Embed Size (px)
Citation preview

Aus dem Friedrich-Loeffler-Institut für Medizinische Mikrobiologie
(Direktor: Univ. - Prof. Dr. med. Ivo Steinmetz)
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
Intrazelluläres Überleben von Staphylococcus aureus in Makrophagen
Inaugural – Dissertation
zur Erlangung des akademischen Grades
Doktor der Medizin (Dr. med.)
der Universitätsmedizin
der Ernst-Moritz-Arndt-Universität
Greifswald
2016
vorgelegt von
Cäcilia Renner
geb. am 24.11.1982
in Dresden

Dekan: Prof. Dr. rer. nat. Max P. Baur
1. Gutachter: Prof. Dr. I. Steinmetz
2. Gutachter: Prof. Dr. J. Buer
Ort der Disputation: Greifswald
Tag der Disputation: 7. Juni 2017

Gewidmet
meinen Eltern und Geschwistern

I
Inhaltsverzeichnis
INHALTSVERZEICHNIS………………………………………………………………………………………....I
ABKÜRZUNGSVERZEICHNIS…………………………………………………………………………………. IV
1 EINLEITUNG ........................................................................................................................ 1
1.1 Staphylokokken ........................................................................................................... 1
1.1.1 Allgemeines und klinische Bedeutung.......................................................................... 1
1.1.2 Virulenzfaktoren von Staphylokokken .......................................................................... 3
1.2 Makrophagen ............................................................................................................... 6
1.2.1 Die Rolle von Makrophagen bei der Abwehr bakterieller Erreger .................................... 6
1.2.2 Oxidative Effektormechanismen von Makrophagen....................................................... 7
1.3 Intrazelluläres Überleben von Staphylokokken ............................................................ 8
1.3.1 Staphylokokken in nicht-professionell phagozytierenden Zellen ..................................... 9
1.3.2 Staphylokokken und Makrophagen.............................................................................10
1.4 Zielstellung der Arbeit ................................................................................................13
2 MATERIAL UND METHODEN ..............................................................................................14
2.1 Material .......................................................................................................................14
2.1.1 Geräte .....................................................................................................................14
2.1.2 Gebrauchsmaterialien ...............................................................................................14
2.1.3 Substanzen ..............................................................................................................15
2.1.4 Lösungen, Puffer und Medien ....................................................................................16
2.1.5 Zellkulturmedien .......................................................................................................16
2.1.6 Bakterienkultur .........................................................................................................17
2.1.7 Software ..................................................................................................................17
2.2 Bakterien ....................................................................................................................17
2.2.1 Bakterienstämme......................................................................................................17
2.2.2 Anlegen der Bakterienstocks .....................................................................................18
2.2.3 Anlage von Bakterienkulturen und Bestimmung der Keimzahl.......................................18
2.3 Makrophagen ..............................................................................................................18
2.3.1 RAW-Makrophagen ..................................................................................................19
2.3.2 Murine Knochenmarkmakrophagen ............................................................................20

II
2.4 Staphylococcus aureus - Infektionsversuche mit murinen Makrophagen: Bakterizidie-
und Phagozytoseassays .................................................................................................22
2.4.1 Makrophagen-Bakterizidie-Assay mit S. aureus ...........................................................22
2.4.2 Makrophagen-Bakterizidie-Assay mit S. epidermidis und S. carnosus ...........................23
2.4.3 Makrophagen-Bakterizidie-Assay mit S. aureus unter IFN-Stimulation .........................23
2.4.4 Makrophagen-Bakterizidie-Assay mit S. aureus unter NO-Inhibition mit Aminoguanidin ..24
2.4.5 Makrophagen-Phagozytose-Assay mit S. aureus unter Fibronektin-freien Bedingungen..24
2.5 NO-Assay ...................................................................................................................24
2.6 Statistik ......................................................................................................................25
3 ERGEBNISSE......................................................................................................................26
3.1 Die Charakterisierung der Phagozytose von Staphylokokken durch murine Makrophagen
...................................................................................................................................26
3.1.1 Die Phagozytose von S. aureus durch murine Makrophagen ........................................26
3.1.2 Der Einfluss von IFN-γ auf die Phagozytose von S. aureus ..........................................28
3.1.3 Die Phagozytose von Koagulase-negativen Staphylokokken ........................................29
3.1.4 Die Rolle von Fibronektin-bindendem Protein A bei der Phagozytose – Untersuchungen an S. carnosus .........................................................................................................30
3.1.5 Die Rolle von Fibronektin bei der Phagozytose von S. aureus Newman.........................32
3.2 Die Charakterisierung des intrazellulären Überlebens von Staphylokokken in murinen
Makrophagen ..................................................................................................................33
3.2.1 Intrazelluläres Überleben von S. aureus Newman in RAW-Makrophagen ......................33
3.2.2 S. aureus Newman in primären Knochenmarkmakrophagen von BALB/c- und C57BL/6-Mäusen ...................................................................................................................35
3.2.3 Vergleich des intrazellulären Überlebens von S. aureus Newman und S. aureus RN 6390 . ...............................................................................................................................37
3.2.4 Vergleich des intrazellulären Überlebens von S. aureus Newman und S. epidermidis ATCC 12228 ............................................................................................................39
3.3 Makrophagen-vermittelte Effektormechanismen bei S. aureus-Infektionen .................41
3.3.1 Die Rolle von Stickoxid (NO)......................................................................................41
3.3.2 Die Rolle von reaktiven Sauerstoffradikalen am Beispiel von C57Bl/6 gp91 phox-/- - Makrophagen ...........................................................................................................46
3.4 Der Einfluss der globalen regulatorischen Systeme accessory-gene-regulator (agr) und
staphylococcal-accessory-element (sae) von S.aureus ...................................................47
3.4.1 Die Rolle von agr für die Interaktion von S. aureus RN 6390 mit Makrophagen ..............48
3.4.2 Die Rolle von sae für die Interaktion von S. aureus Newman mit Makrophagen ..............51

III
4 DISKUSSION.......................................................................................................................55
4.1 Phagozytose und intrazelluläres Überleben von Staphylokokken in Makrophagen .....55
4.1.1 Die bakterielle Regulation der Phagozytose in Makrophagen........................................55
4.1.2 Die bakterielle Regulation des intrazellulären Überlebens in Makrophagen ....................57
4.2 Die Bedeutung von FnBPA und Fn für die Internalisierung von S. aureus...................60
4.3 Die Rolle von iNOS und NADPH-Oxidase in der Abwehr von intrazellulärem S. aureus..
...................................................................................................................................63
4.4 S. aureus - ein fakultativ intrazelluläres Bakterium? ...................................................65
5 ZUSAMMENFASSUNG ........................................................................................................68
6 LITERATURVERZEICHNIS ..................................................................................................70
7 PUBLIKATION.....................................................................................................................92
8 DANKSAGUNG ...................................................................................................................93

IV
Abkürzungsverzeichnis
Abb. Abbildung
AG Aminoguanidin agr Accessory gene regulator AHT Anhydrotetracyclin
ATCC The American Type Culture Collection AtlE Staphylococcus epidermidis Major Autolysin BMM Bone marrow derived macrophages
BSA Bovines Serum-Albumin CFU Colony Forming Unit DMEM Dulbecco´s Modified Eagle Medium
Eap Extracellular Adherence Protein FAK Focal Adhesion Kinase Fc Functional chain
Fg Fibrinogen Fn Fibronektin
FnbP Fibronektin-bindendes Protein Genta Gentamycin GFP Green fluorescent protein
GM-CSF Granulozyten-Makrophagen- Kolonie-stimulierender Faktor
h Stunden
HEK Human Embryonic Kidney Cells hla alpha-Hämolysin hMDM Human Monocyte derived Macrophages
HWI Harnwegsinfektion IFN-γ Interferon-gamma
iNOS Inducible Nitrit Oxide Synthase JNK c-Jun-N-terminale Kinase KNS Koagulase-negative Staphylokokken
Lyso Lysostaphin ml Milliliter µg Mikrogramm
µl Microliter MOI Multiplicity of Infection (Infektionsverhältnis) MRSA Methicillin-resistente S. aureus-Stämme
MSCRAMM Microbial Surface Components Recognizing Adhesive Matrix Molecules
NADPH Nicotinamide adenine dinucleotide phosphate
NK-Zellen Natürliche-Killer-Zellen NO Stickoxid
OD Optische Dichte PBS Phosphatpuffer PFA Paraformaldehyd
p.i. post infectionem PMN Polymorphnuclear Leucocytes PRR Pattern Recognition Receptor
PTK Protein Tyrosin Kinase ROS Reactive Oxygen Species

V
rpm rounds per minute
RPMI Roswell Park Memorial Institute RT Raumtemperatur
s Sekunden sae S. aureus exoprotein expression sar Staphylococcal accessory regulator
sigB alternative Faktor Sigma B SSSS Staphylococcal Scaled Skin Syndrome S. aureus Staphylococcus aureus
t Zeit TCS Two Component Systems TLR Toll-Like-Receptor
TSB Tryptone Soya Broth u.a. unter anderem usw. und so weiter
WT Wildtyp

1
1 EINLEITUNG
1.1 Staphylokokken
1.1.1 Allgemeines und klinische Bedeutung
Die zur Familie der Micrococcaceae zählenden Staphylokokken sind unbewegliche Gram-positive,
Katalase-positive, fakultativ anaerobe Kokken. Ihre Größe beträgt etwa 1 m und sie ordnen sich meist
in Haufen oder in einer Traube (griech. staphylos) an.
Die für die Humanmedizin wichtigste Spezies ist Staphylococcus aureus. Er gehört weltweit zu
den häufigsten Pathogenen, die sowohl innerhalb als auch außerhalb des Krankenhauses Infektionen
hervorrufen können. S. aureus besiedelt bei etwa 20 % der Bevölkerung asymptomatisch Haut- oder
Schleimhäute mit der Nasenschleimhaut als Hauptreservoir (Eiff et al. 2001a; Bera et al. 2006). Die meist
harmlose Kolonisation bildet einen Risikofaktor für eine invasive lokale oder disseminierte S. aureus-
Infektion, die im Extremfall lebensbedrohlich sein kann (Wertheim, Heiman F L et al. 2005; van Belkum
et al. 2009).
Wichtige weitere Gruppe neben S. aureus sind die Koagulase-negativen Staphylokokken (KNS),
die auch in dieser Arbeit betrachtet werden sollen. KNS besitzen im Gegensatz zu S. aureus keine
Plasmakoagulase. Sie besiedeln als natürliche Kommensalen die gesamte Haut und Schleimhaut des
Menschen und sind klassische opportunistische Pathogene. KNS verfügen über ein geringeres Potential
an Pathogenität als S. aureus, wurden jedoch in den letzten Jahrzehnten als wichtige Erreger
nosokomialer Infektionen erkannt. Bisher wurden 32 unterschiedliche Spezies beschrieben, von denen
einige in mikrobiologischen Laboren zu den am häufigsten isolierten Erreger gehören (Eiff et al. 2002; Eiff
et al. 2001b; Keim et al. 2011). Der bedeutendste Vertreter, der 70-80% der KNS-Infektionen ausmacht,
ist Staphylococcus epidermidis (Schoenfelder et al. 2010; Ziebuhr et al. 2006).
1.1.1.1 Klinische Bedeutung von S. aureus
Erster Schritt der S. aureus - Infektion sind meist kleine Haut, - oder Schleimhautläsionen. Diese können
in eine lokale, mit Eiterbildung einhergehende Haut.- oder Weichteilinfektion übergehen, wie z. B.
Wundinfektion, Abszess, Furunkel, Karbunkel. S. aureus kann aber jedes Gewebe infizieren, wenn er in
tiefere Schichten gelangt, oder Zugang zur Blutzirkulation gewinnt (Krishna und Miller 2012). Das
Krankheitsspektrum reicht daher von lokalen bis zu systemischen lebensbedrohlichen Infektionen. Dazu
zählen typischerweise Pneumonie und Sepsis, Osteomyelitis, Spondylodiszitis, Endokarditis. So
verursacht S. aureus neben E. coli zu gleichen Teilen etwa ein Drittel aller Sepsen bei hospitalisierten

EINLEITUNG 2
Patienten (Foster 2005, 2009; Foster et al. 2014). Weiterhin ist S. aureus Erreger spezifischer
Infektionskrankheiten, wie der Impetigo contagiosa, Rhinosinusitis, Otitis media, sowie der Mastitis
puerperalis. Neben den aufgeführten invasiven Infektionen bilden Toxikosen eine weitere Gruppe
S. aureus-verursachter Erkrankungen. Dazu werden das Toxische-Schock-Syndrom (Toxin-1),
Staphylococcal Scaled Skin Syndrome SSSS (Exfoliatintoxin) und Lebensmittelvergiftungen
(Enterotoxine) gezählt, die durch die lokale oder systemische Wirkung der spezifischen Toxine ausgelöst
werden.
1.1.1.2 Klinische Bedeutung von Koagulase-negative Staphylokokken
Koagulase-negative Staphylokokken stellen aufgrund ihrer Fähigkeit der Biofilmbildung die häufigste
Ursache Fremdkörper-assoziierter Infektionen dar (Otto 2012). Durch die Bindung von bakteriellen
Oberflächenproteinen an extrazelluläre Matrixproteine, die die körperfremden Materialien überziehen,
können diese Biofilme entstehen und Gelenkprothesen, Katheter und Gefäßprothesen überziehen.
Bakterien, die in diesem Biofilm leben, sind vor Antibiotika gut geschützt und können daher schlecht
eliminiert werden. (Longauerova 2006; da Silva Domingues, Joana F et al. 2013). Oft ist ausschließlich
nur durch eine weitere chirurgische Intervention eine erfolgreiche Behandlung zu erzielen (Khalil et al.
2007). Biofilme bilden außerdem Foci für systemische S. epidermidis-Infektionen, bevorzugt bei
immunsupprimierten Patienten. Zu den verursachten Krankheitsbildern zählen in diesem Zusammenhang
die Osteomyelitis des Prothesen-umgebenden Knochens, Bakteriämie, Sepsis, sowie Endophtalmitis und
Endokarditis, insbesondere bei künstlichen Herzklappen (Gotz 2002; Rogers et al. 2009; Otto 2014).
1.1.1.3 Therapie von Staphylokokken-Infektionen
Die Behandlung S. aureus-vermittelter Infektionen erfolgt durch penicillinasefeste Penicilline
(Flucloxacillin), da 70-80 % der Stämme Penicillinase bilden. Allerdings nimmt die Resistenzentwicklung
durch Mutation und eine hohe genetische Flexibiltät des Erregers zu und erschwert daher zunehmend
die Behandlung sowohl von S. aureus-Infektionen, als auch KNS-Infektionen (Casey et al. 2007).
Besonders im stationären Bereich stellen Methicillin-resistente Stämme (MRSA, MRSE) mit Resistenz
gegenüber Penicillinase ein ernstzunehmendes Problem dar und machen S. aureus zu einem der
häufigsten Erreger nosokomialer Infektionen. Dies gilt zunehmend auch für S. epidermidis, der sich bei
verstärktem Einsatz von körperfremder Materialien, oder auch immunsupprimierenden Medikamenten zu
einem Haupterreger nosokomialer Infektionen entwickelt hat (Casey et al. 2007; Martins und Cunha,
Maria de Lourdes R S 2007).

EINLEITUNG 3
Die wichtigste Maßnahme zur Vermeidung S. aureus-bedingter Infektionskrankheiten im
Krankenhaus ist neben der Händedesinfektion die Sanierung von Keimträgern durch intranasale
Antibiotikagabe (Wertheim, Heiman F L et al. 2005). Weiterer wichtiger Bestandteil der Prophylaxe ist die
genauere Kenntnis wirtsspezifischer Faktoren, die den Menschen für eine solche Infektion empfänglich
machen und folglich vermieden, oder behandelt werden können (van Belkum et al. 2009).
1.1.2 Virulenzfaktoren von Staphylokokken
1.1.2.1 Virulenzfaktoren von S. aureus
S. aureus verfügt über ein weites Spektrum zahlreich beschriebener Virulenzfaktoren, die seine
Pathogenität ausmachen. Man unterscheidet dabei einerseits zellwandassoziierte Proteine, die an das
Bakterium gebunden sind von solchen Proteinen, die sezerniert werden (sog. Exoproteine, Exoenzyme
und Toxine) (Foster 2005; Asad und Opal 2008; Zecconi und Scali 2013).
Zu den zellwandassoziierten Komponenten zählt die Gruppe der MSCRAMM´s (microbial surface
components recognizing adhesive matrix proteins). Diese umfasst eine Reihe Zellwand -assoziierter
Proteine, z. B. FnbP A und B, Clumpingfactor A und B, das Extrazelluläre Adherence Protein (Eap),
Protein A, oder Kollagenbindendes Protein, die bestimmte extrazelluläre Matrixproteine wie Fibronektin,
Fibrinogen, Kollagen usw. erkennen und an sie binden. S. aureus wird dadurch die Fähigkeit der
Adhärenz an wirtseigene Strukturen vermittelt (Speziale et al. 2009; Foster et al. 2014).
Zu den andererseits von S. aureus sezernierten Proteinen gehören u. a. Koagulase,
Staphylokinase, Katalase sowie verschiedene Leukozidine. Wichtig sind die bereits erwähnten
Hämolysine (α, β, γ, δ), die über die Eigenschaft verfügen Membranen zu lysieren. Außerdem produziert
er Enzyme, wie die Hyaluronidase, Proteasen und Lipasen, die Wirtsstrukturen abbauen können.
S. aureus sezerniert auch eine Reihe von Toxinen (Superantigenen), wie das Toxische-Schock-Syndrom-
Toxin, Exfoliatintoxine und die Enterotoxine (Lowy 1998; Foster 2005; Zecconi und Scali 2013).
Mit Ausnahme der spezifischen Krankheitsbilder, die durch die Wirkung einzelner Toxine
verursacht werden, entstehen S. aureus-Infektionen immer durch ein abgestimmtes Zusammenspiel
verschiedener Virulenzfaktoren (Fournier und Philpott 2005).
1.1.2.2 Virulenzfaktoren von Koagulase-negativen Staphylokokken
Bei vergleichenden Genomanalysen zwischen S. aureus und S. epidermidis stellte sich heraus, dass
Gene von zahlreichen Exotoxinen, die von S. aureus exprimiert werden, bei S. epidermidis fehlen. Dies
mag dafür sprechen, dass S. aureus ein größeres Spektrum an Virulenzfaktoren zur Verfügung steht und
somit aggressivere Möglichkeiten bestehen, der Wirtsabwehr entgegenzuwirken (Gill et al. 2005). Die

EINLEITUNG 4
Mechanismen, über die S. epidermidis verfügt, um im Wirt zu überleben und eine Infektion zu etablieren
sind demgegenüber generell passiver. Die Pathogenität besteht im Wesentlichen in der Fähigkeit der
Biofilm-Bildung (Gotz 2002; Eiff et al. 2002; Mack et al. 2007; Zecconi und Scali 2013; Foster 2005).
Dazu wurden S. epidermidis surface (Ses)-Proteine (u.a. Aap, Bhp, SdrF and SdrG) beschrieben,
die strukturelle Ähnlichkeit mit den bekannten zellwandassoziierten Proteinen von S. aureus aufweisen.
Diese binden Wirtsfaktoren wie Kollagen und Fibronektin und vermitteln somit die Adhärenz an die
Fremdkörper-Oberflächen (Bowden et al. 2005; Arrecubieta et al. 2007). Es wurde darüber hinaus
beschrieben, dass S. epidermidis, ebenso wie S. aureus, FNBP A und B auf seiner Oberfläche exprimiert
und auch im gleichen Maß Fibronektin binden kann (Shinji et al. 1998).
In jüngster Zeit wurden Phenol soluble Modulins (PSM´s) als eine Familie zytolytischer
inflammatorischer Toxine beschrieben, die im Falle von KNS eine entscheidende Rolle in der
Biofilmbildung spielen (Otto 2014, 2012; Peschel und Otto 2013). Es handelt sich um Proteine, die auch
bei S. aureus vorkommen, aber dort mit höheren Expressionsraten und erhöhter zytolytischer Aktivität
(Cheung, Gordon Y C et al. 2010; Otto 2012).
1.1.2.3 Regulation von Virulenzfaktoren
Die Etablierung einer Infektion im Organismus setzt eine koordinierte, auf die Umgebung abgestimmte
Regulation der vielfältigen Virulenzfaktoren voraus. Die Expression von Virulenzfaktoren in S. aureus und
folglich die Synthese von entweder sezernierten oder Zellwand-ständigen Proteinen, wird durch ein
komplexes Zusammenspiel globaler Genregulatoren vermittelt, die so aufeinander abgestimmt werden,
dass sie der Infektionsausbreitung optimal dienen. (Garzoni et al. 2007; Jenkins et al. 2015). Dazu
gehören die Zweikomponentensysteme (TCS) agr, sae, srr, arl und die DNA-bindende SarA-Protein-
Familie, sowie der alternative Faktor sigB (Cheung et al. 2004; Yarwood und Schlievert 2003; Mitchell et
al. 2008).
TCS´s sind so genannte quorum sensing Systeme. Sie ermöglichen dem Erreger eine spezifisch
veränderte Genexpression in Reaktion auf die bestehenden Umweltparameter und folglich eine
kontrollierte Produktion von Proteinen für ein optimales Überleben (Asad und Opal 2008; Cheung et al.
2004; Bronner et al. 2004). Virulenzfaktoren werden somit nur nach Bedarf, vor allem in Abhängigkeit von
der Zelldichte, in bestimmten Phasen des Wachstums gebildet. Die meisten der Zellwand -assoziierten
Proteine werden in der exponentiellen Phase, die sezernierten Proteine vermehrt in der stationären
Wachstumsphase gebildet. Dies vermittelt den Bakterien im Verlauf der Infektion zunächst die Fähigkeit
der Adhärenz an das Wirtszellgewebe und danach die Möglichkeit der Invasion und des intrazellulären

EINLEITUNG 5
Überlebens. Zwei der wichtigsten Regulatorsysteme in der Pathogenese von S. aureus-Infektionen, agr
und sae, sollen im Folgenden genauer vorgestellt werden.
Der globale Regulator agr wird beim Übergang von der exponentiellen in die stationäre
Wachstumsphase der Bakterien durch quorum sensing der Populationsdichte aktiviert. Daraufhin wird die
Synthese Zellwand-assoziierter Proteine (z. B. FnBA, Protein A), wie sie zu Beginn einer S. aureus-
Infektion bevorzugt exprimiert werden, inhibiert und die Produktion von sezernierten Proteine n (z. B. α-
Hämolysin) aktiviert (Novick 2003; Wolz et al. 2000). In-vitro-Studien mit S. aureus zeigten, dass agr-
Mutanten verglichen mit dem Wildtyp in größerem Maß von nicht-professionell phagozytierenden Zellen
internalisiert werden (Wesson et al. 1998). Darüber hinaus wurde auch gesehen, dass die verschiedenen
intrazellulären Wege des Bakteriums, der Wirtsabwehr standzuhalten, mit Defekt im agr-locus nicht mehr
möglich sind. Dazu gehören die Fähigkeit zum Verlassen des Phagosoms und der Übertritt ins Zytosol,
die Unterbindung der phagolysosomalen Verschmelzung, die Apoptoseinduktion der Wirtszelle, oder die
Umgehung der Autophagie (Schnaith et al. 2007; Haslinger-Loffler et al. 2005; Menzies und Kourteva
2000). Entscheidend dabei ist die fehlende Expression von α-Hämolysin in den agr-defizienten Stämmen
(Jarry et al. 2008; Abel et al. 2011; Bantel et al. 2001; Mestre et al. 2010). Aber auch δ- und β-Hämolysin
spielen für die genannten Vorgänge eine Rolle (Giese et al. 2011a). In-vivo zeigten agr-Mutanten eine
attenuierte Virulenz und Zytotoxizität im Vergleich zum Wildtyp (Xiong et al. 2006).
Das Zweikomponentensystem sae ist ein agr-unabhängig arbeitendes regulatorisches System.
Es induziert in der postexponentiellen Wachstumsphase die Synthese von u.a. alpha-Hämolysin (α-
Toxin), beta-Hämolysin, Koagulase, Protein A, Eap und FnBPA und inhibiert dagegen die Transkription
von Kapselpolysaccharid (cap). Wie agr in Reaktion auf die Populationsdichte der Bakterien wirkt,
reguliert sae in Abhängigkeit von bestimmten Stimuli der Umgebung, wie hohe Salzkonzentrationen,
niedriger pH, Glucose, oder Antibiotika in subinhibitorischen Konzentrationen (Rogasch et al. 2006;
Geiger et al. 2008; Harraghy et al. 2005). S. aureus-Stämme mit Mutationen des sae-locus führten in
Endothelzellen in-vitro zu reduzierten Invasionsraten von S. aureus, was auf die verminderte Expression
von fnbA zurückgeführt wurde (Steinhuber et al. 2003). In in-vivo-Modellen von Sepsis bzw. Peritonitis
zeigten sae-Mutanten eine deutlich attenuierte Virulenz im Gesamtorganismus (Watkins et al. 2011;
Voyich et al. 2009).
Die Regulatoren sae und agr wirken also in der Kontrolle einiger Gene entgegengesetzt, so bei
der Expression von FnBP´s (fnbp), Kollagen (coa) und Kapselpolysaccharid (cap), zeigen aber
andererseits gleichsinnige Effekte bei der Expression von α-Toxin (hla), oder Eap (eap), das von beiden
aktiviert wird (Steinhuber et al. 2003; Xiong et al. 2006; Goerke et al. 2001; Harraghy et al. 2005).

EINLEITUNG 6
1.2 Makrophagen
1.2.1 Die Rolle von Makrophagen bei der Abwehr bakterieller Erreger
Makrophagen sind neben den neutrophilen Granulozyten wichtige Zellen der angeborenen Abwehr. Ihre
zentrale Funktion ist die Erkennung von Erregern, deren Phagozytose und Elimination. Es sind langlebige
Zellen, die sich in allen Geweben des Körpers befinden, besonders aber in für Infektionen empfänglichen
Stellen wie Lunge und Darm. Die im Blut zirkulierenden Monozyten sind ihre Vorläuferzellen.
Makrophagen sind als ortsständige Zellen in der Regel die ersten Immunzellen, mit denen bakterielle
Erreger in Kontakt treten. Die im Gegensatz dazu kurzlebigen, nicht im gesunden Gewebe
vorkommenden Neutrophilen werden dann durch sie angelockt (Alberts 2011).
Die Erregererkennung ist der erste Schritt der Wirtsabwehr. Makrophagen erkennen bakterielle
Erreger dabei anhand konservierter mikrobieller Strukturen, wie beispielsweise Lipopolysaccharide und
Flagellin (Pathogen Associate Molecular Patterns, PAMP´s). Als Erkennungsrezeptoren dienen neben
verschiedenen anderen vor allem die so genannten Toll-Like-Receptors (TLR´s), besonders TLR-2. Durch
die Rezeptorbindung wird der Makrophage aktiviert und es erfolgt über die Induktion von intrazellulären
Signalkaskaden die Produktion inflammatorischer Zytokine und Chemokine . Makrophagen bilden dabei
die Hauptquelle für die Produktion der wichtigsten inflammatorischen Parameter wie TNF-α, Il-6 und Il-β,
wie sie für die Abwehr einer Infektion entscheidend sind, als auch an der Entstehung des septischen
Schocks beteiligt sind. Kommt es nämlich zu einer massiven inflammatorischen Antwort, hat das eine
systemische Zytokinwirkung und damit ein septisches Geschehen zur Folge. Grundlage dafür bildet im
Wesentlichen die durch den proinflammatorischen Zytokinüberschuss ausgelöste endotheliale
Überaktivierung und damit mikrovaskuläre Dysfunktion, die letztlich in ein Multiorganversagen mündet
(Fournier und Philpott 2005; Fournier 2012; Sagy et al. 2013).
Die Phagozytose ist ein Prozess, der zur Aufnahme des Erregers ins Phagosom führt, einem neu
gebildeten Organell im Zytoplasma. Die graduelle Veränderung des Phagosoms hin zum Phagolysosom
wird als Phagosomenreifung bezeichnet. Dazu gehören die Ansäuerung des Phagosomeninneren, sowie
die Bildung reaktiver oxidativer Radikale, die durch die Aktivierung des NADPH-Oxidase-Komplexes
entstehen. Zusätzlich entstehen reaktive Stickoxidverbindungen, gebildet durch die Stickoxid-Synthetase
(iNOS). Es handelt sich um zwei essentielle Enzymkomplexe an der Phagosomenwand. Im letzten Schritt
erfolgt die Verschmelzung des Phagosoms mit dem Lysosom, das Lysozyme, Hydrolasen und Defensine
enthält, die den aufgenommenen Mikroorganismus verdauen und abbauen können. In der Regel
überleben Makrophagen die Phagozytose und die Wirkung der von ihnen produzierten reaktiven
bakteriziden Verbindungen. Neutrophile Granulozyten hingegen sterben nach der Phagozytose ab und

EINLEITUNG 7
werden dann wiederum durch Makrophagen abgebaut (Alberts 2011; Rigby und DeLeo 2012; DeLeo et
al. 2009).
Einige bakterielle Erreger haben gelernt, intrazellulär in Makrophagen zu überleben und zu
replizieren. Dazu gehören u.a. Mycobakterium tuberculosis, Salmonella enteritidis, Listeria
monocytogenes, Legionella pneumophila und Burkholderia pseudomallei . Von diesen ist bekannt, dass
sie sich intrazellulär in Makrophagen vermehren können, statt abgebaut zu werden, und dass sie somit
Infektionen aufrecht erhalten (Amer und Swanson 2002 ; Yamamoto 2006). Mycobakterium tuberculosis
und Legionella spp. beispielsweise können die Reifung des Phagosomens verhindern, indem sie die
phagolysosomale Fusion unterbinden. Sie persistieren dann in den frühen Reifungsstadien des s ie
umschließenden Phagosoms, das noch nicht viele bakterizide Komponenten enthält (Rohde et al. 2007;
Xu und Luo 2013). Andere Erreger wie Listeria spp. und B. pseudomallei zerstören das Phagosom in
einem frühen Reifungsstadium und gelangen dabei ins Zytosol (Dewamitta et al. 2010; Gan 2005; Lazar
Adler, Natalie R et al. 2009).
1.2.2 Oxidative Effektormechanismen von Makrophagen
Stickoxid (NO) stellt eine zentrale Komponente für die bakterizide Aktivität von Makrophagen dar,
insbesondere gegenüber intrazellulären Erregern (Fang und Vazquez-Torres 2002; Nathan und Hibbs, J
B Jr 1991; Vazquez-Torres et al. 2008; Nathan und Shiloh 2000; Chakravortty und Hensel 2003). NO und
NO-Derivate werden zusammenfassend als Reactive Nitrogen Intermediates (RNI) bezeichnet und sind
Produkte der Stickoxid-Synthase (NOS). Bisher konnte die Existenz von drei Isoformen dieses Enzyms
nachgewiesen werden, die nach dem Ort ihres jeweiligen Vorkommens als eNOS (in Endothelzellen) und
nNOS (in Neuronen) bezeichnet werden. (Hibbs, John B Jr 2002) Die dritte Form des Enzyms ist die
induzierbare Stickoxid-Synthase iNOS, die vor allem in Makrophagen vorkommt (Nathan 1997;
MacMicking et al. 1997). In ruhenden Makrophagen wird iNOS kaum exprimiert. Erst die Aktivierung von
Makrophagen durch Zytokine, insbesondere IFN-γ, TNF-alpha, Il-1β, oder die Erkennung und Bindung
von Mikroorganismen führt zur Transkription des iNOS Gens (Liew 1994; Kang et al. 2011; Kim et al.
2015).
Die Sauerstoffradikale demgegenüber, gebildet von der NADPH-Oxidase, werden auch unter
dem Begriff Reactive Oxygen Intermediates (ROI) zusammengefasst. Allgemein können diese von allen
aeroben Zellen gebildet werden, doch sind die Phagozyten von Säugern auf eine besonders hohe
Produktion von ROI spezialisiert. Dabei besitzen neutrophile Granulozyten die höchste Kapazität zur
Bildung und Freisetzung von ROI (Nathan und Shiloh 2000). Gendefekte in den verschiedenen
zytosolischen, oder membrangebundenen Untereinheiten des NADPH-Oxidase-Komplexes führen zum

EINLEITUNG 8
Funktionsverlust des Enzyms. Eine bekannte Erkrankungsform diesbezüglich ist die Chronische
Granulomatose, eine Immunschwächeerkrankung mit fehlender Bildung von ROS, die folglich mit ernsten
und rekurrenten Infektionen durch z.B. S. aureus, Aspergillus fumigatus und Salmonella spp. einhergeht
(Vignais 2002).
Neben der bakteriziden Funktion in Phagozyten verfügen ROI und RNI über zusätzliche
regulatorische Funktionen im Immunsystem mit Wirkung über die Zelle hinaus. Dies betrifft die Induktion
von Signalwegen, die Anlockung von Neutrophilen, oder die Aktivierung der Zytokinproduktion (Guzik et
al. 2003; Bogdan et al. 2000a).
1.3 Intrazelluläres Überleben von Staphylokokken
Wenn auch lange als klassischer extrazellulärer Erreger beschrieben (Finlay und Cossart 1997), besteht
inzwischen kein Zweifel daran, dass sich S. aureus auch intrazellulär aufhalten kann. Zahlreiche in-vitro-
Modelle der letzten zwei Jahrzehnte mit nicht-professionell phagozytierenden Zellen, wie Endothelzellen,
Epithelzellen, Enterozyten, Keratinozyten, Osteoblasten (Edwards et al. 2010; Sinha und Fraunholz 2010;
Sinha und Herrmann 2005; Sinha et al. 2000; Kahl et al. 2000; Garzoni und Kelley 2009; Hess et al. 2003;
Edwards et al. 2011; Ellington et al. 2003; Fraunholz und Sinha 2012) zeigten, dass S. aureus in Zellen
eindringen und dort für längere Zeit persistieren kann. Selbst in professionell phagozytierenden Zellen
wie Makrophagen und Neutrophilen wurde in den letzten Jahren beschrieben, dass S. aureus die
Phagozytose überleben und intrazellulär verbleiben kann (Voyich et al. 2005; Gresham et al. 2000; Kubica
et al. 2008; Olivier et al. 2009; Hamza und Li 2014).
Darüberhinaus unterstützen zahlreiche in-vivo-Untersuchungen die Annahme, dass
intrazellulärer S. aureus tatsächlich eine wichtige Rolle in der Pathogenese von S. aureus-Infektionen
spielt. Studien zu chronischer Mastitis (Hebert et al. 2000; Brouillette et al. 2003), Rhinosinusitis (Plouin-
Gaudon et al. 2006; Clement et al. 2005) und Osteomyelitis (Hamza et al. 2013), die häufig chronische,
rekurrente und therapierefraktäre Verläufe zeigen, weisen auf den Zusammenhang von intrazellulärem
S.aureus und der Entstehung einer Infektion des Organismus hin. Gresham et al. beispielsweise
infizierten gesunde Maus-Organismen mit Neutrophilen, die aus erkranktem Gewebe isoliert wurden und
in denen intrazellulärer S. aureus nachweisbar war, wodurch eine Infektion der gesunden Organismen
erzeugt werden konnte (Gresham et al. 2000). S. aureus wurde intrazellulär in Osteoblasten aus
Knochenmaterial von chirurgischen Patienten mit chronischer, therapieresistenter Osteomyelitis
nachgewiesen (Bosse et al. 2005), und es konnte entsprechend am in-vitro-Modell gezeigt werden, dass
infizierte Osteoblasten intrazellulären S. aureus freigeben können und dies zu einer Reinfektionen von
neuen Osteoblasten führt (Ellington et al. 2003). In einer jüngeren in-vivo-Studie konnte am Tiermodell

EINLEITUNG 9
gezeigt werden, dass intrazellulärer S. aureus schwere Knocheninfektionen verursacht, nachdem
S. aureus-infizierte Osteoblasten zu offen frakturiertem Knochen gegeben wurden (Hamza et al. 2013).
Auch S. epidermidis besitzt die Fähigkeit der Internalisierung in eukaryonte nicht-professionell
phagozytierende Zellen, wie an Osteoblasten nachgewiesen werden konnte (Valour et al. 2013; Khalil et
al. 2007). Auch hier ergeben sich Zusammenhänge zwischen intrazellulärem Vorkomen und der
Pathogenese von Erkrankungen, zum Beispiel im Hinblick auf prothesennahe Knocheninfektionen in der
Endoprothetik, oder Biomaterial-assoziierte Infektionen (Broekhuizen, Corine A N et al. 2008;
Broekhuizen, C A N et al. 2010; Valour et al. 2013).
1.3.1 Staphylokokken in nicht-professionell phagozytierenden Zellen
1.3.1.1 Phagozytose
Von zentraler Bedeutung für die Internalisation von S. aureus in nicht-professionell phagozytierende
Zellen sind die Fibronektin-bindenden Proteine (FnBP) auf der Bakterienoberfläche, das extrazelluläre
Matrixprotein Fibronektin, sowie der Wirtszellrezeptor α5β1-Integrin (Sinha et al. 1999; Fowler et al. 2000;
Menzies 2003; Dziewanowska et al. 1999; Hauck und Ohlsen 2006; Hoffmann et al. 2011; Hauck et al.
2012; Ridley et al. 2012). Fibronektin vermittelt dabei die Bindung der FnBP´s an den Integrinrezeptor.
Über weitere Integrin-vermittelte intrazelluläre Signalwege kommt es dann, einem Phagozyten-ähnlichen
Zipper-Mechanismus entsprechend, zur aktiven Internalisation des Erregers durch die Wirtszelle (Agerer
et al. 2003; Sinha et al. 2000; Edwards et al. 2011; Hauck und Ohlsen 2006; Schroder et al. 2006; Agerer
et al. 2005).
Experimente mit einem weiteren typischen zellwandständigen Protein von S. aureus, Clumping
Faktor A, ergaben zwar die Adhärenz an die Wirtszelle, im Gegensatz zu FnBP aber nicht die
Internalisation. Der Integrin-vermittelte aktive Aufnahmeprozess blieb aus (Sinha et al. 2000).
Andererseits konnte eine Internalisierung auch nach Ausschaltung der fnbp-Gene nachgewiesen werden,
so dass vermutlich noch weitere bakterielle Adhäsine die Phagozytose induzieren können (Brouillette et
al. 2003). Bedeutsam wird dies bei dem auch in dieser Arbeit verwendeten S. aureus Stamm Newman,
dessen fnbp-Gene aufgrund struktureller Veränderungen funktionslos sind (Grundmeier et al. 2004). Das
Extrazelluläre Adhärenz Protein (Eap) spielt bei S. aureus Newman eine zentrale Rolle und kompensiert
die Fibronektin-bindende Kapazität, als auch seine Internalisierungsrate (Hussain et al. 2002; Haggar et
al. 2003; Harraghy et al. 2003; Bur et al. 2013b). Der genaue Zell-Rezeptor dafür ist bisher nicht
identifiziert (Alva-Murillo et al. 2014).
Dazu kommen Untersuchungen der jüngsten Zeit, wo das S. aureus Autolysin (Atl), eine
Hydrolase, als ein weiteres Invasin identifiziert wurde. Das Heat-shock-Protein Hsc70 dient dabei als

EINLEITUNG 10
Wirtszellrezeptor, wobei die Bindung mit Atl direkt und ohne Brückenmolekül erfolgt (Hirschhausen et al.
2010). Im Fall von S. epidermidis scheint diese Form der Internalisierung sogar essentie ll zu sein
(Hirschhausen et al. 2010).
1.3.1.2 Intrazelluläres Überleben
S. aureus verfügt je nach Stamm und auch je nach Wirtszelltyp über verschiedenste Strategien, um im
intrazellulären Milieu zu überleben und den Abwehrmechanismen des Wirtszellorganismus zu
entkommen (Sinha und Fraunholz 2010). Grundlegend hierfür ist die Fähigkeit des Bakteriums zur
Anpassung der Genexpression an die intrazelluläre Umgebung (Garzoni et al. 2007). Die Produktion
verschiedener Toxine, wie beispielsweise α-Toxin, ermöglicht S. aureus das Verlassen des
Phagolysosoms und den Übergang ins Zytosol (Jarry et al. 2008; Shompole et al. 2003; Giese et al.
2011b; Sinha und Fraunholz 2010). Es wird weiterhin beschrieben, dass nach Übergang ins Zytosol
S. aureus die Apoptose der Wirtszelle induzieren kann (Bayles et al. 1998; Haslinger-Loffler et al. 2006;
Menzies und Kourteva 2000). Auch die Fähigkeit, die Autophagie für sich zu nutzen, wurde beschrieben
(Schnaith et al. 2007; Pareja, Maria Eugenia Mansilla und Colombo 2013). Ein erwähnenswerter Aspekt
in diesem Zusammenhang ist auch die Bildung von Small Colony Variants (SCV´s). Es handelt sich dabei
um weniger virulente Mutanten, die metabolisch inaktiv über lange Zeiträume intrazellulär verbleiben
können (Garzoni und Kelley 2011; Tuchscherr et al. 2010; Eiff et al. 2006).
Untersuchungen an verschiedenen klinischen S. aureus-Stämmen brachten hervor, dass kurz
nach der Aufnahme in Keratinozyten und Fibroblasten die Mehrheit der Stämme im Phagolysosom
erfolgreich abgetötet wird. Nur wenigen besonders zytotoxischen Stämmen war es vorbehalten, das
Phagolysosom zu verlassen und der Wirtsabwehr zu entgehen. Die gleichen Stämme führten in in-vivo-
Experimenten auch zu deutlich erhöhter Mortalität. (Krut et al. 2003).
Ein Beispiel für das verlängerte intrazelluläre Überleben von S. aureus aufgrund eines Defekts
auf Seiten der Wirtszelle ist die Cystische Fibrose. Im in-vitro-Modell wurde nachgewiesen, dass in CF-
Bronchialepithelzellen S. aureus das Phagosom schneller verlässt als in gesunden Zellen und er den
Abwehrmechanismen der Wirtszelle somit entgeht (Jarry und Cheung 2006; Jarry et al. 2008).
1.3.2 Staphylokokken und Makrophagen
1.3.2.1 Phagozytose von Staphylokokken in Makrophagen
Wie oben dargestellt, kann S. aureus in nicht-professionell phagozytierende Zellen aufgenommen werden
und dort überleben. Zunehmend gibt es auch Hinweise für die Aufnahme und intrazelluläres Überleben
in professionellen Phagozyten, den neutrophilen Granulozyten (Gresham et al. 2000; Voyich et al. 2005)

EINLEITUNG 11
und Makrophagen (Martinez-Pulgarin et al. 2009; Kubica et al. 2008; Das und Bishayi 2009, 2010; Hamza
und Li 2014; Das und Bishayi 2010; Dey und Bishayi 2015).
Ein spezieller Aufnahmemechanismus von S. aureus in Makrophagen ist nicht bekannt. Vielmehr
gibt es aufgrund der Fülle von Rezeptoren auf der Makrophagenoberfläche verschiedene Ansätze, die
Aufnahme von S. aureus in Makrophagen zu erklären. Neben den bekannten Rezeptoren für Komplement
und Fcγ im Rahmen der opsoninabhängigen Phagozytose sind zahlreiche spezifische Rezeptoren
bekannt, die Mikroorganismen direkt erkennen und binden. Dazu zählen die Scavenger Rezeptoren, mit
den Vertretern SRA und CD36 als am meisten untersuchte Rezeptoren im Zusammenhang mit der
S. aureus-spezifischen Abwehr (Stuart et al. 2005; Thomas et al. 2000; Leelahavanichkul et al. 2012).
Die speziellen Liganden auf bakterieller Seite bzw. die konkreten Signalwege, die intrazellulär nach der
Bindung ausgelöst werden, sind nicht einheitlich beschrieben bzw. stammspezifisch sehr unterschiedlich
(DeLoid et al. 2009) (Blanchet et al. 2014). Beide Rezeptoren spielen auch in-vivo eine bedeutsame Rolle
in der Abwehr von S. aureus-Infektionen (Stuart et al. 2005; Thomas et al. 2000; Blanchet et al. 2014),
vermutlich bedingt durch eine erhöhte Phagozytoseleistung des Makrophagen (Thomas et al. 2000).
Zu erwähnen sind ebenso die Rezeptoren der TLR-Familie, mit TLR2 als bedeutendstem
Vertreter für S. aureus-Infektionen. Diese sind entscheidend für die Aktivierung von Signalkaskasen und
die Regulation der Zytokinproduktion, sind selbst aber keine eigentlichen Phagozytoserezeptoren
(Watanabe et al. 2007; Yimin et al. 2013; Fournier 2012; Bishayi et al. 2014).
Zur Bedeutung von FnBP und dem α5β1-Integrin-Rezeptor, wie sie für die Internalisation von
S. aureus in nicht-professionell phagozytierende Zellen vielfach beschrieben ist (Sinha et al. 1999; Alva-
Murillo et al. 2014), gibt es auch bezüglich der Aufnahme in Makrophagen einige Angaben. Wang et al.
konnten beispielsweise zeigen, dass unter den vielen bekannten Makrophagen-Rezeptoren das α5β1-
Integrin für die Phagozytose von S. aureus eine bedeutende Rolle spielt (Wang et al. 2008). Weiterhin
wurde nachgewiesen, dass die FnBP-vermittelte Fibronektinbindung von S. aureus die Phagozytoserate
in Makrophagen eindeutig steigert (Shinji et al. 1998). Interessanterweise wurde in diesem
Zusammenhang beobachtet, dass der KNS-Stamm S. saprophyticus nicht stärker aufgenommen wird,
obwohl er ebenso FnBP´s auf seiner Oberfläche trägt und auch in gleichem Maß an Fn bindet. Ähnliches
wurde auch für die Internalisation des KNS-Vertreters S. epidermidis in nicht-professionell
phagozytierende Zellen beschrieben (Sinha et al. 1999; Agerer et al. 2005). Die Fn-FNBP-Bindung allein
bewirkt keine Phagozytose in Makrophagen. Es wird daher angenommen, dass u.a. die strukturellen
Unterschiede der Fn-FnBP-Bindung im Fall von S. aureus im Gegensatz zu KNS-Stämmem die
Aktivierung des Wirtszellrezeptors α5β1-Integrin auslösen und damit die Internalisierung bewirken (Shinji
et al. 2003; Wang et al. 2008; Shinji et al. 2011).

EINLEITUNG 12
1.3.2.2 Intrazelluläres Überleben von Staphylokokken in Makrophagen
Makrophagen sind die ersten Zellen der angeborenen Abwehr, die durch die Phagozytose bakterielle
Erreger eliminieren und eine Infektion verhindern. Es ist bekannt, dass S. aureus der Makrophagen-
vermittelten Abwehr widersteht, indem er intrazellulär persistieren kann (Kubica et al. 2008; Hamza und
Li 2014; Dey und Bishayi 2015). Auch für Neutrophile wurde dies beschrieben (Gresham et al. 2000;
Voyich et al. 2005; Voyich et al. 2009). Die Mechanismen, durch die er dem intrazellulären Abbau
entgehen kann, sind bisher unvollständig beschrieben. Eine Grundlage ist u.a. eine Veränderung der
Genexpression, um sich an das intrazelluläre Milieu anzupassen und dieses sogar zu nutzen. Diese
Anpassung erfolgt durch die globalen Genregulatoren, vor allem agr und sigB (Kubica et al. 2008). Wie
bereits für nicht-professionell phagozytierende Zellen beschrieben (Jarry et al. 2008), spielt für das
Überleben von S. aureus in Makrophagen das agr-regulierte zytolytische α-Toxin eine essentielle Rolle,
sowie weiterhin die Protease Aureolysin (Kubica et al. 2008), und die antioxidativen Moleküle Katalase
und Superoxiddismutase (SOD) (Das et al. 2008; Das und Bishayi 2009, 2010).
Für Neutrophile wurde beschrieben, dass S. aureus die Phagozytose überlebt und recht schnell
eine Lyse der Wirtszelle hervorruft (Voyich et al. 2005; Lundqvist-Gustafsson et al. 2001), was mit einer
erhöhten Virulenz des Erregers in-vivo korreliert (Voyich et al. 2009). Ähnliches konnte auch bereits in
nicht-professionell phagozytierenden Zellen beobachtet werden (Bayles et al. 1998; Menzies und
Kourteva 1998; Haslinger-Loffler et al. 2006). In Makrophagen überlebt S. aureus hingegen über mehrere
Tage und bewirkt dabei keinen Zelltod (Kubica et al. 2008). Weiterführende Untersuchungen konnten
zeigen, dass durch Phagozytose von S. aureus in Makrophagen sogar antiapoptotische, sowie
zytoprotektive Wege induziert werden (Koziel et al. 2009) und dies wiederum am in-vivo- Modell mit einer
erhöhten Virulenz und Infektionsausbreitung einhergeht (Koziel et al. 2013).
Die Fähigkeit des intrazellulären Überlebens in Makrophagen wurde nicht nur für S. aureus,
sondern auch für Koagulase-negative Staphylokokken nachgewiesen (da Silva Domingues, Joana F et
al. 2013; Boelens et al. 2000). Die klinische Bedeutung fremdkörperassoziierter Infektionen ist erheblich
und gekennzeichnet durch Chronifizierung, Rekurrenz und Therapieresistenz. S. epidermidis konnte ohne
Zeichen des Abbaus in Makrophagen im Fremdkörper-umgebenden Gewebe nachgewiesen werden. Die
intrazelluläre Persistenz von S.epidermidis wurde somit als entscheidend in der Pathogenese von
Fremdkörperinfektionen gesehen (Boelens et al. 2000)

ZIELSTELLUNG 13
1.4 Zielstellung der Arbeit
Es ist bekannt, dass S. aureus die Aufnahme in nicht-professionell phagozytierende Zellen induzieren
kann und dort intrazellulär persistiert. Auch für das intrazelluläre Überleben in Neutrophilen und
Makrophagen als Zellen der angeborenen Immunität gibt es zunehmend Hinweise .
Im Rahmen dieser Arbeit sollte die Erreger-Wirt-Interaktion von S. aureus mit Makrophagen näher
charakterisiert werden. Zu diesem Zweck wurde ein in-vitro-Infektionsmodell mit einer murinen
Makrophagen-Zelllinie, sowie mit primären murinen Knochenmarkmakrophagen etabliert.
Im ersten Teil der Arbeit sollte die Phagozytose und das intrazelluläre Überleben verschiedener
S. aureus-Stämme, sowie Koagulase-negativer Staphylokokken untersucht und verglichen werden. In
diesem Zusammenhang wurde auch der Einfluss von sowohl Fibronektin-bindendem Protein A, als auch
Fibronektin selbst für die Internalisierung von S. aureus untersucht.
Im zweiten Teil der Arbeit sollten bakterizide oxidative Effektormechanismen des Makrophagen
und deren Einflüsse auf die S. aureus-Infektion untersucht werden. Dazu erfolgten Experimente mit
Knochenmarkmakrophagen von Mäusen mit Defekten im iNOS Gen bzw. der gp91 Untereinheit der
NADPH-Oxidase.
Im abschließenden Teil der Arbeit sollte die Bedeutung von zwei bakteriellen Regulatorgenen von
S. aureus im Hinblick auf Phagozytose und intrazelluläres Überleben in Makrophagen betrachtet werden.
Dazu wurden S. aureus-Stämme mit Mutationen in den globalen regulatorischen Systemen agr bzw. sae
für die Infektion in murinen Makrophagen eingesetzt.

14
2 MATERIAL UND METHODEN
2.1 Material
2.1.1 Geräte
Bezeichnung Hersteller
CO2-Brutschrank CB Binder
CO2-Brutschrank S2-Labor Nuaire Zapf Instrumente
ELISA-Reader Multiskan ex Thermo
Inverses Mikroskop Axiovert 40 CF Zeiss
Laser Scanning Mikrosokop Zeiss
Mausschrank Uni Protect Ehret
Photometer Genesys 10UV Thermo
Sicherheitswerkbänke HeraSafe HS 12/2 und K15 Heraeus
Sicherheitswerkbank S2 (Lamina-Flow-Einheit
mit Produktschutz) Clean Air Typ MSC-BSS 4/2
Thermo-Schüttelinkubator KS-15 control Edmund Bühler Labortechnik
Tiefkühlschrank -70°C HFU 686 Basic Heraeus
Tischzentrifuge 5417R Eppendorf
Zentrifuge Multifuge 1L-R Heraeus
Zentrifuge GS15R Beckmann
2.1.2 Gebrauchsmaterialien
Bezeichnung Hersteller
Einmalpipetten Sarstedt
Einmalskalpell Dahlhausen
Falcontubes (50 ml) Becton Dickinson
Glas Cover Slips 12mm Glaswarenfabrik Karl Hecht
Kanülen Dispomed
Küvetten Brand
Objektträger Langenbrinck
Petrischalen Sarstedt
Pipettenspitzen Sarstedt

MATERIAL UND METHODEN 15
Reagiergefäße Sarstedt
Röhrchen (15 ml) Sarstedt
Spritzen Becton Dickinson
Zellkulturflaschen (250 ml) Greiner bio one
Zellkulturplatten Greiner bio one
Zellschaber TPP
2.1.3 Substanzen
Bezeichnung Hersteller
Chloramphenicol Roth
Gentamicin Sigma
Anhydrotetracyclin (AHT) Fischer scientific / Acros
BSA AppliChem
Lysostaphin Sigma
Ethanol Roth
Methanol J. T. Baker
Glycerin Merck
Mercaptoethanol Sigma
Kaliumchlorid Merck
Kaliumdihydrogenphosphat Merck
Caseinhydrolysat Roth
Salzsäure Merck
Natriumdihydrogenphosphat Merck
Natriumhydrogenphosphat J. T. Baker
Natriumnitrit Sigma
N (1-Naphtyl) Ethylendiamin Sigma
Sulfanilsäure Sigma
Natriumchlorid Roth
Kalziumchlorid Sigma
Magnesiumchlorid Sigma
Tween 20 Merck
Ammoniumchlorid AppliChem
Colombia Agar mit 5% Schafsblut Becton Dickinson
Müller-Hinton II Agar-Platten Becton Dickinson

MATERIAL UND METHODEN 16
Müller-Hinton, Pulver Oxoid
Tergitol Fluka
Sevofluran Abbott
2.1.4 Lösungen, Puffer und Medien
Bezeichnung Hersteller/Zusammensetzung
Ammoniumchloridlösung 50mM in NH4Cl
in PBS2+
BSA 10 % in PBS
Natriumnitrit 1 M in Aqua bidest
N (1-Naphtyl) Ethylendiamin 1% in Methanol
Phosphate Buffered Saline (PBS) 0,2 M NaCl
2,5 mM KCl 8 mM Na2PO4 1,5 mM KH2PO4
pH 7,4 PBS2+ 1 mM MgCl,
0,1 mM CaCl2 in PBS pH 7,4
Paraformaldehyd (PFA) 4,95 g Na2HPO4 x 2H2O 0,66 g NAH2PO4 x H 2O
26,31 g NaCl Sulfanilsäure 1 % in HCL (4N)
Tergitol 1 % in PBS mit 0,5 % BSA
2.1.5 Zellkulturmedien
Bezeichnung Hersteller
Dulbecco´s Modified Eagle Medium (DMEM) Biochrom
GM-CSF PAN Biotech
IFN-γ Roche
Panexin (+/-Fibronektin) PAN
PBS-Puffer Sigma
RPMI 1640 Biochrom
Trypanblau (0,4 %) Sigma

MATERIAL UND METHODEN 17
Trypsin-EDTA PAA
2.1.6 Bakterienkultur
Bezeichnung Hersteller/Zusammensetzung
B2 10g Caseinhydrolysat 25g Hefeextrakt 25g NaCl
1,0g K2HPO4
Glucose 0,5 %
Tryptone-Soya-Broth Oxoid
2.1.7 Software
Microsoft Word 8.1
Microsoft Excel 8.1
Ascent Software Version 2.6
GraphPad Prism 4.1
2.2 Bakterien
2.2.1 Bakterienstämme
Bakterienspezies Stammbezeichnung Genotyp Plasmid Resistenzen Herkunft
S. aureus Newman WT AG Hecker,
Engelmann,
Greifswald
Newman saer- saer-
RN 6390 WT
RN 6911 agr- agr-
ATCC 25923 WT
ATCC 12600 WT
ATCC 29213 WT
S. epidermidis ATCC 12228 WT
S. carnosus TM 300 WT AG Fraunholz,
Greifswald TM 300 (GFP) GFP
TM 300 (GFP, FnBA) GFP,FnBA

MATERIAL UND METHODEN 18
2.2.2 Anlegen der Bakterienstocks
Zum Anlegen eines Stocks der in dieser Arbeit verwendeten Staphylokokken-Stämme ohne Plasmide
wurde der Keim am Vortag auf Blutplatte angeimpft, über Nacht bei 37°C bebrütet und eine der Kolonien
am Folgetag von der Platte geerntet und in LB-Medium gebracht. Die Anzucht erfolgte im
Schüttelinkubator bei 37°C und 140 rpm über Nacht. Die Kultur wurde dann mit Glycerin zu einer 30%igen
Lösung versetzt, in 0,5 ml Reagiergefäßen zu je 30 l aliquotiert und bei –70°C eingefroren.
Für S. carnosus erfolgte die Anlage eines Stocks durch Anzucht einer Kolonie von Blutplatte in
B2-Medium über Nacht im Schüttelinkubator. Die Bakterienkultur wurde dann bei 4°C und 9000rpm
abzentrifugiert und das Pellet in 1ml Einfriermedium aufgenommen: 800µl B2 + 200µl Glycerin zu einer
20%igen Lösung. Es wurden 20 µl-Aliquots hergestellt und vor der Lagerung bei -70°C in Stickstoff
schockgefroren.
2.2.3 Anlage von Bakterienkulturen und Bestimmung der Keimzahl
Für alle in-vitro-Infektionsversuche mit S. aureus sowie S. epidermidis und S. carnosus wurden am Vortag
der Infektion 5-10 l eines aufgetauten Aliquots aus dem angelegten Stock in 15-20 ml TSB-Medium,
S carnosus in B2-Medium gegeben und für 16-20 h bei 37°C im Schüttelinkubator bei 140rpm bis zu einer
OD540 von 1,4 kultiviert. Diese Bakterienkultur wurde mit PBS verdünnt auf eine OD540 von 0,4 und je
nach gewünschter MOI im entsprechenden Zellkulturmedium eingestellt. Anhand von vorab
durchgeführten Probemessungen mittels Lebendkeimzahlbestimmungen auf Blutagarplatten wurde von
einer Bakteriendichte von 3,5x108 CFU/ml ausgegangen. Die Dosiskontrolle erfolgte durch CFU-
Bestimmung der Verdünnungsreihe des Infektionsmediums.
2.3 Makrophagen
Die Zellkultivierung erfolgte mit dem Serum-freien Zellkultursystem Panexin BMM®. Dieses wurde
etabliert, um bei der Kultivierung von BMM und folglich in der Charakterisierung von murinen
Makrophagen standardisierte und reproduzierbare Bedingungen zu schaffen (Eske et al. 2009). Mit
diesem standardisierten Medium können Schwankungen in der Konzentration von Serumproteinen
ausgeschlossen und bei der Interpretation von Ergebnissen vernachlässigt werden. Des Weiteren können
im Rahmen der Infektionsversuche bestimmte Faktoren, wie z. B. Fn, kontrolliert eingesetzt werden, um
ihre genaue Funktion zu charakterisieren. Hierzu wurde Panexin BMM® nicht nur für die Zellkultivierung

MATERIAL UND METHODEN 19
von primären Knochenmarkmakrophagen verwendet, sondern auch für die Kultivierung von RAW-
Makrophagen und als Medium für alle mit dieser Fragestellung durchgeführten Experimente.
2.3.1 RAW-Makrophagen
Bei RAW-Makrophagen handelt es sich um eine adhärente murine Makrophagen-Zelllinie, die uns
freundlicherweise von der Arbeitsgruppe von Prof. R. Walther (Institut für Biochemie und
Molekularbiologie, Universität Greifswald) zur Verfügung gestellt wurde. Die Zellkultur erfolgte bei 37°C
und 5% CO2 in DMEM + 4% Panexin BMM® in 250 ml-Zellkulturflaschen zu jeweils 20ml Medium.
Für die Erhaltung der Zellen wurden diese alle 3 bis 4 Tage im alten Medium mittels Zellschaber abgelöst.
Ein bis zwei Milliliter der abgelösten Zellsuspension wurden dann in eine neue Kulturflasche mit frischem
Medium gegeben.
2.3.1.1 Einfrieren und Auftauen von Zellen
Die Zellen einer dicht bewachsenen 250 ml-Zellkulturflasche wurden abgelöst und in ein 50 ml-Röhrchen
überführt, welches bei 150g 10 min abzentrifugiert wurde. Nach Dekantieren des Überstandes wurden
die Zellen in Einfriermedium aufgenommen, das sich aus dem für die Zelllinie entsprechenden
Kulturmedium unter Zusatz von 10% DMSO zusammensetzte. Anschließend wurden die Zellen zunächst
bei -70°C gelagert und nach einigen Tagen in den Stickstofftank überführt. Zum Auftauen wurden die
Zellen im Einfrierröhrchen einige Minuten im Wasserbad bei 37°C inkubiert und anschließend in eine 250
ml-Zellkulturflasche mit 20 ml Medium überführt.
2.3.1.2 Ernten und Aussäen von Zellen
Für die Infektionsversuche wurden die Zellen am Vortag geerntet und auf Zellkulturplatten ausgesät. Dazu
wurden die Zellen zweimal mit je 3 ml PBS gewaschen. Anschließend wurden die Zellen wieder mit 5 ml
frischem Zellkulturmedium versetzt. Mittels Zellschaber wurden die Zellen vom Boden der Kulturflasche
abgelöst, resuspendiert und in ein 50 ml-Röhrchen überführt. Nach nochmaligem Resuspendieren wurde
aus dieser geernteten Zellsuspension die Zellzahl mittels Neubauer-Zählkammer ermittelt:
Dazu wurde ein Aliquot (mindestens 10µl) der Zellsuspension entnommen, mit Trypanblau im Verhältnis
1:2 verdünnt und auf eine Neubauer Zählkammer pipe ttiert. Nach dem Auszählen der Zellen unter dem
Mikroskop, konnte die Zellkonzentration der Suspension nach folgender Formel berechnet werden und
durch Zugabe von Medium eine Suspension mit der gewünschten Zellzahl hergestellt werden:

MATERIAL UND METHODEN 20
Die Färbung mit Trypanblau dient zur Identifizierung von vitalen Zellen, so dass nur diese bei der
Zellzählung berücksichtigt werden. Die Zellen wurden in der gewünschten Dichte im Medium eingestellt
und mit 400µl pro Well auf 48 Well-Zellkulturplatten zu 1x105 Zellen pro Well ausgesät. Bis zum
Experiment wurden die vorbereiteten Platten über Nacht bei 37°C und unter 5%CO2 im Brutschrank
inkubiert.
2.3.2 Murine Knochenmarkmakrophagen
2.3.2.1 Maus-Inzuchtstämme
BALB/cAn
C57Bl/6N
Diese Mausstämme wurden von Charles River (Niederlassung Sulzfeld) bezogen und bis zum Versuch
in Makrokolonkäfigen mit bis zu 10 Mäusen gehalten. Sie wurden mit Haltungsfutter für Versuchstiere und
Wasser ad libitum versorgt.
C57BL/6J
iNOS-/- (auf C57Bl/6J Hintergrund)
gp91phox-/- (auf C57Bl/6J Hintergrund)
Zuchtpärchen dieser Mausstämme wurden von The Jackson Laboratory (USA, Maine) bezogen und im
Institut für Versuchstierkunde der Universität gezüchtet.
2.3.2.2 Präparation und Kultivierung von Stammzellen aus murinem Knochenmark
Murine Knochenmarkmakrophagen wurden aus weiblichen, 10-20 Wochen alten Mäusen gewonnen (Az:
LALLF M-V/TSD/7221.3-2.3-029/05). Die Tötung der Tiere erfolgte durch zervikale Dislokation nach
Sedierung mit Sevofluran.
Vor der Präparation wurde die Maus auf einer Unterlage mit Hilfe von Kanülen fixiert und mit
Alkohol desinfiziert. Das Fell wurde im Bauchbereich geöffnet und von den unteren Gliedmaßen entfernt.
104 Zellzahl x Verdünnungsfaktor Zellzahl x = ml Zahl der Großquadrate

MATERIAL UND METHODEN 21
Es erfolgte nun die Präparation von Femur und Tibia zunächst durch Durchtrennung am Kniegelenk. Die
freigelegten Knochen wurden durch Abschaben unter Verwendung eines Skalpells von Weichteilgewebe
befreit und für mindestens 5 Minuten in eine mit Alkohol gefüllte 10 mm- Petrischale gelegt, danach zum
Spülen in eine weitere Petrischale mit sterilem PBS. Mithilfe eines neuen Skalpells erfolgte die Eröffnung
der Röhrenknochen an beiden Enden innerhalb einer leeren Petrischale und die Spülung des
Markraumes mit einer PBS-gefüllten 5ml-Spritze mit 0,4x12mm-Kanüle. Die Zellsuspensionen aller 4
Knochen wurden in einem 50 ml-Falcon-Röhrchen gesammelt und gut resuspendiert, um die
gewonnenen Zellen zu vereinzeln. Anschließend wurde die Suspension bei 1100 U/Min und 4°C für 10
Min abzentrifugiert. Der Überstand wurde daraufhin vollständig dekantiert und die Zellen in RPMI-Medium
mit Zusatz von 5% Panexin und 50 M Mercaptoethanol resuspendiert. Zur Differenzierung der
Knochenmark-Stammzellen wurde dem Medium des Weiteren Makrophagen-Wachstumsfaktor rmGM-
CSF in einer Konzentration von 2 ng/ml zugesetzt. Die Zellen wurden dann mit jeweils 20 ml in mehrere
250 ml-Zellkulturflaschen ausgesät und bei 37°C und 5% CO2 für 9-11 Tage bebrütet, um sie dann für
Experimente einzusetzen. Am 5. und 7. Tag nach Beginn der Kultivierung wurden die Zellen gefüttert.
Dazu wurden etwa 15 ml des verbrauchten Mediums abgenommen und durch frisches Medium unter
Zugabe von GM-CSF 2 ng/ml ersetzt.
2.3.2.3 Ernten und Aussäen der primären Makrophagen
Am Vortag des Infektionsversuches wurden die primären Makrophagen nach 9-11 Tagen Kultivierung
geerntet. Dazu wurde das alte Medium abgenommen, Reste des Mediums und nicht-adhärente Zellen
mit 3 ml PBS pro Flasche zweimal gewaschen. Im Anschluss wurden die Zellen mit 1,5 ml Trypsin
vollständig benetzt und die Flüssigkeit bis auf wenige Tropfen wieder abgenommen. Nach 10 Min
Inkubation bei 37°C im Brutschrank wurden pro Flasche 5 ml frisches Medium auf die Zellen gegeben,
und diese nochmals mit einem Zellschaber vom Boden abgekratzt, da die Adhärenz der
Knochenmakrophagen auch nach Trypsinbehandlung sehr stark ist. Das Medium mit den abgelösten
Zellen wurde in einem 50ml-Röhrchen aufgenommen. Zum Nachspülen wurde nochmals etwa 5 ml
frisches Medium in jede Kulturflasche gegeben, resuspendiert, wieder abpipettiert und zur Zellsuspension
dazugegeben. Die Zellen wurden dann bei 1100 U/min und 4°C für 10 min abzentrifugiert. Der Überstand
wurde verworfen und die Zellen wiederum in 10 ml PBS gewaschen, um Trypsinrückstände zu entfernen,
da das verwendete Serum-freie Medium Trypsin nicht inhibieren kann. Dann erfolgte die Aufnahme des
Zellpellets in etwa 2-3ml Medium und die Bestimmung der Zellzahl mithilfe der Neubauer Zählkammer
(2.3.1.3). Die Zellsuspension wurde danach in der entsprechenden Menge GM-CSF-haltigen Mediums
eingestellt. Nun erfolgte die Aussaat der Zellen auf 48 Well-Zellkulturplatten mit 400l pro Well in einer

MATERIAL UND METHODEN 22
Dichte von 1x105 Zellen pro Well und die Inkubation über Nacht bei 37°C und 5% CO2 im Brutschrank
bis zur Adhärenz.
2.4 Staphylococcus aureus - Infektionsversuche mit murinen Makrophagen:
Bakterizidie- und Phagozytoseassays
Für die experimentelle Untersuchung der Interaktion von S. aureus mit Makrophagen wurde ein
Infektionsmodell in-vitro etabliert. Murine Makrophagen der Zelllinie RAW bzw. primäre Zellen, gewonnen
aus murinem Knochenmark von C57Bl/6- und BALB/c-Makrophagen, wurden dafür mit Staphylokokken
infiziert, wie im Folgenden beschrieben. Um die Interaktion von Seiten der Wirtszelle, als auch von
bakterieller Seite zu charakterisieren, konnte der Bakterizidie-Assay entsprechend modifiziert werden.
Um die Phagozytose von Bakterien in Makrophagen zu untersuchen, wurde lediglich nur der Zeitpunkt
Null untersucht (sog. „Phagozytose-Assay“).
2.4.1 Makrophagen-Bakterizidie-Assay mit S. aureus
Für den Bakterizidie-Assay wurde das Kulturmedium aus den 48 Well Platten abgenommen und die
Zellen zweimal mit etwa 300µl PBS gewaschen, bevor das vorbereitete Infektionsmedium auf die Zellen
gebracht wurde. Die MOI wurde zwischen etwa 200-500 CFU pro Zelle angesetzt (Beschreibung der
Einstellung des Bakterienmediums unter 2.2.3). Nach 1 h Infektionszeit wurden die Wells erneut zweimal
mit PBS gewaschen und so die nicht-adhärenten, nicht-phagozytierten Bakterien entfernt. Nun erfolgte
die kontinuierliche Inkubation mit Zellkulturmedium unte r Zugabe von Gentamycin in einer Konzentration
von 50 µg/ml, um extrazelluläre Bakterien abzutöten und Reinfektionen der Zellen zu vermeiden.
Gentamycin verbleibt bei der eingesetzten Konzentration extrazellulär und durchdringt nicht die
Zellmembran. Vorab durchgeführte Experimente mit unterschiedlichen Konzentrationen von Gentamycin
im Zellkulturmedium zeigten bei einem Gehalt von 50 µg/ml eine Abnahme der extrazellulären Keimzahl
auf etwa 1 bis 3 % der intrazellulären Keimzahl. Die Abnahme der extrazellulären Keime war auch bei
intrazellulärer Proliferation zu beobachten, so dass von ausreichender Wirksamkeit des eingesetzten
Antibiotikums ausgegangen werden konnte (Daten nicht gezeigt). Weitere Vorexperimente mit dem
zusätzlich eingesetzten Enzym Lysostaphin (20 µg/ml) für 30 Minuten nach der Infektion hatten zum Ziel,
speziell extrazelluläre adhärente Bakterien abzulösen und zu eliminieren und daraufhin die intrazellulären
Keimzahlen zu ermitteln. Diese ergaben rund 5 fach geringere Werte im Vergleich zu Kontrollen nur mit
Gentamycin (Daten nicht gezeigt). Die Experimente dieser Arbeit wurden jedoch stets nur unter Gabe von
Gentamycin (50 µg/ml) durchgeführt.

MATERIAL UND METHODEN 23
Für die Bestimmung der intrazellulären Bakterienzahl zum Zeitpunkt Null (Phagozytosewert)
wurden die Zellen vor der Lyse für 30 Min mit Medium unter Zusatz von Gentamycin inkubiert. Weitere
Bestimmungen der intrazellulären Keimzahl schlossen sich in den darauffolgenden Tagen nach der
Phagozytose an, wie jeweils den einzelnen Experimenten zu entnehmen ist. Das Antibiotika-haltige
Medium wurde nach 2, spätestens 3 Tagen erneuert, um extrazelluläre Bakterien aus dem Überstand zu
eliminieren.
Zur Erfassung der extrazellulären Bakterienzahlen zu den jeweiligen Zeitpunkten wurde der
Zellüberstand vollständig in 1,5 ml-Gefäße überführt und 100 l davon unverdünnt auf Müller-Hinton-
Agarplatten zur Extrazellulärwertbestimmung ausgestrichen. Für die intrazelluläre Keimzahlbestimmung
wurden die Wells anschließend mit 300 l PBS zweimal gewaschen und jeweils 150 l Tergitol (1%) zur
Zelllyse zugefügt. Nach 10 Min Inkubation bei 37°C im Brutschrank wurden die Zellen vom Boden der
Wells mit einer Pipette kräftig abgekratzt, die Zellsuspension gut resuspendiert, abgenommen und in 1,5
ml-Gefäße pipettiert. 100 l davon wurden in einer Verdünnungsreihe bis zu 10000fach verdünnt und in
Doppelwerten auf Müller-Hinton-Agarplatten ausgestrichen. Nach 48 Stunden konnten die Kolonien
ausgezählt und die Keimzahlen nach folgenden Formeln bestimmt werden:
Mittelwert und Standardabweichung wurden jeweils aus Keimzahlbestimmungen von 3 Wells berechnet.
2.4.2 Makrophagen-Bakterizidie-Assay mit S. epidermidis und S. carnosus
Die Durchführung des Bakterizidie-Assays mit den Koagulase-negativen Stämmen S. epidermidis ATCC
12228, sowie S. carnosus TM 300 erfolgte wie unter 2.4.1 beschrieben. Abweichungen in der
Bakterienkultur sind aus den im Kapitel 2.2 (Bakterien) aufgeführten Erläuterungen ersichtlich.
2.4.3 Makrophagen-Bakterizidie-Assay mit S. aureus unter IFN-Stimulation
Zur Stimulation der Makrophagen wurden diese am Vortag des Experiments mit IFNγ versetzt und dann
in Zellkulturplatten ausgesät und über Nacht inkubiert. IFNγ (Roche) wurde dabei in einer Konzentration
Intrazelluläre Keimzahl / Well = CFU x Verdünnung x 1,5
Extrazelluläre Keimzahl / Well = CFU x Verdünnung x 4

MATERIAL UND METHODEN 24
von 100-300 U/ml eingesetzt. Unter Infektion erfolgte kein Zusatz von IFNγ. Weitere Vorgehensweise wie
unter 2.6.1 beschrieben.
2.4.4 Makrophagen-Bakterizidie-Assay mit S. aureus unter NO-Inhibition mit
Aminoguanidin
Aminoguanidin-hemisulfat wurde als 100 mM Stocklösung in H2O angesetzt. Dem Zellkulturmedium
wurde bei Aussaat der Zellen 2 mM Aminoguanidin zugesetzt. Weiterhin wurde es in dieser Konzentration
wiederum dem Infektionsmedium und dem Antibiotikamedium zugesetzt.
2.4.5 Makrophagen-Phagozytose-Assay mit S. aureus unter Fibronektin-freien
Bedingungen
Makrophagen wurden in Zellkulturmedium mit Fn-freiem Panexin über Nacht kultiviert. Im weiteren
Versuchsverlauf wurden sie stets unter diesen Bedingungen inkubiert. Der normale Gehalt an Fn in
Panexin beträgt 2 mg/l, so dass die Konzentration in DMEM mit 4 % Panexin 80 µg/l beträgt.
2.5 NO-Assay
Die Makrophagen werden wie in dem oben beschriebenen in-vitro-Infektionsversuchen geerntet und in
Zellkulturplatten (48-Well) mit 400µl pro Well ausgesät und über Nacht inkubiert. Je nach Versuchsziel
wurden die Zellen dabei mit IFNγ und/oder Aminoguanidin versetzt.
Die Infektion verlief wie unter 2.4.1 beschrieben. Nach der Infektion und zweimaligem Waschen mit 300µl
PBS pro Well wurden die Zellen mit nur 200µl Medium (DMEM +4% Panexin) unter Hinzugabe von 20µg
Gentamycin pro ml versetzt. Aus diesem Zellkulturüberstand wurden dann die Proben für den NO-Assay
entnommen. Auch bei Verwendung von primären Knochenmarkmakrophagen erfolgte nach der Infektion
die Kultivierung in DMEM, da RPMI bereits geringe Mengen an Nitrit enthält.
50µl Proben wurden zu verschiedenen Zeitpunkten doppelt aus jedem Well entnommen und auf einer
96-Well-Platte aufgetragen. Bei längeren Infektionsverläufen wurden die gewonnenen Überstände bis zur
Messung bei -70°C gelagert. 3 Tage Beobachtungszeitraum konnten dabei nicht überschritten werden,
da nach diesem Zeitraum die Vitalität der Zellen durch Verbrauch des Mediums nicht mehr gewährleistet
war.
Die Messung des Nitritgehaltes im Zellkulturüberstand wurde dann wie folgt durchgeführt:

MATERIAL UND METHODEN 25
Pro Well wurden erst 50µl Sulfanilsäure und danach 10 µl Salzsäure 37% zugegeben und die Platte
bedeckt bei 37°C und 5% CO2 im Brutschrank in Dunkelheit inkubiert. Anschließend erfolgte die Zugabe
von 50µl pro Well Naphtyl-Ethylendiamin und die Messung des Nitritgehaltes der Proben bei 540nm im
ELISA Reader.
Als Standard diente dabei Natriumnitrit (1mM), welches 1:2 in Medium aufgenommen wurde und in einer
Verdünnungsreihe ebenfalls in Doppelwerten auf die entsprechende Platte pipettiert wurde
2.6 Statistik
Die statistische Auswertung erfolgte im Graphikprogramm GraphPad Prism 4 mit dem Student-T-Test.
Die errechneten p-Werte als Ausdruck der Signifikanz wurden innerhalb der im Ergebnisteil abgebildeten
Graphen in drei Größenordnungen angegeben:
p ≤0,05-0,01 *; p <0,01-0,001 **; p <0,001 ***
26
3 ERGEBNISSE
3.1 Die Charakterisierung der Phagozytose von Staphylokokken durch murine
Makrophagen
3.1.1 Die Phagozytose von S. aureus durch murine Makrophagen
Mit dem Ziel dieser Arbeit, die Interaktion von Staphylococcus aureus und Makrophagen im in-vitro-
Infektionsmodell zu charakterisieren, sollte als erstes die Phagozytose mithilfe eines Phagozytose-
Assays beschrieben werden. Dazu wurden RAW-Makrophagen und primäre Knochenmarkmakrophagen
von C57BL/6- und BALB/c-Mäusen eingesetzt. Es sollten dabei Unterschiede in der
Phagozytosekapazität zwischen den Makrophagen der genetisch unterschiedlichen Mausstämme, sowie
im Vergleich zu einer murinen Makrophagen-Zelllinie untersucht werden. Die Infektion der Zellen erfolgte
in erster Linie mit S. aureus Newman. Bei S. aureus Newman handelt es sich um einen sequenzierten
Referenzstamm, der sehr häufig für genetische, funktionelle und in-vivo-Studien eingesetzt wird.
Daneben wurde auch S. aureus RN 6390, ein Derivat von dem S. aureus-Stamm 8325-4 verwendet,
sowie die S. aureus-Referenzstämme ATCC 25923, 12600 und 29213.
A B
New
man
WT
RN 6
390
WT
ATC
C 2
5923
ATC
C 1
2600
ATC
C 2
9213
102
103
104
105
106
***
Intr
azellu
läre
Keim
zah
l p
ro W
ell
New
man
RN 6
390
103
104
105
106
**
Intr
azellu
läre
Keim
zah
l p
ro W
ell
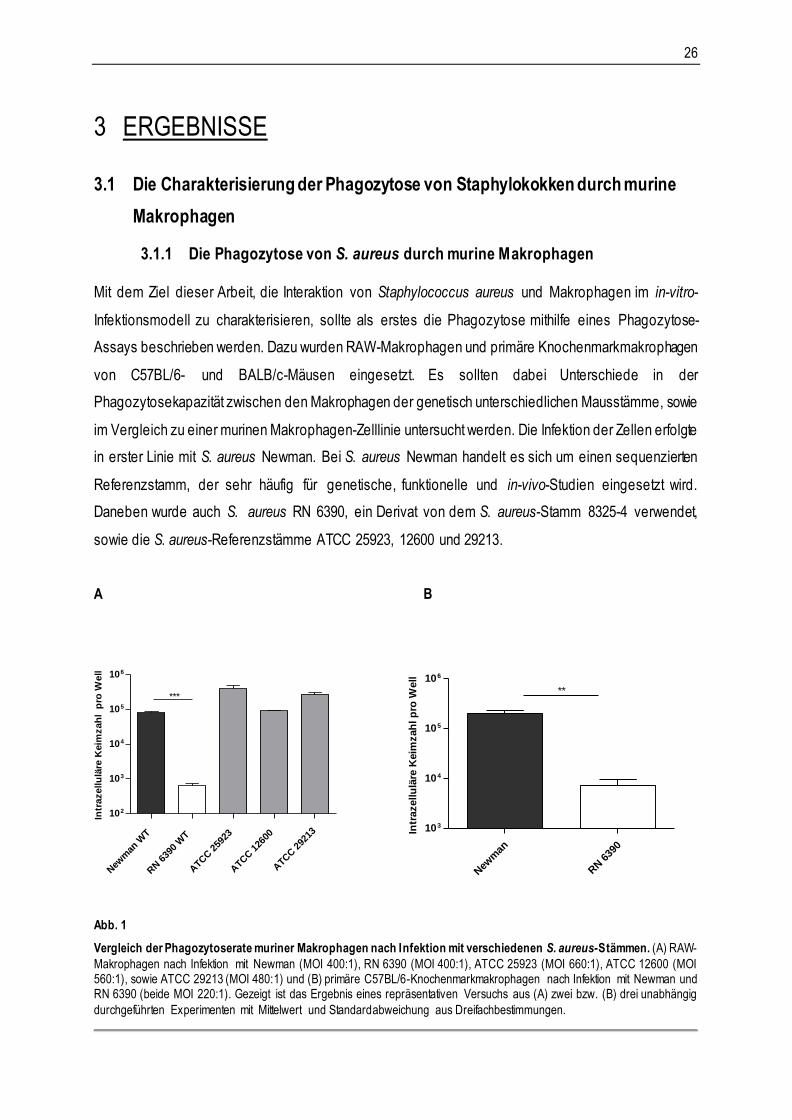
Abb. 1
Vergleich der Phagozytoserate muriner Makrophagen nach Infektion mit verschiedenen S. aureus-Stämmen. (A) RAW-
Makrophagen nach Infektion mit Newman (MOI 400:1), RN 6390 (MOI 400:1), ATCC 25923 (MOI 660:1), ATCC 12600 (MOI 560:1), sowie ATCC 29213 (MOI 480:1) und (B) primäre C57BL/6-Knochenmarkmakrophagen nach Infektion mit Newman und RN 6390 (beide MOI 220:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus (A) zwei bzw. (B) drei unabhängig
durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.

ERGEBNISSE 27
Zunächst wurde die Aufnahme von verschiedenen S. aureus-Stämmen in RAW-Makrophagen untersucht.
Abbildung 1 zeigt, dass zwischen S. aureus Newman und den hier eingesetzten ATCC-Stämmen keine
nennenswerten Unterschiede in der Phagozytose festgestellt werden konnten. Einen hochsignifikanten
Unterschied gab es jedoch beim Vergleich der Internalisierung von Newman und RN 6390 in RAW-
Makrophagen. Dieser zeigte sich auch in primären C57BL/6-Makrophagen (Abb. 1B).
Zahlreich durchgeführte Phagozytose-Assays mit S. aureus Newman (Daten nicht gezeigt) bei
ähnlicher Infektionsdosis (etwa MOI 300:1) zeigten, dass durchschnittlich 100.000 Keime pro Well von
den Zellen phagozytiert wurden (Daten nicht gezeigt). Dies zeigt, dass bei 100.000 ausgesäten Zellen
pro Well mit einem durchschnittlichen Verhältnis von einem Bakterium pro infizierter Wirtszelle zu rechnen
war. Trotz gleicher MOI, gleicher Wirtszellart und standardisiertem Versuchsverlauf unterlag die ermittelte
intrazelluläre CFU in den Experimenten z. T. erheblichen Schwankungen, was mit unterschiedlichen
Wachstumsphasen der Bakterien in der Kultur und folglich veränderter Expression von
Oberflächenproteinen von S. aureus zusammenhängen könnte (Schwankungen zwischen 70.000 bis
300.000 Bakterien pro Well).
Die Zahl intrazellulär detektierter Bakterien bei primären C57BL/6 Makrophagen in Abbildung 1B
verglichen mit den RAW-Makrophagen in Abbildung 1A bei Infektion mit S. aureus Newman, als auch mit
RN 6390 zeigte sich etwa um den Faktor drei erhöht. Eine Ursache mag hier vornehmlich in der Art des
Wirtszellsystems liegen, denn bei primären Makrophagen wurden verglichen mit RAW-Makrophagen
generell, unabhängig vom eingesetzten Bakterienstamm, etwa dreifach höhere intrazelluläre Keimzahlen
ermittelt. Versuche mit BALB/c-Makrophagen zeigten nach Infektion mit Newman (Daten nicht gezeigt)
darüber hinaus in zwei von drei Fällen eine signifikant höhere Phagozytoserate gegenüber C57BL/6-
Makrophagen. In einem Experiment wurde S. aureus Newman hingegen von C57BL/6-Makrophagen
besser phagozytiert, so dass ein reproduzierbarer, signifikanter Unterschied in der Aufnahmekapazität
von BALB/c- und C57BL/6-Makrophagen nicht besteht.
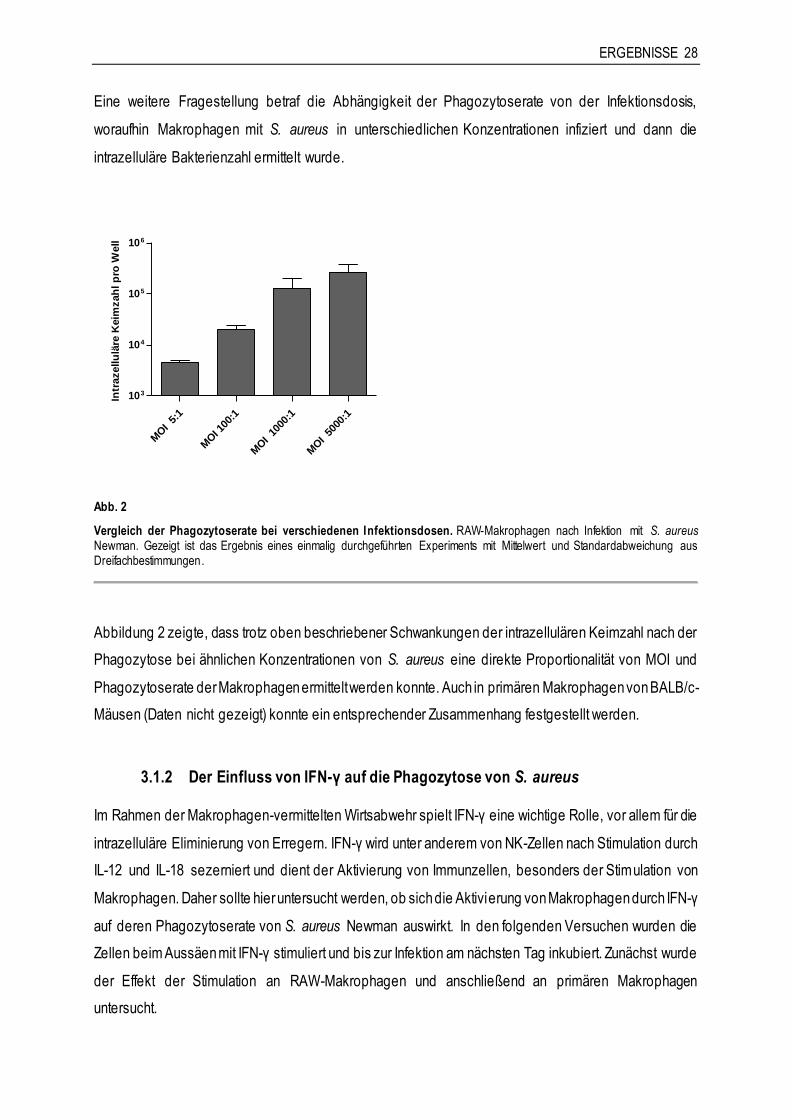
ERGEBNISSE 28
Eine weitere Fragestellung betraf die Abhängigkeit der Phagozytoserate von der Infektionsdosis,
woraufhin Makrophagen mit S. aureus in unterschiedlichen Konzentrationen infiziert und dann die
intrazelluläre Bakterienzahl ermittelt wurde.
MOI
5:1
MOI 1
00:1
MOI
1000
:1
MOI
5000
:1
103
104
105
106
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 2
Vergleich der Phagozytoserate bei verschiedenen Infektionsdosen. RAW-Makrophagen nach Infektion mit S. aureus Newman. Gezeigt ist das Ergebnis eines einmalig durchgeführten Experiments mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Abbildung 2 zeigte, dass trotz oben beschriebener Schwankungen der intrazellulären Keimzahl nach der
Phagozytose bei ähnlichen Konzentrationen von S. aureus eine direkte Proportionalität von MOI und
Phagozytoserate der Makrophagen ermittelt werden konnte. Auch in primären Makrophagen von BALB/c-
Mäusen (Daten nicht gezeigt) konnte ein entsprechender Zusammenhang festgestellt werden.
3.1.2 Der Einfluss von IFN-γ auf die Phagozytose von S. aureus
Im Rahmen der Makrophagen-vermittelten Wirtsabwehr spielt IFN-γ eine wichtige Rolle, vor allem für die
intrazelluläre Eliminierung von Erregern. IFN-γ wird unter anderem von NK-Zellen nach Stimulation durch
IL-12 und IL-18 sezerniert und dient der Aktivierung von Immunzellen, besonders der Stimulation von
Makrophagen. Daher sollte hier untersucht werden, ob sich die Aktivierung von Makrophagen durch IFN-γ
auf deren Phagozytoserate von S. aureus Newman auswirkt. In den folgenden Versuchen wurden die
Zellen beim Aussäen mit IFN-γ stimuliert und bis zur Infektion am nächsten Tag inkubiert. Zunächst wurde
der Effekt der Stimulation an RAW-Makrophagen und anschließend an primären Makrophagen
untersucht.
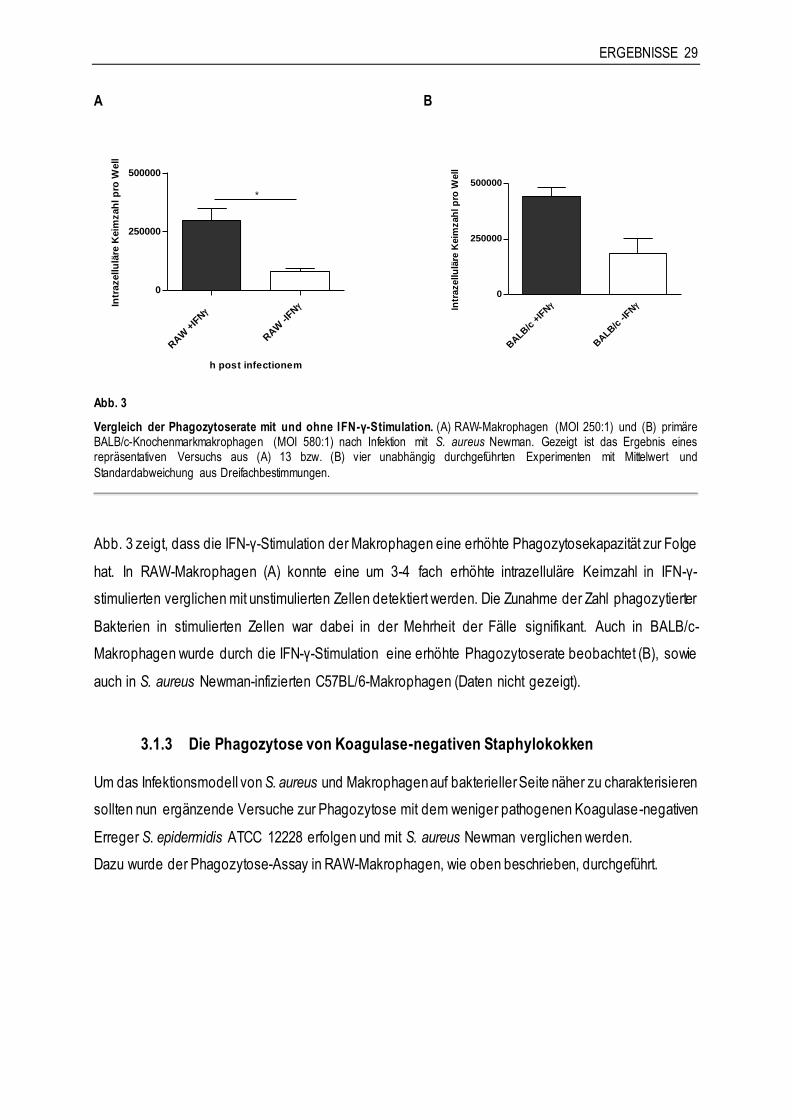
ERGEBNISSE 29
A B
RAW
+IF
N
RAW
-IFN
0
250000
500000
*
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
BALB
/c +
IFN
IFN
-
BALB
/c
0
250000
500000
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 3
Vergleich der Phagozytoserate mit und ohne IFN-γ-Stimulation. (A) RAW-Makrophagen (MOI 250:1) und (B) primäre BALB/c-Knochenmarkmakrophagen (MOI 580:1) nach Infektion mit S. aureus Newman. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus (A) 13 bzw. (B) vier unabhängig durchgeführten Experimenten mit Mittelwert und
Standardabweichung aus Dreifachbestimmungen.
Abb. 3 zeigt, dass die IFN-γ-Stimulation der Makrophagen eine erhöhte Phagozytosekapazität zur Folge
hat. In RAW-Makrophagen (A) konnte eine um 3-4 fach erhöhte intrazelluläre Keimzahl in IFN-γ-
stimulierten verglichen mit unstimulierten Zellen detektiert werden. Die Zunahme der Zahl phagozytierter
Bakterien in stimulierten Zellen war dabei in der Mehrheit der Fälle signifikant. Auch in BALB/c-
Makrophagen wurde durch die IFN-γ-Stimulation eine erhöhte Phagozytoserate beobachtet (B), sowie
auch in S. aureus Newman-infizierten C57BL/6-Makrophagen (Daten nicht gezeigt).
3.1.3 Die Phagozytose von Koagulase-negativen Staphylokokken
Um das Infektionsmodell von S. aureus und Makrophagen auf bakterieller Seite näher zu charakterisieren
sollten nun ergänzende Versuche zur Phagozytose mit dem weniger pathogenen Koagulase-negativen
Erreger S. epidermidis ATCC 12228 erfolgen und mit S. aureus Newman verglichen werden.
Dazu wurde der Phagozytose-Assay in RAW-Makrophagen, wie oben beschrieben, durchgeführt.
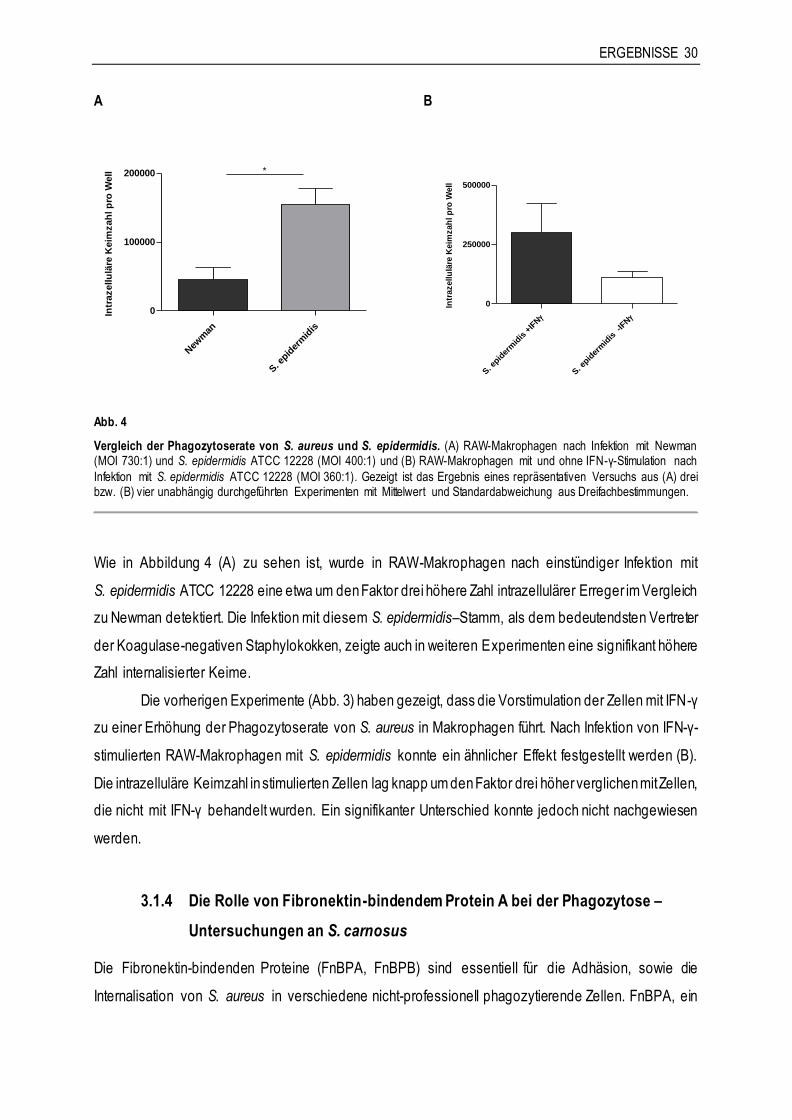
ERGEBNISSE 30
A B
New
man
S. e
pider
mid
is
0
100000
200000 *
Intr
azellu
läre
Keim
zah
l p
ro W
ell
S. epid
erm
idis
+IF
N
S. epid
erm
idis
-IF
N
0
250000
500000
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 4
Vergleich der Phagozytoserate von S. aureus und S. epidermidis. (A) RAW-Makrophagen nach Infektion mit Newman (MOI 730:1) und S. epidermidis ATCC 12228 (MOI 400:1) und (B) RAW-Makrophagen mit und ohne IFN-γ-Stimulation nach
Infektion mit S. epidermidis ATCC 12228 (MOI 360:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus (A) drei bzw. (B) vier unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Wie in Abbildung 4 (A) zu sehen ist, wurde in RAW-Makrophagen nach einstündiger Infektion mit
S. epidermidis ATCC 12228 eine etwa um den Faktor drei höhere Zahl intrazellulärer Erreger im Vergleich
zu Newman detektiert. Die Infektion mit diesem S. epidermidis–Stamm, als dem bedeutendsten Vertreter
der Koagulase-negativen Staphylokokken, zeigte auch in weiteren Experimenten eine signifikant höhere
Zahl internalisierter Keime.
Die vorherigen Experimente (Abb. 3) haben gezeigt, dass die Vorstimulation der Zellen mit IFN-γ
zu einer Erhöhung der Phagozytoserate von S. aureus in Makrophagen führt. Nach Infektion von IFN-γ-
stimulierten RAW-Makrophagen mit S. epidermidis konnte ein ähnlicher Effekt festgestellt werden (B).
Die intrazelluläre Keimzahl in stimulierten Zellen lag knapp um den Faktor drei höher verglichen mit Zellen,
die nicht mit IFN-γ behandelt wurden. Ein signifikanter Unterschied konnte jedoch nicht nachgewiesen
werden.
3.1.4 Die Rolle von Fibronektin-bindendem Protein A bei der Phagozytose –
Untersuchungen an S. carnosus
Die Fibronektin-bindenden Proteine (FnBPA, FnBPB) sind essentiell für die Adhäsion, sowie die
Internalisation von S. aureus in verschiedene nicht-professionell phagozytierende Zellen. FnBPA, ein
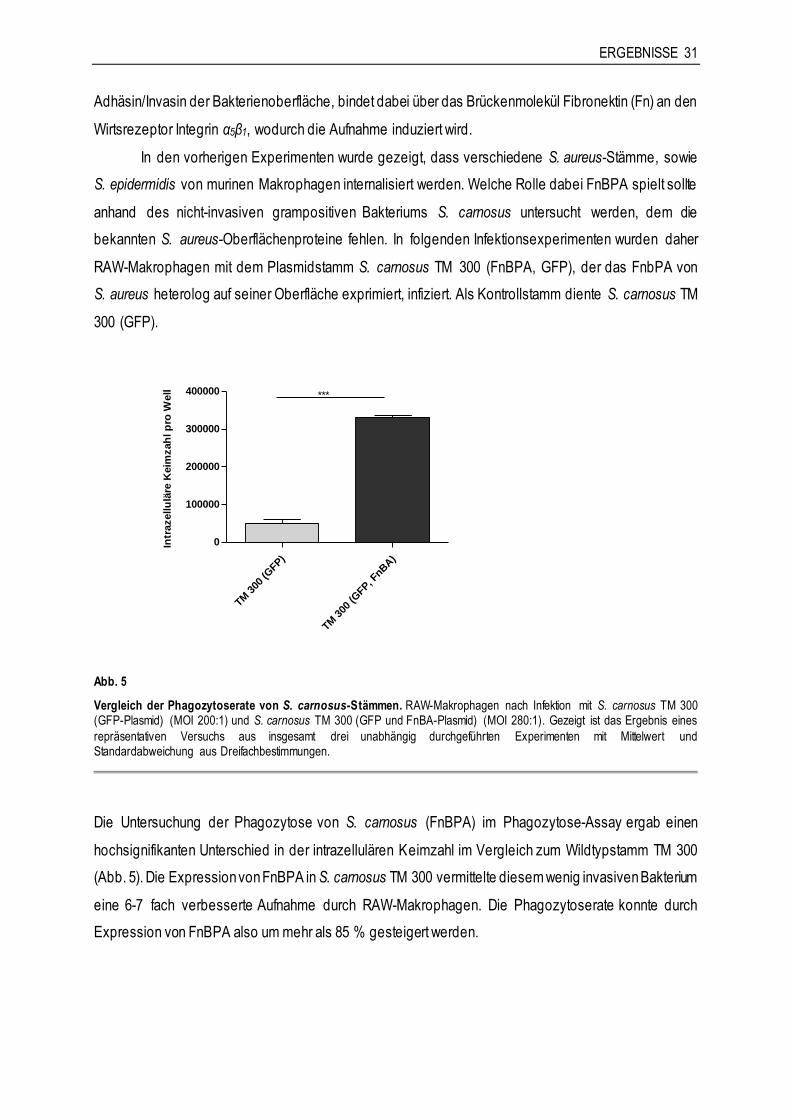
ERGEBNISSE 31
Adhäsin/Invasin der Bakterienoberfläche, bindet dabei über das Brückenmolekül Fibronektin (Fn) an den
Wirtsrezeptor Integrin α5β1, wodurch die Aufnahme induziert wird.
In den vorherigen Experimenten wurde gezeigt, dass verschiedene S. aureus-Stämme, sowie
S. epidermidis von murinen Makrophagen internalisiert werden. Welche Rolle dabei FnBPA spielt sollte
anhand des nicht-invasiven grampositiven Bakteriums S. carnosus untersucht werden, dem die
bekannten S. aureus-Oberflächenproteine fehlen. In folgenden Infektionsexperimenten wurden daher
RAW-Makrophagen mit dem Plasmidstamm S. carnosus TM 300 (FnBPA, GFP), der das FnbPA von
S. aureus heterolog auf seiner Oberfläche exprimiert, infiziert. Als Kontrollstamm diente S. carnosus TM
300 (GFP).
TM 3
00 (G
FP)
TM 3
00 (G
FP, F
nBA)
0
100000
200000
300000
400000 ***
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 5
Vergleich der Phagozytoserate von S. carnosus-Stämmen. RAW-Makrophagen nach Infektion mit S. carnosus TM 300 (GFP-Plasmid) (MOI 200:1) und S. carnosus TM 300 (GFP und FnBA-Plasmid) (MOI 280:1). Gezeigt ist das Ergebnis eines
repräsentativen Versuchs aus insgesamt drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Die Untersuchung der Phagozytose von S. carnosus (FnBPA) im Phagozytose-Assay ergab einen
hochsignifikanten Unterschied in der intrazellulären Keimzahl im Vergleich zum Wildtypstamm TM 300
(Abb. 5). Die Expression von FnBPA in S. carnosus TM 300 vermittelte diesem wenig invasiven Bakterium
eine 6-7 fach verbesserte Aufnahme durch RAW-Makrophagen. Die Phagozytoserate konnte durch
Expression von FnBPA also um mehr als 85 % gesteigert werden.
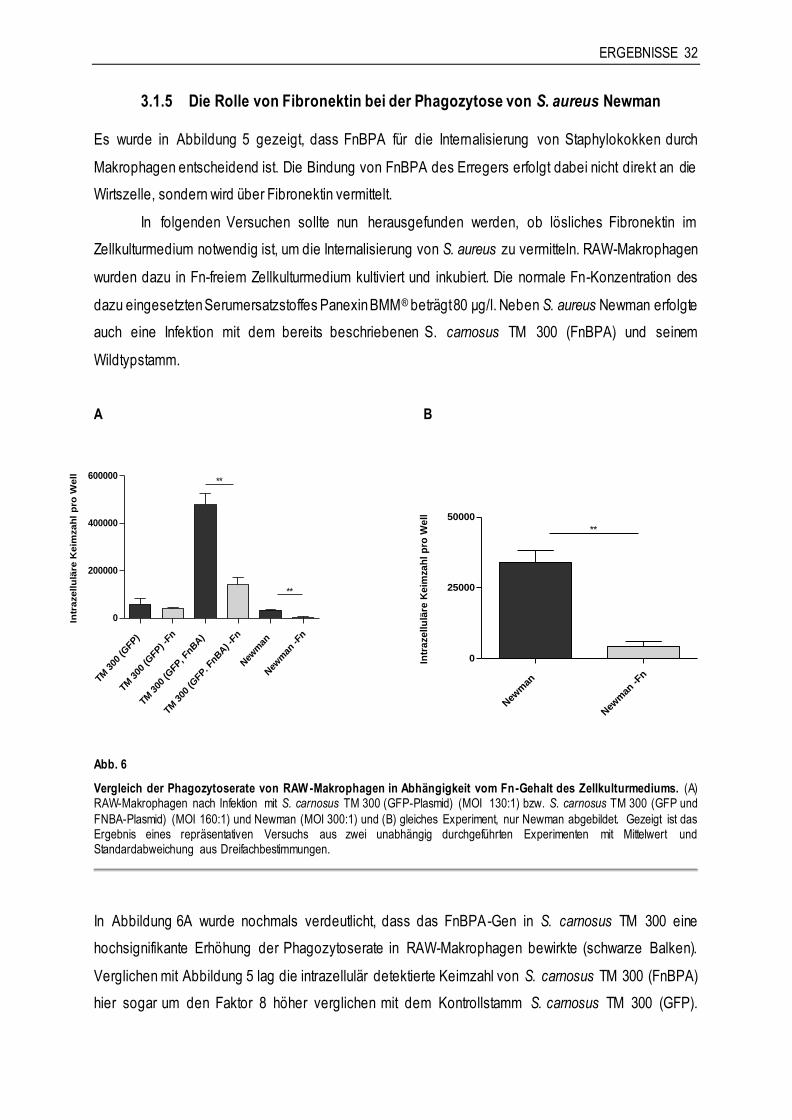
ERGEBNISSE 32
3.1.5 Die Rolle von Fibronektin bei der Phagozytose von S. aureus Newman
Es wurde in Abbildung 5 gezeigt, dass FnBPA für die Internalisierung von Staphylokokken durch
Makrophagen entscheidend ist. Die Bindung von FnBPA des Erregers erfolgt dabei nicht direkt an die
Wirtszelle, sondern wird über Fibronektin vermittelt.
In folgenden Versuchen sollte nun herausgefunden werden, ob lösliches Fibronektin im
Zellkulturmedium notwendig ist, um die Internalisierung von S. aureus zu vermitteln. RAW-Makrophagen
wurden dazu in Fn-freiem Zellkulturmedium kultiviert und inkubiert. Die normale Fn-Konzentration des
dazu eingesetzten Serumersatzstoffes Panexin BMM® beträgt 80 μg/l. Neben S. aureus Newman erfolgte
auch eine Infektion mit dem bereits beschriebenen S. carnosus TM 300 (FnBPA) und seinem
Wildtypstamm.
A B
TM 3
00 (G
FP)
TM 3
00 (G
FP) -
Fn
TM 3
00 (G
FP, F
nBA)
TM 3
00 (G
FP. F
nBA) -
Fn
New
man
New
man
-Fn
0
200000
400000
600000**
**
Intr
aze
llu
läre
Ke
imza
hl
pro
We
ll
New
man
New
man
-Fn
0
25000
50000
**
Intr
aze
llu
läre
Ke
imza
hl
pro
We
ll
Abb. 6
Vergleich der Phagozytoserate von RAW -Makrophagen in Abhängigkeit vom Fn-Gehalt des Zellkulturmediums. (A) RAW-Makrophagen nach Infektion mit S. carnosus TM 300 (GFP-Plasmid) (MOI 130:1) bzw. S. carnosus TM 300 (GFP und
FNBA-Plasmid) (MOI 160:1) und Newman (MOI 300:1) und (B) gleiches Experiment, nur Newman abgebildet. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus zwei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In Abbildung 6A wurde nochmals verdeutlicht, dass das FnBPA-Gen in S. carnosus TM 300 eine
hochsignifikante Erhöhung der Phagozytoserate in RAW-Makrophagen bewirkte (schwarze Balken).
Verglichen mit Abbildung 5 lag die intrazellulär detektierte Keimzahl von S. carnosus TM 300 (FnBPA)
hier sogar um den Faktor 8 höher verglichen mit dem Kontrollstamm S. carnosus TM 300 (GFP).

ERGEBNISSE 33
Kultivierte man RAW-Makrophagen nun in Fn-freiem Medium wurde die Phagozytoserate nach Infektion
mit S. carnosus TM 300 (FnBPA) signifikant um Faktor 3-4 reduziert (A). Sie lag interessanterweise
dennoch immer noch deutlich höher im Vergleich zum Kontrollstamm S. carnosus TM 300 (GFP), auch
ohne Fibronektin im Medium (Abb. 6A).
Auch bei S. aureus Newman konnte ein signifikanter Fn-abhängiger Unterschied in der
Internalisierungsrate festgestellt werden, wie Abbildung 6 (B) des gleichen Experiments zeigte. Die Zahl
internalisierter Bakterien lag bei Fibronektin-haltigem Medium vergleichsweise bereits sehr niedrig (A),
und dennoch wurde sie in RAW-Makrophagen aus Fn-freiem Medium nochmals etwa 8 fach reduziert (B).
3.2 Die Charakterisierung des intrazellulären Überlebens von Staphylokokken in
murinen Makrophagen
Nachdem im ersten Abschnitt die Aufnahme von Staphylokokken durch murine Makrophagen
beschrieben wurde, stellte sich nun die Frage, ob die phagozytierten Erreger durch die bakteriziden
Mechanismen der Wirtszelle eliminiert werden, oder ob Staphylokokken in Makrophagen intrazellulär
persistieren, oder sogar replizieren können. In folgendem Abschnitt sollte daher das intrazelluläre
Überleben von S. aureus, sowie S. epidermidis in murinen Makrophagen beobachtet und charakterisiert
werden. Hierzu wurde der bereits etablierte Phagozytose-Assay zeitlich erweitert und das intrazelluläre
Überleben über einen Zeitraum bis zu 120 h nach der Infektion beobachtet. Zur Vermeidung von
Reinfektionen erfolgte die Zugabe von Gentamycin zum Medium, wie es bekanntermaßen erfolgreich zur
Eradikation extrazellulärer Bakterien eingesetzt wird (Hamza et al. 2013). Als Wirtszellen dienten neben
der Makrophagen-Zellinie RAW wiederum primäre Knochenmarkmakrophagen von C57BL/6- und
BALB/c-Mäusen.
3.2.1 Intrazelluläres Überleben von S. aureus Newman in RAW-Makrophagen
Als erstes wurde das intrazelluläre Überleben von S. aureus Newman in RAW-Makrophagen
charakterisiert. Dabei sollte auch der Effekt von IFN-γ auf die Elimination von intrazellulärem S. aureus
untersucht werden.
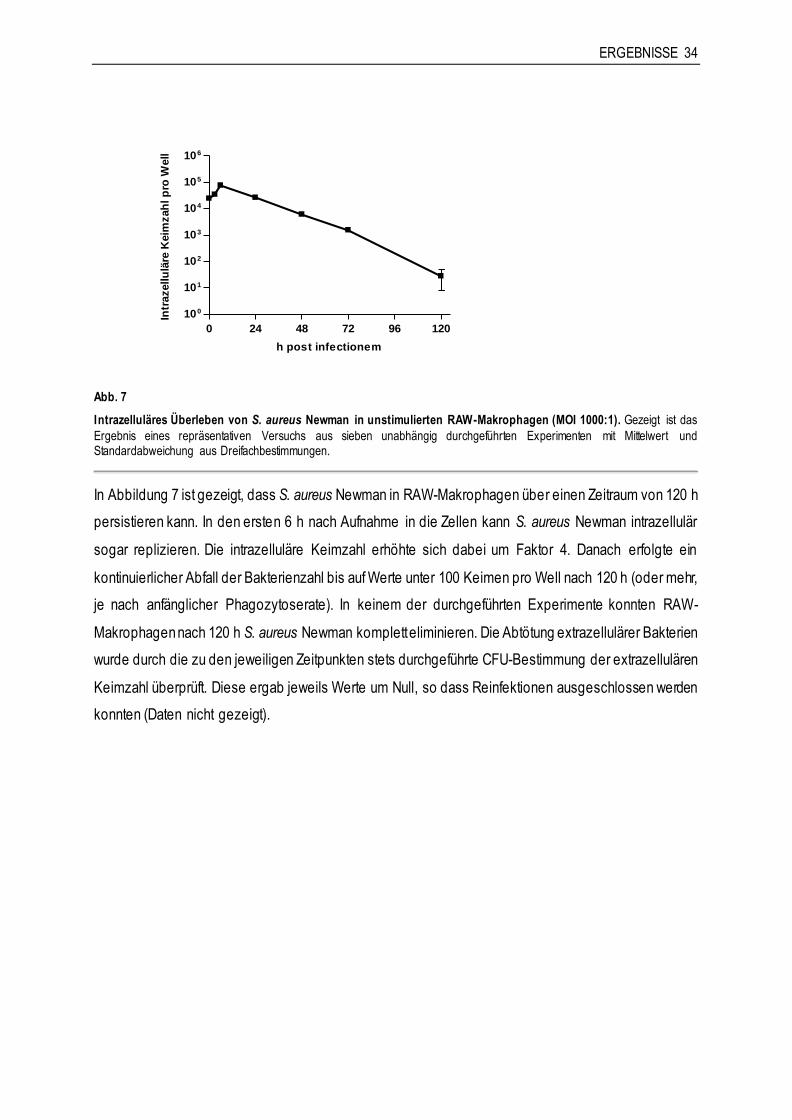
ERGEBNISSE 34
0 24 48 72 96 120
100
101
102
103
104
105
106
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 7
Intrazelluläres Überleben von S. aureus Newman in unstimulierten RAW-Makrophagen (MOI 1000:1). Gezeigt ist das
Ergebnis eines repräsentativen Versuchs aus sieben unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In Abbildung 7 ist gezeigt, dass S. aureus Newman in RAW-Makrophagen über einen Zeitraum von 120 h
persistieren kann. In den ersten 6 h nach Aufnahme in die Zellen kann S. aureus Newman intrazellulär
sogar replizieren. Die intrazelluläre Keimzahl erhöhte sich dabei um Faktor 4. Danach erfolgte ein
kontinuierlicher Abfall der Bakterienzahl bis auf Werte unter 100 Keimen pro Well nach 120 h (oder mehr,
je nach anfänglicher Phagozytoserate). In keinem der durchgeführten Experimente konnten RAW-
Makrophagen nach 120 h S. aureus Newman komplett eliminieren. Die Abtötung extrazellulärer Bakterien
wurde durch die zu den jeweiligen Zeitpunkten stets durchgeführte CFU-Bestimmung der extrazellulären
Keimzahl überprüft. Diese ergab jeweils Werte um Null, so dass Reinfektionen ausgeschlossen werden
konnten (Daten nicht gezeigt).
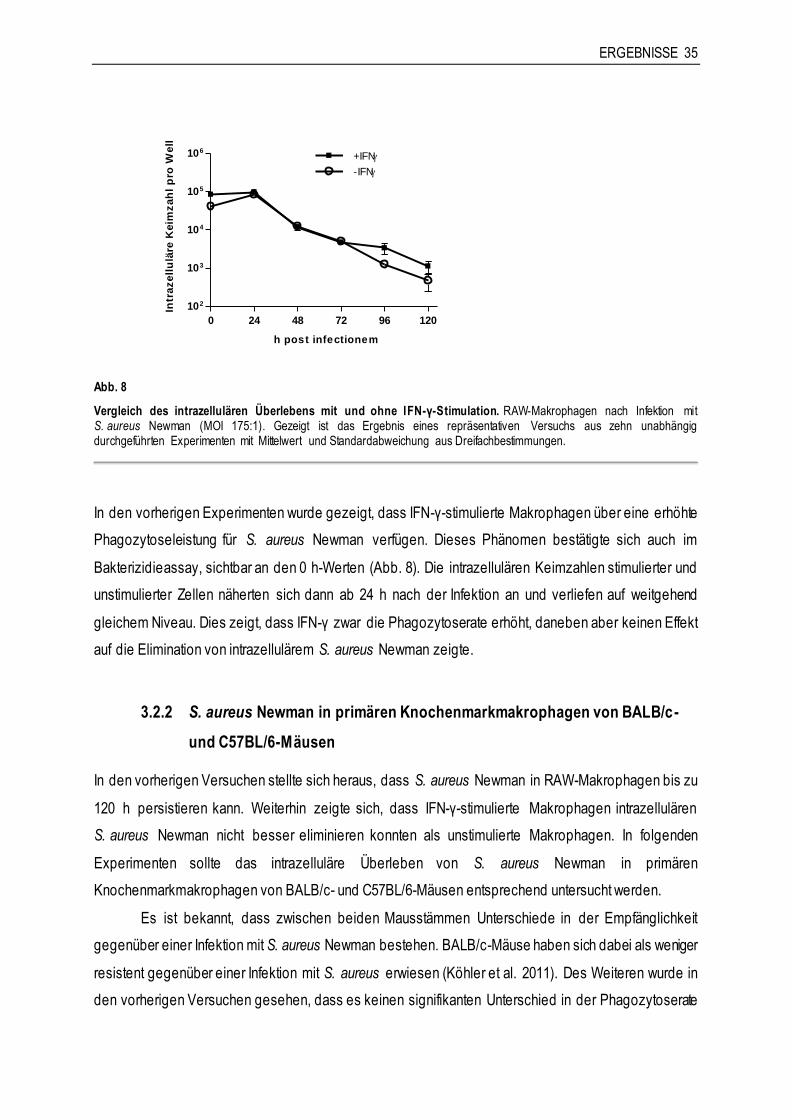
ERGEBNISSE 35
0 24 48 72 96 120
102
103
104
105
106 +IFN
-IFN
h post infectionem
Intr
az
ell
ulä
re K
eim
za
hl
pro
We
ll
Abb. 8
Vergleich des intrazellulären Überlebens mit und ohne IFN-γ-Stimulation. RAW-Makrophagen nach Infektion mit S. aureus Newman (MOI 175:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus zehn unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In den vorherigen Experimenten wurde gezeigt, dass IFN-γ-stimulierte Makrophagen über eine erhöhte
Phagozytoseleistung für S. aureus Newman verfügen. Dieses Phänomen bestätigte sich auch im
Bakterizidieassay, sichtbar an den 0 h-Werten (Abb. 8). Die intrazellulären Keimzahlen stimulierter und
unstimulierter Zellen näherten sich dann ab 24 h nach der Infektion an und verliefen auf weitgehend
gleichem Niveau. Dies zeigt, dass IFN-γ zwar die Phagozytoserate erhöht, daneben aber keinen Effekt
auf die Elimination von intrazellulärem S. aureus Newman zeigte.
3.2.2 S. aureus Newman in primären Knochenmarkmakrophagen von BALB/c-
und C57BL/6-Mäusen
In den vorherigen Versuchen stellte sich heraus, dass S. aureus Newman in RAW-Makrophagen bis zu
120 h persistieren kann. Weiterhin zeigte sich, dass IFN-γ-stimulierte Makrophagen intrazellulären
S. aureus Newman nicht besser eliminieren konnten als unstimulierte Makrophagen. In folgenden
Experimenten sollte das intrazelluläre Überleben von S. aureus Newman in primären
Knochenmarkmakrophagen von BALB/c- und C57BL/6-Mäusen entsprechend untersucht werden.
Es ist bekannt, dass zwischen beiden Mausstämmen Unterschiede in der Empfänglichkeit
gegenüber einer Infektion mit S. aureus Newman bestehen. BALB/c-Mäuse haben sich dabei als weniger
resistent gegenüber einer Infektion mit S. aureus erwiesen (Köhler et al. 2011). Des Weiteren wurde in
den vorherigen Versuchen gesehen, dass es keinen signifikanten Unterschied in der Phagozytoserate
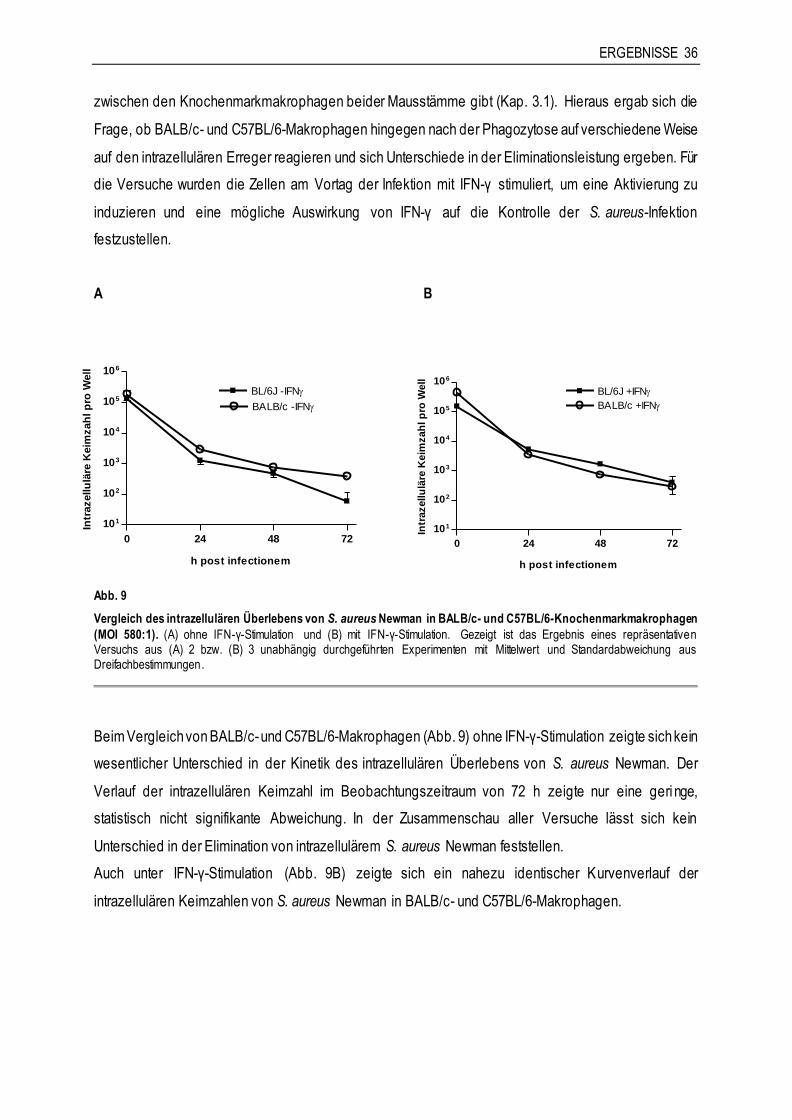
ERGEBNISSE 36
zwischen den Knochenmarkmakrophagen beider Mausstämme gibt (Kap. 3.1). Hieraus ergab sich die
Frage, ob BALB/c- und C57BL/6-Makrophagen hingegen nach der Phagozytose auf verschiedene Weise
auf den intrazellulären Erreger reagieren und sich Unterschiede in der Eliminationsleistung ergeben. Für
die Versuche wurden die Zellen am Vortag der Infektion mit IFN-γ stimuliert, um eine Aktivierung zu
induzieren und eine mögliche Auswirkung von IFN-γ auf die Kontrolle der S. aureus-Infektion
festzustellen.
A B
0 24 48 72
101
102
103
104
105
106
BL/6J -IFN
BALB/c -IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48 72
101
102
103
104
105
106
BL/6J +IFN
BALB/c +IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 9
Vergleich des intrazellulären Überlebens von S. aureus Newman in BALB/c- und C57BL/6-Knochenmarkmakrophagen
(MOI 580:1). (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus (A) 2 bzw. (B) 3 unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Beim Vergleich von BALB/c- und C57BL/6-Makrophagen (Abb. 9) ohne IFN-γ-Stimulation zeigte sich kein
wesentlicher Unterschied in der Kinetik des intrazellulären Überlebens von S. aureus Newman. Der
Verlauf der intrazellulären Keimzahl im Beobachtungszeitraum von 72 h zeigte nur eine geri nge,
statistisch nicht signifikante Abweichung. In der Zusammenschau aller Versuche lässt sich kein
Unterschied in der Elimination von intrazellulärem S. aureus Newman feststellen.
Auch unter IFN-γ-Stimulation (Abb. 9B) zeigte sich ein nahezu identischer Kurvenverlauf der
intrazellulären Keimzahlen von S. aureus Newman in BALB/c- und C57BL/6-Makrophagen.
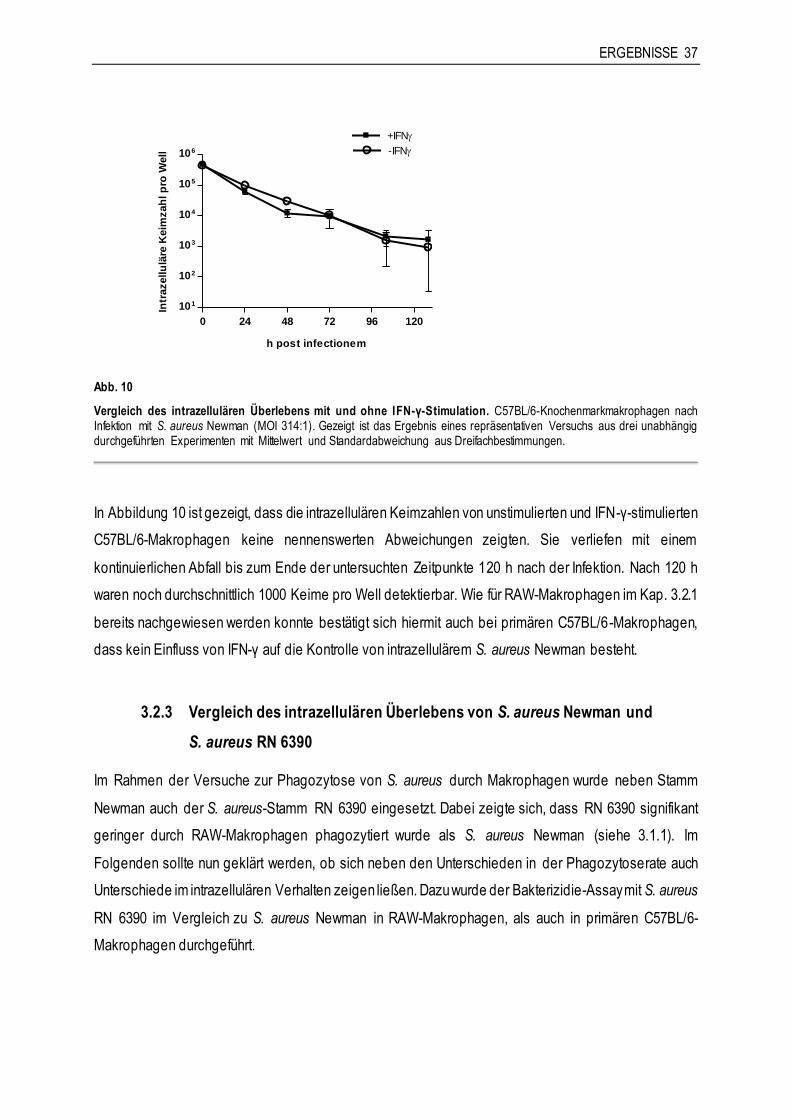
ERGEBNISSE 37
0 24 48 72 96 120
101
102
103
104
105
106
+IFN
-IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 10
Vergleich des intrazellulären Überlebens mit und ohne IFN-γ-Stimulation. C57BL/6-Knochenmarkmakrophagen nach Infektion mit S. aureus Newman (MOI 314:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In Abbildung 10 ist gezeigt, dass die intrazellulären Keimzahlen von unstimulierten und IFN-γ-stimulierten
C57BL/6-Makrophagen keine nennenswerten Abweichungen zeigten. Sie verliefen mit einem
kontinuierlichen Abfall bis zum Ende der untersuchten Zeitpunkte 120 h nach der Infektion. Nach 120 h
waren noch durchschnittlich 1000 Keime pro Well detektierbar. Wie für RAW-Makrophagen im Kap. 3.2.1
bereits nachgewiesen werden konnte bestätigt sich hiermit auch bei primären C57BL/6-Makrophagen,
dass kein Einfluss von IFN-γ auf die Kontrolle von intrazellulärem S. aureus Newman besteht.
3.2.3 Vergleich des intrazellulären Überlebens von S. aureus Newman und
S. aureus RN 6390
Im Rahmen der Versuche zur Phagozytose von S. aureus durch Makrophagen wurde neben Stamm
Newman auch der S. aureus-Stamm RN 6390 eingesetzt. Dabei zeigte sich, dass RN 6390 signifikant
geringer durch RAW-Makrophagen phagozytiert wurde als S. aureus Newman (siehe 3.1.1). Im
Folgenden sollte nun geklärt werden, ob sich neben den Unterschieden in der Phagozytoserate auch
Unterschiede im intrazellulären Verhalten zeigen ließen. Dazu wurde der Bakterizidie-Assay mit S. aureus
RN 6390 im Vergleich zu S. aureus Newman in RAW-Makrophagen, als auch in primären C57BL/6-
Makrophagen durchgeführt.
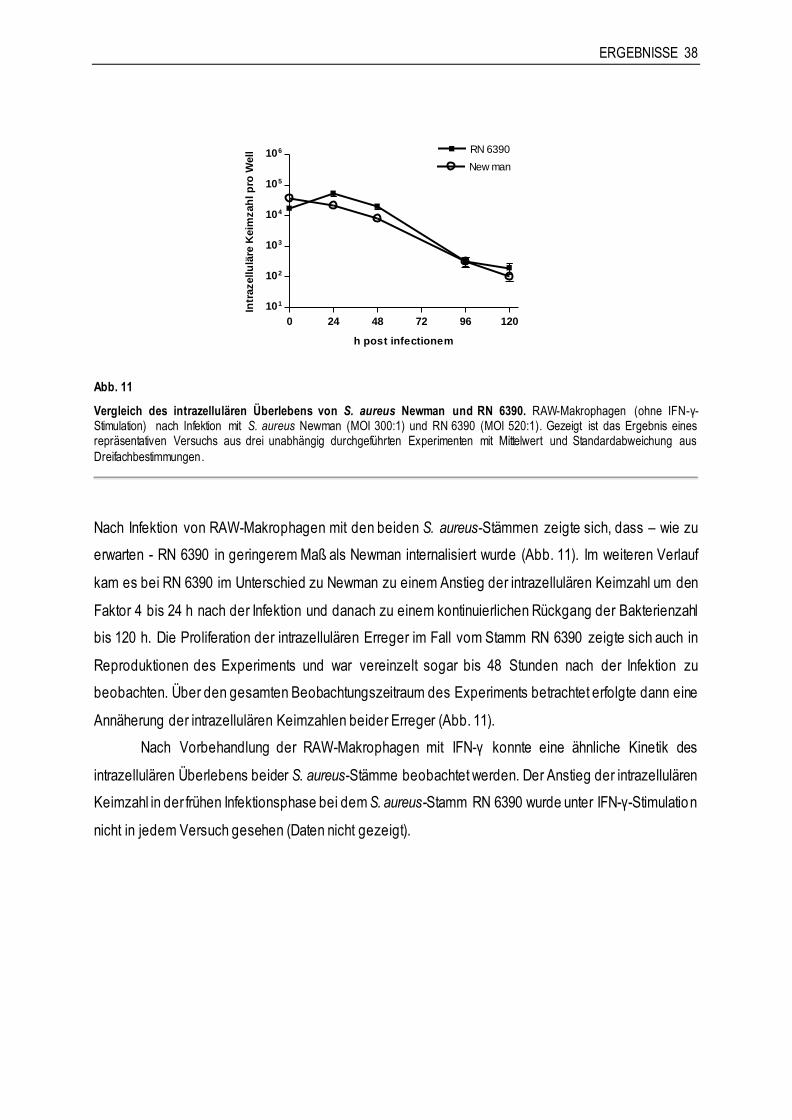
ERGEBNISSE 38
0 24 48 72 96 120
101
102
103
104
105
106 RN 6390
New man
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 11
Vergleich des intrazellulären Überlebens von S. aureus Newman und RN 6390. RAW-Makrophagen (ohne IFN-γ-Stimulation) nach Infektion mit S. aureus Newman (MOI 300:1) und RN 6390 (MOI 520:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus
Dreifachbestimmungen.
Nach Infektion von RAW-Makrophagen mit den beiden S. aureus-Stämmen zeigte sich, dass – wie zu
erwarten - RN 6390 in geringerem Maß als Newman internalisiert wurde (Abb. 11). Im weiteren Verlauf
kam es bei RN 6390 im Unterschied zu Newman zu einem Anstieg der intrazellulären Keimzahl um den
Faktor 4 bis 24 h nach der Infektion und danach zu einem kontinuierlichen Rückgang der Bakterienzahl
bis 120 h. Die Proliferation der intrazellulären Erreger im Fall vom Stamm RN 6390 zeigte sich auch in
Reproduktionen des Experiments und war vereinzelt sogar bis 48 Stunden nach der Infektion zu
beobachten. Über den gesamten Beobachtungszeitraum des Experiments betrachtet erfolgte dann eine
Annäherung der intrazellulären Keimzahlen beider Erreger (Abb. 11).
Nach Vorbehandlung der RAW-Makrophagen mit IFN-γ konnte eine ähnliche Kinetik des
intrazellulären Überlebens beider S. aureus-Stämme beobachtet werden. Der Anstieg der intrazellulären
Keimzahl in der frühen Infektionsphase bei dem S. aureus-Stamm RN 6390 wurde unter IFN-γ-Stimulation
nicht in jedem Versuch gesehen (Daten nicht gezeigt).
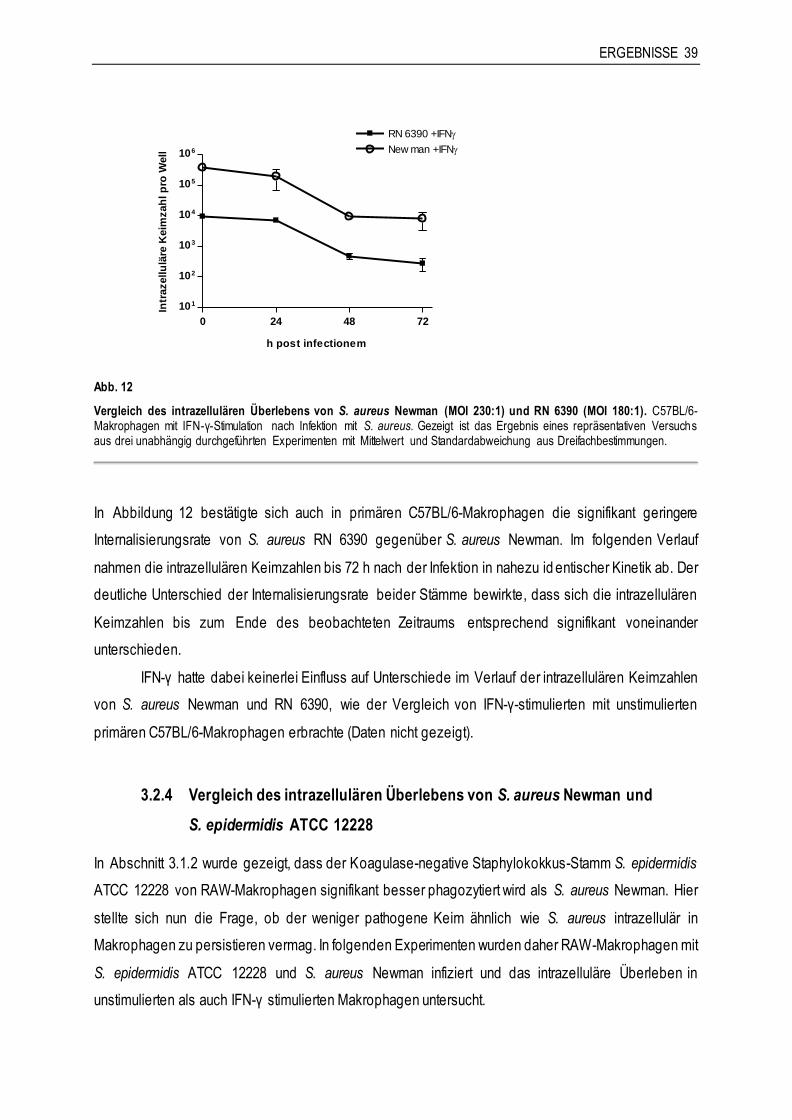
ERGEBNISSE 39
0 24 48 72
101
102
103
104
105
106 New man +IFN
RN 6390 +IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 12
Vergleich des intrazellulären Überlebens von S. aureus Newman (MOI 230:1) und RN 6390 (MOI 180:1). C57BL/6-Makrophagen mit IFN-γ-Stimulation nach Infektion mit S. aureus. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In Abbildung 12 bestätigte sich auch in primären C57BL/6-Makrophagen die signifikant geringere
Internalisierungsrate von S. aureus RN 6390 gegenüber S. aureus Newman. Im folgenden Verlauf
nahmen die intrazellulären Keimzahlen bis 72 h nach der Infektion in nahezu id entischer Kinetik ab. Der
deutliche Unterschied der Internalisierungsrate beider Stämme bewirkte, dass sich die intrazellulären
Keimzahlen bis zum Ende des beobachteten Zeitraums entsprechend signifikant voneinander
unterschieden.
IFN-γ hatte dabei keinerlei Einfluss auf Unterschiede im Verlauf der intrazellulären Keimzahlen
von S. aureus Newman und RN 6390, wie der Vergleich von IFN-γ-stimulierten mit unstimulierten
primären C57BL/6-Makrophagen erbrachte (Daten nicht gezeigt).
3.2.4 Vergleich des intrazellulären Überlebens von S. aureus Newman und
S. epidermidis ATCC 12228
In Abschnitt 3.1.2 wurde gezeigt, dass der Koagulase-negative Staphylokokkus-Stamm S. epidermidis
ATCC 12228 von RAW-Makrophagen signifikant besser phagozytiert wird als S. aureus Newman. Hier
stellte sich nun die Frage, ob der weniger pathogene Keim ähnlich wie S. aureus intrazellulär in
Makrophagen zu persistieren vermag. In folgenden Experimenten wurden daher RAW-Makrophagen mit
S. epidermidis ATCC 12228 und S. aureus Newman infiziert und das intrazelluläre Überleben in
unstimulierten als auch IFN-γ stimulierten Makrophagen untersucht.
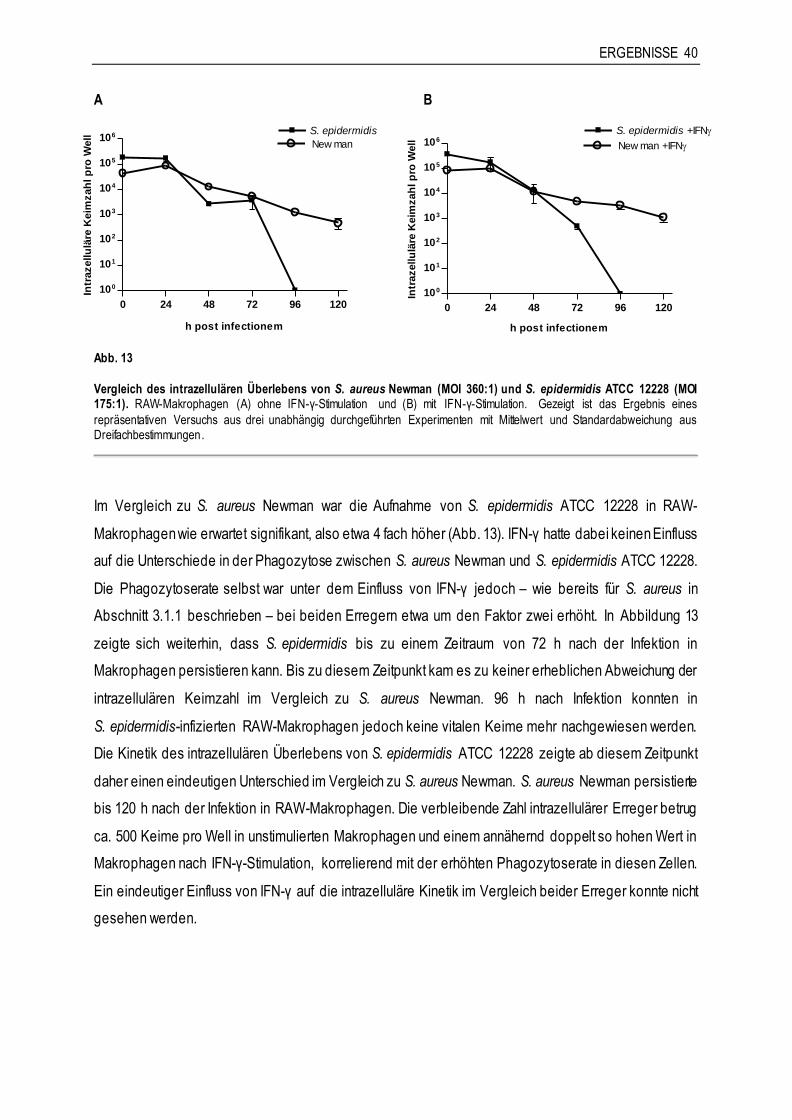
ERGEBNISSE 40
A B
0 24 48 72 96 120
100
101
102
103
104
105
106
New man
S. epidermidis
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48 72 96 120
100
101
102
103
104
105
106 New man +IFN
S. epidermidis +IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 13
Vergleich des intrazellulären Überlebens von S. aureus Newman (MOI 360:1) und S. epidermidis ATCC 12228 (MOI 175:1). RAW-Makrophagen (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist das Ergebnis eines
repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Im Vergleich zu S. aureus Newman war die Aufnahme von S. epidermidis ATCC 12228 in RAW-
Makrophagen wie erwartet signifikant, also etwa 4 fach höher (Abb. 13). IFN-γ hatte dabei keinen Einfluss
auf die Unterschiede in der Phagozytose zwischen S. aureus Newman und S. epidermidis ATCC 12228.
Die Phagozytoserate selbst war unter dem Einfluss von IFN-γ jedoch – wie bereits für S. aureus in
Abschnitt 3.1.1 beschrieben – bei beiden Erregern etwa um den Faktor zwei erhöht. In Abbildung 13
zeigte sich weiterhin, dass S. epidermidis bis zu einem Zeitraum von 72 h nach der Infektion in
Makrophagen persistieren kann. Bis zu diesem Zeitpunkt kam es zu keiner erheblichen Abweichung der
intrazellulären Keimzahl im Vergleich zu S. aureus Newman. 96 h nach Infektion konnten in
S. epidermidis-infizierten RAW-Makrophagen jedoch keine vitalen Keime mehr nachgewiesen werden.
Die Kinetik des intrazellulären Überlebens von S. epidermidis ATCC 12228 zeigte ab diesem Zeitpunkt
daher einen eindeutigen Unterschied im Vergleich zu S. aureus Newman. S. aureus Newman persistierte
bis 120 h nach der Infektion in RAW-Makrophagen. Die verbleibende Zahl intrazellulärer Erreger betrug
ca. 500 Keime pro Well in unstimulierten Makrophagen und einem annähernd doppelt so hohen Wert in
Makrophagen nach IFN-γ-Stimulation, korrelierend mit der erhöhten Phagozytoserate in diesen Zellen.
Ein eindeutiger Einfluss von IFN-γ auf die intrazelluläre Kinetik im Vergleich beider Erreger konnte nicht
gesehen werden.
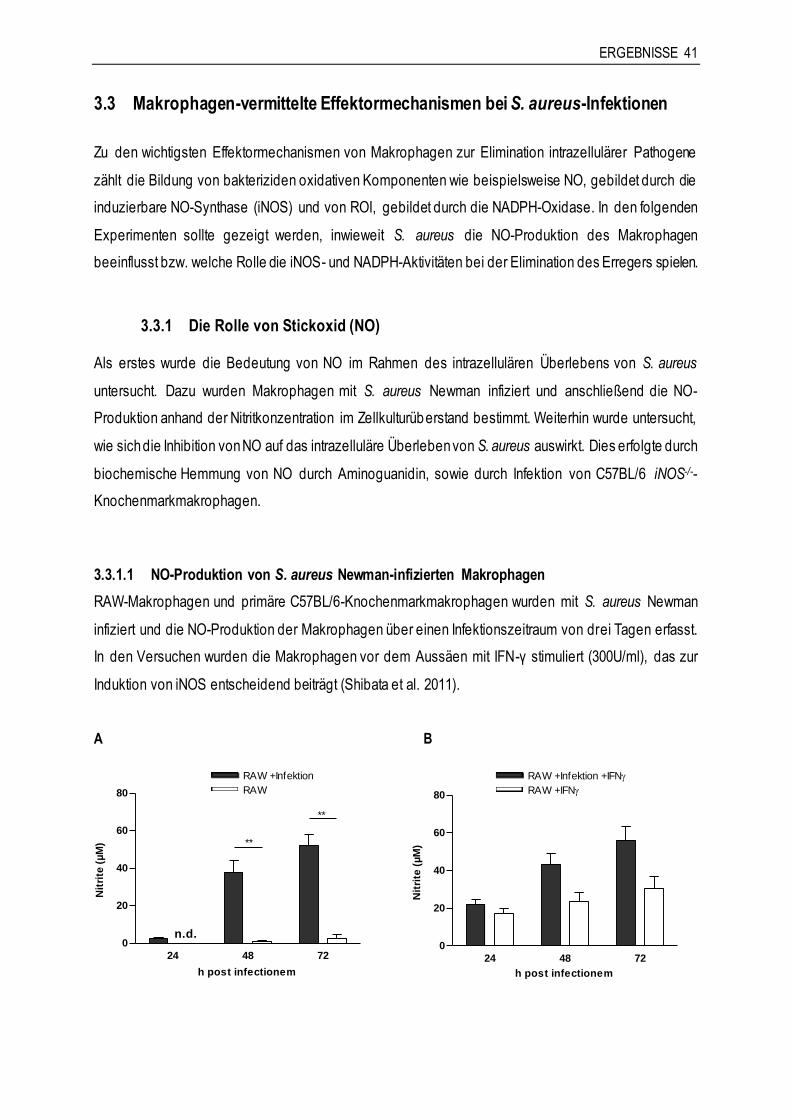
ERGEBNISSE 41
3.3 Makrophagen-vermittelte Effektormechanismen bei S. aureus-Infektionen
Zu den wichtigsten Effektormechanismen von Makrophagen zur Elimination intrazellulärer Pathogene
zählt die Bildung von bakteriziden oxidativen Komponenten wie beispielsweise NO, gebildet durch die
induzierbare NO-Synthase (iNOS) und von ROI, gebildet durch die NADPH-Oxidase. In den folgenden
Experimenten sollte gezeigt werden, inwieweit S. aureus die NO-Produktion des Makrophagen
beeinflusst bzw. welche Rolle die iNOS- und NADPH-Aktivitäten bei der Elimination des Erregers spielen.
3.3.1 Die Rolle von Stickoxid (NO)
Als erstes wurde die Bedeutung von NO im Rahmen des intrazellulären Überlebens von S. aureus
untersucht. Dazu wurden Makrophagen mit S. aureus Newman infiziert und anschließend die NO-
Produktion anhand der Nitritkonzentration im Zellkulturüberstand bestimmt. Weiterhin wurde untersucht,
wie sich die Inhibition von NO auf das intrazelluläre Überleben von S. aureus auswirkt. Dies erfolgte durch
biochemische Hemmung von NO durch Aminoguanidin, sowie durch Infektion von C57BL/6 iNOS-/--
Knochenmarkmakrophagen.
3.3.1.1 NO-Produktion von S. aureus Newman-infizierten Makrophagen
RAW-Makrophagen und primäre C57BL/6-Knochenmarkmakrophagen wurden mit S. aureus Newman
infiziert und die NO-Produktion der Makrophagen über einen Infektionszeitraum von drei Tagen erfasst.
In den Versuchen wurden die Makrophagen vor dem Aussäen mit IFN-γ stimuliert (300U/ml), das zur
Induktion von iNOS entscheidend beiträgt (Shibata et al. 2011).
A B
24 48 72
0
20
40
60
80
RAW +Infektion
RAW
n.d.
**
**
h post infectionem
Nit
rite
(µM
)
24 48 72
0
20
40
60
80
RAW +Infektion +IFN
RAW +IFN
h post infectionem
Nit
rite
(µM
)
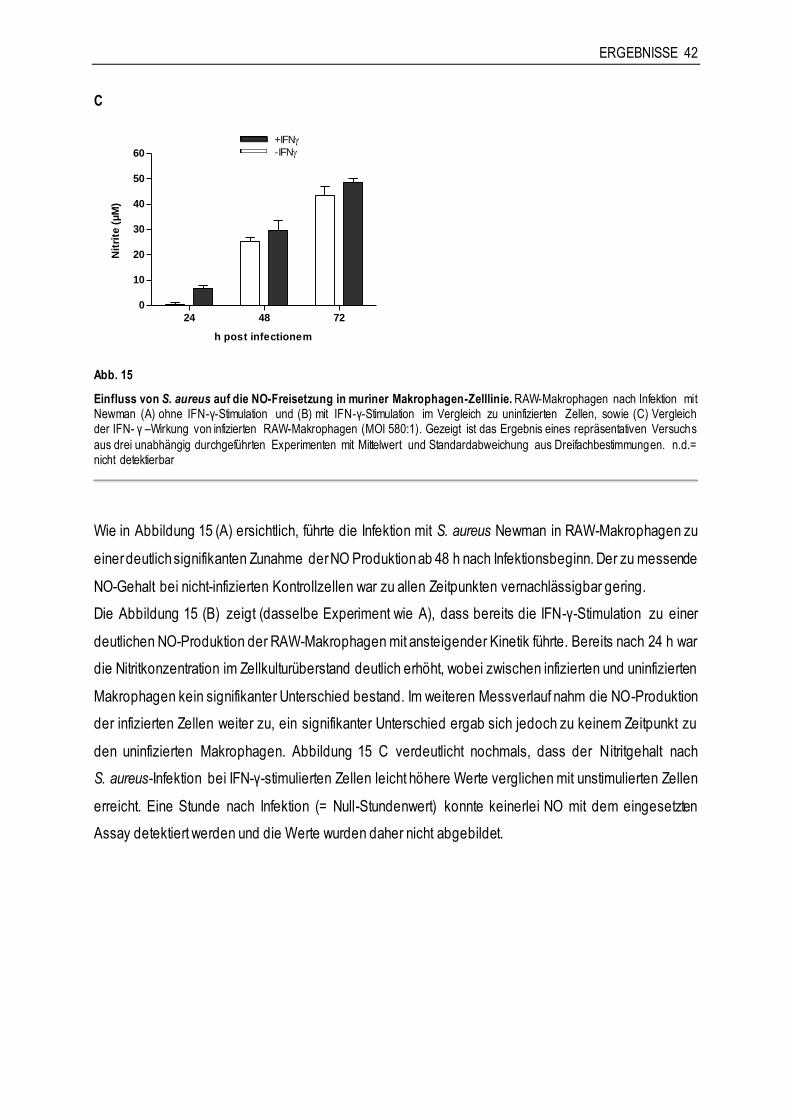
ERGEBNISSE 42
C
24 48 72
0
10
20
30
40
50
60 -IFN
+IFN
h post infectionem
Nit
rite
(µM
)
Abb. 15
Einfluss von S. aureus auf die NO-Freisetzung in muriner Makrophagen-Zelllinie. RAW-Makrophagen nach Infektion mit Newman (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation im Vergleich zu uninfizierten Zellen, sowie (C) Vergleich der IFN- γ –Wirkung von infizierten RAW-Makrophagen (MOI 580:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs
aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen. n.d.= nicht detektierbar
Wie in Abbildung 15 (A) ersichtlich, führte die Infektion mit S. aureus Newman in RAW-Makrophagen zu
einer deutlich signifikanten Zunahme der NO Produktion ab 48 h nach Infektionsbeginn. Der zu messende
NO-Gehalt bei nicht-infizierten Kontrollzellen war zu allen Zeitpunkten vernachlässigbar gering.
Die Abbildung 15 (B) zeigt (dasselbe Experiment wie A), dass bereits die IFN-γ-Stimulation zu einer
deutlichen NO-Produktion der RAW-Makrophagen mit ansteigender Kinetik führte. Bereits nach 24 h war
die Nitritkonzentration im Zellkulturüberstand deutlich erhöht, wobei zwischen infizierten und uninfizierten
Makrophagen kein signifikanter Unterschied bestand. Im weiteren Messverlauf nahm die NO-Produktion
der infizierten Zellen weiter zu, ein signifikanter Unterschied ergab sich jedoch zu keinem Zeitpunkt zu
den uninfizierten Makrophagen. Abbildung 15 C verdeutlicht nochmals, dass der Nitritgehalt nach
S. aureus-Infektion bei IFN-γ-stimulierten Zellen leicht höhere Werte verglichen mit unstimulierten Zellen
erreicht. Eine Stunde nach Infektion (= Null-Stundenwert) konnte keinerlei NO mit dem eingesetzten
Assay detektiert werden und die Werte wurden daher nicht abgebildet.
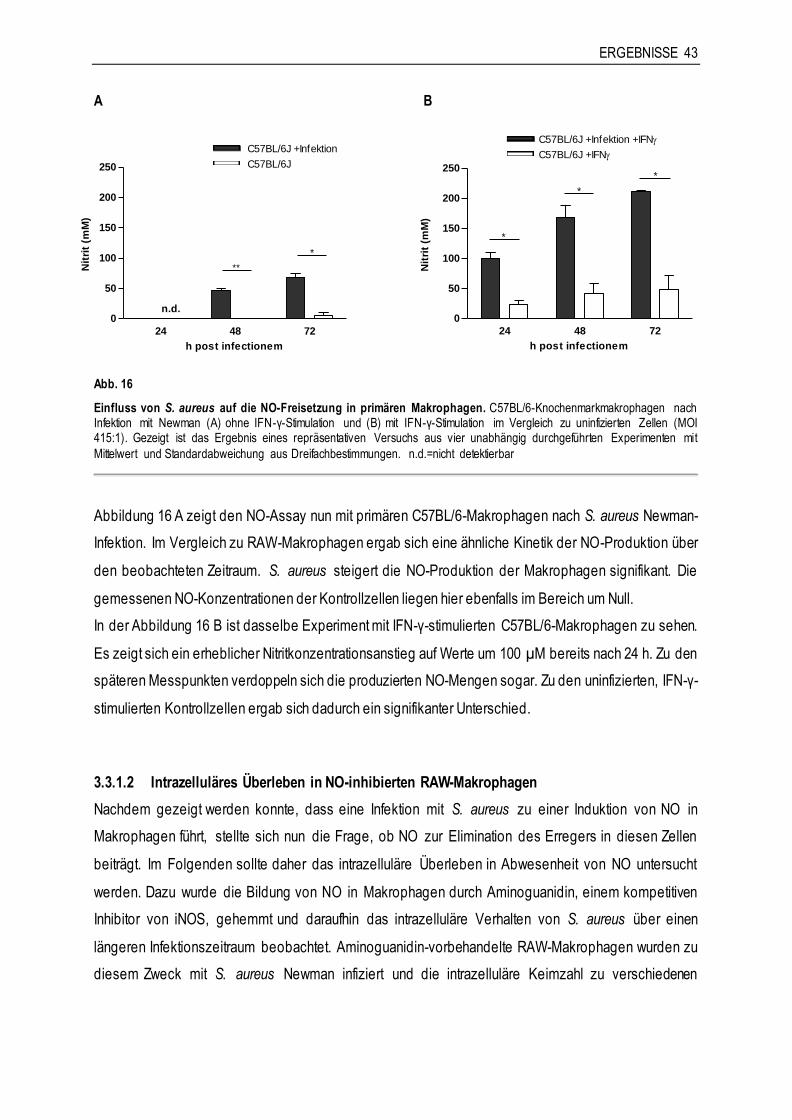
ERGEBNISSE 43
A B
24 48 72
0
50
100
150
200
250
C57BL/6J +Infektion
C57BL/6J
n.d.
**
*
h post infectionem
Nit
rit
(mM
)
24 48 72
0
50
100
150
200
250
C57BL/6J +Infektion +IFN
C57BL/6J +IFN
*
*
*
h post infectionem
Nit
rit
(mM
)
Abb. 16
Einfluss von S. aureus auf die NO-Freisetzung in primären Makrophagen. C57BL/6-Knochenmarkmakrophagen nach Infektion mit Newman (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation im Vergleich zu uninfizierten Zellen (MOI 415:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus vier unabhängig durchgeführten Experimenten mit
Mittelwert und Standardabweichung aus Dreifachbestimmungen. n.d.=nicht detektierbar
Abbildung 16 A zeigt den NO-Assay nun mit primären C57BL/6-Makrophagen nach S. aureus Newman-
Infektion. Im Vergleich zu RAW-Makrophagen ergab sich eine ähnliche Kinetik der NO-Produktion über
den beobachteten Zeitraum. S. aureus steigert die NO-Produktion der Makrophagen signifikant. Die
gemessenen NO-Konzentrationen der Kontrollzellen liegen hier ebenfalls im Bereich um Null.
In der Abbildung 16 B ist dasselbe Experiment mit IFN-γ-stimulierten C57BL/6-Makrophagen zu sehen.
Es zeigt sich ein erheblicher Nitritkonzentrationsanstieg auf Werte um 100 µM bereits nach 24 h. Zu den
späteren Messpunkten verdoppeln sich die produzierten NO-Mengen sogar. Zu den uninfizierten, IFN-γ-
stimulierten Kontrollzellen ergab sich dadurch ein signifikanter Unterschied.
3.3.1.2 Intrazelluläres Überleben in NO-inhibierten RAW-Makrophagen
Nachdem gezeigt werden konnte, dass eine Infektion mit S. aureus zu einer Induktion von NO in
Makrophagen führt, stellte sich nun die Frage, ob NO zur Elimination des Erregers in diesen Zellen
beiträgt. Im Folgenden sollte daher das intrazelluläre Überleben in Abwesenheit von NO untersucht
werden. Dazu wurde die Bildung von NO in Makrophagen durch Aminoguanidin, einem kompetitiven
Inhibitor von iNOS, gehemmt und daraufhin das intrazelluläre Verhalten von S. aureus über einen
längeren Infektionszeitraum beobachtet. Aminoguanidin-vorbehandelte RAW-Makrophagen wurden zu
diesem Zweck mit S. aureus Newman infiziert und die intrazelluläre Keimzahl zu verschiedenen
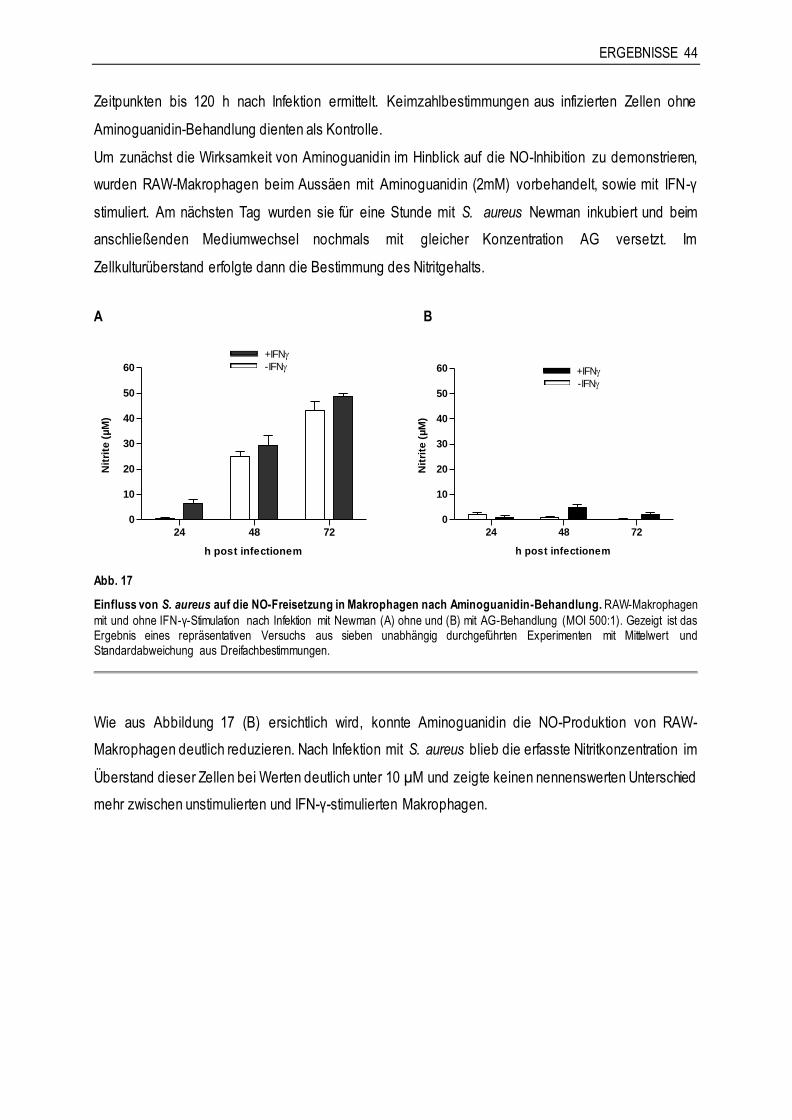
ERGEBNISSE 44
Zeitpunkten bis 120 h nach Infektion ermittelt. Keimzahlbestimmungen aus infizierten Zellen ohne
Aminoguanidin-Behandlung dienten als Kontrolle.
Um zunächst die Wirksamkeit von Aminoguanidin im Hinblick auf die NO-Inhibition zu demonstrieren,
wurden RAW-Makrophagen beim Aussäen mit Aminoguanidin (2mM) vorbehandelt, sowie mit IFN-γ
stimuliert. Am nächsten Tag wurden sie für eine Stunde mit S. aureus Newman inkubiert und beim
anschließenden Mediumwechsel nochmals mit gleicher Konzentration AG versetzt. Im
Zellkulturüberstand erfolgte dann die Bestimmung des Nitritgehalts.
A B
24 48 72
0
10
20
30
40
50
60 -IFN
+IFN
h post infectionem
Nit
rite
(µM
)
24 48 72
0
10
20
30
40
50
60
-IFN
+IFN
h post infectionem
Nit
rite
(µM
)
Abb. 17
Einfluss von S. aureus auf die NO-Freisetzung in Makrophagen nach Aminoguanidin-Behandlung. RAW-Makrophagen
mit und ohne IFN-γ-Stimulation nach Infektion mit Newman (A) ohne und (B) mit AG-Behandlung (MOI 500:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus sieben unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Wie aus Abbildung 17 (B) ersichtlich wird, konnte Aminoguanidin die NO-Produktion von RAW-
Makrophagen deutlich reduzieren. Nach Infektion mit S. aureus blieb die erfasste Nitritkonzentration im
Überstand dieser Zellen bei Werten deutlich unter 10 µM und zeigte keinen nennenswerten Unterschied
mehr zwischen unstimulierten und IFN-γ-stimulierten Makrophagen.
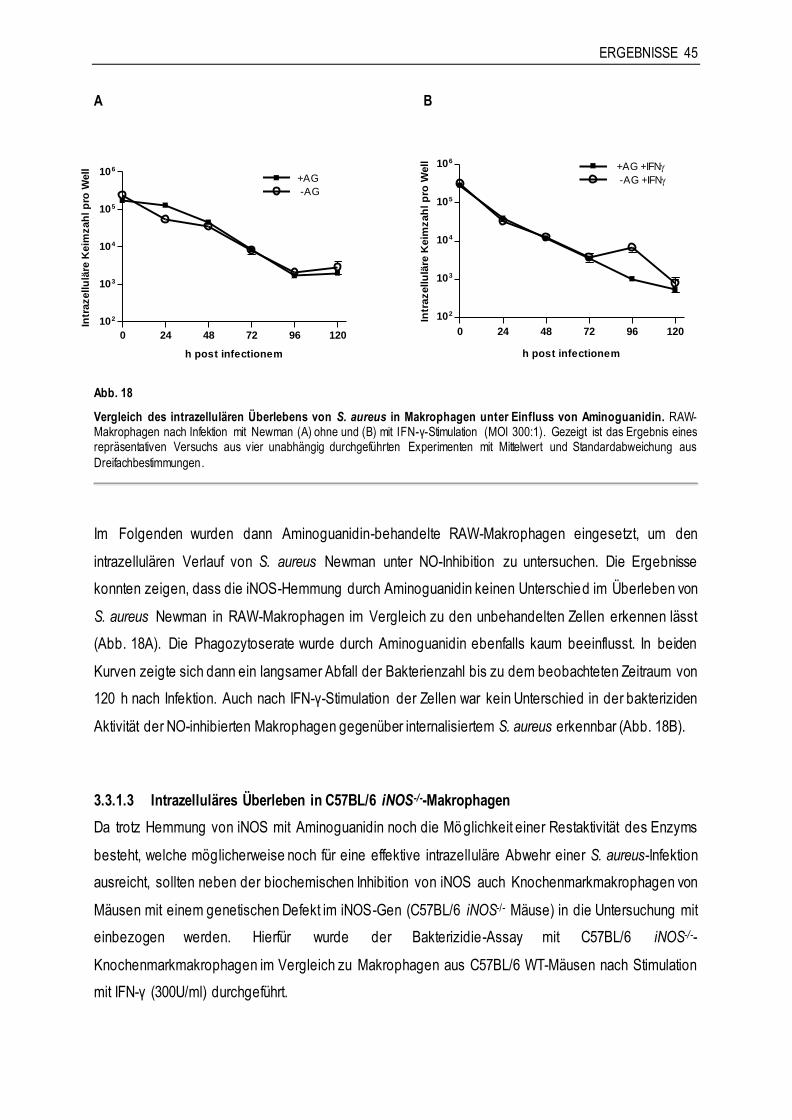
ERGEBNISSE 45
A B
0 24 48 72 96 120
102
103
104
105
106
+AG
-AG
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48 72 96 120
102
103
104
105
106 +AG +IFN
-AG +IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 18
Vergleich des intrazellulären Überlebens von S. aureus in Makrophagen unter Einfluss von Aminoguanidin. RAW-Makrophagen nach Infektion mit Newman (A) ohne und (B) mit IFN-γ-Stimulation (MOI 300:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus vier unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus
Dreifachbestimmungen.
Im Folgenden wurden dann Aminoguanidin-behandelte RAW-Makrophagen eingesetzt, um den
intrazellulären Verlauf von S. aureus Newman unter NO-Inhibition zu untersuchen. Die Ergebnisse
konnten zeigen, dass die iNOS-Hemmung durch Aminoguanidin keinen Unterschied im Überleben von
S. aureus Newman in RAW-Makrophagen im Vergleich zu den unbehandelten Zellen erkennen lässt
(Abb. 18A). Die Phagozytoserate wurde durch Aminoguanidin ebenfalls kaum beeinflusst. In beiden
Kurven zeigte sich dann ein langsamer Abfall der Bakterienzahl bis zu dem beobachteten Zeitraum von
120 h nach Infektion. Auch nach IFN-γ-Stimulation der Zellen war kein Unterschied in der bakteriziden
Aktivität der NO-inhibierten Makrophagen gegenüber internalisiertem S. aureus erkennbar (Abb. 18B).
3.3.1.3 Intrazelluläres Überleben in C57BL/6 iNOS-/--Makrophagen
Da trotz Hemmung von iNOS mit Aminoguanidin noch die Möglichkeit einer Restaktivität des Enzyms
besteht, welche möglicherweise noch für eine effektive intrazelluläre Abwehr einer S. aureus-Infektion
ausreicht, sollten neben der biochemischen Inhibition von iNOS auch Knochenmarkmakrophagen von
Mäusen mit einem genetischen Defekt im iNOS-Gen (C57BL/6 iNOS-/- Mäuse) in die Untersuchung mit
einbezogen werden. Hierfür wurde der Bakterizidie-Assay mit C57BL/6 iNOS-/--
Knochenmarkmakrophagen im Vergleich zu Makrophagen aus C57BL/6 WT-Mäusen nach Stimulation
mit IFN-γ (300U/ml) durchgeführt.
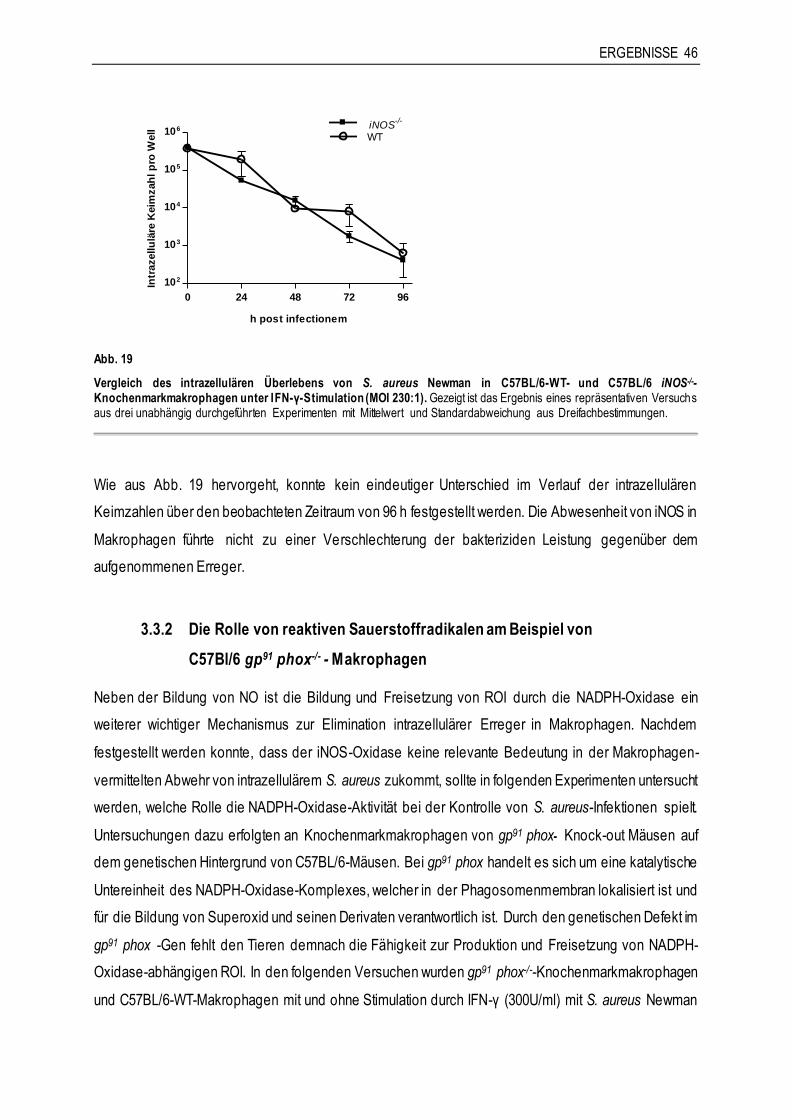
ERGEBNISSE 46
0 24 48 72 96
102
103
104
105
106
WT iNOS
-/-
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 19
Vergleich des intrazellulären Überlebens von S. aureus Newman in C57BL/6-WT- und C57BL/6 iNOS-/-- Knochenmarkmakrophagen unter IFN-γ-Stimulation (MOI 230:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Wie aus Abb. 19 hervorgeht, konnte kein eindeutiger Unterschied im Verlauf der intrazellulären
Keimzahlen über den beobachteten Zeitraum von 96 h festgestellt werden. Die Abwesenheit von iNOS in
Makrophagen führte nicht zu einer Verschlechterung der bakteriziden Leistung gegenüber dem
aufgenommenen Erreger.
3.3.2 Die Rolle von reaktiven Sauerstoffradikalen am Beispiel von
C57Bl/6 gp91 phox-/- - Makrophagen
Neben der Bildung von NO ist die Bildung und Freisetzung von ROI durch die NADPH-Oxidase ein
weiterer wichtiger Mechanismus zur Elimination intrazellulärer Erreger in Makrophagen. Nachdem
festgestellt werden konnte, dass der iNOS-Oxidase keine relevante Bedeutung in der Makrophagen-
vermittelten Abwehr von intrazellulärem S. aureus zukommt, sollte in folgenden Experimenten untersucht
werden, welche Rolle die NADPH-Oxidase-Aktivität bei der Kontrolle von S. aureus-Infektionen spielt.
Untersuchungen dazu erfolgten an Knochenmarkmakrophagen von gp91 phox- Knock-out Mäusen auf
dem genetischen Hintergrund von C57BL/6-Mäusen. Bei gp91 phox handelt es sich um eine katalytische
Untereinheit des NADPH-Oxidase-Komplexes, welcher in der Phagosomenmembran lokalisiert ist und
für die Bildung von Superoxid und seinen Derivaten verantwortlich ist. Durch den genetischen Defekt im
gp91 phox -Gen fehlt den Tieren demnach die Fähigkeit zur Produktion und Freisetzung von NADPH-
Oxidase-abhängigen ROI. In den folgenden Versuchen wurden gp91 phox-/--Knochenmarkmakrophagen
und C57BL/6-WT-Makrophagen mit und ohne Stimulation durch IFN-γ (300U/ml) mit S. aureus Newman
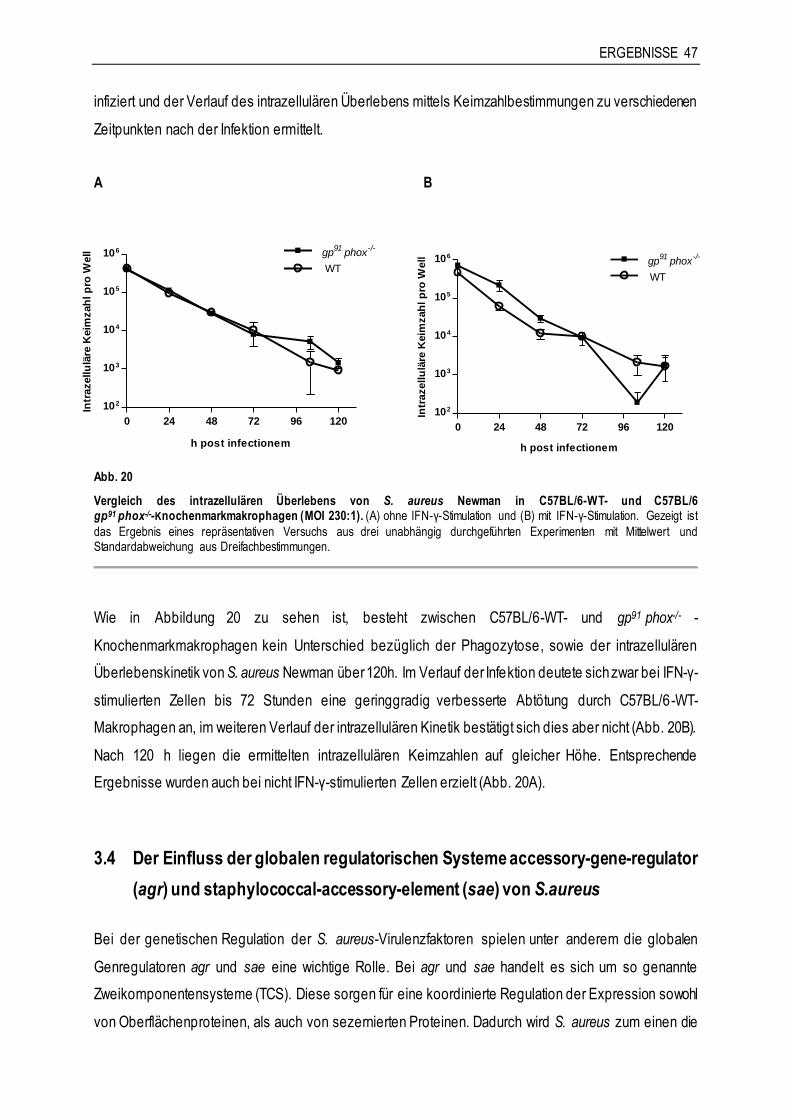
ERGEBNISSE 47
infiziert und der Verlauf des intrazellulären Überlebens mittels Keimzahlbestimmungen zu verschiedenen
Zeitpunkten nach der Infektion ermittelt.
A B
0 24 48 72 96 120
102
103
104
105
106
WT
gp91
phox-/-
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48 72 96 120
102
103
104
105
106
WT
gp91
phox-/-
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 20
Vergleich des intrazellulären Überlebens von S. aureus Newman in C57BL/6-WT- und C57BL/6 gp91 phox-/--Knochenmarkmakrophagen (MOI 230:1). (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist
das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Wie in Abbildung 20 zu sehen ist, besteht zwischen C57BL/6-WT- und gp91 phox-/- -
Knochenmarkmakrophagen kein Unterschied bezüglich der Phagozytose, sowie der intrazellulären
Überlebenskinetik von S. aureus Newman über 120h. Im Verlauf der Infektion deutete sich zwar bei IFN-γ-
stimulierten Zellen bis 72 Stunden eine geringgradig verbesserte Abtötung durch C57BL/6-WT-
Makrophagen an, im weiteren Verlauf der intrazellulären Kinetik bestätigt sich dies aber nicht (Abb. 20B).
Nach 120 h liegen die ermittelten intrazellulären Keimzahlen auf gleicher Höhe. Entsprechende
Ergebnisse wurden auch bei nicht IFN-γ-stimulierten Zellen erzielt (Abb. 20A).
3.4 Der Einfluss der globalen regulatorischen Systeme accessory-gene-regulator
(agr) und staphylococcal-accessory-element (sae) von S.aureus
Bei der genetischen Regulation der S. aureus-Virulenzfaktoren spielen unter anderem die globalen
Genregulatoren agr und sae eine wichtige Rolle. Bei agr und sae handelt es sich um so genannte
Zweikomponentensysteme (TCS). Diese sorgen für eine koordinierte Regulation der Expression sowohl
von Oberflächenproteinen, als auch von sezernierten Proteinen. Dadurch wird S. aureus zum einen die

ERGEBNISSE 48
Adhäsion und Internalisierung ermöglicht, zum anderen werden zytotoxische Eigenschaften bereitgestellt,
um den bakteriziden Mechanismen der Zelle zu entgehen. In diesem Kapitel sollte daher die Bedeutung
der globalen Regulatoren agr und sae für Phagozytose und intrazelluläres Überleben von S. aureus in
Makrophagen untersucht werden. Dies erfolgte unter Einbeziehung zweier Mutanten mit funktionell
inaktivem agr- bzw. sae-Regulator: S. aureus Newman (sae) und S. aureus RN 6911 (agr).
3.4.1 Die Rolle von agr für die Interaktion von S. aureus RN 6390 mit
Makrophagen
Das Zweikomponentensystem agr reguliert viele wichtige Virulenzfaktoren von S. aureus. Es wird in der
späten Wachstumsphase der Bakterien induziert und führt zu einer Produktion sezernierter Proteine, wie
z. B. Hämolysine. Auf der anderen Seite bewirkt es eine verminderte Synthese Zellwand-assoziierter
Moleküle, z. B. FnBP. In Experimenten mit Makrophagen, die mit der agr-Mutante von S. aureus RN 6390
infiziert wurden, sollte die Frage geklärt werden, inwieweit dieser Regulator die Internalisierung und das
intrazelluläre Verhalten von S. aureus in Makrophagen beeinflusst.
3.4.1.1 Einfluss des agr-Regulators auf die Phagozytose
Um die Bedeutung von agr auf Eindringungsvermögen von S. aureus zu untersuchen, wurden die
Phagozytoseraten von RAW-Makrophagen nach Infektion mit S. aureus RN 6911agr - und dem WT-
Stamm RN 6390 verglichen. Der Phagozytose-Assay wurde mit unstimulierten und IFN-γ-stimulierten
Zellen durchgeführt.
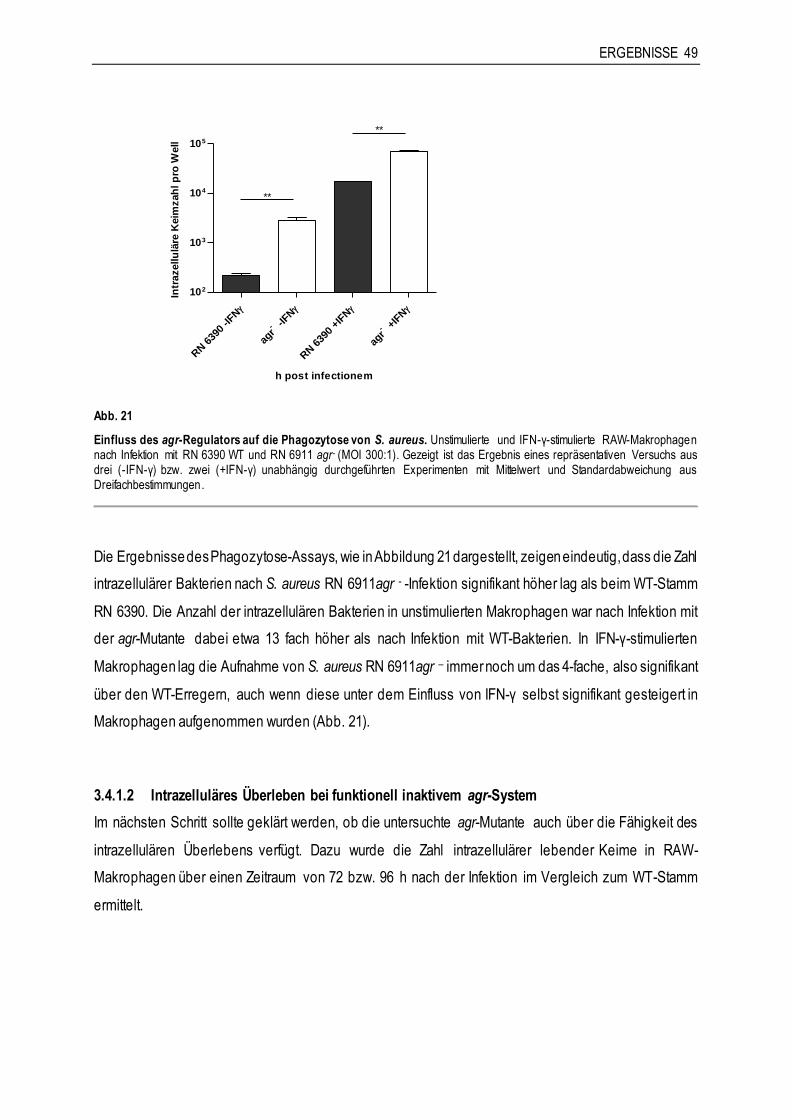
ERGEBNISSE 49
RN 6
390
-IFN
-IF
N-
agr
IFN
+
RN 6
390
+IF
N
-
agr
102
103
104
105
**
**
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 21
Einfluss des agr-Regulators auf die Phagozytose von S. aureus. Unstimulierte und IFN-γ-stimulierte RAW-Makrophagen nach Infektion mit RN 6390 WT und RN 6911 agr- (MOI 300:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei (-IFN-γ) bzw. zwei (+IFN-γ) unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Die Ergebnisse des Phagozytose-Assays, wie in Abbildung 21 dargestellt, zeigen eindeutig, dass die Zahl
intrazellulärer Bakterien nach S. aureus RN 6911agr - -Infektion signifikant höher lag als beim WT-Stamm
RN 6390. Die Anzahl der intrazellulären Bakterien in unstimulierten Makrophagen war nach Infektion mit
der agr-Mutante dabei etwa 13 fach höher als nach Infektion mit WT-Bakterien. In IFN-γ-stimulierten
Makrophagen lag die Aufnahme von S. aureus RN 6911agr – immer noch um das 4-fache, also signifikant
über den WT-Erregern, auch wenn diese unter dem Einfluss von IFN-γ selbst signifikant gesteigert in
Makrophagen aufgenommen wurden (Abb. 21).
3.4.1.2 Intrazelluläres Überleben bei funktionell inaktivem agr-System
Im nächsten Schritt sollte geklärt werden, ob die untersuchte agr-Mutante auch über die Fähigkeit des
intrazellulären Überlebens verfügt. Dazu wurde die Zahl intrazellulärer lebender Keime in RAW-
Makrophagen über einen Zeitraum von 72 bzw. 96 h nach der Infektion im Vergleich zum WT-Stamm
ermittelt.
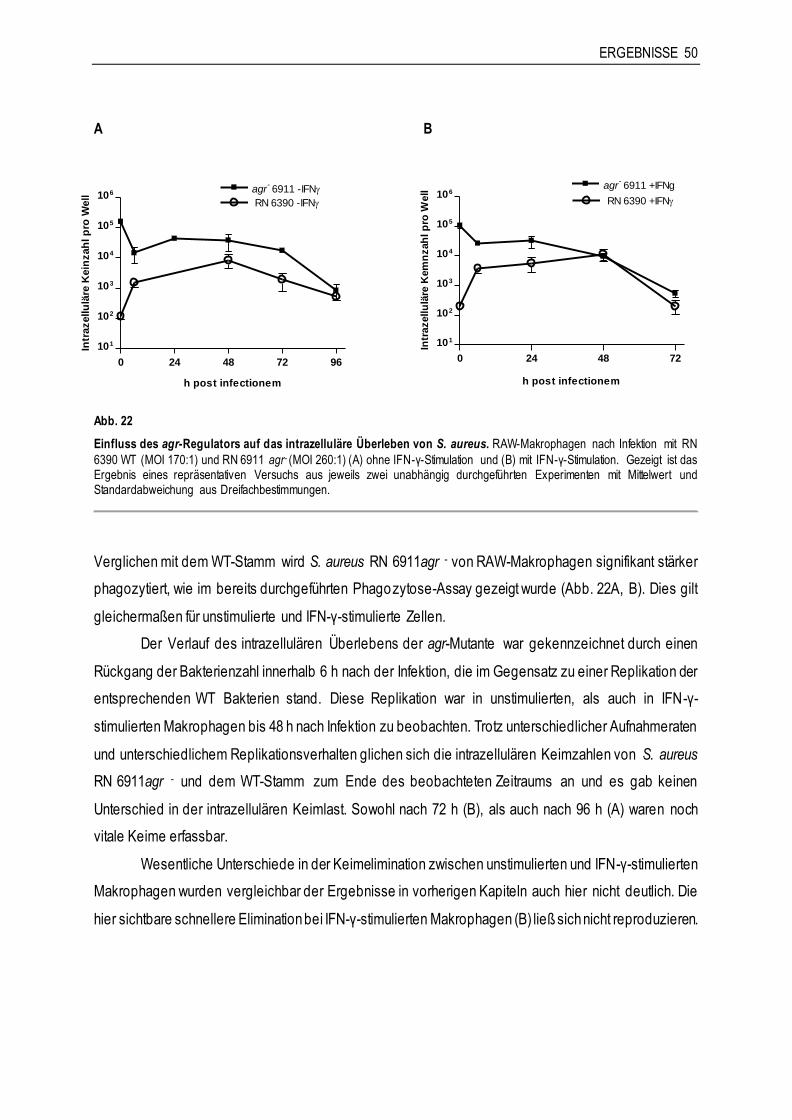
ERGEBNISSE 50
A B
0 24 48 72 96
101
102
103
104
105
106
RN 6390 -IFN
agr-6911 -IFN
h post infectionem
Intr
azellu
läre
Kein
zah
l p
ro W
ell
0 24 48 72
101
102
103
104
105
106
RN 6390 +IFN
agr-6911 +IFNg
h post infectionem
Intr
azellu
läre
Kem
nzah
l p
ro W
ell
Abb. 22
Einfluss des agr-Regulators auf das intrazelluläre Überleben von S. aureus. RAW-Makrophagen nach Infektion mit RN
6390 WT (MOI 170:1) und RN 6911 agr- (MOI 260:1) (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus jeweils zwei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Verglichen mit dem WT-Stamm wird S. aureus RN 6911agr - von RAW-Makrophagen signifikant stärker
phagozytiert, wie im bereits durchgeführten Phagozytose-Assay gezeigt wurde (Abb. 22A, B). Dies gilt
gleichermaßen für unstimulierte und IFN-γ-stimulierte Zellen.
Der Verlauf des intrazellulären Überlebens der agr-Mutante war gekennzeichnet durch einen
Rückgang der Bakterienzahl innerhalb 6 h nach der Infektion, die im Gegensatz zu einer Replikation der
entsprechenden WT Bakterien stand. Diese Replikation war in unstimulierten, als auch in IFN-γ-
stimulierten Makrophagen bis 48 h nach Infektion zu beobachten. Trotz unterschiedlicher Aufnahmeraten
und unterschiedlichem Replikationsverhalten glichen sich die intrazellulären Keimzahlen von S. aureus
RN 6911agr - und dem WT-Stamm zum Ende des beobachteten Zeitraums an und es gab keinen
Unterschied in der intrazellulären Keimlast. Sowohl nach 72 h (B), als auch nach 96 h (A) waren noch
vitale Keime erfassbar.
Wesentliche Unterschiede in der Keimelimination zwischen unstimulierten und IFN-γ-stimulierten
Makrophagen wurden vergleichbar der Ergebnisse in vorherigen Kapiteln auch hier nicht deutlich. Die
hier sichtbare schnellere Elimination bei IFN-γ-stimulierten Makrophagen (B) ließ sich nicht reproduzieren.
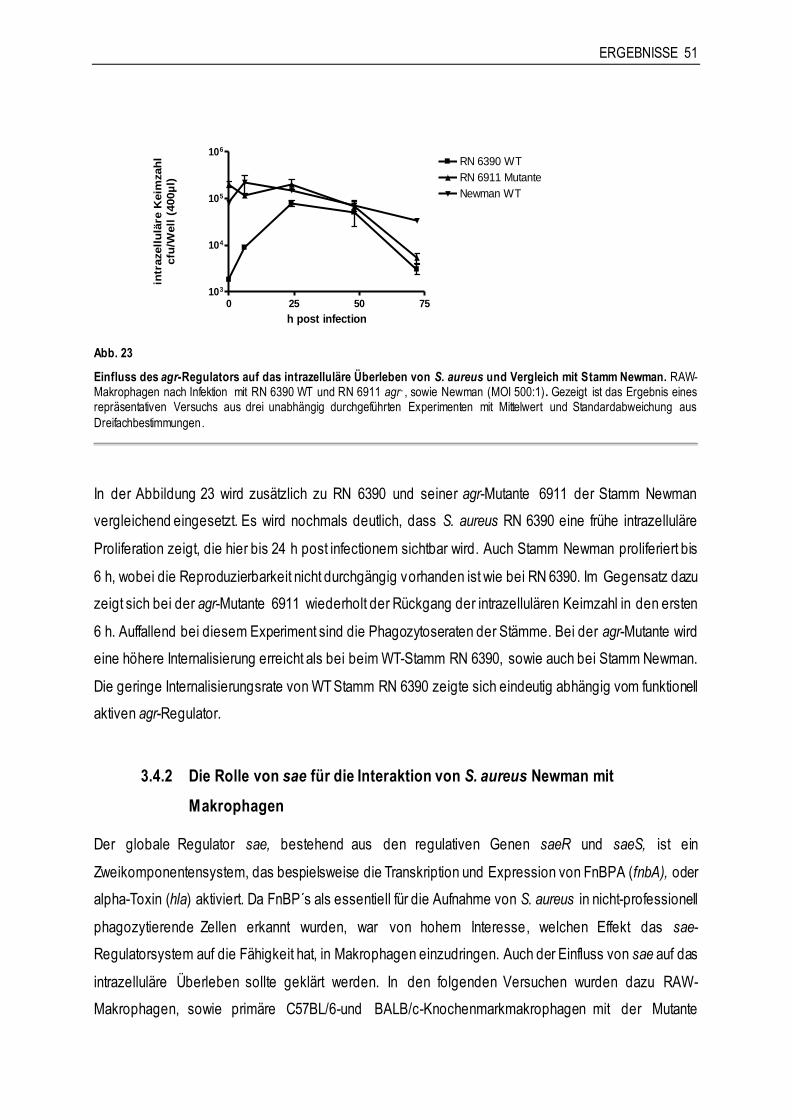
ERGEBNISSE 51
Abb. 23
Einfluss des agr-Regulators auf das intrazelluläre Überleben von S. aureus und Vergleich mit Stamm Newman. RAW-Makrophagen nach Infektion mit RN 6390 WT und RN 6911 agr- , sowie Newman (MOI 500:1). Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus drei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus
Dreifachbestimmungen.
In der Abbildung 23 wird zusätzlich zu RN 6390 und seiner agr-Mutante 6911 der Stamm Newman
vergleichend eingesetzt. Es wird nochmals deutlich, dass S. aureus RN 6390 eine frühe intrazelluläre
Proliferation zeigt, die hier bis 24 h post infectionem sichtbar wird. Auch Stamm Newman proliferiert bis
6 h, wobei die Reproduzierbarkeit nicht durchgängig vorhanden ist wie bei RN 6390. Im Gegensatz dazu
zeigt sich bei der agr-Mutante 6911 wiederholt der Rückgang der intrazellulären Keimzahl in den ersten
6 h. Auffallend bei diesem Experiment sind die Phagozytoseraten der Stämme. Bei der agr-Mutante wird
eine höhere Internalisierung erreicht als bei beim WT-Stamm RN 6390, sowie auch bei Stamm Newman.
Die geringe Internalisierungsrate von WT Stamm RN 6390 zeigte sich eindeutig abhängig vom funktionell
aktiven agr-Regulator.
3.4.2 Die Rolle von sae für die Interaktion von S. aureus Newman mit
Makrophagen
Der globale Regulator sae, bestehend aus den regulativen Genen saeR und saeS, ist ein
Zweikomponentensystem, das bespielsweise die Transkription und Expression von FnBPA (fnbA), oder
alpha-Toxin (hla) aktiviert. Da FnBP´s als essentiell für die Aufnahme von S. aureus in nicht-professionell
phagozytierende Zellen erkannt wurden, war von hohem Interesse, welchen Effekt das sae-
Regulatorsystem auf die Fähigkeit hat, in Makrophagen einzudringen. Auch der Einfluss von sae auf das
intrazelluläre Überleben sollte geklärt werden. In den folgenden Versuchen wurden dazu RAW-
Makrophagen, sowie primäre C57BL/6-und BALB/c-Knochenmarkmakrophagen mit der Mutante
SAM 21 28.08.07Killing-Assay S. aureus 2 Wildtyp-Stämme und eine Agr-Mutante
mit RAW-M über 72h
MOI 500:1
Mediumwechsel nach 48h
0 25 50 75103
104
105
106
RN 6390 WT
RN 6911 Mutante
Newman WT
h post infection
intr
azell
ulä
re K
eim
zah
l
cfu
/Well
(400µ
l)
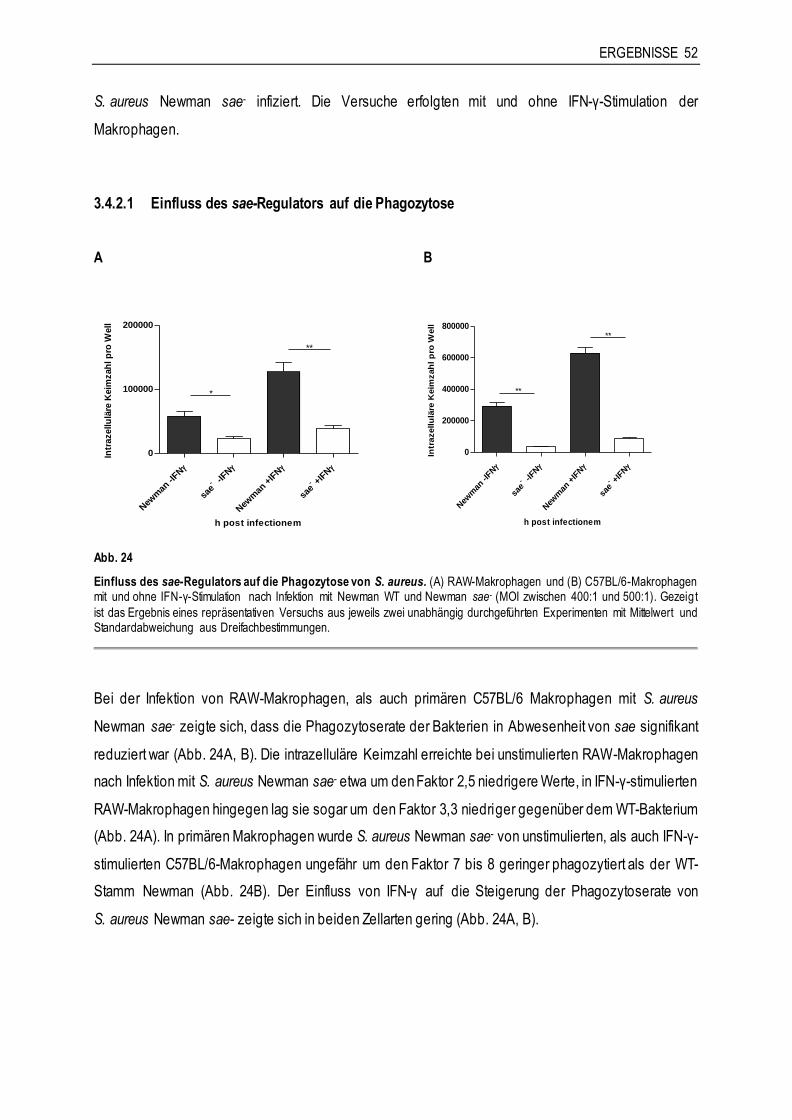
ERGEBNISSE 52
S. aureus Newman sae- infiziert. Die Versuche erfolgten mit und ohne IFN-γ-Stimulation der
Makrophagen.
3.4.2.1 Einfluss des sae-Regulators auf die Phagozytose
A B
New
man
-IFN
-IF
N-
sae
New
man
+IF
N
+IF
N-
sae
0
100000
200000
**
*
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
New
man
-IFN
-IFN
-
sae
New
man
+IF
N
+IF
N-
sae
0
200000
400000
600000
800000
**
**
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 24
Einfluss des sae-Regulators auf die Phagozytose von S. aureus. (A) RAW-Makrophagen und (B) C57BL/6-Makrophagen mit und ohne IFN-γ-Stimulation nach Infektion mit Newman WT und Newman sae- (MOI zwischen 400:1 und 500:1). Gezeigt
ist das Ergebnis eines repräsentativen Versuchs aus jeweils zwei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Bei der Infektion von RAW-Makrophagen, als auch primären C57BL/6 Makrophagen mit S. aureus
Newman sae- zeigte sich, dass die Phagozytoserate der Bakterien in Abwesenheit von sae signifikant
reduziert war (Abb. 24A, B). Die intrazelluläre Keimzahl erreichte bei unstimulierten RAW-Makrophagen
nach Infektion mit S. aureus Newman sae- etwa um den Faktor 2,5 niedrigere Werte, in IFN-γ-stimulierten
RAW-Makrophagen hingegen lag sie sogar um den Faktor 3,3 niedriger gegenüber dem WT-Bakterium
(Abb. 24A). In primären Makrophagen wurde S. aureus Newman sae- von unstimulierten, als auch IFN-γ-
stimulierten C57BL/6-Makrophagen ungefähr um den Faktor 7 bis 8 geringer phagozytiert als der WT-
Stamm Newman (Abb. 24B). Der Einfluss von IFN-γ auf die Steigerung der Phagozytoserate von
S. aureus Newman sae- zeigte sich in beiden Zellarten gering (Abb. 24A, B).
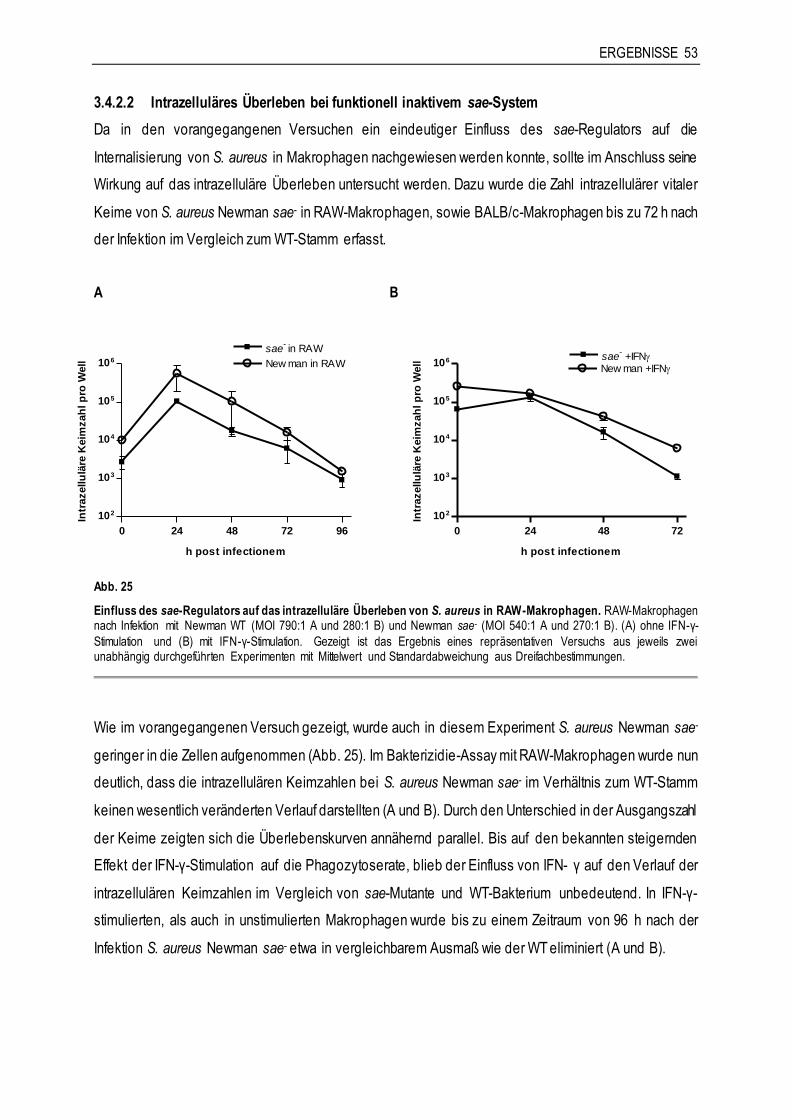
ERGEBNISSE 53
3.4.2.2 Intrazelluläres Überleben bei funktionell inaktivem sae-System
Da in den vorangegangenen Versuchen ein eindeutiger Einfluss des sae-Regulators auf die
Internalisierung von S. aureus in Makrophagen nachgewiesen werden konnte, sollte im Anschluss seine
Wirkung auf das intrazelluläre Überleben untersucht werden. Dazu wurde die Zahl intrazellulärer vitaler
Keime von S. aureus Newman sae- in RAW-Makrophagen, sowie BALB/c-Makrophagen bis zu 72 h nach
der Infektion im Vergleich zum WT-Stamm erfasst.
A B
0 24 48 72 96
102
103
104
105
106
sae-in RAW
New man in RAW
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48 72
102
103
104
105
106
New man +IFNsae
-+IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 25
Einfluss des sae-Regulators auf das intrazelluläre Überleben von S. aureus in RAW-Makrophagen. RAW-Makrophagen nach Infektion mit Newman WT (MOI 790:1 A und 280:1 B) und Newman sae- (MOI 540:1 A und 270:1 B). (A) ohne IFN-γ-
Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist das Ergebnis eines repräsentativen Versuchs aus jeweils zwei unabhängig durchgeführten Experimenten mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
Wie im vorangegangenen Versuch gezeigt, wurde auch in diesem Experiment S. aureus Newman sae-
geringer in die Zellen aufgenommen (Abb. 25). Im Bakterizidie-Assay mit RAW-Makrophagen wurde nun
deutlich, dass die intrazellulären Keimzahlen bei S. aureus Newman sae- im Verhältnis zum WT-Stamm
keinen wesentlich veränderten Verlauf darstellten (A und B). Durch den Unterschied in der Ausgangszahl
der Keime zeigten sich die Überlebenskurven annähernd parallel. Bis auf den bekannten steigernden
Effekt der IFN-γ-Stimulation auf die Phagozytoserate, blieb der Einfluss von IFN- γ auf den Verlauf der
intrazellulären Keimzahlen im Vergleich von sae-Mutante und WT-Bakterium unbedeutend. In IFN-γ-
stimulierten, als auch in unstimulierten Makrophagen wurde bis zu einem Zeitraum von 96 h nach der
Infektion S. aureus Newman sae- etwa in vergleichbarem Ausmaß wie der WT eliminiert (A und B).
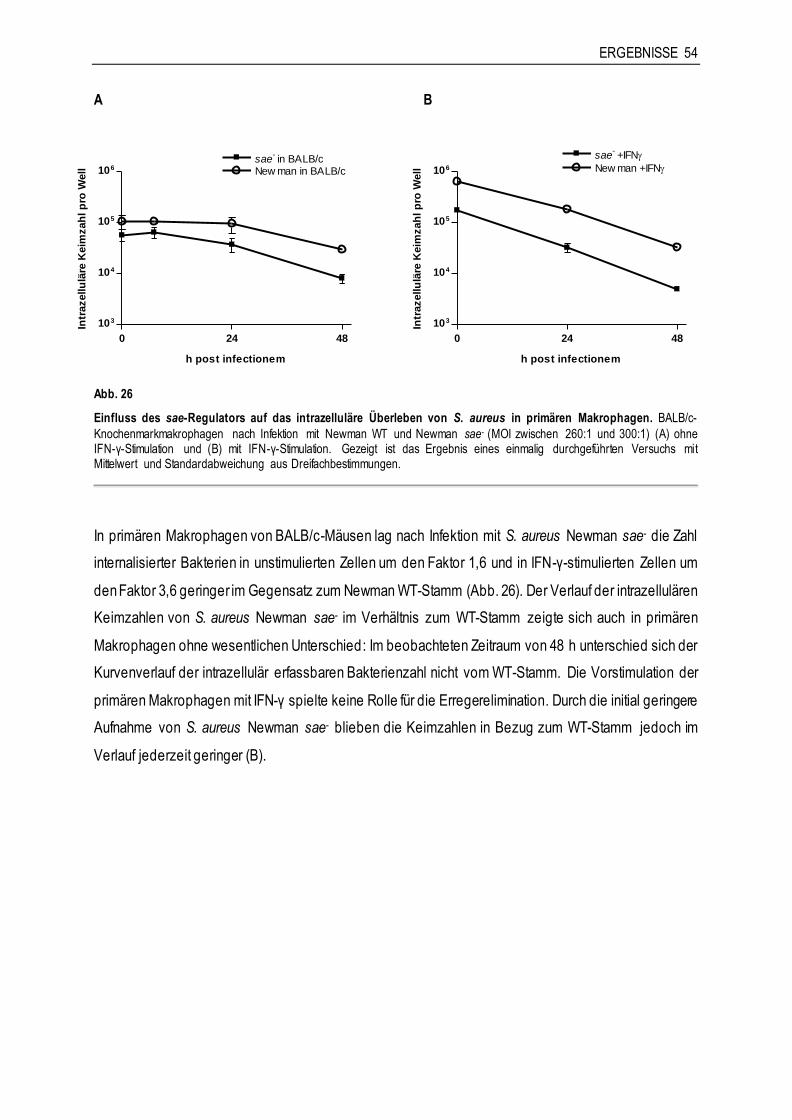
ERGEBNISSE 54
A B
0 24 48
103
104
105
106 New man in BALB/csae
-in BALB/c
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
0 24 48
103
104
105
106 New man +IFN
sae-+IFN
h post infectionem
Intr
azellu
läre
Keim
zah
l p
ro W
ell
Abb. 26
Einfluss des sae-Regulators auf das intrazelluläre Überleben von S. aureus in primären Makrophagen. BALB/c-
Knochenmarkmakrophagen nach Infektion mit Newman WT und Newman sae- (MOI zwischen 260:1 und 300:1) (A) ohne IFN-γ-Stimulation und (B) mit IFN-γ-Stimulation. Gezeigt ist das Ergebnis eines einmalig durchgeführten Versuchs mit Mittelwert und Standardabweichung aus Dreifachbestimmungen.
In primären Makrophagen von BALB/c-Mäusen lag nach Infektion mit S. aureus Newman sae- die Zahl
internalisierter Bakterien in unstimulierten Zellen um den Faktor 1,6 und in IFN-γ-stimulierten Zellen um
den Faktor 3,6 geringer im Gegensatz zum Newman WT-Stamm (Abb. 26). Der Verlauf der intrazellulären
Keimzahlen von S. aureus Newman sae- im Verhältnis zum WT-Stamm zeigte sich auch in primären
Makrophagen ohne wesentlichen Unterschied: Im beobachteten Zeitraum von 48 h unterschied sich der
Kurvenverlauf der intrazellulär erfassbaren Bakterienzahl nicht vom WT-Stamm. Die Vorstimulation der
primären Makrophagen mit IFN-γ spielte keine Rolle für die Erregerelimination. Durch die initial geringere
Aufnahme von S. aureus Newman sae- blieben die Keimzahlen in Bezug zum WT-Stamm jedoch im
Verlauf jederzeit geringer (B).

55
4 DISKUSSION
4.1 Phagozytose und intrazelluläres Überleben von Staphylokokken in
Makrophagen
4.1.1 Die bakterielle Regulation der Phagozytose in Makrophagen
Die Stammabhängigkeit der Phagozytose von S. aureus
Es gibt zahlreiche Studien, die die Internalisation von S. aureus in nicht-professionell phagozytierende
Zellen charakterisieren. Dort werden auch die stammabhängigen Unterschiede in der Invasivität
beschrieben (Sinha et al. 1999). In der vorliegenden Arbeit wurden nun ähnliche Beobachtungen für die
S. aureus-Makrophagen-Interaktion gemacht. Die S. aureus–Referenzstämme Newman und RN 6390
zeigten im Vergleich zu den S. aureus ATCC-Stämmen 25923, 12600 und 29213 deutlich geringere
Phagozytoseraten. Dies ist übereinstimmend mit den Untersuchungen an sämtlichen nicht-professionell
phagozytierenden Zellen, bei denen im Vergleich von verschiedenen S. aureus-Stämmen die
Laborstämme Newman und RN 6390 generell geringere Internalisierungsraten zeigten (Sinha et al. 1999;
Steinhuber et al. 2003).
Die Phagozytose von S. epidermidis
Die im Vergleich zu anderen Stämmen geringere Internalisierungsrate von S. aureus Newman zeigte sich
nicht allein gegenüber den S. aureus-ATCC-Stämmen, sondern sogar gegenüber S. epidermidis, obwohl
es sich um einen KNS-Vertreter handelt. Dies bestätigt nicht nur die geringe Internalisierungsrate von
Stamm Newman, sondern weist darüberhinaus auf das pathogene Potenzial des hier verwendeten
S. epidermidis-Stamms ATCC 12228 hin. Aus anderen Arbeiten ist bekannt, dass S. epidermidis
stammabhängig von Osteoblasten internalisiert werden kann (Khalil et al. 2007; Valour et al. 2013).
Außerdem konnte gezeigt werden, dass S. epidermidis über ein von S. aureus unabhängiges Fibronektin-
bindendes Protein, EmbP, verfügt (Williams et al. 2002).
Die Rolle von FnBP´s für die Phagozytose von S. aureus
Es konnte in vorliegender Arbeit gezeigt werden, dass die reduzierte Expression der FnbP´s im Fall von
den S. aureus-Stämmen Newman bzw. RN 6390 sich in der Internalisierungsrate in Makrophagen
erheblich abbildete. Für S. aureus Newman gilt dabei, dass seine geringe Internalisierungsrate durch eine
Mutation von fnbpA und fnbpB entsteht, die den Funktionsverlust beider Gene bewirkt, wie in nicht-
professionell phagozytierenden Zellen gezeigt werden konnte (Grundmeier et al. 2004).
Interessanterweise konnte in vorliegender Arbeit festgestellt werden, dass der Stamm RN 6390 noch

DISKUSSION 56
deutlich geringer in Makrophagen aufgenommen wurde als der Stamm Newman. Dies deutet zum einen
daraufhin, dass S. aureus Newman den Funktionsverlust seiner FnBP´s durch andere Invasine
kompensieren kann. In diesem Zusammenhang ist erwähnenswert, dass das extrazelluläre Adhärenz -
Protein Eap für die Invasivität von Stamm Newman eine erhebliche Rolle spielt (Haggar et al. 2003). Es
ist außerdem bekannt, dass S. aureus Newman im Vergleich zu anderen Stämmen Eap deutlich stärker
exprimiert (Harraghy et al. 2005).
Zum anderen liegt die Ursache der geringen Aufnahme von S. aureus RN 6390 in Zellen
bekanntermaßen in einem regulatorischen und damit komplexeren Defekt als im Fall von Newman. Dies
gilt neben RN 6390 für alle Derivate von S. aureus 8325-4 (Sinha et al. 1999). Dieser Defekt führt in der
Folge u.a. zu einer verringerten Expression von FnbP´s (Vaudaux et al. 2002; Saravia-Otten et al. 1997;
Herbert et al. 2010) und somit zur verringerten Aufnahme in nicht-professionell phagozytierende Zellen
(Sinha et al. 2000). Es handelt sich um einen Defekt im rsbU-Gen (Giachino et al. 2001; Kullik et al. 1998).
rsbU ist eine Phosphatase, die für die Aktivierung von SigB notwendig ist, von welchem wiederum bekannt
ist, dass es agr negativ reguliert (Giachino et al. 2001). Aufgrund des rsbU-Defektes bleibt die Aktivierung
des SigB-Regulators und folglich die von SigB regulierte Unterdrückung der agr-Expression aus (Bischoff
et al. 2001; Pane-Farre et al. 2009; Horsburgh et al. 2002). In einer kürzlich durchgeführten Studie mit
S. aureus und Makrophagen (Olivier et al. 2009) bestätigte sich dies und ist daher übereinstimmend mit
den geringen Internalisierungsraten von RN 6390 in dieser Arbeit. Im Vergleich zu S. aureus 8325-4
wurde bei Olivier et al. der Stamm mit wieder hergestelltem rsbU-Gen, genannt SH1000, (Horsburgh et
al. 2002) deutlich besser in die Makrophagen-Zelllinie THP-1 aufgenommen (Olivier et al. 2009).
Weiterführend konnte in Experimenten mit der agr-Mutante S. aureus RN 6911 in dieser Arbeit die
Phagozytoserate sogar deutlich über die von Stamm Newman gesteigert werden.
Die Rolle der globalen Regulatoren agr und sae für die Phagozytose von S. aureus
Es wurde bereits in früheren Studien nachgewiesen, dass FnBP´s als auch die Fibronektinbindung selbst
negativ agr-reguliert sind (Wolz et al. 2000; Saravia-Otten et al. 1997), wodurch S. aureus agr-Mutanten
besser in nicht-professionell phagozytierende Zellen eindringen konnten (Wesson et al. 1998). Dies
konnte in dieser Arbeit nun auch für Makrophagen bestätigt werden. Höchstwahrscheinlich ist durch den
Gendefekt neben den FnBP´s die Produktion noch weiterer Proteine beeinflusst, die für die Bindung an
Wirtsfaktoren eine Rolle spielen (Blevins et al. 2002). Zusätzlich ist bekannt, dass durch die fehlende
sigB-Aktivität im Fall von S. aureus RN 6390 nicht nur agr überexprimert, sondern auch der sar-Regulator
geringer exprimiert wird (Gertz et al. 2000; Gertz et al. 1999). Dieser ist im Gegensatz zu agr an der
Expression der FnbP´s beteiligt (Wolz et al. 2000).
Demgegenüber resultierte die Infektion der Makrophagen mit der Mutante S. aureus
Newman sae- in Experimenten dieser Arbeit in deutlich verringerten intrazellulären Keimzahlen, wie

DISKUSSION 57
entsprechend an Endothelzellen und Epithelzellen bereits gezeigt wurde (Steinhuber et al. 2003; Liang et
al. 2006). Es zeigte sich hier der entgegengesetzte Effekt im Verg leich zur Infektion mit der agr-Mutante.
Der sae-abhängigen Phagozytose von S. aureus mag zugrunde liegen, dass der Regulator sae neben
anderen Oberflächenproteinen die Transkription von fnbp aktiviert, und dass agr bekanntermaßen
entgegengesetzt wirkt (Steinhuber et al. 2003; Rogasch et al. 2006; Wolz et al. 2000). Das besonders für
die Invasivität von Stamm Newman entscheidende Eap ist sogar essentiell durch sae reguliert, was mit
den hier gemachten Beobachtungen korreliert (Harraghy et al. 2005). Nach unserem Wissensstand der
gängigen Literatur ist dies die erste Darstellung zum Einfluss von sae und agr für die Aufnahme von
S. aureus in Makrophagen.
4.1.2 Die bakterielle Regulation des intrazellulären Überlebens in Makrophagen
Die fehlende Stammabhängigkeit des intrazellulären Überlebens in murinen Makrophagen
Während sich in vorliegender Arbeit die Phagozytose von S. aureus in Makrophagen deutlich abhängig
vom jeweils eingesetzten Stamm zeigte, konnte diese Beobachtung beim intrazellulären Überleben nicht
gemacht werden. Vielmehr wurde deutlich, dass alle hier eingesetzten S. aureus-Stämme einen
vergleichbaren Verlauf der Überlebenskurve über 5 Tage zeigten, bei dem die intrazelluläre Keimzahl
stetig abnahm. Zwei weitere Studien, die sich mit diesem relativ langen Beobachtungszeitraum von
intrazellulärem S. aureus beschäftigten, konnte ähnliche Überlebenskurven in einer Ratten-
Alveolarmakrophagen-Zellinie (Hamza et al. 2013; Hamza und Li 2014), sowie in humanen Monocyt-
derived-Macrophages (hMDM) beobachten (Kubica et al. 2008). Anders als hier konnte bei Kubica et al.
der Stamm S. aureus 8325-4 (RN 6390 als Derivat) nicht intrazellulär überleben. Dies wurde auf den
rsbU-Defekt zurückgeführt, wie in Experimenten mit dem Stamm SH1000 mit wiederhergestelltem rsbU–
Gen bewiesen werden konnte. Korrelierende Effekte konnten bei Olivier et al. auch an THP1-
Makrophagen gesehen werden, wo sich rsbU als essentiell für das intrazelluläre Überleben erwies (Olivier
et al. 2009). In den hier durchgeführten Experimenten konnte dies nicht bestätigt werden, da der Stamm
RN 6390 mindestens 5 Tage intrazellulär überlebte. Die Diskrepanzen zwischen den Daten dieser Arbeit
und den von Kubica et al. bzw. Olivier et al. gemachten Beobachtungen könnten u.a. darin begründet
sein, dass in beiden Studien humane Makrophagen-Zelllinien (hMDM, THP-1) eingesetzt wurden und
diese sich von der hier eingesetzten murinen Makrophagen-Zelllinie RAW unterscheiden. Es handelt sich
bei hMDM-, sowie THP-1- Zellen nicht um inflammatorische Makrophagen, sondern um Zellen
monozytären Ursprungs. Es bestehen Unterschiede im Differenzierungsgrad und somit eine geringere
Phagozytoseleistung und eine veränderte Zytokinfreisetzung der Zellen im Vergleich zur murinen
Makrophagen-Zelllinie, wie kürzlich untersucht worden ist (da Silva Domingues, Joana F et al. 2013;

DISKUSSION 58
Kubica et al. 2008). Dies macht deutlich, dass die verschiedenen Infektionssysteme nur bedingt
vergleichbar sind, und lässt die hier gewonnenen Ergebnisse an der murinen Makrophagen-Zelllinie
aussagekräftiger erscheinen. Zusätzlich konnten die Ergebnisse durch Untersuchungen mit primären
Knochenmarkmakrophagen von C57BL/6- und BALB/c-Mäusen bestätigt werden. Diese weisen im
Vergleich zur murinen Zelllinie deutlich höhere Expressionsraten sämtlicher Proteine und Zytokine auf
(Chamberlain et al. 2009) und erbringen daher nochmals aussagekräftigere Ergebnisse. Nach unserem
Wissensstand der verfügbaren Literatur sind dies die ersten Ergebnisse zur Kinetik des intrazellulären
Überlebens von S. aureus in primären murinen Knochenmarkmakrophagen.
Intrazelluläres Überleben in genetisch unterschiedlichen primären Makrophagen
In anderen Infektionsmodellen, wie beispielsweise mit dem intrazellulären gramnegativen Stäbchen
B. pseudomallei, lassen die derzeitigen Ergebnisse vermuten, dass die genetisch bedingte
unterschiedliche bakterizide Aktivität von Makrophagen aus BALB/c- und C57BL/6-Mäusen zu der
unterschiedlichen Empfänglichkeit dieser Mausstämme gegenüber dem Erreger beitragen (Breitbach et
al. 2006; Depke et al. 2014; Eske et al. 2009). Ergebnisse unserer Arbeitsgruppe konnten zeigen, dass
auch gegenüber einer Infektion mit S. aureus Unterschiede in der Empfänglichkeit bestehen. Dabei
zeigten sich BALB/c-Mäuse empfänglicher gegenüber einer pulmonalen Infektion mit S. aureus als
C57BL/6-Mäuse (Köhler et al. 2011). Die Ergebnisse der vorliegenden Arbeit zeigten jedoch, dass keine
Unterschiede in der Eliminationsfähigkeit von primären Knochenmarkmakrophagen von C57BL/6- und
BALB/c-Mäusen gegenüber intrazellulärem S. aureus bestehen. Dies deutet daraufhin, dass die
unterschiedliche Empfänglichkeit beider Mausstämme gegenüber einer S. aureus-Infektion vermutlich
nicht auf Unterschieden im intrazellulären Überleben von S. aureus in Makrophagen beruht (Depke et al.
2014). Die Beobachtungen weisen vielmehr daraufhin, dass Makrophagen nicht entscheidend für die
Elimination von S. aureus, jedoch für eine generelle Entzündungsreaktion sind (Fournier 2012; Alberts
2011), wie die entsprechenden in-vivo-Untersuchungen unserer Arbeitsgruppe zeigten (Köhler et al.
2011).
Die Rolle von agr und sae für die Kinetik des intrazellulären Überlebens von S. aureus
Während in der schon erwähnten Arbeit von Kubica et al. lediglich alle 24 h die intrazelluläre Keimzahl
bestimmt wurde, konnte in der vorliegenden Studie durch Messung der Keimzahl bereits 6 h nach der
Infektion der Verlauf der intrazellulären Keimzahl differenzierter charakterisiert werden. Trotz des
kontinuierlichen Rückgangs der intrazellulären Keimzahl im Verlauf über 5 Tage war eine kurzzeitige
Proliferation von intrazellulären S. aureus-Stämmen in den ersten 6 h nach der Phagozytose feststellbar,
die besonders bei dem Stamm RN 6390 konsequent auffällig war. Dieses Phänomen wurde bereits früher
mit RN 6390 in Epithelzellen beschrieben (Wesson et al. 1998). Außerdem konnte in den Experimenten

DISKUSSION 59
vorliegender Studie hin und wieder auch eine Proliferation bis zu 24, oder 48 h nach der Phagozytose
dieses Stamms beobachtet werden. Ursächlich dafür ist wiederum die fehlende sigB-Aktivität in RN 6390.
So produziert RN 6390 im Vergleich zum Stamm Newman größere Mengen an hla, sowie weiterer
Exoproteine, die aus der Über-Regulation von agr resultieren (Kullik et al. 1998; Horsburgh et al. 2002)
und entscheidend für das intrazelluläre Überleben sind (Kubica et al. 2008; Jarry et al. 2008; Shompole
et al. 2003; Sinha und Fraunholz 2010; Fraunholz und Sinha 2012). Bei den Experimenten mit der agr-
Mutante RN 6911 in dieser Arbeit konnte dies bewiesen werden, da die beschriebene frühe Proliferation
dann ausblieb. Das weitere intrazelluläre Überleben war im Vergleich zum Wildtyp ansonsten unverändert
über 5 Tage möglich. Demgegenüber stehen die Ergebnisse der Arbeit von Kubica et al. Sie wiesen nach,
dass das intrazelluläre Überleben von S. aureus in Makrophagen eindeutig agr-abhängig ist. Agr-
Mutanten wurden dort innerhalb von 2 Tagen eliminiert. In vorliegender Arbeit stellte sich agr hingegen
nicht als essentiell für das Überleben von S. aureus in Makrophagen dar. Entsprechend seiner
regulatorischen Funktion verhinderte das Fehlen von agr hier nur die frühe Proliferation intrazellulärer
Keime und führte weiterhin zu erhöhten Phagozytoseraten. Ein möglicher Erklärungsansatz für die
Unterschiede zu den Ergebnissen von Kubica et al. ist die dort eingesetzte Newman agr-Mutante im
Gegensatz zur hier verwendeten agr-Mutante RN 6911. Über die stammbedingten Unterschiede
zwischen S. aureus Newman und RN 6390, insbesondere dem rsbU-Defekt bei S. aureus RN 6390 mit
konsekutiver agr-Überregulation, wurde bereits diskutiert. Eine mögliche Bestätigung findet dieser
Erklärungsansatz durch entsprechende Experimente mit RN 6911 agr- in Epithelzellen, in denen sich das
intrazelluläre Verhalten ähnlich zeigte wie hier in Makrophagen (Wesson et al. 1998). Die Vergleichbarkeit
von Experimenten mit den Referenzstämmen Newman und RN 6390 ist aufgrund ihrer bekannten
regulatorischen Unterschiede begrenzt, wie aus der Literatur bereits hervorgeht (Blevins et al. 2002;
Mainiero et al. 2010; Horsburgh et al. 2002). Untersuchungen mit vielen verschiedenen S. aureus-
Stämmen müssten folgen, um repräsentative und klinisch relevante Ergebnisse zu erhalten.
Der zusätzlich in dieser Arbeit untersuchte sae-Regulator zeigte neben der deutlich positiv
regulierten Phagozytoserate auf den weiteren intrazellulären Verlauf keinen Einfluss, wie Experimente
mit Newman sae- ergaben. Trotz der in dieser Arbeit für Mutantenversuche unterschiedlich eingesetzten
Stämme ergibt sich das Bild, dass das intrazelluläre Überleben von S. aureus in Makrophagen wenig
störanfällig ist und, wie die Ergebnisse vermuten lassen, durch die Beteiligung vieler verschiedener
regulatorischer Einflüsse stabil gehalten wird. Keine der insgesamt in dieser Dissertation durchgeführten
Untersuchungen an murinen Makrophagen zeigte eine frühzeitige Elimination von intrazellulärem
S. aureus.

DISKUSSION 60
Intrazelluläres Überleben von S. epidermidis
Die bei S. aureus gemachten Beobachtungen zeigten sich, mit einigen Abweichungen, auch bei dem hier
verwendeten KNS-Vertreter S. epidermidis ATCC 12228. Dieser weniger virulente Erreger im Vergleich
zur S. aureus zeigte auch zu frühen Zeitpunkten nach der Internalisierung keine Proliferation. Weiterhin
unterschied sich der Verlauf des Überlebens des in dieser Studie eingesetzten S. epidermidis-Stammes
mit dem aller eingesetzten S. aureus-Stämme, da nach 3 Tagen keine intrazellulären Keime mehr
nachgewiesen werden konnten. S. epidermidis ATCC 12228 konnte allerdings für mindestens 48 h in
Makrophagen persistieren und wurde nicht sofort eliminiert. Eine frühere Arbeit mit in-vivo-Experimenten
zu Biomaterial-assoziierten Infektionen konnte S. epidermidis bis zu 60 Tage in Makrophagen in der
Umgebung infizierter Katheter nachweisen (Boelens et al. 2000). Diese lange intrazelluläre Persistenz
erklärte sich aus der lokal reduzierten Abwehrlage des Wirts im Biomaterial-umgebenden Gewebe und
ist eine mögliche Ursache für die gefürchteten therapieresistenten Fremdkö rperinfektionen durch
S. epidermidis. Ein Widerspruch zu der kürzeren Überlebensdauer von S. epidermidis gegenüber
S. aureus in dieser Arbeit ergibt sich damit nicht, da die in-vivo Situation bei Boelens et al. andere
Rahmenbedingungen schafft und das Fremdkörpermaterial eine veränderte Zytokinfreisetzung hervorruft.
Der hier gebrachte Nachweis, dass S. epidermidis in Makrophagen internalisiert und nicht sofort abgebaut
wird, spricht für sein pathogenes Potenzial. Es ist beispielsweise bekannt, dass S. epidermidis über
zytolytische Toxine verfügt, die ihn vor der Abwehr in Neutrophilen schützen können (Cheung, Gordon Y
C et al. 2010; Otto 2012). Eine ähnliche aggressive Expression der Toxine wie bei S. aureus konnte nicht
gesehen werden. Dies würde nicht der kommensalen Art der Interaktion von KNS-Stämmen mit ihrem
Wirt entsprechen, was sich in dieser Arbeit durch die kürzere intrazelluläre Persistenz von S. epidermidis
gegenüber S. aureus verdeutlichen ließ. Einschränkend muss jedoch hinzugefügt werden, dass im
Rahmen dieser Arbeit nur ein S. epidermidis-Stamm untersucht wurde. Weitere Studien mit weiteren
KNS-Stämmen sind nötig, um das intrazelluläre Überleben dieser Erreger in Makrophagen genauer zu
charakterisieren.
4.2 Die Bedeutung von FnBPA und Fn für die Internalisierung von S. aureus
Fnbp erhöht die Internalisierung von S. aureus durch Makrophagen
FnBP´s sind essentiell für die Internalisation von S. aureus in nicht-professionell phagozytierende Zellen.
Über das Brückenmolekül Fibronektin, einem extrazellulären Matrixprotein, stellen sie die Verbindung
zum α5β1-Integrinrezeptor der Wirtszellmembran her und vermitteln damit die Internalisierung (Sinha et
al. 1999; Sinha und Fraunholz 2010; Hauck und Ohlsen 2006; Dziewanowska et al. 1999; Alva-Murillo et
al. 2014). Dem sonst nicht-invasiven KNS-Stamm S. carnosus konnte durch die Expression von

DISKUSSION 61
S. aureus-FnBP´s die volle Invasivität, sowie die Fibronektin-Bindungskapazität übertragen werden
(Sinha et al. 1999; Sinha et al. 2000). S. carnosus fehlen sämtliche bekannte S. aureus-
Oberflächenproteine und er kann kein Fibronektin binden (Sinha et al. 1999).
In vorliegender Arbeit wurde nun gezeigt, dass FnBPA auch für die Aufnahme von S. aureus in
Makrophagen eine Rolle spielt, wie in Experimenten mit FnBPA-exprimierendem S. carnosus gezeigt
werden konnte. Die Internalisierung war dabei abhängig von Fibronektin. Shinji et al. konnten bereits
früher zeigen, dass Fibronektin-gebundener S. aureus deutlich besser durch Makrophagen
aufgenommen wird (Shinji et al. 1998). Später fanden die Autoren heraus, dass diese Aufnahme durch
FnBP´s vermittelt wird (Shinji et al. 2011). Aber auch ohne Fibronektin zeigte sich in unserer Arbeit die
Internalisierung von S. carnosus (FNBPA) im Vergleich zum Wildtyp S. carnosus erstaunlicherweise
deutlich erhöht. In diesem Zusammenhang ist erwähnenswert, dass neben der Fähigkeit der Fibronektin-
Bindung FnBP´s zusätzlich über die Möglichkeit der Fibrinogen-Bindung verfügen, wodurch es vermutlich
auch zu einer Internalisierung kommen kann (Wann et al. 2000). Interessanterweise konnte von Shinji et
al. im in-vivo-Modell gezeigt werden, dass das Vorhandensein von FnBP´s schwere septische Verläufe
und eine erhöhte Mortalität des Gesamtorganismus zur Folge hat. Auch andere Autoren zeigten an in-
vivo-Modellen, dass FnBP´s als Virulenzfaktoren von S. aureus eine Rolle spielen und mit der Entstehung
von systemischen inflammatorischen Reaktionen in Verbindung gebracht werden können (Palmqvist et
al. 2005; Brouillette et al. 2003; Edwards et al. 2010; Jenkins et al. 2015). In diesem Zusammenhang
konnte nachgewiesen werden, dass S. aureus-FnBP´s nicht nur zu gesteigerten Internalisierungsraten
führten, sondern auch eine Entzündungsreaktion im Makrophagen induzierten, was an der deutlich
gesteigerten Produktion proinflammatorischer Zytokine wie TNF-alpha und IL-6 erkennbar war (Shinji et
al. 2011). In nicht-professionell phagozytierenden Zellen wurde die FnBPA-abhängige Internalisation von
S. aureus als mögliche Strategie des Bakteriums gedeutet, der immunologischen Abwehr zu entgehen
und vor der Phagozytose und Abwehr durch Makrophagen und Neutrophile geschützt zu sein (Fraunholz
und Sinha 2012). Im Widerspruch dazu wird in dieser Arbeit deutlich, dass S. aureus auch Makrophagen
auf diese Weise internalisieren kann und er intrazellulär für mehrere Tage überlebt, statt eliminert zu
werden.
Ähnlich wie für nicht-professionell phagozytierende Zellen mehrfach nachgewiesen wurde (Sinha
et al. 1999; Sinha und Fraunholz 2010; Hauck und Ohlsen 2006; Dziewanowska et al. 1999) ist bei
Aufnahme von S. aureus in Makrophagen der α5β1-Integrinrezeptor auf der Wirtszellmembran
entscheidend (Wang et al. 2008; Shinji et al. 2003). Es wird angenommen, dass die Integrin-Aktivierung
durch die FnBP-Bindung erfolgt und es über die damit einhergehenden Zytoskelett-Veränderungen zur
Aufnahme von S. aureus in Makrophagen kommt (Shinji et al. 2003; Shinji et al. 2007). Die α5β1-Integrin-
Aktivierung durch FnBP´s führt konkret auch zu der beschriebenen Induktion einer inflammatorischen

DISKUSSION 62
Antwort im Makrophagen (Shinji et al. 2011). Dies deutet daraufhin, dass auch in einem professionell-
phagozytierenden Wirtszellsystem die Phagozytose durch bakterielle Mechanismen beeinflusst werden
kann, und dass es sich bei der FnBP-Fn-induzierten Internalisation von S. aureus in Makrophagen um
ein pathogenes Potenzial des Erregers handelt. Nicht zuletzt bestätigen dies die Experimente mit agr-
und sae-Mutanten dieser Studie, die den Einfluss bakterieller Komponenten auf das intrazelluläre
Überleben und die Phagozytose von S. aureus in Makrophagen eindeutig zeigten.
S.epidermidis wird FnbP-unabhängig internalisiert
Interessanterweise konnte in dieser Arbeit gezeigt werden, dass auch S. epidermidis und der
apathogene Wildtyp-Stamm S. carnosus in Makrophagen aufgenommen werden. Für S. epidermidis
wurde bereits nachgewiesen, dass die Aufnahme in Osteoblasten unabhängig von FnBP´s, sowie
unabhängig vom α5β1-Integrin ist (Khalil et al. 2007). Shinji et al zeigten in Experimenten mit primären
Makrophagen, dass der KNS-Stamm S. saprophyticus und andere klinische KNS-Stämme Fibronektin in
gleichem Maß wie S. aureus binden konnten, diese Fn-Bindung im Gegensatz zu S. aureus allerdings
keine Steigerung der Phagozytoserate bewirkte (Shinji et al. 1998). Ursächlich dafür wurden die fehlende
Integrinrezeptor-Aktivierung und die folglich ausbleibenden Cytoskelettveränderungen bei den KNS-
Stämmen gesehen, die demgegenüber bei S. aureus stattfanden und die Phagozytose in Makrophagen
bewirkten (Shinji et al. 2003). Darüberhinaus wurde herausgefunden, dass für S. epidermidis das
Autolysin atlE für die Vermittlung der Internalisierung in nicht-professionell phagozytierende Zellen eine
essentielle Rolle spielt. Als Wirtszellrezeptor konnte das heat shock cognate protein Hsc70 identifiziert
werden (Hirschhausen et al. 2010)
Spezialfall S. aureus Newman: Internalisierung Fn-abhängig, aber FnbP-unabhängig
Die Ergebnisse vorliegender Arbeit konnten nachweisen, dass die Aufnahme von S. aureus Newman in
Makrophagen eindeutig Fibronektin-abhängig ist. Überraschender Weise ergab sich weiterhin, dass
Newman etwa in gleichem Maß wie S. carnosus Wildtyp in murine Makrophagen aufgenommen wurde.
Die Phagozytose von S. carnosus TM 300 zeigte sich im Gegensatz zu S. aureus Newman jedoch nicht
Fn-abhängig, womit frühere Untersuchungen bestätigt werden, dass S. carnosus keine
Fibronektinbindung eingehen kann, da ihm entsprechende Oberflächenproteine fehlen (Sinha et al.
2000). Dass S. carnosus dennoch internalisiert wird hängt möglicherweise damit zusammen, dass es sich
bei Makrophagen um ein professionell phagozytierendes Wirtszellsystem handelt. Dieses verfügt über
eine Vielzahl an Erkennungsrezeptoren ohne Notwendigkeit für opsonierende Proteine, oder bestimmte
bakterielle Bindungseigenschaften (Palecanda und Kobzik 2001; Taylor et al. 2005; Pluddemann et al.
2009), so dass sich auch der apathogene KNS-Stamm S. carnosus intrazellulär nachweisen ließ.

DISKUSSION 63
Die niedrigen Internalisierungsraten von S. aureus Newman ergeben sich hingegen, wie oben
erwähnt, durch die Mutation in den fnbp-Genen (Grundmeier et al. 2004). Überträgt man FnBP´s
(Newman) dem Stamm S. carnosus wird diesem weder die Fn-Bindungsfähigkeit, noch die Invasivität
vermittelt (Grundmeier et al. 2004). Ähnliches zeigte sich auch hier an Makrophagen. Die
Phagozytoserate wird durch Versuche mit S. carnosus (FnBPA) erheblich gesteigert, bleibt im Fall von
S. aureus Newman durch den Funktionsverlust der FnBP´s gering. Die Internalisierung erfolgt bei
Newman, als auch bei S. carnosus (FnBPA) trotzdem deutlich Fn-abhängig. Das bei Stamm Newman
stark exprimierte Eap, das neben anderen Liganden auch Fibronektin bindet, spielt eine bedeutende Rolle
in der FnbP-unabhängigen Internalisierung von S. aureus Newman (Bur et al. 2013a; Haggar et al. 2003;
Hussain et al. 2002; Fraunholz und Sinha 2012).
4.3 Die Rolle von iNOS und NADPH-Oxidase in der Abwehr von intrazellulärem
S. aureus
Reaktive Sauerstoffradikale (ROS), sowie NO und seine Derivate (RNI) sind wichtige toxische Moleküle
in der Makrophagen-Abwehr gegenüber intrazellulären Erregern (Chakravortty und Hensel 2003; Nathan
und Shiloh 2000). Der Bildung von NO wird dabei ein wichtiger Teil der Abwehrmechanismen von
Makrophagen zuteil.
Um die Bedeutung von NO in der Abwehr von intrazellulärem S. aureus zu bestimmen konnte
zunächst gezeigt werden, dass S. aureus die NO-Produktion in Makrophagen induziert. Bereits aus
früheren Untersuchungen ist bekannt, dass S. aureus die iNOS-Expression und NO-Produktion auslöst
(Cunha et al. 1993a; Cunha et al. 1993b; Kuo et al. 2003; Watanabe et al. 2007; Sasaki et al. 1998; Kim
et al. 2015). Die hier vorliegenden Ergebnisse brachten hervor, dass d ie Stimulation der NO-Produktion
in Makrophagen nach S. aureus-Infektion deutlich stärker ausfällt als durch eine Vorbehandlung mit IFN-γ
erreicht werden kann, obwohl IFN-γ das Schlüsselmolekül für die iNOS-Induktion in Makrophagen ist
(Shibata et al. 2011; Bogdan et al. 2000b). Überraschenderweise zeigten in dieser Arbeit Experimente
mit Aminoguanidin, einem biochemischen Inhibitor der iNOS-Expression, dass die Eliminationsfähigkeit
der Makrophagen nach S. aureus-Infektion unabhängig von iNOS blieb. In weiterführenden Experimenten
mit Makrophagen von Mäusen mit genetischem Defekt im iNOS-Gen bestätigte sich dieses Ergebnis. Im
Vergleich von C57BL/6 WT-Makrophagen und C57BL/6 iNOS-/--Makrophagen ergaben sich keine
Unterschiede in den Überlebensraten von intrazellulärem S. aureus Newman.
In entsprechenden in-vivo-Experimenten unserer Arbeitsgruppe mit C57BL/6 iNOS-/--Mäusen
ergab sich, dass die iNOS-Oxidase auch für die Abwehr einer pulmonalen S. aureus-Infektion keine Rolle

DISKUSSION 64
spielt (Köhler et al. 2011). Demgegenüber zeigte sich die NADPH-Oxidase als essentiell für das
Überleben der Tiere, wie sich an den deutlich verringerten Überlebensraten von C57BL/6 gp91phox-/--
Mäusen nach pulmonaler Infektion mit S. aureus nachweisen ließ (Köhler et al. 2011). Die Ergebnisse
vorliegender Arbeit legten überraschender Weise an C57BL/6 gp91phox-/--Makrophagen dar, dass sich
die Überlebensraten von intrazellulärem S. aureus im Vergleich zu Wildtyp-Makrophagen nicht
unterschieden. Nach unserem Kenntnisstand sind dies die ersten Ergebnisse zur Interaktion von
S. aureus mit primären Knochenmarkmakrophagen von Mäusen mit Defekten im iNOS-Gen bzw. der gp91
Untereinheit der NADPH-Oxidase.
Es ist anzunehmen, dass die Bedeutung der am in-vivo- Modell beobachteten essentiellen Rolle
von NADPH in der Abwehr einer S. aureus–Infektion nicht Makrophagen zukommt. Ähnlich wurde dies
von Blos et al. in der Kontrolle einer Leishmania-Infektion beobachtet. Es zeigte sich, dass die NADPH-
Oxidase von existenzieller Bedeutung für das Überleben des Gesamtorganismus ist, die
Eliminationsfähigkeit von gp91phox-/--Makrophagen im Vergleich zu WT-Makrophagen sich dabei aber
nicht unterschied (Blos et al. 2003). Ferner wurde an verschiedenen anderen Infektionsmodellen
beschrieben, dass neutrophile Granulozyten für die Kontrolle einer S. aureus-Infektion entscheidend sind
(Robertson et al. 2008; Rigby und DeLeo 2012; Kockritz-Blickwede et al. 2008). Diesen Beobachtungen
entsprechend wurde herausgefunden, dass für die Elimination von S. aureus in Neutrophilen die NADPH-
Oxidase-Funktion eine entscheidende Rolle spielt (Ellson et al. 2006; Schwartz et al. 2009).
In Ergebnissen unserer Arbeitsgruppe konnte bestätigt werden, dass Makrophagen im
Gegensatz zu Neutrophilen für die Abwehr einer pulmonalen S. aureus-Infektion keine Rolle spielen, und
dass die Depletion von Makrophagen sogar zu verlängerten Überlebensraten der Mäuse führte (Köhler
et al. 2011). Dies könnte damit zusammenhängen, dass Makrophagen die Hauptproduzenten
proinflammatorischer Zytokine sind (Fournier und Philpott 2005; Kerdudou et al. 2006; Fournier 2012)
und eine übermäßige Zytokinproduktion auch zu erhöhter Gewebeschädigung und folglich erhöhter
Mortalität führt. Weiterhin ist in diesem Zusammenhang erwähnenswert, dass die FnBP-vermittelte
Internalisierung von S. aureus in Makrophagen nicht nur die Phagozytoserate steigert, wie die
vorliegenden Ergebnisse zeigen konnten, sondern zusätzlich e ine starke proinflammatorische
Zytokinbildung induziert (Shinji et al. 2011). Dies zeigte sich an weniger schweren Krankheitsverläufen
und erhöhten Überlebensraten im murinen Modell nach Infektion mit fnbp-Mutanten (Shinji et al. 2011).
Eine der möglichen Ursachen des fehlenden Einflusses der iNOS- bzw. NADPH-Effektormoleküle
des Makrophagen gegenüber intrazellulärem S. aureus mag die Fähigkeit sein, durch die Vielfalt
bakterieller Enzyme wie beispielsweise Katalase, Superoxid-Dismutase, L-Laktatdehydrogenase, oder
Staphyloxanthin, die oxidative und nitrosative Wirkung der iNOS- und NADPH-Produkte zu neutralisieren
(Gaupp et al. 2012; Richardson et al. 2008; Clauditz et al. 2006). Dies wurde kürzlich für die S. aureus-

DISKUSSION 65
Makrophagen-Interaktion untersucht und zeigte sich entscheidend für die fehlende Erregerelimination der
Makrophagen und das intrazelluläre Überleben von S. aureus (Das et al. 2008; Das und Bishayi 2010,
2009; Bishayi et al. 2014; Gaupp et al. 2012; Olivier et al. 2009; Richardson et al. 2008).
4.4 S. aureus - ein fakultativ intrazelluläres Bakterium?
Die häufigsten und klassischen Manifestationen von S. aureus-Infektionen, wie Haut.- und
Weichteilinfektionen, Abszess, Impetigo, Sepsis, Pneumonie usw. werden durch extrazelluläre Bakterien
verursacht. In der Untersuchung von Gewebeproben findet man S. aureus dabei in extrazellulären
Räumen (Finlay und Cossart 1997; Lowy 2000). Dennoch lassen chronische und rekurrente
Krankheitsverläufe vermuten, dass S. aureus auch als intrazelluläres Bakterium pathogenetisch
bedeutsam ist. Dies zeigt sich an häufig endovaskulären Infektionen, an hohen Metastasierungsraten von
Infektionen, an hohen Rückfallraten trotz antibiotischer Therapie, oder dass eine antibiotische Therapie
wirkungslos bleibt, wie bei langwierigen Osteomyelitiden.
Diese klinischen Beobachtungen werden durch die zahlreichen in-vitro-Untersuchungen
bestätigt, die S. aureus intrazellulär in sämtlichen Wirtszellen nachweisen konnten. Dies wurde als
Fähigkeit gedeutet, einerseits der konventionellen antibiotischen Therapie, aber andererseits auch der
Abwehr durch professionell phagozytierende Zellen zu entgehen (Fraunholz und Sinha 2012; Garzoni
und Kelley 2009). Wie in vorliegender Arbeit nachgewiesen werden konnte, kann S. aureus jedoch auch
professionell phagozytierende Zellen internalisieren und über längere Zeiträume überleben, so dass er
als fakultativ intrazelluläres Bakterium bezeichnet werden könnte. Bei B. pseudomallei, M. tuberkulosis,
Legionella pneumopila, oder Listeria monocytogenes handelt es sich um typische Beispiele fakultativ
intrazellulärer Bakterien (Molmeret et al. 2004; Seveau et al. 2007; Xu und Luo 2013; Rohde et al. 2007;
Gan 2005). Diese zeichnen sich dadurch aus, dass sie intrazellulär replizieren können und über längere
Zeiträume intrazellulär verbleiben, insbesondere in Makrophagen. Der intrazelluläre Aufenthalt ist dabei
Teil ihres pathogenen Zyklus und die Grundlage für die spezifischen Krankheitsbilder, die diese Erreger
hervorrufen.
Davon unterscheidet sich S. aureus, wie in vorliegender Arbeit gezeigt werden konnte. Allenfalls,
wenn überhaupt, war eine kurzzeitige initiale Replikation mit Übergang in eine langsame, aber
konsequente Reduktion der intrazellulären Keimzahlen über etwa 5 Tage zu beobachten. Des Weiteren
fehlt ein definitiver Nachweis, dass intrazellulärer S. aureus tatsächlich pathogenetische Grundlage einer
bestimmten Krankheit ist, auch wenn in-vivo-Studien zu chronischer Mastitis, Rhinosinusitis, septischer
Arthritis und Osteomyelitis den Verdacht nahe legen (Clement et al. 2005; Hebert et al. 2000; Hamza et
al. 2013; Mal et al. 2013; Bosse et al. 2005; Gresham et al. 2000).

DISKUSSION 66
S. aureus stellt damit, unserer Meinung nach und bezogen auf den derzeitigen Wissensstand,
kein typisches fakultativ intrazelluläres Bakterium dar. Dennoch ist der hier gebrachte Nachweis der
intrazellulären Persistenz von S. aureus in Makrophagen möglicherweise eine der Ursachen für schwer
zu therapierende bzw. chronisch-rekurrente Krankheitsverläufe.
Es ist denkbar, dass S. aureus durch den vorübergehenden intrazellulären Aufenthalt im
Makrophagen an andere Stellen des Organismus gelangt, wodurch es zur Disseminierung der Infektion
kommt. Vermutlich verlässt er die Zellen dann wieder und vermehrt sich. Kubica et al. formulierten bereits
diese Vermutung. Sie beobachteten in ihrer Studie eine ähnliche intrazelluläre Überlebenskurve, wie sie
hier gesehen werden konnte. Nach etwa 5 Tagen verließ S. aureus die Makrophagen und proliferierte
massiv extrazellulär (Kubica et al. 2008). Aufgrund des hier eingesetzten Antibiotika-haltigen Mediums im
Vergleich zur Arbeit von Kubica et al. konnte dies in vorliegenden Ergebnissen nicht beobachtet werden.
Es weist aber darauf hin, dass S. aureus die Makrophagen tatsächlich als kurzfristiges
Überlebensreservoir zu nutzen vermag, um dann die Zelle wieder zu verlassen und e xtrazellulär zu
proliferieren.
In diesen Zusammenhang ist erwähnenswert, dass intrazellulärem S. aureus eine zytoprotektive
Wirkung auf Makrophagen zugeschrieben werden konnte (Kubica et al. 2008), die im Gegensatz zu
apoptotischen Effekten in anderen Zellen steht (Hess et al. 2003; Menzies und Kourteva 1998; Haslinger-
Loffler et al. 2006; Baran et al. 2001). Ähnlich wird es bekanntermaßen von obligat- bzw. fakultativ
intrazellulären Erregern zur Erhaltung ihrer intrazellulären Nische angewandt (Fischer et al. 2001; Hacker
et al. 2006; Abu-Zant et al. 2005). S. aureus induziert nach der Phagozytose die Expression
antiapoptotischer Faktoren des Makrophagen und verlängert so dessen Lebenszyklus. Die Induktion der
Veränderung der Genexpression der Wirtszelle erinnert dabei an fakultativ- und obligat intrazelluläre
Bakterien (Koziel et al. 2009; Kubica et al. 2008; Koziel et al. 2013; Garzoni et al. 2007). In vorliegender
Dissertation zeigten sich entsprechende Ergebnisse. Durch Messung der Zytotoxizität (Daten nicht
gezeigt), sowie lichtmikroskopische Beurteilung der Morphologie und Adhärenz der Zellen war erkennbar,
dass die infizierten Makrophagen über den beobachteten Zeitraum von 5 Tagen größtenteils vital blieben.
Ein weiterer Erklärungsansatz im Hinblick auf die beobachteten Überlebenskurven von S. aureus
in Makrophagen ist, dass er nach etwa 5 Tagen der Wirtsabwehr erliegt und sich bis dahin das
Gleichgewicht zwischen bakteriziden Mechanismen des Makrophagen und der daran angepassten
Expression von bakteriellen Virulenzfaktoren über eine kurze Zeit hält.
In jedem Fall kann die Fähigkeit, zumindest über einen gewissen Zeitraum intrazellulär in
Makrophagen persistieren zu können, als pathogenes Potenzial von S. aureus gesehen werden. Diese
Eigenschaft, die bei S. epidermidis in dieser Arbeit nicht in gleichem Maße beobachtet werden konnte,

DISKUSSION 67
könnte auch mitbestimmend sein für den Unterschied in der Pathogenität zwischen S. aureus und
Koagulase-negativen Staphylokokken.

68
5 ZUSAMMENFASSUNG
Im Rahmen vorliegender Dissertation wurde das intrazelluläre Überleben von S. aureus in Makrophagen
charakterisiert. Dabei lag ein Schwerpunkt auf der Verwendung von primären murinen Knochenmark-
makrophagen, deren Interaktion mit S. aureus bisher wenig beschrieben ist. Die Internalisierung und
Persistenz von S. aureus in primären Makrophagen könnte dabei als Pathogenitätsmechanismus für die
therapieresistenten und persistierenden, sowie rekurrenten Infektionen relevant sein, die der Erreger
neben den klassischen Manifestationen einer S. aureus-Infektion hervorruft. In diesem Zusammenhang
sollte auch diskutiert werden, ob es sich bei S. aureus um ein typisches fakultativ intrazelluläres Bakterium
handelt.
Zunächst wurde anhand eines in-vitro-Infektionsmodells gezeigt, dass verschiedene S. aureus–
Stämme in einer murinen Makrophagen-Zelllinie, sowie in primären murinen Knochenmark-Makrophagen
mindestens 5 Tage unter langsamer Reduktion der anfänglichen Keimzahl überleben können. Der als
Kontrollstamm eingesetzte KNS-Vertreter S. epidermidis ATCC 12228 konnte etwa 3 Tage intrazellulär
nachgewiesen werden.
Die gleichen S. aureus-Stämme wurden bezüglich ihrer Internalisierungsrate untersucht. Dabei
zeigte sich, dass die S. aureus-Stämme Newman und RN 6390 im Vergleich zu den S. aureus ATCC-
Stämmen 25923, 12600 und 29213 deutlich geringer internalisiert wurden. Sogar S. epidermidis ATCC
12228 wies ihnen gegenüber eine höhere Phagozytoserate auf. Dies verdeutlicht die Begrenzbarkeit von
Studien, in denen unterschiedliche S. aureus-Stämme untersucht werden. Die geringen
Internalisierungsraten von Newman und RN 6390 lassen sich dabei aus ihren bekannten regulatorischen
bzw. funktionellen Gendefekten erklären.
Im Rahmen dieser Arbeit wurde darüber hinaus ein Infektionsmodell entwickelt, in dem
Makrophagen von BALB/c- und C57BL/6-Mäusen vergleichend bezüglich Phagozytose und
intrazellulärem Überleben von S. aureus untersucht wurden. Anders als uns von in-vivo-
Infektionsmodellen mit S. aureus bekannt ist, zeigten sich in-vitro keine Unterschiede in der
Eliminationsfähigkeit von primären Knochenmarkmakrophagen von C57BL/6- und BALB/c-Mäusen
gegenüber intrazellulärem S. aureus. Die genetisch bedingte unterschiedliche Empfänglichkeit der
verschiedenen Mausstämme gegenüber einer S. aureus-Infektion beruht wahrscheinlich nicht auf den
unterschiedlichen bakteriziden Mechanismen der Makrophagen.
Es erfolgten weiterhin Experimente mit primären Knochenmarkmakrophagen von Mäusen mit
Defekten im iNOS Gen bzw. der gp91 Untereinheit der NADPH-Oxidase, um die Makrophagen-
spezifischen Abwehrmechanismen gegenüber einer S. aureus-Infektion genauer zu charakterisieren.

ZUSAMMENFASSUNG 69
Dabei konnte kein Effekt von bakteriziden oxidativen, oder nitrosativen Effektormechanismen des
Makrophagen auf die Phagozytose, oder das intrazelluläre Überleben von S. aureus festgestellt werden,
obwohl S. aureus die NO-Produktion in Makrophagen induzierte.
Zur Untersuchung der bakteriellen Regulation in der Interaktion mit Makrophagen wurden
schließlich S. aureus-Stämme mit Mutationen in den globalen regulatorischen Systemen agr bzw. sae
eingesetzt. Agr und sae sind essentielle Regulatoren der S. aureus-Pathogenitätsfaktoren. In
vorliegenden Ergebnissen zeigte sich deren Einfluss vor allem bezüglich der Phagozytoserate. Das
intrazelluläre Überleben hingegen wurde nur unwesentlich beeinflusst. Während sae sich als
entscheidend für die Internalisierung erwies, zeigte agr einen gegensätzlichen Effekt. Agr zeigte sich
hingegen verantwortlich für die initiale, kurzzeitige Proliferation von S. aureus in Makrophagen. Weiterhin
stellten sich das S. aureus-Oberflächenprotein FnBPA, als auch das extrazelluläre Matrixprotein
Fibronektin für die Internalisierung in primäre Makrophagen als essentiell heraus. Diese Ergebnisse
zeigten, dass die intrazelluläre Aufnahme von S. aureus in Makrophagen durch verschiedene bakterielle
Komponenten stark beeinflusst wird.

70
6 LITERATURVERZEICHNIS
1. Abel, Jens; Goldmann, Oliver; Ziegler, Christina; Holtje, Claudia; Smeltzer, Mark S.; Cheung,
Ambrose L. et al. (2011): Staphylococcus aureus evades the extracellular antimicrobial activity
of mast cells by promoting its own uptake. In: J Innate Immun 3 (5), S. 495–507. DOI:
10.1159/000327714.
2. Abu-Zant, Alaeddin; Santic, Marina; Molmeret, Maelle; Jones, Snake; Helbig, Jurgen; Abu
Kwaik, Yousef (2005): Incomplete activation of macrophage apoptosis during intracellular
replication of Legionella pneumophila. In: Infect Immun 73 (9), S. 5339–5349. DOI:
10.1128/IAI.73.9.5339-5349.2005.
3. Agerer, Franziska; Lux, Sigrid; Michel, Antje; Rohde, Manfred; Ohlsen, Knut; Hauck, Christof R.
(2005): Cellular invasion by Staphylococcus aureus reveals a functional link between focal
adhesion kinase and cortactin in integrin-mediated internalisation. In: J Cell Sci 118 (Pt 10), S.
2189–2200. DOI: 10.1242/jcs.02328.
4. Agerer, Franziska; Michel, Antje; Ohlsen, Knut; Hauck, Christof R. (2003): Integrin-mediated
invasion of Staphylococcus aureus into human cells requires Src family protein-tyrosine kinases.
In: J Biol Chem 278 (43), S. 42524–42531. DOI: 10.1074/jbc.M302096200.
5. Alberts, Bruce (2011): Molekularbiologie der Zelle. 5., vollst. überarb. Aufl. Weinheim: Wiley -
VCH.
6. Alva-Murillo, Nayeli; López-Meza, Joel Edmundo; Ochoa-Zarzosa, Alejandra (2014):
Nonprofessional phagocytic cell receptors involved in Staphylococcus aureus internalization. In:
BioMed research international 2014, S. 538546. DOI: 10.1155/2014/538546.
7. Amer, Amal O.; Swanson, Michele S. (2002): A phagosome of one's own: a microbial guide to
life in the macrophage. In: Curr Opin Microbiol 5 (1), S. 56–61.
8. Arrecubieta, Carlos; Lee, Mei-Ho; Macey, Alistair; Foster, Timothy J.; Lowy, Franklin D. (2007):
SdrF, a Staphylococcus epidermidis surface protein, binds type I collagen. In: J Biol Chem 282
(26), S. 18767–18776. DOI: 10.1074/jbc.M610940200.
9. Asad, Shadaba; Opal, Steven M. (2008): Bench-to-bedside review: Quorum sensing and the
role of cell-to-cell communication during invasive bacterial infection. In: Crit Care 12 (6), S. 236.
DOI: 10.1186/cc7101.

LITERATURVERZEICHNIS 71
10. Bantel, H.; Sinha, B.; Domschke, W.; Peters, G.; Schulze -Osthoff, K.; Janicke, R. U. (2001):
alpha-Toxin is a mediator of Staphylococcus aureus-induced cell death and activates caspases
via the intrinsic death pathway independently of death receptor signaling. In: J Cell Biol 155 (4),
S. 637–648. DOI: 10.1083/jcb.200105081.
11. Baran, J.; Weglarczyk, K.; Mysiak, M.; Guzik, K.; Ernst, M.; Flad, H. D.; Pryjma, J. (2001): Fas
(CD95)-Fas ligand interactions are responsible for monocyte apoptosis occurring as a result of
phagocytosis and killing of Staphylococcus aureus. In: Infect Immun 69 (3), S. 1287–1297. DOI:
10.1128/IAI.69.3.1287-1297.2001.
12. Bayles, K. W.; Wesson, C. A.; Liou, L. E.; Fox, L. K.; Bohach, G. A.; Trumble, W. R. (1998):
Intracellular Staphylococcus aureus escapes the endosome and induces apoptosis in epithelial
cells. In: Infect Immun 66 (1), S. 336–342.
13. Bera, Agnieszka; Biswas, Raja; Herbert, Silvia; Gotz, Friedrich (2006): The presence of
peptidoglycan O-acetyltransferase in various staphylococcal species correlates with lysozyme
resistance and pathogenicity. In: Infect Immun 74 (8), S. 4598–4604. DOI: 10.1128/IAI.00301-
06.
14. Bischoff, M.; Entenza, J. M.; Giachino, P. (2001): Influence of a functional sigB operon on the
global regulators sar and agr in Staphylococcus aureus. In: J Bacteriol 183 (17), S. 5171–5179.
15. Bishayi, B.; Bandyopadhyay, D.; Majhi, A.; Adhikary, R. (2014): Possible role of Toll-like
receptor-2 in the intracellular survival of Staphylococcus aureus in murine peritoneal
macrophages: involvement of cytokines and anti-oxidant enzymes. In: Scand J Immunol 80 (2),
S. 127–143. DOI: 10.1111/sji.12195.
16. Blanchet, Charlene; Jouvion, Gregory; Fitting, Catherine; Cavaillon, Jean-Marc; Adib-Conquy,
Minou (2014): Protective or Deleterious Role of Scavenger Receptors SR-A and CD36 on Host
Resistance to Staphylococcus aureus Depends on the Site of Infection. In: PLoS One 9 (1), S.
e87927. DOI: 10.1371/journal.pone.0087927.
17. Blevins, Jon S.; Beenken, Karen E.; Elasri, Mohamed O.; Hurlburt, Barry K.; Smeltzer, Mark S.
(2002): Strain-dependent differences in the regulatory roles of sarA and agr in Staphylococcus
aureus. In: Infect Immun 70 (2), S. 470–480.
18. Blos, Martin; Schleicher, Ulrike; Soares Rocha, F Janaina; Meissner, Udo; Rollinghoff, Martin;
Bogdan, Christian (2003): Organ-specific and stage-dependent control of Leishmania major
infection by inducible nitric oxide synthase and phagocyte NADPH oxidase. In: Eur J Immunol
33 (5), S. 1224–1234. DOI: 10.1002/eji.200323825.

LITERATURVERZEICHNIS 72
19. Boelens, J. J.; Dankert, J.; Murk, J. L.; Weening, J. J.; van der Poll, T; Dingemans, K. P. et al.
(2000): Biomaterial-associated persistence of Staphylococcus epidermidis in pericatheter
macrophages. In: J INFECT DIS 181 (4), S. 1337–1349. DOI: 10.1086/315369.
20. Bogdan, C.; Rollinghoff, M.; Diefenbach, A. (2000a): Reactive oxygen and reactive nitrogen
intermediates in innate and specific immunity. In: Curr Opin Immunol 12 (1), S. 64–76.
21. Bogdan, C.; Rollinghoff, M.; Diefenbach, A. (2000b): The role of nitric oxide in innate immunity.
In: Immunol Rev 173, S. 17–26.
22. Bosse, Michael J.; Gruber, Helen E.; Ramp, Warren K. (2005): Internalization of bacteria by
osteoblasts in a patient with recurrent, long-term osteomyelitis. A case report. In: J Bone Joint
Surg Am 87 (6), S. 1343–1347. DOI: 10.2106/JBJS.D.02649.
23. Bowden, M. Gabriela; Chen, Wei; Singvall, Jenny; Xu, Yi; Peacock, Sharon J.; Valtulina, Viviana
et al. (2005): Identification and preliminary characterization of cell -wall-anchored proteins of
Staphylococcus epidermidis. In: Microbiology 151 (Pt 5), S. 1453–1464. DOI:
10.1099/mic.0.27534-0.
24. Breitbach, Katrin; Klocke, Sonja; Tschernig, Thomas; van Rooijen, Nico; Baumann, Ulrich;
Steinmetz, Ivo (2006): Role of inducible nitric oxide synthase and NADPH oxidase in early control
of Burkholderia pseudomallei infection in mice. In: Infect Immun 74 (11), S. 6300–6309. DOI:
10.1128/IAI.00966-06.
25. Broekhuizen, C A N; Sta, M.; Vandenbroucke-Grauls, C M J E; Zaat, S A J (2010): Microscopic
detection of viable Staphylococcus epidermidis in peri-implant tissue in experimental biomaterial-
associated infection, identified by bromodeoxyuridine incorporation. In: Infect Immun 78 (3), S.
954–962. DOI: 10.1128/IAI.00849-09.
26. Broekhuizen, Corine A N; Boer, Leonie de; Schipper, Kim; Jones, Christopher D.; Quadir, Shan;
Vandenbroucke-Grauls, Christina M J E; Zaat, Sebastian A J (2008): Staphylococcus
epidermidis is cleared from biomaterial implants but persists in peri-implant tissue in mice despite
rifampicin/vancomycin treatment. In: J Biomed Mater Res A 85 (2), S. 498–505. DOI:
10.1002/jbm.a.31528.
27. Bronner, Stéphane; Monteil, Henri; Prévost, Gilles (2004): Regulation of virulence determinants
in Staphylococcus aureus: complexity and applications. In: FEMS Microbiol. Rev. 28 (2), S. 183–
200. DOI: 10.1016/j.femsre.2003.09.003.

LITERATURVERZEICHNIS 73
28. Brouillette, Eric; Grondin, Gilles; Shkreta, Lulzim; Lacasse, Pierre; Talbot, Brian G. (2003): In
vivo and in vitro demonstration that Staphylococcus aureus is an intracellular pathogen in the
presence or absence of fibronectin-binding proteins. In: Microb Pathog 35 (4), S. 159–168.
29. Bur, Stephanie; Preissner, Klaus T.; Herrmann, Mathias; Bischoff, Markus (2013a): The
Staphylococcus aureus extracellular adherence protein promotes bacterial internalization by
keratinocytes independent of fibronectin-binding proteins. In: J Invest Dermatol 133 (8), S. 2004–
2012. DOI: 10.1038/jid.2013.87.
30. Bur, Stephanie; Preissner, Klaus T.; Herrmann, Mathias; Bischoff, Markus (2013b): The
Staphylococcus aureus extracellular adherence pro tein promotes bacterial internalization by
keratinocytes independent of fibronectin-binding proteins. In: J Invest Dermatol 133 (8), S. 2004–
2012. DOI: 10.1038/jid.2013.87.
31. Casey, A. L.; Lambert, P. A.; Elliott, T S J (2007): Staphylococci. In: Int J Antimicrob Agents 29
Suppl 3, S. S23-32. DOI: 10.1016/S0924-8579(07)72175-1.
32. Chakravortty, Dipshikha; Hensel, Michael (2003): Inducible nitric oxide synthase and control of
intracellular bacterial pathogens. In: Microbes Infect 5 (7), S. 621–627.
33. Chamberlain, Lisa M.; Godek, Marisha L.; Gonzalez-Juarrero, Mercedes; Grainger, David W.
(2009): Phenotypic non-equivalence of murine (monocyte-) macrophage cells in biomaterial and
inflammatory models. In: J Biomed Mater Res A 88 (4), S. 858–871. DOI: 10.1002/jbm.a.31930.
34. Cheung, Ambrose L.; Bayer, Arnold S.; Zhang, Gongyi; Gresham, Hattie; Xiong, Yan-Qiong
(2004): Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. In:
FEMS Immunol. Med. Microbiol. 40 (1), S. 1–9.
35. Cheung, Gordon Y C; Rigby, Kevin; Wang, Rong; Queck, Shu Y.; Braughton, Kevin R.; Whitney,
Adeline R. et al. (2010): Staphylococcus epidermidis strategies to avoid killing by human
neutrophils. In: PLoS Pathog 6 (10), S. e1001133. DOI: 10.1371/journal.ppat.1001133.
36. Clauditz, Alexandra; Resch, Alexandra; Wieland, Karsten-Peter; Peschel, Andreas; Gotz,
Friedrich (2006): Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its
ability to cope with oxidative stress. In: Infect Immun 74 (8), S. 4950–4953. DOI:
10.1128/IAI.00204-06.
37. Clement, Sophie; Vaudaux, Pierre; Francois, Patrice; Schrenzel, Jacques; Huggler, Elzbieta;
Kampf, Sandy et al. (2005): Evidence of an intracellular reservoir in the nasal mucosa of patients

LITERATURVERZEICHNIS 74
with recurrent Staphylococcus aureus rhinosinusitis. In: J INFECT DIS 192 (6), S. 1023–1028.
DOI: 10.1086/432735.
38. Cunha, F. Q.; Assreuy, J.; Moncada, S.; Liew, F. Y. (1993a): Phagocytosis and induction of nitric
oxide synthase in murine macrophages. In: Immunology 79 (3), S. 408–411.
39. Cunha, F. Q.; Moss, D. W.; Leal, L. M.; Moncada, S.; Liew, F. Y. (1993b): Induction of
macrophage parasiticidal activity by Staphylococcus aureus and exotoxins through the nitric
oxide synthesis pathway. In: Immunology 78 (4), S. 563–567.
40. da Silva Domingues, Joana F; van der Mei, Henny C; Busscher, Henk J.; van Kooten, Theo G
(2013): Phagocytosis of bacteria adhering to a biomaterial surface in a surface thermodynamic
perspective. In: PLoS One 8 (7), S. e70046. DOI: 10.1371/journal.pone.0070046.
41. Das, D.; Saha, S. S.; Bishayi, B. (2008): Intracellular survival of Staphylococcus aureus:
correlating production of catalase and superoxide dismutase with levels of inflammatory
cytokines. In: Inflamm Res 57 (7), S. 340–349. DOI: 10.1007/s00011-007-7206-z.
42. Das, Debaditya; Bishayi, Biswadev (2009): Staphylococcal catalase protects intracellularly
survived bacteria by destroying H2O2 produced by the murine peritoneal macrophages. In:
Microb Pathog 47 (2), S. 57–67. DOI: 10.1016/j.micpath.2009.04.012.
43. Das, Debaditya; Bishayi, Biswadev (2010): Contribution of Catalase and Superoxide Dismutase
to the Intracellular Survival of Clinical Isolates of Staphylococcus aureus in Murine Macrophages.
In: Indian J Microbiol 50 (4), S. 375–384. DOI: 10.1007/s12088-011-0063-z.
44. DeLeo, Frank R.; Diep, Binh An; Otto, Michael (2009): Host defense and pathogenesis in
Staphylococcus aureus infections. In: Infect Dis Clin North Am 23 (1), S. 17–34. DOI:
10.1016/j.idc.2008.10.003.
45. DeLoid, Glen M.; Sulahian, Timothy H.; Imrich, Amy; Kobzik, Lester (2009): Heterogeneity in
macrophage phagocytosis of Staphylococcus aureus strains: high-throughput scanning
cytometry-based analysis. In: PLoS One 4 (7), S. e6209. DOI: 10.1371/journal.pone.0006209.
46. Depke, Maren; Breitbach, Katrin; Dinh Hoang Dang, Khoa; Brinkmann, Lars; Salazar, Manuela
Gesell; Dhople, Vishnu Mukund et al. (2014): Bone marrow-derived macrophages from BALB/c
and C57BL/6 mice fundamentally differ in their respiratory chain complex proteins, lysosomal
enzymes and components of antioxidant stress systems. In: J Proteomics 103C, S. 72–86. DOI:
10.1016/j.jprot.2014.03.027.

LITERATURVERZEICHNIS 75
47. Dewamitta, Sita R.; Nomura, Takamasa; Kawamura, Ikuo; Hara, Hideki; Tsuchiya, Kohsuke;
Kurenuma, Takeshi et al. (2010): Listeriolysin O-dependent bacterial entry into the cytoplasm is
required for calpain activation and interleukin-1 alpha secretion in macrophages infected with
Listeria monocytogenes. In: Infect Immun 78 (5), S. 1884–1894. DOI: 10.1128/IAI.01143-09.
48. Dey, Somrita; Bishayi, Biswadev (2015): Killing of Staphylococcus aureus in murine
macrophages by chloroquine used alone and in combination with ciprofloxacin or azithromycin.
In: J Inflamm Res 8, S. 29–47. DOI: 10.2147/JIR.S76045.
49. Dziewanowska, K.; Patti, J. M.; Deobald, C. F.; Bayles, K. W.; Trumble, W. R.; Bohac h, G. A.
(1999): Fibronectin binding protein and host cell tyrosine kinase are required for internalization
of Staphylococcus aureus by epithelial cells. In: Infect Immun 67 (9), S. 4673–4678.
50. Edwards, Andrew M.; Potter, Ursula; Meenan, Nicola A G; Potts, Jennifer R.; Massey, Ruth C.
(2011): Staphylococcus aureus keratinocyte invasion is dependent upon multiple high-affinity
fibronectin-binding repeats within FnBPA. In: PLoS One 6 (4), S. e18899. DOI:
10.1371/journal.pone.0018899.
51. Edwards, Andrew M.; Potts, Jennifer R.; Josefsson, Elisabet; Massey, Ruth C. (2010):
Staphylococcus aureus host cell invasion and virulence in sepsis is facilitated by the multiple
repeats within FnBPA. In: PLoS Pathog 6 (6), S. e1000964. DOI: 10.1371/journal.ppat.1000964.
52. Eiff, C. von; Becker, K.; Machka, K.; Stammer, H.; Peters, G. (2001a): Nasal carriage as a source
of Staphylococcus aureus bacteremia. Study Group. In: N Engl J Med 344 (1), S. 11–16. DOI:
10.1056/NEJM200101043440102.
53. Eiff, C. von; Proctor, R. A.; Peters, G. (2001b): Coagulase-negative staphylococci. Pathogens
have major role in nosocomial infections. In: Postgrad Med 110 (4), S. 63-4, 69-70, 73-6.
54. Eiff, Christof von; Peters, Georg; Becker, Karsten (2006): The small colony variant (SCV)
concept -- the role of staphylococcal SCVs in persistent infections. In: Injury 37 Suppl 2, S. S26-
33. DOI: 10.1016/j.injury.2006.04.006.
55. Eiff, Christof von; Peters, Georg; Heilmann, Christine (2002): Pathogenesis of infections due to
coagulase-negative staphylococci. In: Lancet Infect Dis 2 (11), S. 677–685.
56. Ellington, J. K.; Harris, M.; Webb, L.; Smith, B.; Smith, T.; Tan, K.; Hudson, M. (2003):
Intracellular Staphylococcus aureus. A mechanism for the indolence of osteomyelitis. In: J Bone
Joint Surg Br 85 (6), S. 918–921.

LITERATURVERZEICHNIS 76
57. Ellson, Chris D.; Davidson, Keith; Ferguson, G. John; O'Connor, Rod; Stephens, Len R.;
Hawkins, Phillip T. (2006): Neutrophils from p40phox-/- mice exhibit severe defects in NADPH
oxidase regulation and oxidant-dependent bacterial killing. In: J Exp Med 203 (8), S. 1927–1937.
DOI: 10.1084/jem.20052069.
58. Eske, Kristin; Breitbach, Katrin; Kohler, Jens; Wongprompitak, Patimaporn; Steinmetz, Ivo
(2009): Generation of murine bone marrow derived macrophages in a standardised serum-free
cell culture system. In: J Immunol Methods 342 (1-2), S. 13–19. DOI: 10.1016/j.jim.2008.11.011.
59. Fang, Ferric C.; Vazquez-Torres, Andres (2002): Nitric oxide production by human
macrophages: there's NO doubt about it. In: Am J Physiol Lung Cell Mol Physiol 282 (5), S.
L941-3. DOI: 10.1152/ajplung.00017.2002.
60. Finlay, B. B.; Cossart, P. (1997): Exploitation of mammalian host cell functions by bacterial
pathogens. In: Science 276 (5313), S. 718–725.
61. Fischer, S. F.; Schwarz, C.; Vier, J.; Hacker, G. (2001): Characterization of antiapoptotic
activities of Chlamydia pneumoniae in human cells. In: Infect Immun 69 (11), S. 7121–7129.
DOI: 10.1128/IAI.69.11.7121-7129.2001.
62. Foster, Timothy J. (2005): Immune evasion by staphylococci. In: Nat Rev Microbiol 3 (12), S.
948–958. DOI: 10.1038/nrmicro1289.
63. Foster, Timothy J. (2009): Colonization and infection of the human host by staphylococci:
adhesion, survival and immune evasion. In: Vet. Dermatol. 20 (5-6), S. 456–470. DOI:
10.1111/j.1365-3164.2009.00825.x.
64. Foster, Timothy J.; Geoghegan, Joan A.; Ganesh, Vannakambadi K.; Höök, Magnus (2014):
Adhesion, invasion and evasion: the many functions of the surface proteins of Staphylococcus
aureus. In: Nat. Rev. Microbiol. 12 (1), S. 49–62. DOI: 10.1038/nrmicro3161.
65. Fournier, Benedicte (2012): The function of TLR2 during staphylococcal diseases. In: Front Cell
Infect Microbiol 2, S. 167. DOI: 10.3389/fcimb.2012.00167.
66. Fournier, Benedicte; Philpott, Dana J. (2005): Recognition of Staphylococcus aureus by the
innate immune system. In: Clin Microbiol Rev 18 (3), S. 521–540. DOI: 10.1128/CMR.18.3.521-
540.2005.
67. Fowler, T.; Wann, E. R.; Joh, D.; Johansson, S.; Foster, T. J.; Hook, M. (2000): Cellular invasion
by Staphylococcus aureus involves a fibronectin bridge between the bacterial fibronectin-binding
MSCRAMMs and host cell beta1 integrins. In: Eur J Cell Biol 79 (10), S. 672–679.

LITERATURVERZEICHNIS 77
68. Fraunholz, Martin; Sinha, Bhanu (2012): Intracellular Staphylococcus aureus: live-in and let die.
In: Front Cell Infect Microbiol 2, S. 43. DOI: 10.3389/fcimb.2012.00043.
69. Gan, Yunn-Hwen (2005): Interaction between Burkholderia pseudomallei and the host immune
response: sleeping with the enemy? In: J INFECT DIS 192 (10), S. 1845–1850. DOI:
10.1086/497382.
70. Garzoni, Christian; Francois, Patrice; Huyghe, Antoine; Couzinet, Sabine; Tapparel, Caroline;
Charbonnier, Yvan et al. (2007): A global view of Staphylococcus aureus whole genome
expression upon internalization in human epithelial cells. In: BMC Genomics 8, S. 171. DOI:
10.1186/1471-2164-8-171.
71. Garzoni, Christian; Kelley, William L. (2009): Staphylococcus aureus: new evidence for
intracellular persistence. In: Trends Microbiol 17 (2), S. 59–65. DOI: 10.1016/j.tim.2008.11.005.
72. Garzoni, Christian; Kelley, William L. (2011): Return of the Trojan horse: intracellular phenotype
switching and immune evasion by Staphylococcus aureus. In: EMBO Mol Med 3 (3), S. 115–
117. DOI: 10.1002/emmm.201100123.
73. Gaupp, Rosmarie; Ledala, Nagender; Somerville, Greg A. (2012): Staphylococcal response to
oxidative stress. In: Front Cell Infect Microbiol 2, S. 33. DOI: 10.3389/fcimb.2012.00033.
74. Geiger, Tobias; Goerke, Christiane; Mainiero, Markus; Kraus, Dirk; Wolz, Christiane (2008): The
virulence regulator Sae of Staphylococcus aureus: promoter activities and response to
phagocytosis-related signals. In: J Bacteriol 190 (10), S. 3419–3428. DOI: 10.1128/JB.01927-
07.
75. Gertz, S.; Engelmann, S.; Schmid, R.; Ohlsen, K.; Hacker, J.; Hecker, M. (1999): Regulation of
sigmaB-dependent transcription of sigB and asp23 in two different Staphylococcus aureus
strains. In: Mol Gen Genet 261 (3), S. 558–566.
76. Gertz, S.; Engelmann, S.; Schmid, R.; Ziebandt, A. K.; Tischer, K.; Scharf, C. et al. (2000):
Characterization of the sigma(B) regulon in Staphylococcus aureus. In: J Bacteriol 182 (24), S.
6983–6991.
77. Giachino, P.; Engelmann, S.; Bischoff, M. (2001): Sigma(B) activity depends on RsbU in
Staphylococcus aureus. In: J Bacteriol 183 (6), S. 1843–1852. DOI: 10.1128/JB.183.6.1843-
1852.2001.
78. Giese, Bernd; Glowinski, Frithjof; Paprotka, Kerstin; Dittmann, Silvia; Steiner, Tobias; Sinha,
Bhanu; Fraunholz, Martin J. (2011a): Expression of delta-toxin by Staphylococcus aureus

LITERATURVERZEICHNIS 78
mediates escape from phago-endosomes of human epithelial and endothelial cells in the
presence of beta-toxin. In: Cell Microbiol 13 (2), S. 316–329. DOI: 10.1111/j.1462-
5822.2010.01538.x.
79. Giese, Bernd; Glowinski, Frithjof; Paprotka, Kerstin; Dittmann, Silvia; Steiner, Tobias; Sinha,
Bhanu; Fraunholz, Martin J. (2011b): Expression of δ -toxin by Staphylococcus aureus mediates
escape from phago-endosomes of human epithelial and endothelial cells in the presence of β-
toxin. In: Cell. Microbiol. 13 (2), S. 316–329. DOI: 10.1111/j.1462-5822.2010.01538.x.
80. Gill, Steven R.; Fouts, Derrick E.; Archer, Gordon L.; Mongodin, Emmanuel F.; Deboy, Robert
T.; Ravel, Jacques et al. (2005): Insights on evolution of virulence and resistance from the
complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a
biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. In: J Bacteriol 187 (7),
S. 2426–2438. DOI: 10.1128/JB.187.7.2426-2438.2005.
81. Goerke, C.; Fluckiger, U.; Steinhuber, A.; Zimmerli, W.; Wolz, C. (2001): Impact of the regulatory
loci agr, sarA and sae of Staphylococcus aureus on the induction of alpha-toxin during device-
related infection resolved by direct quantitative transcript analysis. In: Mol Microbiol 40 (6), S.
1439–1447.
82. Gotz, Friedrich (2002): Staphylococcus and biofilms. In: Mol Microbiol 43 (6), S. 1367–1378.
83. Gresham, H. D.; Lowrance, J. H.; Caver, T. E.; Wilson, B. S.; Cheung, A. L.; Lindberg, F. P.
(2000): Survival of Staphylococcus aureus inside neutrophils contributes to infection. In: J
Immunol 164 (7), S. 3713–3722.
84. Grundmeier, Matthias; Hussain, Muzaffar; Becker, Petra; Heilmann, Christine; Peters, Georg;
Sinha, Bhanu (2004): Truncation of fibronectin-binding proteins in Staphylococcus aureus strain
Newman leads to deficient adherence and host cell invasion due to loss of the cell wall anchor
function. In: Infect Immun 72 (12), S. 7155–7163. DOI: 10.1128/IAI.72.12.7155-7163.2004.
85. Guzik, T. J.; Korbut, R.; Adamek-Guzik, T. (2003): Nitric oxide and superoxide in inflammation
and immune regulation. In: J Physiol Pharmacol 54 (4), S. 469–487.
86. Hacker, Georg; Kirschnek, Susanne; Fischer, Silke F. (2006): Apoptosis in infectious disease:
how bacteria interfere with the apoptotic apparatus. In: Med Microbiol Immunol 195 (1), S. 11–
19. DOI: 10.1007/s00430-005-0239-4.

LITERATURVERZEICHNIS 79
87. Haggar, Axana; Hussain, Muzaffar; Lonnies, Helena; Herrmann, Mathias; Norrby -Teglund,
Anna; Flock, Jan-Ingmar (2003): Extracellular adherence protein from Staphylococcus aureus
enhances internalization into eukaryotic cells. In: Infect Immun 71 (5), S. 2310–2317.
88. Hamza, T.; Dietz, M.; Pham, D.; Clovis, N.; Danley, S.; Li, B. (2013): Intra-cellular
Staphylococcus aureus alone causes infection in vivo. In: Eur Cell Mater 25, S. 341-50;
discussion 350.
89. Hamza, Therwa; Li, Bingyun (2014): Differential responses of osteoblasts and macrophages
upon Staphylococcus aureus infection. In: BMC Microbiol 14, S. 207. DOI: 10.1186/s12866-014-
0207-5.
90. Harraghy, Niamh; Hussain, Muzaffar; Haggar, Axana; Chavakis, Triantafyllos; Sinha, Bhanu;
Herrmann, Mathias; Flock, Jan-Ingmar (2003): The adhesive and immunomodulating properties
of the multifunctional Staphylococcus aureus protein Eap. In: Microbiology 149 (Pt 10), S. 2701–
2707.
91. Harraghy, Niamh; Kormanec, Jan; Wolz, Christiane; Homerova, Dagmar; Goerke, Christiane;
Ohlsen, Knut et al. (2005): sae is essential for expression of the staphylococcal adhesins Eap
and Emp. In: Microbiology 151 (Pt 6), S. 1789–1800. DOI: 10.1099/mic.0.27902-0.
92. Haslinger-Loffler, B.; Wagner, B.; Bruck, M.; Strangfeld, K.; Grundmeier, M.; Fischer, U. et al.
(2006): Staphylococcus aureus induces caspase-independent cell death in human peritoneal
mesothelial cells. In: Kidney Int 70 (6), S. 1089–1098. DOI: 10.1038/sj.ki.5001710.
93. Haslinger-Loffler, Bettina; Kahl, Barbara C.; Grundmeier, Matthias; Strangfeld, Katrin; Wagner,
Britta; Fischer, Ute et al. (2005): Multiple virulence factors are required for Staphylococcus
aureus-induced apoptosis in endothelial cells. In: Cell Microbiol 7 (8), S. 1087–1097. DOI:
10.1111/j.1462-5822.2005.00533.x.
94. Hauck, Christof R.; Borisova, Marina; Muenzner, Petra (2012): Exploitation of integrin function
by pathogenic microbes. In: Curr Opin Cell Biol 24 (5), S. 637–644. DOI:
10.1016/j.ceb.2012.07.004.
95. Hauck, Christof R.; Ohlsen, Knut (2006): Sticky connections: extracellular matrix protein
recognition and integrin-mediated cellular invasion by Staphylococcus aureus. In: Curr Opin
Microbiol 9 (1), S. 5–11. DOI: 10.1016/j.mib.2005.12.002.

LITERATURVERZEICHNIS 80
96. Hebert, A.; Sayasith, K.; Senechal, S.; Dubreuil, P.; Lagace, J. (2000): Demonstration of
intracellular Staphylococcus aureus in bovine mastitis alveolar cells and macrophages isolated
from naturally infected cow milk. In: FEMS Microbiol Lett 193 (1), S. 57–62.
97. Herbert, Silvia; Ziebandt, Anne-Kathrin; Ohlsen, Knut; Schafer, Tina; Hecker, Michael; Albrecht,
Dirk et al. (2010): Repair of global regulators in Staphylococcus aureus 8325 and comparative
analysis with other clinical isolates. In: Infect Immun 78 (6), S. 2877–2889. DOI:
10.1128/IAI.00088-10.
98. Hess, D. J.; Henry-Stanley, M. J.; Erickson, E. A.; Wells, C. L. (2003): Intracellular survival of
Staphylococcus aureus within cultured enterocytes. In: J Surg Res 114 (1), S. 42–49.
99. Hibbs, John B Jr (2002): Infection and nitric oxide. In: J INFECT DIS 185 Suppl 1, S. S9-17. DOI:
10.1086/338005.
100. Hirschhausen, Nina; Schlesier, Tim; Schmidt, M. Alexander; Gotz, Friedrich; Peters, Georg;
Heilmann, Christine (2010): A novel staphylococcal internalization mechanism involves the
major autolysin Atl and heat shock cognate protein Hsc70 as host cell receptor. In: Cell Microbiol
12 (12), S. 1746–1764. DOI: 10.1111/j.1462-5822.2010.01506.x.
101. Hoffmann, Christine; Ohlsen, Knut; Hauck, Christof R. (2011): Integrin-mediated uptake of
fibronectin-binding bacteria. In: Eur J Cell Biol 90 (11), S. 891–896. DOI:
10.1016/j.ejcb.2011.03.001.
102. Horsburgh, Malcolm J.; Aish, Joanne L.; White, Ian J.; Shaw, Les; Lithgow, James K.; Foster,
Simon J. (2002): sigmaB modulates virulence determinant expression and stress resistance:
characterization of a functional rsbU strain derived from Staphylococcus aureus 8325-4. In: J
Bacteriol 184 (19), S. 5457–5467.
103. Hussain, Muzaffar; Haggar, Axana; Heilmann, Christine; Peters, Georg; Flock, Jan-Ingmar;
Herrmann, Mathias (2002): Insertional inactivation of Eap in Staphylococcus aureus strain
Newman confers reduced staphylococcal binding to fibroblasts. In: Infect Immun 70 (6), S. 2933–
2940.
104. Jarry, Todd M.; Cheung, Ambrose L. (2006): Staphylococcus aureus escapes more efficiently
from the phagosome of a cystic fibrosis bronchial epithelial cell line than from its normal
counterpart. In: Infect Immun 74 (5), S. 2568–2577. DOI: 10.1128/IAI.74.5.2568-2577.2006.

LITERATURVERZEICHNIS 81
105. Jarry, Todd M.; Memmi, Guido; Cheung, Ambrose L. (2008): The exp ression of alpha-
haemolysin is required for Staphylococcus aureus phagosomal escape after internalization in
CFT-1 cells. In: Cell. Microbiol. 10 (9), S. 1801–1814. DOI: 10.1111/j.1462-5822.2008.01166.x.
106. Jenkins, Amy; Diep, Binh An; Mai, Thuy T.; Vo, Nhung H.; Warrener, Paul; Suzich, Joann et al.
(2015): Differential expression and roles of Staphylococcus aureus virulence determinants
during colonization and disease. In: mBio 6 (1), S. e02272-14. DOI: 10.1128/mBio.02272-14.
107. Kahl, B. C.; Goulian, M.; van Wamel, W.; Herrmann, M.; Simon, S. M.; Kaplan, G. et al. (2000):
Staphylococcus aureus RN6390 replicates and induces apoptosis in a pulmonary epithelial cell
line. In: Infect Immun 68 (9), S. 5385–5392.
108. Kang, Seok-Seong; Ryu, Young Hee; Baik, Jung Eun; Yun, Cheol-Heui; Lee, Kangseok; Chung,
Dae Kyun; Han, Seung Hyun (2011): Lipoteichoic acid from Lactobacillus plantarum induces
nitric oxide production in the presence of interferon-gamma in murine macrophages. In: Mol
Immunol 48 (15-16), S. 2170–2177. DOI: 10.1016/j.molimm.2011.07.009.
109. Keim, Luiz S.; Torres-Filho, Sylvio R.; Silva, Patricia Vollu; Teixeira, Lenise A. (2011):
Prevalence, aetiology and antibiotic resistance profiles of coagulase negative staphylococci
isolated in a teaching hospital. In: Braz J Microbiol 42 (1), S. 248–255. DOI: 10.1590/S1517-
83822011000100031.
110. Kerdudou, Sylvain; Laschke, Matthias W.; Sinha, Bhanu; Preissner, Klaus T.; Menger, Michael
D.; Herrmann, Mathias (2006): Fibronectin binding proteins contribute to the adherence of
Staphylococcus aureus to intact endothelium in vivo. In: Thromb Haemost 96 (2), S. 183–189.
111. Khalil, Hesham; Williams, Rachel J.; Stenbeck, Gudrun; Henderson, Brian; Meghji, Sajeda; Nair,
Sean P. (2007): Invasion of bone cells by Staphylococcus epidermidis. In: Microbes Infect 9 (4),
S. 460–465. DOI: 10.1016/j.micinf.2007.01.002.
112. Kim, Nam Joong; Ahn, Ki Bum; Jeon, Jun Ho; Yun, Cheol-Heui; Finlay, B. Brett; Han, Seung
Hyun (2015): Lipoprotein in the cell wall of Staphylococcus aureus is a major inducer of nitric
oxide production in murine macrophages. In: Mol Immunol 65 (1), S. 17–24. DOI:
10.1016/j.molimm.2014.12.016.
113. Kockritz-Blickwede, Maren von; Rohde, Manfred; Oehmcke, Sonja; Miller, Lloyd S.; Cheung,
Ambrose L.; Herwald, Heiko et al. (2008): Immunological mechanisms underlying the genetic
predisposition to severe Staphylococcus aureus infection in the mouse model. In: Am J Pathol
173 (6), S. 1657–1668. DOI: 10.2353/ajpath.2008.080337.

LITERATURVERZEICHNIS 82
114. Köhler, Jens; Breitbach, Katrin; Renner, Cäcilia; Heitsch, Anne-Katrin; Bast, Antje; van Rooijen,
Nico et al. (2011): NADPH-oxidase but not inducible nitric oxide synthase contributes to
resistance in a murine Staphylococcus aureus Newman pneumonia model. In: Microbes Infect.
13 (11), S. 914–922. DOI: 10.1016/j.micinf.2011.05.004.
115. Koziel, Joanna; Kmiecik, Katarzyna; Chmiest, Daniela; Maresz, Katarzyna; Mizgalska, Danuta;
Maciag-Gudowska, Agnieszka et al. (2013): The role of Mcl-1 in S. aureus-induced
cytoprotection of infected macrophages. In: Mediators Inflamm 2013, S. 427021. DOI:
10.1155/2013/427021.
116. Koziel, Joanna; Maciag-Gudowska, Agnieszka; Mikolajczyk, Tomasz; Bzowska, Malgorzata;
Sturdevant, Daniel E.; Whitney, Adeline R. et al. (2009): Phagocytosis of Staphylococcus aureus
by macrophages exerts cytoprotective effects manifested by the upregulation of antiapoptotic
factors. In: PLoS One 4 (4), S. e5210. DOI: 10.1371/journal.pone.0005210.
117. Krishna, Sheila; Miller, Lloyd S. (2012): Host-pathogen interactions between the skin and
Staphylococcus aureus. In: Curr Opin Microbiol 15 (1), S. 28–35. DOI:
10.1016/j.mib.2011.11.003.
118. Krut, Oleg; Utermohlen, Olaf; Schlossherr, Xenia; Kronke, Martin (2003): Strain-specific
association of cytotoxic activity and virulence of clinical Staphylococcus aureus isolates. In:
Infect Immun 71 (5), S. 2716–2723.
119. Kubica, Malgorzata; Guzik, Krzysztof; Koziel, Joanna; Zarebski, Miroslaw; Richter, Walter;
Gajkowska, Barbara et al. (2008): A potential new pathway for Staphylococcus aureus
dissemination: the silent survival of S. aureus phagocytosed by human monocyte -derived
macrophages. In: PLoS One 3 (1), S. e1409. DOI: 10.1371/journal.pone.0001409.
120. Kullik, I.; Giachino, P.; Fuchs, T. (1998): Deletion of the alternative sigma factor sigmaB in
Staphylococcus aureus reveals its function as a global regulator of virulence genes. In: J
Bacteriol 180 (18), S. 4814–4820.
121. Kuo, Chen-Tzu; Chiang, Ling-Ling; Lee, Chun-Nin; Yu, Ming-Chih; Bai, Kuan-Jen; Lee, Horng-
Mo et al. (2003): Induction of nitric oxide synthase in RAW 264.7 macrophages by lipoteichoic
acid from Staphylococcus aureus: involvement of protein kinase C- and nuclear factor-kB-
dependent mechanisms. In: J Biomed Sci 10 (1), S. 136–145.
122. Lazar Adler, Natalie R; Govan, Brenda; Cullinane, Meabh; Harper, Marina; Adler, Ben; Boyce,
John D. (2009): The molecular and cellular basis of pathogenesis in melioidosis: how does

LITERATURVERZEICHNIS 83
Burkholderia pseudomallei cause disease? In: FEMS Microbiol Rev 33 (6), S. 1079–1099. DOI:
10.1111/j.1574-6976.2009.00189.x.
123. Leelahavanichkul, Asada; Bocharov, Alexander V.; Kurlander, Roger; Baranova, Irina N.;
Vishnyakova, Tatyana G.; Souza, Ana C P et al. (2012): Class B scavenger receptor types I and
II and CD36 targeting improves sepsis survival and acute outcomes in mice. In: J Immunol 188
(6), S. 2749–2758. DOI: 10.4049/jimmunol.1003445.
124. Liang, Xudong; Yu, Chuanxin; Sun, Junsong; Liu, Hong; Landwehr, Christina; Holmes, David;
Ji, Yinduo (2006): Inactivation of a two-component signal transduction system, SaeRS,
eliminates adherence and attenuates virulence of Staphylococcus aureus. In: Infect Immun 74
(8), S. 4655–4665. DOI: 10.1128/IAI.00322-06.
125. Liew, F. Y. (1994): Regulation of nitric oxide synthesis in infectious and autoimmune diseases.
In: Immunol Lett 43 (1-2), S. 95–98.
126. Longauerova, A. (2006): Coagulase negative staphylococci and their participation in
pathogenesis of human infections. In: Bratisl Lek Listy 107 (11-12), S. 448–452.
127. Lowy, F. D. (1998): Staphylococcus aureus infections. In: N Engl J Med 339 (8), S. 520–532.
DOI: 10.1056/NEJM199808203390806.
128. Lowy, F. D. (2000): Is Staphylococcus aureus an intracellular pathogen? In: Trends Microbiol 8
(8), S. 341–343.
129. Lundqvist-Gustafsson, H.; Norrman, S.; Nilsson, J.; Wilsson, A. (2001): Involvement of p38-
mitogen-activated protein kinase in Staphylococcus aureus-induced neutrophil apoptosis. In: J
Leukoc Biol 70 (4), S. 642–648.
130. Mack, Dietrich; Davies, Angharad P.; Harris, Llinos G.; Rohde, Holger; Horstkotte, Matthias A.;
Knobloch, Johannes K-M (2007): Microbial interactions in Staphylococcus epidermidis biofilms.
In: Anal Bioanal Chem 387 (2), S. 399–408. DOI: 10.1007/s00216-006-0745-2.
131. MacMicking, J.; Xie, Q. W.; Nathan, C. (1997): Nitric oxide and macrophage function. In: Annu
Rev Immunol 15, S. 323–350. DOI: 10.1146/annurev.immunol.15.1.323.
132. Mainiero, Markus; Goerke, Christiane; Geiger, Tobias; Gonser, Christoph; Herbert, Silvia; Wolz,
Christiane (2010): Differential target gene activation by the Staphylococcus aureus two-
component system saeRS. In: J Bacteriol 192 (3), S. 613–623. DOI: 10.1128/JB.01242-09.
133. Mal, Pinky; Dutta, Kallol; Bandyopadhyay, Debasish; Basu, Anirban; Khan, Rajni; Bishayi,
Biswadev (2013): Azithromycin in combination with riboflavin decreases the severity of

LITERATURVERZEICHNIS 84
Staphylococcus aureus infection induced septic arthritis by modulating the production of free
radicals and endogenous cytokines. In: Inflammation research : official journal of the European
Histamine Research Society … [et al.] 62 (3), S. 259–273. DOI: 10.1007/s00011-012-0574-z.
134. Martinez-Pulgarin, Susana; Dominguez-Bernal, Gustavo; Orden, Jose A.; de la Fuente, Ricardo
(2009): Simultaneous lack of catalase and beta-toxin in Staphylococcus aureus leads to
increased intracellular survival in macrophages and epithelial cells and to attenuated virulence
in murine and ovine models. In: Microbiology 155 (Pt 5), S. 1505–1515. DOI:
10.1099/mic.0.025544-0.
135. Martins, Andre; Cunha, Maria de Lourdes R S (2007): Methicillin resistance in Staphylococcus
aureus and coagulase-negative staphylococci: epidemiological and molecular aspects. In:
Microbiol Immunol 51 (9), S. 787–795.
136. Menzies, B. E.; Kourteva, I. (1998): Internalization of Staphylococcus aureus by endothelial cells
induces apoptosis. In: Infect Immun 66 (12), S. 5994–5998.
137. Menzies, B. E.; Kourteva, I. (2000): Staphylococcus aureus alpha-toxin induces apoptosis in
endothelial cells. In: FEMS Immunol Med Microbiol 29 (1), S. 39–45.
138. Menzies, Barbara E. (2003): The role of fibronectin binding proteins in the pathogenesis of
Staphylococcus aureus infections. In: Curr Opin Infect Dis 16 (3), S. 225–229. DOI:
10.1097/01.qco.0000073771.11390.75.
139. Mestre, Maria Belen; Fader, Claudio M.; Sola, Claudia; Colombo, Maria Isabel (2010): Alpha-
hemolysin is required for the activation of the autophagic pathway in Staphylococcus aureus -
infected cells. In: Autophagy 6 (1), S. 110–125.
140. Mitchell, Gabriel; Lamontagne, Charles-Antoine; Brouillette, Eric; Grondin, Gilles; Talbot, Brian
G.; Grandbois, Michel; Malouin, François (2008): Staphylococcus aureus SigB activity promotes
a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small -
colony variants isolated from cystic fibrosis patients. In: Molecular Microbiology 70 (6), S. 1540–
1555. DOI: 10.1111/j.1365-2958.2008.06511.x.
141. Molmeret, Maelle; Bitar, Dina M.; Han, Lihui; Kwaik, Yousef Abu (2004): Cell biology of the
intracellular infection by Legionella pneumophila. In: Microbes Infect 6 (1), S. 129–139.
142. Nathan, C. (1997): Inducible nitric oxide synthase: what difference does it make? In: J Clin Invest
100 (10), S. 2417–2423. DOI: 10.1172/JCI119782.

LITERATURVERZEICHNIS 85
143. Nathan, C.; Shiloh, M. U. (2000): Reactive oxygen and nitrogen intermediates in the relationship
between mammalian hosts and microbial pathogens. In: Proc Natl Acad Sci U S A 97 (16), S.
8841–8848.
144. Nathan, C. F.; Hibbs, J B Jr (1991): Role of nitric oxide synthesis in macrophage antimicrobial
activity. In: Curr Opin Immunol 3 (1), S. 65–70.
145. Novick, Richard P. (2003): Autoinduction and signal transduction in the regulation of
staphylococcal virulence. In: Mol Microbiol 48 (6), S. 1429–1449.
146. Olivier, Aurelie C.; Lemaire, Sandrine; van Bambeke, Francoise; Tulkens, Paul M.; Oldfield, Eric
(2009): Role of rsbU and staphyloxanthin in phagocytosis and intracellular growth of
Staphylococcus aureus in human macrophages and endothelial cells. In: J INFECT DIS 200 (9),
S. 1367–1370. DOI: 10.1086/606012.
147. Otto, Michael (2012): Molecular basis of Staphylococcus epidermidis infections. In: Semin
Immunopathol 34 (2), S. 201–214. DOI: 10.1007/s00281-011-0296-2.
148. Otto, Michael (2014): Staphylococcus epidermidis pathogenesis. In: Methods Mol Biol 1106, S.
17–31. DOI: 10.1007/978-1-62703-736-5_2.
149. Palecanda, A.; Kobzik, L. (2001): Receptors for unopsonized particles: the role of alveolar
macrophage scavenger receptors. In: Curr Mol Med 1 (5), S. 589–595.
150. Palmqvist, Niklas; Foster, Timothy; Fitzgerald, J. Ross; Josefsson, Elisabet; Tarkowski, Andrzej
(2005): Fibronectin-binding proteins and fibrinogen-binding clumping factors play distinct roles
in staphylococcal arthritis and systemic inflammation. In: J INFECT DIS 191 (5), S. 791–798.
DOI: 10.1086/427663.
151. Pane-Farre, Jan; Jonas, Beate; Hardwick, Steven W.; Gronau, Katrin; Lewis, Richard J.; Hecker,
Michael; Engelmann, Susanne (2009): Role of RsbU in controlling SigB activity in
Staphylococcus aureus following alkaline stress. In: J Bacteriol 191 (8), S. 2561–2573. DOI:
10.1128/JB.01514-08.
152. Pareja, Maria Eugenia Mansilla; Colombo, Maria I. (2013): Autophagic clearance of bacterial
pathogens: molecular recognition of intracellular microorganisms. In: Front Cell Infect Microbiol
3, S. 54. DOI: 10.3389/fcimb.2013.00054.
153. Peschel, Andreas; Otto, Michael (2013): Phenol-soluble modulins and staphylococcal infection.
In: Nat Rev Microbiol 11 (10), S. 667–673. DOI: 10.1038/nrmicro3110.

LITERATURVERZEICHNIS 86
154. Plouin-Gaudon, I.; Clement, S.; Huggler, E.; Chaponnier, C.; Francois, P.; Lew, D. et al. (2006):
Intracellular residency is frequently associated with recurrent Staphylococcus aureus
rhinosinusitis. In: Rhinology 44 (4), S. 249–254.
155. Pluddemann, Annette; Mukhopadhyay, Subhankar; Sankala, Marko; Savino, Silvana; Pizza,
Mariagrazia; Rappuoli, Rino et al. (2009): SR-A, MARCO and TLRs differentially recognise
selected surface proteins from Neisseria meningitidis: an example of fine specificity in microbial
ligand recognition by innate immune receptors. In: J Innate Immun 1 (2), S. 153–163. DOI:
10.1159/000155227.
156. Richardson, A. R.; Libby, S. J.; Fang, F. C. (2008): A Nitric Oxide -Inducible Lactate
Dehydrogenase Enables Staphylococcus aureus to Resist Innate Immunity. In: Science 319
(5870), S. 1672–1676. DOI: 10.1126/science.1155207.
157. Ridley, Robert A.; Douglas, Ian; Whawell, Simon A. (2012): Differential adhesion and invasion
by Staphylococcus aureus of epithelial cells derived from different anatomical sites. In: J Med
Microbiol 61 (Pt 12), S. 1654–1661. DOI: 10.1099/jmm.0.049650-0.
158. Rigby, Kevin M.; DeLeo, Frank R. (2012): Neutrophils in innate host defense against
Staphylococcus aureus infections. In: Semin Immunopathol 34 (2), S. 237–259. DOI:
10.1007/s00281-011-0295-3.
159. Robertson, Charles M.; Perrone, Erin E.; McConnell, Kevin W.; Dunne, W. Michael; Boody,
Barrett; Brahmbhatt, Tejal et al. (2008): Neutrophil Depletion Causes a Fatal Defect in Murine
Pulmonary Staphylococcus aureus clearance. In: Journal of Surgical Research 150 (2), S. 278–
285. DOI: 10.1016/j.jss.2008.02.009.
160. Rogasch, Kathrin; Ruhmling, Vanessa; Pane-Farre, Jan; Hoper, Dirk; Weinberg, Christin; Fuchs,
Stephan et al. (2006): Influence of the two-component system SaeRS on global gene expression
in two different Staphylococcus aureus strains. In: J Bacteriol 188 (22), S. 7742–7758. DOI:
10.1128/JB.00555-06.
161. Rogers, Kathie L.; Fey, Paul D.; Rupp, Mark E. (2009): Coagulase -negative staphylococcal
infections. In: Infect Dis Clin North Am 23 (1), S. 73–98. DOI: 10.1016/j.idc.2008.10.001.
162. Rohde, Kyle; Yates, Robin M.; Purdy, Georgiana E.; Russell, David G. (2007): Mycobacterium
tuberculosis and the environment within the phagosome. In: Immunol Rev 219, S. 37–54. DOI:
10.1111/j.1600-065X.2007.00547.x.

LITERATURVERZEICHNIS 87
163. Sagy, Mayer; Al-Qaqaa, Yasir; Kim, Paul (2013): Definitions and pathophysiology of sepsis. In:
Curr Probl Pediatr Adolesc Health Care 43 (10), S. 260–263. DOI:
10.1016/j.cppeds.2013.10.001.
164. Saravia-Otten, P.; Muller, H. P.; Arvidson, S. (1997): Transcription of Staphylococcus aureus
fibronectin binding protein genes is negatively regulated by agr and an agr-independent
mechanism. In: J Bacteriol 179 (17), S. 5259–5263.
165. Sasaki, S.; Miura, T.; Nishikawa, S.; Yamada, K.; Hirasue, M.; Nakane, A. (1998): Protective role
of nitric oxide in Staphylococcus aureus infection in mice. In: Infect Immun 66 (3), S. 1017–1022.
166. Schnaith, Annabelle; Kashkar, Hamid; Leggio, Sonja A.; Addicks, Klaus; Kronke, Martin; Krut,
Oleg (2007): Staphylococcus aureus subvert autophagy for induction of caspase-independent
host cell death. In: J Biol Chem 282 (4), S. 2695–2706. DOI: 10.1074/jbc.M609784200.
167. Schoenfelder, Sonja M.K.; Lange, Claudia; Eckart, Martin; Hennig, Susanne; Kozytska, Svitlana;
Ziebuhr, Wilma (2010): Success through diversity – How Staphylococcus epidermidis
establishes as a nosocomial pathogen. In: International Journal of Medical Microbiology 300 (6),
S. 380–386. DOI: 10.1016/j.ijmm.2010.04.011.
168. Schroder, Andreas; Kland, Raphael; Peschel, Andreas; Eiff, Christof von; Aepfelbacher, Martin
(2006): Live cell imaging of phagosome maturation in Staphylococcus aureus infected human
endothelial cells: small colony variants are able to survive in lysosomes. In: Med Microbiol
Immunol 195 (4), S. 185–194. DOI: 10.1007/s00430-006-0015-0.
169. Schwartz, Jamie; Leidal, Kevin G.; Femling, Jon K.; Weiss, Jerrold P.; Nauseef, William M.
(2009): Neutrophil bleaching of GFP-expressing staphylococci: probing the intraphagosomal fate
of individual bacteria. In: J Immunol 183 (4), S. 2632–2641. DOI: 10.4049/jimmunol.0804110.
170. Seveau, Stephanie; Pizarro-Cerda, Javier; Cossart, Pascale (2007): Molecular mechanisms
exploited by Listeria monocytogenes during host cell invasion. In: Microbes Infect 9 (10), S.
1167–1175. DOI: 10.1016/j.micinf.2007.05.004.
171. Shibata, Takehiko; Nagata, Kisaburo; Kobayashi, Yoshiro (2011): Apoptotic neutrophils and
nitric oxide regulate cytokine production by IFN-γ-stimulated macrophages. In: Cytokine 53 (2),
S. 191–195. DOI: 10.1016/j.cyto.2010.10.003.
172. Shinji, H.; Sakurada, J.; Seki, K.; Murai, M.; Masuda, S. (1998): Different effects of fibronectin
on the phagocytosis of Staphylococcus aureus and coagulase-negative staphylococci by murine
peritoneal macrophages. In: Microbiol Immunol 42 (12), S. 851–861.

LITERATURVERZEICHNIS 88
173. Shinji, Hitomi; Kamada, Minori; Seki, Keiko; Tajima, Akiko; Iwase, Tadayuki; Masuda, Shogo
(2007): Expression and distribution of very late antigen-5 in mouse peritoneal macrophages
upon ingestion of fibronectin-bound Staphylococcus aureus. In: Microbiol Immunol 51 (1), S. 63–
71.
174. Shinji, Hitomi; Seki, Keiko; Tajima, Akiko; Uchida, Atsuko; Masuda, Shogo (2003): Fibronectin
bound to the surface of Staphylococcus aureus induces association of very late antigen 5 and
intracellular signaling factors with macrophage cytoskeleton. In: Infect Immun 71 (1), S. 140–
146.
175. Shinji, Hitomi; Yosizawa, Yukio; Tajima, Akiko; Iwase, Tadayuki; Sugimoto, Shinya; Seki, Keiko;
Mizunoe, Yoshimitsu (2011): Role of fibronectin-binding proteins A and B in in vitro cellular
infections and in vivo septic infections by Staphylococcus aureus. In: Infect Immun 79 (6), S.
2215–2223. DOI: 10.1128/IAI.00133-11.
176. Shompole, Sankale; Henon, Kim T.; Liou, Linda E.; Dziewanowska, Katarzyna; Bohach, Gregory
A.; Bayles, Kenneth W. (2003): Biphasic intracellular expression of Staphylococcus aureus
virulence factors and evidence for Agr-mediated diffusion sensing. In: Mol Microbiol 49 (4), S.
919–927.
177. Sinha, B.; Francois, P.; Que, Y. A.; Hussain, M.; Heilmann, C.; Moreillon, P. et al. (2000):
Heterologously expressed Staphylococcus aureus fibronectin-binding proteins are sufficient for
invasion of host cells. In: Infect Immun 68 (12), S. 6871–6878.
178. Sinha, B.; Francois, P. P.; Nusse, O.; Foti, M.; Hartford, O. M.; Vaudaux, P. et al. (1999):
Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to
integrin alpha5beta1. In: Cell Microbiol 1 (2), S. 101–117.
179. Sinha, Bhanu; Fraunholz, Martin (2010): Staphylococcus aureus host cell invasion and post-
invasion events. In: Int. J. Med. Microbiol. 300 (2-3), S. 170–175. DOI:
10.1016/j.ijmm.2009.08.019.
180. Sinha, Bhanu; Herrmann, Mathias (2005): Mechanism and consequences of invasion of
endothelial cells by Staphylococcus aureus. In: Thromb Haemost 94 (2), S. 266–277. DOI:
10.1267/THRO05020266.
181. Speziale, Pietro; Pietrocola, Giampiero; Rindi, Simonetta; Provenzano, Maria; Provenza, Giulio;
Di Poto, Antonella et al. (2009): Structural and functional role of Staphylococcus aureus surface
components recognizing adhesive matrix molecules of the host. In: Future Microbiol 4 (10), S.
1337–1352. DOI: 10.2217/fmb.09.102.

LITERATURVERZEICHNIS 89
182. Steinhuber, Andrea; Goerke, Christiane; Bayer, Manfred G.; Doring, Gerd; Wolz, Christiane
(2003): Molecular architecture of the regulatory Locus sae of Staphylococcus aureus and its
impact on expression of virulence factors. In: J Bacteriol 185 (21), S. 6278–6286.
183. Stuart, Lynda M.; Deng, Jiusheng; Silver, Jessica M.; Takahashi, Kazue; Tseng, Anita A.;
Hennessy, Elizabeth J. et al. (2005): Response to Staphylococcus aureus requires CD36-
mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. In: J Cell Biol 170
(3), S. 477–485. DOI: 10.1083/jcb.200501113.
184. Taylor, P. R.; Martinez-Pomares, L.; Stacey, M.; Lin, H-H; Brown, G. D.; Gordon, S. (2005):
Macrophage receptors and immune recognition. In: Annu Rev Immunol 23, S. 901–944. DOI:
10.1146/annurev.immunol.23.021704.115816.
185. Thomas, C. A.; Li, Y.; Kodama, T.; Suzuki, H.; Silverstein, S. C.; El Khoury, J. (2000): Protection
from lethal gram-positive infection by macrophage scavenger receptor-dependent phagocytosis.
In: J Exp Med 191 (1), S. 147–156.
186. Tuchscherr, Lorena; Heitmann, Vanessa; Hussain, Muzaffar; Viemann, Dorothee; Roth,
Johannes; Eiff, Christof von et al. (2010): Staphylococcus aureus small -colony variants are
adapted phenotypes for intracellular persistence. In: J INFECT DIS 202 (7), S. 1031–1040. DOI:
10.1086/656047.
187. Valour, Florent; Trouillet-Assant, Sophie; Rasigade, Jean-Philippe; Lustig, Sebastien; Chanard,
Emmanuel; Meugnier, Helene et al. (2013): Staphylococcus epidermidis in orthopedic device
infections: the role of bacterial internalization in human osteoblasts and biofilm formation. In:
PLoS One 8 (6), S. e67240. DOI: 10.1371/journal.pone.0067240.
188. van Belkum, Alex; Melles, Damian C.; Nouwen, Jan; van Leeuwen, Willem B; van Wamel,
Willem; Vos, Margreet C. et al. (2009): Co-evolutionary aspects of human colonisation and
infection by Staphylococcus aureus. In: Infect Genet Evol 9 (1), S. 32–47. DOI:
10.1016/j.meegid.2008.09.012.
189. Vaudaux, Pierre; Francois, Patrice; Bisognano, Carmelo; Kelley, William L.; Lew, Daniel P.;
Schrenzel, Jacques et al. (2002): Increased expression of clumping factor and fibronectin-
binding proteins by hemB mutants of Staphylococcus aureus expressing small colony variant
phenotypes. In: Infect Immun 70 (10), S. 5428–5437.
190. Vazquez-Torres, Andres; Stevanin, Tania; Jones-Carson, Jessica; Castor, Margaret; Read,
Robert C.; Fang, Ferric C. (2008): Analysis of nitric oxide -dependent antimicrobial actions in

LITERATURVERZEICHNIS 90
macrophages and mice. In: Methods Enzymol 437, S. 521–538. DOI: 10.1016/S0076-
6879(07)37026-2.
191. Vignais, P. V. (2002): The superoxide-generating NADPH oxidase: structural aspects and
activation mechanism. In: Cell Mol Life Sci 59 (9), S. 1428–1459.
192. Voyich, Jovanka M.; Braughton, Kevin R.; Sturdevant, Daniel E.; Whitney, Adeline R.; Said-
Salim, Battouli; Porcella, Stephen F. et al. (2005): Insights into mechanisms used by
Staphylococcus aureus to avoid destruction by human neutrophils. In: J Immunol 175 (6), S.
3907–3919.
193. Voyich, Jovanka M.; Vuong, Cuong; DeWald, Mark; Nygaard, Tyler K.; Kocianova, Stanislava;
Griffith, Shannon et al. (2009): The SaeR/S Gene Regulatory System Is Essential for Innate
Immune Evasion by Staphylococcus aureus. In: J INFECT DIS 199 (11), S. 1698–1706. DOI:
10.1086/598967.
194. Wang, Qing-Qing; Li, Hong; Oliver, Tim; Glogauer, Michael; Guo, Jian; He, You-Wen (2008):
Integrin beta 1 regulates phagosome maturation in macrophages through Rac expression. In: J
Immunol 180 (4), S. 2419–2428.
195. Wann, E. R.; Gurusiddappa, S.; Hook, M. (2000): The fibronectin-binding MSCRAMM FnbpA of
Staphylococcus aureus is a bifunctional protein that also binds to fibrinogen. In: J Biol Chem 275
(18), S. 13863–13871.
196. Watanabe, Ikuko; Ichiki, Manami; Shiratsuchi, Akiko; Nakanishi, Yoshinobu (2007): TLR2 -
mediated survival of Staphylococcus aureus in macrophages: a novel bacterial strategy against
host innate immunity. In: J Immunol 178 (8), S. 4917–4925.
197. Watkins, Robert L.; Pallister, Kyler B.; Voyich, Jovanka M.; Diep, Binh An (2011): The SaeR/S
Gene Regulatory System Induces a Pro-Inflammatory Cytokine Response during
Staphylococcus aureus Infection. In: PLoS ONE 6 (5), S. e19939. DOI:
10.1371/journal.pone.0019939.
198. Wertheim, Heiman F L; Melles, Damian C.; Vos, Margreet C.; van Leeuwen, Willem; van Belkum,
Alex; Verbrugh, Henri A.; Nouwen, Jan L. (2005): The role of nasal carriage in Staphylococcus
aureus infections. In: Lancet Infect Dis 5 (12), S. 751–762. DOI: 10.1016/S1473-3099(05)70295-
4.

LITERATURVERZEICHNIS 91
199. Wesson, C. A.; Liou, L. E.; Todd, K. M.; Bohach, G. A.; Trumble, W. R.; Bayles, K . W. (1998):
Staphylococcus aureus Agr and Sar global regulators influence internalization and induction of
apoptosis. In: Infect Immun 66 (11), S. 5238–5243.
200. Williams, Rachel J.; Henderson, Brian; Sharp, Lindsay J.; Nair, Sean P. (2002): Identification of
a fibronectin-binding protein from Staphylococcus epidermidis. In: Infect Immun 70 (12), S.
6805–6810.
201. Wolz, C.; Pohlmann-Dietze, P.; Steinhuber, A.; Chien, Y. T.; Manna, A.; van Wamel, W.; Cheung,
A. (2000): Agr-independent regulation of fibronectin-binding protein(s) by the regulatory locus
sar in Staphylococcus aureus. In: Mol Microbiol 36 (1), S. 230–243.
202. Xiong, Yan Q.; Willard, Julie; Yeaman, Michael R.; Cheung, Ambrose L.; Bayer, Arnold S. (2006):
Regulation of Staphylococcus aureus alpha-toxin gene (hla) expression by agr, sarA, and sae in
vitro and in experimental infective endocarditis. In: J INFECT DIS 194 (9), S. 1267–1275. DOI:
10.1086/508210.
203. Xu, Li; Luo, Zhao-Qing (2013): Cell biology of infection by Legionella pneumophila. In: Microbes
Infect 15 (2), S. 157–167. DOI: 10.1016/j.micinf.2012.11.001.
204. Yamamoto, Tomoko (2006): Bacterial strategies fo r escaping the bactericidal mechanisms by
macrophage. In: Yakugaku Zasshi 126 (12), S. 1235–1243.
205. Yarwood, Jeremy M.; Schlievert, Patrick M. (2003): Quorum sensing in Staphylococcus
infections. In: J Clin Invest 112 (11), S. 1620–1625. DOI: 10.1172/JCI20442.
206. Yimin; Kohanawa, Masashi; Zhao, Songji; Ozaki, Michitaka; Haga, Sanae; Nan, Guangxian et
al. (2013): Contribution of toll-like receptor 2 to the innate response against Staphylococcus
aureus infection in mice. In: PLoS One 8 (9), S. e74287. DOI: 10.1371/journal.pone.0074287.
207. Zecconi, Alfonso; Scali, Federico (2013): Staphylococcus aureus virulence factors in evasion
from innate immune defenses in human and animal diseases. In: Immunol Lett 150 (1-2), S. 12–
22. DOI: 10.1016/j.imlet.2013.01.004.
208. Ziebuhr, Wilma; Hennig, Susanne; Eckart, Martin; Kranzler, Hennes; Batzilla, Christoph;
Kozitskaya, Svetlana (2006): Nosocomial infections by Staphylococcus epidermidis: how a
commensal bacterium turns into a pathogen. In: Int J Antimicrob Agents 28 Suppl 1, S. S14-20.
DOI: 10.1016/j.ijantimicag.2006.05.012.

92
7 PUBLIKATION
Ergebnisse dieser Arbeit wurden veröffentlicht in:
Köhler J, Breitbach K, Renner C, Heitsch AK, Bast A, van Rooijen N, Vogelgesang S, Steinmetz I.
„NADPH-oxidase but not inducible nitric oxide synthase contributes to resistance in an acute
murine Staphylococcus aureus Newman pneumonia model.“
Microbes and Infection 13 (11), S. 914-922, 2011 May 27

93
8 DANKSAGUNG
Ich danke Ivo Steinmetz und Katrin Breitbach für die hervorragende und ausdauernde Betreuung, sowie
die Möglichkeit, dass ich mit ihnen habe wissenschaftlich arbeiten können.





























