Embed Size (px)
Citation preview

1. Gesichtsfeld – Theoretischer Teil Aufbau des Vertebratenauges Der Aufbau des Vertebratenauges ist in Abbildung 1 dargestellt.
Abbildung 1: Aufbau des Vertebratenauges. Die Pupille ist hier nicht beschriftet, stellt aber den Bereich zwischen der Iris dar.
- Sclera (Lederhaut): Die Sclera ist ein hartes, weißes Bindegewebe, das das Auge schützt. - Conjunctiva: Die Conjunctiva ist eine Schleimschicht, die aus zarten Epithelzellen
besteht und die Sclera umgibt, um Feuchtigkeit zu spenden. - Cornea (Hornhaut): Die Cornea ist der Teil der Sclera, der nicht von der Conjunctiva
bedeckt wird. Durch sie fällt das Licht in das Auge. - Chorioidea (Aderhaut): Die Chorioidea liegt zwischen der Sclera und der Retina und
versorgt die Retina mit Nährstoffen. - Retina (Netzhaut): Die Retina (ihr Aufbau wird in einem eigenen Kapitel näher erörtert)
besteht aus mehreren Zellschichten (Sinnes- und Nervenzellen) und dient der Aufnahme und Verarbeitung von Lichtsinnesreizen.
- Ciliarkörper: Die Chorioidea mündet in den Ciliarkörper, der aus Ciliarmuskeln und Zonulafasern besteht. Der Ciliarkörper selbst bildet das Kammerwasser, das sich in der vorderen und hinteren Augenkammer sammelt (bei Verstopfung des Ciliarkörpers kann das Kammerwasser nicht mehr abfließen, wodurch sich der Augeninnendruck erhöht. Die Folge davon ist ein Glaucom mit stark eingeschränktem Sichtfeld). Die Ciliarmuskeln sind über die Zonulafasern mit der Linse verbunden und regeln durch Kontraktion/Entspannung die Form der Linse (siehe dazu das Kapitel über die Akkommodation).

- Linse: Die Linse ist für die Lichtbrechung verantwortlich und ihre Form ist variabel. Sie unterteilt das Auge auch in drei Räume, nämlich die vordere Augenkammer, die hintere Augenkammer und den Glaskörper.
- Iris (Regenbogenhaut): Die Iris ist auch ein Teil der Chorioidea und regelt die Größe der Pupille (siehe dazu das Kapitel über Adaptation). Die Pupille ist der Teil der Cornea, der nicht von der Iris verdeckt wird. Die Iris bestimmt auch die Augenfarbe durch individuelle Pigmentierung.
- Glaskörper: Der Glaskörper ist gallertartig und durchsichtig. Er fungiert als Flüssiglinse (wie das Kammerwasser) und dient so der Lichtbrechung und Fokussierung auf die Retina.
- Papille („Blinder Fleck“, Discus nervus opticus): Die Papille stellt eine Eigenart des Vertebratenauges im Gegensatz zum Cephalopodenauge dar (siehe dazu das Kapitel über die Evolution des Vertebratenauges), da man an diesem Teil des Auges nichts sieht. Das liegt am Fehlen von Photorezeptorzellen, denn hier verdichten sich die Axone der Ganglienzellen aus der Retina zum Nervus opticus, der die Signale vom Auge zum restlichen Teil des Gehirns leitet. Außerdem befinden sich hier die Vena und die Arteria centralis, die die Retina versorgen.
- Fovea centralis: Die Fovea centralis stellt das Zentrum des Gesichtsfeldes dar und ist der Punkt des schärfsten Sehens, da sich hier mehr als 150000 Zapfen pro mm² befinden (die Zapfen und Stäbchen werden später näher erläutert) und dies die größte Dichte an Zapfen auf der gesamten Retina ist (Stäbchen sind auf der Fovea centralis nicht lokalisiert).
Die Evolution des Vertebratenauges Als Besonderheit des Vertebratenauges wurde bereits die Papille genannt, die eine Eigenheit der Vertebraten darstellt. Das prinzipiell gleich aufgebaute und sehr ähnlich erscheinende Cephalopodenauge besitzt diese Besonderheit nicht. Das lässt sich aus der unterschiedlichen Entwicklung der beiden Augentypen erklären. Das Vertebratenauge entstand, als sich das Neuralrohr einfaltete und sich eine Augenblase bildete. Diese Blase verband sich mit dem Diencephalon und stülpte sich ein, so das die Nervenzellen vor (in Bezug zur Richtung aus der das Licht eintrifft) den Sinneszellen liegen. Aufgrund dieser Konstruktion wird das Vertebratenauge als invers bezeichnet und die Papille ist die Folge dieser Entwicklung. Die Cephalopoden besitzen ein everses Auge, das heißt die Sinneszellen liegen vor den Nervenzellen und es gibt keinen blinden Fleck. Ihr Auge entstand einfach durch die Einstülpung lichtempfindlicher Epidermiszellen. Der dioptrische Apparat Mit der Bezeichnung „Dioptrischer Apparat“ werden die Cornea, das Kammerwasser, die Linse und der Glaskörper umschrieben, die alle den Zweck der Lichtbrechung und der Bündelung des Lichts auf der Retina erfüllen. Das Licht wird beim Einfallen gebrochen und spiegelverkehrt auf die Retina geworfen (das Gehirn dreht das Bild dann richtig herum) und wohin es dabei fällt bestimmt der Auftrittswinkel auf den dioptrischen Apparat. Die Komplexität des dioptrischen Apparats lässt sich durch zwei Formeln erklären: Die Lichtempfindlichkeit S hängt vom Linsendurchmesser D, dem Rezeptordurchmesser d und der Brennweite f wie folgt ab: S ~ D² * d² / f² Das Auflösungsvermögen R steht zum optischen Abstandswinkel a im Bezug R ~ 1/a Es wird daraus ersichtlich, dass ein größerer Linsenapparat wichtig ist, um die Sehschärfe effektiv zu erhöhen, denn mehr Licht wird stärker gebündelt. Gemessen wird die Brechkraft D

des dioptrischen Apparats in der Einheit Dioptrien (dpt) und ausgerechnet wird sie über die Formel D = 1/f 1.4. Aufbau und Funktion der Retina Die Retina dient der Aufnahme von Lichtquanten und der Umwandlung derselben in elektrische Reize, die dem Gehirn übermittelt werden. Dazu besteht sie aus mehreren Zellschichten, die in Abbildung 2 gezeigt sind.
Abbildung 2: Rod bezeichnet Stäbchenzellen, Cone bezeichnet Zapfenzellen.
- Pigmentepithel: Das Pigmentepithel versorgt die Zäpfchen und Stäbchen mit Nährstoffen und verhindert Reflexionen des einfallenden Lichts.
- Zapfen und Stäbchen (Cone und Rod): Zapfen und Stäbchen sind verantwortlich für die Reizaufnahme und wandeln Lichtquanten in elektrische Signale um (der Prozess wird in dem Abschnitt über Phototransduktion genauer erklärt).
- Horizontalzellen: Die Horizontalzellen verknüpfen mehrere Stäbchen und Zapfen miteinander und stehen auch mit Bipolarzellen in Kontakt.
- Bipolarzellen: Bipolarzellen verbinden die Stäbchen und Zapfen mit den Ganglienzellen und sind entscheidend für die Auslösung eines Rezeptorpotentials.
- Amakrinzellen: Amakrinzellen verknüpfen Ganglienzellen miteinander, stehen aber auch mit Bipolarzellen querverbunden.
- Ganglienzellen: Ganglienzellen senden Aktionspotentiale über ihre, zum Nervus opticus gebündelten, Axone zum Gehirn.
Die Bedeutung von Horizontalzellen und Amakrinzellen liegt in der lateralen Hemmung (laterale Inhibition). Wird ein Stäbchen oder Zäpfchen erregt, so erregen Horizontalzellen benachbarte Rezeptorzellen, hemmen aber entfernter liegende Rezeptorzellen (dies bewirkt eine Verstärkung des Kontrasts zwischen einem hellen Punkt und seiner dunklen Umgebung) und auch benachbarte Neuronen. Die Amakrinzellen wirken genauso, nur auf Ganglien- statt auf Rezeptorzellen. 1.4.1. Die Phototransduction Die Phototransduction (also die Umwandlung von Lichtquanten in elektrochemische Reize) findet in den Zapfen und Stäbchen statt. Der Prozess soll hier am Beispiel einer Stäbchenzelle erläutert werden.

In den Stäbchen sind viele Disks angeordnet, in deren Membran ein Molekülkomplex namens Rhodopsin (ein Sehpigment) integral eingelagert ist. Rhodopsin besteht aus Retinal und Opsin (es ist dabei zu beachten, dass nur ein Komplex aus Retinal und Opsin Rhodopsin heißt und von anderen Sehpigmenten unterschieden werden muss, so findet sich bei, im Süßwasser lebenden, Fischen und Amphibien Porphyropsin, das statt Retinal 3-Dehydro-Retinal besitzt). Das Retinal ist dabei von entscheidender Bedeutung für die Phototransduction, denn durch Licht ändert es seine Konformation: Von 11-cis-Retinal zu All-trans-Retinal. Abbildungen 3 und 4 zeigen diesen Effekt.
Abbildung 3: 11-cis-Retinal ändert bei Lichteinfall seine Konformation. Isomerasen überführen es dann wieder in seine ursprüngliche Konformation zurück. In diesem Zustand ist es an Opsin gebunden und purpurfarben.
Abbildung 4: All-trans-Retinal ist vom Opsin abgelöst und farblos (Man redet daher von Bleichung). Die Gesamtreaktion:
1. Durch Lichteinfall wird 11-cisRetinal zu All-trans-Retinal und löst sich vom Opsin ab (bleibt anfangs allerdings noch kovalent gebunden), das dadurch aktiviert wird.
2. Das aktive Opsin induziert ein G-Protein namens Transducin (ungefähr 500 Transducin-Moleküle werden von einem Opsin aktiviert). Dieses startet eine Enzymkaskade.
3. Das Transducin aktiviert ein Enzym namens Phospho-di-esterase (PDE). 4. Das Enzym Phospho-di-esterase hydrolisiert cGMP-Moleküle (jedes PDE hydrolisiert
ungefähr 1000 cGMP-Moleküle!) zu GMP, das sich daraufhin von Natriumkanälen ablöst. 5. Die Natriumkanäle sind ligandenabhängig und cGMP ist der Ligand. Ohne ihren Liganden
bleiben die Natrium-Kanäle geschlossen und eine Hyperpolarisation ist die Folge. Das bedeutet, dass keine Depolarisation an die Synapse weitergeleitet und somit die Ausschüttung des Transmitters Glutamat reduziert wird. Glutamat ist ein Inhibitor. Ein Lichtreiz führt also zur Inhibierung einer Inhibition.

Abbildung 5 zeigt das Ganze grafisch dargestellt.
Abbildung 5: Die Phototransduction. Rhodopsin löst dabei durch die Aktivierung von Transducin eine Enzymkaskade aus. 1.4.2. Rezeptive Felder Ein Rezeptives Feld ist definiert als die Anzahl an Rezeptorzellen, die mit einer Ganglienzelle verbunden sind. Rezeptive Felder sind generell rund und überlappen sich teilweise. Die Sehschärfe hängt dabei von der Größe des Rezeptiven Feldes ab (Sie ist indirekt proportional dazu), da ein Reiz irgendwo auf dem Rezeptiven Feld zwar eine Reaktion auslöst, aber nicht genau lokalisiert werden kann, je mehr Photorezeptorzellen eine Ganglienzelle bedienen. Die Rezeptiven Felder lassen sich in zwei Klassen einteilen, die ON- und die OFF-Felder. Die ON-Felder führen zu einer Depolarisation im Zentrum des Feldes und einer Hyperpolarisation in der Peripherie, also zu einer Erregung durch Reiztypen, die heller als der Hintergrund sind. Die OFF-Felder führen zu einer Hyperpolarisation im Zentrum und einer Depolarisation in der Peripherie, also zu einer Erregung durch Reiztypen, die dunkler sind als der Hintergrund. Ausschlaggebend dafür, ob ein Rezeptives Feld nun ON oder OFF ist, sind die Bipolarzellen und ihre verschiedenen Rezeptortypen. Der Vorteil von Rezeptiven Feldern besteht in einem geringeren Energieverbrauch, weil die Ruheaktivität der Retina gering sein kann und in einer Symmetrie bezüglich Hell- und Dunkelreizen. Der Sinn von Rezeptiven Feldern besteht in der Erhöhung des Kontrasts zwischen Hell und Dunkel und der Steigerung der Empfindlichkeit. 1.4.3. Räumliches Auflösungsvermögen Die Höhe der Auflösung besagt, ab wann man zwei nebeneinander liegende Punkte noch als zwei einzelne Punkte unterscheiden kann. Beim Menschen beträgt das Räumliche Auflösungsvermögen 0.1mm.

1.4.4. Skotopisches und Photopisches System und das Purkinje-Phänomen Wie bereits angedeutet wurde gibt es zwei unterschiedliche Typen von Photorezeptoren, die in Abbildung 6 gezeigt sind.
Abbildung 6: Die Stäbchen sind für Hell-/Dunkelsehen, die Zapfen für das Farbensehen zuständig.
- Stäbchen: Die Stäbchen sind für das Skotopische Sehen (Hell/Dunkel) verantwortlich und ihr Empfindlichkeitsmaximum liegt bei 510nm. Wie bereits erläutert wurde, basiert das Skotopische Sehen auf dem Sehpigment Rhodopsin, das bei Licht ausbleicht.
- Zapfen: Die Zapfen sind für das Photopische Sehen (Blau/Grün/Rot) verantwortlich und besitzen ihr Empfindlichkeitsmaximum bei 550nm. Es gibt drei Typen von Zapfen, von denen jedes eine andere Isoform des Photopsin genannten Zapfenopsin besitzt. Jeder der drei Typen ist dadurch gekennzeichnet, dass er Licht einer bestimmten Wellenlänge am stärksten absorbiert. Es gibt dabei Grün-, Blau- und Rot-Zapfen. Die Absorptionsspektren sind in Abbildung 7 dargestellt. Mischfarben entstehen dadurch, dass unterschiedliche Zapfentypen unterschiedlich stark erregt werden (die Farbwahrnehmung hängt dann davon ab, welcher stärker erregt wurde, beispielsweise ob man Gelb oder Orange sieht, wenn Grün und Rot erregt wurden).
- Purkinje-Phänomen: Beim Übergang von Hell- zu Dunkelsehen sieht man die Farben mit unterschiedlicher Intensität. Das hängt damit zusammen, dass die Zapfen hauptsächlich aktiv sind, denn das Rhodopsin der Stäbchen wird durch Licht gebleicht (bei Vertebraten ist die Rückführung des All-trans-Retinals zum 11-cis-Retinal sehr umständlich, teilweise findet gar eine Neusynthese des Retinals aus Vitamin A statt). Wie in Abbildung 7 zu sehen ist, haben die Zapfen ihr Absorptionsmaximum im blau-grünen Bereich, ihr Minimum im roten Bereich. Deshalb sieht man beim Übergang von Hell zu Dunkel blau-grüne Farben intensiver als Rote. Bei Skotopischem Sehen bei Tageslicht liegt das Absorptionsmaximum im Gegensatz dazu im gelben Bereich.
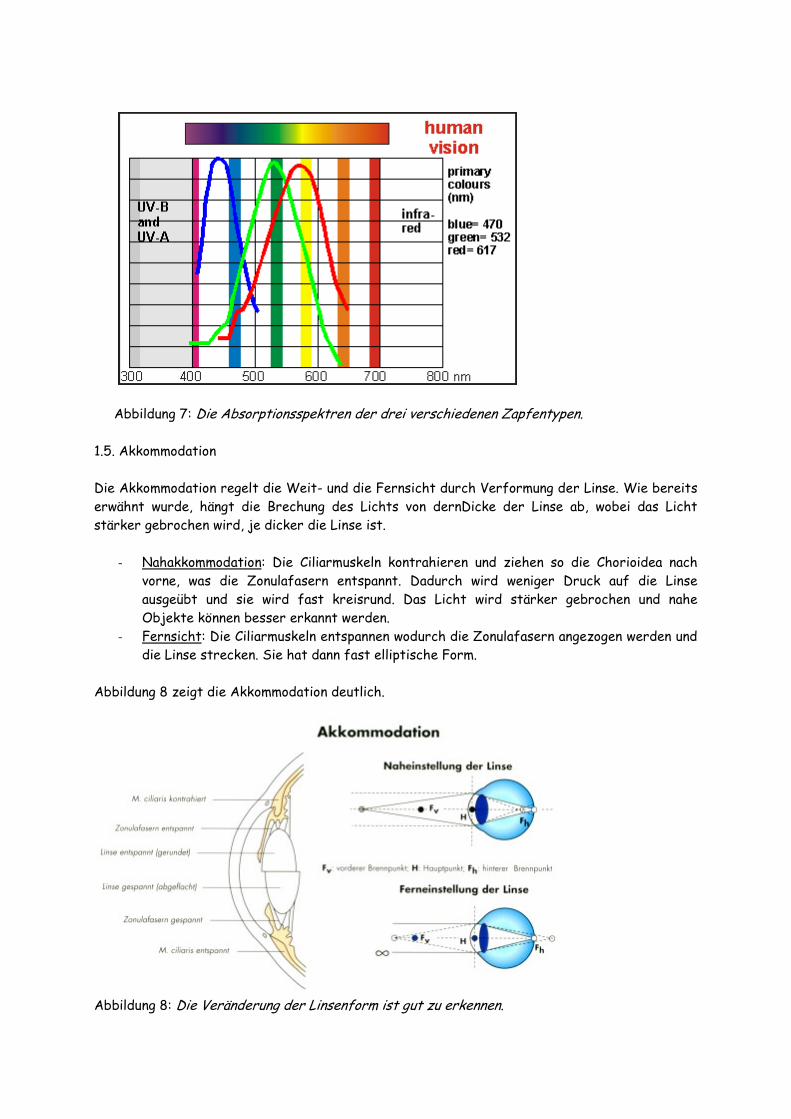
Abbildung 7: Die Absorptionsspektren der drei verschiedenen Zapfentypen. 1.5. Akkommodation Die Akkommodation regelt die Weit- und die Fernsicht durch Verformung der Linse. Wie bereits erwähnt wurde, hängt die Brechung des Lichts von dernDicke der Linse ab, wobei das Licht stärker gebrochen wird, je dicker die Linse ist.
- Nahakkommodation: Die Ciliarmuskeln kontrahieren und ziehen so die Chorioidea nach vorne, was die Zonulafasern entspannt. Dadurch wird weniger Druck auf die Linse ausgeübt und sie wird fast kreisrund. Das Licht wird stärker gebrochen und nahe Objekte können besser erkannt werden.
- Fernsicht: Die Ciliarmuskeln entspannen wodurch die Zonulafasern angezogen werden und die Linse strecken. Sie hat dann fast elliptische Form.
Abbildung 8 zeigt die Akkommodation deutlich.
Abbildung 8: Die Veränderung der Linsenform ist gut zu erkennen.

1.6. Sensorische Adaptation Unter Sensorischer Adaptation versteht man die Anpassung eines sensorischen Systems an den physikalischen Bereich seiner Input-Größen. Das bedeutet, dass sensorische Systeme bestimmte, konstante Reize herausfiltern, um andere Reize besser wahrnehmen zu können. So gewöhnt man sich beispielsweise nach einer bestimmten Zeit an einen Geruch, der permanent verströmt wird. Im visuellen Bereich gibt es für sensorische Adaptation viele Beispiele. So ändert sich der Durchmesser der Pupille je nachdem, wie hell die Umgebung ist (in erregtem Zustand kann die Pupille um das Achtfache vergrößert sein), durch die Kontraktion/Entspannung der Irismuskeln. Dabei wird die Pupille größer, je dunkler die Umgebung ist, um mehr des spärlichen Lichts aufnehmen zu können. Ein weiterer Adaptationsmechanismus besteht in der, bereits angesprochenen, Ausbleichung des Rhodopsins. Bei Menschen dauert die Reisomerisierung des Retinals sehr lange und das Retinal wird teilweise aus Vitamin A neu synthetisiert. Bei großer Helligkeit bleicht das Rhodopsin aus und ist dann inaktiv, so dass die Umgebung hauptsächlich durch die Zapfen wahrgenommen werden kann und die Augen an die Helligkeit angepasst sind. Ein interessantes Beispiel für sensorische Adaptation liefern bestimmte Fische, die zu Retinomotorik befähigt sind. Das bedeutet, dass sie die Rezeptorzellen in das Pigmentepithel verlagern können, um sie vor Licht zu schützen. Als räumliche und zeitliche Summation werden Phänomene bezeichnet, bei denen sich synaptische Potentiale in räumlicher Nähe zueinander an einem Neuron addieren oder präsynaptische Reize schnell hintereinander an der Synapse ankommen und sich addieren und so den Schwellenwert für die Auslösung eines Aktionspotentials überschreiten. Das Umschalten von photopischem zu skotopischem Sehen beim Übergang von Dunkelheit zur Helligkeit (und umgekehrt) gehört auch zur sensorischen Adaptation. Siehe dazu Abschnitt 1.4.4. Ein weiterer Adaptationsmechanismus besteht in der Verringerung der Ca2+ Konzentration im Zellinneren. Bei Belichtung schließen sich die Kationen-Kanäle, so dass Ca2+ nicht mehr in die Zellen einströmen kann, aber weiterhin durch membranständige Ionen-Austauscher in den extrazellulären Raum transportiert. Die niedrige Ca2+ Konzentration im intrazellulären Raum führt zu einer verstärkten Aktivität des Enzyms Guanylatzyklase, das GTP zu cGMP synthetisiert, welches wiederum zur Öffnung der Kationen-Kanäle beiträgt und damit dem Effekt der Beleuchtung entgegenwirkt. 1.7. Farbtheorien Es gibt zwei (scheinbar) sich ausschließende Theorien zu diesem Thema und eine dritte Theorie, die die beiden vorherigen vereint. Die trichromatische Theorie von Young (1804) und Helmholtz (1879) besagt, dass es drei verschiedene Zapfentypen gibt, die jeweils unabhängige Empfängersysteme darstellen. Die Signale dieser drei Empfängersysteme werden aber gemeinsam von einem neuronalen Helligkeitssystem und einem neuronalen Farbensystem verarbeitet. Es sei dazu angemerkt, dass im Tierreich die Spanne von dinochromatischem bis zu tetrachromatischem Sehen (Tauben und Goldfische haben vier Klassen von Zapfen) reicht. Um eine Verrechnung der Absorptionsspektren zu bekommen, werden mindestens zwei verschiedene Rezeptortypen benötigt. Die Gegenfarbentheorie von Hering (1905) besagt, dass es zwei verschiedene antagonistische physiologische Farbsysteme gibt, nämlich Rot-Grün und Gelb-Blau (so würde eine gelbe Erregung blau hemmen). Ein weiteres antagonistisches System besteht in dem Schwarz-Weiß-System. Die Zonentheorie wurde von Johannes Kries Anfang des 19.Jahrhunderts entwickelt und vereinigt die trichromatische Theorie mit der Gegenfarbentheorie. Dabei wird die trichromatische Theorie auf die drei Zapfentypen angewendet und die Gegenfarbentheorie für die nachgeschalteten neuronalen Verarbeitungssysteme angenommen.

1.8. Der Farbkreis und die Farbmischungen Der Mensch kann zwanzig verschiedene Farbtöne sehen und insgesamt 7 Millionen verschiedene Farbmischungen wahrnehmen. Abbildung 9 zeigt den Farbkreis, der 16 verschiedene Farbtöne enthält. Ein Farbkreis kann modifiziert werden, indem man Purpur zwischen Rot und Blau einfügt oder herausnimmt, aber auch durch Einbezug von Schwarz und Weiß, die keine richtigen Farben sind, da bei Weiß Licht des gesamten sichtbaren Spektrums reflektiert wird, während bei Schwarz das Licht des gesamten sichtbaren Spektrums absorbiert wird.
Abbildung 9: Der Farbkreis mit 16 verschiedenen Farbtönen, auch Purpur Es gibt zwei verschiedene Formen der Farbmischung: 1.8.1. Additive Farbmischung: Wenn Lichtphotonen unterschiedlicher Wellenlänge auf die gleiche Stelle der Retina treffen, ergeben sich zwangsläufig Farbmischungen. Es werden dann Farbtöne angezeigt, die anderen Spektralbereichen entsprechen oder gar Purpur, das keine eigene Wellenlänge hat. Abbildung 10 gibt einen Überblick über die Möglichkeiten der Farbmischung.
Abbildung 10: Das Farbdreieck. Gut zu sehen ist, dass Purpur keiner Wellenlänge entspricht.

Die Abbildung 10 zeigt zum Beispiel, dass aus Blau, Grün und Rot Weiß gemischt wird. Purpur entsteht durch Mischung von Rot und Blau. Es sei noch anzumerken, dass dies ein rein physiologischer Prozess und daher nicht subjektiv ist. Das zeigen Farbwahrnehmungen anderer Tierarten, wie der Bienen, die von der menschlichen Farbwahrnehmung entscheidend abweichen. Das hängt einfach von den Absorptionsspektren der Photorezeptoren ab (Bienen können UV-Licht wahrnehmen). 1.8.2. Subtraktive Farbmischung: Dies ist ein physikalischer Prozess bei dem Farbfilter Teile des Farbspektrums absorbieren. Die drei Komplementärfarben Cyan, Magenta und Gelb filtern unterschiedliche Farben heraus, so filtert Cyan Rot, Magenta Grün und Gelb Blau heraus. Angenommen blaues, rotes und grünes Licht träfen auf Cyan, so würde Rot herausgefiltert werden und ein Blau-Grüner Farbton wäre zu sehen. 1.9. Spektraltheorie Licht stellt einen kleinen Teil des Spektrums elektromagnetischer Strahlen dar. Der Mensch kann Licht der Wellenlängen 400nm bis 700nm wahrnehmen. Im gesamten Tierreich reicht Wahrnehmung von 300nm (UV-Licht, z.B. Bienen) bis 750nm (Infrarot, so z.B. Grubenottern mit deren Grubensinnesorganen Wärmestrahlung erkannt werden kann). Licht kann entweder monochromatisch (nur eine Wellenlänge) oder polychromatisch (mehrere Wellenlängen) sein. Eine wichtige Quelle für sichtbares Licht sind Elektronen, die von einem angeregten (energiereichen) Zustand in einen energieärmeren (stabileren) Zustand übergehen und die Energie dabei ein Photon emittiert. Dieser Prozess heißt spontane Emission und dauert ungefähr 10ns. Die Energiedifferenz des Elektrons lässt sich durch die Einsteinsche Gleichung E = hv = hc/(Lambda)U herleiten und lautet |UE| = hv = hc/(Lambda) wobei h das Plancksche Wirkungsquantum, LambdaUdie Wellenlänge und c die Lichtgeschwindigkeit ist.
- Linienspektrum: Die Energieniveaus in Atomen können keinen beliebigen Zwischenwerte annehmen, daher sind auch die Energien emittierter Photonen auf bestimmte Werte beschränkt. Licht zeigt sich dann als Linien ohne Übergang. Als Beispiel soll Natrium dienen, dass bei Verbrennung eine gelblich-orange Flamme erzeugt. Linienspektren sind bei Gasen oft beobachtbar.
- Kontinuierliche Spektren: Atome in Flüssigkeiten oder Festkörpern sind nahe beieinander und können miteinander stark wechselwirken, so dass die Energieniveaus der Atome zu ununterbrochenen Bereichen von Energieniveaus verbreitert sind. Wenn sich diese so genannten Energiebänder überlappen entsteht ein zusammenhängender Bereich von besetzbaren Energieniveaus und das Emissionsspektrum ist kontinuierlich. Als Beispiel soll hier der Regenbogen dienen.

Material und Methoden An Material wurden für die Versuche folgende Gegenstände verwendet: Perimeterapparatur, Augenklappe, Testmarken in den Farben Rot, Grün, Blau, Gelb, Schwarz und Weiß, Landolt’sche Testringe, Projektor mit regulierbarer Helligkeit und verschiedenen Farbfiltern, Schwanenhalslampe. Versuchsdurchführung: Versuch 1: Bei Versuch 1 soll die Winkelabhängige Empfindlichkeit der Retina für bestimmte Farben (Spektrenbereiche) bestimmt werden. Hierfür wird der Versuchsperson mit der Augenklappe ein Auge abgedeckt. Danach Positioniert sie den Kopf so, dass ihr freies Auge im Brennpunkt der Perimeterapparatur liegt (s.Abb.X, Punkt b). Nun kann man, indem man die Bunten Testmarken (Gelb, Grün, Rot, Blau, Schwarz und Weiß) von außen nach innen am Meridian entlang bewegt erkennen, wann die Versuchtpersonden Gegenstand bemerkt und wann sie die Farbe richtig erkennt. Sobald die Versuchsperson die Farbe richtig erkennt, kann man am Meridian den Winkel, vom Null Grad Winkel in der Mitte aus gemessen, ablesen, bei der die Versuchsperson für diese Farbe empfindlich ist. Indem man die Meridiane Verstellt und Winkel von 0°-180°, 45°-215°, 90°-270° und 135°-315° durchmisst, kann man das gesamte Gesichtsfeld Der Testperson Durchmessen. Die Ergebnisse werden in einem Koordinatensystem ausgewertet.
Bei diesem Versuch ist darauf zu achten, dass die Testperson immer den Punkt m (s.Abb.X) im Mittelpunkt des Perimeterbogens fixiert, der gleichmäßig beleuchtet sein muss, und auch den Kopf nicht bewegt. Des weiteren muss sie die unbunten Marken als Schwarz und Weiß erkennen und nicht nur als hell oder dunkel. Es ist auch zu beobachten, ob Regelmäßigkeiten bei den Farbverwechslungen vorliegen und woran dies liegen könnte. Die Messungen werden für jede Farbe und jeden Meridian mindestens doppelt durchgeführt und es ich darauf zu achten, dass es die Farbabfolge für die Testperson keinesfalls vorhersehbar ist.
Abb.X: Perimeterapparatur zur Bestimmung des Gesichtsfelds

Versuch 2: Bei diesem Versuch sollen die Sehschärfe und die Gesichtsfeldgrenzen bei unterschiedlichen Adaptationszuständen des Auges bestimmt werden. Der Versuchsaufbau hierfür ist analog zu Versuch 1, wiederum nimmt die Versuchsperson hinter einer Perimeterapparatur platz und hat ein Auge bedeckt. Allerdings werden nun statt bunter Testmarken Landolt’sche Testringe (s.Abb.Y) von außen am Meridian entlanggeführt (bei diesem Versuch nur der Horizontale also 90°-270°) und die Versuchsperson muss feststellen wo diese geöffnet sind. Landolt-Testringe sind Ringe mit genau definierten Abmessungen und einer Öffnung. Mithilfe dieser Definierten Größen und dem Abstand zwischen Auge und Testring kann man die Sehschärfe des Auges nach folgender Formel bestimmen:
1 1
1 8 0 6 0
S i nW i n k e l m i u t e n
d
E
α
απ
=
× ×=
×
S: Sehschärfe (nach Landolt) d: Breite der Öffnung im Ring E: Abstand Auge-Testring Damit ist dann α die Lücke in Winkelminuten, die die Versuchsperson gerade noch erkennen kann und die Sehschärfe der Kehrwert von α. Für den Versuch stehen zehn verschiedene Testringe zur Verfügung mit jeweils d= 18; 12; 9; 6; 3; 1,5; 0,9; 0,6; 0,3 und 0,15mm. Da bei diesem Versuch auch der Unterschied in der Sehschärfe zwischen Dunkel- und Helladaptation gemessen werden soll, findet jeweils eine Doppelmessung bei Helladaptiertem und Dunkeladaptierten Auge statt (Dunkeladaptation durch ca. halbstündigen Aufenthalt in Dunkelheit). Es werden wieder die Winkelwerte aufgeschrieben, bei der die Versuchsperson die Öffnungsrichtung richtig erkannt hat, und die Werte in ein Koordinatensystem übertragen. Bei diesem Versuch ist wiederum darauf zu achten, dass die Versuchsperson den Fixpunkt in der 0° Position des Meridians fixiert und den Kopf nicht bewegt. Auch ist auf eine gleichmäßige Ausleuchtung der Apparatur zu achten und dass die Reihenfolge der Öffnungsrichtung rein zufällig ist.
Versuch 3: Bei diesem Versuch soll das sogenannte Purkinje-Phänomen Demonstriert werden, dass sich aus den unterschiedlichen Empfindlichkeitsmaxima des Dunkeladaptierten und des Helladaptierten Auges ergibt. Da beim Dunkeladaptierten Auge vor allem mit den Stäbchen gesehen wird, die bei ca. 500nm die höchste Empfindlichkeit haben und beim Helladaptierten Auge mit den Zäpfchen, deren höchste Empfindlichkeit bei ca. 560nm liegt, wird bei einem kontinuierlichen Spektrum jeweils bei Dunkel-und Helladaptierten Auge ein anderer Bereich als der Hellste Wahrgenommen. Dazu wird das Licht einer Glühlampe (die ein kontinuierliches Spektrum aufweist) durch ein
Abb.Y: Prinzip eines Landolt’schen Testrings

Gitter geschickt, wo es sich wie in einem Prisma nach Wellenlänge aufspaltet. Die Versuchsperson markiert nun auf der Wand auf die Projiziert wird die für sie subjektiv hellste stelle und zwar erst im dunkeladaptierten Zustand (mindestens eine halbe stunde Aufenthalt in einem Dunklen Raum) und dann im helladaptierten. Zur Berechung der markierten Wellenlängen werden nun zwei definierte Wellenlängen (mit zwei definierten Farbfiltern erzeugt) auf die Wand projiziert und auch diese stellen markiert. Mit Hilfe der zwei Referenzlinien der definierten Wellenlängen kann man nun die beiden markierten, als am hellsten empfundenen Wellenlängen ausmessen und berechnen. Bei diesem Versuch ist darauf zu achten, dass bei der Messung mit Dunkeladaptierten Augen das Projizierte Spektrum nicht zu hell ist, sodass eine Umstellung auf Helladaptation erfolgt. Versuch 4: Bei diesem Versuch sollen die Adaptationskurven der beiden Sehsysteme erarbeitet werden. Dazu wird folgender Versuchsaufbau verwendet:
Bei diesem Versuchsaufbau wird die Helligkeit einer Lampe über einen Dimmer geregelt und das entstehende Licht durch einen Spalt und evtl. einen Farbfilter auf eine Wand projiziert. Zur Messung der relativen Helligkeit ist noch eine Photozelle im Strahlengang befestigt, die die relative Helligkeit über ein Voltmeter anzeigt. Beim ersten Versuchsteil sollen nur die Zäpfchen Angesprochen werden, daher wird ein Rotfilter in den Strahlengang eingebracht, da die Stäbchen unempfindlich gegen Rotes Licht sind. Zuerst wird die Versuchsperson extrem Helladaptiert durch dreiminütiges betrachten einer von einer sehr hellen Lampe beleuchteten Weißen Fläche. Anschließend Regelt sie die Helligkeit der Lampe über den Dimmer so, dass sie immer gerade noch den Lichtbalken auf der Wand erkennen kann. Dabei werden die Werte des Photometers immer halbminütlich abgelesen und Protokolliert. Wenn sich die Werte auf dem Voltmeter über längere Zeit nicht mehr ändern (die Versuchsperson also die Helligkeit nicht mehr Nachregelt) wird der Versuch abgebrochen, da sich das Zapfensehsystem der Versuchsperson soweit adaptiert hat, wie es kann und keine weitere Veränderung zu erwarten ist. Nun wird der Versuch wiederholt, nur ohne der Rotfilter, sodass das gesamte Sehsystem der Versuchsperson angesprochen wird. Hier wird der Versuch allerdings nicht abgebrochen wenn sich die Werte nicht mehr ändern, sondern mindestens 20min lang protokolliert, da sich nach der Adaptation der Zäpfchen nun auch die Adaptation der Stäbchen bemerkbar macht, die von dem Weißen Licht auch gereizt werden. Es sollte sich eine Adaptationskurve wie in Abb. Z2 ergeben.
Abb.Z: Versuchsaufbau zur Bestimmung der Adaptationskurven der beiden Sehsysteme
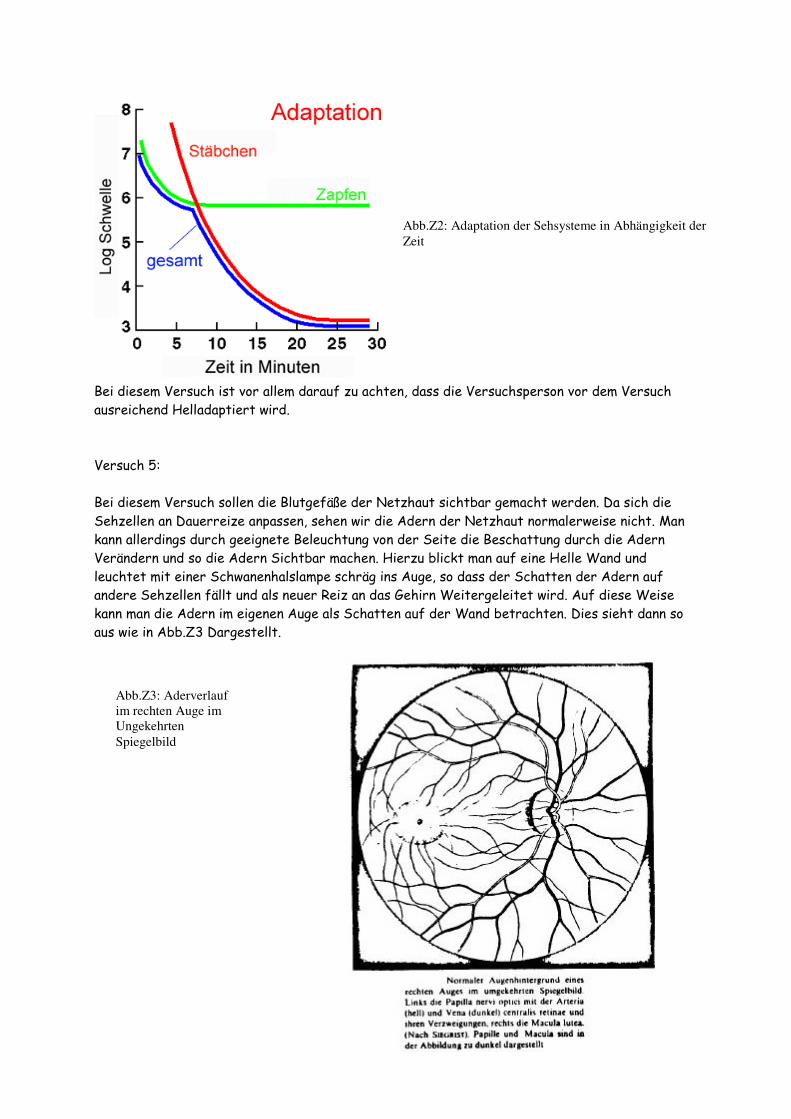
Bei diesem Versuch ist vor allem darauf zu achten, dass die Versuchsperson vor dem Versuch ausreichend Helladaptiert wird. Versuch 5: Bei diesem Versuch sollen die Blutgefäße der Netzhaut sichtbar gemacht werden. Da sich die Sehzellen an Dauerreize anpassen, sehen wir die Adern der Netzhaut normalerweise nicht. Man kann allerdings durch geeignete Beleuchtung von der Seite die Beschattung durch die Adern Verändern und so die Adern Sichtbar machen. Hierzu blickt man auf eine Helle Wand und leuchtet mit einer Schwanenhalslampe schräg ins Auge, so dass der Schatten der Adern auf andere Sehzellen fällt und als neuer Reiz an das Gehirn Weitergeleitet wird. Auf diese Weise kann man die Adern im eigenen Auge als Schatten auf der Wand betrachten. Dies sieht dann so aus wie in Abb.Z3 Dargestellt.
Abb.Z2: Adaptation der Sehsysteme in Abhängigkeit der
Zeit
Abb.Z3: Aderverlauf
im rechten Auge im
Ungekehrten
Spiegelbild

3. Ergebnisse 3.1 Die Gesichtsfeldgrenzen für bunte und unbunte Reize Tabelle1: Bestimmung der Grenzen des Gesichtsfeldes Farben Horizontal 0°-180° Vertikal 90°-270°
temporal-nasal nasal-temporal von oben von unten
rot 70° 20° 17,5° 53,5° blau 77° 31° 20° 45° grün 40,5° 20° 14° 31° schwarz 72° 29,5° 15° 56° weiß 86° 50° 30,5° 57° gelb 82° 42,5° 25° 50,5° Farben Diagonal 45°-215° Diagonal 135°-315°
temporal-nasal nasal-temporal temporal-nasal nasal-temporal
rot 70° 32,5° 39,5° 37° blau 68,5° 36,5° 40° 45° grün 46° 32,5° 30,5° 28° schwarz 75,5° 47,5° 61,5° 49° weiß 78° 46,5° 55° 48° gelb 65° 38° 46° 43° Der jeweilige Durchschnitt der gemessenen Werte (s.Anhang1) wird in ein Koordinatensystem eingetragen, das im Anhang 2 zu finden ist. Dabei wurden die Farben auf zwei Systeme verteilt, da das Ergebnis dadurch übersichtlicher wird. 3.2 Sehschärfe und Gesichtsfeldgrenzen Tabelle 2: Bestimmung der Sehschärfe (Dunkeladaption) Ringgröße temporal-nasal nasal-temporal Sehschärfe Ø 1. 2. Ø 1. 2. 0,15 Nicht erkannt Nicht erkannt 0,3 0° 0° 0° 0° 0° 0° 0,36 0,6 0° 0° 0° 0° 0° 0° 0,18 0,9 19,5° 0° 39° 0° 0° 0° 0,12 1,5 46° 42° 50° 46° 40° 52° 0,073
3 57,5°
52° 63° 46,5° 48° 45° 0,036
6 57,5°
60° 55° 52,5° 55° 50° 0,018
9 62,5°
64° 61° 58,5° 60° 57° 0,012
12 57,5°
60° 55° 53° 48° 58° 0,0092
18 72,5 75° 70° 64° 60° 68° 0,0061

° Die Sehschärfe S wird folgendermaßen berechnet: Die Formel aus Abschnitt 2.2 lautet am Beispiel d =3 mm und E = 38 cm:
14,27380
601803=
×Π
××=α
036,014,27
1==S
Die ermittelte Sehschärfe wird gegen den Blickwinkel im Diagramm 1 aufgetragen. Daraus ergibt sich folgende Kurve.
Diagramm 1: Dunkeladaption
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
72,5
62,5
62,5
19,5 0 0 46
52,5 53
Blickwinkel (°)
Sehschärfe (dpt)
Tabelle 3: Bestimmung der Sehschärfe (Helladaption) Ringgröße temporal-nasal nasal-temporal Sehschärfe Ø 1. 2. Ø 1. 2.
0,15 0° 0° 0° 0° 0° 0° 0,66 0,3 0° 0° 0° 10° 15° 5° 0,33 0,6 17,5° 16° 19° 13,5° 17° 10° 0,16 0,9 23° 21° 25° 30° 25° 35° 0,11 1,5 37,5° 34° 41° 44,5° 40° 49° 0,066 3 42,5° 53° 32° 49° 49° 49° 0,033 6 51° 63° 39° 67,5° 65° 70° 0,016 9 52,5° 50° 55° 68,5° 75° 62° 0,01 12 62° 65° 59° 66° 65° 67° 0,0082 18 62,5° 60° 65° 65,5° 66° 65° 0,0055 Die Sehschärfe bei Helladaption wird analog berechnet:
33,30340
601803=
×Π
××=α
033,0
33,30
1==S
Auch diese Werte werden im Diagramm 2 gegeneinander aufgetragen.
temporal nasal temporal

Diagramm 2: Helladaption
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
62,5
52,5
42,5 23 0 0
13,5
44,5
67,5 66
Blickwinkel (°)
Sehschärfe (dpt)
Leider konnten die Diagramme nicht in einem gemeinsamen Koordinatensystem abgebildet werden, da die Skalen zu unterschiedlich waren. 3.3 Das Purkinje Phänomen Skizze des Versuchs Die zwei Lilienspektren mit definierten Wellenlängen betragen für Linie 1 499nm und für Linie 2 571nm. Die hellste Stelle im kontinuierlichen Spektrum im Hellen wurde von der Versuchsperson mit dem Strich bei H markiert. Nach der Dunkeladaption wurde die hellste Stelle dagegen bei D gekennzeichnet. Anhand der Orientierungspunkte 1 und 2 können nun die Wellenlängen für D und H berechnet werden: Dunkeladaption (D) 507,5nm Helladaption (H) 543,5nm
1 D H 2
L = 17 cm ≈ 72 nm
L = 2 cm ≈ 8,5 nm
L = 10,5 cm ≈ 44,5 nm
temporal nasal temporal
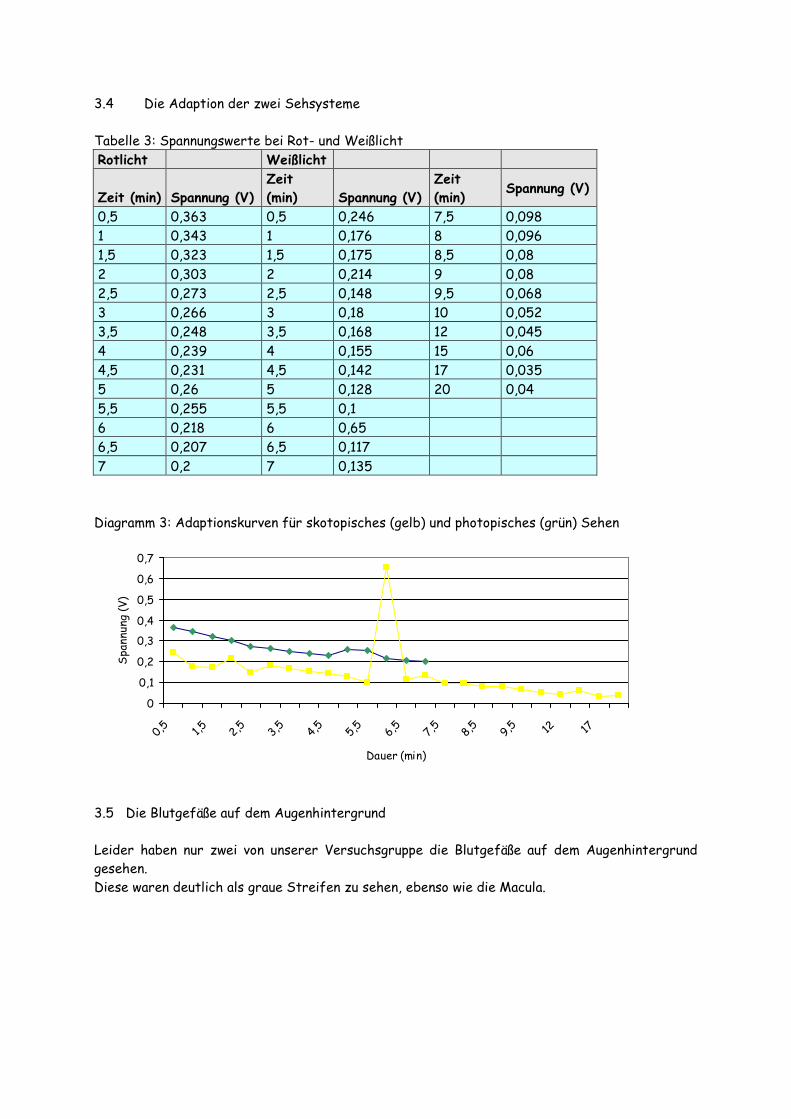
3.4 Die Adaption der zwei Sehsysteme Tabelle 3: Spannungswerte bei Rot- und Weißlicht Rotlicht Weißlicht
Zeit (min) Spannung (V)
Zeit
(min) Spannung (V)
Zeit
(min) Spannung (V)
0,5 0,363 0,5 0,246 7,5 0,098 1 0,343 1 0,176 8 0,096 1,5 0,323 1,5 0,175 8,5 0,08 2 0,303 2 0,214 9 0,08 2,5 0,273 2,5 0,148 9,5 0,068 3 0,266 3 0,18 10 0,052 3,5 0,248 3,5 0,168 12 0,045 4 0,239 4 0,155 15 0,06 4,5 0,231 4,5 0,142 17 0,035 5 0,26 5 0,128 20 0,04 5,5 0,255 5,5 0,1 6 0,218 6 0,65 6,5 0,207 6,5 0,117 7 0,2 7 0,135 Diagramm 3: Adaptionskurven für skotopisches (gelb) und photopisches (grün) Sehen
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,5 1,5 2,
53,5
4,5
5,5
6,5
7,5
8,5
9,5 12 17
Dauer (min)
Spannung (V)
h
3.5 Die Blutgefäße auf dem Augenhintergrund Leider haben nur zwei von unserer Versuchsgruppe die Blutgefäße auf dem Augenhintergrund gesehen. Diese waren deutlich als graue Streifen zu sehen, ebenso wie die Macula.

4.Diskussion
Untersuchung der spektralen Empfindlichkeit der Netzhaut und der Gesichtsfeldgrenzen für bunte und unbunte Testmarken Bei diesem Versuch würde man aufgrund einschlägiger Literatur erwarten, dass auf Grund der Gegenfarbtheorie die Gesichtsfelder jeweils für weiß und schwarz, rot und grün sowie gelb und blau sehr ähnlich sind. Dies trifft bei der Testperson relativ genau zu, bis auf rot und grün. Bei grün hat sie dabei meist mehrere Versuche gebraucht, bis die richtige Farbe erkannt wurde. Allerdings wurde auch bei den anderen Farben dieses Phänomen beobachtet. Die Farberkennung fing dabei oft bei schwarz oder weiß an, ging dann über ein bis mehrere Farben um dann bei der richtigen zu landen. Ein häufiges Beispiel dafür ist schwarz, gelb, grün. Ähnliche Farben wie weiß und gelb wurden bemerkenswerter Weise selten verwechselt, obwohl hier die Gefahr dafür eigentlich am größten ist. Das Gesichtsfeld unserer Testperson ist für weiß am größten, was sich durch die Verteilung der Zapfen und Stäbchen in der Retina erklären lässt. Die vertikale Ausrichtung der Gesichtsfeldgrenzen ist auf der nasalen Seite des Auges geringer als auf der temporalen, da die Nase den Blickwinkel einschränkt. Weitere Fehlerquellen während des Versuchs könnten hier (aber auch bei den folgenden Tests) zum einen die Ablenkung durch andere Personen sein, da im selben Raum noch andere Versuche stattfanden und die Umgebung ziemlich laut war. Zum anderen ist es schwierig ständig nur den Perimetermittelpunkt zu fixieren, da der Blick automatisch auf einen sich bewegenden Gegenstand gerichtet wird und die Augen auf Dauer schmerzen. Bestimmung der Sehschärfe und der Gesichtsfeldgrenzen bei unterschiedlichem Adaptionszustand Beide Kurven zeigen den erwarteten Verlauf, denn aus den Kurven kann man lesen, dass das Auflösungsvermögen in der Peripherie des Gesichtsfeldes geringer ist als in der Mitte. Durch einen Vergleich der erhaltenen Linien lässt sich außerdem schließen, dass die Sehschärfe eines dunkeladaptierten Auges wesentlich geringer ist als die des helladaptierten. Diese Ergebnisse resultieren aus der unterschiedlichen Vernetzung der Photorezeptorzellen in der Retina. Während in der Fovea centralis jeder Zapfen auf ein Folge-Ganglion geschaltet ist, was eine hohe Bildauflösung ermöglicht, sind in der Peripherie viele Stäbchen auf ein Ganglion verschaltet, das Bild ist unscharf. Beim Sehen im Dunkeln nützt aber die Fovea, die Stelle schärfsten Sehens nichts, da die Zapfen von den Wellenlängen des Dämmerungslichts nicht erregt werden. Hier sieht man mit der Peripherie besser. Als Fehlerquelle ist hierbei die Augenklappe zu nennen, denn wenn ein kleiner Landolt´scher Testring von nasal nach temporal bewegt wird und die Öffnung zeigt entgegen der Bewegung, kann man erst relativ spät erkennen, wo die Öffnung ist. Nachweis zweier verschiedener Sehsysteme für das hell- bzw. dunkeladaptierte Auge Wie erwartet liegt zwischen den beiden hellsten Punkten bei Hell- und Dunkeladaption ein Abstand von 36 nm. Diese Erscheinung wird Purkinje-Phänomen bezeichnet (s. Abschnitt 1.4.4). Demnach sollte das Absorptionsmaximum beim skotopischen Sehen bei 500 nm und beim photopischen bei 560 nm liegen.

Bei unserem Versuch sind die Werte sowohl nach oben, als auch nach unten verschoben. Das könnte daher kommen, dass die Testperson schräg neben dem Spektrum stand und dieses deshalb verschoben wahrnahm. Außerdem waren die kennzeichnenden Striche an der Wand nicht dünn und gerade sondern doppelt, wodurch das Abzeichnen erschwert wurde. Und schlussendlich spielt auch die subjektive Farbwahrnehmung der Testperson eine Rolle. Aufnahme der Adaptionskurven für beide Sehsysteme Die Kurven zeigen den typischen Adaptionsverlauf. Die Dunkeladaption der Zapfen erreicht nach ein paar Minuten eine Grenze, von der ab keine weitere Adaption mehr möglich ist, da die Zapfen über eine geringere Rhodopsinmenge als die Stäbchen verfügen. Sie sind daher weit weniger lichtempfindlich. Die Stäbchen können selbst sehr geringe Lichtintensitäten noch wahrnehmen, da sie große Mengen an Rhodopsin haben. Sie sind daher in der Lage selbst wenige Lichtquanten zu absorbieren und in Reize umzuwandeln. Die kleinen Schwankungen der Kurve lassen sich durch die Empfindlichkeit des Voltmeters und der Photozelle erklären. Diese ist so empfindlich, dass sie schon Intensitätsschwankungen angibt, wenn der Tisch auf dem sie steht wackelt. Außerdem ist sie nur mit Tesafilm festgeklebt und verrutscht deshalb leicht. Der Ausreisser bei 6 min (Weißlicht) kommt dadurch zustande, das die Testperson zu heftig am Dimmer gedreht hat. Eigenbeobachtung der Blutgefäße auf dem Augenhintergrund Normalerweise sind die Blutgefäße nicht zu erkennen, da sich die Sehzellen an Dauerreize anpassen. Fällt nun seitlich Licht in das Auge, fällt der Schatten anderer Blutgefäße auf die Retina. Dieser Reiz wird von den Rezeptorzellen registriert und weitergeleitet, man kann das Innere des eigenen Auges sehen. 5. Quellenangaben
Abb.1: http://www.hh.schule.de/ngb/ltb/ltb-03/Hirn/Auge.JPG Abb.2: http://www.catalase.com/retina.gif Abb.3: http://ep.llnl.gov/msds/orgchem/Chem227/Vision/cis-retinal.gif Abb.4: http://ep.llnl.gov/msds/orgchem/Chem227/Vision/retinal.gif Abb.5: http://depts.washington.edu/ophthweb/images/KPgraphic.jpg Abb.6: http://www.accessexcellence.org/AE/AEC/CC/vision_background.html Abb.7: http://www.seafriends.org.nz/phgraph/phdwg45.gif Abb.8: http://www.optik-reiter.de/Start/Produkte/Kontaktlinsen/Firmen/Wissen/Akk.jpg Abb.9: http://www.unimannheim.de/fakul/psycho/irtel/lehre/seminararbeiten/ w96/Farbe/bilder/FARBKRS.GIF Abb.10: http://www.zeiss.de/C12567B000352701/GraphikTitelIntern/Color Triangle/$File/Farbdreieck.jpg Versuchsskript http://www.egbeck.de/skripten/12/bs12-38.htm 6. Anhang
Ergebnisse während der Versuche Diagramme zu 3.1


















![Ein theoretischer Dialog zwischen einem Muslim … · Web viewEin theoretischer Dialog zwischen einem Muslim und einem Christ [ Deutsch - German – ألماني ] islamweb.net Übersetzer:](https://img.pdfslide.org/doc/110x75/5e57fc0cb58bf50c12682b14/ein-theoretischer-dialog-zwischen-einem-muslim-web-view-ein-theoretischer-dialog.jpg)










