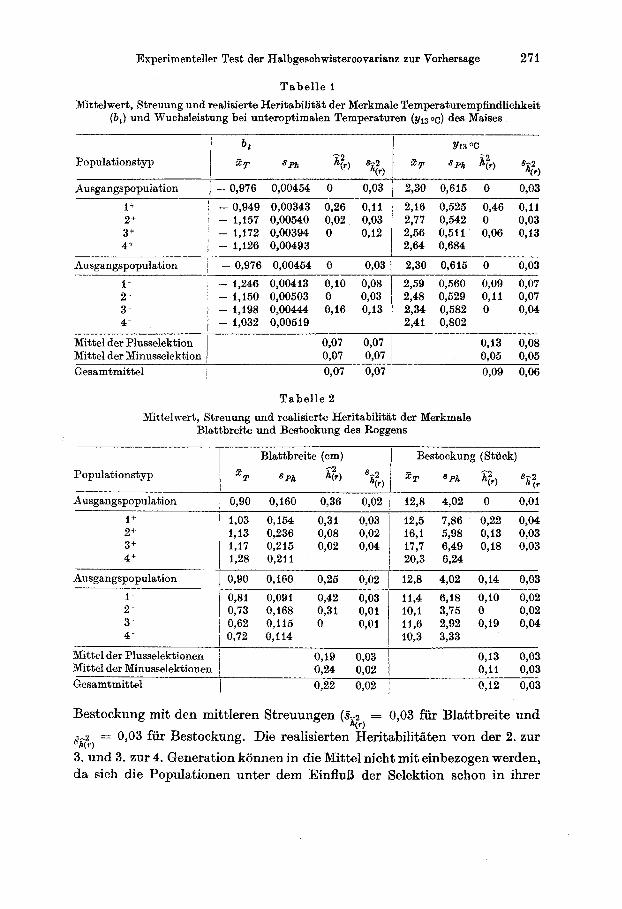
Tabel le 1 Nlittelwert, Streuung und realisierte Heritabilitilt der Merkmale Temperaturempfindlichkeit
( a , ) und Wuchsleistung bei unteroptimalen Temperaturen (yi3 OC) des Maises
11,4 6,18 0,lO 0,02
11,6 2,92 0,19 0,04 10,3 3,33
10,l 3,75 0 0,02
Populationstyp
Ausgangspopulation I - 0,976 0,00454 0 0,03 1 2,30 0,615 0 0,03
1' I - 0,949 0,00343 0,26 0,11 2,16 0,525 0,46 0 , l l 2+ ~ - 1,157 0,00540 0,02 0,03 2,77 0,542 0 0,03 3+ , - 1,172 0,00394 0 0,12 I 2,56 0,511 0,06 0,13 4f I - 1,126 0,00493 1 2,64 0,684
Ausgangspopulation ~ - 0,976 0,00454 0 0,03 1 2,30 0,615 0 0,03
~ - 1,246 0,00413 0,lO 0,08 ~ 2,59 0,560 0,09 0,07
I - 1,150 0,00503 0 0,03 2,48 0,529 0 , l l 0,07
0,13 0,08 0,07 0,07 , 0,05 0,05
0,09 0,06
1' 2 -
4- - 1,032 0,00519 1 2,41 0,802 3 - - 1,198 0,00444 0,16 0,13 234 0,582 0 0,04
Mittel der Plusselektion Mittel der Minusselektion Gesamtmittel ~ 0,07 0,07 1
Tabelle 2
Mittelwert, Streuung und realisierte Heritabilitiit der Merkmale Blattbreite und Bestockung des Roggens
Blattbreite (em) 1 Bestockung (Stuck) I
1
j Populationstyp 1 'T 8Ph $) 8-42 h(?) 1 '!l' ' P h hfr) 'X&
Ausgangspopulation 1 0,90 0,160 0,36 0,02 I 12,s 4,02 0 0,Ol
I+ 2 +-
3f 4+
1,03 0,154 0,31 12,5 7,86 0,22 0,04 1,13 0,236 0,OS :l:i I 16,l 5,98 0,13 0,03 1,17 0,215 0,02 0,04 17,7 6,49 0,18 0,03 1,28 0,211 1 20,3 6,24

272 K. BELLMAXX und I. GRIESS
b , (alle Werte x 10-6) 1964 1965 1966 Mittel
genetischen Struktur verandert haben. Diese 4 mittleren realisierten Heri- tabiIitiiten dienen als Vergleichskriterium fur die h i G , die durch mehrjiihrige Sippenanalysen bestimmt worden sind.
In Tabelle 3 und 4 sind diese Schiitzungen fur h;HG) angegeben. Da eine weitgehende Ubereinstimmung zwischen %& und zu beobachten ist, kann man annehmen, daD sich brauchbare Vorhersagen von A G auf Grund von h2-Schltzungen aus Sippenanalysen auch dann erhalten lassen, wenn *
die vorn genannten Voraussetzungen zum ubergang von den phano- ty pischen zu den genotypischen Varianzkomponenten nicht erfullt sind Es besteht die Tendenz der Uberschiitzung des genetischen Gewinns (z. B. Bestockung). Bemerkenswert ist, daB die Obereinstimmung zwischen
Tabelle 3
Auf der Grundlage von dreijiihrig wiederholten, hierarchisch klassifizierten Sippenanalysen geschatzte Heritabilitat xfRG) und in einem vierjahrigen Selektionsexperiment (doppelte Trunkation) ermittelte realisierte Heritabilitiit (%&$ fur 2 physiologische Wachstums-
parameter
-
y (13 "C) (alle Werte x 10-3) 1964 1965 1966 Mittel
c2Ph
q H a )
14,73 12,39 12,18 13,lO 419,7 81,2 196,O 232,3
0,055 0 0,179 0,076 0,118 0,415 0,233 0,154 0,012 0,006 0,011 0,009 0,025 0,029 0,028 0,027
~ Blattbreite (alle Werte x 10-3) Bestockung j 1967 1968 Mittel I 1967 1968 Mittel
5% ! 0,6 495 1 9 1 3,4 273 z%h 17,6 3491 2::; 1 25,3 38,8 32,l
0,353 0,201
S 0,147 0,220
i?HG) I 0,136 0,528
0,006 0,018 OTao5 I 09174
i
q 7 ) (vgl. Tab. 1 0,07 und Text) 0,07
Sh &?) 1 1 ( 7 )
(vgl. Tab. 2 0,337 1 (vgl. Tab. 2 0,116 und Text) 0,03 ~ und Text) 0,03
(vgl. Tab. 1 0,09 und Text) 0,06

Experimenteller Test der Halbgeschwistercovarianz zur Vorhersage 273
realisierter Heritabilitat und dem entsprechenden Schatzwert aus H G- Analysen nur dann erzielt werden kann, wenn genugend groBe SchLtzpliine (mehrjahrig wiederholt) vorliegen und auch h& sicher genug bestimmt ist (mehrmalige Selektion und/oder genugend groBe Anzahl selektierter Eltern). Beobachtete Nichtubereinstimmung bei Vergleichen von Einzel- ergebnissen (vgl. Tab. 3, 4), d. h. Vergleichen von Einzelschatzung von h& und von h&, die auf geringem Versuchsumfang basieren und durch G-U- Interaktionen belastet sind, konnen zu der wahrscheinlich falschen SchluB- folgerung verleiten, daB nicht erfullte Modellannahmen zu wheblichen Ver- zerrungen von h&, und damit zu unbrauchbaren Vorhersagen des zu erwartenden genetischen Gewinns fuhren. Nach diesen Untersuchungen und aus Simulationsstudien zum gleichen Problem (Vortrag BELLMANN, AHRENS 1968) mit vollkommen gleichen Ergebnissen kann vielmehr fol- gender SchluB gezogen werden :
Bei genugend groBen Versuchsumfangen von Sippenanalysen (min- destens 100 Vatergruppen, Gesamtumfang N = 2000 bis N = 3000) konnen hinreichend sichere Schatzungen des genetischen Gewinns vorgenommen werden. Das stimmt mit statistischen Untersuchungen von HERRENDORPER (1 968) uberein. Dabei ist anzustreben, daB mehrere Sippenanalysen mittleren Umfangs aus verschiedenen Umweltverhaltnissen (Jahre, Orte) zusammen- zufassen sind.
Auf Grund der Ergebnisse der Selektionsversuche erscheint es uns un- gerechtfertigt, die Nichtubereinstimmung von vorhergesagtem und tat- sachlich realisiertem Gewinn im praktischen Zuchtablaufnur auf Fehler bei der Sippenanalyse zuruckzufuhren. R (d. h. der erzielte genetische Gewinn) ist eine ZufallsgroBe mit einem Erwartungswert und einer Varianz (vgl. PROUT, 1962). Hieraus folgt, daB der Zuchter eine vorhergesagte Heri- tabilitat erst im Verlauf mehrerer Selektionsschritte und bei genugend grol3en Populationen (Zahl der selektierten Eltern !) realisiert. Der quanti- tativ-genetisch arbeitende Zuchtungsforscher ist uberfordert, den zu er- wartenden genetischen Gewinn fur die Einzelsituation (einmalige Selektion, ein Jahr, kleine Populationen) vorherzusagen,
A
,
Zusammenf assung
Es werden mehrjahrige Ergebnisse uber einen Vergleich der realisierten Heritabilitat mit der auf Grund von Sippenanalysen vorhergesagten Heritabilitat fur quantitative Merkmale mit kontinuierlicher Variation mitgeteilt. Die in einem Selektionsversuch mit
doppelter Trunkation festgestellte mittlere realisierte Heritabilitiit ( 9) stimmte weit- gehend mit der mittleren, auf der Basis von Covarianzen zwischen Halbgeschwistern ge- schatzten Heritabilitat (%tHG,) uberein.
-
-

274 K. BELLMANN und I. GRIESS
Aus den Ergebnissen, die auch bei Simulationsstudien erhalten wurden, kann man folgern, daf3 nicht erfiillte Modellannahmen, die einer h2-Schiitzung aus Sippenanalysen zugrunde liegen, keine Einschriinkung der Anwendbarkeit dieser Sippenanalysen zu sein brauchen, wenn der Versuchsumfang groD genug ist ( N = 2000 bis 3000) und gepoolte Information aus verschiedenen Jahren vorliegt.
Summary
Results with regard to a comparison of realized heritability (h??,) of characters with continuous variation in maize and rye based on sib analysis replicated over several years are reported. The realized heritability observed during a selection experiment (double truncation) agreed widely with the heritability estimated by means of covariances of half sib family means in a hirarchical classified analysis of variance. The results, supported by adequate Monte-Carlo-simulation, which are previously reported, indicate that non-ful- flment of model assumptions (put as a foundation of h2-estimation by means of sib analy- ses) need not be considered as a restriction of applicability of COMSTOCK and ROBINSON’S experiment I in every case, if the sample size is large enough ( N = 2000 to 3000) and if pooled information from different years is available.
L i t e r a t u r
BELLMANN, K., und H. AHRENS, 1968: Die Schatzung von genetischen Varianzen bei Kopplung und Epistasie. Vortrag auf der intern. biometrischen Konferenz, Budapest, Mhrz 1968.
BELLMANN, K., A. RAEUBER, G. I ~ I N L , E. ABERG, CH. PFEFFER, A. WINKEL, 0. MRLZEK und A. SCETNEKI, 1962: Phiinometrische Untersuchungen am Mais. Tagungsbericht DAL Berlin, Nr. 48, 131-138.
BELLMANN, K., G. MEINL und A. RAEUBER, 1964: Mehrjiihrige phanometrische Unter- suchungen an einem groBeren Maissortiment in GroB-Lusewitz. Zuchter 34, 273-286.
BELLMANN, K., A. RAEUBER, A. WINKEL, CH. PFEFFER, E. ABERG und 0. MR~ZEK, 1967: Ergebnisse eines Phanometrieversuches beim Mais an verschiedenen klimatischen Standorten Nord-, Mittel- und Sudeuropas . (Ein Beitrag zur Klimaresistenzziichtung des Maises). Zeitschr. f . Pflanzenziichtung 67, 293-316.
COMSTOCK, R. E., and H. F. ROBINSON, 1952: Estimation of average dominance of genes. In J. W. Gowen: Heterosis 494-516.
FELSENSTEIN, J., 1965: The effect of linkage on directional selection. Genetics 52,349-363. HERREND~RFER, G., 1968 : Beitrag zur Versuchsplanung der Schiitzung des Heritabilitiits-
koeffizienten von Merkmalen unserer Haustiere (Rind, Schwein, Schaf). Diss. Rostock, 73 S.
MR~ZEK, O., A. RAEUBER, K. BELLMANN, G. MEINL, CH. PFEFFER, A. WINKEL, A. KOVACS, A. SCETNEKI und V. TARAU, 1965 : Internationale phanometrische Untersuchungen am Mais (tschech.). Vedeck6 Prace Vyskumneto Ustave Kukurice v Trnave 1, 127-138.
NEI, M., 1963: Effect of selection on the components of genetic variance. In: Stat. Gen. and Plt. Breed. Ed. Hanson and Robinson NC State Coll. Raleigh. S. 501-515.
PROUT, T., 1962: The error variance of the heritability estimate. Biometrics 18, 404407. RAEUBER, A., K. BELLMANN, G. MEINL, 0. MRBZEK, CH. PFEFFER und A. WINKEL, 1961:
Anwendung nichtlinearer Korrelationen bei phanometrischen Arbeiten beim Mais. Zeitschr. f. Pflanzenzuchtung 56,433442.
Emgang des Manuskripts: 5. 9. 68 Anschrift der Verfasser : Dr. K. BELLMANN 11 14 Berlin-Blankenburg GeorgenstraBe 22 J. GRIESS 2551 GroB-Liisewitz Am Sportplatz











![Zum Problem des Genetischen Lehrens - Martin Wagenscheinmartin-wagenschein.de/en/2/W-172.pdf · Martin Wagenschein Zum Problem des Genetischen Lehrens* [W172] Einige Erfahrungen und](https://img.pdfslide.org/doc/110x75/5b9f242f09d3f2ab0b8cf3f4/zum-problem-des-genetischen-lehrens-martin-wagenscheinmartin-martin-wagenschein.jpg)

















