Embed Size (px)
Citation preview

Bayerische Akademie der Wissenschaften
36Ökologische Rolle der Flechten
Verlag Dr. Friedrich PfeilISSN 0938-5851 · ISBN 978-3-89937-096-6
Rundgesprächeder Kommission für Ökologie


129
Ökologie und Physiologie Borken bewohnender Flechten
Rundgespräche der Kommission für Ökologie, Bd. 36 »Ökologische Rolle der Flechten«, S. 129-141.© 2009 by Verlag Dr. Friedrich Pfeil, München – ISSN 0938-5851 – ISBN 978-3-89937-096-6
Ökologie und Physiologie Borken bewohnender Flechten
Michael Lakatos*, Britta Hartard* und Cristina Máguas**
Zusammenfassung
Flechten sind als poikilohydre, d. h. wechselfeuchte Organismen in der Lage, verschiedene Wasserquellen wie Regen, Nebel, Tau und Wasserdampf für ihre Photosynthese zu nutzen. In einem Fließgleichgewicht zwischen Austrocknung und Wiederbefeuchtung, Nährstoffaufnahme und -auswaschung sowie Photosynthese und Respiration stehen Flechten mit der unmittelbaren Umwelt in Wechselwirkung. Dabei verfügen sie über keine aktiven Kontrollmechanismen wie z. B. Spaltöffnungen zur Transpirationsverminderung und CO2-Aufnahme oder Wurzeln und Leitgefäße zur Wasserversorgung, sondern tauschen Ressourcen passiv über ihre gesamte Oberfläche aus. Während ihrer Stoffwechselaktivität akkumulieren sie aus der Umwelt aufgenommene Stoffe in ihrem organischen Material. Dies gilt nicht nur für umweltbelastende Substanzen wie SO2, Schwermetalle oder ein Übermaß an Stickstoff, sondern auch für CO2- und Wasserressourcen. Unsere Forschungsergebnisse zeigen u. a., dass sie sich insbesondere hinsichtlich der Austauschprozesse mit der Umgebungsfeuchte maßgeblich von Höheren Pflanzen unterscheiden. Im Zuge der aktuellen Klimaforschung ist es u. a. unabdingbar, die komplexen Mechanismen des Wasser- und Gasaustausches im Allgemeinen zu verstehen. Hierbei stößt man jedoch bei der Charakterisierung von Mikro- und Mesohabitaten sowie bei Langzeitmessungen häufig an apparative und ökonomische Grenzen. Des Weiteren sind in aktuellen Modellen insbesondere passive Gas- und Wasseraus-tauschprozesse von poikilohydren Organismen wie Flechten noch nicht berücksichtigt. Neueste Untersuchungen zeigen, dass die spezifische Zusammensetzung natürlich vorkommender stabiler Isotope (13C und 18O) in epiphytischen Flechten als Indikator für eine qualitative Bewertung von CO2- und H2O-Austauschprozessen dienen kann. Anhand eines von uns neu entwickelten Modells können die Mechanismen passiver Wasseraustauschprozesse biologischer Körper sehr präzise studiert werden. Unter Berücksichtigung der Fraktionierungsprozesse können mittels der Isotopensignatur von Flechten neue Erkenntnisse über CO2- und Wasserkreisläufe gewonnen und bestehende Modelle erweitert werden.
Summary
Ecology and physiology of lichens growing on barks. Lichens are poikilohydric organisms that have access to a variety of different water resources such as rain, fog, dew and water vapour to conduct photosynthesis. Lichens directly interact with their adjacent environment within an equilibrium between desiccation and rewetting, nutrient uptake and release, as well as photosynthesis and respiration. They do not feature active control mechanisms such as stomata to reduce the transpiration and to take up CO2 nor roots or vascular systems for permanent water supply. Instead, they passively exchange resources over their entire surface. During meta-bolic activity, they accumulate substances taken up from the environment in their organic material. This applies not only for polluting substances such as SO2, heavy metals or excess of nitrogen, but also for CO2 and water resources. Our investigations highlighted that, particularly with regard to the utilisation of water resources, li-
* Lakatos, Michael, Juniorprof. Dr.; Hartard, Britta, Dr., Technische Universität Kaiserslautern, FB Biologie, Abteilung Experimentelle Ökologie, Postfach 3049, 67653 Kaiserslautern. E-Mail: [email protected]
** Máguas, Cristina, Dr., Stable Isotope Laboratory, Centro de Ecologia e Biologia Vegetal, Faculdade de Ciên-cias, Universität Lissabon, Campo Grande, 1749-016, Lissabon, Portugal.

130
Ökologische Rolle der Flechten
Einführung
Eine der schwierigsten Aufgaben ist die Erfassung lokaler Langzeitveränderungen in der Umwelt: Wie verändern sich CO2 -Konzentrationen, Luft-feuchtigkeit und Wasserverfügbarkeit z. B. bei Veränderungen der Landnutzung , bei Waldfrag-mentierung, Wiederaufforstung oder Städtebau? Um diese Veränderungen beurteilen und ihnen mit geeigneten Maßnahmen begegnen zu können, ist ein Langzeitmonitoring erforderlich, das vor entsprechenden Eingriffen mit einem relativ ho-hen apparativen Aufwand durchgeführt werden muss. Meistens stehen diese Daten jedoch nicht zur Verfügung, um Änderungen wissenschaftlich nachvollziehen zu können. Lösungsansätze hier-zu bieten langsam wachsende Bioindikatoren , die aufgrund bestimmter Eigenschaften Umweltpa-rameter über längere Zeiträume integrieren und deren qualitative Änderungen widerspiegeln. Damit sind sie geeignet, schnell und präzise Aussagen über Umweltveränderungen zu tref-fen. Es wurden bereits Flechten vorgestellt, die Schwermetall - und SO2 -Belastungen sowie Stick-stoffeinträge widerspiegeln (vgl. Hauck (2009) in diesem Band). Bioindikatoren für die komplexen Vorgänge der CO2 - und Wasseraustauschprozesse zu finden, ist jedoch bisher mit großen geogra-phischen und organismischen Einschränkungen verbunden (z. B. Fletcher et al. 2005, Reyes-Garcia et al. 2008). Epiphytische und vor allem corticole, d.h. Borken bewohnende, Flechten können einen sol-chen Bioindikator darstellen, da sie hauptsächlich über atmosphärische Prozesse integrieren. Unsere aktuellen Forschungsergebnisse zeigen, warum und wie epiphytische Flechten als Indikatoren für spezifische Unterschiede in der CO2 -Konzentrati-on oder Wasserverfügbarkeit bei kleinräumigen bis regionalen Fragenstellungen genutzt werden
können. Vor allem CO2 und Wasser sind zwei wesentliche Faktoren, die im Kontext des glo-balen Wandels schwerwiegenden Änderungen unterworfen sind. Daher ist es von besonderer Bedeutung, für diese beiden komplexen Faktoren verlässliche Bioindikatoren zu entwickeln, die fehlende Langzeitmonitoring -Datensätze kom-pensieren und es ermöglichen, lösungsorientierte Maßnahmenkataloge umzusetzen.
Epiphytische Flechten
Epiphyten sind im Allgemeinen Organismen, die andere Pflanzen als Substrat nutzen, ohne aus ihnen Wasser oder Nahrung zu beziehen. Meist siedeln sie auf langlebigen oder sich langsam verändernden Substraten wie Borken, Nadeln oder immergrünen Blättern. In letzteren Fällen nennt man sie Epiphylle , während Organismen auf Borken als corticol bezeichnet werden. Das Interessante an corticolen Epiphyten ist, dass sie kaum von Bodenprozessen, sondern viel mehr von atmosphärischen Prozessen beeinflusst werden. Corticole Flechten finden wir in verschiedenen Wuchsformen : als crustose (Krusten bildende ), foliose (blattartige ) oder fruticose (strauchartige ) Flechten. Auf einem Baum treten diese unter-schiedlichen Formen in verschiedenen Assoziatio-nen auf. Die Vergesellschaftung hängt dabei sehr stark von den mikroklimatischen Unterschieden entlang der verschiedenen Baumzonierungen ab. In den humiden Tropen, in denen die klimatischen Unterschiede zwischen Kronen- und Bodenregi-on extrem groß sind, lassen sich Bäume z. B. in 6 verschiedene Zonie rungen aufteilen (Johansson 1974, siehe auch Zotz 2007). Diese Zonierun-gen unterscheiden sich z.B. hinsichtlich Licht , Wind und relativer Feuchtigkeit und bedingen unterschiedliche Austrocknungsereignisse der
chens follow an alternative strategy in comparison to vascular plants. Within the scope of current investigations on how to counteract climate change, it is important to understand the complex water and gas exchange mech-anisms. However, the characterisation of micro- and macrohabitats as well as long-term changes requires an excessive application of apparative and economic investment. Moreover, passive gas and water exchange processes, as exhibited by cryptogams, are not yet considered in current flux models. Recent studies indicate the suitability of naturally occurring compositions of stable isotopes (13C and 18O) of epiphytic lichens to be used as indicators for a qualitative evaluation of CO2 and water exchange processes. On the basis of our novel model the mechanisms of passive water exchange processes of cryptogams can be studied in more detail. Thus, with the knowledge of the respective isotopic fractionation processes, the isotope signature of lichens can be used to gain new insights into CO2 and water circulations which, prospectively, may be incor-porated in existing flux models.

131
Flechten. Im oberen Kronenraum führen hohe Lichteinstrahlung, starke Windturbulenzen und niedrige Luftfeuchte dazu, dass Flechten regelmä-ßig schnell austrocknen, während sie im unteren Bereich bei geringem Lichteinfall, schwachen Windturbulenzen und permanent hoher Luft-feuchte nur langsam austrocknen und somit lange physiologisch aktiv bleiben (Lakatos et al. 2006). In den unterschiedlichen Zonierungen wurden in den Tropen auf nur einem Baum bereits 173 ver-schiedene Flechtenarten gezählt (Aptroot 1997). Epiphytische Flechten finden wir aber nicht nur in den Tropen, sondern überall dort, wo es Bäume und Sträucher gibt . In den Tropen ist ihre Biomasse mit ca. 100 kg Trockengewicht pro Hektar im Tiefland (z. B. Freiberg & Freiberg 2000) und über 1300 kg/ha in den montanen Regenwäldern (Dalling 1994) vergleichsweise niedrig. In temperaten und borealen Gebieten weisen Flechten habitatsabhängig eine Biomasse mit Spitzen von 2000 bis 3000 kg/ha auf (Rhoades 1981, Pypker et al. 2006). Mit dieser hohen Abun-danz bieten sie u. a. wichtige Lebensräume und Nahrungsquellen für andere Organismen (vgl. Segerer (2009) in diesem Band). Aufgrund ihres häufigen Auftretens lassen sich epiphytische Flechten daher gut als Bioindikatoren verwenden. Bei mikroskalierten Vorgängen und Mechanismen innerhalb der Kronenräume sind allerdings noch viele Fragen offen, welche Kurzzeit- und Lang-zeitmessungen nicht nur im Tieflandregenwald und im temperaten Wald erfordern.
Stabile Isotope als physiologische und ökologische Werkzeuge
Stabile Isotope stellen ein sehr interessantes Forschungsinstrument dar, das zur Untersu-chung sowohl kurzzeitiger und kleinräumiger molekularphysiologischer Vorgänge als auch lang andauernder, großräumiger ökologischer Prozesse verwendet werden kann. Die Ergebnisse können dabei vom Blatt zur Pflanze, weiter zum Ökosystem, auf die Region und bis auf globale Ebenen skaliert werden. Stabile Isotope sind verschiedene Formen des-selben Elements mit gleicher Anzahl von Protonen und Elektronen, aber unterschiedlicher Neutro-nenzahl. Im Gegensatz zu radioaktiven Isotopen sind sie dauerhaft stabil und zerfallen nicht. In der Natur kommen stabile Isotope verschiedener Elemente in unterschiedlichen Häufigkeiten vor (Tab. 1). Ihr Massenunterschied kann zur Identi-fizierung und Untersuchung natürliche Prozesse verwendet werden. Die Isotopenhäufigkeit einer Probe wird dabei durch die so genannte Delta-Notation beschrieben:
⎧ 13C/12C Probe ⎫ δ13C = ⎪ ————————— − 1 ⎪ · 1000 [‰] ⎩ 13C/12C Standard ⎭Das Isotopenverhältnis (z. B. 13C/12C, 18O/16O) einer Probe wird dabei relativ zu einem inter-nationalen Standard dargestellt, für δ13C ist dies z. B. die 13C/12C-Ratio eines fossilen Belemniten der PeeDee-Kalkformation aus North Carolina
Michael Lakatos et al.
13C
12C
δ13C: -8 ‰CO2 der Luft
δ13C: -8 ‰CO2 der Luft
Stomata
Δ13C: 4,4 ‰Δ13
Enzym
RubisCO
Δ13C: 30 ‰
EnzymEnzymδ13C: -28 ‰
C im org. Material
Transport
Δ13C: keine
35%65%
C3:
C4/CAM/CCM:
13C
12C
Stomata
δ13C: -20 ‰
Transport
Δ13C: keine
Enzym
PEPC/CCM
Δ13C: 5,5 ‰
Enzym
RubisCO
Δ13C: 30 ‰
Enzym
Δ
Enzym
Δ
Enzym
Δ
Enzym
Δ
EnzymEnzym
ΔΔ13C: 4,4 ‰ΔΔΔΔ
EnzymEnzymEnzymEnzym
Abb. 1. Isotopendiskriminierung bei wesentlichen Prozessen der CO2-Carboxylierung bei C3-Pflanzen und bei Pflanzen mit C4-Photosynthese, CAM (Crassulaceen Acid Metabolism) oder CCM (CO2 -Konzentrierungsmecha-nismus); Erläuterungen siehe Text S. 132.
Ökologie und Physiologie Borken bewohnender Flechten
132
Ökologische Rolle der Flechten
(V-PDB), für δ18O hingegen ist es Meerwasser (Standard Mean Ocean Water, SMOW). Aufgrund ihrer unterschiedlichen Massen und der Symmetrie der Atomkerne und Elektro-nenhüllen, unterliegen schwere Isotope eines Ele-ments einer so genannten Isotopenfraktionierung. Dieser Fraktionierung liegen verschiede Effekte zugrunde:– Bei den kinetischen Isotopeneffekten handelt
es sich entweder um physikalische Vorgänge (z. B. das schwerere Isotop diffundiert lang-samer als das leichtere) oder um biochemische Vorgänge (z. B. ein Enzym diskriminiert eines der beiden Isotope).
– Thermodynamische oder Gleichgewichts-(Äquilibrierungs-)Isotopeneffekte treten z. B. auf, wenn Wasser in die Gasphase oder wenn CO2 in HCO3
– übergeht.Wenn wir diese der Isotopenfraktionierung zu-grunde liegenden Prozesse kennen, können wir stabile Isotope als natürliche Marker verwenden,
um biochemische Prozesse in der Natur zu cha-rakterisieren.
Diskriminierung von CO2 bei der Carboxylierungsreaktion
Bei der Carboxylierungsreaktion , wie sie in C3-Pflanzen und in Pflanzen mit C4-Photosynthese oder CAM (Crassulaceen Acid Metabolism) auf-tritt, kommt es an verschiedenen Stellen des Stoffwechselweges zu Isotopendiskriminierungen (Abb. 1). Das CO2 der Luft besitzt typischerweise ein δ13C von –8 ‰. Im Blatt kommt es bei der Dif-fusion in den substomatären Raum1 zu einer phy-sikalischen Diskriminierung von 4 ‰ aufgrund des Diffusionswiderstandes (Craig 1954). Auch das Eingangsenzym Ribulose-1,5-bisphosphat-carboxylase/-oxygenase (RubisCO; EC 4.1.1.39), welches das CO2 fixiert, diskriminiert mit ca. 30 ‰ relativ stark gegen das schwere 13CO2 (Farquhar & Richards 1984). Diese Prozesse sind jedoch nicht additiv, da 65 % der im substomatären Raum befindlichen CO2-Moleküle nicht fixiert werden und nur 35 % in organisches Material umgewan-delt werden. Beim Transport nach der Fixierung kommt es zu keiner weiteren Diskriminierung mehr, sodass die Isotopensignatur des gesamten organischen Materials von C3-Pflanzen ungefähr bei –28 ‰ liegt. Bei C4- und CAM-Pflanzen diskriminiert das CO2-vorfixierende Enzym, die Phosphoenolpyruvatcarboxylase (PEPC; EC 4.1.1.31), geringer gegen 13C. Erst anschließend wird das vorfixierte CO2 durch die nachgeschal-tete RubisCO letztendlich fixiert. Dadurch steigt anteilig die Fixierungsrate der im substomatären Raum befindlichen CO2-Moleküle. Dies führt zu einer Verringerung der Gesamtdiskriminierung gegenüber C3-Pflanzen, sodass das organische Material mit einer Isotopensignatur von –20 ‰ weniger abgereichert ist. Ähnliche Effekte kennen wir auch bei Flech-ten, deren Photobiont (Cyanobakterien und Grünalgen) einen so genannten CO2 -Konzen-trierungsmechanismus (CCM) besitzt (Máguas et al. 1995). Das CO2 wird dabei über spezielle Transporter ins Zellinnere aufgenommen, in Bicarbonat (HCO3
–) umgewandelt und in die
1 Bereich unterhalb der regulierbaren Spaltöffnungen (Stomata ) von (Blatt-)Oberflächen, in dem der Gasaustausch mit dem umliegenden pflanzlichen Gewebe stattfindet.
δ13C des organischen Materials [‰]-35 -30 -25 -20 -15 -10
rela
tive
Häu
figke
it [ %
]
30
25
20
15
10
5
0
Photobiont ohne CCM Cyanobakterien mit CCM Grünalgen mit CCM
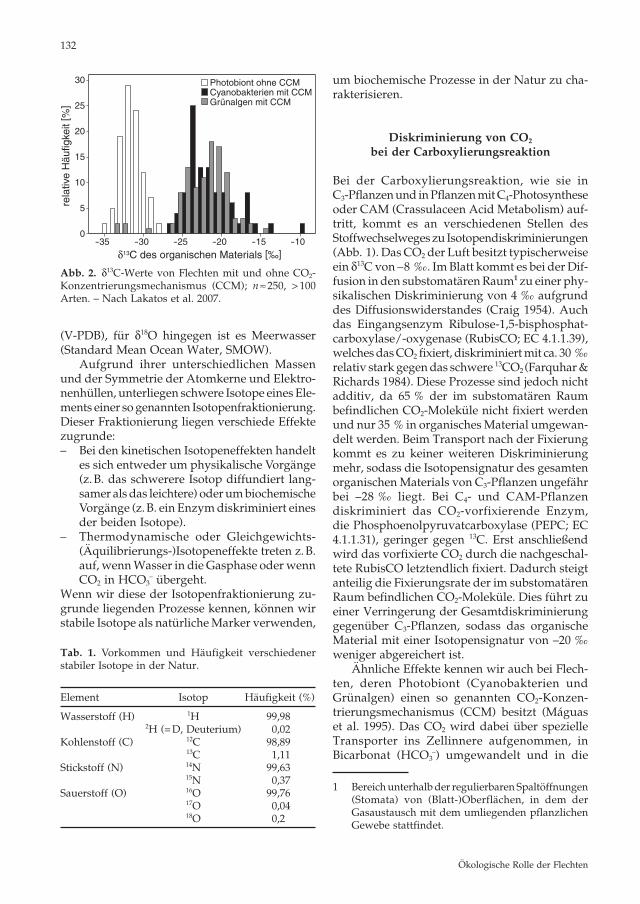
Abb. 2. δ13C-Werte von Flechten mit und ohne CO2-Konzentrierungsmechanismus (CCM) ; n ≈ 250, > 100 Arten. – Nach Lakatos et al. 2007.
Tab. 1. Vorkommen und Häufigkeit verschiedener stabiler Isotope in der Natur.
Element Isotop Häufigkeit (%)
Wasserstoff (H) 1H 99,98 2H (= D, Deuterium) 0,02Kohlenstoff (C) 12C 98,89 13C 1,11Stickstoff (N) 14N 99,63 15N 0,37Sauerstoff (O) 16O 99,76 17O 0,04 18O 0,2

133
Carboxysome bzw. Pyrenoide zur dort befind-lichen RubisCO transportiert (Smith & Griffiths 1996). Auch Hornmoose besitzen Pyrenoide und einen CCM (Smith & Griffiths 2000). Die Carboxysome und Pyrenoide sind jeweils aus Protein - bzw. Stärkeschalen aufgebaut und stellen eine zusätzliche CO2 -Diffusionsbarriere dar, die den CO2-Verlust reduziert. Innerhalb der Flechten zeigen sich daher deutliche Unterschiede zwi-schen solchen, deren Photobionten keinen CCM aufweisen (Mittel: δ13C –32 ‰), und denjenigen, die Cyanobakterien oder Grünalgen mit CCM beinhalten (Mittel: δ13C –22 ‰; Abb. 2).
CO2-Fixierung im Mikroökosystem
Die Heterogenität innerhalb der Flechtengrup-pen mit und ohne CCM ist allerdings recht groß. Einerseits spielen hierbei vom Thallusbau abhängige CO2 -Diffusionsprozesse (Máguas & Brugnoli 1996) sowie häufig auftretende Limi-tierungen der Stoffwechselprozesse von Flech-ten z. B. durch Licht - oder Wassermangel eine Rolle (vgl. Green (2009) in diesem Band). Auf der anderen Seite muss jedoch auch die von der Flechte aufgenommene CO2-Quelle mit in Betracht gezogen werden. Erste Hinweise auf einen Einfluss der Exposition hinsichtlich un-terschiedlicher CO2 -Konzentrationen ergaben sich bei einem Vergleich der terricolen (boden-bewohnenden) Rentierflechten Cladonia stellaris , C. arbuscula und C. submitis sowie der corticolen Bartflechte Usnea filipendula (Abb. 3). Alle vier Arten stammen aus einem mediterranen Sand-dünenhabitat und weisen einen Grünalgenpho-tobionten mit CCM auf. Die δ13C-Werte der vier Arten weisen signifikante Unterschiede zwischen –20,7 und –25,5 ‰ auf. In einer mikroskalierten Profiluntersuchung wurden CO2-Proben aus und über dem Boden sowie in Flechtenpolstern und oberhalb dieser Polster entnommen und deren CO2-Konzentration und δ13C-Signatur analysiert. Es zeigte sich ein linearer Zusammenhang zwi-schen CO2-Konzentration und Isotopensignal. Das heißt, dass bodenwüchsige Flechten das CO2 sowohl aus der Bodenrespiration (ca. –14 ‰) als auch aus der Umgebungsluft (ca. –8 ‰) fixieren, während luftexponierte Flechten nur das CO2 der Umgebungsluft aufnehmen (Abb. 4). Ähnliche Ergebnisse wurden bei corticolen Flechten im tropischen Tieflandregenwald in Panama ge-funden. Auch innerhalb des Waldes findet man
in der Regel einen Gradienten unterschiedlicher CO2-Konzentrationen . Im Unterwuchs liegen die δ13C-Werte des CO2 bei –11 ‰, im mittleren Stammbereich hingegen bei –8 ‰ (Sternberg et al. 1989). Entsprechend wurden auch bei der Ana-lyse der stabilen Isotopensignatur zweier sowohl bodennah als auch luftexponiert wachsender corticoler Flechtenarten signifikante Unterschie-de gefunden (–33 ‰ im Bodenbereich, –30 bzw. –31 ‰ am mittleren Stammbereich). Auch dies weist darauf hin, dass CO2 mit unterschiedlicher Isotopensignatur fixiert wird. In einer weiteren Studie im Tieflandregenwald von Französisch-Guayana wurden die drei eng anliegenden crustosen Flechtenarten Thelotre-ma alboolivaceum , Herpothallon rubrocinctum und Phyllopsora corallina sowie die abstehende Flechte Coenogonium linkii untersucht (Lakatos et al. 2006), die häufig nebeneinander auf der Borke vorkommen (Abb. 5). Im Unterwuchs sind hier lediglich schwache Turbulenzen zu finden, d. h. das CO2, das aus der Rinde respiriert wird und einen abgereicherten δ13C-Wert von ungefähr –29 ‰ besitzt, verbleibt zunächst in der borken-nahen Grenzschicht, bevor es sich anschließend mit der umgebenden Luft (δ13C-Wert: –9 ‰) vermischt. Die eng an die Borke gepressten crustosen Flechten weisen ebenfalls einen abge-reicherten δ13C-Wert von –33 ‰ auf, während die exponierte Flechte, die neben dem CO2 aus der Rindenrespiration auch das der umgebenden Luft aufnimmt , einen Wert von –31 ‰ besitzt. In Abbildung 6 sind diese und weitere Daten aus den unterschiedlichsten Habitaten zusammengestellt (Lakatos et al. 2007). Hierbei kann allgemein un-terschieden werden zwischen Flechten, die dem CO2 aus der Rindenrespiration (δ13C ~ –30 ‰), der Bodenrespiration (δ13C ~ –16 ‰) oder der Umgebungsluft (δ13C ~ –8 ‰) ausgesetzt sind, sowie zwischen Flechten mit und ohne CO2-Kon-zentrierungsmechanismus . Flechten ohne CCM hatten im Median δ13C-Signaturen von jeweils –32, –31 und –27 ‰ und weisen damit eine weitaus deutlichere Inkorporation der beeinflussenden 13CO2-Quelle in ihrem organischen Material auf als diejenigen mit CCM (ca. –23 ‰). Es besteht somit die Möglichkeit, Flechten ohne CCM als mikro-skalierte Indikatoren für kleinräumige und regio-nale Veränderungen über lange Zeiträume zu verwenden (Lakatos et al. 2007). Untersuchungen derselben Flechtenart aus verschiedenem Herbar-material weisen außerdem auf die Anwendbarkeit von Flechten als Langzeitindikator über mehrere
Ökologie und Physiologie Borken bewohnender Flechten

134
Ökologische Rolle der Flechten
a b
Abb. 3. Usnea filipendula (a) und Cladonia arbuscula (b) aus dem Untersuchungsgebiet, einem Sanddünenhabitat in Portugal. – Fotos: M. Lakatos.
C. stellaris C. arbuscula C. submitis
U. filipendula
-25 ‰ -24,3 ‰
-20,7 ‰
-25,5 ‰
Photobiont: Trebouxia mit CCM: δ13C >-20 ‰
(terricol)
(corticol)
Umgebungs-CO2: -8 ‰
Respirations-CO2:ca. -14 ‰
Boden < -16 ‰
CO2
Abb. 4. Zusammenhang zwischen der Isotopensignatur der CO2-Quellen (CO2 aus der Bodenrespiration und aus der Umgebungsluft) und der Isotopensignatur im organischen Material der entsprechend exponierten Flechten Usnea filipendula (luftexponiert) und dreier Cladonia-Arten (bodenexponierte Flechtenpolster).
135
a b
c d
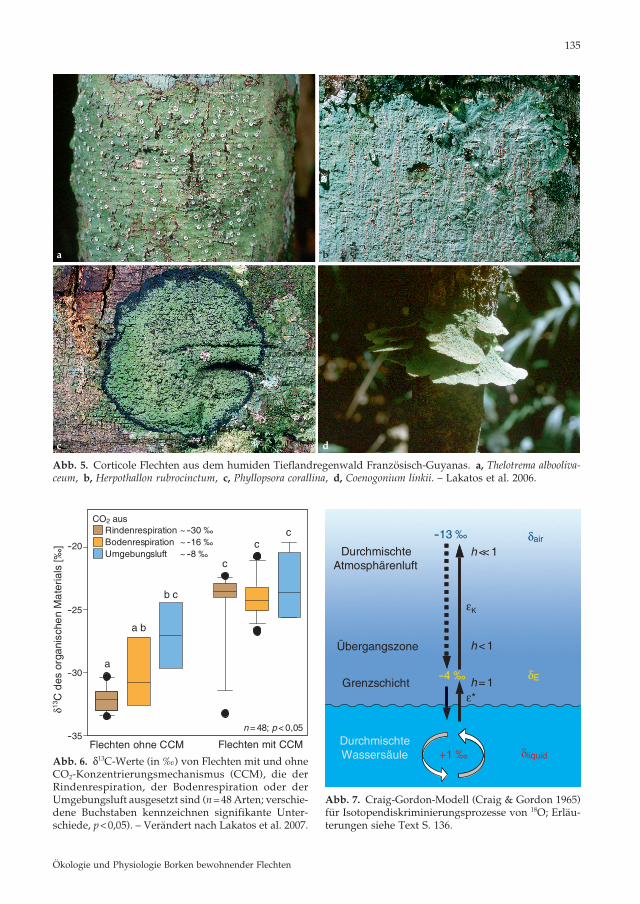
Abb. 5. Corticole Flechten aus dem humiden Tieflandregenwald Französisch-Guyanas. a, Thelotrema albooliva-ceum , b, Herpothallon rubrocinctum , c, Phyllopsora corallina , d, Coenogonium linkii . – Lakatos et al. 2006.
Abb. 7. Craig-Gordon-Modell (Craig & Gordon 1965) für Isotopendiskriminierungsprozesse von 18O; Erläu-terungen siehe Text S. 136.
Durchmischte Wassersäule
Grenzschicht
Übergangszone
Durchmischte Atmosphärenluft
h = 1
h < 1
h << 1
δliquid
δE
ε*
εK
δair
-4 ‰
-13 ‰
+1 ‰ Abb. 6. δ13C-Werte (in ‰) von Flechten mit und ohne CO2-Konzentrierungsmechanismus (CCM ), die der Rindenrespiration , der Bodenrespiration oder der Umgebungsluft ausgesetzt sind (n = 48 Arten; verschie-dene Buchstaben kennzeichnen signifikante Unter-schiede, p < 0,05). – Verändert nach Lakatos et al. 2007.
n = 48; p < 0,05-35
-30
-25
-20
Flechten ohne CCM Flechten mit CCM
a
a b
b c
c
cc
δ13C
des
org
anis
che
n M
ate
rials
[‰]
Rindenrespiration ~-30 ‰Bodenrespiration ~-16 ‰Umgebungsluft ~-8 ‰
CO2 aus
Ökologie und Physiologie Borken bewohnender Flechten

136
Ökologische Rolle der Flechten
Jahrzehnte hin. Die Ergebnisse zeigen, dass die in knapp 150 Jahren stattgefundene Abweichung der atmosphärischen CO2-Isotopensignatur (von 1846: –6,5 ‰ auf 1989: –8,0 ‰) sich tatsächlich in den untersuchten Signaturen der Flechten thalli widerspiegelt (1848: –29,3 ‰; 1989: –30,6 ‰; Máguas & Brugnoli 1996). Das bedeutet, dass Flechten über Mikrohabitatsbedingungen der Vergangenheit Aufschluss geben können und dass das δ13C ihres organischen Materials im Speziellen als Indikator für mikroskalierte CO2-Prozesse verwendet werden kann.
Nutzung unterschiedlicher Wasserressourcen
Der Wasserdampf ist ein maßgeblicher, jedoch wenig verstandener Bestandteil des Wasser-kreislaufs. Konventionell lässt sich die Wasser-dampfmenge in der Luft als Luftfeuchtigkeit messen. Die Isotopensignatur lässt jedoch quali-tativ Rückschlüsse zu, ob diese Luftfeuchtigkeit überwiegend aus Evaporationsprozessen des Bodens, aus Transpirationsprozessen der Pflanzen oder aus dem Regenwasser stammt. Zeitliche und räumliche Verlaufsmuster der Zusammen-setzung dieses Wasserdampfes wurden bisher sehr selten untersucht. So bestehen teilweise Probleme, die einzelnen Prozesse adäquat zu erfassen. Hierbei stellt sich auch die Frage, welche Wasserressourcen Flechten wie und wann nutzen können. Es ist bekannt, dass sie Niederschläge in unterschiedlicher Form, sei es als Regen, Taufall oder Nebel , nutzen können und dass sie auch Bodenwasser, kapillares Wasser oder Stammab-flusswasser aufnehmen können (z. B. Lange et al. 1986, 2001). Wie kann jedoch zwischen den unterschiedlichen Wasserressourcen, die Flech-ten und auch andere Mikroorganismen nutzen, differenziert werden? Hier spielt die Isotopensignatur des 18O eine wichtige Rolle. 18O unterliegt ebenso wie 13C ver-schiedenen Diskriminierungsprozessen (Abb. 7). Wenn Wasser über einer offenen Wasserfläche evaporiert (Übergang flüssig – gasförmig) findet eine Äquilibrierungsfraktionierung (ε*) statt, sodass die Luftfeuchtigkeit im Vergleich zum flüssigen Wasser stets an 18O abgereichert ist. Entfernt sich die Luft von der Wasserfläche, finden sowohl Äquilibrierungs- als auch kine-tische (εk) Diffusionsprozesse statt, welche den Wasserdampf weiter abreichern, bis sich in der
gut durchmischten Luftsäule der Atmosphäre Werte von ca. –13 ‰ einstellen. Die Formel, die den gesamten Prozess integriert, lautet:
δliquid – hδair – ε* – εk δE ≈ ——————————— ≈ –4 ‰ 1 – h
mit h = relative Luftfeuchtigkeit . Dieses so ge-nannte Craig-Gordon-Modell (Craig & Gordon 1965) ist die Basis für sämtliche in der Natur stattfindende Wasseraustauschprozesse .
Wasseraustauschprozesse in Flechten
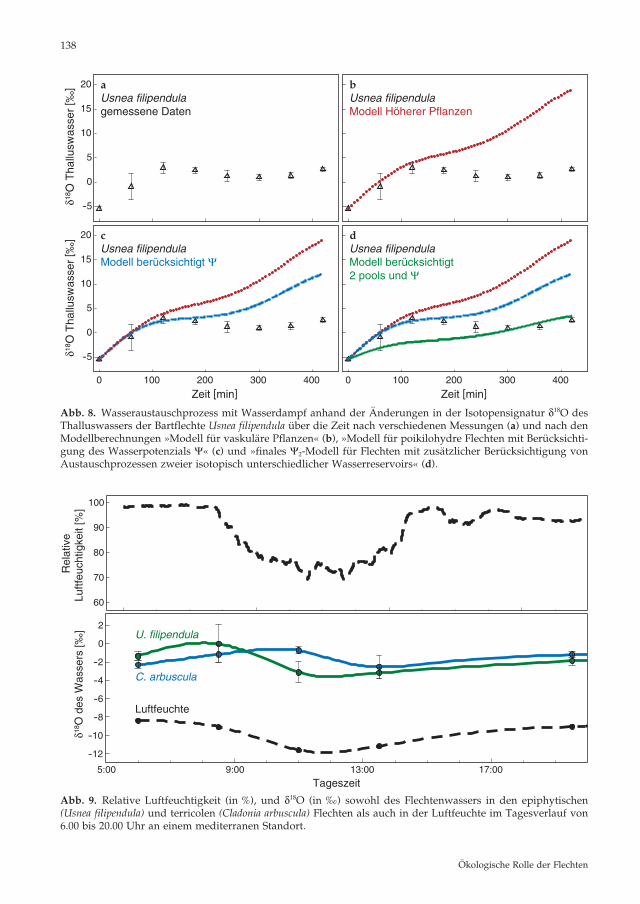
Inwieweit lässt sich jedoch das ursprüngliche Craig-Gordon-Modell, welches die Evaporation an Wasseroberflächen beschreibt, auch für Was-seraustauschprozesse in Flechten anwenden, bzw. welche komplexeren Zusammenhänge müssen dabei zusätzlich berücksichtigt werden? Um zu untersuchen, ob es einen Austauschpro-zess zwischen der Luftfeuchtigkeit und dem angefeuchteten Thallus gibt, wurden feuchte Thalli von Cladonia arbuscula in einer gasdichten Kammer unter kontrollierten Temperatur - und Luftfeuchtigkeitsbedingungen einem Wasser-dampf mit stark abgereicherter Isotopensignatur ausgesetzt. Die Isotopensignatur δ18O sowohl im Thalluswasser als auch in der Luftfeuchte wur-de kontinuierlich bestimmt. Das angereicherte Thalluswasser äquilibrierte in wenigen Stunden mit der Luftfeuchte in der Kammer und erlangte schließlich die abgereicherte Isotopensignatur der Luftfeuchte. Aufgrund der schnellen isoto-pischen Äquilibrierung lassen diese Ergebnisse daher den Schluss zu, dass Flechten(wasser) als Indikator für die Isotopensignatur des Wasser-dampfs verwendet werden kann. Zum Vergleich: Eine Äquilibrierung zwischen dem Blattwasser Höherer Pflanzen und der Luftfeuchte findet im Allgemeinen aufgrund der typischen Blattwas-seranreicherung nicht statt. Wie funktioniert dieser schnelle Austausch von Thalluswasser bei Flechten? Das Transpi-rationsmodell für Höhere Pflanzen (Farquhar & Cernusak 2005, Helliker & Griffiths 2007) trifft auf die gemessene Flechtenevaporation nur unzu-reichend zu (Abb. 8a,b). Einer der maßgeblichen Parameter in dem zugrunde liegenden Modell ist die relative Luftfeuchtigkeit h. Das Transpirati-onsmodell geht davon aus, dass aufgrund der kontinuierlichen Wasserzufuhr und der stoma-tären Regulierung die relative Luftfeuchtigkeit h

137
im substomatären Raum Höherer Pflanzen immer bei ca. 100 % liegt. Dies trifft bei Flechten und vielen anderen poikilohydren Organismen aber nicht zu, da hier das Wasserpotenzial des Thallus (Ψ) – und somit auch die unmittelbar darüberlie-gende, mit Ψ assoziierte Luftfeuchte – stark vom Wassergehalt der Flechte abhängt. Flechten haben außerdem hygroskopische Eigenschaften, sodass in ihrem Thallus Bereiche mit unterschiedlichem Wasserpotenzial und unterschiedlichen Evapo-rationsraten zu finden sind. Somit beeinflusst auch die Umgebungsfeuchtigkeit das Wasser-potenzial des Thallus. Die Luftfeuchtigkeit h an der unmittelbaren Thallusoberfläche ist demnach nicht konstant ca. 100 %, sondern muss stets in Relation zum Wasserpotenzial der Flechte be-rechnet werden (Hartard et al., submitted). Das Wasserpotenzial des Flechtenthallus lässt sich mit so genannten Druck-Volumen-Kurven kalkulie-ren (Proctor et al. 1998, Beckett 1997), während Wasserpotenzial und relative Umgebungsfeuchte durch die Kelvin-Gleichung zueinander in Bezug stehen. Berücksichtigt man diese Abhängigkeiten, so verbessert sich das beschreibende Modell (Abb. 8c). Dennoch verbleibt eine Diskrepanz zwischen modellierten und gemessenen Werten. Woran kann das liegen? In dem Beitrag »Ökologische Aspekte der Wechselbeziehung zwischen Pilz und Alge« wurde bereits darauf hingewiesen, wie verschie-den und komplex morphologische Strukturen in Flechten aufgebaut sind (Honegger (2009) in diesem Band). Die verschiedenen Strukturen unterscheiden sich im Hinblick auf Gasaustausch , Wasserhaltekapazität und Wasserleitfähigkeit (Blum 1973, Rundel 1988). Die Hyphen sind teilweise dichter oder lockerer gepackt und verschiedene Thallusschichten sind mehr oder weniger der Umgebungsfeuchtigkeit ausgesetzt. Daher liegt die Vermutung nahe, dass inner-halb der Flechten verschiedene Wasserreservoirs vorhanden sind, die aufgrund ihrer morpholo-
gischen Struktur entweder »progressiv« oder »konservativ« evaporieren (z. B. Goebel 1926, Jahns 1984, Honegger 1995, Scheidegger et al. 1995, Büdel & Scheidegger 1996) und daher ganz unterschiedliche Isotopenzusammenset-zungen besitzen. Innerhalb eines Zentralstrangs (z. B. bei Usnea filipendula ) kann beispielsweise davon ausgegangen werden, dass Wasser kaum evaporiert bzw. in Kontakt zur evaporierenden Oberfläche tritt, während es im äußeren Bereich zu einem stärkeren Wasseraustausch mit dem Wasserdampf der Außenluft kommt. Wir haben vier morphologisch unterschiedliche Flechten vermessen und ermittelt, welcher strukturelle Anteil der Flechten an den unmittelbaren Eva-porationsprozessen (exponierte Schichten) und welcher indirekt am Austausch mit dem Wasser der Außenluft (introvertierte Schichten) beteiligt ist (Tab. 2). Werden diese Gewichtungen in das Modell integriert, ergeben sich relativ gute Werte für die Simulation im Vergleich zur Mes-sung (Abb. 8d). Auf dieser Basis haben wir ein neues Modell (Ψ2-Modell) entwickelt, welches isotopische Austauschprozesse von Wasser in Flechten beschreibt (Hartard et al., submitted). Es kann auf die physiologisch-physikalischen Austauschprozesse aller bisher von uns unter-suchten epiphytischen und terricolen Flechten angewendet werden. Mit dem Ψ2-Modell konnten wir relevante Mechanismen aufzeigen, welche im Anschluss an weitere Untersuchungen zukünftig auch in andere Modelle, z. B. in ökosystemare Wasserzirkulationsmodelle , integriert werden können.
Differenzierung von Wasserressourcen
In Freilanduntersuchungen wurden parallel zur relativen Luftfeuchtigkeit und zu deren Isotopen-signatur die Wassersignaturen der epiphytischen Flechte Usnea filipendula und der terricolen Flechte
Tab. 2. Geschätzte Anteile verschiedener Flechtenthalli, die am unmittelbaren Wasseraustauschprozess (expo-nierte Schichten) und nur indirekt am Austausch mit dem Wasser der Außenluft (introvertierte Schichten) be-teiligt sind (nach Hartard et al. submitted).
Flechtenart Lebensweise exponierte Schichten introvertierte Schichten
Parmelia caperata corticol 20 % 80 %Usnea filipendula corticol 70 % 30 %Cladonia convoluta terricol 30 % 70 %Cladonia arbuscula terricol 50 % 50 %
Ökologie und Physiologie Borken bewohnender Flechten
138
Ökologische Rolle der Flechten
Rel
ativ
e Lu
ftfeu
chtig
keit
[%]
60
70
80
90
100
Tageszeit5:00 9:00 13:00 17:00
-12
-10
-8
-6
-4
-2
0
2
δ18O
des
Was
sers
[‰]
Luftfeuchte
C. arbuscula
U. filipendula
Abb. 8. Wasseraustauschprozess mit Wasserdampf anhand der Änderungen in der Isotopensignatur δ18O des Thalluswassers der Bartflechte Usnea filipendula über die Zeit nach verschiedenen Messungen (a) und nach den Modellberechnungen »Modell für vaskuläre Pflanzen« (b), »Modell für poikilohydre Flechten mit Berücksichti-gung des Wasserpotenzials Ψ« (c) und »finales Ψ2-Modell für Flechten mit zusätzlicher Berücksichtigung von Austauschprozessen zweier isotopisch unterschiedlicher Wasserreservoirs « (d).
Abb. 9. Relative Luftfeuchtigkeit (in %), und δ18O (in ‰) sowohl des Flechtenwassers in den epiphytischen (Usnea filipendula ) und terricolen (Cladonia arbuscula ) Flechten als auch in der Luftfeuchte im Tagesverlauf von 6.00 bis 20.00 Uhr an einem mediterranen Standort.
δ18O
Tha
llusw
asse
r [ ‰
]δ18
O T
hallu
swas
ser
[ ‰]
-5
0
5
10
15
20 aUsnea filipendulagemessene Daten
bUsnea filipendulaModell Höherer Pflanzen
Zeit [min]0 100 200 300 400
-5
0
5
10
15
20
Zeit [min]0 100 200 300 400
dUsnea filipendulaModell berücksichtigt 2 pools und Ψ
cUsnea filipendulaModell berücksichtigt Ψ
139
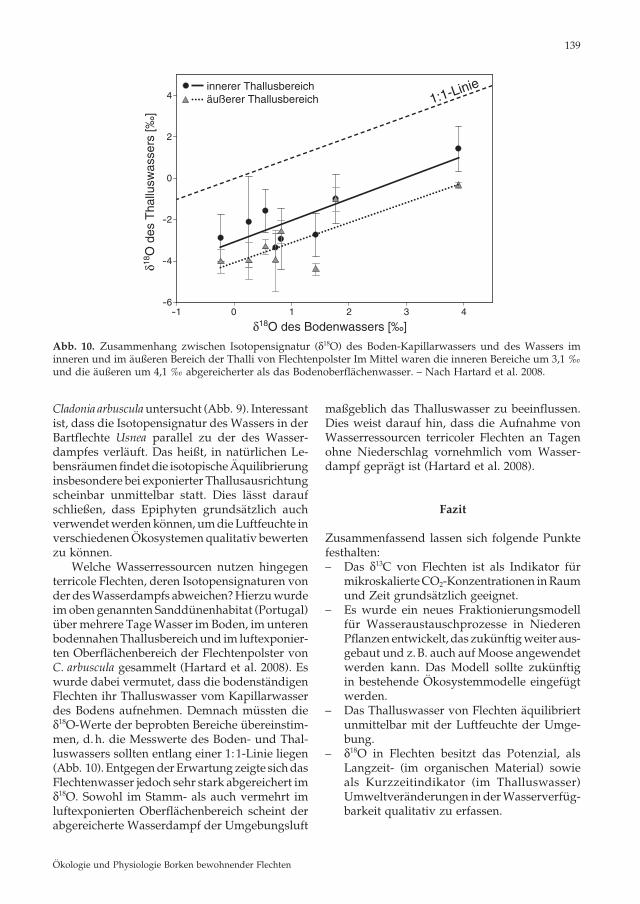
Cladonia arbuscula untersucht (Abb. 9). Interessant ist, dass die Isotopensignatur des Wassers in der Bartflechte Usnea parallel zu der des Wasser-dampfes verläuft. Das heißt, in natürlichen Le-bensräumen findet die isotopische Äquilibrierung insbesondere bei exponierter Thallusausrichtung scheinbar unmittelbar statt. Dies lässt darauf schließen, dass Epiphyten grundsätzlich auch verwendet werden können, um die Luftfeuchte in verschiedenen Ökosystemen qualitativ bewerten zu können. Welche Wasserressourcen nutzen hingegen terricole Flechten, deren Isotopensignaturen von der des Wasserdampfs abweichen? Hierzu wurde im oben genannten Sanddünenhabitat (Portugal) über mehrere Tage Wasser im Boden, im unteren bodennahen Thallusbereich und im luftexponier-ten Oberflächenbereich der Flechtenpolster von C. arbuscula gesammelt (Hartard et al. 2008). Es wurde dabei vermutet, dass die bodenständigen Flechten ihr Thalluswasser vom Kapillarwasser des Bodens aufnehmen. Demnach müssten die δ18O-Werte der beprobten Bereiche übereinstim-men, d. h. die Messwerte des Boden- und Thal-luswassers sollten entlang einer 1:1-Linie liegen (Abb. 10). Entgegen der Erwartung zeigte sich das Flechtenwasser jedoch sehr stark abgereichert im δ18O. Sowohl im Stamm- als auch vermehrt im luftexponierten Oberflächenbereich scheint der abgereicherte Wasserdampf der Umgebungsluft
maßgeblich das Thalluswasser zu beeinflussen . Dies weist darauf hin, dass die Aufnahme von Wasserressourcen terricoler Flechten an Tagen ohne Niederschlag vornehmlich vom Wasser-dampf geprägt ist (Hartard et al. 2008).
Fazit
Zusammenfassend lassen sich folgende Punkte festhalten:– Das δ13C von Flechten ist als Indikator für
mikroskalierte CO2 -Konzentrationen in Raum und Zeit grundsätzlich geeignet.
– Es wurde ein neues Fraktionierungsmodell für Wasseraustauschprozesse in Niederen Pflanzen entwickelt, das zukünftig weiter aus-gebaut und z. B. auch auf Moose angewendet werden kann. Das Modell sollte zukünftig in bestehende Ökosystemmodelle eingefügt werden.
– Das Thalluswasser von Flechten äquilibriert unmittelbar mit der Luftfeuchte der Umge-bung.
– δ18O in Flechten besitzt das Potenzial, als Langzeit- (im organischen Material) sowie als Kurzzeitindikator (im Thalluswasser) Umweltveränderungen in der Wasserverfüg-barkeit qualitativ zu erfassen.
Abb. 10. Zusammenhang zwischen Isotopensignatur (δ18O) des Boden-Kapillarwassers und des Wassers im inneren und im äußeren Bereich der Thalli von Flechtenpolster Im Mittel waren die inneren Bereiche um 3,1 ‰ und die äußeren um 4,1 ‰ abgereicherter als das Bodenoberflächenwasser. – Nach Hartard et al. 2008.
-1 0 1 2 3 4-6
-4
-2
0
2
4
δ18O des Bodenwassers [‰]
δ18O
des
Tha
llusw
asse
rs [‰
]
1:1-Linieinnerer Thallusbereichäußerer Thallusbereich
Ökologie und Physiologie Borken bewohnender Flechten

140
Ökologische Rolle der Flechten
Literatur
Aptroot, A. 1997. Lichen biodiversity in Papua New Guinea, with the report of 173 species on one tree. – Bibliotheca Lichenologica, 68: 203-213.
Beckett, R. P. 1997. Pressure-volume analysis of a range of poikilohydric plants implies the existence of negative turgor in vegetative cells. – Annals of Botany, 79: 145-152.
Blum, O. B. 1973. Water relation. – In: Ahmadjian, V. & M. E. Hale (eds.): The lichens. Academic Press, New York: 381-400.
Büdel, B. & C. Scheidegger. 1996. Thallus morphology and anatomy. – In Nash, T. H. III (ed.): Lichen Biology. Cambridge University Press, Cambridge: 37-64.
Craig, H. 1954. Carbon-13 in plants and the relation-ship between carbon-13 and carbon-14 variations in nature. – J. of Geology, 62: 115-149.
Craig, H. & L. I. Gordon. 1965. Deuterium and oxygen-18 variations in the ocean and the marine atmosphere. – In: Tongiorgi, E. (ed.): Stable isotopes in oceano-graphic studies and paleotemperatures. Laboratorio di Geologia Nucleare, Pisa, Italy: 9-130.
Dalling, J. W. 1994. The Vegetation colonization of landslides in the Blue Mountains, Jamaica. – Bio-tropica, 26: 392-399.
Farquhar, G. D. & L. A. Cernusak. 2005. On the isotopic composition of leaf water in the non-steady state. – Functional Plant Biology, 32: 293-303.
Farquhar, G. D. & R. A. Richards. 1984. Isotopic com-position of plant carbon correlates with water-use efficiency of wheat genotypes. – Australian J. of Plant Physiology, 11: 539-552.
Fletcher, B. J., D. J. Beerling, S. J. Brentnall & D. L. Royer. 2005. Fossil bryophytes as recorders of ancient CO2 levels: Experimental evidence and a Cretaceous case study. – Global Biogeochemical Cycles, 19: GB3012.
Freiberg, M. & E. Freiberg. 2000. Epiphyte diversity and biomass in the canopy of lowland and mon-tane forests in Ecuador. – J. of Tropical Ecology, 16: 673-688.
Goebel, K. von. 1926. Ein Beitrag zur Biologie der Flechten. – Annales du Jardin Botanique de Bui-tenzorg, 36: 1-83.
Green, T. G. A. 2009. Lichens in arctic, antarctic and alpine ecosystems. – In: Bayer. Akademie d. Wissen-schaften (Hrsg.): Rundgespräche der Kommission für Ökologie, 36: Ökologische Rolle der Flechten. Pfeil, München: 45-65.
Hartard, B., C. Maguas & M. Lakatos. 2008. Delta O-18 characteristics of lichens and their effects on evapo-rative processes of the subjacent soil. – Isotopes in Environmental and Health Studies, 44: 111-125.
Hartard, B., M. Cuntz, C. Maguas & M. Lakatos. Submit-ted. Fractionation processes of desiccating lichens: A novel water exchange model for poikilohydric cryptogams. – New Phytologist.
Hauck, M. 2009. Flechten und Schadstoffe. – In: Bayer. Akademie d. Wissenschaften (Hrsg.): Rundgesprä-che der Kommission für Ökologie, 36: Ökologische Rolle der Flechten. Pfeil, München: 83-92.
Helliker, B. R. & H. Griffiths. 2007. Toward a plant-based proxy for the isotope ratio of atmospheric water vapor. – Global Change Biology, 13: 723-733.
Honegger, R. 1995. Experimental studies with foliose macrolichens: Fungal responses to spatial distur-bance at the organismic level and to spatial problems at the cellular level during drought stress events. – In: 5th International Mycological Congress, Vancou-ver, Canada. Canadian J. of Botany, Supplement 1 A-D: S569-S578.
– 2009. Flechten: ökologische Aspekte der Wechsel-beziehung zwischen Pilz und Alge. – In: Bayer. Akademie d. Wissenschaften (Hrsg.): Rundgesprä-che der Kommission für Ökologie, 36: Ökologische Rolle der Flechten. Pfeil, München: 25-41.
Jahns, H. M. 1984. Morphology, reproduction and water relations – a system of morphogenetic interactions in Paremlia saxatilis. – In: Hertel, H. & F. Oberwinkler (eds.): Beiträge zur Lichenologie. Festschrift J. Poelt. Beiheft zur Nova Hedwigia, 79: 715-737.
Johansson, D. 1974. Ecology of vascular epiphytes in West African rain forest. – Acta Phytogeographica Suecica, 59: 1-136.
Lakatos, M., U. Rascher & B. Büdel. 2006. Functional characteristics of corticolous lichens in the un-derstory of a tropical lowland rain forest. – New Phytologist, 172: 679-695.
Lakatos, M., B. Hartard & C. Máguas. 2007. The stable isotopes δ13C and δ18O of lichens can be used as tracers of microenvironmental carbon and water sources. – In: Dawson, T. & R. Siegwolf (eds.): Stable isotopes as indicators of ecological change. Academic Press, London: 77-92.
Lange, O. L., E. Kilian & H. Ziegler. 1986. Water vapor uptake and photosynthesis of lichens: Performance differences in species with green and blue-green algae as phycobionts. – Oecologia, 71: 104-110.
Lange, O. L., T. G. Green & U. Heber. 2001. Hydration-dependent photosynthetic production of lichens: What do laboratory studies tell us about field performance? – J. of Experimental Botany, 52: 2033-2042.
Máguas, C. & E. Brugnoli. 1996. Spatial variation in carbon isotope discrimination across the thalli of several lichen species. – Plant Cell & Environment, 19: 437-446.
Máguas, C, H. Griffiths & M. S. J. Broadmeadow. 1995. Gas exchange and carbon isotope discrimination in lichens – evidence for interactions between CO2-concentrating mechanisms and diffusion limitation. – Planta, 196: 95-102.
Proctor, M. C., Z. Nagy, Z. Csintalan & Z. Takacs. 1998. Water-content components in bryophytes: Analysis of pressure-volume relationships. – J. of Experimental Botany, 49: 1845-1854.

141
Pypker, T. G., M. H. Unsworth & B. J. Bond. 2006. The role of epiphytes in rainfall interception by forests in the Pacific Northwest. II. Field measurements at the branch and canopy scale. – Canadian J. of Forest Research, 36: 819-832.
Reyes-Garcia, C., M. Mejia-Chang, G. D. Jones & H. Griffiths. 2008. Water vapour isotopic exchange by epiphytic bromeliads in tropical dry forests reflects niche differentiation and climatic signals. – Plant Cell Environment, 31: 828-841.
Rhoades, F. M. 1981. Biomass of epiphytic lichens and bryophytes on Abies lasiocarpa on a Mt. Baker lava flow, Washington. – The Bryologist, 84: 39-47.
Rundel, P. W. 1988. Water relation. – In: Galum, M. (ed.): Handbook of lichenology, Vol. 2, CRC Press, Boca Raton: 17-36.
Scheidegger, C., B. Schroeter & F. Frey. 1995. Structural and functional processes during water vapour up-take and desiccation in selected lichens with green algal photobionts. – Planta, 197: 399-409.
Segerer, A. 2009. Die Bedeutung von Flechten als Nahrungsgrundlage für Tiere, insbesondere für Schmetterlinge. – In: Bayer. Akademie d. Wissen-schaften (Hrsg.): Rundgespräche der Kommission für Ökologie, 36: Ökologische Rolle der Flechten. Pfeil, München: 109-127.
Smith, E. C. & H. Griffiths. 1996. The occurrence of the chloroplast pyrenoid is correlated with the activ-ity of a CO2-concentrating mechanism and carbon isotope discrimination in lichens and bryophytes. – Planta, 198: 6-16.
– 2000. The role of carbonic anhydrase in photosyn-thesis and the activity of the carbon-concentrating-mechanism in bryophytes of the class anthocerotae. – New Phytologist, 145: 29-37.
Sternberg, L. da S. L., S. S. Mulkey& S. J. Wright. 1989. Ecological interpretation of leaf carbon isotope ratios: Influence of respired carbon dioxide. – Ecol-ogy, 70: 1317-1324.
Zotz, G. 2007. Johansson revisited: The spatial structure of epiphyte assemblages. – J. of Vegetation Science, 18: 123-130.
Ökologie und Physiologie Borken bewohnender Flechten

142
Ökologische Rolle der Flechten
B. Hoppe: Ihre Ausführungen schienen in die Richtung zu gehen, dass die Problematik von CO2 -Aufnahme und Gasaustausch relativ gesichert ist und auch schon vielfach untersucht wurde. Bei dem Verständnis zum Wasseraustausch und vor allem bei den Möglichkeiten der Bestimmung des Wassergehalts in dem Flechtenthallus entsteht aber der Eindruck, dass es noch schwierig ist, allein die Messgrößen zu gewinnen. Es entstand etwas der Eindruck, dass Sie die Formel mehr-mals nachbessern mussten, fast in Richtung »Schönung«. Gibt es weitere Forschungen, die diese Fragestellung verstärkt absichern in die Richtung, die Sie vorgetragen haben?
M. Lakatos: Vor ungefähr zwanzig Jahren haben Herr Professor Lange und Herr Professor Ziegler Pionierarbeit geleistet und die ersten Ergebnisse zu Fraktionierungsprozessen von 13C unter ande-rem bei Flechten publiziert1. Seitdem wurden in einigen wenigen Veröffentlichungen diese Me-chanismen bezüglich Flechten weiter untersucht und es besteht eine solide Forschungsgrundlage, die jedoch bisher den Effekt verschiedener CO2-Quellen nicht berücksichtigt hat. Die Forschung an Wasseraustauschprozessen mittels 18O hat jedoch erst vor vier Jahren begonnen und steht somit noch am Anfang. Die schrittweise Opti-
mierung des vorgestellten Modells liegt daran, dass bis dahin noch keine Formel für poikilo-hydre Organismen existierte. Wir mussten sie daher erst einmal belegen, beweisen, verändern und anpassen, und uns dabei überlegen, welche Mechanismen poikilohydrer Pflanzen integriert werden müssen. Wir haben inzwischen erfasst, wie der qualitative Wasseraustausch bei Flechten funktioniert, verschiedene Eckpunkte müssen wir jedoch noch abgleichen und verschiedene Mechanismen feiner skalieren, um das Modell dann auch technisch umzusetzen. Vor allem benö-tigen wir auch noch verschiedene Referenzdaten von poikilohydren Organismen, die sich in ihrer Morphologie oder Habitatpräferenz deutlich un-terscheiden. Man muss auch je nach Fragestellung zwischen unterschiedlichen Vorgehensweisen wählen, und für diese unterschiedlichen Bereiche brauchen wir natürlich eine Kalibrierung. Daran arbeiten wir momentan. Des Weiteren weisen Niederschläge und die Luftfeuchte weltweit ganz andere Werte auf. Sie folgen zwar bestimmten Regeln, aber wir müssen trotzdem prüfen, ob tatsächlich Flechten dies langzeitig gleichmäßig integrieren, und insbesondere müssen wir diese Langzeitdaten auch abgleichen, bevor wir am Ende entscheiden, ob wir bestimmte Flechten als Bioindikatoren universell verwenden können.
Diskusssion
1 Lange, O. L., T. G. A. Green & H. Ziegler. 1988. Water status related photosynthesis and carbon isotope discrimination in species of the lichen genus Pseudocyphellaria with green or blue-green photobionts and in photosymbiodemes. – Oecologia, 75: 494-501.
Lange, O. L., E. Kilian & H. Ziegler. 1986. Water vapour uptake and photosynthesis of lichens: Performance differences in species with green and blue-green algae as phycobionts. – Oecologia, 71: 104-110.
Rundel, P. W., W. Stichler, R. H. Zander & H. Ziegler. 1979. Carbon and hydrogen isotope ratios of bryophytes from arid and humid regions. – Oecologia, 44: 91-94.


Rundgespräche der Kommission für ÖkologieHerausgegeben von der Bayerischen Akademie der Wissenschaften
Bestellungen an:
Verlag Dr. Friedrich Pfeil, Wolfratshauser Straße 27, D-81379 MünchenTel.: +49 (0)89 742827-0 • Fax: +49 (0)89 7242772 • E-Mail: [email protected]
www.pfeil-verlag.de





















![Flechtpultrusion & Flechtentwicklungen am ITV Denkendorf Flechten... · Pultrusion (Röchling, Saertex, BAM, Fiberware ITKE, ITV) [Quelle BAM] 10 Faserverbundbauteile von der Faser](https://img.pdfslide.org/doc/110x75/5eae42cc9116ec5e20721f56/flechtpultrusion-flechtentwicklungen-am-itv-flechten-pultrusion-rchling.jpg)







