Embed Size (px)
Citation preview

Protoplasma 102, 141--146 (1980) F IOTOPLASMA �9 by Springer-Verlag 1980
I n d u c e d Cel l F u s i o n in Foraminifera
D. SCHWAB * and HILTRUD SCHWAB-STE3:
Fachridltung Anatomie im Fachbereich Theoretische Medizin, Universit~it des Saarlandes, Homburg (Saar)
Received August t7, 1979 Accepted September 26, 1979
S u m m a r y
The fusion of cells of the monothalamous foraminifer Myxotheca arenilega (Protozoa, Rhizo- poda) is described. The fusion process is induced by the separation of the extracellular organic shell from the cell body. As several cells can fuse either stepwise or simultanously, a giant cell of some millimeters in diameter can be formed.
Keywords: Cell fusion; Foramini[era; Protozoa.
1. I n t r o d u c t i o n
From numerous eukaryotic cells, i.e., myoblasts, macrophages, gametes, erythrocytes, lower and higher fungi, it is known, that, independent of their natural fusion capacity, cell fusion can be induced by a wide range of factors (for references see POSTE and ALLISON 1973, POSTE and NICOLSON 1978, BITTAR 1979). Among the protozoan order Foraminifera cell fusion certainly is a normal process during the development of cells, but within the life cycle cell fusion is restricted exclusively to the very beginning of agamogony, i.e., when gametes fuse in order to form zygotes. Cells in all other stages during agamogony and gamogony do not fuse, even if they are brought into contact with their reticulopodial membranes. The present investigation demonstrates that even a protozoon, the mono- thalamous foraminifer Myxotheca arenilega, is suited for the study of cell fusion, because fusion can be induced in various cells out of the life cycle, independent of their stage of development.
* Correspondence and Reprints: Fachrichtung Anatomie im Fachbereich Theoretische Medi- zin der Universit~t des Saarlandes, D-6650 Homburg (Saar), Federal Republic of Germany.
0033-183X/80/0102/0141/$ 01.20

142 D. SCHWAB and HILTRUD SCHWAB-STEY
2. Material and Methods
The cultivation of M. arenilega has been previously described (ScHwAI~ 1969). For the fusion experiments the cells were transferred into small dishes containing fresh seawater. Then the ceils were carefully squeezed with a pipette, in order to separate the protoplast from the orga- nic shell.
3. Resul t s
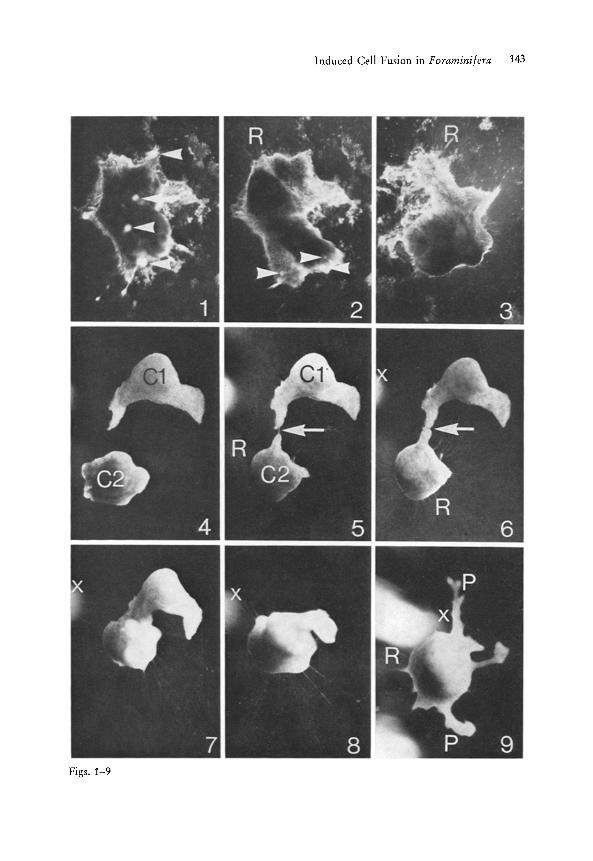
Immediately after the removal of the shell, the cell of M. arenilega shows an irregular shape (Fig. 1), Additionally, small opaque cytoplasmic protrusions covering the total cell surface are temporarily visible (Fig. 1). Within the following 30 minutes the cell is able to round off and finally exhibits a shape similar to a shell-covered celt (Figs. i-3). During the process of plasma- accumulation numerous radially extended reticulopodia are formed at the cell surface contacting the substrate (Figs. 2 and 3). When the shell-free foraminiferan cells are brought into juxtaposition, in order for them to con- tact, cell fusion starts. In Figs. 4-9 a fusion process is demonstrated. One protoplast has already built up its reticulopodial network and is contacting a freshly squeezed off neighbouring protoplast over one small reticulopodium (Fig. 5). This reticulopodium enlarges within 5 minutes (Fig. 6), resulting in the formation of an extended cytoplasmic bridge between the two fusing protoplasts (Fig. 7). During the fusion process both protoplasts show a relatively rapid cytoplasmic streaming and continuously change their cell shapes (compare Figs. 5-8). The fusion process of two cells via reticulopodia is terminated after about
Figs. 1-3. Change of the cell shape of M. arenilega after separation of the cell from the extra- cellular organic shell. Fig. 1. Cell of M. arenilega 1 rain after separation from the organic shell. The naked protoplast shows an irregular shape and numerous opaque protrusions (ar- rows) spread over the total surface. At that time no retriculopodia are visible. Fig. 2. 10 min later the cell has formed short reticulopodia (R), the cell shape has changed, and the cyto- plasmic protrusions (arrows) have nearly disappeared. Fig. 3. Again 10 rain later the cell has almost totally rounded off, showing a smooth surface and at the celI periphery numerous smatl reticulopodia (R). Magn. Fig. 1-3. X 1542.5
Figs. 4-9. Fusion process of two shell-free ceils of M. arenilega. Fig. 4. The two shell-free cells (C 1 and C 2) 5 min after separation from the organic shell Fig. 5. After 20 min the upper ceil (C J) lies still motionless whereas the lower cell (C 2) has already extended numer- ous reticulopodia (R). One of these reticulopodia has come into contact with the upper cell (arrow). Fig. 6. After 30 min the contacting reticulopodium has enlarged and is building an extended cytoplasmic bridge between the neighbouring cells (arrow). Fig. 7. After 60 min the fusion of the two cells expands over a large area. During the fusion process the upper cell which did not form reticulopodia is pulled down towards the lower cell being anchored at the substrate by numerous reticulopodia. Fig. 8. After 90 min the cytoplasmic fusion is almost completed. Fig. 9. After 120 miu the total fusion of the two cells is terminated. Dur- ing the fusion process the lower cell has migrated (compare marker x in Fig. 6-9). The fused cell shows large lateral IobopodiaI protrusions (P) and small reticulopodia (R). Magn. Fig. 4-9. X 71.5
Induced Cell Fusion in l:oraminifera 143
Figs. 1-9

144 D. SCHWAB and HILTRUD SCHWAB-STEY
two hours (Fig. 9). I f two protoplasts are directly brought into contact with their plasmalemmata the fusion time is markedly reduced. It is possible to fuse several protoplasts at the same time (Fig. 10) or stepwise within a few days. In this way a giant foraminiferan cell can be developed reaching a diameter of several millimeters (Fig. 11).
Fig. 10. Four shell-free cells (C 1-C4) of M. arenilega brought into contact with their plasmalemmata during simultanous fusion. R reticulopodia. Magn. • 140 Fig. 11. A normal adult cell of M. arenilega (C 1) covered by its organic shell and a large naked shell-free cell (C 2) produced by fusion of twelve cells. Magn. • 60
After fusion the protoplast shows only small reticulopodia but several large lobopodial protrusions, which can reach several hundred micrometers (Fig. 9). Fusion can also be observed when cells, which were only part ly squeezed off, were brought into contact with their plasmalemmata. The newly formed celt then is covered by the shells of the fused cells. The fusion experiments described here for M. arenilega can also be carried out with other monothalamous Foraminifera, i.e., Allogromia laticollaris Arnold, Allogromia laticollaris strain GF, and Boderia albicollaris Schwab.

Induced Cell Fusion in Foraminifera 145
4. D i scuss ion
The present investigation shows that protozoan cells of the order Forarnini/era can be stimulated for cell fusion. During the life cycle, cells of the mono- thalamous foraminifer Myxotheca arenilega normally do not fuse, except in the stage of gametogenesis (GRELL 1968). The gametes represent the only shell-free stage within the life cycle, in all other phases the cells are covered by an organic shell of different thickness, depending on the stage of develop- ment (ScHwAB 1969). Cell fusion in different developmental stages besides gametogenesis can also be obtained, but only after mechanical treatment, i.e., separation of the extracellular organic shell from the cell body. The fusion process of foraminiferan cells is demonstrated in Figs. 4-10. The shell-free cells do not only fuse at the areas of their plasmalemmata which originally have been covered by the organic shells, but they can also fuse over their reticulopodial membranes. This is remarkable because the reticulo- podia are characteristic components of every growing foraminiferan cell. These naked cytoplasmic strands are extended through an aperture in the organic shell providing the attachment of the cell at the substrate and the food supply. But fusion never occurs over reticulopodia in shell-covered cells. The fusion of foraminiferan cells can be observed in the culture medium without adding any fusogen substances. As the ion concentration in the medium, the pH, and the temperature are not changed, the elimination of the shell must be solely responsible for the altered plasma membrane properties and the changed behaviour of the cells. Investigating the cell pigments of M. arenilega one can demonstrate that carotene wax pigments are synthesized by the foraminiferan cell depending on the light intensity offered during cultivation (ScHwAB and SCH~OBACH 1979). The differences in pigmentation are clearly visible with the naked- eye, because cells cultivated in total darkness are markedly less pigmented than cells grown by daylight in combination with artificial lighting. It is, therefore, no problem to observe the cytoplasmic fusion under the light microscope if one takes differently coloured cells, because in the region of fusion the cytoplasm shows mixed colours, expanding all over the cell during fusion. This phenomenon brings about precise data of the beginning and progress of cytoplasmic fusion. Further investigations will show how fused cells develop and in which way the concentration change of divalent cations in the culture medium and other chemical and physical parameters can influence fusion of foraminiferan cells.
Acknowledgements
This investigation was supported by the Deutsche Forschungsgemeinschaft Schw 225/1.
10 Protoplasma 102/1--~

146 SCHWAB et aI.: Induced Cell Fusion in Foraminifera
References
BITTAR, E. E., 1979: Membrane structure and function, Vol. 3. New York-London-Sidney- Toronto: J. Wiley & Sons.
G~,E~.L, K. G., 1968: Protozoologie. Berlin-GiJttingen-Heidelberg: Springer. POSTE, G., ALLISON, A. C., 1973: Membrane fusion. Biochim. biophys. Acta 300, 421--465. - - NIC~OLSON, G. L., 1978: Cell surface reviews, Vol. 5. Amsterdam-New York-Oxford:
North-Holland Publ. Comp. ScHwa~, D., 1969: Elektronenmikroskopische Untersuchung an der Foraminifere Myxotheca
arenilega Schaudinn. Z. Zellforsch, 96, 295--324. - - SCHLO]3ACH, ~-I., 1979: Pigments in monozhalamous Foraminifera. J. Foram. Res. 9, 141--
146.


![Sendai [Schreibgeschützt] [Kompatibilitätsmodus] · 2014. 10. 21. · Paramyxovirus-Cell Membrane Fusion 2002 Journal of Virology 76 (24)Cell Membrane Fusion. 2002. Journal of Virology](https://img.pdfslide.org/doc/110x75/6094d970401cb65a1550bf0a/sendai-schreibgeschtzt-kompatibilittsmodus-2014-10-21-paramyxovirus-cell.jpg)


























