Embed Size (px)
Citation preview


Selektive Neutralität und Effizienz der EvolutionWas wir aus Evolutionsexperimenten lernen können
Peter SchusterInstitut für Theoretische Chemie und Molekulare
Strukturbiologie der Universität Wien
Seminar des Naturhistorischen Museums
Wien, 28.04.2004

Web-Page for further information:
http://www.tbi.univie.ac.at/~pks
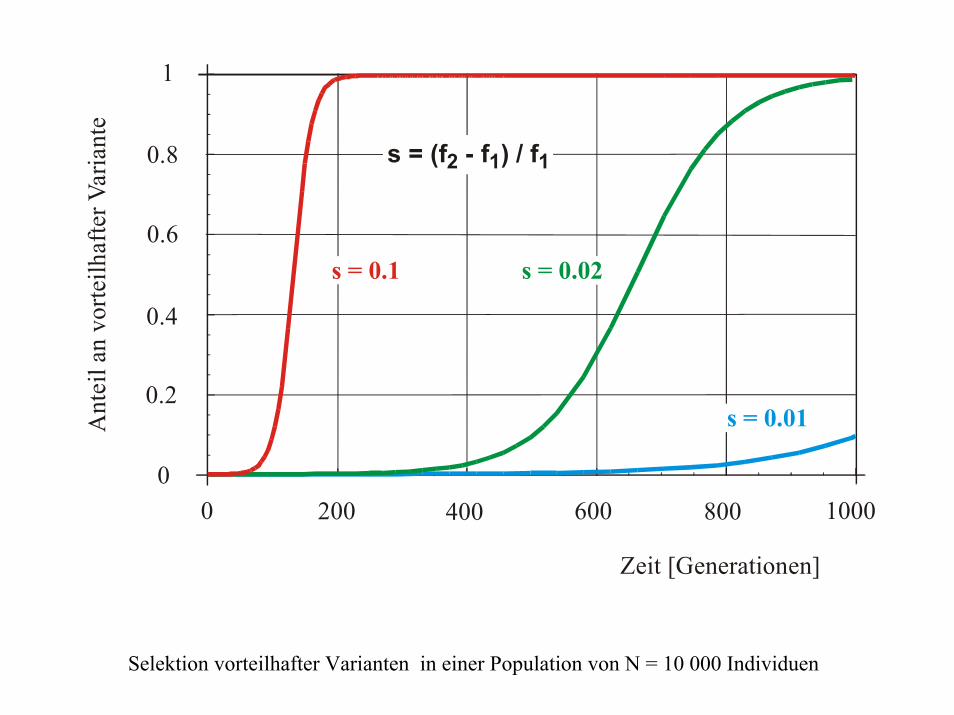
200 400 600 800 1000
0.2
00
0.4
0.6
0.8
1
Zeit [Generationen]
Ant
eil a
n vo
rteilh
afte
r Var
iant
e
s = 0.1
s = 0.01
s = 0.02
s = (f - f ) / f2 1 1
Selektion vorteilhafter Varianten in einer Population von N = 10 000 Individuen

Massif Central
Mount FujiBeispiele glatter Landschaften

Dolomiten
Beispiele zerklüfteter Landschaften Bryce Canyon
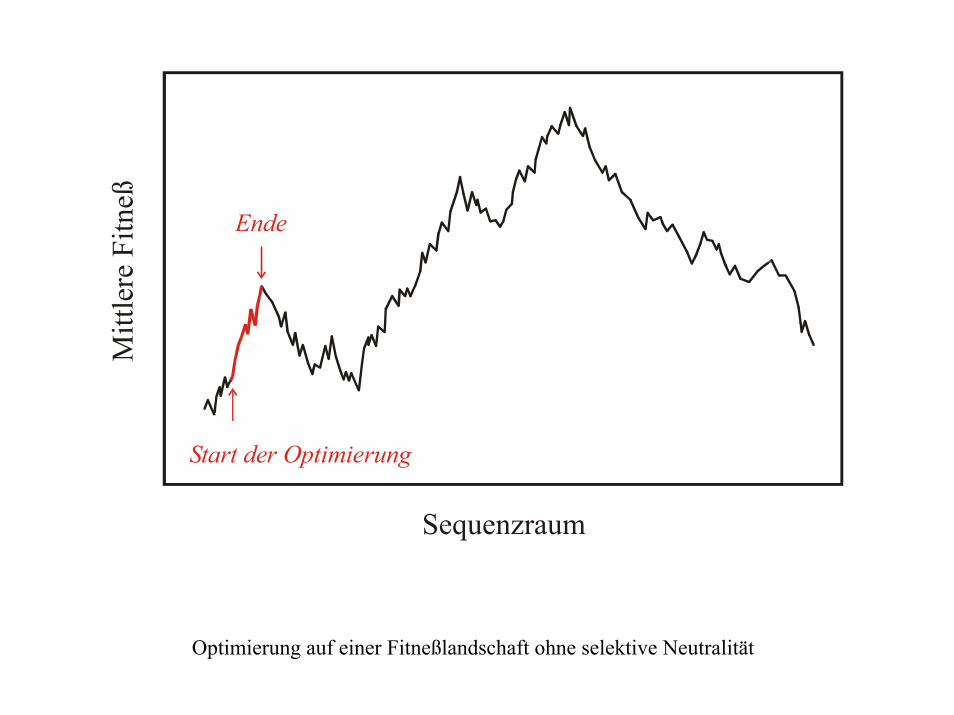
Sequenzraum
Mitt
lere
Fitn
eß
Start der Optimierung
Ende
Optimierung auf einer Fitneßlandschaft ohne selektive Neutralität
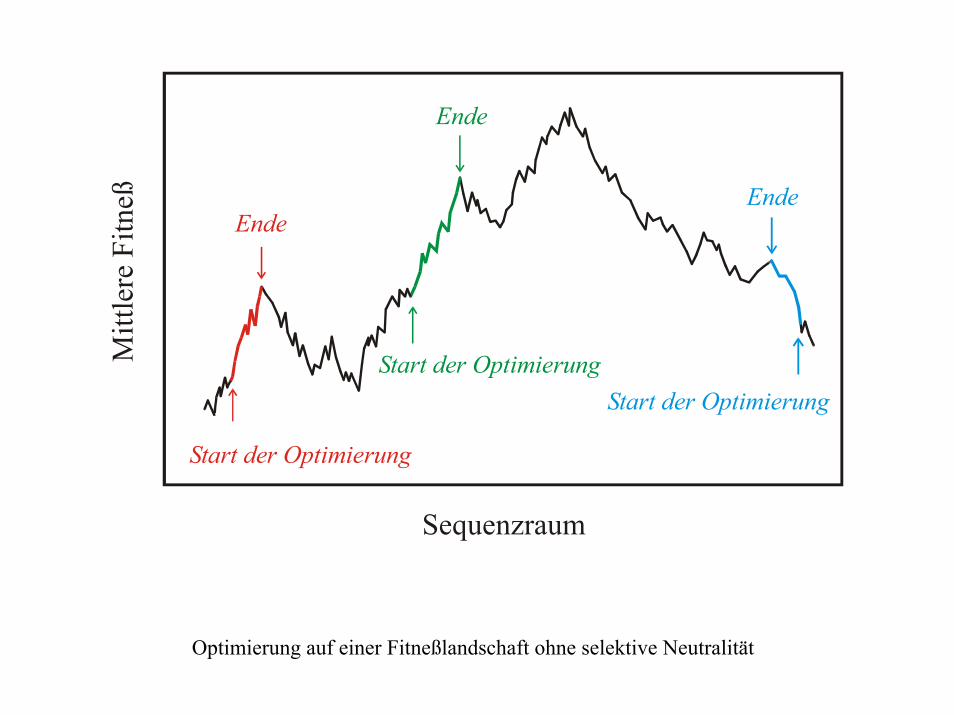
Sequenzraum
Mitt
lere
Fitn
eß
Start der Optimierung
Start der Optimierung
Start der Optimierung
Ende
Ende
Ende
Optimierung auf einer Fitneßlandschaft ohne selektive Neutralität
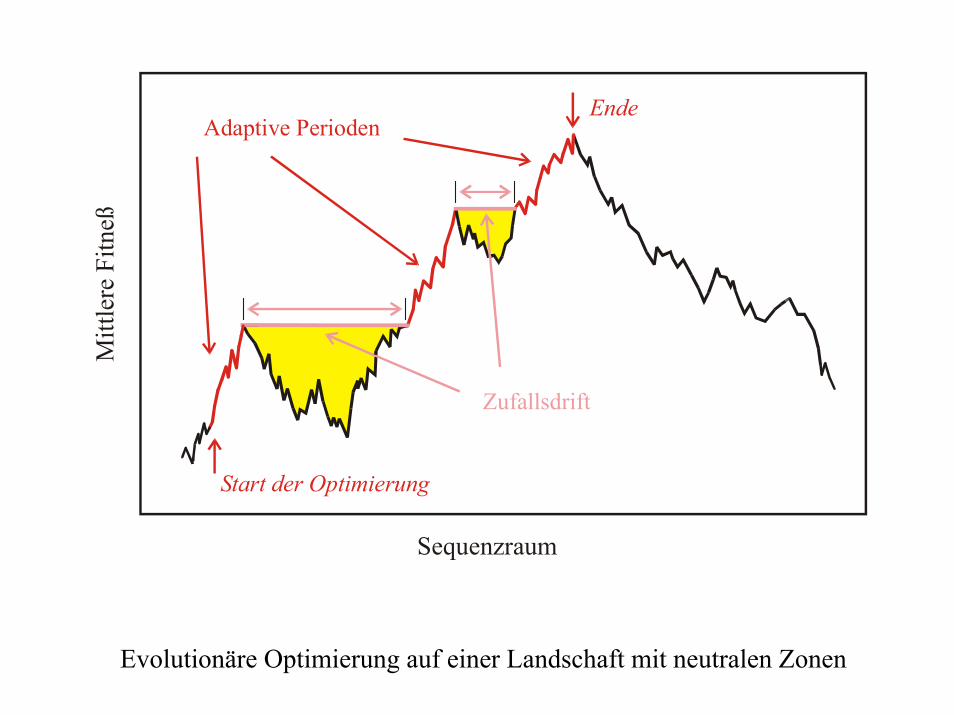
Sequenzraum
Mitt
lere
Fitn
eß
Start der Optimierung
Ende
Zufallsdrift
Adaptive Perioden
Evolutionäre Optimierung auf einer Landschaft mit neutralen Zonen

Grand Canyon
Beispiel einer Landschaft mit neutralen Graten und Plateaus

Neutrale Grate und Plateaus

„...Variations neither useful not injurious would not be affected by natural selection, and would be left either a fluctuating element, as perhaps we see in certain polymorphic species, or would ultimately become fixed, owing to the nature of the organism and the nature of the conditions. ...“
Charles Darwin, Origin of species (1859)

The molecular clock of evolution
Motoo Kimura’s population genetics of neutral evolution.
Evolutionary rate at the molecular level. Nature 217: 624-626, 1955.
The Neutral Theory of Molecular Evolution. Canbridge University Press. Cambridge, UK, 1983.

Molecular evolution through comparison of sequences from different organisms
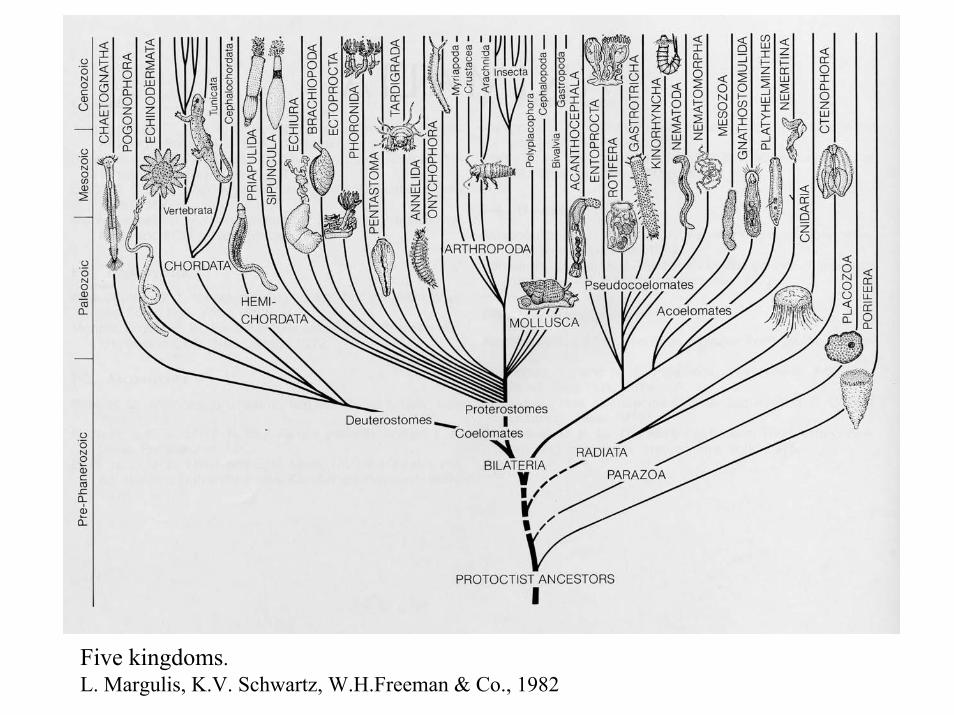
Five kingdoms.L. Margulis, K.V. Schwartz, W.H.Freeman & Co., 1982
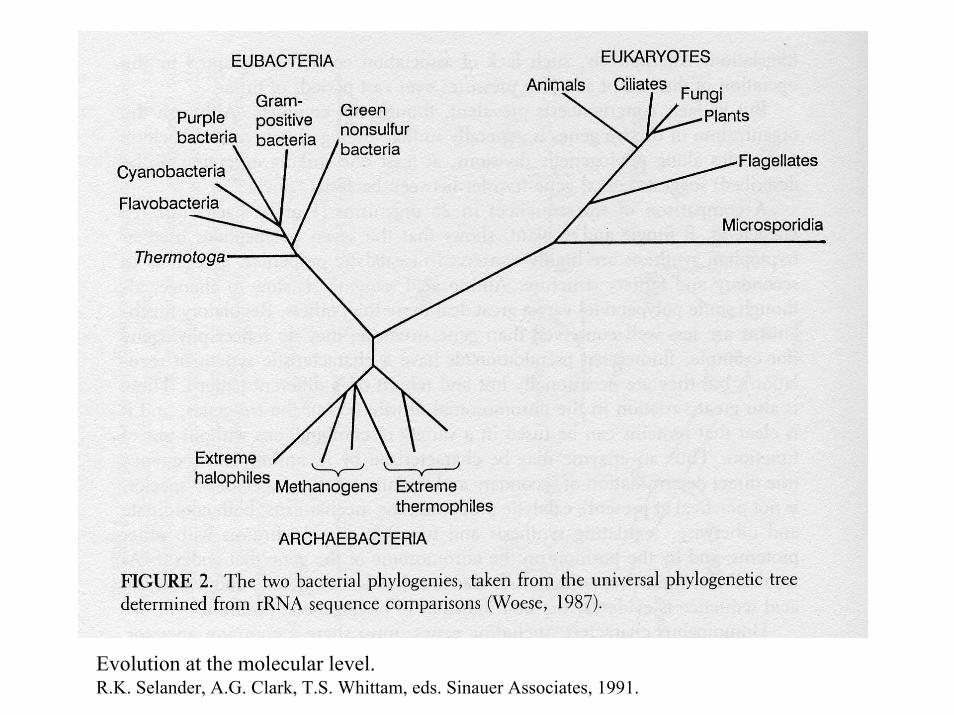
Evolution at the molecular level.R.K. Selander, A.G. Clark, T.S. Whittam, eds. Sinauer Associates, 1991.
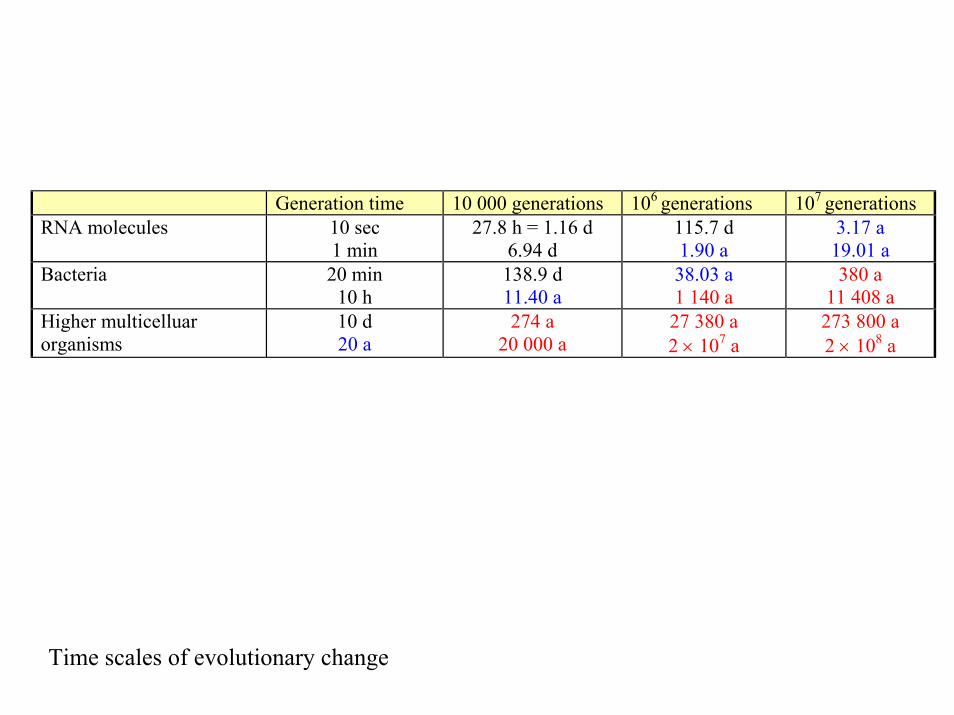
Generation time 10 000 generations 106 generations 107 generations RNA molecules 10 sec
1 min 27.8 h = 1.16 d
6.94 d 115.7 d 1.90 a
3.17 a 19.01 a
Bacteria 20 min 10 h
138.9 d 11.40 a
38.03 a 1 140 a
380 a 11 408 a
Higher multicelluar organisms
10 d 20 a
274 a 20 000 a
27 380 a 2 × 107 a
273 800 a 2 × 108 a
Time scales of evolutionary change

Bacterial Evolution
S. F. Elena, V. S. Cooper, R. E. Lenski. Punctuated evolution caused by selection of rare beneficial mutants. Science 272 (1996), 1802-1804
D. Papadopoulos, D. Schneider, J. Meier-Eiss, W. Arber, R. E. Lenski, M. Blot. Genomic evolution during a 10,000-generation experiment with bacteria. Proc.Natl.Acad.Sci.USA 96 (1999), 3807-3812
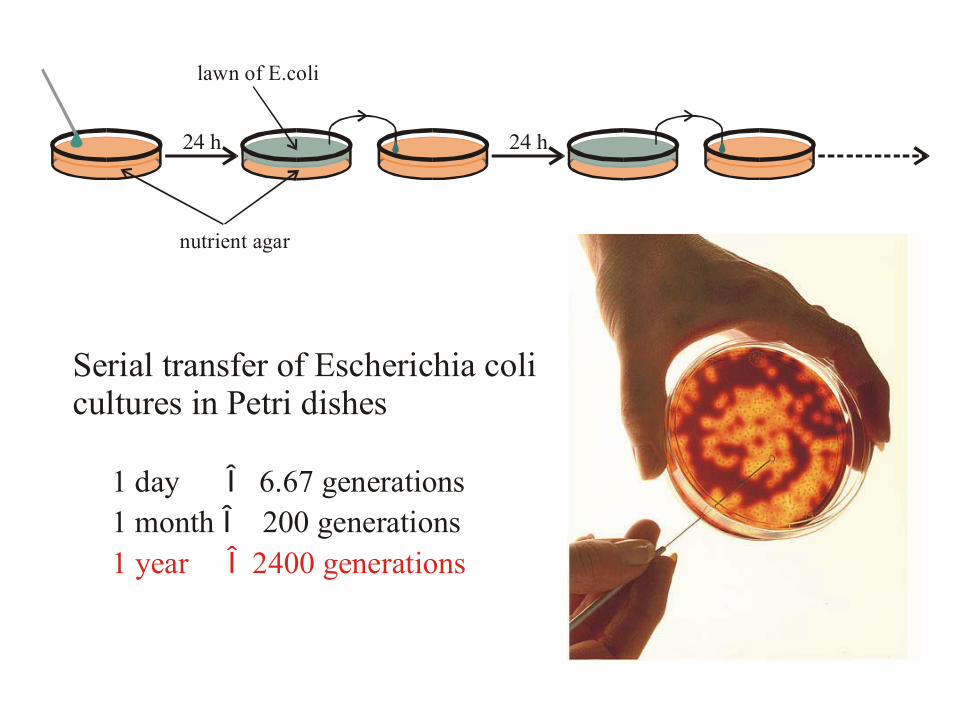
24 h 24 h
Serial transfer of Escherichia colicultures in Petri dishes
1 day 6.67 generations 1 month 200 generations
ÎÎ
1 year 2400 generationsÎ
lawn of E.coli
nutrient agar
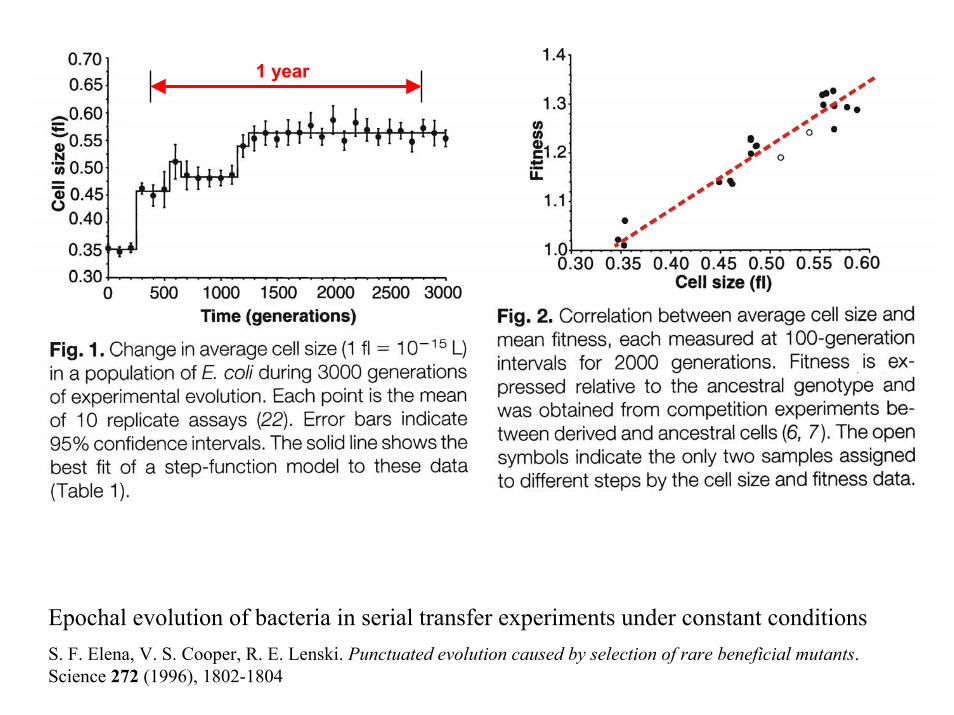
1 year
Epochal evolution of bacteria in serial transfer experiments under constant conditionsS. F. Elena, V. S. Cooper, R. E. Lenski. Punctuated evolution caused by selection of rare beneficial mutants. Science 272 (1996), 1802-1804
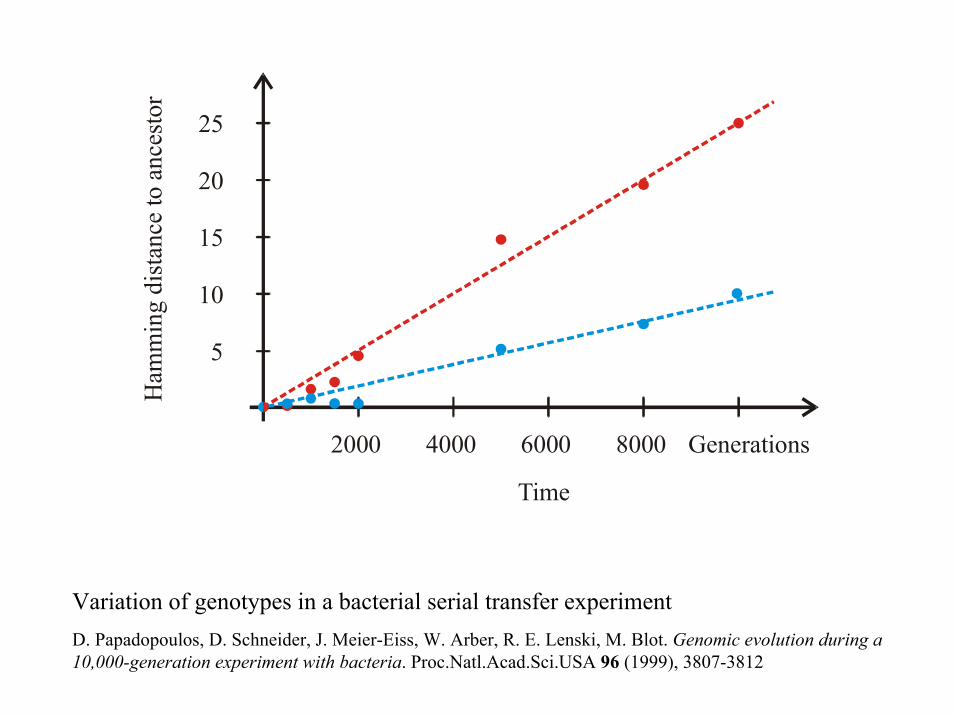
2000 4000 6000 8000
Time
5
10
15
20
25H
amm
ing
dist
ance
to a
nces
tor
Generations
Variation of genotypes in a bacterial serial transfer experimentD. Papadopoulos, D. Schneider, J. Meier-Eiss, W. Arber, R. E. Lenski, M. Blot. Genomic evolution during a 10,000-generation experiment with bacteria. Proc.Natl.Acad.Sci.USA 96 (1999), 3807-3812

Evolution of RNA molecules based on Qβ phage
D.R.Mills, R,L,Peterson, S.Spiegelman, An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule. Proc.Natl.Acad.Sci.USA 58 (1967), 217-224
S.Spiegelman, An approach to the experimental analysis of precellular evolution. Quart.Rev.Biophys. 4 (1971), 213-253
C.K.Biebricher, Darwinian selection of self-replicating RNA molecules. Evolutionary Biology 16 (1983), 1-52
C.K.Biebricher, W.C. Gardiner, Molecular evolution of RNA in vitro. Biophysical Chemistry 66 (1997), 179-192
G.Strunk, T. Ederhof, Machines for automated evolution experiments in vitro based on the serial transfer concept. Biophysical Chemistry 66 (1997), 193-202
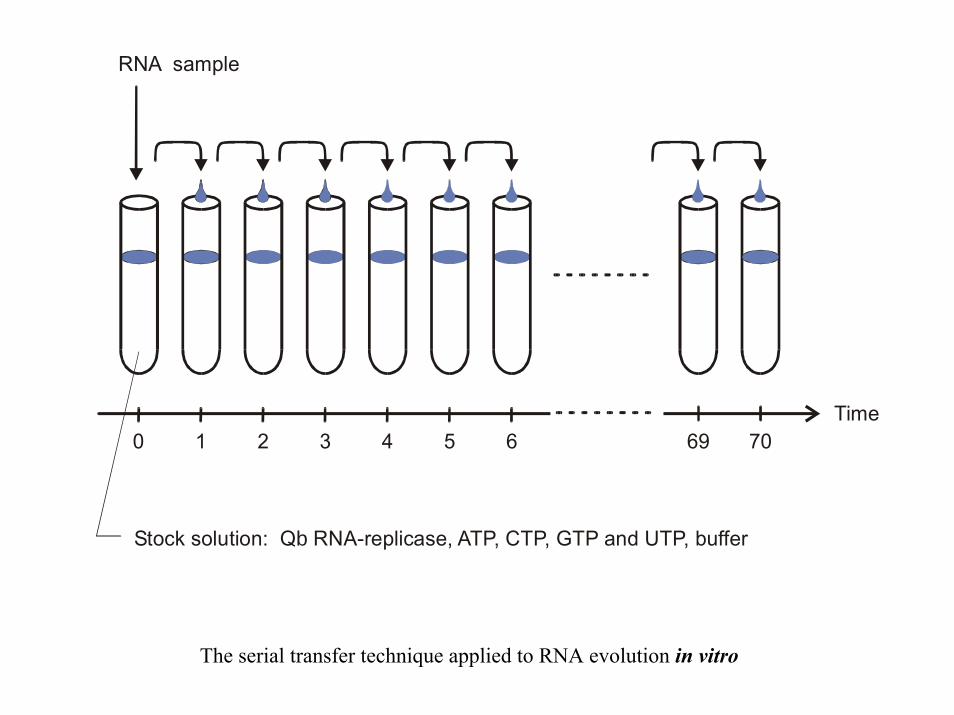
RNA sample
Stock solution: Q RNA-replicase, ATP, CTP, GTP and UTP, bufferb
Time0 1 2 3 4 5 6 69 70
The serial transfer technique applied to RNA evolution in vitro
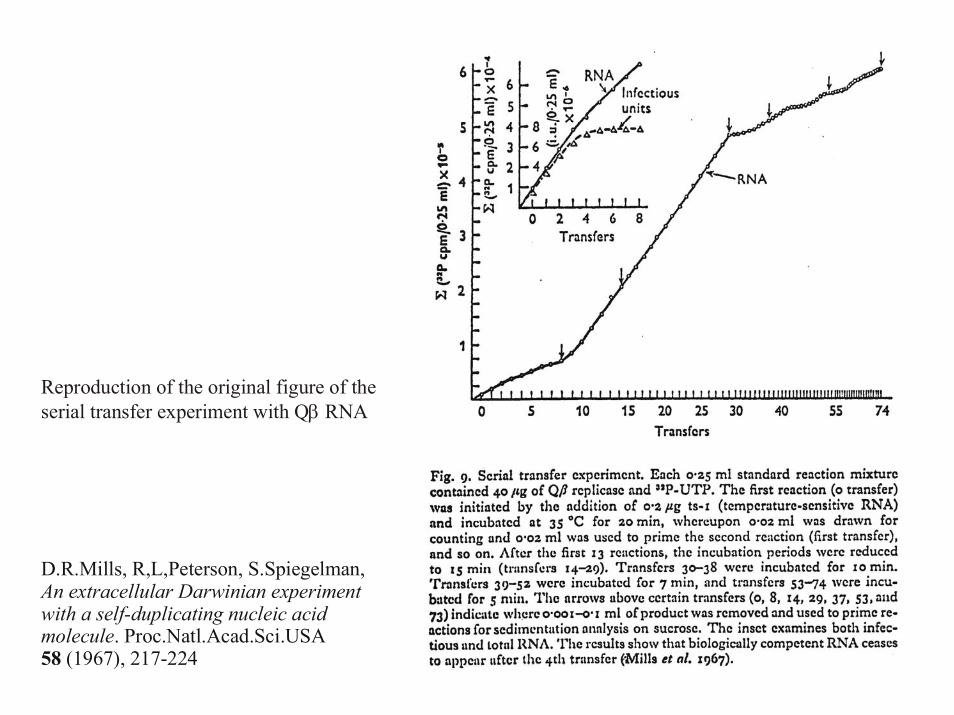
Reproduction of the original figure of theserial transfer experiment with Q RNAβ
D.R.Mills, R,L,Peterson, S.Spiegelman,
. Proc.Natl.Acad.Sci.USA (1967), 217-224
An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule58
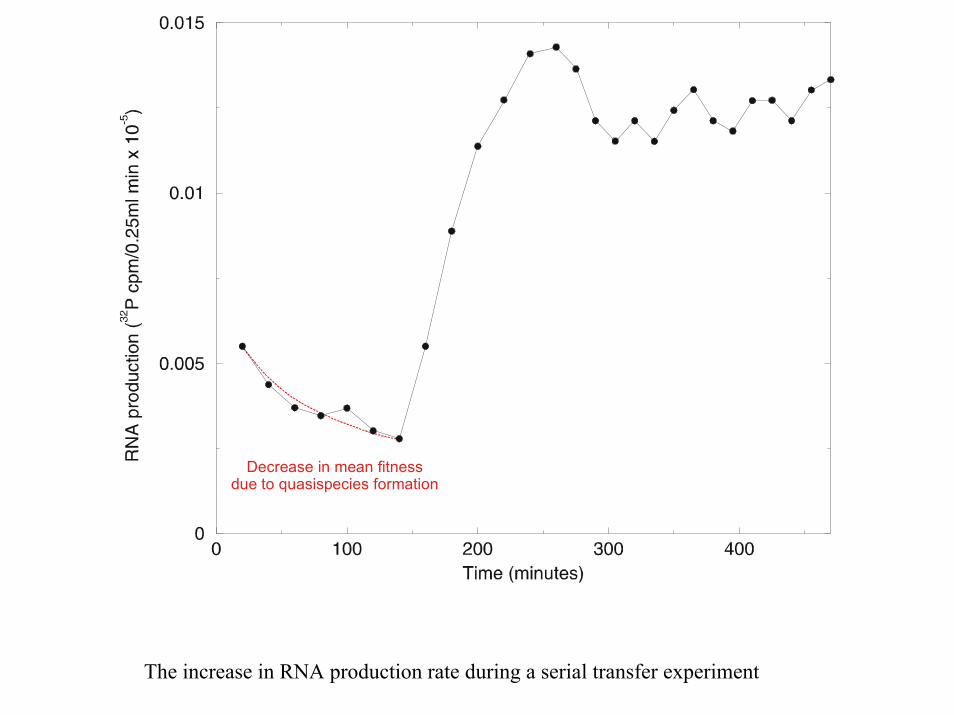
Decrease in mean fitnessdue to quasispecies formation
The increase in RNA production rate during a serial transfer experiment

Evolutionary design of RNA molecules
D.B.Bartel, J.W.Szostak, In vitro selection of RNA molecules that bind specific ligands. Nature 346 (1990), 818-822
C.Tuerk, L.Gold, SELEX - Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 249 (1990), 505-510
D.P.Bartel, J.W.Szostak, Isolation of new ribozymes from a large pool of random sequences. Science 261 (1993), 1411-1418
R.D.Jenison, S.C.Gill, A.Pardi, B.Poliski, High-resolution molecular discrimination by RNA. Science 263 (1994), 1425-1429
Y. Wang, R.R.Rando, Specific binding of aminoglycoside antibiotics to RNA. Chemistry &Biology 2 (1995), 281-290
Jiang, A. K. Suri, R. Fiala, D. J. Patel, Saccharide-RNA recognition in an aminoglycoside antibiotic-RNA aptamer complex. Chemistry & Biology 4 (1997), 35-50

No new principle will declare itself from below a heap of facts.
Sir Peter Medawar, 1985
G
G
G
C
C
C
C
C
C
G
G
G
C
C
C
G
G
G
C
C
C
G
G
G
G
G
G
C
C
C
Plus Strand
Plus Strand
Plus Strand
Minus Strand
Minus Strand
Minus Strand
3'
3'
3'5'
5'
5'
5'
5'
5'3'
3'
3'
+
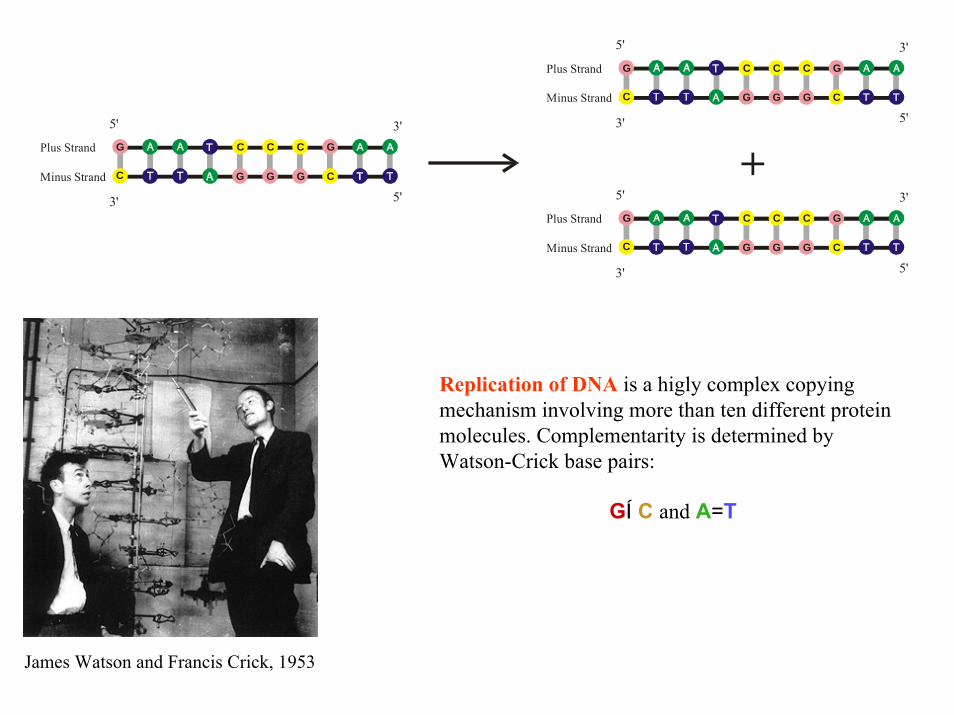
Replication of DNA is a higly complex copying mechanism involving more than ten different proteinmolecules. Complementarity is determined byWatson-Crick base pairs:
GÍC and A=T
James Watson and Francis Crick, 1953
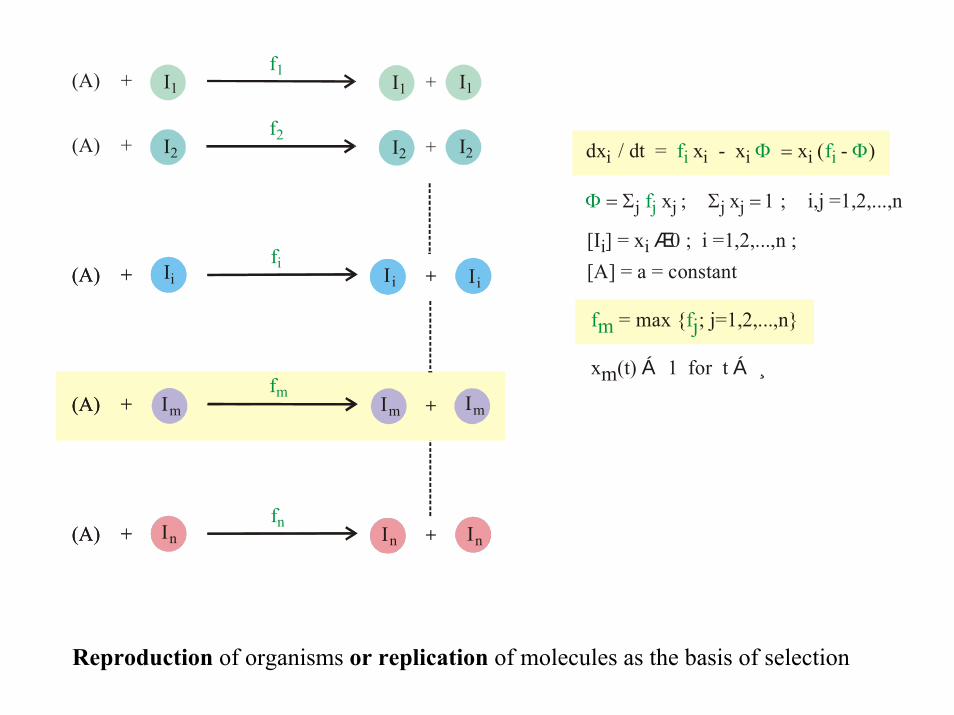
dx / dt = x - x
x
i i i
j j
; Σ = 1 ; i,j
f
f
i
j
Φ
Φ
fi Φ = (
= Σ
x - i )
j jx =1,2,...,n
[I ] = x 0 ; i i Æ i =1,2,...,n ; Ii
I1
I2
I1
I2
I1
I2
I i
I n
I i
I nI n
+
+
+
+
+
+
(A) +
(A) +
(A) +
(A) +
(A) +
(A) +
fn
fi
f1
f2
I mI m I m++(A) +(A) +fm
fm fj= max { ; j=1,2,...,n}
xm(t) 1 for t Á Á¸
[A] = a = constant
Reproduction of organisms or replication of molecules as the basis of selection
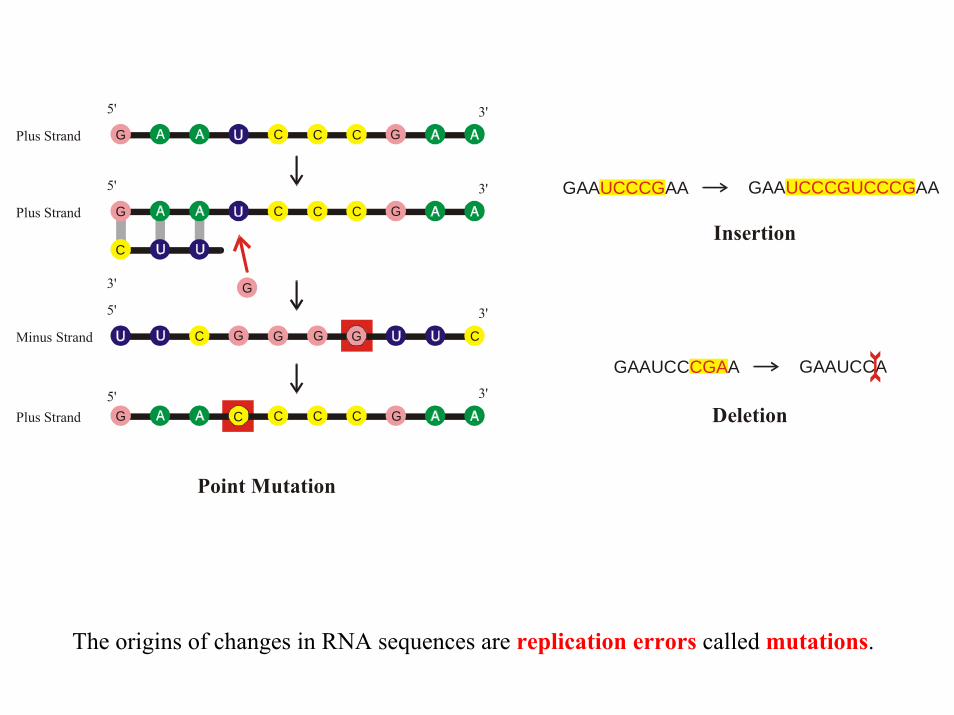
G
G
G
C
C
C
G
C
C
G
C
C
C
G
C
C
C
G
C
G
G
G
G
C
Plus Strand
Plus Strand
Minus Strand
Plus Strand
3'
3'
3'
3'
5'
3'
5'
5'
5'
Point Mutation
Insertion
Deletion
GAA AA UCCCG
GAAUCC A CGA
GAA AAUCCCGUCCCG
GAAUCCA
The origins of changes in RNA sequences are replication errors called mutations.

Theory of molecular evolution
M.Eigen, Self-organization of matter and the evolution of biological macromolecules. Naturwissenschaften 58 (1971), 465-526
C.J. Thompson, J.L. McBride, On Eigen's theory of the self-organization of matter and the evolution of biological macromolecules. Math. Biosci. 21 (1974), 127-142
B.L. Jones, R.H. Enns, S.S. Rangnekar, On the theory of selection of coupled macromolecular systems. Bull.Math.Biol. 38 (1976), 15-28
M.Eigen, P.Schuster, The hypercycle. A principle of natural self-organization. Part A: Emergence of the hypercycle. Naturwissenschaften 58 (1977), 465-526
M.Eigen, P.Schuster, The hypercycle. A principle of natural self-organization. Part B: The abstract hypercycle. Naturwissenschaften 65 (1978), 7-41
M.Eigen, P.Schuster, The hypercycle. A principle of natural self-organization. Part C: The realistic hypercycle. Naturwissenschaften 65 (1978), 341-369
J. Swetina, P. Schuster, Self-replication with errors - A model for polynucleotide replication.Biophys.Chem. 16 (1982), 329-345
J.S. McCaskill, A localization threshold for macromolecular quasispecies from continuously distributed replication rates. J.Chem.Phys. 80 (1984), 5194-5202
M.Eigen, J.McCaskill, P.Schuster, The molecular quasispecies. Adv.Chem.Phys. 75 (1989), 149-263
C. Reidys, C.Forst, P.Schuster, Replication and mutation on neutral networks. Bull.Math.Biol. 63(2001), 57-94

Chemical kinetics of molecular evolution
M. Eigen, P. Schuster, `The Hypercycle´, Springer-Verlag, Berlin 1979
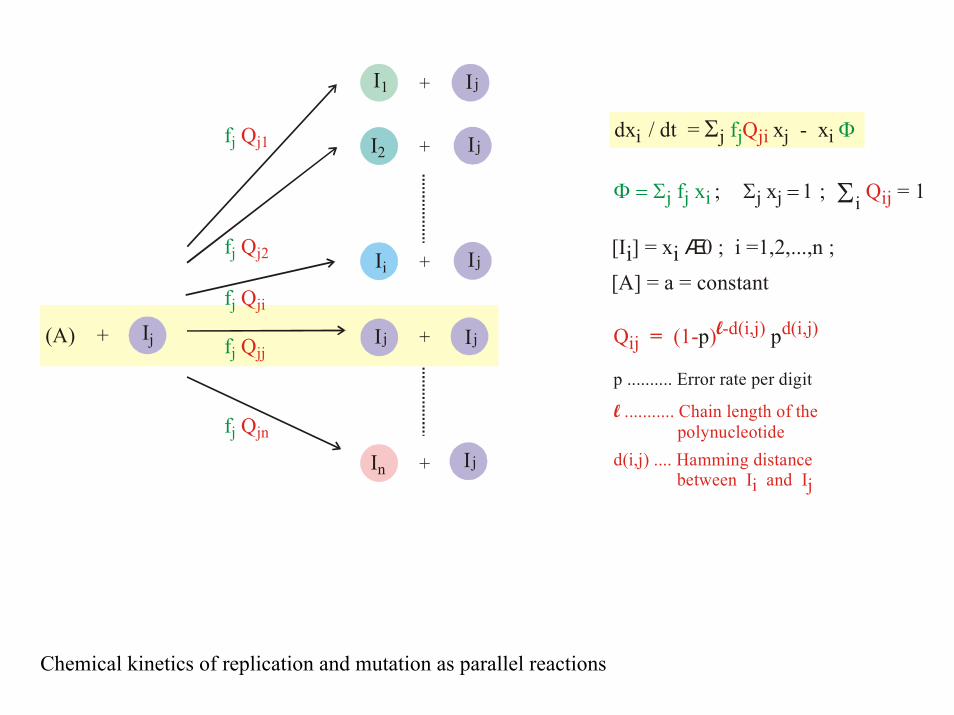
Ij
In
I2
Ii
I1 I j
I j
I j
I j
I j
I j +
+
+
+
+
(A) +
fj Qj1
fj Qj2
fj Qji
fj Qjj
fj Qjn
Q (1- ) ij-d(i,j) d(i,j) = lp p
p .......... Error rate per digit
d(i,j) .... Hamming distance between Ii and Ij
........... Chain length of the polynucleotidel
dx / dt = x - x
x
i j j i
j j
Σ
; Σ = 1 ;
f
f x
j
j j i
Φ
Φ = Σ
Qji
QijΣi = 1
[A] = a = constant
[Ii] = xi 0 ; Æ i =1,2,...,n ;
Chemical kinetics of replication and mutation as parallel reactions
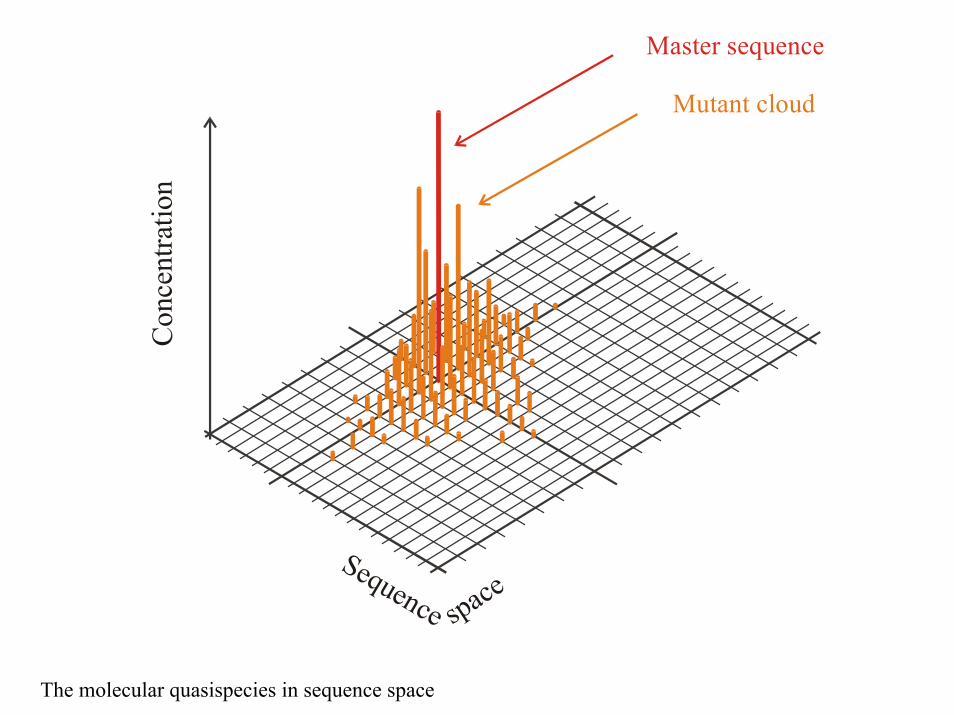
spaceSequence
Con
cent
ratio
n
Master sequence
Mutant cloud
The molecular quasispecies in sequence space
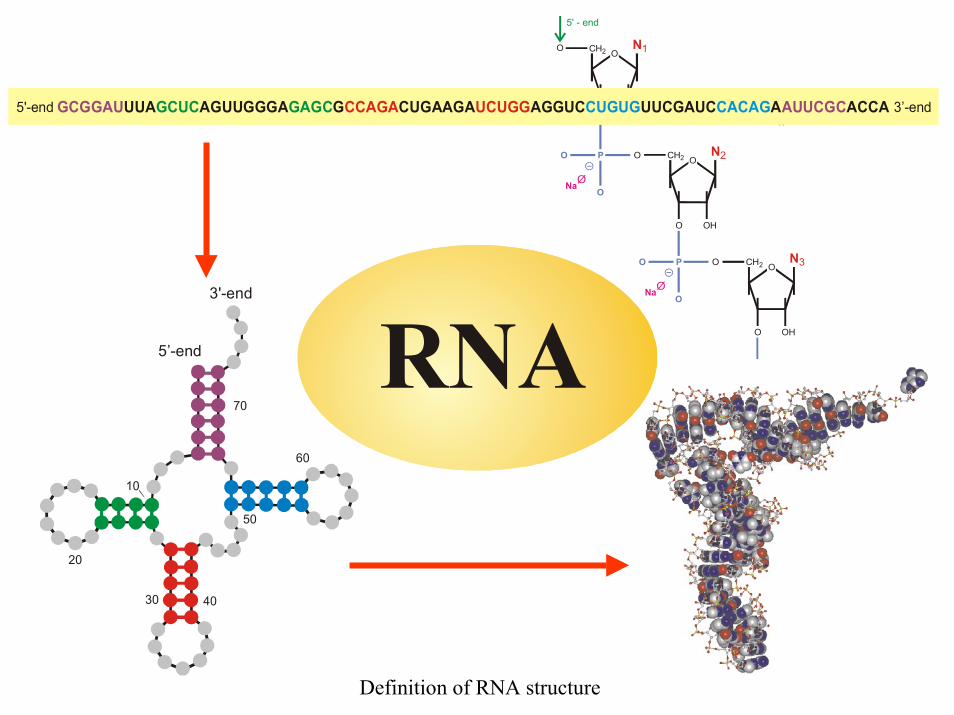
OCH2
OHO
O
PO
O
O
N1
OCH2
OHO
PO
O
O
N2
OCH2
OHO
PO
O
O
N3
OCH2
OHO
PO
O
O
N4
N A U G Ck = , , ,
3' - end
5' - end
NaØ
NaØ
NaØ
NaØ
RNA
nd 3’-endGCGGAU AUUCGCUUA AGUUGGGA G CUGAAGA AGGUC UUCGAUC A ACCAGCUC GAGC CCAGA UCUGG CUGUG CACAG
3'-end
5’-end
70
60
50
4030
20
10
Definition of RNA structure
5'-e

Optimization of RNA molecules in silico
W.Fontana, P.Schuster, A computer model of evolutionary optimization. Biophysical Chemistry 26 (1987), 123-147
W.Fontana, W.Schnabl, P.Schuster, Physical aspects of evolutionary optimization and adaptation. Phys.Rev.A 40 (1989), 3301-3321
M.A.Huynen, W.Fontana, P.F.Stadler, Smoothness within ruggedness. The role of neutralityin adaptation. Proc.Natl.Acad.Sci.USA 93 (1996), 397-401
W.Fontana, P.Schuster, Continuity in evolution. On the nature of transitions. Science 280(1998), 1451-1455
W.Fontana, P.Schuster, Shaping space. The possible and the attainable in RNA genotype-phenotype mapping. J.Theor.Biol. 194 (1998), 491-515
B.M.R. Stadler, P.F. Stadler, G.P. Wagner, W. Fontana, The topology of the possible: Formal spaces underlying patterns of evolutionary change. J.Theor.Biol. 213 (2001), 241-274

Evolution in silico
W. Fontana, P. Schuster, Science 280 (1998), 1451-1455
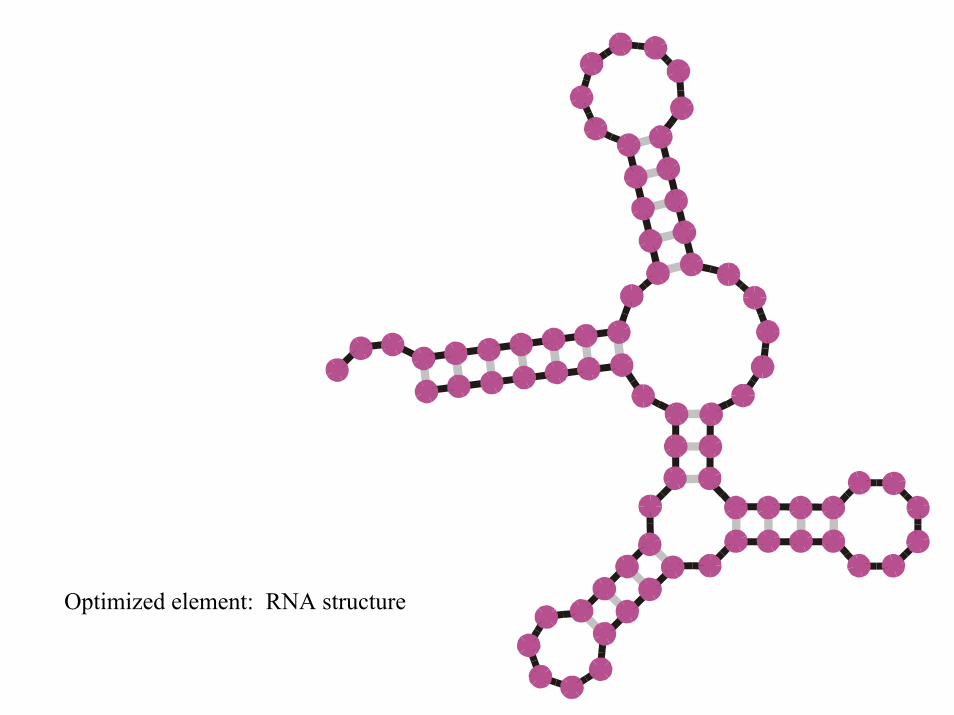
Optimized element: RNA structure
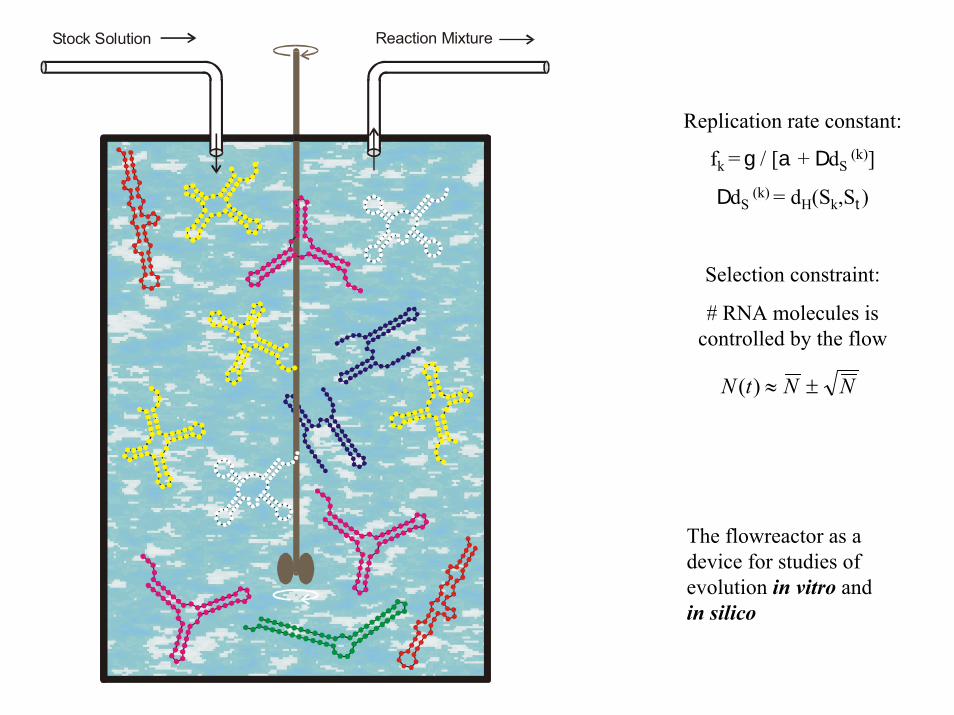
Stock Solution Reaction Mixture
Replication rate constant:
fk = g / [a + DdS(k)]
DdS(k) = dH(Sk,St)
Selection constraint:
# RNA molecules is controlled by the flow
NNtN ±≈)(
The flowreactor as a device for studies of evolution in vitro and in silico
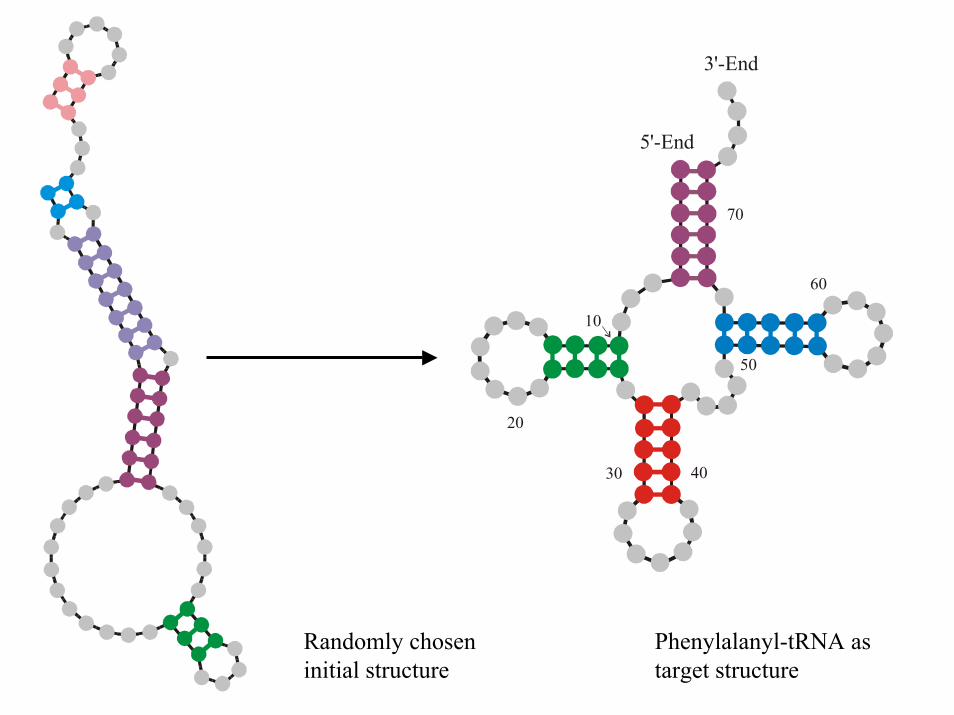
5'-End
3'-End
70
60
50
4030
20
10
Randomly chosen initial structure
Phenylalanyl-tRNA as target structure
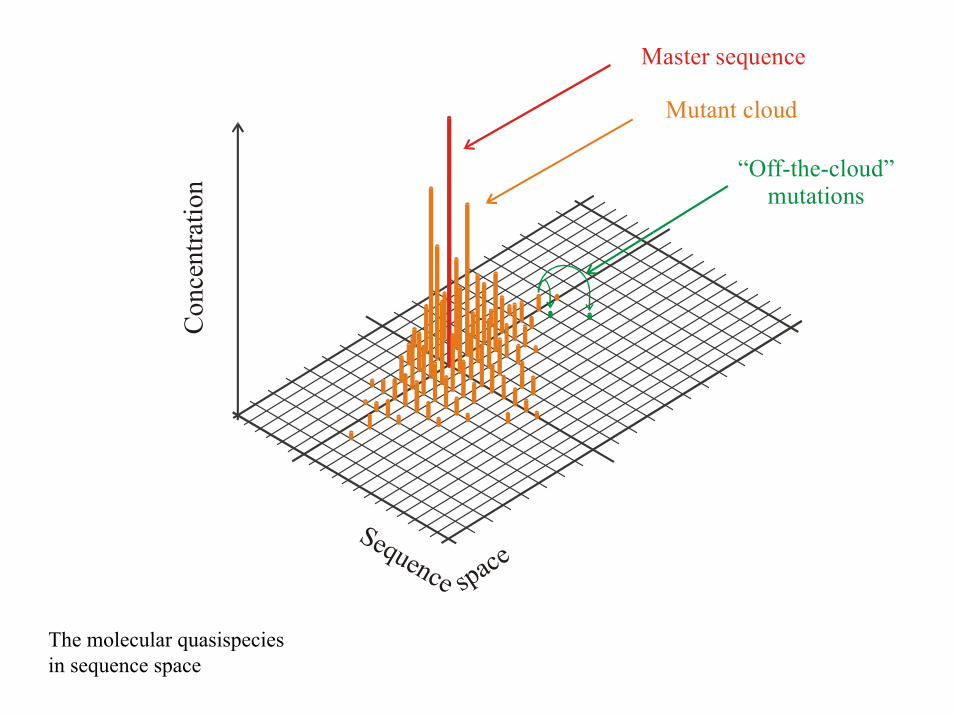
spaceSequence
Con
cent
ratio
n
Master sequence
Mutant cloud
“Off-the-cloud” mutations
The molecular quasispeciesin sequence space
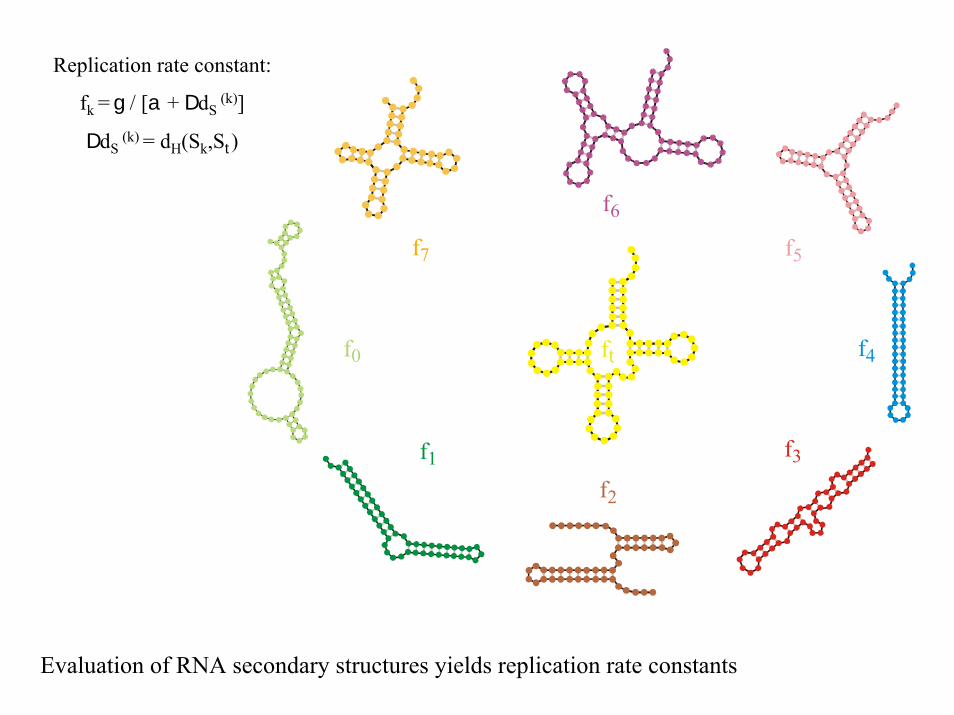
f0 ft
f1
f2
f3
f4
f6
f5f7
Replication rate constant:
fk = g / [a + DdS(k)]
DdS(k) = dH(Sk,St)
Evaluation of RNA secondary structures yields replication rate constants
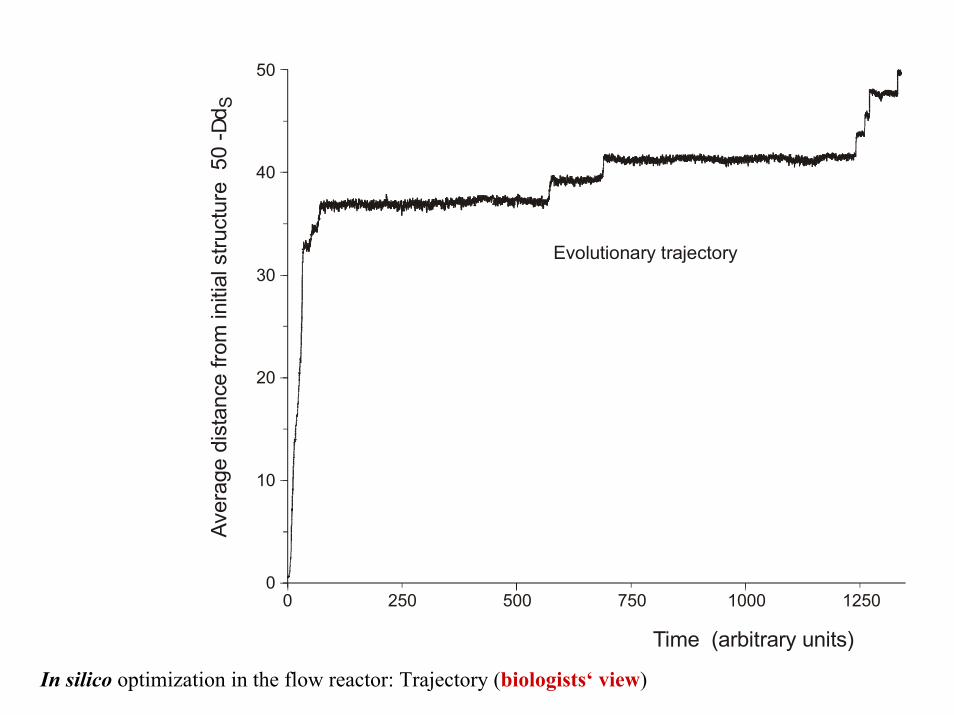
In silico optimization in the flow reactor: Trajectory (biologists‘ view)
Time (arbitrary units)
Aver
age
dist
ance
from
initi
al s
truct
ure
50
-d
D
S
500 750 1000 12502500
50
40
30
20
10
0
Evolutionary trajectory
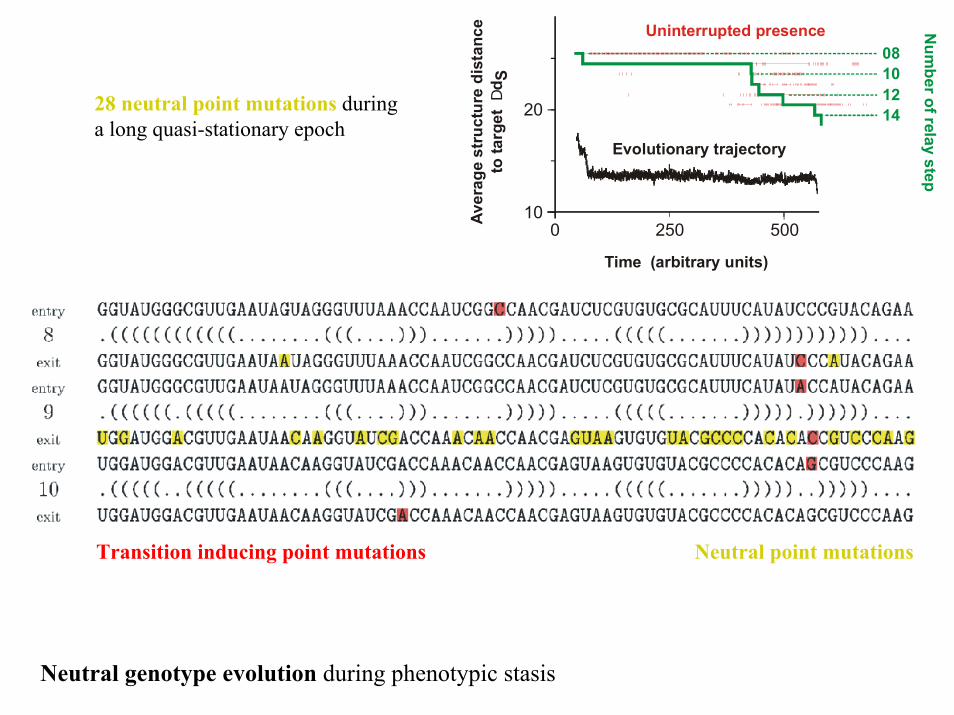
1008
1214
Time (arbitrary units)
Aver
age
stru
ctur
e di
stan
ce
to ta
rget
d S
D
5002500
20
10
Uninterrupted presence
Evolutionary trajectory
Num
ber of relay step
28 neutral point mutations during a long quasi-stationary epoch
Transition inducing point mutations Neutral point mutations
Neutral genotype evolution during phenotypic stasis
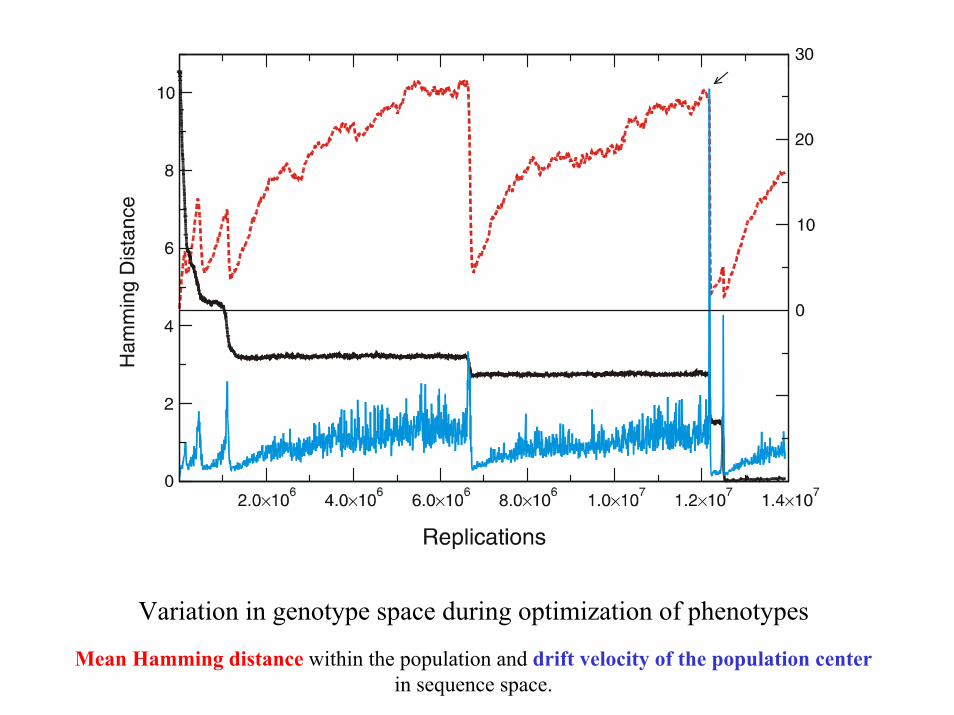
Variation in genotype space during optimization of phenotypes
Mean Hamming distance within the population and drift velocity of the population centerin sequence space.
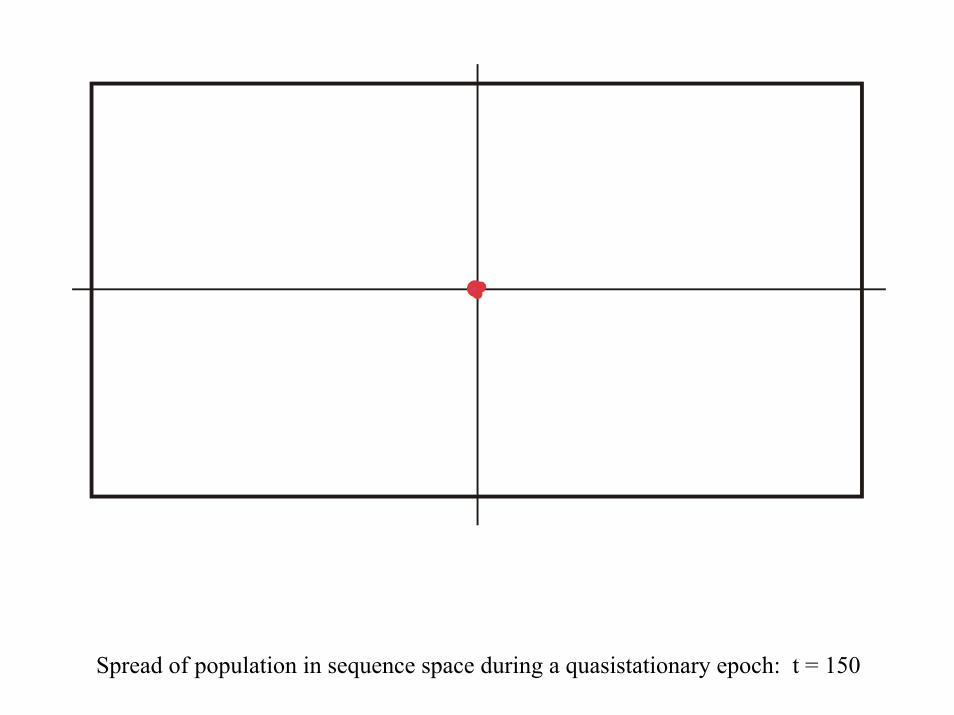
Spread of population in sequence space during a quasistationary epoch: t = 150
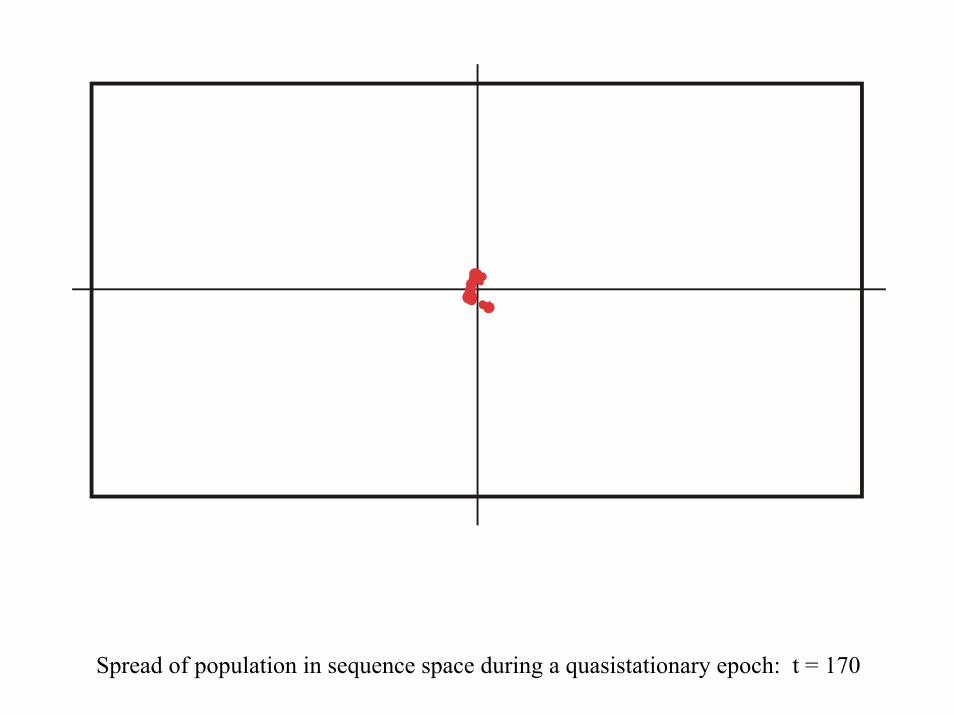
Spread of population in sequence space during a quasistationary epoch: t = 170
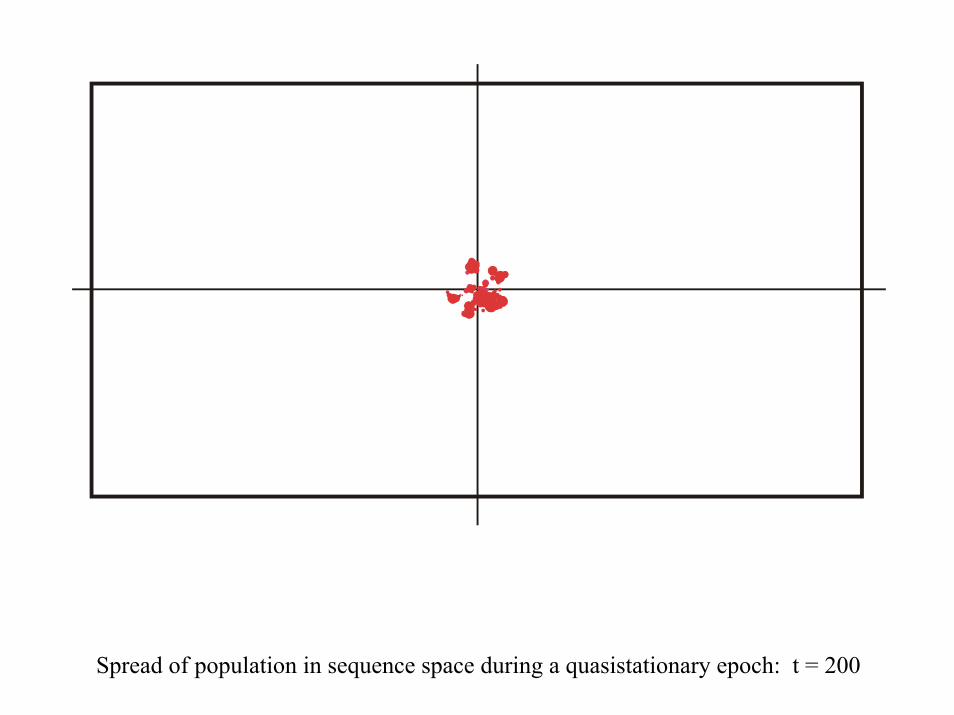
Spread of population in sequence space during a quasistationary epoch: t = 200
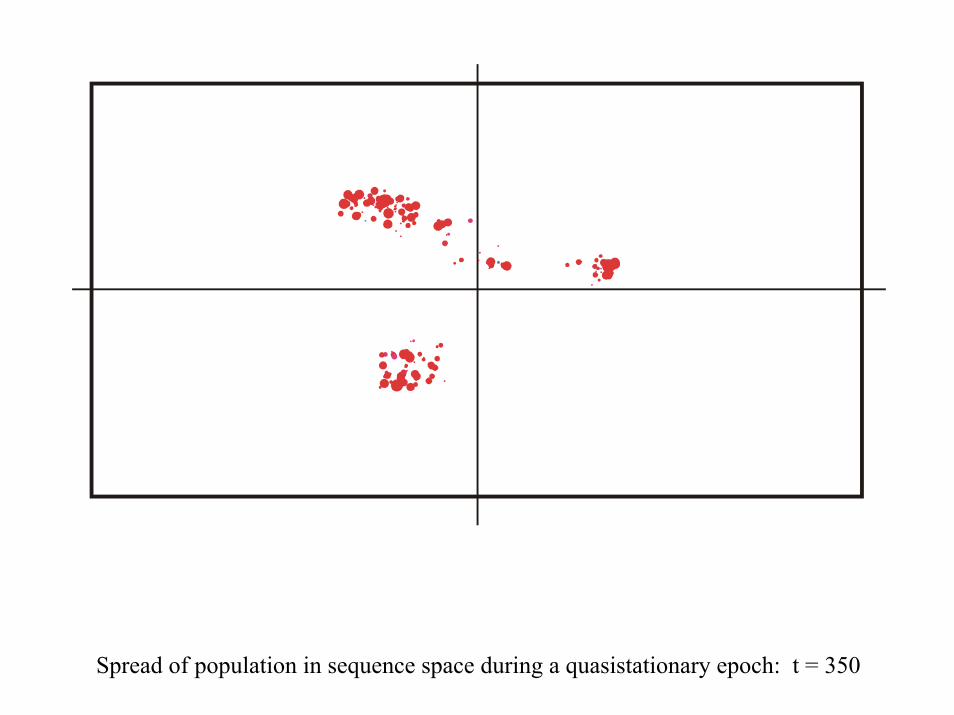
Spread of population in sequence space during a quasistationary epoch: t = 350
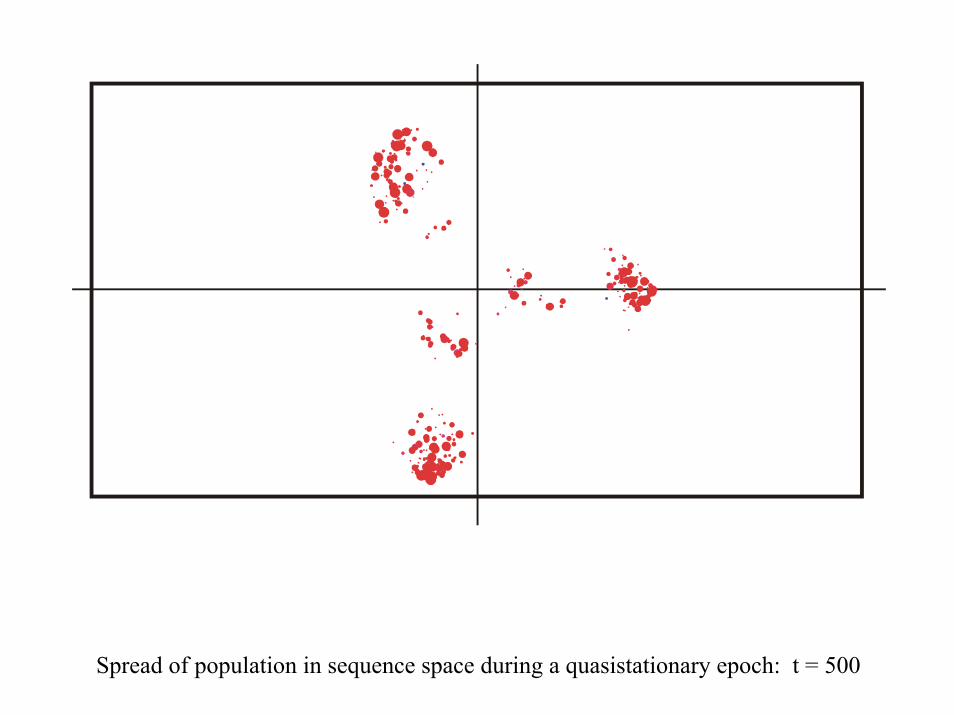
Spread of population in sequence space during a quasistationary epoch: t = 500
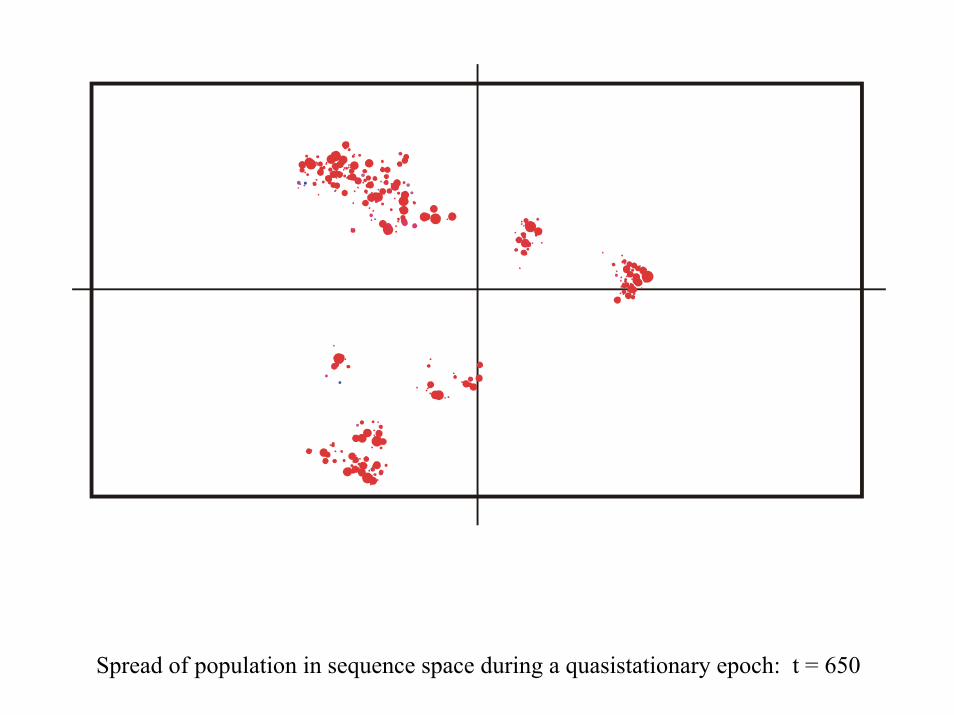
Spread of population in sequence space during a quasistationary epoch: t = 650
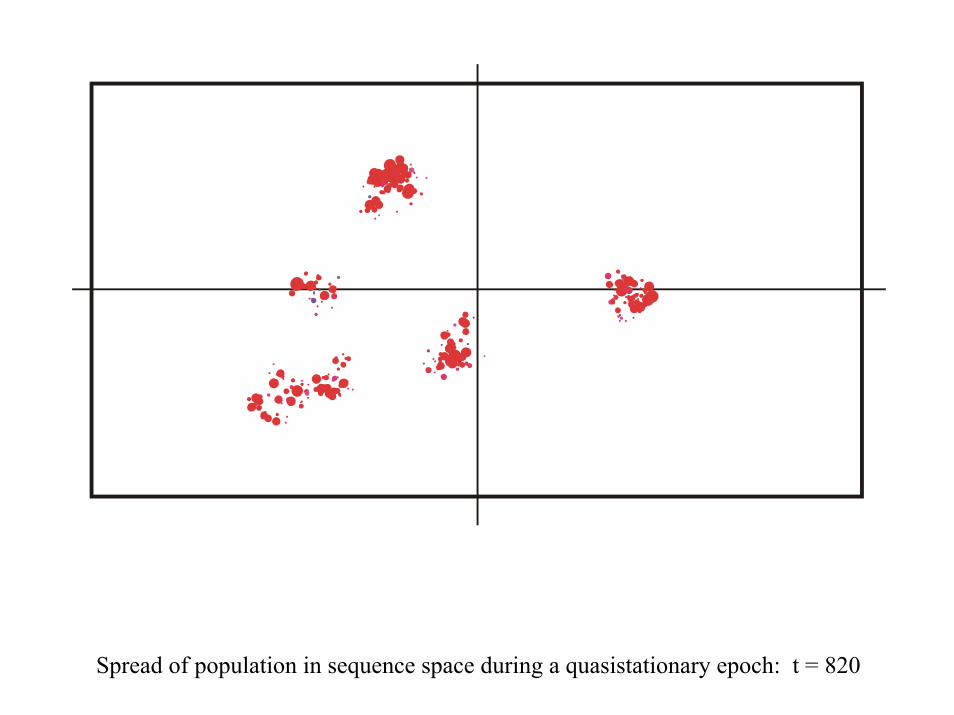
Spread of population in sequence space during a quasistationary epoch: t = 820
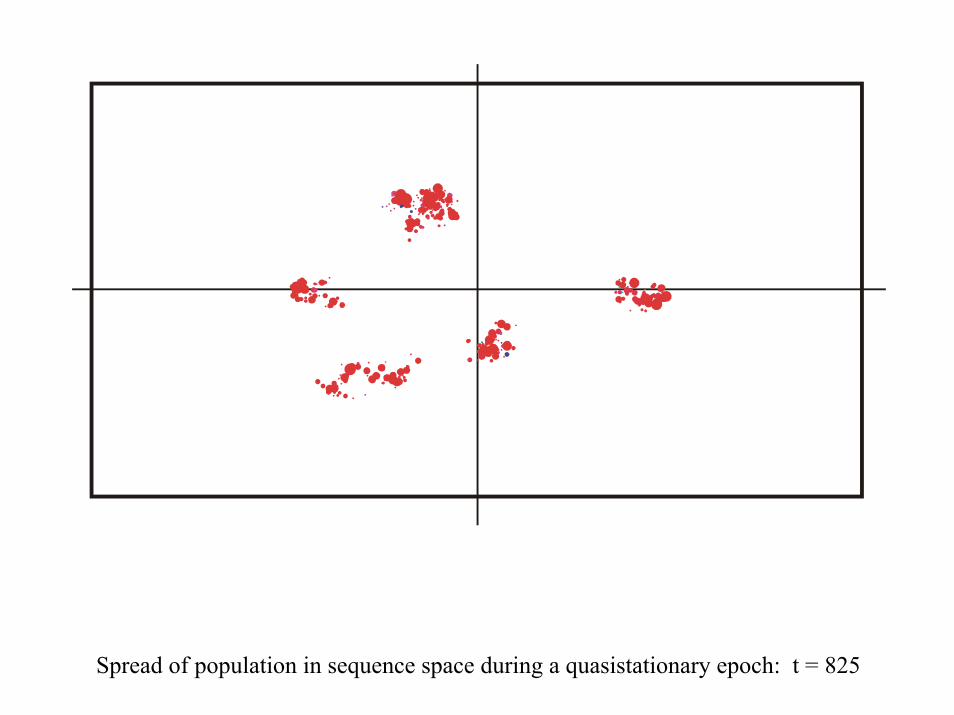
Spread of population in sequence space during a quasistationary epoch: t = 825
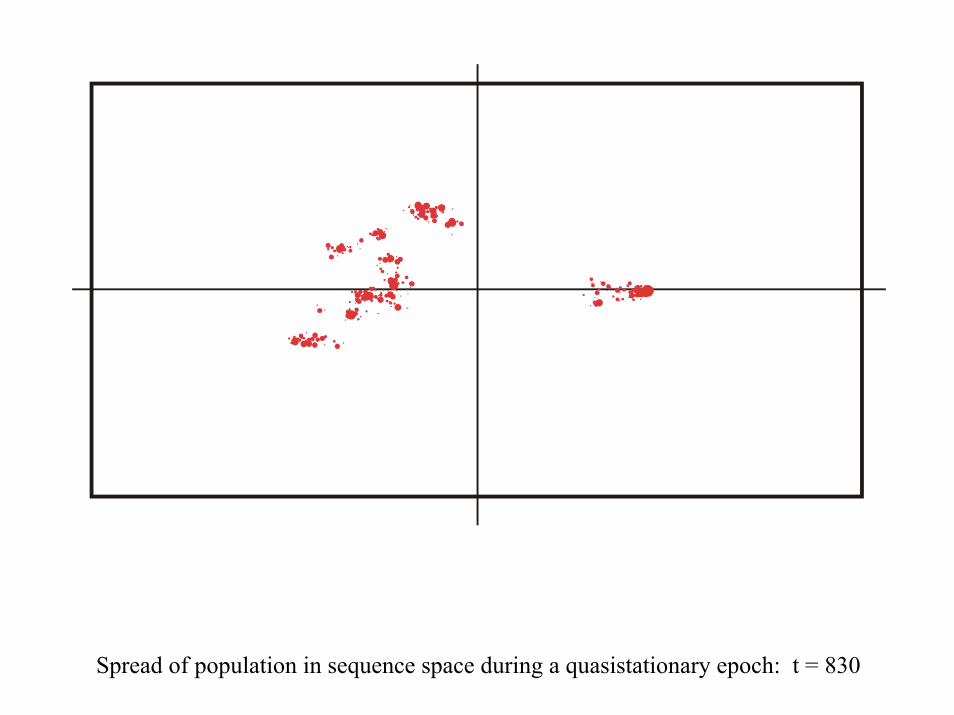
Spread of population in sequence space during a quasistationary epoch: t = 830
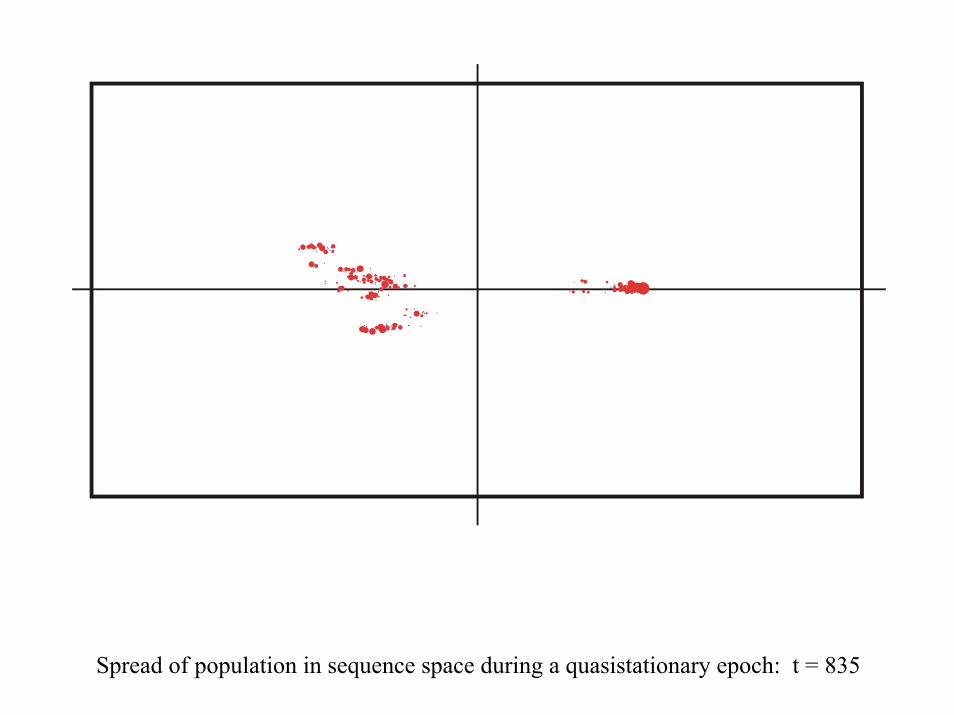
Spread of population in sequence space during a quasistationary epoch: t = 835
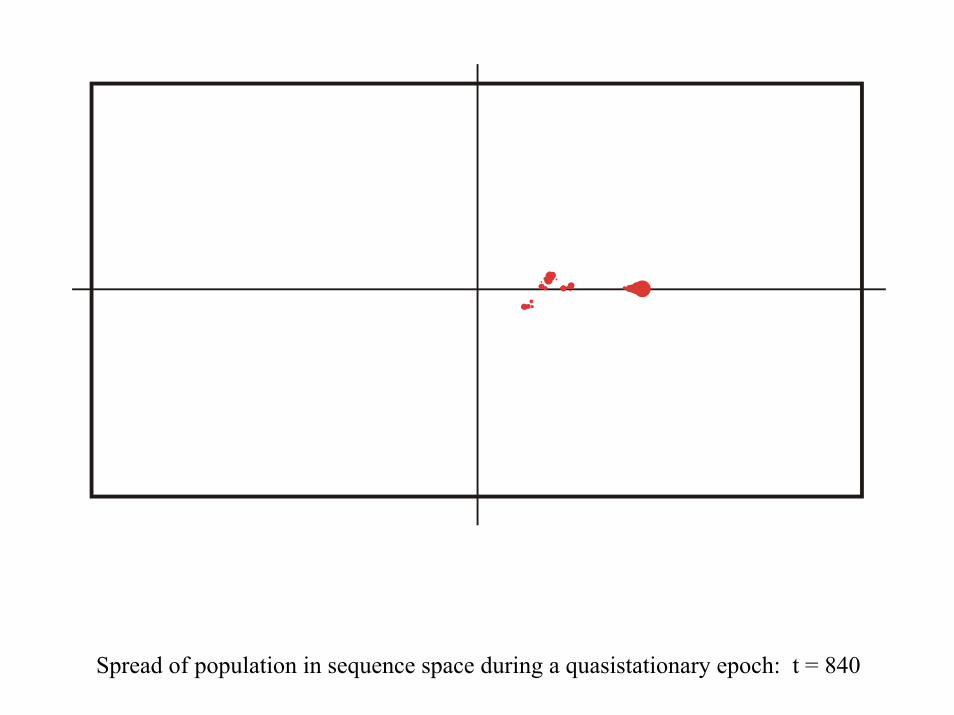
Spread of population in sequence space during a quasistationary epoch: t = 840
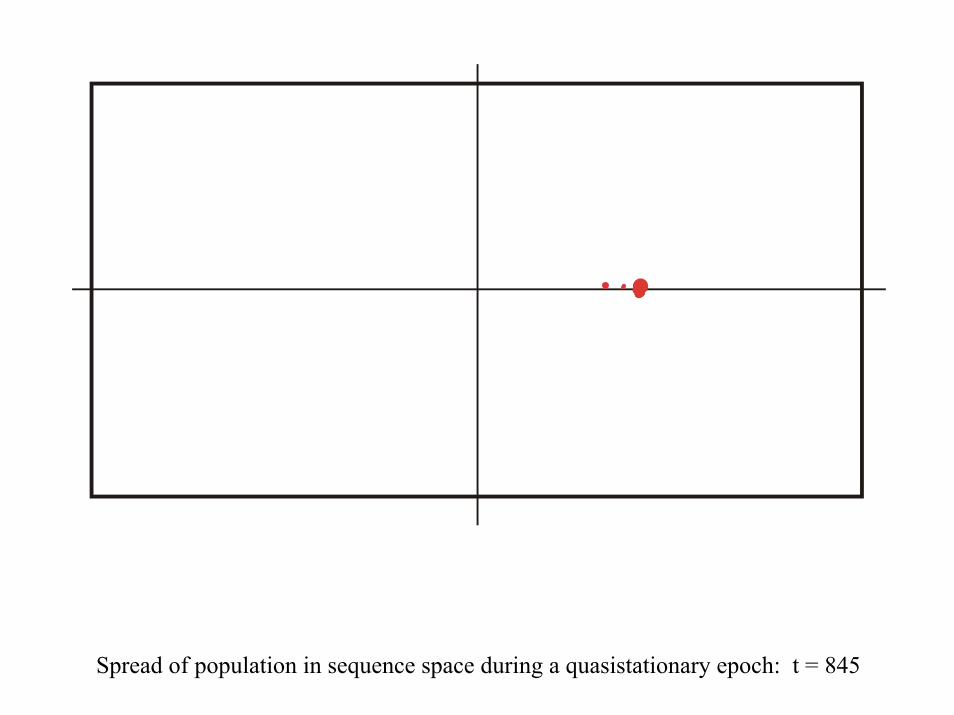
Spread of population in sequence space during a quasistationary epoch: t = 845
Spread of population in sequence space during a quasistationary epoch: t = 850
Spread of population in sequence space during a quasistationary epoch: t = 855
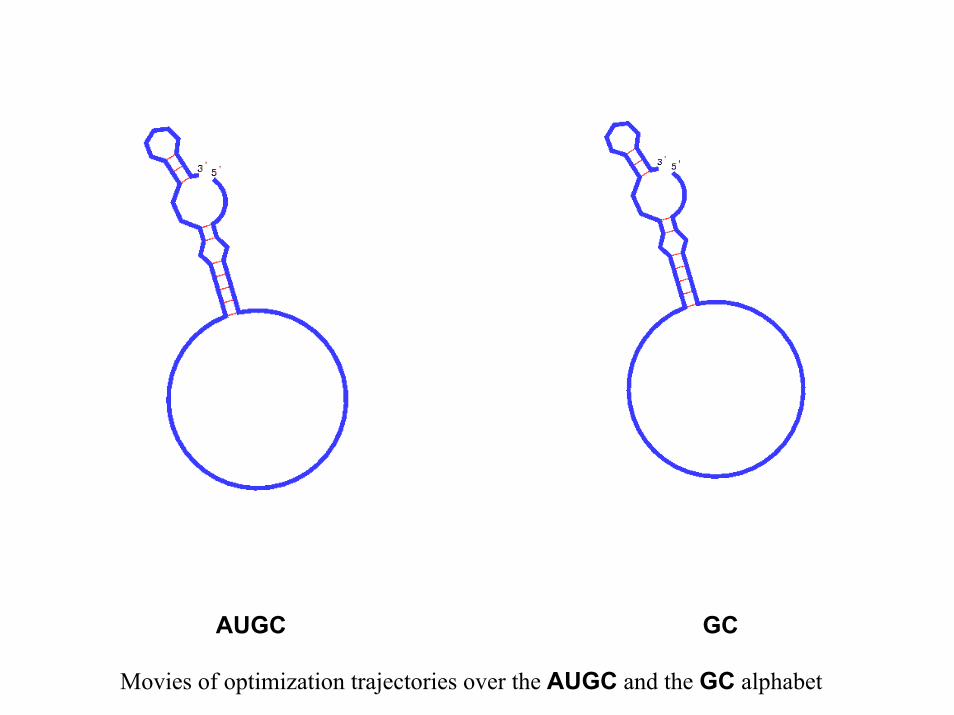
AUGC GC
Movies of optimization trajectories over the AUGC and the GC alphabet
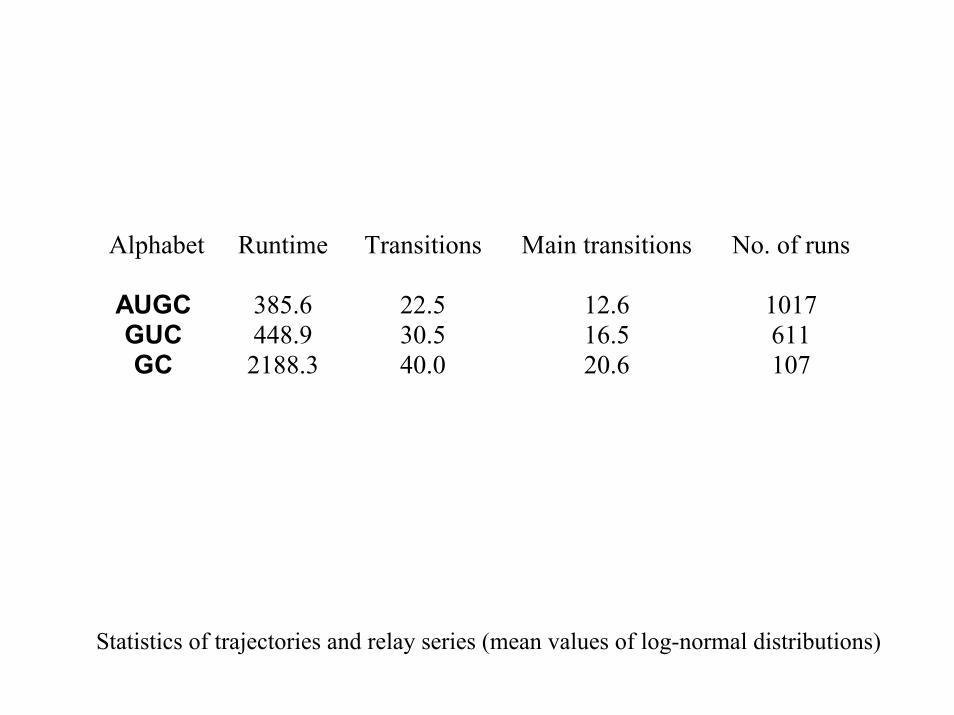
Alphabet Runtime Transitions Main transitions No. of runs
AUGC 385.6 22.5 12.6 1017 GUC 448.9 30.5 16.5 611 GC 2188.3 40.0 20.6 107
Statistics of trajectories and relay series (mean values of log-normal distributions)

Theory of sequence – structure mappings
P. Schuster, W.Fontana, P.F.Stadler, I.L.Hofacker, From sequences to shapes and back: A case study in RNA secondary structures. Proc.Roy.Soc.London B 255 (1994), 279-284
W.Grüner, R.Giegerich, D.Strothmann, C.Reidys, I.L.Hofacker, P.Schuster, Analysis of RNA sequence structure maps by exhaustive enumeration. I. Neutral networks. Mh.Chem. 127 (1996), 355-374
W.Grüner, R.Giegerich, D.Strothmann, C.Reidys, I.L.Hofacker, P.Schuster, Analysis of RNA sequence structure maps by exhaustive enumeration. II. Structure of neutral networks and shape space covering. Mh.Chem. 127 (1996), 375-389
C.M.Reidys, P.F.Stadler, P.Schuster, Generic properties of combinatory maps. Bull.Math.Biol. 59 (1997), 339-397
I.L.Hofacker, P. Schuster, P.F.Stadler, Combinatorics of RNA secondary structures. Discr.Appl.Math. 89 (1998), 177-207
C.M.Reidys, P.F.Stadler, Combinatory landscapes. SIAM Review 44 (2002), 3-54
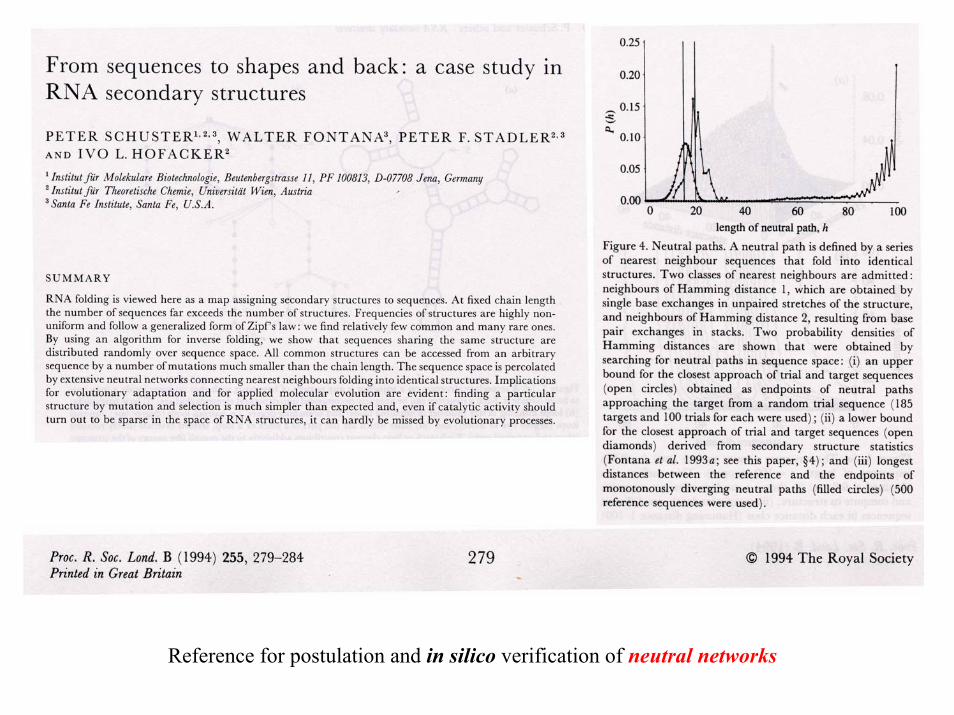
Reference for postulation and in silico verification of neutral networks
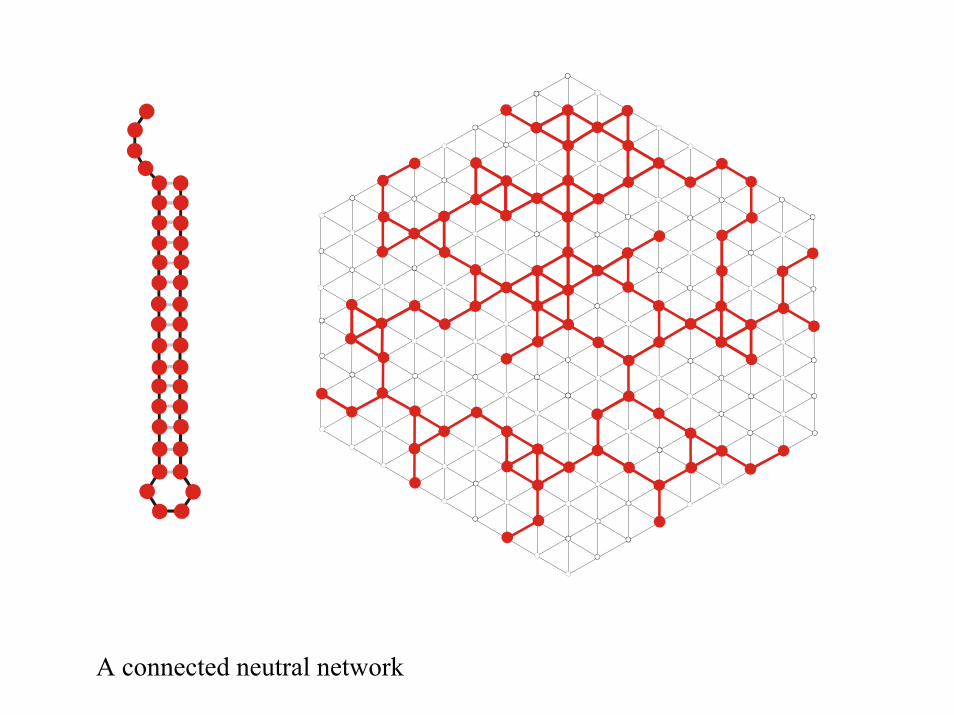
A connected neutral network
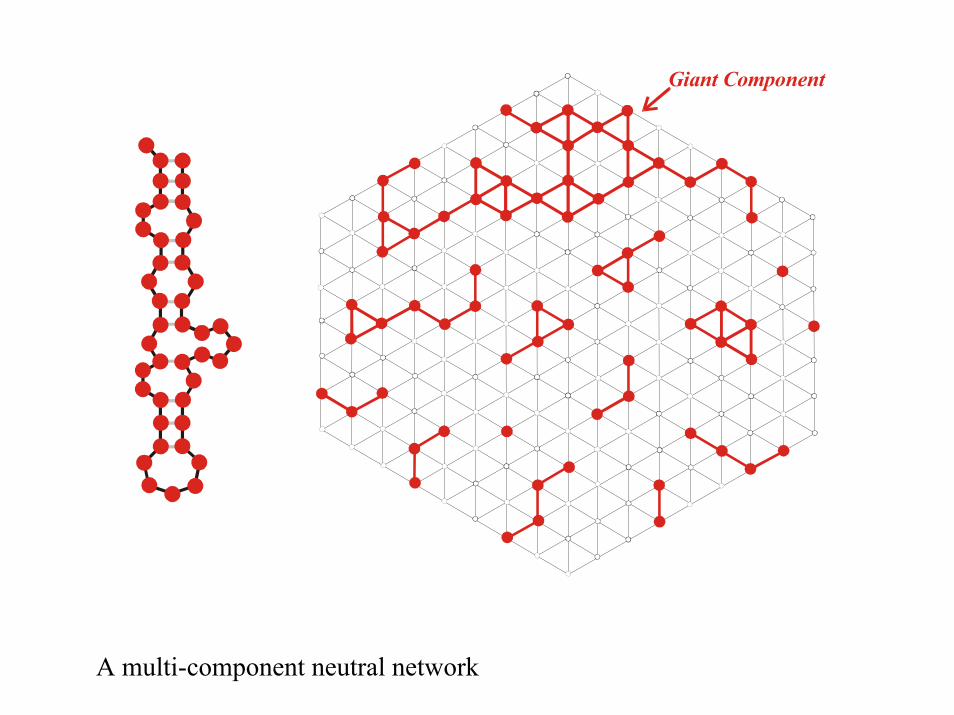
Giant Component
A multi-component neutral network

RNA 9:1456-1463, 2003
Evidence for neutral networks and shape space covering

Evidence for neutral networks and intersection of apatamer functions

Reference for the definition of the intersection and the proof of the intersection theorem
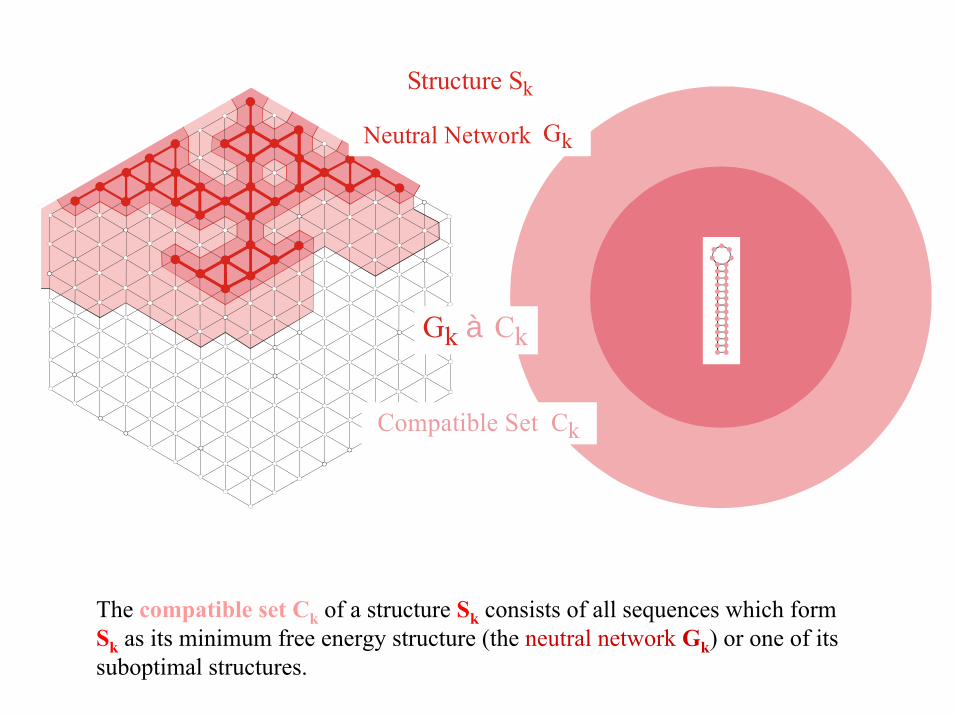
GkNeutral Network
Structure S k
Gk Cà k
Compatible Set Ck
The compatible set Ck of a structure Sk consists of all sequences which form Sk as its minimum free energy structure (the neutral network Gk) or one of itssuboptimal structures.
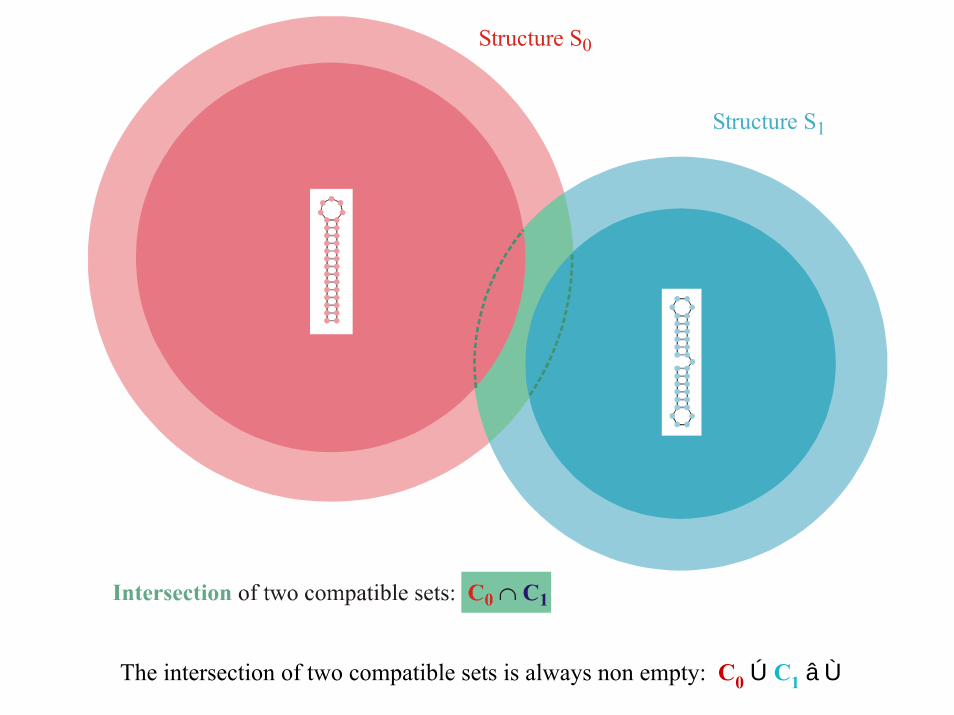
Structure S 0
Structure S 1
The intersection of two compatible sets is always non empty: C0 Ú C1 â Ù

A ribozyme switch
E.A.Schultes, D.B.Bartel, Science 289 (2000), 448-452
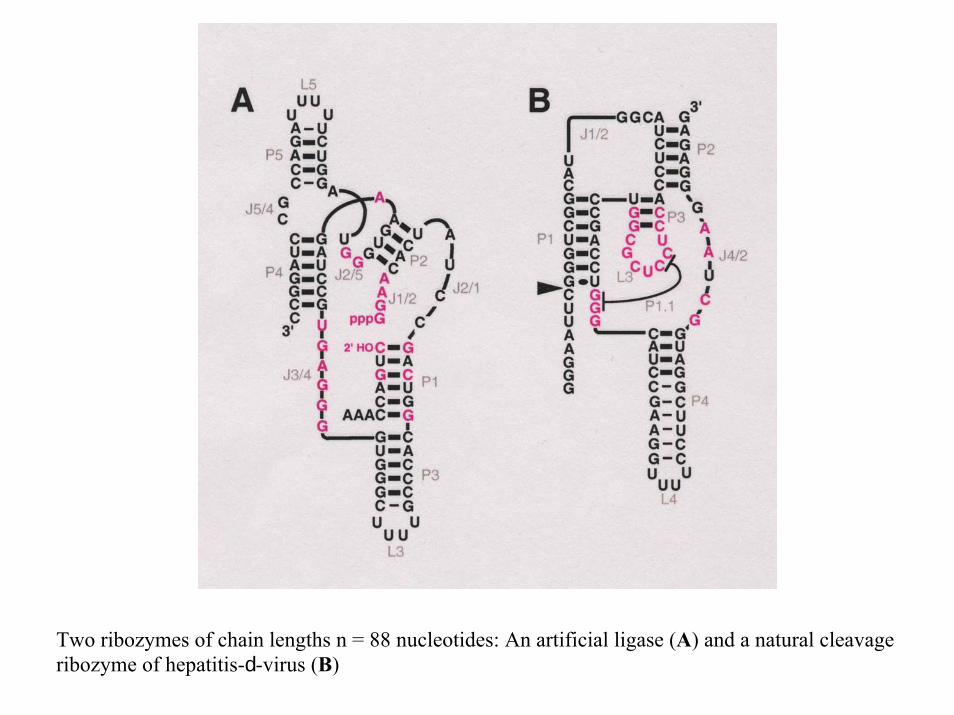
Two ribozymes of chain lengths n = 88 nucleotides: An artificial ligase (A) and a natural cleavage ribozyme of hepatitis-d-virus (B)
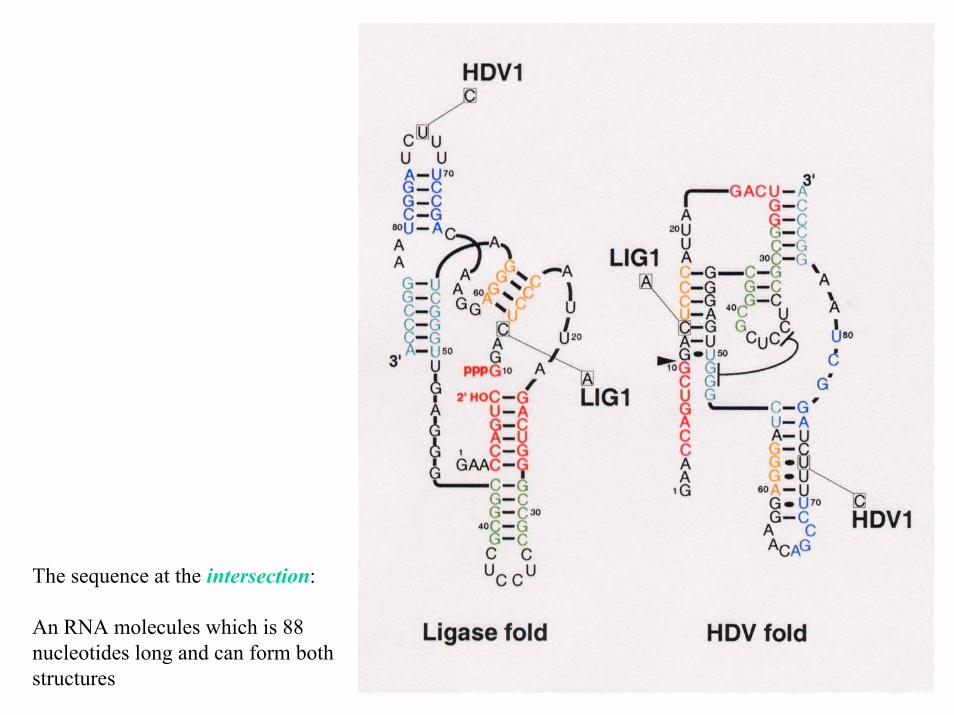
The sequence at the intersection:
An RNA molecules which is 88 nucleotides long and can form both structures
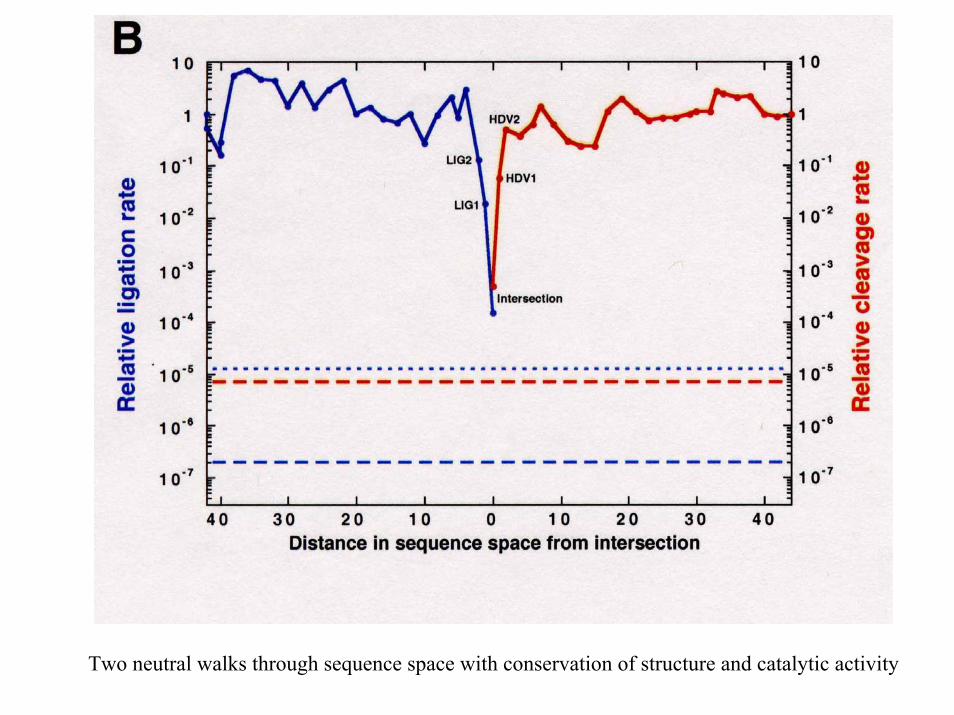
Two neutral walks through sequence space with conservation of structure and catalytic activity
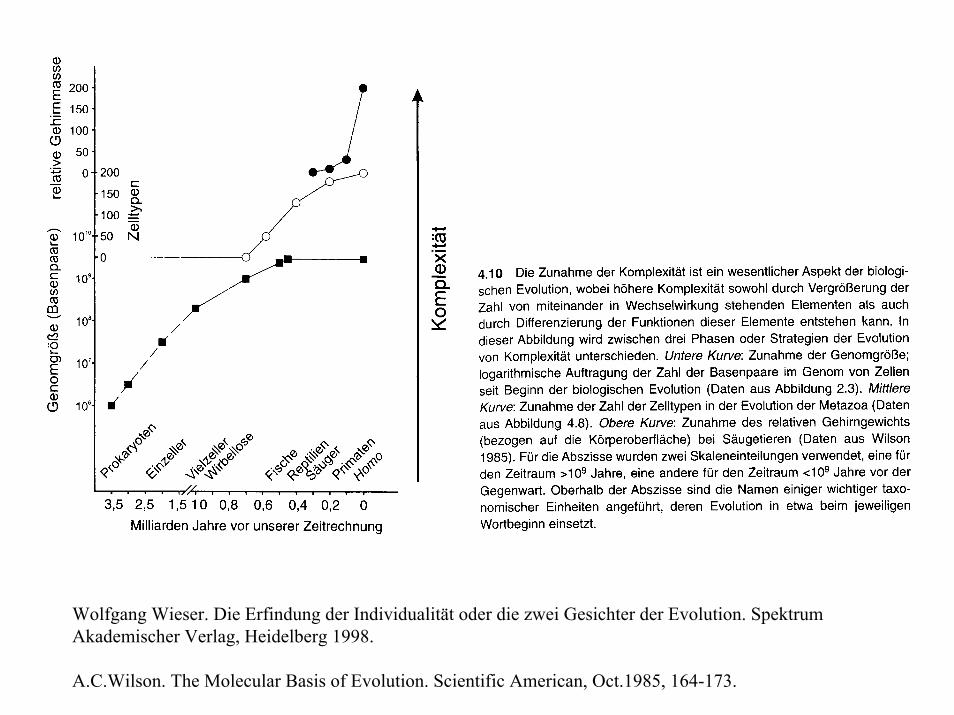
Wolfgang Wieser. Die Erfindung der Individualität oder die zwei Gesichter der Evolution. Spektrum Akademischer Verlag, Heidelberg 1998.
A.C.Wilson. The Molecular Basis of Evolution. Scientific American, Oct.1985, 164-173.

1968
2004Evolution (Cartoon Î 1980)

Acknowledgement of support
Fonds zur Förderung der wissenschaftlichen Forschung (FWF)
Projects No. 09942, 10578, 11065, 1309313887, and 14898
Jubiläumsfonds der Österreichischen Nationalbank
Project No. Nat-7813
European Commission: Project No. EU-980189
Siemens AG, Austria
The Santa Fe Institute and the Universität Wien
The software for producing RNA movies was developed by Robert Giegerich and coworkers at the Universität Bielefeld
Universität Wien

CoworkersUniversität Wien
Walter Fontana, Santa Fe Institute, NM
Christian Reidys, Christian Forst, Los Alamos National Laboratory, NM
Peter Stadler, Bärbel Stadler, Universität Leipzig, GE
Ivo L.Hofacker, Christoph Flamm, Universität Wien, AT
Andreas Wernitznig, Michael Kospach, Universität Wien, ATUlrike Langhammer, Ulrike Mückstein, Stefanie Widder
Jan Cupal, Kurt Grünberger, Andreas Svrček-Seiler, Stefan Wuchty
Ulrike Göbel, Institut für Molekulare Biotechnologie, Jena, GEWalter Grüner, Stefan Kopp, Jaqueline Weber

Web-Page for further information:
http://www.tbi.univie.ac.at/~pks