Embed Size (px)
Citation preview

Inhalt der Vorlesung bis Weihnachten
1
1.Aminosäuren2.Peptide und Proteine3.Enzyme & Cofaktoren4.Kohlenhydrate5.Lipide6.Nukleotide7.Zellorganellen8.Replikation und Transkription9.Translation, Proteinexpression, Gentechnik

1850 Louis Pasteur: Vitalismus 1860-1917 Eduard Buchner:zellfreie alkoholische Gärung
2
Pioniere der Enzymologie2. Enzyme und Cofaktoren

1926 James Batcheller Sumner: UreaseEnzyme sind Proteine
John Burdon Sanderson Haldane:Konzept der enzymatischen Katalyse
3
Pioniere der Enzymologie2. Enzyme und Cofaktoren
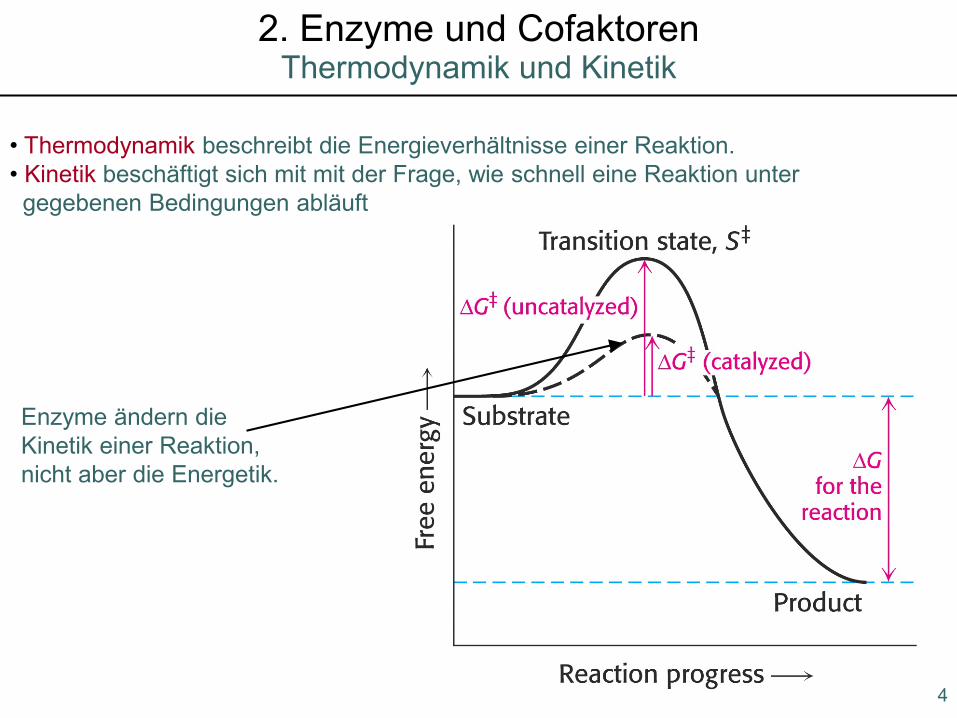
• Thermodynamik beschreibt die Energieverhältnisse einer Reaktion. • Kinetik beschäftigt sich mit mit der Frage, wie schnell eine Reaktion unter gegebenen Bedingungen abläuft
Enzyme ändern dieKinetik einer Reaktion,nicht aber die Energetik.
4
Thermodynamik und Kinetik2. Enzyme und Cofaktoren
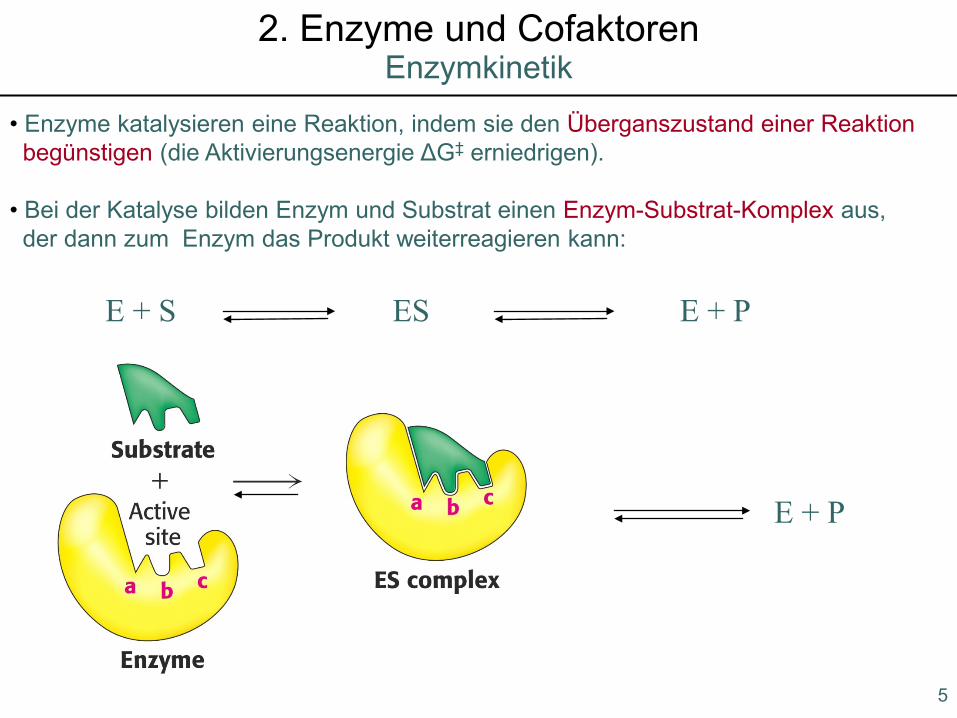
• Enzyme katalysieren eine Reaktion, indem sie den Überganszustand einer Reaktion begünstigen (die Aktivierungsenergie ΔG‡ erniedrigen).
• Bei der Katalyse bilden Enzym und Substrat einen Enzym-Substrat-Komplex aus, der dann zum Enzym das Produkt weiterreagieren kann:
E + S ES E + P
E + P
5
Enzymkinetik2. Enzyme und Cofaktoren

Faustregeln: - alle 10°C Verdoppelung von v0- ∆G‡ um 6 kJ niedriger 10 × schnellere Reaktion
6
Temperatur und Aktivierungsenergie2. Enzyme und Cofaktoren
Geschwindigkeitsgesetz: S → P
empirische Aktivierungsenergie(Arrhenius-Gleichung )
Reaktionskinetik – Einfluss von Temperatur und Aktivierungsenergie
freie Aktivierungsenthalpie(Theorie des Übergangszustandes, Eyring)
][d
][d SktSv =−=
RTEaeAk /−=
RTGeh
kTk /‡∆−=
RTGevv /maxo,o
‡∆−=
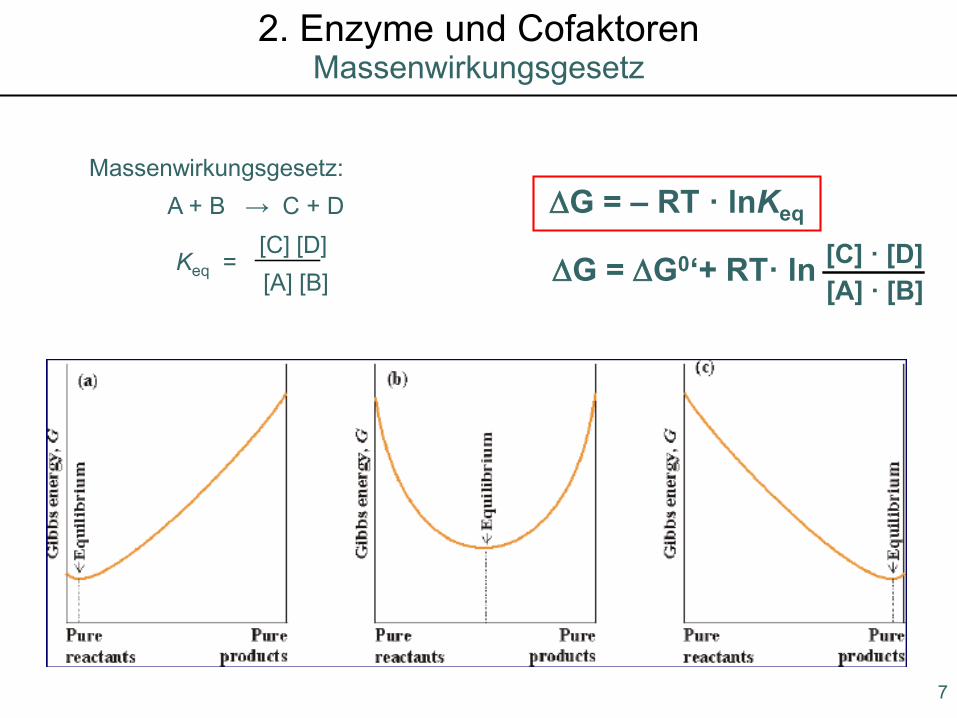
Massenwirkungsgesetz:∆G = – RT · lnKeqA + B → C + D
Keq =[C] [D][A] [B] ∆G = ∆G0‘+ RT· ln [C] · [D]
[A] · [B]
7
Massenwirkungsgesetz2. Enzyme und Cofaktoren
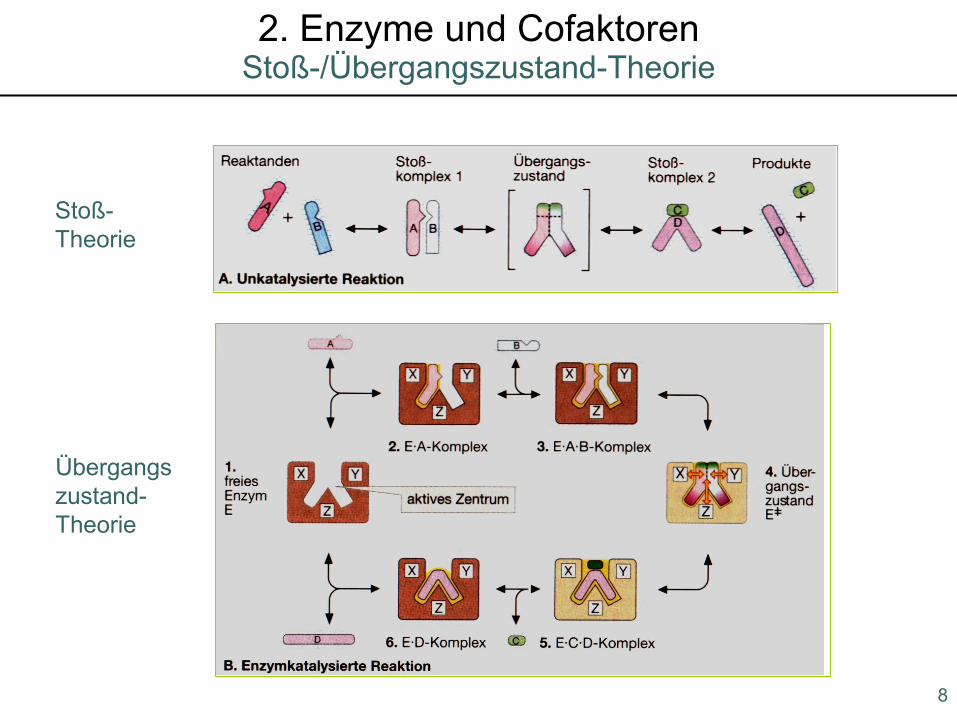
Stoß-Theorie
Übergangszustand-Theorie
8
Stoß-/Übergangszustand-Theorie2. Enzyme und Cofaktoren

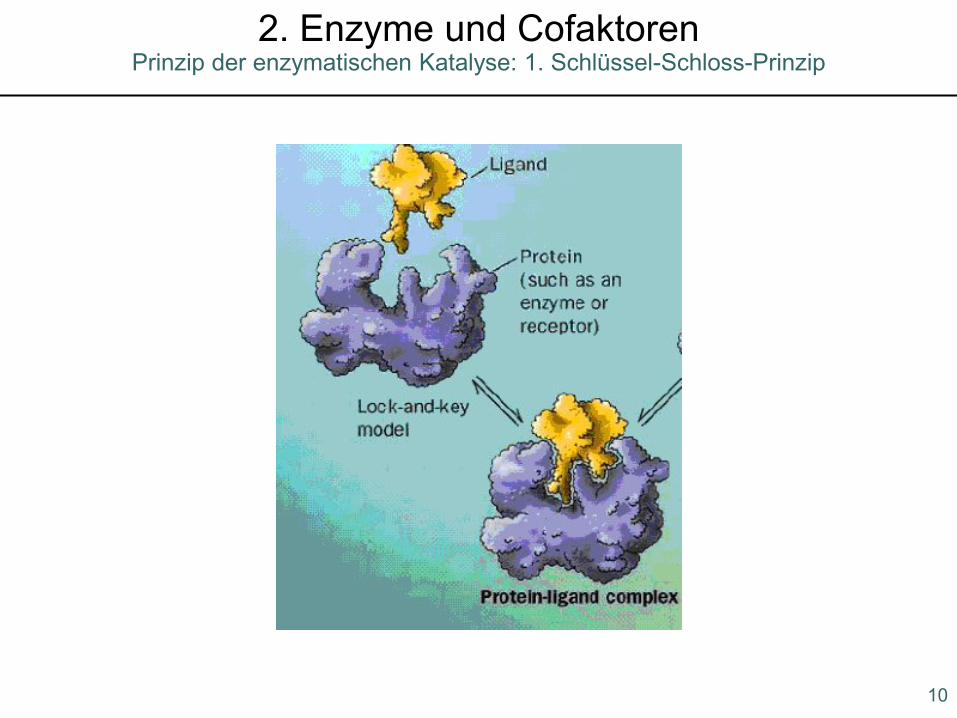
1. Schlüssel-Schloss-Prinzip
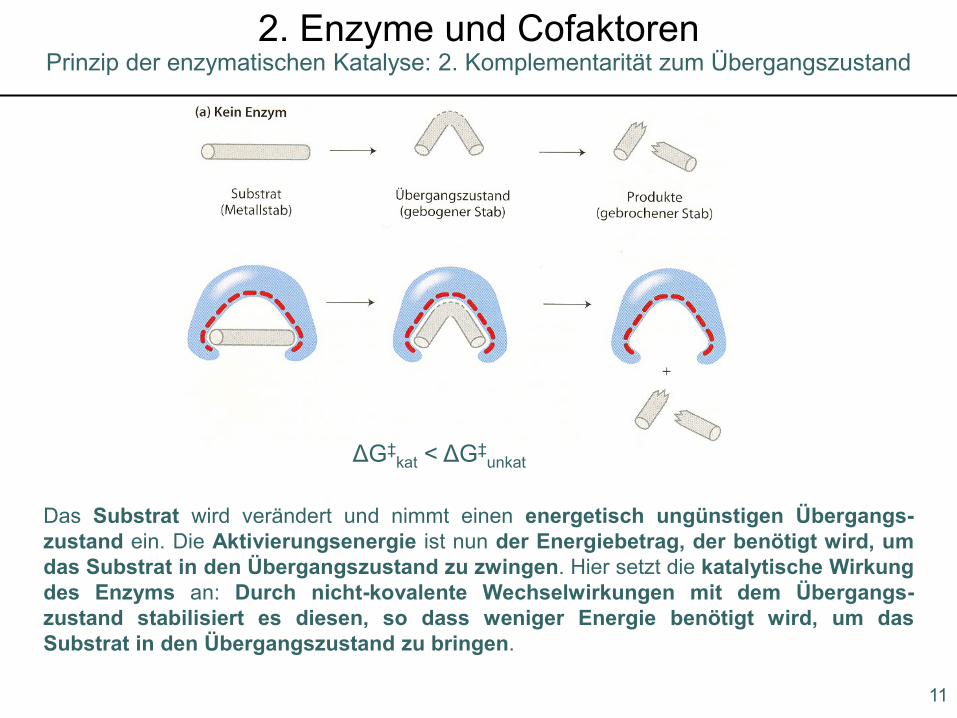
2. Komplementarität zum Übergangszustand
4. Verlust an Entropie
5. Desolvatisierung, Induced fit
3. Bindungsenergie
6. Elementarschritte im katalytischen Zyklus
9
Prinzip der enzymatischen Katalyse2. Enzyme und Cofaktoren
10
Prinzip der enzymatischen Katalyse: 1. Schlüssel-Schloss-Prinzip2. Enzyme und Cofaktoren
11
Prinzip der enzymatischen Katalyse: 2. Komplementarität zum Übergangszustand2. Enzyme und Cofaktoren
ΔG‡kat < ΔG‡
unkat
Das Substrat wird verändert und nimmt einen energetisch ungünstigen Übergangs-zustand ein. Die Aktivierungsenergie ist nun der Energiebetrag, der benötigt wird, umdas Substrat in den Übergangszustand zu zwingen. Hier setzt die katalytische Wirkungdes Enzyms an: Durch nicht-kovalente Wechselwirkungen mit dem Übergangs-zustand stabilisiert es diesen, so dass weniger Energie benötigt wird, um dasSubstrat in den Übergangszustand zu bringen.
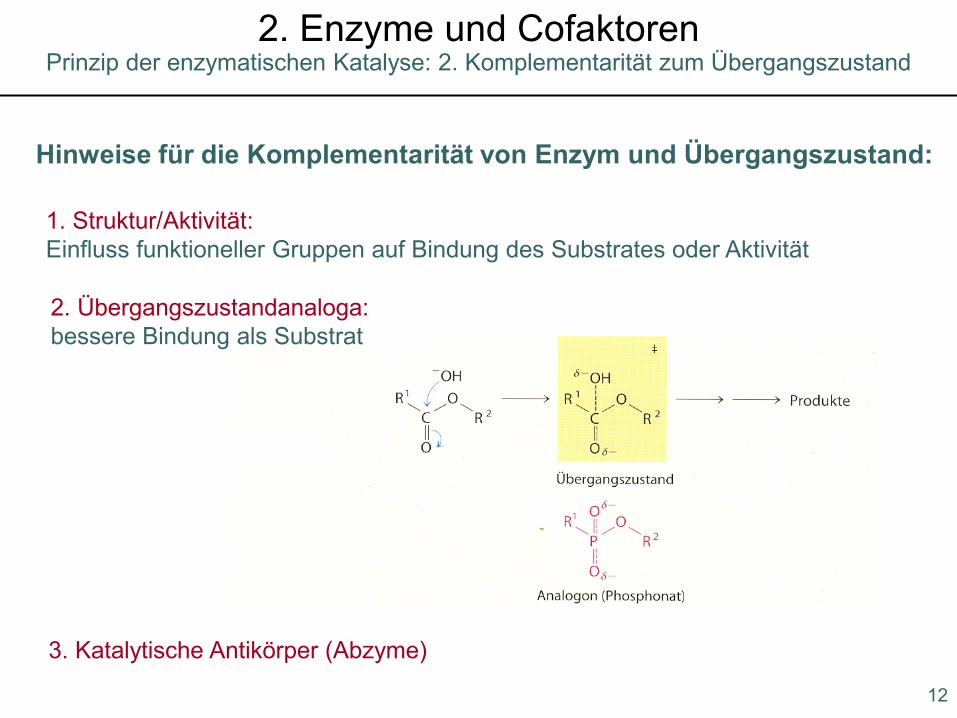
Hinweise für die Komplementarität von Enzym und Übergangszustand:
1. Struktur/Aktivität: Einfluss funktioneller Gruppen auf Bindung des Substrates oder Aktivität
2. Übergangszustandanaloga:bessere Bindung als Substrat
3. Katalytische Antikörper (Abzyme)
12
Prinzip der enzymatischen Katalyse: 2. Komplementarität zum Übergangszustand2. Enzyme und Cofaktoren
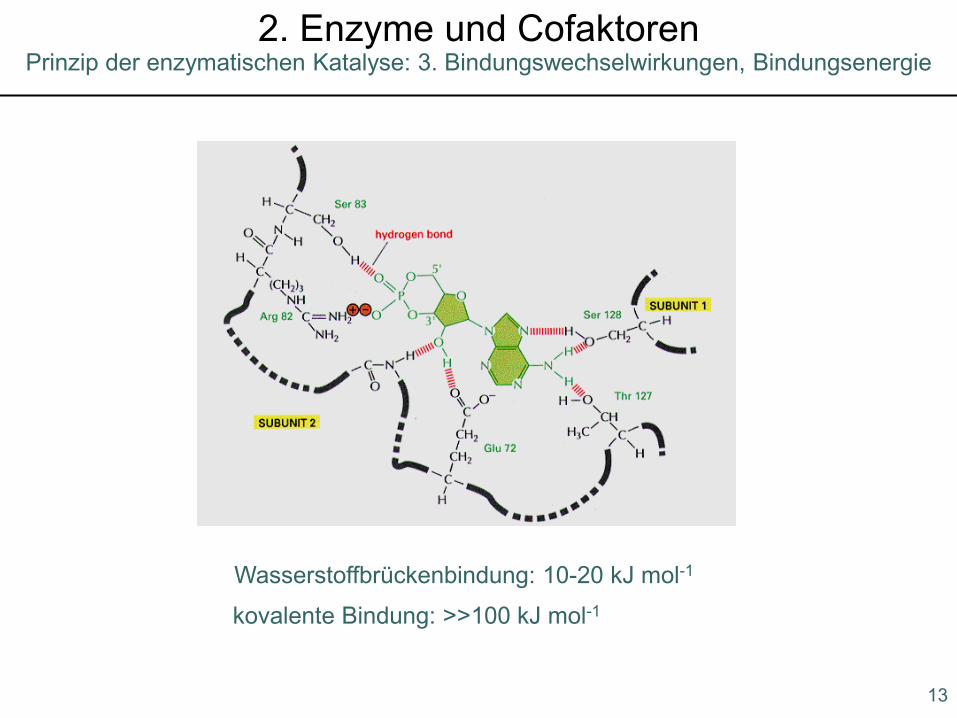
Wasserstoffbrückenbindung: 10-20 kJ mol-1
kovalente Bindung: >>100 kJ mol-1
13
Prinzip der enzymatischen Katalyse: 3. Bindungswechselwirkungen, Bindungsenergie2. Enzyme und Cofaktoren
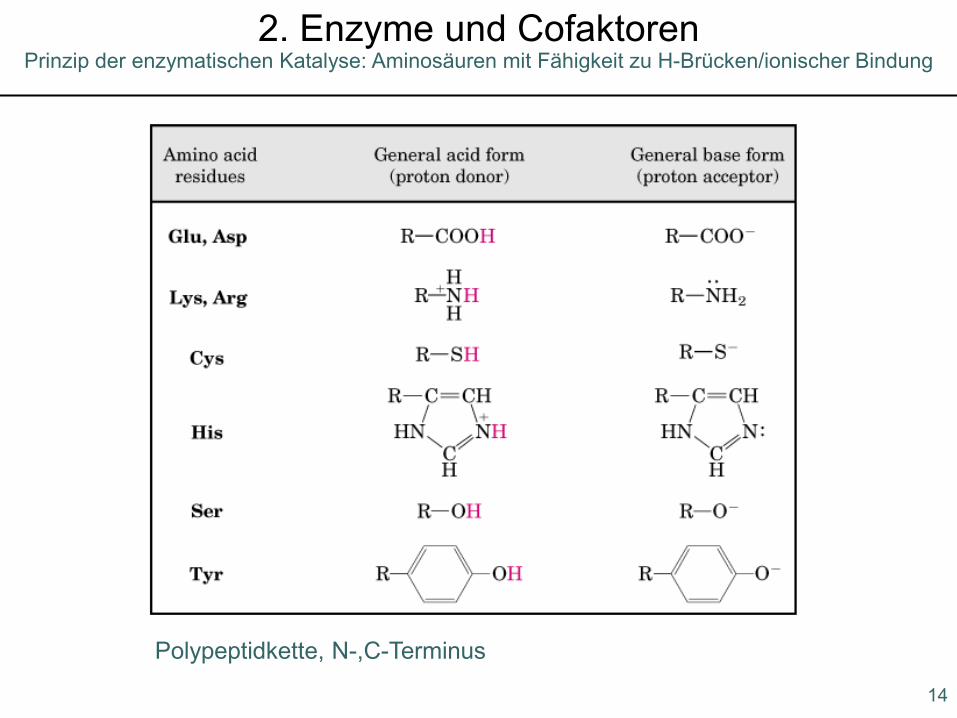
Polypeptidkette, N-,C-Terminus
14
Prinzip der enzymatischen Katalyse: Aminosäuren mit Fähigkeit zu H-Brücken/ionischer Bindung2. Enzyme und Cofaktoren
15
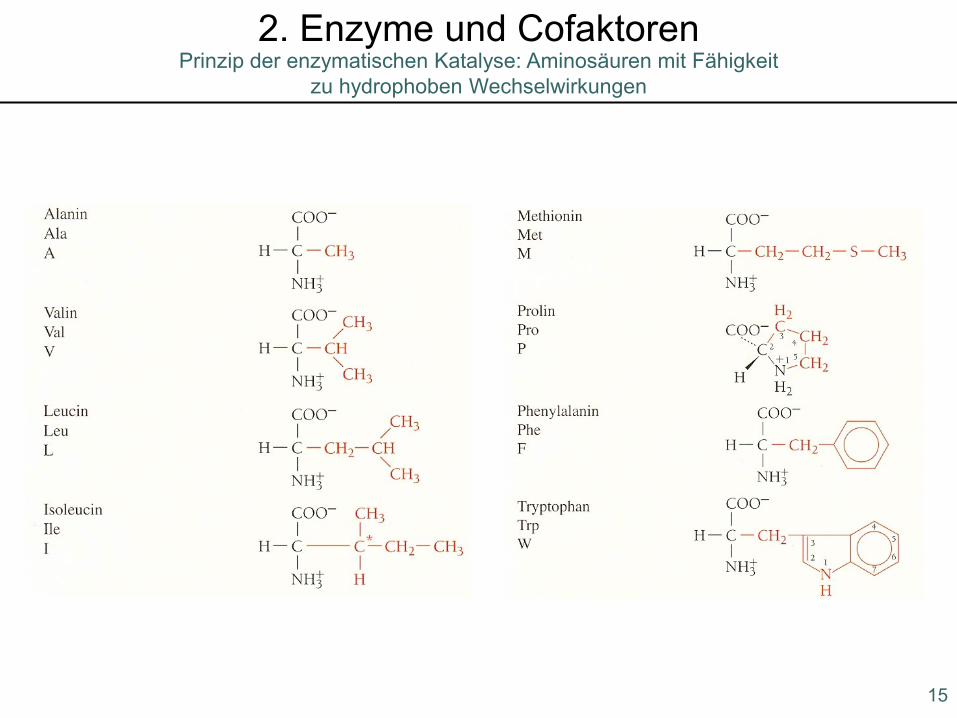
Prinzip der enzymatischen Katalyse: Aminosäuren mit Fähigkeit zu hydrophoben Wechselwirkungen
2. Enzyme und Cofaktoren
16
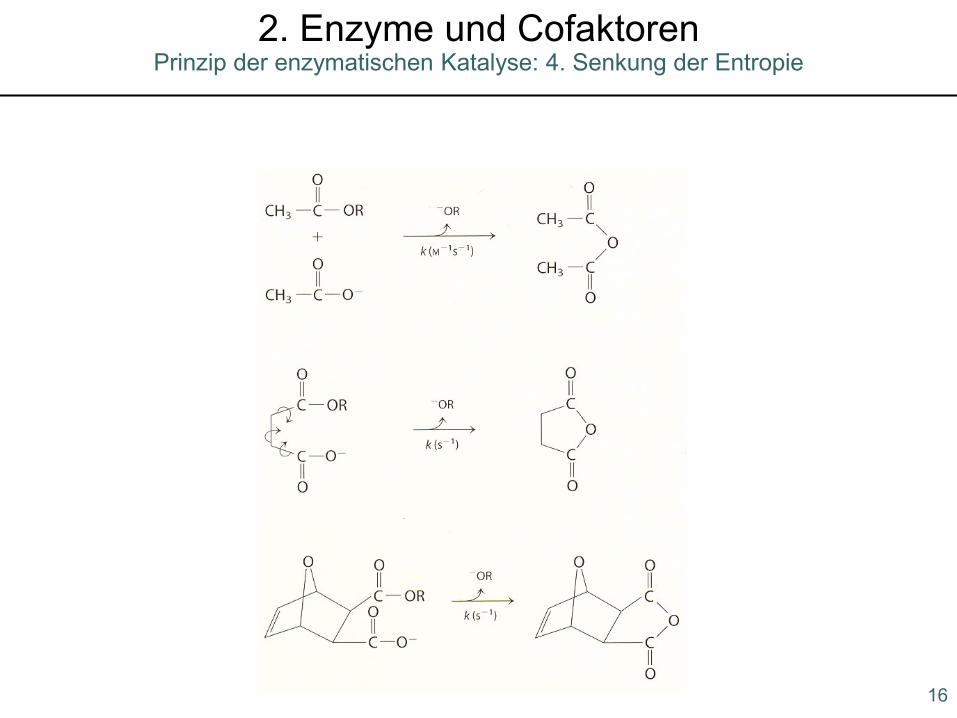
Prinzip der enzymatischen Katalyse: 4. Senkung der Entropie2. Enzyme und Cofaktoren
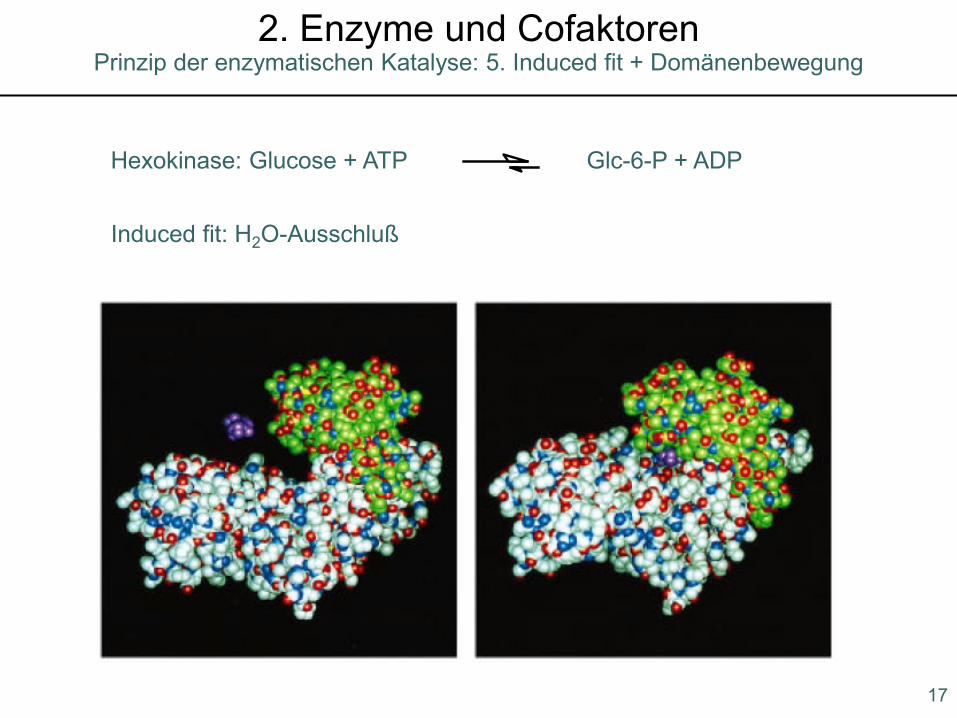
Induced fit: H2O-Ausschluß
Hexokinase: Glucose + ATP Glc-6-P + ADP
17
Prinzip der enzymatischen Katalyse: 5. Induced fit + Domänenbewegung2. Enzyme und Cofaktoren
18
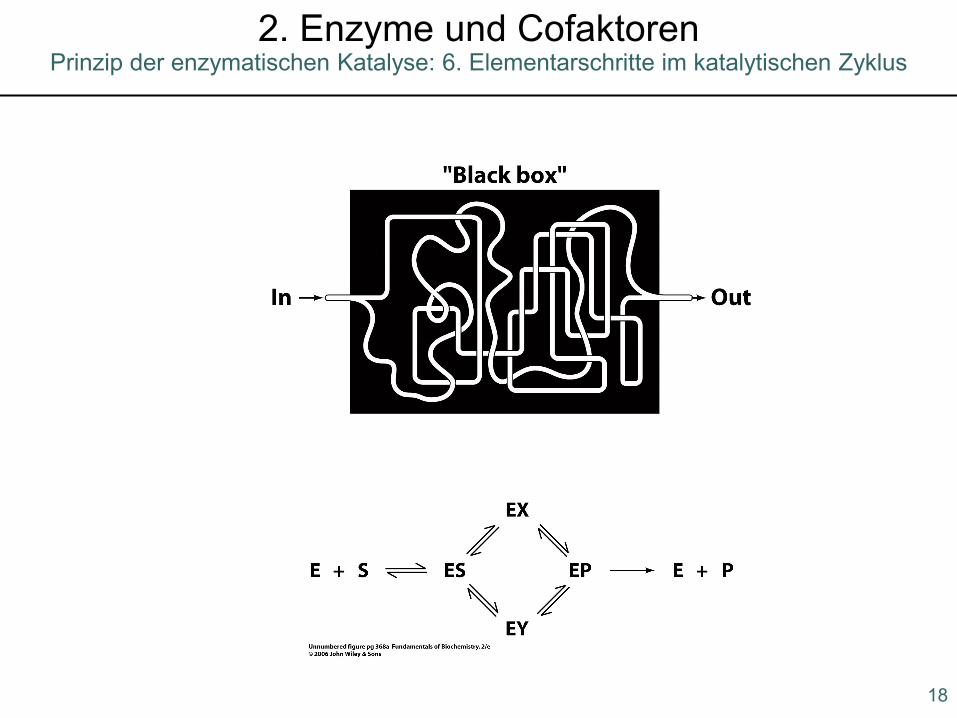
Prinzip der enzymatischen Katalyse: 6. Elementarschritte im katalytischen Zyklus2. Enzyme und Cofaktoren

Beschleunigungsrate einiger Enzyme: (‚Proficiency‘)
19
Enzyme = Biokatalysatoren2. Enzyme und Cofaktoren
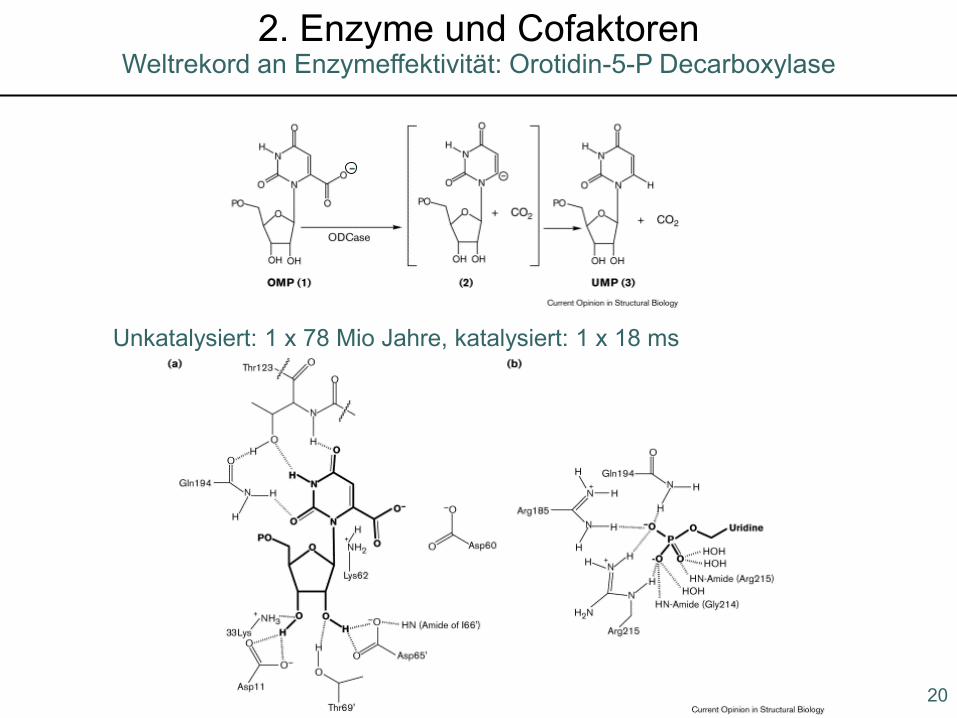
Unkatalysiert: 1 x 78 Mio Jahre, katalysiert: 1 x 18 ms
-
20
Weltrekord an Enzymeffektivität: Orotidin-5-P Decarboxylase2. Enzyme und Cofaktoren

1. Allgemeine Säure/Base-Katalyse: Bsp. Hydrolasen
2. Kovalente Katalyse: Bsp. Transaminasen, Decarboxylasen, Carboxylasen
3. Katalyse an Metallionen: Carboanhydrase, Katalase, Proteasen
21
Enzyme: einige Katalysemechanismen2. Enzyme und Cofaktoren
Aminosäurereste beispielsweise von Histidin reagieren als Säure oder Base, indem sie während einer Reaktion Protonen (H+-Ionen) aufnehmen oder abgeben.
Aminosäurereste oder Coenzyme gehen kovalente Bindungen mit einem Substrat ein und bilden ein kurzlebiges Zwischenprodukt. In der Regel sind bei solchen Reaktionen nukleophile Aminosäure-Seitenketten (beispielsweise Lysin-Seitenketten mit Aminogruppe) oder Coenzyme wie Pyridoxalphosphat beteiligt.
Metallionen können als strukturstabilisierende Koordinationszentren, Redox-Partner (oft Eisen- oder Kupfer-Ionen) oder als Lewis-Säuren (häufig Zink-Ionen) die Katalyse unterstützen. Sie können negative Ladungen stabilisieren bzw. abschirmen oder Wassermoleküle aktivieren.

Reaktionsgeschwindigkeit: schneller
Reaktionsbedingungen: milder, aber begrenzt auf wässriges Milieu
Spezifität: substrat- und stereoselektiv, kaum Nebenreaktionen
Regulation: vielfach
22
Enzyme vs. chemische Katalysatoren2. Enzyme und Cofaktoren
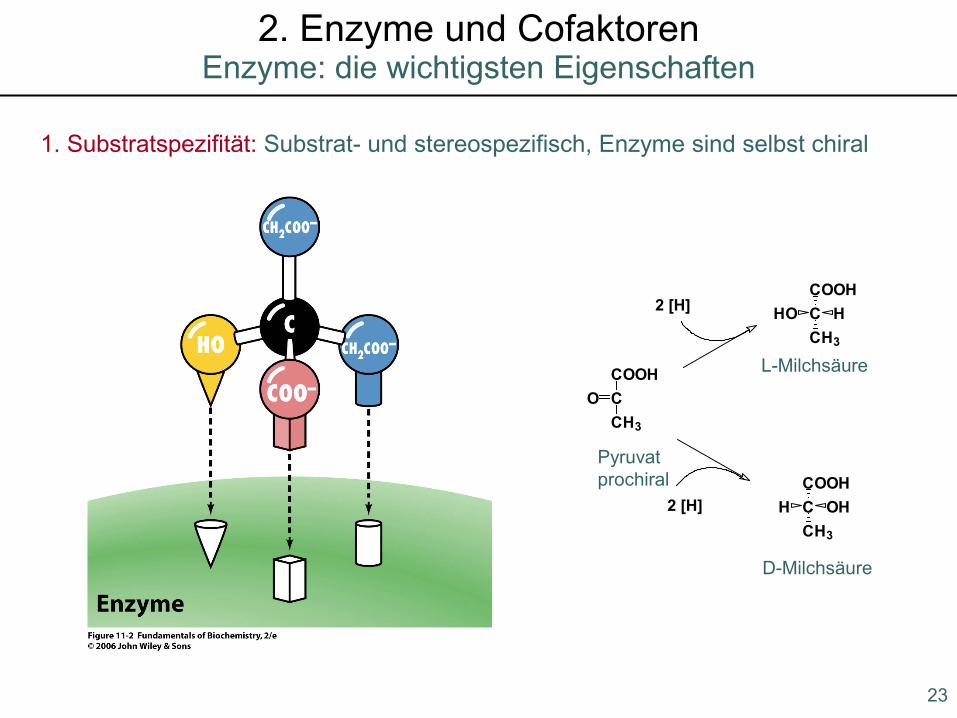
1. Substratspezifität: Substrat- und stereospezifisch, Enzyme sind selbst chiral
Pyruvatprochiral
L-Milchsäure
D-Milchsäure
COOHCCH3
O
COOHCCH3
HO H
COOHCCH3
H OH
2 [H]
2 [H]
23
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
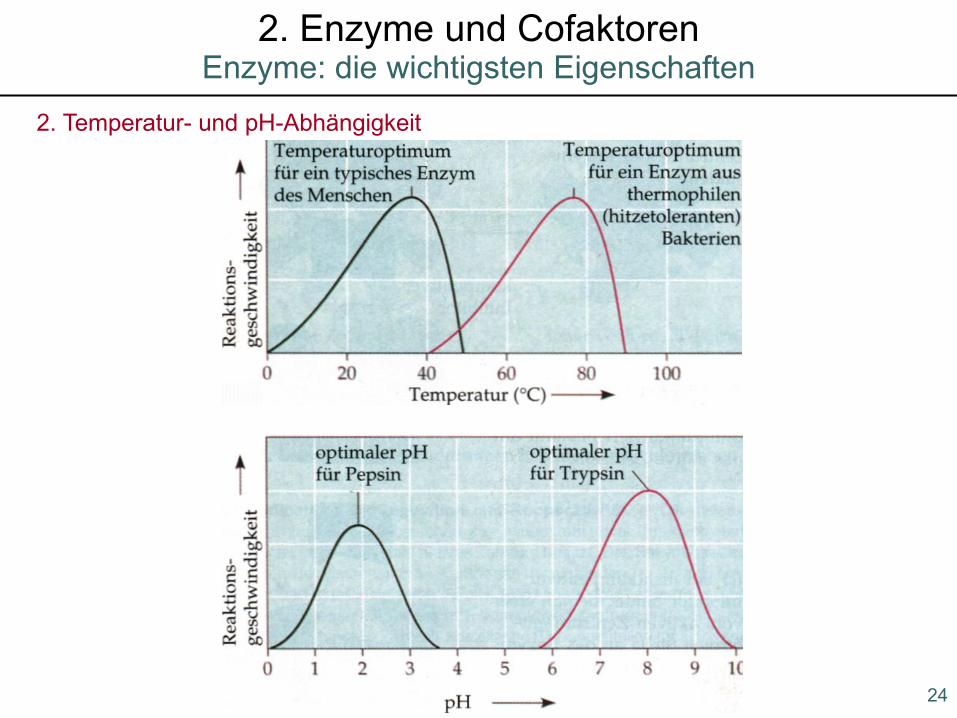
2. Temperatur- und pH-Abhängigkeit
24
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
3. Isoenzyme: Mehrere sehr ähnliche Enzyme → eine Reaktion
Bsp.: Lactat Dehydrogenasen
Funktion: Modulation der Aktivität in verschiedenen Geweben/Organellen
M-Typ H-Typ
- anaerobesGewebe
- aerobesGewebe
- Herzinfarkt-Diagnostik
25
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
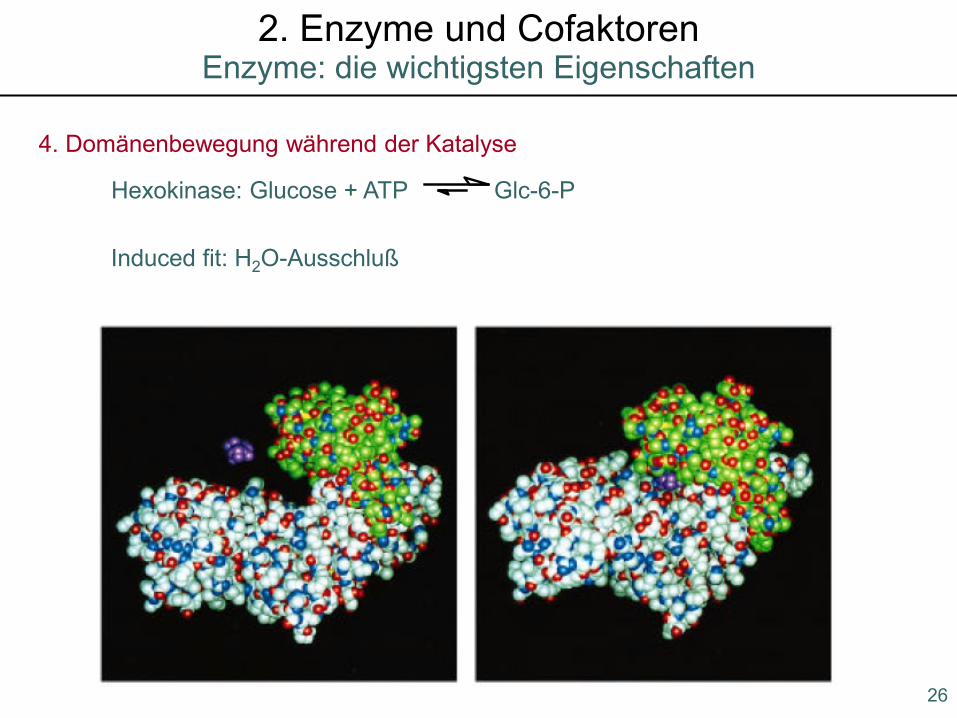
4. Domänenbewegung während der Katalyse
Induced fit: H2O-Ausschluß
Hexokinase: Glucose + ATP Glc-6-P
26
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
27
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
CTP, UTP
5. Allosterische Regulation
Bsp: Aspartat-Transcarbamoylase
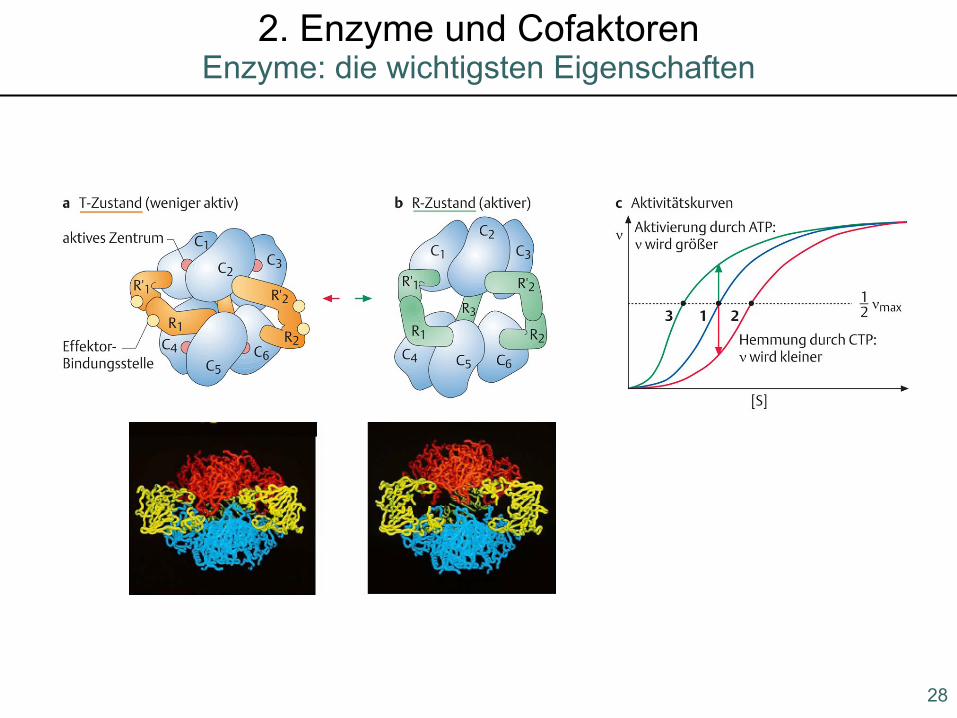
28
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren

29
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren
Hill-Koeffizient (nH):
Mittel, um die Kooperativität der Substratbindung eines Enzyms zu messen
positve Kooperativität:
nH liegt zwischen 1 und der Anzahl der substratspezifischen Bindungsstellen(je größer nH, desto stärker die Kooperativität)
negative Kooperativität:
nH < 1
keine Kooperativität:
nH = 1
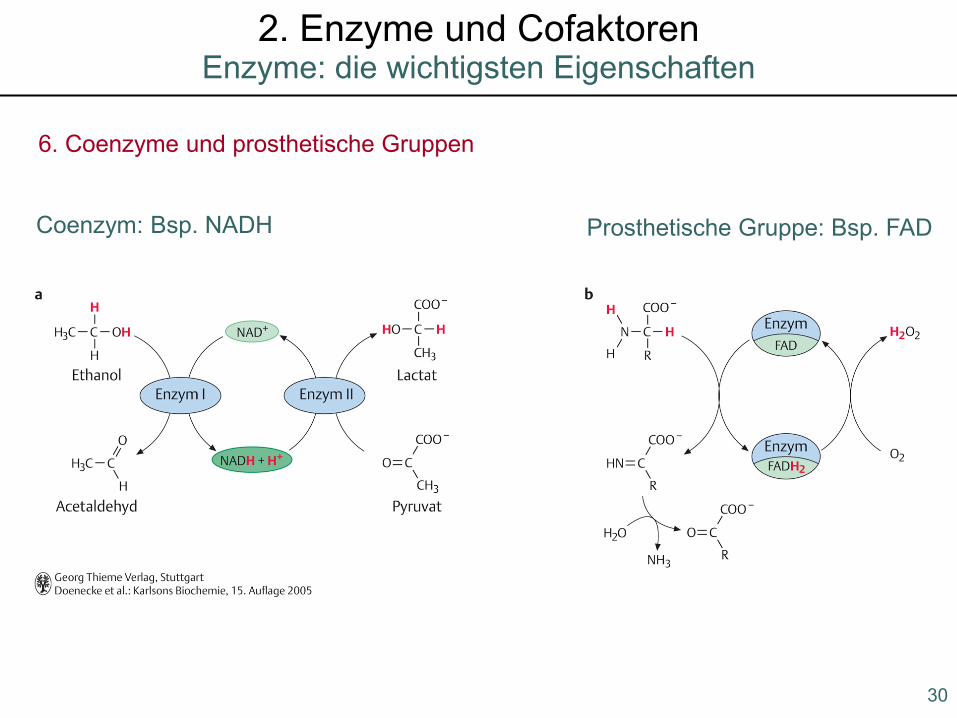
6. Coenzyme und prosthetische Gruppen
Coenzym: Bsp. NADH Prosthetische Gruppe: Bsp. FAD
30
Enzyme: die wichtigsten Eigenschaften2. Enzyme und Cofaktoren

-katalysieren eine chemische Reaktion durch Erniedrigung der Aktivierungsenergie
-stabilisieren den Übergangszustand durch effektive Bindung
-sind substratspezifisch
-sind selbst chiral und stereospezifisch
-haben Temperatur- und pH-Optimum
-zeigen Domänenbewegung während der Katalyse
-besitzen häufig Coenzyme oder prosthetische Gruppen
-sind regulierbar (Hemmung/Aktivierung)
31
Enzyme: Zusammenfassung2. Enzyme und Cofaktoren
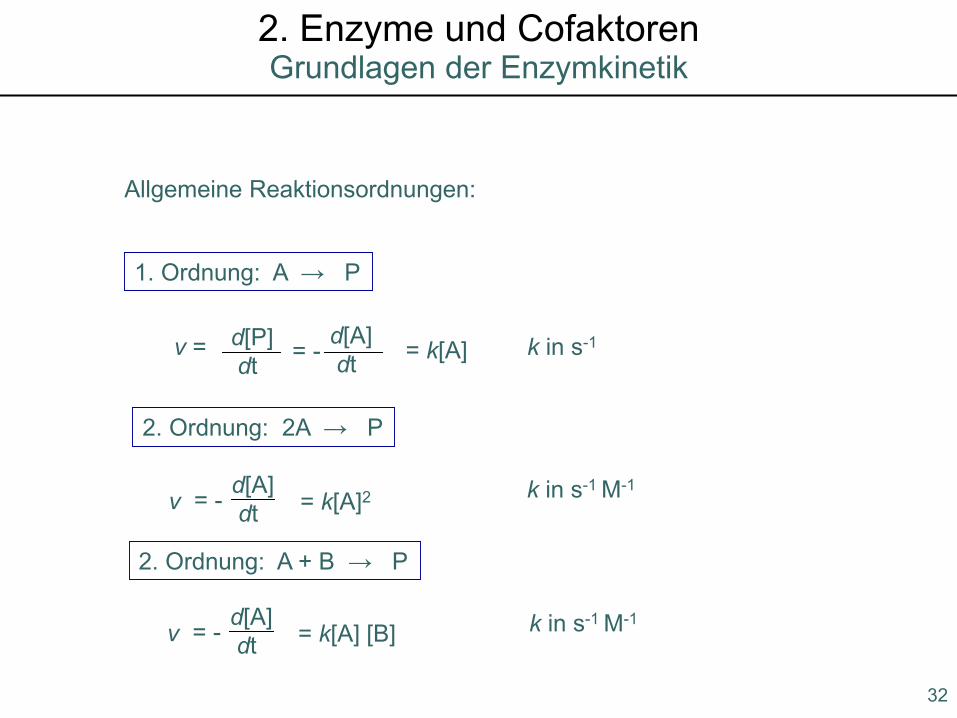
Allgemeine Reaktionsordnungen:
1. Ordnung: A → P
v = d[P]dt
d[A]dt= - = k[A] k in s-1
2. Ordnung: 2A → P
d[A]dt= -v = k[A]2 k in s-1 M-1
2. Ordnung: A + B → P
d[A]dt= -v = k[A] [B] k in s-1 M-1
32
Grundlagen der Enzymkinetik2. Enzyme und Cofaktoren
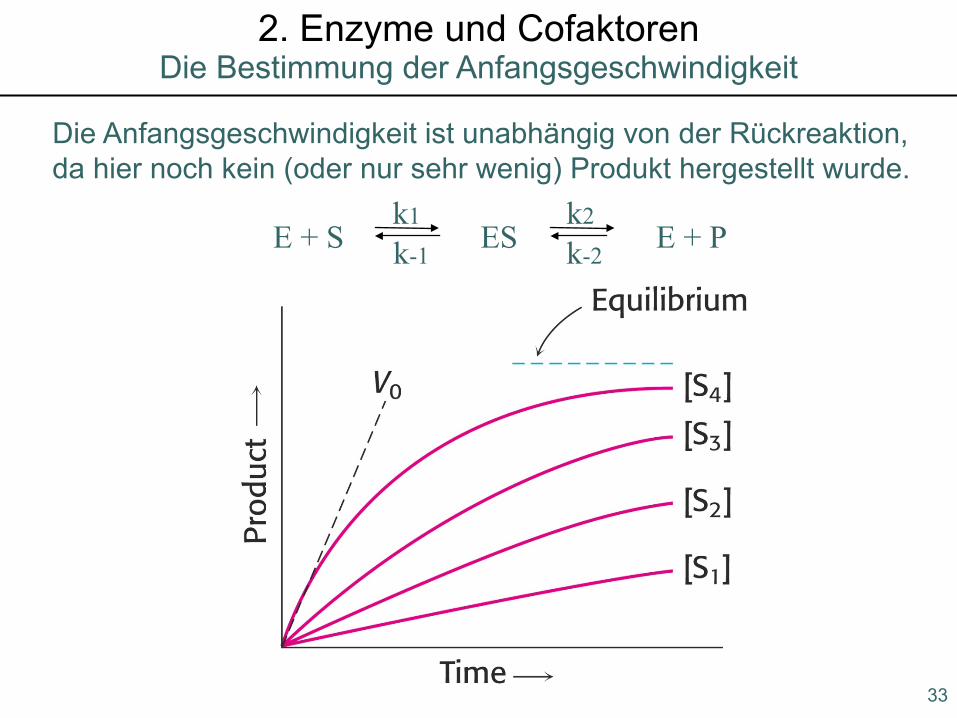
E + S ES E + Pk1 k2
k-1 k-2
Die Anfangsgeschwindigkeit ist unabhängig von der Rückreaktion,da hier noch kein (oder nur sehr wenig) Produkt hergestellt wurde.
33
Die Bestimmung der Anfangsgeschwindigkeit2. Enzyme und Cofaktoren

34
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
1903 Victor Henri: bei einer enzymatischen Katalysation vorübergehend Enzym-Substrat-Komplex
1913 Maud Menten und Leonor Michaelis: Michaelis-Menten-Gleichung
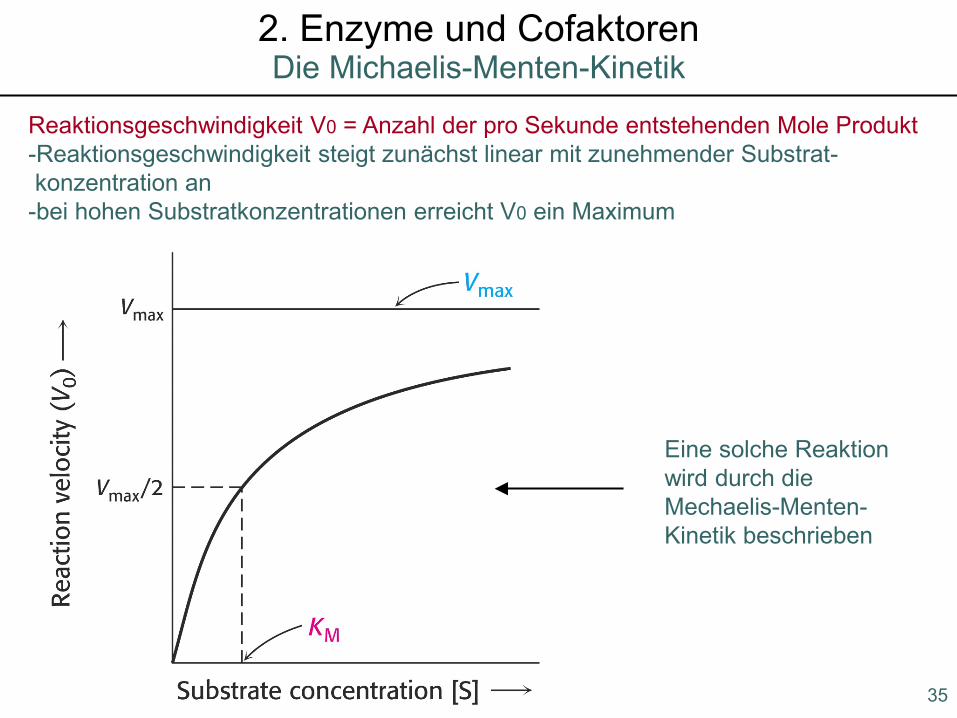
Reaktionsgeschwindigkeit V0 = Anzahl der pro Sekunde entstehenden Mole Produkt-Reaktionsgeschwindigkeit steigt zunächst linear mit zunehmender Substrat-konzentration an
-bei hohen Substratkonzentrationen erreicht V0 ein Maximum
Eine solche Reaktionwird durch die Mechaelis-Menten-Kinetik beschrieben
35
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
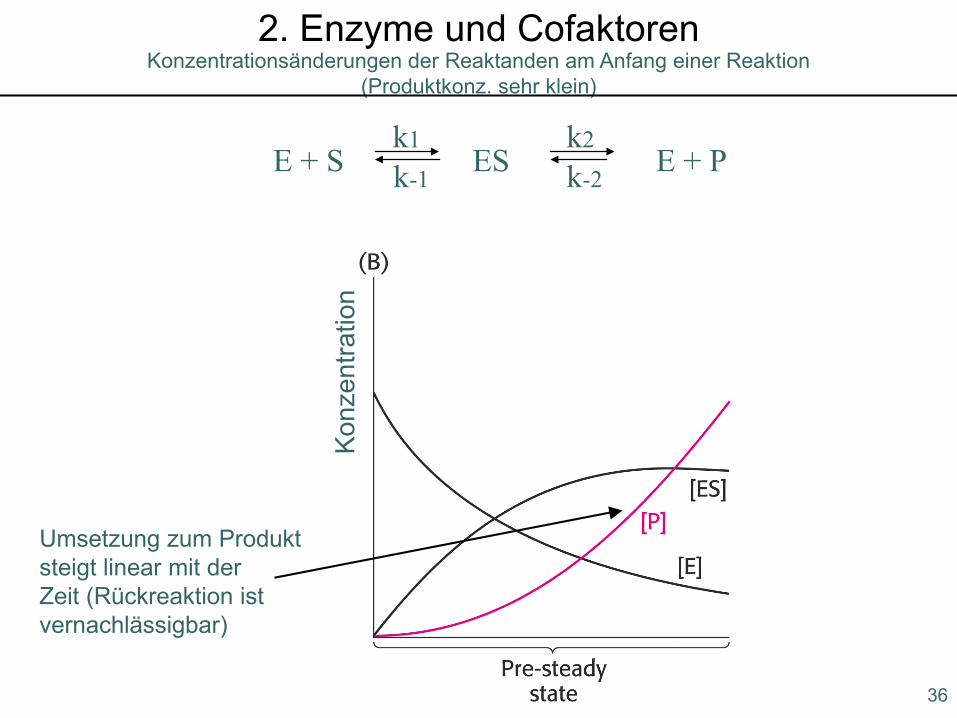
Umsetzung zum Produktsteigt linear mit der Zeit (Rückreaktion istvernachlässigbar)
Konz
entra
tion
E + S ES E + Pk1 k2
k-1 k-2
36
Konzentrationsänderungen der Reaktanden am Anfang einer Reaktion (Produktkonz. sehr klein)
2. Enzyme und Cofaktoren
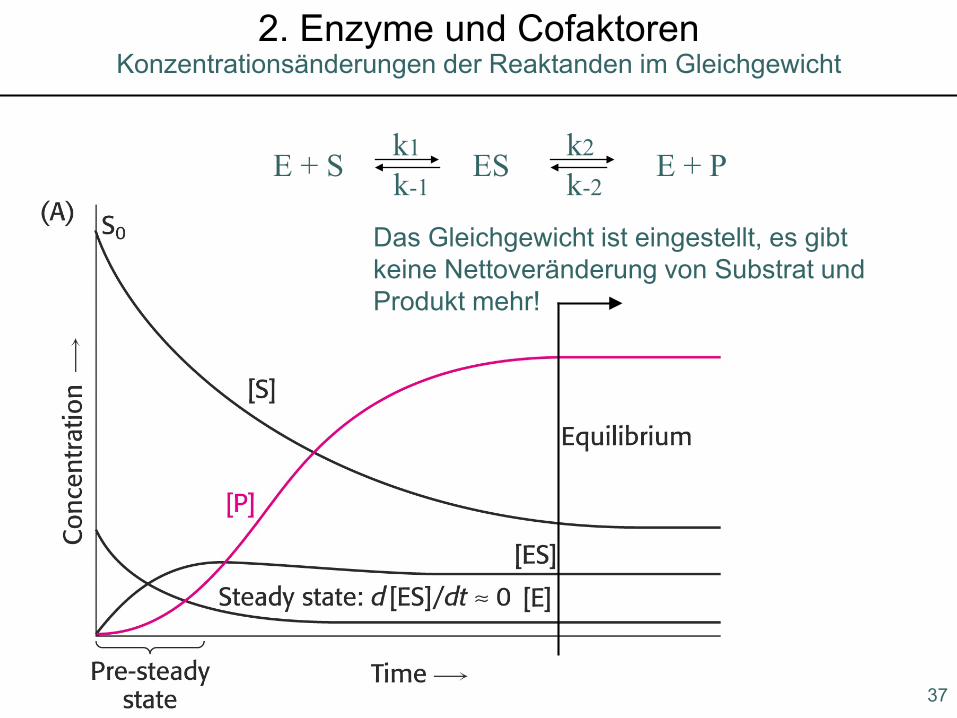
E + S ES E + Pk1 k2
k-1 k-2
Das Gleichgewicht ist eingestellt, es gibtkeine Nettoveränderung von Substrat undProdukt mehr!
37
Konzentrationsänderungen der Reaktanden im Gleichgewicht2. Enzyme und Cofaktoren

Die MM-Kinetik formuliert einen Ausdruck, der die Katalyse-geschwindigkeit mit der Substrat- und Enzymkonzentration verbindet.
Das MM-Modell ist das einfachste, mit dem man die kinetischen Eigenschaften vieler enzymkatalysierter Reaktionen beschreiben kann.
Zentraler Punkt bei dieser Betrachtungsweise ist die Michealis-Menten Gleichung
38
Was macht die Michaelis-Menten-Kinetik?2. Enzyme und Cofaktoren
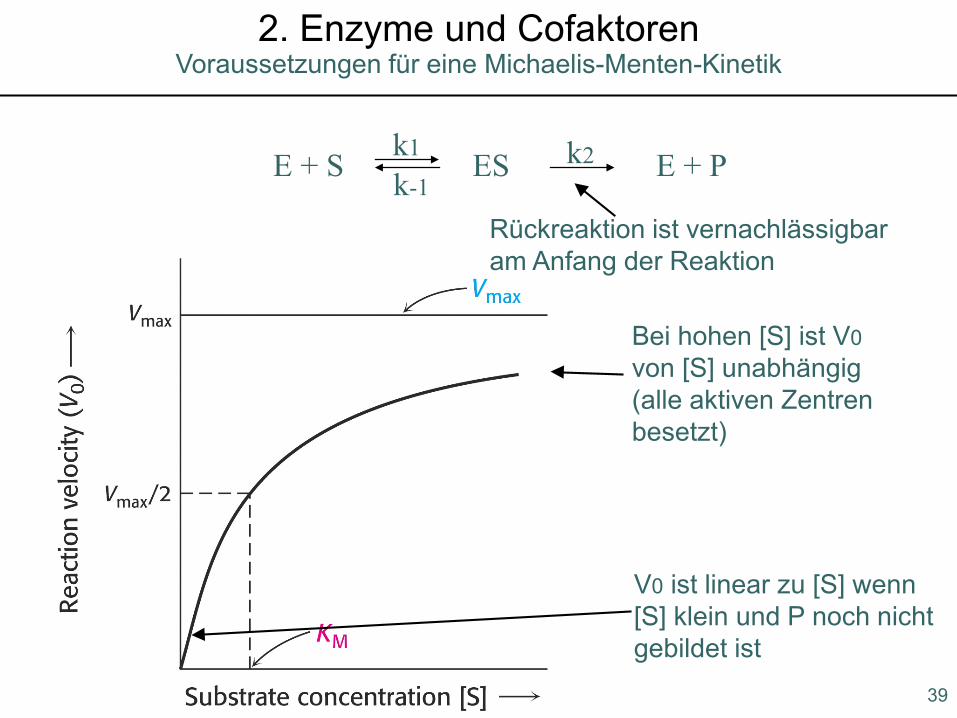
E + S ES E + Pk1 k2k-1
V0 ist linear zu [S] wenn [S] klein und P noch nichtgebildet ist
Bei hohen [S] ist V0von [S] unabhängig(alle aktiven Zentrenbesetzt)
Rückreaktion ist vernachlässigbaram Anfang der Reaktion
39
Voraussetzungen für eine Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
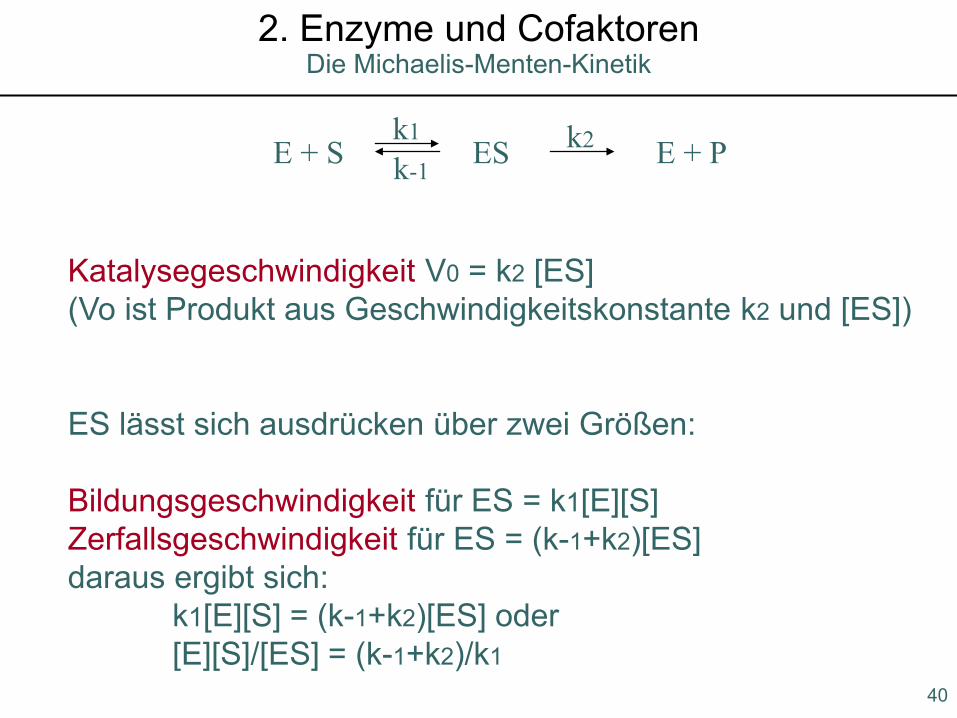
Katalysegeschwindigkeit V0 = k2 [ES](Vo ist Produkt aus Geschwindigkeitskonstante k2 und [ES])
ES lässt sich ausdrücken über zwei Größen:
Bildungsgeschwindigkeit für ES = k1[E][S]Zerfallsgeschwindigkeit für ES = (k-1+k2)[ES]daraus ergibt sich:
k1[E][S] = (k-1+k2)[ES] oder[E][S]/[ES] = (k-1+k2)/k1
40
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
E + S ES E + Pk1 k2k-1

E + S ES E + Pk1 k2k-1
[E][S]/[ES] = (k-1+k2)/k1 = KM
KM ist die Michaelis-Menten Konstante (hat Konzentrations-einheit).Diese Konstante ist ein wichtiges Charakteristikum für E-S-Wechselwirkungen und von der Konzentration dieser beidenunabhängig.
Umformen der obigen Gleichung ergibt:[E][S]/KM = [ES]
41
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
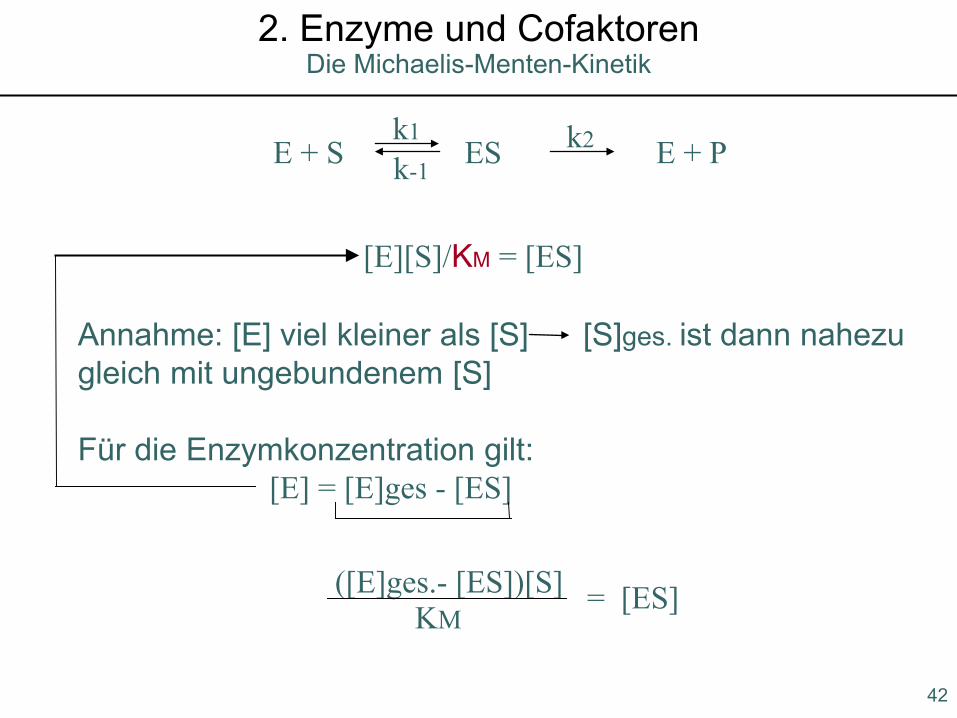
E + S ES E + Pk1 k2k-1
[E][S]/KM = [ES]
Annahme: [E] viel kleiner als [S] [S]ges. ist dann nahezu gleich mit ungebundenem [S]
Für die Enzymkonzentration gilt:[E] = [E]ges - [ES]
([E]ges.- [ES])[S] = [ES]KM
42
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
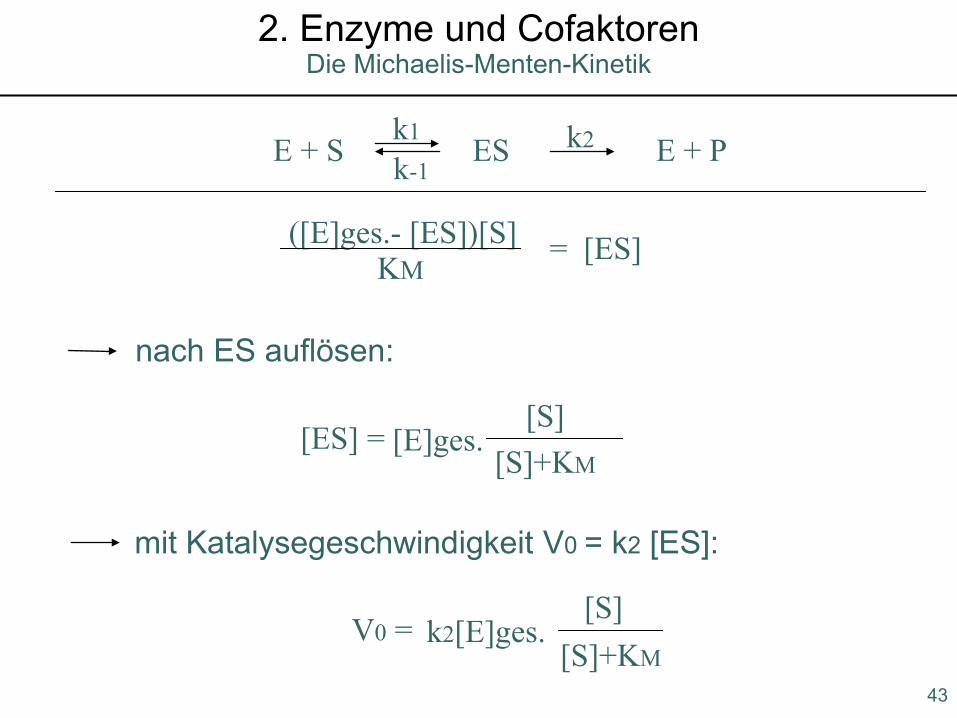
E + S ES E + Pk1 k2k-1
([E]ges.- [ES])[S] = [ES]KM
nach ES auflösen:
[ES] = [E]ges.[S]
[S]+KM
mit Katalysegeschwindigkeit V0 = k2 [ES]:
V0 = k2[E]ges.[S]
[S]+KM43
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
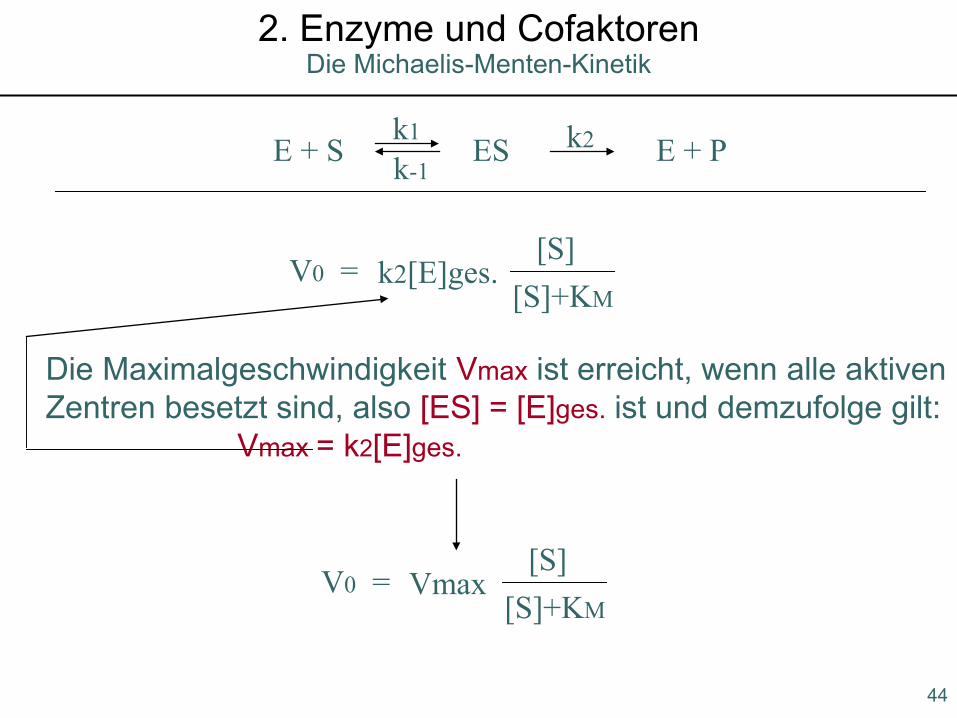
E + S ES E + Pk1 k2k-1
V0 = k2[E]ges.[S]
[S]+KM
Die Maximalgeschwindigkeit Vmax ist erreicht, wenn alle aktivenZentren besetzt sind, also [ES] = [E]ges. ist und demzufolge gilt:
Vmax = k2[E]ges.
V0 = Vmax[S]
[S]+KM
44
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren

V0 =Vmax [S]KM + [S]
Die Michaelis-Menten Gleichung
45
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
-wenn [S] sehr klein ist wird ist V0 direkt Proportional zu [S]-wenn [S] sehr groß ist (viel größer als KM) ist V0=Vmax-wenn [S]=KM wird V0 = Vmax/2d.h. KM ist die Substratkonzentration,bei der die Reaktionsgeschwindigkeithalbmaximal ist!
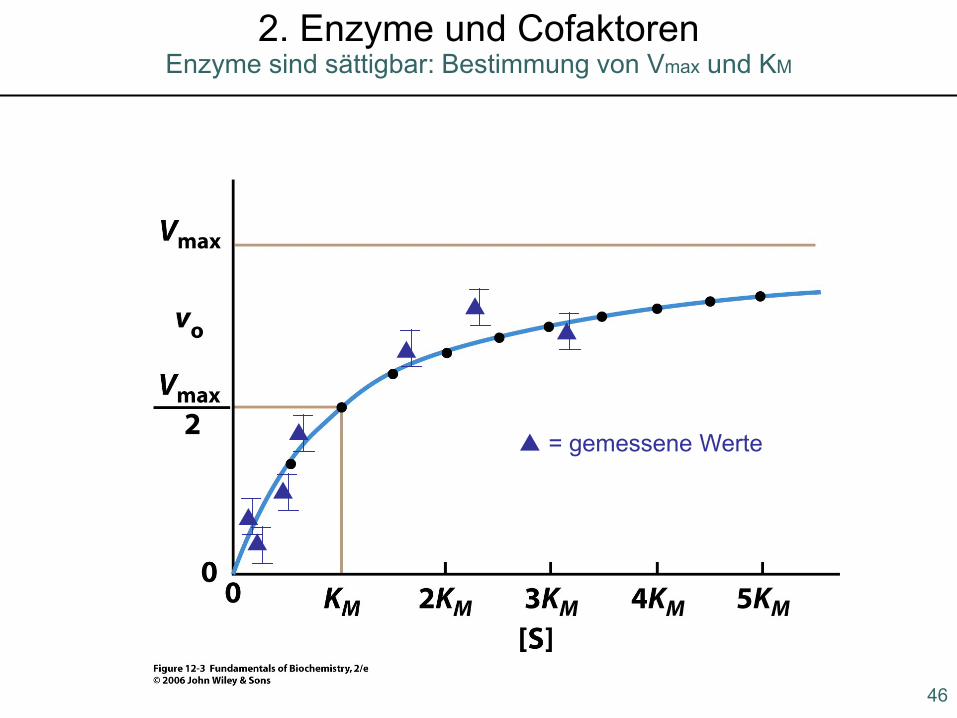
▲
▲
▲
▲
▲▲
▲
▲ = gemessene Werte
46
Enzyme sind sättigbar: Bestimmung von Vmax und KM
2. Enzyme und Cofaktoren
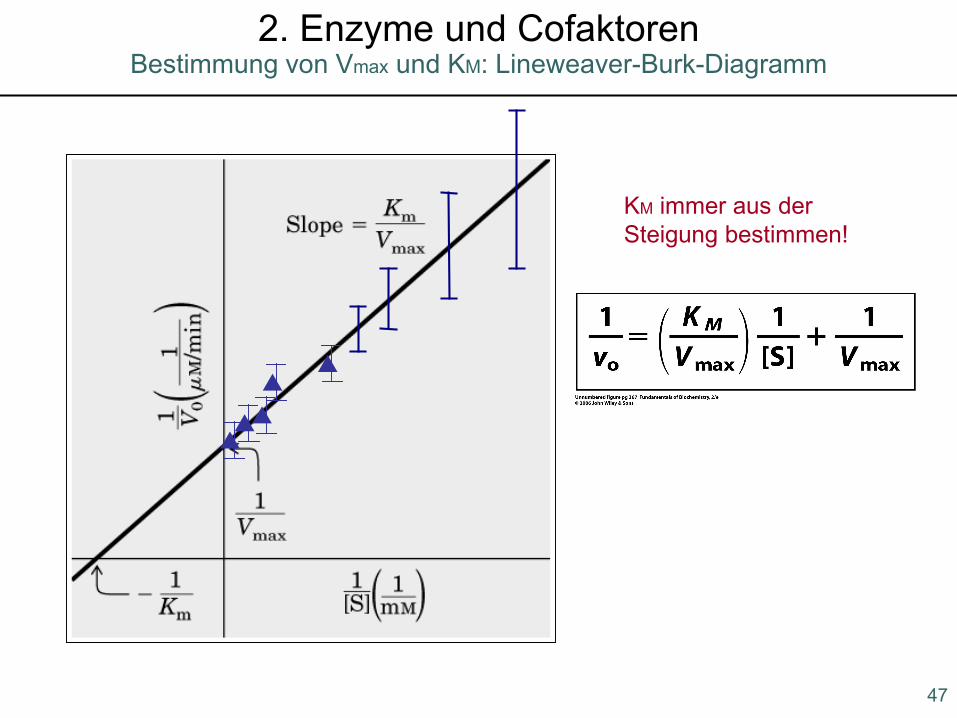
▲▲▲
▲▲
47
Bestimmung von Vmax und KM: Lineweaver-Burk-Diagramm2. Enzyme und Cofaktoren
KM immer aus der Steigung bestimmen!

Ein Beispiel, wie sich unterschiedliche KM-Werte in biologischenSystemen auswirken können:Viele Asiaten vertragen keinen Alkohol. Dieser Effekt wird durch Acetaldehyd hervorgerufen, das durch die AD gebildet wird.
EtOH + NAD+ Acetaldehyd + NADH
Acetaldehyd wird durch eine weitere DH zu Acetat abgebaut. Hiervon hat der Mensch 2 Isozyme: Mitochondriale DH mit niedrigem KM, cytosolische DH mit hohem KM. Bei alkohol-empfindlichen Menschen ist die mitochondriale DH mutiert und daher inaktiv.Aufgrund des hohen KM der cytosolischen DH wird Acetaldehyd nur sehr ineffizient abgebaut und daher ins Blut abgegeben. Dies führt zu den physiologischen Effekten.
AD
48
Die Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
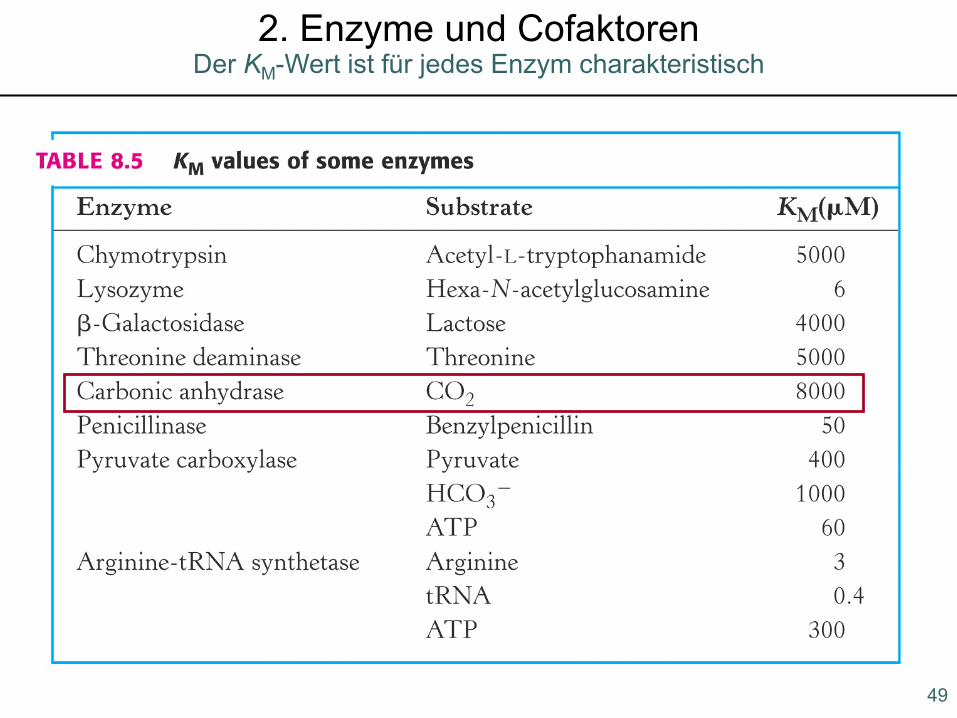
49
Der KM-Wert ist für jedes Enzym charakteristisch2. Enzyme und Cofaktoren

Die Wechselzahl kcat ist die Anzahlvon Substratmolekülen, die beivollständiger Sättigung des Enzymsmit Substrat pro Zeiteinheit umge-setzt werden.
Für die meisten Reaktionen liegt kcatzwischen 1 und 10000/sec.
50
Enzymkinetik: wichtige Definitionen2. Enzyme und Cofaktoren
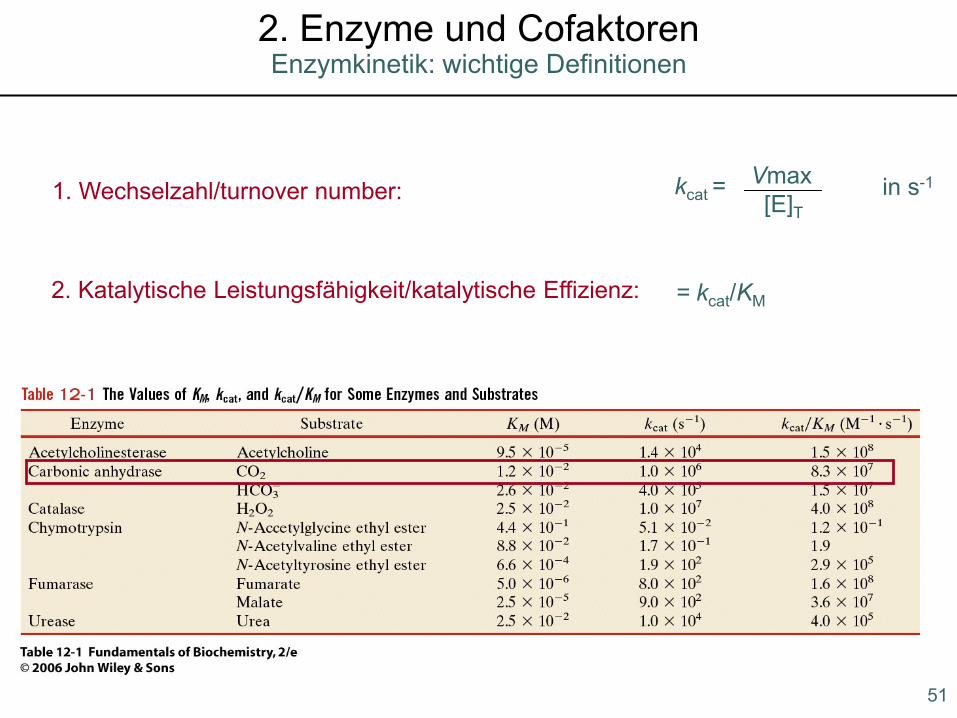
1. Wechselzahl/turnover number: kcat =Vmax[E]T
in s-1
2. Katalytische Leistungsfähigkeit/katalytische Effizienz: = kcat/KM
51
Enzymkinetik: wichtige Definitionen2. Enzyme und Cofaktoren

1. Wechselzahl/catalytic number: kcat =Vmax[E]T
in s-1
2. Katalytische Leistungsfähigkeit: = kcat/KM
3. Einheiten der Enzymaktivität:
- klassisch: 1 Enzymeinheit: 1 U = 1 µmol Substrat min-1
- seit 1972 SI-Einheit : 1 katal = 1 mol Substrat s-1
1 unit = 16,67 nkat
typische spezifische Aktivität: 5-100 Units (mg Enzym)-1
52
Enzymkinetik: wichtige Definitionen2. Enzyme und Cofaktoren
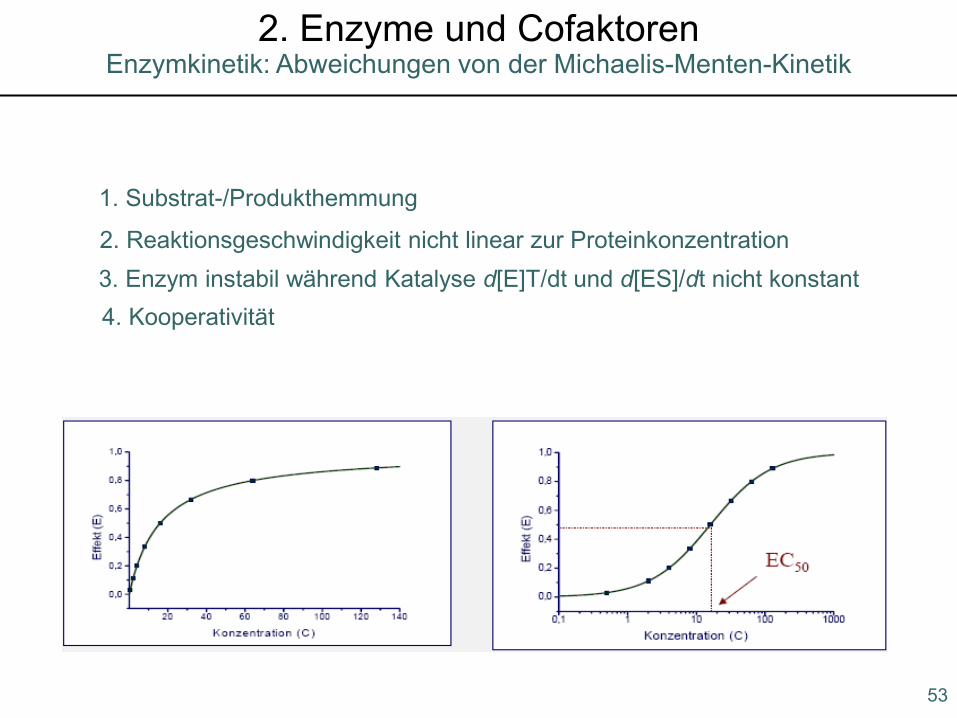
1. Substrat-/Produkthemmung
3. Enzym instabil während Katalyse d[E]T/dt und d[ES]/dt nicht konstant2. Reaktionsgeschwindigkeit nicht linear zur Proteinkonzentration
4. Kooperativität
53
Enzymkinetik: Abweichungen von der Michaelis-Menten-Kinetik2. Enzyme und Cofaktoren
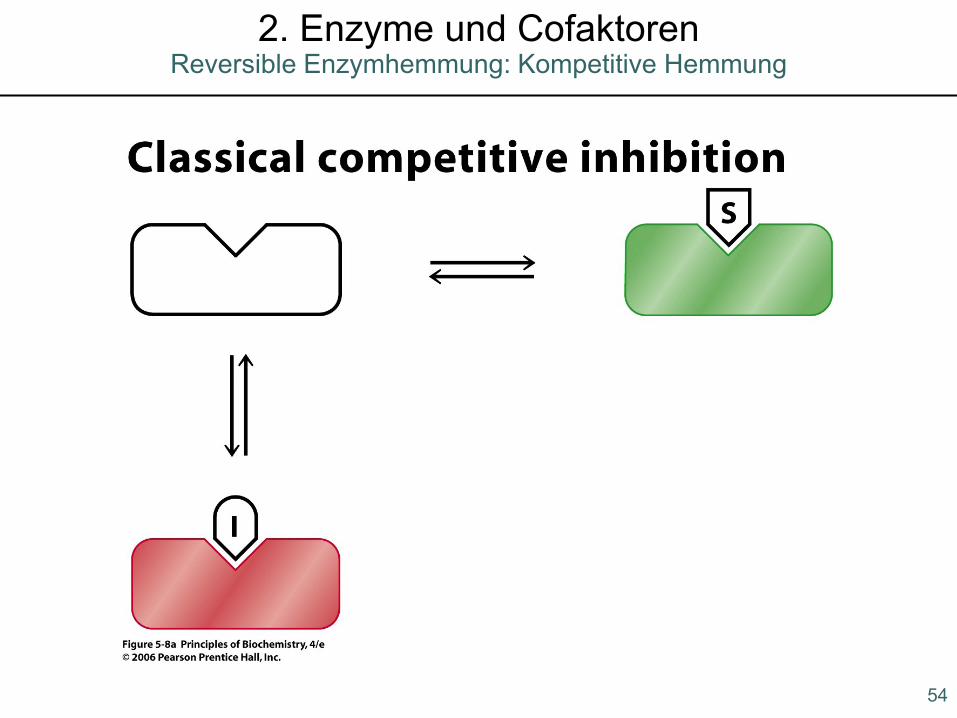
54
Reversible Enzymhemmung: Kompetitive Hemmung2. Enzyme und Cofaktoren
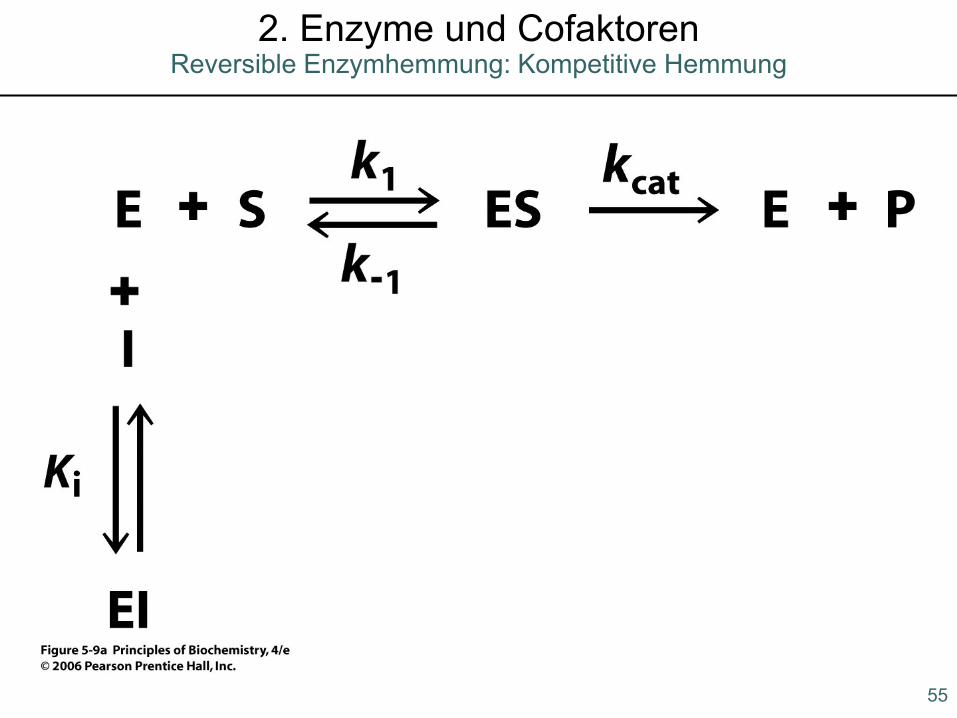
55
2. Enzyme und CofaktorenReversible Enzymhemmung: Kompetitive Hemmung
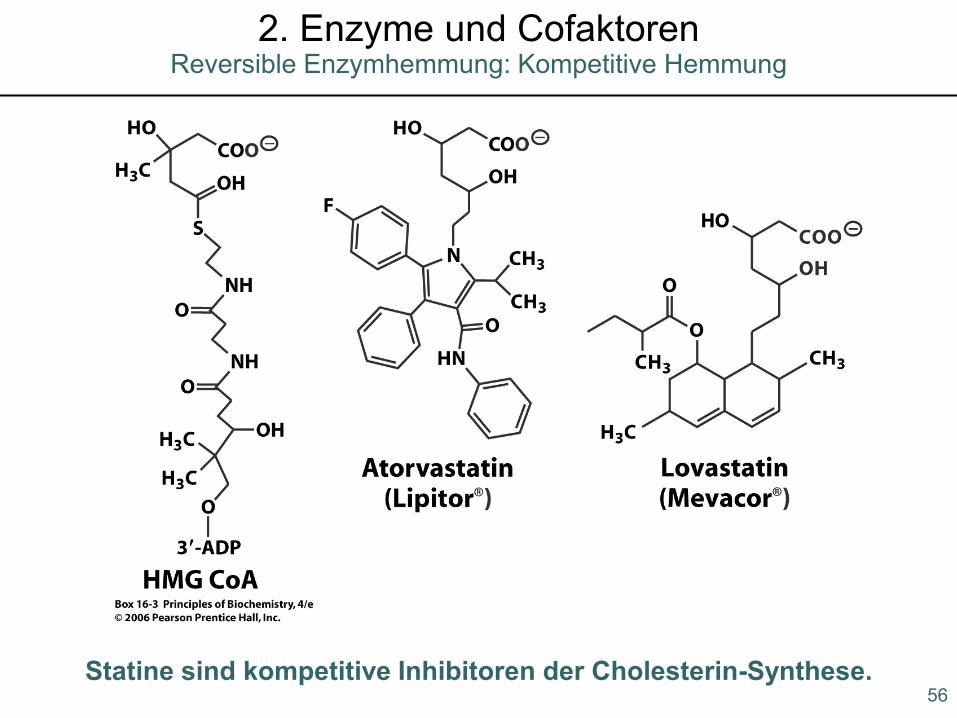
Statine sind kompetitive Inhibitoren der Cholesterin-Synthese.56
2. Enzyme und CofaktorenReversible Enzymhemmung: Kompetitive Hemmung
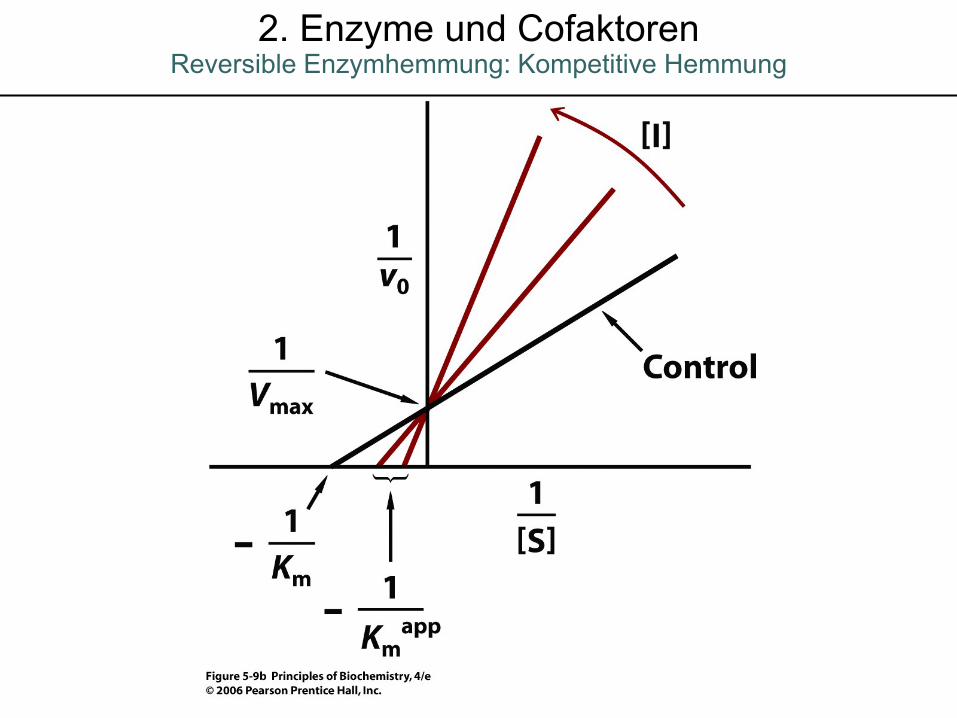
2. Enzyme und CofaktorenReversible Enzymhemmung: Kompetitive Hemmung
58
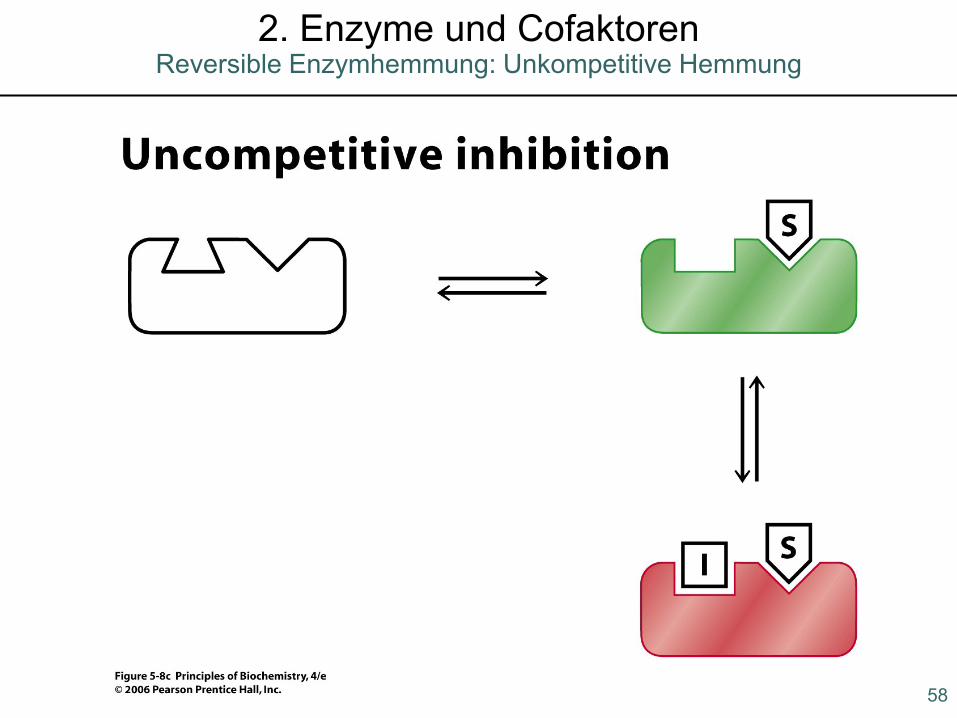
Reversible Enzymhemmung: Unkompetitive Hemmung2. Enzyme und Cofaktoren
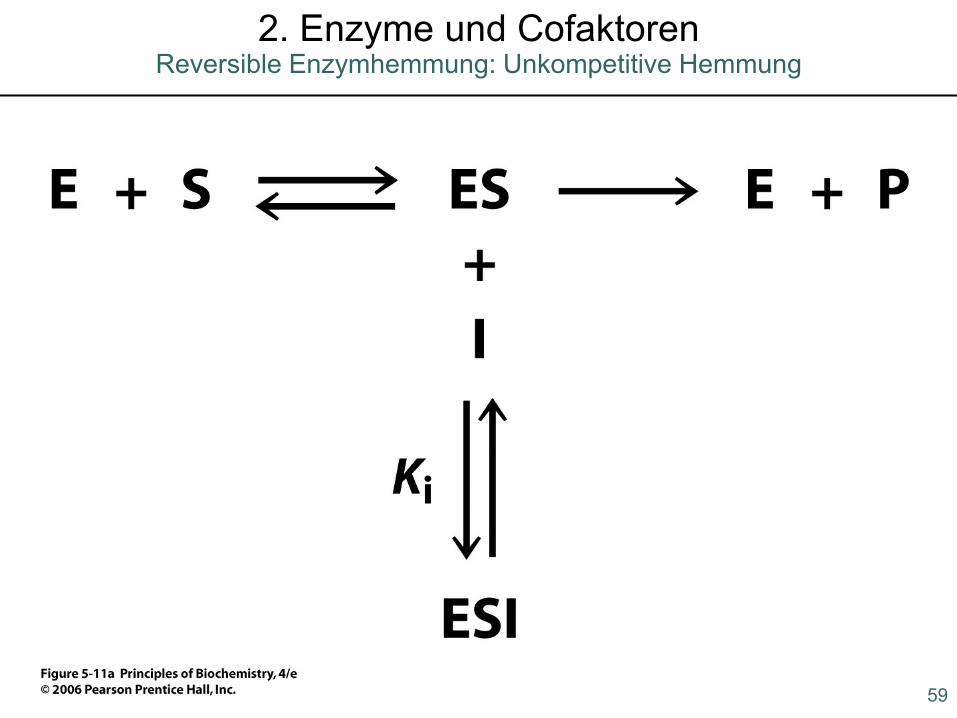
59
2. Enzyme und CofaktorenReversible Enzymhemmung: Unkompetitive Hemmung
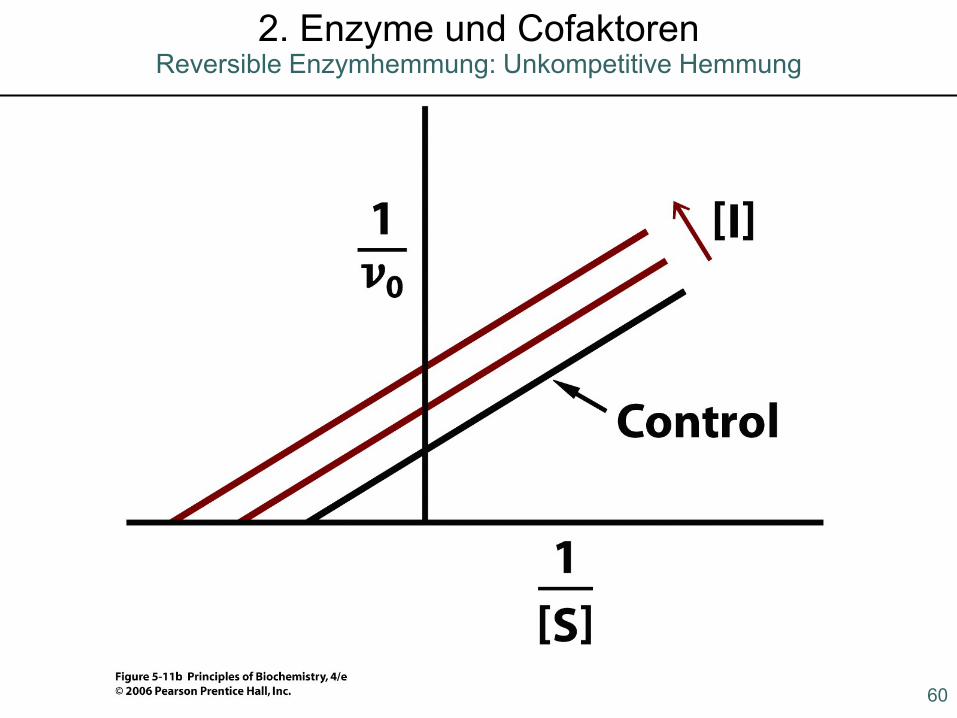
60
2. Enzyme und CofaktorenReversible Enzymhemmung: Unkompetitive Hemmung
61
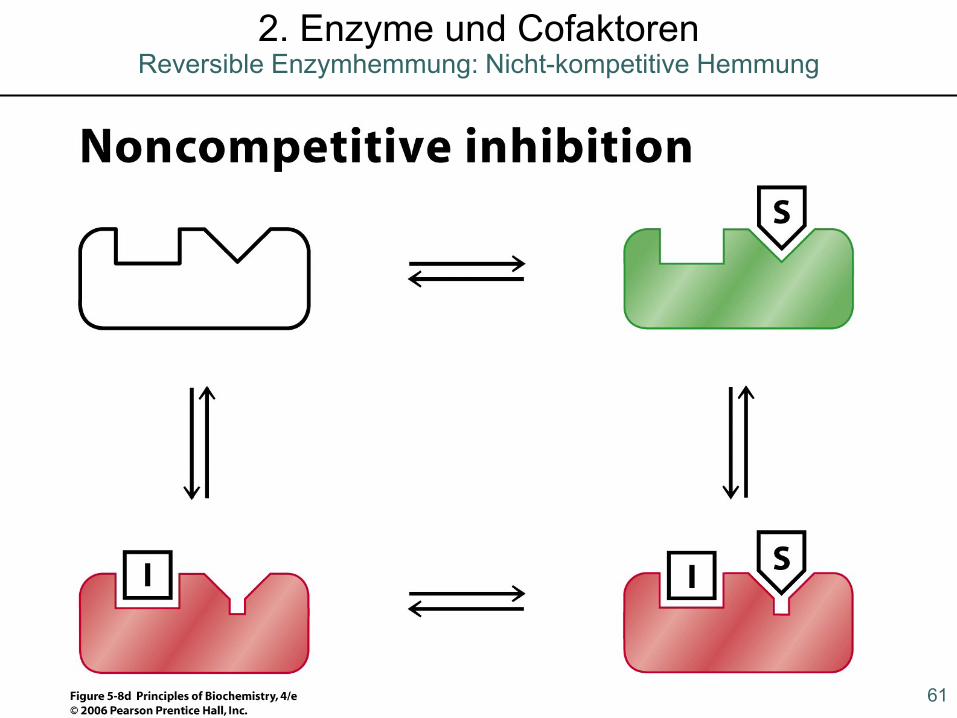
Reversible Enzymhemmung: Nicht-kompetitive Hemmung2. Enzyme und Cofaktoren
62
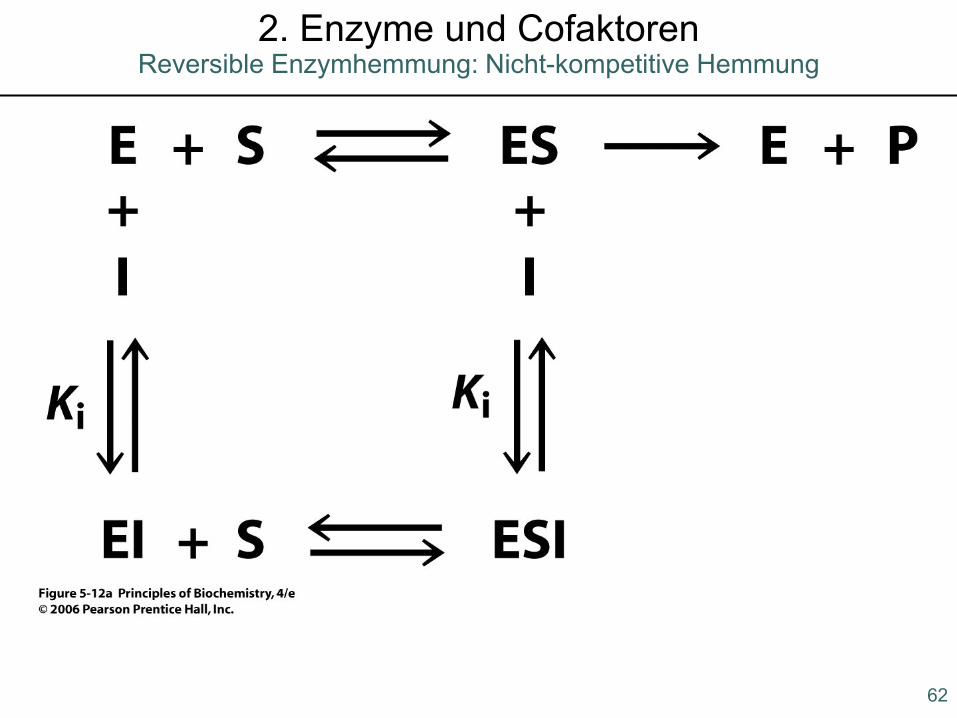
2. Enzyme und CofaktorenReversible Enzymhemmung: Nicht-kompetitive Hemmung
63
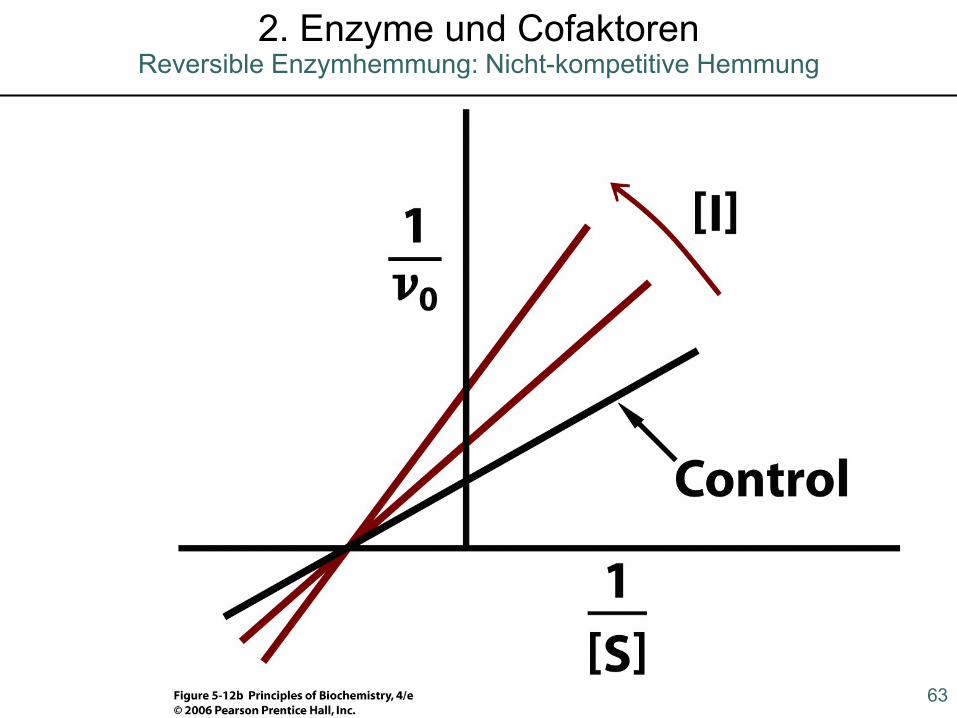
2. Enzyme und CofaktorenReversible Enzymhemmung: Nicht-kompetitive Hemmung

64
Einfluss von Inhibitoren auf vmax und Km
2. Enzyme und Cofaktoren

65
Irreversible Enzymhemmung2. Enzyme und Cofaktoren
Der Inhibitor bleibt „fest“ an der aktiven Stelle gebunden, d.h. eine Dissoziation des Enzym-Inhibitor-Komplexes in freies Enzym und Inhibitor ist nicht möglich. Das Enzym bleibt „vergiftet“. Es muss neues Enzym hergestellt werden.
Beispiele für irreversible Inhibition:
-Alkylphosphate (z.B. Sarin = Acetylcholinesterase-Hemmer)
-CN--Ionen (z.B. Zyankali = Hemmung der Cytochrom-c-Oxidase)
-Schwermetalle (z.B. As2+ = Hemmung der Glycerinaldehyd-3-phosphat-Dehydrogenase
Selbstmord-Substrate:
Pseudosubstrate, die Enzyme durch kovalente Bindung an das aktive Zentrum irreversibel hemmen und darunter selbst funktionsunfähig werden, z.B. Serinprotease-inhibitoren.
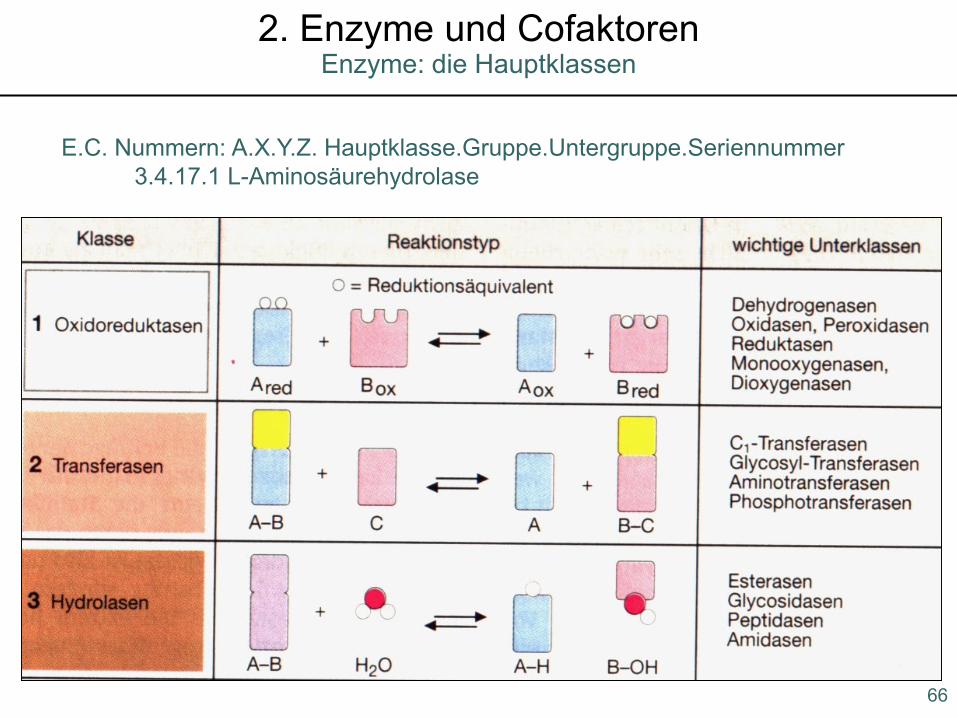
E.C. Nummern: A.X.Y.Z. Hauptklasse.Gruppe.Untergruppe.Seriennummer3.4.17.1 L-Aminosäurehydrolase
66
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
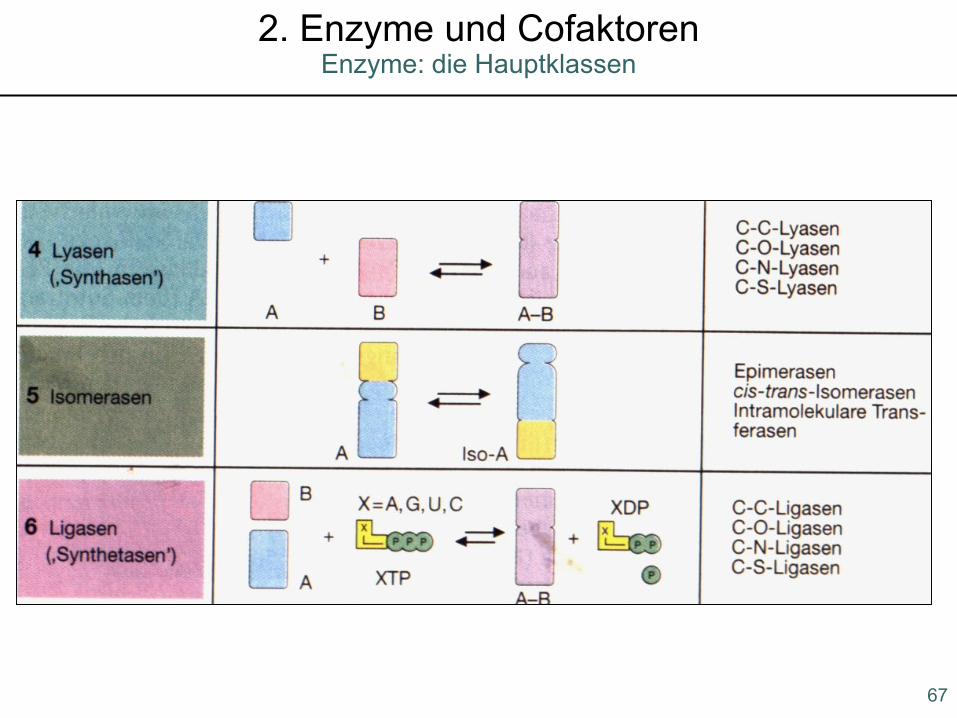
67
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
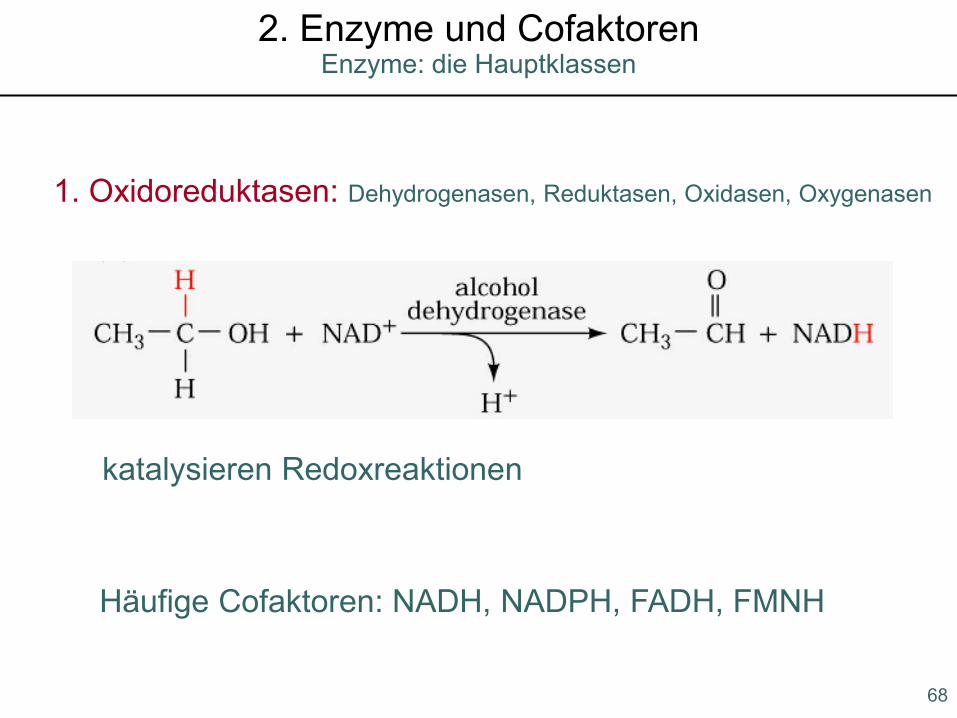
1. Oxidoreduktasen: Dehydrogenasen, Reduktasen, Oxidasen, Oxygenasen
katalysieren Redoxreaktionen
Häufige Cofaktoren: NADH, NADPH, FADH, FMNH
68
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
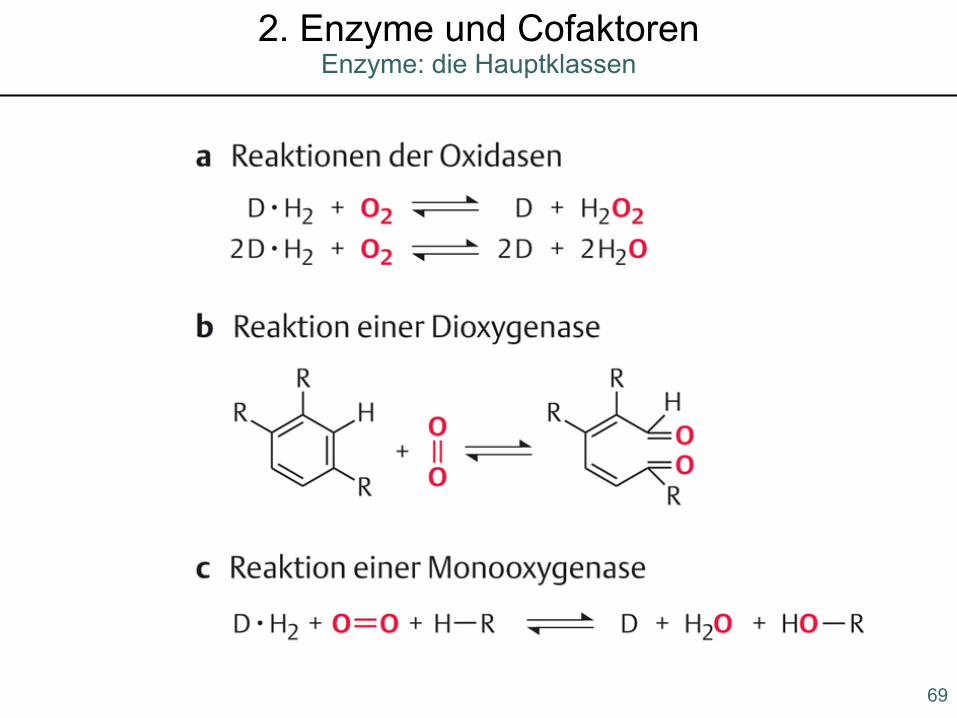
69
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
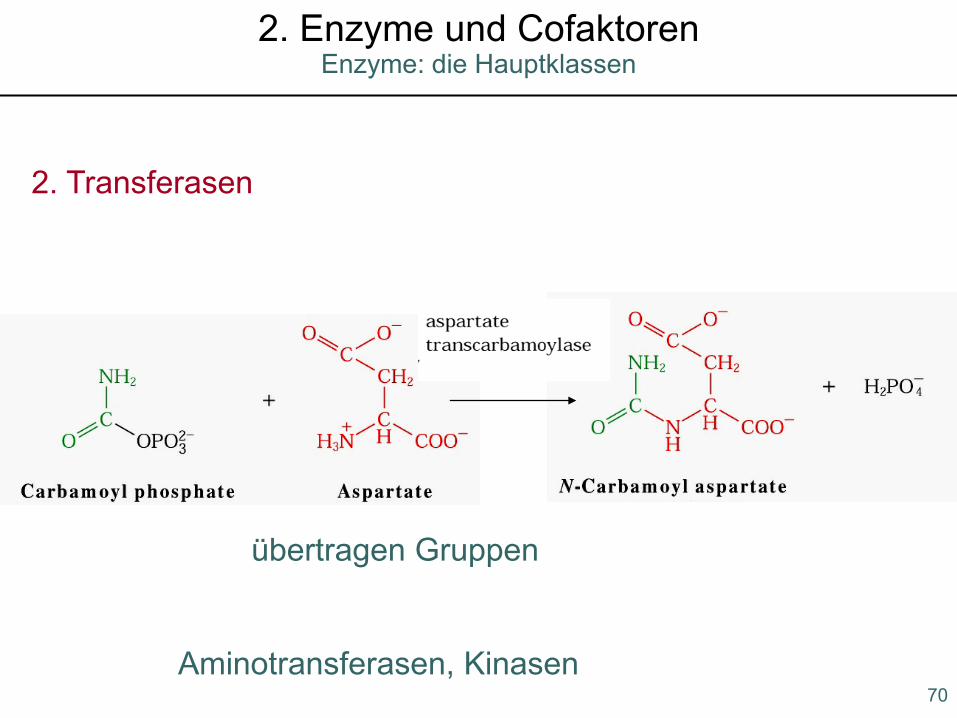
2. Transferasen
übertragen Gruppen
Aminotransferasen, Kinasen70
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
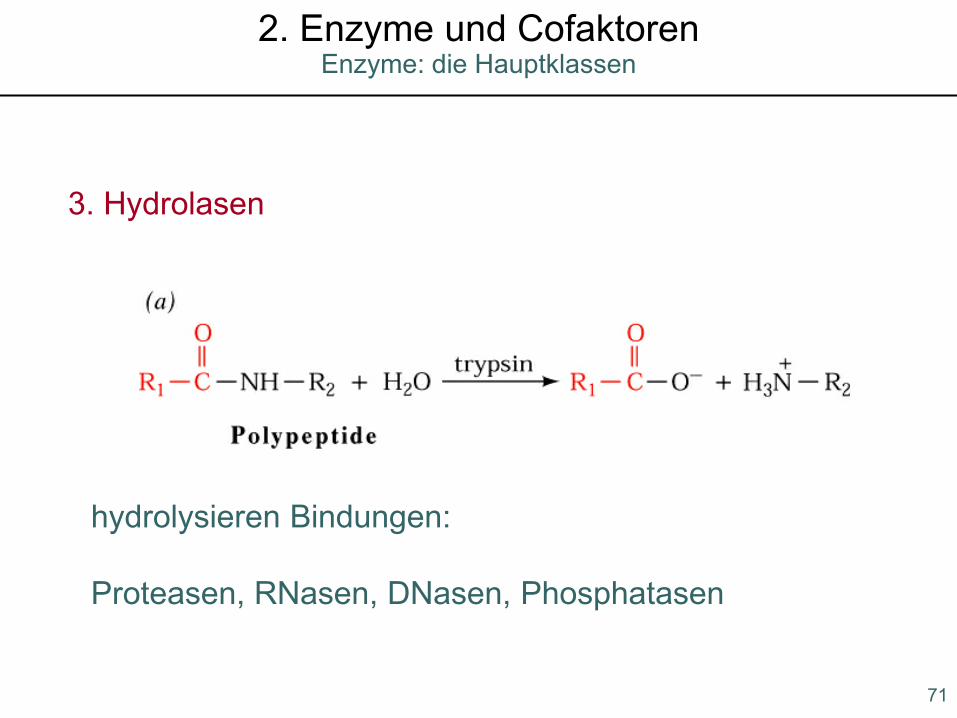
3. Hydrolasen
hydrolysieren Bindungen:
Proteasen, RNasen, DNasen, Phosphatasen
71
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
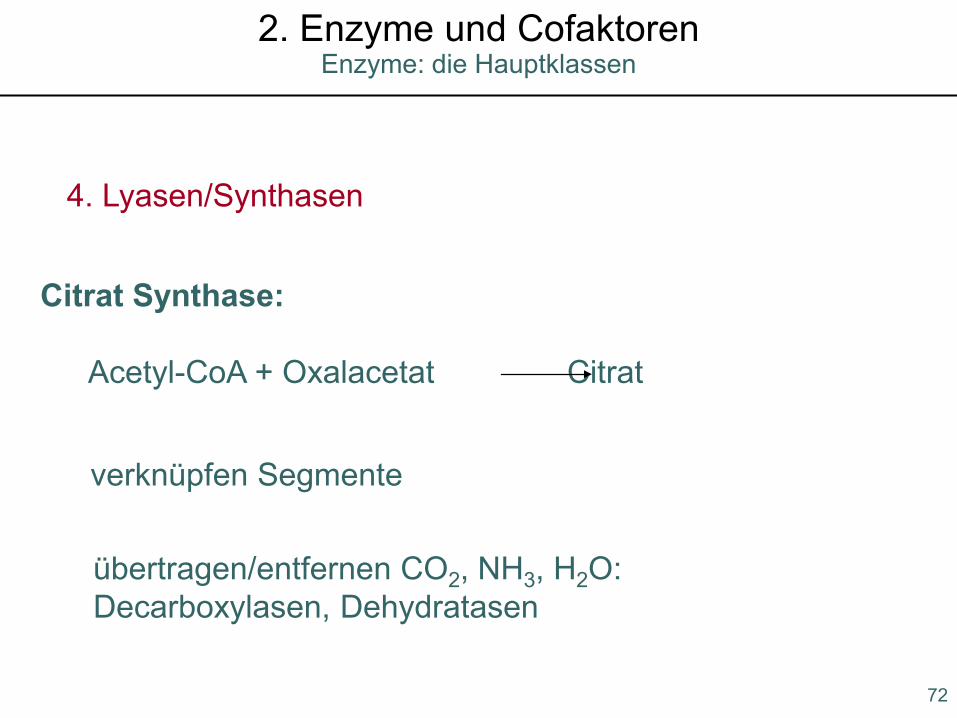
4. Lyasen/Synthasen
verknüpfen Segmente
Citrat Synthase:
Acetyl-CoA + Oxalacetat Citrat
übertragen/entfernen CO2, NH3, H2O:Decarboxylasen, Dehydratasen
72
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
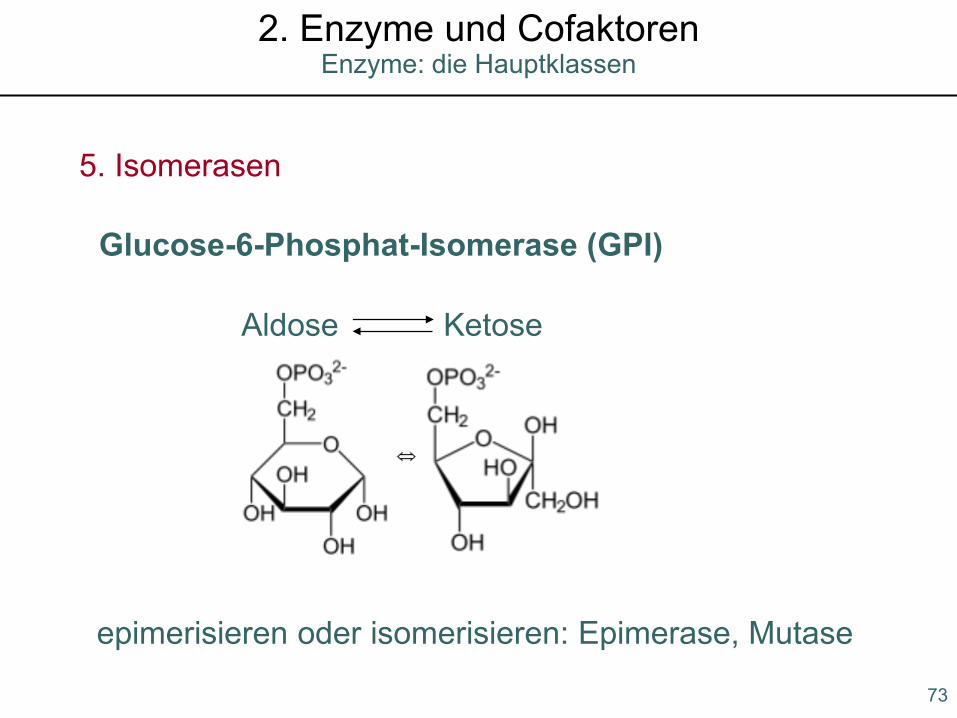
5. Isomerasen
epimerisieren oder isomerisieren: Epimerase, Mutase
Glucose-6-Phosphat-Isomerase (GPI)
Aldose Ketose
73
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
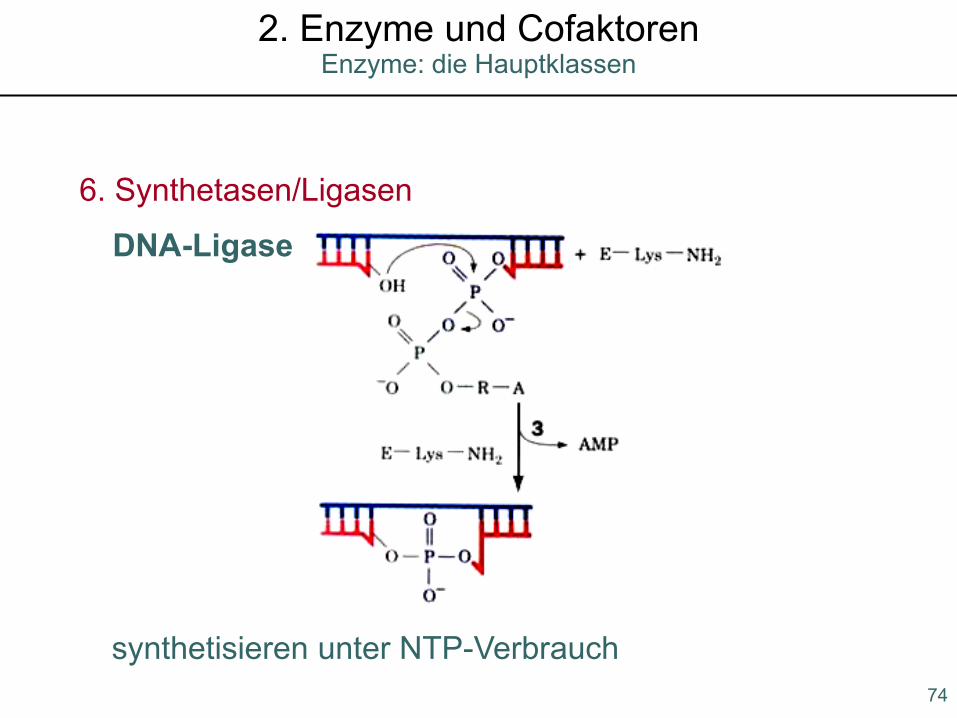
6. Synthetasen/Ligasen
synthetisieren unter NTP-Verbrauch
DNA-Ligase
74
Enzyme: die Hauptklassen2. Enzyme und Cofaktoren
75
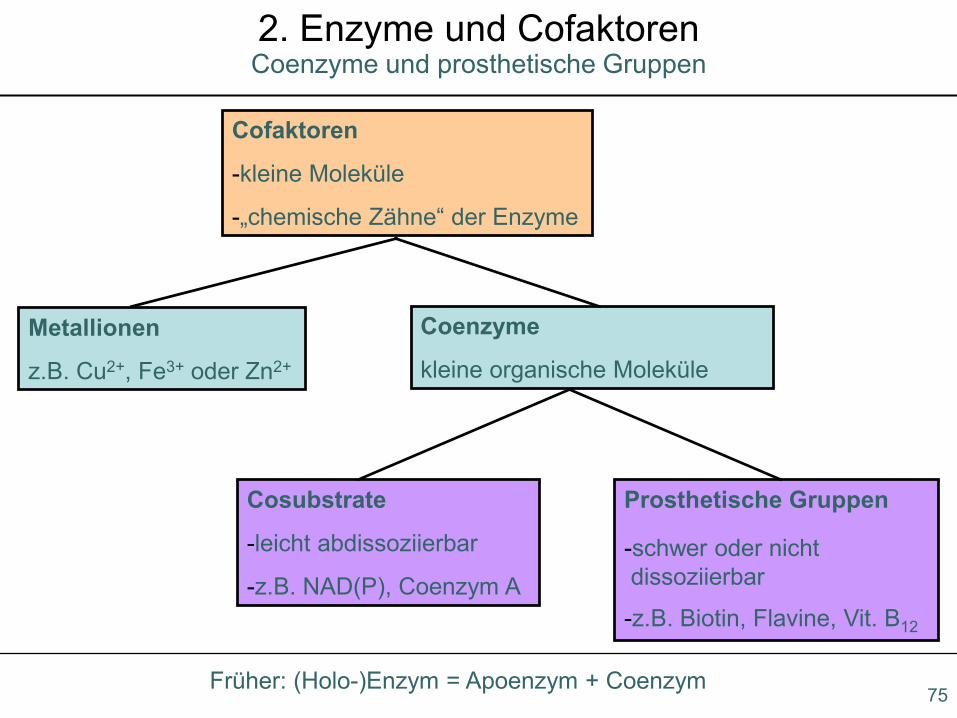
Coenzyme und prosthetische Gruppen2. Enzyme und Cofaktoren
Cofaktoren
-kleine Moleküle
-„chemische Zähne“ der Enzyme
Metallionen
z.B. Cu2+, Fe3+ oder Zn2+
Coenzyme
kleine organische Moleküle
Cosubstrate
-leicht abdissoziierbar
-z.B. NAD(P), Coenzym A
Prosthetische Gruppen
-schwer oder nicht dissoziierbar
-z.B. Biotin, Flavine, Vit. B12
Früher: (Holo-)Enzym = Apoenzym + Coenzym
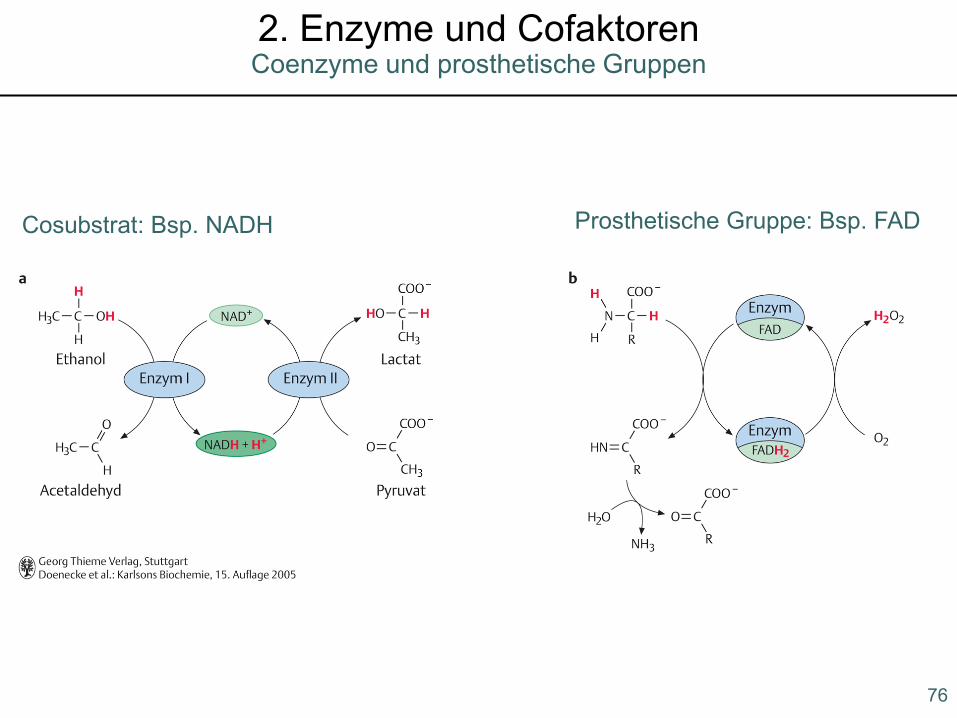
Cosubstrat: Bsp. NADH Prosthetische Gruppe: Bsp. FAD
76
Coenzyme und prosthetische Gruppen2. Enzyme und Cofaktoren

Vitamine: organische Verbindungen, die vom Menschen nicht synthetisiert werden können, und mit der Nahrung aufgenommen werden müssen. Ein Mangel an Vitaminen führt zu Stoffwechselstörungen.
Vitamin Coenzym / prosthetische GruppeB1 Thiamin Thiamindiphosphat TPP
B2-Gruppe
-Riboflavin Flavinadenosin-Di-/Mono-phosphat FAD/FMN
-Nicotinsäureamid Nicotinamidadenin-Dinukleotid-(Phosphat) NAD(P)
´ -Folsäure Tetrahydrofolsäure THF
-Pantothensäure Coenzym A CoA
B6 Pyridoxin Pyridoxal-Phosphat PLP
B12 Cobalamin B12-Enzyme B12
H Biotin Biotin-Carboxylasen
77
Coenzyme, prosthetische Gruppen und Vitamine2. Enzyme und Cofaktoren

Prinzip: Coenzyme können funktionelle Gruppen übertragen
Phosphoryl-Transfer: Glucose + ATP + H2O → Glucose-6-P + ADP
Acetyl-Transfer: Acetyl-Coenzym A + Glucosamin → N-Acetyl-Glucosamin + CoA
Methyl-Transfer: S-Adenosylmethionin + R → S-Adenosylhomocystein + R-CH3
Beispiele:
Coenzyme mit Gruppenübertragungspotential: ATP, CoA, THF, Biotin, PLP, TPP
78
Coenzyme und Gruppenübertragung2. Enzyme und Cofaktoren
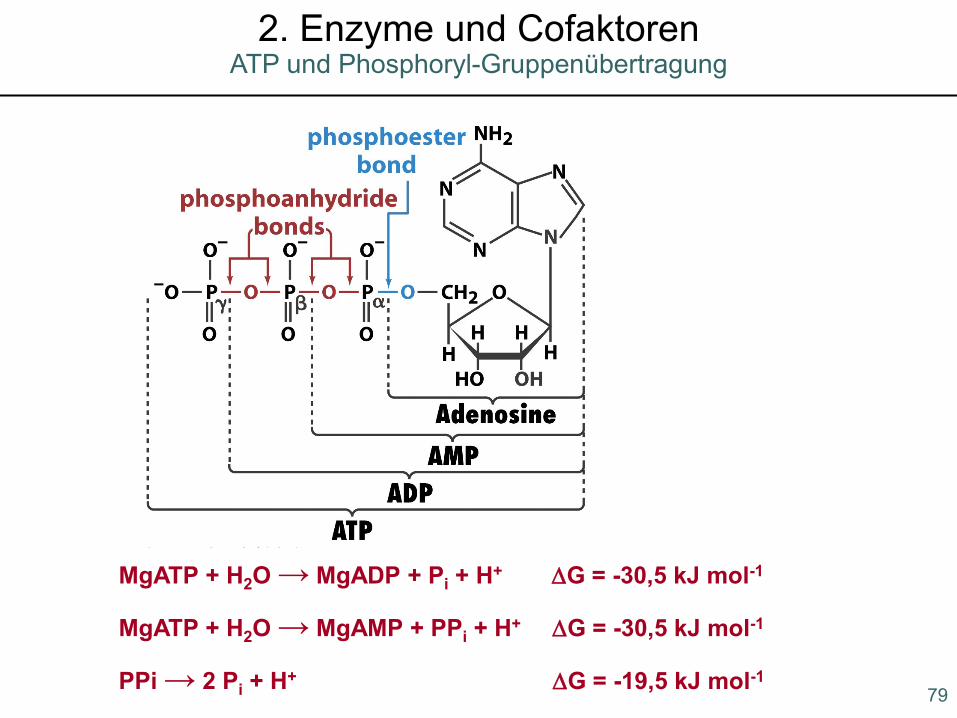
MgATP + H2O → MgADP + Pi + H+ ∆G = -30,5 kJ mol-1
MgATP + H2O → MgAMP + PPi + H+ ∆G = -30,5 kJ mol-1
PPi → 2 Pi + H+ ∆G = -19,5 kJ mol-1 79
ATP und Phosphoryl-Gruppenübertragung2. Enzyme und Cofaktoren
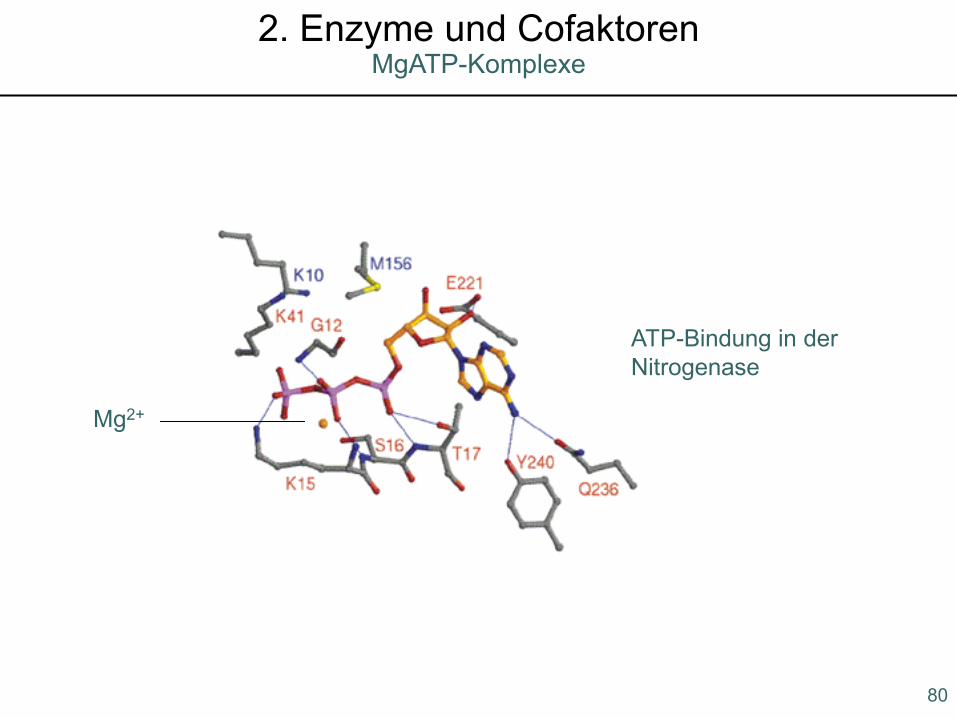
Mg2+
ATP-Bindung in der Nitrogenase
80
MgATP-Komplexe2. Enzyme und Cofaktoren

MgATP + H2O → MgADP + Pi ∆G0‘ = -30,5 kJ mol-1
In der Zelle (aerob, Durchschnitt): [ATP]: 3 mM[ADP]: 0,8 mM[Pi]: 4 mMT = 37°C
[ATP], [ADP], [Pi]: je 1 M, 25°C
∆G = ∆G°‘ + RT ln ([ADP] [Pi] / [ATP])
∆G = -30,5 kJ mol-1 – 17,6 kJ mol-1 = ~ 50 kJ mol-1
81
Zelluläre und Standardenthalpie der ATP-Hydrolyse2. Enzyme und Cofaktoren
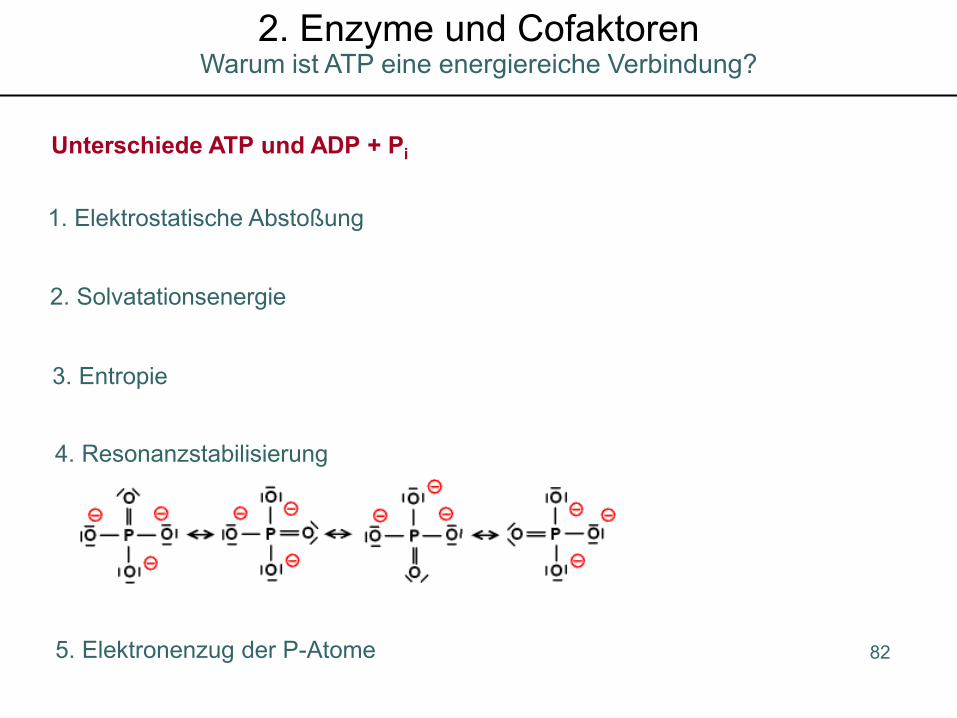
1. Elektrostatische Abstoßung
2. Solvatationsenergie
Unterschiede ATP und ADP + Pi
3. Entropie
4. Resonanzstabilisierung
5. Elektronenzug der P-Atome 82
Warum ist ATP eine energiereiche Verbindung?2. Enzyme und Cofaktoren

83
Vorteile von ATP2. Enzyme und Cofaktoren
Vorteil des ATP gegenüber Verbindungen mit anderen Säureanhydriden:
› Phosphoanhydridbindungen benötigen bei einer normalen Hydrolyse eine hohe Aktivierungsenergie
› bei enzymatischer Hydrolyse minimiert (ATP also energiereich im Sinne der Hydrolyse, nicht der Bindungsspaltung)
› ATP unter physiologischen Bedingungen sehr stabil
› in enzymatischen Reaktionen aber ein schneller Energielieferant

- Energieladung = Maßzahl für den Energiestatus einer Zelle
- beschreibt das Verhältnis aller Adenosylnukleotide
- Engergieladung = 1, wenn nur ATP vorliegt (hypothetischer Fall)
- Realität: Energieladung zwischen 0,7 und 0,95 -›reguliert durch Schlüsselenzyme des Stoffwechsels
(z.B. Phosphofructokinase)
Energieladung (energy charge)2. Enzyme und Cofaktoren
84

-> Bewertung von ADP als ½ ATP
Reaktion der Myokinase (Muskel):
Energieladung (energy charge)2. Enzyme und Cofaktoren
85

Phosphokreatin als Energiespeicher im Muskel!
86
Biologisch relevante energiereiche Phosphate2. Enzyme und Cofaktoren
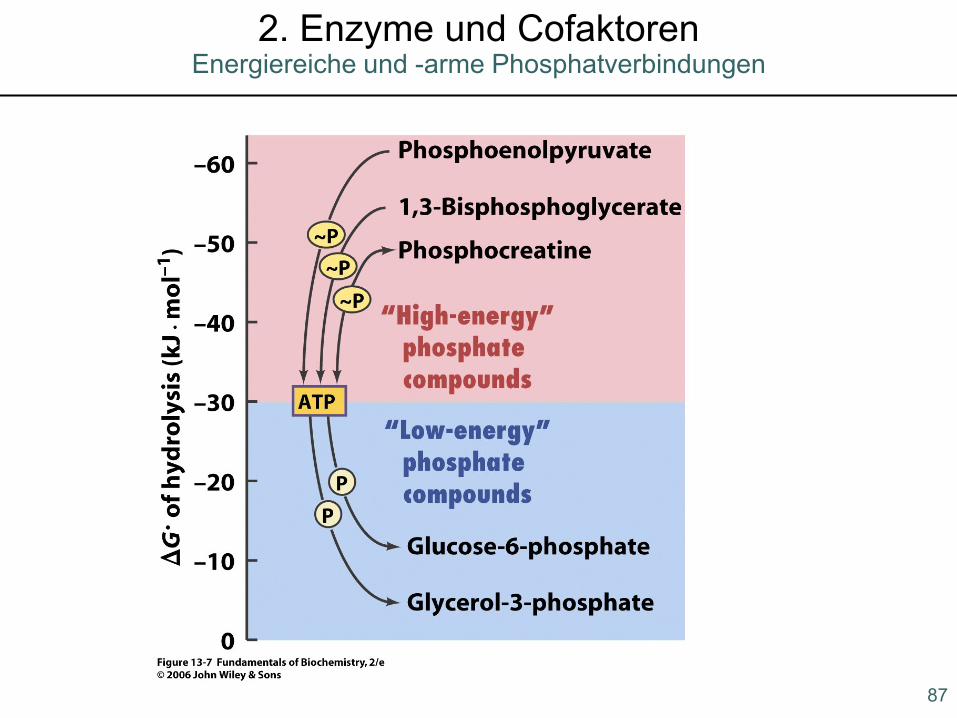
87
Energiereiche und -arme Phosphatverbindungen2. Enzyme und Cofaktoren
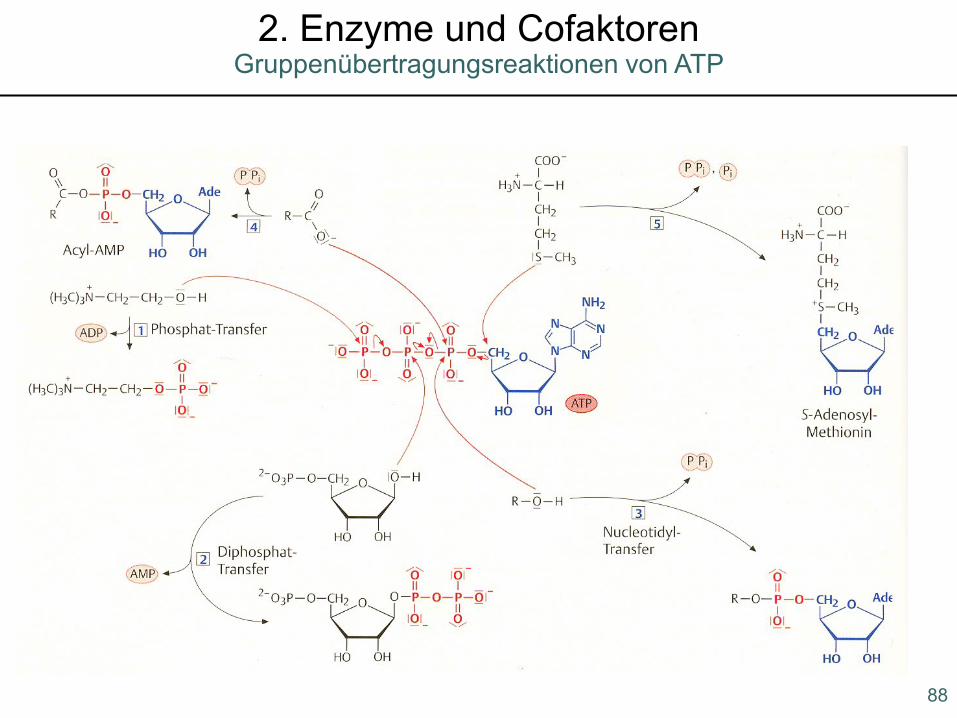
88
Gruppenübertragungsreaktionen von ATP2. Enzyme und Cofaktoren
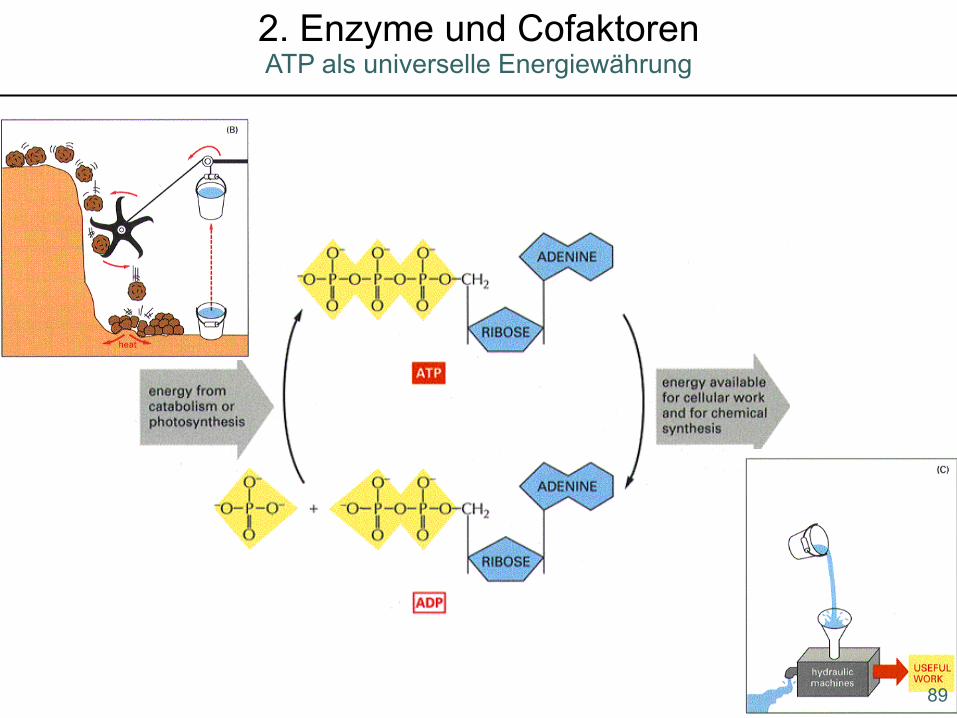
89
ATP als universelle Energiewährung2. Enzyme und Cofaktoren

(1) A + B C + D ∆G1
Gesetz der Additivität der freien Enthalpie!
(2) D + E F + G ∆G2
(1) + (2): A + B + E C + F + G ∆G3 = ∆G1 + ∆G2
Beispiel 1:
90
Kopplung von Reaktionen2. Enzyme und Cofaktoren
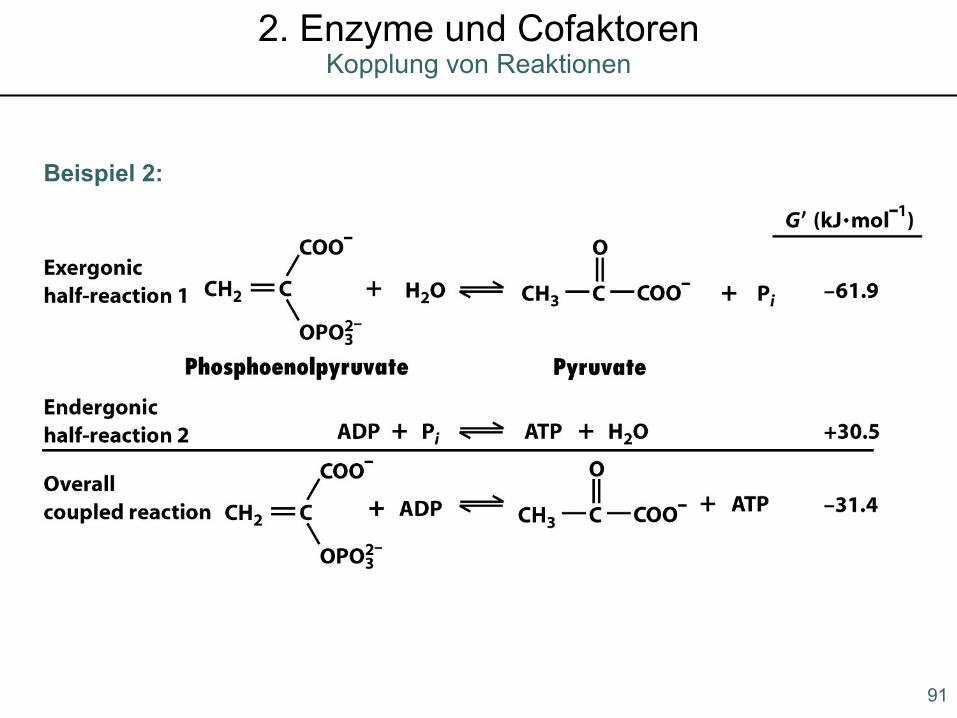
Beispiel 2:
91
Kopplung von Reaktionen2. Enzyme und Cofaktoren
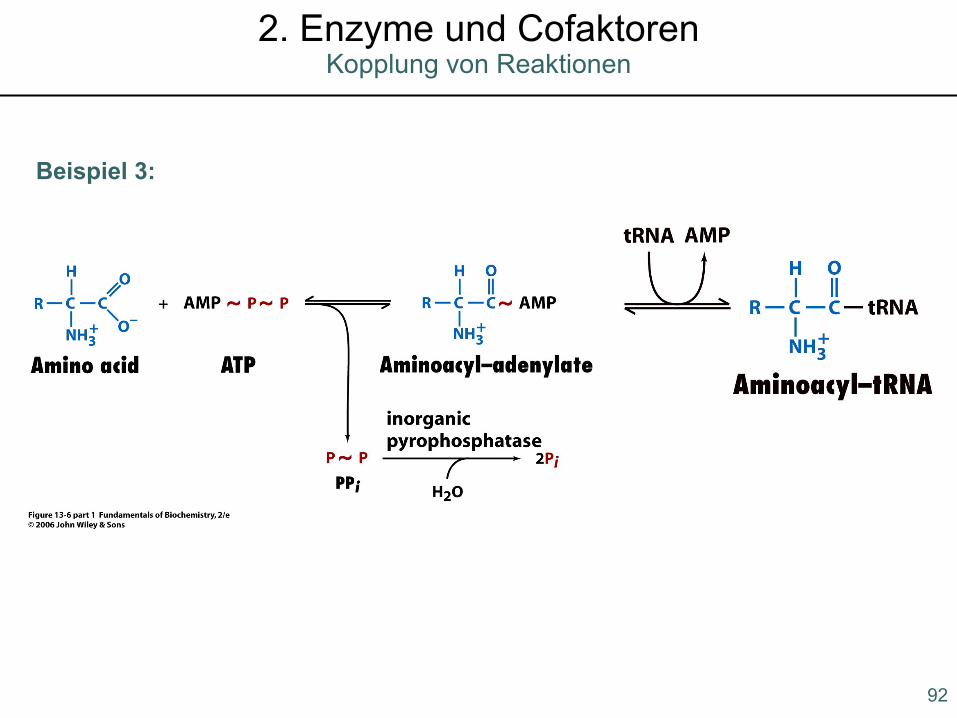
Beispiel 3:
92
Kopplung von Reaktionen2. Enzyme und Cofaktoren
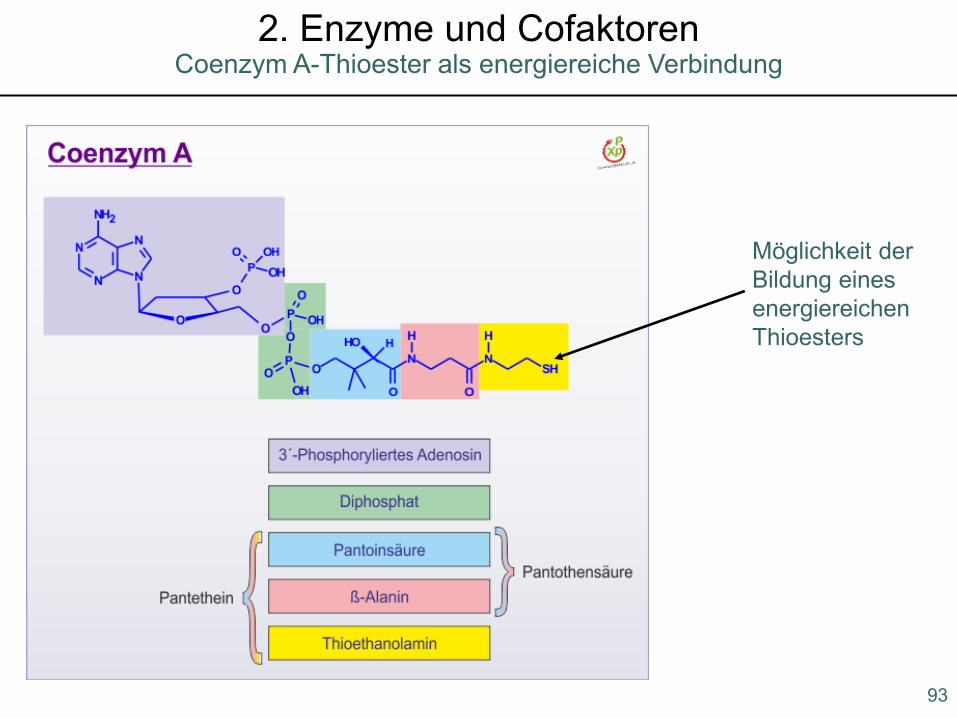
93
Coenzym A-Thioester als energiereiche Verbindung2. Enzyme und Cofaktoren
Möglichkeit der Bildung eines energiereichen Thioesters
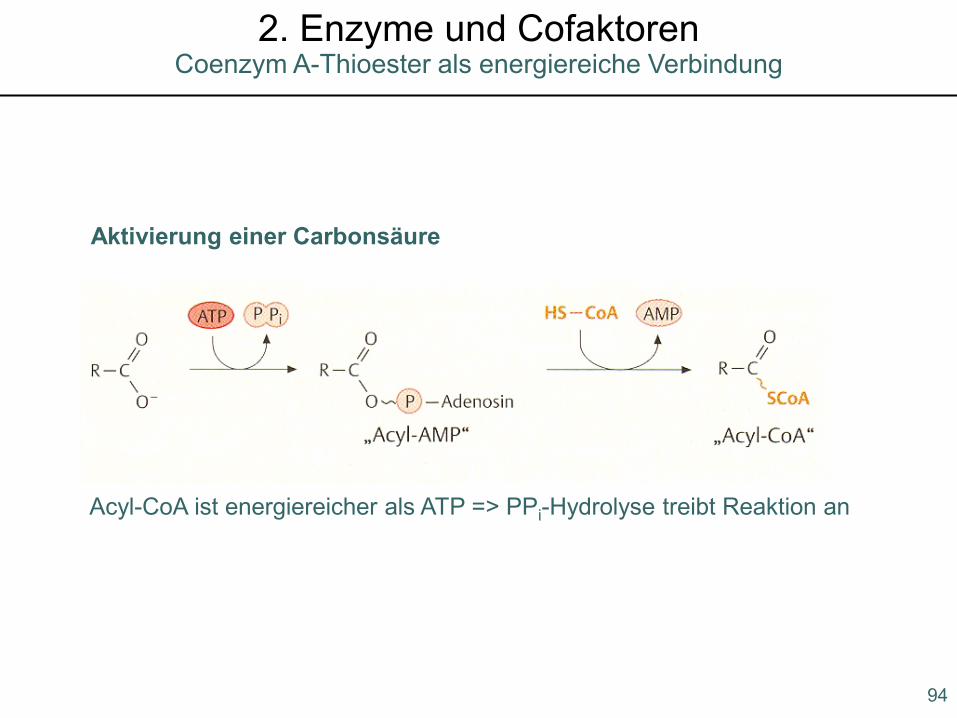
Aktivierung einer Carbonsäure
Acyl-CoA ist energiereicher als ATP => PPi-Hydrolyse treibt Reaktion an
94
Coenzym A-Thioester als energiereiche Verbindung2. Enzyme und Cofaktoren

Typische Reaktionen von Acetyl-CoA
1. Esterbildung (Acetylierungen)Glucosamin + Acetyl-CoA → N-Acetyl-Glucosamin + CoA
2. Kondensationen (CH-acide Methylgruppe)Bsp.: Fettsäurestoffwechsel, Citratsynthase
95
Die aktivierte Essigsäure2. Enzyme und Cofaktoren
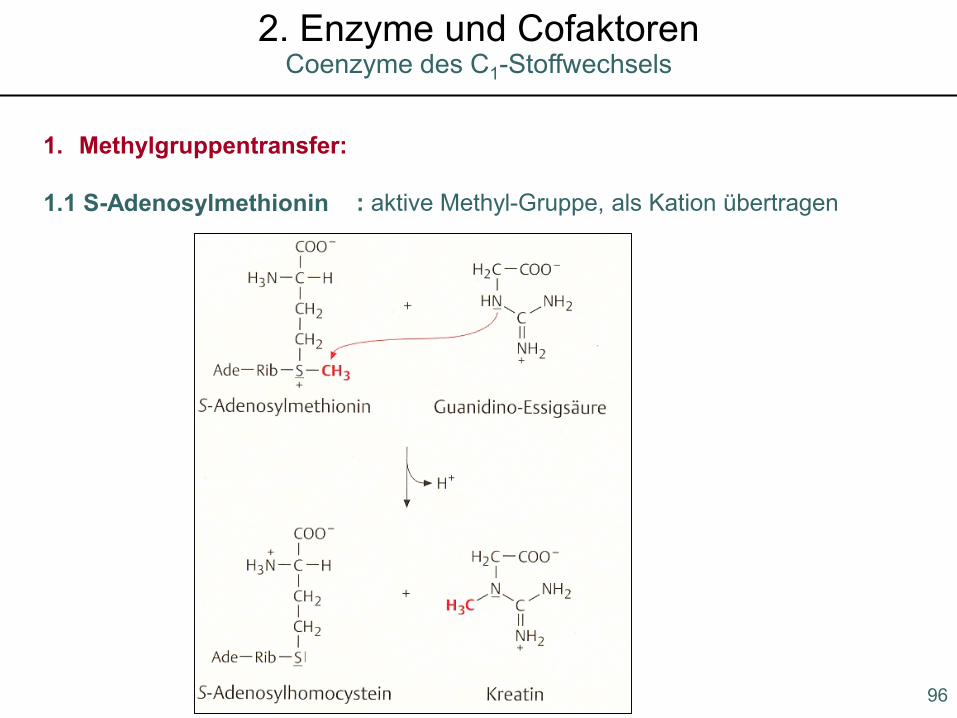
1. Methylgruppentransfer:
1.1 S-Adenosylmethionin : aktive Methyl-Gruppe, als Kation übertragen
96
Coenzyme des C1-Stoffwechsels2. Enzyme und Cofaktoren
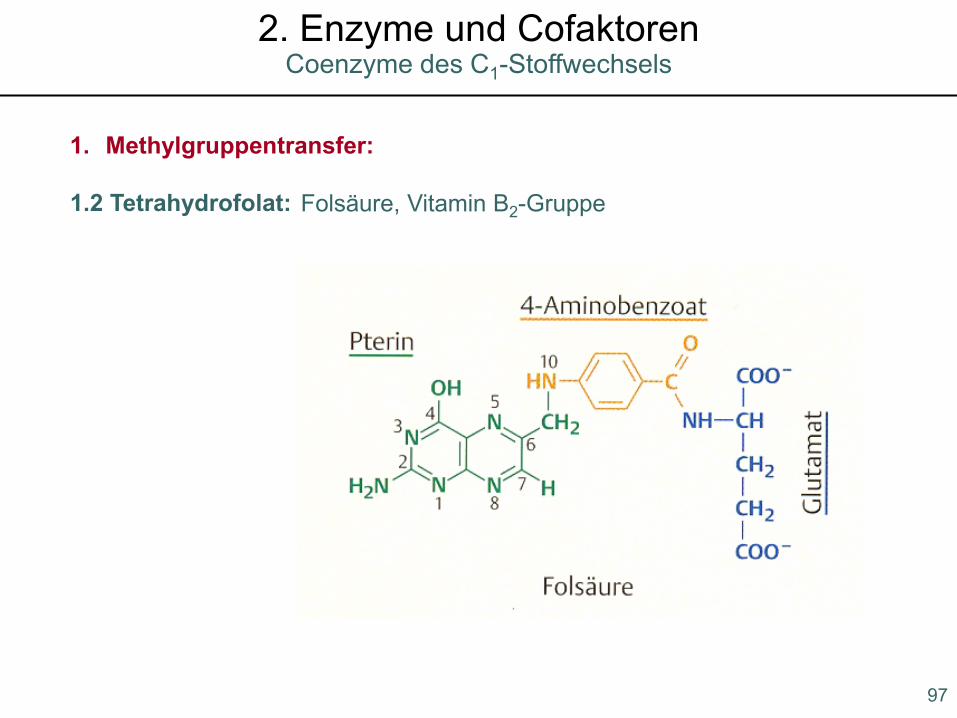
1. Methylgruppentransfer:
1.2 Tetrahydrofolat: Folsäure, Vitamin B2-Gruppe
97
Coenzyme des C1-Stoffwechsels2. Enzyme und Cofaktoren
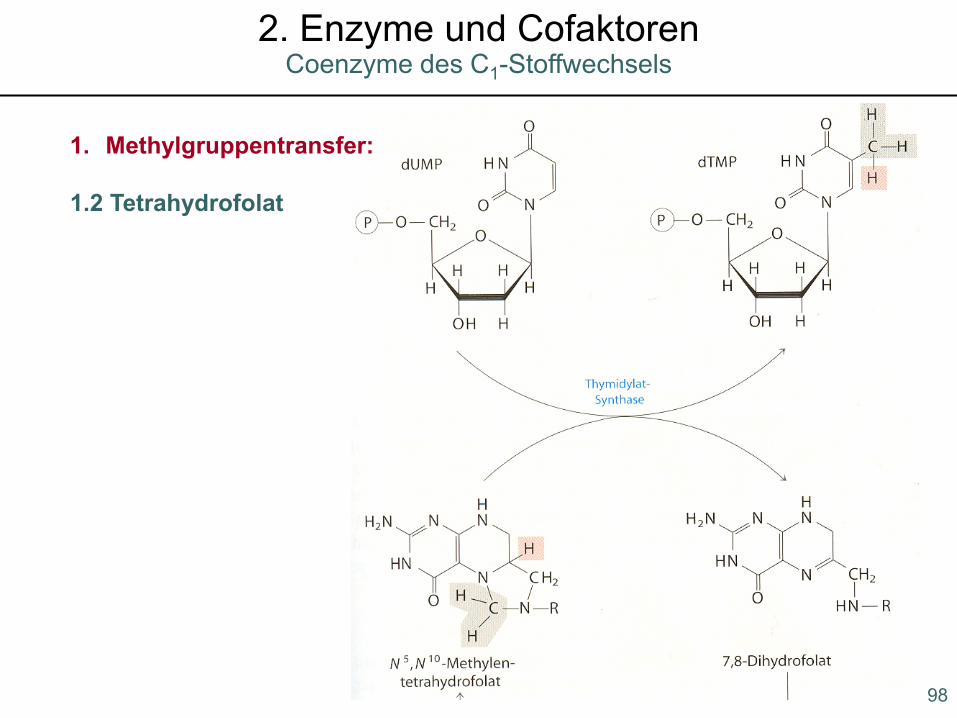
1. Methylgruppentransfer:
1.2 Tetrahydrofolat
98
Coenzyme des C1-Stoffwechsels2. Enzyme und Cofaktoren
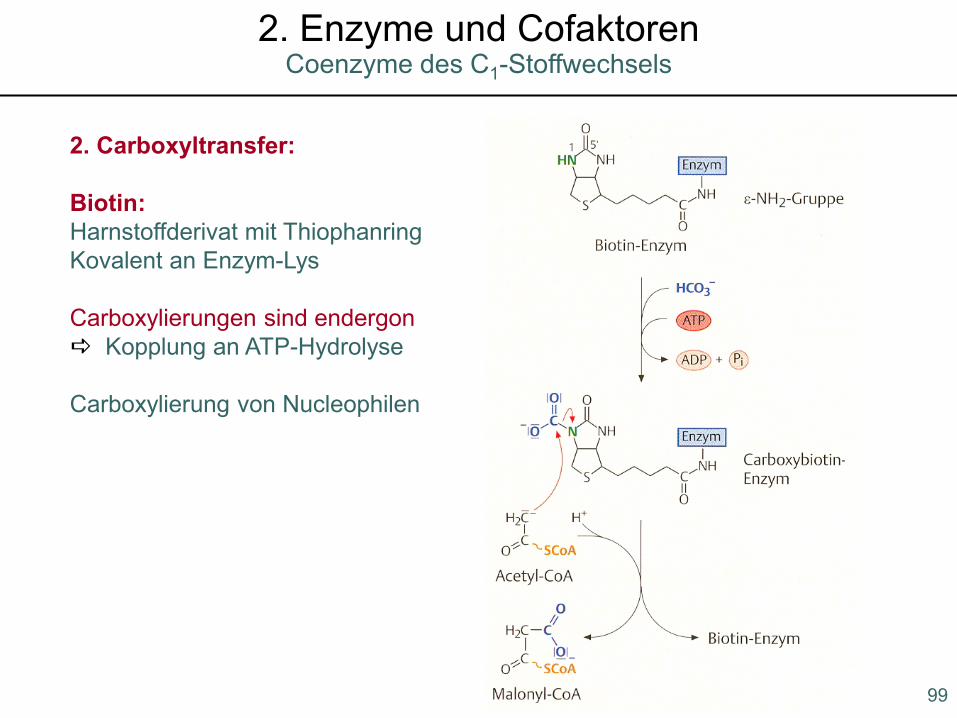
2. Carboxyltransfer:
Biotin: Harnstoffderivat mit ThiophanringKovalent an Enzym-Lys
Carboxylierungen sind endergon Kopplung an ATP-Hydrolyse
Carboxylierung von Nucleophilen
99
Coenzyme des C1-Stoffwechsels2. Enzyme und Cofaktoren
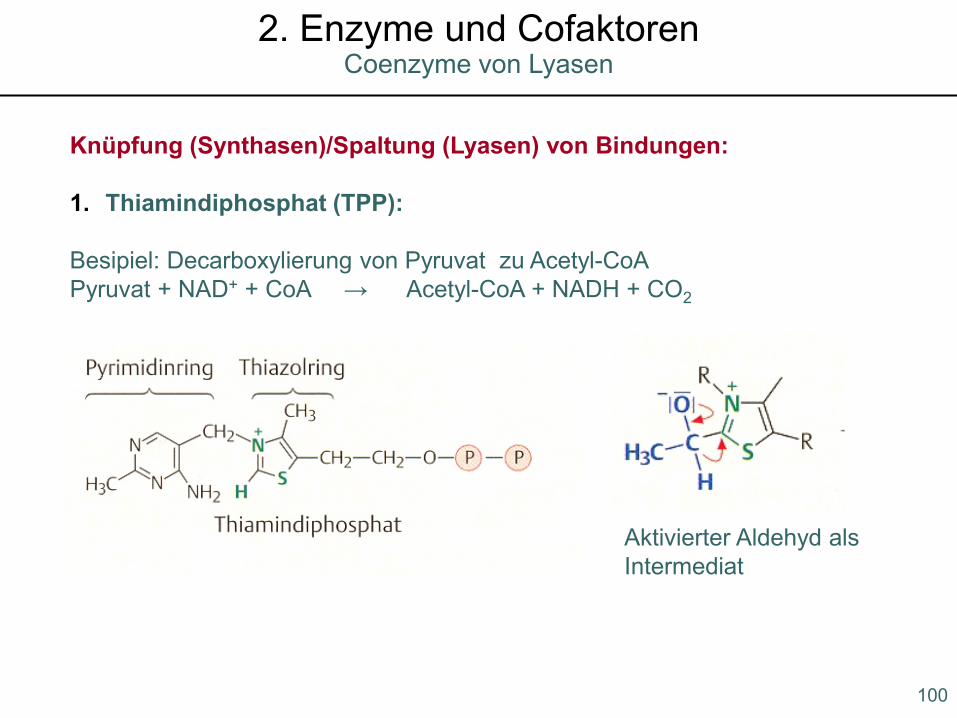
Knüpfung (Synthasen)/Spaltung (Lyasen) von Bindungen:
1. Thiamindiphosphat (TPP):
Besipiel: Decarboxylierung von Pyruvat zu Acetyl-CoAPyruvat + NAD+ + CoA → Acetyl-CoA + NADH + CO2
Aktivierter Aldehyd als Intermediat
100
Coenzyme von Lyasen2. Enzyme und Cofaktoren
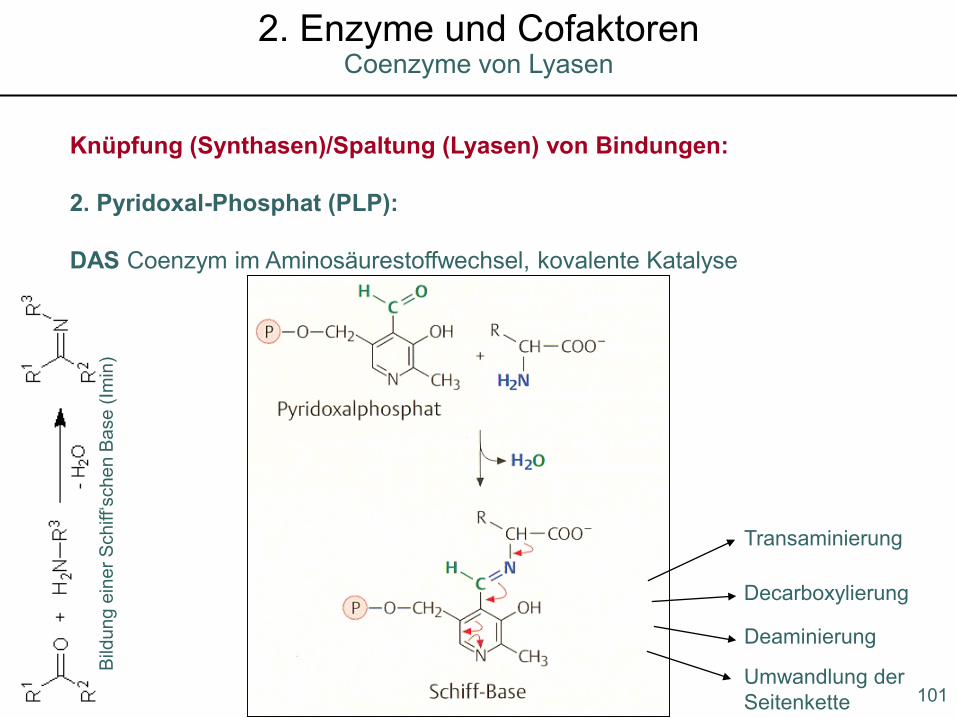
Knüpfung (Synthasen)/Spaltung (Lyasen) von Bindungen:
2. Pyridoxal-Phosphat (PLP):
DAS Coenzym im Aminosäurestoffwechsel, kovalente Katalyse
Transaminierung
Decarboxylierung
Deaminierung
Umwandlung der Seitenkette 101
Coenzyme von Lyasen2. Enzyme und Cofaktoren
Bild
ung
eine
r Sch
iff‘s
chen
Bas
e (Im
in)
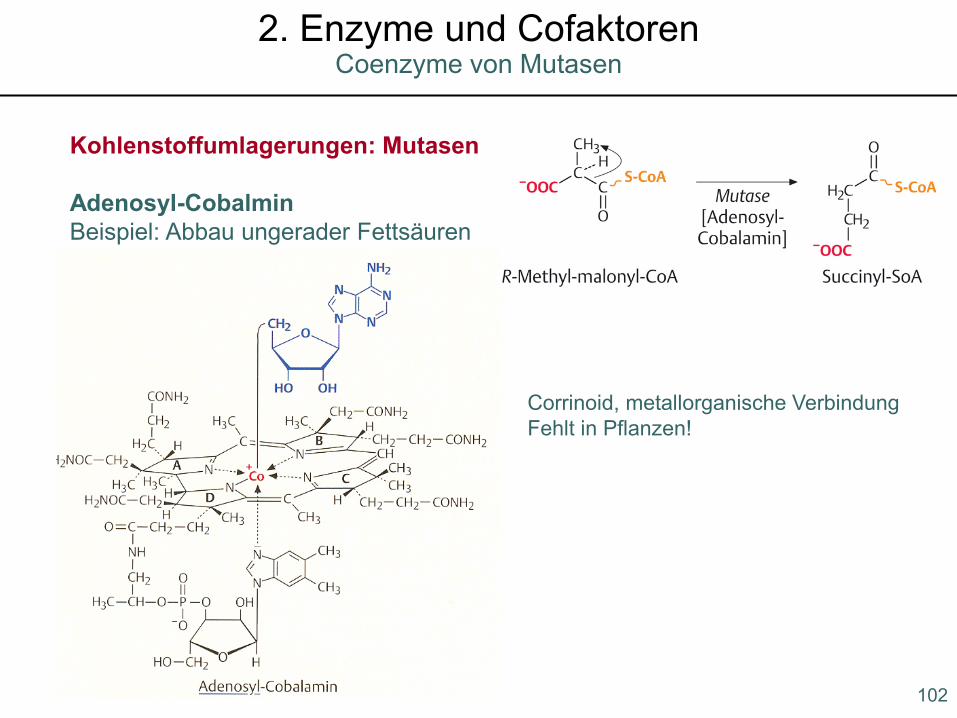
Kohlenstoffumlagerungen: Mutasen
Adenosyl-CobalminBeispiel: Abbau ungerader Fettsäuren
Corrinoid, metallorganische VerbindungFehlt in Pflanzen!
102
Coenzyme von Mutasen2. Enzyme und Cofaktoren
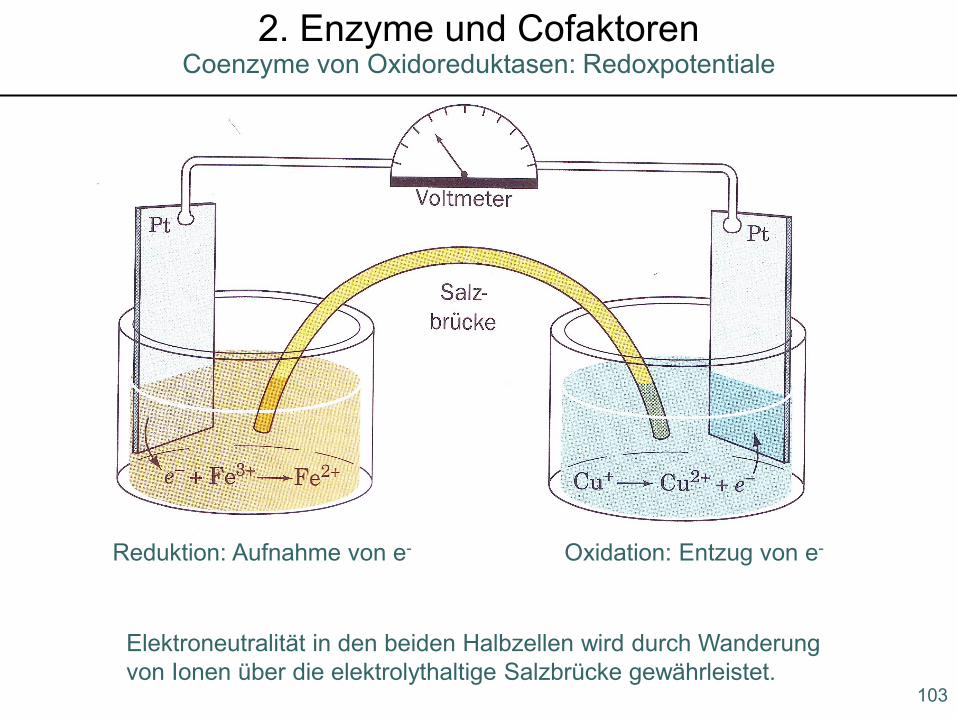
103
Coenzyme von Oxidoreduktasen: Redoxpotentiale2. Enzyme und Cofaktoren
Reduktion: Aufnahme von e- Oxidation: Entzug von e-
Elektroneutralität in den beiden Halbzellen wird durch Wanderung von Ionen über die elektrolythaltige Salzbrücke gewährleistet.

104
Coenzyme von Oxidoreduktasen: Redoxpotentiale2. Enzyme und Cofaktoren
Eh = Redoxpotential eines RedoxpaaresE0 = Redoxpotential eines Redoxpaares 1 M, pH 0E0‘ = Redoxpotential eines Redoxpaares 1 M, pH 7
Standardredoxpotentiale werden verwendet, um Elektronenaffinitäten zu vergleichen
Redoxpotentiale werden auf die Wasserstoffteilreaktion definiert, d.h. der Standardwasserstoffelektrode wird willkürlich ein Redoxpotential von 0 V zugewiesen:
2 H+ + 2 e- H2
Je positiver das Standardredoxpotential, desto höher die Elektronenaffinität der oxidierten Form. Daraus ergibt sich für die oxidierte Form eine umso größere Tendenz, Elektronen aufzunehmen und dadurch in die reduzierte Form über-zugehen.
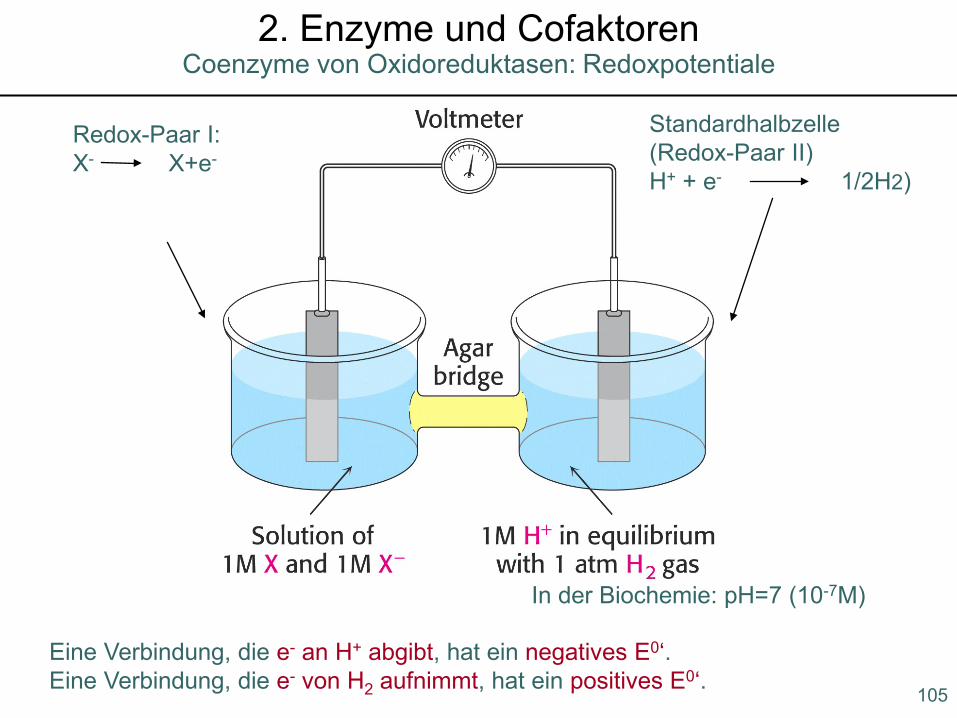
Standardhalbzelle(Redox-Paar II)H+ + e- 1/2H2)
Redox-Paar I:X- X+e-
In der Biochemie: pH=7 (10-7M)
Eine Verbindung, die e- an H+ abgibt, hat ein negatives E0‘.Eine Verbindung, die e- von H2 aufnimmt, hat ein positives E0‘.
Coenzyme von Oxidoreduktasen: Redoxpotentiale2. Enzyme und Cofaktoren
105
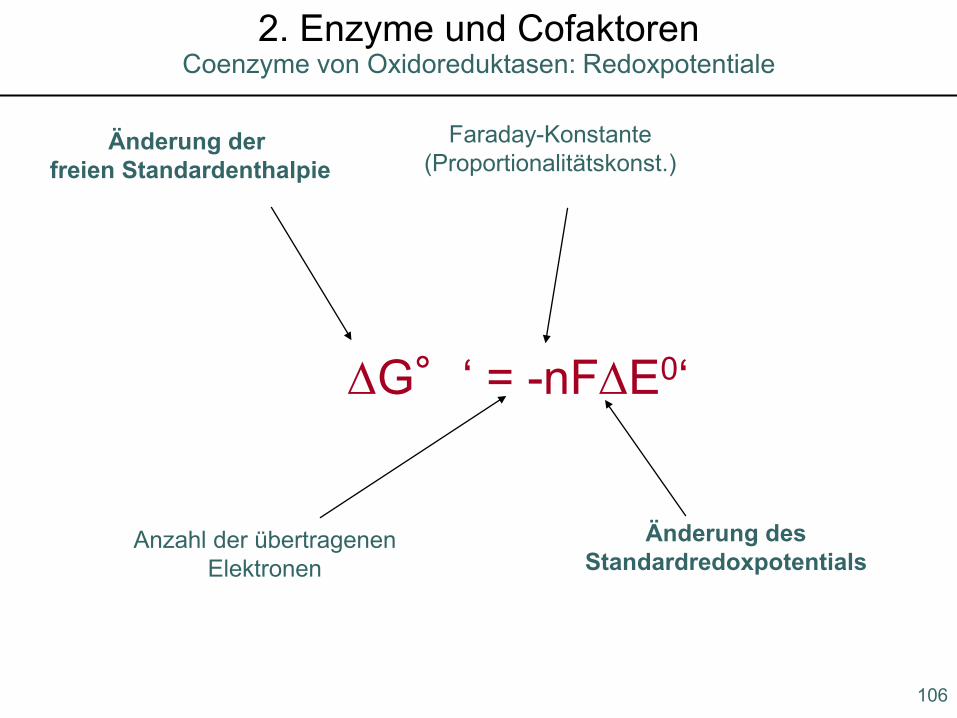
∆G°‘ = -nF∆E0‘
Anzahl der übertragenenElektronen
Faraday-Konstante(Proportionalitätskonst.)
Änderung der freien Standardenthalpie
Änderung desStandardredoxpotentials
Coenzyme von Oxidoreduktasen: Redoxpotentiale2. Enzyme und Cofaktoren
106

Redoxpotentiale von Redox-Cofaktoren
107
Coenzyme von Oxidoreduktasen: Redoxpotentiale2. Enzyme und Cofaktoren
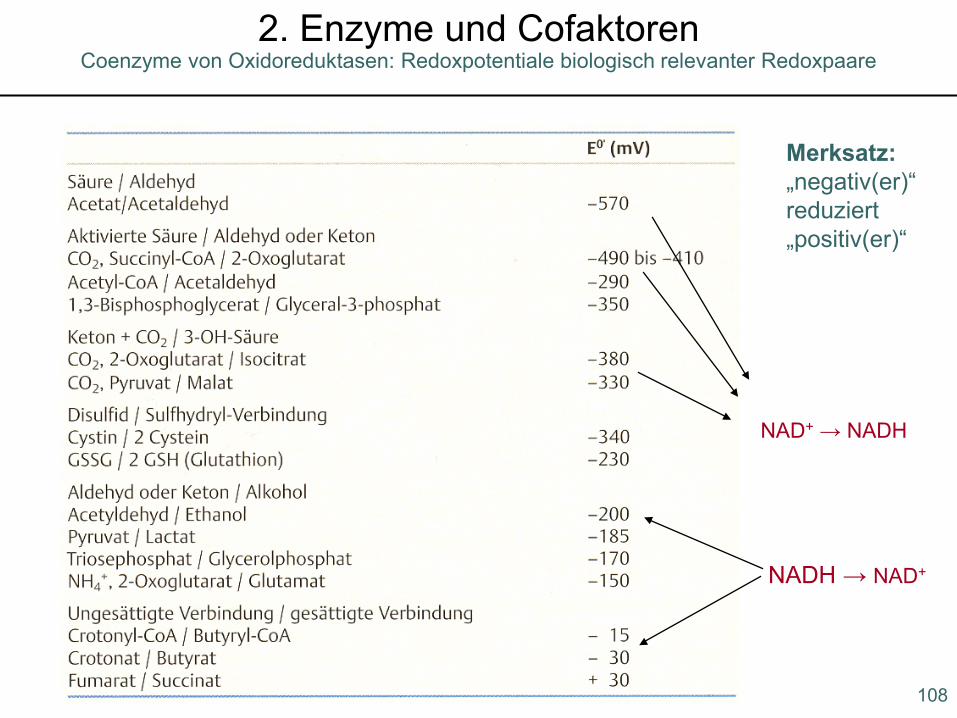
NAD+ → NADH
NADH → NAD+
108
Coenzyme von Oxidoreduktasen: Redoxpotentiale biologisch relevanter Redoxpaare2. Enzyme und Cofaktoren
Merksatz:„negativ(er)“ reduziert „positiv(er)“
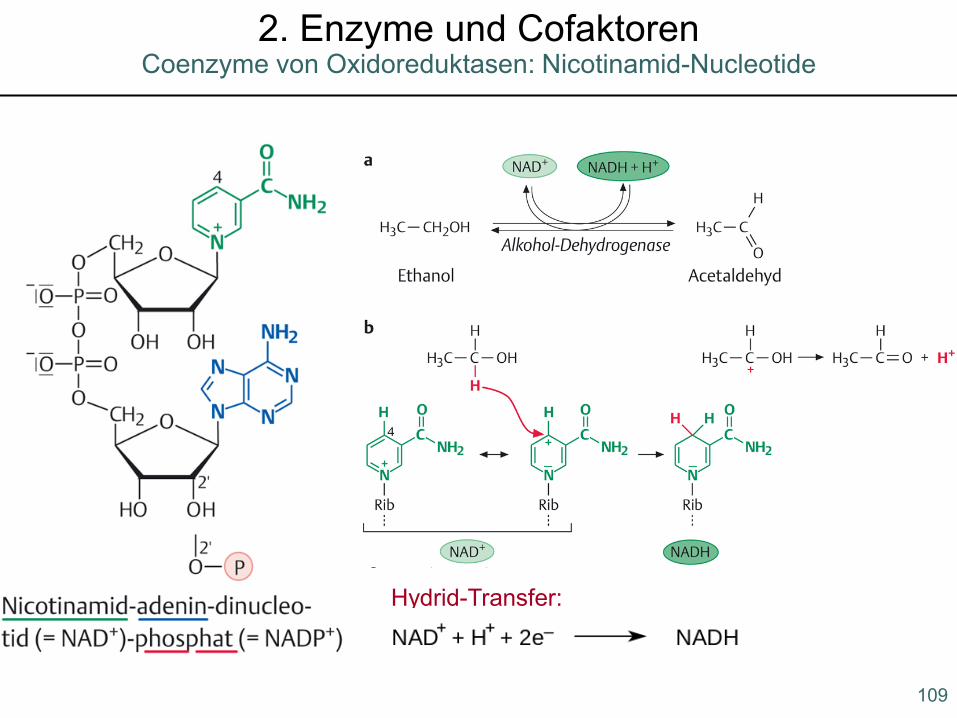
Hydrid-Transfer:
109
Coenzyme von Oxidoreduktasen: Nicotinamid-Nucleotide2. Enzyme und Cofaktoren
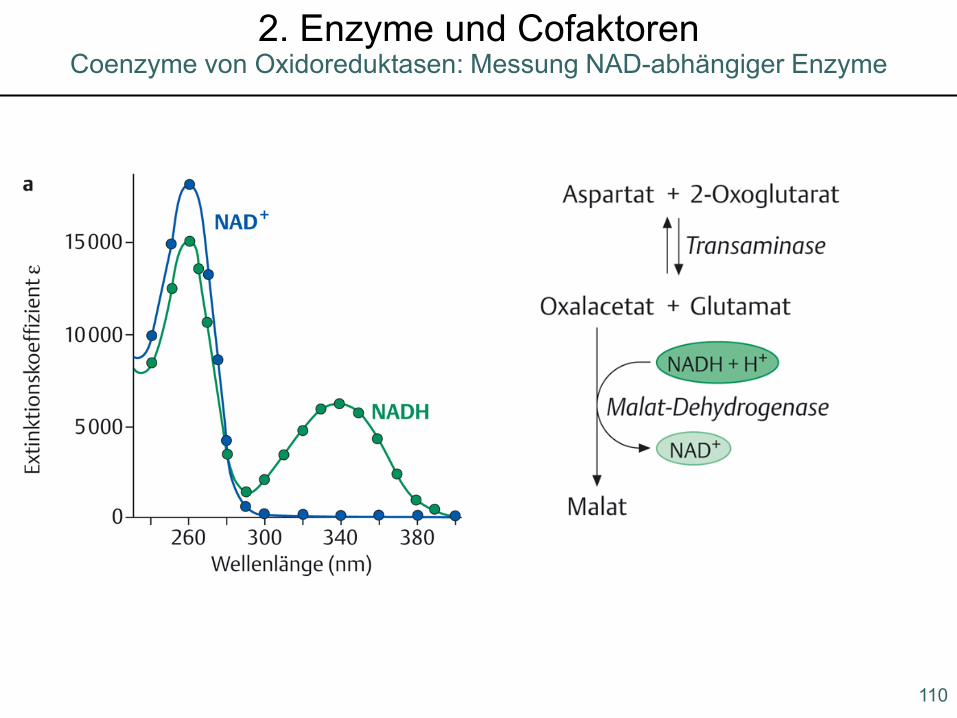
110
Coenzyme von Oxidoreduktasen: Messung NAD-abhängiger Enzyme2. Enzyme und Cofaktoren
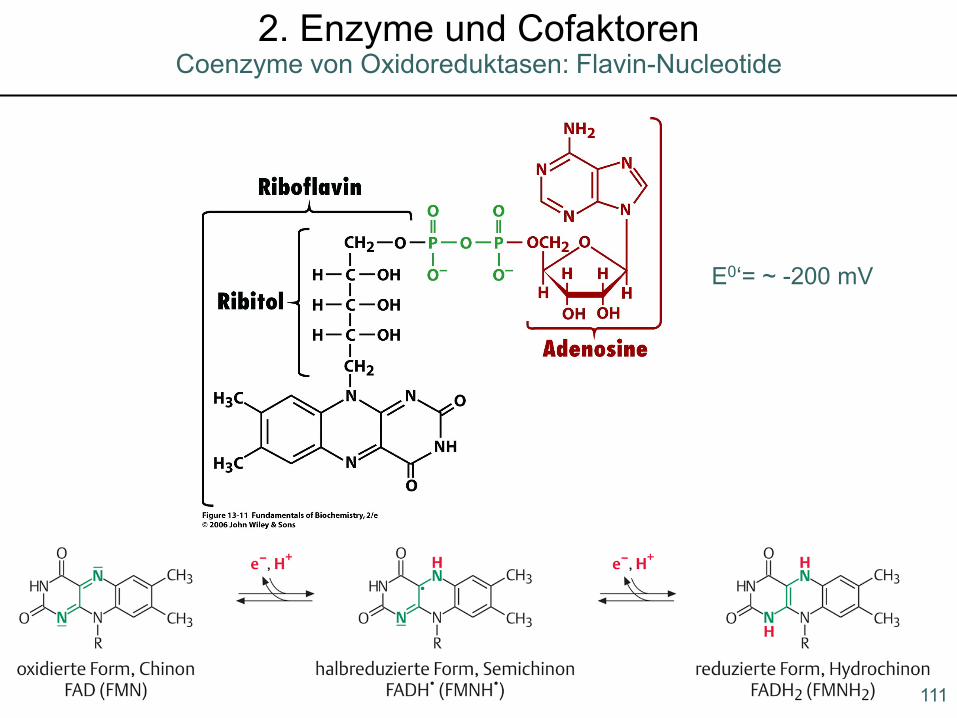
E0‘= ~ -200 mV
111
Coenzyme von Oxidoreduktasen: Flavin-Nucleotide2. Enzyme und Cofaktoren
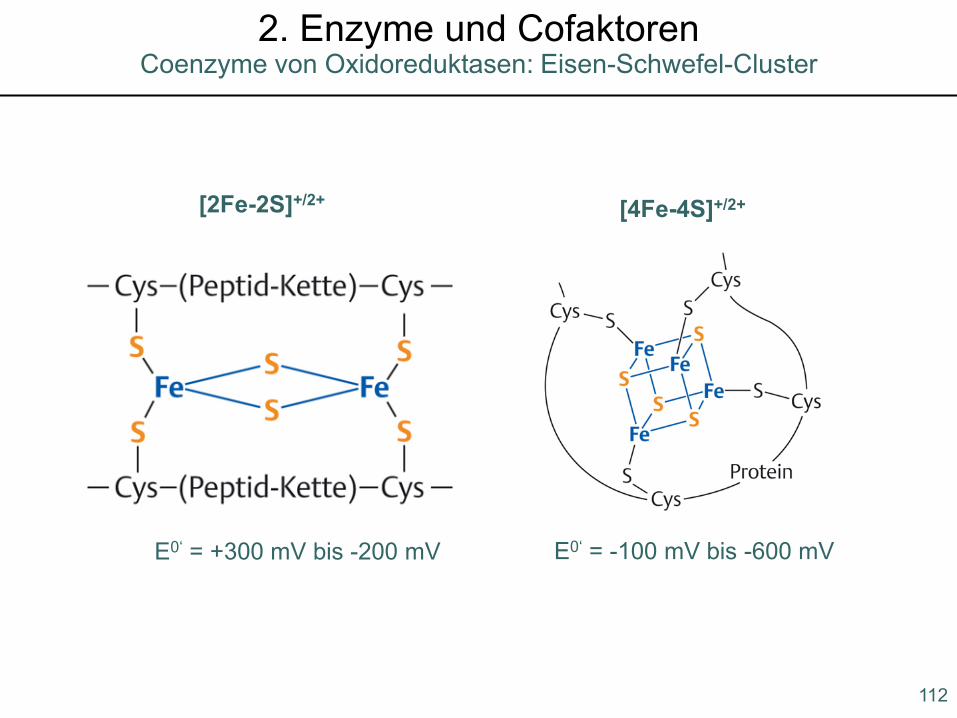
[2Fe-2S]+/2+ [4Fe-4S]+/2+
E0‘ = +300 mV bis -200 mV E0‘ = -100 mV bis -600 mV
112
Coenzyme von Oxidoreduktasen: Eisen-Schwefel-Cluster2. Enzyme und Cofaktoren
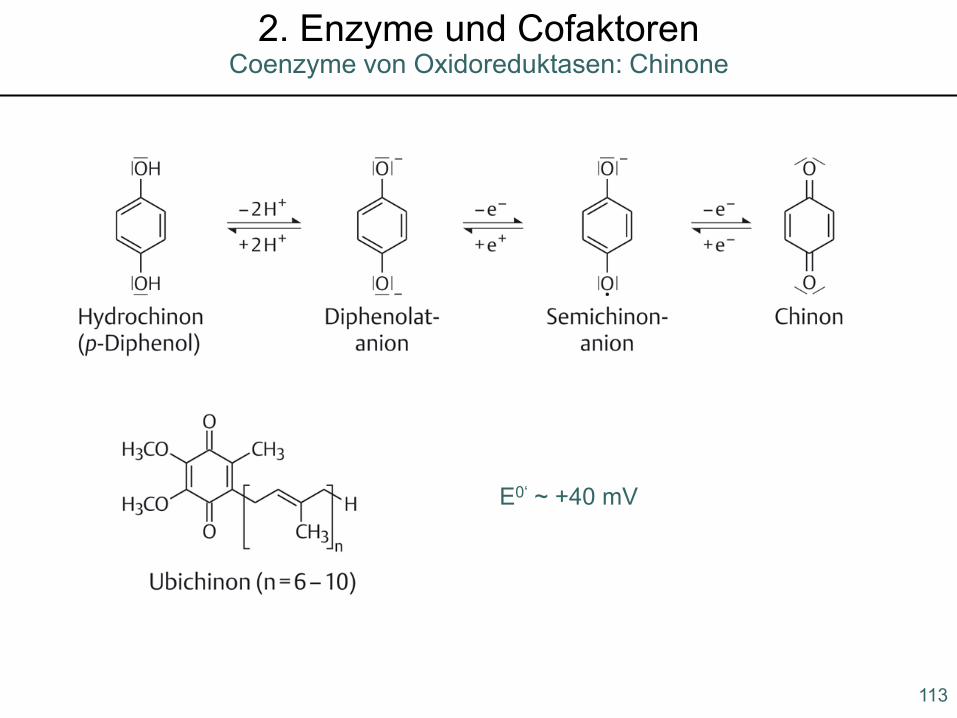
E0‘ ~ +40 mV
113
Coenzyme von Oxidoreduktasen: Chinone2. Enzyme und Cofaktoren
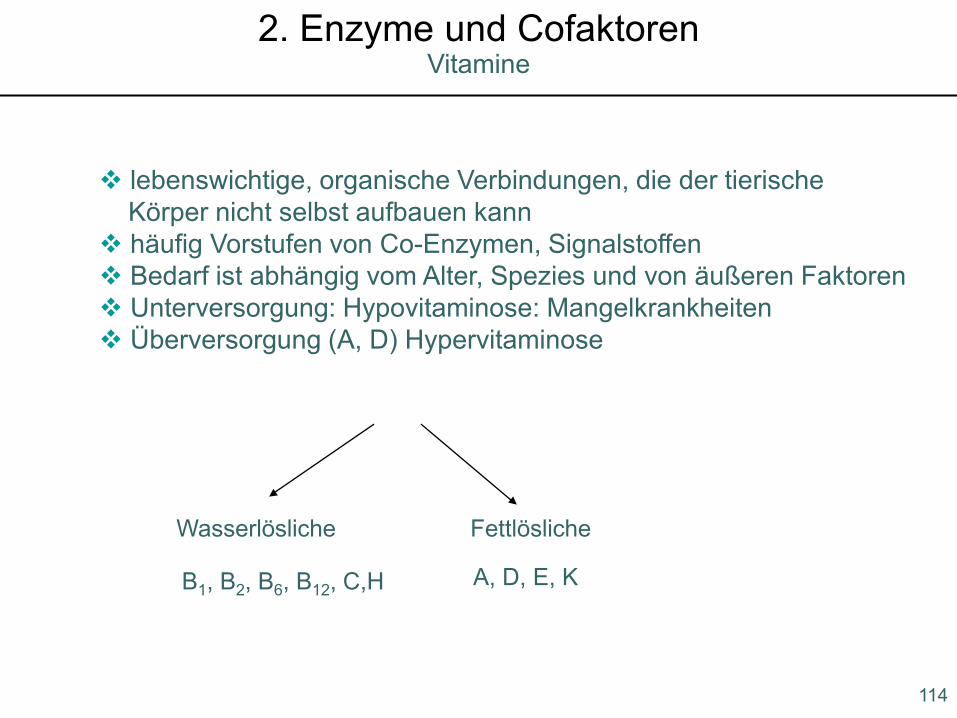
lebenswichtige, organische Verbindungen, die der tierischeKörper nicht selbst aufbauen kann
häufig Vorstufen von Co-Enzymen, Signalstoffen Bedarf ist abhängig vom Alter, Spezies und von äußeren Faktoren Unterversorgung: Hypovitaminose: Mangelkrankheiten Überversorgung (A, D) Hypervitaminose
Wasserlösliche Fettlösliche
B1, B2, B6, B12, C,H A, D, E, K
114
Vitamine2. Enzyme und Cofaktoren
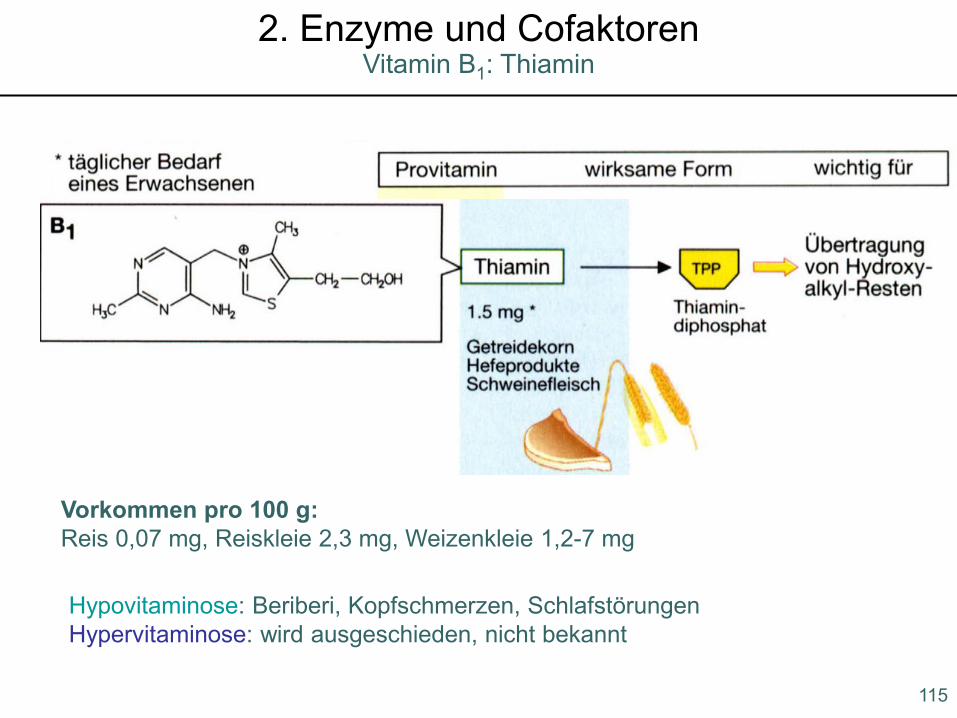
Hypovitaminose: Beriberi, Kopfschmerzen, SchlafstörungenHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Reis 0,07 mg, Reiskleie 2,3 mg, Weizenkleie 1,2-7 mg
115
Vitamin B1: Thiamin2. Enzyme und Cofaktoren
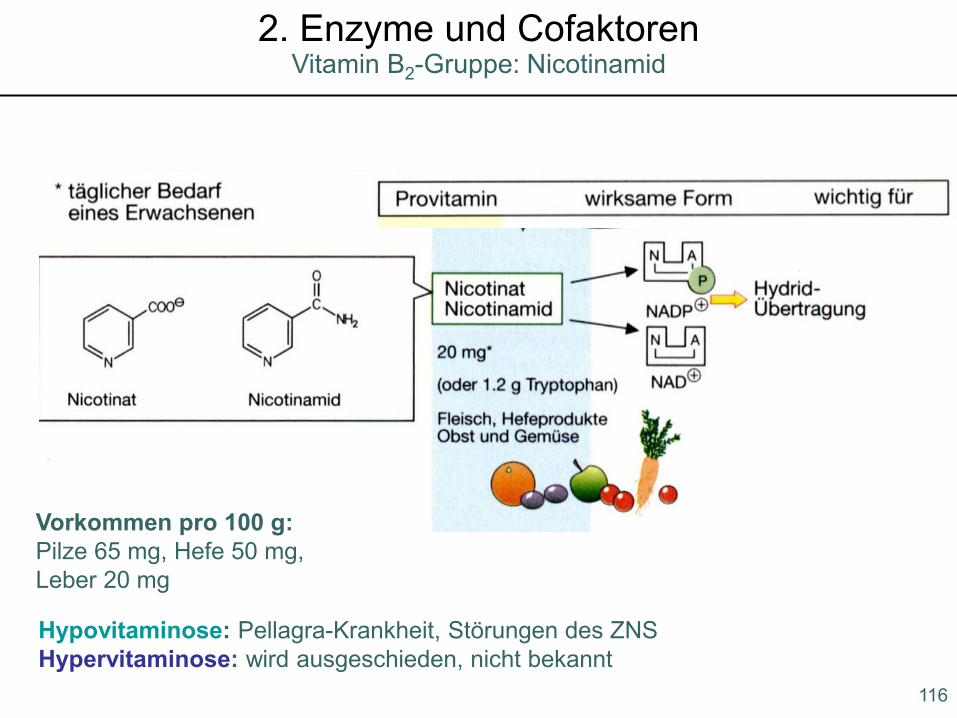
Hypovitaminose: Pellagra-Krankheit, Störungen des ZNSHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g: Pilze 65 mg, Hefe 50 mg, Leber 20 mg
116
Vitamin B2-Gruppe: Nicotinamid2. Enzyme und Cofaktoren
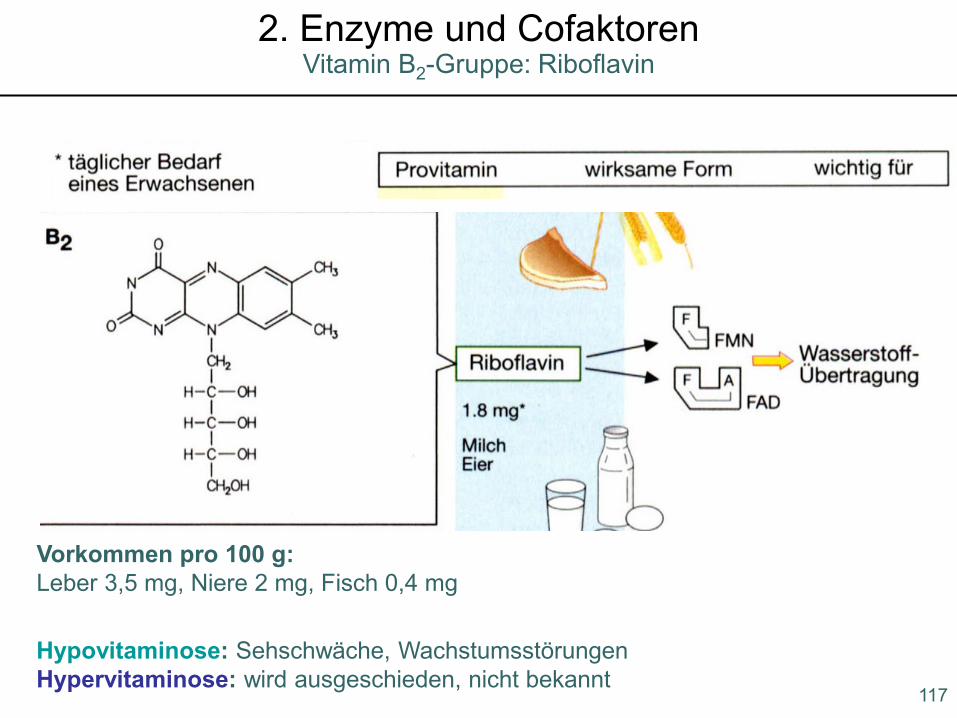
Hypovitaminose: Sehschwäche, WachstumsstörungenHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Leber 3,5 mg, Niere 2 mg, Fisch 0,4 mg
117
Vitamin B2-Gruppe: Riboflavin2. Enzyme und Cofaktoren
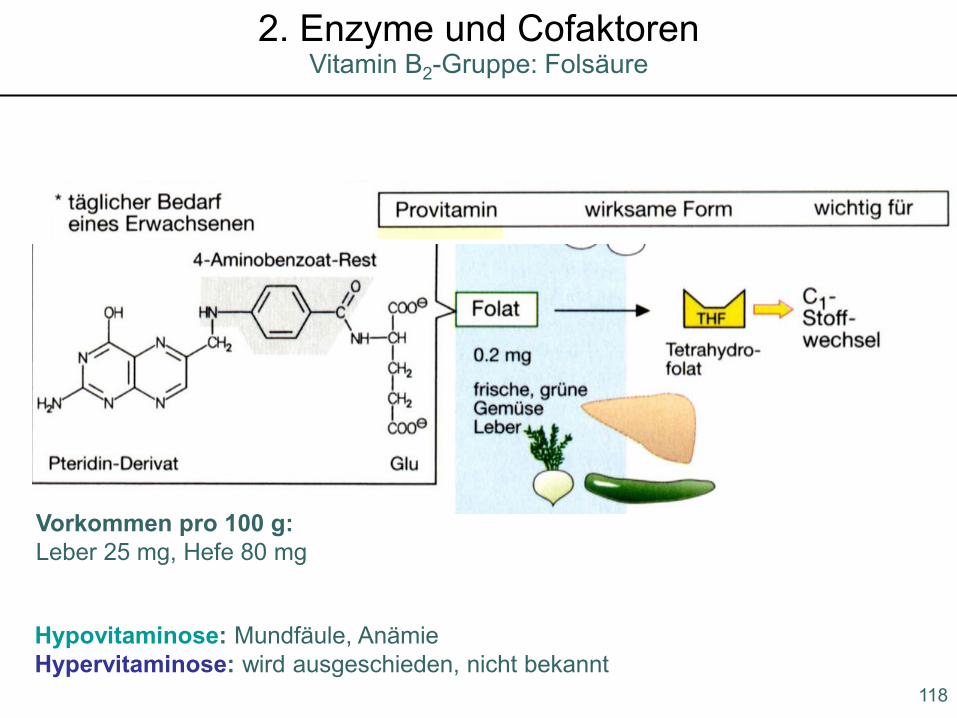
Hypovitaminose: Mundfäule, AnämieHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Leber 25 mg, Hefe 80 mg
118
Vitamin B2-Gruppe: Folsäure2. Enzyme und Cofaktoren
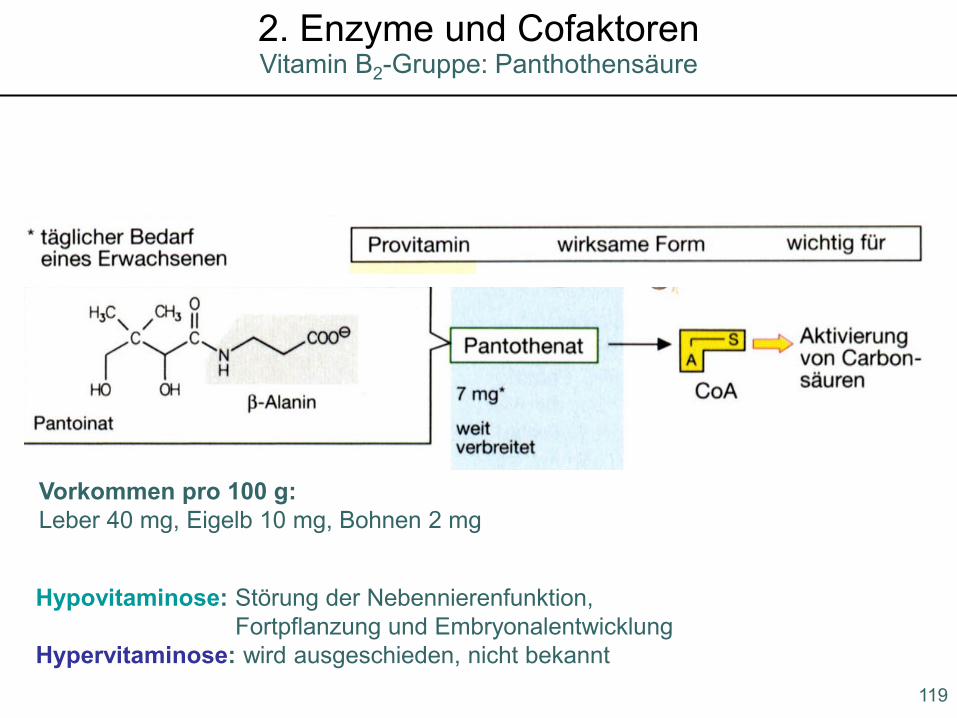
Hypovitaminose: Störung der Nebennierenfunktion, Fortpflanzung und Embryonalentwicklung
Hypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Leber 40 mg, Eigelb 10 mg, Bohnen 2 mg
119
Vitamin B2-Gruppe: Panthothensäure2. Enzyme und Cofaktoren
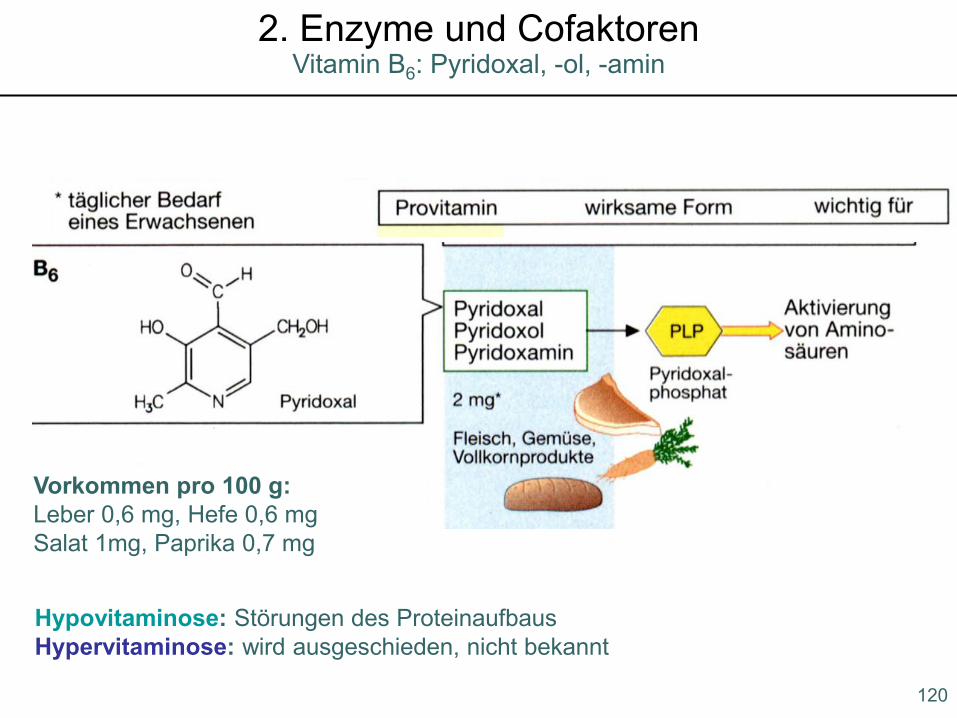
Hypovitaminose: Störungen des ProteinaufbausHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g: Leber 0,6 mg, Hefe 0,6 mgSalat 1mg, Paprika 0,7 mg
120
Vitamin B6: Pyridoxal, -ol, -amin2. Enzyme und Cofaktoren
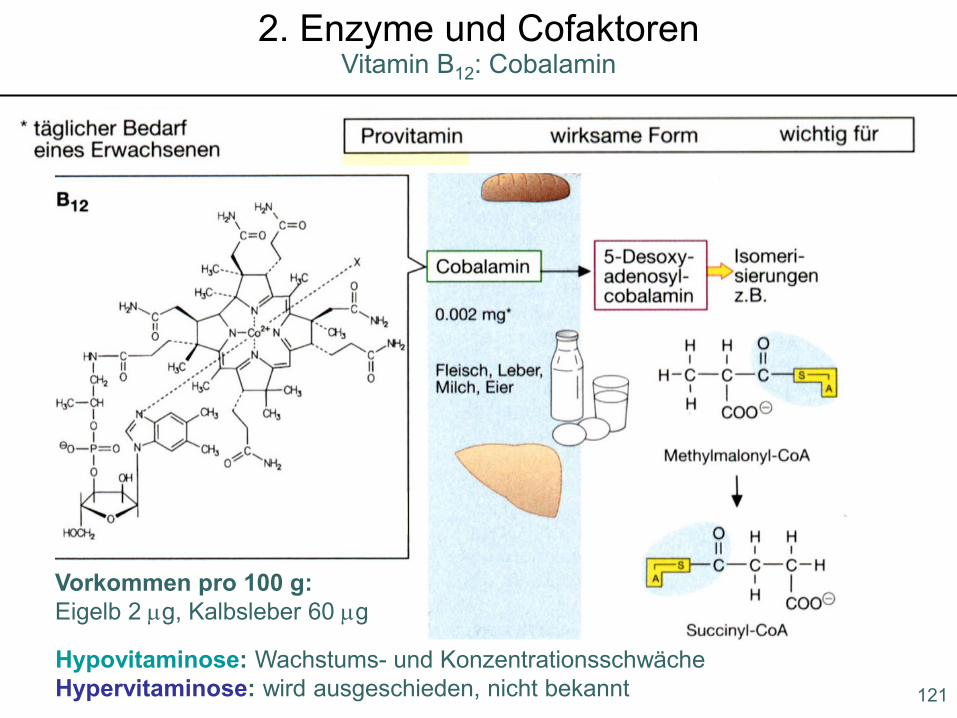
Hypovitaminose: Wachstums- und KonzentrationsschwächeHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Eigelb 2 µg, Kalbsleber 60 µg
121
Vitamin B12: Cobalamin2. Enzyme und Cofaktoren
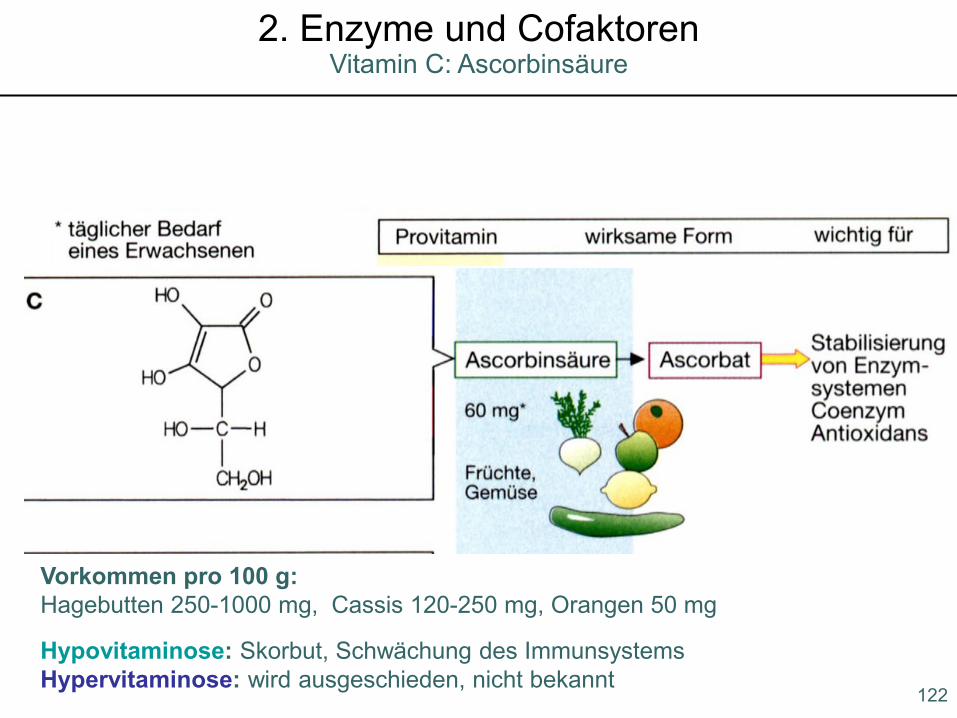
Hypovitaminose: Skorbut, Schwächung des ImmunsystemsHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Hagebutten 250-1000 mg, Cassis 120-250 mg, Orangen 50 mg
122
Vitamin C: Ascorbinsäure2. Enzyme und Cofaktoren
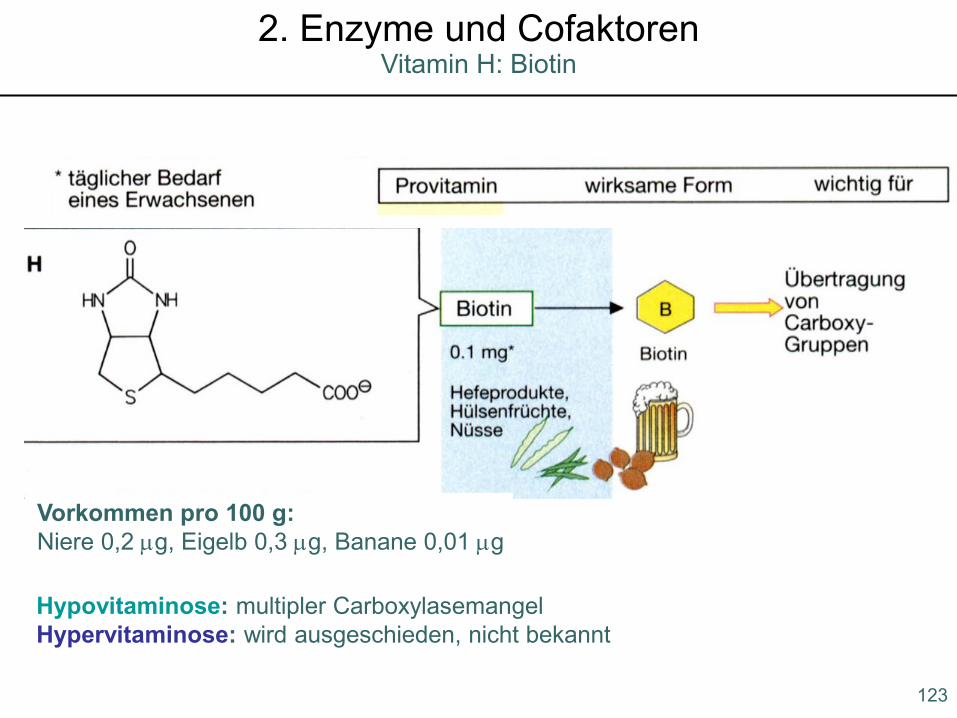
Hypovitaminose: multipler CarboxylasemangelHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Niere 0,2 µg, Eigelb 0,3 µg, Banane 0,01 µg
123
Vitamin H: Biotin2. Enzyme und Cofaktoren
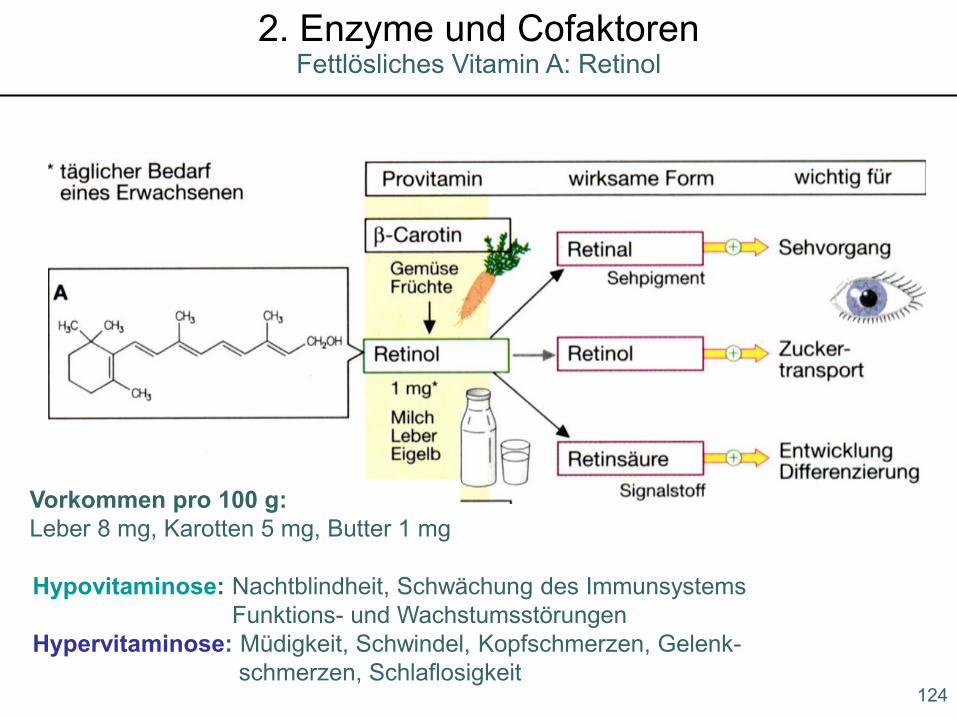
Hypovitaminose: Nachtblindheit, Schwächung des ImmunsystemsFunktions- und Wachstumsstörungen
Hypervitaminose: Müdigkeit, Schwindel, Kopfschmerzen, Gelenk-schmerzen, Schlaflosigkeit
Vorkommen pro 100 g:Leber 8 mg, Karotten 5 mg, Butter 1 mg
124
Fettlösliches Vitamin A: Retinol2. Enzyme und Cofaktoren
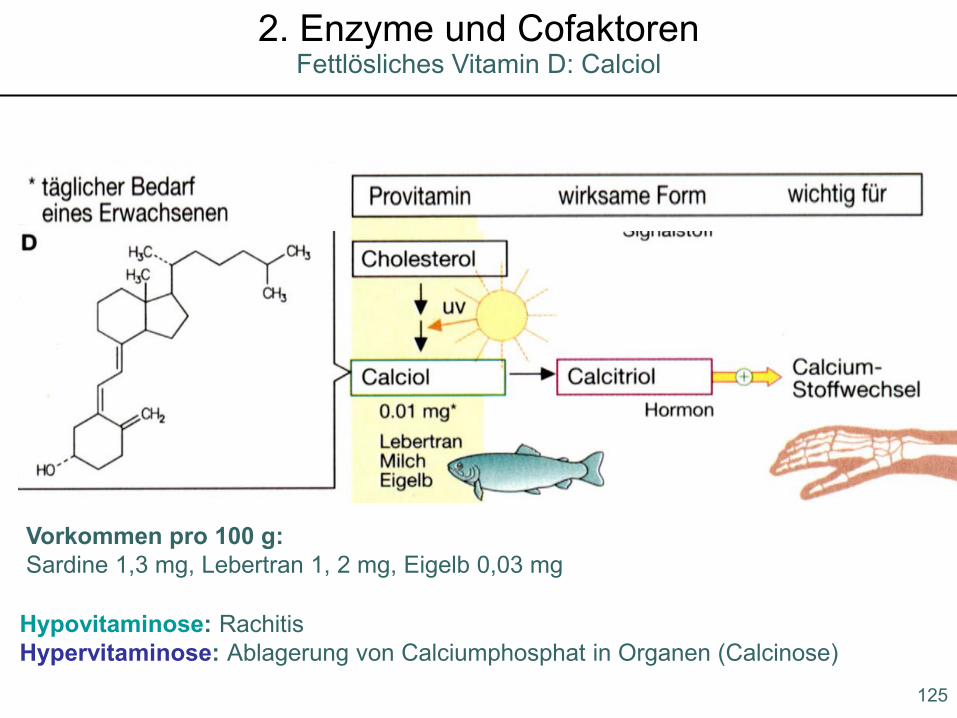
Hypovitaminose: RachitisHypervitaminose: Ablagerung von Calciumphosphat in Organen (Calcinose)
Vorkommen pro 100 g:Sardine 1,3 mg, Lebertran 1, 2 mg, Eigelb 0,03 mg
125
Fettlösliches Vitamin D: Calciol2. Enzyme und Cofaktoren
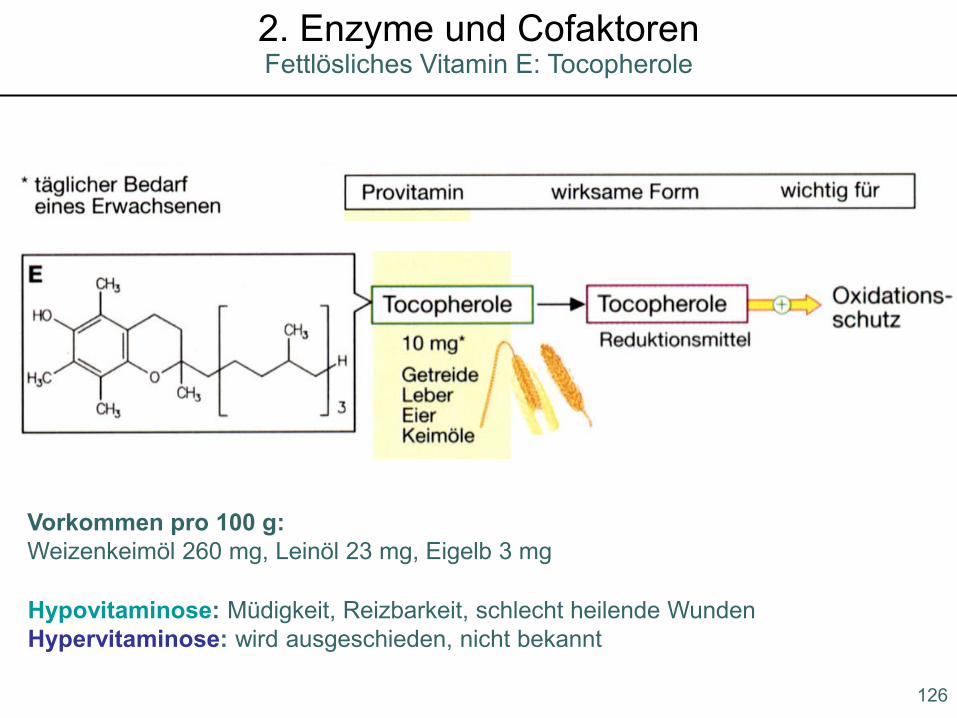
Hypovitaminose: Müdigkeit, Reizbarkeit, schlecht heilende WundenHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Weizenkeimöl 260 mg, Leinöl 23 mg, Eigelb 3 mg
126
Fettlösliches Vitamin E: Tocopherole2. Enzyme und Cofaktoren
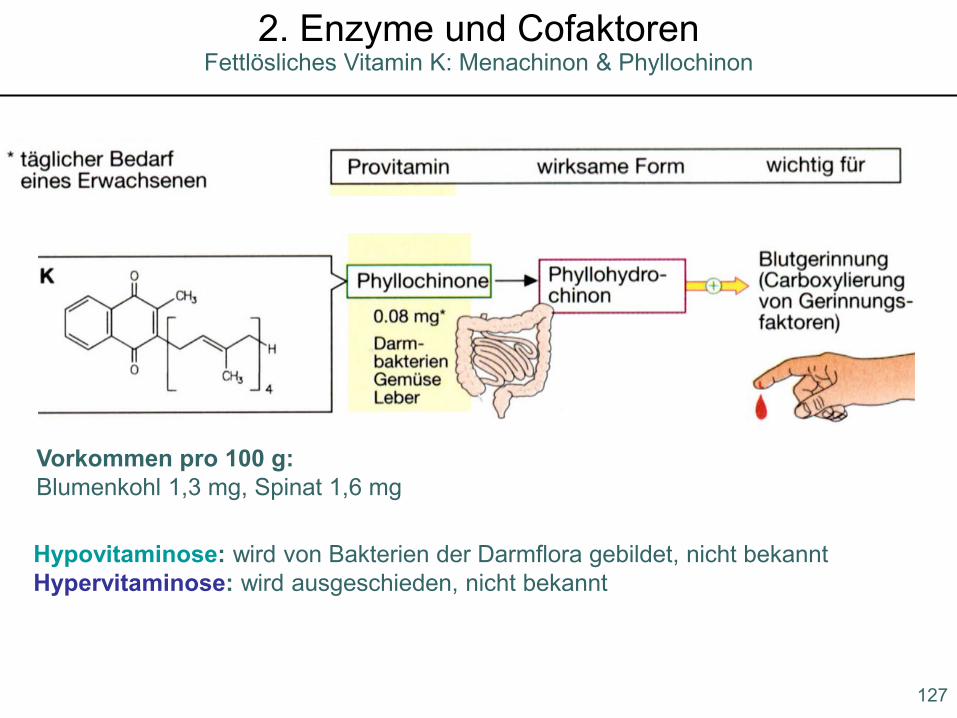
Hypovitaminose: wird von Bakterien der Darmflora gebildet, nicht bekanntHypervitaminose: wird ausgeschieden, nicht bekannt
Vorkommen pro 100 g:Blumenkohl 1,3 mg, Spinat 1,6 mg
127
Fettlösliches Vitamin K: Menachinon & Phyllochinon2. Enzyme und Cofaktoren

• Enzyme (fast immer Proteine) werden in sechs Klassen eingeteilt.
• Sie beschleunigen als Biokatalysatoren biochemische Reaktionen durch Herabsetzung der Aktivierungsenergie.
• Enzyme sind reaktions- und substratspezifisch.
• Sie haben ein Temperatur- und pH-Optimum.
• Ein Modell zur kinetischen Beschreibung einfacher Enzymreaktionen ist die Michaelis-Menten-Theorie.
• Einige Enzyme katalysieren Reaktionen mithilfe von Cofaktoren. Zu diesen gehören organische Coenzyme, von denen viele aus Vitaminen gebildet werden.
• Wichtige katalytische Mechanismen sind Säure/Base-Katalyse, kovalente Katalyse und Metallionenkatalyse.
• Ein besonders wichtiger Mechanismus der enzymvermittelten Katalyse ist die Stabilisierung des Übergangszustands.
• Enzyme sind hemmbar (reversibel oder irreversibel).128
Zusammenfassung2. Enzyme und Cofaktoren