Embed Size (px)
Citation preview

Mémoire pr
Laboratoire D’
Contribution à l’étude de la biologie du
Trachurus mediterraneus
gamétogenèse, période de ponte, indice de condition, fécondité et lecture des otolithes
Mademoiselle BENSAADA Fatima Zohra
Membres du jury :
Président : Mr Baba-
Examinateur: Mr. Taleb M. Z.
Examinateur : Mr Abi
Encadreur : Mr Bensahla Talet A.
Co- encadreur : Mlle Dalouche F.
moire présenté pour l’obtention du diplôme de
MAGISTER
Laboratoire D’aquaculture et Bioremediation – Aquabior
Intitulé :
Contribution à l’étude de la biologie du Chinchard
(Steindachner, 1868) pêché dans la baie d’Oran
gamétogenèse, période de ponte, indice de condition, fécondité et lecture des otolithes
Présenté par:
Mademoiselle BENSAADA Fatima Zohra
Soutenu le 08/05/2012
-Hamed. M. B Professeur Université d’Oran
Taleb M. Z. Maître de conférences
Mr Abi-Ayad S.-M. E.-A. Professeur
Mr Bensahla Talet A. Professeur
: Mlle Dalouche F. Maître-assistante A
Année Universitaire : 2011/2012
Aquabior
pêché dans la baie d’Oran : étude de la
gamétogenèse, période de ponte, indice de condition, fécondité et lecture des otolithes
Université d’Oran
Maître de conférences Université d’Oran
Université d’Oran
rofesseur Université d’Oran
Université d’Oran

Remerciements
Je souhaite tout d’abord remercier mon promoteur Mr Bensahla Talet .A et mon co-
promoteur Melle Dalouche .F, leur relectures attentives, leur conseils et leur correction m’ont
permis tout au long de ce travail d’écriture de faire évaluer fortement ma réflexion et mon
manuscrit.
Je remercie également les membres du jury président Mr Baba-Hamed d’avoir
accepté de rapporter sur mon manuscrit, ainsi que les examinateurs Mr Abi-Ayad S.-
M. E.-A et Mr Taléb. Z.
Je remercie particulièrement l’ensemble de l’équipe AQUABIOR avec lesquels ce fut
toujours un plaisir de travailler. Un grand merci à Gherram .M et Tahari .F Z pour les conseils
qui m’ont donnés, leur chaleur humaine et toute leur qualité de travail, qui m’ont permis
d’initier ce travail de mémoire.
Je remercie Nassima pour sa disposition et sa gentillesse, Mr Bensahla .L pour les
connaissances qu’il m’a apportées dans les statistiques (la pratique des logiciels), ainsi que
Guendouz .M, et tous mes collègues biologistes.
Pour terminer j’embrasse chaleureusement ma famille, en particulier mes parents pour leur
patience et leur inconditionnel soutien, ma sœur et mon frère (qui se connaissent).

Résumé
Résumé
Cette étude a été réalisée sur la biologie d'une espèce de carangidés de la baie d’Oran:
Trachurus mediterraneus (Steindachner, 1868). 1251 individus ont échantillonnés de juillet
2010 à juin 2011. La longueur moyenne totale a été estimée à 20,16cm pour les femelles et
20,42 cm pour les mâles. Les longueurs totales obtenues à partir des échantillons étudiés sont
comprise entre 13,2cm et 29,4cm et un intervalle plus large de poids compris entre 13,2g et
29,4g.
Nous avons examiné la sexe ratio, la taille et l'âge à la première maturité, la fécondité, la
période de reproduction, l'âge et croissance chez cette espèce. La sexe ratio globale chez les
femelles et les mâles est de 0,36 et 0,42 respectivement. Il varie d'un mois à l'autre. La taille à
la première maturité est de15 cm chez les mâles et les femelles soit 1 ans.
La reproduction a lieu pendant la période printemps-été avec une forte activité
reproductrice d’avril à juin. Une étude histologique des gonades et une analyse statistique des
variables biométriques (RGS, RHS, Kc) ont été aussi utilisées pour étudier la biologie du
chinchard, Trachurus mediterraneus. Cinq stades de maturité sexuelle peuvent être décrite
dans les deux sexes mâles et femelles. L'indice gonado-somatic, atteint sa valeur maximale
en mai-juin.
La fécondité varie entre 2063 et 18854 ovocytes. Les diamètres ovocytaires varient entre
38.4μm et704μm avec une moyenne de 262,19μm.
Les paramètres de croissance de Van Bertalanffy pour les deux sexes sont:
Femelles : L∞ =22cm, to = -4,57ans et K = 0,625an
Mâles : L∞ =24cm, to = -5,249 ans et K = 0,625an
Le taux d’allométrie b de l'équation conduit à la valeur 4,64, ce qui indique que la
croissance du poids est allométrique majorante.
Mots clés: Trachurus mediterraneus, Baie d’Oran, sex-ratio, reproduction, fécondité,croissance.

Abstract
Abstract
This study was achieved on the biology of a species of Carangidae fished in the Bay of
Oran: Trachurus mediterraneus (Steindachner, 1868). 1251 individuals were sampled from
July 2010 to june 2011.
The total average length was estimated at 20.16 cm for females and 20.42 cm for males;
the total length obtained from the samples studied are between 13.2 cm and 29.4 cm and a
broader range of weight from 1.7g to 174.1 g. We examined the sex ratio, size and age at first
maturity, fecundity, reproductive period, age and growth in this species.
The overall sex ratio for females and males is 0.36 and 0.42 respectively. It varies from a
month to another. The size at first maturity is 15 cm in males and females which corresponds
to one year of age. Spawning occurs during spring and summer with high reproductive
activity in April-June, a histological study of gonads and a statistical analysis of the biometric
variables (RGS, RHS, Kc) were also used to study the biology of horse mackerel, Trachurus
mediterraneus.
Six stages of sexual maturity can be described in both male and female sexes. Gonado-
somatic index, reaches its maximum value in May-June.
Fertility varies between 2063 and 18,854 eggs. The oocyte diameters ranging from 38.4μm
et704μm with an average of 262.19 μm.
Growth parameters of Von Bertalanffy for both sexes are:
Females: L ∞ = 22 cm, to = -4.57 years, K = 0.625 year
Males: L ∞ = 24 cm, to = -5.249 years, K = 0.625 year
The rate of allometry b of the equation leads to the value 4.64, indicating that growth is
allometric majorant weight.
Keywords: Trachurus mediterraneus, Oran Bay, sex ratio, reproduction, fertility, growth.

الملخص
الملخص
,Trachurus mediterraneus اجریت ھذه الدراسة على نوع من االسماك صید في حوض وھران السكوداالصفر
.2011ویونیو 2010فردمابین یولیو 1251اخذت عینات من (Steindachner, 1868)
مجموع االطوال المحصل علیھامن خالل الدراسة .سم للذكور20,42سم لالناث 20,16یقدر متوسط الطول االجمالي ب
الحجم ,النسبة الجنسیة درسنا. غ174,1غ و 1,7سم ومجموعة من الكتل تتراوح ما بین29,4سم و13,2یتراوح مابین
.ربسن و نمو ھذا الض,فترةالوضع ,والسن اثناء اول نضج جنسي
سم 15طول اول نضج یقدرب ,تتغیرمن شھرالخر,على ھذاالترتیب 0,42, 0,36النسبة الجنسیة الشاملة للذكورواالناث
, الدراسة النسیجیة لالعضاء التناسلیة. فترة الوضع تحدث ما بین فصلي الربیع و الصیف, لدىالجنسین مایوافق سنة واحدة
ومعمل الشرط استعملت ایضا , متوسط النضج الكبدي, متوسط النضج الجنسي, یةالتحلیل االحصائي للمتغیرات البیومتر
یبلغ متوسط النضج الجنسي اعلى , نستطیع وصف ستة مراحل نضج للذكور و لالناث. لدراسة بیولوجیا السكود االصفر
.قیمة لھ خالل شھري مایو ویولیو
میكرومتر اما القیمة 704میكرومتر و 38,4یتراوح قطرھا ما بین, بویضة 18854و 2063الخصوبة تتراوح مابین
.میكرومتر 262,19المتوسطة لالقطار فتقدر ب
: فون برتاالنفي معامالت النمو لدى الجنسین حسب معادلة
∞ L= 22 سنة , سم -4,57= to, سنة 0,625=K : االناث
5,249-سم، ∞ L = 24 سنة = to, 0,625 = K الذكور:
4,64 قیمة یؤدي إلى من المعادلة قیاس التنامي ب معدل الحد األعلى قیاس التنامي لھا زیادة الوزنمشیرا إلى أن
مفتاحیةالكلمات ال
Trachurus mediterraneus ,الخصوبة، النمو, ،التكاثرنسبة الجنس، وھران خلیج

Tables des matières
IntroductionIntroduction
Etude bibliographiqueI. Présentation de l’espèce
1. Présentation des carangidés.2. Espéces de Trachurus abondantes en méditerranée.
2.1. Chinchard commun Trachurus trachurus2.2. Chinchard bleu Trachurus picturatus2.3. Chinchard à queue jaune Trachurs mediterraneus
2.3.1. Caractères morphologiques2.3.2. Distribution géographique2.3.3. Migration et bathymétrie2.3.4. Systématique, synonymie2.3.5. Régime alimentaire du Trachurus mediterraneus
3. Zone de pêcheII. Etude de la reproduction
1. Introduction sur la reproduction2. Ovogenèse et folliculogenèse3. Fécondité4. Développement ovocytaire5. Spermatogenèse et spermiation
5.1. Spermatogenèse5.2. Spermiation
6. Sexe ratio7. Taille à la première maturité sexuelle8. La période de ponte : RGS, RHS
8.1 Rapport gonado-somatique8.2. Rapport hépato-somatique
9. Coefficient de condition KC
III. Matériel et méthodes1. Identification des espèces2. Mesure des paramètres biologiques3. Sexe et stades de maturité4. Fécondité
4.1. Méthode de comptage5. Etude histologique
5.1. Fixation5.2. Inclusion
5.3. Réalisation des coupes
5.4. Coloration
5.5. Déparaffinage
5.6. Réhydratation
5.7. Eclaircissement
01
030303030303040404050607070808080909101011111111111212131313131313141414141415151515

5.8. Montage
5.9. Lecture des lames
IV. Résultats et discussion
IV.1.Résultats
1.1Distribution des trois espèces1.2 Taille à la première reproduction1.3 Sex-ratio
1.3.1 Sex-ratio globale1.3.2 Evolution du sexe ratio en fonction des saisons
1.3.3 Evolution du taux de féminité et de masculinité et du sexe ratio en
fonction des saisons
1.3.4 Evolution de la sex-ratio en fonction des tailles
1.4 Répartition des tailles et calcul de la taille moyenne
1.5 Période de reproduction
1.5.1 Evolution mensuelle du RGS
1.5.2 Evolution mensuelle du RHS
1.5.3 Comparaison entre le RGS et le RHS
1.6 Evolution de l’indice de condition (Kc)1.7 Stades de maturité sexuelle
1.7.1 Maturation des gonades
1.7.2 Répartition des diamètres ovocytaires
1.8 Fécondité
1.9 Etude histologique1.9.1. Histologie des ovaires1.9.2. Histologie des testicules
IV.2 Discussion2.1. Distribution des trois espèces2.2. Taille à la première reproduction2.3. Répartition des tailles et calcul de la taille moyenne2.4. Sex-ratio2.5. Période de ponte2.6. Stades de maturité2.7. Fécondité2.8. Conclusion sur la reproduction
V. Croissance1. Introduction
2. Définition des otolithes3. Description de l’oreille interne4. Composition chimique des otolithes4.1. Sagitta
4.2. Astiriscus
4.3. Lapilli
4.4. Description des otolithes du chinchard à queue jaune
5. Composition chimique des otolithes
6. Intérêt des otolithes
VI. Matériel et méthodes
1. Croissance
16161717172022222323
23242525252627282829292930323239454545454646474849495050505151515151525252535353

1.1. Paramètres de croissance
1.2. Les relations longueurs-poids
1.3. Analyse statistique
VII. Résultats et Discussion
1. Résultats
1.1. Les longueurs
1.1.1 Distribution des fréquences de longueurs
1.1.2 Relation longueurs totales et longueurs à la fourche
1.2. Otolithométrie
1.2.1 Relation avec le poids des otolithes
1.3. Croissance de Trachurus mediterraneus
1.3.1 Paramètres de l’équation de Von Bertalanffy
1.4. Relations longueurs-poids
2. Discussion
3. Conclusion sur la croissance
VIII. Conclusion
Références bibliographiques
Annexe
5455565656565757575762626870707070707275767891

Liste des figures
Figure 01 : Répartition géographique (mondiale, méditerranéenne).
Figure 02 : Distribution mensuelle des proportions de Trachurus mediterraneus,
Trachurus trachurus, Trachurus picturatus.
Figure 03 : répartition des proportions de Trachurus mediterraneus, Trachurus.
trachurus, Trachurus picturatus échantillonnés.
Figure 04 : Les trois espèces de Trachurus échantillonnées, a) : Trachurus.
mediterraneus, b) : Trachurus picturatus, c) : Trachurus trachurus. d) : Scutelles.
Figure 05 : Détermination de la taille de première maturité sexuelle chez les femelles de
Trachurus mediterraneus.
Figure 06 : Détermination de la taille de première maturité sexuelle chez les mâles de
Trachurus mediterraneus.
Figure 07 : Evolution mensuelles du sex-ratio chez le Trachurus mediterraneus.
Figure 08 : Evolution mensuelle du taux de féminité et de masculinité chez
Trachurus mediterraneus.
Figure 09: Variation du taux de féminité et du taux de masculinité en fonction de la taille
chez le Trachurus mediterraneus.
Figure 10 : Courbe d’abondance des mâles et des femelles chez le Trachurus
Mediterraneus.
Figure 11: Evolution mensuelle du RGS moyen chez le Trachurus mediterraneus
femelles et mâles.
Figure 12 : Evolution mensuelle du RHS moyen chez les Trachurus mediterraneus
Mâles.
Figure 13 : Evolution mensuelle du RGS et RHS moyen chez les femelles de Trachurus
Mediterraneus.
Figure 14 : Evolution mensuelle du RGS et RHS moyen chez les mâles de Trachurus
Mediterraneus.
Figure 15: Evolution mensuelle de l’indice de condition des femelles et des mâles de
Trachurus mediterraneus.
Figure 16 : Evolution mensuelle des pourcentages des stades de maturité sexuelle
de Trachurus mediterraneus femelles observés à l’échelle macroscopique.
Figure 17 : Distributions mensuelles des diamètres ovocytaires (µm) Chez les femelles
05
18
18
19
21
22
23
24
24
25
26
26
27
27
28
29

de Trachurus mediterraneus.
Figure 18 : Relation fécondité totale Ŕ longueur à la fourche.
Figure 19 : Relation fécondité relative par gramme de gonade et la longueur à la fourche.
Figure 20 : Variation de la fécondité relative par 100g de poids corporel en fonction de la
longueur à la fourche.
Figure 21 : Relation poids gonade-poids total Chez le Trachurus mediterraneus.
Figure 22: Coupes histologiques des ovaires de Trachurus mediterraneus en début de
maturation coloré à l’hématoxyline.
Figure 23 : Coupe histologique des ovaires matures colorés au trichrome à froid.
Figure 24 : Coupes histologique des ovaires matures colorés au Trichrome à froid.
Figure 25: Coupes histologique des ovaires matures colorés au Trichrome de Masson
(poisson pêché le mois d’Avril). Les flèches représentent les nucléoles surnuméraires.
Figure 26 : Coupes histologique des ovocytes hyalins et atrétiques colorés au
Trichrome de masson (poisson pêché le mois d’Avril).
Figure 27 : Coupes histologiques de testicules prélevés sur un mâle pêché au mois de
juillet.
Figure 28: Coupes histologiques de testicules prélevés sur un mâle pêché au mois d’avril.
Figure 29 : Coupes histologiques de testicules prélevés sur un mâle pêché au mois
d’avril.
Figure 30 : Coupes histologiques colorées au Trichrome à chaud de testicules prélevés
d’un mâle pêché le mois d’avril.
Figure 31: Coupes histologiques colorées à l’hématoxyline de testicules prélevés d’un
mâle pêché le mois de septembre.
Figure 32 : Fréquences mensuelles des T. mediterraneus mâles en fonction des centres
de classe de taille.
Figure 33 : Fréquences mensuelles des T. mediterraneus femelles en fonction des centres
de classe de taille.
Figure 34: Fréquences mensuelles (novembre, avril) des T. mediterraneus indéterminés
en fonction des centres de classe de taille.
Figure 35 : Fréquences mensuelles des T. mediterraneus femelles et mâles en fonction
des centres de classe de taille.
Figure 36 : Otolithe d’un Trachurus mediterraneus mâle (classe 16cm, âge II) ans)
(Gx100).
30
31
31
31
32
34
35
36
37
38
40
41
42
43
44
57
59
60
61
62

Figure 37 : Variations des tailles moyennes de Trachurus mediterraneus en fonction
des âges lus par otolithométrie. 70

Liste des tableaux
Tableau 01 : Synonymie.
Tableau 02 : Effectif et pourcentages des différents stades de maturité des
femelles de Trachurus mediterraneus.
Tableau 03 : Effectif et pourcentages des différents stades de maturité des mâles
De Trachurus mediterraneus.
Tableau 04 : Sexe ratio global chez le Trachurus mediterraneus.
Tableau 05 : Récapitulatif des résultats obtenus sur la fécondité.
Tableau 06: La relation longueur totale et longueur à la fourche utilisée est de la forme.
Tableau 07 : a) Clé âge-longueur totale des femelles de Trachurus mediterraneus.
b) Clé âge-longueur totale des mâles de Trachurus mediterraneus.
c) Clé âge-longueur totale des indéterminés de Trachurus mediterraneus.
d) Clé âge-longueur totale des sexes confondus de Trachurus mediterraneus.
e) Récapitulatif des longueurs moyennes obtenues par lecture des otolithes.
chez Trachurus mediterraneus.
Tableau 08 : relation poids total-poids de l’otolithe.
Tableau 09 : relation longueur total-poids de l’otolithe.
Tableau 10: relation entre poids de l’otolithe et l’âge.
Tableau 11 : récapitulatif des paramètres obtenus de l’équation de Von Bertalanffy.
Tableau 12 : relation mensuelles taille-poids, PT= a LTb.
Tableau 13 : Récapitulatif des paramètres de la relation taille-poids, PT= a LTb.
Tableau 14 : Comparaison de paramètres de croissance de Trachurus mediterraneus.
Tableau 15 : Paramètres de la relation taille-poids (a, b) dans la présente étude et
les autres régions.
06
20
21
22
63
62
64
65
66
67
68
68
69
69
70
71
71
74
75

Liste des abréviations
cm: Centimètre.
K : Taux de croissance.
Kc: Indice de condition.
Lf: Longueur à la fourche.
LS : Longueur standard.
Lt: Longueur totale.
mm: millimètre.
Pf: Poids du foie.
Pg: Poids des gonades.
PT : Poids total de l’individu.
RGS: Rapport gonado-somatique.
RHS: Rapport hépato-somatique.
SG: spermatogonie.
SGA: Spermatogonie A.
SGB: Spermatogonie B.
SPC I: Spermatocyte I.
SPC II: Spermatocyte II.
SPC: spermatocyte.
SPD: spermatide.
SZ: spermatozoïde.
T : Trachurus.
T : tonne
Tm : Trachurus mediterraneus.
TM : taille moyenneTp: Trachurus picturatus.
Tt : Trachurus trachurus.
μm: micromètre.

Introduction
1
Introduction générale
Au niveau du port de pêche d’Oran, les petits pélagiques sont parmi les principales
ressources vivantes exploitées.
Le président de la fédération algérienne de la pêche, a affirmé que la production nationale
de poissons a enregistré une baisse alarmante estimée à 69,5% en 2010 par rapport aux années
précédentes. A ce titre, il a expliqué que la production est passée de 187000 tonnes de
poissons en 2009 à 130000 tonnes en 2010, (Bellout, 2010).
La situation devient de plus en plus préoccupante, car en l’espace d’une année, les secteurs
de la pêche et des ressources halieutiques ont été consolidés par plus d’une centaine de
bateaux de pêche, dont plus d’une vingtaine de sardiniers et autres moyens inhérents au
secteur,( Aït Ouakli ,2011).
Dans cette étude nous nous intéressons au genre Trachurus, précisément le chinchard à
queue jaune, classé dans la famille des carangidés (perciformes), représenté par 33 genres et
140 espèces (Froese et Pauly, 2001), à présent 15 espèces du genre Trachurus sont connues
dans le monde.
Dans le but de contribuer à la connaissance des fonctions biologiques du Trachurus
mediterraneus pêché dans la baie d’Oran, nous nous sommes proposé d’étudier la croissance
et la reproduction de cette espèce. Nous avons fait des coupes histologiques de gonades des
Trachurus mediterraneus afin de tenter de décrire le processus de reproduction et à
développer une gamme de maturité, l’observation microscopique aide à identifier les états de
maturation de cette espèce.
La présente étude contribue aussi à l’étude de l'âge faite par lecture des otolithes, car la
détermination du taux de croissance et l'âge constituent une enquête ichtyologique très
importante pour le poisson ; la croissance est l'un des principaux facteurs qui déterminent les
conditions des stocks (Prodanov et Mikhailov, 1983).
L'analyse statistique des variables biométriques a été utilisée pour étudier les paramètres
suivants : le rapport gonado-somatique (RGS), le rapport hépato-somatique (RHS), l’indice de
condition (Kc), le calcul des paramètres de croissance de Von Bertalanffy et la relation taille
poids.

Introduction
2
Les résultats obtenus sont consignés dans le présent mémoire qui comporte 3 parties
principales. La première partie est consacrée aux généralités sur l’espèce, la deuxième partie
porte sur le cadre de l’étude et la méthodologie et la troisième partie est consacrée aux
résultats obtenus et la discussion de ces derniers.
Ce travail s’intègre dans l’étude de l’ichtyofaune de la baie d’Oran, zone d’une très grande
richesse où plus de 50 espèces de Téléostéens et de Sélaciens sont pêchées et aire de
reproduction de nombreux poissons côtiers (Dalouche, 1980).

Reproduction Etude bibliographique
3
I Présentation de l’espèce
1. Présentation de la famille des carangidés
Les carangidés sont des poissons perciformes à l’aspect extérieur assez variable. La forme
de leur corps varie modérément allongé à haut et fortement compressé. La forme de la tête
varie aussi de façon considérable de longue et basse à courte et haute. Les yeux sont
habituellement protégés par une paupière « adipeuse » transparente immobile, mais non
grasse. Le museau peut être pointu ou arrondi. Les dents sont rangées de petites villiformes, à
grande coniques et sont situées différemment sur le prémaxillaire, le dentaire, le vomer, les
palatins, la langue ou le pharynx (Gunn, 1990). Les membres de cette famille sont aussi
caractérisés par une nageoire anale avec deux épines antérieures séparées du reste de la
nageoire, mais qui s’enfoncent souvent avec l’âge, et la nageoire caudale est profondément
déviée. La nageoire dorsale est généralement divisée en une portion antérieure avec 4 à 8
épines et de 17 à 44 rayons mous. Chez de nombreux carangidés les derniers rayons de la
nageoire dorsale et anale sont détachés et forment 1 à 9 petites pinnules. La nageoire pectorale
est souvent longue et falciforme.
Les carangidés sont principalement marins, mais peuvent aussi se rencontrer en eaux
saumâtres. Certaines espèces pénètrent, de manière opportuniste, en eaux douces ou dans les
lacs. Ils se reproduisent surtout en été, pondent des œufs pélagiques et sont des carnivores
rapides (Smith-Vaniz, 1986). Certains d’entre eux sont d’importants poissons de
consommation.
Dans cette famille il y’a 32 genres connus, comprenant environ 140 espèces selon (Smith-
Vaniz, 2007).
2. Espèces de Trachurus abondantes en méditerranée
2.1. Chinchard commun Trachurus trachurus (LINNE, 1758)
Les chinchards présentent tout le long de leur ligne latérale des scutelles, c’est-à-dire des
écailles hypertrophiées. Chez le chinchard commun, ces scutelles sont presque aussi
développées de part et d’autre de la courbe que dessine la ligne latérale vers la moitié du
corps, la ligne latérale accessoire dépasse la moitié de la seconde dorsale (Smith-Vaniz 1986 ;
Quéro et al.,2003).
2.2. Chinchard bleu Trachurus picturatus (BOWDICH, 1825)
Les chinchards se ressemblent fort entre eux. Les différentes espèces sont souvent l’objet
de confusions. Le chinchard bleu a comme particularité d’avoir le rayon postérieur de ses
nageoires dorsale et anale séparé du précédent par une distance moitié plus grande que celle
des autres rayons entre eux (Guichet, 1966).

Reproduction Etude bibliographique
4
2.3. Chinchard à queue jaune : Trachurus mediterraneus (STEINDACHNER, 1868)
2.3.1 Caractères morphologiques
Corps allongé et légèrement comprimé, grands yeux avec une paupière adipeuse bien
développée; mâchoire supérieure modérément large et s’étendant jusque sous le bord
antérieur de l’œil; dents petites en une seule rangée aux deux mâchoires; branchiospines
(rudiments compris) 13-17 supérieures, 36-41 inférieures sur le premier arc branchial. Deux
nageoires dorsales, la première à 8 épines, la seconde à une épine et 29-35 rayons mous; anale
à 2 épines séparées du reste de la nageoire, suivies d’une épine et 26-39 rayons mous; rayon
mou terminal de la dorsale et de l’anale relié par une membrane au reste de la nageoire mais
séparé de l’avant dernier rayon par une distance au plus égale à 1,5 fois celle qui sépare les
précédents; pectorales à peu près égales à la longueur de la tête. Ecailles modérément petites
et cycloïdes (lisses au toucher) sur tout le corps sauf sur une petite surface située en arrière
des pectorales; écailles de la partie antérieure de la ligne latérale grandes et semblables aux
scutelles de la partie postérieure, leur hauteur maximum atteint 3,3-4,3% de la longueur
standard; écailles et scutelles de la ligne latérale 75-89 dont 35-44 scutelles sur la partie
rectiligne. Ligne latérale accessoire se terminant entre la huitième épine et le troisième rayon
mou de la dorsale. Pas de marques distinctives pour la coloration sauf une petite tache noire
sur le bord de l’opercule près de son angle supérieur. Partie supérieure du corps et de la tête
foncée à noire ou grise à vert bleuâtre, les deux tiers inférieurs du corps et de la tête
généralement plus pâles, blanchâtres à argentés (FAO, 1987).
Cette espèce se différencie de la précédente par les scutelles antérieures à la courbe de la
ligne latérale nettement moins développées que les postérieures et par la ligne latérale
accessoire ne dépassant pas le début de la seconde dorsale (Quéro et al, 2003).
2.3.2 Distribution géographique
Le chinchard à queue jaune est une espèce pélagique, marine et saumâtre, fréquente dans
les eaux subtropicales de l’atlantique de l’est, en méditerranée et en mer noire, vivant à des
profondeurs de 0 à 500m. En mer Noire mer de Marmara et mer d’Azov, a été cité comme
une sous espèce Trachurus mediterraneus ponticus. (Aleev, 1956) d’origine méditerranéenne
(Banarescu, 1964) mais n’est plus reconnue comme valide. Le golfe de Gascogne est la limite
nord de son aire de répartition (Smith-Vaniz, 1986). Elle y est plus est abondante au sud des
pertuis charentais que dans la moitié du golfe (Quéro et al., 1989).

Reproduction Etude bibliographique
5
Figure 1 : Répartition géographique (mondiale, méditerranéenne), (Cardenas et al ., 2004).
2.3.3 Migration et bathymétrie
Le chinchard vit prés du fond, mais aussi dans toute la colonne d’eau. Au printemps, il
migre du sud de la mer noire vers le nord pour se nourrir et se reproduire, et il redescend au
sud en automne. Le chinchard de la cote roumaine de la mer noire passe l’hiver en mer de
Marmara (Maximov et al., 2002).
Le temps minimum de doublement de ses populations est de 1,4-4,4 ans. C’est une espèce
commerciale, importante pour les pêcheurs (Berg et al., 1949), qui a été surexploitée en mer
Noire. Les captures ont drastiquement diminué sur la côte roumaine en peu de temps ; 2660
tonnes en 1988, 165 tonnes en 1990 et de 0-3 tonnes depuis 1997.

Reproduction Etude bibliographique
6
2.3.4 Systématique, synonymie
Phylum : Chordés
Embranchement : Vertébrés
Sous embranchement : Gnathostomes
Super classe : Poissons
Classe : Ostéichtyens
Sous classe : Actinoptérygiens
Super ordre : Téléostéens
Ordre : Perciformes
Sous ordre : Percoïdes
Famille : Carangidés
Genre : Trachurus
Espèce : mediterraneus
Tableau 1: Synonymie
Trachurus mediterraneus (STEINDACHNER, 1868).
Nom commun langue pays
Chinchard à queue jaune Français France
Chourou Arabe Tunisie
Jurel mediterraneo Espagnol Espagne
Sari Turque Turquie
Sugrella Italien Italie
Troûboûles Arabe Liban

Reproduction Etude bibliographique
7
Autres synonymes
Khourir Arabe Algérie
Sekoud asfar Arabe
2.3.5. Régime alimentaire du Trachurus mediterraneus
Le chinchard est un poisson prédateur qui se nourrit de petits poissons et de crustacés
(zooplancton) (Smith-Vaniz, 1986, Bensalem, 1988, Santic et al., 2003). Dans le nord-ouest
de la mer Noire, il consomme principalement des poissons (60-91% sprat, anchois, gobie,
mulet), secondairement des crustacés (7-33% crevettes, copépodes, mysidacés) et
accidentellement des polychètes, diatomées, macrophytes (Bauchot, 1987).
3. Zone de pêche
Les chinchards utilisés dans notre étude ont été péchés dans la baie d’Oran, situé sur la
côte ouest algérienne entre le golfe d’Arzew et la baie des andalouses (Leclaire, 1972), est
compris entre le cap de l’aiguille à l’est et le cap Falcon à l’ouest. La baie d’Oran est baignée
par les eaux d’origine atlantique ; la circulation apparait turbulente le long du continent
africain (Remili et al., 2009).

Reproduction Etude bibliographique
8
II. Reproduction
1. Introduction sur la reproduction
La connaissance de la biologie de la reproduction des poissons est essentielle pour une
bonne gestion des ressources halieutiques. Certains paramètres tels que la taille de
première maturité, la fécondité, le diamètre des œufs et la variation des indices gonado-
somatique et hépato-somatique permettent de mieux caractériser la reproduction ; le stade
de maturation sexuelle et la stratégie de ponte (Heins et al., 2004).
2. Ovogenèse et folliculogenèse
L’ovogenèse est la transformation de l’ovogonie en ovocytes (Selman et Wallace, 1989).
Ce processus regroupe toutes les transformations subies par la cellule germinale primordiale
pour devenir un ovocyte prêt à être fécondé, avec son vitellus, son enveloppe primaire ou
Zona radiata et ses granules corticaux (Mellinger, 2002).
Les ovogonies, issues des cellules germinales primordiales ou gonocytes, pendant
l’embryogenèse, prolifèrent par mitoses (Nagahama, 1983 ; Higashino et al., 2002) ; Patino
et Sullivan, 2002 ; Thiry et Poncin, 2005). Chez les poissons osseux et les amphibiens, à
l’inverse de tous les autres vertébrés, elles continuent à se diviser dans l’ovaire des adultes
(Tyler et Sumpter, 1996; Wallace et Selman, 1990; Jalabert, 2005). Les ovogonies se divisent
par mitoses, certaines maintiennent la population dans l’épithélium germinal, d’autres entrent
en méiose pour devenir des ovocytes qui migrent dans la lumière de l’ovaire. Le processus de
folliculogenèse commence à l’initiation (Grier, 2000 ; Grier et Lo Nostro, 2000).
Les ovocytes primaires, recrutés parmi les ovogonies entrent en méiose (Ravaglia et
Maggese, 2003) mais restent bloqués en prophase de la division réductionnelle, au stade
diplotène (Nagahama, 1983 ; Patino et Sullivan, 2002). Ils subissent alors une phase
d’accroissement cytoplasmique et une différenciation sans division (Wallace et Selman,
1990).
Pendant cette phase, l’ovocyte accumule des réserves nutritives et s’entoure d’une
enveloppe folliculaire composées de deux assises de cellules, les cellules folliculaires formant
la granulosa et les cellules théquales formant la thèque, cette enveloppe cellulaire est séparée
de l’ovocyte d’une couche acellulaire : la zona radiata. L’ovocyte et ses enveloppes forment
un follicule ovarien.

Reproduction Etude bibliographique
9
A la fin de cette longue période d’accroissement, un signal hormonal provoque la reprise
de la méiose, le noyau se brise et la moitié du matériel chromosomique est perdu par
l’expulsion du premier globule polaire. L’ovocyte secondaire ainsi formé est bloqué en
métaphase de la division équationnelle (Wallace et Selman, 1990). La maturation ovocytaire
finale, processus hormonodépendant, permettra l’ovulation, rendra l’ovocyte fertilisable
(Wallace et al., 1993) puis le développement embryonnaire possible (Saat, 1993). La fin de la
deuxième division de la méiose et l’expulsion du deuxième globule polaire se produisent
après la fécondation (Patino et Sullivan, 2002).
3. Fécondité
Plusieurs définitions de la fécondité ont été proposées par plusieurs auteurs ; (Bagenal,
1978) distingue :
Fécondité absolue : qui correspond au nombre d’œufs mûrs prêts à être pondus. Fécondité de
la population globale qui tient compte de la fécondité par classe de taille, de la sex-ratio par
taille et de la taille à la première maturité sexuelle ; et aussi le nombre d’ovules en fonction de
la taille de l’individu (Lt en cm) (Oosthuizen et Daan, 1974).
Fécondité relative : définie comme le nombre d’œufs par gramme de poissons (Nikolsky,
1963).
Fécondité relative moyenne : Correspond d’après (Oosthuizen et Daan, 1974), au nombre
d’ovules par gramme de poids corporel Pen (g).
4. Développement ovocytaire
Ce processus est découpé en six stades en fonction de la croissance ovocytaire. Le premier
stade est l’ovogenèse qui, pour ces auteurs, est la transformation de l’ovogonie en ovocyte
primaire. Le second est celui de la croissance primaire de l’ovocyte, Il est caractérisé par une
synthèse intense d’ARN, une augmentation importante de la taille de l’ovocyte, l’apparition
dans le cytoplasme du noyau de Balbiani et la formation de la zona radiata. C’est à la fin de
cette deuxième période que l’ovocyte quitte les nids où il côtoyait les ovogonies.
Le stade des alvéoles corticales se caractérise par l’apparition de ces inclusions
cytoplasmiques dans l’ovocyte. Ces alvéoles corticales finissent par occuper entièrement
l’ovoplasme, leur contenu a une origine endogène.
Le stade suivant est celui de l’apparition des inclusions lipidiques. Il est suivi par le stade
de vitellogenèse qui, d’après ces auteurs, représente la période de grossissement de l’ovocyte
où des protéines extraovariennes en particulier la vitellogénine, sont séquestrées, transformées
en protéines vitellines et stockées dans l’ovocyte (Rinchard et al., 1998) . Le dernier stade est

Reproduction Etude bibliographique
10
celui de la maturation pendant, lequel la vésicule germinale se rompt, et chez certains
téléostéens, l’œuf s’hydrate ce qui peut considérablement augmenter sa taille (Tyler et
Sumpter, 1996).
5. Spermatogenèse et spermiation
5.1. Spermatogenèse
La spermatogenèse est la succession des divisions cellulaires et des transformations, au
sein du testicule d’une cellule germinale peu différenciée, la spermatogonie en une cellule
germinale fonctionnelle, le spermatozoïde (Billard, 1979) ; (Barnabe, 1991). Les cellules
germinales évoluent toujours au contact des cellules somatiques de soutien appelées cellules
de Sertoli (Billard et al, 1972). Au début du cycle seules les spermatogonies A (SGA) sont
présentes, chaque (SGA) est collée à une ou plusieurs cellules de (Sertoli Grier et al., 1980).
Les multiplications des spermatogonies par mitose, la méiose puis la spermatogenèse se
déroulent à l’intérieur même d’une enveloppe formée par les extensions des cellules de
Sertoli, l’ensemble constitue un spermatocyste ou cyste. Les cellules d’un même cyste sont au
même stade de développement (Turner, 1919); (Billard et al., 1972) ; (Grier et al., 1980);
(Pudney , 1995) ; (Grier et Tylor, 1998) ; (Gusmao et al, 1999).
(Mattei et al., 1993) qualifient la spermatogenèse "cystique " lorsqu’elle se déroule
entièrement à l’intérieur des spermatocystes, et "semi cystique" lorsque ceux ci s’ouvrent
avant la formation des spermatozoïdes. Dans ce cas des cellules germinales à différents stades
de maturation peuvent être présentes dans la lumière des lobules où se terminent la
différenciation (Mattei et al., 1993) ; (Manni et Rasotto, 1997) ; (Yoneda et al., 2001). Les
SGA donnent par mitose les SGB qui restent groupées par deux puis quatre, ce qui initie la
formation d’un cyste. La division des SGB produit des spermatocytes de premier ordre
(SPCI). Les SPCI après une première division de méiose vont se transformer en spermatocyte
de second ordre SPCII (Pudney, 1995 ; Dziewulska et Domagala, 2003).
Les SPCII deviennent des spermatides SPD après la deuxième division de la méiose
(Nagahama, 1986) ; (Dziewulska et Domagala, 2003). Même si les spermatides ont un jeu de
chromosomes haploïdes ce ne sont pas des gamètes mâles. Elles doivent entrer dans une phase
de transformations biochimiques et morphologiques, nommée spermiogenèse, conduisant à
une cellule germinale hautement différenciée, le spermatozoïde (Nagahama, 1986).

Reproduction Etude bibliographique
11
5.2. Spermiation
A la fin de la spermiogenèse la paroi sertolienne des cystes devenue de plus en plus mince
s’ouvre et les spermatozoïdes sont libérés dans la lumière du lobule, c’est la spermiation
(Pudney, 1995) ; (Schulz et al., 2005). Ils se concentrent dans la lumière des lobules
séminifères d’où ils gagnent le système évacuateur. La spermiation correspond à l’émission
du sperme au niveau de l’orifice urogénital après pression abdominale. Elle est généralement
accompagnée d’une hydratation de gonades et du sperme. Lors de l’émission du sperme les
spermatozoïdes sont libres dans le plasma séminal chez les espèces à fécondation externe
(Legendre et Jalabert, 1988). La période de spermiation peut se poursuivre plusieurs mois
mais la qualité du sperme diminue fortement dans le temps du fait des phénomènes de
vieillissement des spermatozoïdes (Billard, 1979).
6. Sexe ratio
Les proportions numériques des sexes sont exprimées en pourcentage respectif des
mâles et des femelles et ont été étudiées en fonction des mois. Nous avons calculé le taux de
masculinité (% mâles = nombre des mâles x 100/ nombre total des mâles et des femelles) et
le taux de féminité (% femelles = nombre des femelles x 100/ nombre total des mâles et
des femelles).
7. Taille à la première maturité sexuelle
La taille de la première maturité sexuelle est la longueur pour laquelle 50% des individus
sont mûrs (Fontana, 1969). Pour l’établissement de la courbe du pourcentage des femelles ou
des mâles mûrs par classe de taille, nous avons compté durant la période de ponte le nombre
de femelles ou de mâles ayant atteint ou dépassé le stade III (début de la vitellogenèse) par
rapport au nombre total des femelles ou des mâles par classe de taille.
8. Période de ponte : RGS, RHS
8.1. Rapport gonado-somatique (RGS)
La détermination de la période de ponte a été réalisée en calculant le rapport gonado-
somatique RGS (Bougis, 1952), basé sur la variation de la masse des gonades au cours du
cycle sexuel.
RGS= Poids de la gonade x 100/poids somatique du poisson

Reproduction Etude bibliographique
12
8.2. Rapport hépato-somatique (RHS)
Les variations pondérales hépatiques, influencées par les conditions génitales, permettent
de définir les périodes de ponte chez les téléostéens ; en effet, l’élaboration des produits
sexuels est en étroite relation avec les phénomènes physiologiques qui ont lieu au niveau du
foie (Thiam, 1980). Le RHS est défini par la relation :
RHS= Poids du foie x 100/ poids somatique du poisson
9. Coefficient de condition Kc
Le coefficient de condition de (Beckman, 1948) permet de comparer les suites d’une même
espèce. La relation allométrique longueur-poids variant peu au cours de la croissance, ce
coefficient permet de suivre les variations physiologiques d’un poisson par rapport à sa valeur
spécifique normale, pour la détermination du facteur de condition Kc, qui correspond au
rapport de la masse réelle observée Mo d’un poisson par la masse théorique Mt (Mo/Mt),
plusieurs expressions sont habituellement employées (Bauchot et Bauchot, 1978) :
Kc= 100 * Pt/Lt3 (M (g) et Lt (mm))

Reproduction Matériel et méthodes
13
III. Matériel et méthodes
L’échantillonnage aléatoire du chinchard provenant des captures des sardiniers et/ou des
chalutiers, effectué au niveau de la baie d’Oran a été réalisé une fois par mois, de juillet 2010
à juin 2011, soit 12 mois, dans le but d’avoir une large gamme de taille de spécimens.
1. Identification des espèces
Regroupées par espèces, les poissons furent identifiés selon la clé de détermination
(FAO, 1987).
2. Mesure des paramètres biologiques
Les individus capturés ont été transférés au laboratoire de biologie marine pour la
dissection. Cet échantillonnage a couvert presque l’ensemble des gammes de taille de
chinchards allant de 130mm à 290mm ; pour chaque poisson, la longueur totale et la longueur
à la fourche ont été mesurées au mm prés, la masse totale a été mesurée à l’aide d’une balance
ayant une précision de 0,1g et de portée de 7000g
Les poissons sont disséqués, les masses des gonades et du foie ont été déterminées à l’aide
d’une balance ayant une précision de 0,1mg et de 82g de portée.
3. Sexe et stades de maturité
Après ouverture de la cavité générale (en partant de l’anus jusqu’au niveau des nageoires
pectorales), le sexe est déterminé par l’examen macroscopique des gonades. La détermination
des stades de maturité sexuelle (Tableau1, Annexe2) a été établie selon l’échelle proposée par
(CNRDPA, 2009).
4. Fécondité
4.1. Méthode de comptage
Les gonades femelles en maturité avancée (stade : IV, V) sont pesées, placées dans des
piluliers contenant le liquide de Gilson sont ensuite agitées quotidiennement pendant 15 à 30
jours ; après cette période d’agitation, le stroma ovarien se dissout et les ovocytes se détachent
facilement les uns des autres.
Pour éviter les comptages longs et fastidieux, nous avons eu recours à un sous
échantillonnage en diluant le contenu de chaque pilulier dans 250 ml d’eau distillée, ensuite
nous prélèvons à l’aide d’une pipette deux ml de la solution, ceux–ci sont placés dans une
cuve de Dollfus à fond quadrillé et examinons au microscope stéréoscopique en lumière
réfléchie. Nous mesurons à l’aide d’un micromètre oculaire (0-12μm) les diamètres des œufs
pour obtenir la distribution de fréquence.

Reproduction Matériel et méthodes
14
La fécondité est ensuite calculée selon la formule :
f = n × Pg / Pech
5. Etude histologique
5.1. Fixation
Elle a pour but de conserver au mieux les constituants cellulaires, et de prévenir la
décomposition post-mortem. Etape essentielle elle doit se faire, idéalement, juste après la mort
de l’animal par immersion de l’organe dans un grand volume de fixateur. La fixation peut être
réalisée à l’aide d’agents chimiques pris isolément (éthanol, formol, acide picrique…) ou
mélangés en proportions adéquates comme dans le liquide de Bouin utilisé dans cette étude
(annexe 4).
5.2. Inclusion
Elle donne aux pièces (gonades) la consistance nécessaire à leur débitage en coupes sans
déformer l’architecture cellulaire. La substance utilisée dans le cas présent est la paraffine. Vu
le caractère hydrophobe de celle-ci l’ensemble du processus d’inclusion comporte tout
d’abord la déshydratation des pièces fixées à l’aide de l’acétone (4 bains, ½ heure pour
chacun), et leur imprégnation au moyen d’un solvant de la paraffine (butanol ou hydrocarbure
benzénique, tel que : le xylène et le toluène) (deux bains successifs, 1heure pour chacun),
cette étape intermédiaire est garante d’une bonne pénétration du tissu par la paraffine, non
miscible aux cétones.
Le prélèvement est ensuite placé dans deux bains de paraffine fondue (étuve à 58-60°C, 1
heure pour chaque bain) qui infiltre tout l’échantillon et remplace totalement le solvant. Le
coulage du bloc a lieu à température ambiante entre deux barres de Leuckart juxtaposées pour
obtenir, après refroidissement, un bloc de paraffine solidifiée, de consistance homogène, à
l’intérieur duquel est incluse la pièce à étudier. La conservation des blocs est excellente
(plusieurs dizaines d’années) et se fait à la température du laboratoire.
5.3. Réalisation des coupes
Le bloc de paraffine est débité en coupe mince (5-8 μm) à l’aide d’un microtome à main.
Les coupes sont prélevées puis étalées sur une lame en verre (côté brillant) placée sur une
plaque chauffante (60°C, 4min), dés le lendemain, les préparations peuvent être colorées ou
rangées (plusieurs mois dans une boite opaque).

Reproduction Matériel et méthodes
15
5.4. Coloration
L’observation microscopique de gonades femelles et males est basée sur la coloration, le
but est d’accentuer les contrastes afin de distinguer et reconnaitre les différents constituants
de la préparation (coupes transversales des gonades).
5.5. Déparaffinage
Avant toute coloration, de par la nature aqueuse des colorants, les coupes doivent être
déparaffinées (par du toluène ou équivalent).
5.6. Réhydratation :
Par des alcools (méthanol, éthanol) de degrés décroissants (100°, 95°, 70°), suivis d’un
rinçage à l’eau courante. Les temps de coloration peuvent varier en fonction de l’épaisseur de
la coupe.
Les colorants utilisés sont : Hématoxyline- éosine : c’est la technique la plus couramment
utilisée en histologie animale, le colorant basique hématoxyline, colore les structure acides en
bleu violacé, les noyaux, les ribosomes, le réticulum endoplasmique rugueux ont une forte
affinité pour ce colorant vu leur richesse en ADN et en ARN, en revanche l’éosine est un
colorant acide qui colore les structures basiques en rouge et en rose, la plupart des protéines
cytoplasmiques sont basiques, le cytoplasme des cellules apparait donc habituellement coloré
en rose ou en rouge, en général lorsque la coloration avec l’HE est appliquée à des cellules
animales, les noyaux sont colorés en bleu violet et les cytoplasmes en rouge rose.
Les trichromes sont des colorations topographiques impliquant l’emploi de trois colorants,
(l’hémalun, phloxine, vert lumière pour le trichrome à froid, et l’hématoxyline de Régaud à la
place de l’hémalun dans le trichrome à chaud), qui ont l’avantage de donner trois teintes
particulières aux diverses structures permettant ainsi de différencier aisément, par exemple,
cytoplasme et noyaux, tissus musculaires et conjonctifs…etc. Après coloration les
préparations sont déshydratées dans des bains d’alcools croissants (70°, 95°, 100°)
5.7. Éclaircissement
Les préparations sont imprégnées et éclaircies par un hydrocarbure bénzénique (xylène,
toluène) ou équivalent (2-3 bains, 3 à 5min) miscible à une résine de synthèse (de type
Eukitt).

Reproduction Matériel et méthodes
16
5.8. Montage
Les coupes colorées sont recouvertes d’une lamelle en verre collée à l’Eukitt, qui permettra
une conservation permanente, vu son indice de réfraction très élevé (n= 1,74) de préférence à
l’abri de la lumière. Le montage des préparations a un triple objectif, à savoir la protection
mécanique, une longue période de conservation et l’obtention d’un bon degré de transparence
essentiel du point de vue de l’observation microscopique.
5.9. Lecture des lames
La lecture des lames au microscope optique se fait aux faibles grossissements (10) pour
avoir une bonne vue d'ensemble des tissus. Nous avons pris des photos à l’aide d’une caméra
(Image Driving soft ware MDCE-5C(E) G108005809).

Reproduction Résultats
17
IV.1. Résultats
1.1 Distribution des trois espèces
A partir d’un échantillonnage mensuel aléatoire nous avons obtenu les résultats suivants :
Au cours de notre sélection nous notons la présence d’une espèce, deux ou les trois en
même temps.
La figure ci-dessus représente les différents pourcentages des trois espèces obtenus chaque
mois:
Au mois de juillet nous soulignons la présence de Trachurus mediterraneus et Trachurus
trachurus, par contre le Trachurus picturatus n’apparait pas.
Au mois de septembre l’échantillonnage a été fait deux fois à cause de l’absence du
chinchard au mois d’aout vu la limitation de sa pêche (période de reproduction).
Le Trachurus picturatus n’apparait qu’au mois de septembre avec un pourcentage faible
(2,33%).
Au mois d’octobre et novembre nous ne retrouvons que le Trachurus mediterraneus, en
revanche les deux autres espèces sont absentes.
Au mois de décembre, janvier et février le Trachurus trachurus prédomine les deux autres
espèces alors qu’au mois de mars nous observons uniquement sa présence.
Au cours de la période printanière le Trachurus mediterraneus présente des pourcentages
plus élevés que les deux autres espèces respectivement au mois d’avril et mais (55% et 57%).
Au mois de juin le Trachurus trachurus est de 69% alors que celui de Trachurus
mediterraneus est seulement de 10%.
Pour conclure, nous remarquons la présence des trois espèces de chinchard (Trachurus
trachurus, Trachurus mediterraneus, Trachurus picturatus) dont les pourcentages présentent
des variations mensuelles.
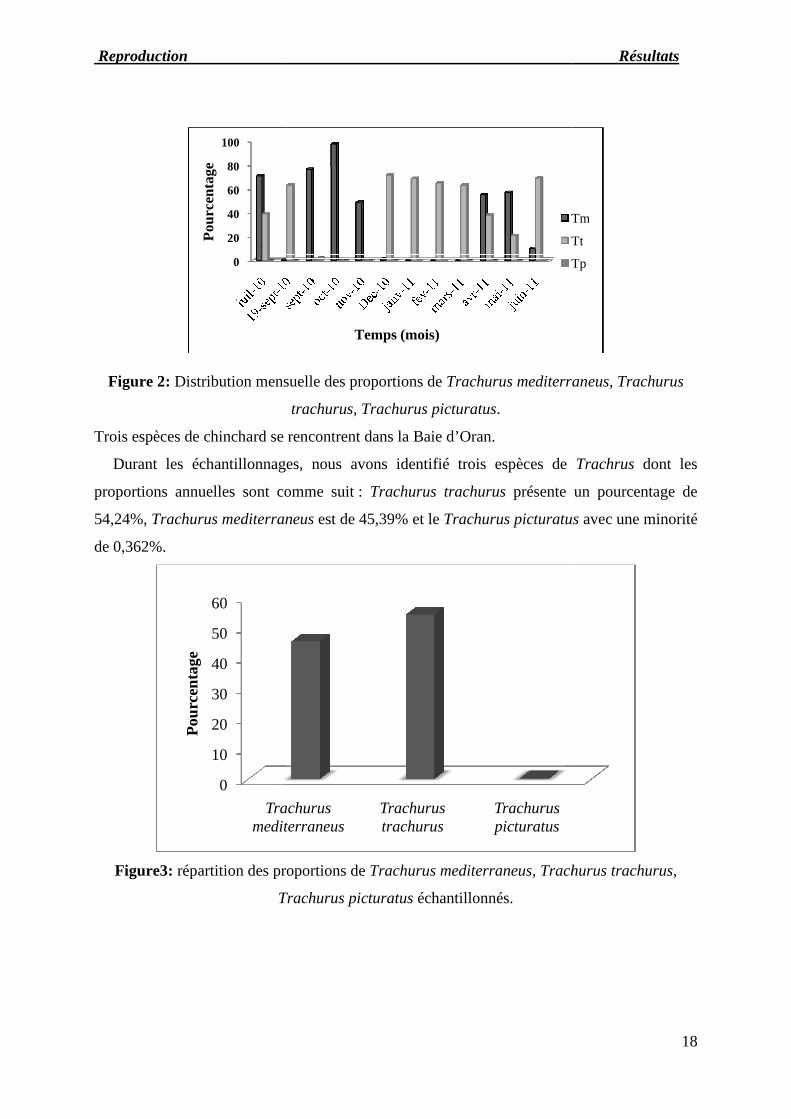
Reproduction Résul
Figure 2: Distribution mensuelle des proportions de
Trois espèces de chinchard se rencontrent dans la Baie d’Oran.
Durant les échantillonnages, nous avons identifié trois espèces de
proportions annuelles sont comme suit
54,24%, Trachurus mediterraneus
de 0,362%.
Figure3: répartition des proportions de
Trachurus picturatus
0
20
40
60
80
100
Po
urc
enta
ge
0
10
20
30
40
50
60
Trachurusmediterraneus
Pou
rcen
tage
Reproduction Résul
Distribution mensuelle des proportions de Trachurus mediterraneus, Trachurus
trachurus, Trachurus picturatus.
Trois espèces de chinchard se rencontrent dans la Baie d’Oran.
Durant les échantillonnages, nous avons identifié trois espèces de
proportions annuelles sont comme suit : Trachurus trachurus présente un pourcentage de
Trachurus mediterraneus est de 45,39% et le Trachurus picturatus
répartition des proportions de Trachurus mediterraneus, Trachurus trachurus,
Trachurus picturatus échantillonnés.
Temps (mois)
Tm
Tt
Tp
Trachurusmediterraneus
Trachurustrachurus
Trachuruspicturatus
Reproduction Résultats
18
erraneus, Trachurus
Durant les échantillonnages, nous avons identifié trois espèces de Trachrus dont les
présente un pourcentage de
Trachurus picturatus avec une minorité
Trachurus mediterraneus, Trachurus trachurus,
Tm
Tt
Tp

Reproduction Résul
a) Trachurus mediterraneus
Figure
19 cm
Reproduction Résul
Trachurus mediterraneus b) Trachurus picturatus
c)Trachurus trachurus
b) Scutelles
Figure 4: Espèces de Trachurus échantillonnés.
20 cm
BENSAADA F Z, 2011
BENSAADA F Z, 2011
18 cm
BENSAADA F Z,
Reproduction Résultats
19
Trachurus picturatus
Reproduction Résultats
20
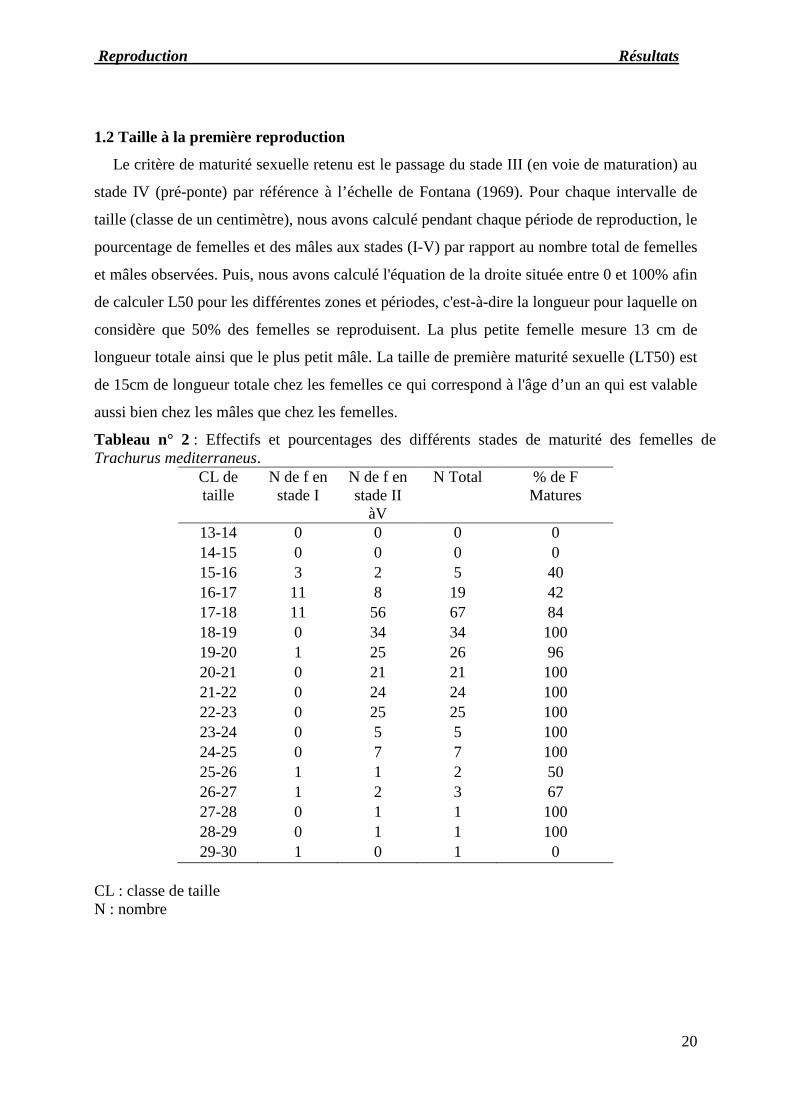
1.2 Taille à la première reproduction
Le critère de maturité sexuelle retenu est le passage du stade III (en voie de maturation) au
stade IV (pré-ponte) par référence à l’échelle de Fontana (1969). Pour chaque intervalle de
taille (classe de un centimètre), nous avons calculé pendant chaque période de reproduction, le
pourcentage de femelles et des mâles aux stades (I-V) par rapport au nombre total de femelles
et mâles observées. Puis, nous avons calculé l'équation de la droite située entre 0 et 100% afin
de calculer L50 pour les différentes zones et périodes, c'est-à-dire la longueur pour laquelle on
considère que 50% des femelles se reproduisent. La plus petite femelle mesure 13 cm de
longueur totale ainsi que le plus petit mâle. La taille de première maturité sexuelle (LT50) est
de 15cm de longueur totale chez les femelles ce qui correspond à l'âge d’un an qui est valable
aussi bien chez les mâles que chez les femelles.
Tableau n° 2 : Effectifs et pourcentages des différents stades de maturité des femelles deTrachurus mediterraneus.
CL detaille
N de f enstade I
N de f enstade II
àV
N Total % de FMatures
13-14 0 0 0 014-15 0 0 0 015-16 3 2 5 4016-17 11 8 19 4217-18 11 56 67 8418-19 0 34 34 10019-20 1 25 26 9620-21 0 21 21 10021-22 0 24 24 10022-23 0 25 25 10023-24 0 5 5 10024-25 0 7 7 10025-26 1 1 2 5026-27 1 2 3 6727-28 0 1 1 10028-29 0 1 1 10029-30 1 0 1 0
CL : classe de tailleN : nombre
Reproduction Résultats
21
12 14 16 18 20 22 24 26 28 30
LT(cm)
0
20
40
60
80
100
Po
urc
enta
ge
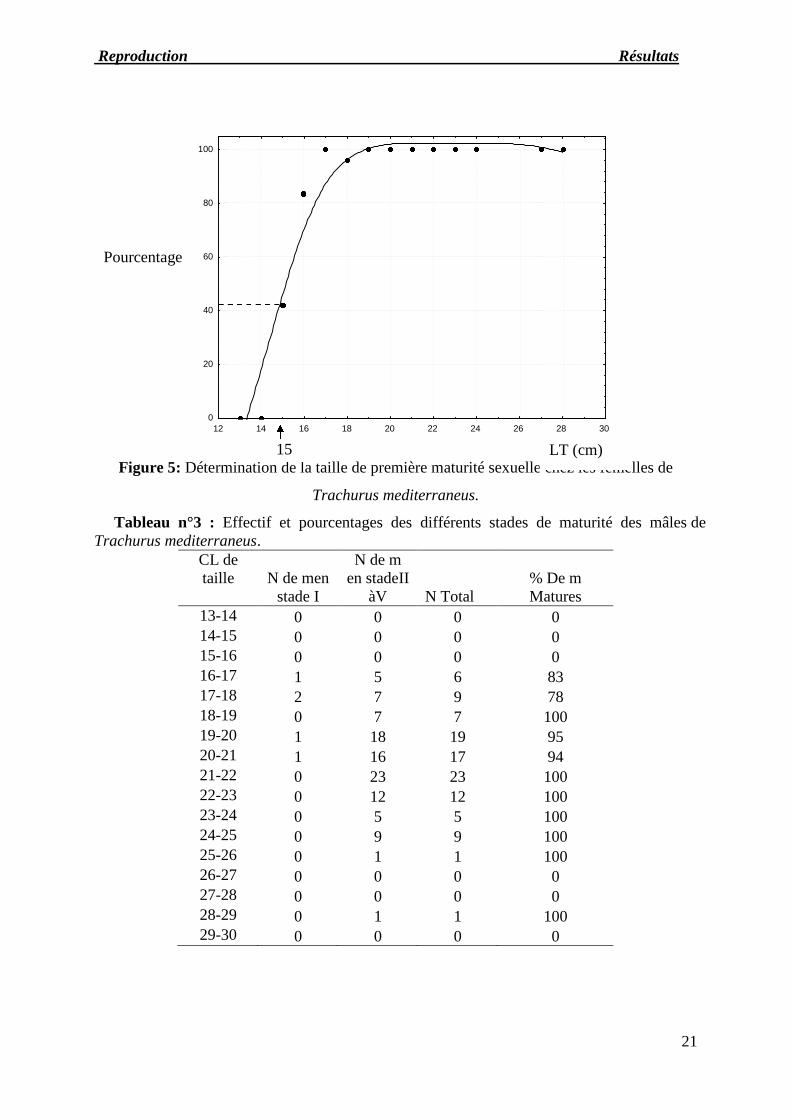
Figure 5: Détermination de la taille de première maturité sexuelle chez les femelles de
Trachurus mediterraneus.
Tableau n°3 : Effectif et pourcentages des différents stades de maturité des mâles deTrachurus mediterraneus.
CL detaille N de men
stade I
N de men stadeII
àV N Total% De mMatures
13-14 0 0 0 014-15 0 0 0 015-16 0 0 0 016-17 1 5 6 8317-18 2 7 9 7818-19 0 7 7 10019-20 1 18 19 9520-21 1 16 17 9421-22 0 23 23 10022-23 0 12 12 10023-24 0 5 5 10024-25 0 9 9 10025-26 0 1 1 10026-27 0 0 0 027-28 0 0 0 028-29 0 1 1 10029-30 0 0 0 0
Pourcentage
LT (cm)15
Reproduction Résultats
22
12 14 16 18 20 22 24 26 28 30
LT(cm)
0
20
40
60
80
100
Po
urc
enta
ge
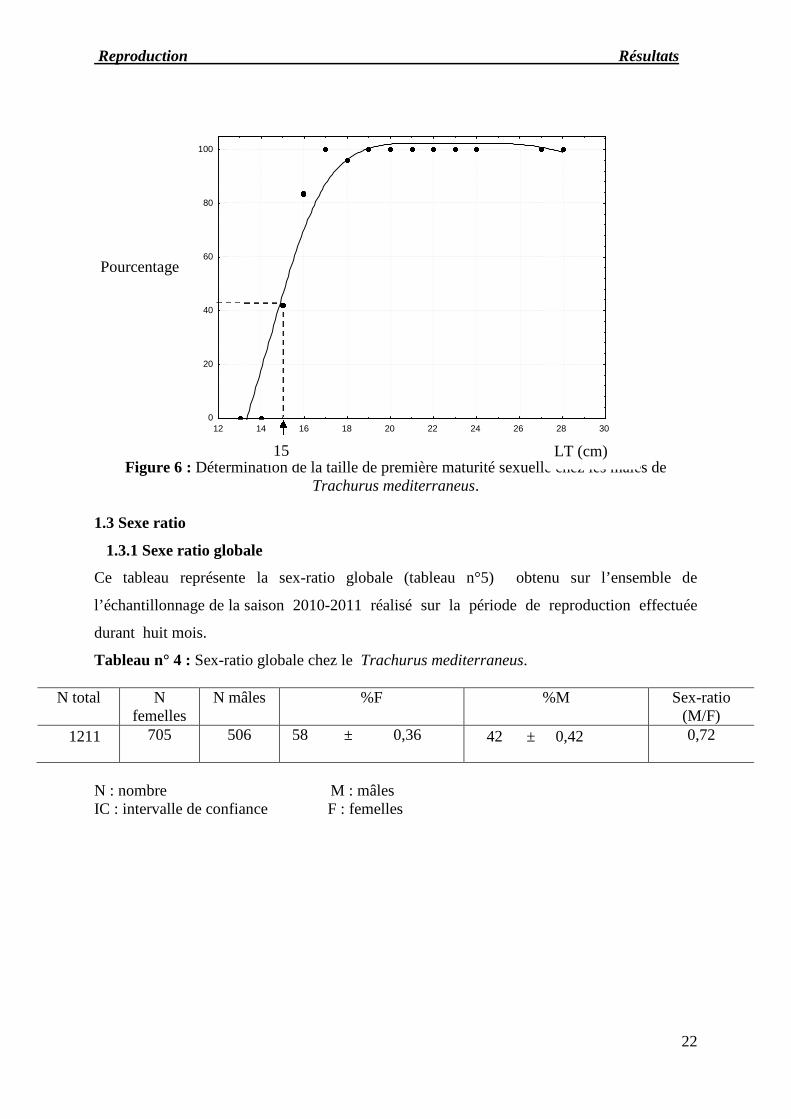
Figure 6 : Détermination de la taille de première maturité sexuelle chez les mâles deTrachurus mediterraneus.
1.3 Sexe ratio
1.3.1 Sexe ratio globale
Ce tableau représente la sex-ratio globale (tableau n°5) obtenu sur l’ensemble de
l’échantillonnage de la saison 2010-2011 réalisé sur la période de reproduction effectuée
durant huit mois.
Tableau n° 4 : Sex-ratio globale chez le Trachurus mediterraneus.
N total Nfemelles
N mâles %F %M Sex-ratio(M/F)
1211 705 506 58 ± 0,36 42 ± 0,42 0,72
N : nombre M : mâlesIC : intervalle de confiance F : femelles
Pourcentage
LT (cm)15
Reproduction Résul
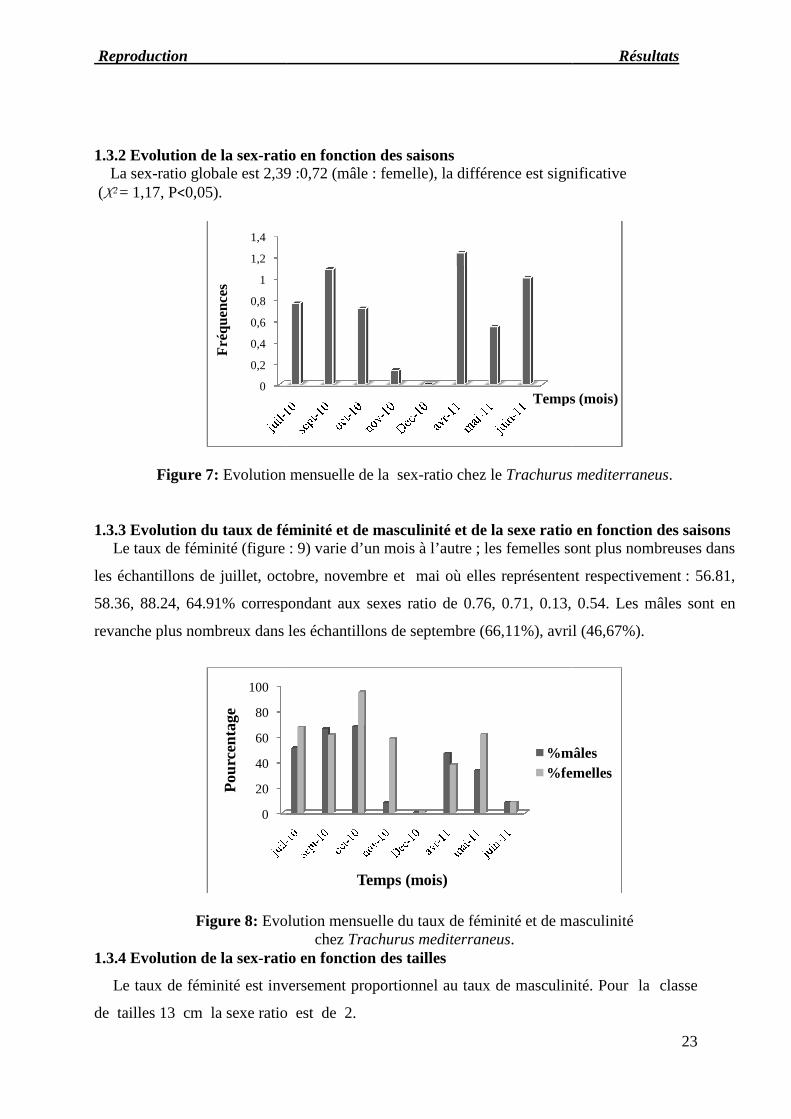
1.3.2 Evolution de la sex-ratio en fonction desLa sex-ratio globale est 2,39
(X2 = 1,17, P<0,05).
Figure 7: Evolution mensuelle de la sex
1.3.3 Evolution du taux de féminité et de masculinité etLe taux de féminité (figure
les échantillons de juillet, octobre, novembre et mai où elles
58.36, 88.24, 64.91% correspondant aux sexes ratio de 0.76, 0.71, 0.13, 0.54. Les
revanche plus nombreux dans les échantillons de septembre (66,11%), avril
Figure 8: Evolution mensuelle du t
1.3.4 Evolution de la sex-ratio en fonction des tailles
Le taux de féminité est inversement proportionnel au taux de masculinité.
de tailles 13 cm la sexe ratio est de
0
0,2
0,4
0,6
0,8
1
1,2
1,4
Fré
qu
ence
s
0
20
40
60
80
100
Pou
rcen
tage
Reproduction Résul
atio en fonction des saisonsratio globale est 2,39 :0,72 (mâle : femelle), la différence est significative
Evolution mensuelle de la sex-ratio chez le Trachurus
Evolution du taux de féminité et de masculinité et de la sexe ratio en fonction des saisons: 9) varie d’un mois à l’autre ; les femelles sont plus nombreuses dans
les échantillons de juillet, octobre, novembre et mai où elles représentent respectivement
58.36, 88.24, 64.91% correspondant aux sexes ratio de 0.76, 0.71, 0.13, 0.54. Les
revanche plus nombreux dans les échantillons de septembre (66,11%), avril
Evolution mensuelle du taux de féminité et de masculinitéchez Trachurus mediterraneus.
ratio en fonction des tailles
Le taux de féminité est inversement proportionnel au taux de masculinité.
ratio est de 2.
Temps (mois)
Temps (mois)
%mâles
%femelles
Reproduction Résultats
23
), la différence est significative
Trachurus mediterraneus.
e ratio en fonction des saisons; les femelles sont plus nombreuses dans
représentent respectivement : 56.81,
58.36, 88.24, 64.91% correspondant aux sexes ratio de 0.76, 0.71, 0.13, 0.54. Les mâles sont en
revanche plus nombreux dans les échantillons de septembre (66,11%), avril (46,67%).
aux de féminité et de masculinité
Le taux de féminité est inversement proportionnel au taux de masculinité. Pour la classe
Temps (mois)
%mâles
%femelles
Reproduction Résultats
24
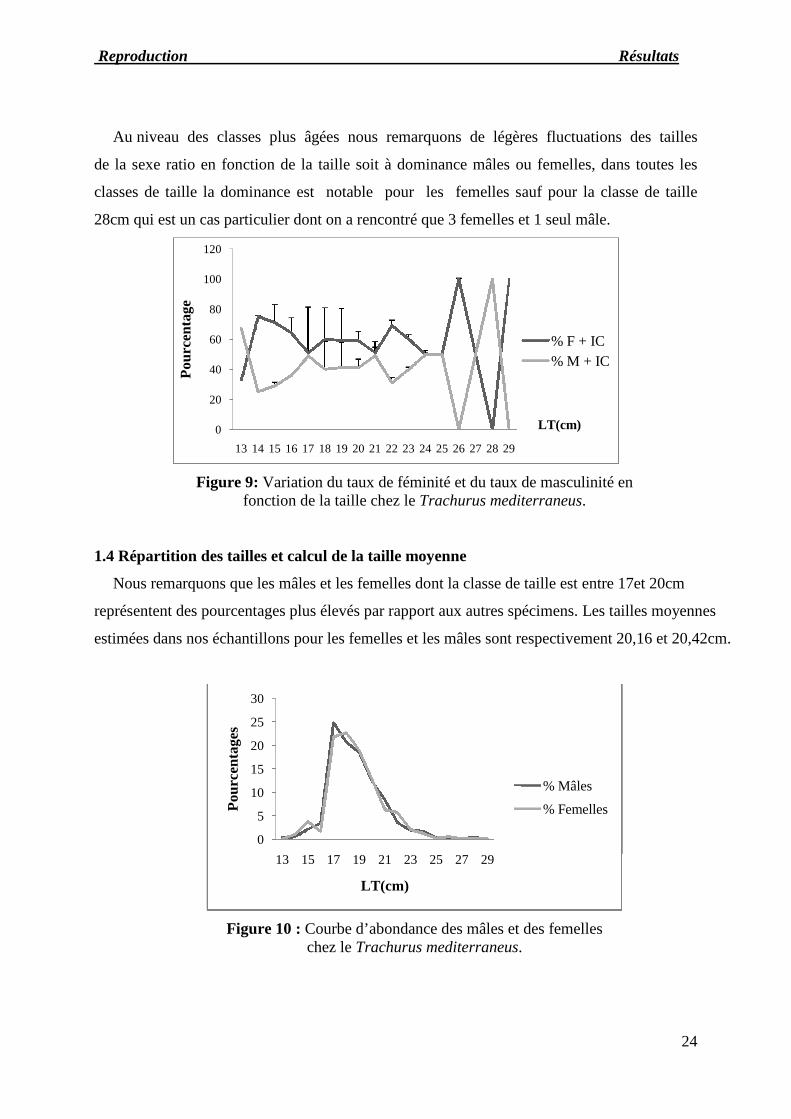
Au niveau des classes plus âgées nous remarquons de légères fluctuations des tailles
de la sexe ratio en fonction de la taille soit à dominance mâles ou femelles, dans toutes les
classes de taille la dominance est notable pour les femelles sauf pour la classe de taille
28cm qui est un cas particulier dont on a rencontré que 3 femelles et 1 seul mâle.
Figure 9: Variation du taux de féminité et du taux de masculinité enfonction de la taille chez le Trachurus mediterraneus.
1.4 Répartition des tailles et calcul de la taille moyenne
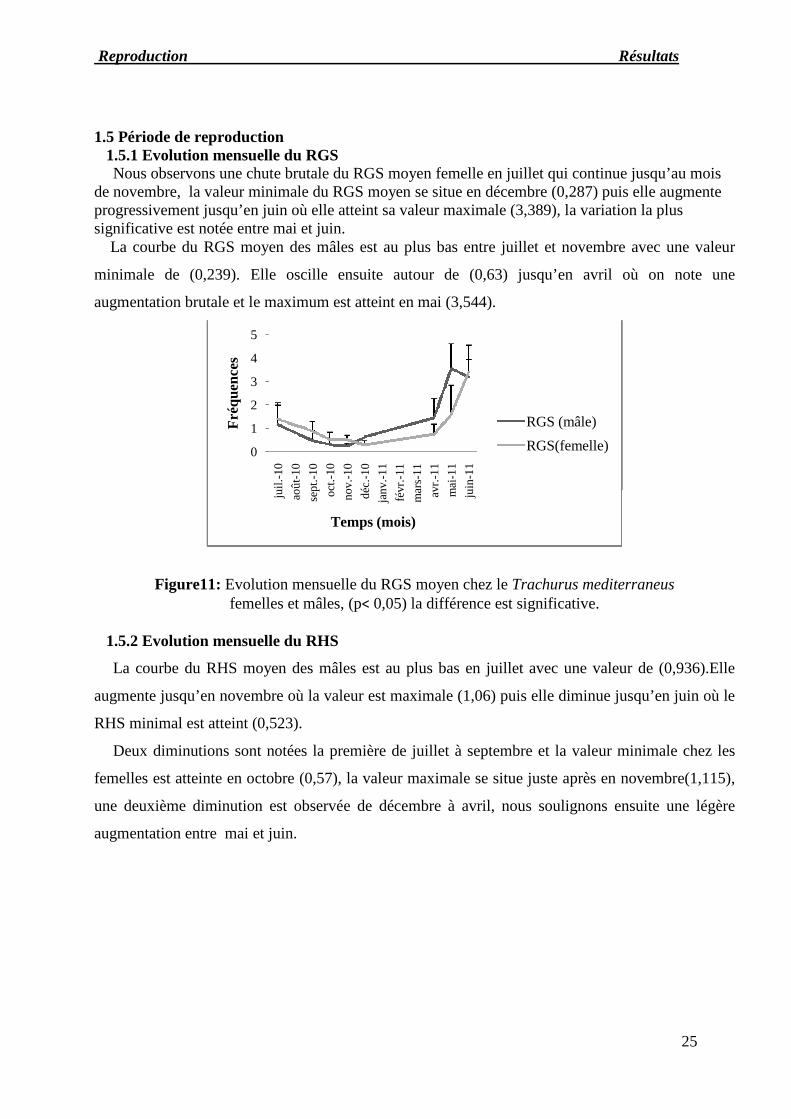
Nous remarquons que les mâles et les femelles dont la classe de taille est entre 17et 20cm
représentent des pourcentages plus élevés par rapport aux autres spécimens. Les tailles moyennes
estimées dans nos échantillons pour les femelles et les mâles sont respectivement 20,16 et 20,42cm.
Figure 10 : Courbe d’abondance des mâles et des femelleschez le Trachurus mediterraneus.
0
20
40
60
80
100
120
13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
Po
urc
enta
ge
% F + IC
% M + IC
LT(cm)
0
5
10
15
20
25
30
13 15 17 19 21 23 25 27 29
Po
urc
enta
ges
LT(cm)
% Mâles
% Femelles
Reproduction Résultats
25
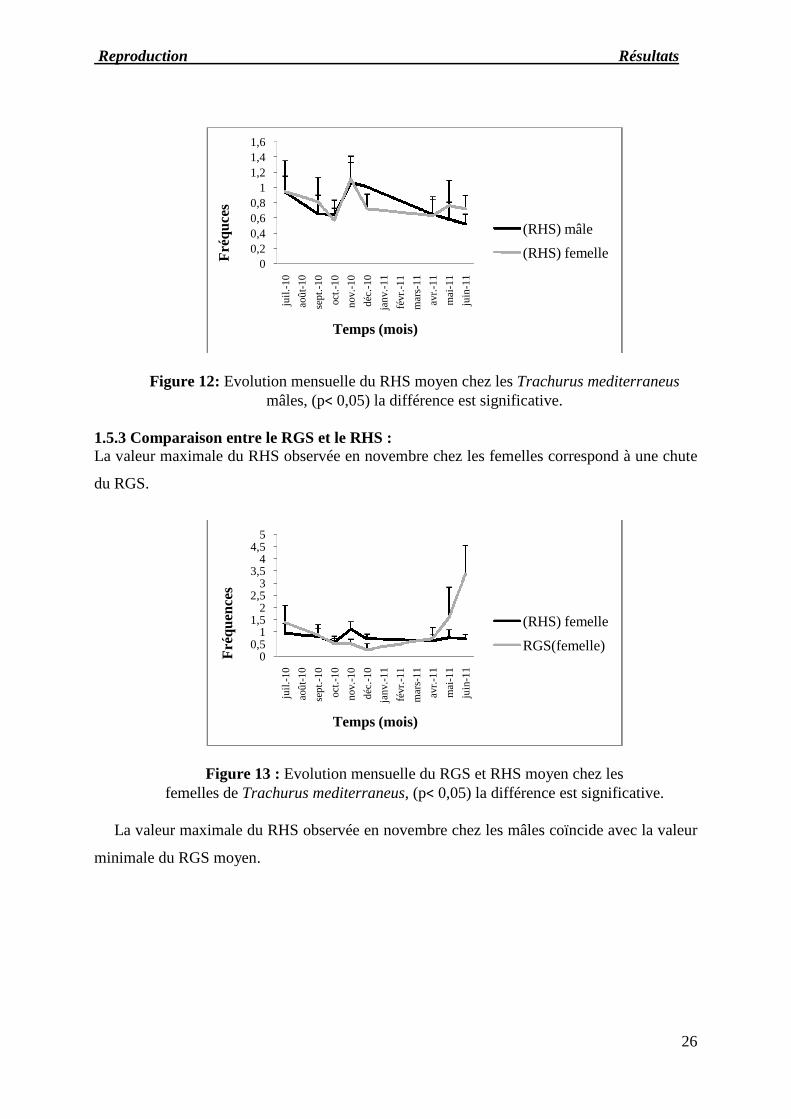
1.5 Période de reproduction1.5.1 Evolution mensuelle du RGSNous observons une chute brutale du RGS moyen femelle en juillet qui continue jusqu’au mois
de novembre, la valeur minimale du RGS moyen se situe en décembre (0,287) puis elle augmenteprogressivement jusqu’en juin où elle atteint sa valeur maximale (3,389), la variation la plussignificative est notée entre mai et juin.
La courbe du RGS moyen des mâles est au plus bas entre juillet et novembre avec une valeur
minimale de (0,239). Elle oscille ensuite autour de (0,63) jusqu’en avril où on note une
augmentation brutale et le maximum est atteint en mai (3,544).
Figure11: Evolution mensuelle du RGS moyen chez le Trachurus mediterraneusfemelles et mâles, (p< 0,05) la différence est significative.
1.5.2 Evolution mensuelle du RHS
La courbe du RHS moyen des mâles est au plus bas en juillet avec une valeur de (0,936).Elle
augmente jusqu’en novembre où la valeur est maximale (1,06) puis elle diminue jusqu’en juin où le
RHS minimal est atteint (0,523).
Deux diminutions sont notées la première de juillet à septembre et la valeur minimale chez les
femelles est atteinte en octobre (0,57), la valeur maximale se situe juste après en novembre(1,115),
une deuxième diminution est observée de décembre à avril, nous soulignons ensuite une légère
augmentation entre mai et juin.
0
1
2
3
4
5
juil
.-1
0
aoû
t-1
0
sep
t.-1
0
oct
.-1
0
no
v.-1
0
déc
.-1
0
jan
v.-1
1
févr
.-1
1
mar
s-1
1
avr.
-11
mai
-11
juin
-11
Fré
qu
ence
s
Temps (mois)
RGS (mâle)
RGS(femelle)
Reproduction Résultats
26
Figure 12: Evolution mensuelle du RHS moyen chez les Trachurus mediterraneusmâles, (p< 0,05) la différence est significative.
1.5.3 Comparaison entre le RGS et le RHS :La valeur maximale du RHS observée en novembre chez les femelles correspond à une chute
du RGS.
Figure 13 : Evolution mensuelle du RGS et RHS moyen chez lesfemelles de Trachurus mediterraneus, (p< 0,05) la différence est significative.
La valeur maximale du RHS observée en novembre chez les mâles coïncide avec la valeur
minimale du RGS moyen.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
juil
.-1
0
aoû
t-1
0
sep
t.-1
0
oct.
-10
nov
.-1
0
déc
.-1
0
jan
v.-1
1
févr
.-1
1
mar
s-1
1
avr.
-11
mai
-11
juin
-11
Fré
qu
ces
Temps (mois)
(RHS) mâle
(RHS) femelle
00,5
11,5
22,5
33,5
44,5
5
juil
.-1
0
aoû
t-1
0
sep
t.-1
0
oct.
-10
nov
.-1
0
déc
.-1
0
jan
v.-1
1
févr
.-1
1
mar
s-1
1
avr.
-11
mai
-11
juin
-11
Fré
qu
ence
s
Temps (mois)
(RHS) femelle
RGS(femelle)
Reproduction Résultats
27
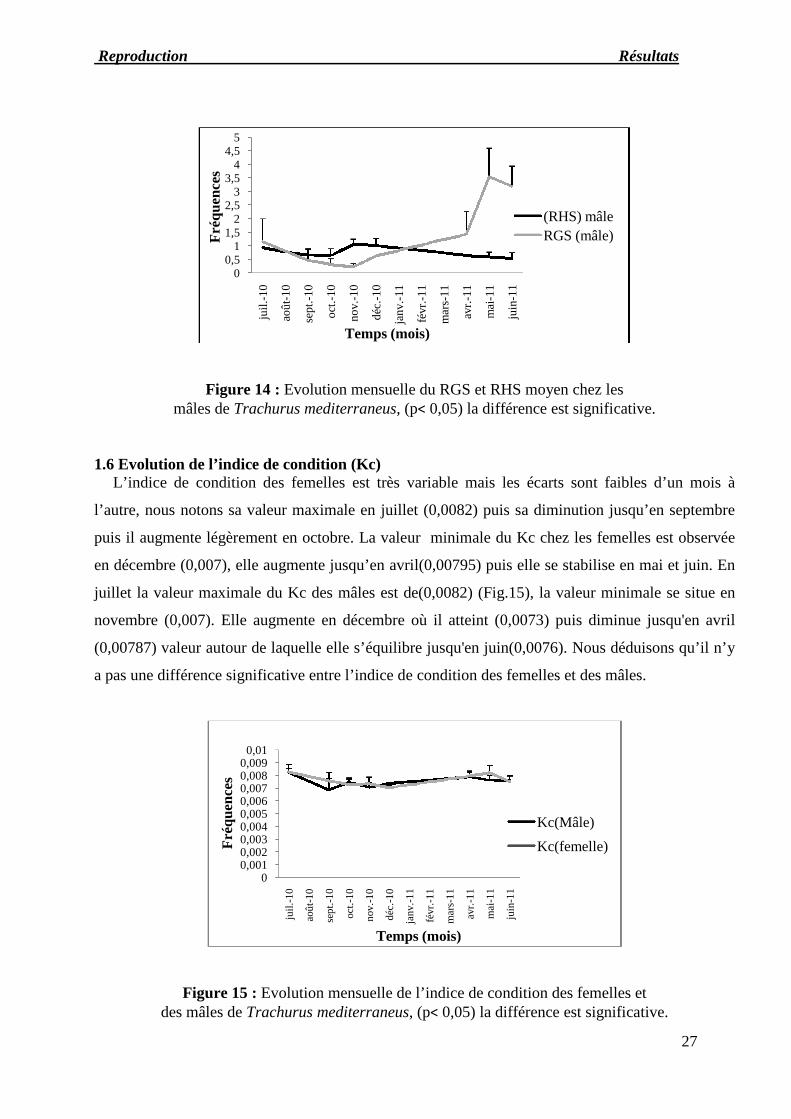
Figure 14 : Evolution mensuelle du RGS et RHS moyen chez lesmâles de Trachurus mediterraneus, (p< 0,05) la différence est significative.
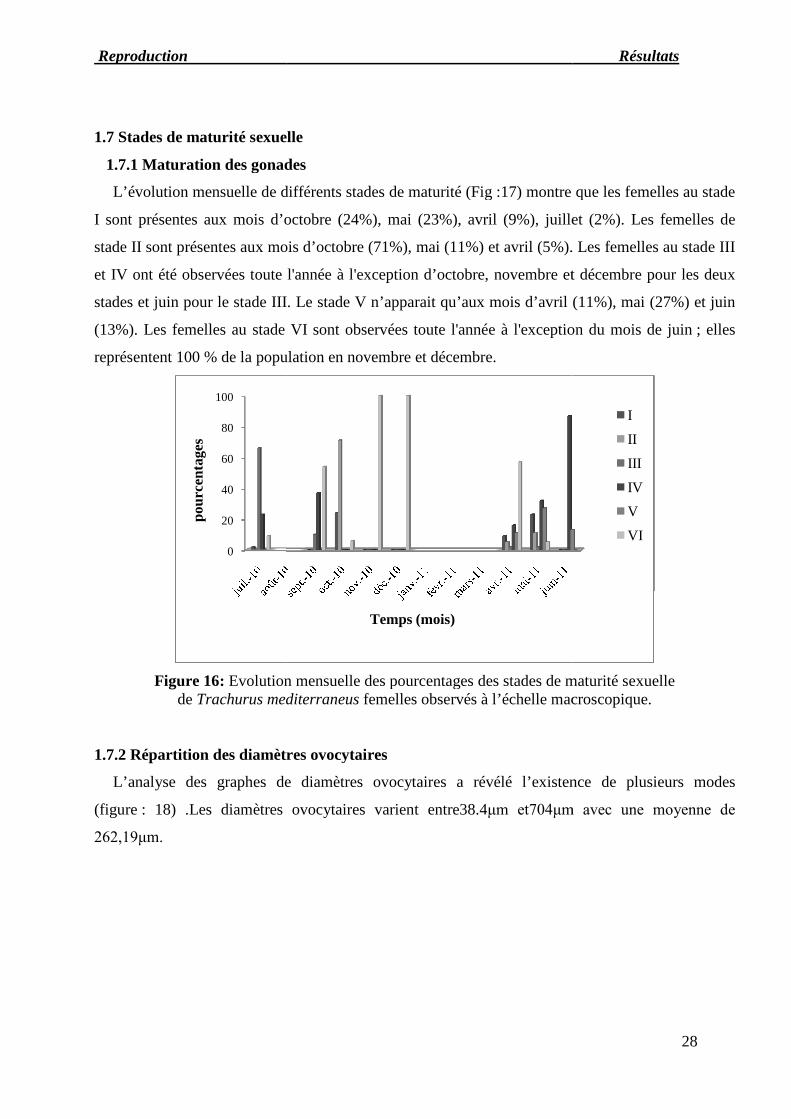
1.6 Evolution de l’indice de condition (Kc)L’indice de condition des femelles est très variable mais les écarts sont faibles d’un mois à
l’autre, nous notons sa valeur maximale en juillet (0,0082) puis sa diminution jusqu’en septembre
puis il augmente légèrement en octobre. La valeur minimale du Kc chez les femelles est observée
en décembre (0,007), elle augmente jusqu’en avril(0,00795) puis elle se stabilise en mai et juin. En
juillet la valeur maximale du Kc des mâles est de(0,0082) (Fig.15), la valeur minimale se situe en
novembre (0,007). Elle augmente en décembre où il atteint (0,0073) puis diminue jusqu'en avril
(0,00787) valeur autour de laquelle elle s’équilibre jusqu'en juin(0,0076). Nous déduisons qu’il n’y
a pas une différence significative entre l’indice de condition des femelles et des mâles.
Figure 15 : Evolution mensuelle de l’indice de condition des femelles etdes mâles de Trachurus mediterraneus, (p< 0,05) la différence est significative.
00,5
11,5
22,5
33,5
44,5
5
juil
.-1
0
aoû
t-1
0
sep
t.-1
0
oct
.-1
0
no
v.-1
0
déc
.-1
0
jan
v.-1
1
févr
.-1
1
mar
s-1
1
avr.
-11
mai
-11
juin
-11
Fré
qu
ence
s
Temps (mois)
(RHS) mâle
RGS (mâle)
00,0010,0020,0030,0040,0050,0060,0070,0080,009
0,01
juil
.-1
0
aoû
t-1
0
sep
t.-1
0
oct.
-10
nov
.-1
0
déc
.-1
0
jan
v.-1
1
févr
.-1
1
mar
s-1
1
avr.
-11
mai
-11
juin
-11
Fré
qu
ence
s
Temps (mois)
Kc(Mâle)
Kc(femelle)
Reproduction Résul
1.7 Stades de maturité sexuelle
1.7.1 Maturation des gonades
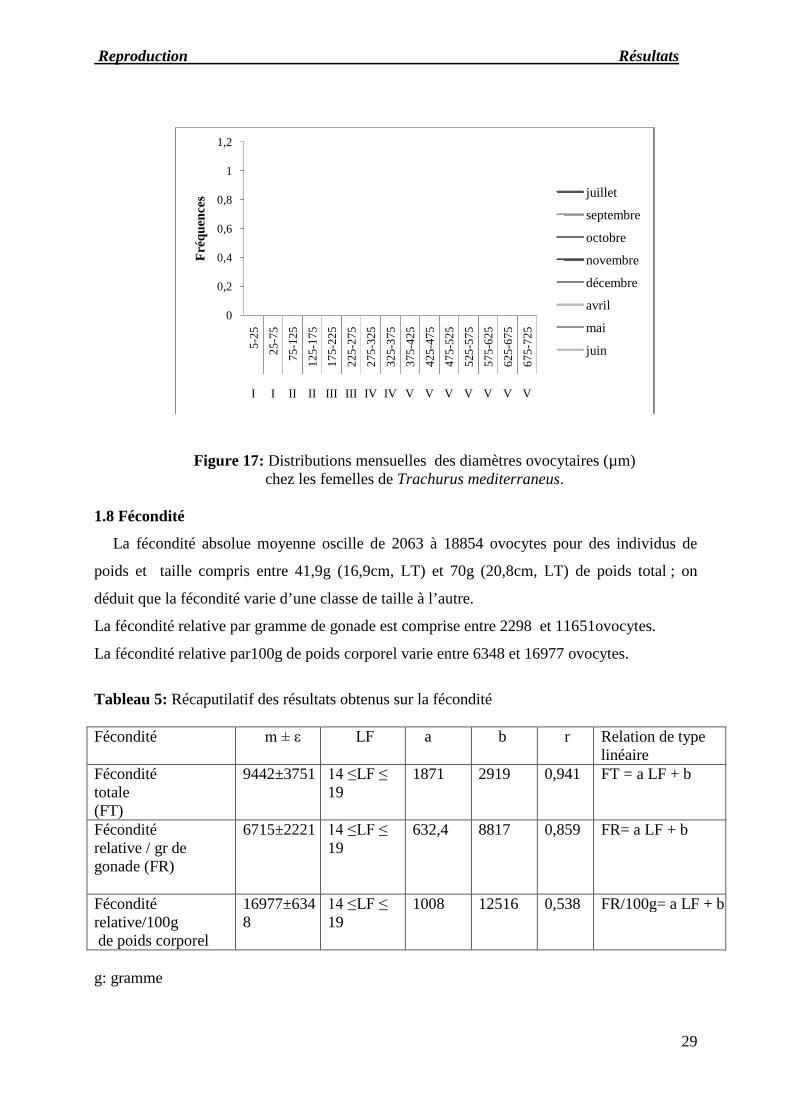
L’évolution mensuelle de différents stades de maturité (Fig
I sont présentes aux mois d’octobre (24%), mai (23%), avril (9%), juillet (2%). Les femelles de
stade II sont présentes aux mois d’octobre (71%), mai (11%) et avril (5%). Les feme
et IV ont été observées toute l'année à l'exception
stades et juin pour le stade III. Le stade V n’apparait qu’aux mois d’avril (11%), mai (27%) et juin
(13%). Les femelles au stade
représentent 100 % de la population en
Figure 16: Evolution mensuelle des pourcentages des stades de maturité sexuellede Trachurus mediterraneus
1.7.2 Répartition des diamètres ovocytaires
L’analyse des graphes de diamètres ovocytaires
(figure : 18) .Les diamètres ovocytaires varient entre38.4
262,19μm.
0
20
40
60
80
100
po
urc
enta
ges
Reproduction Résul
1.7 Stades de maturité sexuelle
1.7.1 Maturation des gonades
de différents stades de maturité (Fig :17) montre que les femelles au stade
I sont présentes aux mois d’octobre (24%), mai (23%), avril (9%), juillet (2%). Les femelles de
stade II sont présentes aux mois d’octobre (71%), mai (11%) et avril (5%). Les feme
ont été observées toute l'année à l'exception d’octobre, novembre et décembre pour les deux
stades et juin pour le stade III. Le stade V n’apparait qu’aux mois d’avril (11%), mai (27%) et juin
Les femelles au stade VI sont observées toute l'année à l'exception d
% de la population en novembre et décembre.
Evolution mensuelle des pourcentages des stades de maturité sexuellemediterraneus femelles observés à l’échelle macroscopique.
ition des diamètres ovocytaires
L’analyse des graphes de diamètres ovocytaires a révélé l’existence de plusieurs modes
: 18) .Les diamètres ovocytaires varient entre38.4μm et704μm avec une moyenne de
Temps (mois)
Reproduction Résultats
28
:17) montre que les femelles au stade
I sont présentes aux mois d’octobre (24%), mai (23%), avril (9%), juillet (2%). Les femelles de
stade II sont présentes aux mois d’octobre (71%), mai (11%) et avril (5%). Les femelles au stade III
d’octobre, novembre et décembre pour les deux
stades et juin pour le stade III. Le stade V n’apparait qu’aux mois d’avril (11%), mai (27%) et juin
servées toute l'année à l'exception du mois de juin ; elles
Evolution mensuelle des pourcentages des stades de maturité sexuellefemelles observés à l’échelle macroscopique.
a révélé l’existence de plusieurs modes
04μm avec une moyenne de
I
II
III
IV
V
VI
Reproduction Résultats
29
Figure 17: Distributions mensuelles des diamètres ovocytaires (µm)chez les femelles de Trachurus mediterraneus.
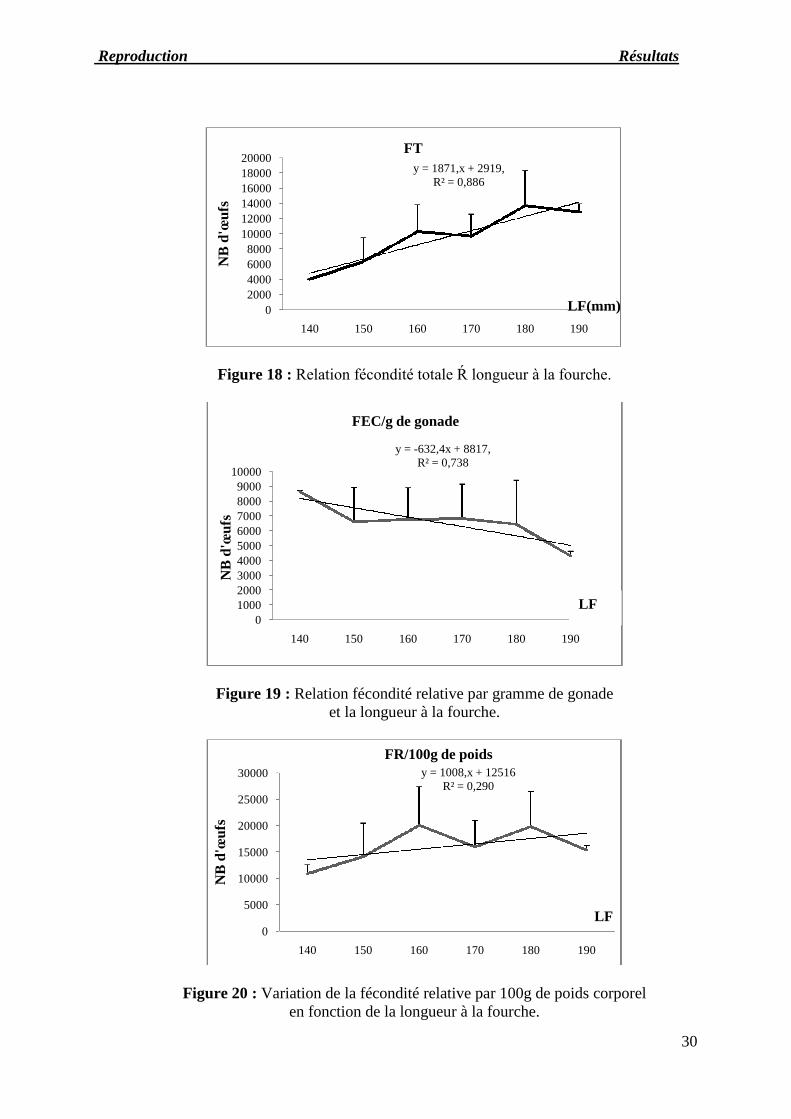
1.8 Fécondité
La fécondité absolue moyenne oscille de 2063 à 18854 ovocytes pour des individus de
poids et taille compris entre 41,9g (16,9cm, LT) et 70g (20,8cm, LT) de poids total ; on
déduit que la fécondité varie d’une classe de taille à l’autre.
La fécondité relative par gramme de gonade est comprise entre 2298 et 11651ovocytes.
La fécondité relative par100g de poids corporel varie entre 6348 et 16977 ovocytes.
Tableau 5: Récaputilatif des résultats obtenus sur la fécondité
Fécondité m ± ε LF a b r Relation de typelinéaire
Féconditétotale(FT)
9442±3751 14 ≤LF ≤ 19
1871 2919 0,941 FT = a LF + b
Féconditérelative / gr degonade (FR)
6715±2221 14 ≤LF ≤ 19
632,4 8817 0,859 FR= a LF + b
Féconditérelative/100gde poids corporel
16977±6348
14 ≤LF ≤ 19
1008 12516 0,538 FR/100g= a LF + b
g: gramme
0
0,2
0,4
0,6
0,8
1
1,2
5-2
5
25
-75
75
-12
5
12
5-1
75
17
5-2
25
22
5-2
75
27
5-3
25
32
5-3
75
37
5-4
25
42
5-4
75
47
5-5
25
52
5-5
75
57
5-6
25
62
5-6
75
67
5-7
25
I I II II III III IV IV V V V V V V V
Fré
qu
ence
s juillet
septembre
octobre
novembre
décembre
avril
mai
juin
Reproduction Résultats
30
Figure 18 : Relation fécondité totale Ŕ longueur à la fourche.
Figure 19 : Relation fécondité relative par gramme de gonadeet la longueur à la fourche.
Figure 20 : Variation de la fécondité relative par 100g de poids corporelen fonction de la longueur à la fourche.
y = 1871,x + 2919,R² = 0,886
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
140 150 160 170 180 190
NB
d'œ
ufs
FT
LF(mm)
y = -632,4x + 8817,R² = 0,738
0100020003000400050006000700080009000
10000
140 150 160 170 180 190
NB
d'œ
ufs
FEC/g de gonade
LF
y = 1008,x + 12516R² = 0,290
0
5000
10000
15000
20000
25000
30000
140 150 160 170 180 190
NB
d'œ
ufs
FR/100g de poids
LF
Reproduction Résultats
31
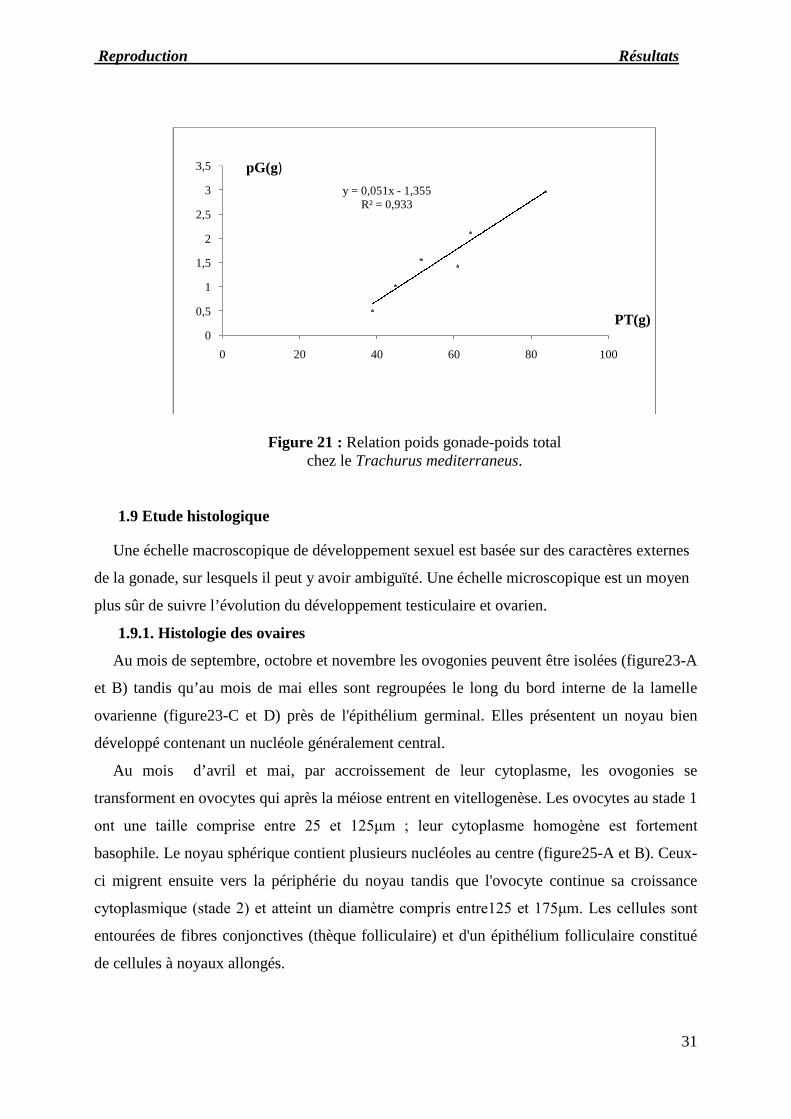
Figure 21 : Relation poids gonade-poids totalchez le Trachurus mediterraneus.
1.9 Etude histologique
Une échelle macroscopique de développement sexuel est basée sur des caractères externes
de la gonade, sur lesquels il peut y avoir ambiguïté. Une échelle microscopique est un moyen
plus sûr de suivre l’évolution du développement testiculaire et ovarien.
1.9.1. Histologie des ovaires
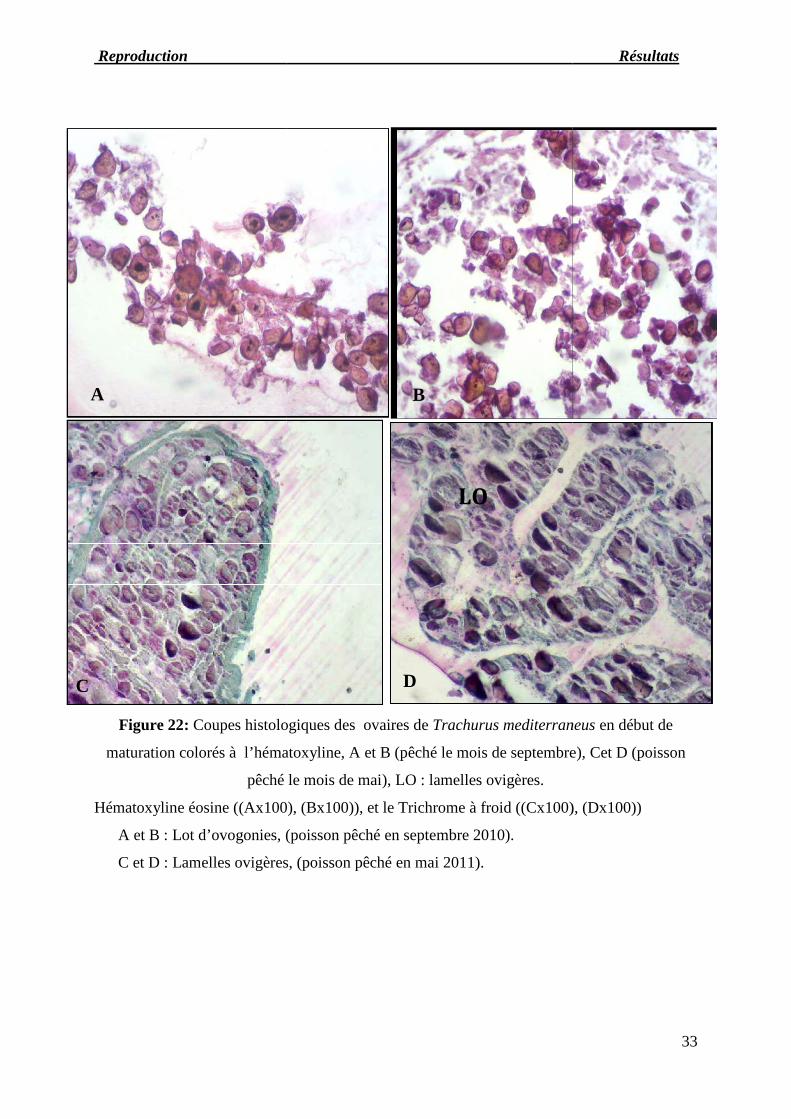
Au mois de septembre, octobre et novembre les ovogonies peuvent être isolées (figure23-A
et B) tandis qu’au mois de mai elles sont regroupées le long du bord interne de la lamelle
ovarienne (figure23-C et D) près de l'épithélium germinal. Elles présentent un noyau bien
développé contenant un nucléole généralement central.
Au mois d’avril et mai, par accroissement de leur cytoplasme, les ovogonies se
transforment en ovocytes qui après la méiose entrent en vitellogenèse. Les ovocytes au stade 1
ont une taille comprise entre 25 et 125μm ; leur cytoplasme homogène est fortement
basophile. Le noyau sphérique contient plusieurs nucléoles au centre (figure25-A et B). Ceux-
ci migrent ensuite vers la périphérie du noyau tandis que l'ovocyte continue sa croissance
cytoplasmique (stade 2) et atteint un diamètre compris entre125 et 175μm. Les cellules sont
entourées de fibres conjonctives (thèque folliculaire) et d'un épithélium folliculaire constitué
de cellules à noyaux allongés.
y = 0,051x - 1,355R² = 0,933
0
0,5
1
1,5
2
2,5
3
3,5
0 20 40 60 80 100
PT(g)
pG(g)

Reproduction Résultats
32
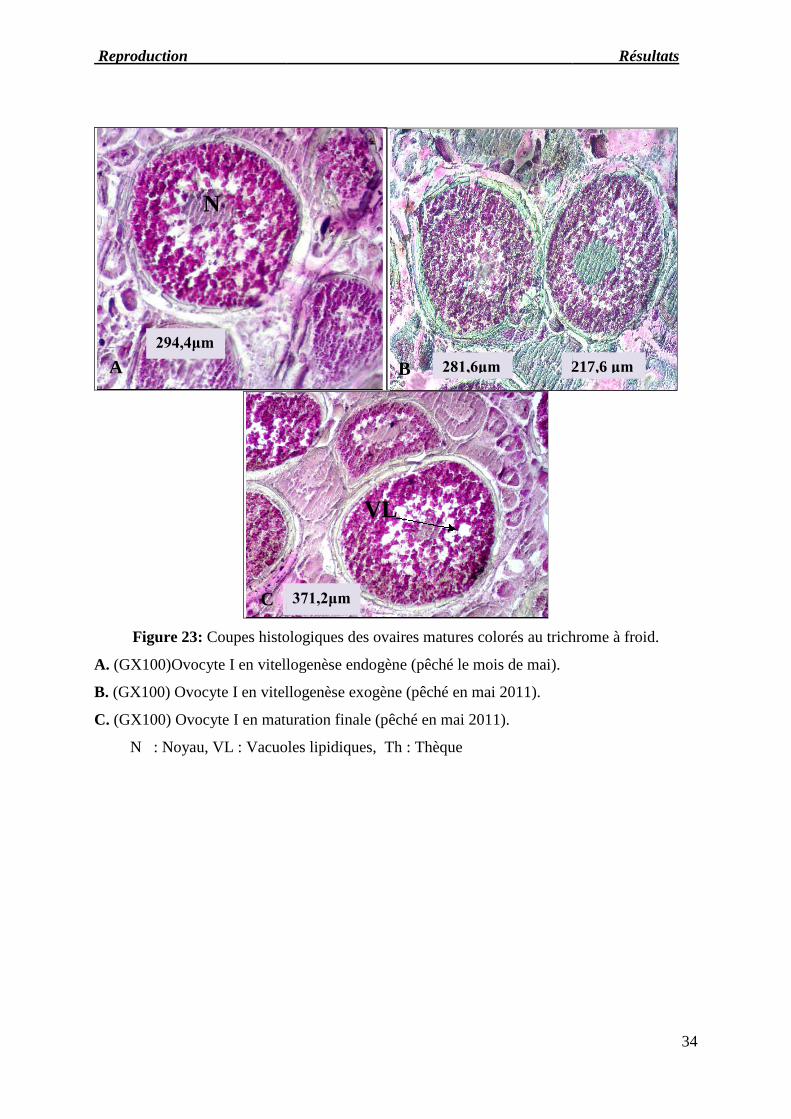
Les ovocytes de stade 3 de diamètre compris entre (175 et 275μm) observés au mois de
mai peuvent être considérés comme entrant en début de maturation; les cellules subissent
d'importantes transformations qui concernent le noyau, le nucléoplasme et les membranes. Le
cytoplasme devient moins basophile.
La membrane nucléaire perd sa régularité et forme des villosités dans lesquelles s'insèrent
les nucléoles; autour du noyau apparaît une plage concentrique transitoire dans laquelle se
forment des vacuoles lipidiques (le stade 3 est souvent décrit comme une phase de
vacuolisation) (figure26-A, B et D). Le cytoplasme à la périphérie (cytoplasme cortical) prend
un aspect radié dû à la présence de microvillosités. L'enveloppe continue formée autour de
l'ovocyte par les cellules de l'épithélium folliculaire devient nette à ce stade et atteint sa taille
maximale; elle est appelée granulosa. Entre cette enveloppe et la membrane cytoplasmique se
développe une zone riche en mucopolysaccharides acides appelée membrane pellucide ou
zona radiata. Elle est en effet traversée de stries radiaires par où passent les microvillosités du
cytoplasme et les prolongements des cellules folliculaires.
Au mois de mai, le stade 4 des ovocytes peut être identifié à une phase de vitellogenèse.
Une couronne de globules vitellins apparaît à la périphérie tandis que les vacuoles lipidiques
restent concentrées autour du noyau. Cet envahissement de l'espace cytoplasmique par les
globules vitellins entraîne un accroissement des ovocytes dont le diamètre atteint 275 à
375μm (figure24-A, B et C). La zona radiata s'épaissie (17,4μm) et la granulosa est entourée
d'une thèque conjonctive.
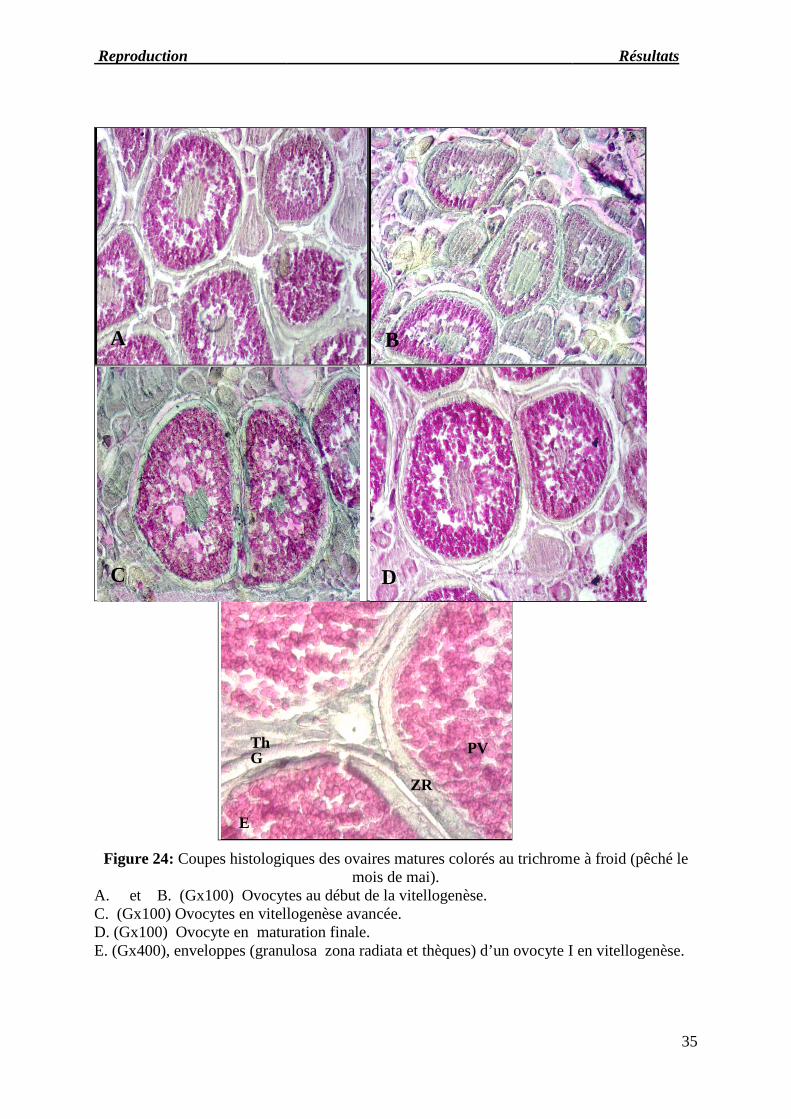
Au stade 5, les ovocytes d'un diamètre de 380 à plus de 475μm présentent, entre l'étroite
bande de cytoplasme cortical et le noyau, une distribution uniforme de vacuoles lipidiques et
de globules vitellins séparés par quelques travées cytoplasmiques. La zona radiata continue de
s'épaissir (25,6μm) par addition de lamelles concentriques. Le noyau central finit par être
repoussé vers la périphérie tandis que les globules vitellins d'une part et les gouttelettes
lipidiques d'autre part confluent en gouttes et masses de plus en plus grandes. Le stade 6
précède la ponte; ce stade d'accroissement rapide est caractérisé par l'aspect transparent que
prennent les ovules qui est dû à une absorption d'eau favorisant l'éclatement des follicules
(figure 27-A, B, C et D). Les ovocytes observés à ce stade ont un diamètre compris entre 480
et 725μm. Certains follicules matures subissent l’atrésie (figure27-E et F)
Reproduction Résul
Figure 22: Coupes histologique
maturation colorés à l’hématoxyline, A et B (pêché le mois de septembre), Cet D (poisson
pêché le mois de mai), LO
Hématoxyline éosine ((Ax100), (Bx100)), et le Trichrome à froid ((
A et B : Lot d’ovogonies, (poisson pêché en septembre 2010).
C et D : Lamelles ovigères, (poisson pêché en mai 2011).
A
C
Reproduction Résul
histologiques des ovaires de Trachurus mediterraneus
ation colorés à l’hématoxyline, A et B (pêché le mois de septembre), Cet D (poisson
pêché le mois de mai), LO : lamelles ovigères.
Hématoxyline éosine ((Ax100), (Bx100)), et le Trichrome à froid ((Cx100), (Dx100))
, (poisson pêché en septembre 2010).
: Lamelles ovigères, (poisson pêché en mai 2011).
B
LO
D
Reproduction Résultats
33
Trachurus mediterraneus en début de
ation colorés à l’hématoxyline, A et B (pêché le mois de septembre), Cet D (poisson
Cx100), (Dx100))
Reproduction Résul
Figure 23: Coupes histologique
A. (GX100)Ovocyte I en vitellogenèse endogène (
B. (GX100) Ovocyte I en vitellogenèse exogène (
C. (GX100) Ovocyte I en maturation finale (
N : Noyau, VL : Vacuoles lipidiques
N
A
C
294,4μm
Reproduction Résul
histologiques des ovaires matures colorés au trichrome à froid.
vitellogenèse endogène (pêché le mois de mai).
vitellogenèse exogène (pêché en mai 2011).
I en maturation finale (pêché en mai 2011).
VL : Vacuoles lipidiques, Th : Thèque
VL
B 281,6μm 217,6
371,2μm
Reproduction Résultats
34
des ovaires matures colorés au trichrome à froid.
217,6 μm
Reproduction Résul
Figure 24: Coupes histologique
A. et B. (Gx100) Ovocytes au début de la vitellogenèse.C. (Gx100) Ovocytes en vitellogenèse avancée.D. (Gx100) Ovocyte en maturation finale.E. (Gx400), enveloppes (granulo
A
C
G
E
Th
Reproduction Résul
histologiques des ovaires matures colorés au trichrome à froidmois de mai).
Ovocytes au début de la vitellogenèse.Ovocytes en vitellogenèse avancée.Ovocyte en maturation finale.nveloppes (granulosa zona radiata et thèques) d’un ovocyte I en vitellogenèse
B
D
ZR
PV
Reproduction Résultats
35
au trichrome à froid (pêché le
hèques) d’un ovocyte I en vitellogenèse.
Reproduction Résul
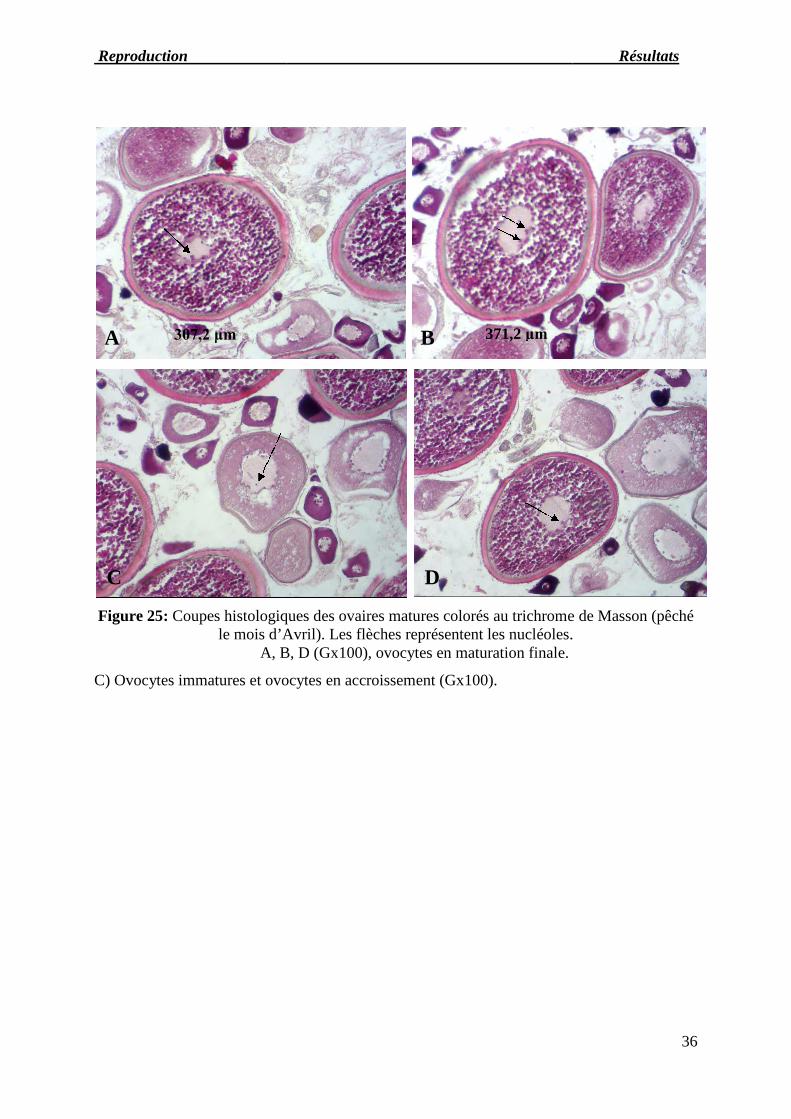
Figure 25: Coupes histologiquele mois d’Avril
A, B, D
C) Ovocytes immatures et ovocytes en accroissement (Gx100).
A
C
307,2 μm
Reproduction Résul
histologiques des ovaires matures colorés au trichrome de Massonmois d’Avril). Les flèches représentent les nucléoles.
A, B, D (Gx100), ovocytes en maturation finale.
C) Ovocytes immatures et ovocytes en accroissement (Gx100).
B
D
371,2 μm
Reproduction Résultats
36
au trichrome de Masson (pêchéLes flèches représentent les nucléoles.
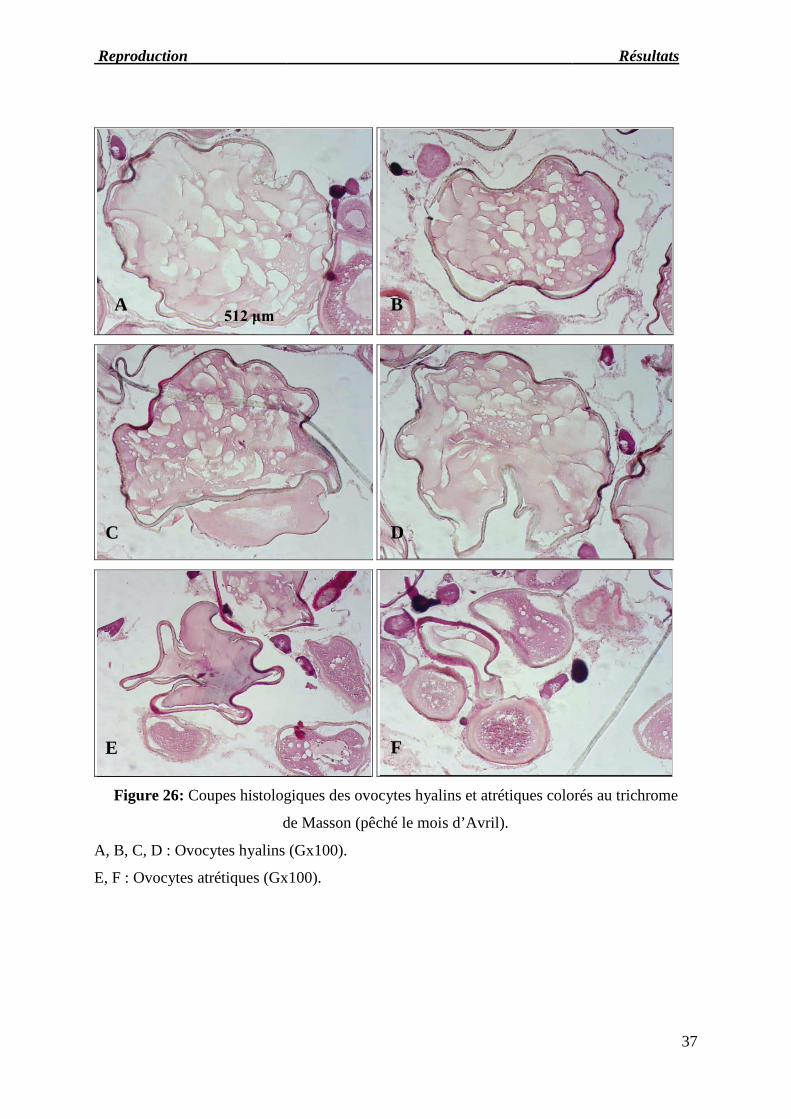
Reproduction Résul
Figure 26: Coupes histologique
de Masson
A, B, C, D : Ovocytes hyalins (Gx100).
E, F : Ovocytes atrétiques (Gx100).
A
C
E
512 μm
Reproduction Résul
histologiques des ovocytes hyalins et atrétiques colorés
de Masson (pêché le mois d’Avril).
: Ovocytes hyalins (Gx100).
: Ovocytes atrétiques (Gx100).
B
D
F
Reproduction Résultats
37
colorés au trichrome

Reproduction Résultats
38
1.9.2. Histologie des testicules
La description histologique des stades de la gamétogenèse est plus difficile à établir chez
les mâles que chez les femelles car les différents stades d'évolution des cellules reproductrices
sont présents presque toute l'année dans les testicules. Toutefois il est possible de dégager des
tendances en fonction des stades dominants.
Au mois de septembre nous observons des individus immatures (stade 1) ou au repos
surtout caractérisés par la présence de cellules germinales et de spermatogonies. Durant la
phase de repos, les cellules germinales constituent une couche entourant la lumière de chaque
lobule; durant la phase de maturation, elles sont isolées dans les tissus conjonctifs entourant
les lobules. L'activité mitotique entraîne la formation de cystes de spermatogonies (figure :
28-A), de plus en plus nombreux au fur et à mesure de la maturation des gonades.
Au mois d’octobre et novembre, les testicules renferment essentiellement des
spermatogonies et des spermatocytes (stade 2) de diamètre plus petits et également regroupés
par cystes (figure : 28-C), toutes les cellules d'un cyste étant aux mêmes stades. Les
spermatocytes résultent de la méiose subie par les spermatogonies; des changements
intracellulaires entraînant une nouvelle diminution du diamètre des cellules les amènent au
stade de spermatides.
Le mois de juillet correspond d’une part à la présence de tous les stades de la
spermatogenèse (stade 3), y compris, dans la lumière centrale des lobules, des
spermatozoïdes formés à partir des spermatides et qui sont libérés par éclatement des cystes.
En fin de maturation, au mois d’avril et mai (stade 4), les spermatogonies sont rares et les
spermatozoïdes ont envahi les canaux collecteurs (figure : 31). L'émission des gamètes
correspond au stade 5 ; les testicules contiennent presque exclusivement des spermatozoïdes
et des cellules germinales. Au mois de juillet, le stade 6, fin de maturation, les testicules
présentent quelques spermatozoïdes résiduels, les lobules sont vides bien distincts, séparés par
les cellules de Leydig et le tissu conjonctif se développe.
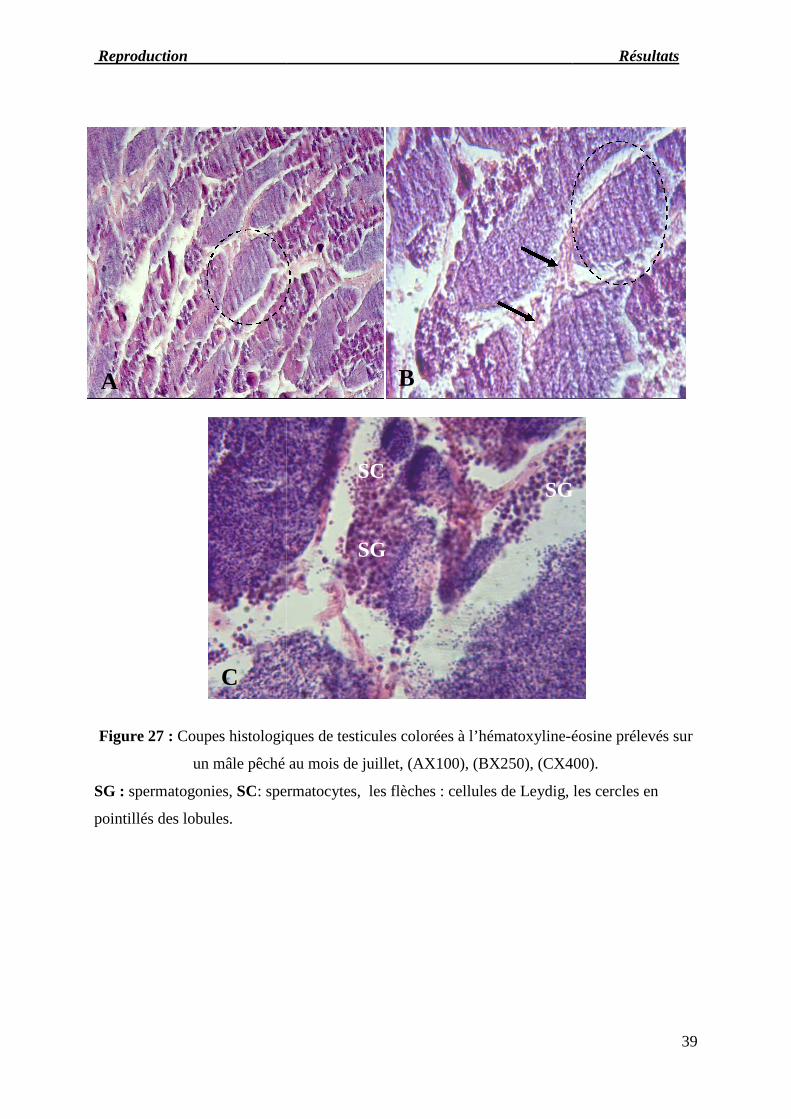
Reproduction Résul
Figure 27 : Coupes histologiques de testicules
un mâle pêché au mois de juillet, (AX100), (BX250),
SG : spermatogonies, SC: spermatocytes, les flèches
pointillés des lobules.
A
C
Reproduction Résul
Coupes histologiques de testicules colorées à l’hématoxyline-éosine
mâle pêché au mois de juillet, (AX100), (BX250), (CX400).
: spermatocytes, les flèches : cellules de Leydig, les cercles en
SG
SG
B
SC
Reproduction Résultats
39
éosine prélevés sur
(CX400).
: cellules de Leydig, les cercles en
SG
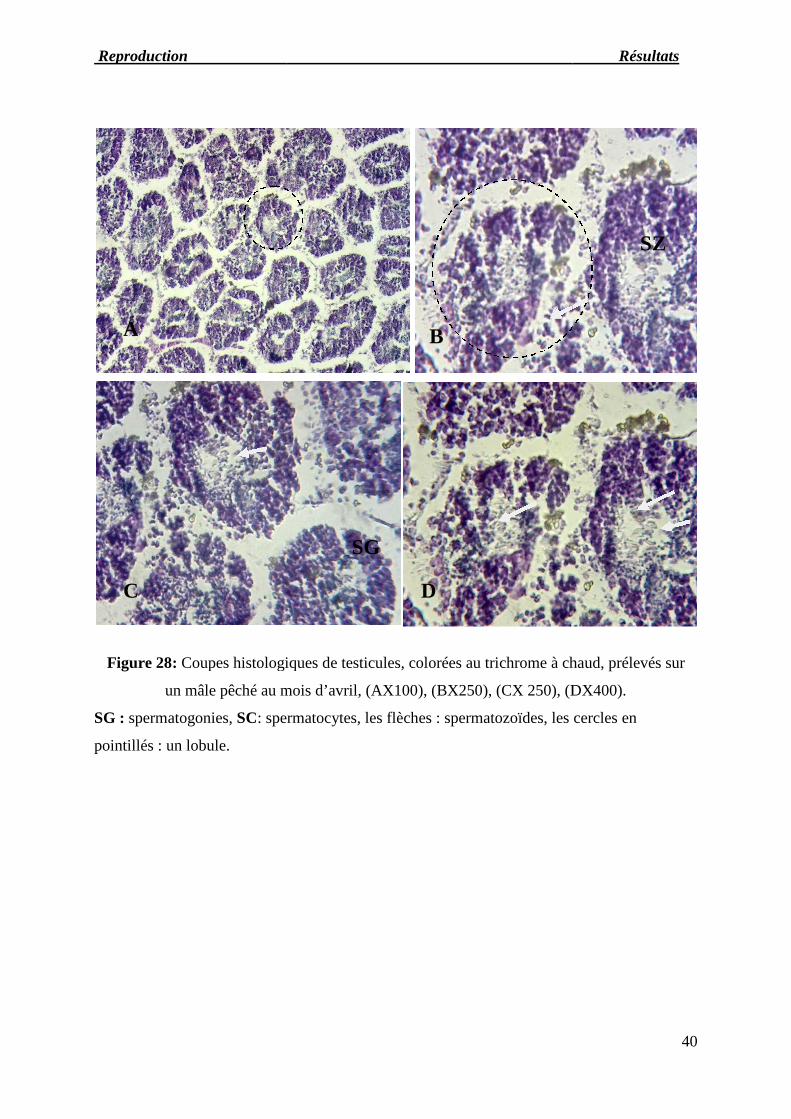
Reproduction Résul
Figure 28: Coupes histologiques de testicules
un mâle pêché au mois d’avril,
SG : spermatogonies, SC: spermatocytes, les flèches
pointillés : un lobule.
A
C
Reproduction Résul
Coupes histologiques de testicules, colorées au trichrome à chaud,
mâle pêché au mois d’avril, (AX100), (BX250), (CX 250), (DX400).
: spermatocytes, les flèches : spermatozoïdes, les cercles en
B
D
SG
Reproduction Résultats
40
, colorées au trichrome à chaud, prélevés sur
(AX100), (BX250), (CX 250), (DX400).
: spermatozoïdes, les cercles en
SZ
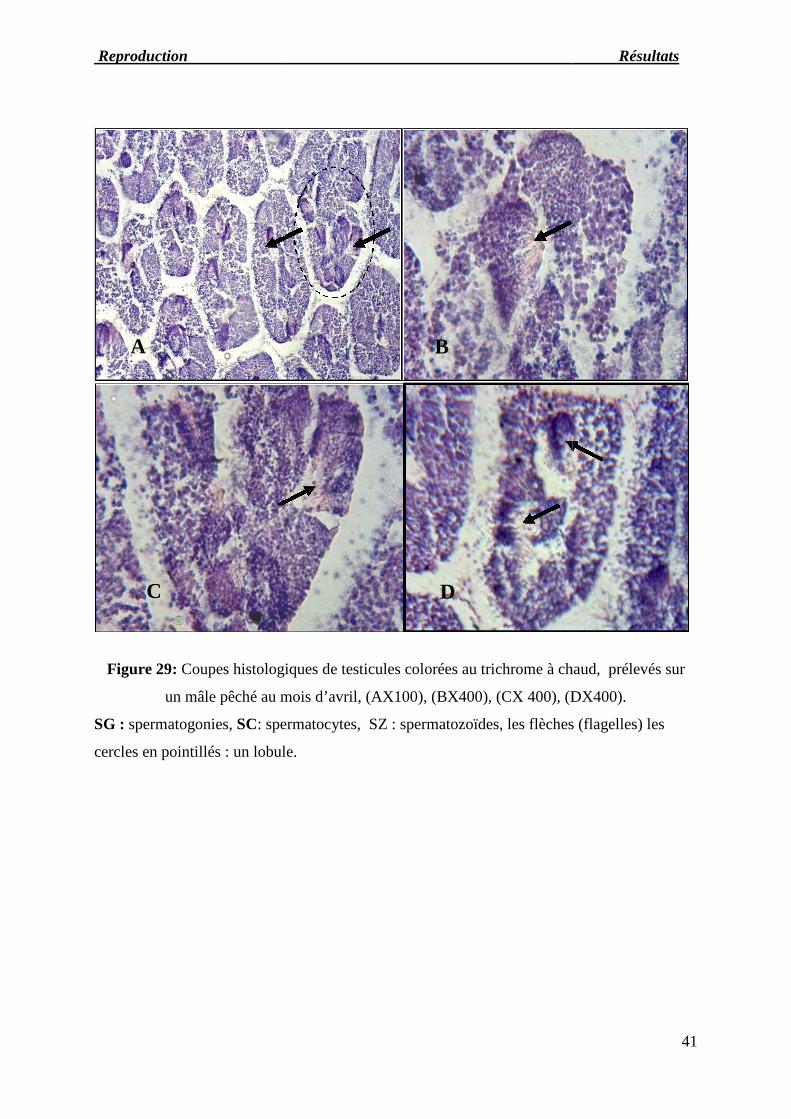
Reproduction Résul
Figure 29: Coupes histologiques de testicules
un mâle pêché au mois d’avril, (AX100), (BX400), (CX 400), (DX400).
SG : spermatogonies, SC: spermatocytes, SZ
cercles en pointillés : un lobule.
A
C
Reproduction Résul
Coupes histologiques de testicules colorées au trichrome à chaud,
mâle pêché au mois d’avril, (AX100), (BX400), (CX 400), (DX400).
: spermatocytes, SZ : spermatozoïdes, les flèches
: un lobule.
B
D
Reproduction Résultats
41
colorées au trichrome à chaud, prélevés sur
mâle pêché au mois d’avril, (AX100), (BX400), (CX 400), (DX400).
les flèches (flagelles) les
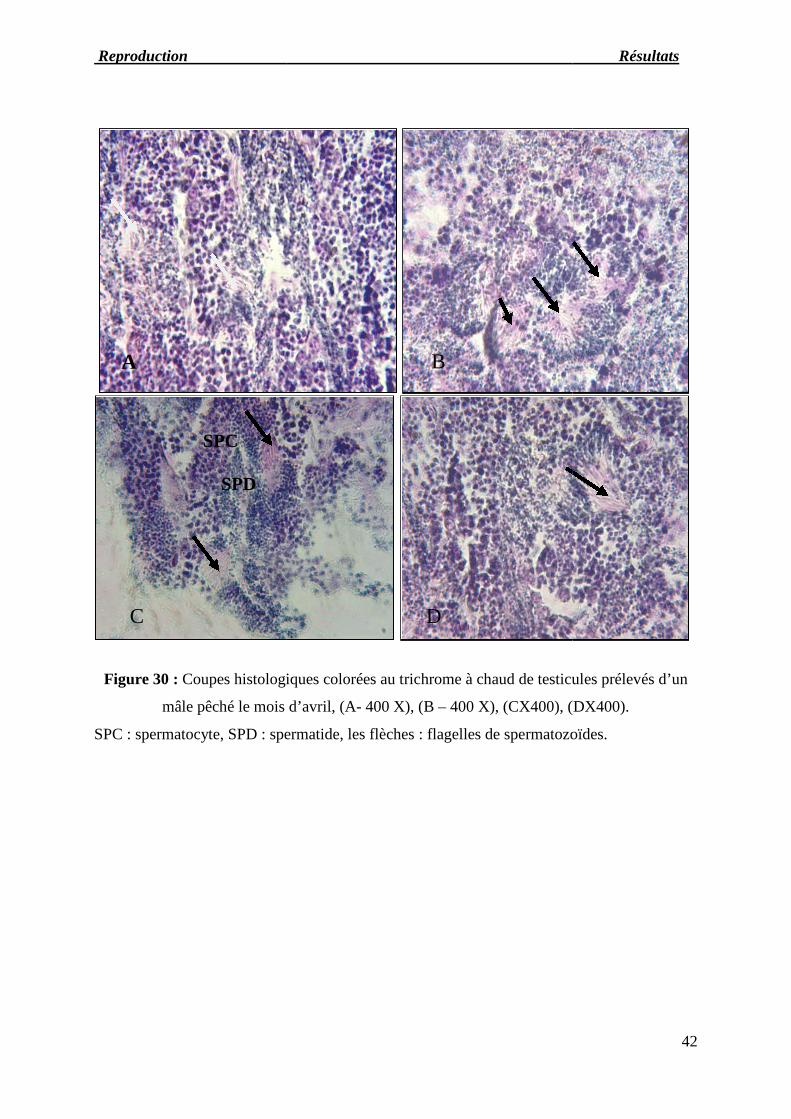
Reproduction Résul
Figure 30 : Coupes histologiques
mâle pêché le mois d’avril,
SPC : spermatocyte, SPD : spermatide,
SPD
SPC
A
C
Reproduction Résul
Coupes histologiques colorées au trichrome à chaud de testicules prélevés d’un
le mois d’avril, (A- 400 X), (B – 400 X), (CX400), (DX400)
SPD : spermatide, les flèches : flagelles de spermatozoïde
B
D
Reproduction Résultats
42
de testicules prélevés d’un
(DX400).
spermatozoïdes.
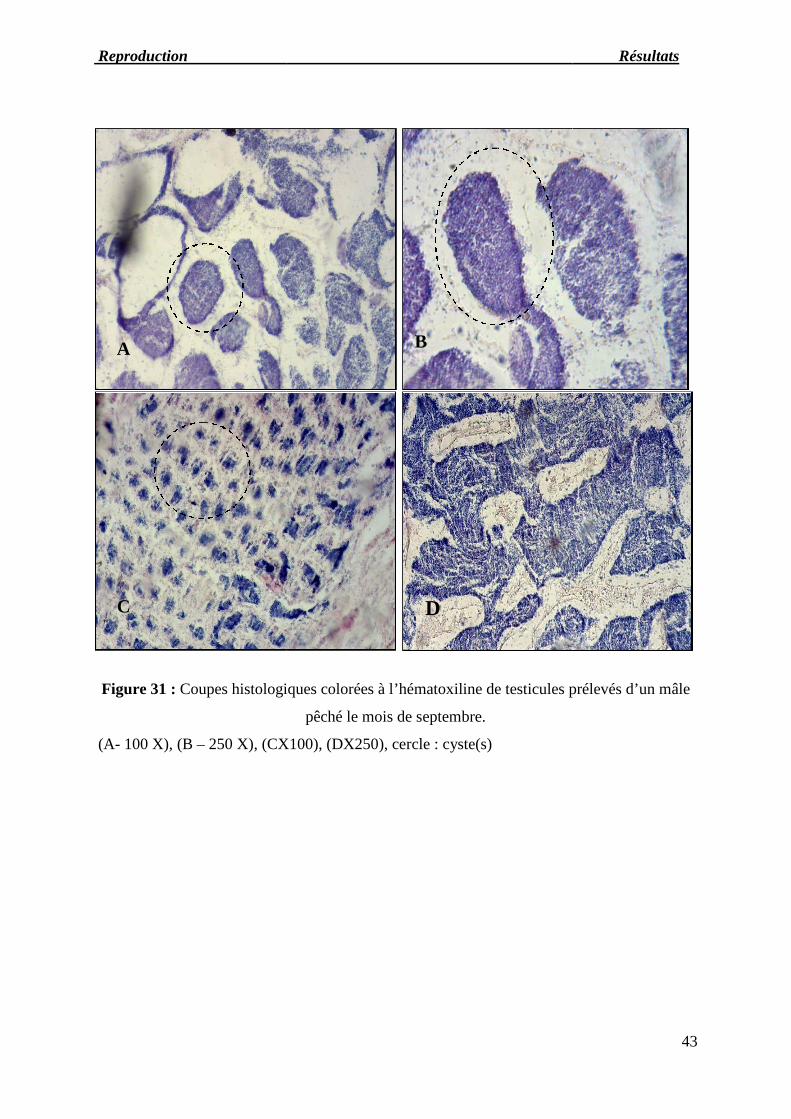
Reproduction Résul
Figure 31 : Coupes histologiques
(A- 100 X), (B – 250 X), (CX
A
C
Reproduction Résul
Coupes histologiques colorées à l’hématoxiline de testicules prélevés d’un mâle
pêché le mois de septembre.
0 X), (CX100), (DX250), cercle : cyste(s)
B
D
Reproduction Résultats
43
de testicules prélevés d’un mâle

Reproduction Discussion
44
IV.2. Discussion
2.1. Distribution des trois espèces
Durant notre échantillonnage annuel nous avons trouvé une grande proportion de
Trachurus trachurus (54,24%), un pourcentage de (45,39%) de Trachurus mediterraneus et
une très faible concentration de Trachurus picturatus (0,362%). Ces résultats contredisent
ceux de (Gherram, 2009) qui a travaillé dans la même zone d’étude et a échantillonné en
grande concentration les Trachurus mediterraneus 56%, une proportion de 33% des
Trachurus trachurus, et 11 % de Trachurus picturatus ;et divergent aussi de ceux de
(Dardignac, 1963) dans le golfe du Lion où le Trachurus mediterraneus, est l’espèce
dominante pour 80% de chinchards issus de chaque pêche.
Cette répartition est similaire à celle de (Korichi, 1988) dans la Baie de Bou Ismail où le
Trachurus trachurus est le plus fréquent.
Selon (Dardignac, 1963), l’aire de distribution de Trachurus mediterraneus est limitée par
l’isohaline 38%0 dans le golfe du Lion.
D’après (Viette et al., 1997) T.mediterraneus, T.trachurus, T. picturatus et T.ponticus
sont répartis simultanément dans la Méditerranée et la mer Noire ainsi que le long des côtes
de l'Atlantique de la Manche jusqu’au Maroc.
Dans la Baie d’Oran nous avons rencontré une minorité de Trachurus picturatus ce qui
rejoint la description de (Dardignac, 1963) sur cette espèce de haute mer qui ne fut rencontrée
qu’à des exceptions prés, sur les fonds supérieurs à 100 m.
2.2. Taille à la première reproductionLa taille de première maturité sexuelle est très variable chez les téléostéens, elle n’est pas
statistiquement différente en fonction du sexe. Toutefois, les mâles atteignent la maturité
sexuelle au même moment que les femelles.
Le Trachurus mediterraneus mûr le plus petit était de 10.4 cm, (Genç et al., 1998). Nous
avons trouvé dans notre étude une taille de 15cm pour les deux sexes cette valeur s’approche
de celle trouvée par (Soliman et al., 1982) dans la Baie d’Aboukir.
La taille minimale de première maturité calculée par (Arnéri, 1983) en adriatique est de
16cm cette valeur ne s’éloigne pas de celle trouvée par (Ben Salem, 1977) qui proposent des
tailles totales de 17 à 18cm dans les eaux tunisienne (Ben Salem et Ktari, 1981). En revanche

Reproduction Discussion
45
notre résultat ne se concorde pas avec celui de (Bini, 1968) et (Fisher et al., 1987), qui
suggèrent que la première reproduction intervient à 23cm et un âge de 3 et 4 ans
respectivement.
2.3. Répartition des tailles et calcul de la taille moyenneSelon (Korichi, 1988), les tailles moyennes du Trachurus mediterraneus pêché dans la baie
de Bou Ismail est de (13.87, 12,96) pour les femelles et les mâles. En revanche les tailles
moyennes estimées dans nos échantillons sont supérieures (20,16 et 20,42cm) femelles et
mâles respéctivement.
Suivant (Korichi, 1988) les tailles moyennes de Trachurus trachurus sont supérieures à
celles de Trachurus mediterraneus, plus côtier. Au contraire des Trachurus de la baie d’Oran
dont les tailles moyennes de notre espèce sont supérieures à celle trouvée par (Gherram, 2009)
sur le Trachurus trachurus de la même région.
D’après (Korichi, 1988), à 20 m les Trachurus mediterraneus sont dominants alors qu’à
70m ce sont les Trachurus trachurus qui sont dominants.
2.4. Sexe ratioLa sexe ratio des téléostéens a fait l’objet de nombreux travaux chez diverses espèces de
poissons de différentes familles. Chez le Trachurus mediterraneus sa valeur de 0.72, montre
qu’il y’a une dominance d’un sexe par rapport à l’autre; globalement les pourcentages sont
comme suit : 58% femelles 42% mâles, nous déduisons que les femelles sont observées plus
fréquemment que les mâles dans toutes les classes de taille.
En revanche le sex-ratio est légèrement en faveur des mâles dans les eaux
britanniques (Kerstan, 1985 in Korichi, 1988).
Selon (Aka et al., 2004), la variation de la sex- ratio dépend de l’état physiologique des
poissons. En général, chez les téléostéens, les mâles sont prédominants en période de
reproduction, dans notre région nous remarquons que le sex-ratio s’approche de 1 au mois
d’avril, alors qu’en période de repos sexuel, ce sont les femelles qui sont prédominantes
(Paugy, 1980 ; Santos et al., 2007).
Plusieurs facteurs tels que la migration pour la recherche de nourriture, la croissance
différentielle et le taux de mortalité par sexe influencent également le sex–ratio chez les
poissons (Mellinger, 2002).
Nos résultats sont similaires à ceux de (Şahin et al., 2008) dans la partie EST de la mer
noire dont les femelles représentent 68% de la population et les mâles 31%.

Reproduction Discussion
46
2.5. Période de ponte
Dans la présente étude, le suivi de la variation du RGS femelle indique un maximum de
(3,39 ± 1,16) au mois de juin, et celle des mâles (3,544± 1,058) au mois de mai, donc la
période de ponte s’étend d’avril à juin chez le Trachurus mediterraneus, ce dernier se
reproduit une seule fois par ans avec une forte activité de reproduction pendant la période
(printemps-été).
D’après (Şahin et al., 2008) dans la mer noire, le RGS et les valeurs de l’indice de
condition indiquent que la période de frai a commencé au début de Juillet et s'est poursuivie
jusqu'au début Septembre, la femelle a atteint une valeur de RGS 2,368 ±0.890 en août et
0.987 ± 0.326. Les périodes de frai ont été très proches les unes des autres.
Dans d'autres études réalisées dans la même région la période de frai s’étale du 15 Mai au
15 août (Slastenenko, 1956; Ivanov et Beverton, 1985) et de Juin à Septembre (Genç et al.,
1998).
Le maximum des valeurs de RGS des femelles ont été 4,90 en Juillet, 3,29 en août, et 1,04
en Septembre (Genç et al., 1998) dans la mer noire.
Selon Viette, (1997) dans le golfe de Trieste, la période de reproduction s’étend de mai à
aout, qui rejoint celle de (Demir, 1961;Cautis, 1979; Arneri and Tangerini, 1983; Arneri,
1984; Fisher et al., 1987).
Le RGS atteint des valeurs plus élevées chez les mâles que chez les femelles, ce qui
confirme les résultats cités précédemment et contredit ce qui est considéré comme la norme
(Macer, 1974; Treasurer et Holliday, 1981).
La période de reproduction des mâles s’étale d’avril à juin comme chez les femelles avec
un seul pic en fin de mai, nous pouvons dire qu’il y a une synchronisation de maturation
gonadique entre les deux sexes et donc une concordance de la période de ponte entre
les femelles et les mâles de Trachurus mediterraneus.
Il est maintenant largement admis chez les poissons téléostéens que les réserves vitellines
élaborées par les hépatocytes sont accumulées dans les ovocytes pendant la vitellogenèse
(Thiaw, 1993 ; Diaye, 1995 ; Wallace & Selman, 1985). Dans ces conditions, la baisse du
RHS d’avril à juin expliquerait la croissance du RGS dans la période correspondante. La
croissance rapide du RGS en un mois s'expliquant par une entrée massive d'eau dans l'ovaire à
la fin de la maturation. S’il ya une croissance du RHS à cette période elle est dû à la synthèse
de réserve vitelline ce qui est contradictoire, chez les espèces qui ont un cycle de reproduction
comme c’est le cas de notre espèce. Les valeurs mensuelles du coefficient de condition Kc

Reproduction Discussion
47
ne présentent pas de variations remarquables d’un mois à l’autre chez les mâles et les
femelles.
2.6. Stades de maturité
Un pourcentage élevé du stade I, s’observe pour les mois de septembre, octobre et
novembre 2010 avec 100%, ce pourcentage diminue encore plus au mois de décembre
2010 à 20% et disparaît brutalement au mois d’avril 2011.
Le satde II est observé jusqu’au cours du mois d’avril ainsi que le stade III le Trachurus
mediterraneus est en phase de préparation à la ponte qui vient par la suite au mois de mai et
juin où nous remarquons l’apparition des stades IV et V, le stade VI apparait pendant toute
l’année sauf au mois de juin et il représente 100% de la population en post ponte.
Ces résultats ne concordent pas avec ceux de (Mellinger, 2002) in (Sylla et al., 2009) où nous
remarquons une hétérogénéité de distribution des diamètres ovocytaires indiquent un
recrutement continu d’ovocytes vitellogéniques, qui ne sont pas tous émis en même temps. La
reproduction est donc asynchrone, fractionnée avec une émission des ovocytes mûrs en petites
quantités durant un long intervalle de temps suivies d’une récupération très rapide de l’ovaire.
Cette reproduction asynchrone, typique des reproducteurs multiples a été rapportée chez
d’autres espèces de Carangidés, Alectis indicus et Caranx ignobilis (Von Westernhagen,
1974), Trachurus trachurus (Karlou-Riga et Economidis, 1997), Parastromateus niger
(Dadzie & al., 2008).
2.7. Fécondité
La fécondité absolue présente une grande variation entre les femelles, elle varie de 2063 à
18854ovocytes pour des femelles de longueur à la fourche comprise entre 151 et 183 mm.
Elle augmente avec la taille des femelles.
En général, la fécondité est très élevée chez les Carangidés. (Dadzie & al., 2008).et
(Dadzie et al., 2008) ont rapporté chez Parastromateus niger des fécondités comprises entre
71 305 et 3 895 449 ovocytes pour des femelles de taille (LS) comprise entre 398 mm et 490
mm. Ces auteurs attribuent cette forte fécondité à une stratégie visant à maximiser la survie de
la progéniture chez les espèces qui ne pratiquent pas la protection parentale comme c’est le
cas chez Trachinotus teraia. La relation taille-fécondité des femelles de poissons n’est pas
absolument générale, surtout chez les téléostéens, où la fécondité peut plafonner à un âge
avancé (Mellinger, 2002). En milieu naturel, la température de l’eau, la disponibilité de proies
influenceraient le processus de maturation des ovocytes, par conséquent, la fécondité des
poissons (Pörtner et al., 2001).

Reproduction Discussion
48
La fécondité relative est liée à la taille du poisson, il existe une relation linéaire (figure 21),
nous observons que les poissons de petites tailles pondent plus d’œufs que les poissons de
grandes tailles 20 et 21cm, cette diminution est dû à une incapacité du poisson à produire de
grandes quantités d’œufs, nous pouvons expliquer cette faiblesse par un vieillissement du
poisson qui n’est plus capable de se reproduire.
2.8. Conclusion sur la reproduction
Une seule période de ponte a été mise en évidence chez Trachurus mediterraneus
de la baie d’Oran, elle s'étend d’avril à juillet. Les femelles de cette espèce
comportent une bonne proportion d'individus matures pendant la saison estivale.
Les tailles et poids de première maturité sexuelle sont différents selon les
sexes et les saisons de reproduction. La maturité intervient plus tôt chez les
mâles.

Croissance Etude bibliographique
49
V. Croissance
1. Introduction : Estimation de l’âge
L’âge des téléostéens est d’une façon générale appréhendé par trois types de méthodes
classiques : la scalimétrie, l’otolithométrie et l’ostéochronologie. Les pièces calcifiées les plus
utilisées pour l’estimation de l’âge sont les écailles, les otolithes et divers éléments du
squelette (Meunier, 1988; Loubens et Panfili, 1992). Le genre Trachurus est dépourvu
d’écailles c’est pourquoi beaucoup d’auteurs ont utilisé les otolithes pour déterminer l’âge.
2. Définition des otolithes
Les otolithes sont des concrétions calcaires situées dans l’oreille interne des poissons
utilisées pour la spatialisation ; ils résultent d’un processus d’accrétion permanant fortement
influencé par les conditions environnementales. Elles sont lues en routine pour les estimations
d’âge et de croissance, la gestion des stocks ou pour divers problèmes écologiques (Anatole
al., 2008).
3. Description et fonction de l’oreille interne
L’oreille interne qui existe chez tous les vertébrés gnathostomes, fonctionne à la fois
comme un système auditif qui détecte les ondes sonores et comme un système vestibulaire qui
détecte les accélérations linéaires et angulaires, permettant aux organismes de maintenir leur
équilibre.
Chez les poissons l’oreille interne est une structure paire noyée dans le crâne, à chaque côté
de la tête, prés de l’encéphale moyen. Chaque oreille est un ensemble complexe de canaux et
de sacs remplis d’endolymphe, un liquide visqueux, l’anatomie de ces labyrinthes et la
structure de mécanorécepteurs labyrinthiques sont connues chez de nombreuses espèces de
poissons (Louverstein, 1971).
Les téléostéens ont trois canaux semi circulaires en position orthogonale les uns par rapport
aux autres ce qui permet la détection des accélérations angulaires. Les canaux s’ouvrent dans
une série de grandes chambres d’interconnexion ou sacs optiques qui contiennent un tissu
sensoriel, la macula qui détecte les accélérations linéaires et les sons.
4. Description des otolithes
Les otolithes des trois paires de sacs otiques différents en taille et en forme. Les différences
morphologiques des otolithes tendent à refléter leur phylogénie et leur développement, bien
qu’il y’ait une variation considérable inter et intra- spécifique de la forme apparaissant être
dues à des influences génétiques et environnementales (Lombarte et Lleonart, 1993 ; Nolf,

Croissance Etude bibliographique
50
1995 ; Torres et al., 2000). Chez les ostéichthyens, il y’a trois otolithes par oreille, la sagitta,
le lapillus et l’asteriscus, renfermés dans trois sacs otiques (Ostacs) remplis d’un liquide aux
propriétés spéciales, l’endolymphe. Les otolithes des trois paires diffèrent en taille et en forme
pour chaque espèce mais sont symétriques (droite-gauche), excepté les poissons plats. Chez la
plupart des espèces, la sagitta est le plus grand des otolithes et c’est donc le plus utilisé dans
les études de sclérochronologie (Wright et al., 2001).
4.1. Sagitta
C’est l’une des trois paires trouvées dans le labyrinthe membraneux des poissons osseux.
Elle se trouve en dessous du succulus (petit sac) des paires inférieures. Elle est habituellement
compressée latéralement et de forme elliptique. Cependant la forme de sagitta varie
considérablement selon les espèces. La sagitta est plus large que l’asteriscus et le lapillus, La
sagitta est l’otolithe la plus fréquemment utilisée pour l’otolithométrie.
4.2. Asteriscus
L’une des trois otolithes trouvées dans le labyrinthe membraneux des poissons osseux, elle
se trouve en dessous du lagena « flask » des paires inférieures chez les poissons non
ostariophysians. L’asteriscus est petite et formée comme un hémisphère aplati ou un quart de
lune. En ostariophysi asteriscus est rudement circulaire et latéralement compressée et
considérablement large que la sagitta.
4.3. Lapilli
L’une des trois otolithes trouvées dans le labyrinthe membraneux des poissons osseux. La
majorité de la partie dorsale de l’otolithe se trouve en dessous de l’utriculus petite poche) des
paires supérieures. Dans la plupart des poissons, les lapilli sont sous forme d’une sphère
aplatie aux pôles et elle est plus petite que la sagitta.
4.4. Description des otolithes du chinchard à queue jaune
L’espèce étudiée se caractérise par ses otolithes robustes et épaisses, pourvues d’un suculus
assez large leur area dorsale est un peu plus étendue que chez la plupart des espèces du genre
Trachurus, elles présentent des annuli (un annulus est formé d’une zone étroite et hyaline qui
correspond à une croissance lente et une zone large et opaque qui correspond à une croissance
rapide).
5. Composition chimique des otolithes
Trente éléments chimiques ont été détectés au total dans les otolithes d’un large éventail
d’espèces, mais à des concentrations très variables (Campana, 1999). Les otolithes sont
composées à plus de 99% de CaCO3, et quatre grandes familles d’éléments se distinguent en

Croissance Etude bibliographique
51
fonction de leur concentration : les éléments majeurs (Ca, C, O), les éléments mineurs ayant
des concentrations supérieures à 100 ppm (Na, Sr, K, S, N, Cl, P), les éléments de traces ayant
des concentrations inférieures à 100 ppm (Mg, Si, Zn, B, Fe, Hg, Mn, Ba, Cu, Al, Br., Li ), et
enfin les éléments ultra-traces ayant des concentrations inférieures à 1ppm (Pb, As…).
6. Intérêt des otolithes
Les otolithes se sont avérées utiles en taxonomie (Hecht, 1979), permettant par exemple,
l’étude de la ration alimentaire à partir des contenus stomacaux partiellement digérés (Suter et
Morel, 1996 ; Olsson et North, 1997 ; Watanabe et Saito, 1998 ; Alonso et al., 1999).
De même, les otolithes, issues de sites archéologiques et paléontologiques permettent de
reconstruire les paléo-environnements et les paléo-faunes (Nolf, 1995).
La morphométrie des otolithes a aussi été utilisé pour l’identification et l’étude des
variations géographiques des populations et des stocks de poissons (Messieh et al., 1989 ;
Castonguay et al., 1991 ; Campana et Casselman, 1993 ; Friedland et Reddin, 1994).

Croissance Matériel et méthodes
52
VII. Matériel et méthodes
L’étude de la croissance a été réalisée à partir de la lecture de 705 paires d’otolithes
prélevées au fil des échantillonnages de telle sorte que toutes les tailles des captures ont été
représentées. Après leur extraction les paires d’otolithes rincées à l’eau du robinet ont été
placées dans des sachets répertoriés. Ensuite les otolithes sont pesés à l’aide d’une balance de
précision puis mesuré à l’aide d’une règle graduée de l’oculaire du microscope utilisé.
La lecture de l’âge du Chinchard à queue jaune a porté sur l’observation sous microscope
stéréoscopique en lumière réfléchie (WILD Type 327616) de stries de croissance de paires de
sagitta entières imprégnées dans l’eau distillée.
En vue de réaliser la meilleure estimation possible de l’âge et réduire les éventuelles erreurs
d’analyse d’otolithes, trois lectures ont été effectuées par trois observateurs différents, l’âge
adopté est celui qui présente une meilleure cordonance entre les différentes lectures.
La lecture se fait sous microscope stéréoscopique en lumière réfléchie, l’otolithe posée sur
fond noir est immergées dans du méthanol 70%, tous les anneaux observés sont comptés ; on
considère qu’il n’ y’a pas de « faux anneaux » chez le chinchard, bien que l’interprétation du
1er anneau reste discutable. Cette méthode présente cependant des inconvénients, notamment
son coût et la toxicité de l’alcool utilisé, c’est pourquoi la lecture est faite sur des otolithes
immergée dans de l’eau distillée. Les deux techniques ont donné le même nombre d’anneaux
pour les paires lues.
1. La Croissance
Elle est considérée comme une résultante des actions simultanées de facteurs anaboliques
proportionnels à la surface et des facteurs cataboliques proportionnels au volume du corps
(Bouaziz., 1992).
Les paramètres de la croissance sont :
• La longueur asymptotique L∞ : C’est la longueur moyenne qu’atteindrait un poisson qui
vivrait indéfiniment
• L’âge au temps t0 : C’est l’âge théorique qu’aurait un poisson de longueur nulle s’il avait
grandit de tout temps selon l’équation en question.
Le plus souvent, t0 prend des valeurs négatives car les jeunes poissons grandissant plus
rapidement que prédit par l’équation de Von Bertallanfy. Dans notre cas, nous avons
emprunté le modèle empirique de (Pauly, 1980).
En effet, il a été démontré que le paramètre K représentant le coefficient de
catabolisme dans la courbe de croissance de (Von Bertalanffy, 1938) est étroitement
lié avec la longévité du poisson (Beverton et Holt, 1959), cette loi, dérivée en fait des

Croissance Matériel et méthodes
53
données de croissance en liaison avec la courbe de croissance de Von Bertalanffy est
plutôt applicable aux poissons de petite et moyenne taille (Pauly 1980) .
1.1. Paramètres de croissance
Le modèle retenu pour décrire la croissance linéaire est celui de Von Bertalanffy, (1938),
son expression est: Lt = L∞ [1– exp (–K(t–to)]
Les paramètres du modèle de croissance absolue de l’équation de Von Bertalanffy :
où L∞ est la longueur maximale théorique, K est le taux avec lequel l’animal évolue à L∞,
et to le temps de recrutement à la population ont été estimés à partir de l’analyse de la
distribution des fréquences de taille en utilisant Excel.
1.2. Les relations longueurs-poidsPour comprendre la relation entre le poids du poisson et sa taille, nous avons établi une
relation du type y= axb où y est le poids P (pour Poids) et x la taille LT. Une relation
d’allométrie du type P= a Lb lie le poids d’un poisson à sa longueur totale ; dans cette
équation :
P : est le poids du poisson
L : est sa longueur
a : est une constante
b : est le coefficient d’allométrie (coefficient de croissance relative entre le poids et la
longueur). Sa valeur est voisine de trois car le poids est assimilé à un volume et il est élevé à
la puissance trois puisque le volume est le cube de trois longueurs. Quand b = 3 il y a
isométrie de croissance (et la densité spécifique de l’animal ne change pas). Si b>3,
l’allométrie est majorante, le poisson grossit plus vite qu’il ne grandit. Si b<3, l’allométrie est
minorante, le poisson grandit plus vite qu’il ne grossit.
1.3. Analyse statistique
Pour l’étude statistique des paramètres de croissance et de reproduction, nous avons utilisé
l’Excel et STATISTICA (version 6) à l’aide de ce dernier nous avons fait la courbe sigmoïde
et les tests Non-paramétrique.

Croissance Discussion
54
VII. Résultats et Discussion
1. Résultats
1.1 Les longueurs
1.1.1 Distribution des fréquences de longueurs
Nous présentons ci-dessous, les distributions de fréquences de taille pour les femelles, les
mâles et les individus à sexes indéterminés. Formés par les sommets des histogrammes, nous
observons des modes en forme de cloche caractéristique de la loi normale.
Chez les mâles, nous remarquons en juillet, un seul mode à 17cm ; en septembre nous
observons deux modes, le premier à 17cm et le second plus important à 18 et19cm ; en
octobre la répartition est bimodale 18 et19cm dominent, au mois de novembre la répartition
est monomodale la classe 21cm domine.
Le mois de décembre est particulier, nous n’avons rencontré qu’un seul individu de taille
29cm. Au mois d’avril la répartition est plurimodale, nous avons observé la dominance de la
classe 17cm, plus loin 20 et21cm. Au mois de mai nous remarquons que la classe 18cm
domine puis 19et 20cm. En juin la répartition est plurimodale, les classes dominantes par
ordre sont : 18,19cm âgés de I à II ans, 20cm et 17cm.
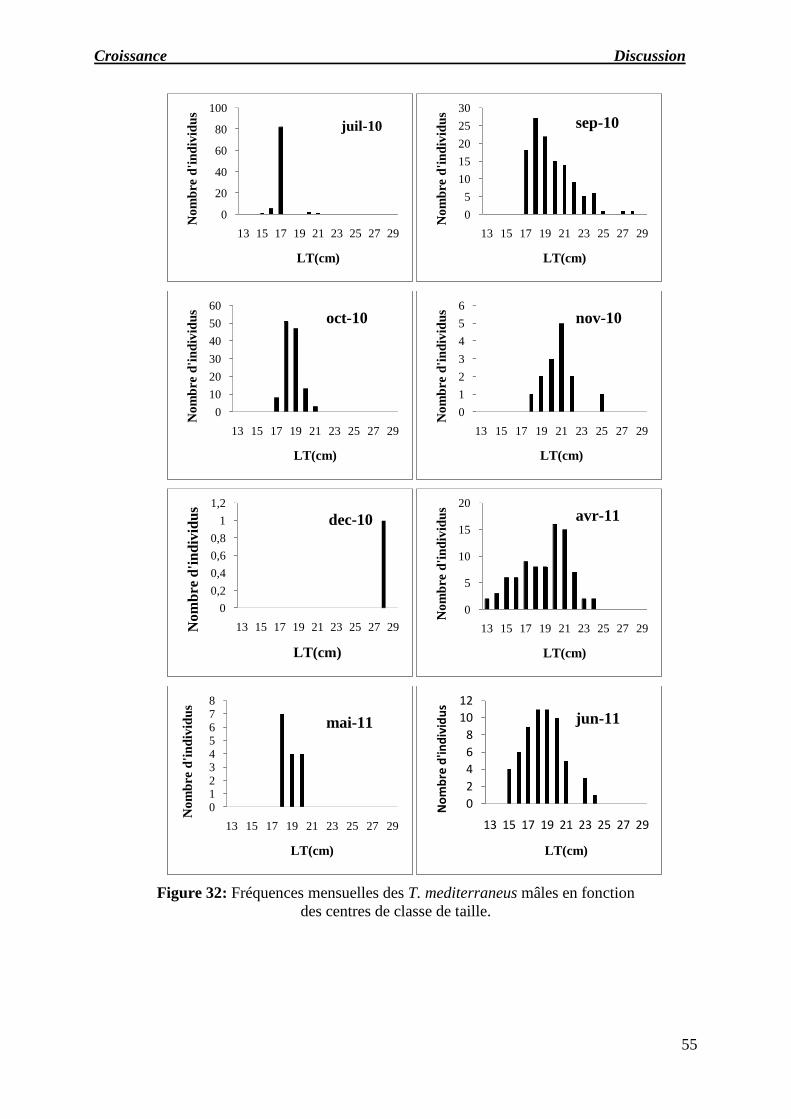
Croissance Discussion
55
Figure 32: Fréquences mensuelles des T. mediterraneus mâles en fonctiondes centres de classe de taille.
0
20
40
60
80
100
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
juil-10
0
5
10
15
20
25
30
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
sep-10
0
10
20
30
40
50
60
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
oct-10
0
1
2
3
4
5
6
13 15 17 19 21 23 25 27 29N
om
bre
d'i
nd
ivid
us
LT(cm)
nov-10
0
0,2
0,4
0,6
0,8
1
1,2
13 15 17 19 21 23 25 27 29No
mb
red
'in
div
idu
s
LT(cm)
dec-10
0
5
10
15
20
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
avr-11
012345678
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
mai-11
0
2
4
6
8
10
12
13 15 17 19 21 23 25 27 29
No
mb
red
'ind
ivid
us
LT(cm)
jun-11

Croissance Discussion
56
Chez les femelles du mois de juillet nous soulignons la dominance de la classe 17cm. Au
mois de septembre la répartition est bimodale, la classe 20cm suivie par 17cm. En octobre ce
sont les classes 18 et 19cm qui dominent, en novembre les classes 19,20 et 21cm âgés de I à
IV ans prédominent. Le mois de décembre est particulier, nous notons la présence de 3
individus 25, 26 et 29cm. Au mois d’avril nous observons un seul mode la classe 20cm qui
domine. En mai la répartition est plurimodale 15/16cm puis 17/18 cm ,au mois de juin nous
notons deux modes 20cm et 18/19cm.
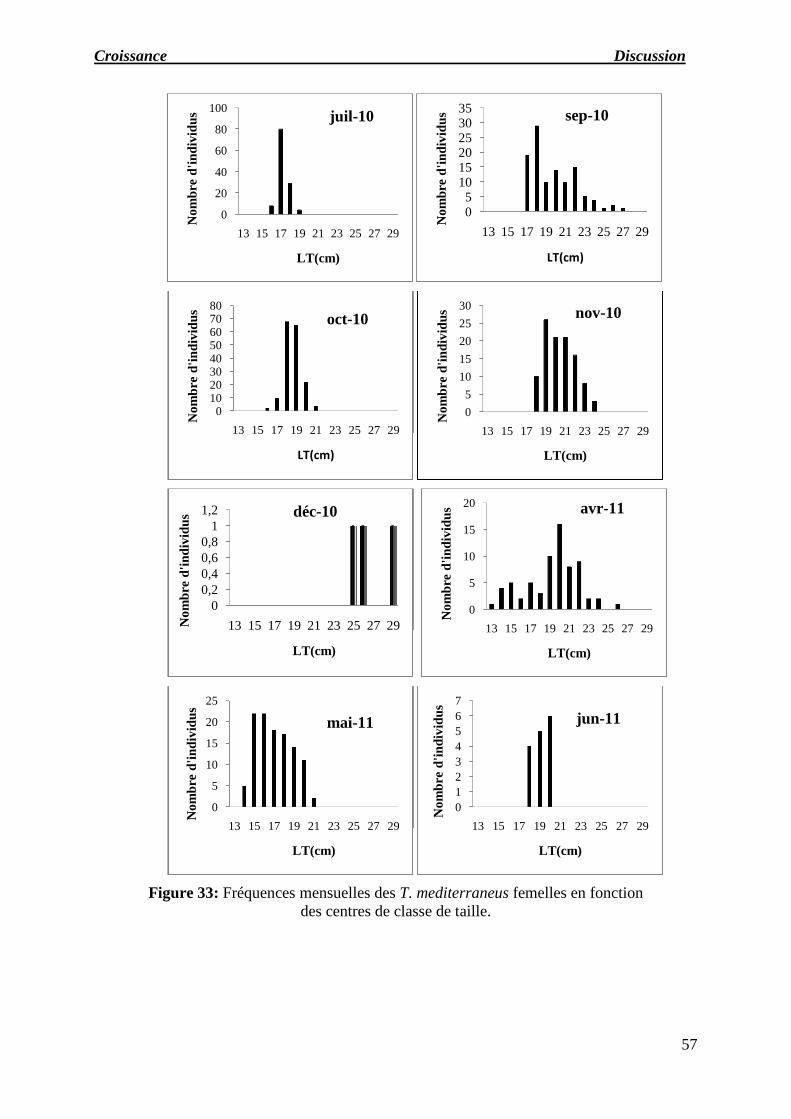
Croissance Discussion
57
Figure 33: Fréquences mensuelles des T. mediterraneus femelles en fonctiondes centres de classe de taille.
0
20
40
60
80
100
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
juil-10
05
101520253035
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
sep-10
01020304050607080
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
oct-10
0
5
10
15
20
25
30
13 15 17 19 21 23 25 27 29N
om
bre
d'i
nd
ivid
us
LT(cm)
nov-10
00,20,40,60,8
11,2
13 15 17 19 21 23 25 27 29No
mb
red
'in
div
idu
s
LT(cm)
déc-10
0
5
10
15
20
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
avr-11
0
5
10
15
20
25
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
mai-11
0
1
2
3
4
5
6
7
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
jun-11
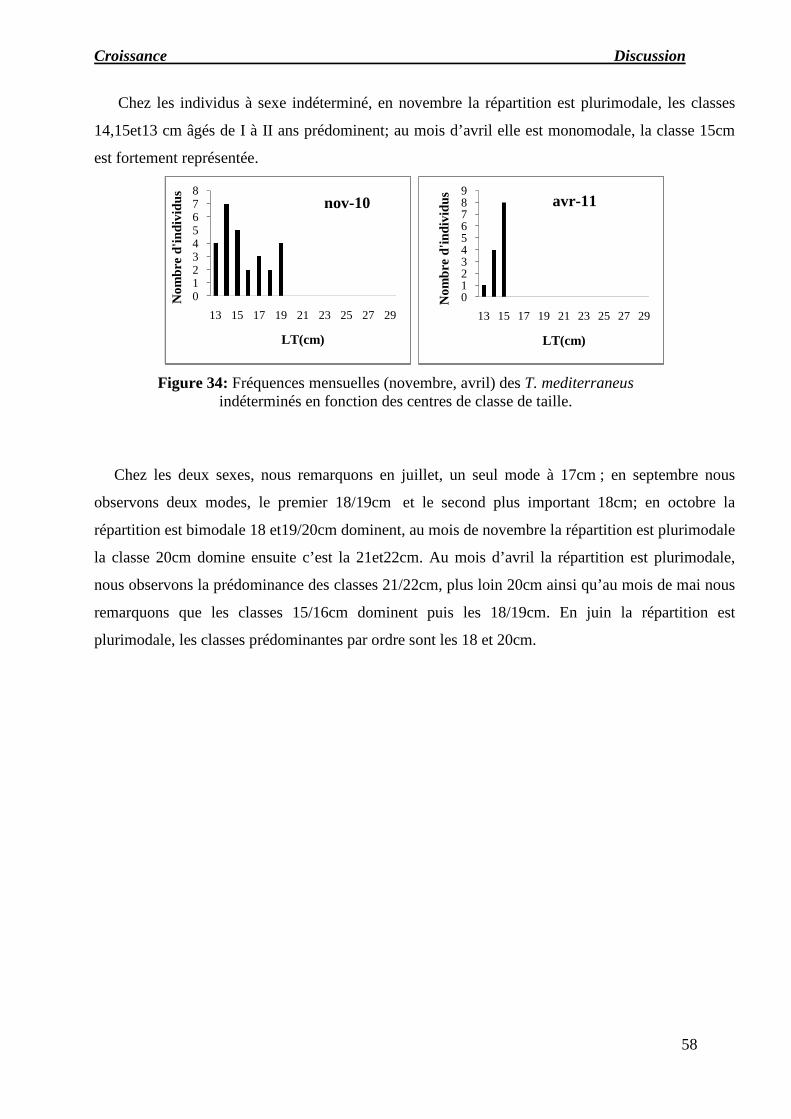
Croissance Discussion
58
Chez les individus à sexe indéterminé, en novembre la répartition est plurimodale, les classes
14,15et13 cm âgés de I à II ans prédominent; au mois d’avril elle est monomodale, la classe 15cm
est fortement représentée.
Figure 34: Fréquences mensuelles (novembre, avril) des T. mediterraneusindéterminés en fonction des centres de classe de taille.
Chez les deux sexes, nous remarquons en juillet, un seul mode à 17cm ; en septembre nous
observons deux modes, le premier 18/19cm et le second plus important 18cm; en octobre la
répartition est bimodale 18 et19/20cm dominent, au mois de novembre la répartition est plurimodale
la classe 20cm domine ensuite c’est la 21et22cm. Au mois d’avril la répartition est plurimodale,
nous observons la prédominance des classes 21/22cm, plus loin 20cm ainsi qu’au mois de mai nous
remarquons que les classes 15/16cm dominent puis les 18/19cm. En juin la répartition est
plurimodale, les classes prédominantes par ordre sont les 18 et 20cm.
012345678
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
0123456789
13 15 17 19 21 23 25 27 29
No
mb
red
'in
div
idu
s
LT(cm)
avr-11nov-10
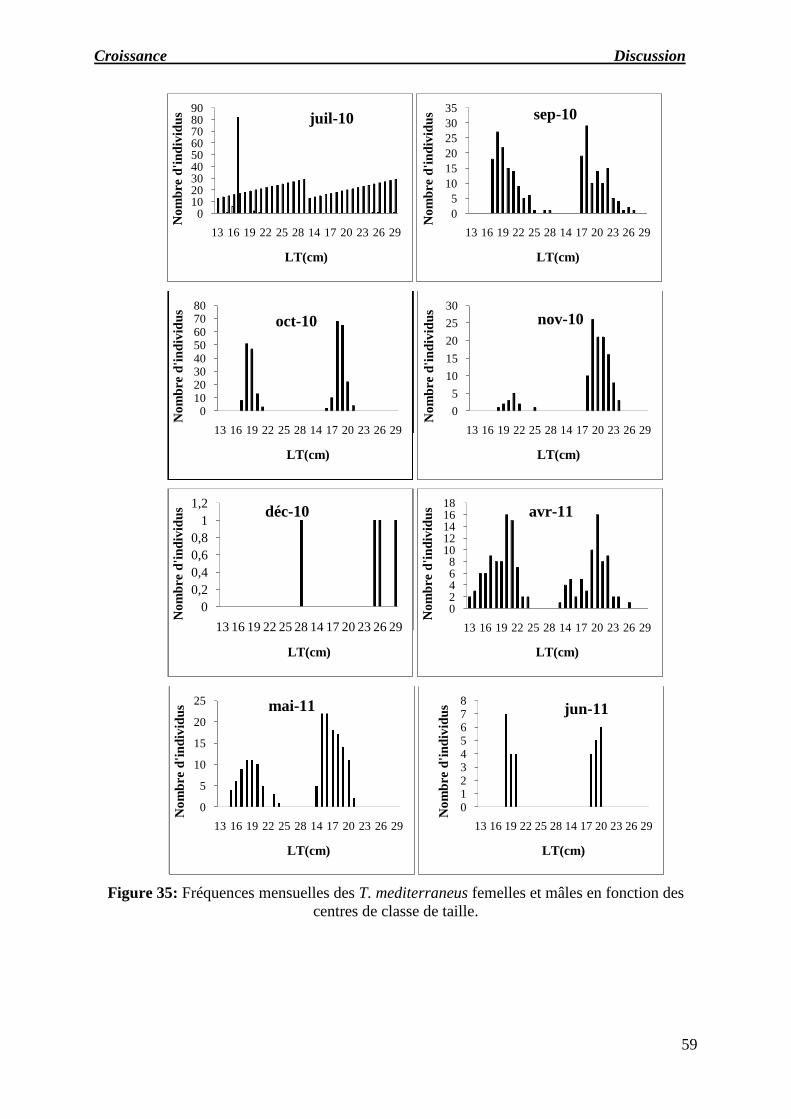
Croissance Discussion
59
Figure 35: Fréquences mensuelles des T. mediterraneus femelles et mâles en fonction descentres de classe de taille.
0102030405060708090
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
juil-10
0
5
10
15
20
25
30
35
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
sep-10
01020304050607080
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
oct-10
0
5
10
15
20
25
30
13 16 19 22 25 28 14 17 20 23 26 29N
om
bre
d'i
nd
ivid
us
LT(cm)
nov-10
0
0,2
0,4
0,6
0,8
1
1,2
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
déc-10
02468
1012141618
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
avr-11
0
5
10
15
20
25
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
mai-11
012345678
13 16 19 22 25 28 14 17 20 23 26 29
No
mb
red
'in
div
idu
s
LT(cm)
jun-11
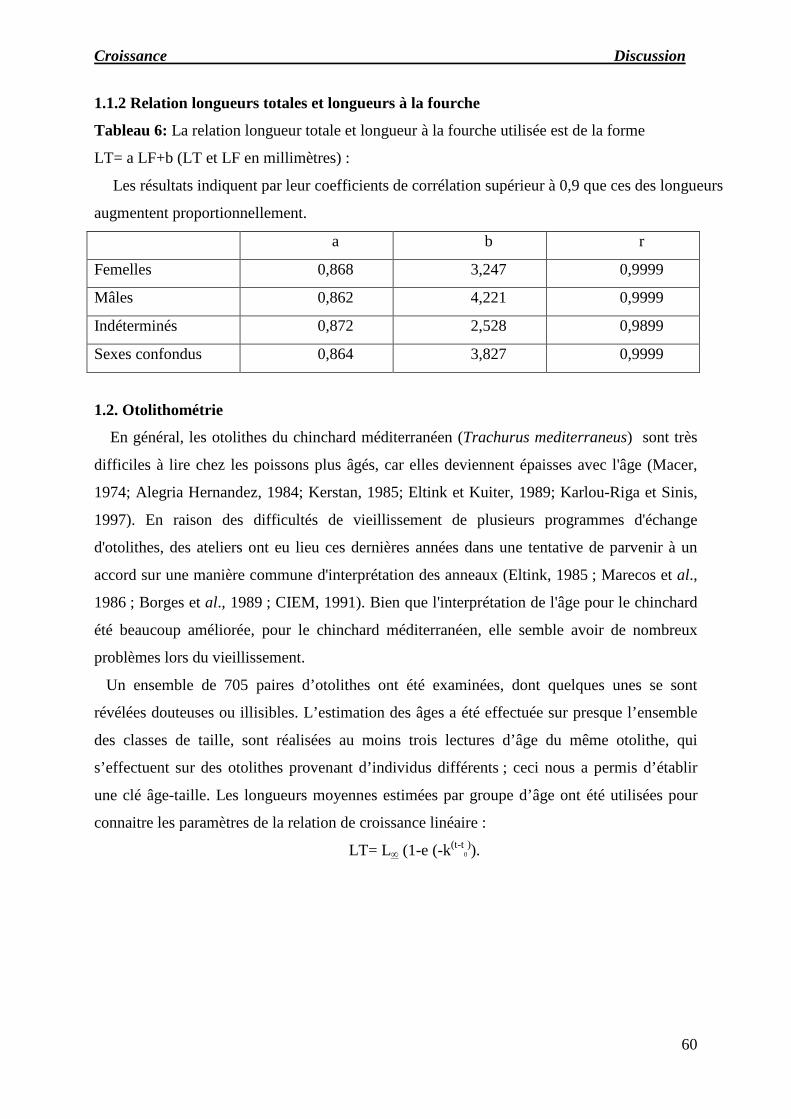
Croissance Discussion
60
1.1.2 Relation longueurs totales et longueurs à la fourche
Tableau 6: La relation longueur totale et longueur à la fourche utilisée est de la forme
LT= a LF+b (LT et LF en millimètres) :
Les résultats indiquent par leur coefficients de corrélation supérieur à 0,9 que ces des longueurs
augmentent proportionnellement.
a b r
Femelles 0,868 3,247 0,9999
Mâles 0,862 4,221 0,9999
Indéterminés 0,872 2,528 0,9899
Sexes confondus 0,864 3,827 0,9999
1.2. Otolithométrie
En général, les otolithes du chinchard méditerranéen (Trachurus mediterraneus) sont très
difficiles à lire chez les poissons plus âgés, car elles deviennent épaisses avec l'âge (Macer,
1974; Alegria Hernandez, 1984; Kerstan, 1985; Eltink et Kuiter, 1989; Karlou-Riga et Sinis,
1997). En raison des difficultés de vieillissement de plusieurs programmes d'échange
d'otolithes, des ateliers ont eu lieu ces dernières années dans une tentative de parvenir à un
accord sur une manière commune d'interprétation des anneaux (Eltink, 1985 ; Marecos et al.,
1986 ; Borges et al., 1989 ; CIEM, 1991). Bien que l'interprétation de l'âge pour le chinchard
été beaucoup améliorée, pour le chinchard méditerranéen, elle semble avoir de nombreux
problèmes lors du vieillissement.
Un ensemble de 705 paires d’otolithes ont été examinées, dont quelques unes se sont
révélées douteuses ou illisibles. L’estimation des âges a été effectuée sur presque l’ensemble
des classes de taille, sont réalisées au moins trois lectures d’âge du même otolithe, qui
s’effectuent sur des otolithes provenant d’individus différents ; ceci nous a permis d’établir
une clé âge-taille. Les longueurs moyennes estimées par groupe d’âge ont été utilisées pour
connaitre les paramètres de la relation de croissance linéaire :
LT= L∞ (1-e (-k(t-t0
)).

Croissance
Figure36: Otolithe d’un Trachurus mediterraneus
Les tableaux : 8 (a, b, c, e) représentent les résultats des
les différentes tailles de poissons, classées par demi
indéterminés et sexes confondus. Pour chaque groupe d’âges on a calculé la taille moyenne
TM.
Chez les femelles sur 372 otolithes nous avons lu des âges de zéro à quatre pour des tailles
allant respectivement de 16,04 à 22cm, avec un maximum d’
Trachurus mediterraneus mâle (classe 16cm, âge II ans) (Gx100),
o : centre du nucleus.
: 8 (a, b, c, e) représentent les résultats des âges lus par otolithométrie, pour
les différentes tailles de poissons, classées par demi-centimètre, pour les femelles,
indéterminés et sexes confondus. Pour chaque groupe d’âges on a calculé la taille moyenne
Chez les femelles sur 372 otolithes nous avons lu des âges de zéro à quatre pour des tailles
allant respectivement de 16,04 à 22cm, avec un maximum d’individus d’âge II.
O
Discussion
61
mâle (classe 16cm, âge II ans) (Gx100),
âges lus par otolithométrie, pour
centimètre, pour les femelles, mâles,
indéterminés et sexes confondus. Pour chaque groupe d’âges on a calculé la taille moyenne
Chez les femelles sur 372 otolithes nous avons lu des âges de zéro à quatre pour des tailles
individus d’âge II.
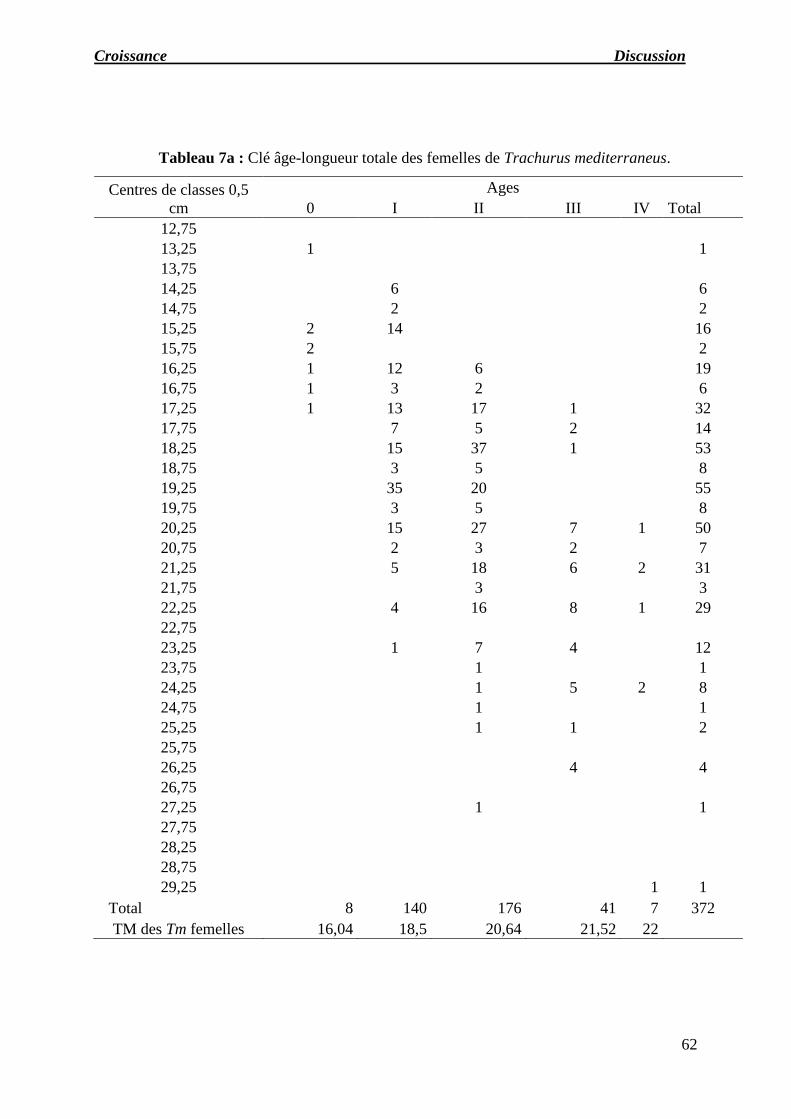
Croissance Discussion
62
Tableau 7a : Clé âge-longueur totale des femelles de Trachurus mediterraneus.
Centres de classes 0,5cm
Ages
0 I II III IV Total
12,7513,25 1 113,7514,25 6 614,75 2 215,25 2 14 1615,75 2 216,25 1 12 6 1916,75 1 3 2 617,25 1 13 17 1 3217,75 7 5 2 1418,25 15 37 1 5318,75 3 5 819,25 35 20 5519,75 3 5 820,25 15 27 7 1 5020,75 2 3 2 721,25 5 18 6 2 3121,75 3 322,25 4 16 8 1 2922,7523,25 1 7 4 1223,75 1 1
24,25 1 5 2 824,75 1 125,25 1 1 225,7526,25 4 426,7527,25 1 127,7528,2528,7529,25 1 1
Total 8 140 176 41 7 372
TM des Tm femelles 16,04 18,5 20,64 21,52 22
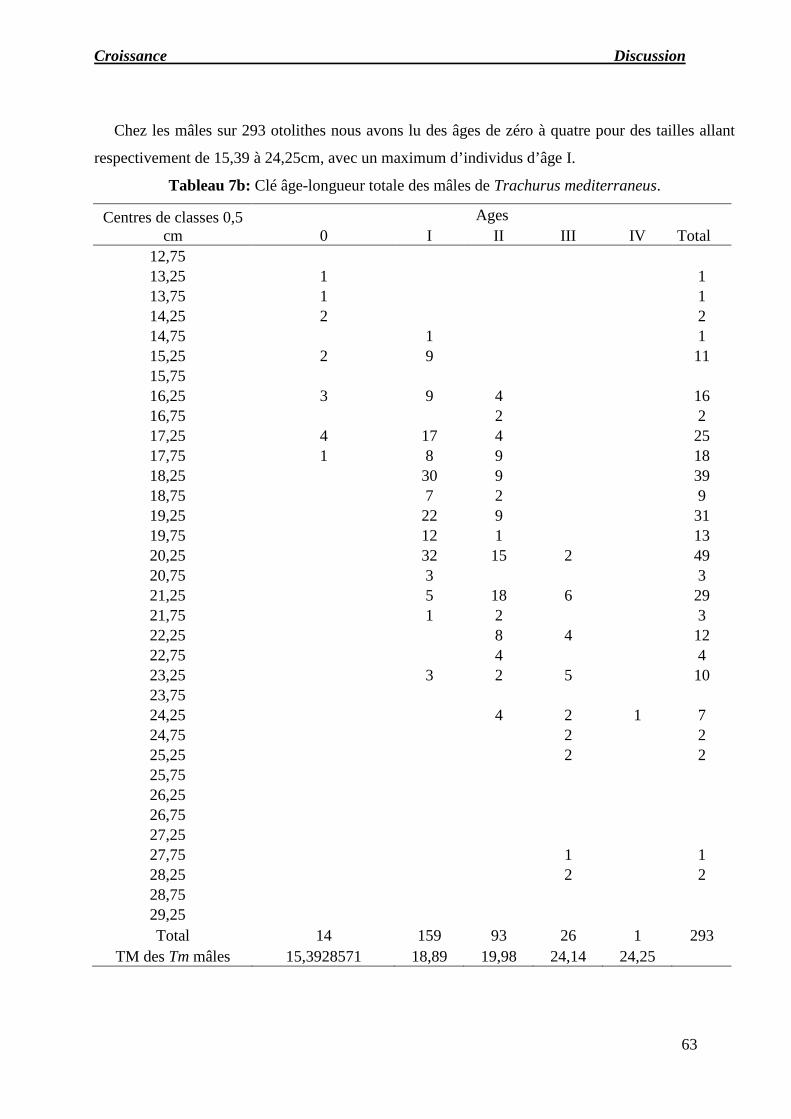
Croissance Discussion
63
Chez les mâles sur 293 otolithes nous avons lu des âges de zéro à quatre pour des tailles allant
respectivement de 15,39 à 24,25cm, avec un maximum d’individus d’âge I.
Tableau 7b: Clé âge-longueur totale des mâles de Trachurus mediterraneus.
Centres de classes 0,5cm
Ages
0 I II III IV Total
12,7513,25 1 113,75 1 114,25 2 214,75 1 115,25 2 9 1115,7516,25 3 9 4 1616,75 2 2
17,25 4 17 4 2517,75 1 8 9 1818,25 30 9 3918,75 7 2 919,25 22 9 3119,75 12 1 1320,25 32 15 2 4920,75 3 321,25 5 18 6 2921,75 1 2 322,25 8 4 1222,75 4 4
23,25 3 2 5 1023,7524,25 4 2 1 724,75 2 225,25 2 225,7526,2526,7527,2527,75 1 128,25 2 228,7529,25
Total 14 159 93 26 1 293
TM des Tm mâles 15,3928571 18,89 19,98 24,14 24,25
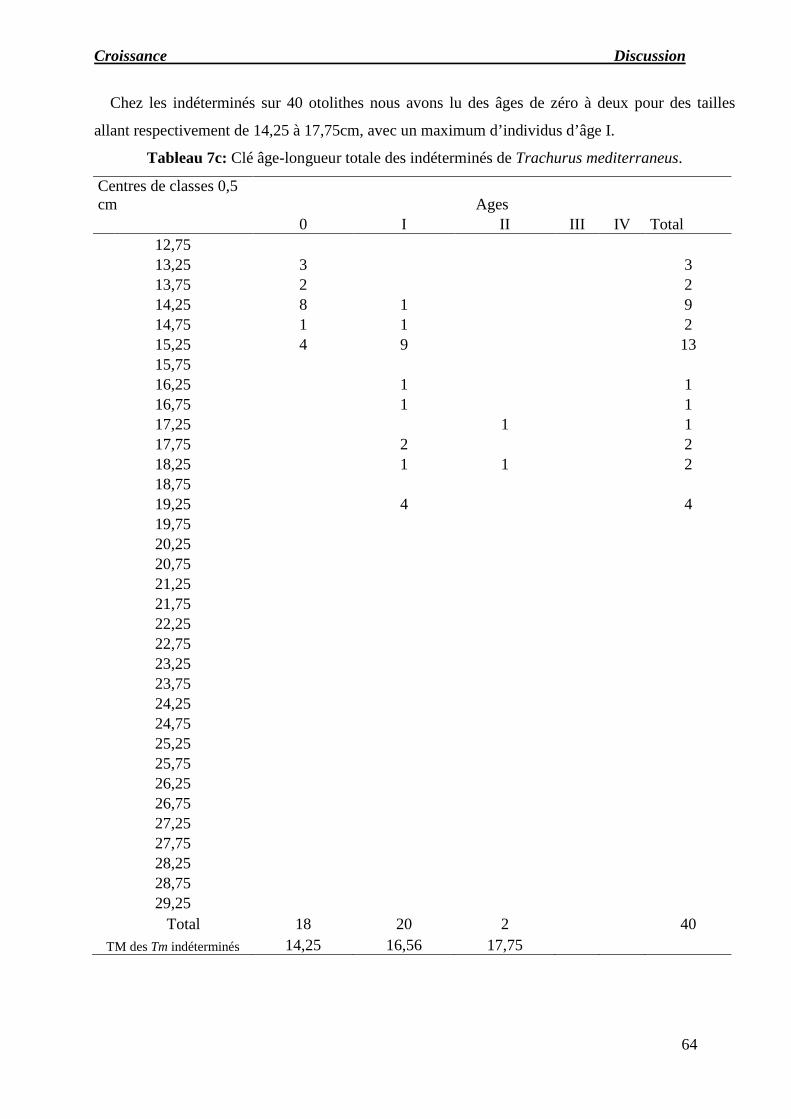
Croissance Discussion
64
Chez les indéterminés sur 40 otolithes nous avons lu des âges de zéro à deux pour des tailles
allant respectivement de 14,25 à 17,75cm, avec un maximum d’individus d’âge I.
Tableau 7c: Clé âge-longueur totale des indéterminés de Trachurus mediterraneus.
Centres de classes 0,5cm Ages
0 I II III IV Total
12,7513,25 3 313,75 2 214,25 8 1 914,75 1 1 215,25 4 9 1315,7516,25 1 116,75 1 117,25 1 117,75 2 218,25 1 1 218,7519,25 4 419,7520,2520,7521,2521,7522,2522,7523,25
23,7524,2524,7525,2525,7526,2526,7527,2527,7528,2528,7529,25
Total 18 20 2 40
TM des Tm indéterminés 14,25 16,56 17,75
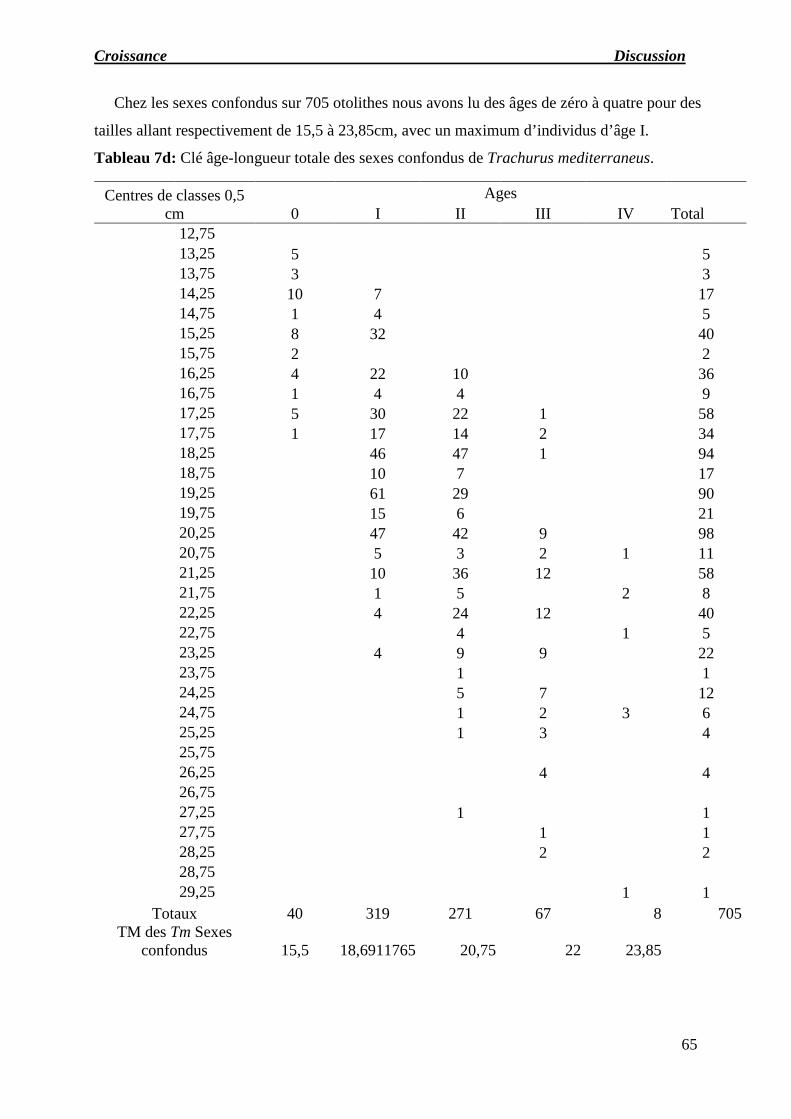
Croissance Discussion
65
Chez les sexes confondus sur 705 otolithes nous avons lu des âges de zéro à quatre pour des
tailles allant respectivement de 15,5 à 23,85cm, avec un maximum d’individus d’âge I.
Tableau 7d: Clé âge-longueur totale des sexes confondus de Trachurus mediterraneus.
Centres de classes 0,5cm
Ages
0 I II III IV Total12,7513,25 5 513,75 3 314,25 10 7 1714,75 1 4 515,25 8 32 4015,75 2 216,25 4 22 10 3616,75 1 4 4 917,25 5 30 22 1 5817,75 1 17 14 2 3418,25 46 47 1 9418,75 10 7 1719,25 61 29 9019,75 15 6 2120,25 47 42 9 9820,75 5 3 2 1 1121,25 10 36 12 5821,75 1 5 2 822,25 4 24 12 4022,75 4 1 523,25 4 9 9 2223,75 1 124,25 5 7 1224,75 1 2 3 625,25 1 3 425,7526,25 4 426,7527,25 1 127,75 1 128,25 2 228,7529,25 1 1
Totaux 40 319 271 67 8 705TM des Tm Sexes
confondus 15,5 18,6911765 20,75 22 23,85
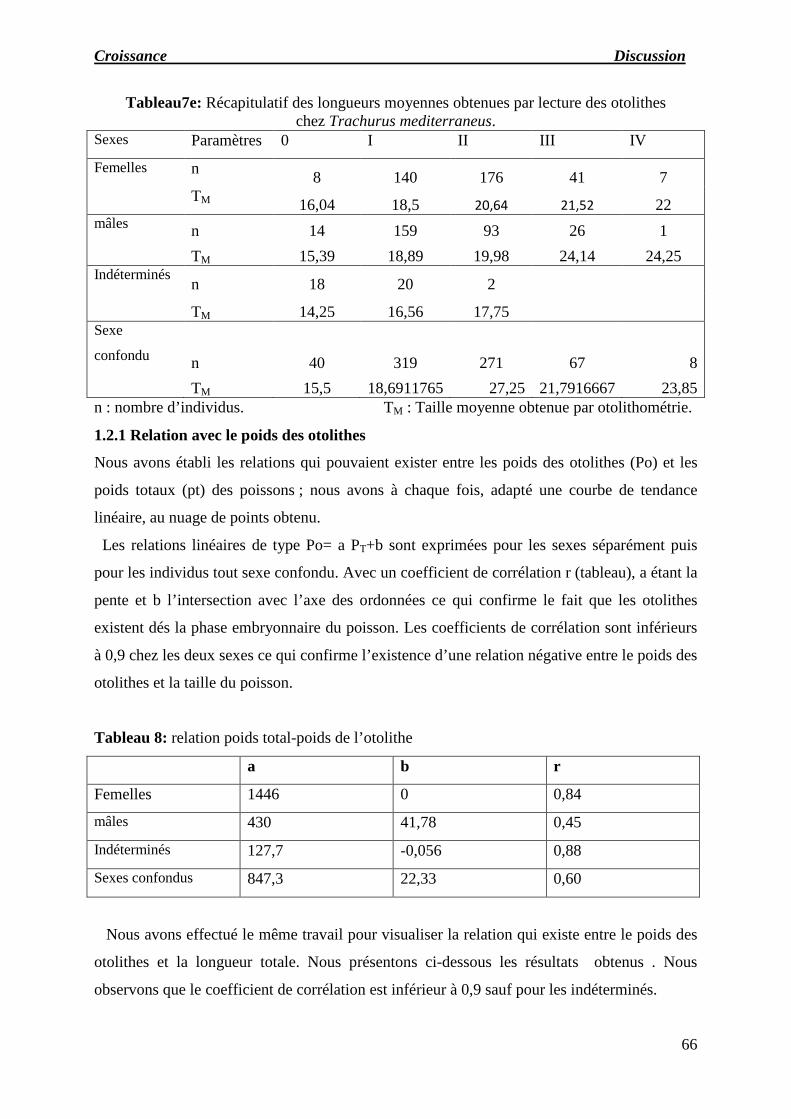
Croissance Discussion
66
Tableau7e: Récapitulatif des longueurs moyennes obtenues par lecture des otolitheschez Trachurus mediterraneus.
Sexes Paramètres 0 I II III IV
Femelles n8 140 176 41 7
TM 16,04 18,5 20,64 21,52 22mâles n 14 159 93 26 1
TM 15,39 18,89 19,98 24,14 24,25Indéterminés
n 18 20 2
TM 14,25 16,56 17,75Sexe
confondu n 40 319 271 67 8
TM 15,5 18,6911765 27,25 21,7916667 23,85n : nombre d’individus. TM : Taille moyenne obtenue par otolithométrie.
1.2.1 Relation avec le poids des otolithes
Nous avons établi les relations qui pouvaient exister entre les poids des otolithes (Po) et les
poids totaux (pt) des poissons ; nous avons à chaque fois, adapté une courbe de tendance
linéaire, au nuage de points obtenu.
Les relations linéaires de type Po= a PT+b sont exprimées pour les sexes séparément puis
pour les individus tout sexe confondu. Avec un coefficient de corrélation r (tableau), a étant la
pente et b l’intersection avec l’axe des ordonnées ce qui confirme le fait que les otolithes
existent dés la phase embryonnaire du poisson. Les coefficients de corrélation sont inférieurs
à 0,9 chez les deux sexes ce qui confirme l’existence d’une relation négative entre le poids des
otolithes et la taille du poisson.
Tableau 8: relation poids total-poids de l’otolithe
a b r
Femelles 1446 0 0,84
mâles 430 41,78 0,45
Indéterminés 127,7 -0,056 0,88
Sexes confondus 847,3 22,33 0,60
Nous avons effectué le même travail pour visualiser la relation qui existe entre le poids des
otolithes et la longueur totale. Nous présentons ci-dessous les résultats obtenus . Nous
observons que le coefficient de corrélation est inférieur à 0,9 sauf pour les indéterminés.
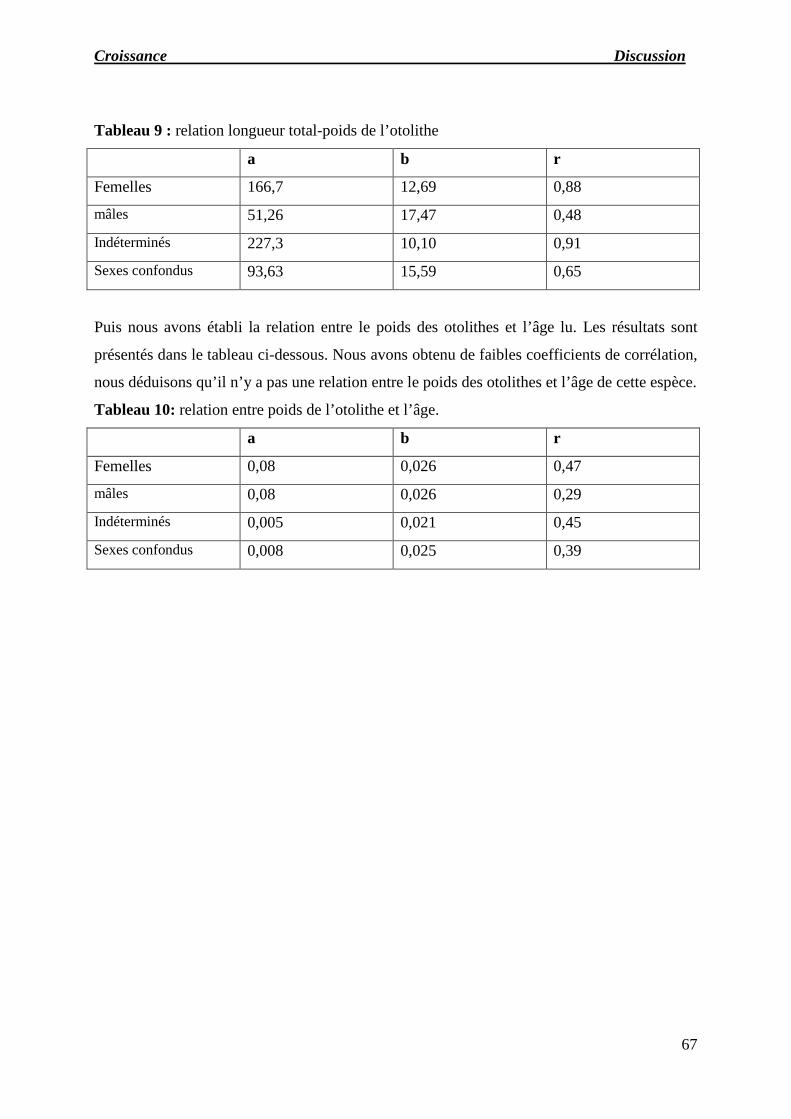
Croissance Discussion
67
Tableau 9 : relation longueur total-poids de l’otolithe
a b r
Femelles 166,7 12,69 0,88
mâles 51,26 17,47 0,48
Indéterminés 227,3 10,10 0,91
Sexes confondus 93,63 15,59 0,65
Puis nous avons établi la relation entre le poids des otolithes et l’âge lu. Les résultats sont
présentés dans le tableau ci-dessous. Nous avons obtenu de faibles coefficients de corrélation,
nous déduisons qu’il n’y a pas une relation entre le poids des otolithes et l’âge de cette espèce.
Tableau 10: relation entre poids de l’otolithe et l’âge.
a b r
Femelles 0,08 0,026 0,47
mâles 0,08 0,026 0,29
Indéterminés 0,005 0,021 0,45
Sexes confondus 0,008 0,025 0,39
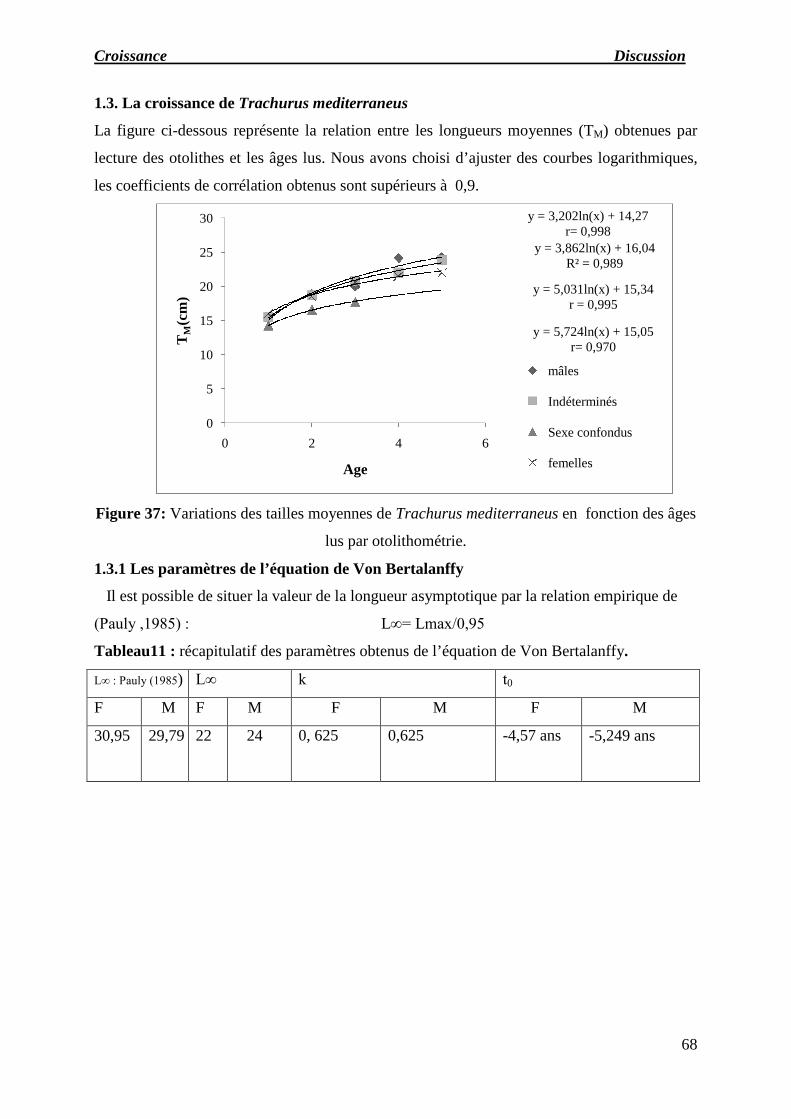
Croissance Discussion
68
1.3. La croissance de Trachurus mediterraneus
La figure ci-dessous représente la relation entre les longueurs moyennes (TM) obtenues par
lecture des otolithes et les âges lus. Nous avons choisi d’ajuster des courbes logarithmiques,
les coefficients de corrélation obtenus sont supérieurs à 0,9.
Figure 37: Variations des tailles moyennes de Trachurus mediterraneus en fonction des âges
lus par otolithométrie.
1.3.1 Les paramètres de l’équation de Von Bertalanffy
Il est possible de situer la valeur de la longueur asymptotique par la relation empirique de
(Pauly ,1985) : L∞= Lmax/0,95
Tableau11 : récapitulatif des paramètres obtenus de l’équation de Von Bertalanffy.
L∞ : Pauly (1985) L∞ k t0
F M F M F M F M
30,95 29,79 22 24 0, 625 0,625 -4,57 ans -5,249 ans
y = 5,724ln(x) + 15,05r= 0,970
y = 5,031ln(x) + 15,34r = 0,995
y = 3,202ln(x) + 14,27r= 0,998
y = 3,862ln(x) + 16,04R² = 0,989
0
5
10
15
20
25
30
0 2 4 6
TM
(cm
)
Age
mâles
Indéterminés
Sexe confondus
femelles
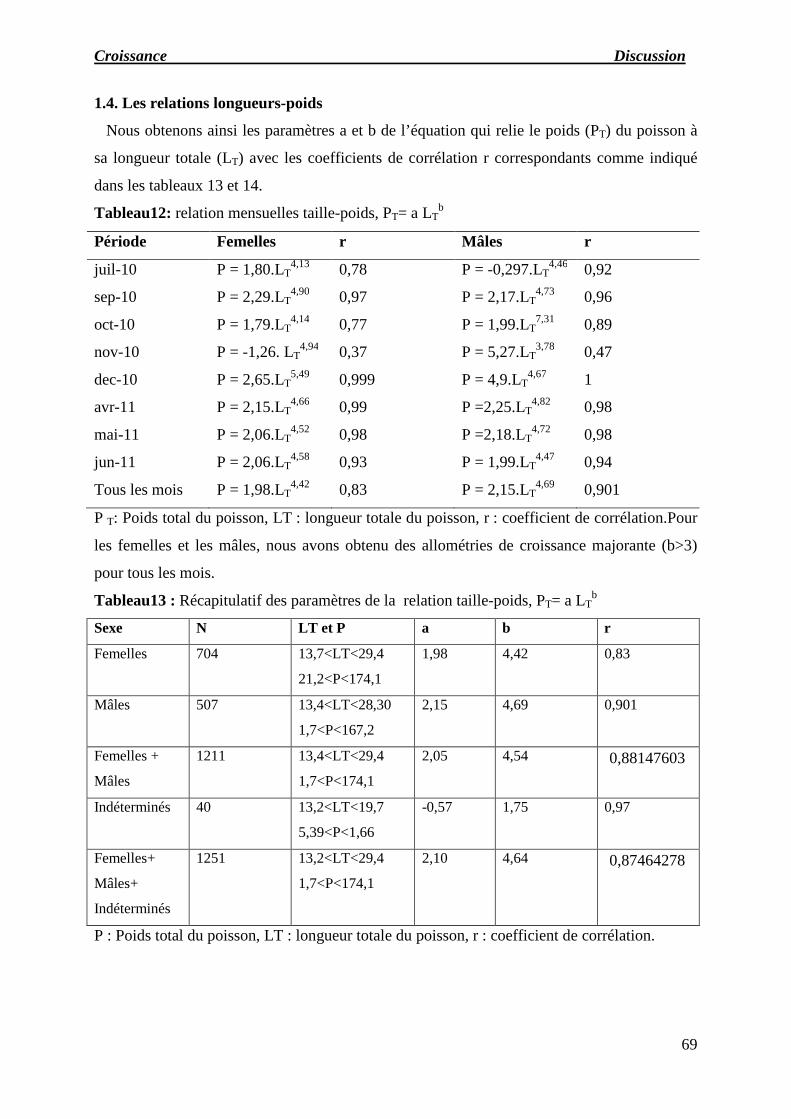
Croissance Discussion
69
1.4. Les relations longueurs-poids
Nous obtenons ainsi les paramètres a et b de l’équation qui relie le poids (PT) du poisson à
sa longueur totale (LT) avec les coefficients de corrélation r correspondants comme indiqué
dans les tableaux 13 et 14.
Tableau12: relation mensuelles taille-poids, PT= a LTb
Période Femelles r Mâles r
juil-10 P = 1,80.LT4,13 0,78 P = -0,297.LT
4,46 0,92
sep-10 P = 2,29.LT4,90 0,97 P = 2,17.LT
4,73 0,96
oct-10 P = 1,79.LT4,14 0,77 P = 1,99.LT
7,31 0,89
nov-10 P = -1,26. LT4,94 0,37 P = 5,27.LT
3,78 0,47
dec-10 P = 2,65.LT5,49 0,999 P = 4,9.LT
4,67 1
avr-11 P = 2,15.LT4,66 0,99 P =2,25.LT
4,82 0,98
mai-11 P = 2,06.LT4,52 0,98 P =2,18.LT
4,72 0,98
jun-11 P = 2,06.LT4,58 0,93 P = 1,99.LT
4,47 0,94
Tous les mois P = 1,98.LT4,42 0,83 P = 2,15.LT
4,69 0,901
P T: Poids total du poisson, LT : longueur totale du poisson, r : coefficient de corrélation.Pour
les femelles et les mâles, nous avons obtenu des allométries de croissance majorante (b>3)
pour tous les mois.
Tableau13 : Récapitulatif des paramètres de la relation taille-poids, PT= a LTb
Sexe N LT et P a b r
Femelles 704 13,7<LT<29,4
21,2<P<174,1
1,98 4,42 0,83
Mâles 507 13,4<LT<28,30
1,7<P<167,2
2,15 4,69 0,901
Femelles +
Mâles
1211 13,4<LT<29,4
1,7<P<174,1
2,05 4,54 0,88147603
Indéterminés 40 13,2<LT<19,7
5,39<P<1,66
-0,57 1,75 0,97
Femelles+
Mâles+ et
Indéterminés
1251 13,2<LT<29,4
1,7<P<174,1
2,10 4,64 0,87464278
P : Poids total du poisson, LT : longueur totale du poisson, r : coefficient de corrélation.

Croissance Discussion
70
Pour les individus à sexes indéterminés, nous avons obtenu une allométrie de croissance
minorante. Pour les mâles et les femelles, nous remarquons une tendance générale à
l’allométrie de croissance majorante ; nous déduisons déduit que le poids croit plus vite que la
taille chez Trachurus mediterraneus.
2. Discussion
Les longueurs totales notées à partir des échantillons étudiés sont comprises entre 13,2cm
et 29,4cm par contre sur la côte bulgare de la mer noire elles varient de 10,5 à 17cm, (Raykov
,2008), cette gamme se distingue de celle de la côte turque de la mer noire, rapportée par
(Şahin et al., 1997), dans la côte turque de la mer noire sont 7,4 et 14,5cm. Dans la même
année (Viette, 1997) observe que la longueur totale de spécimens capturés dans le golfe de
Trieste (9,8 à 38,3 cm) ne correspond pas avec la limite maximale de 10-50 cm proposé par
Fischer et al. (1987) pour la gamme de taille de T. mediterraneus. Le manque de gros
spécimens pourrait être une caractéristique de la section locale de population ou pourrait être
dû à la surpêche, ou à la tendance à de plus gros spécimens se trouvant dans une eau plus que
celle du golfe de Trieste, (Karlovac ,1971). (Genç et al., 1999) pour les populations de
chinchard de la côte Est de la mer noire qui est comprise entre 7,4 à 14,5cm et 6,5à 19cm. Ces
études montrent qu’il y’a une très grande variabilité de longueurs entre les chinchards de la
mer noire, du golfe de Trieste et ceux étudiés dans la baie d’Oran (présente étude).
D’après (Şahin, 2008), dans l’Est de la mer noire les longueurs des poissons variaient entre
9,2cm et 19cm alors que les poids oscillaient entre7, 263 g et 60,812g par contre nous avons
obtenu un intervalle plus large de poids compris entre 1,7g et 174,1g.
Selon (Özaydin et al., 2000), dans le nord de Chypre, l'Est de la mer Méditerranée., ces
différences peuvent être attribuées à la stratégie d’échantillonnage utilisée, tels que la période
ainsi que des variations de température et probable différence entre le potentiel trophique et
diverses localités.
Dans la présente étude les spécimens de Trachurus mediterraneus ont été répartis entre le
groupe d’âge 0 à IV ans, (Sahin et al., 1997) les a classés entre 0 et VI ans, de même le
chinchard de la côte bulgare est compris entre le groupe d’âge 0 à VI ans,( Raykov, 2008) ;
ces résultats concordent avec ceux de (Viette, 1997) où l’âge 0 à VI ans à partir de la lecture
des otolithes et de 0 à V à partir de la lecture des écailles.
Nos observations sur l'âge et la longueur totale à la maturité sexuelle (15cm, I an), ne
confirment pas ceux d’Arneri et al., (1983), et ceux de (Viette, 1997) qui observent que
l’âge et la taille de maturité sexuelle est (16 cm, II ans) .
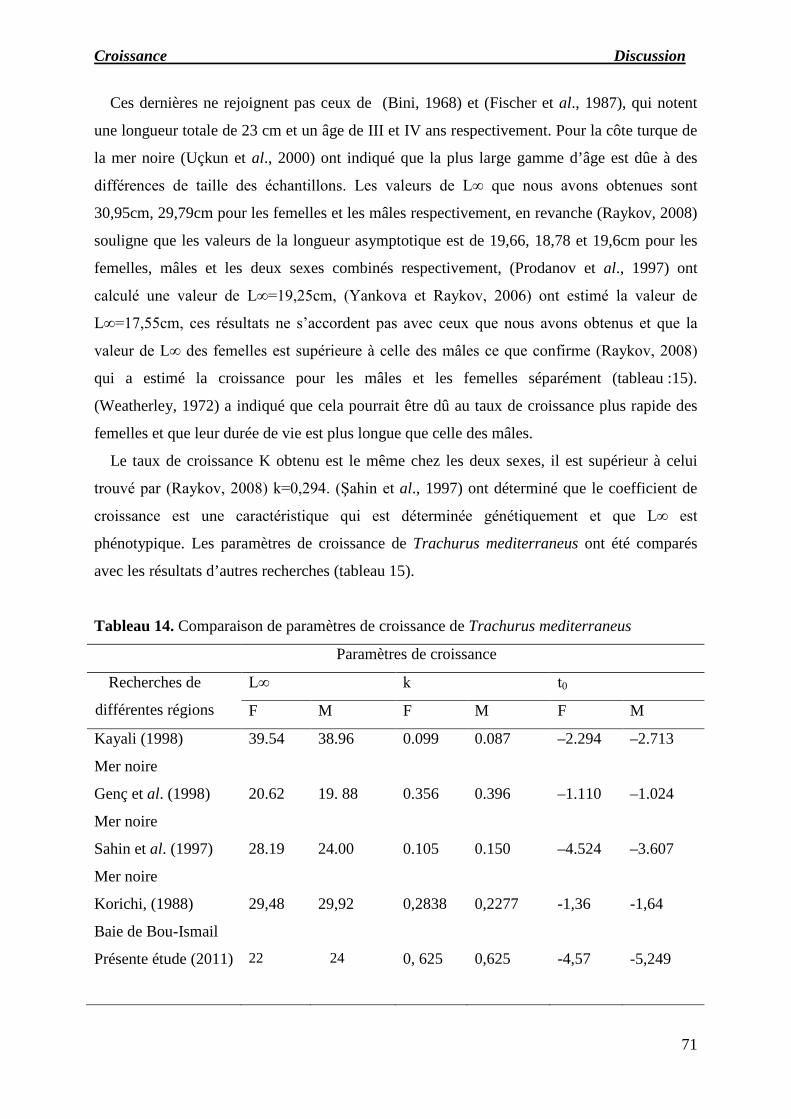
Croissance Discussion
71
Ces dernières ne rejoignent pas ceux de (Bini, 1968) et (Fischer et al., 1987), qui notent
une longueur totale de 23 cm et un âge de III et IV ans respectivement. Pour la côte turque de
la mer noire (Uçkun et al., 2000) ont indiqué que la plus large gamme d’âge est dûe à des
différences de taille des échantillons. Les valeurs de L∞ que nous avons obtenues sont
30,95cm, 29,79cm pour les femelles et les mâles respectivement, en revanche (Raykov, 2008)
souligne que les valeurs de la longueur asymptotique est de 19,66, 18,78 et 19,6cm pour les
femelles, mâles et les deux sexes combinés respectivement, (Prodanov et al., 1997) ont
calculé une valeur de L∞=19,25cm, (Yankova et Raykov, 2006) ont estimé la valeur de
L∞=17,55cm, ces résultats ne s’accordent pas avec ceux que nous avons obtenus et que la
valeur de L∞ des femelles est supérieure à celle des mâles ce que confirme (Raykov, 2008)
qui a estimé la croissance pour les mâles et les femelles séparément (tableau :15).
(Weatherley, 1972) a indiqué que cela pourrait être dû au taux de croissance plus rapide des
femelles et que leur durée de vie est plus longue que celle des mâles.
Le taux de croissance K obtenu est le même chez les deux sexes, il est supérieur à celui
trouvé par (Raykov, 2008) k=0,294. (Şahin et al., 1997) ont déterminé que le coefficient de
croissance est une caractéristique qui est déterminée génétiquement et que L∞ est
phénotypique. Les paramètres de croissance de Trachurus mediterraneus ont été comparés
avec les résultats d’autres recherches (tableau 15).
Tableau 14. Comparaison de paramètres de croissance de Trachurus mediterraneus
Paramètres de croissance
Recherches de
différentes régions
L∞ k t0
F M F M F M
Kayali (1998)
Mer noire
39.54 38.96 0.099 0.087 –2.294 –2.713
Genç et al. (1998)
Mer noire
20.62 19. 88 0.356 0.396 –1.110 –1.024
Sahin et al. (1997)
Mer noire
28.19 24.00 0.105 0.150 –4.524 –3.607
Korichi, (1988)
Baie de Bou-Ismail
29,48 29,92 0,2838 0,2277 -1,36 -1,64
Présente étude (2011) 22 24 0, 625 0,625 -4,57 -5,249
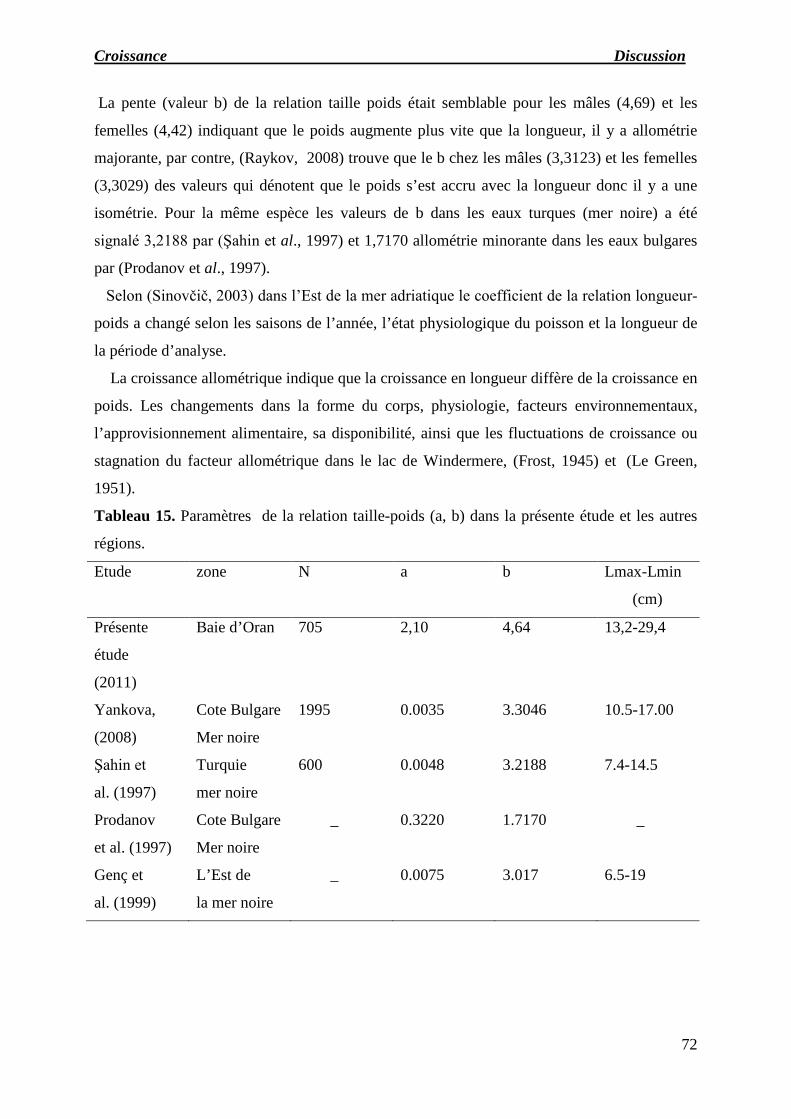
Croissance Discussion
72
La pente (valeur b) de la relation taille poids était semblable pour les mâles (4,69) et les
femelles (4,42) indiquant que le poids augmente plus vite que la longueur, il y a allométrie
majorante, par contre, (Raykov, 2008) trouve que le b chez les mâles (3,3123) et les femelles
(3,3029) des valeurs qui dénotent que le poids s’est accru avec la longueur donc il y a une
isométrie. Pour la même espèce les valeurs de b dans les eaux turques (mer noire) a été
signalé 3,2188 par (Şahin et al., 1997) et 1,7170 allométrie minorante dans les eaux bulgares
par (Prodanov et al., 1997).
Selon (Sinovčič, 2003) dans l’Est de la mer adriatique le coefficient de la relation longueur-
poids a changé selon les saisons de l’année, l’état physiologique du poisson et la longueur de
la période d’analyse.
La croissance allométrique indique que la croissance en longueur diffère de la croissance en
poids. Les changements dans la forme du corps, physiologie, facteurs environnementaux,
l’approvisionnement alimentaire, sa disponibilité, ainsi que les fluctuations de croissance ou
stagnation du facteur allométrique dans le lac de Windermere, (Frost, 1945) et (Le Green,
1951).
Tableau 15. Paramètres de la relation taille-poids (a, b) dans la présente étude et les autres
régions.
Etude zone N a b Lmax-Lmin
(cm)
Présente
étude
(2011)
Baie d’Oran 705 2,10 4,64 13,2-29,4
Yankova,
(2008)
Cote Bulgare
Mer noire
1995 0.0035 3.3046 10.5-17.00
Şahin et
al. (1997)
Turquie
mer noire
600 0.0048 3.2188 7.4-14.5
Prodanov
et al. (1997)
Cote Bulgare
Mer noire
_ 0.3220 1.7170 _
Genç et
al. (1999)
L’Est de
la mer noire
_ 0.0075 3.017 6.5-19

Croissance Discussion
73
3. Conclusion sur la croissance
Dans la présente étude nous avons utilisé la lecture des otolithes pour déterminer l'âge.
Les classes d'âge ont été réparties de 0 à 4. Le taux d’allométrie b de l'équation conduit à la
valeur 4,64, ce qui indique que la croissance du poids est allométrique majorante. Les
paramètres de croissance de l'équation de Von Bertalanffy ont été évaluées séparément pour
les femelles et les mâles comme suit: L ∞ = 22, K = 0, 625, t0= -4,57ans, et L ∞ = 24, K =
0,625, t0= -5,249ans, l'âge maximum chez les mâles et les femelles est 4 ans. Les mâles sont
caractérisés par des taux de croissance supérieurs à ceux des femelles.

Conclusion
74
Conclusion générale
La présente étude a permis de préciser un certain nombre de points de la biologie de
Trachurus mediterraneus pêché dans la baie d’Oran.
La sex-ratio globale est comprise entre 0,36 et 0,42 (mâles: femelles). Les femelles sont
plus nombreuses pendant la période intense de reproduction (mai -juin) et pendant les mois
d’octobre et novembre. Les mâles sont plus nombreux aux mois de septembre et avril.
La reproduction a lieu pendant la période printemps-été avec une forte activité
reproductrice d’avril à juin. Une étude histologique des gonades et une analyse statistique des
variables biométriques (RGS, RHS, Kc) ont été réalisées pour étudier la biologie du
chinchard. Cinq stades de maturité sexuelle peuvent être décrits dans les deux sexes mâles et
femelles. L'indice gonado-somatique atteint sa valeur maximale en mai et juin.
La taille de première maturité sexuelle se situe à 15 cm de longueur totale chez les mâles et
les femelles ce qui correspond à une année d’âge. L'étude de la fécondité a montré que
Trachurus mediterraneus a une fécondité absolue faible; en effet elle oscille entre 2063 et
18854 ovocytes pour des femelles de 17 cm (148 g) à 31 cm (698 g). Le diamètre ovocytaire
est compris entre 38.4μm et704μm avec une moyenne de 262,19μm.
L'étude de l'âge et de la croissance a été réalisée par otolithométrie, l'estimation de l'âge,
nous a permis de distinguer 5 groupes d'âge pour les deux sexes. Les longueurs obtenues par
le modèle de Von Bertalanffy montrent que les mâles ont une longueur asymptotique
légèrement supérieures à celles des femelles. Les valeurs de ces paramètres de croissance
retenues pour les deux sexes sont:
Femelles : L∞ =22cm, to = -4,57ans et K = 0,625an
Mâles : L∞ =24cm, to = -5,249 ans et K = 0,625an
Le taux d’allométrie b de l'équation à la valeur de 4,64 donc majorant, ce qui indique que
la croissance du poids est plus rapide que celle de la taille.
A priori, la différence de croissance entre les deux sexes peut être due au fait que
les femelles se nourrissent plus que les mâles étant donné leur besoin élevé en énergie
indispensable pour le bon déroulement de leurs fonctions métaboliques reproductives.
La variation peut être due aussi à différents stades ontogénétiques de développement, ainsi
que les différences dans les conditions environnementales, la longueur, l’âge, le sexe, le
développement

Conclusion
75
des gonades, la taille, la quantité et la qualité de nourriture, ainsi que la température de l’eau
sont étroitement liés à des paramètres de croissance, (Ricker, 1975).
Au terme de ce travail il semble que la pêche du chinchard à queue jaune du genre
Trachurus en baie d’Oran cible essentiellement des jeunes individus ayant des tailles
comprises entre 130 mm et 290 mm. L’étude de la biologie de la reproduction et de la
croissance du Trachurus mediterraneus, laisse comprendre une situation de surexploitation de
cette espèce dans la zone étudiée. L’augmentation de l’effort de pêche et la capacité motrice
des chalutiers ne font qu’aggraver la situation de cette ressource. Ainsi, le transfert partiel de
l’activité de pêche vers le large, au-delà de 12 milles (1mille=1852m), est recommandé ainsi
que l’effort de pêche de cette ressource côtière devrait être réduit. Cette mesure permettra
certainement une augmentation de la biomasse, une amélioration du niveau de recrutement et
une croissance de la productivité de la ressource.

Références bibliographiques
76
Références bibliographique
Aka Z., Koç H.T., Turan C.2004. A study on the anchovy Engraulis encrasicolus, Linnaeus
(1758) in Turkish Seas, Turkey. Pak. J. Biol Sci. 7 : 1121-1126.
Aleev Yu, G. 1957. Horse mackerel (Trachurus) of the Soviet seas. Tr. Sevastopol. Biol. St.,
9: 167–212. (in Russian).
Alegria Hernandez, V. 1984. Observations on the age and growth of Trachurus trachurus
(L.) in the middle Adriatic. Biljeske-Notes. Inst. Oceanogr. Ribar., Split 58, p. 6.
Alonso M.K., Crespo E.A., Goodall R.N.P., Pedraza S.N., Schiavini A.C.M. 1999.
Stomach contents of false killer whales (Pseu-doria crassidens) stranded on the coasts of the
Strait of Magellan, Tierra dei Fuego. Marine Mammal Science 15, 712-724.
Anatole C., Cao F.2006. Vision par ordinateur et otolithes : vers une modélisation de la
morphogenèse.
Arneri E., Tangerini P. 1983. Biological data, from the ‘‘Pipeta’’ expeditions, on Trachurus
mediterraneus (Steindachner) in the Adriatic Sea. CGPM Troisieme consultation technique
sur l’évaluation des stocks dans l’Adriatique, Fano, Annex O: 127–130.
Arneri E. 1984. Nota preliminare sulla biologia della specie del genere Trachurus (T.
mediterraneus, T. trachurus, T. picturatus) in Adriatico. Nova Thalassia, 6(suppl.): 459–464.
B Chemmam-Abdelkader., El abed A., M. M. Kraiem.2002. Periode de ponte, sex-ratio et
maturite sexuelle de Dentex maroccanus (teleostei, sparidae) des cotes tunisiennes. Rapp int.
INSTM, Salammbô.Vol. 29
Bagenal T. B.1978. Aspects of fish fecundity. In Gerking, S. D. (Ed). Ecology of Freshwater
fish production. Blackwell Scientific Publications, oxford.
Banarescu P.1964. The fauna of the popular Republic of Romania. Pisces – Osteichthyes
XIII Academici Republicii Populare Romine, Bucuresti, 959 pp. (in Romanian).
Barnabe G.1991. Reproduction chez les poissons. In: Bases biologiques et écologiques de
l’aquaculture. Lavoisier Paris (Tech & Doc), 500 p.
Bauchot M.L. 1987. Serranidae. In: W. Fischer, M.L. Bauchot, M. Schneider (Editors).
Fiches FAO d’Identification des Espèces pour les Besoins de la Pêche. (Révision 1),
Méditerranée et Mer Noire. Zones de Pèche 37, II (Vertébrés). FAO-CEE, Rome, 1.
Bauchot R. & M.L. Bauchot .1978. Coefficient de condition et indice pondéral chez les
téléostéens. Cybium 3(4):3-16.

Références bibliographiques
77
Beckman W.C. 1948. The length-weight relationship, factors for conversion between
standard and total lengths, and coeffecients of condition for several Michigan fishes. Trans.
Am. Fish. Soc. 75:237-256.
Ben Salem M.1988. Régime alimentaire de Trachurus trachurus (Linnaeus, 1758) et de
Trachurus mediterraneus (Steindachner, 1868) (Poissons, Téléostéens, Carangidae) de la
province Atlantico-Méditerranéenne. Cybium, 12: 247-253.
Ben Salem M. 1988. Taxinomie numérique des espèces du genre Trachurus Rafinesque 1810
(Poissons, Téléostéens, Carangidae). Cybium, 12: 45–58.
Ben Salem M., Ktari, M. H. 1981. Présentation des espèces du genre Trachurus Rafinesque,
1810 et Caranx Lacépède, 1801 (Poissons, Téléostéens, Carangidae) des côtes tunisiennes:
morphologie et biologie. Bulletin Offciel National des Pêches, Tunisie, 4: 155–168.
Bennoui A. & Neghli L.2009.Rapport de participation à la campagne évaluation des
ressources pélagiques. Centre National de Recherche et de Développement de la Pêche et
d’Aquaculture (CNRDPA).
Berg L., Bogdanov L. S., Editors., Kozhin N. I. et Rass T. S. 1949. Commercial fishes of
the USSR (en russe).
Beverton ., Holt .1959. A review of the lifespans and natural mortality rates of fish in nature
and their relation to growth and other physiological characteristics. In Wolstenholme, G.E.W.
and M. O'Connor (eds.). CIBA Colloquium on ageing, Vol.5:142–180
Billard R., Breton B., Jalabert B.1972. Les cellules de Sertoli des poissons téléostéens. I.
Etude ultrastructurale. Ann. Biol. Anim. Biochim. Biophys., 12(1): 19-32.
Billard R.1979. La gamétogenèse, le cycle sexuel et le contrôle de la reproduction chez les
poissons Téléostéens. Bull. Fr. Pisc., 273: 117-136.
Bini G. 1968. Famiglia Carangidae. In Atlante dei pesci delle coste italiane, pp. 43–55.
Mondo sommerso. Roma, 175 pp.
Bouaziz A. 1992. Le merlu (Merluccius merluccius mediterraneus Cadenat, 1950) de la
baie de Bou Ismaïl : biologie et écologie. Thèse de Magister, ISMAL, Algérie.
Borges, M.F.1989. Results of the horse mackerel (Trachurus trachurus L.) otolith exchange
programme during 1988–1989. ICES, CM/H 13, 10 pp.
Bougis P. 1952. Recherche biométriques sur les rougets (M.barbatus L., M. surmuletus L.).
Archs. Zool. exp. gén., 89, 2,57-174.

Références bibliographiques
78
Campana S.E. & Casselman .M. 1993. Scock discriminarion using orolirh shape analysis.
Canadian Journal of Fisheries and Aquatic Sciences 50, 1062-1083.
Campana S.E. (1999). Chemistry and composition of fish otoliths: pathways, mechanisms
and applications. Marine Ecology Progress Series 188:263-297, in Panfili. 2007.
Les otolithes, intégrateurs de l'histoire individuelle des poissons. Ecole Doctorale Systèmes
intégrés en Biologie, Agronomie, Géosciences, Hydrosciences, Environnement (SIBAGHE).
Université Montpellier 2.
Cardenas L., Cristian E., Hernandez., E Poulin., A Magoulas., I. K. Weld., F Patricio
Ojeda. 2004. Origin, diversification, and historical biogeography of the genus Trachurus
(Perciforms: Carangidae).Molecular Phylogenetics and Evolution, 35, 496-507.
Castonguay M., Simard., P. Gagnon, P. 1991. Usefulness of Fourieranalysis of otolith
shape for Atlantic mackerel (Scomber scombrus) stock discrimination. Canadian Journal of
Fisheries and AquaticSciences 48, 296-302.
Cautis I. 1979. Biologie de la reproduction. In Le chinchard de la Mer Noire (Trachurus
mediterraneus ponticus), pp. 533–547. Ed. by E. Pora. Institut roumain de recherches marines,
Constanta (Roumanie). 753 pp.
Chilton D.E., R.J. Beamish. 1982. Age determination for fishes studied by the
Groundfish Program at the Pacific Biological Station. Canadian Special Publication of
Fisheries and Aquatic Sciences 60.
Dadzie S., Abou-Seedo F., & Manyala J. O. 2008. Length-Length relationship, Length-
weight relationship, size at maturity and fecondity of Parastromateus niger (Carangidae) in
Kuwait waters. J. Appl. Ichthyol. 24: 334-336.
Dalouche F. 1980. La pêche et ses statistiques dans la region oranaise. Etude de quelques
caractères biologiques sur la sardine Sardina pilchardus, (Walb, 1972). Thèse de
magistèr, Université d’Oran: 107p.
Dardignac J. 1963. Les chinchards (Trachurus) des côtes Française de la méditerranée, (note
préliminaire). Rapp.P.V Réuen.CIESM. Vol. XVII.fase. 342-346p.
Demir M. 1961. On the eggs and larvae of the Trachurus trachurus (L.) and Trachurus
mediterraneus (Stdhnr) from the Sea of Marmara and the Black Sea. CIESMM, 16: 317–320.
Diaye N., A. M. 1995. Contribution à la connaissance de la biologie de la reproduction de
deux espèces de Sparidae (Poisson, Téléostéens) des côtes sénégalaises. DEA de Biologie
Animale. Université Cheikh Anta Diop. Dakar, 44 pp.

Références bibliographiques
79
Dieuzeide R., Roland J.1958. Prospection des fonds chalutables des côtes algériennes.
Recherche des nouvelles zones (années 1956-1957), Bull. Sta. Aquic. Pêche, Castiglione,
Nouvelle série (9) 9-69.
Dziewulska K., Domagala J. 2003. Histology of salmonid testes during maturation. Reprod.
Biol., 3(1): 47-61.
Eltink, A.1985. Results of horse mackerel (Trachurus trachurus L.) otolith exchange
programm. ICES, CM/H 40, 17 pp.
Eltink, A., Kuiter, C.J.1989. Validation of ageing techniques on otoliths of horse mackerel
(Trachurus trachurus L.). ICES, CM/H 43, 15 pp.
FAO. 1987. Fiche FAO d’identification des espèces pour les besoins de la pêche.
Fischer W., Bauchot M. L., Schneider M. 1987. Fiche FAO d’identification des
espèces pour les besoins de la pêche. Rév. 1. Méditerranée et mer noire, zone de pêche 37, II:
Vertébrés. 761: 1530p.
Fischer W., Bauchot M. L., Schneider M. 1987. Fiches FAO d’identification des espèces
pour les besoins de la pêche. Méditerranée et Mer Noire, Zone de pêche 37, vol. 2, p. 1028.
FAO, CEE, Rome. 1529 pp.
Fontana A., Le guen J. C. 1969. Etude de la maturité sexuelle et de la fécondité de
Pseudotolithus fonticulus elongatus. Cah. O.R.S.T.O.M. Sér. Océanogr., Vol. VII, N” 3 : 14p.
freshwater fish productions, Oxford, pp. 75-101.
Friedland K.D., Reddin, D.G. 1994. Use of otolith morphology in stock discriminations of
Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and A quatic Sciences 51, 91-
98.
Friedland K.D., Haas R.E., Reddin D.G., Shimizu N.,Youngson A.E .1998.
Strontium:calcium ratios in Atlantic salmon (Salmo salar) otoliths and observations on
growth and maturation. Cana-dian Joltrnal of Fisheries and Aquatic Sciences 55, 1158-1168.
Froese R., Pauly, D., (Editors).2001. FishBase.World Wide Web electronic publication.
Frost W.E. 1945. The age and growth of eels (Anguilla anguilla) from the Windermere
catchment area. Part 2.J. Anim. Ecol., 14: 106-12.
Genç Y., Akbulut B., Başar S., Ceylan B., Çiftçi Y., Şahin. T., Tabak İ ., Üstündağ C.,
Zengin M. 1998. Ekonomik Deniz Ürünleri Araştırma Projesi Sonuç Raporu,
TAGEM/İY/96/17/3/001, Su Ürünleri Merkez Araştırma Enstitüsü, Trabzon, 127s

Références bibliographiques
80
Genç Y., Akbulut B., Başar S., Ceylan N., Çiftçi Y., Şahin T., Tabak I., Üstündağ C.,
Zengin M.1999. The Research Project of Economical Marine Products. TKB, Central
Fisheries Research Institute Trabzon, 157 pp.
Gherram M. 2009. Contribution à l'étude de la reproduction du saurel Trachurus
trachurus (L., 1758) pêché en baie d'Oran : étude de l’ovogenèse, période de ponte, sex-ratio,
indice de condition et fécondité. Thèse de Magister. Université d’Oran : 87p.
Grier H.J., Linton J.R., Leatheland J.F., DE Vlaming V.L. 1980. Structural evidence of
two testicular types in teleost fishes. Am. J. Anat., 159: 331-345.
Grier H.J., Taylor R.G. 1998. Testicular maturation and regression in the common snook.
J. Fish Biol., 53: 521-542.
Grier H.J., LO Nostro F.2000. The teleost germinal epithelium: a unifying concept Proc.
6TH International Symposium Reprod. Physiol. Fish. Bergen, Norway. Norsberg et al., eds.
Grier H.J.2000. Ovarian germinal epithelium and folliculogenesis in the common snook,
Centrapomus undecimalis (Teleostei: Centrapomidae). J. Morphol., 243: 265-281.
Guichet R. 1966. Répartition bathymétrique saisonnière des poissons de chalut dans le golfe
de Gascogne.Cons.Int.Explor.Mer.4, 6 p.
Guillermo C. 1998. L’évolution des colorants à travers les âges, revue de l’AFH.
Gunn J.S.1990. Revision of selected genera of the family carangidae, Records of the
Australian Museum. Supp. 12.
Gusmão P., Foresti F., Quagio-Grassiotto I. 1999. Ultrastructure of spermiogenesis in
Plagioscion squamosissimus (Teleostei, Perciforme, Sciaenidae). Tissue Cell, 31(6): 627-
633.
Hecht T. 1979. The value of otoliths in fresh water fish- eries biology and taxonomy.
University of the North Publication Series A No. 19: 1–29.
Heins D. C., Baker J.A., & Guill J. M. 2004. Seasonal and interannual components of
intrapopulation variation in clutch size and egg size of a darter. Ecol. Freshw. Fish. 13: 258-
265.
Higashino T., Miura T., Miura C., Yamauchi K. 2002. Histological studies on early
oogenesis in barfin flounder (Verasper moseri). Zool. Sci., 19: 557-563.
Ivanov L., Beverton R.J.H. 1985. The Fishery Resources of the Mediterranean Part 2: The
Black Sea, GFCM, Studies and Reviews No.60: 135 s.
Jalabert B.2005. Particularities of reproduction and oogenesis in teleost fish compared to
mammals. Reprod. Nutr. Dev., 45: 261-279.

Références bibliographiques
81
Jardas I., Pallaoro A., Santic M. 2003. Biometric properties of Horse mackerel, Trachurus
trachurus (Osteichthyes: Carangidae) from the middle Adriatic Sea. International Journal of
Marine Sciences, 86-95. Mayr, E., Linsley, E. and Usinger, R. 1956. Methods and principles
of zoological systematics. Moscow: 15-42. (in Russian).
Jensen A.C. 1965. A standard terminology and notation for otolith readers. ICNAF
Research Bulletin 2: 5-7.
Karlou-Riga C. 1995. Biology and dynamics of the species of the genus Trachurus (Pisces,
Carangidae) in the Saronikos Gulf. PHD thesis, Aristotele University of Tessaloniki, School
of Biology, Greece, 296 pp.
Karlou-Riga C., Economidis P. S.1997. Spawning frequency and batch fecundity of horse
mackeret, (Trachurus trachurus L), in the Saronikos Gulf (Greece). J. Appl. Ichthyol. 13:
97-104.
Karlovac J., and Karlovac O. 1971. Contribution à la connaissance de l’écologie de
Trachurus trachurus (L.) au large de l’Adriatique. Izvjesca Institut za Oceanografiju i
Ribarstvo, Split, 5(2A): 24.
Kayalı E. 1998. Doğu Karadeniz’deki İstavrit (Trachurus mediterraneus Steindchner, 1868)
ve Hamsi (Engraulis encrasicolus Linnea, 1758) Balıklarının Bazı Populasyon Parametreleri
ve Beslenme Ekolojileri, Yükesek Lisans Tezi, KTÜ, Fen Bilimleri Enstitüsü, Bal Tekn. Müh.
Anabilim Dalı, Trabzon, 238 pp.
Kerstan M. 1985. Age, growth, maturity, and mortality estimates of horse mackerel
(Trachurus trachurus) from the waters west of Great Britain and Ireland in 1984. Archiv für
Fischereiwissenschaft, 36: 115–154.
Korichi H. S. 1988. Contribution à l’étude biologique de deux espèces de Saurel :
Trachurus trachurus (Linné, 1758), Trachurus mediterraneus (Steindachner, 1868) et de la
dynamique de Trachurus trachurus (L) en baie de Bou Ismail (Alger) Thèse Magister
I.S.M.A.L : 260p.
Langeron M. 1942. Précis de microscopie : technique-expérimentation-diagnostique.
Masson et Cie, Paris, 1339p.
Layachi M., Melhaoui M., Srour A., Ramdani M. 2007. Contribution à l’étude de la
reproduction et de la croissance du Rouget-barbet de vase (Mullus barbatus L., 1758) de la
zone littorale méditerranéenne de Nador (Maroc). Bulletin de l’Institut Scientifique, Rabat,
section Sciences de la Vie, no29.

Références bibliographiques
82
Le Green E.D. 1951. The length-weight relationship and seasonal cycle in gonad weight and
condition in the perch (Perca fluviatilis). J. Anim. Ecol, 20: 530-538.
Leclaire L. 1972. La sédimentation Holocène sur le Versant méridional du bassin
Algéro-Baléares (pré continent Algérien). Mémoire. Muséum National d’Histoire Naturelle.
Paris., 391p.
Leclercq-Smekens M., Leloup R. 1998. Notions de technique histologique et
d’histochimie, Facultés Universitaires Notre-Dame de la Paix Namur.
Legendre M., Jalabert B.1988. Physiologie de la reproduction. In: Biologie et écologie des
poissons d'eau douce africains. Lévèque C., Bruton M.N. et Ssentongo G.W. Eds.
ORSTOM Paris, pp. 153-187.
Lombarte A., Morales-Nin B. 1989. Growth of Nezumia aequalisand Coelorhynchus
fasciatus (Pisces: Macruridae) in Namibian waters. Collection ofScientific Papers of ICSEAF
16, 191-198.
Lombarte A., Lleonart J. 1993. Otolith size changes related with body growth, habitat depth
and temperature. Environmental Bio-logy of Fishes 37, 297-306.
Loubens G., Panfili J. 1992. Estimation de l'âge individuel de Pro-chilodm nigricam
(Teleosrei, Prochilodidae) dans le Béni (Bolivie) : protocole d'étude et application. Aquatic
Living ReJource.r 5, 41-56.
Louverstein O. 1971. The labyrinth. In fish physiology (Hoar, WS. & Randall, D.J., ed.)
New York, USA : Academic Press. pp 207-240.
Macer C. T. 1974. The reproductive biology of the horse mackerel Trachurus trachurus (L.)
in the North Sea and English Channel. Journal of Fish Biology, 6: 415–438.
Manni L., Rasotto M.B. 1997.Ultrastructure and histochemistry of the testicular efferent
duct system and spermiogenesis in Opistognathus whitehurstii (Teleostei, Trachinoidei).
Zoomorphol., 177: 93-102.
Marecos, M.L. 1986. Preliminary analysis of horse mackerel (Trachurus trachurus L.) otolith
(L1) measurements. ICES, CM/H 72, 7 pp.
Mattei X., Siau Y., Thiam D,. Thiaw O.T. 1993. Peculiarities in the organization of
testis of Ophidion sp. (Pisces Teleostei). Evidence of two types of spermatogenesis in teleost
fish. J. Fish Biol., 43: 931–937.
Maximov V., Radu G., Radu E., Butu A.2002. Contributions à la connaissance des
caractéristiques biologiques et biochimiques des principales espèces de poissons du
littoral roumain de la mer Noire au cours de l’année 2000. Recherches Marines,
INCDM, Constanta, 34:239-259 .

Références bibliographiques
83
Mellinger J.2002. Sexualité et Reproduction des poissons. CNRS EDITIONS, Paris. 346 pp.
Messieh S.N., M Dougall, C., Claytor, R. 1989. Separation of Atlantic herring (Clllpea
harengus) stocks in the southern Gulf of St. Lawrence using digitized otolith morphometrics
and discrimi- nant function analysis. Canadicm Techniral Report of Fusheries and Aquatic
ScienceJ 1647.
Meunier F.J.1988. Détermination de l'âge individuel chez les Ostéichthyens à l'aide de la
squelettochronologie : historique et méthodologie. Acta Oecologica. Oemlogitl Generalis 9,
299-329.
Mikhailov K., Prodanov, K. 1983. Approximate assessment of the natural mortality rate of
the anchovy in the Bulgarian Black Sea coast. Pross. IRR, Varna, 20: 173-182.
Nagahama Y. 1986. Testis. In: Vertebrate endocrinology: fundamentals and biomediacal
implication. Pang P.K.T., Schreibman M.P., Gorbman A. eds., vol. I. New York:
Academic Press, pp. 399–437.
Nagahama Y.1983. The functional morphology of teleosts gonads. In: Fish physiology.
Hoar W.S., RandallD.J. and Donalson E.M. eds., vol. IXA. London: Academic Press, pp.
223–275.
Nikolski G. VI .1963. The ecology offishes. London and New-York Academic press. 352 p.
Nolf D. 1995. Studies on fossil otoliths: the state of the art. In Recentdevelopments in fish
otolith research (Secor, D.H., Dean M. & Cam- pana, S.E., ed.), pp. 513-544. Columbia, sc,
USA: University of South Carolina Press.
Olsson O., North A.W. 1997. Diet of the King Penguin Aptenodytes patagonims during three
summers at South Georgia. lbis 139, 504-512.
Oosthuizen E., N. Daan. 1974. Egg fecundity and maturity of North Sea cod, Gadus
morhua. Neth. J. Sea Res., 8(4): 378–397.
Özaydin O., Bilecenoğlu M., Kaya M. 2000. Age and growth of the Curled Picarel
Centracanthus cirrus Rafinesque, 1810 (Osteichthyes: Centracanthidae) in Northern Cyprus,
Eastern Mediterranean Sea. Acta Adriatica, 41: 35-42.
Panfili J., De Pontual H., Troadect H., P.J. Wright.2001. Manuel de sclérochronologie des
poissons.
Patiño R., Sullivan C.V.2002. Ovarian follicle growth, maturation, and ovulation in teleost
fish. Fish Physiol. Biochem., 26: 57-70.
Paugy D.1980. Écologie et biologie des Alestes nurse (Pisces Characidae) des rivières de
Côte d’Ivoire. Cah. ORSTOM ser. Hydrobiol. 13 (3-4): 143-149.

Références bibliographiques
84
Pauly D., 1980.On the interrelationships between natural mortality, growth parameters and
mean environmental temperature in 175 fish stocks. J. Cons. CIEM, 39 (2) : 175-192.
Petit. H.V., F. Contonguay.1994. Growth and carcass quality of prolific cross lambs fed
Pörtner H. O., Berdal B., Blust R., Brix O., Colosimo A., Fischer T., Giuliani Johansen
T., Knust R., Wachter B. 2001. Climate induced temperature effects on growth
performance, fecundity and recruitment in marine fish: developing a hypothesis for cause and
effect relationships in Atlantic cod (Gadus morhua) and common eelpout (Zoarces viviparus).
Cont. Shelf Res. 21: 1975-1997.
Prodanov K., Daskalov G., Mikhailov, K., Maxim K., Chashchin, A., Arkhipov A.,
Shlyakhov V., Ozdamar E. 1997. General fisheries council for Mediterranean, FAO.
Studies and reviews. Environmental management of fish resources in the Black Sea and their
rational exploitation. Studies and reviews, 68: 73-81.
Prolonge C.2007. Étude histologique du développement sexuel de l’apron du Rhône Zingel
asper L., percidé endémique menacé d’extinction, Laboratoire de Reproduction et
Développement Comparés, 25 rue du plat, 69288 Lyon Cedex 02.
Pudney J. 1995. Spermatogenesis in nonmammalian vertebrates. Microsc. Res. Tech., 32:
459-497.
Quéro J.-C., Dardignac J. P.-P. &VAYNE J.-J.1989. Les poissons du golfe de Gascogne.
Ifremer/ Secrétariat de la Faune et de la flore.229p.
Quéro J.-C., Porché P.-P. & VAYNE J.-J. 2003. Guide des poissons de l’Atlantique
européen. Les guides du naturaliste. Delachaux et Niestlé Eds,Lonay/ Paris. 465p.83pl.
Quéro J.-C., Abernot-LE Gac C., DE Casamajor M.-N., Gautier G., Morandeau G.,
Spitz J., Duhamel E., Van Canneyt O., Vayne J.-J. 2007. Faune Française de
l'Atlantique poissons carangidés. Ann. Soc. Sei. nat. Charente-Marit., 9(9).
Ravaglia M.A., Maggese M.C.2003. Ovarian follicle ultrastructure in the teleost
Synbranchus marmoratus (Bloch, 1795), with special reference to the vitelline enveloppe
developpement. Tissue Cell, 33: 9-17.
Raykov V., Yankova M. 2008. Growth dynamics and mortality estimation of the Horse
Mackerel (Trachurus mediterraneus ponticus, Aleev) migrating along the Bulgarian Black
Sea Coast. Proceedings of first Biannual Scientific Conference “Black Sea Ecosystem and
Beyond” 8-10 May 2005, Istanbul, 765-778.
Remili S, Kerfouf .2009. Surveillance environnementale et développement durable côtier
(cas du littoral oranais). Actes du Cinquième Colloque International sur le thème :

Références bibliographiques
85
« Energies, Changements climatiques et développement durable », Hammamet, Tunisie, les
15,16 et 17 juin.
Ricker W. E.1975. Computation and Interpretation of biological statistics of fish populations,
Bulletin No. 191, Department of the Environment, Fisheries and Marine Services, Ottawa,
Ontario.
Rinchard J., Kestemont P., Poncin P. 1998. Croissance ovocytaire et régulation
stéroïdienne chez les poissons à pontes unique et multiple: une revue. Ann. Limnol., 34(2):
211-225.
Selman K., Wallace R.A.1989.Cellular aspects of oocyte growth in teleosts. Zool. Sci., 6:
211-231.
Smith-Vaniz W.F. 1986. Carangidae. In: Fishes of the North-eastern Atlantic and the
Mediterranean / Poissons de l'Atlantique du Nord-est et de la Méditerranée (Whitehead
PJ.P., Bauchot M.-L, Hureau J.-C, Nielsen J. & Tortonese E. Eds), Unesco, Paris vol 2:
815 - 844.
Smith-Vaniz W.F. 2007. Review of the Crevalle jacks, Carnax hippos complex, with
description of new species from West Africa (Teleostei : Carangidae), Carpenter (K.E.), Fish,
Bull. 105 : 207-233.
Saat T.1993.The morphology of oocytes final maturation and fertilization in fish. In:
Physiological and biochemical aspects of fish development (Walther B.T., Fyhn H.J., eds.)
Norway: Univ. of Bergen, pp. 71–85.
Şahin T., Genç, Y. and Okur, H. 1997. Karadeniz’in Türkiye Sularındaki İstavrit
(Trachurus mediterraneus ponticus) Populasyonunun Gelişme ve Üreme Özelliklerinin
İncelenmesi, Turk. J. Zool, 21: 321-327.
Şahin C., N. Kasapoğlu., A. Mutlu Gözler., F. Kalayc., N. Hacimurtazaoğlu., C
Mutlu.2008. Age, Growth, and Gonadosomatic Index (GSI) of Mediterranean Horse
Mackerel (Trachurus mediterraneus Steindachner, 1868) in the Eastern Black Sea,Rize
University, Faculty of Fisheries, Rize - Turkey,Karadeniz Technical University, Faculty of
Marine Science, Trabzon - Turkey,Giresun University, Faculty of Arts and Sciences, Giresun
– TURKEY, 157pp.
Santic et al.2003. Feeding habits of mediterranean horse mackerel, Trachurus mediterraneus
(carangidae), in the central adriatic sea Mer Médit., 35: 452-453.

Références bibliographiques
86
Santos J. N. S., Araùjo F. G., Silva M. A., Vasconcellos R. M.2007. Sex ratio and sexual
dimorphism of the anchovy Anchoa januaria (Actinopterygii, Engraulidae) in a tropical bay
in south-eastern Brazil. J. Fish Biol. 71: 877-888.
Schulz R.W., Bogerd J., França L.R., Menting S., Godinho H.P., Vilela D.A.R. 2005.
Sertoli cell proliferation in adult testis–evidence from two fish species belonging to different
orders. Biol. Reprod., 73: 891-898
Sinovčič G. 2003. The length-weight relationship of anchovy, Engraulis engrasicolus (L.), in
the eastern Adriatic Sea. Acta Adriatica. 44: 181.
Slastenenko E. 1956. Karadeniz Havzası Balıkları, E.B.K.Umum Müdürlüğü, İstanbul, 711s.
Soliman I.A., Hasnem M.T., Al-Sayes A.A. 1982. Thes biological investigation on some
pelagic fish species. Bull.Inst.Océanogr. & fish, ARE, 8 (é) : 257-270
Steindachner F. 1868. Ichthyol. Bericht. über· eine nach. Spannien und, Portugal
unternommere Reise. Sitzs-Akad.' Wiss. Wien, LVII-l, p. 382.
Staicu I., Radu G., Maximov V., Radu E., Anton E.2004. Etat des populations des
principales espèces de poisson à valeur marchande du secteur marin roumain (1990-2002).
Recherches Marines, INCDM, Constanta, 35:153-172.
Suter W., Morel, P. 1996. Pellet analysis in the assessment of Great Cormorant
Phalacrocorax carbo diet: teducing biases from orolith wear when reconstructing fish length.
Colonial waterbirds 19,280-284.
Sylla S., Boua C., Atse., N’Guessan., J. Kouassi. 2009. Stratégie de Reproduction du
Carangidae Trachinotus teraia Cuvier, 1832 dans la lagune Ebrié. Centre de Recherches
Océanologiques (CRO), B.P.V 18 Abidjan (Côte d’Ivoire). Sciences & Nature Vol.6 N°1 : 83
– 94.
Thiam M. 1980. Biologie, écologie et dynamique des cynoglosses du plateau continental
sénégambien. Reproduction, biométrie et croissance de Cynoglosses canariensis
(Steind.1882). Doc. Scient. CRODJ, ISRA, 65, 58p.
Thiaw O. T. 1993. Etude ultrastructurale de la gamétogenèse comparée des Cyprinodontidae
Myers, 1955 (poissons, Téléostéens). Relation entre la morphologie des gamètes, la
phylogénie et la taxonomie. Doct. d'Etat es Sei. Nat. UniV1. C. A. D. Dakar,
Thiry M., Poncin P. 2005. Morphological changes of the nucleolus during oogenesis in
oviparous teleost fish, Barbus barbus. J. Struct. Biol., 152: 1-13.

Références bibliographiques
87
Torres G. ., Lombarte, A. & Morales-Nin, B. 2000. Sagittal otolith size and shape
variability to identify geographical intraspecific differences in three species of the genus
Merluccius. journal of the Marine Biological Association of the United-Kingdom, Plymouth
80,333-342.
Treasurer J. W., Holliday, F. G. T. 1981. Some aspects of the reproductive biology of perch
Perca fluviatilis L. A histological description of the reproductive cycle. Journal of Fish
Biology, 18: 359–376.
Turner C.L. 1919. The seasonal cycle in the spermary of the perch. J. Morphol., 32: 681-711.
Tyler C.R., Sumpter J.P. 1996. Oocyte growth and development in teleosts. Rev. Fish Biol.
Fisch., 6: 287-318.
Uçkun D., Toğulga, M. and Taşakavak, E. 2000. A preliminary study on the growth of the
Common hake (Merluccius merluccius L., 1978) in Izmir Bay, Aegean Sea. Acta Adriatica,
41: 25-34.
Viette M., Ferrero E.A., Giulianini P. G. 1997. Reproductive biology of scad,
Trachurus mediterraneus (Teleostei, Carangidae), from the Gulf of Trieste. – ICES
Journal of Marine Science, 54: 267–272.
Von Bertalanffy. 1938. A quantitative theory or organic growth laws. Hum. Biol., 10: 181-
213.
Wallace R. A., et Selman, K.1985. Major protein changes during vitellogenesis and
maturation of Fundulus heterclitus. Develp. Biol. 110: 492-498.
Wallace R.A., Selman K. 1990. Ultrastructural aspects of oogenesis and oocyte growth in
fish and amphibians. J. Electron microsc. Tech., 16: 175-201.
Wallace R.A., Boyle S.M., Grier H.J., Petrino T.R., Selman K. 1993. Preliminary
observations on oocyte maturation and other aspects of reproductive biology in captive female
snook, Centropomus undecimalis. Aquaculture, 116(2-3): 257-273.
Watanabe y ., Saito, H. 1998. Feeding and growth of early juvenile Japanese sardines in the
Pacifie waters off central Japan. Journal of Fish Biology 52, 519-533.
Weatherley A.H. 1972. Growth and ecology of fish ecology of fish populations. Academic
Press, London, pp. 293.
Westernhagen V.H. 1974. Observations on the natural spawning of Alectis indicus (Rüppell)
and Caranx ignobilis (Forsk.)(Carangidae). J. Fish Biol.6:513-516.
Wilson C.W., R.J. Beamish, E.B. Brothers K.D. Carlander J.M. Casselman, J.M.
Dean, A. Jearld, Jr., E.D. Prince., A. Wild. 1987. Glossary, p. 527-530. In R.C.

Références bibliographiques
88
Summerfelt, and G.E. Hall (eds.), Age and Growth of Fish, Iowa State University Press,
Ames, Iowa.
Wright P.J., Armstrong J. D., Fallon-Cousins P. 2001. The rela-tionship between otolith
accretion and resting metabolic rate in juvenile Atlantic salmon during a change in
temperature.journa/ of Fish Biology 59, 657-666.
Yankova M., Raykov V. 2006. Approximate assessment of the horse mackerel natural
mortality rate, Trachurus mediterraneus ponticus, Aleev in the Bulgarian Black Sea
territorial waters. Secretary marine I.N.C.D.M. 36, 341-348.
Yoneda M., Fujita H., Matsuura S., Matsuyama M., Tokimura M., Takeshita N.,
Takeshita K .2001. Reproductive cycle, fecundity, and seasonal distribution of the angler fish
Lophius litulon in the East China and Yellow seas. Fish Bull., 99: 356-370.
Autre références :
Ministère de la pêche. 2010.
Direction de la pêche d’Oran.2011.
Centre National de Recherche et de Développement de la Pêche et d’Aquaculture
(CNRDPA) .2009.
Aït Ouakli W.2011. l’expression le Quotidien actualité.

Annexes
89
Annexe 1Fiche d’échantillonnage de poissonDate d’échantillonnageNom de l’espèceDate de pêcheLieu de pêcheSexe
N° LT LF PT PG PF Stadesde
maturité
Otolithe Estomac Muscle
01
02
03
04
05
06
07
08
09
10
11
12
13
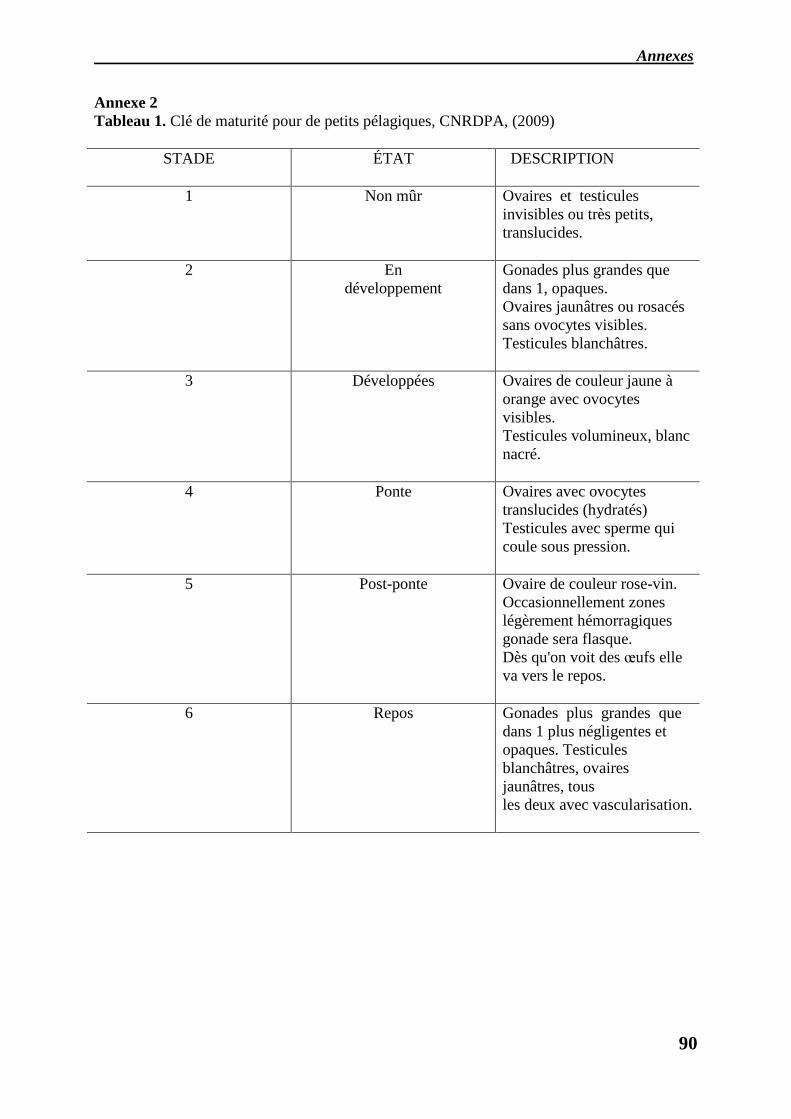
Annexes
90
Annexe 2Tableau 1. Clé de maturité pour de petits pélagiques, CNRDPA, (2009)
STADE ÉTAT DESCRIPTION
1 Non mûr Ovaires et testiculesinvisibles ou très petits,translucides.
2 Endéveloppement
Gonades plus grandes quedans 1, opaques.Ovaires jaunâtres ou rosacéssans ovocytes visibles.Testicules blanchâtres.
3 Développées Ovaires de couleur jaune àorange avec ovocytesvisibles.Testicules volumineux, blancnacré.
4 Ponte Ovaires avec ovocytestranslucides (hydratés)Testicules avec sperme quicoule sous pression.
5 Post-ponte Ovaire de couleur rose-vin.Occasionnellement zoneslégèrement hémorragiquesgonade sera flasque.Dès qu'on voit des œufs elleva vers le repos.
6 Repos Gonades plus grandes quedans 1 plus négligentes etopaques. Testiculesblanchâtres, ovairesjaunâtres, tousles deux avec vascularisation.

Annexes
91
Annexe 3Liquide de Gilson
solution Volume et quantité
Alcool 60° 100 ml
Eau distillée 888 ml
Acide nitrique 15 ml
Chlorure de mercure 20 g
Acide acétique glacial 18 ml
Annexe 4Composition des solutions utilisées dans l’histologie
Bouin : solution mère (1L)Solution Volume et quantité Conservation
Alcool 95° 1L illimité
Acide picrique cristallisé10 g
Bouin alcoolique (100ml)
Solution mère 45 ml 2 Semaines
Formol 26 ml
Eau distillée 22 ml
Acide acétique glacial 7 ml

Annexes
92
Annexe 5Hématoxyline (100ml)
solution Volume et quantité conservation
Solution (a) :
Hématoxyline 0,5 g
Ethanol absolu 5 ml
Solution (b) :
Alun de K 10 g
Eau distillée 100 mlOxyde de mercure 0,25 g
Acide acétique 2 ml
Solution (a) +(b) Ajout de 2 % d’acideacétique pour conserver
Eosine B (100 ml)
solution Volume et quantité
Eosine B 1g
Ethanol 70° 100 ml
Annexe 6Solution albumineuseComposants Quantités
Albumine d’œuf 1g
Eau distillée 100ml

Annexes
93
Annexe 7
Technique :
Tableau I : présentation des différentes étapes de déshydratation et inclusion (Leclercq-
Smekens, 1998).
Etape Solution Durée
I- la fixation Bouin alcoolique 15 jours
II- Déshydratation :
4 bains d’acétone : Acétone 1Acétone 2Acétone 3Acétone 4
30 min30 min30 min30 min
2 bains de toluéne : Toluéne 1Toluéne 2
30 min30 min
III- inclusion et mise enbloc :
2 bains de paraffine Paraffine 1Paraffine 2
1 h1h
Annexes
94
Annexe 8Technique:Tableau: présentation des différentes étapes de coloration à l’hématoxyline éosine(Guillermo, 1998).Etapes Bains Durée
Déparaffinage 2 bains de xylène10 mn chacun
Rinçage 1 bain d'alcool absolu1 bain d'alcool à 95°
3 mn3 mn
Hydratation Eau courante 6 mn
Coloration Hématoxyline 1 mn
Rinçage Eau de robinet 6 mn
Coloration Eosine 10 secondesRinçage Eau courante 6 mn
Déshydratation 3 bains d’alcool1 bain alcool 70°1 bain d’alcool absolu1bain d’alcool 95°
1 mn 30 secondes2 mn2 mn
Eclaircissement 2 bains de xylène 5mn chacun
Montage milieu de montage (Eukitt)Pose d'un couvre objet(lamelle)
Annexe 9Trichrome à froid : hémalun-phloxine-vert lumière (Langeron, 1942).
Mode opératoire :Déparaffinage Toluol (2 à 3 minutes)
Méthanol absolue (2 à 3 minutes)
Méthanol 95 % (3 minutes)
Méthanol 70 % (3 minutes)
Eau courante (10 minutes) pour blanchir
ColorationHémalun (10 minutes)Eau couranteEthanol-HCl (2 minutes) pour différencier

Annexes
95
Eau courante (20 minutes) pour bleuirPhloxine (5 minutes)Eau acétifiée à 1% (3 bains successifs)Acide phosphotungstique (1 minute) pour différencierEau courante (5 minutes) pour rincerVert lumiere (30 secondes)Eau acétifiée à 1% (3 bains successifs) pour rincerEau acétifiée à 1% (3 bains successifs) pour rincer
DéshydraterIsopropanol (3 bains successifs)Toluol (3 bains successifs)Monter la préparation au DPX.
Préparation des différentes solutions de colorationHémalun :
Porter à ébullition un litre d’une solution saturée d’alun de potassium (Merck 1047)
dans l’eau distillée ; Retirer de la source de chaleur et ajouter lentement (et par petite dosesurtout au début) 3g d’Hématein Standard Fluka ;Laisser bouillir ;Refroidir ;Filtrer et ajouter 20 ml d’acide acétique glacial (Merck 60) ;Toujours filtrer avant l’emploiNe pas conserver la solution plus de deux moisEthanol-HClAlcool éthylique 95° dénaturé à l’éther 100ml ;HCL fumant (Merck 317) 0,4 mlPhloxinePhloxine B à 5% dans de l’eau distillée.Acide phosphotungstique (Mecrk) 5g ;Eau distillée 100 mlVert lumièreDissoudre 1g de light green SF yellowish (National Aniline Division) dans 100 ml d’eauacitifiée à 1% ;A l’emploi, diluer 10 fois cette solution.Eau acétifiée à 1%
1 ml d’acide acétique dans 99 ml eau distillée.
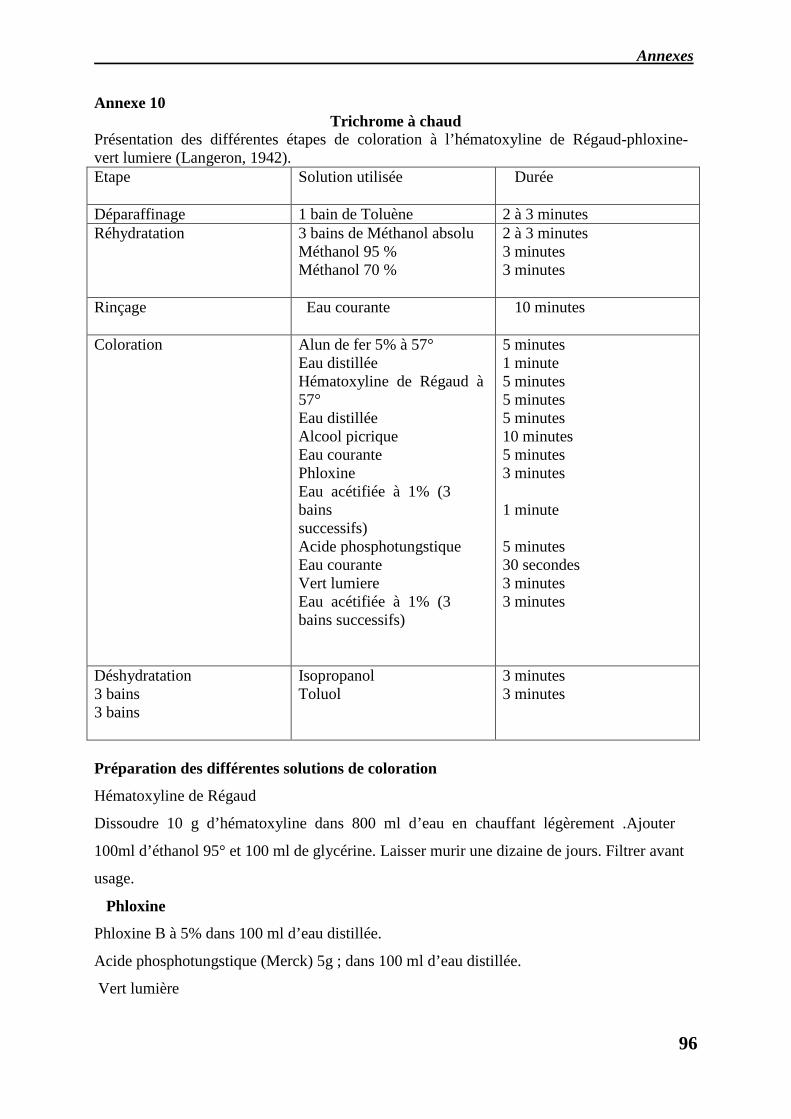
Annexes
96
Annexe 10Trichrome à chaud
Présentation des différentes étapes de coloration à l’hématoxyline de Régaud-phloxine-vert lumiere (Langeron, 1942).Etape Solution utilisée Durée
Déparaffinage 1 bain de Toluène 2 à 3 minutesRéhydratation 3 bains de Méthanol absolu
Méthanol 95 %Méthanol 70 %
2 à 3 minutes3 minutes3 minutes
Rinçage Eau courante 10 minutes
Coloration Alun de fer 5% à 57°Eau distilléeHématoxyline de Régaud à57°Eau distilléeAlcool picriqueEau courantePhloxineEau acétifiée à 1% (3bainssuccessifs)Acide phosphotungstiqueEau couranteVert lumiereEau acétifiée à 1% (3bains successifs)
5 minutes1 minute5 minutes5 minutes5 minutes10 minutes5 minutes3 minutes
1 minute
5 minutes30 secondes3 minutes3 minutes
Déshydratation3 bains3 bains
IsopropanolToluol
3 minutes3 minutes
Préparation des différentes solutions de coloration
Hématoxyline de Régaud
Dissoudre 10 g d’hématoxyline dans 800 ml d’eau en chauffant légèrement .Ajouter
100ml d’éthanol 95° et 100 ml de glycérine. Laisser murir une dizaine de jours. Filtrer avant
usage.
Phloxine
Phloxine B à 5% dans 100 ml d’eau distillée.
Acide phosphotungstique (Merck) 5g ; dans 100 ml d’eau distillée.
Vert lumière

Annexes
97
Dissoudre 1g de light green SF yellowish (National Aniline Division) dans 100 ml d’eau
acétifiée à 1% .A l’emploi, diluer 10 fois cette solution.
Eau acétifiée à 1%
1 ml d’acide acétique dans 99 ml eau distillée.
Annexe 11Trichrome de Masson
1) Coloration des noyaux
Premier temps : Temps de mordançage dans une solution d’Alun de fer 15mn à une
heure
Deuxième temps : Sans laver allonger les lames dans l’hématoxiline de Régaud, de
15mn à 1 heure.
Laver à l’eau courante
2) Différenciation
Un bain d’alcool à 95° de quelques minutes
Un bain d’alcool picrique de 10mn
Un bain d’alcool à 95°
Lavage à l’eau courante pendant 2 à 3 mn
3) Bleuissement des noyaux
Un bain de solution de carbonate de lithiumLavage à l’eau courante
4) Coloration des cytoplasmes
Fuchsine acide + ponceau de xylidine 5mnRinçage rapide à l’eau courante (obligatoire)
5) Différenciation
6) Acide phosphomolybidique 5mn
Pas de rinçage
7) Coloration du tissu conjonctif
Vert lumière 6mn
Lavage à l’eau du robinet
8) Différenciation du vert lumière
Un bain d’eau acétifiée
Un bain d’alcool à 75°
Deux bains d’alcool à 100°
Trois bains de toluène
Montage au DPX

Résume Cette étude a été réalisée sur la biologie d'une espèce de carangidés de la baie d’Oran: Trachurus mediterraneus (Steindachner, 1868). 1251 individus ont échantillonnés de juillet 2010 à juin 2011. La longueur moyenne totale a été estimée à 20,16cm pour les femelles et 20,42 cm pour les mâles.Les longueurs totales obtenues à partir des échantillons étudiés sont comprise entre 13,2cm et 29,4cm et un intervalle plus large de poids compris entre 13,2g et 29,4g.Nous avons examiné la sexe ratio, la taille et l'âge à la première maturité, la fécondité, la période de reproduction, l'âge et croissance chez cette espèce. La sexe ratio globale chez les femelles et les mâles est de 0,36 et 0,42 respectivement. Il varie d'un mois à l'autre. La taille à la première maturité est de15 cm chez les mâles et les femelles soit 1 ans. La reproduction a lieu pendant la période printemps-été avec une forte activité reproductrice d’avril à juin. Une étude histologique des gonades et une analyse statistique des variables biométriques (RGS, RHS, Kc) ont été aussi utilisées pour étudier la biologie du chinchard, Trachurus mediterraneus. Cinq stades de maturité sexuelle peuvent être décrite dans les deux sexes mâles et femelles.L'indice gonado-somatic, atteint sa valeur maximale en mai-juin. La fécondité varie entre 2063 et 18854 ovocytes. Les diamètres ovocytaires varient entre 38.4 Micro m et704Micro m avec une moyenne de 262,19 Micro m.Les paramètres
de croissance de Van Bertalanffy pour les deux sexes sont: Femelles : L Infini =22cm, to
= -4,57ans et K = 0,625an Mâles : L Infini =24cm, to = -5,249 ans et K =
0,625an Le taux d’allométrie b de l'équation conduit à la valeur 4,64, ce qui indique que la croissance du poids est allométrique majorante.
Mots clés: Trachurus mediterraneus; Baie d’Oran; Sex-ratio; Reproduction; Fécondité; Croissance; RGS; RHS; Kc.












![MR-Kontrastmittel-Untersuchungen bei eingeschränkter ... · Die alternativ empfohlene kontrastmittelverstärkte MR-Angiographie (ceMRA) kann […]das gesamte Gefäßsystem […]](https://img.pdfslide.org/doc/110x75/5ccefd3b88c993b6568b9cd1/mr-kontrastmittel-untersuchungen-bei-eingeschraenkter-die-alternativ-empfohlene.jpg)
















