Embed Size (px)
Citation preview

RedaktionF. Moosig, Lübeck/Bad Bramstedt O. Distler, Zürich
Z Rheumatol 2011 · 70:198–204DOI 10.1007/s00393-010-0692-3Online publiziert: 26. Januar 2011© Springer-Verlag 2011
J.U. Holle1 · S. Wieczorek2 · J.T. Epplen2 · W.L. Gross1
1 Poliklinik für Rheumatologie, Vaskulitiszentrum, Universitätsklinikum Schleswig-Holstein, Campus Lübeck, Klinik für Rheumatologie und Klinische Immunologie, Klinikum Bad Bramstedt2 Abteilung für Humangenetik, Ruhr-Universität, Bochum
Genetischer Hintergrund der Vaskulitiden
Leitthema
Die Kenntnis der genetischen Risiko-faktoren, die zu Autoimmunerkran-kungen wie Vaskulitiden beitragen, könnten wertvolle Hinweise auf die Pathogenese liefern und damit auch gezielte therapeutische Ansätze er-öffnen. Erste interessante Variatio-nen, vor allem Einzelbasenaustau-sche (so genannte „single nucleoti-de polymorphisms“, SNPs), sind in der Vergangenheit durch den so genann-ten Kandidatengen-Ansatz für ver-schiedene Vaskulitiden identifiziert worden. Die Ergebnisse von Kandi-datengen-Studien zeigen hier insbe-sondere für die ANCA-assoziierten Vaskulitiden, dass neben bestimm-ten „Autoimmunitätsgenen“, die zu mehreren Autoimmunerkrankungen prädisponieren, einzelne Risikogene existieren, die spezifisch für eine AN-CA-assoziierte Vaskulitis (AAV; Z. B. Wegener-Granulomatose/WG, aber nicht Churg-Strauss-Syndrom/CSS) oder auch Unterformen der verschie-denen AAV (Z. B. lokalisiert, nicht ge-neralisiert) sind. In der Zukunft sol-len genomweite Assoziationsstudien (GWAS) mehr Aufschluss über die ge-netischen Risiken einer Erkrankung geben, erfordern aber zum Erzielen statistisch signifikanter Ergebnisse eine deutlich höhere Fallzahl. Dieser Artikel gibt eine Übersicht über heu-te verfügbare Ansätze zur Identifizie-rung genetischer Risikofaktoren so-wie die bisher wichtigsten identifi-zierten Risikogene bei den Vaskuli-tiden und ihre pathophysiologische Bedeutung.
Kandidatengen-Ansatz versus genomweite Assoziationsstudie
Beide heute verfügbare Ansätze zur Iden-tifizierung genetischer Risiken haben Vor- und Nachteile [16, 24]: Der Kandi-datengen-Ansatz hat den Vorteil, dass im Vergleich zur GWAS eine statistische Si-gnifikanz mit einer geringeren Proben-zahl erreicht wird, ist aber dadurch limi-tiert, dass stets nur einzelne Gene bzw. Polymorphismen untersucht werden, die der Forscher als möglicherweise relevant für die Erkrankung einstuft. Die GWAS hingegen ist nicht Hypothesen-getrie-ben, erfährt aber – bedingt durch den Ansatz einer simultanen Untersuchung einer enormen Anzahl genetischer Varia-tionen – einen im Vergleich zum Kandi-datengen-Ansatz deutlichen Verlust sta-tistischer „Power“ (Signifikanzlevel typi-scherweise bei p<10−5), sodass eine valide Analyse nur mit großen Fallzahlen mög-lich sind.
Risikogene bei Großgefäßvaskulitiden
Sowohl bei der Riesenzellarteriitis (RZA) als auch bei der Takayasu-Arteriitis (TA) konnten bisher nur Assoziationen mit der HLA-Region im Rahmen von Kandida-tengen-Analysen als verlässlich replizier-bar herausgearbeitet werden. Dabei be-steht für die TA eine sehr starke Assozi-ation mit HLA-B52 (vor allem mit dem Allel HLA-B*5201; „Odds Ratio“/OR 4,7–10,2; [21]) und für die RZA eine Assozi-ation mit dem HLA-DRB1*04-Allel (OR 2,18–3,84; [9]). HLA-Assoziationen wur-
den für diverse Erkrankungen beschrie-ben.
E Man nimmt an, dass bestimmte HLA-Allele zu einer Konformation der MHC-Peptide führen, welche die Präsentation distinkter (Auto-)Antigene erleichtern.
Für die RZA sind weitere sieben Poly-morphismen in insgesamt zwei Kohor-ten durch Kandidatengen-Analyse iden-tifiziert worden, die eine Assoziation mit einem weiteren HLA-Locus sowie mit Genpolymorphismen von Zytokinen, Adhäsionsmolekülen und Wachstums-faktoren beschreiben (HLA-DRB1*04, ICAM1 241, IL-6 174, IL-10–1082/-592, ENOS 298, VEGF 634; [16]), ohne je-doch in der jeweils anderen Kohorte re-pliziert werden zu können. Die Ergebnis-se sind daher als vorläufig zu betrachten (. Tab. 1).
Risikogene bei Kleingefäßvaskulitiden
Für die AAV konnten in den letzten Jah-ren mehrere relevante Genpolymorphis-men identifiziert und repliziert werden. Dabei zeigt sich, dass die drei verschiede-nen Entitäten WG, mikroskopische Poly-angiitis (MPA) und CSS, die als AAV zu-sammengefasst werden, sowohl gemein-same genetische Risikofaktoren haben als auch solche, die nur jeweils eine oder zwei dieser Erkrankungen auszeichnen (. Tab. 1). Nachfolgend sind Ergebnis-se aus Kandidatengen-Analysen zusam-mengefasst. Eine GWAS, die auf der In-
198 | Zeitschrift für Rheumatologie 3 · 2011


itiative einer internationalen Koopera-tion der Europäischen Vaskulitis Stu-diengruppe (EUVAS) und des ameri-kanischen VCRC („Vasculitis Research Consortium“) basiert und Patienten mit WG und MPA eingschlossen hat, ist der-zeit in Auswertung. Ergebnisse werden in 2011 erwartet.
PTPN22-Allel
So konnte das PTPN22*620W-Allel als Risikofaktor für die WG (OR 1,76 für alle WG-Patienten, OR 2,01 in ANCA-positi-ver WG; [13]) und die MPA (OR 2,55; [5]) identifiziert werden, nicht jedoch für das CSS (unpublizierte Daten). Interessant in diesem Zusammenhang ist, dass die-se genetische Assoziation nur für die AN-CA-positive WG gefunden wurde. Auch die MPA, die in der Regel ANCA-positiv ist, weist diese Assoziation auf, während dieses Allel für CSS-Patienten, die in der Mehrzahl ANCA-negativ sind, keinen Ri-sikofaktor darstellt. Betrachtet man nur ANCA-positive CSS-Patienten, so scheint das PTPN22-Allel hier eine ähnliche Ef-fektgröße zu haben wie in ANCA-positi-ven WG- und MPA-Patienten (unpubli-zierte Daten).
>PTPN22 fungiert als Autoimmunitätsgen und ist mit einem positiven Auto-antikörperstatus assoziiert
Das PTPN22*620 W-Allel konnte auch für andere Autoimmunerkrankungen als Risikogen identifiziert werden („Auto-immunitätsgen“) und scheint auch bei anderen Erkrankungen mit einem posi-tiven Autoantikörperstatus assoziiert zu sein (z. B. mit anti-CCP-positiver rheu-matoider Arthritis/RA; [19]). Das Gen-produkt von PTPN22 entspricht einer Tyrosinkinase (LYP), die u. a. in T-Zel-len exprimiert wird und physiologi-scherweise das „T-Zell-Signaling“ sowie die Produktion des Zytokins Interleu-kin- (IL-)2 inhibiert. Der Austausch von Arginin zu Tryptophan an der Position 620 (PTPN22*620 W) induziert ein ver-mindertes „T-Zell-Signaling“, wobei bis-her nicht genau verstanden ist, auf wel-che Weise diese Mutation zur Autoim-munität prädisponiert. Möglicherweise führt dies zu einer verminderten negati-ven T-Zell-Selektion im Thymus oder zu einer verminderten Aktivität regulatori-scher T-Zellen.
Tab. 1 Übersicht über die wichtigsten, bisher identifizierten genetischen Risikofaktoren bei Vaskulitiden
Vaskulitis Gen/Allel/Poly-morphismus
Funktion des Genprodukts Odds Ratio Litera-tur
Riesenzellarteriitis HLA-DRB1*04 Antigenpräsentation 2,18–3,84 [9]
Takayasu-Arteriitis HLA-B*5201 Antigenpräsentation 4,7–10,2 [21]
Kawasaki-Syndrom ITPKC, rs28493229 Signaltransduktion (TCR) 1,74 [26]
Wegener-Granulo-matose
HLA-DPB1*0401 Antigenpräsentation 3,91 [10, 12]
HLA-DR4 1,7 [18]
Pi*Z Proteinaseinhibitor (auch von PR3) 1,47–14,62 [7, 15]
PTPN22*620 W Tyrosinphosphatase, Signaltrans-duktion (TCR)
1,76–2,01 [13]
Mikroskopische Polyangiitis
PTPN22*620 Signaltransduktion 2,55 [5]
Churg-Strauss-Syn-drom
HLA-DRB1 Antigenpräsentation [20, 23]
HLA-DRB3 0,54–0,61
HLA-DRB4 1,87–2,45
IL10.2-Haplotyp IL-10-Produktion 2,16 [22]
M. Behçet HLA-B51 Antigenpräsentation 5,78 [27]
ENOS, E298D Produktion von NO durch Endo-thelzellen
0,82–4,3 [3, 6]
MEFV (M694 V u. a.)
IL-1-Produktion (Aktivierung des „innate“ Immunsystems)
2,06–5,2 [2, 11]
TCR T-Zell-Rezeptor, PR3 Proteinase 3, IL Interleukin, NO Stickstoffmonoxid.

HLA-DPB1-Allele
Neben dem Autoimmunitätsgen PTPN22*620 W besteht für die WG eine noch stärkere Assoziation mit einem HLA-Allel (HLA-DPB1*0401; [12]) und dem Defizienzallel Pi*Z des Alpha-1-Antitrypsin-Gens [7, 15]. Diese HLA-As-soziation findet sich weder bei der MPA [1] noch dem CSS ([1, 23], . Abb. 1) und ist ebenfalls wie PTPN22 mit einem posi-tiven Autoantikörperstatus assoziiert [10]. Weitere bereits replizierte HLA-Assozia-tionen sind der HLA-DR4-Serotyp für die WG sowie HLA-DRB1, -DRB3 und -DRB4 für das CSS [18, 20, 23].
Die Daten aus . Abb. 1 fassen für WG und CSS mehrere unabhängige Arbeiten zusammen, welche übereinstimmende Ergebnisse lieferten [1, 10, 12, 23]. In einer britischen Kohorte fand sich ein weitge-hend übereinstimmendes Häufigkeits-muster für die jeweiligen Entitäten [1]. HLA-DPB1*0401 scheint somit ein relativ spezifischer genetischer Risikofaktor zu sein, welcher die WG von anderen AAV abgrenzt. Die Assoziation mit der WG be-trifft dabei insbesondere die PR3-ANCA-positiven Patienten (nicht dargestellt).
Alpha-1-Antitrypsin
Alpha-1-Antitrypsin ist in der Lage, PR3, das Autoantigen der WG, zu binden und zu neutralisieren. Da eine erhöhte „Ver-fügbarkeit“ von PR3 (z. B. erhöhte Mem-bran-PR3-Expression auf Neutrophilen) mit der WG und einer erhöhten Rezidiv-rate bei WG in Verbindung gebracht wer-den konnte [17], könnte funktionsfähiges Alpha-1-Antitrypsin durch Neutralisation von PR3 einen protektiven Effekt haben, eine Alpha-1-Antitrypsin-Defizienz aber möglicherweise begünstigend im Hin-blick auf die „Verfügbarkeit“ von PR3 und damit der WG sein.
IRF5-Gen
Ein Haplotyp des IRF5- („interferon regu-latory factor 5“-) Gens hat einen protek-tiven Effekt bei der WG (OR 0,73). Die-ser Effekt ist bei systemischer WG größer als bei lokalisierter WG [25]. Obwohl die Anzahl der Patienten mit lokalisierter WG in der Studie klein war und der Replika-
Zusammenfassung · Abstract
Z Rheumatol 2011 · 70:198–204 DOI 10.1007/s00393-010-0692-3© Springer-Verlag 2011
J.U. Holle · S. Wieczorek · J.T. Epplen · W.L. GrossGenetischer Hintergrund der Vaskulitiden
ZusammenfassungGenetische Assoziationsstudien haben auf dem Gebiet der chronischen Entzündungs- und Autoimmunerkrankungen wesentliche Erkenntnisse für die zugrundeliegende Pa-thophysiologie erbracht. So sind genetische Risikofaktoren identifiziert worden, welche die Suszeptibilität für bestimmte Erkrankun-gen (oft mehrere) erhöhen, ohne notwendi-gerweise direkt in die Pathogenese oder pa-thogenetischen Abläufe involviert zu sein. Für die Autoimmunvaskulitiden konnten erste Ri-sikogene bzw. -polymorphismen identifiziert werden. Einen limitierenden Faktor stellt die oft geringe Anzahl an Probanden dar. Ferner sind ethnische Unterschiede bei der Analyse ein Problem. Nur wenige der detektierten Ri-sikogene bzw. -polymorphismen wurden da-her an einer großen Kohorte und/oder auch verlässlich repliziert, z. B. PTPN22*620 W für Wegener-Granulomatose (WG) und mikros-kopische Polyangiitis (MPA), die Alpha-1-Anti-
trypsin-Defizienz sowie das HLA-DPPB*0401 für die WG oder der HLA-DRB3/DRB4 für das Churg-Strauss-Syndrom. Genomweite Asso-ziationsstudien (GWAS) bieten die Möglich-keit, das gesamte Genom auf genetische Ri-sikofaktoren zu screenen, erfordern aber die Untersuchung noch größerer Kohorten zum Erreichen statistisch signifikanter Ergebnisse. Ergebnisse aus GWAS zu M. Behçet und dem Kawasaki-Syndrom konnten neue, bisher nicht durch den Kandidatengen-Ansatz iden-tifizierte Polymorphismen identifizieren, de-ren pathophysiologische Bedeutung teilwei-se aber (noch) völlig unklar ist und die einer Replikation bedürfen.
SchlüsselwörterANCA-assoziierte Vaskulitiden · Großgefäßvaskulitiden · Kawasaki-Syndrom · Morbus Behçet · Kandidatengene · Genomweite Assoziationsstudie
The genetics of vasculitides
AbstractGenetic association studies have been of great value in the past by contributing to the understanding of pathophysiological mech-anisms of chronic inflammatory and auto-immune diseases. Many genetic risk fac-tors have been identified which confer sus-ceptibility for one or several (autoimmune) disease(s). Using a candidate-gene approach, the first genetic risk factors and polymor-phisms of vasculitides have been identified. Due to the rarity of autoimmune vasculiti-des often only small sample numbers have been generated and analysed, leading to in-consistent results. Furthermore, differences in ethnic background may complicate analy-sis. Only few of the detected risk factors have been reliably replicated in larger cohorts, such as the association of the PTPN22*620W allele with WG and MPA, the deficiency al-lele Pi*Z of the alpha1 antitrypsin gene and
the HLA-DPB*04041 allele with WG and the HLA-DRB3/DRB4 with CSS. Genome-wide as-sociation studies (GWAS) offer the advan-tage of screening the whole genome for risk factors rather than relying on disease mod-els postulated by the investigator; however, they require even larger sample sizes. Initial results from GWA studies are available for Be-hçet’s disease and Kawasaki syndrome, which identified new genetic associations but re-quire replication, especially since some of the identified risk factors could not be linked to pathophysiological pathways to date.
KeywordsANCA-associated vasculitides · Large vessel vasculitides · Kawasaki syndrome · Behçet’s disease · Candidate genes · Genomwide association study
201Zeitschrift für Rheumatologie 3 · 2011 |
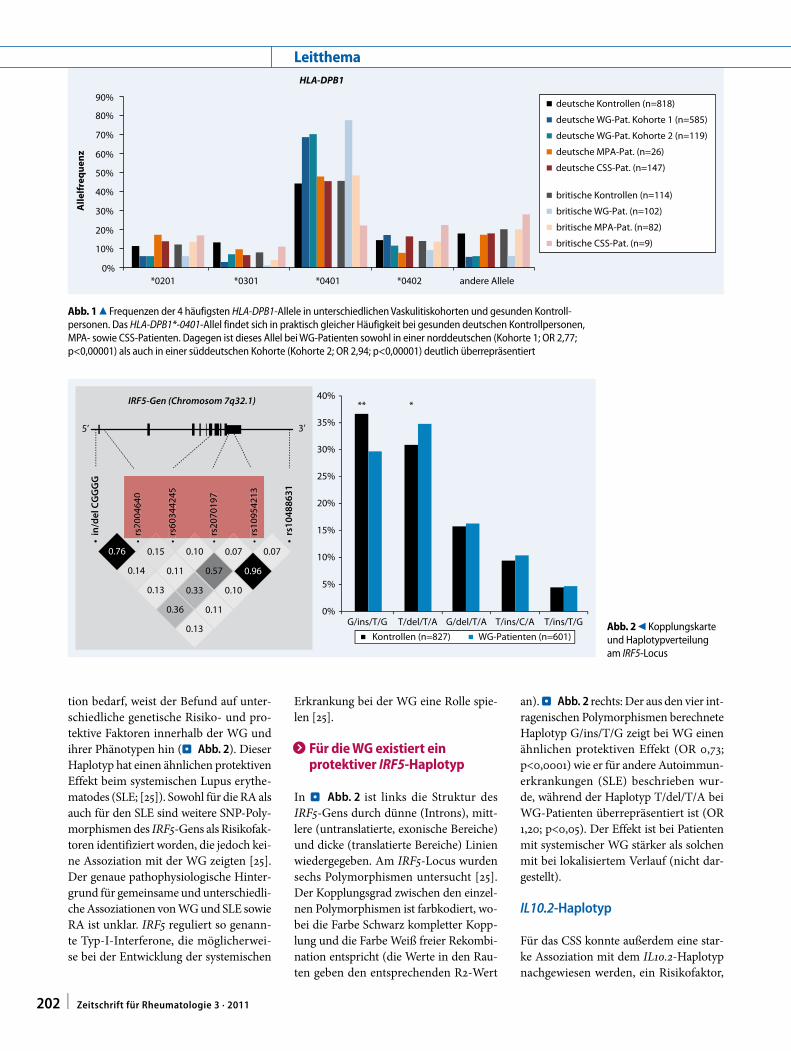
tion bedarf, weist der Befund auf unter-schiedliche genetische Risiko- und pro-tektive Faktoren innerhalb der WG und ihrer Phänotypen hin (. Abb. 2). Dieser Haplotyp hat einen ähnlichen protektiven Effekt beim systemischen Lupus erythe-matodes (SLE; [25]). Sowohl für die RA als auch für den SLE sind weitere SNP-Poly-morphismen des IRF5-Gens als Risikofak-toren identifiziert worden, die jedoch kei-ne Assoziation mit der WG zeigten [25]. Der genaue pathophysiologische Hinter-grund für gemeinsame und unterschiedli-che Assoziationen von WG und SLE sowie RA ist unklar. IRF5 reguliert so genann-te Typ-I-Interferone, die möglicherwei-se bei der Entwicklung der systemischen
Erkrankung bei der WG eine Rolle spie-len [25].
> Für die WG existiert ein protektiver IRF5-Haplotyp
In . Abb. 2 ist links die Struktur des IRF5-Gens durch dünne (Introns), mitt-lere (untranslatierte, exonische Bereiche) und dicke (translatierte Bereiche) Linien wiedergegeben. Am IRF5-Locus wurden sechs Polymorphismen untersucht [25]. Der Kopplungsgrad zwischen den einzel-nen Polymorphismen ist farbkodiert, wo-bei die Farbe Schwarz kompletter Kopp-lung und die Farbe Weiß freier Rekombi-nation entspricht (die Werte in den Rau-ten geben den entsprechenden R2-Wert
an). . Abb. 2 rechts: Der aus den vier int-ragenischen Polymorphismen berechnete Haplotyp G/ins/T/G zeigt bei WG einen ähnlichen protektiven Effekt (OR 0,73; p<0,0001) wie er für andere Autoimmun-erkrankungen (SLE) beschrieben wur-de, während der Haplotyp T/del/T/A bei WG-Patienten überrepräsentiert ist (OR 1,20; p<0,05). Der Effekt ist bei Patienten mit systemischer WG stärker als solchen mit bei lokalisiertem Verlauf (nicht dar-gestellt).
IL10.2-Haplotyp
Für das CSS konnte außerdem eine star-ke Assoziation mit dem IL10.2-Haplotyp nachgewiesen werden, ein Risikofaktor,
40%
35%
30%
25%
20%
15%
10%
5%
0%G/ins/T/G T/ins/C/A T/ins/T/GT/del/T/A G/del/T/A
IRF5-Gen (Chromosom 7q32.1)
5’
• in
/del
CG
GG
G
• rs
2004
640
• rs
6034
4245
• rs
2070
197
• rs
1095
4213
• rs
1048
8631
3’
** *
0.76
0.14
0.15 0.10
0.11
0.13
0.36
0.13
0.11
0.10
0.96
0.07
0.33
0.57
0.07
rs20
0464
0
rs60
3442
45
rs20
7019
7
rs10
9542
13
Kontrollen (n=827) WG-Patienten (n=601)Abb. 2 9 Kopplungskarte und Haplotypverteilung am IRF5-Locus
90%
80%
70%
60%
50%
Alle
lfreq
uenz
40%
30%
20%
10%
0%*0201 *0301 *0401 *0402 andere Allele
HLA-DPB1
britische CSS-Pat. (n=9)
britische MPA-Pat. (n=82)
britische WG-Pat. (n=102)
britische Kontrollen (n=114)
deutsche MPA-Pat. (n=26)
deutsche WG-Pat. Kohorte 2 (n=119)
deutsche WG-Pat. Kohorte 1 (n=585)
deutsche Kontrollen (n=818)
deutsche CSS-Pat. (n=147)
Abb. 1 8 Frequenzen der 4 häufigsten HLA-DPB1-Allele in unterschiedlichen Vaskulitiskohorten und gesunden Kontroll-personen. Das HLA-DPB1*-0401-Allel findet sich in praktisch gleicher Häufigkeit bei gesunden deutschen Kontrollpersonen, MPA- sowie CSS-Patienten. Dagegen ist dieses Allel bei WG-Patienten sowohl in einer norddeutschen (Kohorte 1; OR 2,77; p<0,00001) als auch in einer süddeutschen Kohorte (Kohorte 2; OR 2,94; p<0,00001) deutlich überrepräsentiert
202 | Zeitschrift für Rheumatologie 3 · 2011
Leitthema

der allein das CSS auszuweisen scheint [22, 23] und generell mit erhöhten IL-10-Serumspiegeln assoziiert ist, wie sie auch beim CSS nachweisbar sind. Ein direk-ter Zusammenhang muss beim CSS aber noch gezeigt werden. Anzumerken ist, dass dieser Haplotyp insbesondere mit dem ANCA-negativen CSS assoziiert ist, sodass auch hier möglicherweise Unter-schiede im Genotyp für die Ausprägung unterschiedlicher Phänotypen innerhalb des CSS verantwortlich zu sein scheinen.
Bisher identifizierte Risikogene bei weiteren Vaskulitiden
Für das Kawasaki-Syndrom (mukoku-tanes Lymphknotensyndrom) ist auf-grund der stark unterschiedlichen Prä-valenz mit deutlicher Häufung in Japan und Korea ein starker genetischer Hin-tergrund anzunehmen. Mehrere Kandi-datengen-Analysen haben jedoch in den letzten zwei Jahren divergente Ergebnisse in verschiedenen Kohorten ergeben. Le-diglich für mehrere assoziierte SNPs im Gen der Inositol-1,4,5-triphopshatase-3-Kinase C (ITPKC), einem Enzym, das in die Signaltransduktion u. a. auch des T-Zell-Rezeptors (TCR) involviert ist, konn-te eine replizierbare Assoziation gefunden werden [26], die möglicherweise eine Stö-rung des „TCR-Signaling“ induziert. Die erste GWAS konnte fünf potenzielle Lo-ci als Risikofaktoren identifizieren, die of-fensichtlich einem gemeinsamen Signal-weg angehören. Die Studie wurde jedoch an einer kleinen Patientenzahl durchge-führt (n=109) und bisher nicht repliziert, sodass die Ergebnisse als vorläufig zu wer-ten sind [4].
> Mutationen im MEFV-Gen könnten zu einer überschießenden IL-1-Produktion führen
Neben der sicherlich überzeugenden As-soziation für den M. Behçet mit HLA-B51 [14, 27] konnten Assoziationen mit einem Polymorphismus der endothelialen NO-Synthetase (ENOS), allerdings mit stark variierenden Ergebnissen (OR 0,82–4,3; [2, 5]), und von Mutationen im MEFV-Gen (OR 2,96–4,55; [2, 11]) identifiziert und auch repliziert werden. Die Kandi-
datengen-Analyse des MEFV-Gens beim M. Behçet beruhte auf der Beobachtung, dass Patienten ähnliche klinische Befun-de aufweisen können wie Patienten mit dem familiären Mittelmeerfieber (FMF), für das Mutationen im MEFV-Gen cha-rakteristisch sind. Das Genprodukt des MEFV-Gens, Pyrin, hat eine supprimie-rende Funktion bei der Synthese von IL-1β. Mutationen könnten zu einem Wegfall der supprimierenden Funktion und da-mit zu einer überschießenden IL-1-Pro-duktion führen. Die Rolle von IL-1β beim M. Behçet ist allerdings unklar.
Eine in 2009 durchgeführte GWAS konnte fünf potenzielle Risikogene iden-tifizieren, deren Funktion unklar ist [8]. Kein der bisher durch Kandidatengen-Analyse identifizierter Polymorphis-mus wurde durch die GWAS bestätigt, was in klarer Weise die Limitationen ei-ner GWAS aufzeigt: Die möglicherweise zu kleine Probenzahl (152 Patienten, 172 Kontrollen) ermöglicht keine Identifikati-on geringer oder mäßiger genetischer Ri-sikofaktoren; die Replikation der Ergeb-nisse steht aus.
Fazit für die Praxis
F Die HLA-Region zeigt nach wie vor die überzeugendsten Assoziationen für verschiedene Vaskulitiden.
F Nur wenige identifizierte Kandidaten-gene konnten bisher verlässlich rep-liziert werden; erste Ergebnisse zei-gen, dass bei Vaskulitiden (vor allem bei den AAV) mit klinischen und/oder serologischen Gemeinsamkeiten ge-meinsame genetische Risikofaktoren, welche eine verwandte Gruppe von Erkrankungen definieren, existieren sowie weitere genetische Assoziatio-nen zu finden sind, die spezifisch für nur eine Erkrankung sind.
F Der Ansatz der GWAS ist viel verspre-chend im Hinblick auf die Gewinnung umfassender Daten, die ein geneti-sches Risiko für eine Erkrankung ent-halten können.
F Erste durchgeführte GWAS zeigen je-doch eher ernüchternde Ergebnisse und untermauern die Notwendigkeit großer Fallzahlen, um verlässliche Er-gebnisse zu generieren.
KorrespondenzadresseDr. J.U. Holle
Poliklinik für Rheumatologie, Vaskulitiszentrum, Universitätsklinikum Schleswig-Holstein, Campus Lübeck, Klinik für Rheumatologie und Klinische Immunologie, Klinikum Bad Bramstedt Oskar-Alexander-Str. 26, 24576 Bad [email protected]
Interessenkonflikt. Die korrespondierende Autorin gibt an, dass kein Interessenkonflikt besteht.
Literatur
1. Arning L, Holle JU, Harper L et al (2010) Are there specific genetic risk factors for the different forms of ANCA-associated vasculitis? Ann Rheum Dis [Epub ahead of print Aug 10]
2. Ayesh S, Abu-Rmaileh H, Nassar S et al (2008) Mo-lecular analysis of MEFV gene mutations among Palestinian patients with Behcet’s disease. Scand J Rheumatol 37:370–374
3. Ben Dhifallah I, Houman H, Khanfir M et al (2008) Endothelial nitric oxide synthase gene polymor-phism is associated with Behcet’s disease in Tunsi-sian population. Hum Immunol 69:661–665
4. Burgner D, Davila S, Breunis WB et al (2009) A ge-nome-wide association study identifies novel and functionally related susceptibility Loci for Kawasa-ki disease. PLoS Genet 5:e1000319
5. Carr EJ, Niederer HA, Williams J et al (2009) Con-firmation of the genetic association of CTLA4 and PTPN22 with ANCA-associated vasculitis. BMC Med Genet 10:121
6. Dursun A, Durakbasi-Dursun HG, Dursun R et al (2009) Angiotensin-converting enzyme gene and endothelial nitric oxide synthase gene polymor-phism in Behcet’s disease with or without ocu-lar involvement. Inflamm Res [Epub ehead of print Mar 3]
7. Esnault VL, Testa A, Audrian M et al (1993) Alpha 1-antitrypsin genetic polymorphism in ANCA-posi-tive systemic vasculitis. Kidney Int 43:1329–1332
8. Fei Y, Webb R, Cobb BL et al (2009) Identification of novel genetic susceptibility loci for Behcet’s di-sease using a genome-wide associaction study. Arthritis Res Ther 11:R66
9. Gonzalez-Gay MA, Amoli MM, Garcia-Porrua C et al (2003) Genetic markers of disease susceptibility and severity in giant cell arteritis and polymyalgia rheumatic. Semin Arthritis Rheum 33:38–48
10. Heckmann M, Holle JU, Arning L et al (2008) The Wegener’s granulomatosis quantative trait lo-cus on chromosome 6p21.3 as characterized by tagSNP genotyping. Ann Rheum Dis 67:972–979
11. Imirzalioglu N, Dursun A, Tastan B et al (2005) MEFV gene is a probable susceptibility gene for Behcet’s disease. Scand J Rheumatol 34:56–58
12. Jagiello P, Gencik M, Arning L et al (2004) New ge-nomic region for Wegener’s granulomatosis as re-vealed by an extended association screen with 202 apoptosis-related genes. Hum Genet 114:468–477
13. Jagiello P, Aries P, Arning L et al (2005) The PTPN22 620 W allele is a risk factor for Wegener’s granulo-matosis. Arthritis Rheum 12:4039–4043
14. Krause I, Weinberger A (2008) Behcet’s disease. Curr Opin Rheumatol 20:82–87
203Zeitschrift für Rheumatologie 3 · 2011 |

15. Mahr AD, Edberg J, Stone J et al (2009) Alpha 1-an-titrypsin deficiency and the risk for Wegener’s gra-nulomatosis. In: Proceedings of the 14th Inter-national Vasculitis and ANCA Workshop. APMIS 117(Suppl 127):157 [abstract N1]
16. Monach P, Merkel PA (2010) Genetics of vasculitis. Curr Opin Rheumatol 22:157–163
17. Rarok AA, Stegeman CA, Limburg PC et al (2002) Neutrophil membrane expression of proteinase 3 (PR3) is related to relapse in PR3-ANCA associated vasculitis. J Am Soc Nephrol 13:1224–1233
18. Stassen PM, Cohen-Tervaert JW, Leemsp SP et al (2009) HLA-DR4, DR13(6) and the ancestral haplo-type A1 B8DR3 are associated with ANCA-asso-ciated vasculitis and Wegener’s granulomatosis. Rheumatology 48:622–625
19. Szodoray P, Szabo Z, Kapitány A et al (2010) Anti-citrullinated protein/peptide autoantibodies in as-sociation with genetic and environmental factors as indicators of disease outcome in rheumatoid ar-thritis. Autoimm Rev 9:140–143
20. Vaglio A, Martorana D, Maggiore U et al (2007) HLA-DRB4 as a genetic risk factor for Churg-Strauss syndrome. Arthritis Rheum 56:3159–3166
21. Vargas-Alacron G, Hernandez-Pacheco G, Soto ME et al (2005) Comparative study of the residues 63 and 67 on the HLA-B molecule in patients with Takayasu’s arteritis. Immunol Lett 96:225–229
22. Wieczorek S, Hellmich B, Arning L et al (2008) Functionally relevant variations of the interleu-kin-10 gene associated with antineutrophil cyto-plasmic antibody-negative Churg-Strauss syndro-me. Arthritis Rheum 58:1839–1848
23. Wieczorek S, Hellmich B, Gross WL et al (2008) As-sociations of Churg-Strauss syndrome with the HLA-DRB1 locus, and relationship to the genetics of antineutrophil cytoplasmic antibody-associated vasculitides: comment on the article by Vaglio et al. Arthritis Rheum 58:329–330
24. Wieczorek S, Holle JU, Epplen JT (2010) Recent pro-gress in the genetics of Wegener’s granulomatosis and Churg-Strauss syndrome. Curr Opin Rheuma-tol 22:8–14
25. Wieczorek S, Holle JU, Müller S et al (2010) A func-tionally relevant IRF5 haplotype is associated with reduced risk to Wegener’s granulomatosis. J Mol Med 88:413–421
26. Onouchi Y, Gunji T, Burns JC et al (2008) ITPKC functional polymorphism associated with Kawasa-ki disease susceptibility and formation of coronary artery aneurysms. Nat Genet 40:35–42
27. De Menthon M, Lavalley MP, Maldini C et al (2009) HLA-B51/B5 and the risk of Behcet’s disease: a sys-tematic review and meta-analysis of case-con-trol genetic association studies. Arthritis Rheum 61:1287–1296
Maßnahmen zur Infektionsprävention haben einen zunehmend hohen Stellen-wert. Infektiöse Komplikationen durch medizinische Maßnahmen sollen soweit irgend möglich vermieden werden. Neben dem qualifizierten Hygienefach-personal in den Kliniken ist es wichtig, dass auch in den Abteilungen erfahrene und in Hygiene fortgebildete Ärztinnen und anerkannte (Fach-)Ärzte Ansprech-partner für ihre Kollegen und Mitarbei-ter sind. Aufgrund ihrer besonderen Sachkenntnis in Hygienefragen können sie das Bewusstsein für die Relevanz der Einhaltung der geltenden Hygiene-standards immer wieder schärfen und die Umsetzung fördern.
Neues Kursformat
Mit dem von der MEDILYS Laborgesell-schaft mbH und den Asklepios Kliniken ent wickelten Kurs nach den RKI-Richt-linien kann nun die Qualifikation eines Hygienebeauftragten Arzt/Ärztin erworben werden. Der Kurs ist zweiteilig als Blended Learning Kurs aufgebaut.
Der erste Schritt zum Zertifikat ist die Teilnahme an einer interaktiven Elearning einheit, mit dem man sich im
Selbststudium die Grundlagen erarbeitet. Das Modul steht ein halbes Jahr zum Lernen und Nach schlagen online zur Verfügung. Die erfolgreiche Absolvierung der Lern-kontrollfragen zeigt für jedes Kapitel, dass die Inhalte beherrscht werden.
Am 14. und 15. April sowie am 10. und 11. November 2011 finden dann in der Asklepios Klinik Barmbek in Ham-burg der zweite Teil, die abschließende Präsenzveranstaltung statt, bei der die Mitglieder des wissenschaftlichen Redaktionsteams und weitere Experten anwesend sind, um im kollegialen Dialog die wichtigsten Aspekte der Infektions-prävention zu vertiefen. Der Kurs ist durch
die Ärztekammer Hamburg und durch die Staatliche Zentralstelle für Fernunterricht (ZFU) mit 40 CME-Punkten zertifiziert.
Weitere InformationenAsklepios Kliniken Hamburg GmbH – Ärzteakademie, [email protected]
Fachnachrichten
KursankündigungHygienebeauftragter Arzt/Ärztin
Erster Blended Learning Kurs Hygiene beauftragte Arzt/Ärztin nach den neuen RKI-Richtlinien ab 1. Januar 2011
© p
icpr
ofi / f
otol
ia.c
om
204 | Zeitschrift für Rheumatologie 3 · 2011





























