Embed Size (px)
Citation preview
6 Aminosäuren
Lothar Eggeling
6.1 Einleitung 110
6.1.1 Bedeutung und Anwendungsbereiche 110
6.1.2 Aminosäuren in der Futtermittelindustrie 111
6.1.3 Wirtschaftliche Bedeutung 111
6.2 Herstellung von Aminosäuren 112
6.2.1 Klassische Entwicklung von Produktionsstämmen 113
6.2.2 Moderne Entwicklung von Produktionsstämmen 113
6.3 L-Glutamat 115
6.3.1 Syntheseweg und Regulation 115
6.3.2 Produktionsprozess 117
6.4 L-Lysin 118
6.4.1 Syntheseweg und Regulation 118
6.4.2 Produktionsstämme 119
6.4.3 Produktionsprozess 120
6.5 L-Threonin 122
6.5.1 Syntheseweg und Regulation 122
6.5.2 Produktionsstämme 122
6.5.3 Produktionsprozess 123
6.6 L-Phenylalanin 124
6.6.1 Syntheseweg und Regulation 124
6.6.2 Produktionsstämme 125
6.6.3 Produktionsprozess 125
6.7 Ausblick 126
H. Sahm, G. Antranikian, K.-P. Stahmann, R. Takors (Hrsg.), Industrielle Mikrobiologie, DOI 10.1007/978-3-8274-3040-3_6,© Springer-Verlag Berlin Heidelberg 2013

110 Kapitel 6 ⋅Aminosäuren
6.1 Einleitung
6.1.1 Bedeutung undAnwendungsbereiche
Aminosäuren sind die Bausteine der Proteineund damit für alle Lebewesen lebensnotwendig.Allerdings besitzen Mensch und Tier nur be-grenzte Synthesemöglichkeiten dafür. So kannder Mensch acht der 20 proteinbildenden Ami-nosäuren nicht synthetisieren. Diese essenziellenAminosäuren sind die drei verzweigtkettigenAminosäuren L-Leucin, L-Isoleucin und L-Valin,des Weiteren L-Threonin, L-Lysin, L-Methionin,L-Phenylalanin und L-Tryptophan. Der Bedarf
� Tabelle 6.1 Die Mengen an Aminosäuren, die jährlich hergestellt werden, und ihre Hauptverwendung
Aminosäure Menge (t/a) Hauptverwendung
L-Glutamat 2 300 000a „Würzmittel“
L-Lysin 1 300 000a Tierfutter
D,L-Methionin 850 000a Tierfutter
L-Threonin 190 000a Tierfutter
Glycin 16 000 Süßstoff
L-Aspartat 14 000 Süßstoff
L-Phenylalanin 14 000 Süßstoff
L-Cystein 6000 Pharmaka
L-Tryptophan 4500a Tierfutter
L-Cystin 3500 Pharmaka
L-Arginin 2000 Pharmaka
L-Alanin 1500 Süßstoff
L-Leucin 1200 Pharmaka
L-Valin 1000 Pharmaka
L-Isoleucin 500 Pharmakaa Weltmarkt 2009, sonst 2004
dieser Aminosäuren wird normalerweise durchden Verzehr von Eiweißprodukten gedeckt. Erkann aber je nach Lebensalter und Gesundheits-zustand recht unterschiedlich sein. So hat einSchulkind einen dreifach höheren Bedarf an L-Lysin als ein Erwachsener, und im Krankheitsfallkann die gezielte Versorgung mit Aminosäurennotwendig sein. Deswegen werden Aminosäu-ren z. B. für Infusionslösungen benötigt, die beichronisch entzündlichen DarmerkrankungenAnwendung finden, oder auch in der postope-rativen Phase. Als Beispiel für eine Infusions-lösung zur parenteralen Ernährung sei Amino-plasmal B genannt (Zusammensetzung unterwww.pharmazie.com/graphic/A/29/1-26529.pdf). Auch bei angeborenen Störungen des Ami-nosäurehaushalts, wie der Phenylketonurie, sind

6.1 ⋅ Einleitung 111
6
Aminosäuren Bestandteil spezieller diätischerLebensmittel.
Neben der Nutzung von Aminosäuren imPharmabereich gibt es weitere vielfältige An-wendungsgebiete (� Tab. 6.1). So wird in derNahrungsmittelindustrie Natriumglutamat be-nötigt. Dieses bewirkt den typischen „Umami“-Geschmack, der zusammen mit süß, salzig, sauerund bitter zu den fünf elementaren Geschmacks-empfindungen zählt. Das Wort „umami“ kommtaus dem Japanischen und bedeutet „herzhaft“,„wohlschmeckend“, „pikant“. Weiterhin werdenbeispielsweise die Aminosäuren L-Alanin undGlycin benötigt, weil sie Getränken zum Süßenzugesetzt werden. In der chemischen Industriewerden Aminosäuren als Bausteine benutzt, wiebeispielsweise L-Valin zur Synthese von Pesti-ziden oder L-Phenylalanin und L-Aspartat zurSynthese des Süßstoffs Aspartam.
6.1.2 Aminosäuren in derFuttermittelindustrie
Von herausragender wirtschaftlicher Bedeutungsind Aminosäuren in der Futtermittelindustrie.Die Zugabe von Aminosäuren zu Futtermittelnführt zu einer ausgewogenenAminosäurezusam-mensetzung, die dem tatsächlichen Bedarf ange-passt ist. Sie� reduziert die Kosten für Futtermittelrohstof-
fe,� steigert die Effizienz der Futtermittelverwer-
tung und� reduziert die umweltbelastende Stickstoffaus-
scheidung.Weizen als Futtermittel hat z. B. einen niedrigenL-Lysin-Anteil, sodass die anderen Aminosäu-ren nicht vollständig genutzt werden können(�Abb. 6.1). Die Zugabe von L-Lysin erhöhtden Anteil der Verwertung der anderen Ami-nosäuren. Nach der Zugabe von L-Lysin istals nächste Aminosäure L-Threonin limitie-rend – und danach L-Tryptophan. Durch Zugabedieser drei Aminosäuren wird der Nährwert vonWeizen nahezu verdoppelt, was bei steigendem
Phe
ThrLys Trp
Met
LeuVal
Ile
�Abb. 6.1 Das Liebig’sche Fass illustriert die Limitationdurch die niedrigste Fassdaube. So begrenzt bei Wei-zen als Proteinquelle zunächst L-Lysin den Nährwert desProteins. Nach der Zugabe von L-Lysin ist L-Threonin li-mitierend und danach L-Tryptophan. Die Zugabe dieserdrei Aminosäuren verdoppelt nahezu den Nährwert vonWeizen
Fleischkonsum einer Schonung der Resourcengleichkommt. Bei Nutzung von Sojamehl alsProteinquelle ist L-Methionin die zuerst limi-tierende Aminosäure, sodass auch hierfür eingroßer Bedarf besteht.
6.1.3 Wirtschaftliche Bedeutung
Aufgrund des zunehmenden Fleischkonsums,insbesondere in Asien und Südamerika, werdenvermehrt Aminosäuren für die Bereitstellungals Futtermittelzusatz benötigt. In der Haupt-sache sind dies L-Lysin, D,L-Methionin undL-Threonin, deren Bedarf jährlich um 5 bis 7%steigt.
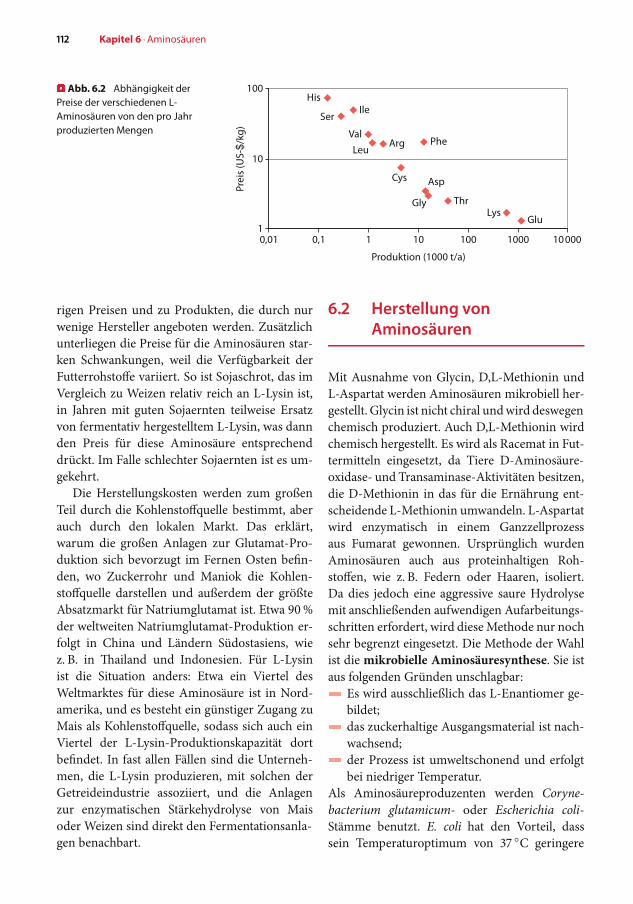
Die in den größten Mengen hergestelltenAminosäuren sind auchdie billigsten (� Abb. 6.2).Dies liegt daran, dass bei größerem Bedarf zu-nächst mehr Anbieter konkurrieren. Das hat dieEntwicklung effizienterer Fermentationsprozes-se zur Folge, was die Kosten reduziert und somitgünstigere Produkte liefert. Dies führt letztlichbei den großvolumigen Produkten zu sehr nied-
112 Kapitel 6 ⋅Aminosäuren
� Abb. 6.2 Abhängigkeit derPreise der verschiedenen L-Aminosäuren von den pro Jahrproduzierten Mengen
10,01
Produktion (1000 t/a)
Prei
s (U
S-$/
kg)
0,1 1 10
Glu
Thr
PheArg
Ile
AspCys
LysGly
Leu
Val
Ser
His
100 1000 10 000
10
100
rigen Preisen und zu Produkten, die durch nurwenige Hersteller angeboten werden. Zusätzlichunterliegen die Preise für die Aminosäuren star-ken Schwankungen, weil die Verfügbarkeit derFutterrohstoffe variiert. So ist Sojaschrot, das imVergleich zu Weizen relativ reich an L-Lysin ist,in Jahren mit guten Sojaernten teilweise Ersatzvon fermentativ hergestelltem L-Lysin, was dannden Preis für diese Aminosäure entsprechenddrückt. Im Falle schlechter Sojaernten ist es um-gekehrt.
Die Herstellungskosten werden zum großenTeil durch die Kohlenstoffquelle bestimmt, aberauch durch den lokalen Markt. Das erklärt,warum die großen Anlagen zur Glutamat-Pro-duktion sich bevorzugt im Fernen Osten befin-den, wo Zuckerrohr und Maniok die Kohlen-stoffquelle darstellen und außerdem der größteAbsatzmarkt für Natriumglutamat ist. Etwa 90%der weltweiten Natriumglutamat-Produktion er-folgt in China und Ländern Südostasiens, wiez. B. in Thailand und Indonesien. Für L-Lysinist die Situation anders: Etwa ein Viertel desWeltmarktes für diese Aminosäure ist in Nord-amerika, und es besteht ein günstiger Zugang zuMais als Kohlenstoffquelle, sodass sich auch einViertel der L-Lysin-Produktionskapazität dortbefindet. In fast allen Fällen sind die Unterneh-men, die L-Lysin produzieren, mit solchen derGetreideindustrie assoziiert, und die Anlagenzur enzymatischen Stärkehydrolyse von Maisoder Weizen sind direkt den Fermentationsanla-gen benachbart.
6.2 Herstellung vonAminosäuren
Mit Ausnahme von Glycin, D,L-Methionin undL-Aspartat werden Aminosäuren mikrobiell her-gestellt. Glycin ist nicht chiral undwird deswegenchemisch produziert. Auch D,L-Methionin wirdchemisch hergestellt. Es wird als Racemat in Fut-termitteln eingesetzt, da Tiere D-Aminosäure-oxidase- und Transaminase-Aktivitäten besitzen,die D-Methionin in das für die Ernährung ent-scheidende L-Methionin umwandeln. L-Aspartatwird enzymatisch in einem Ganzzellprozessaus Fumarat gewonnen. Ursprünglich wurdenAminosäuren auch aus proteinhaltigen Roh-stoffen, wie z. B. Federn oder Haaren, isoliert.Da dies jedoch eine aggressive saure Hydrolysemit anschließenden aufwendigen Aufarbeitungs-schritten erfordert, wird dieseMethode nur nochsehr begrenzt eingesetzt. Die Methode der Wahlist die mikrobielle Aminosäuresynthese. Sie istaus folgenden Gründen unschlagbar:� Es wird ausschließlich das L-Enantiomer ge-
bildet;� das zuckerhaltige Ausgangsmaterial ist nach-
wachsend;� der Prozess ist umweltschonend und erfolgt
bei niedriger Temperatur.Als Aminosäureproduzenten werden Coryne-bacterium glutamicum- oder Escherichia coli-Stämme benutzt. E. coli hat den Vorteil, dasssein Temperaturoptimum von ○C geringere

6.2 ⋅Herstellung von Aminosäuren 113
6
L-Lysin S-Aminoethylcystein
NH2
HO
O
H2N
NH2
HO
O
H2N
S
�Abb. 6.3 Vergleich der Strukturen von L-Lysin undS-Aminoethylcystein, einem Analogon von L-Lysin
Kühlungskosten bei der Fermentation erfor-dert als bei der Kultivierung von C. glutami-cum, dessen Temperaturoptimum bei etwa ○Cliegt. Demgegenüber sind Fermentationen mitC. glutamicum unempfindlich gegenüber Pha-geninfektionen, welche bei E. coli, besonders intropischen Ländern, ein großes Problem seinkönnen.
6.2.1 Klassische Entwicklung vonProduktionsstämmen
Die Wildtyp-Isolate von E. coli und C. glutami-cum bilden jeweils nur die Mengen an Amino-säure, die sie selbst zum Wachstum benötigen.Dies bewirken effiziente Kontrollmechanismen,wie die allosterische Kontrolle von Syntheseen-zymen durch die entsprechenden Aminosäuren,weiterhin die Kontrolle der Transkriptionsinitia-tion durch den Beladungszustand der tRNA oderauch globale Regulationsmechanismen. Deshalbwerden Mutantenstämme gewonnen, die dieAminosäure verstärkt synthetisieren. In der klas-sischen Stammentwicklung werden die Zellendazu ungerichtet, z. B. durch UV-Bestrahlung,mutagenisiert und in groß angelegten Screeningsder Stamm mit der besten Aminosäureausschei-dung ausgewählt. In den Screenings werden oftAminosäureanaloga eingesetzt, die die Aktivi-tät von Schlüsselenzymen hemmen. So hemmtbeispielsweise das Lysinanalogon L-Aminoethyl-
cystein (AEC) wie L-Lysin selbst die Aspartat-kinase als Schlüsselenzym der Lysinsynthese,ermöglicht aber kein Wachstum (�Abb. 6.3 und6.6). Mutanten, die trotz Anwesenheit von L-Aminoethylcystein wachsen, scheiden häufigL-Lysin aus. Ausgehend von der isolierten Mu-tante wird die gesamte Prozedur mit weiterenAnaloga, oder auch nach anderen Selektionskri-terien, wiederholt (� Tab. 6.2), um schließlich zueinem industriell geeigneten Stamm zu gelangen.
Die klassische Stammentwicklung hat jedochNachteile:� Sie ist äußerst arbeitsaufwendig, da sehr viele
Klone gescreent werden müssen.� Sie ist auf Zufallstreffer in derMutagenese an-
gewiesen.� Es akkumulieren Mutationen, die zu schlech-
temWachstum führen.Schlechtes Wachstum ist unerwünscht, weil esden Produktionsprozess verlangsamt und so-mit ineffizient macht. Die Stammentwicklungmit ungerichteter Mutagenese und Screening aufProduktbildung hat aber auch Vorteile:� Es liegen die für die Produktbildung relevan-
ten Mutationen vor.� Es ist keine Kenntnis der molekularen Funk-
tion von Genorten erforderlich.Tatsächlich sind durch ungerichtete Mutagene-se und Screening auf erhöhte ProduktbildungStämme mit hervorragenden Produktbildungs-eigenschaften entstanden. Der Wert der unge-richteten Mutagenese besteht ferner darin, dassaus solchen Ansätzen mutierte Gene resultieren,die z. B. für nicht mehr allosterisch regulierteBiosyntheseenzyme codieren. Diese sind aberVoraussetzung zur modernen Entwicklung vonProduktionsstämmen mit rekombinanten Me-thoden.
6.2.2 Moderne Entwicklung vonProduktionsstämmen
Heutzutage bieten molekulare rekombinanteTechniken dieMöglichkeit, gezieltMutationen indas Chromosom einzuführen oder Gene überzu-

114 Kapitel 6 ⋅Aminosäuren
� Tabelle 6.2 Genealogie eines klassisch entwi-ckelten L-Lysin-Produktionsstamms. Der Wildtyp vonCorynebacterium glutamicumwurde mutiert und ineinem ersten Schritt unter AEC-resistenten Klonen derbeste Lysinproduzent ausgewählt. Durch fünf weitereMutagenese- und Selektionsschritte erfolgte jeweilseine Steigerung der Produktausbeute
Stamm Eigenschaft Lysin-Ausbeute(%)
AJ 1511 Wildtyp 0
AJ 3445 AECr 16
AJ 3424 AECr Ala− 33
AJ 3796 AECr Ala− CCLr 39
AJ 3990 AECr Ala− CCLr MLr 43
AJ 1204 AECr Ala− CCLr MLr FPs 50
AECr, resistent gegenüber S-(β-Aminoethyl)-L-cystein;Ala− , auxotroph für L-Alanin; CCLr, resistent gegen-über α-Chlorocaprolactam; MLr, resistent gegenüberμ-Methyllysin; FPs , sensitiv gegenüber β-Fluoropyruvat.
exprimieren. Dies zielt darauf ab, den gesamtenStofffluss einschließlich der Transportvorgän-ge in Richtung der gewünschten Aminosäurezu erhöhen, die Bildung unerwünschter Neben-produkte zu verringern und globale Kontroll-mechanismen zu überwinden (�Abb. 6.4). Diessetzt natürlich Kenntnisse zu den jeweiligen mo-lekularen Mechanismen voraus, die erforschtwerden müssen. Zusätzlich verwenden mole-kulare Techniken oft Gene, die wie erwähntaus klassisch gewonnenen Stämmen stammen.Darüber hinaus können klassisch gewonneneStämme auch die Basis bilden, um durch mole-kulare rekombinante Techniken eine zusätzlicheProduktsteigerung zu erreichen.
Weitere Ansätze zur Entwicklung von Pro-duktionsstämmen bieten die unter „omics“ zu-sammengefassten globalen Analysen (BoxS. 115). So wird die Bestimmung und Analysevon gleichzeitig möglichst vielen Metaboliten,dem Metabolom, als Metabolomik (Metabolo-mics) bezeichnet. Durch Vergleich des Meta-boloms von zwei Stämmen lässt sich erkennen,
Zucker
Zucker
Vorstufen Neben-produkte
Reduktions-äquivalenteAbbau-
wege
Synthese-wege
Aminosäure Abbau
Aminosäure
globaleRegulations-
mechanismen
Feedback-Hemmung
1
2
3
4
5
6
7
8
9
�Abb. 6.4 Mögliche Ansätze zur Verbesserung derAminosäure-Produktion bestehen in (1) verbesserterZuckeraufnahme, (2) erhöhter Vorstufenbereitstellung,(3) vermehrter Bildung von Reduktionsäquivalenten,(4) Überwindung der Feedback-Hemmung von Enzymen,(5) Aufhebung limitierender Biosyntheseschritte, (6) Ver-hinderung der Nebenproduktbildung, (7) verbessertemExport, (8) Verhinderung des Abbaus und (9) Optimierungder globalen Regulationsmechanismen für die Aminosäu-resynthese
wo sich ein Metabolit im Stoffwechsel anstaut.Dies ist ein Hinweis darauf, dass der nachfol-gende Schritt im Syntheseweg eine Limitationdarstellt, die dann durch Genüberexpressionüberwunden werden kann. Eine weitere globaleAnalyse ist die Bestimmung aller Transkrip-te einer Zelle, des Transkriptoms, welche alsTranskriptomik (Transkriptomics) bezeichnetwird. Dies kann hilfreich sein, um z. B. durchVergleiche verschiedener Fermentationen miteinem Produktionsstamm zu prüfen, ob einbestimmtes Expressionsmuster mit einem be-sonders guten Prozessverlauf korreliert. Fallsdies der Fall ist, besteht die Möglichkeit, diegefundenen Gene gezielt für eine Stammopti-mierung einzusetzen. Auch genomweite Studienauf Sequenzebene, die unter dem Begriff Geno-mik (Genomics) zusammengefasst sind, werdenzur Stammoptimierung eingesetzt. Sie ermögli-chen es, vollständige Genomsequenzen klassisch

6.3 ⋅ L-Glutamat 115
6
Die verschiedenenomics-Techniken
Zu den omics-Techniken zählenkomplette Analysen des Ge-noms (Genomics), Transkriptoms(Transkriptomics), Proteoms (Pro-teomics) und des Metaboloms(Metabolomics). Während reduk-tionistische Ansätze einzelne
Gene, Transkripte, Proteine oderMetaboliten betrachten, ver-suchen omics-Ansätze einenmöglichst weitreichendenÜberblick zu geben. Die Sys-tembiologie vereint die nötigenbiologischen, physikalischen
und mathematischen Ansätze.Sie versucht, die erfassten ver-schiedenen Ebenen und ihreInteraktion zu beschreiben, umso die Zelle in ihrer Gesamtheitzu erfassen.
gewonnener Mutanten zu erhalten und durchVergleich dieser Sequenzen mit dem Wildtyp-Genom Mutationen zu identifizieren, die für dieProduktion wichtig sind. Die so aus Genomver-gleichen gewonnenen Mutationen können dannin das Wildtyp-Genom eingebaut werden, so-dass nur diejenigen Mutationen vorliegen, dieeine erhöhte Produktbildung bewirken, und da-gegen diejenigen, die das Wachstum verzögern,ausgemerzt sind.
6.3 L-Glutamat
6.3.1 Syntheseweg und Regulation
Das Natriumsalz von L-Glutamat ist Trägerder spezifischen Geschmackskomponente „u-mami“, wobei die molekulare Erkennung vonNatriumglutamat durch spezifische Rezeptorenunserer Zunge ausgelöst wird. Die Empfindung„umami“ wird noch zusätzlich durch die Ribo-nukleotide Inosinmonophosphat und Guanosin-monophosphat verstärkt und entsprechend alsGeschmacksverstärker eingesetzt. Während dieRibonukleotide mit Mutanten von Corynebacte-rium ammoniagenes produziert werden, erfolgtdie Produktion von L-Glutamat ausschließlichmit Corynebacterium glutamicum. Die Entde-ckung dieses Bakteriums in Japan Mitte desletzten Jahrhunderts, zusammen mit der Beob-achtung, dass es L-Glutamat ausscheiden kann,war der Beginn der mittlerweile zu großer Blü-
te gelangten mikrobiellen Aminosäureindustrie(Kap. 1).
Der aufgenommene Zucker wird in C. gluta-micum über die Glykolyse und den Pentosephos-phatweg zu Phosphoenolpyruvat bzw. Pyruvatverstoffwechselt. Pyruvat wird zu Acetyl-CoAdecarboxyliert, was anschließend zusammen mitOxalacetat zuCitrat umgesetzt und in denTricar-bonsäurezyklus eingeschleust wird (� Abb. 6.5).Dessen Intermediat α-Ketoglutarat wird durchdie Glutamat-Dehydrogenase reduktiv mit Am-monium zu L-Glutamat umgesetzt. Das beidieser Reaktion benötigte NADPH + H+ stehtdurch die vorhergehende Decarboxylierung vonIsocitrat zur Verfügung. L-Glutamat ist der zen-trale Metabolit, um Aminogruppen für Bio-synthesezwecke zur Verfügung zu stellen, wiez. B. für Transaminase-Reaktionen, und liegt imCytosol von C. glutamicum in hoher Konzen-tration von etwa 150mmol/l vor. Neben derGlutamat-Dehydrogenase besitzt C. glutamicumals weiteres System zur Ammoniumassimila-tion zwar noch die Glutamin-Synthetase undGlutamin-α-Ketoglutarat-Aminotransferase, wieMarkierungsexperimente aber zeigten, trägt denHauptanteil der Glutamatbildung die Glutamat-Dehydrogenase, die in C. glutamicum eine hohespezifische Aktivität von 1,5 μmol pro min undmg Protein hat. Da α-Ketoglutarat bei der Bil-dung von L-Glutamat dem Tricarbonsäurezyklusentzogen wird, muss Oxalacetat durch Carboxy-lierung der C-Bausteine Pyruvat und Phospho-enolpyruvat nachgeliefert werden. Wie sich her-ausgestellt hat, besitzt C. glutamicum dafür zwei
116 Kapitel 6 ⋅Aminosäuren
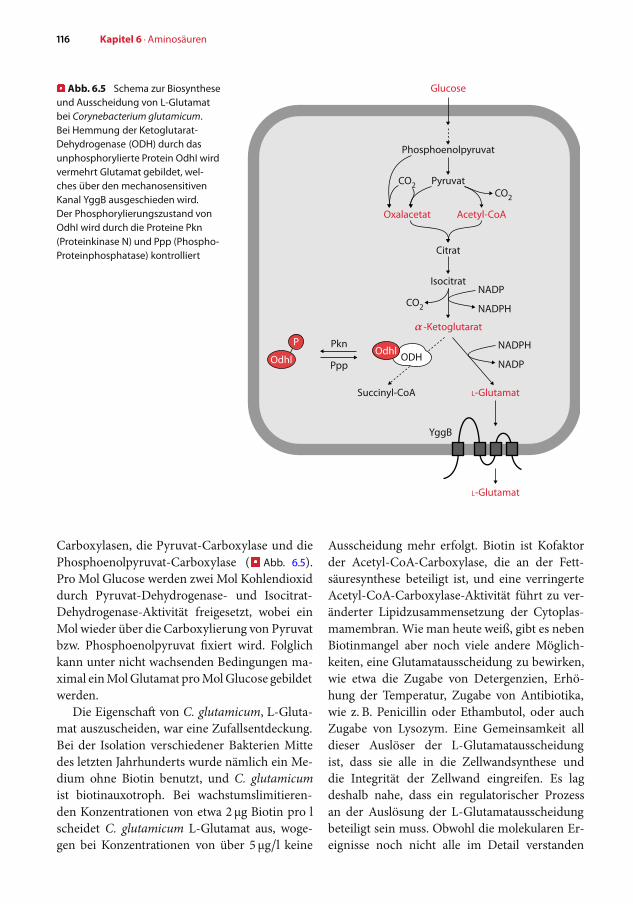
� Abb. 6.5 Schema zur Biosyntheseund Ausscheidung von L-Glutamatbei Corynebacterium glutamicum.Bei Hemmung der Ketoglutarat-Dehydrogenase (ODH) durch dasunphosphorylierte Protein OdhI wirdvermehrt Glutamat gebildet, wel-ches über den mechanosensitivenKanal YggB ausgeschieden wird.Der Phosphorylierungszustand vonOdhI wird durch die Proteine Pkn(Proteinkinase N) und Ppp (Phospho-Proteinphosphatase) kontrolliert
Glucose
Phosphoenolpyruvat
Pyruvat
Oxalacetat
CO2CO2
CO2
NADP
NADPH
Acetyl-CoA
Citrat
Isocitrat
Succinyl-CoA
YggB
α -Ketoglutarat
L-Glutamat
L-Glutamat
Pkn
Ppp
NADPH
NADPODHOdhl
Odhl
P
Carboxylasen, die Pyruvat-Carboxylase und diePhosphoenolpyruvat-Carboxylase (� Abb. 6.5).Pro Mol Glucose werden zwei Mol Kohlendioxiddurch Pyruvat-Dehydrogenase- und Isocitrat-Dehydrogenase-Aktivität freigesetzt, wobei einMol wieder über die Carboxylierung von Pyruvatbzw. Phosphoenolpyruvat fixiert wird. Folglichkann unter nicht wachsenden Bedingungen ma-ximal einMolGlutamat proMolGlucose gebildetwerden.
Die Eigenschaft von C. glutamicum, L-Gluta-mat auszuscheiden, war eine Zufallsentdeckung.Bei der Isolation verschiedener Bakterien Mittedes letzten Jahrhunderts wurde nämlich ein Me-dium ohne Biotin benutzt, und C. glutamicumist biotinauxotroph. Bei wachstumslimitieren-den Konzentrationen von etwa 2 μg Biotin pro lscheidet C. glutamicum L-Glutamat aus, woge-gen bei Konzentrationen von über 5 μg/l keine
Ausscheidung mehr erfolgt. Biotin ist Kofaktorder Acetyl-CoA-Carboxylase, die an der Fett-säuresynthese beteiligt ist, und eine verringerteAcetyl-CoA-Carboxylase-Aktivität führt zu ver-änderter Lipidzusammensetzung der Cytoplas-mamembran. Wie man heute weiß, gibt es nebenBiotinmangel aber noch viele andere Möglich-keiten, eine Glutamatausscheidung zu bewirken,wie etwa die Zugabe von Detergenzien, Erhö-hung der Temperatur, Zugabe von Antibiotika,wie z. B. Penicillin oder Ethambutol, oder auchZugabe von Lysozym. Eine Gemeinsamkeit alldieser Auslöser der L-Glutamatausscheidungist, dass sie alle in die Zellwandsynthese unddie Integrität der Zellwand eingreifen. Es lagdeshalb nahe, dass ein regulatorischer Prozessan der Auslösung der L-Glutamatausscheidungbeteiligt sein muss. Obwohl die molekularen Er-eignisse noch nicht alle im Detail verstanden

6.3 ⋅ L-Glutamat 117
6
sind, haben neueste Untersuchungen gezeigt,dass tatsächlich ein regulatorisches Protein ei-ne Schlüsselrolle spielt. Es handelt sich dabeium ein kleines Protein, OdhI, das phospho-ryliert oder unphosphoryliert vorliegen kann(�Abb. 6.5). Der Phosphorylierungszustand vonOdhI wird durch Proteinkinasen, Pkn (Protein-kinase N), bzw. eine Phosphatase, Ppp (Phospho-Proteinphosphatase), kontrolliert, die jeweilsin der Cytoplasmamembran lokalisiert sind.Diese Enzyme detektieren offensichtlich den Zu-stand der Zellwand. Bei desintegrierter Zellwandliegt weniger unphosphoryliertes OdhI vor. Indieser unphosphorylierten Form bindet OdhIan die OdhA-Untereinheit der Ketoglutarat-Dehydrogenase, wodurch die Aktivität des En-zyms bis auf eine geringe Restaktivität reduziertwird. Dadurch wird α-Ketoglutarat nicht mehrzu Succinyl-CoA umgewandelt, sondern stehtvermehrt zur Glutamatbildung zur Verfügung.
Der Export von Glutamat erfolgt durch einenmechanosensitiven Kanal. Dieser Kanal, YggB,besitzt vier transmembrane Helices. Er dient ver-mutlich wie vergleichbare Kanäle in E. coli zurAufrechterhaltung des Zellinnendrucks und derOsmoregulation. C. glutamicum kommt im Bo-den vor und muss deswegen in der Lage sein,auf plötzliche Veränderungen der osmotischenKonzentration zu reagieren. Setzt z. B. nach Tro-ckenheit Regen ein, strömt Wasser in die Zelle,was zu ihrem Platzen führen könnte, wenn nichtübermechanosensitive Kanäle ein schneller Aus-strom osmotisch aktiver Substanzen möglich wä-re. Da bekannt ist, dass mechanosensitive Kanälein Abhängigkeit von der Membranspannung so-wie auch von der Lipidumgebung öffnen undschließen, scheinen die verschiedenen Auslöserder Glutamatausscheidung auch den Kanal zubeeinflussen. Die Deletion des mechanosensiti-ven Kanals YggB in C. glutamicum verringert dieGlutamatausscheidung stark, und Mutanten vonC. glutamicum, die sehr gut Glutamat ausschei-den, tragen Mutationen in diesem Protein. Zu-sammengenommen wird inC. glutamicumdurchBeeinflussung der Zellwand eine Kette von Er-eignissen ausgelöst, die sowohl die Synthese vonGlutamat als auch dessen Export begünstigen.
6.3.2 Produktionsprozess
Die wichtigsten Prozessparameter, welche dieGlutamat-Produktion beeinflussen, sind nebender Biotinkonzentration der pH-Wert, die Tem-peratur sowie die Sauerstoff- und Ammonium-versorgung. Zu Beginn der Fermentation wirdder pH-Wert im Medium mit Ammoniak auf8,5 eingestellt. Da durch die Glutamatausschei-dung das Medium angesäuert wird, muss Basezugeführt werden. Dies erfolgt im Verlauf derFermentation durch kontinuierliche Zufuhr vonAmmoniak. Dadurchwird der pH-Wert konstantauf 7,8 gehalten und gleichzeitig der notwen-dige Stickstoff zugeführt. Auch Glucose wirdbei der großtechnischen Glutamat-Herstellungweitgehend kontinuierlich zugegeben. Für eineoptimale Glutamatausbeute ist auch eine ausrei-chende Sauerstoffversorgung erforderlich. UnterSauerstoff-limitierenden Bedingungen werdennämlich neben Glutamat noch Lactat und Succi-nat gebildet. Generell kommtder Prozessführungin der großtechnischen Produktion eine großeRolle zu, wie dies in Kap. 2 dieses Buches darge-stellt wird.
Die Glutamat-Produktion mit C. glutami-cum wird in gerührten Bioreaktoren mit einemVolumen bis zu 500m ausgeführt, die mit ver-schiedenen Mess- und Regeleinheiten ausgestat-tet sind. Nach der Wachstumsphase wird dieProduktion z. B. durch die Zugabe von Deter-genzien oder durch Erhöhung der Temperaturvon ○C auf ○C induziert. Nach etwa zweiTagen wird die Fermentation beendet, und dieGlutamatausbeute liegt dann bei 60 bis 70%bezogen auf die umgesetzte Glucose. Nach Ab-trennung der Zellen wird der Kulturüberstand,der das Ammoniumsalz des L-Glutamats enthält,zur Aufarbeitung auf einen Anionenaustauschergegeben. Glutamat bindet, und das frei werden-de Ammonium wird nach Destillation erneut indie Fermentation eingesetzt. Vom Ionenaustau-scher wird mittels NaOH das Natriumsalz desGlutamats eluiert. Es wird kristallisiert und liegtnach weiteren Schritten wie Bleichen und Siebenin Lebensmittelqualität vor.

118 Kapitel 6 ⋅Aminosäuren
� Abb. 6.6 Schema zur L-Lysin-synthese in C. glutamicum. In demverzweigten Syntheseweg wird dasSchlüsselenzym Aspartatkinasedurch die Endprodukte L-Lysin plusL-Threonin in seiner Aktivität ge-hemmt (punktierte Linie). Der Exportvon L-Lysin durch die Cytoplas-mamembran erfolgt durch denTransporter LysE
L-Aspartat
Dihydrodipicolinat
D, L-Diaminopimelat
L, L-Diaminopimelat
Succinyldiaminopimelat
Succinylaminoketopimelat
L-Lysin
L-Lysin
L-Methionin L-Threonin
L-Isoleucin
L-Piperidin-2,6-dicarboxylat
Dihydro-dipicolinat-
Synthase
Aspartat-semialdehyd
Transaminase
Dehydrogenase
Aspartatkinase
Succinylase
Desuccinylase
Epimerase
Exporter(lysE)
Transaminase
Homoserin- Dehydrogenase (hom)
Oxalacetat
Pyruvat
6.4 L-Lysin
6.4.1 Syntheseweg und Regulation
L-Lysin ist nach L-Glutamat die wichtigstefermentativ hergestellte Aminosäure auf demWeltmarkt. Es wird hauptsächlich für Schwei-ne- und Hühnerfutter-Formulierungen benutzt(� Tab. 6.1). Die Aminosäure wird mit C. glutami-cum, zum Teil aber auch mit E. coli produziert.Bausteine des Kohlenstoffskeletts von L-Lysinsind Oxalacetat und Pyruvat, die im Zentral-stoffwechsel bereitgestellt werden. Oxalacetatwird zunächst durch Transaminierung zu L-Aspartat umgewandelt (� Abb. 6.6). Aspartatist Substrat der Aspartatkinase, die ein sehrwichtiges Kontrollelement der Synthese dar-
stellt. Im Wildtyp von C. glutamicum ist dieAspartatkinase-Aktivität bei gleichzeitiger An-wesenheit von L-Lysin plus L-Threonin in einerKonzentration von je etwa 1mmol/l nahezuvollständig gehemmt. Eine weitere wichtige Kon-trolle innerhalb des verzweigten Synthesewegesfindet auf der Stufe des Aspartatsemialdehydsstatt. Aspartatsemialdehyd wird einerseits mitPyruvat durch die Dihydrodipicolinat-Synthasein den spezifischen L-Lysinsyntheseweg ein-geschleust, andererseits für die Synthese vonL-Threonin, L-Methionin und L-Isoleucin ver-wendet. An dieser Verzweigung wird der Flussin die Lysinsynthese durch eine begrenzendeDihydrodipicolinat-Synthase-Aktivität limitiert.Der Fluss in Richtung Threoninsynthese wirddurch Hemmung der Homoserin-Dehydrogena-se-Aktivität durch L-Threonin kontrolliert und

6.4 ⋅ L-Lysin 119
6
zusätzlich noch durch Repression des entspre-chenden hom-Gens durch L-Methionin. Ausge-hend von L-Piperidin-2,6-dicarboxylat erfolgtin der weiteren L-Lysinsynthese der Einbau derzweiten Aminogruppe. Dies geschieht entwederüber den Dehydrogenase- oder den Transami-naseweg. Im Dehydrogenaseweg wird in einerEinschrittreaktion die Aminogruppe direkt überdie Dehydrogenase eingebaut. Im Transamina-seweg dagegen wird Glutamat als Aminodonorbenutzt, und derWeg erfolgt insgesamt über vierSchritte und ist deswegen aufwendiger. Die pa-rallelen Wege ermöglichen C. glutamicum einesehr flexible Anpassung an eine unterschiedlicheStickstoffverfügbarkeit, wie es z. B. im Boden derFall ist, da die Dehydrogenase nur eine geringeAffinität zuAmmoniumhat. Bei hoherAmmoni-umkonzentration erfolgt die Lysinsynthese überden Dehydrogenaseweg, wohingegen bei niedri-ger Ammoniumkonzentration der aufwendigereTransaminaseweg benutzt wird.
Das in der Zelle synthetisierte L-Lysin wirddurch das Exportprotein LysE aktiv ausgeschie-den. Da L-Lysin positiv geladen ist, ist dessenpassive Diffusion über die Cytoplasmamem-bran nicht möglich. Das Exportprotein ist miteinem Molekulargewicht von 25 081 relativ kleinund besitzt sechs Transmembranhelices. Höchst-wahrscheinlich ist der Exporter als Dimer aktiv,da bekannt ist, dass Transporter zur Funktionmindestens zehn Transmembranhelices besit-zen müssen und für verwandte Proteine vonLysE der Aufbau als Dimer nachgewiesen ist.Die Synthese von LysE wird durch den RegulatorLysG kontrolliert. Erst bei erhöhter zellinter-ner L-Lysinkonzentration von über 20mmol/lbewirkt dieser Regulator die Transkription desExportergens und damit die Synthese des Ex-portproteins. Wie weitergehende Untersuchun-gen zeigten, dient dieses System im Wildtypdazu, eine Akkumulation von L-Lysin in derZelle zu verhindern, wenn es aus extern ver-fügbaren Peptiden stammt. Wie viele Bakteriennimmt C. glutamicum nämlich Peptide auf undhydrolysiert sie in ihre Aminosäurebausteine.Ist aber zu viel L-Lysin aus solchen Quellenvorhanden, kann es zellintern zu hohen wachs-
tumshemmenden Konzentrationen von übereinem Mol L-Lysin kommen, da C. glutamicumdiese Aminosäure nicht abbauen kann. Folg-lich trägt LysE zur Regulation der cytosolischenL-Lysinkonzentration unter natürlichen Bedin-gungen bei, wenn Peptide verwertet werden,wohingegen andere Bakterien dies durch Abbauvon L-Lysin erreichen.
6.4.2 Produktionsstämme
Eine signifikante L-Lysinbildung von etwa 20 g/lwurde schon vor vielen Jahren mit Mutantenvon C. glutamicum erhalten, die Homoserin-auxotroph sind. In diesen Stämmen kann dasZwischenproduktAspartatsemialdehyd nur nochzu Lysin umgesetzt werden. Bei Zugabe nied-riger Threoninkonzentrationen zum Mediumwird dann die Aspartatkinase trotz hoher Ly-sinkonzentration nicht gehemmt. Eine Über-produktion von Lysin erhält man außerdem mitMutanten von C. glutamicum, bei denen die As-partatkinase nicht mehr gehemmt wird. SolcheMutanten kann man unter Nutzung des Lysin-Antimetaboliten S-Aminoethylcystein (AEC)isolieren (�Abb. 6.3). AEC bindet im Wild-typ wie Lysin an das allosterische Zentrum derAspartatkinase, wodurch es zurWachstumshem-mung kommt. In AEC-resistenten Mutanten istdagegen das allosterische Zentrum der Aspartat-kinase so verändert, dass weder AEC noch Lysinbinden. Eine so veränderte Aspartatkinase un-terliegt somit keiner Feedback-Hemmung mehr,und entsprechend scheiden diese Mutanten vonC. glutamicum unter optimalen Kulturbedingun-gen bis zu 50 g/l Lysin in das Medium aus.
Wie bereits erwähnt, hat die Stammentwick-lung durch Genomanalysen von klassisch ge-wonnenen Produzenten eine neue Dimensi-on erreicht. Im Falle der L-Lysinbildung wur-den durch Genomsequenzierungen von klas-sisch gewonnenen Produzenten fünf für dieL-Lysinbildung entscheidende Mutationen iden-tifiziert. Durch spezifischenEinbau dieser Punkt-mutationen in dasWildtyp-Chromosomkann ei-
120 Kapitel 6 ⋅Aminosäuren
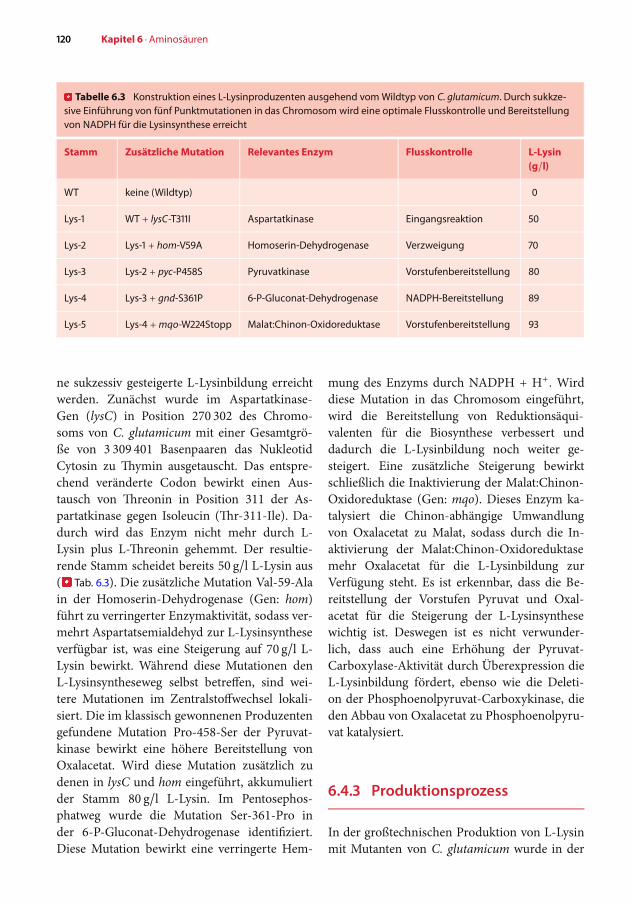
� Tabelle 6.3 Konstruktion eines L-Lysinproduzenten ausgehend vomWildtyp von C. glutamicum. Durch sukkze-sive Einführung von fünf Punktmutationen in das Chromosom wird eine optimale Flusskontrolle und Bereitstellungvon NADPH für die Lysinsynthese erreicht
Stamm Zusätzliche Mutation Relevantes Enzym Flusskontrolle L-Lysin(g/l)
WT keine (Wildtyp) 0
Lys-1 WT + lysC-T311I Aspartatkinase Eingangsreaktion 50
Lys-2 Lys-1 + hom-V59A Homoserin-Dehydrogenase Verzweigung 70
Lys-3 Lys-2 + pyc-P458S Pyruvatkinase Vorstufenbereitstellung 80
Lys-4 Lys-3 + gnd-S361P 6-P-Gluconat-Dehydrogenase NADPH-Bereitstellung 89
Lys-5 Lys-4 +mqo-W224Stopp Malat:Chinon-Oxidoreduktase Vorstufenbereitstellung 93
ne sukzessiv gesteigerte L-Lysinbildung erreichtwerden. Zunächst wurde im Aspartatkinase-Gen (lysC) in Position 270 302 des Chromo-soms von C. glutamicum mit einer Gesamtgrö-ße von 3 309 401 Basenpaaren das NukleotidCytosin zu Thymin ausgetauscht. Das entspre-chend veränderte Codon bewirkt einen Aus-tausch von Threonin in Position 311 der As-partatkinase gegen Isoleucin (Thr-311-Ile). Da-durch wird das Enzym nicht mehr durch L-Lysin plus L-Threonin gehemmt. Der resultie-rende Stamm scheidet bereits 50 g/l L-Lysin aus(� Tab. 6.3). Die zusätzliche Mutation Val-59-Alain der Homoserin-Dehydrogenase (Gen: hom)führt zu verringerter Enzymaktivität, sodass ver-mehrt Aspartatsemialdehyd zur L-Lysinsyntheseverfügbar ist, was eine Steigerung auf 70 g/l L-Lysin bewirkt. Während diese Mutationen denL-Lysinsyntheseweg selbst betreffen, sind wei-tere Mutationen im Zentralstoffwechsel lokali-siert. Die im klassisch gewonnenen Produzentengefundene Mutation Pro-458-Ser der Pyruvat-kinase bewirkt eine höhere Bereitstellung vonOxalacetat. Wird diese Mutation zusätzlich zudenen in lysC und hom eingeführt, akkumuliertder Stamm 80 g/l L-Lysin. Im Pentosephos-phatweg wurde die Mutation Ser-361-Pro inder 6-P-Gluconat-Dehydrogenase identifiziert.Diese Mutation bewirkt eine verringerte Hem-
mung des Enzyms durch NADPH + H+. Wirddiese Mutation in das Chromosom eingeführt,wird die Bereitstellung von Reduktionsäqui-valenten für die Biosynthese verbessert unddadurch die L-Lysinbildung noch weiter ge-steigert. Eine zusätzliche Steigerung bewirktschließlich die Inaktivierung der Malat:Chinon-Oxidoreduktase (Gen: mqo). Dieses Enzym ka-talysiert die Chinon-abhängige Umwandlungvon Oxalacetat zu Malat, sodass durch die In-aktivierung der Malat:Chinon-Oxidoreduktasemehr Oxalacetat für die L-Lysinbildung zurVerfügung steht. Es ist erkennbar, dass die Be-reitstellung der Vorstufen Pyruvat und Oxal-acetat für die Steigerung der L-Lysinsynthesewichtig ist. Deswegen ist es nicht verwunder-lich, dass auch eine Erhöhung der Pyruvat-Carboxylase-Aktivität durch Überexpression dieL-Lysinbildung fördert, ebenso wie die Deleti-on der Phosphoenolpyruvat-Carboxykinase, dieden Abbau von Oxalacetat zu Phosphoenolpyru-vat katalysiert.
6.4.3 Produktionsprozess
In der großtechnischen Produktion von L-Lysinmit Mutanten von C. glutamicum wurde in der
6.4 ⋅ L-Lysin 121
6
010 20
Zeit [h]0 30 40
0
50
100
150
200
10
20
30
L-Ly
sin
[g/l]W
achs
tum
Zuck
er [g
/l]
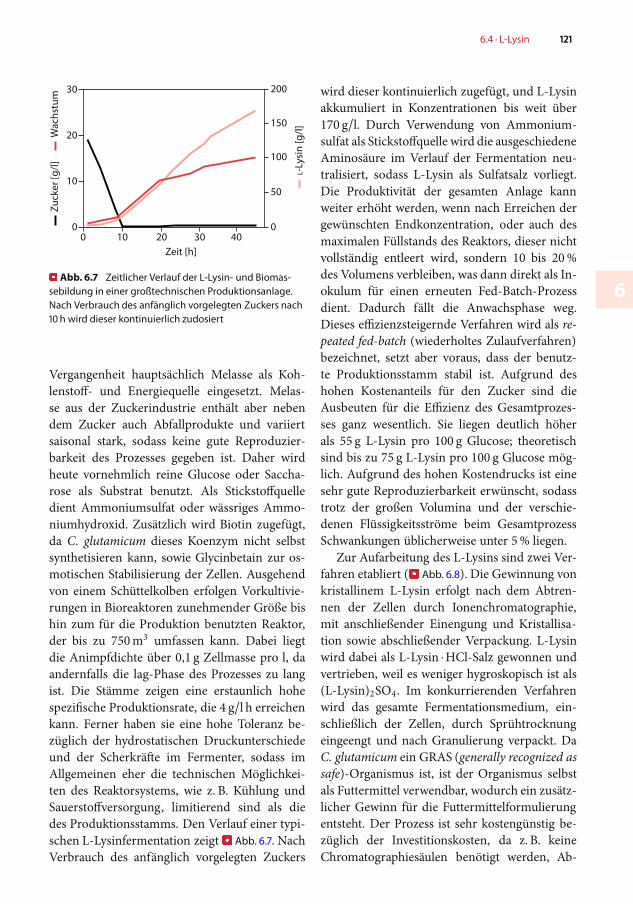
�Abb. 6.7 Zeitlicher Verlauf der L-Lysin- und Biomas-sebildung in einer großtechnischen Produktionsanlage.Nach Verbrauch des anfänglich vorgelegten Zuckers nach10 h wird dieser kontinuierlich zudosiert
Vergangenheit hauptsächlich Melasse als Koh-lenstoff- und Energiequelle eingesetzt. Melas-se aus der Zuckerindustrie enthält aber nebendem Zucker auch Abfallprodukte und variiertsaisonal stark, sodass keine gute Reproduzier-barkeit des Prozesses gegeben ist. Daher wirdheute vornehmlich reine Glucose oder Saccha-rose als Substrat benutzt. Als Stickstoffquelledient Ammoniumsulfat oder wässriges Ammo-niumhydroxid. Zusätzlich wird Biotin zugefügt,da C. glutamicum dieses Koenzym nicht selbstsynthetisieren kann, sowie Glycinbetain zur os-motischen Stabilisierung der Zellen. Ausgehendvon einem Schüttelkolben erfolgen Vorkultivie-rungen in Bioreaktoren zunehmender Größe bishin zum für die Produktion benutzten Reaktor,der bis zu 750m umfassen kann. Dabei liegtdie Animpfdichte über 0,1 g Zellmasse pro l, daandernfalls die lag-Phase des Prozesses zu langist. Die Stämme zeigen eine erstaunlich hohespezifische Produktionsrate, die 4 g/l h erreichenkann. Ferner haben sie eine hohe Toleranz be-züglich der hydrostatischen Druckunterschiedeund der Scherkräfte im Fermenter, sodass imAllgemeinen eher die technischen Möglichkei-ten des Reaktorsystems, wie z. B. Kühlung undSauerstoffversorgung, limitierend sind als diedes Produktionsstamms. Den Verlauf einer typi-schen L-Lysinfermentation zeigt�Abb. 6.7. NachVerbrauch des anfänglich vorgelegten Zuckers
wird dieser kontinuierlich zugefügt, und L-Lysinakkumuliert in Konzentrationen bis weit über170 g/l. Durch Verwendung von Ammonium-sulfat als Stickstoffquelle wird die ausgeschiedeneAminosäure im Verlauf der Fermentation neu-tralisiert, sodass L-Lysin als Sulfatsalz vorliegt.Die Produktivität der gesamten Anlage kannweiter erhöht werden, wenn nach Erreichen dergewünschten Endkonzentration, oder auch desmaximalen Füllstands des Reaktors, dieser nichtvollständig entleert wird, sondern 10 bis 20%des Volumens verbleiben, was dann direkt als In-okulum für einen erneuten Fed-Batch-Prozessdient. Dadurch fällt die Anwachsphase weg.Dieses effizienzsteigernde Verfahren wird als re-peated fed-batch (wiederholtes Zulaufverfahren)bezeichnet, setzt aber voraus, dass der benutz-te Produktionsstamm stabil ist. Aufgrund deshohen Kostenanteils für den Zucker sind dieAusbeuten für die Effizienz des Gesamtprozes-ses ganz wesentlich. Sie liegen deutlich höherals 55 g L-Lysin pro 100 g Glucose; theoretischsind bis zu 75 g L-Lysin pro 100 g Glucose mög-lich. Aufgrund des hohen Kostendrucks ist einesehr gute Reproduzierbarkeit erwünscht, sodasstrotz der großen Volumina und der verschie-denen Flüssigkeitsströme beim GesamtprozessSchwankungen üblicherweise unter 5% liegen.
Zur Aufarbeitung des L-Lysins sind zwei Ver-fahren etabliert (� Abb. 6.8). Die Gewinnung vonkristallinem L-Lysin erfolgt nach dem Abtren-nen der Zellen durch Ionenchromatographie,mit anschließender Einengung und Kristallisa-tion sowie abschließender Verpackung. L-Lysinwird dabei als L-Lysin ⋅HCl-Salz gewonnen undvertrieben, weil es weniger hygroskopisch ist als(L-Lysin)SO. Im konkurrierenden Verfahrenwird das gesamte Fermentationsmedium, ein-schließlich der Zellen, durch Sprühtrocknungeingeengt und nach Granulierung verpackt. DaC. glutamicum ein GRAS (generally recognized assafe)-Organismus ist, ist der Organismus selbstals Futtermittel verwendbar, wodurch ein zusätz-licher Gewinn für die Futtermittelformulierungentsteht. Der Prozess ist sehr kostengünstig be-züglich der Investitionskosten, da z. B. keineChromatographiesäulen benötigt werden, Ab-

122 Kapitel 6 ⋅Aminosäuren
Biomasse
H2SO4Ernte,
Inaktivierung
Mutter-lauge
Kondensat
Fermentationsmedium
(NH4)2SO4
NH3
HCl
Ionen-austausch
Evaporation
Kristallisation
Trocknung,Kühlung
L-Lysin-HClAusbeute 90 %
Verpackung
Ernte,Inaktivierung
Evaporation
Granulation
Trocknung,Kühlung
BiolysAusbeute 95 %
Verpackung
� Abb. 6.8 Zwei verschiedene Wege zur Aufarbeitungdes L-Lysins aus dem Fermentationsmedium
fallprodukte verhindert werden und geringeVerluste garantiert sind (� Abb. 6.8). Das gewon-nene Produkt wird als Biolys gehandelt.
6.5 L-Threonin
6.5.1 Syntheseweg und Regulation
L-Threonin wird als Futtermitteladditiv benutztund mit E. coli produziert. Die Synthese von L-Threonin erfolgt über einen nur kurzen Weg vonfünf Schritten (�Abb. 6.9). Allerdings wird dasIntermediat Aspartatsemialdehyd auch für dieL-Lysinsynthese benötigt und das IntermediatHomoserin für die L-Methioninsynthese. Dar-über hinaus ist L-Threonin auch Vorstufe der L-Isoleucinsynthese. Es ist einleuchtend, dass die-ser Syntheseweg, der in die Synthese von mehre-ren Aminosäuren eingebettet ist, einer besonde-ren Regulation bedarf. Dies ist in E. coli so gelöst,dass drei IsoenzymemitAspartatkinase-Aktivität
vorliegen, die jeweils spezifisch gehemmt wer-den: eines durch L-Threonin, eines durch L-Lysin und eines durch L-Methionin. Weiterhingibt es auch zwei Homoserin-Dehydrogenase-Aktivitäten, vondenendie eine durchL-Threoninund die andere durch L-Methionin gehemmtwird. Zusätzlich sind auch die entsprechen-den Gene in Transkriptionseinheiten angeord-net, sodass auch auf der Stufe der Genexpres-sion eine von der Aminosäureverfügbarkeitabhängige Synthese gewährleistet ist. Das fürdie L-Threonin-Produktion relevante Operon istthrABC. Dabei codiert thrA für ein bifunktionel-les Enzym mit Aspartatkinase- und Homoserin-Dehydrogenase-Aktivität, thrB für eine Ho-moserinkinase und thrC für eine Threonin-Synthase. Dieses Operon unterliegt einer starkenKontrolle der Transkription durch Attenuation.Das entsprechende Leaderpeptid vor thrABCist Thr-Ile-Thr-Thr-Thr-Ile-Thr-Ile-Thr-Thr. Inder Zelle wird über die Synthesegeschwindig-keit dieses Leaderpeptids der Beladungszustandvon tRNAThr und tRNAIle detektiert. Wenn dietRNAs nicht mit ihrer zugehörigen Aminosäurebeladen sind, kann keine Leaderpeptid-Bildungerfolgen, und die Transkription des Operonsist dann mindestens zehnfach erhöht. Dage-gen kommt es zum vorzeitigen Abbruch derTranskription, wenn viel beladene tRNAs vorlie-gen.
6.5.2 Produktionsstämme
Schlüssel der L-Threonin-Produktion mit E. colisind: L-Threonin!Produktionsstämme (1) die Ex-pression des thrABC-Operons mit nicht mehrhemmbaren Enzymaktivitäten; (2) Verhinde-rung der Verstoffwechselung von L-Threoninzu L-Isoleucin; (3) die Verhinderung des L-Threoninabbaus; und (4) der Export von L-Threonin.
Das Gen thrA codiert für eine Aspartat-kinase und Homoserin-Dehydrogenase. DieHemmung dieser Enzymaktivitäten durch L-Threonin wurde aufgehoben. Um auch eine

6.5 ⋅ L-Threonin 123
6
Aspartat-semialdehyd
HomoserinL-Met
L-Lys
L-Isoleucin
Homoserin-phosphat
L-Threonin
Aspartatkinase
Homoserin-dehydrogenasethrA
Homoserin-kinasethrB
L-Aspartat
Aspartyl-phosphat
Threonin-synthasethrC
Threonin-dehydratase
�Abb. 6.9 Schema zur L-Threoninsynthese in E. coliund ihre Verknüpfung mit der Synthese von L-Lysin,L-Methionin und L-Isoleucin. Parallele Pfeileweisen aufIsoenzyme hin. Feedback-Hemmungen von Enzymaktivi-täten sind durch punktierte Linien angegeben
hohe Expression zu gewährleisten, wurde derAttenuatorbereich des Operons deletiert, wasden vorzeitigen Abbruch der Transkription desthrABC-Operons verhindert. Da bei starkerL-Threoninsynthese auch L-Isoleucin gebildetwurde, wurde eine Mutante mit verringerterSubstrataffinität der Threonin-Dehydratase iso-liert. Diese Mutante benötigt zwar bei nied-rigen L-Threoninkonzentrationen die Zugabevon L-Isoleucin zum Medium; bei hohen L-Threoninkonzentrationen bildet sie aber geradeausreichend L-Isoleucin zum Wachstum. E. coliist in der Lage, L-Threonin abzubauen, und ver-wendet hierfür eine Threonin-Dehydrogenase.Inaktivierung des entsprechenden Gens, tdh,verbessert weiter die L-Threoninakkumulation.Eine kürzlich erfolgteweitere Stammentwicklungbetrifft die Beobachtung, dass Stämme, die sehrviel L-Threonin bilden, auch schlechter wach-
sen. Es gelang, Mutanten zu isolieren, die beiZugabe sehr hoher L-Threoninkonzentrationenwieder etwas besser wachsen als der Ausgangs-stamm. Wie sich herausgestellt hat, liegt diesan einem Basenaustausch im Promotorbereichdes Gens rhtA. Dieses Gen codiert für einenExportcarrier der DME (drug/metabolite expor-ter)-Familie mit zehn transmembranen Helices.Die Promotormutation führt nahezu zu einerVerdoppelung der L-Threoninexportrate, sodasszellinterne hohe L-Threoninkonzentrationenvermieden werden, die offensichtlich nachteiligfür die Zelle sind. Der imVergleich zuC. glutami-cum sehr effektive L-Threoninexport beiE. coli istmaßgeblicher Grund dafür, dass zur L-Threonin-Produktion Stämme von E. coli benutzt werden.
6.5.3 Produktionsprozess
Die Kultivierung des L-Threoninproduzentenerfolgt in einem einfachen Salzmedium mitGlucose oder Saccharose als Substrat. Nachdem Beimpfen und Verbrauch des vorgelegtenZuckers erfolgt dessen kontinuierliche Zugabe.Über pH-Kontrolle wird zusätzlich Ammoniakoder Ammoniumhydroxid als Stickstoffquellezugeführt. Die Fütterungsstrategie ist im Ver-gleich zur L-Lysinbildung einfach, da kein ba-sisches Produkt akkumuliert und ein Gegenionwie Sulfat nicht benötigt wird. Nach etwa zweibis drei Tagen ist die Fermentation beendet.L-Threonin liegt im Kulturmedium in Konzen-trationen von über 100 g/l vor, sodass es bereitsteilweise kristallisiert. Die auf den Zucker be-zogenen Ausbeuten betragen etwa 60%. DieAufarbeitung des Produktes ist wegen der ge-ringen Löslichkeit und der geringen Salzfrachtrelativ einfach. Nach Abtrennung der Zellenwird die L-Threonin-enthaltende Lösung etwaseingeengt und die Kristallisation der Amino-säure durch Abkühlen eingeleitet. Abtrennungund Trocknung der Kristalle liefert L-Threoninmit einer Reinheit von über 90%. Eine Rekris-tallisation wird durchgeführt, wenn hochreinesL-Threonin benötigt wird.

124 Kapitel 6 ⋅Aminosäuren
L-Cystein
L-Cystein ist eine schwefelhalti-ge Aminosäure, die bis Anfangdieses Jahrhunderts noch aus-schließlich aus Haar, das reichan L-Cystein ist, isoliert wurde.Durch Metabolic Engineeringist es gelungen, den Stoffwech-sel von E. coli so anzupassen,dass derzeit bereits etwa einViertel des produzierten L-Cysteins mikrobiell hergestelltwird. Eine besondere Heraus-
forderung war es dabei, diezellinterne Verfügbarkeit desSchwefels zu gewährleisten. AlsSchwefelquelle der mikrobiel-len Synthese dienen Sulfat oderThiosulfat, an deren Aufnahmeund Reduktion zum Sulfid zwölfGene beteiligt sind. Im Produk-tionsprozess mit E. coli wird eineFütterungsstrategie mit Thiosul-fat als Schwefelquelle benutzt.L-Cystein wird dabei in hohen
Konzentrationen von über 30 g/lgebildet, und es fällt unter denaeroben Kultivierungsbedin-gungen in Form des Dimers alsCystin aus. Dieses wird gereinigtund durch Elektrolyse wieder zuL-Cystein reduziert.(Weitere Informationen un-ter www.wacker.com/cms/de/wacker_group/services/special_cysteine/cysteine.jsp.)
6.6 L-Phenylalanin
6.6.1 Syntheseweg und Regulation
L-Phenylalanin gehört wie L-Tyrosin und L-Tryptophan zu den aromatischen Aminosäuren,die den Syntheseweg bis zum Chorismat ge-meinsam haben. Wie aus � Abb. 6.10 zu ersehenist, sind die Vorstufen für die Aromatenbiosyn-these Erythrose-4-phosphat und Phosphoenol-pyruvat, welche im ersten Schritt zu 3-Desoxy-D-arabinoheptulonsäure-7-phosphat (DAHP)umgesetzt werden. Sechs weitere Enzyme kata-lysieren die Umsetzung dieses ersten Zwischen-produktes zum Chorismat. Dieses Intermediatwird dann über Anthranilat zu L-Tryptophanoder über Prephenat zu L-Phenylalanin bzw. L-Tyrosin umgesetzt.
Dieser Aromatenbiosyntheseweg unterliegtin E. coli einer komplexen Regulation. So be-sitzt dieses Bakterium drei DAHP-Synthase-Isoenzyme, welche durch die Gene aroF, aroGund aroH codiert werden, wobei das IsoenzymAroG etwa 80% der gesamten DAHP-Synthese-Aktivität ausmacht. Die Synthese und Aktivi-tät jedes dieser drei Isoenzyme wird jeweilsdurch eine der drei aromatischen Aminosäu-ren reguliert, sodass in der Regel E. coli diese
Chorismat
Prephenat
L-Phenylalanin
DAHP-SynthasearoF, G, H
Chorismat-mutasepheA
Prephenat-dehydratasepheA
Erythrose-4-phosphat+ Phosphoenolpyruvat
L-Tyr
L-Trp
�Abb. 6.10 Schema zur Synthese der aromatischenAminosäuren und ihrer Regulation in E. coli. Feedback-Hemmungen von Enzymaktivitäten sind durch punktierteLinien angegeben. Das Schlüsselenzym 3-Desoxy-D-arabinoheptulonsäure-7-phosphat-Synthase ist alsDAHP-Synthase abgekürzt
Aminosäuren nicht überproduziert. Ein wei-terer wichtiger Schritt in der Regulation desPhenylalaninsynthese-Endzweigs ist das bifunk-tionale Enzym PheA mit Chorismatmutase- undPrephenat-Dehydratase-Aktivität, da diese Akti-vitäten durch L-Phenylalanin gehemmt werden.Zusätzlich wird die pheA-Genexpression auch

6.6 ⋅ L-Phenylalanin 125
6
durch den Beladungszustand der tRNAPhe durchAttenuation kontrolliert.
6.6.2 Produktionsstämme
Produzentenstämme haben Feedback-resistentedurch aroG oder aroF codierte DAHP-Aktivitätund Feedback-resistente durch pheA codierteEnzymaktivitäten. Generell sind ProduzentenL-Tyrosin-auxotroph. Ein Grund dafür ist, dasswegen des nur zwei Schritte umfassenden Wegesvon Prephenat zu L-Tyrosin bei sehr starkerL-Phenylalaninbildung auch geringe MengenL-Tyrosin gebildet würden, was natürlich un-erwünscht ist. Ein weiterer Grund ist, dass we-gen der L-Tyrosin-Auxotrophie das Wachstumüber die Zugabe von L-Tyrosin gezielt gesteuertwerden kann (Abschn. 6.6.3). In einigen E. coli-Stämmen wird über einen Temperatur-sensitivenRepressor des Bakteriophagen λ zusammen mitdem λ PL-Promotor eine induzierbare Expressi-on von pheA und aroF erreicht. Somit wird esmöglich, durch Temperaturerhöhung eine sehrhohe Enzymaktivität in der Produktionspha-se zu erreichen, wodurch Stabilitätsproblemeaufgrund hoher Enzymkonzentrationen und-aktivitäten reduziert werden. Dadurch könnendie Kultivierungsschritte bis zum Animpfen desBioreaktors bei niedriger Expression der Schlüs-selgene erfolgen und erst in der tatsächlichenFermentation die Gene stark exprimiert wer-den.
6.6.3 Produktionsprozess
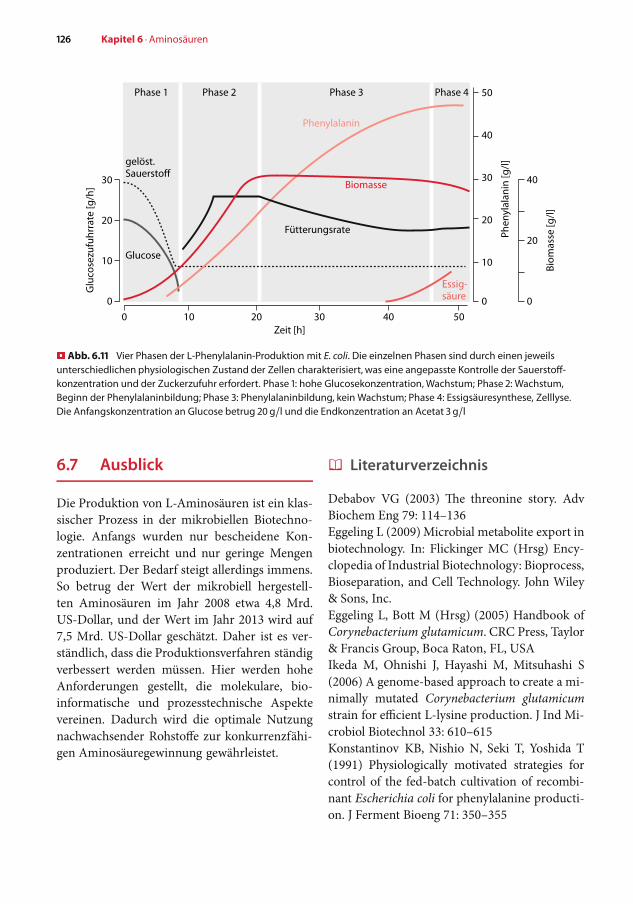
Für eine hohe L-Phenylalaninbildung ist eineausgeklügelte Kontrolle des Stoffwechsels in derFermentation nötig. Dies hat zwei Gründe: Zumeinen muss der Kohlenstofffluss optimal auf dievier Produkte der Glucose-Verstoffwechslungverteilt werden – L-Phenylalanin, die Zellmas-se, Essigsäure und CO. Zum anderen ändertsich die Aktivität der Zellen im Verlauf der Kul-tur ständig, sodass die Prozessparameter bei
der Kultivierung fortwährend angepasst wer-den müssen. � Abbildung 6.11 zeigt den typischenVerlauf einer L-Phenylalanin-Produktion. DasHauptproblem ist, dass E. coli bei Glucoseüber-schuss oder Sauerstoffmangel dazu neigt, Essig-säure auszuscheiden. Um dies zu verhindern,wurde eine adaptive Zuckerzufuhr-Strategieentwickelt, die on-line Daten und Flüsse zumSauerstoffverbrauch, der Zuckerkonzentrationund der Biomassekonzentration erfasst. Die-se Daten dienen der Anpassung während desProzesses an die optimale Zuckerzufuhr. NachVerbrauch des anfänglich vorgelegten Zuckers inPhase 1 treten die Zellen in Phase 2 ein, in derder Zucker kontinuierlich zugegeben wird. Dadie Biomasse zunimmt, steigt auch die Rate derZuckerzufuhr in dieser Phase. Durch die adap-tive Zuckerzufuhr-Strategie wird eine zu hoheGlucosekonzentration vermieden, da dies wieerwähnt zur Essigsäurebildung führen würde.Gleichzeitig wird aber auch eine bestimmte Glu-cosekonzentration nicht unterschritten, da sonsteine starke CO-Bildung auftreten würde. Wenndas anfangs vorhandene L-Tyrosin verbrauchtist, treten die Zellen in Phase 3 ein. Wie erwähnt,sind alle L-Phenylalanin-Produzenten auxotrophfür L-Tyrosin. Die zu Beginn der Fermentationvorgelegte L-Tyrosinkonzentration bestimmt so-mit die Konzentration der Biomasse in Phase 3,mit der nun Glucose effizient zu L-Phenylalaninumgewandelt wird. In Phase 3 geht die Stoff-wechselkapazität der Zellen teilweise zurück,was folglich auch zu verringerter Zuckerzufuhrführt. Wegen weiter nachlassender Stoffwech-selaktivität der Zellen beginnt schließlich einesignifikante Ausscheidung von Essigsäure. DieZellen befinden sich in Phase 4, in der kei-ne wesentliche L-Phenylalanin-Akkumulationmehr erfolgt, sodass der Prozess schließlich ab-gebrochenwird.Dieses Beispiel derAminosäure-Produktion zeigt, dass durch eine ausgeklügelteFütterungsstrategie mit adaptiver Kontrolle einesehr hohe L-Phenylalaninkonzentration inner-halb von nur 2,5 Tagen erreicht werden kann.Werte von 50 g/l L-Phenylalanin bei einer Aus-beute von 28%, bezogen auf den eingesetztenZucker, sind berichtet.
126 Kapitel 6 ⋅Aminosäuren
Glucose
Phase 1 Phase 2 Phase 3 Phase 4
gelöst.Sauerstoff
Fütterungsrate
Biomasse
Phenylalanin
Essig-säure0 0 0
20
40
10
20
30
40
50
10
20
30
100 20Zeit [h]
Glu
cose
zufu
hrra
te [g
/h]
Phen
ylal
anin
[g/l]
Biom
asse
[g/l]
30 40 50
� Abb. 6.11 Vier Phasen der L-Phenylalanin-Produktion mit E. coli. Die einzelnen Phasen sind durch einen jeweilsunterschiedlichen physiologischen Zustand der Zellen charakterisiert, was eine angepasste Kontrolle der Sauerstoff-konzentration und der Zuckerzufuhr erfordert. Phase 1: hohe Glucosekonzentration, Wachstum; Phase 2: Wachstum,Beginn der Phenylalaninbildung; Phase 3: Phenylalaninbildung, kein Wachstum; Phase 4: Essigsäuresynthese, Zelllyse.Die Anfangskonzentration an Glucose betrug 20 g/l und die Endkonzentration an Acetat 3 g/l
6.7 Ausblick
Die Produktion von L-Aminosäuren ist ein klas-sischer Prozess in der mikrobiellen Biotechno-logie. Anfangs wurden nur bescheidene Kon-zentrationen erreicht und nur geringe Mengenproduziert. Der Bedarf steigt allerdings immens.So betrug der Wert der mikrobiell hergestell-ten Aminosäuren im Jahr 2008 etwa 4,8 Mrd.US-Dollar, und der Wert im Jahr 2013 wird auf7,5 Mrd. US-Dollar geschätzt. Daher ist es ver-ständlich, dass die Produktionsverfahren ständigverbessert werden müssen. Hier werden hoheAnforderungen gestellt, die molekulare, bio-informatische und prozesstechnische Aspektevereinen. Dadurch wird die optimale Nutzungnachwachsender Rohstoffe zur konkurrenzfähi-gen Aminosäuregewinnung gewährleistet.
b Literaturverzeichnis
Debabov VG (2003) The threonine story. AdvBiochem Eng 79: 114–136Eggeling L (2009)Microbial metabolite export inbiotechnology. In: Flickinger MC (Hrsg) Ency-clopedia of Industrial Biotechnology: Bioprocess,Bioseparation, and Cell Technology. John Wiley& Sons, Inc.Eggeling L, Bott M (Hrsg) (2005) Handbook ofCorynebacterium glutamicum. CRC Press, Taylor& Francis Group, Boca Raton, FL, USAIkeda M, Ohnishi J, Hayashi M, Mitsuhashi S(2006) A genome-based approach to create a mi-nimally mutated Corynebacterium glutamicumstrain for efficient L-lysine production. J Ind Mi-crobiol Biotechnol 33: 610–615Konstantinov KB, Nishio N, Seki T, Yoshida T(1991) Physiologically motivated strategies forcontrol of the fed-batch cultivation of recombi-nant Escherichia coli for phenylalanine producti-on. J Ferment Bioeng 71: 350–355