Embed Size (px)
Citation preview

Herstellung sicherer und wirksamer Lebendvakzine gegen die Koi Herpesvirus
Infektion von Karpfen
I n a u g u r a l d i s s e r t a t i o n
zur
Erlangung des akademischen Grades eines
Doktor der Naturwissenschaften
(Dr. rer. nat.)
der Mathematisch-Naturwissenschaftlichen Fakultät
der
Universität Greifswald
vorgelegt von
Lars Schröder
geboren am 25.02.1988
in Schwerin
Greifswald, 18.04.2019

Dekan: Prof. Dr. Werner Weitschies 1. Gutachter: Prof. Dr. Dr. h.c. Thomas C. Mettenleiter 2. Gutachter: Prof. Dr. Dieter Steinhagen Tag der Promotion: 12.04.2019

„Um an die Quelle zu kommen, muss man gegen den Strom schwimmen.“
- Konfuzius
Ich widme diese Arbeit meiner kleinen Tochter Neele,
die ich über alles liebe und für die ich immer da sein werde
und allen Menschen, die an mich geglaubt und mich so weit gebracht haben.

Inhaltsverzeichnis
1. Einleitung .................................................................................... 1
1.1. Das Koi-Herpesvirus ................................................................... 2
1.1.1. KHV als Vertreter der Familie Alloherpesviridae ........................... 2
1.1.2. Übertragung und Ausbreitung von KHV ....................................... 3
1.1.3. Krankheitsbild einer KHV-Infektion ............................................ 4
1.1.4. Wirtsspektrum von KHV .......................................................... 5
1.2. Molekulare Struktur und Replikation von KHV .................................... 5
1.2.1. Struktur und Genomaufbau von KHV .......................................... 5
1.2.2. Replikationszyklus von KHV ..................................................... 6
1.2.3. Virale Enzyme für den Nukleotidstoffwechsel als Virulenzfaktoren ..... 9
1.2.4. Membran-Glykoproteine ....................................................... 10
1.3. Herstellung von Impfstoffen gegen virale Infektionen ........................ 11
1.3.1. Lebendvakzine .................................................................. 11
1.3.2. DIVA-Vakzine .................................................................... 11
1.3.3. Genetechnik in der Impfstoffentwicklung .................................. 12
1.4. Das Immunsystem der Fische ...................................................... 13
2. Zielsetzung ................................................................................ 19
3. Publikationen .............................................................................. 21
3.1. Publikation I .......................................................................... 21
3.2. Publikation II ......................................................................... 35
3.3. Pubilkation III ........................................................................ 67
4. Eigenanteil an den zur Dissertation eingereichten Publikationen ................ 77
4.1. Publikation I .......................................................................... 77
4.2. Publikation II ......................................................................... 79
4.3. Publikation III ........................................................................ 81
5. Zusammenfassende Diskussion der Ergebnisse ....................................... 85
6. Quellen ..................................................................................... 93
7. Zusammenfassung der Dissertation .................................................... 99
8. Summary .................................................................................. 101
9. Anhang .................................................................................... 103
9.1. Eigenständigkeitserklärung ....................................................... 103
9.2. Publikationen und Tagungsbeiträge ............................................. 104
9.2.1. Tagungsbeiträge ............................................................... 104

9.2.2. Publikationen .................................................................. 105
9.3 Lebenslauf ........................................................................... 107
9.4 Danksagung .......................................................................... 109


Einleitung
1
1. Einleitung
Der Karpfen zählt neben den Regenbogenforellen zu den bedeutendsten in
Deutschland gezüchteten Speisefischarten. Insbesondere in Bayern, Sachsen und
Brandenburg spielt die Aquakultur von Karpfen eine große Rolle. Weltweit werden
nach Angaben der Ernährungs- und Landwirtschaftsorganisation der Vereinten
Nationen (FAO) 4,15 Millionen Tonnen Karpfenfleisch pro Jahr erzeugt (FAQ, 2014;
Füllner, Pfeifer, & Langner, 2007). Die Karpfen-Teichwirtschaft wird jedoch seit
Ende der 90iger Jahre durch das Koi-Herpesvirus (KHV oder Cyprinid herpesvirus 3)
bedroht, welches Massensterben bei Koi und Nutzkarpfen verursacht (Ayana
Perelberg et al., 2003; Hedrick et al., 2000). Eine KHV-Infektion führt zur schwerer
klinischer Symptomatik mit Mortalitätsraten zwischen 70 und 80 % (Bretzinger,
Fischer-Scherl, Oumouna, Hoffmann, & Truyen, 1999). Deshalb wurde die KHV
Infektion in Deutschland im Dezember 2005 in die Verordnung über anzeigepflichtige
Krankheiten aufgenommen. Diese Anzeigepflicht beschränkte sich zunächst nur auf
den Virusnachweis bei Nutzkarpfen, wurde dann aber im Januar 2006 auch auf
Zierfisch-Zuchtformen der Karpfen, die sogenannten Kois, erweitert (TierSG, §1 VO
über anzeigepflichtige Tierseuchen; Fischseuchen-VO, Richtlinie 91/67/EWG,
Anhang A). Durch die Fischseuchen-Verordnung in Verbindung mit der Verordnung
über anzeigepflichtige Krankheiten auf der Grundlage der EU-Richtlinie 2006/88/EG,
ist die Bekämpfung und Diagnose von KHV in Deutschland streng geregelt. KHV ist
mittlerweile weltweit verbreitet und verursacht Ausbrüche in Deutschland,
Frankreich, Italien, Österreich, dem Vereinigten Königreich, Polen Belgien,
Dänemark, den Niederlanden, der Schweiz, Indonesien, Japan, Südafrika, Taiwan,
den USA und Thailand (Haenen, Way, Bergmann, & Ariel, 2004; Sano et al., 2004).
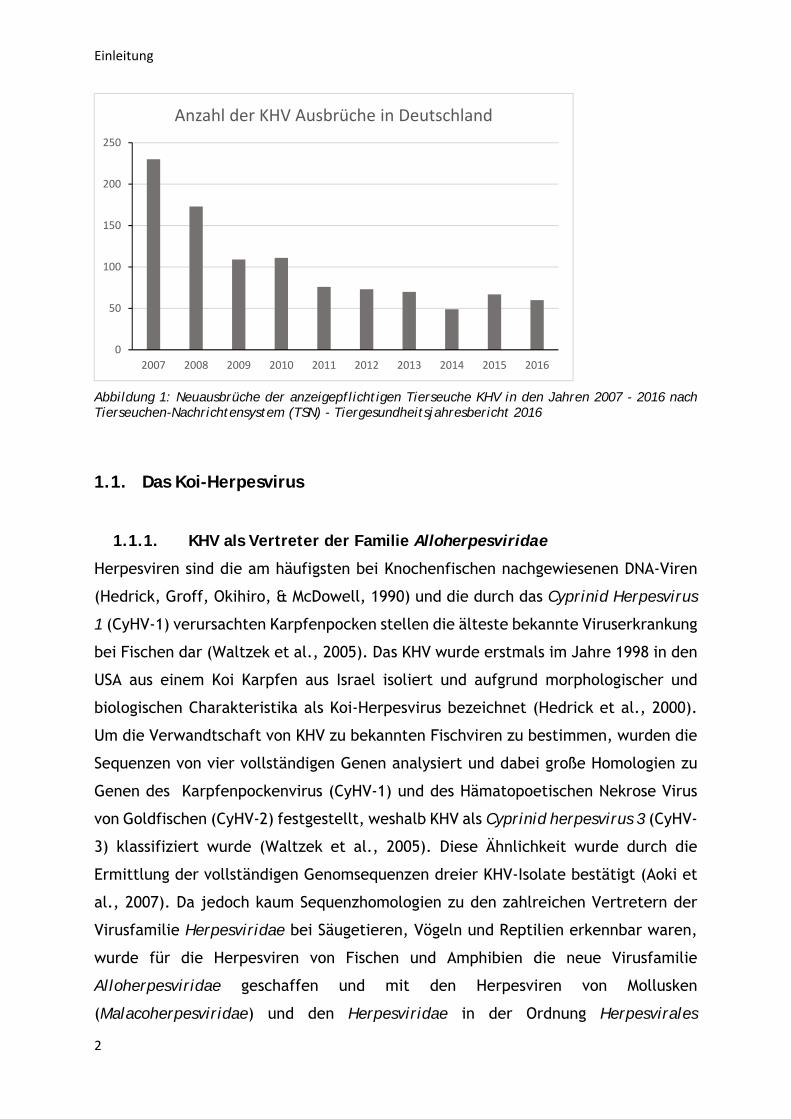
Im Jahre 2016 wurden alleine in Deutschland wieder 60 KHV- Ausbrüche angezeigt
(siehe Abbildung 1). Aus diesem Grund ist die Entwicklung schützender Vakzinen
dringend notwendig und steht auch im Mittelpunkt dieser Promotionsschrift.
Einleitung
2
Abbildung 1: Neuausbrüche der anzeigepflichtigen Tierseuche KHV in den Jahren 2007 - 2016 nach Tierseuchen-Nachrichtensystem (TSN) - Tiergesundheitsjahresbericht 2016
1.1. Das Koi-Herpesvirus
1.1.1. KHV als Vertreter der Familie Alloherpesviridae
Herpesviren sind die am häufigsten bei Knochenfischen nachgewiesenen DNA-Viren
(Hedrick, Groff, Okihiro, & McDowell, 1990) und die durch das Cyprinid Herpesvirus
1 (CyHV-1) verursachten Karpfenpocken stellen die älteste bekannte Viruserkrankung
bei Fischen dar (Waltzek et al., 2005). Das KHV wurde erstmals im Jahre 1998 in den
USA aus einem Koi Karpfen aus Israel isoliert und aufgrund morphologischer und
biologischen Charakteristika als Koi-Herpesvirus bezeichnet (Hedrick et al., 2000).
Um die Verwandtschaft von KHV zu bekannten Fischviren zu bestimmen, wurden die
Sequenzen von vier vollständigen Genen analysiert und dabei große Homologien zu
Genen des Karpfenpockenvirus (CyHV-1) und des Hämatopoetischen Nekrose Virus
von Goldfischen (CyHV-2) festgestellt, weshalb KHV als Cyprinid herpesvirus 3 (CyHV-
3) klassifiziert wurde (Waltzek et al., 2005). Diese Ähnlichkeit wurde durch die
Ermittlung der vollständigen Genomsequenzen dreier KHV-Isolate bestätigt (Aoki et
al., 2007). Da jedoch kaum Sequenzhomologien zu den zahlreichen Vertretern der
Virusfamilie Herpesviridae bei Säugetieren, Vögeln und Reptilien erkennbar waren,
wurde für die Herpesviren von Fischen und Amphibien die neue Virusfamilie
Alloherpesviridae geschaffen und mit den Herpesviren von Mollusken
(Malacoherpesviridae) und den Herpesviridae in der Ordnung Herpesvirales
0
50
100
150
200
250
2007 2008 2009 2010 2011 2012 2013 2014 2015 2016
Anzahl der KHV Ausbrüche in Deutschland

Einleitung
3
zusammengefasst (Davison et al., 2009). Die Familie Alloherpesviridae umfasst
zurzeit 13 Spezies, die aufgrund ihrer Genomsequenzen und Wirtsorganismen in 4
Gattungen Batrachovirus (Viren der Ranidae), Cyprinivirus (Viren der Cyprinidae und
Anguillidae), Ictalurivirus (Viren der Ictaluridae und Acipenseridae) und
Salmonivirus (Viren der Salmonidae) aufgeteilt wurden
(https://talk.ictvonline.org/taxonomy/).
1.1.2. Übertragung und Ausbreitung von KHV
Im Allgemeinen werden KHV-bedingte Krankheitsausbrüche nur bei
Wassertemperaturen über 17°C beobachtet, doch gibt es Hinweise darauf, dass das
Virus auch bei einer Temperatur von 10°C noch vermehrt werden kann (Baumer,
Fabian, Wilkens, Steinhagen, & Runge, 2013; Gilad et al., 2004). Bei
Wassertemperaturen unter 13°C scheint KHV symptomlos im Wirt zu überdauern und
erst im optimalen Temperaturbereich beginnen die Tiere infektiöse Viruspartikel
auszuscheiden (St-Hilaire et al., 2005). Die Übertragung von KHV erfolgt über das
Wasser, in dem das Virus über mehrere Stunden lang infektiös bleiben kann (Ayana
Perelberg et al., 2003). Normalerweise beträgt die Inkubationszeit in Abhängigkeit
von Wassertemperatur, Virulenz des Erregers und Empfänglichkeit des Fisches
zwischen 4 bis 21 Tagen, jedoch kann die KHV-Infektion (KHV-I) bei latent infizierten
Fischen Jahre zurückliegen (Hedrick et al., 2000; Ayana Perelberg et al., 2003). Über
symptomfreie, latent infizierte Fische konnte das Virus unerkannt weltweit in
zahlreiche Karpfenbestände eingeschleppt werden (Walster, 1999). Lebenslange,
latente Infektionen sind typisch für Herpesviren. Während dieses Stadiums ist die
virale Replikation und Genexpression erheblich herunterreguliert, was dazu führt,
dass latente Herpesviren nur sehr schwer nachweisbar sind. In Stresssituationen
können die Viren jedoch jederzeit reaktiviert und auf neue Wirte übertragen werden
(Fraser, Lawrence, Wroblewska, Gilden, & Koprowski, 1981; Galloway, Fenoglio, &
McDougall, 1982). Die ersten KHV Infektionen wurden in den Jahren 1998 und 1999
in Israel, USA und Deutschland dokumentiert (Bretzinger et al., 1999; Hedrick et al.,
2000; Ayana Perelberg et al., 2003). Nach der Veröffentlichung dieser Daten wurden
auch in vielen anderen Ländern in Europa, Asien und Nordamerika Ausbrüche der
KHV-I gemeldet (Haenen et al., 2004; Sano et al., 2004; Walster, 1999).

Einleitung
4
1.1.3. Krankheitsbild einer KHV-Infektion
Das KHV ist hochinfektiös und wird über direkten Hautkontakt oder das Wasser
übertragen. Die Virusausscheidung erfolgt über Haut und Kiemen und wahrscheinlich
auch über den Darm. Dabei löst die Infektion eines Bestandes unter natürlichen
Bedingungen eine Morbidität von 80-100% und eine Mortalität von 70-80% aus
(Bretzinger et al., 1999; Walster, 1999). Die Symptomatik kann von Tier zu Tier stark
variieren, äußert sich aber oftmals im Anfangsstadium durch vermehrte
Schleimproduktion auf der Haut und an den Kiemen. In einigen Fällen wurden aber
auch Enophthalmus und Hämorrhagien der Haut beobachtet. Im fortgeschrittenen
Stadium treten dann Schwellungen des Kiemengewebes auf, die bis hin zur
Ausbildungen großflächiger Nekrosen führen können. Des Weiteren verringert sich
die Schleimproduktion und es kommt zur Ausbildung rauer „Sandpapier“-artiger
Haut. Typische Verhaltensänderungen infizierter Tiere sind das Aufsuchen von
strömungsschwachen Wasserzonen, Anorexie, Koordinationsverlust und Apathie (S.
M. Bergmann et al., 2010; Hedrick et al., 2000; Walster, 1999). Ein Hauptproblem
bei der KHV-Infektion ist die Latenz-Etablierung in überlebenden Tieren, die
lebenslang gesunde Tiere im Umfeld infizieren können (Gilad et al., 2004; St-Hilaire
et al., 2005). KHV persistiert in den Leukozyten und überdauert im latenten Zustand
in den Kiemen, Haut und Nieren der Tiere bis zum nächsten Ausbruch (S. Bergmann
& Kempter, 2011).
Abbildung 2: Typisches Krankheitsbild einer KHV Infektion. Gezeigt sind Kiemennekrose und Hautläsionen. Fotos von Dr. Sven Bergmann

Einleitung
5
1.1.4. Wirtsspektrum von KHV
Wie viele andere Herpesviren besitzt auch KHV ein sehr enges Wirtsspektrum
bezüglich der Auslösung einer Erkrankung (Davison, 2002; Walster, 1999). Dieses
beschränkt sich auf Koi- und Karpfenpopulationen, also Fische der Spezies Cyprinus
carpio. Dabei sind Karpfen aller Altersklassen gleichermaßen für eine KHV-Infektion
empfänglich (Hedrick et al., 2000). Jedoch wurden deutliche Unterschiede in der
Mortalitätsrate bei den verschiedenen Altersgruppen festgestellt. So reagieren Tiere
mit einem Gewicht von unter 10g, was etwa Karpfen der Altersgruppe K0 bis K1
entspricht, empfindlicher auf eine KHV-Infektion als Karpfen mit einem Gewicht von
über 200g (≥ K2) (Ayana Perelberg et al., 2003). In den Altersklassen werden die
Karpfen anhand der durchlebten Sommer eingeteilt; so steht K2 für einen
„zweisömmerigen Karpfen“. Mehrfach wurden Versuche durchgeführt, in denen KHV-
infizierte Karpfen mit anderen Fischspezies zusammen kultiviert wurden, bei denen
jedoch bisher niemals Anzeichen einer Erkrankung festgestellt wurden (Ayana
Perelberg et al., 2003; Hedrick et al., 2000; Hedrick, Waltzek, & Mcdowell, 2006;
Walster, 1999). Allerdings konnte das Virus in anderen Fischspezies (Wie unter
anderem Hecht, Goldfische und Karauschen) nachgewiesen werden. Diese Fische
zeigten selbst keinerlei Symptome, waren aber dennoch in der Lage, Karpfen mit
KHV zu infizieren (Meyer, 2007).
1.2. Molekulare Struktur und Replikation von KHV
1.2.1. Struktur und Genomaufbau von KHV
Wie erwähnt, wurde das komplette KHV Genom erstmals 2007 basierend auf drei
Virusisolaten aus Israel, Japan und den USA charakterisiert (Aoki et al., 2007). Das
lineare, doppelsträngige DNA Genom ist etwa 295 kbp groß und codiert für ca. 156
Gene, von denen 8 in zwei Kopien innerhalb direkter Sequenz-Repetitionen an
beiden Genomenden vorliegen (Aoki et al., 2007). Ähnliche DNA-
Sequenzwiederholungen sind charakteristisch für die meisten Herpesviren, wobei
interne und terminale, sowie direkte und inverse Repetitionen vorkommen
(McGeoch, Rixon, & Davison, 2006). Anhand von Anzahl und Lage der
Sequenzwiederholungen werden die Genome der Herpesviren in sechs Klassen
eingeteilt (A-F), wobei das KHV-Genom der Klasse A entspricht (Aoki et al., 2007;

Einleitung
6
Brooks, Carroll, Butel, Morse, & Mietzner, 2013). Wie bei allen Herpesviren wird das
KHV-Genom in ein ikosaedrisches Kapsid verpackt, welches von zwei proteinhaltigen
Tegumentschichten umgeben und mit einer Lipidmembran zellulären Ursprungs
umhüllt wird, in welche verschiedene virale Glykoproteine eingelagert sind (Aoki et
al., 2007; Davison et al., 2009). Das ikosaedrische Kapsid der Triangulationszahl 16
besteht aus insgesamt 161 Kapsomeren (150 Hexone und 11 Pentone). Darüber hinaus
befindet sich an einem Kapsidvertex ein Portalkomplex, durch welchen die virale
DNA ins Kapsid geschleust wird (Homa et al., 2013). Das Tegument enthält zahlreiche
virale und einige zelluläre Proteine und besteht aus einer inneren, Kapsid-nahen und
einer äußeren Schicht (Bechtel, Winant, & Ganem, 2005; Johannsen et al., 2004;
Mettenleiter, 2008). Die molekulare Zusammensetzung der Tegumentschichten ist
bei den Alloherpesviridae noch weitgehend unbekannt. Besser charakterisiert sind
hingegen die Membranproteine der Virushülle des KHV, von denen durch
massenspektrometrische Proteomanalysen 13 Stück nachgewiesen wurden (Michel et
al., 2010).
1.2.2. Replikationszyklus von KHV
Der Replikationszyklus von humanen und anderen Säuger-Herpesviren wurde mit
Hilfe ultrastruktureller und molekularbiologischer Analysen bereits detailiert
untersucht (Granzow et al., 1997; Mettenleiter, 2002; Mettenleiter, Klupp, &
Granzow, 2009). Die Infektion beginnt mit der Virusadsorption an Rezeptoren der
Wirtszellmembran und der anschließenden Fusion von Virushülle und
Plasmamembran mit Hilfe viraler Fusionsproteine (Eisenberg et al., 2012). Bei den
Alphaherpesvirinae erfolgt die Rezeptorbindung meist über das Glykoprotein gD, und
die Fusionsmaschinerie besteht aus den viralen Glykoproteinen gB, gH und gL
(Eisenberg et al., 2012). Die funktionshomologen Proteine der Alloherpesviren
wurden bislang nicht eindeutig identifiziert. Nach erfolgreicher Penetration und
damit dem Eintritt des Kapsides in das Zytoplasma löst sich die äußere
Tegumentschicht ab, während die innere Tegumentschicht mit dem Kapsid
verbunden bleibt und für den Transport der Kapside entlang der Mikrotubuli zum
Zellkern benötigt wird (Antinone et al., 2006; Granzow, Klupp, & Mettenleiter,
2005). Das virale Genom wird durch Kernporen in den Zellkern entlassen wo die die
virale DNA zirkularisiert und repliziert wird (Strang & Stow, 2005). Voraussetzung
hierfür ist allerdings zunächst die Expression viraler Gene durch die zelluläre

Einleitung
7
Transkriptions- und Translationsmaschinerie. Die herpesvirale Genexpression ist
kaskadenartig reguliert, wobei je nach Zeitpunkt ihrer Transkription sehr frühe
(immediate early, IE), frühe (early, E) und späte (late, L) Gene unterschieden
werden. Die E-Gene werden zusätzlich in early und delayed early bzw. early-late
Gene unterteilt (Honess & Roizman, 1974, 1975; Roizman, 2001; Tombacz, Toth,
Petrovszki, & Boldogkoi, 2009). Die Expression der IE-Gene wird häufig durch virale
Tegumentproteine induziert, was jedoch nicht essenziell ist, da auch gereinigte
Herpesvirus DNA infektiös ist (Feldman, Demarchi, Ben-Porat, & Kaplan, 1982; Fuchs,
Granzow, Klupp, Kopp, & Mettenleiter, 2002; Ihara, Feldman, Watanabe, & Ben-
Porat, 1983; Katan, Haigh, Verrijzer, van der Vliet, & O'Hare, 1990). Die E-Gene
codieren vor allem für Proteine, welche für die DNA-Replikation (Helikase, Primase,
DNA-Polymerase) und den Nukleotidstoffwechsel (Thymidinkinase, dUTPase,
Ribonukleotidreduktase) benötigt werden (Cheung, 1991; Huang & Wu, 2004;
Taharaguchi et al., 1994; Tombacz et al., 2009; Watanabe, Ono, Nikami, & Kida,
1998). Die L-Gene, die als letztes exprimiert werden, kodieren vor allem für virale
Strukturproteine (Johnson & Everett, 1986; Tombacz et al., 2009). Aus einigen davon
werden in einem autokatalytischen Prozess die Kapside im Zellkern gebildet, in
welche dann die replizierte Virus-DNA eingeschleust und auf Genomlänge
zugeschnitten wird (Mettenleiter, 2008). Die entstandenen Nukleokapside lagern
sich an die innere Kernmembran an und gelangen durch Knospung in den
perinukleären Spalt (Mettenleiter, 2008). Die primäre Virushülle fusioniert dann mit
der äußeren Kernmembran und die Kapside werden ins Zytoplasma freigesetzt
(Mettenleiter, 2008). Dort lagern sich die inneren Tegumentproteine an, während
die äußerenen Tegumentproteine erst zusammen mit den Glykoproteinen während
der Knospung im Trans-Golgi-Netzwerk eingebaut werden (Mettenleiter, 2008). Die
sekundär umhüllten, reifen Virionen werden in Transportvesikeln zur
Plasmamembran transportiert und gelangen dort durch Fusion der Vesikelmembran
mit der Plasmamembran in den extrazellulären Raum (Mettenleiter, 2008).
Einleitung
8
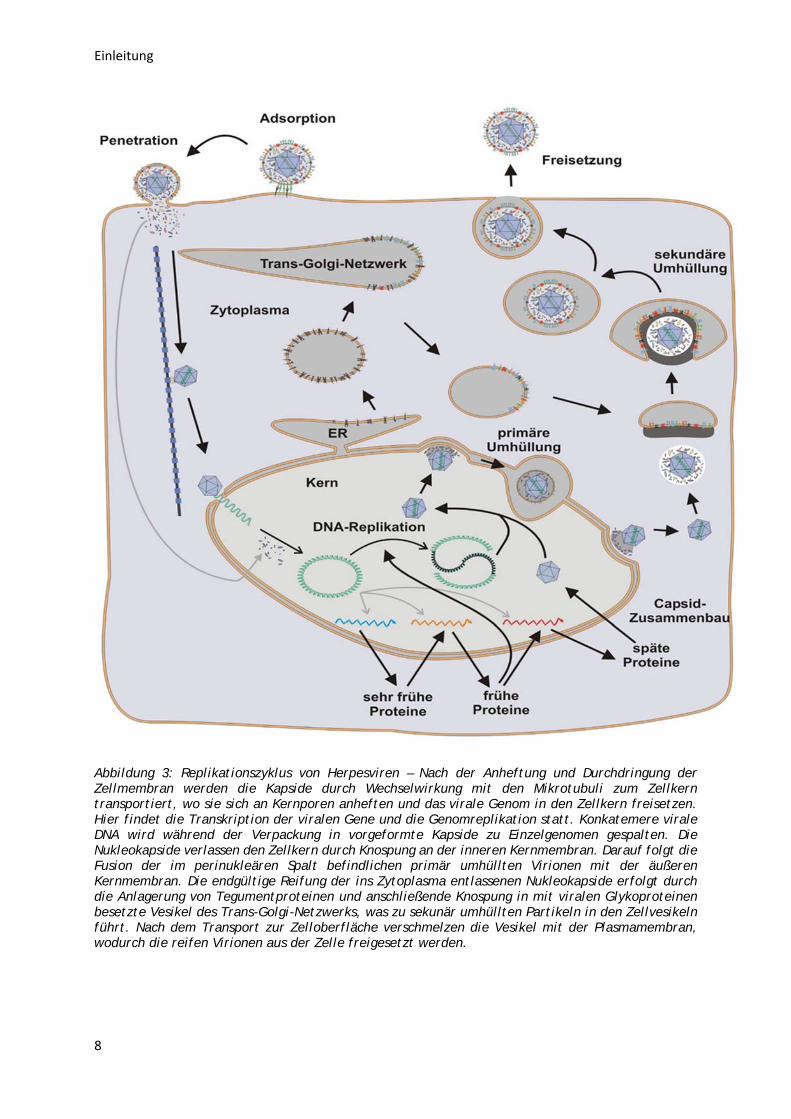
Abbildung 3: Replikationszyklus von Herpesviren – Nach der Anheftung und Durchdringung der Zellmembran werden die Kapside durch Wechselwirkung mit den Mikrotubuli zum Zellkern transportiert, wo sie sich an Kernporen anheften und das virale Genom in den Zellkern freisetzen. Hier findet die Transkription der viralen Gene und die Genomreplikation statt. Konkatemere virale DNA wird während der Verpackung in vorgeformte Kapside zu Einzelgenomen gespalten. Die Nukleokapside verlassen den Zellkern durch Knospung an der inneren Kernmembran. Darauf folgt die Fusion der im perinukleären Spalt befindlichen primär umhüllten Virionen mit der äußeren Kernmembran. Die endgültige Reifung der ins Zytoplasma entlassenen Nukleokapside erfolgt durch die Anlagerung von Tegumentproteinen und anschließende Knospung in mit viralen Glykoproteinen besetzte Vesikel des Trans-Golgi-Netzwerks, was zu sekunär umhüllten Partikeln in den Zellvesikeln führt. Nach dem Transport zur Zelloberfläche verschmelzen die Vesikel mit der Plasmamembran, wodurch die reifen Virionen aus der Zelle freigesetzt werden.

Einleitung
9
1.2.3. Virale Enzyme für den Nukleotidstoffwechsel als Virulenzfaktoren
Die Genome vieler Herpesviren codieren für verschiedene Nukleotid-
Stoffwechselenzyme Thymidinkinase (TK), Thymidylatkinase,
Desoxyuridintriphosphat-Pyrophosphatase (dUTPase, DUT) und
Ribonukleotidreduktase. Da es sich hierbei um ubiquitäre Enzyme handelt, die in
allen teilungsaktiven Zellen exprimiert werden, sind die viralen Homologen für die
Replikation in Zellkultur meist entbehrlich. Allerdings gelten die entsprechenden
Gene bei vielen Herpesviren, einschließlich des Katzenwels Virus (channel catfish
virus, CCV), als wichtige Virulenzfaktoren (Boivin, Coulombe, & Rivest, 2002;
Efstathiou, Kemp, Darby, & Minson, 1989; Freuling, Müller, & Mettenleiter, 2017;
Jöns, Gerdts, Lange, Kaden, & Mettenleiter, 1997; Jons & Mettenleiter, 1996; Zhang
& Hanson, 1995). Bei KHV wurde die Rolle der TK für die Virulenzausprägung in
früheren Studien kontrovers diskutiert.
Die Thymidinkinase (TK) ist ein bei fast allen Organismen mit Ausnahme von Pilzen
vorhandenes Enzym des salvage pathway (Bergungsweg) der Nukleotid-Biosynthese,
welches Desoxythymidin zu Desoxythymidinmonophosphat (dTMP) umwandelt. Auch
die meisten DNA-Viren, von Bakteriophagen bis zu humanpathogenen Viren, wie z.B.
die Herpes-simplex-Viren (HSV-1/2), codieren für eigene Thymidinkinasen, die sich
hinsichtlich ihrer Struktur und Reaktivität teilweise von den zellulären Enzymen
unterscheidet (Wintersberger, 1997). Auch KHV besitzt ein TK-Gen, das für ein
Polypeptid von 224 Aminosäuren mit einer berechneten Molekülmasse von 24,6 kDa
kodiert, und dessen Kinaseaktivität in vitro nachgewiesen werden konnte (Bercovier
et al., 2005).
Die Enzyme der dUTPase-Familie spielen eine wichtige Rolle sowohl für die
Genomintegrität als auch bei der De-novo-Thymidylat-Biosynthese. Die dUTPasen
katalysieren die Hydrolyse von dUTP zu dUMP und anorganischem Pyrophosphat.
Diese Reaktion dient zum einen der Bereitstellung eines Vorläufers (dUMP) für die
Synthese von Thymin-Nukleotiden, die für die DNA-Replikation benötigt werden, und
zum anderen dazu, die intrazelluläre Konzentration von dUTP zu begrenzen. Ein
erhöhter dUTP-Spiegel führt zu einer verstärkten Inkorporation von Uracil in die DNA,
die eine durch Uracil-DNA Glycosylase vermittelte Reparatur durch Exzision
induziert. Dieser Reparaturprozess kann zur DNA-Fragmentierung und damit zum
Zelltod führen (Rona et al., 2013).

Einleitung
10
1.2.4. Membran-Glykoproteine
Die Hülle von Herpesviren wird von Vesikeln des Trans-Golgi-Netzwerks gebildet,
welche die tegumentierten Kapside am Ende der Virusmorphogenese umschließen
(Granzow et al., 1997; Mettenleiter et al., 2009; Turcotte, Letellier, & Lippe, 2005).
In diese Lipidhülle sind virale, häufig glykosylierte Oberflächenproteine eingelagert
(Mettenleiter et al., 2009; Reske, Pollara, Krummenacher, Chain, & Katz, 2007), die
vielfältige Funktionen haben. So erfolgt das Andocken von Herpesvirus-Partikeln an
die Oberfläche der Wirtszelle meist durch die Bindung viraler Glykoproteine (bei
Alphaherpesviren v.a. des Glykoproteins gC) an Heparansulfat-tragende
Proteoglykane (Eisenberg et al., 2012; Rauh, 1990). Für die stabile Virusadsorption
an die Wirtszelle und die anschließende Fusion der Virushülle mit der
Plasmamembran sind weitere virale Glykoproteine notwendig (bei Alphaherpesviren
gD, gB, gH und gL) (Eisenberg et al., 2012; Heldwein & Krummenacher, 2008).
Weitere Glykoproteine sind an der Virusreifung und Freisetzung beteiligt (bei
Alphaherpesviren gE, gI, gM) (Mettenleiter et al., 2009). Die funktionshomologen
Proteine der Alloherpesviridae sind bislang nicht bekannt. Darüber hinaus sind die
viralen Hüllproteine wichtige Ziele der Wirtsimmunantwort.
Als erste Voraussetzung für die Identifizierung und Charakterisierung der
Hüllproteine von KHV wurden neben massenspektrometrischen Proteomanalysen
(Michel et al., 2010) auch monospezifische Antiseren gegen die von den Genen
ORF25, ORF65, ORF81, ORF99, ORF136, ORF138, ORF146, ORF148 und ORF149
codierten vorherhergesagten integralen Membranproteine hergestellt (Fuchs,
Granzow, Dauber, Fichtner, & Mettenleiter, 2014; Rosenkranz et al., 2008). Mittels
dieser Seren konnten die Typ I-Membranproteine pORF25, pORF65, pORF99, pORF136
und pORF149, sowie vor allem das Typ III-Membranprotein pORF81 in KHV-infizierten
Zellen eindeutig nachgewiesen werden. Umgekehrt reagierten in indirekten
Immunfluoreszenztests (IIFT) Seren von natürlich oder experimentell mit KHV
infizierten Karpfen und Koi mit Plasmid-transfizierten Zellen, die die Proteine
pORF25, pORF65, pORF148 oder pORF149 exprimierten (Fuchs et al., 2014). In einer
weiteren Studie konnte gezeigt werden, dass die von ORF32, ORF59, ORF81, ORF83,
ORF99, ORF106, ORF115, und ORF131 codierten KHV-Membranproteine für die
Virusreplikation essenziell sind, während ORF25, ORF64, ORF65, ORF132, ORF136,
ORF148 oder ORF149 in Zellkultur entbehrlich sind. Allerdings führte die Deletion der

Einleitung
11
Gene ORF148 oder ORF25 zu einer Abschwächung der Virulenz von KHV in vivo
(Vancsok et al., 2017).
1.3. Herstellung von Impfstoffen gegen virale Infektionen
1.3.1. Lebendvakzine
Lebendimpfstoffe sind attenuierte Viren, die in der Lage sind innerhalb des Wirts zu
replizieren, aber keine oder allenfalls eine sehr milde Erkrankung verursachen.
Wichtig dabei ist, dass der Verlust der Virulenz, die immunogenen Eigenschaften der
Viren nicht beeinträchtigt (Keystone, Kozarsky, Freedman, Nothdruft, & and Conno,
2013). In den meisten Fällen zeigen Lebendimpfstoffe eine signifikant höhere
Immunogenität als inaktivierte Impfstoffe, da sie eine natürliche Infektion
nachahmen und so ein breiteres Spektrum an sowohl humoralen (B-Zellen) als auch
zellulären (CD8 + und CD4 + T-Zellen) Immunreaktionen hervorgerufen wird
(Keystone et al., 2013). Eine einmalige Impfung reicht meist aus, um einen
langfristigen, manchmal sogar lebenslangen Schutz zu erreichen. Ein Nachteil dieses
Impfstofftyps kann jedoch das Risiko einer Virulenzsteigerung durch Rückmutation
des abgeschwächten Virus sein. Bei neueren Lebendimpfstoffen, die genau
definierte und stabile Deletionen aufweisen, besteht jedoch kein Risiko für
Rückmutationen (Keystone et al., 2013).
1.3.2. DIVA-Vakzine
Als DIVA-Vakzine (engl. differentiation of infected and vaccinated animals) oder
Markervakzine werden Impfstoffe bezeichnet, die es erlauben, geimpfte Tiere von
Feldvirus-infizierten Tieren zu unterscheiden (van Oirschot, 1999). Dabei geht es
jedoch nicht nur darum, dass sich das Impfvirus genetisch vom Wildtyp-Virus
unterscheidet, sondern dass die Antikörperantwort von infizierten Tieren von der von
geimpften Tieren unterschieden werden kann (van Oirschot, Rziha, Moonen, Pol, &
van Zaane, 1986). Die durch eine Virusinfektion induzierten Antikörper können mit
Hilfe serologischer Tests, wie zum Beispiel einem ELISA (Enzyme-linked
Immunosorbent Assay), in der Regel weitaus länger nachgewiesen werden als das
Virus selbst. Bei DIVA-Vakzinen werden deshalb meist ausgewählte immunogene

Einleitung
12
Proteine deletiert, die für den Immunschutz der Tiere nicht essenziell sind. So kann
anhand des Fehlens oder Vorhandenseins von Serumantikörpern gegen das
entsprechende Protein unterschieden werden, ob ein Tier lediglich geimpft, oder
entweder ausschließlich oder zusätzlich durch das Wildtypvirus infiziert wurde (M.,
2006; van Oirschot, 1999). Zum Einsatz kommt diese Art von Impfstoffen vor allem
in der Nutztierhaltung, wo im Hinblick auf die Tilgung von Seuchen die Verbreitung
der Erreger ständig kontrolliert werden muss.
1.3.3. Genetechnik in der Impfstoffentwicklung
Die Verfügbarkeit hochwirksamer und sicherer Impfstoffen für die Veterinärmedizin
ist eine Grundvoraussetzung für die Tiergesundheitskontrolle (Krishnan, 2000;
Meeusen, Walker, Peters, Pastoret, & Jungersen, 2007). Mittels revers genetischer
Systeme ist es möglich, Pathogene wie Viren gezielt zu verändern und so eine neue
Generation von attenuierten Lebendvakzine-Kandidaten zu entwickeln (Freuling et
al., 2017; van Oirschot, 1999). Gentechnische Methoden erleichtern auch die
Herstellung von DIVA-Vakzinen, welche eine genetische und/oder serologische
Differenzierung von Feldvirus-infizierten und geimpften Tieren ermöglichen (siehe
oben). Auch zur Bekämpfung der KHV-Infektion wurden neben mittels klassischer
Methoden abgeschwächten Lebendimpfstoffen aus virulenten Feldisolaten (A.
Perelberg, Ronen, Hutoran, Smith, & Kotler, 2005; Ronen et al., 2003) auch bereits
gentechnisch hergestellte Gendeletionsmutanten erprobt. Grundlage hierfür war die
Kenntnis der Genomsequenz des KHV und der Funktionen und strukturellen Merkmale
einzelner Virusproteine (Aoki et al., 2007; Boutier et al., 2015; Costes et al., 2008;
Fuchs, Fichtner, Bergmann, & Mettenleiter, 2011; Vancsok et al., 2017). Es konnte
bereits in früheren Studien belegt werden, dass virale Enzyme des
Nukleotidstoffelwechsels, wie die TK und die dUTPase (DUT), Virulenzfaktoren von
Herpesviren einschließlich des KHV sind (Freuling et al., 2017; Fuchs et al., 2011;
Jons & Mettenleiter, 1996; Kit, Kit, & Pirtle, 1985). Deshalb wurden auch in der
vorliegenden Arbeit die TK- und DUT-Gene mittels homologer Rekombination einzeln
und in Kombination aus dem KHV-Genom deletiert. Außerdem wurden die Gene der
nicht essenziellen Glykoproteine pORF 148 und pORF 149 deletiert, da diese hoch
immunogen sind (Fuchs et al., 2014) und somit als Marker für eine serologische DIVA
Vakzine dienen könnten. Um die Herstellung der Mutanten und eventuell auch die

Einleitung
13
spätere Impfstoffproduktion zu erleichtern, wurde der Virusstamm KHV-T (Taiwan)
verwendet, da dieser an die Zellkultur adaptiert ist und deutlich höhere Titer als
andere KHV-Isolate erreicht (Gao et al., 2018; Hedrick et al., 2000; Mletzko et al.,
2017).
Ein ebenfalls vielversprechender gentechnischer Ansatz sind sogenannte DNA-
Impfstoffe. Diese basieren auf Expressionsplasmiden für einzelne immunogene
Virusproteine, im Falle von KHV die Membranproteine pORF25 und pORF81, welche
den Fischen meist mehrmals injiziert werden (Zhou et al., 2014b). Solche DNA-
Vakzinen unterstützen ebenfalls die DIVA-Diagnostik und könnten zum Schutz
wertvoller Zierfische (Kois) durchaus geeignet sein. Wegen des großen
Arbeitsaufwands und der relativ hohen Produktionskosten dürfte ein Einsatz bei
Nutzfischen jedoch unwirtschaftlich sein, weshalb dieser Ansatz in der vorliegenden
Arbeit nicht verfolgt wurde.
1.4. Das Immunsystem der Fische
Das Immunsystem umfasst biologische Strukturen und Prozesse innerhalb eines
Organismus, die eine Vielzahl von Noxen erkennen und eliminieren können. Es
schützt insbesondere vor Krankheitserregern und ist in der Lage fremde Zellen, sowie
infizierte eigene Zellen von eigenen gesunden Zellen zu unterscheiden. Alle
Lebewesen verfügen über Immunsysteme, wobei man bei Vertebraten und so auch
bei Fischen neben einem angeborenen zusätzlich ein klassisches „lernfähiges“,
adaptives Immunsystem findet (Beck, 2007; Murphy, 2012; Pradipta, Bismita, &
Surajit, 2012). Wie neuere Studien jedoch zeigen, verfügen bereits Bakterien über
adaptive Immunmechanismen zur Phagenabwehr (z.B. das CRISPR/Cas9 System), die
inzwischen auch vielfältig für molekularbiologische Studien in Eukaryoten eingesetzt
werden (Jinek et al., 2012). Im Grunde unterscheidet sich die Immunantwort bei
Fischen nicht wesentlich von der höherer Vertebraten (Pradipta et al., 2012). Fische
können in ihrem aquatischen Lebensraum, insbesondere bei hoher Bestanddichte und
schlechten Umweltbedingungen über ihre Schleimhäute einer hohen Erregerlast
ausgesetzt sein. Dies erfordert ein effektives Immunsystem, das auch in der Lage
sein muss, bei unterschiedlichen Wassertemperaturen eine Abwehr aufrecht zu
halten (Ellis, 2001).

Einleitung
14
Wie bei höheren Wirbeltieren besteht das Immunsystem von Fischen aus einer
angeborenen (engl. innate) und einer adaptiven Komponente, die sich wiederum in
humorale und zelluläre Kompartimente unterteilen. Zu den angeborenen humoralen
Komponenten des Immunsystems gehören antimikrobielle Peptide, Lysozyme und das
Komplement-System. Zu den zellulären Komponenten des angeborenen
Immunsystems zählen u.a. natürliche Killer- (NK-) Zellen, Makrophagen und
Granulozyten (Murphy, 2012). Bei den adaptiven humoralen Komponenten handelt es
sich um die von B-Zellen gebildeten Immunglobuline (Antikörper), während T-
Lymphozyten die zelluläre Komponente darstellen (Luttmann, Bratke, Küpper, &
Myrtek, 2006; Murphy, 2012). Angeborene Komponenten des Immunsystems werden
u.a. durch physikalische Barrieren, z.B. die mukosalen Oberflächen der Haut bzw.
der Kiemen repräsentiert. Diese verhindern, dass Krankheitserreger wie Bakterien
und Viren in den Organismus gelangen (Alberts, 2002; Boyton & Openshaw, 2002).
Substanzen im Schleim wie antimikrobielle Peptide, Lysozym und Komplement-
Komponenten, welche von den Epithelzellen abgesondert werden, gehören somit zu
den initialen Abwehrmechanismen von Fischen (Nakao, Tsujikura, Ichiki, Vo, &
Somamoto, 2011). Eine weitere physikalisch-chemische Barriere bildet die
Magensäure (Secombes, 1996). Nach der Infektion von Fischen mit Viren produzieren
infizierte Zellen verschieden Interferone, die antivirale Abwehrmechanismen
induzieren. Für die angeborene zelluläre Immunantwort ist eine Vielzahl von
unterschiedlichen Leukozyten-Subpopulationen verantwortlich, wie Makrophagen,
Granulozyten und NK-Zellen (Ellis, 2001; Litman, Cannon, & Dishaw, 2005).
Makrophagen dienen als zentrale Schaltstelle des Immunsystems, da sie
Krankheitserreger aufnehmen und deren Abbauprodukte an Zellen des adaptiven
Immunsystems präsentieren. Dabei produzieren sie eine Vielzahl von Botenstoffen
die zur Regulation der angeborenen und adaptiven Immunantwort beitragen (Vallejo
et al., 1992). Gelingt es den Krankheitserregern, die Barrieren der angeborenen
Immunität zu überwinden, greift das adaptive Immunsystem. Hierzu gehören die von
Plasmazellen gebildeten Antigen-spezifischen Immunglobuline (humorale Immunität)
sowie zytotoxische T-Zellen und T-Helferzellen mit ihren Antigen-spezifischen T-
Zellrezeptoren (zellvermittelte Immunität). Zytotoxische T-Zellen sind in der Lage,
virusinfizierte Zellen, die virale Peptide an der Oberfläche präsentieren, zu lysieren
und somit die Virusvermehrung zu hemmen (Ellis, 2001). B- und T-Zellen spielen
Einleitung
15
durch die Generierung von Gedächtniszellen auch eine wichtige Rolle beim Schutz
vor wiederkehrenden Infektionen (Ellis, 2001).
Immunglobuline oder Antikörper sind große, Y-förmige Proteine, die hauptsächlich
von Plasmazellen produziert werden und mit ihren variablen Regionen an antigene
Strukturen von Krankheitserregern wie Bakterien und Viren binden. Damit können sie
Erreger einerseits direkt oder im Zusammenspiel mit Komplement neutralisieren
bzw. für deren weitere Degradation aufbereiten (Luttmann et al., 2006; Murphy,
2012).
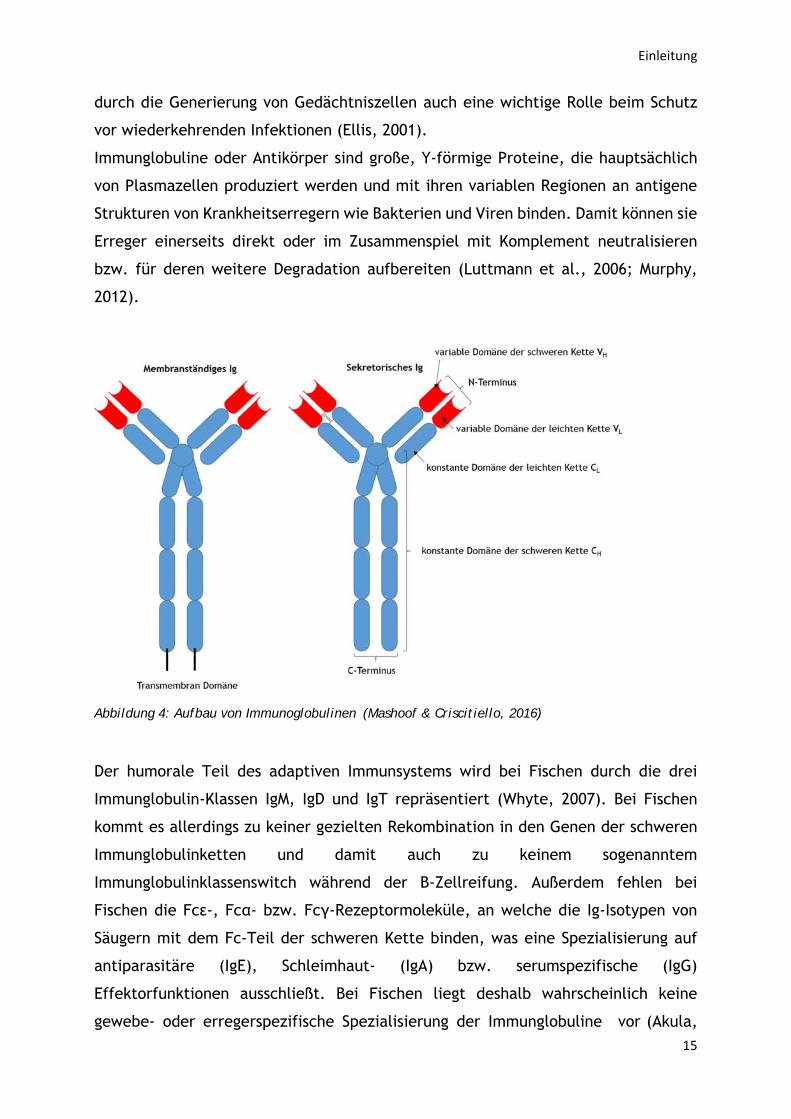
Abbildung 4: Aufbau von Immunoglobulinen (Mashoof & Criscitiello, 2016)
Der humorale Teil des adaptiven Immunsystems wird bei Fischen durch die drei
Immunglobulin-Klassen IgM, IgD und IgT repräsentiert (Whyte, 2007). Bei Fischen
kommt es allerdings zu keiner gezielten Rekombination in den Genen der schweren
Immunglobulinketten und damit auch zu keinem sogenanntem
Immunglobulinklassenswitch während der B-Zellreifung. Außerdem fehlen bei
Fischen die Fcε-, Fcα- bzw. Fcγ-Rezeptormoleküle, an welche die Ig-Isotypen von
Säugern mit dem Fc-Teil der schweren Kette binden, was eine Spezialisierung auf
antiparasitäre (IgE), Schleimhaut- (IgA) bzw. serumspezifische (IgG)
Effektorfunktionen ausschließt. Bei Fischen liegt deshalb wahrscheinlich keine
gewebe- oder erregerspezifische Spezialisierung der Immunglobuline vor (Akula,

Einleitung
16
Mohammadamin, & Hellman, 2014; Wakae et al., 2006). IgM ist eine oligomere Form
von Antikörpern die über Disulfidbrücken bei Säugern Pentamere und bei Fischen
Tetramere bilden. IgM ist der initiale Antikörpertyp, der nach einer Infektion als
Reaktion auf die Antigen-Exposition auftritt (Luttmann et al., 2006). Zunächst galt
IgM als die einzige Immunglobulinklasse bei Knochenfischen, bis bei
Regenbogenforellen und Zebrafischen neuartige, als IgT bzw. IgZ bezeichnete
Immunglobuline entdeckt wurden (Hansen, Landis, & Phillips, 2005; Sunyer, 2013).
Spätere Studien zeigten, dass IgT eine Schlüsselrolle bei der Immunreaktion der Haut
und der Kiemenschleimhaut spielt (Parra, Korytar, Takizawa, & Sunyer, 2016). Das
Immunglobulin D (IgD) ist ein membranständiger Antikörper-Isotyp, der während der
Reifung von B-Zellen durch alternatives mRNA-Spleißen mit IgM koexprimiert wird
(Chen et al., 2009). Die genaue Funktion von IgD ist bislang noch nicht bekannt.
Abbildung 5: Aufbau von Immunoglobulin M - Nach (Hordvik, 2015; Luttmann et al., 2006)
? = Art der Wechselwirkungen zwischen den Untereinheiten ist nicht bekannt.
IgM und IgT werden durch sekretierende B-Zellen bzw. Plasmazellen produziert und
über polymere Immunoglobulinrezeptoren aus dem Epithel in die luminale
Schleimschicht v. a. des Darmes transportiert (Parra et al., 2016) wo es mit der
Darmflora interagiert. In der Lamina propria der Schleimhäute werden Pathogene
oder deren Bestandteile insbesondere von Makrophagen aufgenommen und an T-

Einleitung
17
Helferzellen präsentiert. Gleichfalls können B-Zellen über ihre
Oberflächenimmunglobuline solche Antigene binden und bei Fischen auch
phagozytieren (Sunyer, 2013). Dadurch und durch die stimulierende Wirkung von
Antigen-spezifischen T-Helferzellen beginnen B-Zellen sich zu vermehren und zu
plasmazellenähnlichen Zellen zu differenzieren, die antigenspezifische
Immunglobuline sezernieren (Parra et al., 2016).
Die bedeutendsten immunkompetenten Organe bei Fischen sind Thymus, Niere, Milz,
Darm assoziertes lymphatisches Gewebe (GALT, gut associated lymphoid tissue) und
das interbranchiale lymphatische Gewebe (interbranchial lymphoid tissue) (Fischer,
Koppang, & Nakanishi, 2013). Der Thymus ist ein weiteres lymphatisches Organ und
produziert vor allem T-Lymphozyten, die zu zytotoxischen T-Zellen und T-
Helferzellen differenzieren (Bowden, Cook, & Rombout, 2005). Die Kopfniere enthält
ein breites Repertoire an Immunzellen, darunter die höchste Konzentration an sich
entwickelnden B-Lymphozyten und auch geringe Mengen an Antikörper-
sekretierenden Zellen (Whyte, 2007; Zwollo, Cole, Bromage, & Kaattari, 2005). Die
Milz fungiert wie bei Säugern hauptsächlich als sekundäres Immunorgan, das viele
reife B-Zellen und, IgM-produzierende Zellen enthält (Zwollo et al., 2005; Zwollo,
Haines, Rosato, & Gumulak-Smith, 2008). Da Fische keine Lymphknoten haben, spielt
wahrscheinlich die Milz eine zentrale Rolle beim sogenannten Antigen-Trapping
(Soleto, Fischer, Tafalla, & Granja, 2018). Als Antigen-Trapping wird die
Konzentrierung von Antigenen an der Oberfläche von dendritischen Zellen der
sekundären lymphatischen Organe bezeichnet, sodass eine Erkennung durch
immunkompetente Zellen erfolgen kann. Dendritische Zellen sind auch bei Fischen
beschrieben worden (Soleto et al., 2018).


Zielsetzung
19
2. Zielsetzung
In den 1990er Jahren wurde die KHV-infektion erstmals in Israel und Europa als eine
neuartige Infektionskrankheit von Kois und Karpfen beobachtet und hat sich
mittlerweile weltweit ausgebreitet. Auch in Deutschland wurde die KHV-I im Jahre
2005 in die Verordnung über anzeigepflichtige Tierseuchen aufgenommen und stellt
eine ständige Bedrohung für die hiesige Karpfen-Population dar, die immer wieder
zu erheblichen Verlusten führt.
Das Hauptziel dieser Promotionsschrift war es daher, eine stabil abgeschwächte,
schützende und kostengünstige KHV-Lebendvakzine mithilfe gentechnischer
Verfahren herzustellen. Darüber hinaus sollten Methoden etabliert werden, die eine
genetische und serologische Differenzierung von immunisierten und Wildtyp-KHV
infizierten Fischen erlaubten (DIVA-Prinzip). Die hergestellten Virusrekombinanten
sollten in vitro und in vivo charakterisiert und getestet werden. Hierbei kam es
besonders darauf an, dass durch die eingefügten Deletionen die Vermehrbarkeit in
Zellkultur nicht signifikant beeinträchtigt, die Virulenz in Karpfen jedoch deutlich
reduziert wurde. Neben diesen praktischen Aspekten sollten die im Rahmen dieser
Arbeit gewonnenen Erkenntnisse auch zu einem besseren allgemeinen Verständnis
der Biologie der bislang wenig charakterisierten Familie der Alloherpesviridae
beitragen.


Publikationen
21
3. Publikationen
3.1. Publikation I
“Generation of a potential koi herpesvirus live vaccine by simultaneous deletion of the viral thymdine kinase and dUTPase genes”
Lars Schröder, Sandro Klafack, Sven M. Bergmann, Dieter Fichtner, Yeonhwa Jin, Pei-Yu Lee, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs
Publiziert in Journal of General Virology

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
Generation of a potential koi herpesvirus live vaccine bysimultaneous deletion of the viral thymidine kinase anddUTPase genes
Lars Schröder,1 Sandro Klafack,2 Sven M. Bergmann,2 Dieter Fichtner,2 Yeonhwa Jin,2 Pei-Yu Lee,3 Dirk Höper,4
Thomas C. Mettenleiter1 and Walter Fuchs1,*
Abstract
Koi herpesvirus (KHV, Cyprinidherpesvirus 3) causes a fatal disease of koi and common carp. To obtain safe and efficacious
live vaccines, we generated deletion mutants of KHV lacking the nonessential genes encoding two enzymes of nucleotide
metabolism, thymidine kinase (TK, ORF55) and deoxyuridine-triphosphatase (DUT, ORF123). Since single-deletion mutants
based on a KHV isolate from Israel (KHV-I) only exhibited partial attenuation (Fuchs W, Fichtner D, Bergmann SM,
Mettenleiter TC. Arch Virol 2011;156 : 1059–1063), a corresponding double mutant was generated and tested in vivo, and
shown to be almost avirulent but still protective. To overcome the low in vitro virus titres of KHV-I (�105 p.f.u. ml�1), single
and double TK and DUT deletions were also introduced into a cell culture-adapted KHV strain from Taiwan (KHV-T). The
deletions did not affect in vitro virus replication, and all KHV-T mutants exhibited wild-type-like plaque sizes and titres
exceeding 107 p.f.u. ml�1, as a prerequisite for economic vaccine production. Compared to wild-type and revertant viruses,
the single-deletion mutants of KHV-T were significantly attenuated in vivo, and immersion of juvenile carp in water containing
high doses of the double mutant caused almost no fatalities. Nevertheless, the deletion mutants induced similar levels of
KHV-specific serum antibodies to the parental wild-type virus, and conferred solid protection against disease after challenge
with wild-type KHV. For the convenient differentiation of DNA samples prepared from gill swabs of carp infected with wild-
type and TK-deleted KHV we developed a triplex real-time PCR. Thus, KHV-TDDUT/TK might be suitable as a genetic DIVA
vaccine in the field.
INTRODUCTION
In the late 20th century a previously unknown virus infec-tion led to mass mortality of koi and common carp (Cypri-nus carpio) in Israel and Europe. The causative agent wasidentified as a herpesvirus and designated as carp nephritisand gill necrosis virus (CNGNV) or koi herpesvirus (KHV)[1–4]. During the following years this virus spread acrossmajor parts of the world and caused considerable losses infood and ornamental fish [5]. DNA sequencing of threevirus isolates from the United States, Israel and Japanrevealed an approximately 295 kbp type A herpesvirusgenome containing 22 kbp direct repeat sequences at bothtermini and 156 different open reading frames (ORFs) [6].Sequence analyses further demonstrated a close relationshipto carp pox virus (Cyprinid herpesvirus 1) and goldfish
haematopoietic necrosis virus (Cyprinid herpesvirus 2), and,therefore, KHV was classified as Cyprinid herpesvirus 3(CyHV-3) within the genus Cyprinivirus in the family Allo-herpesviridae and the order Herpesvirales [7] (https://talk.ictvonline.org/taxonomy/).
Because of the economic relevance of KHV disease, safe,efficacious and, at least for common carp, affordable vac-cines are urgently needed. In the first attempts, inactivatedKHV preparations and attenuated live virus vaccinesobtained after cell culture passage and/or UV irradiation ofvirulent KHV were evaluated [4, 8]. Although the latterproved to be useful, the molecular basis for their attenuationremained unclear, which meant that there was a risk ofreversion to a more virulent phenotype. Furthermore, thesevaccines did not support the differentiation of naturally
Received 27 June 2018; Accepted 23 August 2018Author affiliations:
1Institute of Molecular Virology and Cell Biology, Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany; 2Institute ofInfectology, Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany; 3GeneReach Biotechnology Corporation, Taichung, Taiwan, ROC; 4Institute ofDiagnostic Virology, Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany.*Correspondence: Walter Fuchs, [email protected]: koi herpesvirus; Cyprinid herpesvirus 3; vaccine; thymidine kinase; dUTPase.Abbreviations: CCB, common carp brain cells; Ct, cycle threshold; CyHV-3, Cyprinid herpesvirus 3; DIVA, differentiation of infected from vaccinated ani-mals; DUT, dUTPase; FBS, foetal bovine serum; IIF, indirect immunofluorescence; KHV, koi herpesvirus; KHV-T, KHV strain Taiwan; MEM, minimumessential medium; qPCR, quantitative real-time PCR; TK, thymidine kinase.
RESEARCH ARTICLESchröder et al., Journal of General Virology
DOI 10.1099/jgv.0.001148
001148 ã 2018 The Authors
1

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
infected from vaccinated animals (the DIVA principle [9]).Therefore, candidate subunit or DNA vaccines containingor encoding single envelope glycoproteins of KHV, such aspORF25 or pORF81, have been developed, and have beenshown to induce protective immune responses [10–12]. Onthe other hand, defined gene deletions have been introducedby targeted mutagenesis of the KHV genome cloned as abacterial artificial chromosome in Escherichia coli [13], orby homologous recombination in permissive carp cell cul-tures [14], to obtain engineered live vaccines. Interestingly,the deletion of several genes encoding nonessential envelopeglycoproteins of KHV did not lead to sufficient attenuationof the virus in carp, while in the case of pORF25 it signifi-cantly reduced its protective efficacy against wild-type chal-lenge [15]. In contrast, the deletion of two nonessentialKHV genes of unknown function, ORF56 and ORF57, or ofORF57 alone, led to complete attenuation and efficient pro-tection against challenge [16, 17].
In many mammalian and avian herpesviruses homologuesof cellular enzymes involved in nucleotide metabolism suchas thymidine kinase (TK) and deoxyuridine triphosphatase(DUT) have been shown to be dispensable for virus replica-tion in vitro, but identified as important virulence factors invivo [18–22]. Attenuation was also demonstrated for a TKgene-deleted mutant of channel catfish virus, which is amember of the Alloherpesviridae [23]. TK (ORF55) andDUT (ORF123) gene-deleted KHV recombinants were alsoshown to be unaffected in cell culture and attenuated invivo. However, attenuation was incomplete, and single-deletion mutants still caused significant mortality in juvenilecarp [13, 14]. We therefore generated a TK and DUT genedouble-deletion mutant of a virulent KHV isolate fromIsrael (KHV-I) [2], which retained wild-type-like in vitroreplication [14]. However, the parental virus and its deriva-tives, like most field isolates of KHV, only reached titres ofless than 105 p.f.u. ml�1 in cell culture, and the stability ofthe infectious virus at 25
�C, as well as after freeze-thawing,
was very limited [14].
Since these properties would not permit economical vaccineproduction, we introduced the same TK and DUT genedeletions into the genome of a cell culture-adapted, but stillconsiderably virulent, KHV strain from Taiwan (KHV-T)[24]. The in vitro replication properties of the new single-and double-deletion mutants of KHV-T were analysed, andanimal experiments were performed to determine virulenceand protective efficacy of the KHV-T-derived viruses, aswell as of the KHV-I-derived double-deletion mutant incommon carp. Sera collected after vaccination and challengeinfection were investigated for KHV-specific antibodiesusing a recently developed KHV antibody enzyme-linkedimmunosorbent assay (ELISA) [25]. We further developed anew TK gene-specific quantitative real-time PCR (qPCR)that could be combined with the described ORF89-specific[26] and control qPCRs [27]. Using this triplex qPCR thesuitability of TK plus DUT gene-deleted KHV recombinantsas genetic DIVA vaccines was evaluated.
RESULTS AND DISCUSSION
Generation and genomic characterization of KHVTK and DUT gene mutants
We have previously generated and characterized TK
(ORF55) and DUT (ORF123) single- and double-gene dele-
tion mutants of one of the first described KHV isolates from
Israel (KHV-I) [14, 28]. In the present study, we introduced
identical deletions into the genome of the cell culture-
adapted strain KHV-T [24]. To this end, a non-neuronal
common carp brain (CCB) cell line [3] was cotransfected
with genomic KHV-T DNA and transfer plasmids contain-
ing PCR-amplified and subsequently mutagenized genome
fragments of the same virus strain (Fig. 1a–c). To facilitate
the selection of virus mutants resulting from homologous
recombination, a GFP reporter gene cassette was inserted
instead of the deleted viral sequences, and fluorescent foci
of cells infected with the transfection progenies were iso-
lated. To allow additional gene deletions using the same
reporter gene and to avoid possible adverse effects of the
transgene, DNA of the plaque-purified GFP-expressing
KHV mutants was prepared and used for cotransfections of
CCB cells together with TK and DUT gene deletion plas-
mids lacking foreign sequences (Fig. 1b, c), and non-
fluorescent progeny virus plaques were selected. In a similar
manner, TK and DUT rescuants were generated from the
GFP-expressing single-gene deletion mutants. To obtain the
TK and DUT double-deletion mutants of KHV-T and
KHV-I, the GFP cassette was consecutively inserted and
removed at both loci. All of the desired virus recombinants
could be purified to homogeneity after a few (�4) rounds of
plaque isolation. To verify the desired mutations of the TK
and/or DUT genes, the affected genome regions were ampli-
fied from viral DNA by PCR and sequenced using the
primer pairs KTK-PSF/PSR and KDUT-PSF/PSR (Table 1)
[14]. The obtained PCR products of all KHV mutants
exhibited the expected sizes (Fig. 1d, e) and sequences.
These studies confirmed that all of the respective deletion
mutants lacked codons 36 to 270 of the DUT gene (Fig. 1c),
encoding major parts of the conserved functional domain of
trimeric dUTPases (amino acids 30 to 119) [29]. The dele-
tion from the TK gene included codons 43 to 187 and, thus,
the predicted (https://prosite.expasy.org) ATP-binding site
(amino acids 9 to 16) was retained, but the highly conserved
C-terminal TK signature domain (amino acids 162 to 175)
was missing (Fig. 1b). Furthermore, after removal of the
GFP-cassette, the short retained 3¢ parts of both ORFs were
fused out of frame to the 5¢ parts. Therefore, no functional
thymidine kinase and/or dUTPase could be expressed by
any of the generated KHV mutants. The presence of the
desired mutations and the genetic homogeneity of the KHV
recombinants was further confirmed by restriction endonu-
clease digestions of genomic DNA and Southern blot
hybridization with TK and DUT gene-specific probes
(results not shown).
Schröder et al., Journal of General Virology
2
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
Genome sequence of KHV-TDDUT/TK
Next-generation sequencing of the genome of KHV-
TDDUT/TK confirmed that it is a derivative KHV-T, whose
complete genome sequence has been published recently
(GenBank accession # MG925491) [30]. Whereas the
terminal direct repeat sequences of both viruses proved tobe identical, few alterations or ambiguities were foundwithin the central unique region (Table 2). Two of themaffected the length of presumably noncoding homopoly-meric G stretches (positions 102598 and 124177), and sevenothers represented altered copy numbers of perfect or
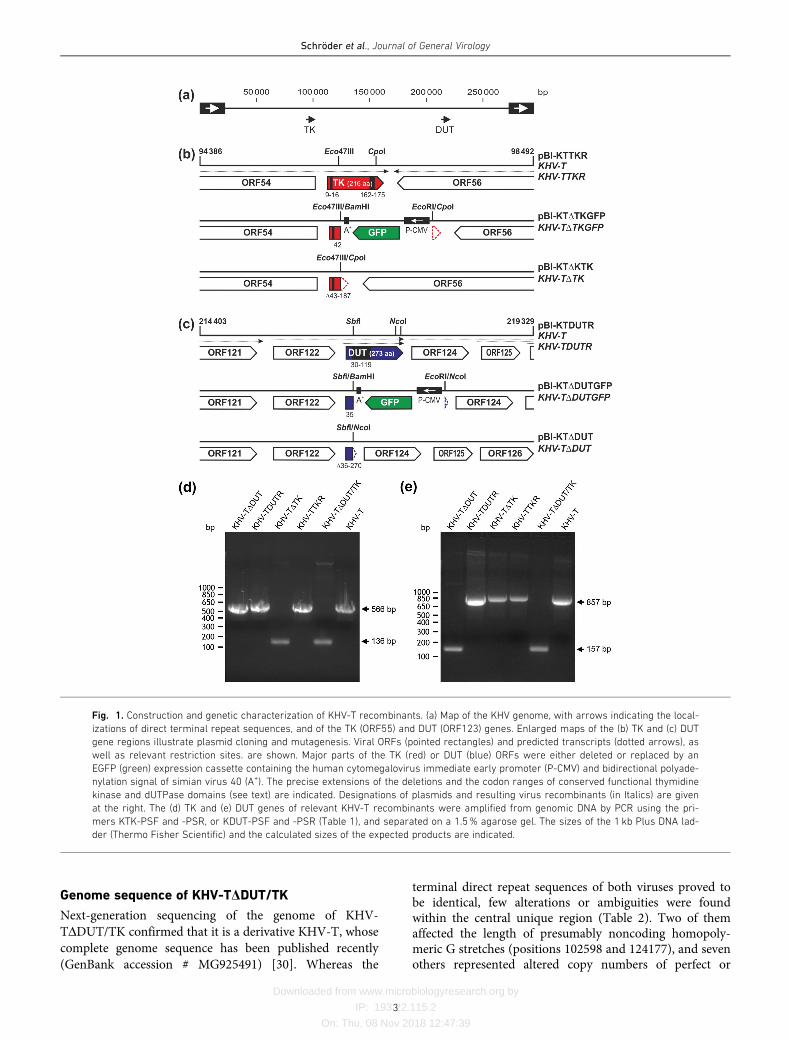
Fig. 1. Construction and genetic characterization of KHV-T recombinants. (a) Map of the KHV genome, with arrows indicating the local-
izations of direct terminal repeat sequences, and of the TK (ORF55) and DUT (ORF123) genes. Enlarged maps of the (b) TK and (c) DUT
gene regions illustrate plasmid cloning and mutagenesis. Viral ORFs (pointed rectangles) and predicted transcripts (dotted arrows), as
well as relevant restriction sites. are shown. Major parts of the TK (red) or DUT (blue) ORFs were either deleted or replaced by an
EGFP (green) expression cassette containing the human cytomegalovirus immediate early promoter (P-CMV) and bidirectional polyade-
nylation signal of simian virus 40 (A+). The precise extensions of the deletions and the codon ranges of conserved functional thymidine
kinase and dUTPase domains (see text) are indicated. Designations of plasmids and resulting virus recombinants (in Italics) are given
at the right. The (d) TK and (e) DUT genes of relevant KHV-T recombinants were amplified from genomic DNA by PCR using the pri-
mers KTK-PSF and -PSR, or KDUT-PSF and -PSR (Table 1), and separated on a 1.5% agarose gel. The sizes of the 1 kb Plus DNA lad-
der (Thermo Fisher Scientific) and the calculated sizes of the expected products are indicated.
Schröder et al., Journal of General Virology
3
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
imperfect tandem repetitions of short sequence elements
(Table 2). Three of these alterations were located within the
viral ORFs 45, 52 and 149, and therefore enlarged or short-
ened the deduced proteins. However, since the length of the
inserted or deleted repeat elements was always a multiple of
three, no frameshifts occurred. It has long been known that
variable numbers of tandem repeats (VNTRs) are present in
many herpesvirus genomes and they have also been found
by the comparison of different KHV strains and isolates [6,
30]. The analysis of individual sequence reads from the
KHV-TDDUT/TK genome strongly indicated that the used
template DNA prepared from a plaque-purified virus
recombinant already contained different variants of most of
these repetitive sequences. Thus, the suitability of such poly-
morphisms for the differentiation of KHV isolates might be
limited, although the predominant repeat copy numbers
appear to be stable over several in vitro or in vivo virus pas-
sages [31, 32]. In contrast to the ambiguities regarding tan-
dem repeats, two single base substitutions were
unambiguously found in KHV-TDDUT/TK when
Table 1. PCR and sequencing primers and TaqMan probes used in this study
Sequences refer to the sequences of the KHV TUMST1 genome [6] (GenBank accession no. AP008984) or of pEGFP-1 [43] (GenBank accession no.
U55761). Reverse strand primers (r) and modifications are indicated.
Name Sequence Genome position Modifications
KTKR-F 5¢-TAGCCGTACAGGGGACAC-3¢ 94 386–94 403 –
KTK-PSF 5¢-TGGAGCGTCTGTCCTACAGC-3¢ 96 073–96 092 –
KTKR-R 5¢-AGGTGATTTCGGTCATGAGC-3¢ 98 473–98 492 (r) –
KTK-PSR 5¢-ACAAGAACGAGGTGGAGCG-3¢ 96 620–96 638 (r) –
KDUTR-F 5¢-CCTACACCGCTCTGTTCG-3¢ 214 403–214 420 –
KDUT-PSF 5¢-AGTTTTCAATGTGGCAGGC-3¢ 216 565–216 583 –
KDUTR-R 5¢-AGAAACTGAGATCATCGCGG-3¢ 219 310–219 329 (r) –
KDUT-PSR 5¢-AATACAGCTACAATTGCGGG-3¢ 217 402–217 421 (r) –
KHV-86F 5¢-GACGCCGGAGACCTTGTG-3¢ 165 351–165 368 –
KHV-163R 5¢-CGGGTTCTTATTTTTGTCCTTGTT-3¢ 165 405–165 428 (r) –
KHV-109P 5¢-CTTCCTCTGCTCGGCGAGCACG-3¢ 165 374–165 395 (r) 5¢ : FAM, 3¢ : TAMRA
KHVTKQ-F 5¢-ACTTTATGCAGCAGCCCTTC-3¢ 96 340–96 359 –
KHVTKQ-R 5¢-CACTTCATGCACACCGCC-3¢ 96 410–96 427 (r) –
KHVTKQ-S 5¢-CCCATGGCGGACAAGCTGGACAAG-3¢ 96 381–96 404 5¢ : Cy 5, 3 : BHQ-2
EGFP15-F 5¢-GAGCAAGGGCGAGGAGC-3¢ 102–118 –
EGFP10-R 5¢-CTTGTACAGCTCGTCCATGC-3¢ 794–813 (r) –
EGFP-HEX 5¢-AGCACCCAGTCCGCCCTGAGCA-3¢ 703–724 5¢ : HEX, 3¢ : BHQ-1
Table 2. Sequence analysis KHV-TDDUT/TK compared to KHV-T
Nucleotide positions of alterations in KHV-TDDUT/TK refer to the sequence of the KHV-T genome (GenBank accession no. MG925491) [30]. The
deleted nucleotide rages are indicated, and the two flanking nucleotides are provided at insertion sites. The designations of affected ORFs, nucleotide
sequences of insertions (+) or deletions (D) and consequences for deduced amino acid sequences, including codon positions, are given. Targeted
mutations that are unique to KHV-TDDUT/TK are in bold.
Position in KHV-T ORF Alteration
37986–38 007 – D GTGTGTGTGTGTGTGTGTGTGT
75907–75 915 45 D AGGAGGTGG (DGGG at codon 336)
91247/91248 52 +AACCACCGAGGA (+TTEE at codon 158)
96099–96 532 55 D TK gene (from codon 43)
102598 – +G
124177 – +G
138261 72 A fi C (K fi N at codon 103)
177575–177594 – D AGAGAGAGAGAGAGAGAGAG
216478/216479 – +CTCAGCACCTTCAGCACCTTCAGCACCTTCAGCACCTTCAGCAC
216585–217288 123 D DUT gene (from codon 36)
256452/256453 149 +TGTCGTGCTTGGTGTGGTTGGGGTCGAAGTCGTACTTGG (+PSTTSTPTTPSTT at codon 513)
257991 – T fi C
270671–270685 – D CGACACAGACTACAG
Schröder et al., Journal of General Virology
4

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
compared to the published KHV-T sequence (positions1238261 and 257991), with one of them leading to an aminoacid substitution in the ORF72 product (Table 2). Remark-ably, the same bases that had been found in KHV-TDDUT/TK were also found in other KHV-T derivatives in our labo-ratory, and in the closely related Japanese isolate KHV-J orTUMST1 (GenBank accession no. AP008984) [6]. Thus, thebase substitutions obviously occurred during propagation ofKHV-T by Guo and coworkers. Nevertheless, the only sig-nificant differences between KHV-TDDUT/TK and thepublished sequence of KHV-T were the expected deletionsof ORFs 55 and 123, encoding TK or DUT, respectively(Table 2). Taken together, our sequence analyses did notprovide any evidence for unwanted mutations affecting thein vitro or in vivo phenotype of KHV-TDDUT/TK com-pared to KHV-T. Furthermore, we were able to confirmmost of the gene alterations in KHV-T acquired during cellculture adaptation, including the frameshift in ORF27,which is considered to be relevant for its lower virulencecompared to certain field isolates [30]. However, since sev-eral other KHV strains and isolates possess similar muta-tions, the precise reason for the exceptional replication ofKHV-T in cell culture remains to be elucidated.
In vitro replication properties of KHV TK and DUTgene mutants
To investigate the cell-to-cell spread of the KHV-T derivedmutants compared to the parental strain, CCB cells weregrown to monolayers in 24-well plates, infected in parallelwith serial dilutions of all viruses and incubated for 4 daysunder plaque assay conditions at 25
�C. The infected cells
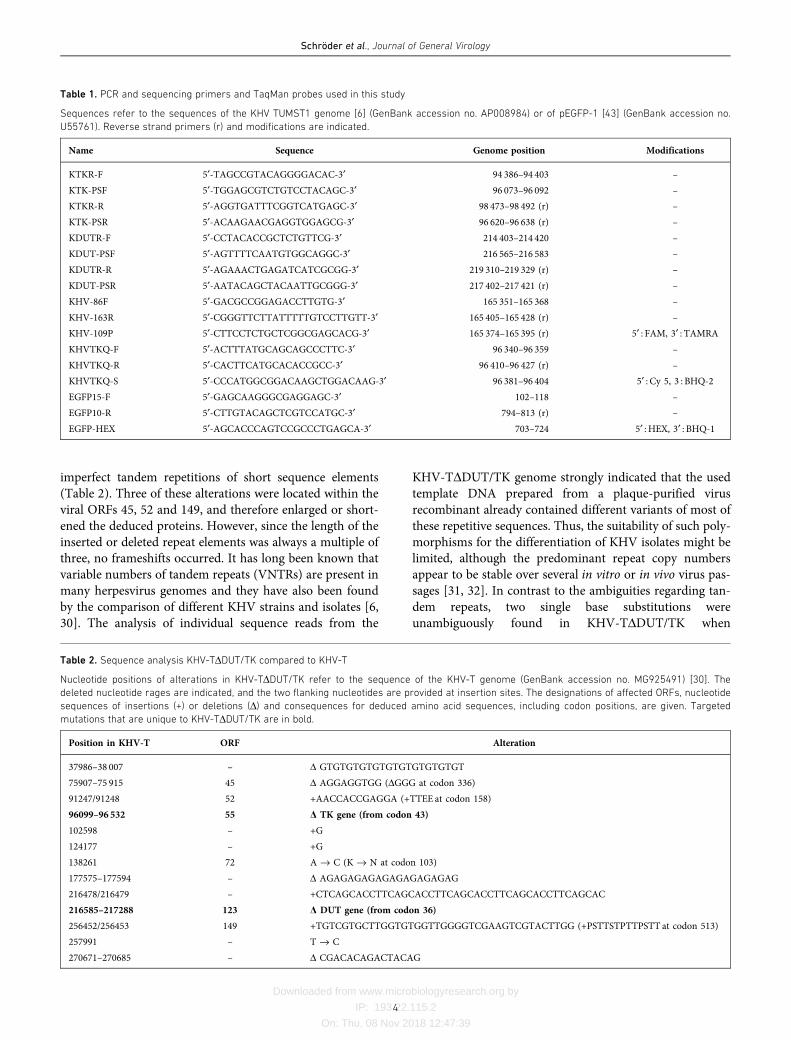
were identified by indirect immunofluorescence (IIF) testsand the plaque areas were determined microscopically,revealing that all TK and/or DUT deletion mutants, irre-spective of the presence or absence of the GFP reportergene, as well as the rescuants, formed plaques ranging fromapproximately 85 to 95% of the wild-type size (Fig. 2a). Theobserved minor differences were not statistically significant.
After the synchronized infection of CCB cells at an m.o.i. of0.1 the replication kinetics also showed wild-type-likecourses for all of the investigated KHV-T recombinants andresulted in maximum titres of between 5�106 and 2�107 p.f.u. ml�1 in total cell lysates (Fig. 2b). Thus, in line with pre-vious in vitro investigations of TK and DUT gene-deletedKHV mutants [13, 14], our present studies confirmed thatboth genes are fully dispensable for virus replication in tis-sue culture. The newly prepared GFPless TK and DUT genedouble-deletion mutant of KHV-I also exhibited similargrowth properties to the corresponding wild-type virus andthe described deletion mutants (results not shown). How-ever, the maximum titres of all KHV-I-derived viruses onCCB cells were only approximately 105 p.f.u. ml�1, anddecreased rapidly if incubation at 25
�C was further contin-
ued [14]. The molecular reasons for these differences in rep-lication rates and virion stability in vitro remain to beelucidated. However, for economic reasons, KHV-T derived
recombinants are apparently more suitable as putative livevaccines than derivatives of KHV-I.
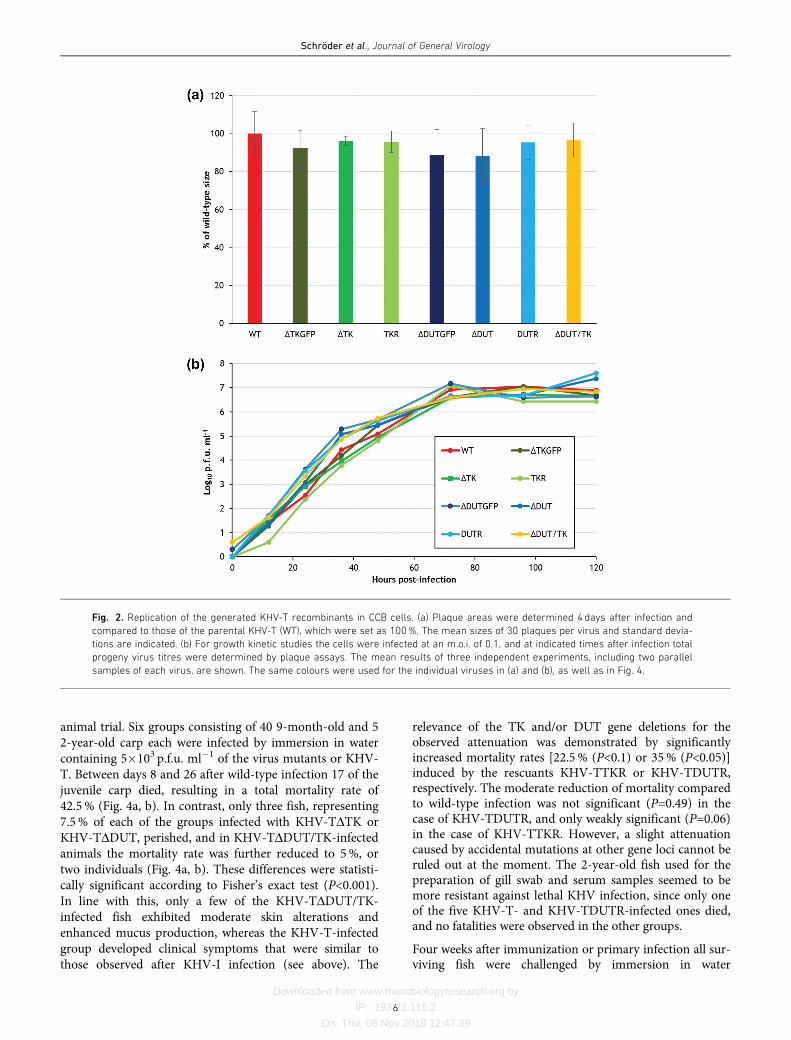
Virulence and protective efficacy of TK and DUTgene deleted KHV mutants
Previous animal experiments have shown that single dele-tions of the TK or DUT genes of KHV lead to significantbut insufficient attenuation [13, 14]. Since both genesseemed to contribute to the virulence of KHV-I, we tested acorresponding double-deletion mutant of KHV-I in vivo.To this end, groups of 48 1-year-old specific pathogen-freecarp were infected either by immersion in water containing1.5�102 plaque-forming units (p.f.u.) ml�1 or by intraperi-toneal (i.p.) administration of 2�103 p.f.u. of either KHV-IDTK/DUT or KHV-I per animal. Whereas no clinical signswere observed after immersion, few fish developed moder-ate symptoms (mainly skin lesions) after the injection ofKHV-IDTK/DUT, and three of them (6%) died (Fig. 3a). Incontrast, control animals infected with similar doses of viru-lent KHV-I developed severe clinical signs, including gillnecrosis, haemorrhages at the fins and neurological disor-ders, and 48 or 60% of the carp succumbed to infection(Fig. 3a). As expected, these differences between the mortal-ity rates caused KHV-IDTK/DUT and wild-type virusproved to be statistically significant according to Fisher’sexact test (P<0.001). After 50 days, all carp that had sur-vived primary infection, as well as the mock-infected con-trol animals, were challenged by immersion in watercontaining 1.5�102 p.f.u. ml�1 KHV-I. Whereas the naïvecarp groups developed severe disease leading to mortalityrates of 50%, the wild-type-primed fish were widely pro-tected, and only one, representing 4 or 5.5% of each group,died after challenge, possibly still as a consequence of theprimary infection with KHV-I (Fig. 3b). The resultsobtained with the KHV-IDTK/DUT-vaccinated animalswere more inhomogeneous. Whereas the i.p. immunizedcarp were completely protected, the group immunized byimmersion exhibited nearly 15% mortality. Although theprotection level of this group compared to non-vaccinatedcontrol animals was still significant (P<0.001), it was alsosignificantly (P<0.05) lower than that of the i.p.-immunizedgroup. Thus, the virus dose used for vaccination by immer-sion might have been too low to achieve a sufficientimmune response in all animals. However, in view of thelow titres of KHV-I-derived viruses in cell culture (seeabove), production of the virus for the administration ofhigher live vaccine doses to huge carp populations byimmersion would be prohibitively expensive. On the otherhand, individual administration of the vaccines by injectionrequires less virus, but is more laborious.
Therefore, we tested the novel TK and DUT gene double-deletion mutant based on the cell culture-adapted strainKHV-T (KHV-TDDUT/TK). To verify the previouslydescribed effects of single deletions of TK or DUT on thevirulence of KHV [13, 14] corresponding deletion mutants(KHV-TDTK, KHV-TDDUT) and rescuants (KHV-TTKR,KHV-TDUTR) of KHV-T were also included in the second
Schröder et al., Journal of General Virology
5
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
animal trial. Six groups consisting of 40 9-month-old and 52-year-old carp each were infected by immersion in watercontaining 5�103 p.f.u. ml�1 of the virus mutants or KHV-T. Between days 8 and 26 after wild-type infection 17 of thejuvenile carp died, resulting in a total mortality rate of42.5% (Fig. 4a, b). In contrast, only three fish, representing7.5% of each of the groups infected with KHV-TDTK orKHV-TDDUT, perished, and in KHV-TDDUT/TK-infectedanimals the mortality rate was further reduced to 5%, ortwo individuals (Fig. 4a, b). These differences were statisti-cally significant according to Fisher’s exact test (P<0.001).In line with this, only a few of the KHV-TDDUT/TK-infected fish exhibited moderate skin alterations andenhanced mucus production, whereas the KHV-T-infectedgroup developed clinical symptoms that were similar tothose observed after KHV-I infection (see above). The
relevance of the TK and/or DUT gene deletions for theobserved attenuation was demonstrated by significantlyincreased mortality rates [22.5% (P<0.1) or 35% (P<0.05)]induced by the rescuants KHV-TTKR or KHV-TDUTR,respectively. The moderate reduction of mortality comparedto wild-type infection was not significant (P=0.49) in thecase of KHV-TDUTR, and only weakly significant (P=0.06)in the case of KHV-TTKR. However, a slight attenuationcaused by accidental mutations at other gene loci cannot beruled out at the moment. The 2-year-old fish used for thepreparation of gill swab and serum samples seemed to bemore resistant against lethal KHV infection, since only oneof the five KHV-T- and KHV-TDUTR-infected ones died,and no fatalities were observed in the other groups.
Four weeks after immunization or primary infection all sur-viving fish were challenged by immersion in water
Fig. 2. Replication of the generated KHV-T recombinants in CCB cells. (a) Plaque areas were determined 4 days after infection and
compared to those of the parental KHV-T (WT), which were set as 100%. The mean sizes of 30 plaques per virus and standard devia-
tions are indicated. (b) For growth kinetic studies the cells were infected at an m.o.i. of 0.1, and at indicated times after infection total
progeny virus titres were determined by plaque assays. The mean results of three independent experiments, including two parallel
samples of each virus, are shown. The same colours were used for the individual viruses in (a) and (b), as well as in Fig. 4.
Schröder et al., Journal of General Virology
6
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
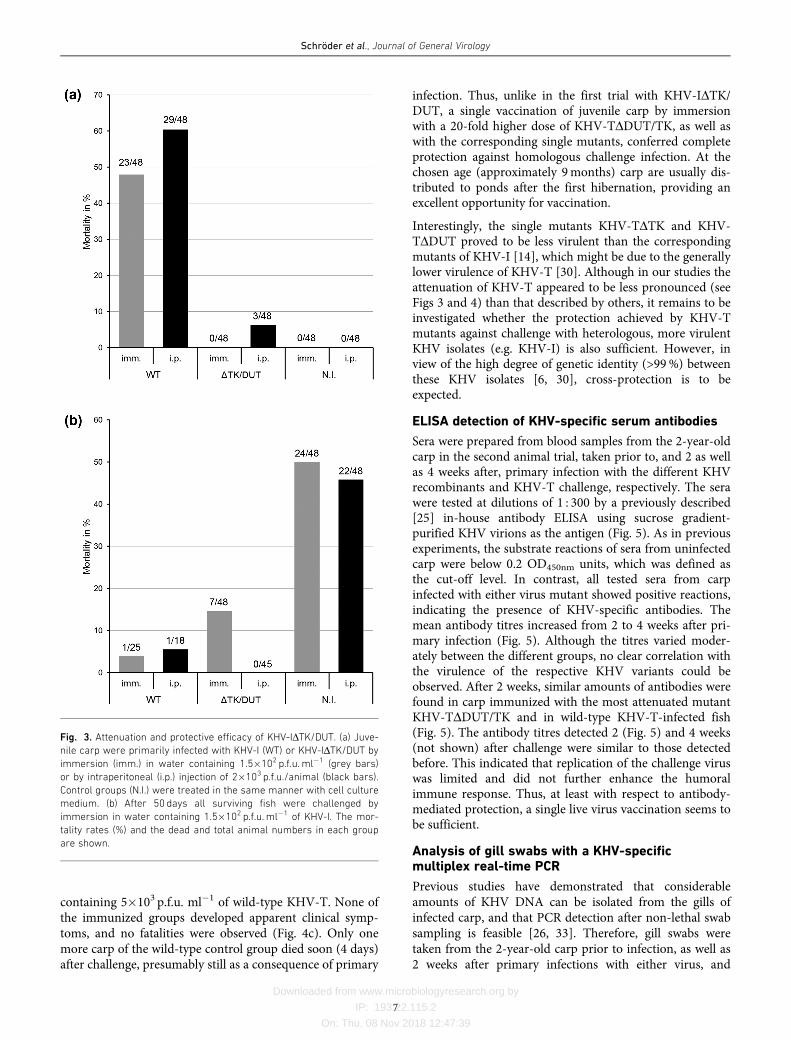
containing 5�103 p.f.u. ml�1 of wild-type KHV-T. None ofthe immunized groups developed apparent clinical symp-toms, and no fatalities were observed (Fig. 4c). Only onemore carp of the wild-type control group died soon (4 days)after challenge, presumably still as a consequence of primary
infection. Thus, unlike in the first trial with KHV-IDTK/DUT, a single vaccination of juvenile carp by immersionwith a 20-fold higher dose of KHV-TDDUT/TK, as well aswith the corresponding single mutants, conferred completeprotection against homologous challenge infection. At thechosen age (approximately 9months) carp are usually dis-tributed to ponds after the first hibernation, providing anexcellent opportunity for vaccination.
Interestingly, the single mutants KHV-TDTK and KHV-TDDUT proved to be less virulent than the correspondingmutants of KHV-I [14], which might be due to the generallylower virulence of KHV-T [30]. Although in our studies theattenuation of KHV-T appeared to be less pronounced (seeFigs 3 and 4) than that described by others, it remains to beinvestigated whether the protection achieved by KHV-Tmutants against challenge with heterologous, more virulentKHV isolates (e.g. KHV-I) is also sufficient. However, inview of the high degree of genetic identity (>99%) betweenthese KHV isolates [6, 30], cross-protection is to beexpected.
ELISA detection of KHV-specific serum antibodies
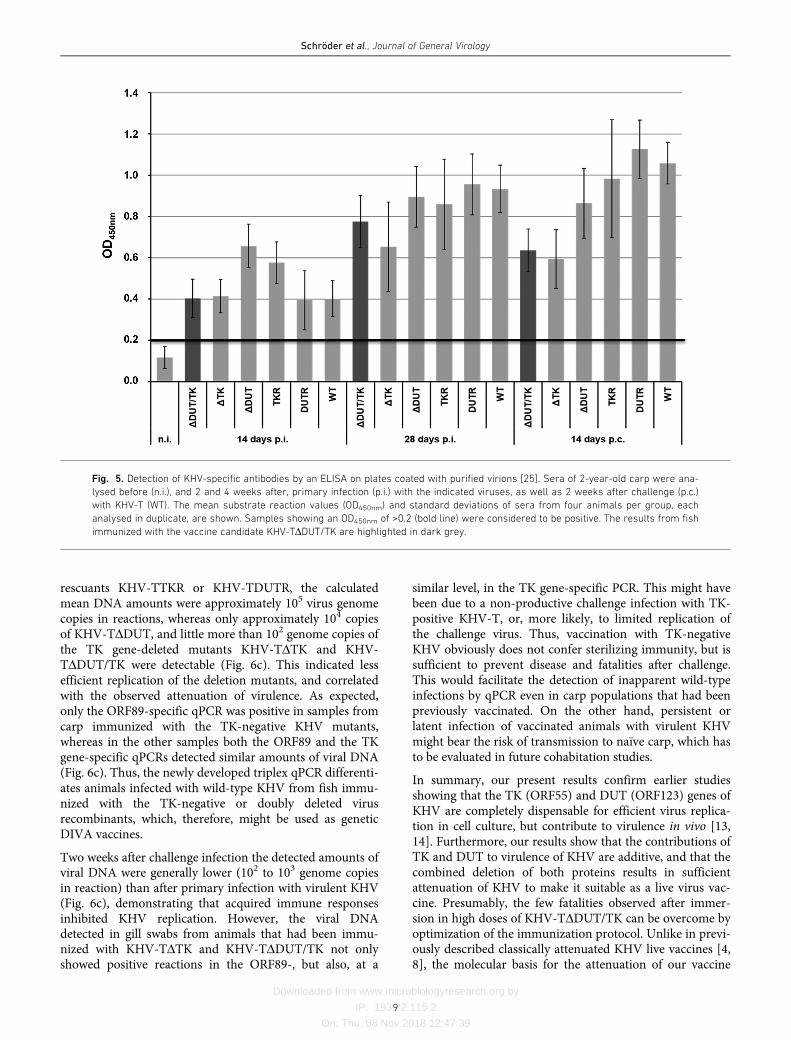
Sera were prepared from blood samples from the 2-year-oldcarp in the second animal trial, taken prior to, and 2 as wellas 4 weeks after, primary infection with the different KHVrecombinants and KHV-T challenge, respectively. The serawere tested at dilutions of 1 : 300 by a previously described[25] in-house antibody ELISA using sucrose gradient-purified KHV virions as the antigen (Fig. 5). As in previousexperiments, the substrate reactions of sera from uninfectedcarp were below 0.2 OD450nm units, which was defined asthe cut-off level. In contrast, all tested sera from carpinfected with either virus mutant showed positive reactions,indicating the presence of KHV-specific antibodies. Themean antibody titres increased from 2 to 4 weeks after pri-mary infection (Fig. 5). Although the titres varied moder-ately between the different groups, no clear correlation withthe virulence of the respective KHV variants could beobserved. After 2 weeks, similar amounts of antibodies werefound in carp immunized with the most attenuated mutantKHV-TDDUT/TK and in wild-type KHV-T-infected fish(Fig. 5). The antibody titres detected 2 (Fig. 5) and 4 weeks(not shown) after challenge were similar to those detectedbefore. This indicated that replication of the challenge viruswas limited and did not further enhance the humoralimmune response. Thus, at least with respect to antibody-mediated protection, a single live virus vaccination seems tobe sufficient.
Analysis of gill swabs with a KHV-specificmultiplex real-time PCR
Previous studies have demonstrated that considerableamounts of KHV DNA can be isolated from the gills ofinfected carp, and that PCR detection after non-lethal swabsampling is feasible [26, 33]. Therefore, gill swabs weretaken from the 2-year-old carp prior to infection, as well as2 weeks after primary infections with either virus, and
Fig. 3. Attenuation and protective efficacy of KHV-IDTK/DUT. (a) Juve-
nile carp were primarily infected with KHV-I (WT) or KHV-IDTK/DUT by
immersion (imm.) in water containing 1.5�102 p.f.u.ml�1 (grey bars)
or by intraperitoneal (i.p.) injection of 2�103 p.f.u./animal (black bars).
Control groups (N.I.) were treated in the same manner with cell culture
medium. (b) After 50 days all surviving fish were challenged by
immersion in water containing 1.5�102 p.f.u.ml�1 of KHV-I. The mor-
tality rates (%) and the dead and total animal numbers in each group
are shown.
Schröder et al., Journal of General Virology
7
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
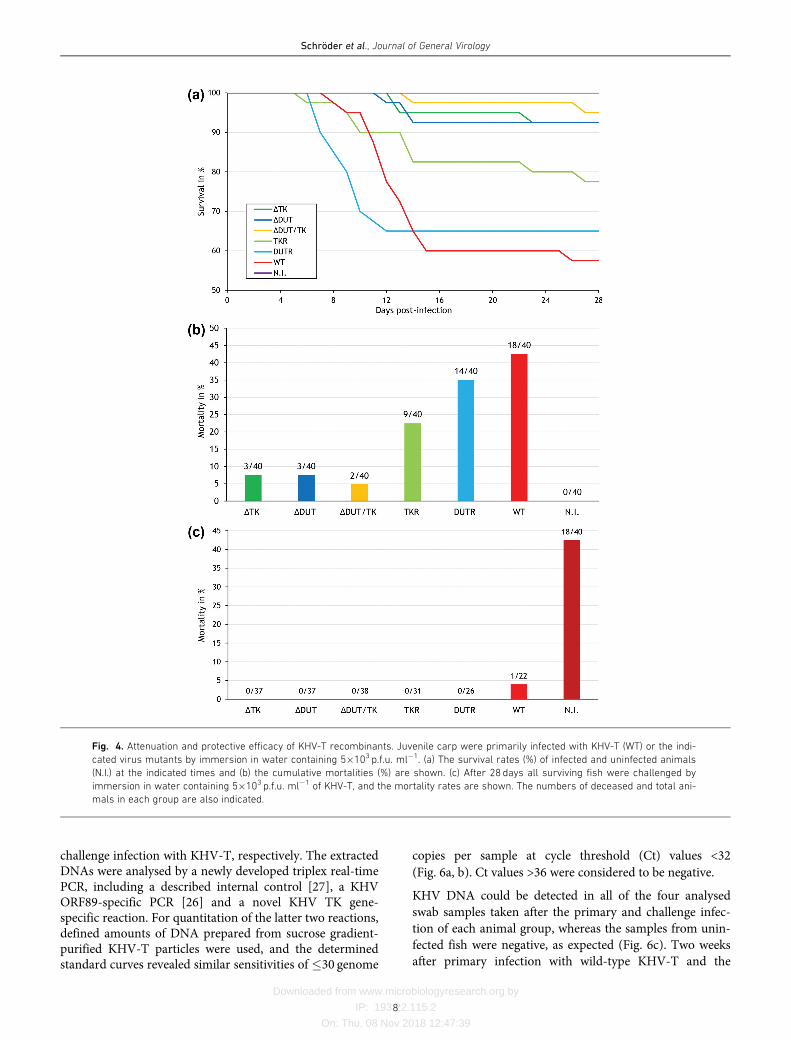
challenge infection with KHV-T, respectively. The extractedDNAs were analysed by a newly developed triplex real-timePCR, including a described internal control [27], a KHVORF89-specific PCR [26] and a novel KHV TK gene-specific reaction. For quantitation of the latter two reactions,defined amounts of DNA prepared from sucrose gradient-purified KHV-T particles were used, and the determinedstandard curves revealed similar sensitivities of �30 genome
copies per sample at cycle threshold (Ct) values <32
(Fig. 6a, b). Ct values >36 were considered to be negative.
KHV DNA could be detected in all of the four analysed
swab samples taken after the primary and challenge infec-
tion of each animal group, whereas the samples from unin-
fected fish were negative, as expected (Fig. 6c). Two weeks
after primary infection with wild-type KHV-T and the
Fig. 4. Attenuation and protective efficacy of KHV-T recombinants. Juvenile carp were primarily infected with KHV-T (WT) or the indi-
cated virus mutants by immersion in water containing 5�103 p.f.u. ml�1. (a) The survival rates (%) of infected and uninfected animals
(N.I.) at the indicated times and (b) the cumulative mortalities (%) are shown. (c) After 28 days all surviving fish were challenged by
immersion in water containing 5�103 p.f.u. ml�1 of KHV-T, and the mortality rates are shown. The numbers of deceased and total ani-
mals in each group are also indicated.
Schröder et al., Journal of General Virology
8
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
rescuants KHV-TTKR or KHV-TDUTR, the calculatedmean DNA amounts were approximately 105 virus genomecopies in reactions, whereas only approximately 104 copiesof KHV-TDDUT, and little more than 102 genome copies ofthe TK gene-deleted mutants KHV-TDTK and KHV-TDDUT/TK were detectable (Fig. 6c). This indicated lessefficient replication of the deletion mutants, and correlatedwith the observed attenuation of virulence. As expected,only the ORF89-specific qPCR was positive in samples fromcarp immunized with the TK-negative KHV mutants,whereas in the other samples both the ORF89 and the TKgene-specific qPCRs detected similar amounts of viral DNA(Fig. 6c). Thus, the newly developed triplex qPCR differenti-ates animals infected with wild-type KHV from fish immu-nized with the TK-negative or doubly deleted virusrecombinants, which, therefore, might be used as geneticDIVA vaccines.
Two weeks after challenge infection the detected amounts ofviral DNA were generally lower (102 to 103 genome copiesin reaction) than after primary infection with virulent KHV(Fig. 6c), demonstrating that acquired immune responsesinhibited KHV replication. However, the viral DNAdetected in gill swabs from animals that had been immu-nized with KHV-TDTK and KHV-TDDUT/TK not onlyshowed positive reactions in the ORF89-, but also, at a
similar level, in the TK gene-specific PCR. This might havebeen due to a non-productive challenge infection with TK-positive KHV-T, or, more likely, to limited replication ofthe challenge virus. Thus, vaccination with TK-negativeKHV obviously does not confer sterilizing immunity, but issufficient to prevent disease and fatalities after challenge.This would facilitate the detection of inapparent wild-typeinfections by qPCR even in carp populations that had beenpreviously vaccinated. On the other hand, persistent orlatent infection of vaccinated animals with virulent KHVmight bear the risk of transmission to naïve carp, which hasto be evaluated in future cohabitation studies.
In summary, our present results confirm earlier studiesshowing that the TK (ORF55) and DUT (ORF123) genes ofKHV are completely dispensable for efficient virus replica-tion in cell culture, but contribute to virulence in vivo [13,14]. Furthermore, our results show that the contributions ofTK and DUT to virulence of KHV are additive, and that thecombined deletion of both proteins results in sufficientattenuation of KHV to make it suitable as a live virus vac-cine. Presumably, the few fatalities observed after immer-sion in high doses of KHV-TDDUT/TK can be overcome byoptimization of the immunization protocol. Unlike in previ-ously described classically attenuated KHV live vaccines [4,8], the molecular basis for the attenuation of our vaccine
Fig. 5. Detection of KHV-specific antibodies by an ELISA on plates coated with purified virions [25]. Sera of 2-year-old carp were ana-
lysed before (n.i.), and 2 and 4 weeks after, primary infection (p.i.) with the indicated viruses, as well as 2 weeks after challenge (p.c.)
with KHV-T (WT). The mean substrate reaction values (OD450nm) and standard deviations of sera from four animals per group, each
analysed in duplicate, are shown. Samples showing an OD450nm of >0.2 (bold line) were considered to be positive. The results from fish
immunized with the vaccine candidate KHV-TDDUT/TK are highlighted in dark grey.
Schröder et al., Journal of General Virology
9
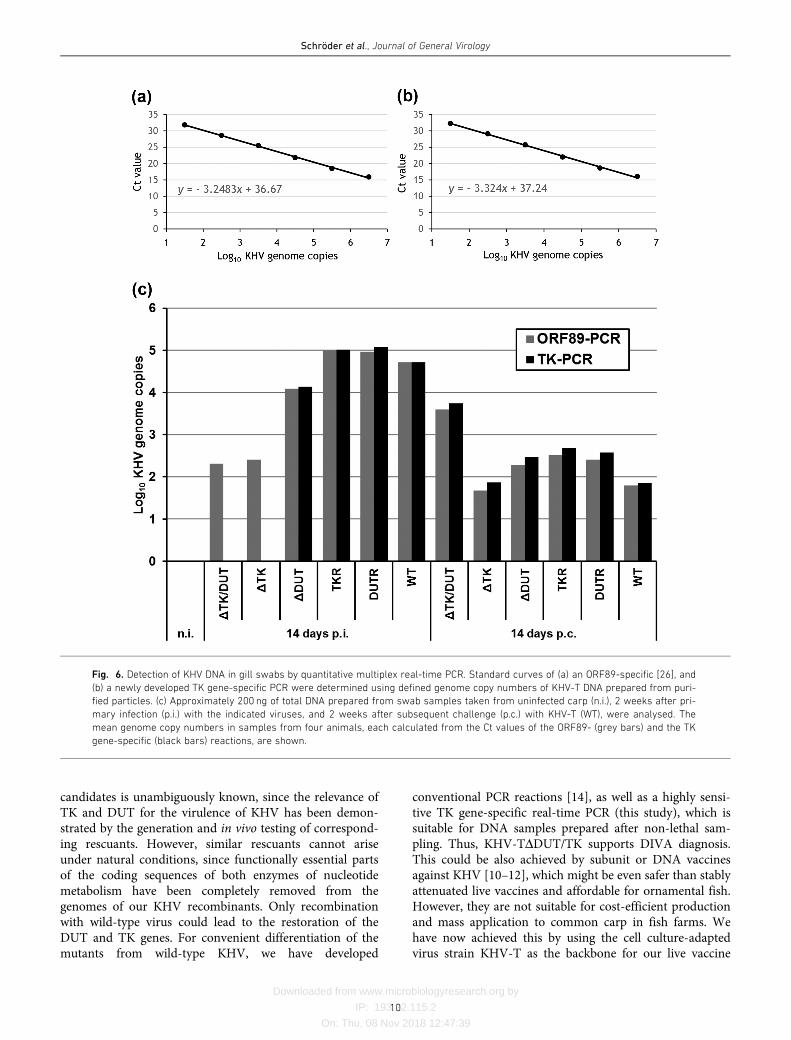
Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
candidates is unambiguously known, since the relevance ofTK and DUT for the virulence of KHV has been demon-strated by the generation and in vivo testing of correspond-ing rescuants. However, similar rescuants cannot ariseunder natural conditions, since functionally essential partsof the coding sequences of both enzymes of nucleotidemetabolism have been completely removed from thegenomes of our KHV recombinants. Only recombinationwith wild-type virus could lead to the restoration of theDUT and TK genes. For convenient differentiation of themutants from wild-type KHV, we have developed
conventional PCR reactions [14], as well as a highly sensi-tive TK gene-specific real-time PCR (this study), which issuitable for DNA samples prepared after non-lethal sam-pling. Thus, KHV-TDDUT/TK supports DIVA diagnosis.This could be also achieved by subunit or DNA vaccinesagainst KHV [10–12], which might be even safer than stablyattenuated live vaccines and affordable for ornamental fish.However, they are not suitable for cost-efficient productionand mass application to common carp in fish farms. Wehave now achieved this by using the cell culture-adaptedvirus strain KHV-T as the backbone for our live vaccine
Fig. 6. Detection of KHV DNA in gill swabs by quantitative multiplex real-time PCR. Standard curves of (a) an ORF89-specific [26], and
(b) a newly developed TK gene-specific PCR were determined using defined genome copy numbers of KHV-T DNA prepared from puri-
fied particles. (c) Approximately 200 ng of total DNA prepared from swab samples taken from uninfected carp (n.i.), 2 weeks after pri-
mary infection (p.i.) with the indicated viruses, and 2 weeks after subsequent challenge (p.c.) with KHV-T (WT), were analysed. The
mean genome copy numbers in samples from four animals, each calculated from the Ct values of the ORF89- (grey bars) and the TK
gene-specific (black bars) reactions, are shown.
Schröder et al., Journal of General Virology
10

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
candidates, which grow to significantly higher titres thanother KHV isolates in different cell culture systems [24].Thus, the production of sufficient amounts of e.g. KHV-TDDUT/TK for vaccination in the field of juvenile carp byimmersion should be no problem. Recently, other nonessen-tial KHV genes contributing to virulence have been identi-fied [15–17]. Although the attenuation achieved by thesingle deletion of several genes encoding highly immuno-genic viral envelope glycoproteins [34, 35] was not sufficient[15], it is conceivable that the additional deletion of thesegenes from KHV-TDDUT/TK would abolish its minorresidual virulence, and make it suitable as a serologicalDIVA vaccine by the use of available monoclonal antibodies[34, 36].
METHODS
Viruses and cells
KHV-I [2], KHV-T [24] and their derivatives were propa-gated in CCB cells [3] at 25
�C in minimum essential
medium (MEM) supplemented with 10% foetal bovineserum (FBS). After the appearance of cytopathogenic effects,the infected cultures were lysed by freeze-thawing andstored in aliquots at �80
�C. For plaque assays, CCB cell
monolayers were incubated with serial virus dilutions for2 h. Then, the inoculum was replaced by MEM containing5% FBS and 6 g of methylcellulose per litre, and incubationat 25
�C was continued for 3 to 7 days.
Construction of deletion plasmids and virusrecombinants
Genomic KHV-T DNA was prepared from infected celllysates as described previously [37]. For the generation oftransfer plasmids the TK (ORF55) and DUT (ORF123)gene regions were amplified from genomic KHV-T DNA byPCR using Pfx DNA polymerase (Thermo Fisher Scientific)and the primers KTKR-F and KTKR-R or KDUTR-F andKDUTR-R, respectively (Table 1). The obtained 4107 and4927 bp PCR products were treated with Klenow polymer-ase and T4 polynucleotide kinase, and inserted into theSmaI-digested vector pBluescript SK(�) (Agilent Technolo-gies). For inactivation of the TK gene (ORF55, 217 codons),codons 43 to 187 were deleted by CpoI/Eco47III digestion ofpBl-KTTKR (Fig. 1b). Subsequently, the plasmid was eitherdirectly religated, or modified by the insertion of an expres-sion cassette for enhanced green fluorescent protein (GFP),which had been isolated as an 1607 bp BamHI/EcoRI frag-ment from the plasmid pBl-GFP [38], resulting in pBl-KTDTK and pBl-KTDTKGFP, respectively (Fig. 1b). Toremove an unwanted NcoI restriction site, the insert of pBl-KTDUTR (Fig. 1c) was shortened by 133 nucleotides byHindIII/BspEI double digestion and religation. Subse-quently, the DUT gene (ORF123, 274 codons) was deletedfrom codon 36 to 270 by double digestion with NcoI andSbfI, and deletion plasmids with and without reporter geneinsertion (pBl-KTDDUTGFP, pBl-KTDDUT) were gener-ated as above (Fig. 1c). In all cloning experiments, non-
compatible ends were blunted by Klenow treatment prior toligation.
CCB cells were cotransfected (K2 Transfection System,Biontex) with genomic KHV-T DNA and the deletion plas-mids pBl-KTDTKGFP or pBl-KTDDUTGFP, and homolo-gous recombination led to the GFP-expressing virusmutants KHV-TDTKGFP or KHV-TDDUTGFP, whichwere plaque-purified to homogeneity. Genomic DNA fromthese mutants was then used for cotransfection withreporter geneless deletion (pBl-KTDTK, pBl-KTDDUT) orrescue plasmids (pBl-KTTKR, pBl-KTDUTR), resulting inthe isolation of KHV-TDTK, KHV-TDDUT, KHV-TTKRand KHV-TDUTR from non-fluorescent virus plaques(Fig. 1). To obtain the double-mutant KHV-TDDUT/TK,KHV-TDDUT was subsequently mutated as above with theplasmids KHV-TDTKGFP and KHV-TDTK. From thedescribed KHV-I mutant KHV-DTKDUTG [14] the GFPinsertion at the DUT gene locus was removed by mutagene-sis with a transfer plasmid derived from pBl-KB5836NB[14] by deletion of the above mentioned NcoI/SbfI fragment,resulting in KHV-IDTK/DUT. Genomic DNA of all gener-ated KHV recombinants was prepared, and the presence ofthe desired mutations was confirmed by restriction analysesand Southern blot hybridization (not shown), as well as byPCR amplification and sequencing of the mutated TK andDUT gene regions with the primers KTK-PSF, KTK-PSR,KDUT-PSF and KDUT-PSR (Table 1) as described previ-ously [14].
In vitro replication studies
For the comparative analysis of virus spread, CCB cellsgrown in 24-well plates were infected with serial dilutions ofKHV-T, KHV-TDDUTGFP, KHV-TDTKGFP, KHV-TDDUT/TKGFP, KHV-TDDUT, KHV-TDTK, KHV-TDDUT/TK, KHV-TDUTR, or KHV-TTKR and incubatedunder semisolid medium for 96 h at 25
�C. Then the cells
were washed with phosphate-buffered saline (PBS) andfixed with 50% methanol/50% acetone for 30min at�20
�C. The infected cells were visualized by IIF reactions
of a monospecific rabbit antiserum raised against the viralenvelope protein pORF81 as described previously [39].Areas of 30 plaques per virus were measured by fluores-cence microscopy (Eclipse Ti-S with software NIS-elements,version 4, Nikon), and the average sizes as well as standarddeviations were calculated as percentages of the mean pla-que areas induced by KHV-T.
For the investigation of replication kinetics, CCB cellsgrown in 24-well plates were infected with the same virusmutants at an m.o.i. of 0.1 at 4
�C to permit virus adsorption
and, after 1 h, shifted to 25�C for penetration. After an addi-
tional 2 h non-penetrated virus was inactivated by treatmentwith citric acid [40], and incubation at 25
�C was continued.
At different times after the temperature shift the infectedcell cultures were frozen at �80
�C. For the determination
of progeny virus titres the cell lysates were thawed at 25�C
and plaque assays followed by IIF tests were performed asabove. The mean titres of two parallel wells per virus in
Schröder et al., Journal of General Virology
11

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
three independent experiments were calculated. Student’s t-test was applied to evaluate the statistical significance of dif-ferences between the virus titres and plaque sizes of theKHV mutants.
Animal trials
Prior to the animal experiments, the used carp were con-firmed to be free of cyprinid herpesviruses (CyHV-1, -2 and-3), carp edema virus (CEV) and the causative agent ofspring viraemia of carp (SVCV) by PCR or RT-PCR analysisof swab and blood samples. Furthermore, the animals werenegatively checked for ectoparasites.
To test the in vivo attenuation and protective efficacy ofKHV-IDTK/DUT, 2 groups of 48 1-year-old (approximately10 g/6 cm) specific pathogen-free carp (obtained from S.H.Leenstra, Wageningen, The Netherlands) were infectedeither by immersion in a bucket with 10 l water containing1.5�106 p.f.u. of virus for 2 h, or by i.p. administration of2�103 p.f.u. of virus per animal. Control groups wereinfected with the same doses of wild-type KHV-I, or mock-infected with cell culture medium. The six groups were keptin separate tanks at a constant temperature of 22
�C and
observed daily for clinical symptoms. Fifty days after pri-mary or mock infection, all surviving fish were challengedby immersion in 10 l water containing 1.5�106 p.f.u. KHV-I, returned to their aquaria and observed for an additional61 days.
In a second trial, 7 groups consisting of 40 9-month-old(approximately 8 g/5–7 cm) and 5 2-year-old (approxi-mately 200 g/20–23 cm) carp from a commercial fish farmin Germany (Fischzucht Traßdorf 1967 e.V., Traßdorf) eachwere kept in individual aquaria at a constant temperature of20
�C. To avoid cannibalism, the small fish were separated
from the larger ones by permeable grids. The carp wereinfected by immersion for 1 h in 5 l water containing2.5�107 p.f.u. of either KHV-T, KHV-TDDUT, KHV-TDTK, KHV-TDDUT/TK, KHV-TDUTR, KHV-TTKR orcell culture medium. After 28 days all carp that had survivedprimary KHV infection were challenged by immersion for1 h in 5 l water containing 2.5�107 p.f.u. of KHV-T,returned to their aquaria and observed for another 28 days.Two and 4 weeks after primary, as well as after challengeinfection, blood samples and gill swabs were taken from theolder carp, whereas only the young ones were consideredfor the evaluation of mortality rates. The statistical signifi-cance of the differences was determined using a two-sidedFisher’s exact test [41].
Enzyme-linked immunosorbent assay (ELISA)
Blood samples from infected and uninfected carp werestored overnight at 4
�C and centrifuged at 600 g for 15min,
and sera were taken from the supernatant. Nunc-Immu-noTM MircoWellTM PolySorpTM F96 plates (Thermo FisherScientific) were coated with sucrose gradient-purified KHVparticles, blocked and incubated with the 1 : 300 diluted serain two replicas, an anti-carp IgM monoclonal antibody(Aquatic diagnostics) and horseradish peroxidase-
conjugated anti-mouse IgG (Jackson ImmunoResearch) asdescribed previously [25]. Substrate reactions were mea-sured in an Infinite F200 microplate reader (Tecan), andOD450nm values >0.2 were considered to be positive.
Quantitative real time PCR (qPCR)
Total DNA from the gill swabs of infected and control carpwas prepared using the QIAamp DNA Mini kit (Qiagen).To this end, the Q-tips used for sampling were incubatedfor �1 h at 56
�C in 180 µl buffer ATL until the samples
were completely lysed and further processed as recom-mended by the manufacturer. The DNA was finally elutedin 50 µl of water. The 25 µl TaqMan triplex qPCR reactionmixes using the QuantiTect Multiplex PCR NoROX kit(Qiagen) contained approximately 200 ng of swab DNAsample, 2�103 copies of plasmid pEGFP-1 (Clontech) as aninternal control, a control-specific primer/probe set (10 µMEGFP15-F, 10 µM EGFP10-R and 1.5 µM EGFP-HEX) [27],a KHV ORF89-specific primer/probe set (10 µM KHV-86F,10 µM KHV-163R and 1.25 µM KHV-109P) [26] and anewly designed KHV TK gene (ORF 55)-specific set, con-sisting of 10 µM KHVTKQ-F, 10 µM KHVTKQ-R and 2 µMKHVTKQ-P (Table 1). For quantitation, reaction mixescontaining 200 ng swab DNA sample from an uninfectedfish and 1 fg (3.14 genome copies) to 1 ng (3.14�106
genome copies) of DNA from sucrose gradient-purified[39] KHV-T particles were used. The approximate genomecopy number per µg virion DNA was calculated (http://cels.uri.edu/gsc/cndna.html) based on a randomly composeddouble-stranded 295 kbp molecule. Reactions were per-formed in an AriaMx Real-Time PCR System (AgilentTechnologies) for 15min at 95
�C, followed by 40 cycles of
1min 95�C, 1min 95
�C, 30 s 60
�C and 30 s 72
�C, and the
KHV genome copy numbers in the samples were calculatedfrom Ct values based on the standard curve.
Sequence analyses
The inserts of transfer plasmids and the modified genomeregions of virus recombinants were analysed by sequencingusing the BigDye Terminator v1.1 cycle sequencing kit anda 3130 Genetic Analyzer (Applied Biosystems). The resultswere evaluated with the Geneious software package in ver-sion 10.2.3 (Biomatters). Furthermore, genomic DNA ofKHV-TDDUT/TK was prepared [37] from infected CCBcells and used to prepare shotgun sequencing libraries forsequencing with the Ion Torrent PGM as described previ-ously [42] with minor modifications. In brief, DNA wasfragmented using the Covaris ultrasonicator M220 and sub-sequently converted to sequencing libraries with the aid of aGeneRead Library L core kit (Qiagen) according to themanufacturer’s instructions using barcoded IonXpressadapters (Thermo Fisher). Before clonal amplification andsequencing with HiQ reagents following the manufacturer’sinstructions, the libraries were size selected as described.The data were analysed by mapping the obtained sequencereads along the reference sequence (GenBank accession no.AP008984) [6] using the Genome Sequencer software suite(v3.0; Roche) with a minimum overlap identity of 99% and
Schröder et al., Journal of General Virology
12

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
a minimum alignment between the reads and the referenceof 95%. In total, >700 000 KHV-specific reads were consid-ered. The few remaining ambiguities were clarified by theexamination of individual reads and/or by Sanger sequenc-ing PCR-amplified DNA fragments (see above).
Funding information
This study was supported by the German Federal Ministry of Food andAgriculture (BMEL).
Acknowledgements
The authors thank G. Strebelow and Patrick Zitzow for help withsequence analyses. The technical assistance of A. Landmesser, I.Werner and J. Lange is greatly appreciated.
Conflicts of interest
The authors declare that there are no conflicts of interest.
Ethical statement
The carp used in this study were kept in accordance with the Germanprotection of animals act. The infection trials were approved by theanimal experiment commission of the federal state Mecklenburg-WestPomerania (Az 7221.3-2-002/15, FLI TV 02/15 KHV-att.). The sufferingof the fish was minimized.
References
1. Bretzinger A, Fischer-Scherl T, Oumouna M, Hoffmann R, Truyen
U. Mass mortalities in koi, Cyprinus carpio, associated with gill andskin disease. Bull Eur Ass Fish Pathol 1999;5:182.
2. Hedrick RP, Marty GD, Nordhausen Rw MJK, Bercovier H et al. Aherpesvirus associated with mass mortality of juvenile and adultkoi, a strain of a common carp. Fish Health Newsletter 1999;7:44–57.
3. Neukirch M, Böttcher K, Bunnajirakul S. Isolation of a virus fromkoi with altered gills. Bull Eur Assoc Fish Pathol 1999;19:221–224.
4. Ronen A, Perelberg A, Abramowitz J, Hutoran M, Tinman S et al.
Efficient vaccine against the virus causing a lethal disease in cul-tured Cyprinus carpio. Vaccine 2003;21:4677–4684.
5. Gotesman M, Kattlun J, Bergmann SM, El-Matbouli M. CyHV-3:the third cyprinid herpesvirus. Dis Aquat Organ 2013;105:163–174.
6. Aoki T, Hirono I, Kurokawa K, Fukuda H, Nahary R et al. Genomesequences of three koi herpesvirus isolates representing theexpanding distribution of an emerging disease threatening koi andcommon carp worldwide. J Virol 2007;81:5058–5065.
7. Davison AJ, Eberle R, Ehlers B, Hayward GS, McGeoch DJ et al.
The order Herpesvirales. Arch Virol 2009;154:171–177.
8. Perelberg A, Ronen A, Hutoran M, Smith Y, Kotler M. Protectionof cultured Cyprinus carpio against a lethal viral disease by anattenuated virus vaccine. Vaccine 2005;23:3396–3403.
9. van Oirschot JT. Diva vaccines that reduce virus transmission. JBiotechnol 1999;73:195–205.
10. Cui LC, Guan XT, Liu ZM, Tian CY, Xu YG. Recombinant lactobacil-lus expressing G protein of spring viremia of carp virus (SVCV)combined with ORF81 protein of koi herpesvirus (KHV): a promis-ing way to induce protective immunity against SVCV and KHVinfection in cyprinid fish via oral vaccination. Vaccine 2015;33:3092–3099.
11. Zhou J, Xue J, Wang Q, Zhu X, Li X et al. Vaccination of plasmidDNA encoding ORF81 gene of CJ strains of KHV provides protec-tion to immunized carp. In Vitro Cell Dev Biol Anim 2014;50:489–495.
12. Zhou JX, Wang H, Li XW, Zhu X, Lu WL et al. Construction of KHV-CJ ORF25 DNA vaccine and immune challenge test. J Fish Dis
2014;37:319–325.
13. Costes B, Fournier G, Michel B, Delforge C, Raj VS et al. Cloningof the koi herpesvirus genome as an infectious bacterial artificialchromosome demonstrates that disruption of the thymidine
kinase locus induces partial attenuation in Cyprinus carpio koi. JVirol 2008;82:4955–4964.
14. Fuchs W, Fichtner D, Bergmann SM, Mettenleiter TC. Generationand characterization of koi herpesvirus recombinants lacking viralenzymes of nucleotide metabolism. Arch Virol 2011;156:1059–1063.
15. Vancsok C, Peñaranda MMD, Raj VS, Leroy B, Jazowiecka-Rakus
J et al. Proteomic and functional analyses of the virion transmem-brane proteome of cyprinid herpesvirus 3. J Virol 2017;91:e01209-17.
16. Boutier M, Ronsmans M, Ouyang P, Fournier G, Reschner A et al.
Rational development of an attenuated recombinant cyprinid her-pesvirus 3 vaccine using prokaryotic mutagenesis and in vivo bio-luminescent imaging. PLoS Pathog 2015;11:e1004690.
17. Boutier M, Gao Y, Vancsok C, Su�arez NM, Davison AJ et al. Identi-fication of an essential virulence gene of cyprinid herpesvirus 3.Antiviral Res 2017;145:60–69.
18. Efstathiou S, Kemp S, Darby G, Minson AC. The role of herpessimplex virus type 1 thymidine kinase in pathogenesis. J Gen Virol
1989;70:869–879.
19. Jöns A, Gerdts V, Lange E, Kaden V, Mettenleiter TC. Attenuationof dUTPase-deficient pseudorabies virus for the natural host. VetMicrobiol 1997;56:47–54.
20. Kit S. Recombinant-derived modified-live herpesvirus vaccines.Adv Exp Med Biol 1989;251:219–236.
21. Pavlova SP, Veits J, Keil GM, Mettenleiter TC, Fuchs W. Protectionof chickens against H5N1 highly pathogenic avian influenza virusinfection by live vaccination with infectious laryngotracheitis virusrecombinants expressing H5 hemagglutinin and N1 neuramini-dase. Vaccine 2009;27:773–785.
22. Schnitzlein WM, Winans R, Ellsworth S, Tripathy DN. Generationof thymidine kinase-deficient mutants of infectious laryngotrachei-tis virus. Virology 1995;209:304–314.
23. Zhang HG, Hanson LA. Deletion of thymidine kinase gene attenu-ates channel catfish herpesvirus while maintaining infectivity.Virology 1995;209:658–663.
24. Mletzko A, Amtmann A, Bergmann S, Lee P, Christian J et al.
Inoculation of cyprinid herpesvirus 3 (CyHV-3) on common carpbrain cells-influence of process parameters on virus yield. In Vitro
Cell Dev Biol Anim 2017;53:579–585.
25. Bergmann SM, Wang Q, Zeng W, Li Y, Wang Y et al. Validation of aKHV antibody enzyme-linked immunosorbent assay (ELISA). J Fish
Dis 2017;40:1511–1527.
26. Gilad O, Yun S, Zagmutt-Vergara FJ, Leutenegger CM, Bercovier
H et al. Concentrations of a Koi herpesvirus (KHV) in tissues ofexperimentally infected Cyprinus carpio koi as assessed by real-time TaqMan PCR. Dis Aquat Organ 2004;60:179–187.
27. Hoffmann B, Depner K, Schirrmeier H, Beer M. A universal heter-ologous internal control system for duplex real-time RT-PCRassays used in a detection system for pestiviruses. J Virol
Methods 2006;136:200–209.
28. Hedrick RP, Gilad O, Yun S, Spangenberg JV, Marty GD et al. Aherpesvirus associated with mass mortality of juvenile and adultkoi, a strain of common carp. J Aquat Anim Health 2000;12:44–57.
29. Marchler-Bauer A, Bo Y, Han L, He J, Lanczycki CJ et al. CDD/SPARCLE: functional classification of proteins via subfamilydomain architectures. Nucleic Acids Res 2017;45:D200–D203.
30. Gao Y, Su�arez NM, Wilkie GS, Dong C, Bergmann S et al. Genomicand biologic comparisons of cyprinid herpesvirus 3 strains. VetRes 2018;49:40.
31. Avarre JC, Madeira JP, Santika A, Zainun Z, Baud M et al. Investi-gation of cyprinid herpesvirus-3 genetic diversity by a multi-locusvariable number of tandem repeats analysis. J Virol Methods
2011;173:320–327.
32. Klafack S, Wang Q, Zeng W, Wang Y, Li Y et al. Genetic variabilityof koi herpesvirus in vitro-a natural event? Front Microbiol 2017;8:982.
Schröder et al., Journal of General Virology
13

Downloaded from www.microbiologyresearch.org by
IP: 193.22.115.2
On: Thu, 08 Nov 2018 12:47:39
33. Monaghan SJ, Thompson KD, Adams A, Bergmann SM. Sensitivity
of seven PCRs for early detection of koi herpesvirus in experimen-tally infected carp, Cyprinus carpio L., by lethal and non-lethalsampling methods. J Fish Dis 2015;38:303–319.
34. Fuchs W, Granzow H, Dauber M, Fichtner D, Mettenleiter TC.
Identification of structural proteins of koi herpesvirus. Arch Virol
2014;159:3257–3268.
35. Michel B, Leroy B, Stalin Raj V, Lieffrig F, Mast J et al. Thegenome of cyprinid herpesvirus 3 encodes 40 proteins incorpo-rated in mature virions. J Gen Virol 2010;91:452–462.
36. Vrancken R, Boutier M, Ronsmans M, Reschner A, Leclipteux T
et al. Laboratory validation of a lateral flow device for the detec-
tion of CyHV-3 antigens in gill swabs. J Virol Methods 2013;193:679–682.
37. Fuchs W, Mettenleiter TC. DNA sequence and transcriptional anal-
ysis of the UL1 to UL5 gene cluster of infectious laryngotracheitisvirus. J Gen Virol 1996;77:2221–2229.
38. Fuchs W, Mettenleiter TC. DNA sequence of the UL6 to UL20genes of infectious laryngotracheitis virus and characterization ofthe UL10 gene product as a nonglycosylated and nonessentialvirion protein. J Gen Virol 1999;80:2173–2182.
39. Rosenkranz D, Klupp BG, Teifke JP, Granzow H, Fichtner D et al.
Identification of envelope protein pORF81 of koi herpesvirus. J Gen
Virol 2008;89:896–900.
40. Mettenleiter TC. Glycoprotein gIII deletion mutants of pseudora-bies virus are impaired in virus entry. Virology 1989;171:623–625.
41. Agresti A. A survey of exact inference for contingency tables.Statistical Science 1992;7:131–153.
42. Höper D, Freuling CM, Müller T, Hanke D, von Messling V et al.
High definition viral vaccine strain identity and stability testingusing full-genome population data–The next generation of vaccinequality control. Vaccine 2015;33:5829–5837.
43. Cormack BP, Valdivia RH, Falkow S. FACS-optimized mutants ofthe green fluorescent protein (GFP). Gene 1996;173:33–38.
Schröder et al., Journal of General Virology
14
Five reasons to publish your next article with a Microbiology Society journal
1. The Microbiology Society is a not-for-profit organization.
2. We offer fast and rigorous peer review – average time to first decision is 4–6 weeks.
3. Our journals have a global readership with subscriptions held in research institutions aroundthe world.
4. 80% of our authors rate our submission process as ‘excellent’ or ‘very good’.
5. Your article will be published on an interactive journal platform with advanced metrics.
Find out more and submit your article at microbiologyresearch.org.

Publikationen
22

Publikationen
35
3.2. Publikation II
“Characterization of gene deletion mutants of Cyprinid herpesvirus 3 (koi herpesvirus) lacking the immunogenic envelope glycoproteins pORF25, pORF65, pORF148 and pORF149” Lars Schröder, Sandro Klafack, Sven M. Bergmann, Pei-Yu Alison Lee, Kati Franzke, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs Publiziert in Virus Research

Accepted Manuscript
Title: Characterization of gene deletion mutants of Cyprinidherpesvirus 3 (koi herpesvirus) lacking the immunogenicenvelope glycoproteins pORF25, pORF65, pORF148 andpORF149
Authors: Lars Schroder, Sandro Klafack, Sven M. Bergmann,Pei-Yu Alison Lee, Kati Franzke, Dirk Hoper, Thomas C.Mettenleiter, Walter Fuchs
PII: S0168-1702(18)30474-XDOI: https://doi.org/10.1016/j.virusres.2018.12.004Reference: VIRUS 97537
To appear in: Virus Research
Received date: 3 August 2018Revised date: 7 December 2018Accepted date: 8 December 2018
Please cite this article as: Schroder L, Klafack S, Bergmann SM, Lee P-YuA,Franzke K, Hoper D, Mettenleiter TC, Fuchs W, Characterization of gene deletionmutants of Cyprinid herpesvirus 3 (koi herpesvirus) lacking the immunogenic envelopeglycoproteins pORF25, pORF65, pORF148 and pORF149, Virus Research (2018),https://doi.org/10.1016/j.virusres.2018.12.004
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

1
2nd revision of Virus Research manuscript VIRUS_2018_394
Characterization of gene deletion mutants of Cyprinid
herpesvirus 3 (koi herpesvirus) lacking the immunogenic
envelope glycoproteins pORF25, pORF65, pORF148 and
pORF149
Lars Schrödera, Sandro Klafackb, Sven M. Bergmannb, Pei-Yu Alison Leed, Kati
Franzkeb, Dirk Höperc, Thomas C. Mettenleitera, Walter Fuchsa*
a Institute of Molecular Virology and Cell Biology, Friedrich-Loeffler-Institut,
Greifswald-Insel Riems, Germany
b Institute of Infectology, Friedrich-Loeffler-Institut, Greifswald-Insel Riems, Germany
c Institute of Diagnostic Virology, Friedrich-Loeffler-Institut, Greifswald-Insel Riems,
Germany
d GeneReach Biotechnology Corporation, Taichung, Taiwan
*Corresponding author: Walter Fuchs
e-mail: [email protected]
phone: +49 38351 71258
Keywords: Cyprinid herpesvirus 3, Koi herpesvirus, Envelope glycoproteins, Gene
deletion mutants, Virus replication in vitro, Virulence in carp
Highlights:
CyhV-3 glycoproteins pORF25, pORF65, pORF148, pORF149 are
dispensable for replication
pORF25 and pORF149 are relevant for efficient virus entry and cell-to-cell
spread
ACCEPTED MANUSCRIP
T

2
The adverse effect of ORF149 deletion is compensated by additional ORF148
deletion
Deletion of ORF148 and/or ORF149 does not significantly reduce virulence of
CyHV-3
Deletion of ORF148 and/or ORF149 does not affect protective efficacy of
CyHV-3
Abstract
Cyprinid herpesvirus 3 (CyHV-3) or koi herpesvirus is a global pathogen causing
mass mortality in koi and common carp, against which improved vaccines are
urgently needed. In this study we investigated the role of four nonessential, but
immunogenic envelope glycoproteins encoded by members of the ORF25 gene
family (ORF25, ORF65, ORF148 and ORF149) during CyHV-3 replication. Single
deletion of ORF65 did not affect in vitro replication, and deletion of ORF148 even
slightly enhanced virus growth on common carp brain (CCB) cells, deletions of
ORF25 or ORF149 led to reduced plaque sizes and virus titers, which was due to
delayed entry into host cells. An ORF148/ORF149 double deletion mutant exhibited
wild-type like growth indicating opposing functions of the two proteins. Electron
microscopy of CCB cells infected with either mutant did not indicate any effects on
virion formation and maturation in nucleus or cytoplasm, nor on release of enveloped
particles. The ORF148, ORF149 and double deletion mutants were also tested in
animal experiments using juvenile carp, and proved to be insufficiently attenuated for
use as live virus vaccines. However, surviving fish were protected against challenge
with wild-type CyHV-3, demonstrating that these antibody inducing proteins are
dispensable for an efficient immune response in vivo.
1. Introduction
Cultivation of common carp has a long tradition in Europe and Asia, and in several
regions provides a key source of protein for human consumption. Furthermore, the
breeding of colored carp varieties (koi) as ornamental fish has become a popular
ACCEPTED MANUSCRIP
T

3
hobby, and a profitable business worldwide (Balon, 1995). In the 1990ies, a
previously unknown virus infection led to mass mortality of common and koi carp in
Israel and Europe. The causative agent was identified as a herpesvirus and
designated as carp interstitial nephritis and gill necrosis virus (CNGV), or koi
herpesvirus (KHV) (Bretzinger et al., 1999; Hedrick et al., 1999; Neukirch et al., 1999;
Ronen et al., 2003). Due to the severity of the infection, the development of
attenuated vaccines was rapidly initiated (Bretzinger et al., 1999; Hedrick et al., 1999;
Neukirch et al., 1999; Ronen et al., 2003).
Analyses of genome sequences demonstrated a close relationship to carp pox virus
(Cyprinid herpesvirus 1), and goldfish haematopoietic necrosis virus (Cyprinid
herpesvirus 2), leading to classification as Cyprinid herpesvirus 3 (CyHV-3) within the
genus Cyprinivirus in the family Alloherpesviridae and the order Herpesvirales
(Davison et al., 2009). The CyHV-3 genome has a size of 295 kbp including 22 kbp of
direct repeat sequences at both ends, and contains approximately 155 open reading
frames (ORFs) predicted to encode proteins (Aoki et al., 2007). Some of these ORFs
are conserved in other herpesviruses of fish or amphibians, but homologies to
mammalian and avian herpesviruses (family Herpesviridae) are very limited. Analysis
of purified virus particles by mass spectrometry identified 40 different CyHV-3
encoded proteins, including 3 capsid components, and 13 membrane proteins
(Michel et al., 2010).
The integral membrane proteins pORF25, pORF65, pORF81, pORF99, pORF136,
pORF148 and pORF149 were also identified and characterized by specific antisera
or monoclonal antibodies (mAb), and, except pORF81 and pORF136, proved to be
modified by N-linked glycosylation (Rosenkranz et al., 2008; Vrancken et al., 2013).
ORFs 25, 65, 148, and 149 constitute members of a family of distantly related genes
which is conserved in cyprinid herpesviruses, and might have evolved from a
common ancestor (Andrew J. Davison et al., 2013; Aoki et al., 2007). Indirect
immunofluorescence (IF) tests of eukaryotic cells transfected with expression
plasmids for ORF25, ORF65, ORF99, ORF148 or ORF149 revealed specific
reactions of the respective glycoproteins with serum antibodies from experimentally
and naturally KHV-infected carp (Fuchs et al., 2014). Therefore, vaccines based on
KHV mutants lacking these genes might be suitable for serological differentiation
between infected and vaccinated animals (DIVA) (van Oirschot, 1999). On the other
hand, candidate subunit or DNA vaccines containing or encoding single CyHV-3
ACCEPTED MANUSCRIP
T

4
envelope proteins like pORF25 or pORF81 have been shown to induce protective
immune responses (Cui et al., 2015; Zhou et al., 2014a; Zhou et al., 2014b),
suggesting that their deletion might affect the protective efficacy of CyHV-3 live
vaccines. Recent studies demonstrated, that the four ORF25 family genes including
ORF65, ORF148, and ORF149 are not essential for CyHV-3 replication in cell
culture, and that deletion of ORF25 attenuated the virus in vivo, but significantly
reduced its protective efficacy against wild-type challenge (Vancsok et al., 2017). In
contrast, single deletions of ORF65, ORF148, or ORF149 did not lead to sufficient
attenuation of a virulent CyHV-3 strain in carp (Vancsok et al., 2017). Thus, other
CyHV-3 recombinants possessing e.g. double deletions of the viral thymidine kinase
(TK, ORF55) and deoxyuridine triphosphatase (dUTPase, ORF123) genes or of the
adjacent ORFs 56 and 57 appeared to be more promising (Boutier et al., 2017;
Schröder et al., 2018).
Whereas the previous functional analyses of CyHV-3 envelope proteins were
performed with a member of the European virus lineage isolated in Belgium (Vancsok
et al., 2017), we reanalyzed the potential of ORF25, ORF65, ORF148, and ORF149
deletions for vaccine development by using a less virulent, but highly cell-culture
adapted East Asian type CyHV-3 strain from Taiwan (KHV-T) (Mletzko et al., 2017;
Gao et al., 2018). To investigate possible additive effects on virulence, or functional
redundancy we also generated an ORF148 and ORF149 double deletion mutant.
Furthermore, the cell culture adaptation of KHV-T facilitated in vitro investigation of
individual viral gene functions. The crucial roles and interactions of envelope
(glyco)proteins during virus entry, virion formation, spread and release have been
extensively investigated for mammalian herpesviruses (reviewed by (Mettenleiter,
2002; Sathiyamoorthy et al., 2017), but little is known about their functional
homologues in Alloherpesviruses. We therefore studied the roles of pORF25,
pORF65, pORF148, and pORF149 in entry and virus morphogenesis by penetration
kinetics and electron microscopy of infected common carp brain (CCB) cells
(Neukirch et al., 1999).
2. Materials and methods
ACCEPTED MANUSCRIP
T

5
2.1. Viruses and cells
KHV-T (Mletzko et al., 2017), and its derivatives were propagated in CCB cells
(Neukirch et al., 1999), which were grown in minimum essential medium (MEM)
supplemented with 10 % fetal bovine serum (FBS) at 25°C. Infected cells were
incubated in medium containing only 5 % FBS, and, after appearance of
cytopathogenic effects, lysed by freeze-thawing and stored in aliquots at -80°C. For
plaque assays, CCB cell monolayers were incubated with serial virus dilutions for 2 h.
Subsequently, the inoculum was replaced by MEM containing 5 % FBS and 6 g/l of
methylcellulose, and incubation at 25°C was continued for 3 to 7 days.
2.2. Construction of deletion plasmids and virus recombinants
Genomic KHV-T DNA was prepared from infected cell lysates as described (Fuchs
and Mettenleiter, 1996). For the generation of deletion mutants, transfer plasmids
were prepared after amplification of the targeted glycoprotein gene regions from
KHV-T DNA by PCR using Pfx DNA polymerase (Thermo Fisher Scientific), with the
respective primer pairs (Table 1). The obtained 6005 bp (ORF148/149), 3629 bp
(ORF25), 3210 bp (ORF65), and 6005 bp (ORF148/149), PCR products were
digested at engineered EcoRI and HindIII sites, and inserted into the correspondingly
digested vector pUC19 (New England Biolabs), resulting in plasmids pUC-KT25R,
pUC-KT65R, and pUC-KT149R (Fig. 1B-D) From the latter plasmid, major parts of
ORF148 and ORF149 were deleted either singly, or in combination (Fig. 1D). For the
double deletion a 3048 bp HpaI/PstI fragment of pUC-KT149R was either removed or
replaced by an expression cassette for enhanced green fluorescent protein (EGFP),
which had been isolated as a 1607 bp BamHI/EcoRI fragment from plasmid pBl-GFP
(Fuchs and Mettenleiter, 1999). Single deletions of ORF148 or ORF149 were
introduced by removal or EGFP-substitutions of 635 bp HpaI/NotI, or 1252 bp
XcmI/PstI fragments, respectively, resulting in transfer plasmids pUC-KT∆148(G),
pUC-KT∆149(G), pUC-KT∆148/149(G). In a similar manner, major parts of ORF25
(Fig. 1B) and ORF65 (Fig. 1C) were removed from the original plasmids by digestion
with BamHI or BsiWI, resulting in deletions of 1220 bp or 1169 bp, respectively.
Optionally, the deleted fragments were replaced by the EGFP expression cassette,
yielding plasmids pUC-KTΔ25(G) and pUC-KTΔ65(G). In all cloning experiments
ACCEPTED MANUSCRIP
T

6
non-compatible fragment ends were blunted by treatment with Klenow polymerase
prior to ligation.
In a first series of experiments CCB cells were cotransfected (K2 Transfection
System, Biontex) with genomic KHV-T DNA and the GFP-gene containing deletion
plasmids Homologous recombination led to the corresponding GFP-expressing virus
mutants KHV-T∆148G, KHV-T∆149G, KHV-T∆148/149G, KHV-TΔ25G and KHV-
TΔ65G (Fig. 1) which could be purified to homogeneity by plaque assays on CCB
cells, and subsequent aspiration of green fluorescent cell foci. Subsequently,
genomic DNA of these KHV-T recombinants was prepared and used for
cotransfections with deletion or rescue plasmids lacking a reporter gene. From the
resulting non-fluorescent virus plaques deletion mutants KHV-TΔ148, KHV-TΔ149,
KHV-TΔ148/149, KHV-TΔ25, and KHV-TΔ65, as well the revertants KHV-T148R,
KHV-T149R, KHV-T148/149R, KHV-T25R and KHV-T65R (Fig. 1) were isolated.
Genomic DNA of all generated CyHV-3 recombinants was analyzed by restriction
and Southern blot analyses (not shown), and the modified genome regions were
further characterized by PCR and sequencing using the specific primers described
here (Table 1), and previously (Fuchs et al., 2014).
2.3. In vitro replication studies and indirect immunofluorescence (IIF) tests
For determination of replication kinetics CCB cells were grown in 24well plates and
infected with KHV-T and the generated virus recombinants at a multiplicity (MOI) of
0.1 at 4°C to permit virus adsorption. After 1 h, pre-warmed medium was added and
incubation was continued for 2 h at 25°C to allow penetration. Subsequently, non-
penetrated virus was inactivated by treatment with citric acid-buffered saline (pH 3.0)
(Mettenleiter, 1989), and incubation at 25°C was continued under fresh medium. At
different times after the temperature shift, individual plates were frozen at -80°C. For
determination of progeny virus titers, the plates were thawed at 25°C, and serial
dilutions of the cell lysates were analyzed by plaque assays on CCB cells. After 4 d
at 25°C, the cells were washed with phosphate-buffered saline (PBS), and fixed with
50 % methanol / 50 % acetone for 30 min at -20°C. The fixed cells were incubated
with a monospecific rabbit antiserum detecting the envelope protein pORF81 of
CyHV-3, and fluorochrome-conjugated secondary antibodies as described
(Rosenkranz et al., 2008), and investigated by fluorescence microscopy (Eclipse Ti-S
ACCEPTED MANUSCRIP
T

7
with software NIS-elements, version 4, Nikon). Mean titers of four replicates per virus
mutant were calculated. Furthermore, the areas of 30 plaques per virus were
measured, and average sizes as well as standard deviations were calculated as
percentages of the mean plaque areas induced by KHV-T. Unpaired two-sided
Student’s t-tests were applied to evaluate the statistical significance of differences
between plaque sizes and titers of the CyHV-3 mutants.
2.4. In vitro penetration kinetics
CCB cells grown in 6well plates were infected with approximately 250 plaque-forming
units (PFU) of the GFP-expressing virus mutants KHV-TΔ148G, KHV-TΔ149G, KHV-
TΔ148/149G, KHV-TΔ25G, and KHV-TΔ65G, or, as wild-type like control, KHV-
TΔDUTGFP (Schröder et al., 2018). After 1 h at 4°C, prewarmed medium was
added, and incubation was continued at 25°C. Before and 10, 20, 30, 60 and 120
minutes after the temperature shift, remaining extracellular virus particles were
inactivated by low pH treatment (Mettenleiter, 1989). Subsequently, the cells were
washed with PBS, overlaid with semisolid medium, and incubated for 3 d at 25°C.
The plaques of the penetrated viruses after different times were counted by
fluorescence microscopy, and compared to plaque numbers obtained in wells which
were not acid-treated, and overlaid with semisolid medium after 120 minutes at 25°C
(Vallbracht et al., 2017). Mean percentage values, and standard deviations were
determined in three independent experiments. Unpaired two-sided Student’s t-tests
were applied to evaluate the statistical significance of differences between
penetration rates of the CyHV-3 mutants.
2.5. Western blot analyses
CCB cells were infected with KHV-T or the generated virus recombinants at an MOI
of 1, and incubated for two days at 25°C. The cells were lysed and proteins were
separated by discontinuous sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). Transfer to nitrocellulose membranes, and subsequent
incubation of the blots was done as described (Pavlova et al., 2009). Monospecific
rabbit antisera against CyHV-3 pORF25, pORF149, and the major capsid protein
pORF92 (Fuchs et al., 2014) were used at dilutions of 1 : 10,000. Binding of
peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch) was
ACCEPTED MANUSCRIP
T

8
detected and recorded (Clarity Western ECL Substrate, and VersaDoc 4000 MP, Bio-
Rad).
2.6. Ultrastructural analyses
CCB cells were infected with KHV-T or the generated virus recombinants at an MOI
of 1, and incubated for 20 h at 25°C. The samples were fixed and processed for
transmission electron microscopy as described previously (Klupp et al., 2000), and
examined using a Tecnai Spirit electron microscope (FEI) and an acceleration
voltage of 80 kV.
2.7. Sequence analyses
The inserts of transfer plasmids, and the modified genome regions of virus
recombinants were analyzed by sequencing using the BigDye Terminator v1.1 cycle
sequencing kit, and a 3130 Genetic Analyzer (Applied Biosystems). Results were
evaluated with the GeneiousTM software package in version 10.2.3 (Biomatters).
Furthermore, the full genome sequence of KHV-T148/149 was determined as
described recently (Höper et al., 2015; Schröder et al., 2018).
2.8. Animal trial
In vivo attenuation and protective efficacy of the generated ORF148 and ORF149
mutants of KHV-T was tested using five groups of carp, each consisting of 40 nine
months-old (approximately 8 g / 5-7 cm) and 5 two years-old (approximately 200 g /
20-23 cm) fish (Fischzucht Traßdorf 1967 e.V., Traßdorf, Germany). The animals
were kept at a constant temperature of 20°C, and infected by immersion for 1 h in 5 l
water containing 2.5 x 107 PFU of either KHV-T, KHV-TΔ148, KHV-TΔ149, KHV-
TΔ148/149, or cell culture medium. After 35 d, all carp that had survived primary
CyHV-3 infection were challenged by immersion for 1 h in 5 l water containing 2.5 x
107 PFU of KHV-T. The fish were examined for additional 28 days for clinical
symptoms. Blood samples and gill swabs were taken from the two years-old carp
before, and two and four weeks after primary infection, as well as after challenge.
The young carps were considered for evaluation of the mortality rates. Statistical
significance of differences between the groups was evaluated using two-sided
Fisher’s exact tests (Agresti A., 1992).
ACCEPTED MANUSCRIP
T

9
2.9. Enzyme-linked immunosorbent assay (ELISA)
Blood samples from infected and uninfected carp were centrifuged at 600 x g for 15
min and the supernatant was used in the ELISA. NUNC POLYSORP immuno-plates
(Thermo Fisher Scientific) were coated with sucrose-gradient purified KHV-T
particles, blocked, incubated with the 1 : 300 diluted sera in two replicates. An anti-
carp IgM monoclonal antibody (Aquatic diagnostics), and horseradish peroxidase-
conjugated anti-mouse IgG (Jackson ImmunoResearch) were added as described
(Bergmann et al., 2017). Substrate reactions were measured in an Infinite F200
microplate reader (Tecan), and OD450nm values > 0.2 were considered as positive.
ACCEPTED MANUSCRIP
T

10
3. Results and discussion
3.1. Generation and genetic characterization of glycoprotein deletion mutants
of KHV-T
Previous studies have shown that sera from naturally or experimentally CyHV-3
infected carp and koi specifically reacted with cells transfected with expression
plasmids encoding pORF25, pORF65, pORF148, and pORF149, which are distantly
related members of a family of type I envelope glycoproteins (Fuchs et al., 2014).
Despite their immunogenicity, the four glycoproteins have recently been shown to be
dispensable for in vitro replication of a highly virulent CyHV-3 isolate from Belgium,
which has been cloned and engineered as a bacterial chromosome (Costes et al.,
2008; Vancsok et al., 2017). In the present study, we generated similar gene deletion
mutants of the cell culture-adapted strain KHV-T (Mletzko et al., 2017).
Like in our previous studies isolation of the gene deletion mutants, and of
corresponding revertants was facilitated by transient insertion of a GFP reporter
cassette (Schröder et al., 2018). The deletion of ORF25 (609 codons, Fig. 1B)
encompassed codons 138 to 545, whereas from ORF65 (595 codons, Fig. 1C),
codons 36 to 425 were removed, and the remaining 5’ and 3’ ends were fused out-of-
frame in the final deletion mutants KHV-T25 and KHV-T65. The single deletions of
ORF148 (602 codons) and ORF149 (700 codons), removed codons 70 to 281 and 17
to 434, respectively (Fig. 1D), and frameshifted the retained parts of the ORFs in
KHV-T148 and KHV-T149. Consequently, the double deletion ranged from codon
17 of ORF149 to codon 281 of ORF148, but in this case the deletion was in frame.
However, expression of a chimeric membrane protein by KHV-T148/149 is unlikely,
since the predicted signal peptide of pORF149 was affected by the deletion (Fig. 1D).
Genomic DNA of all generated CyHV-3 recombinants was analyzed by restriction
enzyme digestions and Southern blot hybridizations, which confirmed the desired,
and indicated no unwanted alterations (see supplementary figure S1). Furthermore,
the mutated genes were amplified from viral DNA by PCR and subsequently
sequenced using the primer pairs CyHV-3O25-F and -R, CyHV-3O65-F and -R, or
CyHV-3O148-R and CyHV3-O149-F, respectively (Fuchs et al., 2014). The PCR
products of all CyHV-3 mutants exhibited the expected sizes (Fig. 2) and sequences.
Analysis of the complete genome sequence was performed only for the recombinant
ACCEPTED MANUSCRIP
T

11
KHV-T148/149. As expected, this sequence was very similar to that of the parental
virus strain KHV-T (Gao et al., 2018), except the deletion within ORFs 148 and 149
(Table 2). Nine of the 11 other observed alterations affected homopolymeric
nucleotide stretches or tandem repetitions of short motifs which tend to be highly
variable (highlighted in grey). Although few of these regions were localized in ORFs
45 and 52, no frameshifts or stop codons were introduced (Table 2). We found similar
differences to the published sequence, including two single base substitutions, and a
considerable elongation of ORF52 also in another recently sequenced recombinant
of KHV-T which lacked the viral thymidine kinase and dUTPase genes (Schröder et
al., 2018). Thus, these alterations are presumably present in the KHV-T variants
used in different labs, and not due to accidental mutations which occurred during
preparation of the virus recombinants.
3.2. Protein expression of the CyHV-3 mutants
Protein expression of the generated glycoprotein gene deletion mutants of KHV-T
was analyzed using available antisera (Fuchs et al., 2014). CCB cells were harvested
for Western blotting 2 d after infection at an MOI of 1 with the respective CyHV-3
mutants. Glycoprotein pORF25 was detectable in cells infected with KHV-T or the
revertant KHV-T25R, but not in cells infected with KHV-T25 (Fig. 3A). Like the
corresponding protein of KHV-I (Fuchs et al., 2014), pPORF25 of KHV-T (calculated
mass of the primary translation product of 66.1 kDa) ,exhibited an apparent molecular
mass of approximately 125 kDa, indicating presence of several carbohydrate chains,
i.a. at 5 predicted N-linked glycosylation sites (asparagine-X-serine/threonine). In
contrast, pORF149 (calculated mass 72.0 kDa) contains only one potential N-
glycosylation site, but three protein bands of approximately 100, 120 and 160 kDa
were detected by the antiserum (Fig. 3B). The specificity of these reactions was
confirmed by the absence of these signals in cells infected with the deletion mutants
KHV-T149 and KHV-T148/149, and their reappearance in ORF149 rescuants (Fig.
3B). It remains to be elucidated whether the large forms of pORF149, which contains
74 serine (10.6 %) and 143 threonine (20.5 %) residues, are due to O-linked
glycosylation, other post-translational modifications, or stable protein-protein
interactions. An antiserum against the major capsid protein pORF92 of CyHV-3
(Fuchs et al., 2014) served as control, and similar amounts of the approximately 140
ACCEPTED MANUSCRIP
T

12
kDa gene product (calculated mass 139.6 kDa) were detected in all infected samples
(Fig. 3, lower panels). Our available pORF65- and pORF148-specific sera (Fuchs et
al., 2014) did not permit unambiguous detection of their target proteins by Western
blot or IF analyses of CyHV-3 infected cells, but expression of pORF25 and
pORF149 was demonstrated to be unaffected in ORF65 and ORF148 mutants of
KHV-T (results not shown).
Although we cannot identify pORF148 directly, previous studies showed that its
expression in another CyHV-3 strain was not affected by deletion of the upstream
ORF149 (Vancsok et al., 2017), and, therefore, it appears likely that ORF148 is also
expressed by KHV-T149. This assumption is supported by the different phenotypes
of our ORF148, ORF149, and double deletion mutants (see below).
The absence of pORF65 from cell lysates infected with KHV-T65 or KHV-65G, as
well as its rescue in KHV-T65R, could be demonstrated by lateral flow tests
(FASTest® KOI HV, Megacor) based on pORF65-specifc mAb (Vrancken et al.,
2013) (see supplementary figure S2). This test might also permit fast differentiation of
potential CyHV-3 vaccines lacking ORF65 from wild-type virus in carp.
3.3. In vitro replication properties of the CyHV-3 mutants
Cell-to-cell spread of KHV-T and the derived mutants was compared by plaque
assays on CCB cells, which were evaluated 4 d after infection (Fig. 4). Infected cells
were identified by IIF tests and plaque areas were determined microscopically. The
plaque sizes of the ORF65 and ORF148 mutants ranged between 95 % and 99 % of
wild-type levels, which was statistically not significant. In contrast, plaque sizes of
KHV-T∆25G, KHV-T∆25, KHV-T∆149G, and KHV-T∆149 were significantly (p <
0.005) reduced by approximately 50 % compared to wild type KHV-T. These
spreading defects were corrected in the rescue mutants KHV-T25R and KHV-T149R.
Interestingly, the double gene deletions in KHV-T∆148/149G and KHV-T∆148/149
also resulted in almost wild-type sized plaques (Fig. 4), indicating that the adverse
effect of ORF149 deletion is compensated by the additional removal of ORF148.
Although plaques of KHV-T∆148/149 appeared significantly (p < 0.05) smaller than
those of KHV-T, no significant size differences were observed between the deletion
mutant and the rescuant KHV-T148/149R.
ACCEPTED MANUSCRIP
T

13
Growth kinetics analyzed after synchronized infection at a MOI of 0.1 confirmed that
ORF25, ORF65, ORF148 and ORF149 are dispensable for productive replication of
CyHV-3 in CCB cells (Fig. 5A). Single deletions of ORF25 (blue bars) or ORF149
(red bars) resulted in delayed replication compared to wild-type KHV-T and the
corresponding revertants. The observed differences were statistically significant (p <
0.05) at nearly all times after infection. The ORF25-deleted, but not the ORF149-
deleted virus also exhibited a significant, approximately 10fold reduction of final titers.
Whereas deletion of ORF65 (orange bars), and ORF148/149 (yellow bars) had no
distinct effect, single deletion of ORF148 (green bars) slightly enhanced replication.
This enhancement was significant (p < 0.05) at 6 of the 7 analyzed times after
infection when compared to wild-type KHV-T, and at 5 time points when compared to
the revertant KHV-T148R (Fig. 5A). Apart from few outliers at single times, all
revertant viruses exhibited wild-type like growth properties (Fig. 5A). The GFP-
expressing gene deletion mutants exhibited similar replication kinetics as the
corresponding reporter-less mutants (results not shown), and were omitted to reduce
complexity of the diagram. However, to confirm the effects of ORF148 and ORF149
deletion, the corresponding subset of CyHV-3 recombinants was reanalyzed in a
second series of growth kinetic studies at lower MOI (0.01) (see supplementary figure
S3). These studies confirmed that, independent of presence or absence of a GFP
reporter gene, the ORF149 deletion delayed, and the ORF148 deletion enhanced
replication of KHV-T. Again the effects appeared additive, resulting in wild-type like
growth of the double deletion mutants.
In general, these results are in line with those of previous investigations of similar
single gene deletion mutants generated by mutagenesis of a BAC-cloned CyHV-3
isolate from Belgium (Vancsok et al., 2017). These studies also showed wild-type like
growth of the ORF65-deleted virus, but delayed replication and cell-to-cell spread of
ORF25- and ORF149-deleted mutants on CCB cells, as well as reduced final titers of
ORF25-negative virus. However, unlike in our experiments a beneficial effect of
ORF148 deletion was not observed, and ORF148/149 double mutants were not
investigated in the previous study (Vancsok et al., 2017).
To further elucidate the functions of the ORF25 gene family members of CyHV-3, we
analyzed in vitro penetration kinetics of the different deletion mutants using acid-
inactivation protocols established for another herpesvirus, Pseudorabies virus (PrV,
Suid alphaherpesvirus 1) (Mettenleiter, 1989; Vallbracht et al., 2017). To facilitate
ACCEPTED MANUSCRIP
T

14
evaluation by direct fluorescence microscopy, the GFP-expressing CyHV-3
glycoprotein deletion mutants were used for these experiments, and a GFP-
expressing dUTPase- deletion mutant of KHV-T (KHV-TDUTG) served as “wild
type” control. This virus had been shown to exhibit wild type-like replication properties
in CCB cells (Schröder et al., 2018). After synchronized adsorption to CCB cells on
ice, infectious particles of KHV-TDUTG, KHV-T65G and KHV-T148G were
shown to enter cells almost completely within two hours at 25°C (Fig. 5B). In
contrast, only 58 % of the KHV-T25G and 36 % of the KHV-T149G particles
became insensitive to acid treatment within this time, and also at earlier times lower
proportions of these two mutants had penetrated into the cells (Fig. 5B). For
ORF149-deleted CyHV-3 the observed penetration defect is statistically highly
significant (p < 0,005). These findings explain the delayed replication (Fig. 5A), and
the reduced plaque sizes (Fig. 4) of ORF25 and ORF149-deleted CyHV-3.
Interestingly, the double mutant KHV-T148/149G exhibited wild type-like penetration
kinetics (Fig. 5B).
To exclude relevance of the envelope glycoproteins pORF25, pORF65, pORF148,
and pORF149 for virion morphogenesis, CCB cells were analyzed by electron
microscopy 20 h after infection with KHV-T or the single and double mutants at a MOI
of 1 (Fig. 6). As exemplarily shown for KHV-T25 and KHV-T149, all stages of
morphogenesis, including nucleocapsid formation in the nucleus (Fig. 6A, 6C, 6E,
asterisks), nuclear egress by transit through the nuclear membrane (Fig. 6A, 6C, 6E,
arrows), final envelopment of nucleocapsids in the cytoplasm (Fig. 6B, 6D, 6F,
squares), and released mature virus particles (Fig. 6A, 6B, 6E, 6F, triangles) were
found in cells infected with either virus. Thus, we found no evidence for a role of the
investigated glycoproteins in maturation or egress of CyHV-3 virions, although it
cannot be excluded considering a potential functional redundancy within the
paralogous ORF25 family proteins.
However, based on our results the ORF25 family members pORF25 and pORF149
play an accessory role during virus entry into host cells. This process and the
involved proteins have been extensively investigated for mammalian
alphaherpesviruses like herpes simplex virus type 1 (HSV-1, Human
alphaherpesvirus 1) (Eisenberg et al., 2012; Sathiyamoorthy et al., 2017), whereas
little is known about the corresponding gene products of alloherpesviruses. In most
ACCEPTED MANUSCRIP
T

15
alphaherpesviruses four envelope glycoproteins (gB, gD, gH, gL) are required for
specific receptor binding and subsequent fusion of the viral envelope with the host
cell plasma membrane. However, except gD, the former proteins are conserved
throughout the Herpesviridae, whereas the ORF25 family seems to be restricted to
the Cyprinivirus genus of the Alloherpesviridae (Aoki et al., 2007; Davison et al.,
2013). Moreover, at least the core fusion protein gB, and gH are absolutely essential
for productive replication of the Herpesviridae, whereas the ORF25 family members
of CyHV-3 are dispensable (Vancsok et al., 2017; this study). Therefore, the proteins
investigated in this study are presumably not required for the membrane fusion
process per se. However, it appears conceivable that the structurally related gene
products of the ORF25 family represent redundant receptor binding proteins of
CyHV-3, which are able to bind the same or different host cell membrane proteins,
and to activate the hitherto unknown alloherpesvirus fusion protein(s). Therefore, it
has to be tested whether combined deletion of all ORF25 family members from the
CyHV-3 genome, i.e. of ORF25, ORF65, ORF148, ORF149, and the often
spontaneously deleted or frameshifted ORFs 26 and 27, still results in a replication
competent virus.
An interesting finding was the positive effect of ORF148 deletion, which compensated
the adverse effects of ORF149 deletion in the double deletion mutants. Although
KHV-T148/149 might express a chimeric membrane protein containing the C-
terminal part of pORF148, and a modified N-terminal signal sequence originating
from pORF149, this was not possible in KHV-T148/149G, where the two gene
remnants were separated by the GFP cassette (Fig. 1D). Nevertheless both double
mutants exhibited similar, almost wild type like in vitro replication properties, which
were clearly improved compared to those of the ORF149 single deletion mutants
(Fig. 4, 5). Furthermore, full genome sequencing of KHV-T148/149 revealed no
accidental mutations compared to KHV-T which could compensate for ORF149
deletion. However, pORF148 and pORF149 exhibit an amino acid sequence identity
of 37.5 %, suggesting related “functions”. Possibly, pORF148 represents a
nonfunctional “copy” of pORF149, which is expressed and able to bind cellular
receptors, but not to trigger subsequent virus entry. Thus, absence of pORF148
might facilitate receptor binding of pORF149, which seems to be the most relevant
ORF25 family member for efficient entry (Fig. 5). In the absence of both proteins,
other receptor binding proteins, e.g. pORF25 or pORF65, might be able to
ACCEPTED MANUSCRIP
T

16
compensate. This hypothesis is supported by the moderately delayed penetration of
ORF25-deleted CyHV-3 (Fig. 5).
3.4. Virulence and protective efficacy of ORF148 and ORF149 gene deleted
CyHV-3 mutants in carp
Previous experiments have shown that deletion of ORF25 leads to attenuation of
CyHV-3, but interferes with protection against subsequent wild type challenge
(Vancsok et al., 2017). This might have been either due to the relevance of pORF25-
specific immune reactions for protection (Zhou et al., 2014b), or to an inefficient in
vivo replication of the deletion mutant. In contrast, ORF65, ORF148, and ORF149
single gene deletion mutants of the BAC-cloned Belgian CyHV-3 isolate were not
sufficiently attenuated (Vancsok et al., 2017). However, previously we have shown
that combined deletion of the TK and dUTPase genes of CyHV-3 leads to an
improved attenuation compared to the corresponding single deletions without
affecting protective efficacy of the candidate vaccine (Fuchs et al., 2011; Schröder et
al., 2018). Therefore, we now examined the virulence of, and the immune responses
induced by the double mutant KHV-T148/149. The corresponding single mutants
KHV-T148 or KHV-T149 were also evaluated, and compared to the parental strain
KHV-T, which has been considered to be much less virulent than the virus used for
previous in vivo testing of glycoprotein deletion mutants of CyHV-3 (Gao et al., 2018).
In the present animal trial, five groups consisting of 40 nine months- and 5 two years-
old carp, were infected by immersion in 5 l water containing 2.5 x 107 PFU of the
virus mutants, KHV-T or cell culture medium. Between day 6 and 31 after wild-type
infection, 17 of the juvenile carp died, resulting in a total mortality rate of 42.5 % (Fig.
7A). This rate was twice as high as observed by others using the corresponding virus
strain (Gao et al., 2018), but in line with the results of previous animal experiments
with KHV-T in our institute (Schröder et al., 2018). It remains to be elucidated
whether this difference is due to different susceptibilities of the used carp breeds,
different virus doses or water temperatures, or to the few determined sequence
alterations between the KHV-T preparations used in the different laboratories (Table
2)(Schröder et al., 2018).
Infection with KHV-T∆148 or KHV-T∆149 led to loss of 9 (22.5 %) or 8 (20 %)
animals, respectively (Fig. 7A). Mortality caused by the double deletion mutant KHV-
ACCEPTED MANUSCRIP
T

17
T∆148/149 was slightly higher (10 of 40 carp or 25 %). According to Fisher’s exact
tests the observed reductions of mortality rates compared to wild type infection were
not significant (p > 0.05), and the animals of all infected groups developed one or
more signs of CyHV-3 infection like fatigue, skin lesion and excess mucus
production. This applied also to the two year-old fish, which, however, survived
infection with wild-type or mutant KHV-T. Thus, in line with previous results (Vancsok
et al., 2017) deletion of ORF148 and ORF149 did not sufficiently attenuate CyHV-3
for use as a live vaccine. Unlike the TK and dUTPase double deletions (Schröder et
al., 2018), combined removal of ORF148 and ORF149 did not improve in vivo
attenuation of the virus, correlating with the enhanced in vitro growth of KHV-
T∆148/149 compared to KHV-T∆149 (Figs. 4, 5).
Five weeks after primary infection all surviving fish and naïve control animals were
challenged by immersion in 5 l water containing 2.5 x 107 PFU of wild-type KHV-T.
The mock-immunized carp developed severe clinical signs, and 15 of them (37.5 %)
died (Fig. 7A). In contrast, no symptoms were observed in any of the immunized
carp, and all of them survived challenge infection (Fig 7A). Although responses to
heterologous, more virulent, challenge virus strains remain to be evaluated, the
present results indicate that even combined deletion of the highly immunogenic
envelope glycoproteins pORF148 and pORF149 (Fuchs et al., 2014) did not
substantially affect protective efficacy of CyHV-3. In line with this, an ELISA using
plates coated with purified CyHV-3 virions (Bergmann et al., 2017), indicated similar
or even higher titers of specific serum antibodies in animals infected with the deletion
mutants than in wild type infected carp (Fig. 7B). We have shown previously that
several other CyHV-3 proteins, including glycoproteins pORF25, pORF65, pORF99,
and the major capsid protein pORF92 also induce humoral immune responses in
carp (Fuchs et al., 2014), which might have covered the lack of pORF148 and/or
pORF149 specific antibodies in the present animal experiments. Since all
glycoproteins encoded by the ORF25 gene family of CyHV-3 are obviously
dispensable for in vitro virus replication (Vancsok et al., 2017, and this study), their
deletion might support the development of DIVA vaccines (van Oirschot, 1999) which
permit serological differentiation between immunized and wild type-infected fish.
Conflicts of interest
ACCEPTED MANUSCRIP
T

18
The authors declare that there are no conflicts of interest
Ethical statement
The carp used in this study were kept in accordance with the German protection of
animal act. The experiments were approved by the animal experiment commission of
the federal state Mecklenburg-West Pomerania. Suffering of animals was minimized.
Acknowledgments
The studies were supported by the German Federal Ministry of Food and Agriculture.
The authors thank G. Strebelow for help with sequence analyses, and M. Jörn for
graphical design of the EM micrographs. The technical help of A. Landmesser, I.
Werner, J. Lange, P. Zitzow, and P. Meyer is greatly appreciated.
ACCEPTED MANUSCRIP
T

19
References
Agresti A., 1992. A Survey of Exact Inference for Contingency Tables. Statistical Science 7(1), 131-177. Andrew J. Davison, Tomofumi Kurobe, Derek Gatherer, Charles Cunningham, Ian Korf, Hideo Fukuda,
Ronald P. Hedrick, Waltzek, T.B., 2013. Comparative Genomics of Carp Herpesviruses. Journal of Virology 87, 2908-2922.
Aoki, T., Hirono, I., Kurokawa, K., Fukuda, H., Nahary, R., Eldar, A., Davison, A.J., Waltzek, T.B., Bercovier, H., Hedrick, R.P., 2007. Genome sequences of three koi herpesvirus isolates representing the expanding distribution of an emerging disease threatening koi and common carp worldwide. J Virol 81(10), 5058-5065.
Balon, E.K., 1995. Origin and domestication of the wild carp, Cyprinus carpio: from Roman gourmets to the swimming flowers. Aquaculture 129, 3–48.
Bergmann, S.M., Wang, Q., Zeng, W., Li, Y., Wang, Y., Matras, M., Reichert, M., Fichtner, D., Lenk, M., Morin, T., Olesen, N.J., Skall, H.F., Lee, P.Y., Zheng, S., Monaghan, S., Reiche, S., Fuchs, W., Kotler, M., Way, K., Brauer, G., Bottcher, K., Kappe, A., Kielpinska, J., 2017. Validation of a KHV antibody enzyme-linked immunosorbent assay (ELISA). J Fish Dis 40(11), 1511-1527.
Boutier, M., Gao, Y., Vancsok, C., Suarez, N.M., Davison, A.J., Vanderplasschen, A., 2017. Identification of an essential virulence gene of cyprinid herpesvirus 3. Antiviral Res 145, 60-69.
Bretzinger, A., Fischer-Scherl, T., Oumouna, M., Hoffmann, R., Truyen, U., 1999. Mass mortalities in koi, Cyprinus carpio, associated with gill and skin disease. Bull. Eur. Ass. Fish Pathol 5(19), 182.
Costes, B., Fournier, G., Michel, B., Delforge, C., Raj, V.S., Dewals, B., Gillet, L., Drion, P., Body, A., Schynts, F., Lieffrig, F., Vanderplasschen, A., 2008. Cloning of the koi herpesvirus genome as an infectious bacterial artificial chromosome demonstrates that disruption of the thymidine kinase locus induces partial attenuation in Cyprinus carpio koi. J Virol 82(10), 4955-4964.
Cui, L.C., Guan, X.T., Liu, Z.M., Tian, C.Y., Xu, Y.G., 2015. Recombinant lactobacillus expressing G protein of spring viremia of carp virus (SVCV) combined with ORF81 protein of koi herpesvirus (KHV): A promising way to induce protective immunity against SVCV and KHV infection in cyprinid fish via oral vaccination. Vaccine 33(27), 3092-3099.
Davison, A.J., Eberle, R., Ehlers, B., Hayward, G.S., McGeoch, D.J., Minson, A.C., Pellett, P.E., Roizman, B., Studdert, M.J., Thiry, E., 2009. The order Herpesvirales. Arch Virol 154(1), 171-177.
Davison, A.J., Kurobe, T., Gatherer, D., Cunningham, C., Korf, I., Fukuda, H., Hedrick, R.P., Waltzek, T.B., 2013. Comparative genomics of carp herpesviruses. J Virol 87(5), 2908-2922.
Eisenberg, R.J., Atanasiu, D., Cairns, T.M., Gallagher, J.R., Krummenacher, C., Cohen, G.H., 2012. Herpes virus fusion and entry: a story with many characters. Viruses 4(5), 800-832.
Fuchs, W., Fichtner, D., Bergmann, S.M., Mettenleiter, T.C., 2011. Generation and characterization of koi herpesvirus recombinants lacking viral enzymes of nucleotide metabolism. Arch Virol 156(6), 1059-1063.
Fuchs, W., Granzow, H., Dauber, M., Fichtner, D., Mettenleiter, T.C., 2014. Identification of structural proteins of koi herpesvirus. Arch Virol 159(12), 3257-3268.
Fuchs, W., Mettenleiter, T.C., 1996. DNA sequence and transcriptional analysis of the UL1 to UL5 gene cluster of infectious laryngotracheitis virus. J Gen Virol 77 ( Pt 9), 2221-2229.
Fuchs, W., Mettenleiter, T.C., 1999. DNA sequence of the UL6 to UL20 genes of infectious laryngotracheitis virus and characterization of the UL10 gene product as a nonglycosylated and nonessential virion protein. J Gen Virol 80 ( Pt 8), 2173-2182.
Gao, Y., Suarez, N.M., Wilkie, G.S., Dong, C., Bergmann, S., Lee, P.A., Davison, A.J., Vanderplasschen, A.F.C., Boutier, M., 2018. Genomic and biologic comparisons of cyprinid herpesvirus 3 strains. Vet Res 49(1), 40.
ACCEPTED MANUSCRIP
T

20
Hedrick, R.P., Marty, G.D., Nordhausen, R.W., M.J., K., Bercovier, H., Eldar, A., 1999. A herpesvirus associated with mass mortality of juvenile and adult koi, a strain of a common carp. Fish Health Newsletter 7(27), 44-57.
Hoper, D., Freuling, C.M., Muller, T., Hanke, D., von Messling, V., Duchow, K., Beer, M., Mettenleiter, T.C., 2015. High definition viral vaccine strain identity and stability testing using full-genome population data--The next generation of vaccine quality control. Vaccine 33(43), 5829-5837.
Klupp, B.G., Granzow, H., Mettenleiter, T.C., 2000. Primary envelopment of pseudorabies virus at the nuclear membrane requires the UL34 gene product. J Virol 74(21), 10063-10073.
Mettenleiter, T.C., 1989. Glycoprotein gIII deletion mutants of pseudorabies virus are impaired in virus entry. Virology 171(2), 623-625.
Mettenleiter, T.C., 2002. Herpesvirus assembly and egress. J Virol 76(4), 1537-1547. Michel, B., Leroy, B., Stalin Raj, V., Lieffrig, F., Mast, J., Wattiez, R., Vanderplasschen, A.F., Costes, B.,
2010. The genome of cyprinid herpesvirus 3 encodes 40 proteins incorporated in mature virions. J Gen Virol 91(Pt 2), 452-462.
Mletzko, A., Amtmann, A., Bergmann, S., Lee, P., Christian, J., Buchholz, R., Becker, A., 2017. Inoculation of cyprinid herpesvirus 3 (CyHV-3) on common carp brain cells-influence of process parameters on virus yield. In Vitro Cell Dev Biol Anim 53(7), 579-585.
Neukirch, M., Böttcher, K., Bunnajirakul, S., 1999. Isolation of a virus from koi with altered gills. Bulletin of the European Association of Fish Pathologists 19, 221–224.
Pavlova, S.P., Veits, J., Keil, G.M., Mettenleiter, T.C., Fuchs, W., 2009. Protection of chickens against H5N1 highly pathogenic avian influenza virus infection by live vaccination with infectious laryngotracheitis virus recombinants expressing H5 hemagglutinin and N1 neuraminidase. Vaccine 27(5), 773-785.
Ronen, A., Perelberg, A., Abramowitz, J., Hutoran, M., Tinman, S., Bejerano, I., Steinitz, M., Kotler, M., 2003. Efficient vaccine against the virus causing a lethal disease in cultured Cyprinus carpio. Vaccine 21(32), 4677-4684.
Rosenkranz, D., Klupp, B.G., Teifke, J.P., Granzow, H., Fichtner, D., Mettenleiter, T.C., Fuchs, W., 2008. Identification of envelope protein pORF81 of koi herpesvirus. J Gen Virol 89(Pt 4), 896-900.
Sathiyamoorthy, K., Chen, J., Longnecker, R., Jardetzky, T.S., 2017. The COMPLEXity in herpesvirus entry. Curr Opin Virol 24, 97-104.
Schröder, L., Klafack, S., Bergmann, S., Fichtner, D., Lee, P., Mettenleiter, T.C., Fuchs, W., 2018. Generation of potential koi herpesvirus live vaccines by simultaneous deletion of the viral thymdine kinase and dUTPase genes. J Gen Virol, doi: 10.1099/jgv.0.001148.
Vallbracht, M., Rehwaldt, S., Klupp, B.G., Mettenleiter, T.C., Fuchs, W., 2017. Functional Relevance of the N-Terminal Domain of Pseudorabies Virus Envelope Glycoprotein H and Its Interaction with Glycoprotein L. J Virol 91(9).
van Oirschot, J.T., 1999. Diva vaccines that reduce virus transmission. J Biotechnol 73(2-3), 195-205. Vancsok, C., Penaranda, M.M.D., Raj, V.S., Leroy, B., Jazowiecka-Rakus, J., Boutier, M., Gao, Y., Wilkie,
G.S., Suarez, N.M., Wattiez, R., Gillet, L., Davison, A.J., Vanderplasschen, A.F.C., 2017. Proteomic and Functional Analyses of the Virion Transmembrane Proteome of Cyprinid Herpesvirus 3. J Virol 91(21).
Vrancken, R., Boutier, M., Ronsmans, M., Reschner, A., Leclipteux, T., Lieffrig, F., Collard, A., Melard, C., Wera, S., Neyts, J., Goris, N., Vanderplasschen, A., 2013. Laboratory validation of a lateral flow device for the detection of CyHV-3 antigens in gill swabs. J Virol Methods 193(2), 679-682.
Zhou, J., Xue, J., Wang, Q., Zhu, X., Li, X., Lv, W., Zhang, D., 2014a. Vaccination of plasmid DNA encoding ORF81 gene of CJ strains of KHV provides protection to immunized carp. In Vitro Cell Dev Biol Anim 50(6), 489-495.
Zhou, J.X., Wang, H., Li, X.W., Zhu, X., Lu, W.L., Zhang, D.M., 2014b. Construction of KHV-CJ ORF25 DNA vaccine and immune challenge test. J Fish Dis 37(4), 319-325.
ACCEPTED MANUSCRIP
T

21
Figure legends
Fig. 1
Construction of CyHV-3 recombinants. (A) Map of the KHV-T genome with
localizations of ORF25, ORF65, ORF148, ORF149 and direct terminal repeat
sequences (rectangles). Enlarged maps of the (B) ORF25, (C) ORF65, and (D)
ORF148/ORF149 gene regions illustrate cloning and mutagenesis. Viral ORFs
(pointed rectangles) and relevant restriction sites are shown. Non-functional,
fragmented or frameshifted ORFs are indicated by dotted lines and different shading.
Engineered restriction sites are printed in Italics. Black rectangles indicate encoded
signal peptides and transmembrane domains of the CyHV-3 glycoprotein genes.
Major parts of them were either replaced by a GFP expression cassette containing
the human cytomegalovirus immediate early promoter (P-CMV), and a
polyadenylation signal (A+), or deleted without substitution. The deleted codon
ACCEPTED MANUSCRIP
T

22
ranges are given. Designations of plasmids and resulting virus recombinants (printed
in Italics) are given at the right.
ACCEPTED MANUSCRIP
T

23
Fig. 2
PCR products of the authentic or mutated CyHV-3 genes (A) ORF25, (B) ORF65,
and (C) ORF148/ORF149 amplified from genomic DNA of the generated
recombinants obtained with primer pairs CyHV-3O25-F/R, CyHV-3O65-F/R, or
CyHV-3O148-R and CyHV-3O149-F (Fuchs et al., 2014). Marker DNAs (1 kb Plus
DNA ladder, Thermo Fisher Scientific), and calculated product sizes are indicated at
left and right, respectively.
ACCEPTED MANUSCRIP
T

24
Fig. 3
Western blot analyses of CCB cells infected with the indicated CyHV-3 recombinants
(MOI 1, 2 d p.i.). Blots of infected and uninfected cells were probed with monospecific
rabbit antisera against (A) pORF25, (B) pORF149, and the major capsid protein
pORF92 (lower panels). Marker proteins (PageRuler Plus Prestained Protein Ladder,
Thermo Fisher Scientific), and specifically detected viral proteins are indicated at the
left and right, respectively.
ACCEPTED MANUSCRIP
T
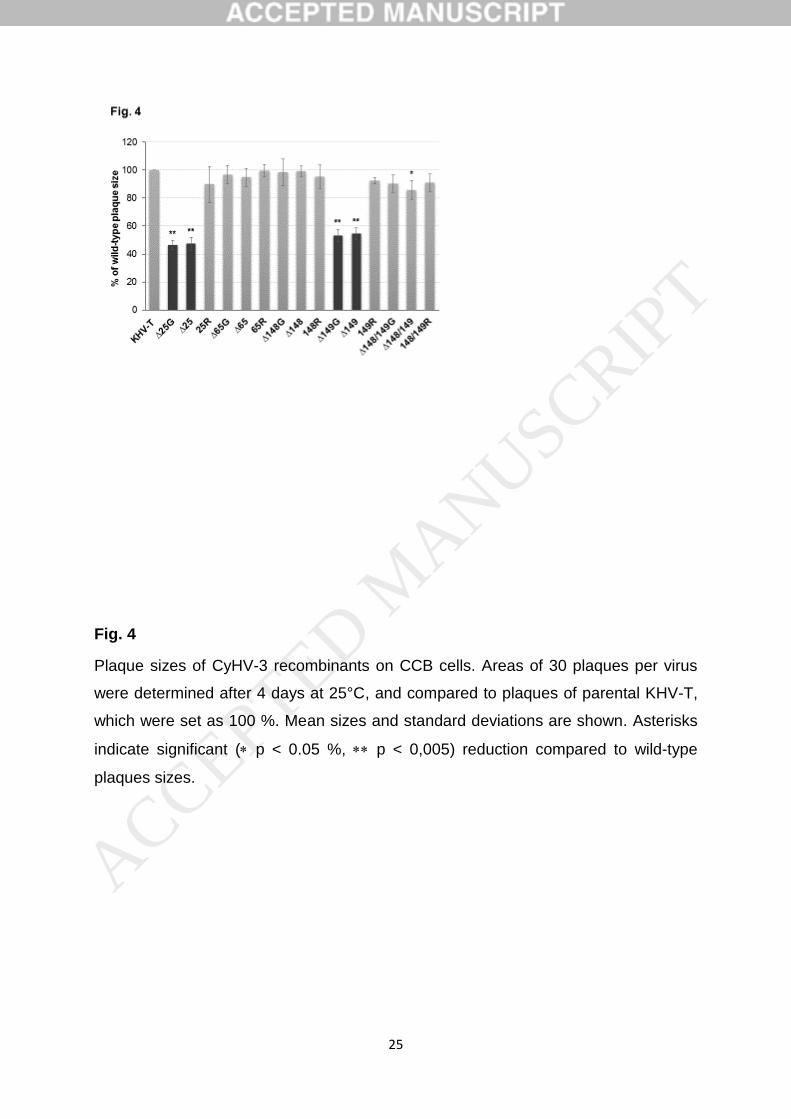
25
Fig. 4
Plaque sizes of CyHV-3 recombinants on CCB cells. Areas of 30 plaques per virus
were determined after 4 days at 25°C, and compared to plaques of parental KHV-T,
which were set as 100 %. Mean sizes and standard deviations are shown. Asterisks
indicate significant ( p < 0.05 %, p < 0,005) reduction compared to wild-type
plaques sizes.
ACCEPTED MANUSCRIP
T
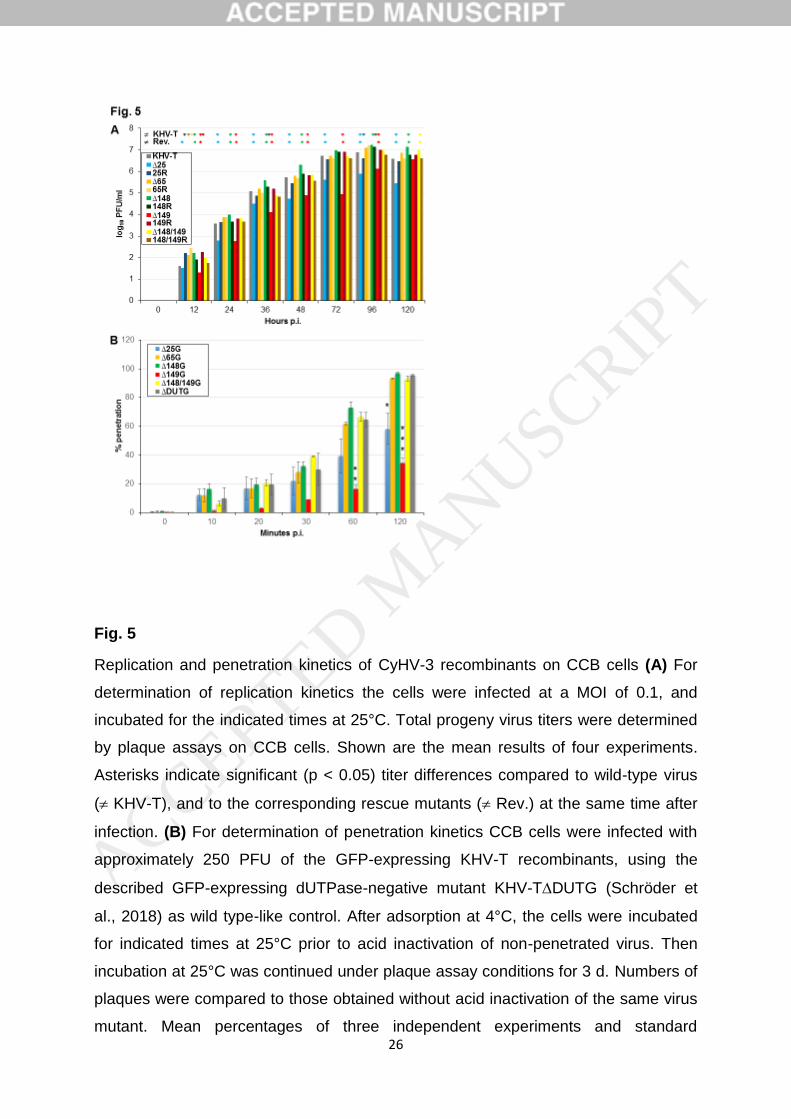
26
Fig. 5
Replication and penetration kinetics of CyHV-3 recombinants on CCB cells (A) For
determination of replication kinetics the cells were infected at a MOI of 0.1, and
incubated for the indicated times at 25°C. Total progeny virus titers were determined
by plaque assays on CCB cells. Shown are the mean results of four experiments.
Asterisks indicate significant (p < 0.05) titer differences compared to wild-type virus
( KHV-T), and to the corresponding rescue mutants ( Rev.) at the same time after
infection. (B) For determination of penetration kinetics CCB cells were infected with
approximately 250 PFU of the GFP-expressing KHV-T recombinants, using the
described GFP-expressing dUTPase-negative mutant KHV-TDUTG (Schröder et
al., 2018) as wild type-like control. After adsorption at 4°C, the cells were incubated
for indicated times at 25°C prior to acid inactivation of non-penetrated virus. Then
incubation at 25°C was continued under plaque assay conditions for 3 d. Numbers of
plaques were compared to those obtained without acid inactivation of the same virus
mutant. Mean percentages of three independent experiments and standard
ACCEPTED MANUSCRIP
T

27
deviations are shown. Asterisks indicate significantly reduced ( p < 0.1, p < 0.05,
p < 0.005) penetration rates.
ACCEPTED MANUSCRIP
T
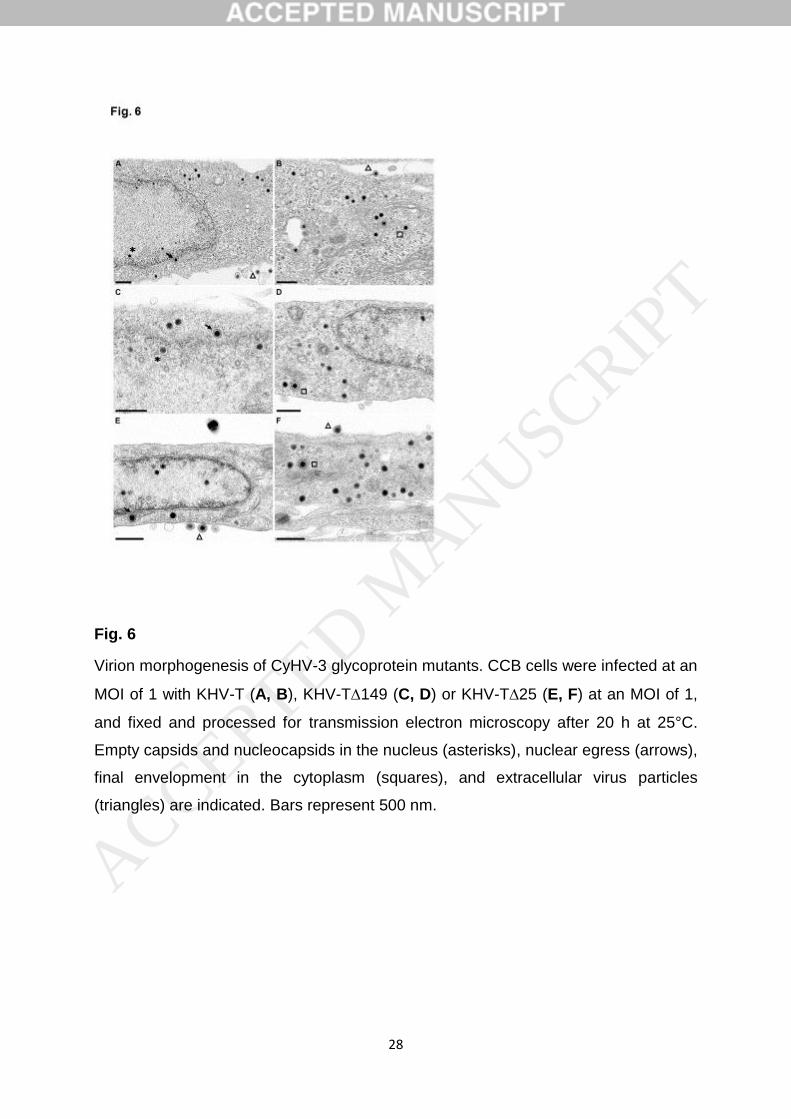
28
Fig. 6
Virion morphogenesis of CyHV-3 glycoprotein mutants. CCB cells were infected at an
MOI of 1 with KHV-T (A, B), KHV-T149 (C, D) or KHV-T25 (E, F) at an MOI of 1,
and fixed and processed for transmission electron microscopy after 20 h at 25°C.
Empty capsids and nucleocapsids in the nucleus (asterisks), nuclear egress (arrows),
final envelopment in the cytoplasm (squares), and extracellular virus particles
(triangles) are indicated. Bars represent 500 nm.
ACCEPTED MANUSCRIP
T
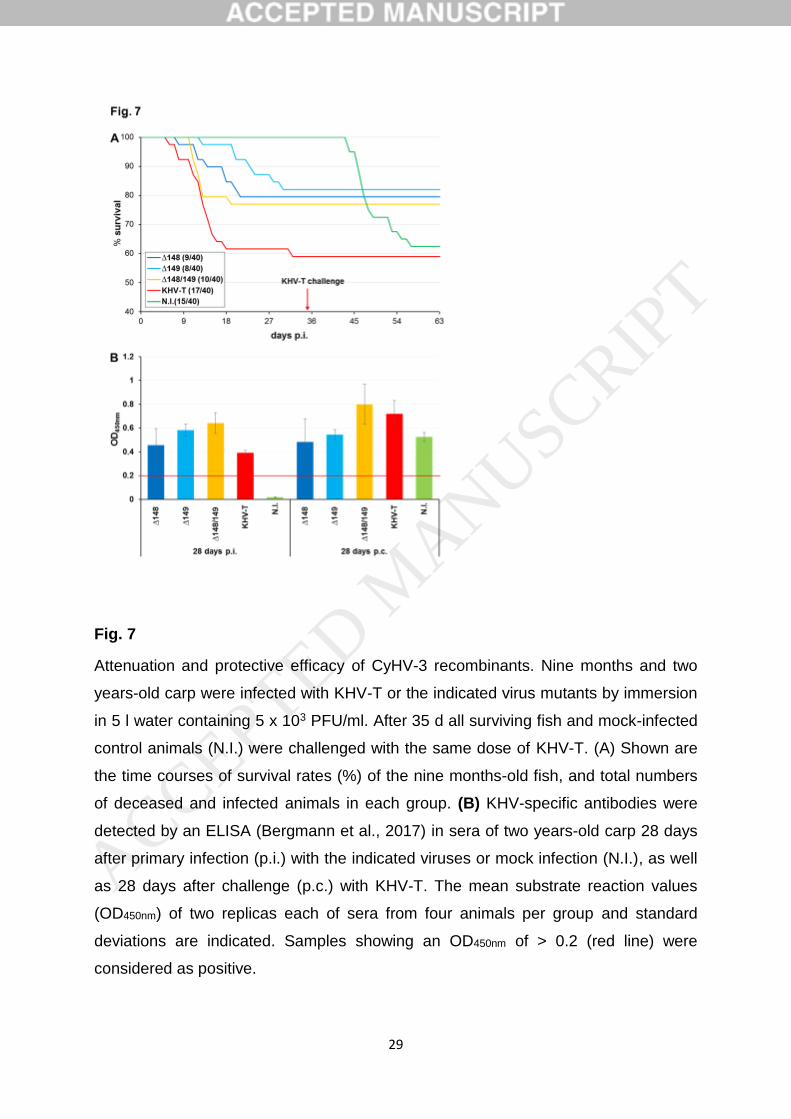
29
Fig. 7
Attenuation and protective efficacy of CyHV-3 recombinants. Nine months and two
years-old carp were infected with KHV-T or the indicated virus mutants by immersion
in 5 l water containing 5 x 103 PFU/ml. After 35 d all surviving fish and mock-infected
control animals (N.I.) were challenged with the same dose of KHV-T. (A) Shown are
the time courses of survival rates (%) of the nine months-old fish, and total numbers
of deceased and infected animals in each group. (B) KHV-specific antibodies were
detected by an ELISA (Bergmann et al., 2017) in sera of two years-old carp 28 days
after primary infection (p.i.) with the indicated viruses or mock infection (N.I.), as well
as 28 days after challenge (p.c.) with KHV-T. The mean substrate reaction values
(OD450nm) of two replicas each of sera from four animals per group and standard
deviations are indicated. Samples showing an OD450nm of > 0.2 (red line) were
considered as positive.
ACCEPTED MANUSCRIP
T

30
Tables
Table 1: PCR and sequencing primers used in this study
name Sequence nucleotide position
KTO25F 5‘-CACGAATTCTCATGTACGAGTACCTGCG-3‘ 44,755 - 44,774
KTO25R 5’-CACAAGCTTCGGAGAGCATGATGAAGAGG-3’ 48,347 - 48,366 (r)
KTO65F 5´-CACGAATTCTTTCAGTTCAACGACCCG-3´ 121,758 - 121,777
KTO65R 5´-CACAAGCTTTACCTTCTACGGCGTGG-3´ 124,935 - 124,954 (r)
KTO149R-F 5´-CACGAATTCCGAGGGAGATCAGAGTTC-3´ 253,076 - 253,094
KTO149R-R 5`-CACAAGCTTGGGATGATGTCAGCGTTG-3´ 259,046 - 259,064 (r)
Nucleotide positions refer to the sequence of the CyHV-3 TUMST1 genome
(GenBank accession # AP008984) (Aoki et al., 2007). Additional, nonmatching
nucleotides (in Italics), engineered restriction sites (bold), and reverse strand primers
(r), are indicated.
ACCEPTED MANUSCRIP
T
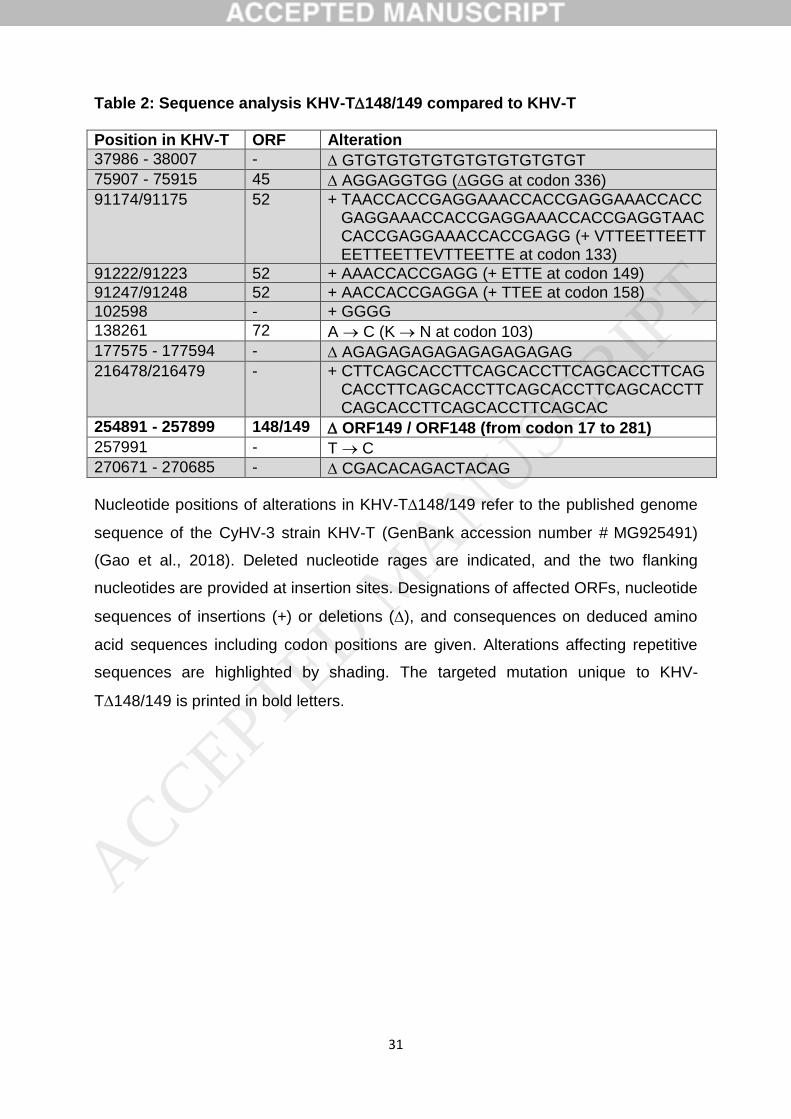
31
Table 2: Sequence analysis KHV-T148/149 compared to KHV-T
Position in KHV-T ORF Alteration
37986 - 38007 - GTGTGTGTGTGTGTGTGTGTGT
75907 - 75915 45 AGGAGGTGG (GGG at codon 336)
91174/91175 52 + TAACCACCGAGGAAACCACCGAGGAAACCACC GAGGAAACCACCGAGGAAACCACCGAGGTAAC CACCGAGGAAACCACCGAGG (+ VTTEETTEETT EETTEETTEVTTEETTE at codon 133)
91222/91223 52 + AAACCACCGAGG (+ ETTE at codon 149)
91247/91248 52 + AACCACCGAGGA (+ TTEE at codon 158)
102598 - + GGGG
138261 72 A C (K N at codon 103)
177575 - 177594 - AGAGAGAGAGAGAGAGAGAG
216478/216479 - + CTTCAGCACCTTCAGCACCTTCAGCACCTTCAG CACCTTCAGCACCTTCAGCACCTTCAGCACCTT CAGCACCTTCAGCACCTTCAGCAC
254891 - 257899 148/149 ORF149 / ORF148 (from codon 17 to 281)
257991 - T C
270671 - 270685 - CGACACAGACTACAG
Nucleotide positions of alterations in KHV-T148/149 refer to the published genome
sequence of the CyHV-3 strain KHV-T (GenBank accession number # MG925491)
(Gao et al., 2018). Deleted nucleotide rages are indicated, and the two flanking
nucleotides are provided at insertion sites. Designations of affected ORFs, nucleotide
sequences of insertions (+) or deletions (), and consequences on deduced amino
acid sequences including codon positions are given. Alterations affecting repetitive
sequences are highlighted by shading. The targeted mutation unique to KHV-
T148/149 is printed in bold letters.
ACCEPTED MANUSCRIP
T

Publikationen
36

Publikationen
67
3.3. Pubilkation III
“In cellulo evolution of Cyprinid herpesvirus 3 seems to follow a viral quasispecies-like model”
Sandro Klafack, Anna-Sophie Fiston-Lavier, Sven M. Bergmann, Saliha Hammoumi, Lars Schröder, Walter Fuchs, Angela Lusiastuti, Pei-Yu Lee, Master students, Anne-Sophie Gosselin-Grenet and Jean-Christophe Avarre
Eingereicht bei Viruses - Animal Viruses

Viruses 2018, 10, x; doi: FOR PEER REVIEW www.mdpi.com/journal/viruses
Article 1
In cellulo evolution of Cyprinid herpesvirus 3 seems 2
to follow a viral quasispecies‐like model 3
Running title: Evolution of CyHV‐3 in vitro 4
Sandro Klafack1, Anna‐Sophie Fiston‐Lavier2, Sven M. Bergmann1*, Saliha Hammoumi2, Lars 5 Schröder3, Walter Fuchs3, Angela Lusiastuti4, Pei‐Yu Lee5, Master students6**, Anne‐Sophie 6 Gosselin‐Grenet7 and Jean‐Christophe Avarre2* 7
1 Institute of Infectology, Friedrich‐Loeffer‐Institut, Federal Research Institute for Animal Health, 8 Greifswald‐Insel Riems, Germany 9 2 ISEM, IRD, CNRS, EPHE, University of Montpellier, Montpellier, France 10 3 Institute of Molecular Virology and Cell Biology, Friedrich Loeffer Institut, Federal Research Institute for 11 Animal Health, Greifswald‐Insel Riems, Germany 12 4 Research Institute for Freshwater Aquaculture and Fisheries Extension, Bogor, Indonesia 13 5 GenReach Biotechnology, Taichung City, Taiwan 14 6 Master Interactions Microorganisms/Hosts and Environment (IMHE), and Master of Sciences and Digital 15 Technology for Healthcare (SDTH), specialty Bioinformatics, Knowledge and Data (BKD), University of 16 Montpellier, Montpellier, France 17 7 DGIMI, INRA, University of Montpellier, Montpellier, France 18 * Corresponding authors: Jean‐Christophe Avarre;Institut des Sciences de l’Evolution de Montpellier,19 University of Montpellier, Place Eugène Bataillon, cc065 34095 Montpellier cedex 5, France;Jean‐20 [email protected] 21 Sven Bergmann;Federal Research Institute for Animal Health,Südufer 10, 17493 Greifswald‐Insel Riems, 22 Germany;[email protected] 23 **Master IMHE: Charles Amoyal, Hajar Amraoui, Audrey Bigourdan, Inès Boujandir, Oceane Bueno, Betty 24 Cotteux, Noëmie De San Nicolas, Romain Delattre, Mame Boucar Diouf, Rémy Dussaut, Kévin Gawron, 25 Jean‐Baptiste Imbert, Halima Mouilhi, Emilie Nifaut, Anais Pages, Lucien Platon, Sandrine Sanchez, Lena 26 Simon, Hélène Sobry, Mathilde Souche, Boris Taillefer. 27 **Master SDTH‐BKD: Edith Ndiesseu Guemgne, Olivier Sacchi, Amad Diouf, Amirouche Labib Ouzerdine, 28 Faustine Durand, Julie Brooke, Julie Cremaschi, Mathieu Massaviol, Mickael Hamouma, Mohammad 29 Salma, Morgan Soulié, Quentin Delorme, Thimothée Virgoulay, Valentin Klein, Yannick Antoine, Aurore 30 Berne, Camille Gaal. 31 32
Abstract:Viruses are able to evolve in vitro by mutation after serial passages on cell cultures, which 33 can lead to either a loss or an increase of virulence. Cyprinid herpesvirus 3 (CyHV‐3), a 295‐kb 34 double stranded DNA virus, is the etiological agent of the koi herpesvirus disease (KHVD). To 35 assess the influence of serial passages, an isolate of CyHV‐3 (KHV‐T) was passaged 99 times onto 36 common carp brain (CCB) cells, and virus virulence was evaluated during passages through 37 experimental infections of common carp. After 78 CCB passages, the isolate was much less virulent 38 than the original form; however, it partially recovered its virulence after 99 passages. A comparative 39 genomic analysis of these three forms of KHV‐T (P0, P78 and P99) revealed a limited number of 40 variations. The largest one was a deletion of 1 363 bp in the predicted ORF150, which was detected 41 in P78 but not in P99. This unexpected finding was confirmed by PCR. ORF150 thus appears as a 42 potential key player for the virulence of CyHV‐3. In addition, the results presented here primarily 43 suggest that CyHV‐3 evolves, at least in vitro, through an assemblage of haplotypes that 44 alternatively become dominant or under‐represented. 45
Keywords: Cyprinid herpesvirus 3; cell culture passages; genome evolution; haplotype 46 47

Viruses 2018, 10, x FOR PEER REVIEW 2 of 10
1. Introduction 48
Common carp (Cyprinus carpio L.) is the most produced fish in the world. With its ornamental 49 breed, the koi, it is also one of the most expensive fish. This fish is highly threatened by the Cyprinid 50 herpesvirus 3 (CyHV‐3), also called Koi Herpesvirus (KHV). Since its first report in the late 1990s [1], 51 KHV disease (KHVD) has spread to many countries worldwide [2], and is recognized as a significant 52 problem for the common carp and koi aquaculture industries [3, 4]. Together with carp pox virus 53 (Cyprinid herpesvirus 1), goldfish herpesvirus (Cyprinid herpesvirus 2) and eel herpesvirus 54 (Anguillid herpesvirus 1), CyHV‐3 clusters as a member of the genus Cyprinivirus in the family 55 Alloherpesviridae [5]. It causes mass mortality in carp aquaculture, often with more than 80% losses 56 and severe symptoms. Most common symptoms are gill necrosis, skin hemorrhage, a massive mucus 57 production, later on sandpaper skin and / or sunken eyes [1, 6]. Once infected with KHV, carps bear 58 the virus lifelong and will therefore act as carriers [7‐9]. Moreover, persistent infected carp can shed 59 the virus for the rest of their life. Then those carp or koi can infect naive fish and spread the virus. 60
The genome of CyHV‐3 was sequenced multiple times [10‐13]. With 156 open reading frames 61 (ORF) and a length of 295 kb, KHV has the biggest genome known in herpesviruses [14]. 62 Unfortunately, little is known about the function of the 156 ORFs or their importance for the 63 virulence. Because of this knowledge gap, it is difficult to target a particular ORF for vaccine 64 production [15, 16]. Thus, classical methods, like attenuation along cell culture passages [17], were 65 used to generate vaccine strains. Although virus attenuation by successive passages on cell culture 66 has proven efficient [18], no genomic and/or transcriptomic comparisons between original and 67 attenuated viral strains have been carried out so far in view of understanding the molecular 68 mechanisms of virus attenuation. It was recently shown that mutations occurring in the genome of 69 CyHV‐3 during cell culture passages could be reverted after additional passages [19]. The occurrence 70 of multiple haplotypes within a same viral specimen is now well documented [12, 20, 21], and these 71 genetic reversions could be the result of a switching dominance of one or several haplotypes over the 72 others. The present study aimed at testing this hypothesis, by the phenotypic and genomic 73 comparison of a same specimen before passage on cell cultures (P0) and after 78 passages (P78) and 74 99 passages (P99). The results presented below prove, for the first time, that CyHV‐3 evolves, at least 75 in vitro, through an assemblage of haplotypes that alternatively become dominant and under‐76 represented. This finding may have important consequences for the understanding of the evolution 77 of this virus, as well as for the development of a vaccine virus. 78
2. Materials and Methods 79
2.1. CyHV‐3 propagation onto CCB cells and virus harvest and storage 80
Common carp brain (CCB) cells [22] were grown at 20°C in minimal essential medium with 81 Earls’ salts (Invitrogen) supplemented with 10% FBS, 10mM HEPES, 2.2 g/l NaHCO (Roth), 1% non‐82 essential amino acids (Biochrom) and 0.12 g/l pyruvic acid sodium salt (Merck). Twenty four‐hour 83 old CCB cell monolayers were adsorbed for 1 hour at 20°C with an isolate of CyHV‐3 collected from 84 an infected koi in Taiwan (KHV‐T) [19]. Culture medium was then added to the KHV‐T inoculated 85 cells, which were subsequently incubated at 20°C. KHV‐T was serially passaged over 100 times. Viral 86 suspensions were collected after consecutive passages when 90% of cells showed cytopathic effects 87 (CPE), usually after 7 days. For virus harvest, flasks were frozen at ‐80°C and thawed once at room 88 temperature. The remaining suspensions were stored at ‐80°C or used directly for new inoculation 89 and adsorption. 90
2.2. Experimental infections of carp 91
The wild‐type KHV‐T (P0) and the resulting viruses after 78 (P78) and 99 (P99) passages were 92 examined for their ability to elicit both mortality and morbidity in common carp. For this purpose, 93 forty 6‐month old carp were placed in 250‐L aquaria. Prior to challenge, fish were adapted to the re‐94 circulating systems for 14 days at 20°C (+/‐ 1°C). First, samples consisting of gill swap and blood for 95

Viruses 2018, 10, x FOR PEER REVIEW 3 of 10
serum preparation were collected before infection. Carp were then immersed for 1 hour in 10‐L tanks 96 containing the respective virus at a dose of 105 TCID50/ml. After that, they were transferred back to 97 their original aquaria. Gill swaps and sera were collected every 14 days from five fish per aquarium. 98 Mortality and morbidity was recorded daily by counting dead fish and carp with symptoms, mainly 99 increased mucus production and local necrosis on the skin and the gill, respectively. Dead fish were 100 immediately examined for the presence of KHV by PCR. All experiments on animals were proceeded 101 under the strict German rules for animal welfare and legally authorized by the animal protection 102 commission of Mecklenburg, Western Pomerania, with the allowance no. 7221.3‐1‐008/18 (TV 02/18 103 KHV‐att 4). 104
2.3. Extraction of viral DNA, library preparation and genome sequencing of P78 and P99 105
Genomic DNA was extracted from both the cell pellets and cell supernatants, using the 106 Nucleospin virus extraction kit (Macherey Nagel). Cell cultures were thawed on ice, aliquoted in 107 volumes of 450 μL and centrifuged at 3000 rpm for 1 min. Supernatants were transferred into new 108 tubes and mixed with 400 μL of lysis buffer supplemented with 10 μL of proteinase K, whereas cell 109 pellets were resuspended in 400 μL of lysis buffer supplemented with 10 μL of proteinase K. Viral 110 lysis was obtained by a 15‐min incubation at 70°C. DNA was then purified following the 111 manufacturerʹs instructions, and eluted in 30 μL of PCR‐grade H2O. Purity of the obtained DNAs 112 was checked by spectrophotometry (Nanodrop 2100) while their quantity was measured using a 113 Qubit (ThermoFisher Scientific). 114
DNA libraries were prepared with the Nextera XT DNA Library Prep kit (Illumina), using 1 ng 115 of input DNA. DNA tagmentation, amplification and purification were carried out exactly as 116 specified by the provider. The average size distribution of DNA fragments was verified with a 117 BioAnalyzer 2100 (Agilent Technologies), and the concentration of each DNA library was measured 118 by qPCR, according to Illumina’s recommendations. Six replicates of samples P78 and P99 were 119 prepared by the IMHE Master students. Among these replicates, two from P78 cell pellets (P78‐1c 120 and P78‐2c), two from P78 cell supernatants (P78‐1s and P78‐2s) and two from P99 cell pellets (P99‐121 1c and P99‐2c), chosen according to their comparable final concentration (comprised between 11.3 122 and 33.7 nM), were sequenced at Montpellier Genomix platform (Montpellier, France) on a single 123 lane of a MiSeq instrument with MiSeq Reagent Kit v2 (Illumina), using 500 cycles (2 x 250 bases, 124 paired‐ends). 125
2.4. Preparation and genome sequencing of KHV‐T (P0) 126
The wild‐type form of KHV‐T had already been sequenced in a previous project (unpublished 127 data), and was used here for genome comparisons. After extraction with the Wizard Genomic DNA 128 purification kit (Promega), DNA was subjected to a specific target enrichment, as described in [12], 129 and sequenced on a HiSeq2000 platform (Illumina) together with 7 other specimens in a paired‐end 130 (2x100 bases) format, at Montpellier Genomix platform (Montpellier, France). 131
2.5. Genomic sequence analysis 132
The seven sequence datasets (P78‐1c, P78‐2c, P78‐1s, P78‐2s, P99‐1c, P99‐2c and P0) were first 133 analyzed by the students, with the help of the authors. Quality of the raw sequencing data was 134 evaluated using fastQC (version 0.11.8; https://github.com/s‐andrews/FastQC). Potential adaptor 135 sequences were removed with Trimmomatic (version 0.35) [23] and bases with a quality score > 30 136 were selected using SeqTk (version 1.3‐r106; ‐q 0.001; https://github.com/lh3/seqtk). The reads were 137 then mapped to the KHV‐J strain reference genome (accession number AP008984) with BWA‐mem 138 (version bwa V0.7.9a) [24], using the default parameters. Reads aligning with a quality lower than 30 139 (except those aligning twice in the terminal repeats of the reference genome) were filtered out. Since 140 the mapping quality of P78 and P99 replicates was comparable, replicate data were merged to end 141 up with three unique sets: P78, P99 and P0. To search for differences between the 3 genomes, calling 142 of variants was performed for the three datasets. Single nucleotide polymorphisms (SNPs) and short 143

Viruses 2018, 10, x FOR PEER REVIEW 4 of 10
indels (< 100 bp were called combining Picard (version 1.61) and GATK (version 3.5), with a minimum 144 quality filter at Q20. The effect of each variant was then assessed using the ʺbedtools intersectʺ 145 (v2.27.0) and SnpEffʺ (v4.3) tools. For both tools, the annotation version 1 from the reference genome 146 (AP008984), downloaded from https://www.ncbi.nlm.nih.gov/nuccore/AP008984.1/, was used. For 147 large variant calling (> 100 bp), read depth along the reference genome was first computed using 148 Samtools with the command “depth” (version 1.9) [25] and then compared between P78, P99 and P0. 149 Detected large variations were confirmed by visualization of the read mapping with the Integrative 150 Genomics Viewer (IGV, V2.4.7) [26], and manual curation allowed to precise their boundaries. 151 Regions of interest were finally annotated using KHV‐J reference genome (annotation release 1), and 152 putative protein motives within predicted open reading frames (ORFs) were searched with the NCBI 153 Conserved Domains (CD) search tool (https://www.ncbi.nlm.nih.gov/cdd/) [27]. 154
2.6. PCR assays 155
Two PCR assays targeting the regions comprised between nt 258055‐260647 (encompassing the 156 whole ORF150) and between nt 258428‐258927 (inside the deletion) were carried out using the 157 following primer pairs designed from P0 sequence: 5’‐GCGTCGACGGAGCATG‐3’ / 5’‐158 CGAAAGAGTAAGCCGTTGCC‐3’ and 5´‐CACAAGAGATGGACGCTCAG‐3´ / 5´‐159 GTTCTCGCCCAGCACCA‐3´. DNA extracted from P0, P78 and P99 was amplified using the GoTaq 160 G2 kit (Promega), under the following conditions: 95°C for 5 minutes, 30 cycles of 95°C for 30 seconds, 161 60°C for 30 seconds and 75°C for 3 minutes, followed by a final extension at 75°C for 5 minutes. PCR 162 products were run onto agarose gels, visualized with ethidium bromide under a UV transilluminator 163 and recovered from the gel for further Sanger sequencing. 164
2.7. Nucleotide sequence accession numbers 165
Raw sequences (fastq files) were stored in the public Sequence Read Archive (SRA) repository 166 and can be accessed under the SRP study accession XXX. 167
3. Results and discussion 168
In view of obtaining a natural attenuated live vaccine, KHV‐T was serially passaged on CCB 169 cells over 100 times at 20°C, and selected passages were tested for attenuation with common carp. 170 Among the selected passages, P78 and P99 exhibited the most interesting features. Firstly, P78 elicited 171 lower mortality and morbidity rates than the P0 (wild‐type) or P99 viruses (Figure 1). Indeed, the 172 mortality induced by P78 during an experimental infection dropped from 90% (P0) to 18% with an 173 infection dose of 105 TCID50/ml and to 0 with 104 TCID50/ml infection dose (Fig. 1A & 1C). Likewise, 174 symptoms were completely absent in the P78 group at 105 TCID50/ml, while the morbidity induced 175 by P0 at the same dose reached 90% and was mostly associated with severe symptoms (Fig. 1B & 1D), 176 consistent with what is usually observed for naturally occurring CyHV‐3 isolates [28]. The mortality 177 induced by P99 (28%) was higher than that obtained with P78. Likewise, morbidity was also much 178 higher than that recorded for P78, with a peak of 100% at 9 dpi; however, the symptoms caused by 179 P99 did not last long and essentially consisted of increased mucus (Fig. 1F). Even though in the 180 absence of replicates, it is not possible to infer whether these differences are significant, it seems that 181 P99 has partially recovered its virulence ability. 182
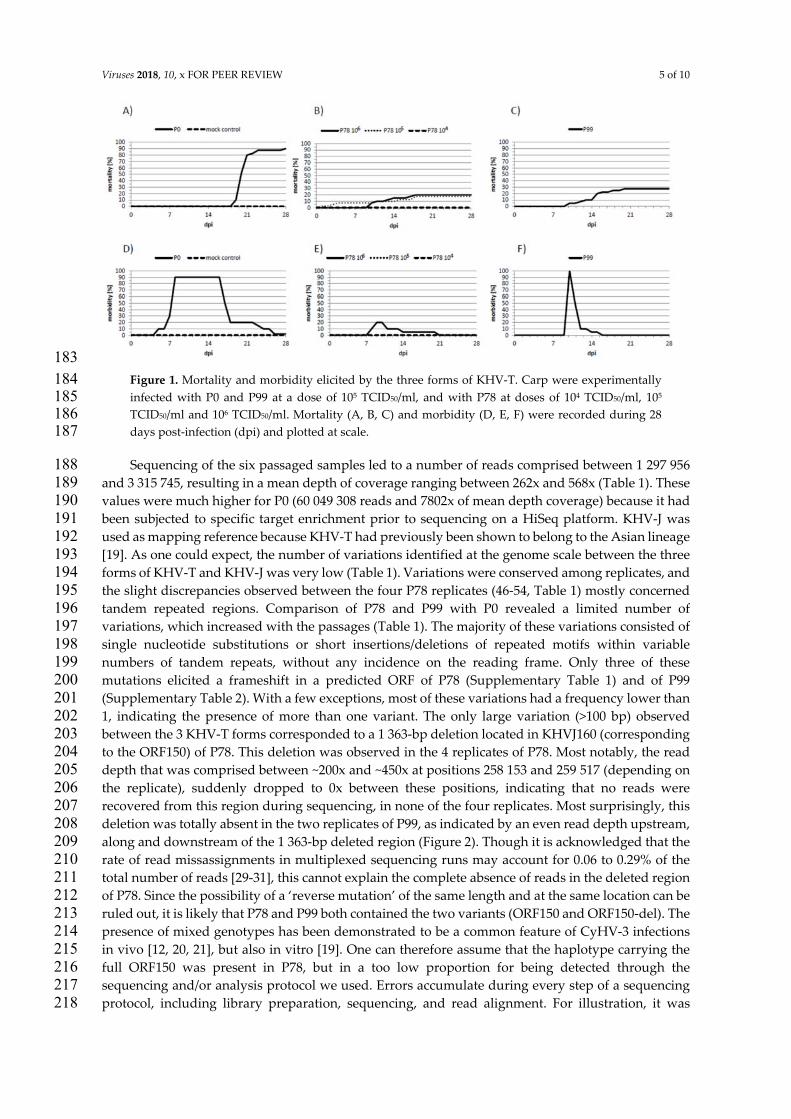
Viruses 2018, 10, x FOR PEER REVIEW 5 of 10
183 Figure 1. Mortality and morbidity elicited by the three forms of KHV‐T. Carp were experimentally 184 infected with P0 and P99 at a dose of 105 TCID50/ml, and with P78 at doses of 104 TCID50/ml, 105 185 TCID50/ml and 106 TCID50/ml. Mortality (A, B, C) and morbidity (D, E, F) were recorded during 28 186 days post‐infection (dpi) and plotted at scale. 187
Sequencing of the six passaged samples led to a number of reads comprised between 1 297 956 188 and 3 315 745, resulting in a mean depth of coverage ranging between 262x and 568x (Table 1). These 189 values were much higher for P0 (60 049 308 reads and 7802x of mean depth coverage) because it had 190 been subjected to specific target enrichment prior to sequencing on a HiSeq platform. KHV‐J was 191 used as mapping reference because KHV‐T had previously been shown to belong to the Asian lineage 192 [19]. As one could expect, the number of variations identified at the genome scale between the three 193 forms of KHV‐T and KHV‐J was very low (Table 1). Variations were conserved among replicates, and 194 the slight discrepancies observed between the four P78 replicates (46‐54, Table 1) mostly concerned 195 tandem repeated regions. Comparison of P78 and P99 with P0 revealed a limited number of 196 variations, which increased with the passages (Table 1). The majority of these variations consisted of 197 single nucleotide substitutions or short insertions/deletions of repeated motifs within variable 198 numbers of tandem repeats, without any incidence on the reading frame. Only three of these 199 mutations elicited a frameshift in a predicted ORF of P78 (Supplementary Table 1) and of P99 200 (Supplementary Table 2). With a few exceptions, most of these variations had a frequency lower than 201 1, indicating the presence of more than one variant. The only large variation (>100 bp) observed 202 between the 3 KHV‐T forms corresponded to a 1 363‐bp deletion located in KHVJ160 (corresponding 203 to the ORF150) of P78. This deletion was observed in the 4 replicates of P78. Most notably, the read 204 depth that was comprised between ~200x and ~450x at positions 258 153 and 259 517 (depending on 205 the replicate), suddenly dropped to 0x between these positions, indicating that no reads were 206 recovered from this region during sequencing, in none of the four replicates. Most surprisingly, this 207 deletion was totally absent in the two replicates of P99, as indicated by an even read depth upstream, 208 along and downstream of the 1 363‐bp deleted region (Figure 2). Though it is acknowledged that the 209 rate of read missassignments in multiplexed sequencing runs may account for 0.06 to 0.29% of the 210 total number of reads [29‐31], this cannot explain the complete absence of reads in the deleted region 211 of P78. Since the possibility of a ‘reverse mutation’ of the same length and at the same location can be 212 ruled out, it is likely that P78 and P99 both contained the two variants (ORF150 and ORF150‐del). The 213 presence of mixed genotypes has been demonstrated to be a common feature of CyHV‐3 infections 214 in vivo [12, 20, 21], but also in vitro [19]. One can therefore assume that the haplotype carrying the 215 full ORF150 was present in P78, but in a too low proportion for being detected through the 216 sequencing and/or analysis protocol we used. Errors accumulate during every step of a sequencing 217 protocol, including library preparation, sequencing, and read alignment. For illustration, it was 218
Viruses 2018, 10, x FOR PEER REVIEW 6 of 10
shown that a variant present at a frequency of 2.5% in the initial sample will not be accurately 219 detected at a read depth lower than 1000x [32, 33]. 220
221 Figure 2. IGV screenshot of the region containing the large deletion for the seven sequenced samples. 222 Each horizontal track corresponds to the reads mapped on the KHV‐J reference genome (AP008984). 223 White areas correspond to an absence of reads and dashed lines indicate the boundaries of the 224 deletion. Gene annotation of KHV‐J is shown at the bottom. KHVJ159 corresponds to ORF149 and 225 KHVJ160 to ORF150, according to Aoki et al [10]. 226
Table 1. Main features of genome comparisons. 227
Sample # readsa % mapped reads Mean coverage
[1st-3rd quartile] # variants against KHV-Jb # variants against P0b
P0 60 049 308 98.62 7802 [7868-7932] 80 - P78-1c 1 297 956 37.86 262 [202-292] 46 21 P78-2c 2 179 255 37.07 418 [301-466] 54 26 P78-1s 3 002 298 27.08 356 [248-398] 50 25 P78-2s 3 315 745 32.28 568 [448-634] 49 23 P99-1c 3 280 716 30.30 559 [404-633] 103 58 P99-2c 2 227 886 31.94 395 [302-451] 103 57
a All reads with a quality < Q30 were removed 228 b Only variations < 100 bp were considered here. 229
To verify these sequencing results, two PCR assays designed to target the full ORF150 and its 230 deleted region were applied on the three KHV‐T samples. Using the first set of primers targeting the 231 full ORF150, a unique band was observed around 1200 bp for P78, whereas for P0 and P99, the PCR 232 product had the expected length around 2593 bp (Fig. 3). Additionally, the second set of primers did 233 not yield any amplification in P78 while a 500‐bp fragment was amplified in P0 and P99, as was 234 expected with this primer set (Figure 3). Sanger sequencing of all the obtained amplicons showed 235 100% identity with P0 sequence. These PCR results clearly confirmed that this deletion did not result 236 from a sequencing artefact. Though often overlooked, the extent of information available in a sample 237 is limited by the quantity of biological material the sample contains prior to its amplification [34]. 238 Moreover, it is also known that amplification by PCR may distort the content of a sample through 239 different sources of errors such as efficiency biases, stochasticity, template switches or polymerase 240 errors [35]. These errors may have a significant impact on sequence representation, especially for 241 sequences that are present at very low copy numbers [35, 36]. As reflected by the weak percent of 242 mapped reads, the amount of viral sequences in the input DNA (ratio of CyHV‐3 / host reads) was 243 relatively low. Taking all these elements together, the results obtained here indicate that the deleted 244

Viruses 2018, 10, x FOR PEER REVIEW 7 of 10
variant was very abundant in P78 but under the detection threshold in P99. In other words, the main 245 haplotype of P78 carried the deletion of 1 363 bp whereas the dominant haplotype in P99 was the 246 same as in P0, the wild‐type KHV‐T. However, whether this deleted variant was already present in 247 P0 or arose by mutation during the successive passages needs to be further investigated. 248
249 Figure 3. PCR results using the primer set covering the whole ORF150 (1) and the primer set designed 250 inside the deletion (2). The expected amplicon sizes according to P0 are 2593 bp and 500 bp using the 251 two primer set, respectively. The two external lanes were loaded with a 100‐bp molecular weight 252 marker, and the corresponding sizes are indicated on the left. . 253
The 1 363‐bp deletion starts between genes KHVJ159 (ORF149) and KHVJ160 (ORF150), and 254 spans more than half of ORF150 (see Figure 2). As a result, the original initiation codon is missing, 255 and an alternative ORF150 could potentially begin at position 259570, providing this resulting 256 truncated ORF is transcribed. As already identified by Aoki et al [10], and confirmed by a new search 257 against the conserved domains database, ORF150 contains a RING‐HC (Really Interesting New Gene) 258 finger domain in its N‐terminal region (e‐value = 1.23x10‐7; 259 https://www.ncbi.nlm.nih.gov/protein/129560669). RING fingers, and especially of HC (C3HC4) 260 types, bind two zinc cations and are involved in ubiquitination, a potent regulator of cellular protein 261 functions such as oncogenesis, viral replication or apoptosis [37‐39]. As this RING motif is missing in 262 the truncated form of ORF150, and since this deletion is the major difference between P78 and P99, it 263 is tempting to associate this absence to the drop of virulence, or partial attenuation, observed for P78. 264 Further studies need to be done to evaluate the potential role of ORF150 in CyHV‐3 virulence. 265
To conclude, our results demonstrate that an in vitro infection by CyHV‐3 is the result of a 266 mixture of haplotypes that co‐occur, and that the ratio between these latter may dramatically varies 267 along infection cycles. To study what drives the haplotype composition during an infection will imply 268 to use other computational and/or sequencing approaches, as for instance those developed for 269 haplotype reconstruction in RNA viruses [40‐42]. Finally, our results also support increasing 270 evidence that large DNA viruses, and especially herpesviruses, may behave like viral quasispecies 271 [43] in the way that they are also composed of a mixture of haplotypes that evolve during infections. 272 This study lays the basis for better understanding the mechanisms of in vitro and in vivo evolution 273 of this very successful virus. 274
Supplementary Materials: The following are available online at www.mdpi.com/xxx/s1, Table S1: Nature and 275 position of variants between P78 and P0. All variations located in the terminal repeats appear only once. 276

Viruses 2018, 10, x FOR PEER REVIEW 8 of 10
Predicted ORFs were deduced from the genome annotation of KHV‐J (AP008984). Missense variant indicates a 277 nucleotide substitution that modifies the amino acid; synonymous variant denotes a nucleotide substitution that 278 does not modify the amino acid; frameshift variant indicates an insertion or a deletion that changes the reading 279 frame; intergenic variant points to a mutation located between two predicted open reading frames. When more 280 than two variants co‐occur at a single position, only the frequency of the most abundant alternate (from all 281 replicates) is indicated. Only variations <100 bp were listed. 282 Table S2: Nature and position of variants between P99 and P0. All variations located in the terminal repeats 283 appear only once. Predicted ORFs were deduced from the genome annotation of KHV‐J (AP008984). Missense 284 variant indicates a nucleotide substitution that modifies the amino acid; synonymous variant denotes a 285 nucleotide substitution that does not modify the amino acid; frameshift variant indicates an insertion or a 286 deletion that changes the reading frame; intergenic variant points to a mutation located between two predicted 287 open reading frames. Only variations <100 bp were listed. 288
Acknowledgments:Part of this work was supported by the ERA‐NET EMIDA funded project MOLTRAQ 289 (ANR‐11‐EMIDA‐006). Data presented in this work were produced through the genotyping and sequencing 290 facilities of ISEM (Institut des Sciences de l’Evolution ‐Montpellier), the MBB platform and Labex CeMEB (Centre 291 Méditerranéen Environnement Biodiversité). Part of the data presented here was generated during a training 292 session of the IMHE Master students of Montpellier University. The first bioinformatics analyses were realized 293 by the students from the Master Sciences and Digital Technology for Healthcare, specialty Bioinformatics, 294 Knowledge, Data of Montpellier University during the Montpellier Omics Days 2018. This is publication IRD‐295 DIVA‐ISEM 2018‐XXX. 296
Conflicts of Interest:Authors declare no conflict of interest. 297
References 298
1. Hedrick, R. P.; Gilad, O.; Yun, S.; Spangenberg, J. V.; Marty, G. D.; Nordhausen, R. W.; Kebus, M. J.; 299 Bercovier, H.; Eldar, A., A herpesvirus associated with mass mortality of juvenile and adult koi, a strain of 300 common carp. Journal of Aquatic Animal Health 2000, 12, (1), 44‐57. 301
2. Pokorova, D.; Vesely, T.; Piackova, V.; Reschova, S.; Hulova, J., Current knowledge on koi herpesvirus 302 (KHV): a review. Veterinarni Medicina 2005, 50, (4), 139‐147. 303
3. Donohoe, O. H.; Henshilwood, K.; Way, K.; Hakimjavadi, R.; Stone, D. M.; Walls, D., Identification and 304 Characterization of Cyprinid Herpesvirus‐3 (CyHV‐3) Encoded MicroRNAs. PLoS ONE 2015, 10, (4), 305 e0125434. 306
4. Haenen, O. L. M.; Way, K.; Bergmann, S. M.; Ariel, E., The emergence of koi herpesvirus and its significance 307 to European aquaculture. Bull. Eur. Assoc. Fish Pathol. 2004, 24, (6), 293‐307. 308
5. Waltzek, T. B.; Kelley, G. O.; Alfaro, M. E.; Kurobe, T.; Davison, A. J.; Hedrick, R. P., Phylogenetic 309 relationships in the family Alloherpesviridae. Dis. Aquat. Org. 2009, 84, (3), 179‐194. 310
6. Bretzinger, A.; Fischer‐Scherl, T.; Oumouna, M.; Hoffmann, R.; Truyen, U., Mass mortalities in Koi carp, 311 Cyprinus carpio, associated with gill and skin disease. Bull. Eur. Assoc. Fish Pathol. 1999, 19, (5), 182‐185. 312
7. Eide, K.; Miller‐Morgan, T.; Heidel, J.; Bildfell, R.; Jin, L., Results of total DNA measurement in koi tissue 313 by Koi Herpes Virus real‐time PCR. Journal of virological methods 2011, 172, (1‐2), 81‐4. 314
8. Eide, K. E.; Miller‐Morgan, T.; Heidel, J. R.; Kent, M. L.; Bildfell, R. J.; Lapatra, S.; Watson, G.; Jin, L., 315 Investigation of koi herpesvirus latency in koi. Journal of virology 2011, 85, (10), 4954‐62. 316
9. Reed, A. N.; Putman, T.; Sullivan, C.; Jin, L., Application of a nanoflare probe specific to a latency associated 317 transcript for isolation of KHV latently infected cells. Virus research 2015, 208, 129‐35. 318
10. Aoki, T.; Hirono, I.; Kurokawa, K.; Fukuda, H.; Nahary, R.; Eldar, A.; Davison, A. J.; Waltzek, T. B.; 319 Bercovier, H.; Hedrick, R. P., Genome sequences of three koi herpesvirus isolates representing the 320 expanding distribution of an emerging disease threatening koi and common carp worldwide. J. Virol. 2007, 321 81, (10), 5058‐5065. 322
11. Gao, Y.; Suárez, N. M.; Wilkie, G. S.; Dong, C.; Bergmann, S.; Lee, P.‐Y. A.; Davison, A. J.; Vanderplasschen, 323 A. F. C.; Boutier, M., Genomic and biologic comparisons of cyprinid herpesvirus 3 strains. Veterinary 324 Research 2018, 49, (1), 40. 325
12. Hammoumi, S.; Vallaeys, T.; Santika, A.; Leleux, P.; Borzym, E.; Klopp, C.; Avarre, J. C., Targeted genomic 326 enrichment and sequencing of CyHV‐3 from carp tissues confirms low nucleotide diversity and mixed 327 genotype infections. Peerj 2016, 4. 328

Viruses 2018, 10, x FOR PEER REVIEW 9 of 10
13. Li, W.; Lee, X.; Weng, S.; He, J.; Dong, C., Whole‐genome sequence of a novel Chinese cyprinid herpesvirus 329 3 isolate reveals the existence of a distinct European genotype in East Asia. Veterinary Microbiology 2015, 330 175, (2–4), 185‐194. 331
14. Davison, A. J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R. P.; Waltzek, T. 332 B., Comparative genomics of carp herpesviruses. Journal of virology 2013, 87, (5), 2908‐22. 333
15. Costes, B.; Fournier, G.; Michel, B.; Delforge, C.; Raj, V. S.; Dewals, B.; Gillet, L.; Drion, P.; Body, A.; Schynts, 334 F.; Lieffrig, F.; Vanderplasschen, A., Cloning of the koi herpesvirus genome as an infectious bacterial 335 artificial chromosome demonstrates that disruption of the thymidine kinase locus induces partial 336 attenuation in Cyprinus carpio koi. Journal of virology 2008, 82, (10), 4955‐64. 337
16. Boutier, M.; Ronsmans, M.; Ouyang, P.; Fournier, G.; Reschner, A.; Rakus, K.; Wilkie, G. S.; Farnir, F.; 338 Bayrou, C.; Lieffrig, F.; Li, H.; Desmecht, D.; Davison, A. J.; Vanderplasschen, A., Rational development of 339 an attenuated recombinant cyprinid herpesvirus 3 vaccine using prokaryotic mutagenesis and in vivo 340 bioluminescent imaging. PLoS pathogens 2015, 11, (2), e1004690. 341
17. Ronen, A.; Perelberg, A.; Abramowitz, J.; Hutoran, M.; Tinman, S.; Bejerano, I.; Steinitz, M.; Kotler, M., 342 Efficient vaccine against the virus causing a lethal disease in cultured Cyprinus carpio. Vaccine 2003, 21, 343 (32), 4677‐4684. 344
18. Enzmann, P.‐J.; Fichtner, D.; Schütze, H.; Walliser, G., Development of vaccines against VHS and IHN: oral 345 application, molecular marker and discrimination of vaccinated fish from infected populations. Journal of 346 Applied Ichthyology 1998, 14, (3‐4), 179‐183. 347
19. Klafack, S.; Wang, Q.; Zeng, W. W.; Wang, Y. Y.; Li, Y. Y.; Zheng, S. C.; Kempter, J.; Lee, P. Y.; Matras, M.; 348 Bergmann, S. M., Genetic Variability of Koi Herpesvirus In vitro‐A Natural Event? Front Microbiol 2017, 349 8. 350
20. Avarre, J. C.; Santika, A.; Bentenni, A.; Zainun, Z.; Madeira, J. P.; Maskur, M.; Bigarré, L.; Caruso, D., Spatio‐351 temporal analysis of Cyprinid herpesvirus 3 genetic diversity at a local scale. Journal of Fish Diseases 2012, 352 35, (10), 767‐774. 353
21. Sunarto, A.; McColl, K. A.; Crane, M. S.; Sumiati, T.; Hyatt, A. D.; Barnes, A. C.; Walker, P. J., Isolation and 354 characterization of koi herpesvirus (KHV) from Indonesia: identification of a new genetic lineage. Journal 355 of Fish Diseases 2011, 34, (2), 87‐101. 356
22. Neukirch, M.; Böttcher, K.; Bunnajirakul, S., Isolation of a virus from koi with altered gills. Bull. Eur. Assoc. 357 Fish Pathol. 1999, 19, (5), 221‐224. 358
23. Bolger, A. M.; Lohse, M.; Usadel, B., Trimmomatic: a flexible trimmer for Illumina sequence data. 359 Bioinformatics 2014, 30, (15), 2114‐2120. 360
24. Li, H.; Durbin, R., Fast and accurate short read alignment with Burrows‐Wheeler transform. Bioinformatics 361 2009, 25, (14), 1754‐60. 362
25. Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 363 Genome Project Data Processing, S., The Sequence Alignment/Map format and SAMtools. Bioinformatics 364 2009, 25, (16), 2078‐2079. 365
26. Thorvaldsdóttir, H.; Robinson, J. T.; Mesirov, J. P., Integrative Genomics Viewer (IGV): high‐performance 366 genomics data visualization and exploration. Briefings in Bioinformatics 2013, 14, (2), 178‐192. 367
27. Marchler‐Bauer, A.; Bryant, S. H., CD‐Search: protein domain annotations on the fly. Nucleic Acids Res. 368 2004, 32, (suppl_2), W327‐W331. 369
28. Bergmann, S. M.; Sadowski, J.; Kiełpiński, M.; Bartłomiejczyk, M.; Fichtner, D.; Riebe, R.; Lenk, M.; 370 Kempter, J., Susceptibility of koi × crucian carp and koi × goldfish hybrids to koi herpesvirus (KHV) 371 and the development of KHV disease (KHVD). Journal of Fish Diseases 2010, 33, (3), 267‐272. 372
29. D’Amore, R.; Ijaz, U. Z.; Schirmer, M.; Kenny, J. G.; Gregory, R.; Darby, A. C.; Shakya, M.; Podar, M.; 373 Quince, C.; Hall, N., A comprehensive benchmarking study of protocols and sequencing platforms for 16S 374 rRNA community profiling. BMC Genomics 2016, 17, (1), 55. 375
30. MacConaill, L. E.; Burns, R. T.; Nag, A.; Coleman, H. A.; Slevin, M. K.; Giorda, K.; Light, M.; Lai, K.; Jarosz, 376 M.; McNeill, M. S.; Ducar, M. D.; Meyerson, M.; Thorner, A. R., Unique, dual‐indexed sequencing adapters 377 with UMIs effectively eliminate index cross‐talk and significantly improve sensitivity of massively parallel 378 sequencing. BMC Genomics 2018, 19, 30. 379
31. Wright, E. S.; Vetsigian, K. H., Quality filtering of Illumina index reads mitigates sample cross‐talk. BMC 380 Genomics 2016, 17, 876. 381

Viruses 2018, 10, x FOR PEER REVIEW 10 of 10
32. Spencer, D. H.; Tyagi, M.; Vallania, F.; Bredemeyer, A. J.; Pfeifer, J. D.; Mitra, R. D.; Duncavage, E. J., 382 Performance of Common Analysis Methods for Detecting Low‐Frequency Single Nucleotide Variants in 383 Targeted Next‐Generation Sequence Data. The Journal of Molecular Diagnostics : JMD 2014, 16, (1), 75‐88. 384
33. Xu, C.; Nezami Ranjbar, M. R.; Wu, Z.; DiCarlo, J.; Wang, Y., Detecting very low allele fraction variants 385 using targeted DNA sequencing and a novel molecular barcode‐aware variant caller. BMC Genomics 2017, 386 18, (1), 5. 387
34. Liu, S.‐L.; Rodrigo, A. G.; Shankarappa, R.; Learn, G. H.; Hsu, L.; Davidov, O.; Zhao, L. P.; Mullins, J. I., 388 HIV Quasispecies and Resampling. Science 1996, 273, (5274), 415‐416. 389
35. Kebschull, J. M.; Zador, A. M., Sources of PCR‐induced distortions in high‐throughput sequencing data 390 sets. Nucleic Acids Res. 2015, 43, (21), e143‐e143. 391
36. Illingworth, C. J. R.; Roy, S.; Beale, M. A.; Tutill, H.; Williams, R.; Breuer, J., On the effective depth of viral 392 sequence data. Virus Evolution 2017, 3, (2), vex030. 393
37. Freemont, P. S., Ubiquitination: RING for destruction? Current Biology 2000, 10, (2), R84‐R87. 394 38. Lorick, K. L.; Jensen, J. P.; Fang, S.; Ong, A. M.; Hatakeyama, S.; Weissman, A. M., RING fingers mediate 395
ubiquitin‐conjugating enzyme (E2)‐dependent ubiquitination. Proceedings of the National Academy of 396 Sciences of the United States of America 1999, 96, (20), 11364‐11369. 397
39. Zhang, Y.; Li, L.‐F.; Munir, M.; Qiu, H.‐J., RING‐Domain E3 Ligase‐Mediated Host–Virus Interactions: 398 Orchestrating Immune Responses by the Host and Antagonizing Immune Defense by Viruses. Frontiers in 399 Immunology 2018, 9, (1083). 400
40. Nkili‐Meyong, A. A.; Bigarré, L.; Labouba, I.; Vallaeys, T.; Avarre, J. C.; Berthet, N., Contribution of Next‐401 Generation Sequencing to Aquatic and Fish Virology. Intervirology 2016, 59, (5‐6), 285‐300. 402
41. Posada‐Cespedes, S.; Seifert, D.; Beerenwinkel, N., Recent advances in inferring viral diversity from high‐403 throughput sequencing data. Virus Research 2017, 239, 17‐32. 404
42. Xue, K. S.; Moncla, L. H.; Bedford, T.; Bloom, J. D., Within‐Host Evolution of Human Influenza Virus. 405 Trends Microbiol. 2018, 26, (9), 781‐793. 406
43. Andino, R.; Domingo, E., Viral quasispecies. Virology 2015, 479‐480, 46‐51. 407 408
© 2018 by the authors. Submitted for possible open access publication under the 409 terms and conditions of the Creative Commons Attribution (CC BY) license 410 (http://creativecommons.org/licenses/by/4.0/). 411

Publikationen
68

Eigenanteil an den zur Dissertation eingereichten Publikationen
77
4. Eigenanteil an den zur Dissertation eingereichten Publikationen
4.1. Publikation I
“Generation of a potential koi herpesvirus live vaccine by simultaneous deletion
of the viral thymdine kinase and dUTPase genes”
Lars Schröder, Sandro Klafack, Sven M. Bergmann, Dieter Fichtner, Yeonhwa Jin, Pei-
Yu Lee, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs
Publiziert in Journal of General Virology
Lars Schröder
Herstellung der Deletionsplasmide
Herstellung der KHV-T Mutanten
Durchführung der in vitro Replikationsstudien
Durchführung des Tierversuches mit den KHV-T Mutanten unter fachlicher Anleitung
Probennahme und Auswertung der Tierversuchsproben mittels qPCR und ELISA von den Tierversuchsproben
Erstellung der Arbeitsversion und Fertigstellung des Manuskripts
Sandro Klafack
Beteiligung am Tierversuch mit den KHV-T Mutanten und der Probennahme
Hilfe beim Generieren der qPCR und ELISA Daten
Sven M. Bergmann
Versuchsleiter und Koordinator des Tierversuches mit den KHV-T Mutanten
Beteiligung an der Auswertung des Tierversuchs der KHV-T Mutanten

Eigenanteil an den zur Dissertation eingereichten Publikationen
78
Dieter Fichtner
Versuchsleiter und Koordinator des Tierversuches mit den KHV-I Mutanten
Auswertung des Tierversuchs der KHV-I Mutanten
Yeonhwa Jin
Beteiligung beim Tierversuch der KHV-T Mutanten
Pei-Yu Alison Lee
Bereitstellung des parentalen Virusisolates KHV-T
Dirk Höper
Durchführung und Auswertung der Sequenzanalyse des Virusgenoms der TK- und dUTPase-negativen KHV-T Mutante
Thomas C. Mettenleiter
Grundkonzeption des Forschungsprojektes
Mitarbeit an der Fertigstellung der Publikation
Walter Fuchs
Herstellung und in vitro Charakterisierung der KHV-I Mutanten
Planung des Forschungsprojektes
Mitarbeit an der Auswertung der Ergebnisse
Mitarbeit an der Fertigstellung der Publikation

Eigenanteil an den zur Dissertation eingereichten Publikationen
79
4.2. Publikation II
“Characterization of gene deletion mutants of Cyprinid herpesvirus 3 (koi herpesvirus) lacking the immunogenic envelope glycoproteins pORF25, pORF65, pORF148 and pORF149”
Lars Schröder, Sandro Klafack, Sven M. Bergmann, Pei-Yu Alison Lee, Kati
Franzke, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs
Publiziert in Virus Research
Lars Schröder
Herstellung der Deletionsplasmide und Virusmutanten
Durchführung der in vitro Replikationsstudien
Durchführung des Tierversuches unter fachlicher Anleitung
Probenentnahme und Durchführung des ELISA von den Tierversuchsproben
Erstellung der Arbeitsversion und Fertigstellung des Manuskripts
Sandro Klafack
Beteiligung beim Tierversuch und der Probennahme
Sven M. Bergmann
Versuchsleiter und Koordinator des Tierversuches
Beteiligung an der Auswertung des Tierversuchs
Pei-Yu Alison Lee
Bereitstellung des parentalen Virusisolates KHV-T
Kati Franzke
Elektronenmikroskopische Untersuchungen

Eigenanteil an den zur Dissertation eingereichten Publikationen
80
Dirk Höper
Durchführung und Auswertung der Sequenzanalyse
Thomas C. Mettenleiter
Grundkonzeption des Forschungsprojektes
Mitarbeit an der Fertigstellung der Publikation
Walter Fuchs
Planung des Forschungsprojektes
Mitarbeit an der Auswertung der Ergebnisse
Mitarbeit an der Fertigstellung der Publikation

Eigenanteil an den zur Dissertation eingereichten Publikationen
81
4.3. Publikation III
“In cellulo evolution of Cyprinid herpesvirus 3 seems to follow a viral quasispecies-like model”
Sandro Klafack, Anna-Sophie Fiston-Lavier, Sven M. Bergmann, Saliha Hammoumi,
Lars Schröder, Walter Fuchs, Angela Lusiastuti, Pei-Yu Lee, Master students, Anne-
Sophie Gosselin-Grenet and Jean-Christophe Avarre
Eingereicht bei Viruses - Animal Viruses
Sandro Klafack
Durchführung der Experimente
Generierung der Daten
Validierung der Sequenzierungsergebnisse
Mitgestaltung der Arbeit
Erstellung der Arbeitsversion des Manuskriptes
Fertigstellung des Manuskripts
Anne-Sophie Gosselin-Grenet
Durchführung der Experimente
Analyse der Sequenzdaten
Fertigstellung des Manuskripts
Sven M. Bergmann
Analyse der Sequenzdaten
Auswertung der Daten
Mitgestaltung der Arbeit
Fertigstellung des Manuskripts
Saliha Hammoumi
Analyse der Sequenzdaten
Bearbeitung der Sequenzdaten

Eigenanteil an den zur Dissertation eingereichten Publikationen
82
Lars Schröder
Durchführung der Experimente
Generierung der Daten
Mitgestaltung der Arbeit
Walter Fuchs
Mitgestaltung der Arbeit
Generierung der Daten
Angela Lusiastuti
Charakterisierung von KHV
Durchführung der Experimente
Pei-Yu Lee
Bereitstellung des parentalen Virusisolates KHV-T
Fertigstellung des Manuskripts
Master students
Hilfe bei der Durchführung der Experimente
Analyse der Sequenzdaten
Anna-Sophie Fiston-Lavier
Analyse der Sequenzdaten
Bearbeitung der Sequenzdaten
Fertigstellung des Manuskripts
Jean-Christophe Avarre
Durchführung der Experimente
Auswertung der Daten
Bearbeitung der Sequenzdaten
Erstellung der Arbeitsversion des Manuskriptes
Fertigstellung des Manuskripts

Eigenanteil an den zur Dissertation eingereichten Publikationen
83
____________________________________
Prof. Dr. Dr. h.c. Thomas C. Mettenleiter Datum:


Zusammenfassende Diskussion der Ergebnisse
85
5. Zusammenfassende Diskussion der Ergebnisse
Durch den unkontrollierten Handel mit infizierten Kois konnte sich das KHV weltweit
ausbreiten und stellt zunehmend eine Bedrohung für die Produktion von Nutzkarpfen,
Kois als auch für Wildfische dar. Aus diesem Grund ist die Herstellung eines
Impfstoffes von großer Bedeutung. Bislang gibt es keinen in der EU zugelassen
Impfstoff gegen die KHV Infektion. Generell ist zwar eine Impfung gegen KHV
möglich, jedoch wird die Sicherheit der verfügbaren, auf konventionell attenuierten
KHV-Isolaten basierenden Lebendimpfstoffen kritisch beurteilt. Deshalb waren wir
daran interessiert, ein kostengünstiges, schützendes und genetisch stabiles Impfvirus
herzustellen (siehe Publikation I). Der Grundgedanke bestand darin, die für den
Nukleotidstoffwechsel relevanten Gene der viralen Thymidinkinase (TK) und
Desoxyuridintriphosphatase (DUT) aus dem KHV-Genom mittels homologer
Rekombination zu deletieren, um dadurch die Virulenz des Erregers zu vermindern.
In vergleichbaren Studien mit anderen Herpesviren konnte gezeigt werden, dass TK
und DUT einen erheblichen Einfluss auf die Virulenz in vivo haben, aber entbehrlich
für die Virusreplikation in Zellkultur sind (Efstathiou et al., 1989; Jöns et al., 1997;
Kit, 1989; Schnitzlein, Winans, Ellsworth, & Tripathy, 1995). In Publikation I konnten
wir zeigen, dass auch bei KHV die Deletion der TK- und/oder DUT-Gene zu einer
signifikanten Attenuierung führt. Obwohl die generierten Virusmutanten in Zellkultur
keinerlei Replikationsdefizite im Vergleich zum Wildtypvirus zeigten, erwiesen sie
sich in Tierversuchen mit Jungkarpfen als deutlich abgeschwächt. Vor allem die mit
der Doppel-Deletionsmutante zweier unterschiedlicher KHV-Isolate, KHV-I∆TK/DUT
oder KHV-TDUT/TK, infizierten Karpfen zeigten nur noch vereinzelt schwache
klinische Symptome wie Hautläsionen und allenfalls minimale Mortalitätsraten (<
5%). Im Gegensatz dazu entwickelten Kontrolltiere, die mit den entsprechenden
Wildtypviren infizierten worden waren, fast durchweg schwere Symptome der KHV-
Infektion, die zu Mortalitätsraten von 40-60% führten. Nach anschließenden
Belastungsinfektionen mit dem Wildtypvirus erwiesen sich die immunisierten Karpfen
als geschützt und zeigten keinerlei Anzeichen einer akuten KHV-Infektion. Mit einem
für den Nachweis KHV-spezifischer Antikörper entwickelten ELISA (S. M. Bergmann
et al., 2017) konnte in den getesteten Seren aus mit den abgeschwächten
Virusmutanten immunisierten und mit Wildtyp-KHV infizierten Karpfen eine

Zusammenfassende Diskussion der Ergebnisse
86
vergleichbar starke humorale Immunantwort nachgewiesen werden, was mit der
guten Schutzwirkung der Impfung korrelierte.
Darüber hinaus wurde aus Kiemenabstrichen DNA extrahiert und mittels einer KHV-
spezifischen quantitativen real time PCR (qPCR) untersucht. Mit dieser früher
beschriebenen Methode wird ein Teil des KHV-Gens ORF89 detektiert und
amplifiziert (Gilad et al., 2004), wodurch wir in der Lage waren, nicht infizierte
Karpfen von infizierten zu unterscheiden. Hierbei fiel auf, dass in den Abstrichen von
Fischen, die mit TK-negativen Deletionsmutanten infiziert waren, weniger
Virusgenome detektiert werden konnten, als in den Abstrichen von Fischen, die mit
Wildtypvirus oder TK-Revertanten infiziert waren. Dies wies auf eine weniger
effiziente Replikation der Deletionsmutanten hin und korrelierte mit der
beobachteten Abschwächung der Virulenz. Mittels einer Multiplex-qPCR, die
zusätzlich einen zweiten, neu generierten Primer-Mix zur Amplifikation eines
Abschnitts des TK-Gens ORF55 enthielt, sollte eine Differenzierung der mit den KHV-
Mutanten immunisierten Karpfen von Wildtyp-infizierten Tieren ermöglicht werden
(genetische DIVA). Wie erwartet waren in dieser Testkomponente alle DNA-Proben
aus mit den TK- oder Doppelmutanten infizierten Fischen negativ, während in den
anderen Proben vergleichbare Virusgenom-Mengen wie bei der ORF89 Amplifikation
detektiert wurden. Zwei Wochen nach der Belastungsinfektion mit dem Wildtypvirus
waren die nachgewiesenen Mengen an viraler DNA geringer als nach einer
Primärinfektion mit virulentem KHV, was belegte, dass die Virusreplikation durch das
adaptive Immunsystem gehemmt wurde. Jedoch zeigten nach der
Belastungsinfektion auch die mit TK-negativem KHV immunisierten Karpfen positive
Reaktionen in denORF89- und TK-Gen-spezifischen PCR-Komponenten. Dieses
Resultat zeigte, dass die Impfung mit TK-negativem KHV keine sterile Immunität
vermittelte, aber ausreichte, um eine Erkrankung und Todesfälle nach der
Belastungsinfektion zu verhindern. Somit sollte die neu entwickelte Multiplex-qPCR
auch den Nachweis inapparenter Wildtyp-Infektionen in geimpften
Karpfenpopulationen erlauben. Dies ist im Hinblick auf die Kontrolle und eine
mögliche Tilgung der Seuche sehr wichtig, da persistierende oder latente Infektionen
geimpfter Tiere mit virulentem KHV das Risiko einer Übertragung auf naive Karpfen
beinhalten. Zusammenfassend zeigen die Ergebnisse aus Publikation I, dass die Gene
ORF55 (TK) und ORF123 (DUT) des KHV-Genoms für eine effiziente Virusreplikation
in der Zellkultur nicht benötigt werden, aber wichtige in vivo Virulenzfaktoren

Zusammenfassende Diskussion der Ergebnisse
87
darstellen. Darüber hinaus zeigten unsere Untersuchungen, dass die kombinierte
Deletion beider Gene zu einer ausreichenden Abschwächung des KHV führt, um die
entsprechende Mutante als Lebendvirusimpfstoff einzusetzen. Durch Nutzung des
Zellkultur-adaptierten KHV-T als Ausgangsstamm könnte ein solcher Impfstoff auch
kostengünstig produziert werden.
Zur weiteren Verbesserung der in Publikation I beschriebenen KHV-Mutanten wurde
versucht, auch eine serologisch differenzierbare Markervakzine gegen die KHV-
Infektion zu generieren. Hierfür wurden die Gene immunogener Hüllglykoproteine
der ORF25-Genfamilie ausgewählt (siehe Publikation II). Die Gene ORF25, 26, 27, 65,
148 und 149 stellen Mitglieder einer Familie von entfernt verwandten Genen dar, die
in den Cyprinid Herpesviren konserviert sind und sich vermutlich aus einem
gemeinsamen Vorfahren entwickelt haben (Aoki et al., 2007; Davison et al., 2013).
Die Gene ORF26 und ORF27 sind jedoch in manchen KHV-Isolaten durch
Leserasterverschiebungen inaktiviert (Gao et al., 2018) und wurden deshalb nicht
weiter untersucht. In Vorversuchen zeigten eukaryotische Zellen, die mit
Expressionsplasmiden für die von ORF25, ORF65, ORF148 oder ORF149 codierten
integralen TypI-Membranproteine transfiziert waren, in indirekten
Immunfluoreszenztests spezifische Reaktionen mit verschiedenen Seren aus
experimentell und natürlich KHV-infizierten Karpfen und Kois (Fuchs et al., 2014).
Aus diesem Grund wurde versucht, KHV-Deletionsmutanten für die vier Gene
herzustellen, die dann auf ihre Eignung als Markerimpfstoffe (serologische DIVA)
geprüft werden sollten. Die Versuche belegten, dass keines der vier Gene für die
Vermehrung des KHV in Zellkultur essentiell ist, da jedes von ihnen zumindest einzeln
aus dem Virusgenom deletiert werden konnte. In vitro Replikationsstudien zeigten,
dass Deletionen der Gene ORF25 oder ORF149 eine verzögerte Virusreplikation und
eine verlangsamte Ausbreitung von Zelle zu Zelle zur Folge hatten, während die
Deletion von ORF65 keinen erkennbaren Effekt hatte. Diese Ergebnisse stimmten im
Wesentlichen mit denen früherer Untersuchungen ähnlicher Mutanten überein, die
durch Mutagenese eines als artifizielles Bakterienchromosom (BAC) klonierten KHV-
Isolates aus Belgien hergestellt worden waren (Vancsok et al., 2017). Ein vorher
nicht beschriebenes interessantes Ergebnis dieser Arbeit war jedoch die
replikationsfördernde Wirkung der ORF148-Deletion, die in einer Doppel-
Deletionsmutante die nachteiligen Effekte der ORF149-Deletion fast vollständig
kompensierte. Beide Proteine weisen eine Aminosäuresequenzidentität von 37,5%

Zusammenfassende Diskussion der Ergebnisse
88
auf, was auf ähnliche Funktionen hindeutet und die Erklärung der gegensätzlichen
Deletions-Effekte erschwert. Um die Funktionen der Mitglieder der ORF25-
Genfamilie des KHV weiter aufzuklären, analysierten wir die in vitro
Penetrationskinetik der verschiedenen Deletionsmutanten unter Verwendung von
Säureinaktivierungsprotokollen, wie sie für ein anderes Herpesvirus, das
Pseudorabiesvirus, etabliert wurden (Mettenleiter, 1989). Diese Untersuchungen
zeigten, dass die Deletion der ORF25-Familienmitglieder pORF25 oder pORF149,
nicht jedoch von pORF65 oder pORF148 zu einem deutlich verlangsamten Eindringen
der Viren in die Wirtszellen führt, was auch die verzögerte Replikation und die
reduzierten Plaquegrößen von ORF25- und ORF149-negativem KHV erklären könnte.
Auch der beobachtete Penetrationsdefekt der ORF149-Deletionsmutante wurde
durch die zusätzliche Deletion von ORF148 korrigiert, was eventuell auf eine
kompetitive Hemmung von Virus-Wirtszellinteraktionen durch pORF148 hindeuten
könnte. Bei Säugetier-Alphaherpesviren wie dem Herpes-Simplex-Virus Typ 1 (HSV-
1) wurden der Viruseintritt in die Zelle durch Membranfusion und die daran
beteiligten viralen Glycoproteine (gB, gD, gH, gL) bereits intensiv untersucht
(Eisenberg et al., 2012; Sathiyamoorthy, Chen, Longnecker, & Jardetzky, 2017). Bei
den Alloherpesviren hingegen sind die beteiligten Proteine bislang nicht bekannt.
Eine zentrale Rolle der Produkte der ORF25-Genfamilie bei der Membranfusion
erscheint unwahrscheinlich, da sie im Gegensatz zu den hochkonservierten
Fusionsproteinen der Herpesviridae auf die Cyprinivirus-Gattung der
Alloherpesviridae beschränkt zu sein scheinen (Aoki et al., 2007; Davison et al.,
2013) und für die produktive Virusreplikation entbehrlich sind (Vancsok et al., 2017).
Es erscheint jedoch denkbar, dass die strukturell verwandten Genprodukte der
ORF25-Familie redundante Rezeptorbindungsproteine des KHV darstellen, die in der
Lage sind, die gleichen oder unterschiedliche Wirtszellmembranproteine zu binden
und die bisher unbekannten Alloherpesvirus-Fusionsproteine zu aktivieren. Die
Hüllglykoproteine pORF148 und pORF149 könnten möglicherweise um denselben
zellulären Rezeptor konkurrieren, wobei nur pORF149 zur anschließenden
Aktivierung der Fusionsproteine in der Lage ist. Daher könnte die Abwesenheit von
pORF148 den Eintritt in die Wirtszelle erleichtern. In Abwesenheit beider Proteine
könnten eventuell andere Rezeptor-Bindungsproteine, wie pORF25 oder pORF65, die
Funktion von pORF149 übernehmen.

Zusammenfassende Diskussion der Ergebnisse
89
Kürzlich veröffentlichte Studien zeigten, dass eine ORF25-Deletionsmutante zwar
eine deutlich reduzierte Virulenz, jedoch nicht gegen Belastungsinfektionen mit
Wildtyp-KHV schützte. Im Gegensatz dazu erwiesen sich ORF65-, ORF148- und
ORF149-negative Deletionsmutanten als unzureichend attenuiert (Vancsok et al.,
2017). In der vorliegenden Arbeit wurde nun die Virulenz und Schutzwirkung der
Doppeldeletionsmutante KHV-T∆148/149 und der entsprechenden Einzelmutanten
KHV-T∆148 und KHV-T∆149 vergleichend in Karpfen untersucht. Leider stellte sich
dabei heraus, dass auch die kombinierte Deletion von pORF148 und pORF149 keine
ausreichende Abschwächung der Virulenz von KHV bewirkte, sondern im Vergleich zu
den Einzelmutanten sogar zu einer leichten Virulenz-Steigerung führte. Ein positives
Resultat war jedoch, dass die beiden Hüllglykoproteine für die Induktion einer
schützenden Immunantwort gegen eine nachfolgende KHV-Belastungsinfektion der
überlebenden Fische entbehrlich waren. Auch im ELISA konnte keine Reduktion der
KHV-spezifischen Serumantikörpertiter nach Infektion mit KHV-T148/149 oder den
Einzelmutanten im Vergleich zur Wildtypinfektion festgestellt werden. Aufgrund
dieser Daten erscheint es möglich, dass eine kombinierte Deletion von immunogenen,
aber für die Schutzwirkung nicht essentiellen Glykoproteingenen der ORF25 Familie
mit echten Virulenzfaktoren wie den TK- und DUT-Genen die Entwicklung
funktionierender DIVA-Impfstoffe erlaubt. Für die differentielle serologische
Diagnostik könnten eventuell kompetitive ELISA Tests unter Nutzung verfügbarer
monoklonaler Antikörper (mAb) gegen pORF65 oder pORF149 (Fuchs et al., 2014)
entwickelt werden. Ob solche Mehrfachmutanten immer noch eine schützende
Immunität gegen die KHV-Infektion vermitteln, und ob die Entwicklung zuverlässiger
und praxistauglicher Tests für eine serologische Differenzierung von geimpften und
Wildtypvirus-infizierten Karpfen prinzipiell möglich ist, muss durch weitere
Tierversuche und in vitro Experimente geklärt werden.
Parallel zu den Versuchen, mit gentechnischen Methoden einen abgeschwächten
KHV-Lebendimpfstoff zu generieren, wurde auch versucht, das gleiche Ziel durch
serielle Zellkulturpassage des virulenten Wildtypvirus zu erreichen. Hierzu wurde in
der in Publikation III dargestellten Studie ein Isolat von KHV-T über hundertmal bei
20°C in einer aus dem Karpfengehirn gewonnenen, nicht-neuronalen Zelllinie (CCB =
common carp brain) passagiert. Anschließend wurde die Virulenz des Virus nach
unterschiedlicher Passagenzahl durch experimentelle Infektionen von Karpfen
getestet. Bedingt durch die Homogenität der Zellkulturen und die Abwesenheit einer

Zusammenfassende Diskussion der Ergebnisse
90
Immunabwehr können sich Viren in vitro durch spontane Mutationen anders
entwickeln als im natürlichen Wirtsorganismus, was zu einem Verlust oder in
Einzelfällen auch zu einer Erhöhung der Virulenz führen kann. Durch eine
vergleichende genomische Analyse verschiedener Passagen sollten solche
Veränderungen festgestellt und so mögliche, bislang unbekannte Virulenzfaktoren
des KHV identifiziert werden. Schon in früheren Studien wurde diese klassische
Methode zur Attenuierung von KHV und anderen Fischpathogenen durch
Zellkulturpassagen angewandt, um potenzielle Impfstoffe gegen die entsprechenden
Erreger zu erhalten (Enzmann, Fichtner, Schütze, & Walliser, 1998; Ronen et al.,
2003). Allerdings wurden bislang kaum Genomvergleiche zwischen den
ursprünglichen und den nach Passage abgeschwächten KHV Isolaten durchgeführt,
um die molekularen Mechanismen der Attenuierung zu verstehen. Von den in
Publikation III durch Infektion von Karpfen geprüften CCB-Zellpassagen von KHV-T
zeigten P78 und P99 die interessantesten phänotypischen Veränderungen. Die
Passage 78 verursachte deutlich niedrigere Mortalitäts- und Morbiditätsraten als das
Ausgangsisolat P0 (Wildtypvirus). In einem Parallelversuch lag durch P78 induzierte
Mortalität bei 18% gegenüber 90% mit P0. Allerdings verursachte P99 wieder eine
höhere Mortalität von 28 %, sowie auch eine erhöhte Morbidität der Fische, was auf
eine Virulenzsteigerung hindeutete. Die vergleichende Genomanalyse dieser drei
KHV-T-Passagen (P0, P78 und P99) zeigte eine begrenzte Anzahl von Veränderungen
gegenüber dem virulenten Ausgangsvirus. Am auffälligsten war eine Deletion von
1363 bp, die einen großen Teil des ORF150 einschloss, und in P78, aber nicht in P99
oder P0 nachgewiesen wurde. Dieser unerwartete Befund der NGS (next generation
sequencing) Analyse des KHV Genoms konnte durch PCR-Amplifikation des
authentischen und mutierten ORF150 aus Virus DNA der jeweiligen Passagen bestätigt
werden. Vor kurzem konnte schon einmal gezeigt werden, dass Mutationen, die im
Genom von KHV während Zellkulturpassagen aufgetreten waren, nach weiteren
Passagen wieder verschwanden (Klafack et al., 2017). Da im aktuellen Fall jedoch
eine irreversible Deletion singulärer KHV-Genomsequenzen festgestellt wurde, ist
davon auszugehen, dass auch in der Viruspopulation von Passage 78 noch geringe,
nicht detektierbare Mengen des Wildtypgenoms vorhanden waren, welches in der
Folge wieder die Oberhand gewann. Dennoch deuten die Ergebnisse darauf hin, dass
ORF150 für einen Virulenzfaktor von KHV codieren könnte. Die Funktion des
vorhergesagten ORF150 Proteins ist bislang nicht bekannt, jedoch enthält es eine

Zusammenfassende Diskussion der Ergebnisse
91
RING Finger-Domäne (Aoki et al., 2007), wie sie vor allem in regulatorischen
Proteinen gefunden wird, die über ihren Zinkfinger mit Nukleinsäuren oder anderen
Proteinen interagieren. Daher erscheint eine Rolle von ORF150 bei der
Virusreplikation von KHV ebenso vorstellbar wie eine Interaktion mit der
Immunabwehr der Wirtszelle. Da die das RING-Motiv umfassende ORF150Deletion der
markanteste Unterschied zwischen P78 und P99 von KHV-T ist, liegt die Vermutung
nahe, dass die Funktion des Proteins mit der Virulenz von KHV korreliert. Um dies zu
prüfen, sollen nun wieder mit gentechnischen Methoden definierte ORF150 Mutanten
von KHV hergestellt und sowohl in vitro als auch in vivo charakterisiert werden.

Zusammenfassende Diskussion der Ergebnisse
92

Quellen
93
6. Quellen
Akula, S., Mohammadamin, S., & Hellman, L. (2014). Fc receptors for immunoglobulins and their
appearance during vertebrate evolution. PLoS One, 9(5), e96903. doi:10.1371/journal.pone.0096903
Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K. and Walter, P. (2002). Molecular biology of the cell (Vol. 4). New York: Garland Science.
Antinone, S. E., Shubeita, G. T., Coller, K. E., Lee, J. I., Haverlock‐Moyns, S., Gross, S. P., & Smith, G. A. (2006). The Herpesvirus capsid surface protein, VP26, and the majority of the tegument proteins are dispensable for capsid transport toward the nucleus. J Virol, 80(11), 5494‐5498. doi:10.1128/JVI.00026‐06
Aoki, T., Hirono, I., Kurokawa, K., Fukuda, H., Nahary, R., Eldar, A., . . . Hedrick, R. P. (2007). Genome sequences of three koi herpesvirus isolates representing the expanding distribution of an emerging disease threatening koi and common carp worldwide. J Virol, 81(10), 5058‐5065. doi:10.1128/JVI.00146‐07
Ayana Perelberg, Margarita Smirnov, Marina Hutoran, Ariel Diamant, Yitzhak Bejerano, & Kotler, M. (2003). Epidemiological description of an new viral disease afflicting cultured Cyprinus carpio in Israel. The Israeli Journal of Aquaculture, 1(55), 5‐12.
Baumer, A., Fabian, M., Wilkens, M. R., Steinhagen, D., & Runge, M. (2013). Epidemiology of cyprinid herpesvirus‐3 infection in latently infected carp from aquaculture. Dis Aquat Organ, 105(2), 101‐108. doi:10.3354/dao02604
Bechtel, J. T., Winant, R. C., & Ganem, D. (2005). Host and viral proteins in the virion of Kaposi's sarcoma‐associated herpesvirus. J Virol, 79(8), 4952‐4964. doi:10.1128/JVI.79.8.4952‐4964.2005
Beck, G. a. G. S. H. (2007). Immunity and the invertebrates (pp. 60‐66). Bercovier, H., Fishman, Y., Nahary, R., Sinai, S., Zlotkin, A., Eyngor, M., . . . Hedrick, R. P. (2005).
Cloning of the koi herpesvirus (KHV) gene encoding thymidine kinase and its use for a highly sensitive PCR based diagnosis. BMC Microbiol, 5, 13. doi:10.1186/1471‐2180‐5‐13
Bergmann, S., & Kempter, J. (2011). Detection of koi herpesvirus (KHV) after re‐activation in persistently infected common carp (Cyprinus carpio L.) using non‐lethal sampling methods. Bulletin‐ European Association of Fish Pathologists, 31(3), 92‐100.
Bergmann, S. M., Sadowski, J., Kielpinski, M., Bartlomiejczyk, M., Fichtner, D., Riebe, R., . . . Kempter, J. (2010). Susceptibility of koi x crucian carp and koi x goldfish hybrids to koi herpesvirus (KHV) and the development of KHV disease (KHVD). J Fish Dis, 33(3), 267‐272. doi:10.1111/j.1365‐2761.2009.01127.x
Bergmann, S. M., Wang, Q., Zeng, W., Li, Y., Wang, Y., Matras, M., . . . Kielpinska, J. (2017). Validation of a KHV antibody enzyme‐linked immunosorbent assay (ELISA). J Fish Dis, 40(11), 1511‐1527. doi:10.1111/jfd.12621
Boivin, G., Coulombe, Z., & Rivest, S. (2002). Intranasal herpes simplex virus type 2 inoculation causes a profound thymidine kinase dependent cerebral inflammatory response in the mouse hindbrain. Eur J Neurosci, 16(1), 29‐43.
Boutier, M., Ronsmans, M., Ouyang, P., Fournier, G., Reschner, A., Rakus, K., . . . Vanderplasschen, A. (2015). Rational development of an attenuated recombinant cyprinid herpesvirus 3 vaccine using prokaryotic mutagenesis and in vivo bioluminescent imaging. PLoS Pathog, 11(2), e1004690. doi:10.1371/journal.ppat.1004690
Bowden, T. J., Cook, P., & Rombout, J. H. (2005). Development and function of the thymus in teleosts. Fish Shellfish Immunol, 19(5), 413‐427. doi:10.1016/j.fsi.2005.02.003
Boyton, R. J., & Openshaw, P. J. (2002). Pulmonary defences to acute respiratory infection. Br Med Bull, 61, 1‐12.

Quellen
94
Bretzinger, A., Fischer‐Scherl, T., Oumouna, M., Hoffmann, R., & Truyen, U. (1999). Mass mortalities in koi, Cyprinus carpio, associated with gill and skin disease. Bull. Eur. Ass. Fish Pathol, 5(19), 182.
Brooks, G. F., Carroll, K. C., Butel, J. S., Morse, S. A., & Mietzner, T. A. (2013). Jawetz, Melnick and Adelberg's Medical Microbiology (Vol. 26).
Chen, K., Xu, W., Wilson, M., He, B., Miller, N. W., Bengten, E., . . . Cerutti, A. (2009). Immunoglobulin D enhances immune surveillance by activating antimicrobial, proinflammatory and B cell‐stimulating programs in basophils. Nat Immunol, 10(8), 889‐898. doi:10.1038/ni.1748
Cheung, A. K. (1991). Cloning of the latency gene and the early protein 0 gene of pseudorabies virus. J Virol, 65(10), 5260‐5271.
Costes, B., Fournier, G., Michel, B., Delforge, C., Raj, V. S., Dewals, B., . . . Vanderplasschen, A. (2008). Cloning of the koi herpesvirus genome as an infectious bacterial artificial chromosome demonstrates that disruption of the thymidine kinase locus induces partial attenuation in Cyprinus carpio koi. J Virol, 82(10), 4955‐4964. doi:10.1128/JVI.00211‐08
Davison, A. J. (2002). Evolution of the herpesviruses. Vet Microbiol, 86(1‐2), 69‐88. Davison, A. J., Eberle, R., Ehlers, B., Hayward, G. S., McGeoch, D. J., Minson, A. C., . . . Thiry, E. (2009).
The order Herpesvirales. Arch Virol, 154(1), 171‐177. doi:10.1007/s00705‐008‐0278‐4 Davison, A. J., Kurobe, T., Gatherer, D., Cunningham, C., Korf, I., Fukuda, H., . . . Waltzek, T. B. (2013).
Comparative genomics of carp herpesviruses. J Virol, 87(5), 2908‐2922. doi:10.1128/JVI.03206‐12
Efstathiou, S., Kemp, S., Darby, G., & Minson, A. C. (1989). The role of herpes simplex virus type 1 thymidine kinase in pathogenesis. J Gen Virol, 70 ( Pt 4), 869‐879. doi:10.1099/0022‐1317‐70‐4‐869
Eisenberg, R. J., Atanasiu, D., Cairns, T. M., Gallagher, J. R., Krummenacher, C., & Cohen, G. H. (2012). Herpes virus fusion and entry: a story with many characters. Viruses, 4(5), 800‐832. doi:10.3390/v4050800
Ellis, A. E. (2001). Innate host defense mechanisms of fish against viruses and bacteria. Dev Comp Immunol, 25(8‐9), 827‐839.
Enzmann, P.‐J., Fichtner, D., Schütze, H., & Walliser, G. (1998). Development of vaccines against VHS and IHN: oral application, molecular marker and discrimination of vaccinated fish from infected populations. Journal of Applied Ichthyology, 14((3‐4)), 179‐183.
FAQ. (2014). Fishery and Aquaculture Statics. Feldman, L. T., Demarchi, J. M., Ben‐Porat, T., & Kaplan, A. S. (1982). Control of abundance of
immediate‐early mRNA in herpesvirus (pseudorabies)‐infected cells. Virology, 116(1), 250‐263.
Fischer, U., Koppang, E. O., & Nakanishi, T. (2013). Teleost T and NK cell immunity. Fish Shellfish Immunol, 35(2), 197‐206. doi:10.1016/j.fsi.2013.04.018
Fraser, N. W., Lawrence, W. C., Wroblewska, Z., Gilden, D. H., & Koprowski, H. (1981). Herpes simplex type 1 DNA in human brain tissue. Proc Natl Acad Sci U S A, 78(10), 6461‐6465.
Freuling, C. M., Müller, T. F., & Mettenleiter, T. C. (2017). Vaccines against pseudorabies virus (PrV). Vet Microbiol, 206, 3‐9. doi:10.1016/j.vetmic.2016.11.019
Fuchs, W., Fichtner, D., Bergmann, S. M., & Mettenleiter, T. C. (2011). Generation and characterization of koi herpesvirus recombinants lacking viral enzymes of nucleotide metabolism. Arch Virol, 156(6), 1059‐1063. doi:10.1007/s00705‐011‐0953‐8
Fuchs, W., Granzow, H., Dauber, M., Fichtner, D., & Mettenleiter, T. C. (2014). Identification of structural proteins of koi herpesvirus. Arch Virol, 159(12), 3257‐3268. doi:10.1007/s00705‐014‐2190‐4
Fuchs, W., Granzow, H., Klupp, B. G., Kopp, M., & Mettenleiter, T. C. (2002). The UL48 tegument protein of pseudorabies virus is critical for intracytoplasmic assembly of infectious virions. J Virol, 76(13), 6729‐6742.
Füllner, D. G., Pfeifer, M., & Langner, N. (2007). Karpfenteichwirtschaft‐Bewirtschaftung von Karpfenteichen. Gute fachliche Praxis: Sächsische Landesanstalt für Landwirtschaft.

Quellen
95
Galloway, D. A., Fenoglio, C. M., & McDougall, J. K. (1982). Limited transcription of the herpes simplex virus genome when latent in human sensory ganglia. J Virol, 41(2), 686‐691.
Gao, Y., Suarez, N. M., Wilkie, G. S., Dong, C., Bergmann, S., Lee, P. A., . . . Boutier, M. (2018). Genomic and biologic comparisons of cyprinid herpesvirus 3 strains. Vet Res, 49(1), 40. doi:10.1186/s13567‐018‐0532‐z
Gilad, O., Yun, S., Zagmutt‐Vergara, F. J., Leutenegger, C. M., Bercovier, H., & Hedrick, R. P. (2004). Concentrations of a Koi herpesvirus (KHV) in tissues of experimentally infected Cyprinus carpio koi as assessed by real‐time TaqMan PCR. Dis Aquat Organ, 60(3), 179‐187. doi:10.3354/dao060179
Granzow, H., Klupp, B. G., & Mettenleiter, T. C. (2005). Entry of pseudorabies virus: an immunogold‐labeling study. J Virol, 79(5), 3200‐3205. doi:10.1128/JVI.79.5.3200‐3205.2005
Granzow, H., Weiland, F., Jons, A., Klupp, B. G., Karger, A., & Mettenleiter, T. C. (1997). Ultrastructural analysis of the replication cycle of pseudorabies virus in cell culture: a reassessment. J Virol, 71(3), 2072‐2082.
Haenen, O., Way, K., Bergmann, S., & Ariel, E. (2004). The emergence of koi herpesvirus and its significance to European aquaculture. Bulletin‐ European Association of Fish Pathologists, 6(24).
Hansen, J. D., Landis, E. D., & Phillips, R. B. (2005). Discovery of a unique Ig heavy‐chain isotype (IgT) in rainbow trout: Implications for a distinctive B cell developmental pathway in teleost fish. Proc Natl Acad Sci U S A, 102(19), 6919‐6924. doi:10.1073/pnas.0500027102
Hedrick, R. P., Gilad, O., Yun, S., Spangenberg, J. V., Marty, G. D., Nordhausen, R. W., . . . Eldar, A. (2000). A Herpesvirus Associated with Mass Mortality of Juvenile and Adult Koi, a Strain of Common Carp. J Aquat Anim Health, 12(1), 44‐57. doi:10.1577/1548‐8667(2000)012<0044:AHAWMM>2.0.CO;2
Hedrick, R. P., Groff, J. M., Okihiro, M. S., & McDowell, T. S. (1990). Herpesviruses detected in papillomatous skin growths of koi carp (Cyprinus carpio). J Wildl Dis, 26(4), 578‐581. doi:10.7589/0090‐3558‐26.4.578
Hedrick, R. P., Waltzek, T. B., & Mcdowell, T. S. (2006). Susceptibility of Koi Carp, Common Carp, Goldfish, and Goldfish3 Common Carp Hybrids to Cyprinid Herpesvirus‐2 andHerpesvirus‐3. Journal of Aquatic Animal Health, 18, 26‐34.
Heldwein, E. E., & Krummenacher, C. (2008). Entry of herpesviruses into mammalian cells. Cell Mol Life Sci, 65(11), 1653‐1668. doi:10.1007/s00018‐008‐7570‐z
Homa, F. L., Huffman, J. B., Toropova, K., Lopez, H. R., Makhov, A. M., & Conway, J. F. (2013). Structure of the pseudorabies virus capsid: comparison with herpes simplex virus type 1 and differential binding of essential minor proteins. J Mol Biol, 425(18), 3415‐3428. doi:10.1016/j.jmb.2013.06.034
Honess, R. W., & Roizman, B. (1974). Regulation of herpesvirus macromolecular synthesis. I. Cascade regulation of the synthesis of three groups of viral proteins. J Virol, 14(1), 8‐19.
Honess, R. W., & Roizman, B. (1975). Regulation of herpesvirus macromolecular synthesis: sequential transition of polypeptide synthesis requires functional viral polypeptides. Proc Natl Acad Sci U S A, 72(4), 1276‐1280.
Hordvik, I. (2015). Immunoglobulin isotypes in Atlantic salmon, Salmo salar. Biomolecules, 5(1), 166‐177. doi:10.3390/biom5010166
Huang, C., & Wu, C. Y. (2004). Characterization and expression of the pseudorabies virus early gene UL54. J Virol Methods, 119(2), 129‐136. doi:10.1016/j.jviromet.2004.03.013
Ihara, S., Feldman, L., Watanabe, S., & Ben‐Porat, T. (1983). Characterization of the immediate‐early functions of pseudorabies virus. Virology, 131(2), 437‐454.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., & Charpentier, E. (2012). A programmable dual‐RNA‐guided DNA endonuclease in adaptive bacterial immunity. Science, 337(6096), 816‐821. doi:10.1126/science.1225829

Quellen
96
Johannsen, E., Luftig, M., Chase, M. R., Weicksel, S., Cahir‐McFarland, E., Illanes, D., . . . Kieff, E. (2004). Proteins of purified Epstein‐Barr virus. Proc Natl Acad Sci U S A, 101(46), 16286‐16291. doi:10.1073/pnas.0407320101
Johnson, P. A., & Everett, R. D. (1986). DNA replication is required for abundant expression of a plasmid‐borne late US11 gene of herpes simplex virus type 1. Nucleic Acids Res, 14(9), 3609‐3625.
Jöns, A., Gerdts, V., Lange, E., Kaden, V., & Mettenleiter, T. C. (1997). Attenuation of dUTPase‐deficient pseudorabies virus for the natural host. Vet Microbiol, 56(1‐2), 47‐54. doi:10.1016/S0378‐1135(96)01353‐3
Jons, A., & Mettenleiter, T. C. (1996). Identification and characterization of pseudorabies virus dUTPase. J Virol, 70(2), 1242‐1245.
Katan, M., Haigh, A., Verrijzer, C. P., van der Vliet, P. C., & O'Hare, P. (1990). Characterization of a cellular factor which interacts functionally with Oct‐1 in the assembly of a multicomponent transcription complex. Nucleic Acids Res, 18(23), 6871‐6880.
Keystone, J. S., Kozarsky, P. E., Freedman, D. O., Nothdruft, H. D., & and Conno, B. A. (2013). Travel Medicine (Vol. 3).
Kit, S. (1989). Recombinant‐derived modified‐live herpesvirus vaccines. Adv Exp Med Biol, 251, 219‐236.
Kit, S., Kit, M., & Pirtle, E. C. (1985). Attenuated properties of thymidine kinase‐negative deletion mutant of pseudorabies virus. Am J Vet Res, 46(6), 1359‐1367.
Klafack, S., Wang, Q., Zeng, W. W., Wang, Y. Y., Li, Y. Y., Zheng, S. C., . . . Bergmann, S. M. (2017). Genetic Variability of Koi Herpesvirus In vitro‐A Natural Event? . Front Microbiol, 8.
Krishnan, B. R. (2000). Current status of DNA vaccines in veterinary medicine. Adv Drug Deliv Rev, 43(1), 3‐11.
Litman, G. W., Cannon, J. P., & Dishaw, L. J. (2005). Reconstructing immune phylogeny: new perspectives. Nat Rev Immunol, 5(11), 866‐879. doi:10.1038/nri1712
Luttmann, W., Bratke, K., Küpper, M., & Myrtek, D. (2006). Der Experimentator: Immunologie (Vol. 2). München: Elsevier Gmbh.
M., J. H. a. M. (2006). Tierärtzliche Impfpraxis (Vol. 3): Joachim H. Mashoof, S., & Criscitiello, M. F. (2016). Fish Immunoglobulins. Biology (Basel), 5(4).
doi:10.3390/biology5040045 McGeoch, D. J., Rixon, F. J., & Davison, A. J. (2006). Topics in herpesvirus genomics and evolution.
Virus Res, 117(1), 90‐104. doi:10.1016/j.virusres.2006.01.002 Meeusen, E. N., Walker, J., Peters, A., Pastoret, P. P., & Jungersen, G. (2007). Current status of
veterinary vaccines. Clin Microbiol Rev, 20(3), 489‐510, table of contents. doi:10.1128/CMR.00005‐07
Mettenleiter, T. C. (1989). Glycoprotein gIII deletion mutants of pseudorabies virus are impaired in virus entry. Virology, 171(2), 623‐625.
Mettenleiter, T. C. (2002). Herpesvirus assembly and egress. J Virol, 76(4), 1537‐1547. Mettenleiter, T. C. (2008). Pseudorabies Virus. Encyclopedia of Virology (B. W. J. Mahy & M. H. V. van
Regenmortel Eds. Vol. 3). Oxford: Academic Press. Mettenleiter, T. C., Klupp, B. G., & Granzow, H. (2009). Herpesvirus assembly: an update. Virus Res,
143(2), 222‐234. doi:10.1016/j.virusres.2009.03.018 Meyer, K. (2007). Untersuchungen zur Übertragung von Koi‐Herpesvirus‐Infektionen durch
symptomlose Carrierfische. Dissertation Tierärztlichen Hochschule Hannover. Michel, B., Leroy, B., Stalin Raj, V., Lieffrig, F., Mast, J., Wattiez, R., . . . Costes, B. (2010). The genome
of cyprinid herpesvirus 3 encodes 40 proteins incorporated in mature virions. J Gen Virol, 91(Pt 2), 452‐462. doi:10.1099/vir.0.015198‐0
Mletzko, A., Amtmann, A., Bergmann, S., Lee, P., Christian, J., Buchholz, R., & Becker, A. (2017). Inoculation of cyprinid herpesvirus 3 (CyHV‐3) on common carp brain cells‐influence of process parameters on virus yield. In Vitro Cell Dev Biol Anim, 53(7), 579‐585. doi:10.1007/s11626‐017‐0170‐1

Quellen
97
Murphy, K. (2012). Janeway´s ‐ Immuno Biology (Vol. 8): Garland Science. Nakao, M., Tsujikura, M., Ichiki, S., Vo, T. K., & Somamoto, T. (2011). The complement system in
teleost fish: progress of post‐homolog‐hunting researches. Dev Comp Immunol, 35(12), 1296‐1308. doi:10.1016/j.dci.2011.03.003
Parra, D., Korytar, T., Takizawa, F., & Sunyer, J. O. (2016). B cells and their role in the teleost gut. Developmental and Comparative Immunology, 64, 150‐166.
Perelberg, A., Ronen, A., Hutoran, M., Smith, Y., & Kotler, M. (2005). Protection of cultured Cyprinus carpio against a lethal viral disease by an attenuated virus vaccine. Vaccine, 23(26), 3396‐3403. doi:10.1016/j.vaccine.2005.01.096
Perelberg, A., Smirnov, M., Hutoran, M., Diamant, A., Bejerano, Y., & Kotler, M. (2003). Epidemiological description of an new viral disease afflicting cultured Cyprinus carpio in Israel. The Israeli Journal of Aquaculture, 1(55), 5‐12.
Pradipta, R. R., Bismita, N., & Surajit, D. (2012). Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunology Letters, 148, 23‐33.
Rauh, T. C. M. a. I. (1990). A glycoprotein gX‐ß‐galactosidasee fusion gene as insertional marker for rapid identification of pseudorabies virus mutants.pdf. Journal of Virological.
Reske, A., Pollara, G., Krummenacher, C., Chain, B. M., & Katz, D. R. (2007). Understanding HSV‐1 entry glycoproteins. Rev Med Virol, 17(3), 205‐215. doi:10.1002/rmv.531
Roizman, B., and Knipe, D.M. (2001). Herpes simplex viruses and their replication. Fields Virology, 4th ed 2, 2399–2460.
Rona, G., Marfori, M., Borsos, M., Scheer, I., Takacs, E., Toth, J., . . . Vertessy, B. G. (2013). Phosphorylation adjacent to the nuclear localization signal of human dUTPase abolishes nuclear import: structural and mechanistic insights. Acta Crystallogr D Biol Crystallogr, 69(Pt 12), 2495‐2505. doi:10.1107/S0907444913023354
Ronen, A., Perelberg, A., Abramowitz, J., Hutoran, M., Tinman, S., Bejerano, I., . . . Kotler, M. (2003). Efficient vaccine against the virus causing a lethal disease in cultured Cyprinus carpio. Vaccine, 21(32), 4677‐4684. doi:10.1016/s0264‐410x(03)00523‐1
Rosenkranz, D., Klupp, B. G., Teifke, J. P., Granzow, H., Fichtner, D., Mettenleiter, T. C., & Fuchs, W. (2008). Identification of envelope protein pORF81 of koi herpesvirus. J Gen Virol, 89(Pt 4), 896‐900. doi:10.1099/vir.0.83565‐0
Sano, M., Ito, T., Kurita, J., Yanai, T., Watanabe, N., Miwa, S., & Iida, T. (2004). First Detection of Koi Herpesvirus in Cultured Common Carp Cyprinus carpio in Japan. Fish Pathology, 3(39), 165‐167.
Sathiyamoorthy, K., Chen, J., Longnecker, R., & Jardetzky, T. S. (2017). The COMPLEXity in herpesvirus entry. Curr Opin Virol, 24, 97‐104. doi:10.1016/j.coviro.2017.04.006
Schnitzlein, W. M., Winans, R., Ellsworth, S., & Tripathy, D. N. (1995). Generation of thymidine kinase‐deficient mutants of infectious laryngotracheitis virus. Virology, 209(2), 304‐314. doi:10.1006/viro.1995.1262
Secombes, C. J. (1996). The nonspecific immune system: celluar defens. In the Fish Immune System: Organism, Pathogen, and Enviroment (Vol. 15).
Soleto, I., Fischer, U., Tafalla, C., & Granja, A. G. (2018). Identification of a Potential Common Ancestor for Mammalian Cross‐Presenting Dendritic Cells in Teleost Respiratory Surfaces. Front Immunol, 9, 59. doi:10.3389/fimmu.2018.00059
St‐Hilaire, S., Beevers, N., Way, K., Le Deuff, R. M., Martin, P., & Joiner, C. (2005). Reactivation of koi herpesvirus infections in common carp Cyprinus carpio. Dis Aquat Organ, 67(1‐2), 15‐23. doi:10.3354/dao067015
Strang, B. L., & Stow, N. D. (2005). Circularization of the herpes simplex virus type 1 genome upon lytic infection. J Virol, 79(19), 12487‐12494. doi:10.1128/JVI.79.19.12487‐12494.2005
Sunyer, J. O. (2013). Fishing for mammalian paradigms in the teleost immune system. Nat Immunol, 14(4), 320‐326. doi:10.1038/ni.2549

Quellen
98
Taharaguchi, S., Inoue, H., Ono, E., Kida, H., Yamada, S., & Shimizu, Y. (1994). Mapping of transcriptional regulatory domains of pseudorabies virus immediate‐early protein. Arch Virol, 137(3‐4), 289‐302.
Tombacz, D., Toth, J. S., Petrovszki, P., & Boldogkoi, Z. (2009). Whole‐genome analysis of pseudorabies virus gene expression by real‐time quantitative RT‐PCR assay. BMC Genomics, 10, 491. doi:10.1186/1471‐2164‐10‐491
Turcotte, S., Letellier, J., & Lippe, R. (2005). Herpes simplex virus type 1 capsids transit by the trans‐Golgi network, where viral glycoproteins accumulate independently of capsid egress. J Virol, 79(14), 8847‐8860. doi:10.1128/JVI.79.14.8847‐8860.2005
Vallejo, A. N., Miller, N. W., Harvey, N. E., Cuchens, M. A., Warr, G. W., & Clem, L. W. (1992). Cellular pathway(s) of antigen processing and presentation in fish APC: endosomal involvement and cell‐free antigen presentation. Dev Immunol, 3(1), 51‐65.
van Oirschot, J. T. (1999). Diva vaccines that reduce virus transmission. J Biotechnol, 73(2‐3), 195‐205. van Oirschot, J. T., Rziha, H. J., Moonen, P. J., Pol, J. M., & van Zaane, D. (1986). Differentiation of
serum antibodies from pigs vaccinated or infected with Aujeszky's disease virus by a competitive enzyme immunoassay. J Gen Virol, 67 ( Pt 6), 1179‐1182. doi:10.1099/0022‐1317‐67‐6‐1179
Vancsok, C., Penaranda, M. M. D., Raj, V. S., Leroy, B., Jazowiecka‐Rakus, J., Boutier, M., . . . Vanderplasschen, A. F. C. (2017). Proteomic and Functional Analyses of the Virion Transmembrane Proteome of Cyprinid Herpesvirus 3. J Virol, 91(21). doi:10.1128/JVI.01209‐17
Wakae, K., Magor, B. G., Saunders, H., Nagaoka, H., Kawamura, A., Kinoshita, K., . . . Muramatsu, M. (2006). Evolution of class switch recombination function in fish activation‐induced cytidine deaminase, AID. Int Immunol, 18(1), 41‐47. doi:10.1093/intimm/dxh347
Walster, C. I. (1999). Clinical observations of severe mortalities in koi carp, Cyprinus carpio, with gill disease. Fish Veterinary Journal, 3, 54‐58.
Waltzek, T. B., Kelley, G. O., Stone, D. M., Way, K., Hanson, L., Fukuda, H., . . . Hedrick, R. P. (2005). Koi herpesvirus represents a third cyprinid herpesvirus (CyHV‐3) in the family Herpesviridae. J Gen Virol, 86(Pt 6), 1659‐1667. doi:10.1099/vir.0.80982‐0
Watanabe, S., Ono, E., Nikami, H., & Kida, H. (1998). Promoter activity of sequence located upstream of the pseudorabies virus early protein 0 gene. Vet Microbiol, 61(1‐2), 7‐19.
Whyte, S. K. (2007). The innate immune response of finfish‐‐a review of current knowledge. Fish Shellfish Immunol, 23(6), 1127‐1151. doi:10.1016/j.fsi.2007.06.005
Wintersberger, E. (1997). Regulation and biological function of thymidine kinase. Biochem Soc Trans, 25(1), 303‐308.
Zhang, H. G., & Hanson, L. A. (1995). Deletion of thymidine kinase gene attenuates channel catfish herpesvirus while maintaining infectivity. Virology, 209(2), 658‐663. doi:10.1006/viro.1995.1300
Zhou, J. X., Wang, H., Li, X. W., Zhu, X., Lu, W. L., & Zhang, D. M. (2014b). Construction of KHV‐CJ ORF25 DNA vaccine and immune challenge test. J Fish Dis, 37(4), 319‐325. doi:10.1111/jfd.12105
Zwollo, P., Cole, S., Bromage, E., & Kaattari, S. (2005). B cell heterogeneity in the teleost kidney: evidence for a maturation gradient from anterior to posterior kidney. J Immunol, 174(11), 6608‐6616.
Zwollo, P., Haines, A., Rosato, P., & Gumulak‐Smith, J. (2008). Molecular and cellular analysis of B‐cell populations in the rainbow trout using Pax5 and immunoglobulin markers. Dev Comp Immunol, 32(12), 1482‐1496. doi:10.1016/j.dci.2008.06.008

Zusammenfassung der Dissertation
99
7. Zusammenfassung der Dissertation
Herstellung sichererer und wirksamer Lebendvakzine gegen die Koi Herpesvirus Infektion von Karpfen
Vorgelegt von Lars Schröder
Das Koi Herpesvirus (KHV, Cyprinid herpesvirus 3) verursacht eine tödliche
Erkrankung bei Kois und Karpfen. Um sichere und wirksame Lebendvirusimpfstoffe
zu erhalten, haben wir Einzel- und Doppeldeletionsmutanten von KHV erzeugt, aus
deren Genom die für die beiden Nukleotidstoffwechselenzyme Thymidinkinase (TK,
ORF55) und Desoxyuridin-Triphosphatase (DUT, ORF123) codierenden Leserahmen
gezielt entfernt worden waren. Die Mutationen wurden durch homologe
Rekombination in den zellkulturadaptierten aber noch virulenten Stamm KHV-T
eingeführt. Umfangreiche in vitro Tests zeigten, dass die Deletion der TK- und DUT-
Gene die KHV-Replikation in Zellkultur (CCB Zellen) nicht erkennbar beeinträchtigt.
In vivo Tests an Jungkarpfen zeigten jedoch eine im Vergleich zum Ausgangsvirus
signifikant reduzierte Virulenz der Einzelgen-Deletionsmutanten eine fast
vollständige Attenuierung der Doppelmutante. Dennoch waren alle immunisierten
Karpfen gegen eine letale Belastungsinfektion mit virulentem KHV geschützt. Mittels
einer neu entwickelten Triplex-Real-Time-PCR und aus Kiementupferproben
isolierter DNA war es möglich, mit TK-negativem KHV immunisierte und Wildtyp-
infizierte Karpfen zu differenzieren. Daher könnte die Doppelmutante KHV-
TΔDUT/TK als genetischer Marker-Impfstoff geeignet sein.
In einer zweiten Studie wurde die Funktion von vier immunogenen
Hüllglykoproteinen der ORF25-Genfamilie (ORF25, ORF65, ORF148 und ORF149) von
KHV untersucht. Hierbei wurde festgestellt, dass alle vier Gene für die
Virusreplikation in Zellkultur entbehrlich sind. Während die Deletion von ORF65
keinen erkennbaren Einfluss auf die Virusvermehrung hatte, führte die Deletion von
ORF148 sogar zu einer leicht erhöhten Replikationsrate. Im Gegensatz dazu
bewirkten Deletionen von ORF25 oder ORF149 einen verzögerten Eintritt in die
Wirtszellen und damit auch eine verlangsamte Vermehrung und Ausbreitung der
Viren. Interessanterweise führte die gemeinsame Deletion der Gene ORF148 und
ORF149 zu einem wildtypähnlichen Wachstumsverhalten, das auf gegensätzliche

Zusammenfassung der Dissertation
100
Funktionen der beiden Proteine hindeutete. Elektronenmikroskopische
Untersuchungen von CCB-Zellen, die mit den verschiedenen
Glykoproteindeletionsmutanten infiziert waren, zeigten keine Auswirkungen auf die
Bildung und Reifung der Virionen im Zellkern oder im Zytoplasma, oder die
Virusfreisetzung. Im Tierversuch erwiesen sich KHV-Mutanten mit Deletionen der
Gene ORF148 und/oder ORF149 als geringfügig, aber für eine Verwendung als
Lebendvirus-Impfstoff nicht ausreichend abgeschwächt. Überlebende Fische waren
jedoch gegen Belastungsinfektionen ebenso gut geschützt wie Wildtyp-infizierte
Karpfen, so dass die Deletion dieser antikörperinduzierenden Proteine zur
Entwicklung von KHV-Markerimpfstoffen beitragen könnte, die eine serologische
Differenzierung von Wildtyp-infizierten und geimpften Fischen erlauben (DIVA-
Prinzip). In einer dritten Studie wurden durch serielle Zellkulturpassage von
virulentem KHV und anschließende in vivo Infektionsversuche Hinweise darauf
gefunden, dass das bislang nicht näher charakterisierte, neben dem ORF149 Gen
lokalisierte ORF150 für einen weiteren Virulenzfaktor von KHV codiert.
Möglicherweise könnte also durch eine kombinierte Deletion der im Rahmen dieser
Arbeit untersuchten KHV-Gene ein sicherer und wirksamer, genetisch und serologisch
differenzierbarer Markerimpfstoff hergestellt werden.

Summary
101
8. Summary
Koi herpesvirus (KHV, Cyprinid herpesvirus 3) causes a fatal disease of koi and
common carp. To obtain safe and efficacious live virus vaccines, we generated single
and double deletion mutants of KHV lacking the genes encoding the nucleotide
metabolism enzymes thymidine kinase (TK, ORF55) and deoxyuridine triphosphatase
(DUT, ORF123). The mutations were introduced by homologous recombination in the
cell culture adapted, but still virulent strain KHV-T. In vitro tests showed that
deletion of the TK and DUT genes does not affect KHV replication in CCB cell cultures.
In vivo tests using juvenile carp revealed that virulence of the single deletion
mutants was significantly reduced compared to parental wild type virus, and that
the double mutant was almost completely attenuated. Nevertheless, all immunized
carp were protected against lethal challenge infections with virulent KHV. Using a
novel triplex-real-time PCR and isolated DNA from gill swab samples, carp immunized
with TK-deleted KHV could be differentiated from wild-type infected fish. Therefore,
the double mutant KHV-TΔDUT/TK might be suitable as a genetic marker vaccine.
In a second study the functions of four immunogenic envelope glycoproteins,
encoded by the ORF25 gene family (ORF25, ORF65, ORF148, and ORF149) of KHV. It
was observed that the four genes are not essential for in vitro virus replication.
Whereas deletion of ORF65 did not detectably affect replication, deletion of ORF148
even slightly enhanced virus growth. In contrast, deletions of ORF25 or ORF149 led
to delayed entry into host cells, resulting in decelerated replication and spread of
the virus. Interestingly, double deletion of the genes ORF148 and ORF149 restored
wild-type-like growth properties, indicating opposite functions of the two proteins.
Electron microscopy of CCB cells infected with the different glycoprotein gene
deletion mutants showed no effect on the formation and maturation of the virions in
the cell nucleus or in the cytoplasm, nor on virus release. In animal experiments the
virus mutants lacking ORF148 and/or ORF149 proved to be slightly but insufficiently
attenuated for use as a live virus vaccines. However, surviving carp were completely
protected against lethal challenge infections, and, thus, deletion of these antibody-
inducing proteins might contribute to development of vaccines which permit
serological differentiation of immunized from wild-type infected animals (DIVA
principle).

Summary
102
In a third study serial cell culture passages of virulent KHV-T and subsequent animal
experiments indicated that the uncharacterised ORF150, which is located upstream
of ORF149, might encode another virulence factor of KHV. Thus, combined deletion
of the KHV genes analysed in the different parts of this thesis might result in a safe
and efficacious genetic, und serological DIVA vaccine.

Anhang
103
9. Anhang
9.1. Eigenständigkeitserklärung
Hiermit erkläre ich, dass diese Arbeit bisher von mir weder an der Mathematisch-Naturwissenschaftlichen Fakultät der Universität Greifswald noch einer anderen wissenschaftlichen Einrichtung zum Zwecke der Promotion eingereicht wurde.
Ferner erkläre ich, dass ich diese Arbeit selbstständig verfasst und keine anderen als die darin angegebenen Hilfsmittel und Hilfen benutzt und keine Textabschnitte eines Dritten ohne Kennzeichnung übernommen habe.
___________________
Lars Schröder

Anhang
104
9.2. Publikationen und Tagungsbeiträge
9.2.1. Tagungsbeiträge
14.03.-17.03.2018 28th Annual Meeting of the Society for Virology, Würzburg
“Generation of potential live vaccines against koi herpesvirus infection by deletion of nonessential viral genes”
(Vortrag)
29.07.-02.08.2017 42nd International Herpesvirus Workshop, Ghent
“Generation of potential live vaccines against koi herpesvirus by deletion of nonessential genes encoding enzymes of nucleotide metabolism”
(Vortrag)
20.09-22.09.2017 Junior Scientist Symposium FLI, Braunschweig
“Generation of potential live vaccines against koi herpesvirus by deletion of nonessential genes encoding enzymes of nucleotide metabolism”
(Poster)
28.09.-30.09.2016 1st SUMMER SCHOOL “INFECTION BIOLOGY”, Greifswald
“Development of recombinant live virus vaccines against koi herpesvirus”
(Poster)
21.09.-23.09.2016 Junior Scientist Symposium FLI, Jena
“Development of recombinant live virus vaccines against koi herpesvirus”
(Poster)

Anhang
105
9.2.2. Publikationen
Generation of a potential koi herpesvirus live vaccine by simultaneous deletion
of the viral thymdine kinase and dUTPase genes”
Lars Schröder, Sandro Klafack, Sven M. Bergmann, Dieter Fichtner, Yeonhwa Jin, Pei-
Yu Lee, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs
Publiziert in Journal of General Virology
Characterization of gene deletion mutants of Cyprinid herpesvirus 3 (koi herpesvirus) lacking the immunogenic envelope glycoproteins pORF25, pORF65, pORF148 and pORF149”
Lars Schröder, Sandro Klafack, Sven M. Bergmann, Pei-Yu Alison Lee, Kati
Franzke, Dirk Höper, Thomas C. Mettenleiter, Walter Fuchs
Publiziert in Virus Research
In cellulo evolution of Cyprinid herpesvirus 3 seems to follow a viral quasispecies-like model
Sandro Klafack, Anna-Sophie Fiston-Lavier, Sven M. Bergmann, Saliha Hammoumi,
Lars Schröder, Walter Fuchs, Angela Lusiastuti, Pei-Yu Lee, Master students, Anne-
Sophie Gosselin-Grenet and Jean-Christophe Avarre
Eingereicht bei Viruses - Animal Viruses

Anhang
106

Anhang
107
9.3 Lebenslauf
Lars Schröder
Friedrich-Loeffler-Straße 24
17489 Greifswald
Geboren am 25.02.1988 in Schwerin
Akademische Ausbildung
Seit 09/2015 Doktorand am Friedrich - Loeffler- Institut Riems
Institut Molekulare Virologie und Zellbiologie
In der Arbeitsgruppe von Dr. Walter Fuchs
Doktorvater: Prof. Dr. Dr. h.c. Thomas C. Mettenleiter
10/2012 - 03/2015 Molekular Biologie Master of Science
Universität Hamburg
In der Arbeitsgruppe von Prof. Dr. Wolfgang Streit
Mikrobiologie
10/2009 - 09/2012 Biologie Bachelor of Science
Universität Hamburg
In der Arbeitsgruppe von Prof. Dr. Wolfgang Streit
Mikrobiologie
Schulausbildung
09/2001 - 06/2008 Sportgymnasium Schwerin
09/1998 - 06/2001 J.-R.-Becher Schule Schwerin
09/1994 - 06/1998 Heinrich Heine, Grundschule Schwerin

Anhang
108
Sonstiges
10/2010 - 03/2015 Studentische Hilfskraft
Im Fachbereich Mikrobiologie, Universität Hamburg
Aufgabenbereiche:
• Vorbereitung der Versuchsprotokolle
• Betreuung und Beaufsichtigung der Versuche
• Anlegen von Tabellen und Übersichten zur Auswertung
11/2013 - 02/2014 Hilfswissenschaftler
Im Heinrich-Pette-Institute Hamburg
Aufgabenbereiche:
• Protein/Protein Interaktionsstudien
• Herstellung kompetenter Zellen
• Transfektion/Transformation
01/2012 - 02/2012 Praktikum
Im Fachbereich Mikrobiologie, Helios Klinikum Schwerin
Aufgabenbereiche:
• ELISA-Techniken
• Resistenzbestimmung von Erregern
• Auftragserfassung mikrobiologischer
Untersuchungsproben

Anhang
109
9.4 Danksagung
Ich danke Herrn Professor Dr. Dr. h.c. Thomas C. Mettenleiter für die Möglichkeit,
meine Dissertation am Friedrich-Loeffler-Institut anzufertigen und für die
Überlassung des spannenden Themas.
Dr. Walter Fuchs möchte ich für die stets hilfreichen Besprechungen, die sehr gute
Betreuung und herzliche Aufnahme in das Labor Fuchs bedanken.
Ein großes Dankeschön geht an Dr. Dr. habil. S. M. Bergmann für die ausgezeichnete
Betreuung bei allen Tierversuchen und für die sehr gute Zusammenarbeit zwischen
den Laboren, sowie für manches offene Ohr bei Problemstellungen.
Ein großes Dankeschön geht auch an Anja Landmesser für die hervorragend fachliche
Betreuung und Unterstützung im Laboralltag, sowie die vielen hilfreichen
Anmerkungen und wertvollen Anregungen. Danke für deine Geduld und deine
aufmunternden Worte.
Allen Mitarbeitern des Labor Fuchs (Anja Landmesser, Alexandra Hübner und Tonny
Kabuuka) danke ich für die super Arbeitsatmosphäre, sowie für die praktische
Unterstützung! Danke, dass ihr immer ein offenes Ohr für mich hattet und mir mit
Rat und Tat zur Seite standet.
Weiterhin möchte ich allen Mitarbeiten der 2. Etage in Gebäude 40.1 danken: für die
Versorgung mit Ratschlägen, Chemikalien und Zelllinien in jedem Labor!
Meiner „Mittagsrunde“ danke ich für die exzellente Pausenbespaßung! Vor allem
Vicky, Stephie, Svenja und Melina danke ich für die Erfahrung, sowie die teils sehr
spannenden und anregenden Gesprächsthemen, die so manchen schlechten Tag
wieder gut gemacht haben.
Ich danke Sandro für seine fachliche Unterstützung im Tierversuch und für so manche
Aushilfe speziell am Wochenenden oder beim Austausch von Erfahrungen und
Anregungen für neue Ideen.
Mein größter Dank gilt natürlich meiner Freundin Vicky. Ich danke dir von ganzem
Herzen für deine unendliche Geduld und Unterstützung, sowohl auf privater Ebene,
als auch bei der Anfertigung dieser Doktorarbeit und den Publikationen.
Zu guter Letzt möchte ich meiner Familie danken, die mir in all den Jahren den
Rücken gestärkt hat und immer an mich geglaubt haben. Und ich möchte hier speziell
meiner kleinen Tochter Neele danken, die mich inspiriert und mir die Motivation gibt
immer weiter zu machen, in der Hoffnung, dass sie mal stolz auf ihren Papa sein
kann.





























