Embed Size (px)
Citation preview

Band XVII. Heft 3 . O r i g i n a l a u f s a t t e .
Studien zur Okologie und Epiderniologie der Insekten . 1 . Die Kieferneule. Panolis flammea Schiff .
W . Zwglfer. Miinchen . Von
(Aus dem Institut fur angewandte Zoologie.)
(Mit 2 Abbildungen und 24 Diagrammen.)
Inhaltsiibcrsicht . Vorbemerkungen . . . . . . . . . . . . . . . . . . . . . . . . 476 Allgemeines zu Material und Methode . . . . . . . . . . . . . . . . 477
b) Der Gesamtwiderstand . . . . . . . . . . . . . . . . . . 484 c) Die Populationsgleichung . . . . . . . . . . . . . . . . . 487
493 Beispiel t’: Prognosestellung . . . . . . . . . . . . . . . . 496 Beispiel 3: Mindestwirkungsgrad einer BekampfungsmaPnahme . . . . 497
von Temperatur und Luftfeuchtigkeit . . . . . . . . . . . . . . 500 a) Spezielles zur Methode . . . . . . . . . . . . . . . . . . 500 b) Die absolute Eizahl der Kieferneule . . . . . . . . . . . . . 501 c) Die ovariale Eientwicklung im Puppen- und Imaginalstadium . . . . 504 d) Das Schlupfen der Falter . . . . . . . . . . . . . . . . . 508 e) Geschlechtliche Aktivitat und Eiablage . . . . . . . . . . . . 512 f) Lebensdauer der Imago . . . . . . . . . . . . . . . . . . 521
Temperatur und Luftfeuchtigkeit . . . . . . . . . . . . . . . . 525 a) Spezielles zur Methode . . . . . . . . . . . . . . . . . . 525 b) DasEi . . . . . . . . . . . . . . . . . . . . . . . 528
d) Larve 11-V . . . . . . . . . . . . . . . . . . . . . 536 e) Puppe . . . . . . . . . . . . . . . . . . . . . . . . 537
IV . Epidemiologische SchluPfolgerungen . . . . . . . . . . . . . . . 540 V . Die Entwicklungsdauer der praimaginalen Stadien . . . . . . . . . . 544
VI . Hucger- und Fiitterungsversuche . . . . . . . . . . . . . . . 553 Zusammenfassung . . . . . . . . . . . . . . . . . . . . . 557 Literatur . . . . . . . . . . . . . . . . . . . . . . . . 559
Seito
. . . . . . . . . . . . . . . . . . . . . . . . I Zur Theorie 479 a) Der Gleichgewichtswiderstand . . . . . . . . . . . . . . . . 480
Beispiel 1: Priifuog einer Analyse . . . . . . . . . . . . . .
I1 . Die Zeugungsfahigkeit von Pmzolis flarrimea Sohiff . in ihrer Abhangigkeit
I11 . Die Vitalitat der praimaginalen Entwicklungsstufen in ihrer Abhangigkeit von
c) LarveI . . . . . . . . . . . . . . . . . . . . . . . 533
Zeitschrift fur angewandte Entomologie . XVJI. 3 . 32

47 6 Zwolfer:
Vorbemerkungen. AnlaB zur vorliegenden Untersuchung bot die letzte Kieferneulen-
kalamitat, die im Sommer 1930 in Hittelfranken und der Oberpfalz zur Eruption gelangte. l)
Nachdern bereits Ra tzeburg (52, 53) einen Zusammenhang zwischen Rlima und Eulengradation vermutete, und durch Berwigs (3) statistisch- meteorologische Arbeiten ein solcher in der Tat nacbgewiesen werden konnte, lag es nahe, mit Hilfe neuer Untersuchungsmethoden, wie sie von P i e r c e (50), Shel ford (61-64), Bodenhe imer (8-13) und anderen in die entomologische Porschung eingefuhrt morden sind, den Versiich zu unternehmen, diese Beziehungen im einzelnen zu ergrunden. 2) Damit ist der Gegenstand der vorliegenden Arbeit gekennzeichnet: Sie gibt die Er- gebnisse theoretischer Uberlegungen und experimenteller Untersuchungen wieder, von denen die letzteren in der Hauptsache sich rait dem EinfluB beschattigen, den Temperatur und Luftfeuchtigkeit - zwei okologisch be- sondars wichtige Klimafaktoren - auf verschiedene Entwicklungsstufen der Kieferneule ausuben. Im theoretischen Teil wird versucht, die epide- miologische Bedeutungen dieser Beziehungen i m Verhaltnis zu derjenigen anderer Umweltseinflusse in allgemein gultiger Form zu erortern.
DaB bei dem Nangel entsprechender Vorarbeiten uber die Kiefern- eule eine restlose Klarung der erwahnten Zusammenhange durch die an einer einzigen Generation gefuhrten Untersuchungen nicht moglich war, sei ausdrucklich erwahnt. Viele im Verlauf der Arbeit sich ergebende Einzelfragen muBten unberucksichtigt bleibeu, andere konnten nur provi- sorisch beantwortet werden, und sind in ihren Ergebnissen durch kunftige Porschungen zu revidieren und zu ergauzen.
Resultate experimenteller Untersuchungen der vorliegenden Art be- durfen zu ihrer Verallgemeinerung der Bestatigung durch Beobachtungen in freier Natur. Da detaillierte Freilanduntersuchungen uber die Kiefern- eule im Sinne dieser Arbeit bisher fehlten - das hieruber bislang bekannte ist in Sacht lebens (56) Monopaphie ubersichtlich ZusammengefaBt - wurden gleichzeitig durch E. Meyer entsprechende Beobachtungen im mittelfrankischen Gradationsgebiet ausgefuhrt, woruber vom Genannten an
Die Anregung zu ihr gab Herr Geheimrat Prof. Dr. K. E s c h e r i c h , dem ich fur die stets gewahrte verstaudnisvolle Unterstutzung und das groBe Interesse, daB er meinen Arbeiton entgegenbrachte, herzlichst danken mochte. - Dem grolzugigen Ent- gegenkonmen der Notgemeinschaft d e r D e u t s c h e n Wissenschaf t , welche die zu den experimentellen Arbeiten erforderliohen Einrichtungen zur Verfugung stellte, und jenem des Reichsernahrungsminis te r iums ist es zu danken, daB die Untersuchungen im geplanten Umfang ausgefuhrt werden konnten. Dank schulde ich fcrner meiner Frau: die mir bei der praktischen Durchfuhrung derselben als Mitarbeiterin tatig zur Seite stand.
') Die Bedeutung dieser Forschungsrichtung speziell fur das Studium der Forst- insekten voll erkannt und erstmals nachdriicklichst auf sie verwiesen zu haben, ist E s c h e r i c h s Verdienst (25).

Studien zur Okologie und Epidemiologie der Insekten. 477
anderer Stelle berichtet wird (46). Diese bezweckten im besonderen genaue phanologische Unterlagen und Anhaltspunkte fiber die mikroklimatischen Verhaltnisse zu liefern, wie sie im naturlichen Biotop der Kieferneule wahrend deren Entwicklungsperiode herrschen. Sie erganzen dergestalt die im Laboratorium auf experimentellem Wege gewonnenen Ergebnisse. Im einzelnen wird auf sie jm Laufe der Arbeit mehrfach zuruckgekommen
' werden.
Allgenaeines zw Material wnd k thode . Das zu den Versuchen verwendete Kieferneulenmaterial entstammte dem mittel-
frankischen und oberpfalzischen Gradationsgebiet, wo es im Laufe des Winters 1929/30 im Puppenstadium eingesammelt worden war.I) Vor dieser Zeit waren die Puppen den Einflussen ihres naturlichen Biotops unterworfen, die je nach Lage der Fundorte bis zu einem gewissen Grade differiert haben werden. Dieser Einweis ist notwendig, da gewisse Abweichungen in den Versuchsresultaten, die sich speziell bei den Puppenversuchen ein- stellten, wohl dem erwahnten Umstand zuzuschreiben und als Nachwirkungen zeitlich zuruckliegender schadlicher Einflusse zu deuten sind.
Auch das MiBlingen der Versuche zur Bestimmung des vitalen Optimums der 11. bis V. Larvenstufe hat ahnliche Ursachen. Dieses Material, das aus technischen Griinden nicht in den notwendigen Mengen im Laboratorium herangezuchtet werden konnte, wurde im Juni 1930 aus dem mittelfrankischen Gradationsgebiet bezogen. Bier war es in der voraufgehenden Zeit offenbar schiidigenden Umweltseinflussen in hohem MaBe ausgesetzt (Ansteckungsmoglichkeit durch Infektionskrankheiten !). Deren Nachwirkungen storten die entsprechenden Versuche in einer Weise, daB ihre Resultate fur den angegebenen Zweck unverwendbar blieben. Lediglich uber die Temperaturabhangigkeit der Entwicklungsdauer dieser Stadieu lieferlen sie Anhaltspunkte.
Demgegenuber wies das zu den Versuchen verwandte Falter-, Ei- und Junglarven- material, das aus Reservefreilandpuppen unter Laboratoriumsbedingungen gezuchtet wurde, hinsichtlich der Umweltseinflusse gleichartige - und wie sich spaterhin zeigte - dem vitalen Optimum dieser Stadien gut genaherte Vorbehandlung auf. Die Ergebnisse der hierhergehorigen Einzelversuche zeigten entsprechende befriedigende Obereinstimmung. a)
Bezuglich dieser Ergebnisse bedarf es noch eines weiteren allgemeinen und grund- satzlichen Hinweises: Sie fuBen auf Beobachtuugen an Versuchsmaterial, daB einer im ,,Eruptionsstadium" befindlichen Kieferneulengeneration entstammte. s, Die Moglichkeit, daB andere Generationen der Kieferneule, wie etwa solche, die einem Vorbereitungs- oder Krisenjahr angehoren, bezuglich ihrer Vitalitiit abweichende Verhaltnisse aufweisen, bleibt damil offen, und muBte durch kunftige Untersuchungen nachgepriift werden. Streng ge- nommen gelten somit die vorliegenden Resultate nur fur Kieferneulengenerationen im Eruptionsstadium einer Kalamitat, ohne vorerst verallgemeinert werden zu durfen. Wenn trotzdem in den folgenden Ausfuhrungen ein solcher Versuch unternommen worden ist. so desbalb, weil bislang noch bei keinem Insekt der exakte Nachweis eines Schwankens der , ,Reakt ionsnorm' von Generation zu Generation auf experimentellem Wege er- bracht norden ist. Lediglich theoretische MutmaDungen sind in dieser Richtung geauaert ~ o r d e n . ~ ) -
I) Seine Beschaffung verdanke ioh dem freundlichen Entgegenkommen verschiedener bayrisoher Forstiimter, insbesonders Eerrn Forstmeister Dr. Ku hn, Heideck.
2, In ahnlichem Sinn auBert sich J a n i s c h (35) gelegentlich seiner Untersuchungen uber die Baumwolleule Prodenza littoralis Boisd. uber die Wichtigkeit optimal vor- behandelten Versuchsmaterials.
Vgl. E s c h e r i c h (24). ") Horn (32).
32 *
478 Zwol fe r :
Die Einhaltung konstanter Temperaturen, wie sie zu den hier in Frage kommenden Experimenten erforderlich sind, wuide fur die Stufen zwischen 4 O - 2 2 O C durch einel; rnit Eiskuhlung arbeitenden, fiinfzelligen L a u t e n s c h lagerschen Reihenthermostaten be- werkstelligt, der in einem wahrend der kiihlen Jahreszeit geheizten Raume untergebracht war. Zu deu hoheren Temperaturstufen dienten mit Gasheizung versehene Einzelthermo- staten der ublichen Form. Samtliche Zellen, auch jene der Einzelthermostaten, waren wahrend der Versuchsdauer einseitig durch Glas abgeschlonsen. Alle Versuche liefen demnach unter diffusem Licht und erfullten somit die Forderung, die sich als Eonsequenz aus der diesbeziiglichen Untersuchung von F r i e d e r i c h s und S t e i n e r (29) ergibt.
Speziell fur die Puppenversuche - unter natiirlichen Verhaltnissen liegt die Kiefern- eulonpuppe bekanntlich in der Waldstreu unter Lichtabschluh - ware an sich d i e An-
%
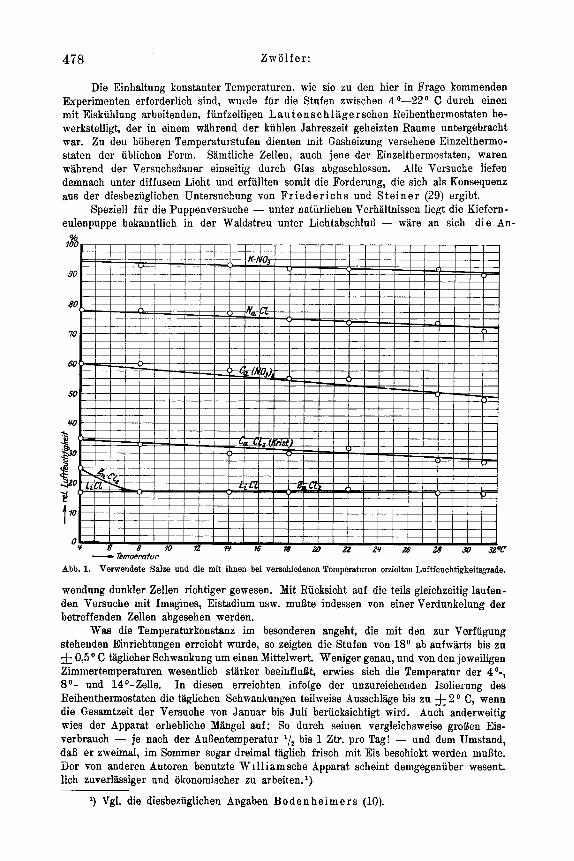
Abb. 1. Verwendete Sake nnd die rnit ihnen bei verscliiedenen Temperaturen erzielten Luftfeuchtigkeitsgrade.
wendung dunkler Zellen richtiger gewesen. Yit Rucksicht auf die teils gleichzeitig laufen- den Versuche mit Imagines, Eistadium usw. multe indessen van einer Verdunkelung der betreffenden Zellen abgesehen werden.
Was die Temperaturkonstanz im besonderen angeht, die mit den zur Verfugung steheuden Einrichtungen erreicht wurde, so zeigten die Stufen van 18O ab aufwarts bis zu & 0,5O C ttiglicher Schwankung um einen Mittelwert. Weniger genau, und von den jeweiligen Zimmertemperaturen wesentlich stiirlier beeinfluat, envies sich die Temperatur der 4 O-,
8O- und 14O- Zelle. In diesen erreichten infolge der unzureicheiiden Isolierung des Reihenthermostaten die tiiglichen Schwankungen teilweise Ausschlie bis zu & 2 O C, wenn die Gesamtzeit der Versuche van Januar bis Juli beriicksichtigt wird. Auch anderweitig wies der Apparat erhebliche Mange1 auf: So durch seinen vergleichsweise grolen Eis- verbrauch - je nach der AuPentemperatur 1/2 bis 1 Ztr. pro Tag! - und dem Umstand, daB er zweimal, im Sommer sogar dreimal ti5glich frisch mit Eis beschickt werden mul3te. Der von anderen Autoren benutzte W ill iamsche Apparat scheint demgegenuber wesent. lich zuverksiger und okonomischer zu arbeiten. I )
I) Vgl. die diesbeziiglichen Angaben Bod e n h e i m e r s (10).

Studien zur Okologie und Epidemiologie der Insekten. 47 9
Im ubrigen wurden wahrend der gesamten Versuchstatigkeit die Zellentemperaturen zweimal tlglich abgelesen - einmal vor, ein andermal ca. 1 Stunde nacb der Eisfiillung - und aus den gefundenen Verten, die annahernd den Naximal- und Minimalzellentempe- raturen entsprechen, das Mittel gebildet. So ergaben sich fur die verschiedenen Tempe- raturstufeu im Durchschnitt Mitteiwerte von ruud 4 O , 8O, 14O, 18O, 22O, (2S0), (27O), 28O, (32O) C.l)
Zur Erzielung und Erhaltung bestimmter relativer Luftfeuchtigkeitsgrade fand in den einzelnen Versuchen das Prinzip konstanter Dampfspannung konzentrierter Salz- losungen Verwendung, das bereits von amerikanischen Autoren 2, und neuerdings auch YOU J a n i s c h (35) und A n d e r s e n (1) zu ahnlichen Zwecken benutzt wurde.
Uber seine Anwendungsweise bei der vorliegenden Untersuchung im besonderen ist alles Nahere aus der Seite 526-527 gegebenen Schilderung zu entnehmel;. Hier sei nur eine Zusammenstellung der in feuchtem Zustand verwendeten Salze gegeben und der relativen Luftfeuchtigkeitsgrade: die rnit ihnen bei den in Betracht kommenden Tempe- raturen erreicht wurden. Bus dem Diagramm Abb. 1 sind alle erforderlichen Einzel- heiten zu ersehen. Bemerkt sei, daB die Messung der Feuchtigkeitsgrade mit dem kleinen L a m p r e chtschen Haarhygrometer unter Verwendung der weiter unten beschriebenen ,,Hygrostaten" ausgefuhrt worden ist. Das MePinstrument wurde vor jeder Benutzung frisch justiert und verblieb im ubrigen zu jeder Nessung ruod zwei Stunden im ,,Zucht- raumcL des mit dem feuchten Salz beschickten Hygrostaten. Seine Genauigkeit betragt nach Angaben der Berstellungsfirma & 3 o/o.
Die 100 Oi0 Fenchtigkeitsstufe wurde mit H,O dest. erzeugt, die O-lOo/o Stufe mit Phosphorpentoxyd in trockenem Zustand. Vollig trocken gibt letzteres 0 o/io relative Luftfeuchtigkeit. Bei Wasseraufnahme, wie sie sich im Laufe der Versuche notwendiger- weise einstellt, steigt dieser Feuchtigkeitsgrad stets etwas an. Dasselbe gilt auch fur einige andere stark hygroskopische Salze, wie LiCl und CaCI,. Die Salzfullungen der Hygrostaten miissen daher wahrend der Versuchsdauer von Zeit zu Zeit erneuert werden. Nieht ge- nugende Beachtung dieses Umstandes kann zu Fehlresultaten fuhren.
Zur anschaulichen Darstellung der gefundenen Ergebnissa fand in der Regel die schon von P i e r c e (50), S h e l f o r d (61, 63), B o d e n h e i m e r (9, 10) und anderen be- nutzte Methode Verwendung, die im Prinzip einer dreidimensionalen graphischen Dar- stellung entspricht und gegeniiber anderen Versuchen dieser Art 2, wie auch rnit der ein- fach zweidimensionalen Darstellungsweise verglichen, wesentlich ubersichtlicher ist.
I. Znr Theorie.4) Der Besprechung der speziellen Versuchsergebnisse seien jene theore-
tischen Uberlegungen vorausgeschickt, die den Gang der experimentellen Untersuchungen wesentlich beeinflufiten. Sie bilden im ubrigen einen Versuch, Hilfsmittel zu einer quantitativen Behandlung des Massenwechsels der Insekten zu schaffen.
I ) Die in Elammer gesetzten Zahlen sind Temperaturmittel, die nur bei einzelnen
2, Vgl. F r i e d e r i c h s (27), S h e l f o r d (64). s, Vgl. W e b e r (72). 4, Der folgende Abschnitt gibt in der Hauptsache die Zusammenfassung von Er-
gebnissen einer an anderer Stelle (77) ausfuhrlich veroffentlichten Studie. Einige der in ihm enthdtenen Gedankengange entspringen neuen Gesichtspunkten und erweitern - ge- nauer gesagt ,,vereinfachenU - das bei friiherer Gelegenheit Mitgeteilte. Neu und zum Teil auf den Erfahrungen der verflossenen bayeriechon Eulenkalamitat fuPend, sind auch die am SchluP des Abschnitts gegebenen 3 Beispiele fur die praktische Anwendung der Populationsgleichung.
Veisuchen erreicht wurden.

480 Zwolfer :
Quantitative Porschungsmethoden setzen MeQbarkeit der untersuchten Erscheinungen voraus. 1st diese Voraussetzung erfullt, und sind die Er- scheinongea gesetzmafiig verknupft, dann muQ auch eine mathematische Behandlung ihrer gegenseitigen Beziehungen moglich seio.
Wohl von ahnlichen Erwiigungen ausgehend, ist von seiten medixinischer Entomo- logen speziell in der Malariaforschung eine mathematische Formulierung epidemiologischer Probleme schon seit langem angestrebt worden: Mannern wie R. Ro13 (54), S e l l a (60), Mart in i (38-40, 42) gebuhrt das Verdienst, auf dem Gebiet bahnbrechend gewirkt zu haben. In den letzten Jahren versuchts Vol te r ra (70, 71) das Bevolkerungsproblem im Tierreich unter rein biozonotischen Gesichtspunkten in ahnlicher Weise zu bearbeiten. Auch die von der menschlichen Bevolkerungsstatistik ausgehenden Untersuchungen von P e a r 1 (48) zielen auf eine mathematische Formulierung der in Betracht kommenden Fragen. Auf wirtschaftlich entomologischem Gebiet hat diese Behandlungsart des Stoffes neuerdings in Thompson (66, 67), C h a p m a n (15-17) und B r e m e r (14) Bearbeiter gefunden.
Auf die Gedankengange der genannten Autoren kann im Rahmen dieser Arbeit nur soweit kurz eingegangen werden, als sich unmittelbar Beriihrungspunkte mit den hier vorgetragenen Uberlegungen ergeben. Die im folgenden mitgeteilte Gleichung (1) wurde in anderer Form und etwas anderem Zusammenhang schon von B r e m e r (14) aufgestellt. Was den Ausdruck ,,Widerstand der Umwelt" angeht, so ist er bereits von Chapman (15-17) fur den in Frage kommenden Begriff verwendet worden.
a) Der Gleichgewichtswiderstand. Zwei Krafte sind es, deren Wirken das organische Gleichgewicht in
der Natur regelt: Die Zeugungskra f t der Organismen auf der einen - der Wide r s t and d e r Umwel t auf der anderen Seite. Das Resultat des Spieles dieser Krafte ist die Popu la t ionsd ich te der einzelnen Organismenarten, d. h. die Zahl der Individuen einer Art auf irgend eine Einheit ihres Lebensraumes bezogen.
Theoretisch sind die genannten 3 GroBen mit beliebiger Genauigkeit meBbar: Die ,,Zeugungskraft" durch die von einem Weibchen durchschnitt- lich produxierte Ei- oder Nachkommenzahl, der , ,Widerstand de r Um- welt" durch den Anteil vernichteter Nachkommen am ZweckmaBigsten in Prozenten der erzeugten Nachkommenschaft ausgedruckt, die ,,Po pu- l a t ionsdichte" endlich durch die auf irgend eine Lebensraumeinheit redueierte Individuenzahl der Art.
Man kann gegen diese Betrachtungsweise einwenden, daB bei ihr kein ,,Messen" im eigentlichen Sinn, sondern nur ein ,,Zahlen'' vorliegt. ,,Messen" setzt gleiche Be- schaffenheit der zu messenden Dinge voraus. ,,Zahlen" abstrahiert von ihren Eigen- schaften, sieht auch verschiedene Dinge als gleichartig an, und ordnet sie einander zu. Dieser Gesichtspunkt ist von M a r t i n i (44) in anderem Zusammenhang in der Tat geltend gemacht worden: ,,Man hat nicht bedacht, da13 die einzelnen Entwicklungsstadien unter sich verschieden sind und sie als gleichwertig behandelt. Sie mussen aber mit ver- schiedenen Werten eingesetzt werden. Man muP sie wiigen nicht zahlen."
Er richtet sich zunachst - und dies mit vollem Recht - gegen die mehrfach vertretene Buffassung, die epidemiologische Bedeutung eines Umweltfaktors komme erst dann richtig zum AUS- druck, wenn seine Wirkung, statt auf die jeweils vorhandene Population, auf die Ausgangs- eizahl umgerechnet worden ist. In diesem Sinn ist der Einwand in den folgenden Aus- fuhrungen speziell in der weiter unten mitgeteilten Populationsgloichung insofern beriick-
Die Berechtigung dieses Einwandes SOU nicht bestritten werden.

Studien zur Okologie und Epidemiologie der Insekten. 48 1
sichtigt, als die einzelnen Widerstandskomponenten sich auf die jeweils vorhandene Population beziehen, auf die sie unmittelbar einwirktsn.
Wenn im obigen Zusammenhang von einem ,,Messen" gesprochen worden ist, so wird dabei die Gleichheit der Individuen in d e r Zugehor igke i t zu e in u n d der- selben Ar t erblickt, unter Abutrahieruog von den morphologischen und okologischen Verschiedenwertigkeiten der einzelnen Entwicklungsstufen. -
hderungen eines im Gleichgewicht befindlichen Systems setzen hderungen der wirkenden Krafte voraus: jede Mehrung oder Minderung des ,,Widerstandes der Urnwelt'' und ebenso jede Mehrung oder Minde- rung der Zeugungskraft kann theoretisch eine Verschiebung des Systems bedingen. Ausdruck dieser Verschiebung sind die von Generation zu Generation beobachtbaren Schwankungen der Popuiationsdichte einer Art an einem bestimmten Biotop. Verschiebungen in der einen Richtung wahrend einer oder mehrerer Generationen werden in den folgenden erfahrungsgemaB durch Verschiebungen in entgegengesetzter Richtung aus- geglichen. Unter der Voraussetzung, daS innerhalb einer gegebenen Popula- tion keine grundsatzliche und dauernde Anderung einer der beiden wirkenden Krafte eintritt, wird daher im Laufe zahlreicher Generationen ein Gleichgewichtszustand angestrebt. Seiner wahren Natur nach stellt er ein dynamisches Sys tem vor.
Ep idemien s ind ze i twei l ige Verschiebungen d ieses Sys t ems i m S i n n e e i n e r e rhebl ichen Zunahme d e r Populat ionsdichte . Bus dem Gesagten folgt, daB sie durch h d e r u n g e i n e r der beiden wirkenden Krafte (Zeugung und Widerstand) oder durch e i n e gleich- s i n n i g e A n d e r u n g be ide r e u s a m m e n bedingt sein konnen.
Durchschnittliche Nachkommenzahl einer Art und Widerstand der Umwelt sind von Art eu Art und Ort eu Ort verschieden. Auch von Generation zu Generation konnen sie innerhalb gewisser Grenzen Schwan- kungen unterworfen sein. Letzteres ist fur den Widerstand der Umwelt hinlanglich bekannt. Was die Nachkommeneahl angeht, so mebren sich in neuerer Zeit Beobachtungen, die fur die Richtigkeit des Satzes sprechen. Neben artspezifischen Eipenschaften hat EinfluB auf sie das Geschlechter- verhaltnis, der Ernahrungseustand der Elterntiere, klimatische Ein wirkungen am Aufenthaltsort der Tiere wahrend der Zeugungsperiode, ja sogar bio- tische Paktoren, wie Sterilitat der Weibchen infolge Parasitierung.
Streng genommen miil3te demnach bei mathematischer Formulierung der Beeiehungen ewischen ,,Zeugungskraft" und , ,Widerstand d e r U m w elt" mit dnderung beider Krafte von Generation zu Generation gerechnet werden.1) lndessen erweist es sich als zweckmafiig und liil3t
l) Dieser Gedankengang ist in der anderen Orteri veroffentlichten Studie (77) konsequent durchgefuhrt. Der hier eingeschlagene in einigen Punkten von der dortigen Darstellung etwas abweichende Weg, andert nichts am Wesen der Formeln, die in jener Abhandlung abgeleitet sind, und deren wichtigste hier iibernommen wurden. Er gestattet durch die Erweiterung des Begriffes ,,Widerstand der Urnwelt" vereinfachte Anwendung der Glei- chungen.

482 Zwolfer :
sich auch theoretisch begriinden, die eine der beiden wirkenden Krafte, die . ,Zeugungskraf t" als kons t an t und ar t spez i f i sch anzusehen, wie das bisher in der Xegel auch schon geschehen ist. Darnach wurde jeder Insektenart eine ,,ideale" oder Zeugungskraft zukommen, der gegenijber die in freier lu'atur oder unter wechselnden Versuchsbedingungen erzielten Werte der Ilurchsohnittsnachkommenzahl in den meisten Piillen nur ,,relative" Bedeutung besitzen. Der Wer t d e r , , re la t iven Durch- schn i t t snachkommenzah l " i s t v o n den o b e n g e n a n n t e n u n d viel- l e i c h t auch noch von a n d e r e n Umwel t se in f lus sen abhangig . D ie , , a b s ~ l u t e ' ' Z e u g u n g s k r a f t e i n e r Ar t kommt d e m g e g e n u b e r n u r u n t e r dem Opt imum samt l i che r m a S g e b e n d e r Bed ingungen z u r v o 11 e n A u s wi r k u ng.
Die Differenz der Werte der , ,absoluten" und der , , relativenb' Durchschnittsnachkommeuzahl 1aSt sich als eine Reduktion der artspezifi- schen Zeugungsfahigkeit auffassen, die durch Urn weltseinflusse bedingt ist. Dami t e r f a h r t d e r oben d e f i n i e r t e Begriff d e s W i d e r s t a n d e s d e r U m w e l t e i n e E r w e i t e r u n g : Neben dem Ante i l v e r n i c h t e t e r I n d i v i d u e n d e r Genera t ion e i n e r A r t umfal j t e r auch d i e Redu'x- t i on i h r e r abso lu t en Nachkommenzahl . MiRt man die Reduktion in Prozenten der absoluten Zeugungskraft der Art, dann kann dieser Wert rechnerisch in derselben Weise behandelt werden, wie jener Umwelts- widerstand, der in Prozent vernichteter Individuen einer Generation ge- messen wurde.
Dies laLlt sich leicht nachweisen: Angenommen, die absolute Zeugungskraft einer Art sei e. Durch widrige Umweltseinflusse moge sie wahrend einer Generation um w0/,, reduziert werden. Dann gelangen in der betreffenden Generation im Durchschnitt pro
Weibchen nur e - -- '" oder e (1 - 5) Eier zur Ablage.
EMte sich die absolute Zeugungskraft voll auswirken konnen, dann waren im Durch- schnitt e Eier j e Weibchen zur Ablage gelangt. Wenn n a c h erfolgter Eiablage vun diesen durch abiotische oder biotische Umweltseinfliisse w vernichtet worden waren,
80 wiirde ein Rest von e - - oder e 1 - - an ifberlebenden zurucligeblieben sein.
B U S der Gleichartigkeit beider Ausdriicke ist zu ersehen, daB AS rechnerisch belanglos bleibt. ob ein Widerstand vor oder nach erfolgter Eiablage wirkte, wenn er in der an- gegebenen Weise gemessen worden ist.
So ist beispielsweise die absolute Zeugungskraft der Kieferneule mit 190 anzusetzen.1) An Hand von Meyer s (46) Freilandbeobachtungen, die durch eigene Untersuchungen 2) eine gewisse Bestiitigung finden, war im Porstamt Heideck im Eruptionsjahr 1930 die durchschnittlich von einem Weibchen abgelegte Eizahl 130. Der Umweltswiderstand, der eine Reduk- tion der absoluten Zeugungskrsft von 190 auf 130 zur Polge hatte, war demnach im vorliegenden Fall gleich 60 Eier je Weibchen oder in Prozent
100
e w 100 ( ;0)
l) Vgl. S. 501ff. dieser Arbeit. Vgl. S. 520 dieser Arbeit.

Studien zur ekologie und Epidemiologie der Insekten. 183
der absoluten Zeugungskraft der Art ausgedriickt gleich 32 %. - Sacht - l e bens (56) Beobachtungen an der Kieferneule im Zossener Revier ergaben 1925 eine durchschnittliche Eiablage je Weibchen von 30 Eiern. In dieseni Fall war der Umweltswiderstaud, der die Reduktion der idealen Eizahl von 190 auf 30 bedingte in Prozent ausgedruckt gleich 84%. Die beiden Beispiele zeigen gleichzeitig, welch erheblichen Schwankungen die Umweltswiderstande unterliegen, die die absolute Eizahl in den einzelnen Generationen reduzieren. -
Die Beziehungen, die zwischen ,,Z e ug ungs k raf t" und , ,Wider s t an d d e r Um we 1 t" bestehen, wenn der Gleichgewichtszustand der Art in auf- einanderfolgenden Generationen gewahrt bleiben soll, sind bereits von B r e m e r (14) allgemein formuliert worden. I n der im folgenden benntzten Fassung lautet der Aiisdruck 1) :
111 + 100 (e - ,-! e w, = . . . . . . . (1)
Hierin stellt W, den Prozentsatz for, welcher von der theoretisch irn Optimum aller Bedingungen moglichen Nachkommenschaft durch den Widerstand der Umwelt ausgemerzt werden mud, um den Gleichgewichts- zustand zu erhalten. e ist der Wert der absoluten Zeugungskraft der Art, m : f deren Geschlechterverhaltnis. Nach den obigen E r l a u t e r u n g e n umfadt somi t d e r W i d e r s t a n d s w e r t W, n i c h t n u r d e n A n t e i l d i r ek t v e r n i c h t e t e r Nachkommen , s o n d e r n a u c h d i e Reduk t ion d e r abso lu t en Zeugungskra f t d e r Art.
Dieser der obigen Gleichung entsprechende Widerstandswert W,, der fur den Fall der Erhaltung des organisohen Gleichgewichtes besteht, sei kurz als , ,Gleichgewicbtswiderstand" bezeichnet. Nach dem Gesagten kommt ihm bis zu einem gewissen Grad ebenso wie der ,,absoluten" Zeugungskraft a r t sp ezif i sc h e Bed eu t un g zu.
Fur den ,,Gleichgewichtswiderstand'l gilt dies genau genommen, nnr fur den Fall, dsB das Geschlechterverhaltnis der Art von Generation zu Generation konstant bleibt. 1st letzteres Schwankungen unterworfen - nnd es liegen Beobachtungen vor, die fur eine derartige Moglichkeit sprechen, - dann muBte auch der Gleichgewichtswiderstand einer Art als variable GrdPe angesehen werden. - Doch 1aBt sich demgegenuber auch die Anschauung vom artspezifisohen Geschlechterverhaltnis vertreten, ahnlich wie dies bei der ,,abso1utenLb Zeugungskraft naher ausgefuhrt worden ist. Etwaige Schwankungen des Geschlechterverhaltnisses in aufeinauderfolgenden Generationen wurden dann sinngemaB auf Umweltswiderstande zuriickgefuhrt werden und sich in einer Steigerung oder Ver- minderung der relativen Durchschnittsnachkommenzahl je Weibchen auBern, je nachdem das Verhaltnis zugunsten der Weibchen oder der Mannchen eine Verschiebung erfuhr.
Fur die Kieferneule beispielsweise mit einem Geschlechterverhaltnis von rund 1 : 1 und einer absoluten Eizahl von 190 ist der Wert des Gleich- gewichtswiderstandes:
l) Wegen der Ableitung dieser nnd der weiter unten mitgeteiltqn Gleichnagen mu13 auf die ausfuhrliche Arbeit (77) verwiesen werden.

484 Z w o l f er:
100 (190 - F) 190---- W, =
wo = 98,95.
Nit anderen Worten : 98,95 O//o der theoretisch im Optimum aller Bedingungen moglichen Nachkommenschaft der Kieferneule muB von eigener Fortpflanzung ausgeschaltet werden, wenn der Gleichgewichts- zustand erhalten bleiben soll. Dabei kann dieser Prozentsatz teils durch Reduktion der absoluten Eizahl, teils durch direkte Vernichtung der Nach- kommenschaft als Auswirkung abiotischer oder biotischer Widerstands- komponenten erreicht werden.
In derselben Weise laBt sich fur jede Art, deren ,,absolute" Zeugungs- kraft und deren Geschlechterverhaltnis bekannt sind, der Wert ihres Gleich- gewichtswiderstan desermitteln. Zunachst mehr von theoretischem Interesse, erlangt er im weiteren Zusammenhang betrachtet praktische Bedeutung. -
b) Der Gesamtwiderstand. Fur die Berechnung des gesamten Umweltswiderstandes, der auf
irgend eine Generation einer Insektenart einwirkte, lassen sich zwei Gleichungen ableiten, die im folgenden mitgeteilt seien.
Bezeichnet man mit PI die Populationsdichte einer Insektenart an einem gegebenen Biotop zu Beginn einsr Generation, mit P, die Popu- lationsdichte zu E n d e der Generation, nennt man ferner die ,,absolute" Zeugungskraft einer Art e, und ihr Geschlechterverhaltnis m : f, dann gilt f iir den Gesamtwiderstand W,, der auf die betreffende Generation ein- wirkte. die Formel
100 ( P ~ . e - p2 . m + f -i-> PI . e (2) W, = e . . . . .
Hiernach ist der Gesamtwiderstand W,, der auf die Generation einwirkte, bestimmbar, wenn Ausgangs- und Endpopulationsdichte der Generation durch Beobachtung ermittelt worden, und absolute Nachkommen- zahl sowie das Geschlechterverhiiltnis der Art bekannt sind. N a c h d i e s e m A u s d r u c k b e r e c h n e t g i b t W, den jen igen P rozen t sa t z a n , d e r Ton d e r im Opt imum a l l e r Bed ingungen moglichen Nachkommen- s c h a f t d e r Ausgangspopula t ion i n d e r be t r e f f enden Genera t ion v e r n i c h t e t wurde. Er urnfafit demnach sowohl d i e Reduk t ion der absolu ten E izah l a l s auch d.en d i r ek t v e r n i c h t e t e n An te i l g ez e u g t e r Nac h kommen.
Fur die praktische Anwendung ist eine Modifikation des obigen Ausdruckes zur Bestimmung des Gesamtwiderstandes vorteilhaft. Sie kann durch algebraische Umformung aus Gleichung (2) abgeleitet werden und lautet :
(3) . . . . . . 1Oo(P, - Pz) + w o P2
P I W, =

Studien zur Okologie und Epidemiologie der Insekten. 485
Hiernach ist der Gesamtwiderstand W, in gleicher Weise wie oben bestimmbar, wenn bekannt bezw. durch Beobachtung ermittelt worden sind, Ausgangs- und Endpopulationsdichte der Generation , sowie der ,,Gleichgewichtswiderstand" Wo der betreffenden Art.
Ein Beispiel moge die praktische Anwendung des Gesagten erlautern: S c hwerd t- f e g e r (59, der den Kiefernspanner in der Letzlinger Heide untersuchte, gibt eine Zu- sammenstellung der Durchschnittspuppenzahlen, die bei Probesammlungen in einzelnen aufeinanderfolgenden Jahrgiingen je Quadratmeter Bodenstreu festgestellt wurden. I) In der folgenden Tabelle sind seine Werte zusammengestellt :
Eerbst, Jahrgang 1924 1925 1926 1W7 1928 1929 Puppenzahl je qm
(im Gesamtdurchsch.) 0,14 0,92 1,11 8,71 33,04 30,OO Wie groB war in den einzelnen Generationen der Gesamtwiderstand der Umwelt?
- Das Geschlechterverhaltnis in der Generation 1927/28 betrug nach Schwerdt f e g e r m : f = 63 : 37, also rund m : f = 2 : 1. Die Durchschnittszahl pro Weibchen (,,absolute Zeugungskraft") sei nach NuSl in (47) mit e = 120 angesetzt. Nach Gleichung (1) be- tragt dann in diesem Fall der Gleichgewichtswiderstand:
I 1. Vor- 2. Vor- 2. Eruptions- / Prodromal- 1. Eruptions- jahr, Beginn
jahr jahr i jahr jahr der Krise bereitungs- bereitungs-
180. (120 - 'T) 120 - = 97,5. w o =
Generation . . . I 1924/25 1 1925126 i 1926/27 I 1927/28 Gesamtwiderstand . 83,57% 96,98 "//, 80,38O/, ~ 90,52"/,
1928.29 97.73
l) Mit Rucksicht auf den besonderen Gegenstand dieser Arbeit ware es naheliegend gewesen , ein Beispiel aus der Kieferoeulenliteratur heranzuziehen. Leider ist keine derartig vollstiindige Zahlenreihe, wie sie S c h w e r d t f e g e r fur den Kiefernspanner gibt, in der Eulenliteratur vorhanden. Dolies (18) hat zwar durch 12 Jahrgange hindurch Probesammlungen nach Forleulenpuppen veranstaltet, teilt aber nur fur 3 derselben nurchschnittspuppenzahlen mit. Da es im obigen Zusammenhang mehr darauf ankam die prinzipielle Seite der Methode zu zeigen, wurde die vollstlindigere Zahlenreihe Sc h w er d t- f e g e r s gewahlt. Die Puppenzahl fur 1929, die bei S c h w e r d t f e g e r fehlt, verdanke ich Herrn Dr. Schot te . Berlin.

486 Z w o l f e r :
Wahrend in den Generationen 1924-1928 die Werte k s Gesamtwiderstandes u n t e r h a1 b des Gleichgemichtswiderstandes lagen, was den lAusbruch einer Kalamitat bedingte, iibertraf er ihn im Jahrgang 1928E9 urn 0,23O/,. Damit war die Erise der Gradation einkeleitet.
Welche Folgsrungen ergeben sich hieraus ? Die mitgeteilten Werte fur den Gesamtwiderstand Iassen erkenneii, in welchem AusmaB dieser schwanken kann. Sie zeigen ferner zahlenmaBig, daB der AnstoR zu der Gradation in eine Zeit fallt, in der die Zunahme der Population des Schadlings sich noch in keiner Weise durch auffallend vermehrten FraB bemerkbar machte. Des weiteren ist ihnen zu entnehmen, daI3 die groRte Zunahme der Schadlingspopulation keineswegs im Eruptionsjahr einer Kalamitat liegt, wie vielfach irrig angenornmen wird, sondern in den vorhergehenden Jahrgangen. Endlich geht aus den mitgeteilten Wider- standswerten noch hervor, daB - um e i n e Erup t ion zu e rz i e l en - s t e t s mehre re Genera t ionen au fe inande r folgen mussen , in denen de r Gesamtwiders tand un t e rha lb des Gle ichgewichtswiders tandes f u r d i e be t re f fende A r t liegt.
Weitere systematisch durch Jahre hindurch fortgefuhrte Freiland- untersuchungen in dieser Richtung wurden zu einer Kenntnis der n i ede r s t en und hochs ten Widerstandswerte sowie ihrer samtlichen Zwischenstufen fuhren, die innerhalb eines gegebenen Biotops fur einen bestimmten Schadling herrschen. Der Vergleich der gefundenen Werte mit den Klima- verhaltnissen der zugehorigen Jabrgange wiirde vermutlich ergeben, daB einer bestimmten Klimakonstellation im allgemeinen ein bestimmter Wert des Gesamtwiderstandes zugeordnet ist. 1) 1st noch die Populationsdichte der betreffenden Schadlingsart fur normale Jahre bekannt, dann liefie sich die Steigerung oder Verniinderung der Population von Jahr zu Jahr rein rechnerisch mit Hilfe der Widerstandswerte und der meteorologischen Daten verfolgen , ohne daB dazu weiterhin umstandliche Einzeluntersuchungen von Fall zu Fall notwendig waren. Uberraschungen fur die Praxis wurden auf diese Weise ausgeschlossen sein.
Die Richtigkeit dieser Uberlegungen wird bis zu einem gewissen Grad bestatigt durch die Untersuchungen von B e r w i g (3) uber Klima und Kieferneulengradation und entsprechende von Ecks te in (19) iiber den Kiefernspanner. Beide Untersuchungen lassen die klimatische Be- dingtheit der Massenvermehrungen dieser Schadlinge erkennen, beide zeigen, daB das erste Jahr mit klimatischen Besonderheiten der Eruption um mehrere Jahre vorausgeht, die Arbeit des letztgenannten Autors deutet ferner daranf hin, daB mehrere Jahre mit gunstigen Kiimabedingungen einander folgen miissen, ehe das Eruptionsjabr erreicht wird. Unter Be- rucksichtigung obiger U berlegungen wiirden derartige Untersuchungen eine noch prazisere Fassung dieser Verhaltnisse gestatten. -
l) Auch Shelford (61) iiuBert sich in diesem Sinne.

Studien zur Okologie und Epidemiologie der Insekten. 487
Der Chapmanschen (15-17) Formel C = Bp (,,the number of insects is equal to
the potential rate of production dividet by the resistance of the environment") liegt ein ahnlicher Gedanke zu Grunde, wie der obigen Bestimmung des Gesamtwiderstandes der auf die Generation einer Insektenart einwirkt. Bus C h a p m a n s Anwendungsweise seiner Formel (17) ist zu entnehmen, da13 er unter C die Endpopulationsdichte versteht, die im obigen [GLeichung (2) nnd (3)] mit P, bezeichnet wurde. Der Wert des , ,Biot ischen Po ten t i a l s 'L seiner Gleichung entspricht der Nachkommenzahl, die von der Ausgangs-
population gezeugt wurde. Es ist demnach Bp = PI . e . ~~ -- C h a p m a n s Widerstand m +f '
R endlich. ist der reziprolie Wert des Bruches, der angibt wieviel von der gezeugten Nachkommenschaft vernichtet wurde. Im Gegensatz hierzu gibt der oben gebrauchte Widerstandswert Wx den P r o z e n t s a t z von Individuen an, der von der im Optimum aller Bedingungen moglichen Nachkommenschaft vernichtet wurde. Unter Berucksichtigung dieser Uberlegungen laBt sich zwischen dem Chapmanschen Widerstandswert R und dem hier gebrauchten Wx folgende Beziehnng ableiten, die ermoglicht den einen Wert in den anderen zu uberfuhren.
R
Es ist namlich: 100 (R - 1)
R * R=-- loo und WX =
100 - Wx
c) Die Populationsgleichung. Die Vorausbestimmung der Zu- und Abnahme der Populationsdichte
einer Insektenart ist eines der Hauptziele epidemiologischer Borschung. Sie setzt voraus, dal3 die wesentlichen Umstiinde die das Schwanken der Populationsdichte der Art bedingen, bekannt und zahlenmaBig meBbar sind. 1st dies der Fall, dann muD eine mathematische Formulierung der gegenseitigen Beziehungen der angedeuteten GroBan moglich sein. Das Folgende ist ein Versuch in dieser Richtung.
I n s e i n e r Gesamthe i t e r s c h e i n t d e r Wide r s t and de r Umwel t a l s Resu l t an te e ines Sys tems v o n Wide r s t andskomponen ten oder Einze lwiders tanden . So werden bekanntlich unter den Einfliissen der Umwelt, die auf das Leben der Insekten einwirken, solche abiotischer und biotischer Art unterschieden. Erstere unifassen - um nur einige der wesentlichsten zu nennen - Klima, Boden usw., letztere jene der Er- n,%hrung, Krankheiten, Feinde, Parasiten, Konkurrenz usf. 1) Wie schon aus dieser Zusammenstellung hervorgeht, sind diese Widerstandskomponenten ihrem Wesen nach auBerordentlich mannigfaltig. In ihrer W i rkun g indessen - sofern man diese in der Vernichtung lebender Individuen erblickt - stimrnen sie iiberein. Wie der Gesamtwiderstand in Prozent der theoretisch moglichen Nachkommenschaft gemessen wurde, so sind die Einzelwiderstande in Prozent des jeweils vernichteten lndividuenauteils einer Population meDbar.
Neben diesen direkt wirkenden Einzelwiderstanden miissen noch solche unterschieden werden, welche durch Beeintrachtigung der Zeugungsfahig keit eine gegebene Insektenpopulation indirekt beeinflussen. Ihr Wesen wurde
l) Ausfiihrlich sind sie bei F r i e d e r i c h s (27) und Shelf ord (64) zusammengestellt.

488 Zwolfer:
bereits eingangs kurz angedeutet. Was ihren Wirkungsgrad angeht, so kommt er in der Reduktion der absoluten Eizahl der betreffenden Insektenart zum Ausdruck. MiBt man diese Reduktion in Prozent der absoluten Ei- zahl, dann lassen sich auch die indirekt wirkenden Widerstandskomponenten mit den direkt wirkenden vergleichen. Beide konnen alsdann - wie friiher gezeigt wurde - in gleicher Weise rechnerisch behandelt werden.
Unter Beriicksichtigung dieser Uberlegungen ist es moglich einen allgemeinen Ausdruck aufzustellen, der alle fur die Schwankungen der Populationsdichte einer Art unmittelbar maBgebenden Glieder vereinigt. Er gestattet die Berechnung der Populationsdichte zu Ende einer Generation (Pg), wenn bekannt sind Ausgangspopulationsdichte (PI) der Art, ihr Geschlechter- verhaltnis (m : f), ihre absolute Eizahl (e) sowie samtliche Widerstands- komponenten (wl, wz, w3 . . . . wn), die nacheinander auf die betreffende Generation einwirkten. Dabei ist vorausgesetzt, daB die direkt wirkenden Einzelwiderstande in Prozent des jeweils vernichteten Individuenanteils der 2opulation gemessen wurden, die indirekt wirkenden in Prozent der Redubtion , welche die absolute Eizahl in der betreffenden Generation erfuhr. Hiernach lautet die Formel zur Bestimmung der Endpopulations- dichte (Px) un ter Beriicksichtigung der Einzelwiderstiinde :
p--l': ( ") ( ") (1-s) . . . . ( 1 - 5 ) (4) " - m + f * 100 100 100
Es ist ohne weiteres einzusehen, daB der auf Grund der Gleichung (4) ermittelte Wert der Populationsdichte P, um so genauer sein und den wirklichen Verhaltnissen entsprechen wird, je vollstandiger die Zahl der wirkenden Widerstandskomponenten erfaBt, und je genauer deren Werte ermittelt wurden. Alle in der angegebenen W eise meBbaren Einzelwider- stande lassen sich der Gleichung einfugen. Es gilt dies beispielsweise fur die Ernahrung, wenn durch Hunger eir bestimmter Prozentsatz von Individuen vernichtet wurde, fur alle Bekampfungsmahahmen - gleichviel ob technischer oder biologischer Art - fiir Anderungen des Lebensraumes durch IGingriffe der Natur oder des Menschen in das von der Art be- wohnte Biotop usw. Auch die durch Abwanderung der Population ver- loren gegangenen Individuen konnen in der Gleichung Beriicksichtigung finden, vorausgesetzt, daB sie prozentual erfaBbar sind. Ihr Prozentwert ware sinngemZiB wie die Wirkung einer einzelnen Widerstandskomponente zu behandeln. -
Der Vollstandigkeit wegen sei kurz darauf hingewiesen, daB Gleichung (4) sich in vereinfachter Form auch folgendermaIden schreiben 1aBt :
PI . E . f . U, . u,. us. . . . . Un . . . . -__ (m + f ) . lO@ Px = . . (5)
Hierin sind u,, u2, us, . . . . . Un, die Prozentsatze der nach Einwirkung von w,, w,, w3, . . . . . Wn Wideistandskomponenten jeweils uberlebenden Individuen einttr Population.

Studien zur Okologie und Epidemiologie der Insekten. 489
Da namlich u, + wI = 100 ist, und somit w, = 100 - u, gesetzt werden kann,
geht der Ausdruck (1 - w l ) durch Umformung in (&I iiber. Entsprechend laBt sich
in der Populationsgleichung der Ausdruck ( 1 - ~~~ 2) a - durch (&) ersetzen usw., woraus
schlieBlich die vereinfachte Form (5) der Populationsgleichung resultiert. Aus didaktischen Griinden wurde indessen in dieser und auch in der anderen Ortes gegebenen Darstellung der Form (4) der Populationsgleichung der Vorzug gegeben. -
Unter Zugrundelegung von Gleichiing (4) ist es moglich - wie bei friiherer Ge- legenheit gezeigt wurde - einen allgemeinen Ausdrucli aufzustellen, der die Populations- dichte nach Ablauf beliebig vieler (q) Generationen angibt, ausgedruckt in der absoluten Zeugungskraft der Art, ihrem Geschlechterverbaltnis, ihrer Ausgangspopulationsdichte und den samtlichen Einzel- oder Gesamtwiderstiinden, die wiihrend der q-Generation wirkten.
Wird in dieser erweiterten Populationsgleichung angenommen , daB die Gesamt- widerstande wahrend q aufeinanderfolgender Generationen unterhalb des Gleichgewichts- widerstandes der Art liegen, so wiirde fur q = a die Endpopulationsdichte der Art theo- retisch ebenfalls den Wert Q) erreichen. Die Unmoglichkeit des letzteren ist leicht ein- zusehen, und im einzelnen durch Untersuchungen von P e a r l (48) und Chapman (15 bis 17) nacbgewiesen, welche gezeigt haben, daB ein bestimmter S a t t i g u n g s g r a d d e r Bevolkerungsdich te einer Art in einem gegebenen Lebensraum nicht uberschritten werden kann.
Theoretisch folgt aus diesen Uberlegungen, daB bei standig wachsender Bevolkerunps- dichte einer Art, in dem Augenblick, wo diese ihren Sattigungsgrad erreicht, zwangslaufig neue Umweltswiders~nde in Erscheinung treten mussen, die durch die Ubervolkerung selbst bedingt sind, und deren WirkungsgroBe nunmehr jene der iihrigen Einzelwider- stande ubertrifft. Vol t e r r a s (70, 71) Untersuchungen behandeln im wesentlichen diesen Sonderfall, der bei Erreichung des Sattigungsgrades der Bevolkerungsdichte einer Art gegeben ist und der bei Schadinsekten 'in der Regel nur gelegentlich und vorubergehend in der Natur verwirklicht wird. -
Die wichtigsten theoretischen Polgerungen, die sich aus der Diskussion der Populationsgleichung (4) ergeben, lassen sich folgendermaBen zu- sammenfassen: 1)
1. I m Laufe einer einzigeu Generation ist die eeitliche Reihenfolge, in der die Komponenten des Widerstandes der Uxnwelt wirken, rein rech- nerisch betrachtet, fur das Endresultat belanglos. Entscheidend ist ledig- iich ihre prozentuale Hohe.
2. Im Laufe mehrerer Generationen betrachtet, lassen sich unter den Widerstandskomponenten +nab h an g ig v e rande r l i c h e'' und ,,a b h an g i g verand er l iche" unterscheiden.2) Erstere besitzen auf die Dauer groBeren
100
Auch Bodenhein ier (13) hat neuerdings auf ihn verwiesen.
I) Wegen Einzelheiten mull auf die ausfuhrliche Arbeit [Z w olf er (77)] verwiesen werden.
Ahnlich unterscheidet btrreits Thom pson (66, 67), mit dessen diesbeziiglichen Arbeiten ich im Laufe der Korrektur bekannt wurde, wie auch M a r t i n i (44) ,,all- gemeine und u n a b h a n g i g e F a k t o r e n " von , , individuel len u n d abhangigen Faktoren" - beides Begriffe, die sich im groBen ganzen mit den obigen der +nab- hhingig veranderlichen" und ,,abhangig veranderlichen" Widerstandstomponenten decken. Ersterer Autor versucht im ubrigen an Hand mathematischer Ableitungen die Unterschiede der WirkungsgroBe , ,schein b a r e r " und , ,wirkl icher" Sterblichkeit (,,apparent mor- tality" und ,,real mortality") bei allgemeinen und individuellen Faktoren zu erortern - eine Frage, dic im obigen Zusammenhang nicht beriihrt ist.

490 Zwol fe r :
EinfluS auf Anderungen der Populationsdichte als letztere. In diesem Sinn kann von Widerstandskomponenten mit p r imare r und s e k u n d a r e r epidemiologischer Bedeutung gesprochen werden.
Zu den Einzelwiderstanden mit seknndarer epidemiologischer Be- deutung gehoren solche, wie sie durch den EinfluB von Parasiten, Feinden, Krankheiten usw. bedingt werden. Diese sind in der Starke ihres Auftretens neben anderem wesentlich von der jeweiligen Populationsdichte des Wirts- tieres abhangig.
Primare epidemiologische Bedeutung besitzen demgegenuber in der u n b e r u h r t e n Natur Kl ima und phys iographische A n d e r u n g e n geologischer Ar t , soweit sie direkt oder indirekt an der Dezimierung der Population eines Insektes beteiligt sind. I m Gegensatz zu den sekun- daren Widerstandskomponenten ist ihre WirkungsgroBe von der Populations- dichte des Schadlings ganzlich unabhangig. U n t e r d e n Verha l tn i s sen des Ku l tu r l andes kommen e i n e Reihe we i t e re r , d u r c h d i e Tatig- k e i t d e s Menschen b e d i n g t e r Wide r s t andskomponen ten mi t p r i - m a r e r ep idemiologischer B e d e u t u n g hinzu: wie jene der Ernahrung (Fruchtwechsel in der Landwirtschaft und waldbauliche Eingriffe in der Borstwirtschaft) der Beeinflussung des Lebensraumes durch KulturmaB- nahmen (Trockenlegung von Sumpfen, hinderungen des Grundwasserhori- zontes, Bodenbearbeitung usw.), der Bekampfung mit technischen oder biologischen Hilfsmitteln. Hier sind sie im einzelnen oder in ihrer Ge- samtheit fur die Schwankungen der Populationsdichte einer Insektenart von Generation zu Generaticn verantwortlich. In der unberuhrten Natur ist dies fast ausschlieBlich das Klima.
Was den praktischen An wendungsbereich der Populationsgleichung angeht, so vereinfacht sie das in entsprechenden Ballen bisher gebrauch- liche Rechenverfahren. Im ubrigen sind bei ihrer Anwendung folgende Uberlegungen zu beriicksichtigen:
Die Ableitung der Populationsgleichung geht von dem Gedanken aus, daS die einzelnen Widerstandskomponenten nacheinander folgend auf den jeweils verbleibenden Rest der uberlebenden Individuen einwirken. Tritt der Pall ein, daB 2 oder mehr Widerstandskomponenten g 1 e i c h z e i t i g n e b e n e i n a n d e r wirken, dann muB ihre Einzelwirkung den Voraus- setzungen der Formelableitung entsprechend summiert, und die Summe als e inhe i t l i ch e Komponente behandelt merden. Hat beispielsweise im Eistadium ein Widerstand w1 % der Eier vernichtet, im Junglarvenstadium ein zweiter Widerstand wp % der Uberlebenden getotet, und ist festgestellt worden, daB im vorgeschrittenen Larvenstadium w a x der noch vorhandenen Individuen beispielsweise durch Mykosen, und gleichzeitig wb % der Tor- handenen Individuen durch Parasiten vernichtet wurden, dann wurde in diesem Falle wa den Wert annehmen: wQ = (w, + wh) und entsprechend in die Populationsgleichung einzusetzen sein. - Dasselbe Ziel wird er-

Studien zur okologie und Epiderniologie der Iosekten. 491
reicht durch die (theoretisch culassige) Annahme, da13 die beiden Wider- stande kurz nache inande r wirkten. I n diesem Ball darf allerdings der Wert des zweiten Widerstandes nicht auf die Gesamtzahl vorhandener Individuen bezogen werden, sondern nur auf den Rest der Uberlebenden nach Abzug des durch den Widerstand wa vernichteten Anteiles.
Ein weiterer Punkt bedarf der Erorterung: Angenommen, die Aus- gangspopulationsdichte sei im Berbst - etwa als Puppenzahl je Quadrat- meter Waldstreuflache - festgestellt worden. Die Eiablage der Art hin- gegen beginnt im Priihjahr. Wahrend des Winters seien von den Puppen w1 % klimatischen Einflusseo erlegen, und w2 % des Restes Mykosen zum Opfer gefallen. Konnen die Werte dieser Widerstande, die vo r der Ei- ablage wirkten, ebenso behandelt werden, wie Widerstande, die nach er- folgter Eiablage einsetzen?
Hat die Populationsdichte im Herbst PI Individuen betragen, dann
Der pi -w1 ' ist durch den Widerstand w, der Anteil ~- rernichtet worden.
verbleibende Rest betragt P
den zweiten Widerstitnd w2 % der Individuen vernichtet worden. Dieser
100
-xvl-pi' ( zo). Hiervon sind durch 1-m-
Anteil betragt demnach P, . 1 - 1 .L. Der verbleibende Rest der ( G o ) G o Uberlebenden ist dann P,
(1 - %). Zu Beginn der Eiablage hat mithin die Populationsdichte
diesen letzteren Wert Pi . 1 - - 1 - - und ist dementsprechend
in die Populationsgleichung einzusetzen. An deren Wesen wird dadurch nichts geandert. Es mu13 lediglich beriicksichtigt werden, da8 nunmehr nach Ablage der Eier die weiterhin wirkenden Widerstande als w3, w4, w5 usw. fortlaufend in die Gleichung einzustellen sind.
Bus dem Gesagten geht hervor, daS die Populationsgleichung zeit- 1 i c h u n b egr enz t anwendbar ist. J e d e s E n t w i c k lun gss t a d iu m, g le ichvie l o h E i , La rve , P u p p e ode r Imago kann i n i h r a l s Be- g i n n bezw. E n d e e i n e r Genera t ion angenommeli werden. Nit anderen Worten: Die auf eine Generation bezogene Populationsgleichung ist anwendbar, gleicbgiiltig, ob die Generation vom Eistadium bis zum nachstfolgenden Eistadium, von Larve bis zur nachstfolgenden Larve, von Puppe bis zur nbhstfolgenden Puppe, oder der Imago bis zur nachst- folgenden Imago gerechnet wird.
Auch die Populationsdichte der Art zu irgend einem beliebigen Zeit- punkt der Generation - also beispielsweise fur irgend ein Entwicklungs- stadium, das praktisch besonders bedeutungsvoll ist - kann nach ihr er-
( 100 w l j . ( z0)
Zeitschrift tiir angewandta Entomologie. XVII, 3. 33

492 Zwol fe r :
mittelt werden, wenn Ausgangspopulationsdichte, absolute Eizahl, Ge- schlechterverhaltnis und diejenigen Widerstiinde bekannt sind, die bis zu dem betreffenden Zeitpunkt gewirkt haben. Letzteres ist fur die Prognose- stellung von Wichtigkeit.
Bei quantitativ analytischen Untersuchungen von Massenwechsel- erscheinungen gibt die Populationsgleichung die Moglichkeit, sich rasch
. uber die Vollstandigkeit und Kichtigkeit der empirisch gefundenen Daten zu vergewissern. 1) Sind namlich bei einer solchen Untersuchnng Aus- gangs- und Endpopulationsdichte der Generation durch Beobachtung fest- gestellt worden, und samtliche wesentlichen Einzelwiderstande ihrer WirkungsgroBe nach empirisch ermittelt, dann miissen die gefundenen Werte, in die Populationsgleichung eingesetzt, diese befriedigen. 1st dies nicht der Pall, und weicht der berechnete Wert der Endpopulationsdichte P, erheblich vom beobachteten Wert ab, so zeigt dies, daB ein oder mehrere wichtige Umweltseinflusse, die auf die betreffende Generation wirkten, der Beobachtung entgangen sind.
Endlich gestattet die Populationsgleichung, wenn bei einer quantitativ analytiscben Untersiichung alle wesentlichen Einzelwiderstande bis auf einen empirisch ermittelt werden konnten, die WirkungsgroBe dieses einen unbekannt gebliebenen in einfacher Weise zu bestimmen. Es ist hierzu lediglich erforderlich, in der Gleichung die gesuchte Widerstandskompo- nente als ,,Unbekannte" (wx) zu behandeln, uad die Gleichung entsprechend nach w, aufzulosen. 2)
So betrachtet, ist die Populationsgleichung ein einfaches Hilfsmittel fur quantitativ analytische Massenwechseluntersuchungen. Bei den empi- risch ermittelten Daten konnen naturlich nur Durchschnittswerte beruck- sichtigt werden. Dementsprechend stellen auch die mit Hilfe der Gleichung gefundenen Ergebnisse nur Mittelwerte vor, die allerdings den wirklichen Verhaltnissen um so naher kommen werden: je umfangreicher das Material ist, auE dem die empirischen Daten fuBen.
Die Anwendung der Pormel fur Prognosezwecke im besonderen setzt voraus, daB die WirkungsgroBe der wesentlichen Einzelwiderstande bis zu dem Zeitpunkt zuverlassig vorausgeschatzt werden kann, fur den die Prognose gestellt werden SOU. Wie an anderer Stelle 3, irn einzelnen naher ausgefuhrt worden ist, kann dieser Zeitpunkt theoretisch sich selbst uber mehrere Generationen erstrecken. 1st eine derartige Schatzung der Einzelwiderstande moglich - und die neuere epidemiologische Unter- suchungsniethodik gibt hierfur eine Handhabe - dann ist kunftig auch eine zuverlassige Prognosestellung durchfuhrbar.
Die folgenden Beispiele zeigen einige der Anwendungsmoglichkeiten der Populationsgleichung bei quantitativen Massenwechseluntersuchungen.
l) Vgl. hierzu Beispiel 1 S. 493 dieser Arbeit. z, Vgl. hierzu Beispiel 3 S. 497 dieser Arbeit. a) Z w o l f e r (77) S. 752ff.

Studien zur dkologie und Epidemiologie der Insekten. 493
Beispiel 1. P r u f u n g e i n e r Ana lyse : An Hand von M e y e r s Beobachtungen 1) lafit sich eine Vorstellung von der WirkungsgroBe der- jenigen Widerstandskomponenten gewinnen, die wiihrend der Eulen- generation 1930 im mittelfrankischen PraSgebiet lokal in Erscheinung Re- treten sind. Die Daten beziehen sich auf einen naher untersuchten Stangenholzbestand des Porstamts Heideck, der schon wahrend der Gene- ration 1929 stark befressen worden war und 1930 n ich t der Arsen- bekampfung unterlag.
Irn Narz 1930 murden hier durchschnittlich 174 Puppen auf 5 qm gefunden. Die Ausgangspopulationsdichte war mithin 34,8 Puppen je Quadratmeter; demnach PI = 34,s. Insgesamt gingen rund 50% dieser Puppen an Parasitierung (Schlupfwespen) iind Verjauchung (Bakteriosen? Mykosen) zugrunde, was einer Widerstandskomponente wl = 50 entspricht.
Die in der Folgezeit von einem Weibchen dieser Eulengeneration durchschnittlich abgelegte Eizahl betrug 130. Nach fruheren Erorterungen ist dies bei der Kieferneule gleichbedeutend mit einer Reduktion der absoluten Eizahl um 31%, mithin w,=31. Im Hinblick auf die weiter unten mitgeteilten experimentellen Untersuchungsergebnisse diirfte diese 31 prozent. Reduktion der absoluten Eizahl in erster Linie der 10 tagigen Regenperiode zuzuschreiben sein, die in Mittelfranken im Mai wahrend der Hauptflugzeit der Falter eintrat.
In diesem Zusammenhang sei daran erinnert, da13 die angegebene Reduktion der hochstmoglichen Eiproduktion von 31 o/o fu r die Verminderung der Nachkommenschaft der betreffcnden Generation rein rechnerisch dieselbe Bedeutung hat wie etwa eine 31 prozent. Eiparasitierung nach erfolgter Eiablage. Es folgt dies aus Absatz 1 der oben mitgeteilten theoretischen SchluBfolgerungen der Populationsgleichung.
2,6% der in diesem Bestand abgelegten Eier wurden parasitiert (l’richogramma eaanescens Westw.). 7,6 % gingen hier wahrend der Ent- wicklung xahrscheinlich infolge abiotischer Einwirkungen zugrunde. Weitere 5,6 % der standig kontrollierten Eier verschwanden spurlos. Ver- mutlich sind die letzteren der rauberischen Tatigkeit von Vogeln und von Peinden aus dem Insektenreich zum Opfer gefallen.2) I m ganzen ent- spricht dies einer Widerstandskomponente w3 = 2,6 + 7,4 + 5,6 = 15,6.
I m Junglarvenstadium wurden naoh Meyers Ermittlungeu im Stangen- hok rund 20 % des vorhandenen Raupenbestandes vernichtet, wohl groSten- teils infolge abiotischer Einwirkungen. Demnach w4 c= 20.
l) Die im folgenden verwendeten Daten sind teils der Meyerschen (46) Arbeit 9nt- nommen, teils verdanlie ich sie seinen mir freundlichst zur Verfiigung gestellten dies- bezuglichen Notizen.
z, Eier, Larven und Adultis der Coccinellide Anatis ooellata L. fanden sich wahrend der Eiablageperiode der Kieferneule in diesem Gebiet allenthalben in auffallend groBer Anzahl. Ob die Art auch Eier und Junglarven der Kieferneule annimmt, steht nicht einwandfrei fest. Nach Erfahrungen bei verwandten Gattungen ist zum mindesten der Verdacht einer derartigen Xebennahrung berechtigt [ E s c h e r i c h (23), S c h i l d e r (58), Z w o l f e r (75)]. Nach R a t z e b u r g (51) sol1 die Art Schmetterlingspuppen verzehren.
33 *

494 Zwol fe r :
I m Altlarvenstadium wurde auf Grund BuBerer Merkmale der Raupen eine Tachinierung von rund 55 % festgestellt. Die durch Krankheiten (Balcferiosetz?, Enapitsa und vielleicht auch direkte klimatische Einflusse) verursachte Verniinderung des Altraupenbestandes IaSt sich nur schatzungs- weise angeben. Ein Vergleich der auf einer Kotfangflache in dieser Ab- teilung sich ansamrnelnden Raupenkadaver mit den bei Probezahlungeu urspriinglich in der Krone festgestellten Raupenzahlen fuhrt auf ca. 36 durch Krankheiten vernichteter Altraupen. Durch Tachinierung und Krank- heiteu zusammen waren demnach 55 % + 35% = 90 4; des vorhandenen Altraupenbestandes ausgemerzt worden. Mithin ergibt sich ein weiterer Widerstandswert w5 = 90.
Wahrend der gesamten FrsBzeit der Altraupen (111.-V. Stadium) wurden auf 1 qm Kotfangflache in diesem Bestand 450 Raupenkadaver abpolesen. Bei einer Schirmflache von rund 2 qm entspricht dies einem Abgang je Stamm von ca. 900 Raupen. Unter Zugrundelegung eines urspriinglichen Raupen besatzes von rund 1000 Stuck je Stamm - wie er sich bei voraufgehenden Probezahlungen in diesen Lagen ergab - entspricht dieser Abgang einer Altraupenverminderung von Hierin sind die durch Tachinose und anderweitige Krankheiten zugrundegegangenen Raupen einbegriffen, und zwar in einem Verhaltnis, das annahernd den oben mitgeteilten Prozentsatzen entspricht. -
Noch schwieriger gestaltet sich die Schatzung der in diesem kahl- gefressenen Bestand durch Hunger zugrunde gegangenen Raupenmengen. Vorausgesetzt, daB sie derjenigen Zahl gleichkommt, die aus dem Bestand in benachbarte ebenfalls kahlgefressene Schlage abzuwandern versuchte, und hierbei in die allseitig aogelegten Fanggraben geriet, ware dieser An- teil schatzungsweise mit 98 % des Restes der Altraupen zu verauscblagen. Dies wurde einer durch mangelhafte Ernahrung hedingten Widerstands- komponente w6 = 98 entsprechen.
Letzteres argibt sich aus folgender Uberlegung: Der in Fragcr kommende Bestand von rund 1/4 ha GroBe war allseitig von einem Fanggraben umgeben, dessen Gesamtlange rund 200 m betrug. Im Durchschnitt fingen sich je laufender Meter nach M e y e r s Schatzung 250 Raupen. Demnach waren insgesamt rund 50000 Raupen durch den Graben abgefangen worden. Wird noch berucksichtigt, daIj aus dem abgegrenzten Gebiet aller Voraussicht nach ebenso viele Raupen auszuwandern, wie umgekehrt aus den benach- barten Bestanden in dieses einzuwandern rersuchteu, so folgt. daB rund die Ealfte der genannten Zahl wandernder Raupen dem abgegrenzten Gebiet entstammt haben durfte. Das waron demnach 25000 Stuck. Bei einem Baumbestand von 250-260 Stuck auf dieser Flache (nach Schatzung von forstlicher Seite) wurden somit pro Stamm im Durchschnitt 96-100 Raupen abgewandert sein. Da nach Abzug der auf den Kotfangflachen ge- sammelten Kadaver mit einem restlichen Altraupenbesatz von ca. 100 je Krone gerechnet werden' muB, hatte der abgewanderte und verhungerte Raupenteil ungefahr den oben ge- nannten Prozentsatz des Restbestandes ausgemacht. -
I m vorstehenden ist nur die Wirkung der hauptsachlichsten Einzel- widerstande berucksichtigt, denen gegenuber Umweltseinflusse von unter- geordnetem Wirkungsgrad kaum ins Gewicht fallen, wie beispielsweise jene, die durch die Tatigkeit der Puppenrauber, Orabwespen, Vogel usw. bedingt waren. Da die Wirkung einiger wichtiger Schlupfwespenparasiten sich bei der Kieferneule erst wahrend des Puppenstadiums auBerr, welches

Studien zur Okologie und Epidemiologie der Insekten. 49.5
in die gegebene Analyse nicht mehr einbezogen werden konnte, muRten diese in der obigen Darstellung unberucksichtigt bleiben. -
Wenn diese Uberlegungen zutreffen, und die wesentlichsten Wider- standskomponenten richtig erfaflt bezw. richtig geschatzt worden sintl, miissen nach fruheren Erorterungen die gefundenen Werte in die Popu- lationsgleichung eingesetzt, eine Endpopulationsdichte ergeben, die rnit der zu Ende der aktiven Lebensperiode dieser Generation an Ort und Stelle ermittelten iibereinstimmt. Uber letztere teilt Meyer mit, daB im fraglichen Bestand Anfang Juli auf 10 q m S verpuppungsreife Raupen und 6Puppen gefunden wurden. Mithin ist dort die beobachte te End- popiilationsdicbte 1,4 Individuen pro Quadratmeter gewesen.
Zur r e c h n e r i s c h e n Ermittlung der Endpopulationsdichte P, mit Hilfe der Populationsgleichung stehen nun folgende Werte zur Verfiigung:
Ferner sind zu beriicksichtigen die ideale Eizahl der Kieferneule mi t e = 190, und das Geschlechterverhaltnis rnit m : f = 1 : 1, somit m + f = 2, f = 1. Hieraus folgt fur P,:
P 1- -34,8; ~ , = 5 0 ; w2=31; w3=15,6; w4=20; ~ , = 9 0 ; ~ , = 9 8 .
34,8.160.1( 50)( 2 100 100 100 100 100 100
31)( I--' 15B)i I--- 20)( - I--- 90)( I--- 98) P, =
= 17,4. 190.0,Fj. 0,69.0,844.0,8.0,1.0,02 = 1,540. Die gute Ubereinstimmung des e r r echne ten Wertes fur die End-
populationsdichte von rund 1,54 Individuen je Quadratmeter mit dem be- o bach te t en von 1,4 Individuen je Quadratmeter, bestatigt bis zu einem gewissen Grad die Richtigkeit der obigen Analyse und zeigt gleichzeitig, daB die WirkungsgroBe der Hauptwiderstandskomponenten gut erfaBt bezw. zuverlassig geschatzt worden ist.
In der gegebenen Darstellung blieb die Wirkung solcher Umweltseinfliisse unberuck- sichtigt, die nur einen geringen Prozentsatz des jeweils vorhandenen Individuenanteiles vernichten. Tatsachlich uben derartige Widerstandskomponenten keinen erheblichen Ein- fluP auf die Hohe der Endpopulatiousdichte aus: Wird beispielsweise in der obigen Ana- lyse angenommen, daP zu irgend einem Zeitpunkt der Tatigkeit der Puppenrauber 1% des vorhandenen Raupenbestandes zum Opfer fiel, so wurde sich der errechnete Wert der Endpopulationsdichte auf 1,53 Individuen je Quadratmeter vermindern, und selbst eine Widerstandskomponente von 5 O/o hatte nur eine Reduktion des Endergebnisses auf 1,46 zur Folge gebabt. Das sind Anderungen. die innerhalb der FehlergroBen liegen, mit welchen bei Bestimmung der Populationsdichte durch Feststellung des Puppenbelages je Quadratmeter Waldstreu ebenfalls gerechnet werden muP. Es erhellt hieraus, daP Widerstandskomponenten der angedeuteten GroBenordnung, wenn sie in geringer Anzahl in Erscheinung treten, unberiicksichtigt bleiben konnen.
Noch ein weiterer Punkt bedarf der Erlauterung: Bezuglich der tachinierten und kranken Altraupen wurde oben fur e r m r e eiu Prozentsatz von 55, fur letztere ein solcher von 35 angegeben. Genau genommen ist diese Trennung nicht scharf durchfuhrbar, da auch von den tachinierten Raupen eine groBe Anzahl von den Infektionskrankheiten er- griffen wurde, und mitsamt der innewohnenden Tachinenlarven zugrunde ging. Die Sektion der Raupenkadaver bat dies wiederholt ergeben. Es folgt auch aus dem Umstand. daB sich zu Ende der Generation nur 1,2 Tachinentonnchen je Quadratmeter am frag- lichen Beobachtungsort fanden, wahrend nach der absoluten Zahl tachinierter Raupen zu

196 Z w o l f e r ;
urteilen (wie sie auf den Kotfangflachen ermittelt wurde) 100-200 Taohinen -Tonnchen je Quadratmeter erwartet werden multen. Mit aus diesem Grunde wurden in der obigen Darstellung die beiden Einzelwiderstande zu einem einzigen Widerstaodswert w6 = 90 zusammengefalt.
Beispiel 2. Prognoses t e l lung : Im einzelnen Fall lassen sich mit Hilfe der Populationsgleichung rasch ungefahre Anhaltspunkte uber die Hohe der fur einen Bestand jeweils kritischen Puppenzahl finden. Nach den Erfahrungen der letzten Kalamitat in Mittelfranken, hat in etwa dreifiigjahrigem Stangenholz ein Belag von rund 500 Altraupen je Stamm KahlfraB zur Polge. Wird die absolute Eizahl der Eule mit 190 angesetzt, ihr Geschlechterverhaltnis rnit 1 : 1, und rechnet man ferner mit gunstigen Witterungsverhaltnissen fur die Kieferneule, und einer Eireduktion von nur 20 % und einer Junglarvensterblichkeit im Stangenholz von ebenfalls 20%, dann laBt sich auf Grund der Populationsgleichung der Ansatz aufstsllen :
Pi. 1 9 0 . 1 Y 100 100 500= F,
worin nach fruheren Erorterungen Pi die Ausgangspopulationsdichte, also im vorliegenden Fall die Zahl gesunder Puppen j e S tamm zu Beginn der Generation vorstellt. (Auf 1 qm reduziert gibt sie die gesuchte kritische Puppenzahl an.)
2 . 5 0 0 100 P -
Bus obiger Gleichung folgt fur P,: - ~ = ca. 8,4. 1- 190.0,8.0,8=- 12
Bei 100 prozent. KronenschluB und rund 2 qm Schirmflache wurde dies einern durchschnittlichen Puppenbelag von rund 4 gesunden Puppen je Quadratmeter gleichkommen, der in diesem Ball als kritische Puppenzahl anzuseheo wke. Dabei mu13 berucksichtigt werden, daB die Wirkung der Wetterlage in der obigen Gleichung geschatzt worden ist. Pallt die Witterung fur die Eule etwas ungunstiger aus als angenommen, dann wurde sich sine entsprechend hohere kritische Puppenzahl ergeben. Be rne r f u h r t e i n KronenschluB d e r k l e ine r ist a l s d e r im Be i sp ie l an- genommene von 1 0 0 % auf k l e i n e r e k r i t i s c h e Pnppenzahlen .
Fur das auf die Eruption folgende Jahr einer Kalamitat im besonderen muB die Eireduktion erheblich hoher angesetzt werden,1) da Ubervolkerung und Ernahrungsverhaltnisse wahrend des voraufgehenden Eruptionsjahres sich in diesem Sinn ungunstig auswirken. Doch fehlen einstweilen noch genauere Unterlagen uber den Umfang, welchen die Eireduktion in diesem Sonderfall erreicht.
In entsprechender Weise laBt sich auch fur Altholzer die kritische Puppenzahl naherungsweise bestimmen, wobei in Rechnung gestellt werden muB, daB bei einem ca. 80jahrigen Bestand erst ein Altraupenbesatz von 4-5000 Stuck je Krone zum KahlfraB fiihrt und die Junglarvensterblich-
- ') Vgl. hierzu Volkels (69) vorlaufige Mitteilung und Sachtleben (55).

Studien zur Okologie und Epiderniologie der Inseliten. 49 7
lichkeit nach Meyer (46) im Altholz - selbst bei einer fur die Eule gunstigen Wetterlage - erheblich hoher ist (ca. 40-50 %).
Beispiel 3. Mindes twirkungsgrad e i n e r BekampfungsmaB- nahme: Die Frage, wie groB der Mindes twirkungsgrad einer tech- nischen oder biologischen BekampfungsmaBnahme sein mu13, wenn der erwunschte Erfolg wahrend einer oder mehrerer Generationen eines Schad- lings anhalten soll, 1aBt sich nur von Fall zu Fall entscheiden. Auch hierfur gibt die Populationsgleichung eine Handhabe, ihn rasch naherungs- weise zu ermitteln. Man muP sich dabei lediglich vor Augen halten, daB jener Indiriduenanteil, der durch die geplante BekampfungsmaBnahme ver- nichtet werden soll, in der Populationsgleichung als eine Komponente des Widerstandes der Umwelt in Erscheinung tritt. Bexeichnet man den Wirkungsgrad derselben mit w,, so 1aBt sich aus der genannten Gleichung fur seine Bestimmung der Ausdruck ableiten:
100. (m + f ) . P2
f . e . ~ , ( 1 - 100 -W&-)(I- 100 wz)(~-:g). . . . . ( I -%) w, = 100 -
I n Gleichung (6 ) stellt Pa jene Endpopulationsdichte vor, bis zu welcher der aus P, Ausgangsindividuen normalerweise sich entwickelnde Schadlingsbestand durch kiinstliche Eingriffe herabgedruckt werden mull, um jeden Schaden zu verhuten. wlr wp, w, . . . . . w, sind naturliche Widerstandskomponenten, die bis zum Zeitpunkt der schadigenden Ent- wicklungsstufe der betreffenden A r t in Wirksamkeit treten, fur sich allein aber nicht ausreichen wurden, den erforderlichen Effekt zu erzielen. m : f ist das Geschlechterverhaltnis. e die absolute Eizahl der Art. -
Angenornmen in einem Kiefernstangenholzbestand habe ein Probe- zahlen nach Eulepuppen einen Belag von 8 gesunden Puppen je Quadrat- meter ergeben. Die Zahl liegt uber dam kritischen Wert. Eine BekampfungsmaBnahme wird geplant. Wie groB muB mindestens deren Wirkung, sein um einen Schaden in der betreffenden Generation zu verhuten? Nach fruheren Erorterungen fiihrt bei der Kieferneule im Stangenholz eine Altraupenzahl von rund COO je Stamm zum KahlfraB. Etwa 200 Altraupen je Stamm wurden hier ungefahr den Grenzwert fur die schadliche Altraupenzahl vorstellen. Durch die geplante Bekampfungs- maBnahme.mul3te also zu i r g e n d e i n e m vor dem s c h a d l i c h e n Alt- r a up e n s tad i u m 1 i e g end en Z e i t p u n k t eine Verminderung der Popu- lationsdichte angestrebt werden, die zur Folge hzt, daB der Altraupen- besatz je Stamm die Zahl 200 nicht ubersteigt.
1st der Standbereich eines Baumes im Durchschnitt 4 qm, so ergibt . letztere auf dieselbe Lebensraumeinheit von 1 qm wie oben umgerechnet
eine anges t r eb te Endpopulationsdichte Pz = 4= 50. Wird noch be-
rucksichtigt, da13 selbst bei einer fur die Eule relativ gunstigen Wetterlage
Dann ware Pl=8.
200

498 Zwolfer :
die Reduktion der absoluten Eizahl rund 200/,, und die Verminderung des Junglarvenstadiums durch abiotische Eiuflusse ebenfalls rund 20% betragt, so entspricht dies 2 Widerstandwerten w1 = 20 und w2 = 20. Der Aus- gangspuppenbelag betiagt 8 gesunde Puppen je Quadratmeter, mithin PI = 8. Fur die Bestimmung von w, kann damit der Ansatz aufgestellt werden [nach Gleicbung (6)l:
100. (1 + 1). 50 w, = 100 - - -
1 . 1 9 0 . 8 . ( 1 - ;;o)(l- $) 10 000
190.8 . 0,s .0,8 = 100-
= 100 - 10,278 = S9,7 22 Es ergibt sich somit als Mindes twirkungsgrad , de r von d e r
gep lan ten Bekampfungsakt ion ge fo rde r t werden mull, e in e Ver- m i n d e r u n g des Schad l ingsbes t andes urn rund goo/, zu i r g e n d e inem vor dem s c h a d l i c h e n Al t l a rvens t ad ium l i egenden Zeit- punkt . Gleichgi i l t ig i s t dabe i , m i t welchen Mit te ln t echn i sche r oder b io logischer A r t de r Kampf ge fuhr t wird und ob s i ch der- s e lbe gegen d i e Puppe , den Pa l t e r , das E i s t ad ium ode r gegen d ie J u n g r a u p e n r ichtet . Stets mu13 unter den angegebenen Voraus- setzungen - Ausgangsbelag 8 gesunde Puppen je Quadratmeter und Stangenholz mit rund 4 qm Bodenflache je Stamm! - dieser Wirkungsgrad einer Bekampfung als MindestmaS gefordert werden, um Schaden zu verhiiten.
In ahnlicher Weise la& sich mit Hilfe der auf mehrere Generationen erweiterten Populationsgleichung 1) der Mindest wirkungsgrad ermitteln, den eine KampfmaBnahme haben mull, wenn ihre Wirkung sich uber 2, 3 oder noch mehr Generationen erstrecken soll. Hierfur ergibt sich durch einfache algebraische Ableitung aus der erweiterten Populationsgleichung zur Bestimmung des Mindestwirkungsgrades w, der Ausdruck:
Hierin stellt q die Anzahl der Generationen Tor, uber die hinaus die Bekampfungsmaflnahme wirken SOU, P, ist der Grenzwert der Popu- lationsdichte des schadlichen Stadiums einer Insektenart, der nach q Gene- rationen nicht uberschritten sein darf , wenn Schaden vermieden werden soll, Pi ist die Busgangspopnlationsdichte derjenigen Generation, in welche die BekampfungsmaIjnahme eingreift; m : f und e sind Geschlechterverhaltnis und ideale Eizahl: W, W', W", . . . W"' 1 * * q-1 sind die Werte der Gesamt- w i d e r s t l n d e , die anch ohne den geplanten Kampf auf den betreffenden
') Vgl. Zwolfer (77) S. 752ff.

Studien zur i)liologie und Epidemiologie der Insekten. 499
Scbadling in den q aufeinanderfolgenden Generationen einwirken wiirden. Ihre Kenntnis ist wie aus der Gleichung hervorgeht, Voraussetzung fur die Bestininlung des ~~indestwirkungsgrades w,. Fur den praktischen Gebrauch wird es im allgemeinen geniigen, den geringsten Wert des Gesamt- widerstandes zu kennen, der an einem gegebenen Biotop unter Bedingungen eintretsn kann, die fur den Scbadling giinstig sind und diesen in Itechnung zu stellen. Seine Ermittlung ist nur auf empirischem Wege moglich, und setzt mehrjahrige Beobachtungen der Bevolkerungsbewegungen der betreffenden Schadlingsart an einem gegebenen Biotop voraus. 1)
Nach obiger Gleichung (7) errechnet, gibt w, denjenigen Prozentsatz von Individuen einer Population an, der in der Ausgangsgeneration zu i rgend einem v o r d e r schadl icb en Entwicklungsstufe der betreffenden Art liegmden Stadium durch kiinstliche Eingriffe technischer oder bio- logischer Art vernichtet werden muB, wenn die Wirkung der MaQnahme q Generationen hindurch anhalten soll. -
Im vorstehenden wurde versucht zu zeigen, wie die verschiedenen Einfliisse der Urn welt mit dem Schicksal der Generationen zahlenmaBig verkniipft sind. In theoretischer Hinsicht ergab die Diskussion eine Be- statigung dessen, was von anderer Seite auf induktivem Wege bereits erkannt war. Neben einigen anderen Umweltseinfliissen den des Elimas als einer primaren Widerstandskomponente im epidemiologischen 9’ inn. In welchem Busman sich dieser geltend macht, ob er durch Reduktion der absoluten Zeugungskraft oder direkte Vernichtung eines Teiles der gezeugten Nachkommenschaft , ob er durch Anderung der Ernahrungs- verhaltnisse oder anderweitiger Widerstandskomponenten, wie sie beispiels- weise durch Biophagen und Krankheiten reprasentiert werden, am starksten in Erscheinung tritt, dariiber vermag nur Beobachtung und Experiment zu entscheiden.
Die folgenden Ausfiihrungen geben die Ergebnisse in diesem Sinne durchgefiihrter experimenteller Untersuchungen wieder. Sie beschiiftigen sich zunachst mit dem EinfluB der beiden okologisch wichtigsten Klima- faktoren 2) - Temperatur und Luftfeuchtigkeit - auf Eiproduktion und einige damit zusammenhangende Erscheinungen , des weiteren mit jenem auf die Vitalitat (Lebensfahigkeit) der Jugendstadien der Kiefernenle. Im Zusammenhang mi t diesen experimentell okologischen und epidemio- logischen Arbeiten wurde gleichzeitig versuch t, das Problem der Ent- wicklungsdauerabhangigkeit der Kieferneule zu fordern.
I) Vgl. hierzu S. 481-485 dieser Arbeit. z, Ahnlichauiulern sich Bodenheimer (8-13), Shelford (Sl-64), Cvarov(68)u.a.

500 Zwolfer :
IT. Die Zeugungsflhigkeit von PufioMs flamrneu Schiff. in ihrer AbhHngigkeit von Temperatur und Luf't'tfeuchtigkeit.
a) Speeielles zur Metliode.
Zur Feststellung der Temperaturabhangigkeit des Entwicklungszustandes der Ovarien suhliipfender Falter wurden jeweils 50 Puppen gleicher tIerkunft (Material: Forstamt Heroldsbaoh, Mittelfranken) und gleicher Vorbehandluug in flachen Petrischalen von 18 cm Durchmesser rnit angefeuchtetern Moos zusamniengebracht und der 4O, So, 14O, 18O und 22' C Temperaturstufe ausgesetzt. Soweit die weiblichen Falter nach dem Schliipfen nicht anderweitig Verwendung fanden, wurden sie in verschiedenen Zeit- abstanden seziert und unter dem Binokular auf den Entwicklungszustand ihrer Ovarien und ihres Fettkorpers hin untersucht. Beziiglich der Unterscheidung von legereifen Eiern und Eianlagen in den Ovarien, die keinerlei Schwierigkeiten bereitet, sei auf Eidnians (21) diesbezugliche Untersuchung verwiesen. Die bei der vorliegenden Arbeit in dieser Richtung Eemachten Erfahrungen decken sich rnit seinen Angaben, so daB auf ihre be- soudere Yitteilung verziahtet werden kann.
Fur die Versuche iiber den Einflul3 von Temperatur und Luftfeuchtigkeit auf geschlechtliche AktivitAt und Eiablage fanden Akkumulatorenglaser von 6 x 15 x 18 cm InneumaB Verwendung, die rnit Glasdeckel abgeschlossen waren. In jedes GefaB wurden 1 oder 2 kleinere Kiefernzweige eingesetzt , deren Schnittflache zur langeren Frisch- haltung ParaffinabschluB erhielt. Die urspriiiigliche Absicht, die Luftfeuchtigteit in diesen GefaBen rnit angefeuchteten Salzen zu regulieren - analog wie bei den Versuchen uber Ei- und Junglarvenmortalitat - erwies sich als undurohfiihrbar. Die Hohe der GefaBe, die - um den Faltern die notige Bewegungsfreiheit zu geben - nicht niedriger gewahlt werden konnte, hat zur Folge, drl3 die feuchtigkeitsregulierende Wirkung der feuchten Salze in den oberen Luftschichten sich nicht mehr ansreichend bemerkbar macht. Nur Luftschichten, die wenige Zentimeter uber der Salzschicht liegen, stellen sich bei un- bewegter Luft in relativ kurzer Zeit auf die entsprechende Luftfeuchtigkeit ein, wie mehrere diesbeziigliche Vorversuche zeigten. Naturlich spielt hierbei auch die Ver- dunstung des eingestellten Kiefernzweiges eine Rolle. A11 diese MiBstAnde sind im Escherichschen Thermohygrostaten (%), der mit bewegter Luft arbeitet, beseitigt. Leider war dessen Konstruktion wahrend der Laufzeit der Versuche noch nicht soweit abgeschlossen, daB er hierfur hatte in Benutzung genommen werden konnen. Es muBte daher bei den entsprechenden Versuchen behelfsmailig mit zwei Feuchtigkeitsstufen gearbeitet werden, von denen die 1000/,- Stufe durch Einlegen mehrerer Blatter gut angefeuchteten Filtrierpapiers auf den Boden des ZuchtgefaBes und dnrch ofteres Ver- nabeln von H,O konstant gehalten wurde. Bei der ,,Trockenluftstufo" (80-90°,'o rel. Luftfeuchtigkeit) fie1 der Wasserzusatz fort. In beiden Versuchsreihen wurde die Luft- feuchtigkeit vou Zeit zu Zeit mit dern Lamprechtschen Hygrometer kontrolliert. Bei der 80-90D/o-Stufe war sie pewissen Schwankungen ausgesetzt (in den Extremen 70 o/io und 90°/o), lag aber hier im allgemeinen zwischen 80-900/,.
Was die Versuchstiere angeht, so wurden zur Vermeidung einer Kopula v o r Be- ginn der Versuche mannliche und weibliche Puppen nach den bekannten Merkmalen [vgl. S a o h t l e b e n (56)] getrennt und bei Zimrnertemperatur in Zuchtkafigen unter feuchtem Moos bis zum Schliipfen aufbewahrt. Die Versuche wurden jeweils am h'achmittag bezw. Abend des Schliipftages angesetzt und mehrere Mannchen und Weibchen (im Hochstfalle insgesamt 12 Veruuchstiere) in entsprechend vorbereitete VersuchsgefSil3e gesetzt. Urn den Versuchstieren eine direkte Aufnahme von Wasser zu ermoglichen, ist in den GefaBen der Trockenluftversuchsreihe je ein kleiner angefeuchteter Wattebausch eingehangt worden, an denen im wejteren Verlauf sowohl Mannchen wie Weibuhen des ofteren begierig saugend beobachtet wurden.

Studien zur Okologie und Epidemiologie der Insekten. 502
lo) Die absolute Eizahl der Kieferneule. Die Bestimmung der absoluten Nachkommenzahl der Kieferneule,
worunter nach friiheren Ausfiihrungen jene Durchschnittseizahl zu ver- stehen ist, die von einem Weibchen im Optimum aller maBgebenden Be- dingungen abgelegt wird, ist mit gewissen Schwierigkeiten verkniipft: Der Ernahrungszustand der Generation, der die Versuchstiere entstammen, und alle Momente, die ihn beeinflussen, die Einwirkung der AuBenwelt auf die Puppe, und endlich jene auf den Falter nach dem Schliipfen konnen - theoretisch betrachtet - hierfur bedeutungsvoll werden. Das Optimum all dieser Bedingungen ist fur die Kieferneule noch nicht bekannt. Fur einige derselben ergab es sich naherungsweise erst im Laufe der vor- liegenden Untersuchung. Einfache Zwingerversuche mit Auszahlung der je Waibchen abgelegten Eier hatten demnach unter AuBerachtlassung dieser Uberlegungen nur r e l a t ive Werte fur die Zeugungskraft gegeben. Demgegebenuber setzt die Bestirnmung der idealen Eizahl auf diesem d i r e k t e n Wege voraus, daW sowohl Vorbehandlung des Materials (Er- nahrungszustand, Einflusse auf die Puppe) als auch die Bedingungen des Zwingerversuches selbst dem jeweiligen Optimum entsprechen.
Bei E'eststellung der idealen Eizahl auf i n d i r e k t e m Wege 1aBt sich die letztgenannte Schwierigkeit vermeiden : Wird namlich der Ge- samtzahl Eier, die ein Weibchen wahrend seiner Lebensdauer ablegte, jene der ,,legereifen" hinzugezahlt, die nach dem natiirlichen Tode des Versuchstieres in seinen Ovarien zuriickbleibt, so stellt die Summe die Gesarntzahl legereifer Eier vor, die das Weibchen wahrend seines ganzen Lebens produzierte und - optimale Bedingungen wahreod der Lege- periode vorausgesetzt - auch hatte ablegen konnen. Wird eine hin- reichende Anzahl von Versuchstieren gewahlt, so lassen sich auf diesem Wege brauchbare Unterlagen zur Ermittlung der idealen Eizahl gewinnen. 1) So wurde in den Versuchen rorgegangen, deren Resultate in der folgenden Tabelle zusammengefa5t sind: (Siehe Tabelle auf S. 502.)
Das Optimum bezuglich der V o r b e h a n d 1 u n g des Materials (Ernahrung, Ein- flusse auf die Puppe), auf dessen Bedeutung oben ebenfalls hingewiesen worden ist, muPte bei diesen Untersuchungen zunachst unberiicksichtigt bleibeu. Was den Er- nahrungszustand der Generation anlangt, der die Versuchstiere entstammten, so fie1 deren FraPperiode in das Prodromrljahr einer Kalamiyat. Nahrung ist in diesem ganz allgemein i m OberfluB vorhanden. Bezuglich des Ernahrungszustandes durfte daher daa Versuchs- material dem Optimum entsprochen haben;
Was die auf die Puppe vor Versuchsbeginn wirkenden Einflusse angeht, so hat, * wie im folgenden Abschnitt gezeigt wird, von den verschiedenen UmweltseinfliisRen zum
mindesten die Temperatur in dieser Richtung untergeordnete Bedeutung. Im ubrigen hatten die Puppen den groBten Teil ihrer Eotwicklung in ihrem naturlichen Winterlager verbracht. Sie wurden demselben wenige Wochen vor Beginn der Versuche, die sich von Marz bis Eude Mai erstreckten, entnommen.
I) Die Yethode gilt selbstverstiindlich zunachst nur fur die Kieferneule, als einem Insekt mit postmetaboler Eientwicklung [vgl. E i d m a n n (21)J das keinerlei ,,Reifungs- fraB'' im Imaginalleben durchmacht.
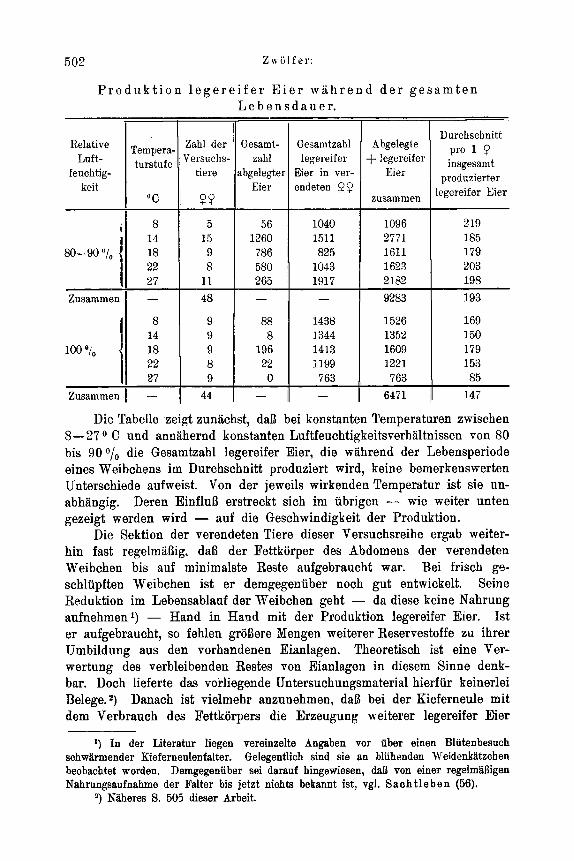
502 Z w 6 I f e r:
P r o d u k t i o n l e g e r e i f e r E i e r w a h r e n d d e r g e s a m t e n
Relative Luft-
feuchtig- keit
I
i 80-90 "lo
I
I Zusammen
looeio I Zusanimen
- - L e b e n s d a u er .
Durchschnitt
insgesam t produzierter
legereifer Eier
pro 1 0
219 185 179 203 198 193
169 150 179 153 85
147
Die Tabelle zeigt zunachst, daB bei konstanten Temperaturen zwischen 8-27 0 C und annahernd konstanten Luftfeuchtigkeitsverhaltnissen von 80 bis 90 die Gesamtzahl legereifer Eier, die wahrend der Lebensperiode eines Weibchens im Durchschnitt produziert wird, keine bemerkenswerten Unterschiede aufweist. Von der jeweils wirkenden Temperatur ist sie un- abhangig. Deren EinfluB erstreckt sich im iibrigen - wie weiter unten gezeigt werden wird - auf die Geschwindigkeit der Produktion.
Die Sektion der verendeten Tiere dieser Versuchsreihe ergab weiter- hin fast regelmaBig. daS der Fettkorper des Abdomens der verendeten Weibchen bis auf minimalste Reste aufgebraucht war. Bei frisch ge- schlupften Weibchen ist er demgegeniiber noch gut entwickelt. Seine Reduktion im Lebensablauf der Weibchen geht - da diese keine Nahrung aufnehmenl) - Hand in Hand mit der Produktion legereifer Eier. 1st er aufgebraucht, so fehlen grol3ere Mengen weiterer Reservestoffe zu ihrer Umbildung aus den vorhandenen Eianlagen. Theoretisch ist eine Ver- wertung des verbleibenden Restes von Eianlagen in diesem Sinne denk- bar. Doch lieferte das vorliegende Untersuchungsmaterial hierfur keinerlei Relege.2) Danach ist vielmehr anzunehmen, daB bei der Kieferneule mit dem Verbrauch des Pettkorpers die Erzeugung weiterer legereifer Eier
I) In der Literatnr liegen vereinzelte Angaben vor uber einen Bliitenbesuch schwarmender Kieferneulenfalter. Gelegentlich sind sie an bluhenden Weidenkatzchen beobachtet worden. Demgegenuber sei darauf hingewiesen, daB von einer regslmaBigen Nahrungsaufnahme der Falter bis jetzt nichts bekannt ist, vgl. Sachtle ben (56).
*) Naheres S. 505 dieser Arbeit.

Studien zur ijkologie und Epidemiologie der Insekten. 50::
eingestellt wird. Die fur diese gefundenen Durchschnittszahlen geben dementsprechend bei der Kieferneule einen brauchbaren Wert fur ihre ,,absolute" Eizahl.
Im Mittel der 48 Versuchstiere obiger Zusammenstellung ergi bt sich auf diese Weise als D u r c h s c h n i t t s z a h l p r o Weibchen p r o d u z i e r t e r l ege re i f e r E i e r i n d e r T rocken lu f t s tu fe d e r Wer t 1 9 0 (abgorundet). E r i s t a l s d i e abso lu t e E izah l d e r Kieferneule a n z u s e h e n , w a h r e n d d e n u n t e r v e r s c h i e d e n e n V e r s u c h s b e d i n g u n g e n ode r a u c h in f r e i e r N a t u r abge leg ten Durchschn i t t s e imengen i n den nieis ten Fa l l en n u r r e l a t i v e B e d e u t u n g zukommt.
Demgegenuber lieferten die mit extremen Luftfeuchtigkeitsverhalt- nissen von 100 o/o kombinierten Temperaturversuche wesentlich andere Werte fur die Durchschnittszahl pro Weibchen produzierter legereifer Eier. Sowohl im Gesamtdurchschnitt, als auch in den einzelnen Temperatur- stufen (mit Ausnahme jener von 180) bleiben diese Zahlen erheblich unter der GroBenordnung jener zuruck, die bei 80-90 O/o relativer Luftfeuchtig- keit erzielt wurden. Am deutlichsten kommt dies in der Kombination der Temperaturstufe von 27 O mit 100 o/o relativer Luftfeuchtigkeit zum Ausdruck. Die Durchschnittszahl pro Weibchen produzierter legereifer Eier betrug hier 85. Bemerkenswerterweise und in deutlichem Gegensatz zu den Trockenluftversuchen ergaben die Sektionen der bei 100 O i 0 Luft- feuchtigkeit verendeten Weibchen in der Regel noch groBere oder ge- ringere Mengen von Pettkorpersubstanz in deren Abdomen. Offenbar liegen hier durch die Kombination mit feuchtigkeitsgesattigter Luft schadigende Einflusse vor, die ein vorzeitiges Absterben der Tiere zur Folge haben. I m ubrigen betrug im G e s a m t d u r c h s c h n i t t d i e Z a h l p r o W e i b c h e n b e i T e m p e r a t u r e n z w i s c h e n 8-27O C u n d 1 0 0 O / o L u f t f e u c h t i g k e i t p r o d u z i e r t e r l e g e r e i f e r E i e r r u n d 150.
Das Gesagte laBt sich wie folgt zusammenfassen: die absolute Zeugungskraft von Punolis flummea Schiff. gemessen an der Eizahl, die ein Weibchen im Optimum aller maBgebender Bedingungen ablegen kann, betragt auf Grund der Untersuchung von 48 Weibchen 190.
Die Gesamtzahl legereifer Eier, die ein Weibchen wahrend seines Lebensablaufes im Durchschnitt erzeugt, erweist s k h bei einer Luftfeuchtig- keit von 80-90 O / o innerbalb eines Temperaturbereiches zwischen 8 bis 27 0 C als unabhangig von den jeweiligen Temperaturen. Peuchtigkeits- gesattigte Atmosphare ubt schadigenden EinfluB auf die Palter aus, inso- fern sie die Produktion legereifer Eier merklich vermindert, letzteres be- sonders bei hoheren Temperaturen (27 0 C).
Ob und inwieweit die Zahl der von einem Weibchen eweugten lege- reifen Eier zur Ablage gelangt, ist eine Prage fiir sich, auf die weiter unten eingegangen werden wjrd.

504 Z w o l f e r :
Uber die Eizahl von I'anolis flamniea Snhiff. liegen in der Literatur Angaben vor von E s c h e r i c h und B a e r (t 'Z), E c k s t e i n (ZO), S a c i i t l e b e n ( 5 5 ) , S p r e n g e l (65), E id - m a n n (21). Dic beidrn erstgenannten Butoren haben auf Grund YOU Zwingerbeobach- tungen an drei eierlegenden Weibchen und von Sektionen mehrerer Tiere fur die pro Weibchen abpelegte Eizahl Werte ermittelt, die um 200 liegen. lhre Angabe kommt dem oben mitgeteilten Wert der idealen Eizahl der Kieferneule am nahesten. - E c k s t e i u und anfanglich auch S a c h t l e b e n vertreten die Auffassung, dalj unter giinstigen Be- dinqungen aus simtlichen Eianlagen der Ovarialschiaiiche legereife Eier hervorgeheu. Eid m a n n hat indessen gezeigt - und die vorliegenden Beobachtungen bestatigen dies vollkommeu - daQ stets nur ein Teil der Eianlagen der Ovarien seine Entwicklung bis zum legereifen Ei durchlauft. Die Mehrzahl derselben bleibt zuriicli ohne ihre Ent- wicklung abzuschlielen. S p r e n g e l nahm auf Grund der Sektion von 6 Weibchen 300 bis 400 als Gesamtzahl legereifer Eier an, die von einem Weibchen erzeugt werden konnen. Diese Zahl als D u r c h s c h n i t t s w e r t ist vermutlich zu hoch gegriffen. S a c h t l e b e n f a l t neuerdings (56) seine eigenen und die in der Literatur vorliegenden Angaben uber die Eizahl der Kieferneule dahin zusammen, d a l die ,,von einem Forl- eulenweibchen abgelegte Eizahl im Durchschnitt etma 150 betriigt", die Hochstzahl liegt nach ihm bei 2.50-260 Stuck.
c) Die ovariale Eientwicklung in1 Puppen- und Imaginalstadium.
Welchen Zustand weisen die Ovarien schliipfender Weibchen auf, und wie wird deren weitere Entwicklung durch aul3ere VerhaItnisse be- einfluBt?
Wird zur Reurteilung des Entwicklungsznstandes der Ovarien schlupfender Weibchen die Zahl der in dieseu vorhandenen legereifen Eier gewahlt, so lassen sich bekanntlich bei den Lepidopteren 2 bezw. 3 Gruppen unterscheiden [ E i d m a n n (21)]. Die Ovarien der Weibchen der ersten Gruppn enthalten beim Schliipfen der Falter keine oder sehr wenig legereife Eier. Deren Entwicklung verlauft postmetabol. Bei der zweiten Gruppe finden sich in den Ovarien schlupfender Weibchen legereife Eier in mehr oder rninder groBer Zahl. Diese letztere Abteilung u m f 9 t nach E i d m a n n Formen, bei denen die Eientwiclilung mit dem Schliipfen der Weibchen bereits abgeschlossen ist und postmetabol keine weitern Umbildung von Eianlagen zu legereifen Eiern stattfindet. Sie enthalt ferner solche Arten, bei deren schlupfenden Weibchen nur ein TeiL des spiiterhin abzulegenden Eivorrates legereif entwickelt ist und postmetabol wahrend des Imaginallebens noch eine weitere Erzengung legereifer Eier stattfindet.
Was im besonderen Panolis flammea Schiff. angeht, so stellt E i d m a n n auf Grund der Untersuchung von 6 frisch geschliipften Weibchen fest, dalj diese Art der zweiten Gruppe zugehort, ihre Weibchen rnit durchschnittlich 86 legereifen Eiern die Puppenhulle verlassen, und ihre Eischlauche im iibrigen postmetabol eine weitere Entwicklung durch- laufen.
Es war zunachst naheliegend anzunehmen, daB die Temperaturen, die vor dem Schliipfen auf die Puppen einwirkec, einen gewissen EinflulS auf diese Verhaltnisse ausuben. So wurde ein Teil der in einem Puppen- temperatiirversuch schliipfenden Weibchen kurz nach dem Schliipfen ein weiterer Tell, der im Imaginalstadium in den betreffenden Temperaturstufen verblieb, in verschiedenen Zeitabstanden nach dem Schliipfeu seziert und auf den Entw icklungszustand seiner Ovarien hin untersucht. Die Ternperaturen dieser Versuchsreihe lageu bei 40, 8 0 , 140, 180 und 220 C. Samtliche Puppenversuche waren mit 100 O / o relativer Luftfeuchtigkeit kombiniert.
Ahnliche Befunde teilt S p r e n g e l (65) mit.
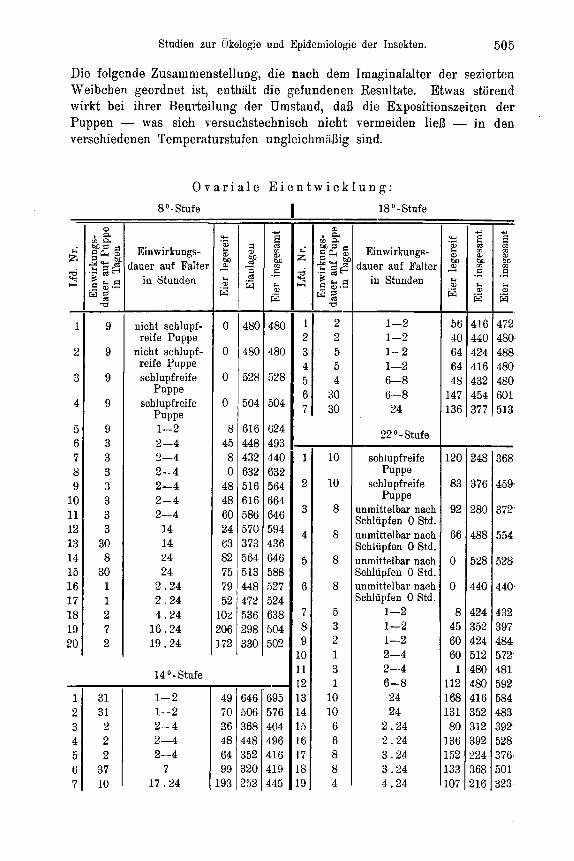
Studien zur Okologie uud Epiderniologie der Insekten. 505
Die folgende Zusammenstellung, die nach dem Imaginalalter der eezierten Weibchen geordnet ist, enthalt die gefundenen Resultate. Etwas storend wirkt bei ihrer Beurteilung der Umstand, da13 die Expositionszeiten der Puppen - was sich Fersuchstechnisch nicht vermeiden lie13 - in den verschiedenen Temperaturstufen ungleichm613ig sind.
1 2
6 7
-
6 3 - 1
2
3
4
5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20
31 1- 2 31 1-2
3 2 2- 4 4 2 2-4 5 2 2-4
37 7 10 17.24
O v a r i a l e 8O- Stufe
Einwirkungs- lauer auf Faltei
in Stunden
nicht sehlupf- reife Puppe
nicht schlupf- reife Puppe schlupfreife
schlupfreife
1-2 2-4 2-4 2-4 2-4 2-4 2-4
PuPPe
PUPpe
14 14 24 24
2 .24 2 .24 4.24
16.24 19.24
14 O - Etufe
- - u .* al m 2
i4
0
0
0
0
3
& al
-
tr 4.5
8 0
48 48 60 24 63 82 75 79 52 oz 106 72
- 49 70 36 48 64 99
193
- - 0
e H 0 m 6
- C80
4 80
528
j04
516 C48 L32 B 2 )16 i16 )86 )70 I73 b64 )13 L48 172 136 198 130
- 146 106 68 48 52 20 52
E i e n t w i c k l u n g :
- - 3 m ar bo m
.- c aJ E
480
480
V'L8
504
624 493 440 632 564 664 646 594 436 646 588 527 524 638 504 502
-
-- 695 576 404 496 416 419 445
d 4
- 1 2 3 4 5 6 7
- 1
2
3
4
5
6
7 8 9 0 1 2 3 4 5 6 7 8 9
--
2 2 5 5 4
30 30
- 10
10
8
8
8
8
5 3 2 1 3 1
10 10 6 6 8 8 4
18O- Stufe - Einwirkungs-
dauer auf Fake: in Stunden
1-2 1-2 1-2 1-2 6-8 6-8 24
22 O - Stufe
schlupfreife Puppe
schlupfreife PWPe
inmittelbar nack 3chliipfen 0 Std inmittelbar nact 3chliipfen 0 Std inmittelbar nact 3chlupfen 0 Std inmittelbar nach 3chlupfen 0 Std
1-2 1-2 1-2 2-4 2-4 6- 8 24 24
2.24 2.24 3.24 3.24 4.24
- - v .- E b
G
56 40 64 64 48
147 136
ar
& a -
- 120
83
92
66
0
0
8 45 60 60 1
112 ,68 .31 80 .36 -52 33 07
416 472 440 480 424 488 416 480 432 480 454 601 377 513
- 248
376
280
488
528
440
424 352 424 512 480 480 416 352 312 392 224 368 216
- 368
4.59.
372
554
528
440
432 397 484 572 481 592 584 483 392 538 376, 501 323

506 Zwolfer :
Bus diesen Daten geht zunachst hervor, daB das Schlupfen der Falter im allgemeinen an einen fortgeschrittenen Entwicklungszustand der Ovarien gebunden ist, insofern eu dessen Beurteilung die Zahl legereifer Eier herangezogen wird. Stets sind solche in groBerer oder geringerer Menge vorhanden. Ausnahmen von dieser Regel kommen vor - so fanden sich unter jnsgesamt 47 untersuchten Weibchen kurz naah dem Schlupfen noch 3 Tiere, deren Ovarien keine legereifen Eier enthielten - doch kommt ihnen kaum grundsatzliche Bedeutung zu. Man kann hierbei an ein verfruhtes Schliipfen denken, da13 vielleicht auf mechanischen Storungen beruht, wie solche sich bei haufiger Kontrolle der Versuchs- tiere nicht ganz vermeiden lassen.
Was die Zahl legereifer Eier bei schlupfenden Weibchen angeht, so ist sie allerdings etwas starkeren Schwankungen uoterworfen, als dies aus den Angaben der oben zitierten Autoren hervorgeht. Die Extreme liegen in dieser Hinsicht bei 0 und 120 Stuck. Eine gewisse Abhangig- keit dieser Zahlen von den Temperaturen, die auf die Puppen einwirkten, kommt wenigstens fur die 8 0 C-Stufe zum Ausdruck, wenn in obiger Zusammenstellung die legereifen Eier schlupfreifer Puppen,I) frisch geschlupfter und 1-4 Stunden alter Palter ins Auge gefal3t werden: Wahrend in der 80-Stufe die Ovarien der Weibchen im Mittel 24,l legereife Eier enthalten, fanden sich in der 140-, 180- und 22 O-Stufe im Mittel 53,4, 56 bezw. 48,6 solcher Eier. Die Zahl l ege re i f e r E i e r i s t demnach bei Weibchen , die a l s P u p p e i n der Zei t vor dem Sch lupfen du rchweg n iede ren Tempera tu ren b i s zu 8 0 C aus- gese tz t waren, r u n d ha lb so groB a ls be i solchen, auf welche i n d i e se r Zei t hohe re Tempera turen e inwirk ten . Die folgende Tabelle laBt dies im einzelnen ubersichtlich erkennen :
(Siehe Tabelle auf 8. 507 oben.) Die uber die postmetabole Eientwicklung vorliegenden Daten sind
sparlich, lassen aber immerhin gut erkennen, da13 im Lauf des Imaginal- lebens der Weibchen, wenn keine Eiablage erfolgt, die Zahl d e r lege- r e i f e n E i e r i n den Ovarien zu- und j e n e de r Eian lagen abnimmt. Bei hoheren Temperaturen erfolgt dieser ProzeB rascher als bei niederen. Die von anderer Seite geauBerte Ansicht, nach der wahrend dieser Zeit eine Zunahme der Gesamteizahl (legereife Eier + Eianlagen) stattfinden soll, laBt sich an Hand des vorliegenden umfangreicheren Materials nicht bestatigen. Beziiglich des Pettkorpers im Abdomen der untersuchten
l) Uber die Merkmale ,,schlupfreifer" Puppen vgl. P i c t e t (49), dessen diesbeziigliche Angaben nach den Erfahruugen bei der vorliegenden Untersuchung auch fur die Forl- eule gelten. - Es sei bei dieser Gelegenheit erwahnt, da13 im Rahmen dieser Arbeit durchgefuhrte Versuche zur Feststellung eines etwaigen Einflusses des atmospharischen Druokes auf das Schlupfen der Puppen von P. flammea kein eindeutiges Ergebnis lieferten. Ein solcher wird von Wolf f und ErauPe (73) wohl in Anlehnung an P i c t e t s Resul- tate mit anderen Lepidopteren auch fur die Forleule angegeben.
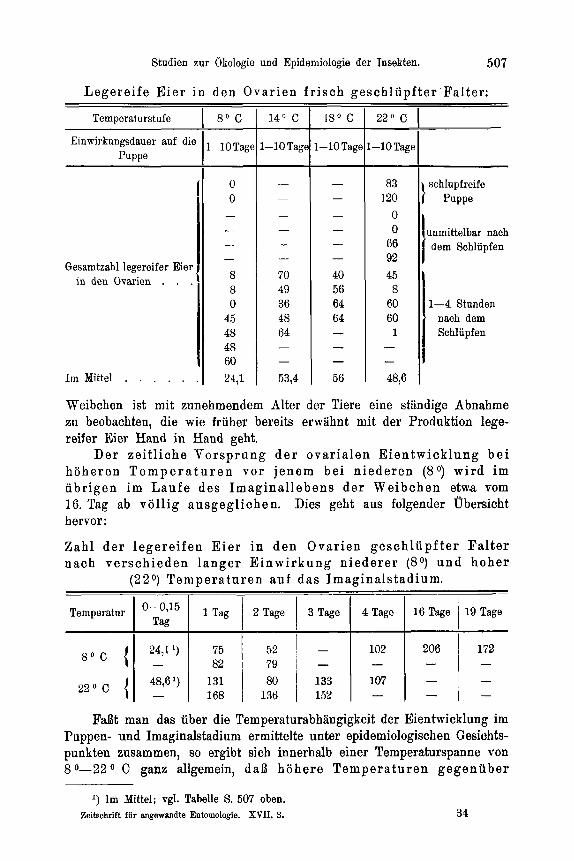
Studien zur Okologie und Epidemiologie der Insekten.
Legereife Eier i n den O v a r i e n f r i s ch geschliipfter Falter:
507
Temperatur 0-0,15 I Tag
24,l L, - 8OC { 48,6') 2 2 o c { -
-
Temperaturstufe I S o C
-
1 Tag 1 2 Tage 1 3 Tage 4 Tago 16 Tage 19 Tage
102 206 172 - - - -
75 1 5 2 , - I 82 79
- - 131 80 1 133 107 168 136 I 1YL - - -
Einwirkungsdauer auf die Puppe I
I
{ Gesamtzahl legereifer Eier
in den Ovarien . . .
I
~
0 0 - - - -
8 8 0
45 48 48 60
Im Mittel . . . . . . I 24,l
14O C
.-lo Tag
- 1 8 O C
1-10Tagt
- 22" c
-10 Tagc
83 120
0 0
66 92 45
8 60 60
1 - - 48,6
schlupfreife PWP9
inmittelbar nach dem Schliipfen
1-4 Stunden nach dem Schliipf en
I) Im Mittel; vgl. Tabelle S. 507 oben. Zeitsohrift f i r angewandte Eotomologie. XVII, 3. 34

508 Z w 6 l f e r :
t i e f e r e n led ig l ich e inen b e s c h l e u n i g e n d e n E i n f l u B auf d ie B i ldung l e g e r e i f e r E i e r ausiiben: d ie Gesamtprodukt ion d e r l e t z t e ren mird h i e rdu rch n i ch t beeinflufit, wenn schiidliche Um- weitseinfliisse (100 O l 0 rel. Luftfeuchtigkeit) wahrend des Imaginallebens ausgeschaltet sind.
(1) Das Soliliipfm der Falter.
Was das Schliipfen der Falter in den einzelnen Temperaturstufen angeht, so erfolgte es in der 220-Stufe am raschesten und war hier etwa 10 Tage nach Versuchbeginn abgeschlossen. In der 8 0- Stufe wiihrte es am langsten - ca. 21/2 Monate - und war auch irn ganzen Verlauf am ungleichmaBigsten. Die 18 0- und 14 0- Stufe lieferte dazwischen- liegende Werte.
Von besonderem Interesse ist das Verhalten der Puppen in der 40-Stufe: Trotz vier Monate langer Ausdehnung dieses Versuches - er wahrte vom 2. Februar bis 5. Juni - schlupften aus 50 gesunden Puppen in dieser Zeit nur 1 Mannchen; letzteres gleich zu Versuchsbeginn. Die Uberfiihrung der Puppen nach Abbruch des Versuches aus der 40-Stufe in Temperaturen von 20-220 C ergab in den folgenden 2 Wochen aus samtlichen Puppen normale Falter. Zwischen 4 0 und SO C muB dem- nach ein Temperaturgrenzwert liegen, der fiir die Erlangung der Schlupf- reife uberwinternder Puppen der Kieferneule bedeutungsvoll ist.
In scheinbarem Gegensatz hierzu standen Beobachtungen, die zeigen, daB das Schliipfen der Palter auch noch bei 2 0-3 0 C erfolgen kann: Wurden namlich Puppen, die im Dezember dem Winterlager entnommen und langere Zeit anschliel3end bei Zimmertemperatur (18 0 ) gehalten waren, bei welcher sie schlieBlich zu schliipfen begannen , zur Verhinderung meiteren Schliipfens den angegebenen Temperaturen ausgesetzt, so blieb die schliipfhemmeude Wirkung der niederen Temperaturen aus. Das Schliipfen erfolgte zwar langsamer, verteilte sich iiber langere Zeit als es bei Zimmertemperatur der Fall war, indessen trat kein volliger Still- stand ein. Wie erklaren sich diese gegensatzlichen Beobachtungen?
-4uBerlich ist bekanntlich der Falter von Panolis flammea in der Puppe schon im Spatsommer entwickelt. Sacht leben (56) erwahnt, dal3 rund 4 Wochen nach Beginn der Verpuppung diese Umwandlung voll- zogen sei. Dies gilt fur Freilandbedingungen und konnte an Puppen- material, das im August und September 1930 von verschiedenen Forst- amtern zur Untersuchung eingeschickt worden war, bestatigt werden.
Die inneren Organe indessen - zum mindesten das generative Organsystem der Weibchen - haben um diese Zeit noch nicht d e n Entwicklungsgrad erreicht, wie er bei schliipfreifen Puppen und schliipfen- den E’altern angetroffen wird. Weibliche Puppen, die im November und Dezember dem Winterlager entnommen und unmittelbar anschlieBend seziert wurden, zeigten dies deutlich: Von legereifen Eiern, die in den

Studien zur Okologie und Epidemiologie der Insekten. 509
Ovarien schlupfender Weibchen fast stets in grofierer oder geringerer Zahl rorhanden sind (vgl. S. 505), ist hier noch nichts zu bemerken. Die Ovarialschlauche zeigen in perlschnurformiger Anordnung und gegen das blinde Ende zu sich verjungend nur Eianlagen in einem sehr unent- wickelten Zustaud. Die einzelnen ,,Perled' sind vergleichsweise aufier- ordentlich klein. I m ubrigen sind die Schlauche vollig im Pettkorper eingebettet, der auf dieser Entwicklungsstufe fast das ganze Abdomen ausfullt. Seine Lappen liegen vielfach den OvarialscblHuchen so fest an, dafi sie beim Praparieren nur schwer von diesen zu trennen sind.
Was die Zahl der Eianlagen angeht, die in den Ovarien den1 Winterlager im November entnommener Puppen gefunden wurden, so betriig sie bei 6 sezisrten Puppen: 463, 464, 504, 512, 576, 600; im Durchschnitt 519. In derselben Grofienordnung bewegen sich - wie fruher gezeigt wurde - die Gesamteizahlen (legereife Eier + Eianlagen) geschlupfter Weibchen. E i n e V e r m e h r u n g de r Gesamteizahl f i n d e t demnach im Anschluf i a n d i e Uberwin te rung n i c h t s ta t t . Hin- gegen erfolgt bei der weiblichen Piippe nach der Uberwinterung auf Kosten des Bettkorpers eine weitere Entwicklung der vorhandenen Ei- anlagen, von denen ein Teil beim schlupfenden Palter die Legereife er- reicht. Entsprechende Um wandlungen werden wahrscheinlich auch die Geschlechtsorgane der mannlichen Puppen nach der Uberwinterung durch- laufen, an denen in dieser Richtung noch keine Untersuchungen vor- genommen worden sind. Bus dem Gesagten geht zusammenfassend her- vor, dafi die Puppen von Yunolis pammea i m Fruhjahr zur Erlangung der Schlupfreife eine weitere Eutwicklung durchmachen, die sich auf innere Organe bezieht.
Damit erklart sich der scheinbare Gegensatz der oben mitgeteilten Beobachtungen : Zu jedem Entwicklungsvorgang wird eine gewisse Tempe- ratur benotigt. Unterhalb einer bestimmten artspezifischen Temperatur- stufe werden Entwicklungsvorgange unterbrocben. Diese Vorstellung liegt der Blunck schen Wlirmesummenregel zu Grunde.1) Ubertragt man sie auf den vorliegenden Fall, so ergibt sich als u n t e r e Tempera tur - g renze (Entwicklungsnul lpunkt ) z u r E r l a n g u n g d e r S c h l u p f r e i f e bei den i i be rwin te rnden Puppen v o n Pan,olis flamrnea Schiff . e in Wer t , d e r zmischen 40 bis 80 C l iegt .
Wird die Formel der Warmesummenregel T (t-tol = k auf die oben iiber das Schliipfen der Falter in der 8 O- und der 22 O - Temperaturstufe mitgeteilten Zeiten aogewendet, so erhalt man fur den Entwicklungsnullpunkt einen Wert von to = 5,s und fur die Thermal- konstante k = 160. Ohne auf diese beiden Zahlen besonderen Wert zu legen - die zu ihrer Ermittlung verwendeten Daten sind zu unvollstandig - sei in diesem Zusammen- hang n u darauf hingewiesen, da13 sich die Tempeiatnrstufe von rund 60 C auch spater- hin bei allen weiteren Beobachtungen an den verschiedenen Stadien der Kieferneule als ausgezeichneter Wert erwies. Unterhalb dieser Temperaturgrenze unterbleiben die Ent- micklungsvorgange oder werden dooh auf ein Minimum reduziert.
l) Vgl. B l u n c k (6), B o d e n h e i m e r (8, 9, l l ) ? M a r t i n i (41), S h e l f o r d (63) u a. 34 *

Zwolfer : 510
Man wird daher nicht fehlgehen in der Annahme, daB die Temperatur- grenze von rund 6 0 C auch i m Leben der Puppe die Rolle eines Ent- wickliingsnullpunktes spielt. Dauernder Verbleib der Puppen im AnschluB an die Uberwinterung in Temperaturen unterhalb dieser Stufe wurden demnach die Entwicklungsvorgange, die zur Schlupfreife der Puppe fuhren und damit das Schlupfen der Falter verhindern. Steigt die Umgebungs- temperatur zeitweise uber den angegebenen Grenzwert, so seteen die Vorgange der inneren Entwicklung ein, die zur Schlupfreife fuhren. Sinkt die Temperatur vorubergehend unter den Grenzwert, dann wird dieser Vorgang unterbrochen.
Der oben mitgeteilte Befund, dalj das Schlupfen der Eulenpuppen auch unterhalb der angegebenen Temperaturgrenze erfolgen kann, wider- spricht keineswegs dem Gesagten : Die betreffenden Puppen waren zuvor bei Zimmertemperatur gehalten und hatten bier ihre Entwicklung bis zur Schlupfreife durchlaufen. Die anschlieaend vwgenominene Erniedrigung ihrer Umgebungstemperatur auf 2 O - 3 O C reichte nicht aus, um den Vorgang des Schlupfens selbst, den Schlupfakt, ganzlich zu verhindern. Dieser letztere ist somit von jeneni Temperaturgrad, der durch den Ent- wicklungsnullpunkt vorgestellt wird, unabhangig. Fur ihn ist vermutlich der Aktivitatsnullpunkt der Falter die mallgebende 'Temperaturgrenze. Uber diesen liegen noch keine eingehenderen Untersuchungen vor. Einige orientierende Versuche an 4 Faltern (2 $2 d) zeigten, daB die Eiiltestarre bei der Kieferneule bei Temperaturen zwischen - 2 O bis + 40 C ein- treten kann. Oberhalb dieser Temperaturen waren die Versuchstiere noch aktiv beweglich. Ein Auskommen der Palter aus der schlupfreifen Puppe bei Temperaturen zwischen - 2 0 bis + 4 0 C ware demnach durchaus denkbar.
Theuerkauf (zitiert nach S a c h t l e b e n 56) beriohtet sogar, daIj er Forleulenfalter bei Schnee und Lufttemperaturen von - 5 O C in Bewegung angetroffen hat.
Das Gesagte laBt sich wie folgt zusammenfassen: Zur Er l angung d e r Sch lupf re i f e bedurfen d i e u b e r w i n t e r n d e n Kie fe rneu lenpuppen e i n e r gewissen Warmesunime der Umgebung. Als wirksam er- miesen s ich h ie r f ur Tempera turen oberha lb e i n e s Grenzwer tes , d e r zwischen 4-80 C liegt. Beobachtungen uber den Ze i t r aum, d e n das Schlupfen d e r Puppen i n verschiedenen kons tan ten Tem- pe ra tu ren beansprucht und theore t i sche Erwagungen sp rechen dafiir, dalj d ieser Grenzwer t annahe rnd bei 60 C liegt. Das Sch lupfen se lbs t i s t v o n d ieser Tempera tu r unabhang ig : F u r d i e u n t e r e Tempera tu rg renze des Schlupfaktes ist ve rmut l i ch d e r u n t e r e Ak t iv i t a t snu l lpunk t maljgebend, d. h. jene Temperatur, hei der die Eiiltestarre der Falter eintritt. -
Welche theoretischen Folgerungen ergeben sich ails diesen Uber- legungen fur das Schlupfen der Puppen im Freiland?

Studien zur dliologie und Epidemiologie der Insekten. 511
Jene Puppen, die an Platzen mit geringem Warmeschutz liegen, welche im Fruhjahr der raschesten Erwartnung ausgesetzt sind, werden im allgemeinen ihre Schliipfreife fruher erlangen als solche, die an Orten mit gutem Wlrmeschutz uberwintern. Dies gilt allerdings nur dann, wenn sowohl a n den Pllitzen mi t g e r i n g e m a ls a u c h a n j enen mit s ta rkem Wiirmeschutz die Abkuhlung wahrend kalter Zwischen- perioden u n t e r den ~ntwicklungsnullpunkt von 60 C sinkt. 1st dies n i c h t der Pall oder sinkt n u r an den schlecht warrnegeschutzten Orten die Temperatur unter den genannten Grenzwert, wahrend sie an gut ge- schutzten gleichzeitig auch wahrend der kiihlen Perioden (nachtliche Ab- kuhlung !) oberhalb des Grenzwertes verbleibt, dann kann ein Ausgleich erfolgen : Der etwa vorhandene Entwicklungsvorsprung der Puppen an Orten mit rascher Erwarmungsmoglichkeit wurde in diesem Fall von Puppen, die an gut geschiitzten Orten liegen, wieder ausgeglichen werden.
Als Uberwinterungsplatze mit geringem Warmeschutz im obigen Sinn haben Boden mit dunner Streudecke und dunnem Humusbelag zu gelten, insbesondere wenn sie in erhohtem MaBe der Insolation ausgesetzt sind. Auch streuberechte Flachen waren hierher zu zahlen. Guten Warme- schutz bieten demgegenuber Boden mit dicker Humusschicht, mit hohem Moosbelag dichter Beerkrautdecke usw. Mey e r s (46) diesbezugliche Tem- peraturmessungen bestatigen dies.
Es durfte hiermit zusammenhangen, daB Beobachtungen aus dem Freiland vorliegen, die sowohl von einem zeitigeren Schlupfen der Puppen bei dunnem Streubelag sprechen [streuberechte Wachen: vgl. Sacht- leben (56), J u d e i c h - N i t s c h e (37)] als auch andere, nach denen ein Unterschied der Schlupftermine bei gut warrnegeschutzten Boden und solcheu mit geiingem Warmeschutz nicht in Erscheinung tritt (Me yer). Welche Bedeutung im ubrigen ein vorzeitiges Schlupfen der Falter fur diese und ihre Nachkommenschaft haben kann, wird in anderem Zusammen- hang erortert werden.
Auch die in der Literatur vorliegenden Angaben uber zeitlich oft erheblich weit auseinander liegende Schlupftermine der Falter an ein und demselben Biotop, lassen sich nit dem geschilderten Warmebedarf der Puppe bei der Pruhjahrsentwicklung in Einklang bringen: Da die Puppe der Kieferneule sowohl in den obexen, wie auch in den mi t t l e r en und den u n t e r e n Schichten der Bodendecke uberwintert, an all diesen Platzen aber die verschiedenartigsten Warmeschutzverhaltnisse herrschen, so erscheint es nach den obigen Darlegungen verstiindlich, daB die ge- samte Schlupfperiode in freier Natur sich stets uber einen langeren Zeitraum erstreckt: hier beginnt das Schlupfen der Palter in der Regel Ende BSarz und dauert bis Anfang Juni an. Bre t tmann [zit. nach Sach t l eben (56)] terichtet sogar von einem Auskommen der Falter Mitte Februar. Die genannten Zeitpunkte sind Extreme. Die Hauptschlupfzeit wird bald fruher bald spater liegen - je nach den Witternngsverhaltnissen, die

512 Zwotfer :
in den einzelnen Jahrgangen herrschen, und unter denen nach der vor- liegenden Untersuchung die Temperatur den HaupteinfluQ auszuiiben scheint. -
e) Geschlechtliclie dktivitfit und Eiabpdge. Neben dem Geschlechterverhaltnis, das an dem vorliegenden Material
auf Grund der Untersuchung der Merkmale von 150 Puppen sich auf rund 1 : 1 belief, 1) ist eine Abhangigkeit der geschlechtlichen Aktivitat von Umweltseinflussen, soweit sie in der Begattung ihren Ausdruck findet, epidsmiologisch von Interesse. Die nachfolgende Zusammenstellung, die die Resultate von 23 Zuchtversuchen bei 5 verschiedenen Temperatur- und 2 Luftfeuchtigkeitsstufen umfaBt, zeigt in der Tat, daB eine solche in erheblichem AusmaB besteht. 2) MaBgebend fur die Beurteilung des Zustandekommens der Kopula war der Sektionsbefund der Weibchen nach ihrem naturlichen Lebensablauf. Wahrend die Bursa copulatrix samt- licher begatteter Versuchsweibchen regelmlBig nur eine Sperinatophore enthielt, wies sie bei mehreren im Freiland gesammelten, naturlich ver- endeten Weibchen 0, 1, 2 und selbst 3 Spermatophoren auf. Letzteres zeigt, daB mehrmalige Begattung der Kieferneulenweibchen vorkommen kann und bestatigt gleichzeitig diesbezugliche Befunde von E c ks t e in (20) und Jnzen tkowsk i lnach Sacht leben (5611.
Gegen die Veisuche laBt sich einwenden, da13 die Behinderung des freien Fluges, im verhaltnismaflig engen Lebensraum der VersuchsgefaBe das teilweise Ausbleiben der Kopula zur Folge hatte. Dies mag zutreffen. Da indessen die ZuchtgefaBe in allen Fiillen gleiche Innenmasse und annahernd gleiche Bevolkerungsdichte hatten, wiirde da- durch nur die absolute Hohe der Zahlen betroffen, die die zustandegekommenen Kopu- lationen angeben. Die gefundenen Werte wurden auf jeden Fall relative Gultigkeit be- halten und damit eine ungefahre Vorstellung der optimalen Temperatur-Luftfeuchtigkeits- kombination geben, wie sie fur die geschlechtliche Aktivitat erforderlich ist. Bei dieser Gelegenheit sei darauf verwiesen, da13 unter dem oben erw%hnten sezierten Freiland- material sich ebenfalls ein unbegattetes Weibchen befand, ivoraus zu entnehmen ist, daB ein Ausbleiben der Kopula auch unabhkgig von beengtem Lebensraum vorkommt. Auch eine Zimmerzucht mit einer ZwingergroBe von rund cbm, die den Tieren geniigend Spielraum bot, bestatigte dies : Von 20 naturlich verendeten Weibchen dieser Zucht erwiesen sich bei der Sektion nur 12 als begattet, dabei standen Mannchen in hin- reichender Zahl zur Begattung zur Verfiigung.
Anschaulich sind diese Verhaltnisse in den Diagrammen Abb. 2 und Abb. 3 zum Ausdruck gebracht. Die in der letzteren den durch Kreise markierten Punkten zugeordneten Zahlen geben die Prozente zustande- gekommener Eopulationen an. Extreru hohe Luftfeuchtigkeitswerte von 1000/, zeigen auch bier - wenn sie wahrend der ganzen Lebensdauer
l) In einer besonders kontrollierten Aufzucht schlupften aus 64 Puppen 35 Mannchen und 29 Weibchen. Das Geschlechterverhaltnis ist ahnlich wie bei der Puppenunter- suchung rund 1: 1. - Zu entsprecbenden Resultaten gelangten Ecks te in (20) und S a c h t l e b e n (55). Es sol1 damit nicht gesagt sein, daB das Geschlechterverhaltnis der Kieferneule konstant diesen Wert bat. Wolff und E r a u s e (73) beispielsweise geben fur die Art ein Geschlechterverhalhis von 2 : 1 an.
21 Siehe Tabelle auf S. 513.
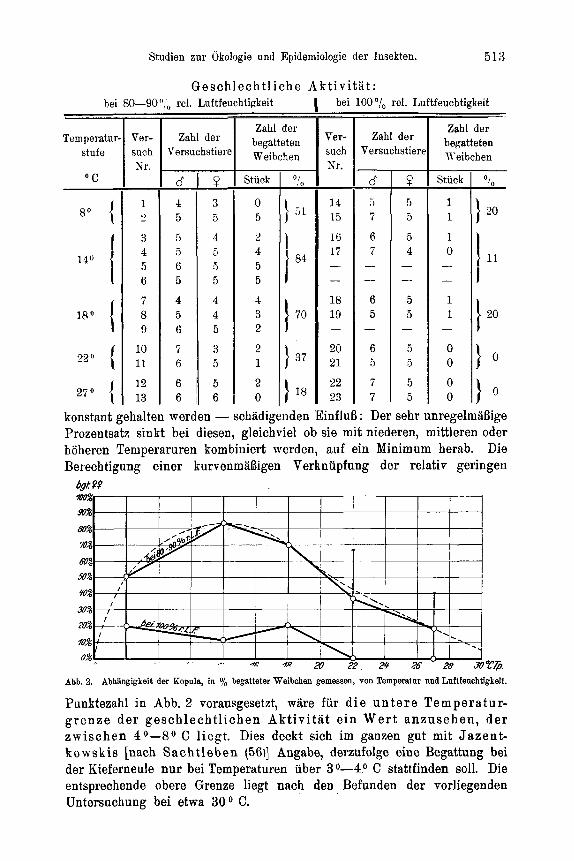
Studien zur dkologie und Epiderniologie der Insekten.
Geschlecht l iche Ak t iv i t a t :
513
bei 80-90",', rel. Luftfeuchtigkeit
Temperatur stufe
OC
go c I I
14') {
180 [ 220 { 27O (
- - Ver- such Sr .
1 2
3 4 5 ti
7 8 9
10 11
12 13
, ~,
Zahl der Versuchstiert
d - 4 5
5
6 5
4 5 6
7 6
6 6
>
0 - 3 5
4
.?I
5
4 4 5
3 5
5 6
3
Zahl der begatteteu Weibchen
Stuck
0 5
2 4 5 5
4 3 2
2 1
2 0
bei 100 '/,, rel. Luftfeuchtigkeit - - Ver- such Kr.
- 14 15
16 17 - -
16 19 -
20 21
22 23
Zahl der Versuchstiere
-
Zahl der begatteten Teibchen
Stuck
1 1
1 0
-
- -
1 1 -
0 0 0 0
konstant gehalten werden - schadigenden EinfluB : Der sehr unregelmaBige Prozentsatz sinkt bei diesen, gleichviel ob sie mit niederen, mittleren oder hoheren Temperaruren kombiniert werden, auf ein Minimum herab. Die Berechtigung einer kurvenmaBigen Verknupfung der relativ geringen
bgk??
' 46 ?R A9 22 24 26 28 30rq
Abb. 2. Abhingigkeit der Kopula, in yo begatteter Weibcheu gemessen, von Temperafur nod Luftfenchtigkeit.
Punktezahl in Abb. 2 vorausgesetzt, ware fur d i e ixntere Temperatur- g renze der geschlecht l ichen Akt iv i ta t e in W e r t anzusehen , der zwischen 4O-So C liegt. Dies deckt sich i m ganzen gut mit Jazent - kowskis [nach Sacht leben (56)J Angabe, derzufolge eine Begattung bei der Kieferneule nur bei Temperaturen uber 30-4O C stattfinden 5011. Die entsprechende obere Grenze liegt nach den Befunden der vorliegenden Untersuchung bei etwa 30° C.

514 Zwol fe r :
Uber das Temperatur - Luftfeuchtigkeitsoptimum fur das Zustande- kommen der Kopulationen gibt dss Diagramm Abb. 3 Anhaltspunkte. Nach ihm i ieg t d e r op t imale Bere i ch d e r geschlecht l ichen Ak t iv i t a t bei de r Kieferneule zwischen 120-160 und goo/, b i s scha tzungsweise 60 Oi0 Luf t f euch t ig kei t. Zu seiner genaueren Fixierung reichen die zur Verfiigung stehenden Versuchsergebnisse nicht aus, deren Erweiterung sich mit den vorhandenen Hilfsmitteln nicht durch- fiihren lie&
Das allgemeine Verhalten der Versuchstiere in den einzelnen Tem- perittur- und Feuchtigkeitsstufen
"r Tp
LT
Abb. 3. Diagramm der geschlechtlichen AktivitBt zur Bestimmung ihres optimalen Temperatur- 1,uftleuchti~keitsbereiches. (Die den Ereis- punkten zueeordoeten Zahlen geben den Durch- schnittsprozentsatz zustande gekommoner Kopu-
lationen an.)
gesattigter Atmosphare als eine
ist insofern bemerkenswert, als es sich zeigte, daR b e i s amt l i chen Tempe- r a t u r s t u f e n , d ie mit dem e x t r e m e n Luf t f euch t igke i t sg rad von 1000/o ve rknupf t waren , sowohl Mann- chen wie Weibchen i m a l lgemeinen n u r wenige u n d schmerfa l l ige Re- wegungen ausfuhr ten . Sie hingen groBtenteils bis zu ihrem Tode in einer Art S t a r r ezus t and an den Nadeln der eingestellten Kiefernzweige, von denen sie bei Entnahme abgeschuttelt werden konnten, ohne dabei irgendwelche Platter- versuche zu unternehmen. In den Tem- peraturstufen, die mit 80-90 ')/o Luft- feuchtigkeit verbunden waren, zeigten die Falter bei 18O-27O lebhafte bis sehr starke Beweglichkeit, in der 140-Stufe mar diese maBig, bei der 80-Stufe end- lich zeigten die Versuchstiere zumeist ein schwerfalliges an Starrezustande er- innerndes Verhalten.
Es liegt nahe, den eigenartigen Zu- stand der Versuchstiere in feuchtigkeits-
Art Feuchtigkeitsstarre aufzufassen. So betrachtet bildet er ein Gegenstuck zur Trocken- bezw. Warmestarre zahlreicher Insekten, die mehrfach bekannt geworden ist und in tropischen und subtropischen Gebieten wahrend der Trockenzeit bei der Ubersomme- rung der Tiere vielfach normalerweise eintritt. 1)
DaB in freier Natur anhaltend extrem hohe Luftfeuchtigkeit, wie sie beispielsweise bei Dauerregen herrscht, in der Tat erheblichen EinfluB auf die Aktivitat der Falter ausubt, ist den Beobachtungen versahiedener mittelfrankischer und oberpfalzischer Porstamter wahrend der letzten
l) Vgl. Uvarov (68), Zwolfer (76).
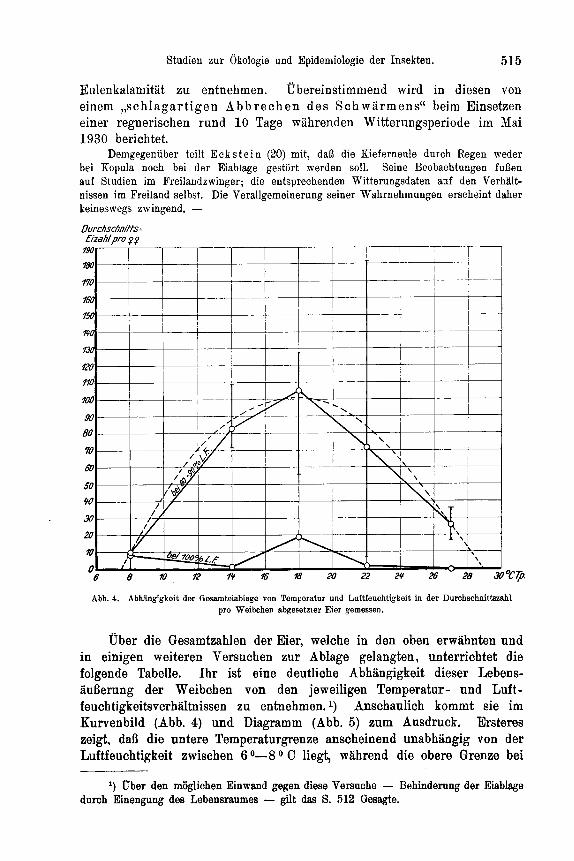
Studien zur &ologie und Epidemiologie der Insekten. 515
Enlenkalamitat zu entnehmen. Ubereinstimmend wird in diesen von einem , ,schlagart igen Abbrechen des Schwarmens" beim Einsetzen einer regnerischen rund 10 Tage wlhreiiden Wittemngsperiode im Nai 1930 berichtet.
Demgegenuber teilt E c k s t e i n (20) mit, daIj die Kieferneule durch Regen weder bci Kopula noch bei der Eiablage gestort werden soll. Seine Beobachtungen fu5en auf Studien im Freilandzwinger; die entsprechenden Witterungsdaten auf den Verhalt- nissen im Freiland selbst. Die Verallgemeinerung seiner Wahrnehmungen erscheint daher keineswegs zwingend. -
Abb. 4. Abhlngigkeit der Gesamteiablage von Temperator und Loftfeochtigkeit in der Durchschnittszahl pro Weibchen abgesetzter Eier gemessen.
Uber die Gesamtzahlen der Eier, welche in den oben erwahnten und in einigen weiteren Versuchen zur Ablage gelangten, unterrichtet die folgende Tabelle. Lhr ist eine deutliche Abhangigkeit dieser Lebens- auBerung der Weibchen von den jeweiligen Temperatur- und Luft- feuchtigkeitsverhaltnissen zu entnehmen. I) Anschaulich kommt Pie im Kurvenbild (Abb. 4) und Diagramm (Abb. 5) zum Ausdruck. Ersteres zeigt, da13 die untere Temperaturgrenze anscheinend unabhangig von der Luftfeuchtigkeit zwischen 6 0-8 0 C liegt, wahrend die obere Grenze bei
I) Qber den moglichen Einwand gegen diese Versuche - Behinderung der Eiablage durch Einengung des Lebensraumes - gilt das S. 512 Gesagte.
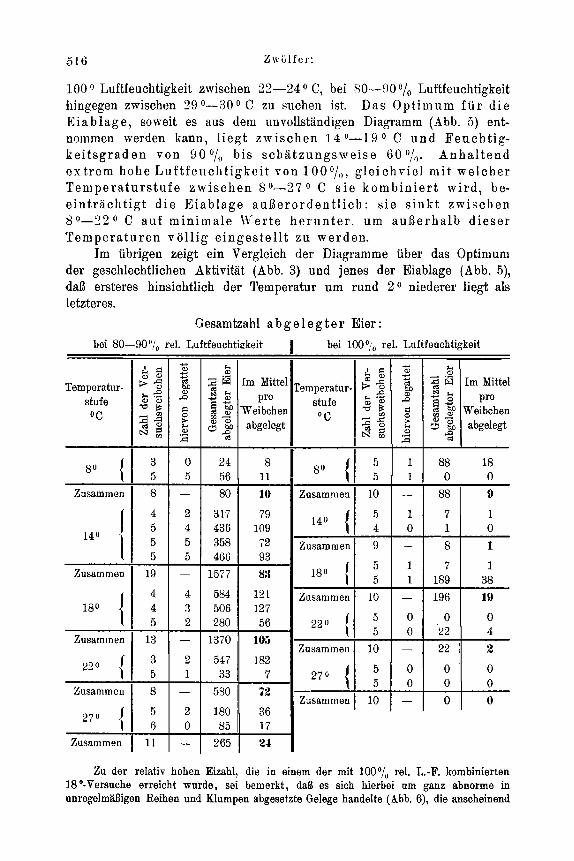
516 Zwolfer :
100 0 Luftfeuchtigkeit zwischen 22-240 C, bei SO-900/o Luftfeuchtigkeit hingegen zwischen 2 0 ° - 3 0 0 C zu suchen ist. Das Opt imum f u r d i e E iab lage , soweit es aus dem unvollstandigen Diagramm (Abb. 5 ) ent- nommen werden kann, l i eq t zwischen 14O--1990 C u n d Feucht ig- ke i t sg raden von 90 O / ( , bis s cha tzungswe i se GOO/, . Anha l t end e x t r e m hohe Luf t f euch t igke i t v o n l O O O / , , g l e i chv ie l mi t welcher Tempera tu r s tu fe zwischen So-27 0 C s i e kombin ie r t wird, be- e in t r ach t ig t d i e Eiab lage auf ie rordent l ich : s i e s ink t zwischen So-22 O C auf minimale TTerte h e r u n t e r . um auBerha lb d i e se r Tempera tu ren vo l l ig e inges t e l l t zu werden.
Im iibrigen zeigt ein Vergleich der Diagramme uber das Optimum der geschlechtlichen Aktivitat (Abb. 3) und jenes der Eiablage (Abb. 5), daB ersteres hinsichtlich der Temperatur urn rund 2 O niederer liegt als letzteres.
Gesamtzahl a b g e l eg t e r Eier : bei SO-90°/, rel. Luftfeuchtigkeit bei 100°;, rel. Luftfeuchtigkeit
Zusarnrnenl 10 1 - 1 196 I 19
Zu der relativ hohen Eizahl, die in einem der mit 100°/, rel. L.-F. kombinierten 18O-Versuche erreicht wurde, sei bomerkt, da13 es sich hierbei um ganz abnorme in unregelmafligen Reihen und Klumpen abgesetzte Gelege handelte (Abb. 6), die anscheinend

Studien zur dkologie und Epideniiologie der Insekten. 517
im Todeskampf von den betreffenden Weibchen abgestolen worden waren. Von diesen insgesamt 189 Eiern hatten sich - unter optimale Entwicklungsbedingungen verbracht - spaterhiu nur 6 Stuck = rund 30,',, bis zum Schliipfen der Jungraupen entwickelt. -
Der Berechnung der Durchschnittszahl abgelegter Eier pro Weibchen in obiger Tabelle und ebenso im Kurvenbild und Diagramm (Abb. 4 und Abb. 5) ist stets die G esam t zah 1 Weibchen der betreffenden Versuchsserie zugrunde gelegt worden. Bei Berucksichtigung ledigiich der begatteten Tiere hatten sich in einzelnen Trockenluft- versuchen hohere Werte fur die Durchschnittseizahlen ergeben (so bei 14O, 18O, 22 O), was eine geringe Verschiebung des Optimums gegen die hohere Temperatur bedeuten wurde. Von einer derartigen Bestimmung der Mittelwerte wurde indessen Abstand ge- nommen, da auch die nicht begatteten Weibchen an der Eiablage bis zu einem gewissen Grad beteiliat waren, der sich in den vorliegen- den Versuchen nicht einwaodfrei bestimmen lie& Bei einer etmaigen Nachprufung und Wieder- holung derselben empfiehlt es sich mit Versuchs. tieren zu arbciten, die ihre Kopula bereits voll- zogen haben.
Es darf aua dem Gesagten noch nicht gefolgert werden, da13 ein voriiber- gehende r Aufenthalt der Palter in un- giinstigen Witterungsverhaltnissen die Ei- ablage ganzlich verhindert. Die oben mitgeteilten Ergebnisse fuBen auf Ver- suchen mit kons tan ten Temperatur- und Luftfeuchtigkeitsverhaltnissen wahrend der gesamten Lebensdauer der Tiere! Ein Wechsel dieser Verhaltnisse fuhrt - wie das durch einen orientierten Ver- such belegt werden kann - zu anderen Resultaten : 5 Versuchspiirchen, die 25 Tage lang einer Temperatur von S o C verbunden mit 80-90 O/o Luftfeuchtigkeit ausgesetzt waren, und in dieser Zeit nur 2 Eier ablegten , setzten anschliellend
Tp Y. 30
tW 70 60 .50 % r!L.F
Abb 5. Diagramm der Gesamteiablage zur Bestimmnng ihres op~imalen Temperatur- und Lnftte~~chti..keitsbereiches. (Die den einzelnen Pnnkten zngeordneten Ziffern geben die im Dnrrhschnitt pro Versuchsweibchen wzhrend seiner gesamter. Lebenspuriodo abgelegten Et-
mengen an.)
unter optimale Bedingungen verbracht (18 0 C / S O "lo Luftfeuchtigkeit) in den folgenden 7 Tagen bis zu ihrern naturlichen Tode rund 340 Eier ab. Die Weibchen ver l ieren demnach i h r e Legefahigkei t t ro t z re la t iv 1 an ge n A u fen t h 1 t e s i n ung ii n s t i ge n B ed in g unge n k e in e s w egs.
Von den 5 Weibchen dieses Versuches erwiesen sich 4 als begattet. Pro Weibchen waren im Gesamtdurchschnitt 68 Eier abgelegt worden, was einer Eireduktion von rund SO0/, entspricht, die wohl auf die anfanglich wirkeuden ungunstigen Einflusse zuruck- geht. Von den Eiern entwickelten sich im ubrigen - unter gunstige Bedingungen verbracht - 83°/0 bis zum Schlupfen der Junglarven, 12O/, starben wahrend der Embryonalentwicklung ab, 5 erwiesen sich als ,,taubLL d. h. zeigten keinerlei Ver- farbung wie sie fur die Entwicklungsvorgange des Eies charakteristisch sind. Der 12tiigige Aufenthalt der Elterntiere bei 80/80-900/, Luftfeuchtigkeit hat demnach keine ausgesprochene Unterdruckung der Entwicklungsfahigkeit der Eier zur Folge, wenn die Versuchstiere anschlieWend in gunstige Bedingungen verbracht werden. -

518 Z w o l f e r :
I m Zusammenhang hiermit ist die Kenntnis der Abhangigkeit der mittleren Lebensdauer der Palter von Temperatur und Luftfeuchtigkeit praktisch von Bedeutung. Auf sie wird weiter unten noch zuriick- gekomtnen werden. -
Uber den ze i t l i chen Ablauf d e r Eiablage bei 80-900/, rel. Luftfeuchtigkeit in den verschiedeaen Temperaturstufen unterrichtet die
folgende Tabelle, die auf einigen der oben genannten taglich kontrollierten Ver- suche beruht:
(Siehe Tabelle auf S. 519.) Abgesehen von der extrem
niederen Temperaturstufe begann somit die Eiablage im groBen ganzen gleich- maBig am 2.-4. Tage nach Tersuchsbeginn, d. h. nach dem Schliipfen der Weib- chen. Die Praovipositions- periode weist also innerhalb eines vitalen Temperatur- bereiches keine deutliche Temperaturabhangigkeit auf. Es darf hieraus geschlossen werden, daB auch die Be- gattung in der Regel in den ersten 2-3 auf das Schlupfen der Weibchen folgenden Tagen stattfindet. In der Tat boten die vorliegenden Ter- suche dreimal Gelegenheit in der Zeit zwischen 18 bis 21 Uhr am 2.Tage nach Be- ginn der betreffenden Ver- suche das Zustandekommen der Kopula zu beobachten.
Die Partner sitzen dabei mit abgewandten oder gleichgerichteten Hopfen an einer Kiefernnadel, ahnlich wie dies bereits von anderen Beobachtern beschrieben worden ist.
I m Gegensatz &urn Beginn der Eiablage ist ihre Beendigung weit- gehend von der Temperatur der betreffenden Versuchsreihe abhangig : So erfolgte der AbschluB der Eiablage in der 140-Stufe nach dem 15. bezw. 17. Tage (vom Schlupfen der Weibchen an gerechnet) bei 18 0 nach 8 und
Abb. 6. Abnorme Eiablagen der Kieferneole bei feuchtigkeits- gesattigter Atmosphiire abgesetzt. (Vergr.)
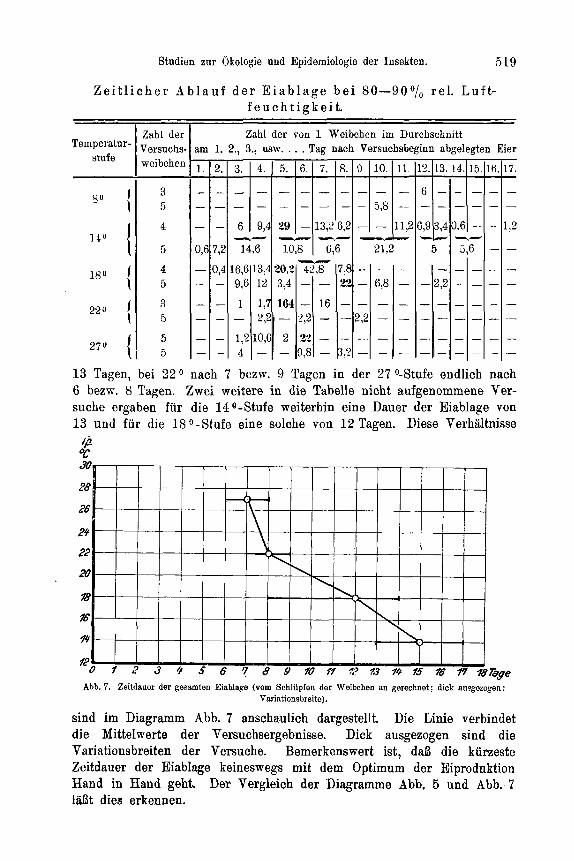
Studien zur Okologie und Epidemiologie der Insekten.
Z e i t l i c h e r A b l a u f d e r E i a b l a g e b e i SO-900/o re l . Luf t - f e u c h t i g k e i t .
519
Temperatur- stufe
80 {
18" {
27* {
1 4 0 {
22" I
Zahl der Jersuchs- weibchen
3 5
4
5
4 5
3 6
5 5
Zahl der von 1 Weibchen im Durchschnitt am 1. 2., 3., usw. . . . Tag nach Versuchsbeginn abgelegten Eier
0,6[7,21 14,6 I 10,6 I 6,6 I 21,2 1 0,4 -
- -
I - - I - -
- I - - I -
13 Tagen, bei 320 nach 7 bezw. 9 Tagen in der 27 O-Stufe endlich nach 6 bezw. 8 Tagen. Zwei weitere in die Tabelle nicht aufgenommene Yer- suche ergaben fur die 140-Stufe weiterhin eine Dauer der Eiablage von 13 und fur die 180-Stufe eine solche von 12 Tagen. Diese Verhaltnisse 4. Dc
Abb. 7. Zeitdaucr der geaarnten Eiablage (vom Schliipfen der Weibchen an gerechnet; dick awgozogen: Variationsbreito).
sind im Diagramm -4bb. 7 anschaulich dargestellt. Die Linie verbindet die Mittelwerte der Versuchsergsbnisse. Dick ausgezogen sind die Variationsbreiten der Versuche. Bemerkenswert ist, daB die kurzeste Zeitdauer der Eiablage keineswegs mit dem Optimum der Eiproduktion Hand in Hand geht. Der Vergleich der Diagramme Abb. 5 und Abb. 7 1aBt dies erkennen.

520 ZwGlfer:
I;;;a;Im;.l 1 , 1 1 1 I ?l Zahl d. Spermatopb. i n
1
Zabl der legereifen Eier in den Ovarien . . ca. 180 0 0 73
- 5 6
3 2
2 25
I) In der Tabelle durch Fettdruck gekennzeicbnet.

Studien zur <)kologie und Epidemiologie der Insekten. 52 1
gesunder Puppen zu einer Dwcbschnittseizahl, die erheblich unter der fiir die Kieferneule idealen liegt. D i e auf G r u n d se iner Angaben bes t immte E izah l r eduk t ion von 30°/, ist nach den im vorstehenden geschilderten Befunden uber den Kinflul3 von Temperatur und Luft- feuchtigkeit auf die Eiablage der Kieferneule im wesent l ichen a l s Folge d e r v o r u b e r g e h e n d ung i ins t igen Wet t e r l age im Mai 1930 anzusehen . I m einzelnen geht dies auch aus den entsprecbenden phanologiscben Kurven M e y e r s deatlich hervor.
Da13 die Methode des Eiauszahlens gefallter Probestiimme Unzulanglichkeiten besitzt, - worauf von anderen Autoren ofters hingewiesen wird - soll nicht bestritten werden. Berucksichtigt man iudessen die obigen Sektionsbefunde, denen Material aus Me y e r s Beobachtungsgebiet zugrunde liegt, und die seine Resultate bis zu einem gewissen Grad bestatigen, so ware es verfeh!t, die ,,30 prozent. Eizahlreduktion" allein durch die Mangel der Bestimmungsmethodik erklaren zu wollen.
In der Literatur liegen 'uislang nur sparliche Beobachtungen iiber den Einflul der Witterung auf Falterflug und Eiablage vor. R a t z e b u r g (52) bemerkt mit Bezug auf die Falter, da13 sie infolge ihres fruhzeitigen Fluges Jeicht von der Witterung leiden". Nach B u r o (zit. nach J u d e i c h - N i t s c h e ) soll Kalte und Schnee die Falter niassenhaft toten. Demgegenuber bemerkt E c k s t e i n (20), d a l die Falter ,,gegen Regen unempfindlichCc sind, der sie weder bei der Kopula noch bei der Eiablage stort. Ob diese SchluP- folgerung , die wie schon oben bemerkt, auf Freilandzwingerbeobachtungen ful3t, vzrall- gemeinert werden kann, mag dahin gestellt bleiben. M e y e r s exakte Freilandbeob- achtungen , die sich aufs beste mit den Resultaten der Laboratoriumwersuche decken, sind nicht mit ihr zu vereinen.
f ) Lebensdauer der Imago.
Im Zusammenhang rnit den erorterten Fragen ist - worauf oben schon hingewiesen wurde - die Kenntnis der Lebensdauer der Palter und ihrer Abhangigkeit yon Witterungseinflussen praktisch von Bedeutung.
Bereits aus E c k s t e i n s (20) Beobaohtungen geht hervor, d a l sie von der Temperatur des Aufenthaltsortes der Falter mitbestirnnit wird. So betragt sie nach Angaben dieses Autors im Januar im ungeheizten Raum beini Weibchen im Mittel 32 Tage, beim Mannohsn 28 Tage; im Februar wurden entsprechend 29 und 24 Tage, im Marz 28 und 24 festgestellt. S a c h t 1 e b e n (56) fa& seine Erfahrungen dahin zusammen, da13 d e r Falter ),im Walde bis zu 4 Wochen leben kann".
Genauere Anbaltspunkte uber die Lebensdauer der Falter und ibre Abhangigkeit von Temperatur und Luftfeuchtigkeitsverhaltnissen gab die taglicbe Kon trolle der im vorigen A bscbnitt besprochenen Versuche. Darnach wird die Lebensdauer der Palter sowohl von der Temperatur wie auch von der Luftfeuchtigkeit der Umgebung wesentlich beeinflul3t. Im b e s o n d e r e n bes i t z t e i n e kons tan t ex t r em hohe L u f t f e u c h t i g k e i t v o n 1000/, e r h e b l i c h l e b e n s v e r k u r z e n d e Wirkung. W i e schon d ie V e r m i n d e r u n g d e r P roduk t ion l ege re i f e r B ie r , wie j e n e d e r gesch lech t l i chen Ak t iv i t a t und d i e d e r E iab lage i n feucht igke i t s - g e s a t t i g t e r A t m o s p h i r e , so we i s t a u c h d i e se E r s c h e i n u n g auf d i e s c h a d ig e n d e n E i n f 1 u s s e e x t r e m h o h e r Lu E t f e 11 c h t i g k e i t s- g r a d e f u r das Imag ina l s t ad ium der K ie fe rneu le hin. In diesem
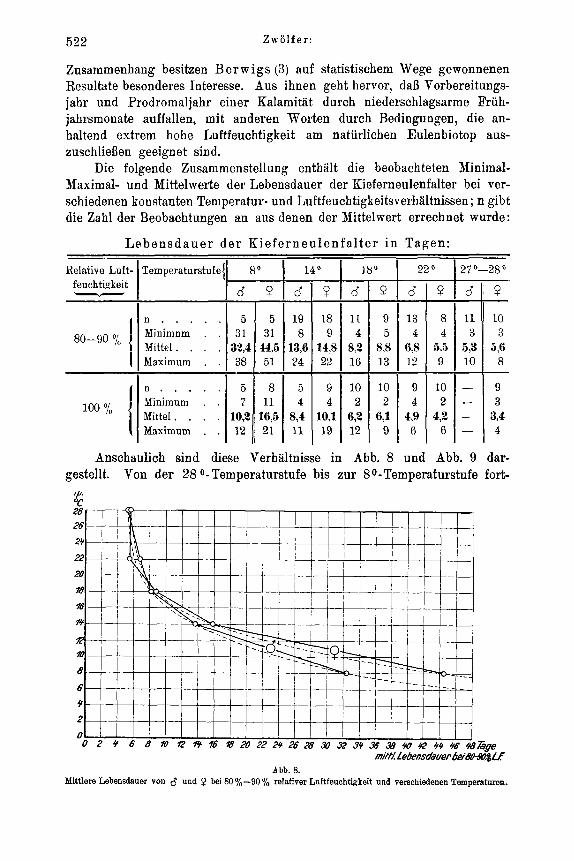
522 Zwol fer :
Zusammenhang besitzen B e r w ig s (3) auf statistischem Wege gewonnenen Resultate besonderes Interesse. Bus ihnen geht hervor, da13 Vorbereitungs- jahr und Prodromaljahr einer Kalamitat durch niederschlagsarme Fruh- jahrsmonate auffallen, mit anderen Worten durch Bedingungen, die an- haltend extrem hohe Luftfeuchtigkeit am naturlichen Eulenbiotop aus- zuschlieflen geeignet siad.
Die folgende Zusammenstellung enthalt die beobachteten Ninimal- Maximal- und Mittelwerte der Lebensdauer der Kieferneulenfalter bei ver- schiedenen konstanten Temperatur- und Luftfeuchtigkeitsverhaltnissen ; n gibt die Zahl der Beobachtungen an aus denen der Mittelwert errechnet wurde:
Lebensdaue r der K ie fe rneu len fa l t e r i n Tagen:
n . . . . . Xinimum . . Mittel. . . . i Maximum . .
100 :/,
P d Q d
18 11 9 13
14,s 8,2 ~ 8,s 6,s 22 16 13 12
9 10 10 9 4 2 2 4
10,l 6,2 G,1 4,9 19 12 9 6
9 4 5 5 4
Anschaulich sind diese Verhaltnisse in Abb. 8 und Abb. 9 dar- Von der 28 0- Temperaturstufe bis zur 80-Temperaturstufe fort- gestellt.
%
28 3 32 34 36 38 40 42 44 y6 Mgge r n j ~ L e b e n s d a u e e r b - ~ ~ L ~
A bb. 8. Mittlere Lebensdauer von 3 und P bei 80 %-90 yo relativer Luftfeuchtigkoit und verschiedenen Temperatmen.
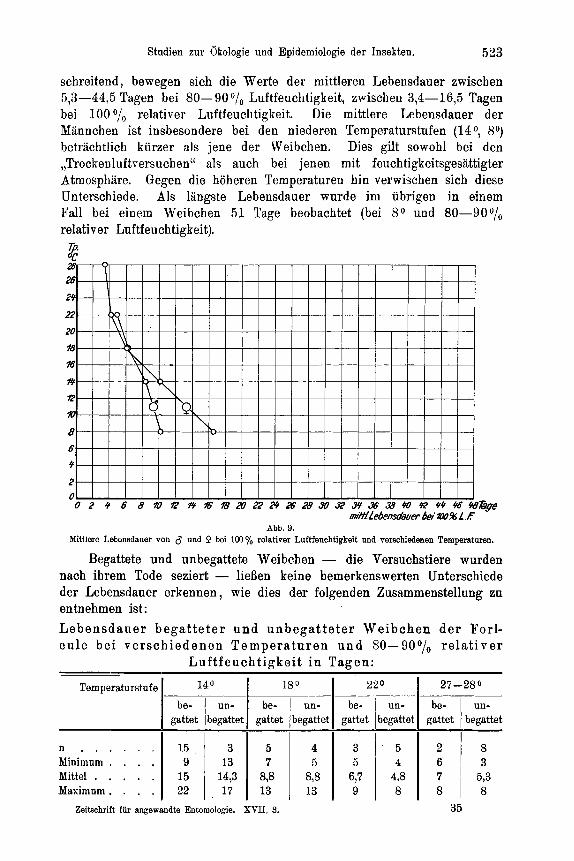
Studien zur Okologie und Epidemiologie der Insekteo. 523
n . . . . . . Minimum . . . . Mittel . . . . . Maximum. . . .
schreitend, bewegen sich die Werte der mittleren Lebensdauer zwischen 5,3-44,5 Tagen bei 80-900/o Luftfeuchtigkeit, zwischeu 3,4-16,5 Tagen bei 100 O/,, relativer Luftfeuchtigkeit. Die mittlere Lebensdauer der Mannchen ist insbesondere bei den niederen Temperaturstufen (140, 80) betrachtlich kiirzer als jene der Weibchen. Dies gilt sowohl bei den ,,Trockenluftversuchen" a h auch bei jenen mit feuchtigkeitsgesattigter Atmosphare. Gegen die hoheren Temperaturen hin verwischen sich diese Unterschiede. Als langste Lebensdauer wurde im ubrigen in eiriem Fall bei einem Weibchen 51 Tage beobachtet (bei 8 0 und 80-900/, relativer Luftfeuchtigkeit).
5 4 15 9 13 7 5
15 14,3 8,8 8,8 22 17 13 13
1 3
Begattete und unbegattete Weibchen - die Versuchstiere wurden nach ihrem Tode seziert - liel3en keine bemerkenswerten Unterschiede der Lebensdauer erkennen , wie dies der folgenden Zusammenstellung zu entnehmen ist:
915 5 4 6,7 I 4.8 9 8
Lebensdaue r bega t t e t e r und unbega t t e u l e be i verschiedenen Tempera tu ren
Luf t feucht igke i t i n ~ __
Temueraturstufel 14' I 18"
2 8 6 3 7 593 8 8
t e r Weibchen d e r Forl- u n d SO-900/0 r e l a t i v e r 'agen:
_ _ _ _ ~ ~
220 I 27-280
Zeitschrift fur angewandte Entomologie. XVII, 3. 35

524 Zwolfer :
Was im iibrigen die Lebensdauerwerte der begatteten n'eibchen dieser Tabelle anbetrifft, so sei in diesem Zusammenhang daran erinnert, daB diese Tiere wahrend der Versuchsdauer ihr Fortpflanzungsgeschaft ausubten.
Das Diagramm Abb. 10 bildet einen Versuch, das Optimum der mitt- leren Lebensdauer der Falter festzustellen. Es wurde danach etwa zwischen 40-80 C und bei Luftfeuchtigkeitswerten kleiner als liegen. Ein Vergleich der Abbildungen 3, 5 und 10 zeigt deutlich, da13 das Optimum der mittleren Lebensdauer keineswegs mit jenem der geschlechtlichen Aktivitat und jenem der Eiablage zusammeufdlt.
Bode n heim e r (9,ll-13) gibt fur den Begriff des vitalen Optimums zwei Definitionrn. Einerseits wird es durch diejenige TemperatwLuftfeuchtigkeitskom bination gekennzeichnet
T 3-
28
a( a-
- p om
n(#' 0x5
bei der die geringste Sterblichkeit auftritt, mit anderen Worten der groBte Prozentsatz von Individuen der Eot- wicklungsstufe eines Insektes die nachstfolgende er- reicht. Seine zweite Definition besagt, da13 das vitale Optimum such durch jane Temperatur-Luftfeuchtigkeits- kombination reprasentiert wird, in der die Mehrzahl der Tiere am langsten iiberlebt, oder was dasselbe besagt, die langste mittlere Lebensdauer einer bestimmten Rnb wicklungstufe vorliegt.
Betrschtet man als die Uberlebenden der Imaginal- generation die Eier eines Insektes - sie sind die auf die Imago folgende Entwicklungsstufe - so ware nach der ersten Definition das Optimum der Eiablage als das vitale Optimum der Falter anzusprechen. Nach der zweiten Definition hingegen ware letzteres identisch mit der h g s t e n mittleren Lebensdauer. Beide Optima- Werte brauohen jedoch, wie die vorstehend mitgeteilten Be- obachtuI;gen zeigen, keineswegs zusammenzufallen. Ge- legentlich und zwar bei den Latenzstadien eines Insektes wird dies der Fall sein (vgl. das fur O ~ J C U T ~ U S hyalzai- pennis Costa von Bodenheimer gegebene Diagramm !) Immerhin gestattet dieser Sonderfall nicht beide Defi- nitionen auf ein und denselben Begriff anzuwenden. Y i t Rucks ich t auf diest, Uber legung wird i n d e r vorl iegenden Arbei t der Begriff d e s v i ta len
# -
e-
6 -
r - 2 -
0
Opt imums n n r in d e r e r s t e n d e r beiden-obigen von Bodenheimer gegebenen Definition en ge br a u c h t.
Neuerdings vertritt J a n i s c h (36) die Auffassucg. daB es fur jede Art ein ,,absolutes OptimumL' gebe, welches ihr vitales und ihr Entwicklungsoptimum u m f d t - zwei Begriffe, die von anderen Autoren scharf getrennt werden. ,,Auf den gauzen Individual- cyklus bezogen" - sagt J a n i s c h - ,,sind beide identisch". Bei einigen Arten mogen sie in der Tat zusammenfallen. Die Regel ist es indes nicht. Dies geht schon aus der hinliinglich bekannten Tatsache hervor, daS fur verschiedene Entwicklungsstufen ein und derselben Art das vitale Optimum auBerordentlich variieren kann : Die uberwinternde Forleulenpllppe beispielsweise verlangt, wie spater gezeigt werden wird, grundsatzlich andere Temperatur- und Feuchtigkeitsbedingungen als das Ei-, Larven- oder Imaginal- stadium dieser Art. Fur die Forleule ein ,,absolutes Optimum" im obigen Sinn aufzustellen, erscheint schon aus diesem Grunde unmoglich. -
Von den Werten der mittleren Lebensdauer der Kieferneulenfalter sind jene praktisch von besonderer Bedeutung, die angeben wie lange
*(&
:G I I I I

Studien zur Okologie und Epidemiologie der Insekten. 525
ein Falter unter niederen Temperaturen verbunden mit trockener oder extrem feuchter Luft leben kann. Ihre Kenntnis gibt gewisse Anhalts- punkte zur Beurteilung, inwieweit ungunstige Witterungverhaltnjsse die Eiablage der Falter zu beeintrachtigen vermogen. Danach miiljte .- wenn die in Abb. 8 und 9 niedergelegten Daten zum Vergleich herangezogen werden - u n m i t t e l b a r im AnschluB an d a s Schl i ipfen d e r Haupt - masse d e r Fa l t e r e i n e t rockene Wi t t e rungspe r iode m i t anha l t end n i ede ren Tempera tu ren ( u n t e r 8 0 C) Ton mindes tens 3-4 wochent- l i che r Dauer e inse tzen , wenn d iese e i n e n groBeren Einf luB auf d e n Gang d e r E iab lage gewinnen soll. Regenwet t e r ve rbunden mi t n i ede ren Tempera tu ren mu5 te demen t sp rechend m i n d e s t e n s 2-3 W o c h en u n un t e r bro c h en a n ha1 ten, u m e r he blic he W i r k ung i n d i e se r R i c h t u n g z u zeigen.
Wurde das Schliipfen der Falter in freier Natur gleichmHBig erfolgen und sich auf eine kurze Zeitspanne von wenigen Tagen beschranken, so ware die Gefahr einer Ausschaltung des groBten Teiles der Eulenpopulation vom Fortpflanzungsgeschaft durch ungunstige Witterungsverhaltnisse wesent- lich pro5er als bei einer Verzettelung des Scblupfens uber einen langeren Zeitraum. Letzteres wird wie schon fruher erwahnt wurde, bei der Kiefern- eule als Regel beobachtet. Unter obigen Gesichtspunkten betrachtet, stellt es gleichsam eine Sicherung vor, die selbst in ganz ungunstigen Jahrgangen zum mindesteu einem Teile der Population noch die Ausiibung seines Fort- pflanzungsgeschaftes ermoglichen wird. Es tragt damit zur Sicherung des Bestandes der Art bei.
111. Die Vitalitlt der prtlimaginalen Entwicklungsstufen in ihrer Abhlngigkeit yon Temperstur und Luftfeuchtigkeit.
Wohl noch grol3ere Bedeutung fur die Verminderung der Nachkommen- schaft einer Eulenpopulation als durch die Reduktion der Eierzeugung besitzen die klimatischen Faktoren durch die direkte Vernichtung eines Teiles der gezeugten Nachkommenschaft. DaB eine derartige direkte Wirkung tatsachlich stattfinden kann, geht aus den Resultaten hervor, die in den folgenden Ausfuhrungen niedergelegt sind. Ihnen liegt die Untersuchung von rund 12000 Eiern, 6600 Eiraupen und ca. 1000 iilteren Raupen uud Puppen der Kieferneule auf ibre Lebensfahigkeit unter ver- schiedenen Temperatur-Luftfeuchtigkeitskombinationen zugrunde.
a) Spezielles zur Methode. Das Grundsatzliche uber die Erhaltung konstanter Temperatur- und Luftfeuchtigkeits-
verhaltnisse, wie sie in den Versuchen angestrebt wurde, uber die in diesem Abschnitt berichtet wird, ist bereits im allgemeinen methodischen Teil mitgeteilt. Es bedarf hier noch einiger Worte uber die technische Durchfuhrung dieser Versnche.
Bei den ersten Experimenten mit der Eizucht unter konstanten Luftfeuchtigkeits- verhaltnissen fanden kleine Glastuben von 1 x 6,5 cm InnenmaB Verwendung. Der untere
35 *

526 Zwolf e r :
Tubenteil enthielt das angefeuchtete Salz und war vom ,,Zuchtraum" durch einen Watte- bausch getrenut. Nach au len ist derselbe im ubrigeu durch einen Korkstopfen abgeschlossen. Die Methode zeigte indessen bei leicht zerflieSlichen Salzen, wie LiCI, CaCI,, ZnCl,, P,05, zahlreiche Mangel. Vor allem storte das Bescbmutzen der Watte durch das zerflielende Salz. Auch wurde letzteres gelegentlich durch Kapillarwirkung der Watte bis in den Zuchtraum hinaufgesogen. Besser bewahrten sich die spaterhin verwandten Glastuben wie Abb. 11 eine solche im Querschnitt in natiirlicher Gro8e darstellt. Zuchtraum (Z) und Salzraum (S) sind hier durch ein Stuck Glasbatist (B) getrennt, welches vermittelst eines dicken Crummiringes (G) an den inneren Wandungen der Glastube (T) festgespannt ist.
Was die Versuchsdauer speziell bei den Eiexperi- menten angeht, SO wurde sie stets 8-10 Tage uber das Schlupfen des letzten Eies fortgesetzt urn Fehlresultate durch etwa verspatetes Schlupfen einzelner Eier zu ver- meiden. Als weitere allerdings nur geringfugige Fehler- quelle komm t Kannibalismus der Junglarven insbesondere in den Trockenluftversuchen in Prage: Da bei diesen Versuchen den schliipfenden Larven kein Futter zur Verfugung stand, fraBen sie zunachst die leeren Ei- schalen, vorgriffen sich dabei aber auch noch gelegent- lich an noch nicht geschliipften Geschwistereiern, deren Embryo sie ausfralen. Um auch diese Fehlerquelle moglichst auszuschalten, wurden bei der taglichen Knn- trolle des Schlupfens die jeweils ausgekommenen Larven entfernt. -
Abb. 11. Tubenfhniger Hygrostat. Abb. 13. (uuetschnitt in natiirlicher G-r6lle ; Schaleuftirmiger Eygrostat. (Querschnitt 2/s natiirlicher GrtJBe;
Erklarung im Text.) Erklsrung im Text.j
Obschon fur die Aufzucht von Eiern brauchbar, erwies sich die oben geschilderte Art der Hggrostaten f u r die Aufzucht von Eiraupen ganzlich ungeeignet: Die Reinigung der Gefale vom anfallenden Raupenkot ist schwierig und aulerordentlich zeitraubend. Zudem haben die ausgesprochen positiv photophilen, anf die scbwachsten Lichtreize reagiorenden Jungraupen die Gewohnheit, den hellsten Stellen des ZuchtgefaBes zuzustreben, die in diesem Falle im oberen Tubenteil liegen. So blieb vielfach das E'utter, welches sich im unteren Teil des Zuchtraums befand, vollig unberiicksichtigt. Im ubrigen wird der Luft- feuchtigkeitsausgleich bei der zum engen Innendurchmesser relativ grolen Hohe des Zucht- raums erschwert, wenn Futterstucke, die im unteren Zuchtraumteil liegen, das Batiststiick verschlieBen
Unter Beriicksichtigung dieser nberlegungen wurde versucht, die angedeuteten Mangel mit Hilfe flacher Doppelglasschalen (Petrischalen) von 10 cm Durchmesser und 2 om Hohe abzustellen. Abb. 12 gibt einen Querschnitt durch einen fertig montierten Hygrostaten dieser Art: Der Deckel (PI) der Petrischale dient zur Aufnahme des an- gefeuchteten Salzes (S). Der Zuchtraum (Z), dor von der kleineren Schalenhalfte (P,)

Studien zur Okologie und Epidemiologie der Insekten. 527
gebildet wird, ist vom Salzraum durch ein Stuck Glasbatist (B) getrennt, welches mit Hilfe eines Leukoplaststreifens (L.) an den AuPenrand der Salzschale festgeklebt wird. Dm bei gewissen leicht zerfliel3lichen Salzen wahrend des Hantierens mit dem Gerat ein Ver- schmutzen des Zuchtraums zu vermeiden, befinden sich unterhalb des angefeuchteten Salzes mehrere Lagen Filtrierpapier (F.), wvolches alle zerfliePenden Bestandteile aufsaugt. Eine kleine Modifikation dieses Hygrostaten, die spaterhin vorgenonimeu wurde, erwies sich als zweckmaQig: Bei ihr ist das Batiststuck nicht direkt an den Salzschalenrand fest- gekittet, sondern an einem genau uber diesen passenden Kartonring, den man sich aus mehreren Streifen steifen Papieres leicht zusammenkleben kann. Der Hygrostat dieser Art besteht somit aus drei losen Teilen: Zuchtschale, Batistsieb und Salzschale, die wie oben beschriebcn zusammengesetzt sind und wahrend der Versuchsdauer durch ein kraftiges Gummiband zusammengehalten werden. Der Vorteil dieser Anderung der ursprunglichen Form bestett in der Yoglichkeit, das Salz in der Salzschale jederzeit leicht erneuern, bezw. auswechseln zu konnen.
Die niedrige Form der Zucht- und Salzschale bei dieser Art der Hygrostaten be- wirkt einen relativ raschen Ausgleich von Luftfeuchtigkeitsunterschieden , wie sie beim Offnen der GefaPe wahrend des Futterns usw. leicht entstehen. Durch den seitlich etwas ubergreifenden Batistrand der Salzschale ist gleichzeitig eine gewisse Durch- luftungsmoglichkeit des Zuchtraunis geboten. Da von auPen eindringende Luft erst uber dio feuchte Salzmasse streichen muP, ehe sie in den Zuohtraum gelangt, ist hierdnrch eine Storung der Feuchtigkeitskonstmz in diesem ausgeschaltet. Das Einbringen von Futter, dau in kleinen Gaben gersicht wird, hat - wie mehrere Kontrollmessnngen er- gaben - keine bemerkenswerte Storung der im Zuchtraum vorhandenen Luftfeuchtigkeit zur Folge. Alles in allem erwies sich der Apparat fur die Aufzucht junger Kieferneulen- larven durchaus brauchbar.
Eine gewisse Schwierigkeit bot die Beschaffung geeigneten Futters fur die Eiraupen: Bekanntlich nehmen diese in der Natur in der Regel die knospenden hfaitriebe der Kiefer als Nahrung an. Vorjahrige Nadeln werden verschmaht. Zu Beginn der Versuche be- fanden sich indessen die Knospen noch auf keinem soweit fortgeschritttenen Entwicklungs- zustand, daS sie den Jungraupen direkt als Nahrung hatten gereicht werden konnen. Aufschub des Versuchsbeginnes auf einen entsprechend spateren Termin, hatte zur 'ijber- haufung mit laufenden Versuchen gefdhrt , die im Interesse einer punktlichen Kontrolle vermieden werden m d t e . Um diese Schwierigkeit zu beheben, wnrden die Knospen in noch unentwickeltem Zustand gereicht, nachdem sie zuvor durch Abschaben von ihren harzigen Deckschuppen befreit Torden waren. Die Junglarven nehmen so vorbereitetes Futter anstandslos an, und gediehen dabei auflerlich ebenso gut als bei den spateren Ver- suchen, zu denen normal ausgetriebene Kiefernknospen als Futter Verwendung fanden. Im ubrigen wnrde das Futter taglich einmal frisch gereicht.
Als Merkmal zur Feststelluug, ob in den einzelnen Versuchen eine Larve die niiohst- folgende Entwicklungsstufe erreichte, diente deren KopfkapselgroPe. Bei einiger 'ijbung ist meist ohne besondere Messung die Unterscheidung der Stadien auf diesem Wege moglich. Da in der Literatur noch keine genaueren Angaben iiber die Kopfkapselgrofle der verschiedenen Larvenstufen von P. llammea vorliegen, seien im folgenden die eigenen Untersuchungsbefande mitgeteilt:
Was die Kopfkapselbreite, angeht, so kann sie fur jedes einzelne Stadium variieren. Besonders zwischen dem 3. und 4. Stadium (Zwei- und Dreihanter) treten auf der einen Seite gelegentlich Maximal-, anf der anderen Seite MinimalgroSen auf, die fast ineinander ubergehen und eine sichere Unterscheidung dieser Entwicklungsstufen zuweilen erschweren. Genauere Messungsergebnisse unter Beriicksichtigung der Variationsbreite sind in der folgenden Zusammenstellung enthalten :
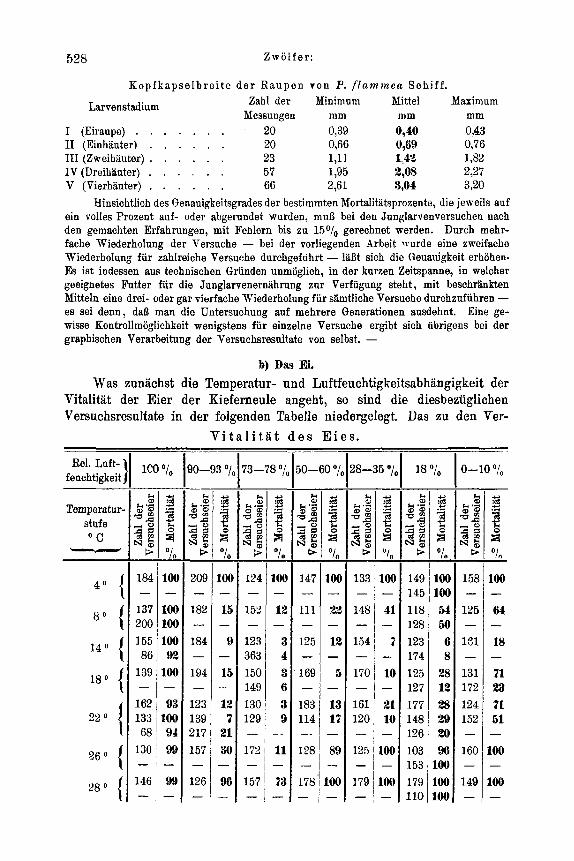
528 Zwolfer:
c
g.8 u’a”
N $ + 184
137 200 155 86
139
;;
- -
K o p f k a p s e l b r e i t e d e r Raupen r o n P. f l a m m e a Schiff .
a s
3
“ in
1OU
100 100 100 92 100
.-
g
- -
Larvenstadium Zahl der Minimum Mittel Maximum Messnngen mm mm mm
I (Eiraupe) . . . . . . . 20 0,39 0,40 0,43 I1 (Einhauter) . . . . . . 20 0,66 0,69 0,76 111 (Zweihauter) . . . . . . 23 1,l I 1 ,4t 1,82 IV(Dreibauter) . . . . . . 57 1,95 2,08 2,27 V (Vierhauter) . . . . . . 66 2,61 3,01 3,20
Hinsichtlich des Genauigkeitsgrades der bestimmten Mortalitatsprozente, die jeweils auf ein volles Prozent auf- oder abgerundet wurden. mu13 bei den Junglarvenversuchen nach den gemachten Erfahrungen, mit Fehlern bis zu 1.5% gerechnet werden. Durch mehr- fache Wiederholung der Versuche - bei der vorliegenden Arbeit w r d e eine zweifache Wiederholung fur zahlreiche Versuche durchgefuhrt - 1a13t sich die Geuauigkeit erhohen. Es ist indessen aus technischen Griinden unmoglich, in der kurzen Zeitspanne, in we!oher geeignetes Futter fur die Junglarvenernahrung zur Verfiigung steht , mit beschrankten Mitteln eine drei- oder gar viwrfache Wiederholung fur samtliche Versuche durchzufuhren - es sei denn, da13 man die Untersuchung auf mehrere Generationen ausdehnt. Eine ge- wisse Eontrollmoglichkeit wenigstens fur einzelne Versuche ergibt sich iibrigens bei der graphischen Verarbeitung der Versuchsresultate von selbst. -
b) Das Ei. Was zunachst die Temperatur- und Luftfeuchtigkeitsabhangigkeit der
Vitalitat der Eier der Kieferneule angeht, so sind die diesbezuglichen Versuchsresultate in der folgenden Tabelle niedergelegt. Das zu den Ver-
V i t a l i t a t d e s E i e s .
Rel. Luft- fenchtigkeit
Temperatur, stufe OC - 8 o i
14O { 18O { 220 { 26O { 28O {
1co 01,
~-
73-78 ‘I,, 50-60 ‘1, I - c
8 .f
G g - 2
+ 124
- c s t -
- -
152
123 363 150 140 130 129
172
157
-
-
-
-
- 18-35 ‘1, 18 “0
- F
5 ’f - 5 st15
b
149 145 118 128 123 174 125 127 177 148 126 103 153 179 110
-
- P c 2
‘ I n
lO(1 lO(1 54 5a 6 8 28 12 28 29 20 96
LOO 100 LOO
-
___.
0-10
c al 2 ’8 =s - 0 d l
P
158
125
lei
131 172 124 152
160
149
G g - -
-
-
-
-
-
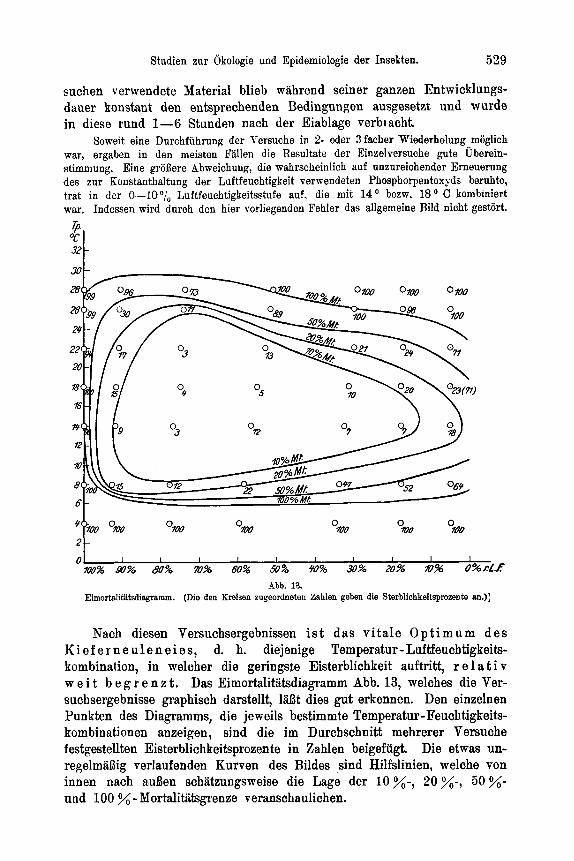
Studien zur Okologie und EpidemioIogie der Insekten. 529
suchen verwendete Material blieb wahrend seiner ganzen Entwicklungs- dauer konstart den entsprechenden Bedingungen ausgesetzt und wurde in diese rund 1-6 Stunden nach der Eiablage verbiacht.
Soweit eine Durchfiihrung der Tersuche in 2- oder 3 facher Wiederholung moglich war, ergaben in den meisten Fallen die Resultate der Einxelversuche gute Uberein- stimmung. Eine grol3ere Abweichung, die wahrscheinlich auf unzureichender Erneuerung des zur Eonstanthaltung der Luftfeuchtigkeit verwendeten Phosphorpentoxyds beruhte, trat in der O--lOo/, Luftfeuchtigkeitsetufe auf, die mit 14 O bezw. 18 O C kombiniert war. Indessen wird durch den hier vorliegenden Fehler das allgemeine Bild nicht gestort.
.R Abb. 1%
Eimortalitiitsdiagramm. (Die den Kreisen zugeordneten Zahlen geben die Sterblichkeitsprozente an.):
Nach diesen Versucbsergebnissen ist das v i ta le O p t i m u m d e s Kie f e r n e u l e n e i e s , d. h. diejenige Temperatur-Luftfeuchtigkeits- kombination, in welcher die geringste Eisterblichkeit auftritt, r e I a t i v w e i t b e g r e n z t. Das Eimortalitatsdiagramm Abb. 13, welches die Ver- suchsergebnisse graphisch darstellt, 1aBt dies gut erkennen. Den einaelnen Punkten des Diagramms, die jeweils bestimmte Temperatur-Peuchtigkeits- kombinationen anzeigen, sind die im Durcbschnitt mehrerer Versuche festgestellten Eisterblichkeitsprozente in Zahlen beigefugt. Die etwas un- regelmal3ig verlaufenden Kurven des Bildes sind Hilfslinien, welche von innen nach auBen schatzungsweise die Lage der lo%-, 20%-, 50%- und 100 % - Mortalitatsgrenze veranschaulichen.

530 Zwolfer:
Darnach liegt das r i t a l e O p t i m u m f u r d a s E u l e n e i zwischen 120-220 C u n d z w i s c h e n L u f t f e u c h t i g k e i t s g r a d e n v o n 65 %--85 %. Es umfafit also eine ziemlich weite Temperatiir und Luft- feuchtigkeitszone. Hierin kornmt die relativ grofie Widerstandsfahigkeit des Euleneies gegen klimatische Einflusse zum Ausdruck. (Die in Abb. 13 in diesem Bereich angegebenen Mortalitatsprozente von 3 und 4 sind auf unvermeidliche Versuchsfehler zuriickzufiihren, wie sie durch unbefruchtete Eier entstehen.)
A b s o l u t t o d l i c h w i r k t d a u e r n d e r A u f e n t h a l t d e r E i e r i n T e m p e r a t u r e n u n t e r h a l b 6O C, g l e i c h v i e l m i t w e l c h e r L u f t f e u c h t i g k e i t s i e k o m b i n i e r t w e r d e n . D i e o b e r e t 6 d l i c h e T e m p e r a t u r g r e n z e 1 i e g t f i i r L u f t f e u c h t i g k e i t s- w e r t e v o n 60-100% b e i e t w a 29-300 C. I n V e r b i n d u n g m i t g e r i n g e r e n L u f t f e u c h t i g k e i t s g r a d e n s i n k t s i e a u f r u n d 240 C h e r a b . Letzteres hangt mit der grofieren Verdunstungs- kraft zusammen, welche die entsprechenden Temperatur - Feuchtigkeits- kombinationen besitzen. Innerhalb eines v i t a 1 e n Temperaturbereiches ist im ubrigen die verhaltnismaBig geringe Sterblichkeit bemerkenswert, welche die Eier bei niederen Luftfeuchtigkeitsgraden von 20 - 60 % zeigen. Es erhellt hieraus, daB die Eier der Kieferneule gegen iiber- mal3ige Verdunstung im grol3en ganzen gut geschutzt sind, was wiederum mit ihrer relativ derben Eischale zusammenhangen durfte.
Die Eier des Maisziinslers beispielsweise, die, wie bei friiherer Gelegenheit (75) ge- zeigt werden konnte, unter siiddeutschen Verhaltnissen im Freien eine abiotische Mortalitat von rund 50 yo aufweisen, besitzen vergleichsweise eine aulerst diinne Schale.
Auffallend ist demgegenuber d i e g r o B e E i s t e r b l i c h k e i t v o n f a s t d u r c h w e g 90-100%, d i e i n g e s a t t i g t e r L u f t f e u c h t i g - k e i t (100%) b e i a l l e n T e m p e r a t u r e n f e s t g e s t e l l t w e r d e n k o n n t e. Diese Empfindlichkeit der Euleneier gegen extrem hohe Luftfeuchtigkeit macht das friiher erwahnte Verhalten der weiblichen Falter in feuchtigkeitsgesattigter Atmosphare verstandlich, bei welcher die Eiablage auf ein Minimum reduziert wird. Biologisch kommt darin das Bestreben der Weibchen zum Ausdruck, ihre Nachkommenschaft nicht unmittelbar schadlichen Umweltseinflussen auszusetzen.
Bus dem Gesagten darf noch nicht geschlossen werden, fdalj voruber- gehende k u r z e Regenperioden im natiirlichen Biotop der Kieferneule wahrend der Entwicklungsxeit der Eier einen hohen Prozentsatz derselben vernichten. lj Die oben mitgeteilten Ergebnisse bezieben sich auf das Verhalten der Eier bei d a u e r n d e m Aufenthalt wahrend ihrer gesamten Entwicklung in feuchtigkeitsgesiittigter Atmosphare Vorubergehender Aufenthalt hat k e i n e s w dieselbe groBe Sterblichkeit zur Rolge. I m
I) Einen ahnlichen Gedanken veitritt Janisch (35). Auch Blunck (6) weist auf diesen Gesichtspunkt hin.
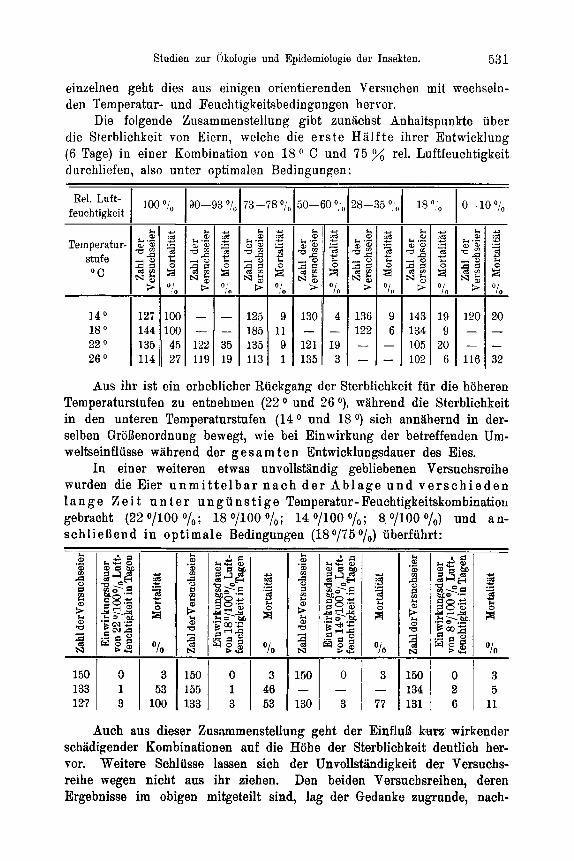
9 Z 0
EP 9P E
E I 0
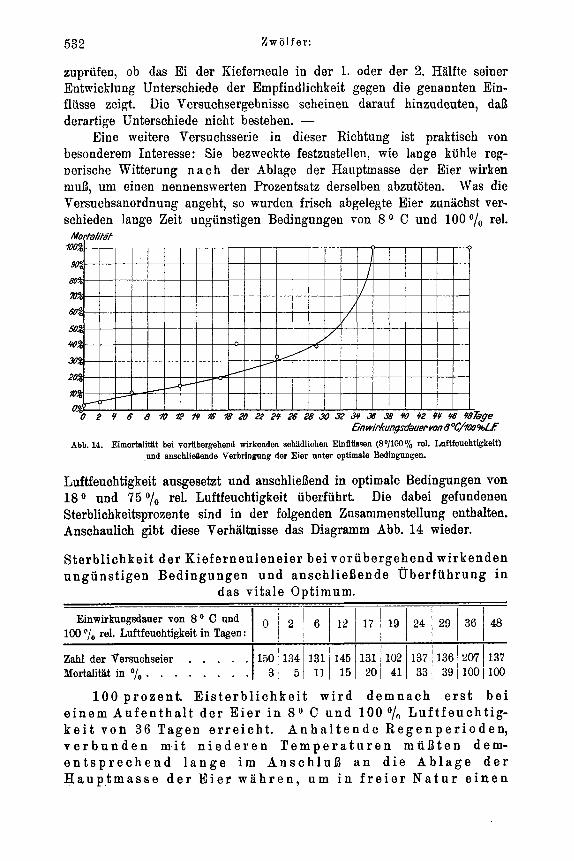
532 Zwol fe r :
zuprufen, ob das Ei der Kieferneule in der 1. oder der 2. Halfte seiner Entwicklung Unterschiede der Empfindlichkeit gegen die genannten Ein- flusse zeigt. Die Versuchsergebnisse scheinen darauf hinzudeuten, daB derartige Unterschiede nicht bestehen. -
Eine weitere Versuchsserie in dieser Richtung ist praktisch von besonderem Interesse: Sie bezweckte festzustellen, wie lange kuhle reg- aerische Witterung n a c h der Ablage der Hauptmasse der Eier wirken mull, urn einen nennenswerten Prozen tsatz derselben abzutoten. Was die Versuchsanordnung angeht, so wurden frisch abgelegte Eier zunachst ver- schieden lange Zeit ungunstigen Bedingungen von 80 C und 1000/o rel.
Abb. 14. Eimortalitiit bei voriibergehend wirkenden schildlichen Einfliissen ( S ~ / l O O ~ O rel. Lnftfenchtigkeit) nnd anschliebnde Verbringong der Eier nnter optimale Bedingungen.
Luftfeuchtigkeit ausgesetzt und anschliefiend in optimale Bedingungen von 18 0 und 76 rel. Luftfeuchtigkeit iiberfuhrt. Die dabei gefundenen Sterblichkeitsprozente sind in der folgenden Busammenstellung enthalten. Anschaulich gibt diese Verhaltnisse das Diagramm Abb. 14 wieder.
S t e rb l i chke i t d e r K ie fe rneu lene ie r be i vorubergehend wirkenden unguns t igen Bed ingungen und anschl ieBende Uberf i ihrung i n
das vi ta le Optimum.
Einwirkungsdauer von 8 C und 100 @/o rel. Luftfeuchtigkeit in Tagen :
Zahl der V'ersuchseier . . . . . 150 134 131 I 145 131 102 137 j 136 207 137 Mortalittit in o/o . . . . . . . .I 3 I 5 1 11 1 15 I 20 1 41 1 33 1 39 I l O O I 100
1 0 0 p r o z e n t . E i s t e r b l i c h k e i t w i r d d e m n a c h e r s t b e i e i n e m A u f e n t h a l t d e r E i e r i n 80 C u n d 100 O/,, L u f t f e u c h t i g - k e i t v o n 3 6 T a g e n e r r e i c h t . d n h a l t e n d e R e g e n p e r i o d e n , v e r b u n d e n m i t n i e d e r e n T e m p e r a t u r e n m u f i t e n d e m - e n t s p r e c h e n d l a n g e i m A n s c h l u B a n d i e A b l a g e d e r H a u p t m a s s e d e r E i e r w a h r e n , u m i n f r e i e r N a t u r e i n e n
I I I

Studien zur Okologie und Epiderniologie der Insekten. 533
h o h e n P r o z e n t s a t z d e r s e l b e n z u v e r n i c h t e n . Unterunseren Breiten sind solche anhaltend ungunstigen Wetterlagen wahrend der Ent- wicklungszeit der Eier (Mai) wohl kaum moglich oder doch auBerordent- lich selten. Kurzere Dauer ungunstiger Einflusse der genannten Art haben aber bereits einen erheblichen Ruckgang der Eisterblichkeit zur Folge. So wurden beispielsweise 1 7 Tage anhaltendes kaltes Regenwetter rund 20 der Eier vernichten und eine 6 tagige Periode dieser Art etwa 8-10 Ole. Weitere Werte konnen naherungsweise durch graphische Interpolation aus Abb. 14 entnommen werden. Sie gibt fur die Praxis Anhaltspunkte, um im einzelnen Fall die Wirkung einer derartig ungunstigen Witterungs- periode auf die Hohe der Eisterblichkeit schatzungsweise zu ermitteln.
Zusainmenfassend laBt sich iiber die Abhangigkeit der Lebensfahigkeit des Euleneies von klimatischeri Einflussen auf Grund der Versuche fest- stellen, da13 dieses gegen Witterungseinfliisse im grol3en und ganzen recht widerstandsfahig ist. Selbst gro13e Schwankungen der Wetterlage werden von der Mehrzahl der Eier ohne Schaden uberstanden werden. Emp- findlichkeit besteht in der Hauptsache gegen hohe Luftfeuchtigkeit, wie sie bei Regenwetter gegeben ist. Doch mussen kalte Regenperioden un- unterbrochen iiber einen Monat wahren, urn eine erhebliche Eisterblich- keit zur Folge zu haben.
In der Tat liegen auch in der Literatur keinerlei Angaben uber Freilandbeobachtungen vor, denen eine grolle Sterblichkeit der Euleneier durch abiotische Faktoren zu entnehmen ware. Sacht leben (55) stellte bei- spielsweise 1925 bei seinen Untersuchungen’ eine nichtparasitiire Ei- sterblichkeit von 200/, fest. - Meyer (46) fand 1930 im Forstamt Heideck neben einer 0-2,6 prozent. Eiparasitierung ein A bsterben der Eier durch anderweitige Ursachen in Hiihe von 1,4-7,4°/0. - Ton rund 5000 Eulen- eiern aus dem Forstamt Schwabach (Mittelfranken), die Ende Mai 1930 untersucht wurden, waren 2,8 O l 0 durch Parasiten vernichtet, 0,4 O/o er- wiesen sich als taub, wahrend 2,3% im Laufe ihrer Entwicklung ab- gestorben waren. Letateres wohl infolge klimatischer Einflusse. -
0) Larve I. Bezuglich der Vitalitat der Eiraupen (Larve I) geben Tabelle und
Mortalitatsdiagramm (Abb. 15) die Ergebnisse der diesbeziiglichen Unter- suchungen wieder. Die Lage der Hilfskurven in letzterem, welche hier etwa ellipsenformig sind, und von innen nach auBen fortschreitend schatzungsweise den Verlauf der 20-, 40-, 50-, 70-, 80- und 1000/o- Mortalitatsgrenze veranschaulichen, zeigt auf den ersten Blick, daB die Verhaltnisse bei der Eiraupe wesentlich anders liegen als beim Ei.
Der opt imale Temperatur-Peuchtigkeitsbereich - die ge- strichelte Linie in Abb. 15 gibt die mutmaflliche 200/o-Mortalitatsgrenze wieder - ist f u r d i e E i r aupe au l le rordent l ich e n g begrenzt . Auf Grund des Diagramms liegt er zwischen 170-18° C und 80-90°/0
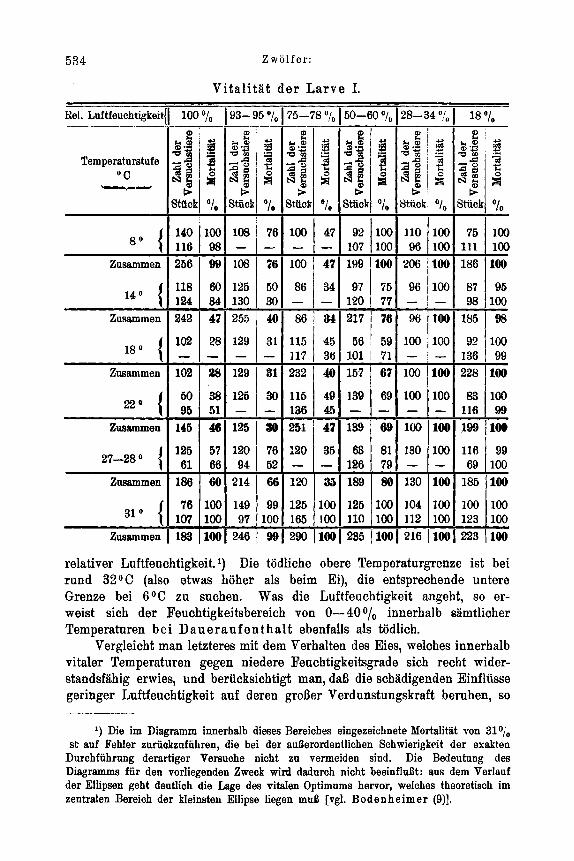
534 Zwolfer:
Vitali tat de r Larve I.
%I. Luftfeuchtigkeitjl 100 o/o I 93- 95 u/o I 75-78 I 50-60 "/" 128-34 O!,, I 18 o/o
3 3 o/o Stuck o/o Stuck1 o/o
140 100 108 1 76 8o ( 1 116 I 981 - 1 -
Zusammen 1 256 I W ) 108 1 76
118 60 125 50 {I 124 1 341 130 1 30
125 57 120 76 27-280 tl 61 I 661 94 i 52
100 i 471 199
86 34 97 - I - I120 86 1 !34 217
115 45 56 117 I 36 101 232 I 401 157
- I - I 126 120 35 189
125 100 126 I 165 I 0 0 110
~ 290 ll00l 235 -
69 100
81 130
iGpG
100 186 100
100 87 95 - 98 100 100 185 !N3
100 92 100 - 136 99 100 228 100
100 8 100 - 116 99
100 199 1100
100 116 99 - 69 100
loo loo 100 100 123 1100 1001 223 I 100
relativer Luftfeuchtigkeit. 1) Die todliche obere Temperaturgrenze ist bei rund 32OC (also etwas hoher als beim Ei), die entsprechende untere Grenze bei 6 0 C zu suchen. Was die Luftfeuchtigkeit angeht, so er- weist sich der Feuchtigkeitsbereich von 0-40 O l 0 innerhalb samtlicher Temperaturen bei Daueraufenthal t ebenfalls als todlich.
Vergleicht man letzteres mit dem Verhalten des Eies, welches innerhalb vitaler Temperaturen gegen niedere Feuchtigkeitsgrade sich recht wider- standsfahig erwies, und berucksichtigt man, daB die schadigenden Einflusse geringer Luftfeuchtigkeit auf deren groIer Verdunstungskraft beruhen, so
l) Die im Diagramm innerhalb dieses Bereiches eingezeichnete Mortalitat von 31 O/b
st auf Fehler zuruakzufiihren, die bei der auBerordentlichen Schwierigkeit der exakten Durchfiihrung derartiger Versuche nicht zu vermeiden sind. Die Bedeutuog des Diagramms fur den vorliegenden Zweck w i d dadurch nicht beeinfluBt: aus dem Vorlauf der Ellipsen geht deutlich die Lage des vitalen Optimums hervor, welches theoretiach im zentralen Bereich der kleineten Ellipse liegen muB [vgl. Bodenheimer (9)].
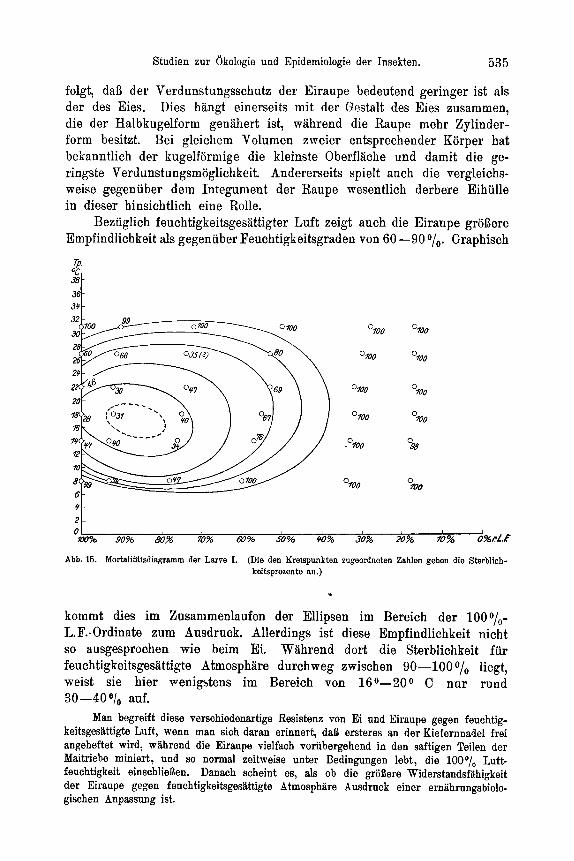
Studien zur Okologie und Epidemiologie der Insekten. 635
folgt, daB der Verdunstungsschutz der Eiraupe hedeutend geringer ist als der des Eies. Dies hangt einerseits mit der Gestalt des Eies zusammen, die der Halbkugelform genahert ist, wahrend die Raupe mehr Zylinder- form besitzt. Bei gleichem Volumen zweier entsprechender Korper hat beiranntlich der kugelformige die kleinste Oberflache und damit die ge- ringste Verdunstungsmoglichkeit. Andererseits spielt auch die vergleichs- weise gegeniiber dem Integument der Raupe wesentlich derbere Eihulle in dieser hinsichtlich eine Rolle.
Bezuglich feuchtigkeitsgesattigter Luft zeigt auch die Eiraupe groBere Empfindlichkeit als gegenuber Feuchtigkeitsgraden von 60 -90 ('lo. Graphisch
"700
'700
'700
'700
.'lo0
'700
'700
"roo
"700
'700
'M
0 700
700% 90% 80% 70% 60% 50% W% 30% 20% TO% O%dE
Abb. 15. MortalitZitsdiagramm der Larve I. (Die den Kreispnnkten zngeordneten Zahlen geben die Sterblich- keitspromnte an.)
kommt dies im Zusammenlaufen der Ellipsen im Bereich der 1000/o- L. I?.- Ordinate zum Ausdruck. Allerdings ist diese Empfindlichkeit nicht so ausgesprochen wie beim Ei. Wahrend dort die Sterblichkeit fur feuchtigkeitsgesattigte Atmosphare durchweg zwischen 90-100~/o liegt, weist sie hier wenigstens im Bereich von 16O--200 C nur rund 30-40 O l 0 auf.
Man begreift diese verschiedenartige Resistenz von Ei und Eiraupe gegen feuchtig- keitsgesattigte Luft, wenn man sich daran erinnert, daB ersteres an der Kiefernnadel frei angeheftet wird, wahrend die Eiraupe vielfach vorubergehend in den saftigen Teilen der Maitriebe miniert, und so normal zeitweise unter Bedingungen lebt, die 10Oo/, Luft- feuchtigkeit einsohlielen. Daoach scheint es, als ob die groBere Widerstandsfahigkeit der Eiraupe gegen feuchti,okeitsgesattitigte Atmosphare Ausdruck einer ernahrungsbiolo- gischen Anpassung ist.
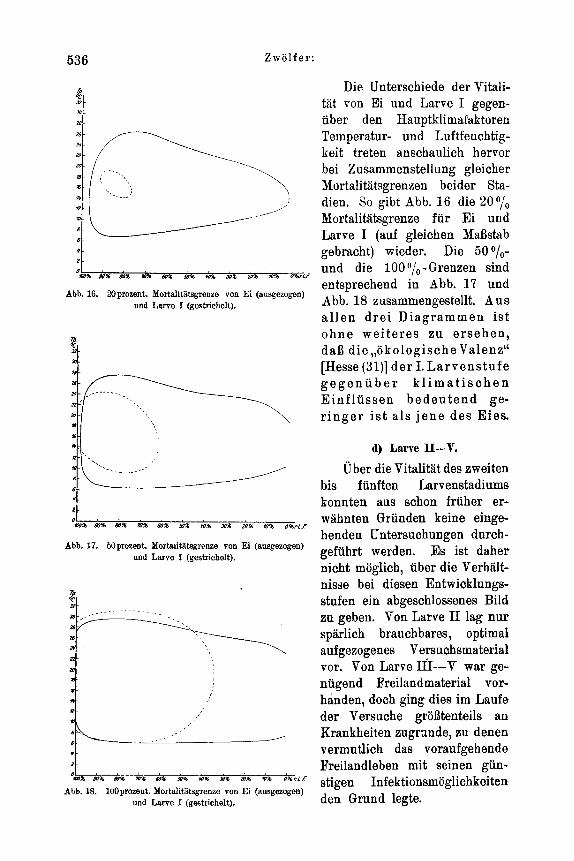
Zwol fe r : 536
Y 2
o , . . . W% ma all W% a0 9% W% a% 10% *% O%"lf
Abb. 16. 2Oprozent. MortaliCatsgrenze von Ei (ausgezogen) und Lame I (gestrichclt).
i , , , , , , , , , , 0 m% &V% U% ill% Y)% 9% W% 3% a% 70% 0L4r1.F
Abb. 17. b0prozent. Yortaiitiitsgrcnze von Ei (ausgezogen) nnd Lave I (gestrichelt).
F TI
i L & ma &VL a% Jb% 8% W% d % P L F
Abb. 18. 100prozent. Mortalit%tsgrenze von Ei (nnsgezogen) und Larve I (gestrichelt).
Die Unterschiede der Vitali- tlit von Ei und Larve I gegen- uber den Hauptklimafaktoren Temperatur- und Luftfeuchtig- keit treten anschaulich hervor bei Zusammenstellung gleicher Mortalitatsgrenzen beider Sta- dien. So gibt Abb. 16 die 20°/o Mortalitatsgrenze fur Ei urid Larve I (auf gleichen MaBstab gebracht) wieder. Die 500/0- und die 100 o/o - Grenzen sind entsprechend in Abb. 17 und Abb. 18 zusammengestellt. A u s a l l e n d r e i Diagrammen ist o h n e wei te res zu e r sehen , daB die, ,okologische Valenz" [Hesse(Sl)] d e r L L a r v e n s t u f e g e g e n i i b e r k l i m a t i s c h e n E i nf 1 us se n b e d e u te n d ge- r i n g e r ist als j e n e des Eies.
a) Lame 11-V.
Uber die Vitalitat des zweiten bis funften Larvenstadiums konnten aus schon friiher er- wahnten Grunden keine einge- henden Untersuchungen durch- gefuhrt werden. Es ist daher nicht moglich, uber die Verhalt- nisse bei diesen Entwicklungs- stufen ein abgeschlossenes Bild zu geben. Von Larve I1 lag nur sparlich brauchbares, optimal aufgezogenes Versuchsmaterial vor. Von Larve III--V war ge- niigend Preilandmaterial vor- handen, doch ging dies im Laufe der Versuche groBtenteils an Krankheiten zugrsnde, zu denen vermutlich das voraufgehende Freilandleben mit seinen gun- stigen InfektionsmSglichkeiten den Grund legte.

Studien zur Okologie uiid Epidemiologie der Insekten. 537
Die in den Versuchen mit Larve II-V abgestorbenen Tieren wurden einzeln mikro- skopisch untersucht. Dabei ergaben sich vorwiegend Bakteriosen, zum Teil aber auch Polyederkrankheiten als Todesursache. Hinsichtlich der Bakteriosen konnte allerdings im Rahmen dieser Untersuchung der exalite Nachweis, ob s i e die primare Todesursache bildeten, nicht erbracht werden. Was die Polyederkrankheiten im besonderen anbetrifft, so scheint sie bai der Kiefarneule keineswegs die groBe Bedeutung zu besitzen wie bei der Nonne: unter rund 500 aus verschiedenen mittelfrankischen und oberpfalzischen Forst- amtern stammenden, an Krankheiten zugrunde gegangenen Raupen, deren Kadaver einzeln untersucht wurden, konnte nur bei 2 Stuck das Vorliegen einer Polyederkrankheit mit Sicherheit festgestellt werden, obschon ein groBer Teil der untersuchten Tiere aus so- genannten ,,Eulenwipfeln'. stammte. Demgegenubor trat die Krankheit in dea Labo- ratoriumszuchten etwas starker auf, ohne aber auch hier im Gegensatz zu Bakteriosen epidemische Formen anzunehmen. Eine deutliche Abhangigkeit der Sarke des Auf- tretens der Erankheit von Temperatur- und Luftfeuchtigkeitsverhaltnissen - auf die be- sonders geachtet wurde - konnte wie beilaufig bemerkt sei, in den Laboratoriumszuchten weder bei der Polyederkrankheit noch bei Bakteriosen festgestellt werden. -
Von 49 Larven des IT. Stadiums (Einhauter), die vom Tage ihrer HIutung ab konstant 180 C/750/,, relativer L.B. ausgesetzt wurden, erreichten 45 Stuck die nachsthohere Entwicklungsstufe. Das kommt einer Mortalitat von rund SO/, gleich. Der entsprechende Versuch mit Larve I ergab Sterblichkeitsprozente von 450/, und 3 6 0 / 0 (im Mittel 40°/o).
In weiteren Versuchen mit Larve 11 bei 184/100U/, L.F; 18u/920/0 L. F; 220/750/, L. F.; und 28U/75 L.F. wurde ebenfalls Sterblichkeit beobachtet. Doch sind die Zahlen der Versuchstiere, die hierzu verwandt wurden, zu gering, urn brauchbare Anhaltspunkte fur die Vitalitit zu geben. Lediglich fur die Bestimmung der Entwicklungsdauer dieses Stadiums konnten sie Verwendung finden.
Das oben mitgeteilte Versuchsergebnis, das zunachst nur orientierenden Charakter besitzt, deutet darauf hin, daB L a r v e I1 e rheb l i cb w i d e r - s tan d s f a h i g e r g e g e n en t sp r e c h e n d e T e m per a t u r- P e u c h t i g k e i t s- kombina t ionen i s t , a l s La rve I. Hiernach erscheint die Annahme berechtigt, daB auch die Larven III-V der Kieferneule grol3ere Wider- standsfahigkeit in diesem Sinn besitzen und daB un te r d e n ver- sch iedenen Larvens tad ien d ieses Insek te s d i e E i r a u p e (Larve I) d ie ger ings te okologische Valenz aufweist.
e) Pnppe. Auch bezuglich der Puppe sind die gewonnenen Untersuchungs-
ergebnisse zu unvollstandig, um die Aufstellung eines allgemeinen Mortalitats- diagrammes zu gestatten, wie es fur Ei und Larve I gegeben wurde.
Es hangt dies mit der Beschranktheit der zur Verfugung stehenden technischen Einrichtungen zusammen. Wie sich im Laufe der Unter- suchung ergab, ist fur die Bearbeitung eines Mortalitatsdiagrammes der Kieferneulenpuppe die Ausdehnung der Versuche auf Temperaturen unter- 00 C erforderlich. Die den vorhandenen Einrichtungen fehlenden niederen Temperaturstufen spielen - wie leicht einzusehen ist -- gerade fur das Leben der Puppe als fur das uberwinternde Latenzstadium eine wesent- lich groBere Rolle als hohere Temperaturstufen. Versuche, die Abhangig-
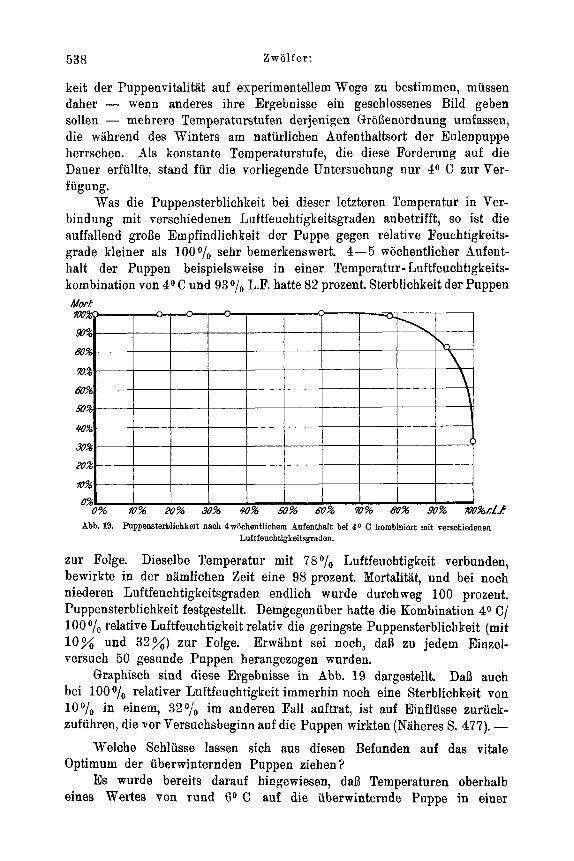
5 3 s Zwal fe r :
keit der Puppenvitalitat auf experimentellem Wege zu bestimmen, miissen daher - wenn anderes ihre Ergebnisse ein geschlossenes Bild geben sollen - mehrere Temperaturstufen derjenigen Grbfienordnung umfassen, die wahrend des Winters am natiirlichen Aufenthaltsort der Eulenpuppe herrschen. Als konstante Temperaturstufe, die diese Porderung auf die Dauer erfiillte, stand fiir die vorliegende Untersuchung nur 40 C zur Ver- fiigung.
Was die Puppensterblichkeit bei dieser letzteren Temperatur in Ver- bindung mit verschiedenen Liiftfeuchtigkeitsgraden anbetrifft, so ist die auffallend groBe Empfindlichkeit der Puppe gegen relative Peuchtigkeits- grade kleiner als 1000/, sehr bemerkenswert. 4-5 wochentlicher Aufent- halt der Puppen beispielsweise in einer Temperatur- Luftfeuchtigkeits- kombination von 4 0 C und 930/, L.F. hatte 8 2 prozent. Sterblichkeit der Puppen MOft
0 % ?0% 20% .N% 40% 50% 60% 70% 80% 90% 7tW%d.L Abb. 19. Poppensterbliohkeit nach 4 wiichentlichem Anfenthalt bei 40 C kombiniert mit verschiedenen
Luftfeuchtigkeitsgraden.
zur Folge. Dieselbe Temperatur mit 78 o/o Luftfeuchtigkeit verbunden, bewirkte in der namlichen Zeit eine 98 prozent. Mortalitiit, und bei noch niederen Luftfeuchtigkeitsgraden endlich wurde durchweg 100 prozent. Puppensterblichkeit festgestellt. Demgegenuber hatte die Kombination 40 C/ 100 o/o relative Luftfeuchtigkeit relativ die geringste Puppensterblichkeit (mit 10% und 32%) zur Folge. Erwabnt sei noch, daB zu jedem Einzel- versuch 50 gesiinde Puppen herangezogen wurden.
Graphisch sind diese Ergebnisse in Abb. 19 dargestellt. DaB auch bei 1000/, relativer Luftfsuchtigkeit immerhin noch eine Sterblichkeit von loo/, in einem, 32OjO im anderen Pall auftrat, ist auf Einflusse zuriick- zufuhren, die vor Versuchsbeginn auf die Puppen wirkten (Naheres S. 477). -
Welche Schliisse lassen sich aus diesen Befunden auf das vitale Optimum der uberwinternden Puppen ziehen ?
Es wurde bereits darauf hingewiesen, da13 Temperaturen oberhalb eines Wertes von rund 6O C auf die iiberwinternde Puppe in einer
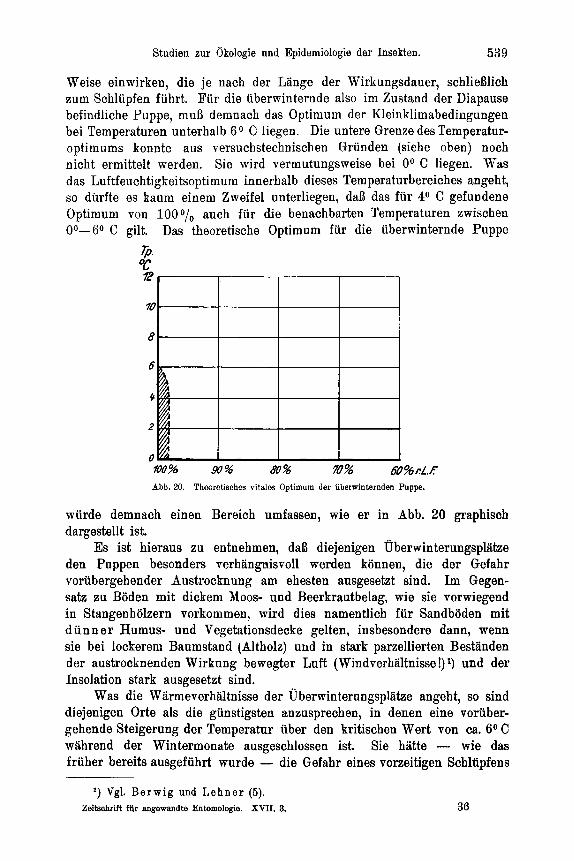
Studien zur Okologie und Epidemiologie der Jnsekten. 539
Weise einwirken, die je nach der Lange der Wirkungsdauer, schliefllich zum Schlupfen fuhrt. Fur die uberwinternde also im Zustand der Iliapause befindliche Puppe, mu13 demnach das Optimum der Kleinklimabedingungen bei Temperaturen unterhalb 6 0 C liegen. Die untere Grenze des Temperatur- optimums konnte aus versuchstechnischen Grunden (siehe oben) noch nicht ermittelt werden. Sie wird vermutungsweise bei O o C liegen. Was das Luftfeiichtigkeitsoptimurn innerhalb dieses Temperaturbereiches angeht, so durfte es kaum einem Zweifel unterliegeu, daB das fur 40 C gefiindene Optimum von 1000/o auch fur die benachbarten Temperaturen zwischen 00-60 C gilt. Das theoretische Optimum fur die uberwinternde Puppe
% OC Iz
70
8
6
4
2
0 700 % 90% 80 % 70% l W 6 P f . f Abb. 20. Theoretisches vital- Optimum der iiberwinternden Puppe.
wurde demnach einen Bereich umfassen, wie er in Abb. 20 graphisch dargestellt ist.
Es ist hieraus zu entnehmen, da13 diejenigen Uberwinterungsplatze den Puppen besonders verhangnisvoll werden konnen, die der Gefahr vorubergehender Austrocknung am ehesten ausgesetzt sind. Im Gegen- satz zu Boden mit dickem Moos- und Beerkrautbelag, wie sie vorwiegend in Stangenholzern vorkommen, wird dies namentlich fur Sandboden mit d u n n e r Humus- und Vegetationsdecke gelten, insbesondere dann, wenn sie bei lockerem Baumstand (Altholz) und in stark parzellierten Bestanden der austrocknenden Wirkung bewegter Luft (Windverhaltnisse !)I) und der Insolation stark ausgesetzt sind.
Was die Warmeverhdtnisse der Uberwinterungsplatze angeht, so sind diejenigen Orte als die gunstigsten anzusprechen, in denen eine voruber- gehende Steigerung der Temperatur iiber den kritischen Wert von ca. 6 0 C wahrend der Wintermonate ausgeschlossen ist. Sie hatte -- wie das friiher bereits ausgefiihrt wurde - die Gefahr eines vorzeitigen Schliipfens
I) Vgl. B e r w i g und L e h n e r (5). Zeitschrift fiir angewandte Entomologie. XVII. 3. 36

540 Z w o l f e r:
der Falter zur Polge, was nach den Erorterungen der vorausgehenden Abschnitte fur das Fortpflanzungsgeschaft und dje Kachkommenschaft ver- hangnisvoll werden konnte. Theoretisch bieten auch in dieser Hinsicht Standorte mit dicker Bodendecke (hohem Humus- und Vegetationsbelag) fur die Puppe die giinstigsten Uberwinterungsplatze. In der Tat wird dieser Schlul durch Mey e r s (46) eingehende Freilandbeobachtungen und durch Erfahrungen, die in der Literatur vorliegen, weitgehend bestiitigt. -
IV. Epidemiologische SohluBfolgerungen. Welche Polgerungen in epidemiologischer Hinsicht ergeben sich aus
all diesen Befunden ? Das gesetzmalrge Eingreifen der okologischen Valenz in die Ver-
breitung der Tiere hat R. Hesse (31) folgendermalen ausgedruckt : ,,Das daiiernde Vorkommen einer Tierart in einem Lebensraum hangt von jener Entwicklungsstufe des Tieres ab, die die geringste okologische Valenz hat". Der Satz beziehe sich zunachst nur auf die geographische Verbreitung der Tiere. SinngemaB 1 Z t er sich auch auf das zahlenmalige Vorkommen einer Tierart erweitern: Piir d i e B e v o l k e r u n g s d i c h t e e i n e r A r t i n a u f e i n a n d e r f o l g e n d e n G e n e r a t i o n e n i s t d a s j e n i g e i h r e r E n t w i c k l u n g s s t a d i e n v o n g r o B t e r B e d e u t u n g , w e l c h e s r e 1 a t i v d i e g e r i n g s t e o k o 1 o g i s c h e V a1 e n z b e s i t z t. 1) Handelt es sich im besonderen um die Beurteilung klimatischer Einflusse auf den Wechsel der Populationsdichte einer Art in aufeinanderfolgenden Generationen, so ist hierfur die geringste okologische Valenz gegeniiber Klimafaktoren malgebend.
Die En twick lungss tu fe m i t g e r i n g s t e r okologischer Valenz gegenuber Kl imafaktoren s t e l l t epidemiologisch be t r ach te t das kr i t i sche S tad ium itn Entwicklungsgang e ine r A r t vor. Die Zei t spanne , i n we lche r es hanptsachl ich in Er sche innng tritt, ist s i n n g e m a l d i e k r i t i s che Zeit fu r die be t re f fende Genera t ion d e r Art . Auf de ren Schicksa l muB i m ubr igen d i e Wi t t e rungs - kons te l la t ion wahrend d e r k r i t i s chen Zei t en t sche idenden Ein- fluIj haben.2)
Die vorstehend mitgeteilten theoretischen Erorterungen lassen ei n e s der Ziele der neueren epidemiologischen Arbeitsmethodik erkennen : Si e ges t a t t e t auf exper imente l lem Wege d a s k r i t i s che S tad ium bezw. d ie k r i t i s chen S tad ien e ine r I n s e k t e n a r t zu ermit te ln . Sind die phanologischen Daten fur die einzelnen Entwicklungsstufen der betreffenden Art hinreichend bekannt, so gibt der Vergleich der Witterungsverhaltnisse
I) Denselben SchluP zieht F r i e d r i c h s (277, allerdings ohue die hier gemachte und weiter unten nliher begriindete Einscbrankung, daB es hauptsachlich auf das Stadium mit ,,relativ" geringster okologischer V a h z aokomnit.
.. . ~
*) Vgl. Zwol fer (75, 76).

Studien zur Okologie und Epidemiologie der Insekten. 541
wahrend der kritischen Zeit aufeinanderfolgender Generationen AufschluB uber die Bedingungen des GroQklimas, die eine Gradation der Art zur Folge haben kiinneu.
Was nun die okologische Valenz der verschiedenen Stadien der Kieferneule aabetrifft, so geben die mitgeteilten Untersuchungsergebnisse ein Bild von der Abhangigkeit der Vitalitat des Eies, der Eiraupe, der Puppe und der Imago von den Klimafaktoren Temperatur und relative Luftfeuchtigkeit. Fur das Raupenstadium 11-V wurde bereits auf die Berechtigung der Annahme hingewiesen, daB die okologische Valenz dieser Stufen groBer ist als jene der Eiraupe. Zur Feststellung, welche der erstgenannten Entwicklungsstufen das kritische Stadium der Kiefern- eule vorstellt, sind demnach die Vitalithtsdiagramme dieser Stufen (Abb. 5, 13, 15, 20) zu vergleichen.
Bezuglich des Imaginalstadiums 1aBt sich zunachst einwenden, daB ein Diagramm, aus dem das Optimum der Eiablage hervorgeht, nicht mit den Mortalitatsdiagrammen der iibrjgen Entwicklungsstufen verglichen werden kann. Demgegenuber sei daran erinnert, daR das vitale Optimum als diejenige Temperatur-Peuchtigkeitskombination definiert wurde, in welcher die geringste. Sterblichkeit auftritt oder was gleichbedeutend ist, der groBte Prozentsatz des betreffenden Stadiums die nachste Entwicklungs- stufe erreicht. Die auf das Imaginalstadium folgende Entwicklungsstufe ist das Ei. Die groBtmopliche Eizahl entspricht hier der groBten Zahl Uberlebender, die geringste Reduktion der absoluten Eizahl sinngemaB der geringsten Mortalitat ! Unter diesem Gesichtspunkt betrachte t steilt das Optimum fur die Eiablage der Palter gleichzeitig das vitale Optimum des Imaginalstadiums vor.
Noch ein weiterer Uinstand muB in diesem Zusammenhang erortert werden : Ein Vergleich der okologischen Valenzen verschiedener Ent- wicklungsstufen einer Art ist nur dann zulassig, wenn die letzteren annahernd ein- und denselben Wohnort innerhalb des Biotops der Art einnehmen. Vergleicht man namlich im Falle der Kieferneule den Bereich des vitalen Optimum fur Ei, Larve I, Puppe und Imago, so zeigt derjenige der Puppe (Abb. 20) sehr engste Grenzen. Doch ergibt eine einfache Uberlegung, daB diese i m vorliegenden Fall aus der Qegenuberstellung auszuscheiden hat: Die Puppe der Kieferneule ist dank ihrer Uberwinterung in der Bodenstreu den d i r e k t en Einflussen der Witterung entzogen. Vergleichs- weise siud die Schwankungen des Kleinklimas am normalen Verpuppungs- orte aullerordentlich gering. Hierdurch ist ein Ausgleioh fur die geringe okologische Valenz der Puppe geschaffen. Wiirde dieselbe im Freien oder unzureichend geschiitzt in Rindenritzen uscv. uberwintern, wie etwa jene des KohlweiBlings (Pieris brassicae L.) dann ware eine Gegenuber- stellung im obigen Binn mit den anderen Entwioklungsstufen zuliissig. So ergibt sich, dab die Puppe der Kieferneule trotz ihrer geringen okologischen Valenz zufolge der ihr eigentiimlichen Gewohnheit an gut
36 *

542 Zwol fe r :
geschutzten Platzen zu uberwintern, nicht als das kritische Stadium der Kieferneule bezeichnet werden kann. I h r e E igena r t i s t f u r loka le Un te r sch iede des Massenwechsels von Bedeutung, n i c h t a b e r f u r d i e Aus losung e i n e r Gradat ion en tsche idend.
Die Angaben von D o l l e s (18) uber den Verjauchungsgrad der Eulenpuppen wSihrend seiner 12 jahrigen Beobachtungen belegen die Richtigkeit des Gesagten: Nach diesem dutor schwankte der Prozentsatz verjauchter, also vermutlich durch klimatische Einflusse zugrunde gegangener Puppen in den 12 aufeinanderfolgenden Generationen; zwischen 10-20 O/,. Ware die Puppe das kritische Stadium der Eule im epidemiologischen Sinn. dann hatten e r h e b l i c h g r o l e r e S c h w a n k u n g e n der Prozentsatze verjauchter Puppen in den einzelnen Jahrgangen erwartet werden miissen. Insbesonders hatten die Generationen der ,,Normaljahre(( und um solche handelte es sich bei den Dollesschen Uutersuchungen vorwiegend - durch wesentlich hohere Verjauchungsprozente gekennzeichnet sein miissen.
Demgegenuber sind Ei, Eiraupe und Imago der Kieferneule den Witterungseinfliissen unmi t t e lb a r und annahernd in gleicher Weise aus- gesetzt. Unter diesen drei Entwicklungsstufen besitzt das vitale Optimum der Eiraupe (Abb. 15) engste Begrenzung. Dasjenige des Eies (Abb. 13) ist am weitesteu, wahrend das vitale Optimum der Falter (Abb. 5) eine mittlere Stellung einnimmt.l) Dementsprechend s t e l l t d i e E i r a u p e das k r i t i s che S t a d i u m d e r K i e f e r n e u l e im epidemiologischen S i n n e vor. E i n e g e w i s s e Bedei i tung i n d i e se r H i n s i c h t wi rd auch d e m I m a g i n a l s t a d i o m n o c h z u g e s p r o c h e n werden mussen , wahrend das Ei infolge seines weitbegrenzten vitalen Optimums epidemio- logisch weniger bedeutuugsvoll ist.
Exakte phanologische Daten uber das Auftreten der Hauptmasse der Eiraupen, die in diesen Zusammenhang von besonderem Interesse sind, fehlen in der alteren Kieferneulenliteratur. Bus den vorhandenen Unter- lagen ist lediglich zu entnehmen, da13 die Hauptzeit fur das Auftreten der Eiraupe etwa von Nitte Mai bis Mitte Juni liegen durfte und je nach Lage und Jahrgang der Beobachtung Unterschiede aufweist. Dies Fare demnach die kritische Zeit in der Entwicklung der Kieferneule. Ent- sprechend lie& die Hauptzeit fur den Palterflug (Eiablageperiode), der nach obigem epidemiologisch ebenfalls eine gewisse Bedeutung zu- gesprochen werden muB , ungefahr zwischen Mitte April bis Nitte Mai.
Nach dem ganzen Gang der Uberlegungen mussen die im Kiefern- eulenbiotop wahrend dieser Zeit herrschenden Kleinklimaverhaltnisse ent-
l) Letzteres unter der Voraussetzung, daB die im Diagramm Abb. 5 niedergelegten Daten nicht nur relative, sondern absolute Giiltigkeit haben. Vergleicho hierzu die Beinerkung Seite 512 dieser Arbeit. Bei der Verschiedenartiglieit der Anordnung der Versuohe mit Imagines einerseits, Eiern und Eiraupen andererseits, die clurch Unvoll- kommenheiten der technischen Einrichtungen bedingt waren, sind Zweifel an der Zuliissigkeit der Gegenuberstellung berechtigt. Es kam in diesem Zusamrnenhang jedoch mehr darauf an, die prinzipielle Bedeutung derartiger Versuche und die Moglichkeit ihrer Auswertung darzulegen. Aufgabe kiinftiger Forschunpen mit vollkommeneren Bilfsmittdn wird es sein, etwaige Versuchsfehler zu berichtigen.

Studien zur Okologie und Epidemiologie der Insektea. 543
scheidenden EinfluB auf Entstehung und Verlauf einer Kieferneulen- kalamitat ausuben. Nun ist das Kleinklima in seinen Schwankungen wesentlich vom GroBklima abhlngig. Die Bewegungen des ersteren mussen dementsprechend in den meteorologischen Daten fur das GroBklima zum Aus- druck kommen. In der Tat ergaben Berwigs (3) statistisch-klimatologische Studien, die auf GroBklimawerten aufgebaut sind, fur die fragliche Zeit- spanne jener Jahre, die einer Eruption vorausgehen, ein von der Norin abweichendes, durch unternormale Niederschlagsmengen ausgezeichnetes Verhalten. Dasselbe macht sich nach Berwig namentlich in den Vor- bereitungsjahren ubereinstimmend bemerkbar.
Noch prkiser lieBen sich diese Verhaltnisse fassen, wenn lediglich die GroBklimadaten fur die ,,kritische Zeit" des Eiraupenstadiums verglichen werden wurden. Ein derartiges Studium setzt allerdings langjahrige e x a k t e phhnologische Daten uber die Kieferneule voraus. Vorerst besitzen wir solche nur fur 1930 und das mittelfrankische Qebiet durch die eingehenden Untersuchungen M e y e r s (46). Nach diesem fie1 hier die Hauptzeit fur das Auftreten der Larve I im genannten Jahre in die Zeit vom 1.-10. Juni. Je nach dem Temperaturverlauf der Fruhjahrswitterung wird naturlich diese kritische Zeit in verschiedenen Jahrgangen bald etwas fruher, bald etwas spater liegen, was die praktische Auswertung der phanologischen Beobachtungen Meyers etwas erschwert. Nach allem Vorhergesagten mul3te jedenfalls die Witterung dieser Zeitspanne im Laufe verschiedener Jahrgange einer Gradation durch Besonderheiten gekennzeichnet sein.
Wird ein derartiger Vergleich fur die drei von Berwig genannten mittelfrankischen Kalamitaten 1899/01, 1911/13, 1918/20 und fur die ver- flossene 1928-1930 an Hand der vorhandenen meteorologischen Daten durchgefuhrt,l) so zeigt sich in der Tat, daB in den fraglichen Jahrgangen wahrend der Zeit vom 25. Mai bis etwa Mitte Juni j e w e i l s e i n e m e i s t i n d a s e r s t e J u n i d r i t t e l f a l l e n d e D e k a d e a u f t r i t t , d e r e n N i e d e r s c h l a g s s u m m u n z w i s c h e n 0 b i s 12,2 m m , i m M i t t e 1 b e i 5,5 m m 1 i e g e n. Die dazwischenliegenden Perioden weisen demgegenuber im ersten Junidrittel Niederschlagsmengen von 14,6 bis 56, l m m , im Mittel 31,O mm Regenmengen auf. Lediglich die auf das Krisenjahr der verschiedenen Kalamitaten folgenden 1 oder 2 Jahrgange und zwei weitere vollig isoliert stehende besitzen wahrend der 38 Be- obachtungsjahre in der ,,kritischen Zeit" geringe Niederschlagsmengen. Ersteres weist gleichzeitig darauf hin, daB der Zusammenbruch einer Euleneruption nicht klimatisch bedingt zu sein braucht.
l) Meteorol. Tagesaufzeichnungen fur Nurnberg fur 1893-1913 im Deutschen Meteorol Jahrbuch (45). Tiigliche Njederschlagsmengen fur 1901-1926 fur Wendel- s t e in , Hipoltstein, Eiliansdorf im Reichswald (Mittelfranken) im ,,Jahrbuch Bayer. Landesstelle f u r Gewasserkunde Bd. 3-28", Munchen 1901-1926. Fur 1927-1930 wurden hierzu die entsprechenden Akten der Landesstelle durchgesehen. Altere Extenso-Publikationen aus dem Gebiet fehlen.

544 Z w o 1 f e r :
DaB stets drei bezw. vier aufeinanderfolgende Jahrgange mit gunstiger Wetterlage wahrend der kritischen Zeit die Eulengradation einleiten, ist bei einem Insekt mit einjahriger Generation begreiflich: ein einziges Jahr mit gunstiger Wetterlage reicht bei einem solchen nicht aus, u m eine Zu- nahme der Populationsdichte zu bewirken, wie sie zu einer Massen- vermehrung erforderlich ist. Hierzu bedarf es m e h r e r e r aufeinander- folgender Generationen. 1)
Das Gesagte gilt zunachst nur fur Mittelfranken. Fiir andere Gebiete mit anderen phanologischen Daten werden sinngemal3 andere kritische Zeiten in Frage kommen. Sie mussen von Pall zu Fall ermittelt werden. I m Zusammenhang mit dieser Untersuchung kam es lediglich darauf an, zu zeigen, daB die kritische Zeit in der Entwicklung eines Insektes relativ kure bemessen sein kann, und daB sie im epidemiologischen Sinn fur das Schicksal einer Eulengeneration von wesentlicher Bedeutung ist. Z u- s a m m e n f a s s e n d e r g i b t s ich spez ie l l fu r das mi t t e l f r ank i sche Gebie t e i n e Niede r sch lagsmenge von 0 b i s r u n d 1 2 mm wahrend e i n e r i n d ie Zei t vom 25. Mai b is Mi t te J u n i fa l lenden Dekade a l s g rada t ions fo rde rnd . E i n e hohere! von ca. 15 b is 20 mm a n a u f w a r t s , ub t g rada t ionshemmenden Einf luB aus , d e r je nach d e r Hohe der Xiederschlagsmenge g r a d u e l l verschieden s e i n wird. In1 i ib r igen f i ihren au fe inande r fo lgende Eulengene- r a t i o n e n m i t g l e i chs inn ig g u n s t i g e r W e t t e r l a g e w a h r e n d d e r k r i t i s c h e n Zei t j e nach d e r Hohe d e r dusgangspopu la t ions - d i c h t e d e s S c h a d l i n g s f r u h e r ode r s p a t e r z u r Erupt ion . A u c h d u r c h g r a d u e l l e Un te r sch iede d e r , ,guns t igen Wet t e r l age" wahrend d e r k r i t i s c h e n Zeit auf e i n a n d e r f o l g e n d e r Genera t ionen kann Zeipun k t und AusmaB d e r E r u p t i o n bes t immend bee in - f luBt werden. -
V. Die Entwicklungsdauer der prttimaginalen Stadien. Die tagliche Kontrolle der Vitalitatsversuche, uber deren Ergebnisse
in den voraufgehenden Abschnitten berichtet wurde, gab Gelegenheit, auch die Prage der Abhangigkeit der Entwicklungsdauer des Eies und der Lame von den Witterungsfaktoren Temperatur und Luftfeuchtigkeit eingehender zu untersuchen. Die diesbezuglichen Ergebnisse sind im folgenden zu- sammenfassend wiedergegeben. 2)
Uber Beeinflussung der Entwicklungsdauer des Eies durch die herrschenden Tempe- raturen liegen in der Literatur Angaben vor von R a t z e b u r g (52, 53), Wolff und
l) Vgl. hierzu das Seite 486 uber den Gesamtwiderstand Gesagte. Entsprechendes gilt fur die Massenvermehrung von Schistocerca greguria Forsk. [Boden h e i m e r (lo)] Und fur jene von Eurygaster integriceps Put. [Z w 61 f e r (76)]
2, l i t Rucksicht auf den mehr epidemiologischen Charakter der vorliegenden Unter- suchung ist auf ausfuhrliche Darstellung unter Mitteilung aller Einzelheiten verzichtet worden.

Studien zur Okologie und Epidemiologie der Insekten. 545
KrauBe (73), E c k s t e i n (20) und S a c h t l e b e n (55). Es handelt sich dabei mehr um gelegentliche Beobachtungen , wie sie wahrend der Zuchtversuche von den genannten Autoren gemacht wurden. Im einzelnen sind sie in S a c h t l e b e n s Monographie (56) zu- sammengestellt, so d a l hier auf ihre Wiedergabe verzichtet werden kann.')
Bus den Versuchen iiber die Entwicklungsdauer des Eies unter konstanten Temperatur- und Luftfeuchtigkeitsverhaltnissen, denen rund 2700 Einzelbeobachtungen zuprunde liegen, geht zunachst hervor, daB die untere Grenze der fur die Eientwicklung wirksamen Temperaturen bei einem Wert zwischen 40 bis 80 C liegt, - ahnlich wie dies auch schon fur die fruher geschilderten LebensauBerungen der Kieferneule gilt: konstanter Aufenthalt der Eier durch 41/, Monate hindurch bei einer Temperatur von 4 0 C hat - gleichviel mit welcher Luftfeuchtigkeit sie kombiniert wird - deren allmahliches Absterben zur Polge, ohne daB ein einziges Ei zum Schlupfen gelangt. Demgegenuber schlupfte hei Tempera- turen von 80 C ein relativ hoher, von den betreffenden Feuchtigkeits- kombinationen abhangiger Prozentsatz im Durchschnitt nach 47,8 Tagen.
Die relative Luftfeuchtigkeit, mit verschiedenen Temperaturen ver- bunden, hat nur auf die Lebensfahigkeit der Eier einen erheblichen Ein- fluB. Eine auffallige Wirkung derselben auf die Entwicklungsdauer der Eier, wie das bei einigen anderen Insekten bereits beobachtet worden ist [P i e rce (50), Andersen (l)] konnte fur das Kieferneulenei nicht festgestellt werden. Seine Eutwicklungsdauer scheint allein von der Temperatur aus- schlaggebend bestimmt zu werden.2) Im ubrigen zeigen die Versuche, daB die Variationsbreite der Entwicklungsdauer bei den tieferen Ternperaturen (80) erheblich grol3er ist als bei den hoheren (220-280). Dies gilt selbst fur ein und dasselbe Gelege, also fur Geschwistereier: Wahrend in den oberen Temperaturstufen das Schlupfen der Eier eines Geleges fast stets innerhalb eines Tages erfolgte, lag es bei der Temperatur von 8 O C haufig mehrere Tage auseinander. Vermutlich hangt diese Erscheinung mit der sehr verminderten Aktivitat der Eiraupen bei niederen Temperaturen zu- sarnmen.
Die folgende Tabelle enthalt die Minimal-Mittel- und Maximalwerte fur die Entwicklungsdauer des Forleuleneies bei verschiedenen Tempera- turen zwischen 40-280 C. Da sich kein nennenswerter EinfluB der Luftfeuchtigkeitsverhaltnisse auf die Entwicklungsdauer der Eier fest- stellen IieB, sind in derselben die Beobachtungen bei verschiedener Luft-
Was die Temperaturahhaogigkeit der Entwicklungsdauer boi Insekten im all- gemeinen angeht, so liegt bieriiber eine umfangreicbe Literatur vor, auf die hier nicht im einzelnen eingegangen werden kann. Es sei iu diesem Zusammenhang nur auf die Arbeiten verwiesen von A n d e r s e n (1), B a c h m e t j e w (a), Blunckffi) , B o d e n h e i m e r ( 8 bis 12), E s c h e r i c h (23), F r i e d e r i c h s (27, 28), H e a d l e e (30), J a n i s c h (33--35), M a r t i n i (40), P i e r c e (50), S a n d e r s o n (57), S h e l f o r d (61-64), U v a r o v (68).
z, Zu demselben Ergebnis gelangte H e a d l e e (30) fur Toxoplera grarninum Rod.: Relative Luftfeuchtigkeitsgrade zwischen 37- 100°/o hatten bei konstsnter Temperatur keinen Einflul3 auf die Entwicklungsdauer dieser Art.
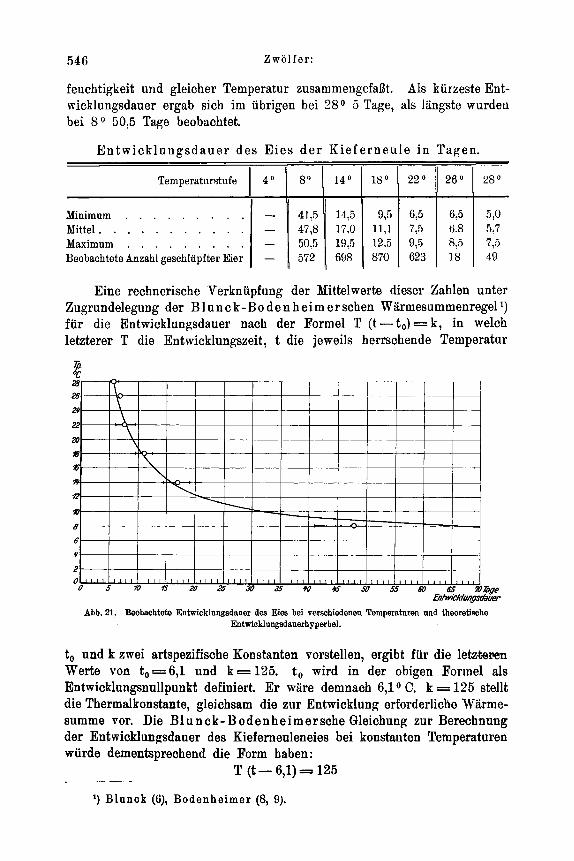
546 Z w o l f e r :
Ternperatiirstufe 4" 8"
Minimum . . . . . . . . . - ~ 413 Mittel . . . . . . . . . . . - 47,8 Maximum . . . . . . . . . - 50.5 Beobachtete Anzahl geschliipfter Eier - 572
I __ - -
14" 18" 22O ' 26" 28"
' 14.5 9,5 1 6,5 6,5 5,O 17.0 11,l 7.5 6.8 5,7 19.5 12.5 9.5 8.5 7,5 698 870 623 18 49
to und k zwei artspezifische Konstanten vorstellen, ergibt fur die letzhmen Werte von to=6,1 und k = 125. t o wird in der obigen Porinel als Entwicklungsnullpunkt definiert. Er ware demnach 6,lO C. k = 125 stellt die Thermalkonstante, gleichsam die zur Entwicklung erforderliche Warme- summe vor. Die Blunck-Bodenheimersche Gleichung zur Berechnung der Entwicklungsdauer des Kieferneuleneies bei konstauten Tetnperaturen wurde dementsprechend die Form baben:
T (t - 6,l) = 125
Y
2 0
l) Blunck (6), Bodenheimer (8, 9).
I I I I 1 1 1 1 I I I I I l l ! I I I I 1 1 1 1 I I I I I l l 1 I l l 1 I l l 1 I I I I I I I I I I I I

Studien zur Okologie und Epidemiologie der Insekten. 547
Temperatur I 8O 1 14O 18O 1 22"
Errechoet fur to = 6,l und k = 125 I 66,O 1 15,8 10,5 ' 7,8 Beobachteter Mittelwert . . . . . 47,8 17,O 11,l I 7,5
1 26O I 28"
6,2 5,7 6,8 5,7
I) Vgl. hierzu Mart in i (41).
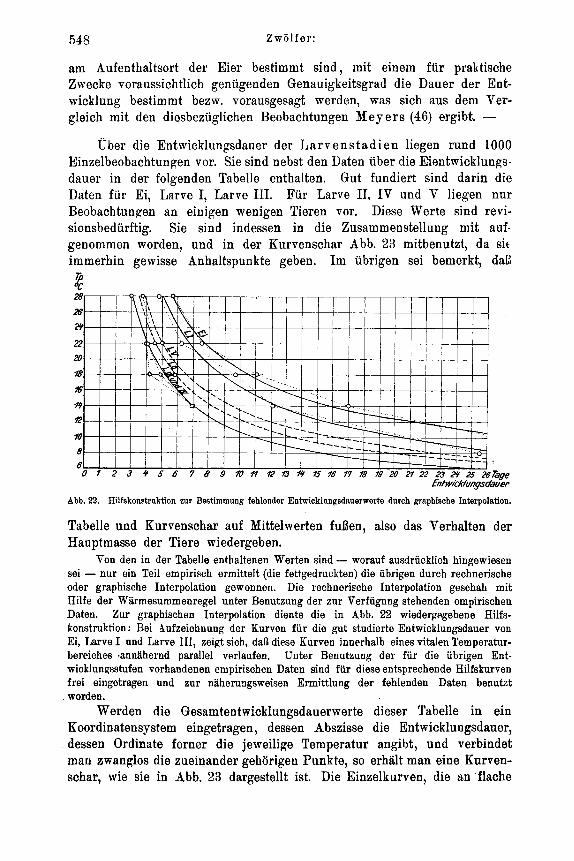
548 Zw6l fe r :
am Aufenthaltsort der Eier bestimmt sind, mit einem fur praktische Zwecke voraussichtlich geniigenden Genauigkeitsgrad die Dauer der Ent- wicklung bestimrnt bezw. vorausgesagt werden, was sich aus dem Ver- gleich mit den diesbezuglichen Beobachtungen Mey e r s (46) ergibt. -
Uber die Entwicklungsdauer der La rvens t ad ien liegen rund 1000 Einzelbeobachtungen vor. Sie sind nebst den Daten uber die Eientwicklungs- dauer in der folgenden Tabelle enthalten. Gut fundiert sind darin die Daten fur Ei, Larve I, Larve 111. Fur Larve 11, IV und V liegen nur Beobachtungen an einigen wenigen Tieren vor. Diese Werfe sind revi- sionsbedurftig. Sie sind indessen in die Zusammenstellung mit auf- genommen worden, und in der Kurvenschar Abb. 23 mitbenutzt, da sit immerhin gewisse Anhaltspunkte geben. Im ubrigen sei bemerkt, daE
8 28 26 w 22 20 78
b 74 n 70 8 6 0 7 2 3 4 5 6 7 8 9 70 77 12 73 74 75 76 77 78 79 20 21 22 23 B 3 MTqe
Fnhwckhngsdauer
Abb. 22. Hilfskonstmktion zuc Bestimmung fehlender Eutwicklungada~erwe~e durch graphiche Interpolation.
Tabelle und Kurvenschar auf Mittelwerten fuBen, also das Verhalten der Hauptmasse der Tiere wiedergeben.
Von den in der Tabelle enthaltenen Werten sind - worauf ausdriicklich hingewiesen sei - nur ein Teil empirisch ermitrelt (die fettgedruckten) die ubrigen durch rechnerische oder graphische Interpolation gewonnen. Die rechnerische Interpolation geschah mit Hilfe der Warmesummenregel unter Benutzung der zur Verfugnng stehenden empirischen Daten. Zur graphischen Interpolation diente die in Abb. 22 wiedergegebene Eilfs- konstruktion: Bei hufzeichnung der Kurven fur die gut studierte Entwicklungsdauer von Ei, h r v e I und Larve 111, zeigt sich, daB diese Kurven innerhalb eines vitalen Temperatur- bereiches -annahernd parallel vwlaufen. Uuter Benutzung der fur die ubrigen Ent- wicklunqsstufen vorhandenen empirischen Daten sind fur diese entsprechende Hilfskurven frei eingetragen und zur nahoruggsweisen Ermittlung der fehlenden Daten benutzt worden.
Werden die Gesamtentwicklungsdauerwerte dieser Tabelle in ein Koordinatensystem eingetragen, dessen Abszisse die Entwicklungsdauer, dessen Ordinate ferner die jeweilige Temperatur angibt, und verbindet mau zwanglos die zueinander gehoripen Punkte, so erhalt man eine Kurven- schar, wie sie in Abb. 23 dargestellt ist. Die Einzelkurven, die an flache
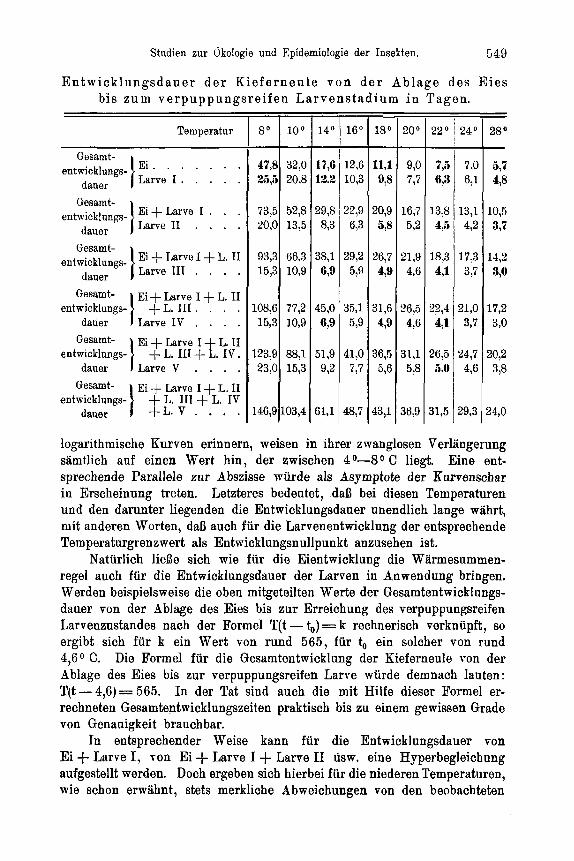
Studien zur Okologie und Epidemiologie der Insekten. 549
24O
En twick lungsdaue r d e r K ie fe rneu le von d e r Ablage des E i e s bis z u m verpuppungs re i f en La rvens t ad ium i n Tagcn.
28O Temperatur
47,s 25,5
73,5 20,O
93,3 15,3
.08,6 15,3
.23,9 23,O
Ei . . . . . . . Larve I . . . . .
Gesamt- entwicklungs-
dauer
32,O 20,8
52,8 13,5
66,3 10,9
77,2 10,9
88,l 15,3
Ei+ Larve I . . . Larve I1 . . . .
Ei + Larvc! I + L. I1 Larve 111 . . . .
Gesamt- entwicklungs-
dauer Gesamt-
entwicklungs- dauer
7,O 6,l
13,l 4,2
17,3 3,7
21,O 3,7
Gesamt- Ei + Larve I + L. I1 entwicklungs- +L. 111. . , .
Ei + Larve I + L. 11 f L. 111 + L. IV .
Ei + Larve I + L. IT + 1,. 111 + L. I V
dauer I Larve IB . . . .
dauer 1 Larve V . . . .
dauer I + L . V . . . .
Gesamt- entwickluogs-
Gesamt- entwicklungs-
5,7 4,s
10,5 3,7
142 3,O
17,2 3,O
24,7 4,6
29,s
17,6 12.2
29,8 873
38,l 6,9
45,O 679
51,9 972
61,l
20,2 3,8
24,O
- __ - 16O
12,6 10,3
22,9
-
6,3
29,2 5,9
35,l 579
41,O 717
48,7 .46,9
- - 18 O
11,l 978
20,9 5,s
-
26,7 4,9
31,6 439
36,5 5,6
43,l 103,4
- - 200
9,o 7 77
-
16,7 532
21,9 4.6
26,5 476
31,l 578
38,9
7,5 673
13,s 4,5
18,3 471
2 2 4 431
26,5 5.0
31,5
logarithmische Kurven erinnern, weisen in ihrer zwanglosen Verlangerung samtlich auf einen Wert hin, der ewischen 4O--80 C liegt. Eine ent- sprechende Parallele zur Abszisse wurde als Asymptote der Kurvenschar in Erscheinung treten. Letzteres bedeutet, dalj bei diesen Temperaturen und den darunter liegenden die Entwicklungsdauer unendlich lange wahrt, mit anderen Worten, daB auch fur die Larvenentwicklung der entsprechende Temperaturgrenzwert als Entwicklungsnullpunkt anzusehen ist.
Naturlich liefie sich wie fur die Eientwicklung die Warmesummen- regel auch fur die Entwicklungsdauer der Larven in Anwendung bringen. Werden beispielsweise die oben mitgeteilten Werte der Gesamtentwicklungs- dauer von der Ablage des Eies bis zur Erreichung des verpuppungsreifen Larvenzustandes nach der Formel T(t - to) = k rechnerisch verknupft, so ergibt sich fur k ein Wert von rund 565, fur to ein solcher von rund 4,60 C. Die Bormel fur die Gesamtentwicklung der Kieferneule ron der Ablage des Eies bis zur verpuppungsreifen Larre wiirde demnach lauten : T(t - 4,6) = 565. In der Tat sind auch die mit Hilfe dieser Formel er- rechneten Gesamtentwicklungszeiten praktisch bis zu einem gewissen Grade von Genauigkeit brauchbar.
In entsprechender Weise kann fur die Entwicklungsdauer von Ei + Larve I, Ton Ei + Larre I + Larve I1 usw. eine Hyperbegleichung aufgestellt werden. Doch ergeben sich hierbei fiir die niederen Temperaturen, wie schon erwahnt, stets merkliche Abweichungen von den beobachteten
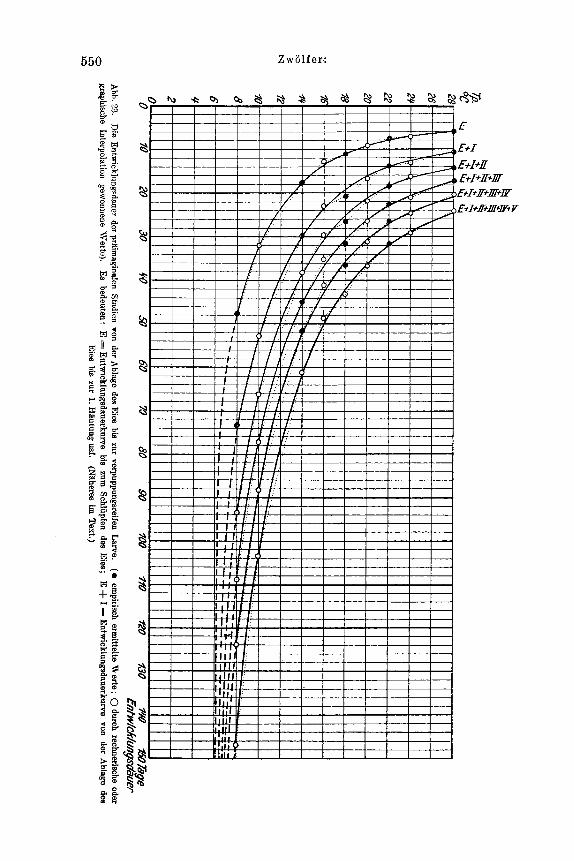
550 Zwolfer:

Studien zur okologie und Epidemiologie der Insekten. 55 1
Entwicklungsdauerwerten, so da13 im ganzen die in Abb. 23 uiedergelegte, empirisch gewonnene Kurvenscbar eine bessere Moglichkeit darstellt, die Entwicklungszeiten der einzelnen Stadien bei verschiedenen Temperaturen auf graphischem Wege naherungsweise zu bestimmen.
Auffallend ist, daB die Gesamtentwicklungsdauer bis zur verpuppungs- reifen Larve fur den Entwicklungsnullpunkt einen Wert fur to von 4,60 C lieferte, wahrend die Eientwicklung allein einen solchen von 6,l O C ergab. Ob dieser geringen Verschiedenheit des fur den Entwicklungsnullpunkt gefundenen Wertes eine grundsatzliche Bedeutung zukommt, ob sie die Folge einer gewissen Unvollkommenheit der Hyperbelgleichung als mathematischer Ausdruck der Entwicklungsdauer bei Insekten ist, oder ob unvermeidliche Pehler der Versuchstechnik an ihr schuld sind, lafit sich einstweilen nicht entscheiden.
Zur praktischen Benutzung der Abb. 23 ist kurz folgendes zu sagen: E bedeutet in ihr die Entwicklungsdauerkurve des Eies; E + I jeue von der Ablage des Eies bis zur ersten Hautung; E + I + 11 entsprechend bis zur zweiten H&utung usw. . . . E + 1 + 11 -/- I11 + LV + V endlich stellt die Kurve der Entwicklungsdauer von der Ablage des Eies bis zum verpuppungsreifen Zustand d e r h r v e vor.
sol1 beispielsweise die Uauer der Entwicklung bei einer bestimroten Temperatur von der Ablage des Eies bis zur 11. Hautung festgestellt werden, dann ist lediglich er- forderlich, vom betreffenden Temperaturpunkt der Ordinate aus, eine Parallele zur Absziwee bis zum Schnittpunkt mit der Kurve E + 1 + I1 zu ziehen. Der Schnittpunkt auf die Abszisse projeziert, ermoglicht die unmittebare Ablesung der gesuchten Entwicklungs- zeit. Wird statt dessen die Entwicklungsdauer eines bestimmten Lamenstadiums dlein bei einer gegebenen Temperatur gewiinscht, etwa die Zeitdauer von der 1. bis zur IT. Hautung bei 160 C, dann ist entsprechend zu verfahren mit dem Unterschied lediglich, da13 nunrnehr die Entfernung der beiden Schnittpunkte der Parallele mit den Eurven E + I, sowie E + I + I1 den gesuchten Wert ergibt.
Auf diese Weise laat sich auf graphischem Wege die Entwicklungs- dauer fur jedes beliebige Stadium von der Ablage des Eies an bis zur Erreichung der Verpuppungsreife feststellen, mit einer fur praktische Zwecke ausreichenden Genauigkeit. Die A bweichungen von den beobachteten Werten betreffen im allgemeinen nur Bruchteile eines Tages. Ausdruck- lich sei darauf hingewiesen, da13 die Kurvenschar auf Grund von Mittel- werten aufgestellt ist. Die Variationsbreiten der Entwicklungszeiten , die im ubrigen bei niederen Temperaturen nicht nur fur das Ei, sondern auch fur die Larven erheblich zunehmen, sind der Ubersichtlichkeit wegen aus dem Bilde fortgelassen. Die Kurvenschar gibt also das Ver- halten der Hauptmasse der Ejer bezw. Larven bezuglich der Entwicklungs- dauer wieder, auf die es ja in der Praxis allein ankommt.
Wie schon erwahnt, fuBen die hier mitgeteilten Beobachtungen auf Untersuchungen bei kons t an te r Temperatur. Ihre Ubertragnng auf die schwankenden Warmeverbaltnisse des Freilandes ist nur bedingt zulassig. I m einzelnen gelten hierfur die bereits oben bai Besprechung der Ei- entwicklung ausgefuhrten Uberlegungen. -
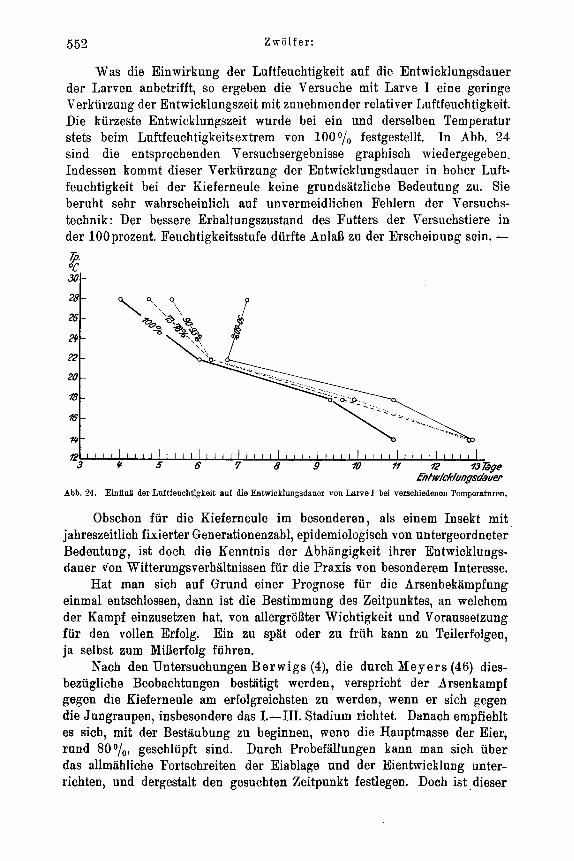
552 ZIT o I f er:
Was die Einwirkung der Luftfeuchtigkeit auf die Entwicklungsdauer der Larven anbetrifft, so ergeben die Versuche mit Larve I eine geringe Verkurzung der Entwicklungszeit mit zunehmender relativer Luftfeuchtigkeit. Die kurzeste Entwicklungszeit wurde bei ein und derselben Temperatur stets beim Luftfeuchtigkeitsextrem von 100 o/o festgestellt. In Abb. 24 sind die entsprechenden Versuchsergebnisse graphisch wiedergegeben. Indessen kommt dieser Verkurzung der Entwicklungsdauer in hoher Luft- feuchtigkeit bei der Kieferneule keine grundsatzliche Bedeutung zu. Sie beriiht sehr wahrscheinlich auf unvermeidlichen Pehlern der Versuchs- technik : Der bessere Erhaltungszustand des Futters der Versuchstiere in der 100prozent. Peuchtigkeitsstufe durfte AnlaB zu der Erscheinung sein. - @. OC
28
2 6
24 -
22
20
W~
76 .
14
RL 3 I I I I 1 I I I I 5 6 ' I I I ' ' I 7 I I I I 8 ' I I I 9 ' I I I m I I ' I I1 ' ' I I Q ' I I I I 73Gge ' Entwibkhgsdauer
Abb. 24. EinfluD der Luftfeucht:gkeit a d die Entwicklungsdauer von Lawe I bei versehiedenen Temperaturen.
Obschon fur die Kieferneule im besonderen, als einem Insekt mit jahreszeitlich fixierter Generationenzahl, epidemiologisch von untergeordneter Bedeutung, ist doch die Kenntnis der Abhangigkeit ihrer Entwicklungs- dauer von Witterungsverhaltnissen fur die Praxis von besonderem Interesse.
Hat man sich auf Qrund einer Propose fur die Arsenbekampfung einmal entschlossen, dann ist die Bestimmung des Zeitpunktes, an welchem der Kampf einzusetzen hat, von allergroljter Wjchtigkeit und Voraussetzung fur den vollen Erfolg. Ein zu spat oder zu fruh kann zu Teilerfolgen, ja selbst zum Miljerfolg fuhren.
Nach den Untersuchungen Berwigs (4), die durch Meyer s (46) dies- bezugliche Beobachtungen bestatigt werden, oerspricht der Arsenkampf gegen die Kieferneule am erfolgreichsten zu werden, wenn er sich gegen die Jungraupen, insbesondere das 1.-111. Stadium richtet. Danach empfiehlt es sich, mit der Bestaubung zu beginnen, wenn die Hauptmasse der Eier, rund SOO/,,, geschlupft sind. Durch Probefallungen kann man sich uber das allmahliche Fortschreiten der Eiablage und der Eientwicklung unter- richten, und dergestalt den gesuchten Zeitpunkt festlegen. Doch ist dieser

Studien zur Okologie und Epidemiologie der Imekten. 553
Weg umstandlich und nicht absolut zuverlassig. ZweckmaBig erscheint es, den fraglichen Zeitpunkt durch Verfolgung des Temperaturverlaufes im Freien unter Berucksichtigung der im vorstehenden dargestellten Ent- wicklungsdauerkurven zu bestimmen, bezw. wenn die Wetterlage den Temperaturverlauf einigermaBen vorauszusehen gestattet, den fraglichen Zeitpunkt an Hand der Kurren sohatzungsweise zu ermitteln.
Um brauchbare Unterlagen fur die Haoptzeit des Falterfluges, der Eiablage usw. zu gewinnen, wird am einfachsten die von Meyer benutzte Methode angewendet: Aussetzen einer bestimmten, ku rz zuvor ein- gesammelten Anzahl gesunder Puppen unter moglichst naturgetreuen Be- dingungen im Walde an einem durch Holzleisten und Drahtgaze ab- geschlossenen Raum. Durch tagliches Absammeln der hier schliipfenden Falter IaBt sich der Zeitpunkt, an dem die Hauptmasse der Falter aus- gekommen ist (rand SOO/,,) ohne Schwierigkeit ermittelo. Berucksichtigt man nuu das Seite 519 und 522 dieser Arbeit uber die Temperaturabhiingigkeit von imaginaler Lebensdauer und Eiablageperiode Gesagte, dann ist auch der Zeitpunkt, bis zu dem die Hauptmasse der Eier abgesetzt worden ist, leicbt und fur praktische Zwecke ausreichend genau bestimmbar. Das weitere uber die Dauer der Eientwicklung ergibt sich, wie bereits erwahnt, aus dem Gang der Temperatur im Freien unter Berucksichtiguug der Entwicklungsdauerkurven. -
VI. Hunger- und Fiitterungsversache. Uber die Hungerfahigkeit frisch geschlupfter Eiraupen galt bislang,
daB derartige Larven im Maximum 3 Tage ohne Nahrungsaufnahme zu leben vermogen. Die Anschauung geht auf Beobachtungen von Wolff und KrauBe (74) sowie Sach t l eben (55) zuruck.
Mit Rucksicht auf das Unvermogen der Eiraupe, vorjahrige Kiefern- nadeln zu befressen - was ubrigens auch noch fur den Einhauter gilt! -- haben die beiden erstgenannten Autoren die erwiihnte Beobacbtung im epidemiologischen Sinn zu deuten versucht: Nach ihnen wurde in ,,nor- malen" Jahren die Mehrzahl der Eiraupchen aus Nahrungsmangel zugrunde gehen, da sie nach Annahme der Genannten in solchen zu einem Zeit- punkt schlupfen, an den] die Kiefernknospen noch nicht entfaltet sind. Demgegenuber sollen Jahre der Gradation durch ,,verspateten Falterflug" und in Zusammenhang damit durch ein gunstigeres Zusammentreffen des Eiraupenschlupfens mit dem Entwicklungsgang der Kiefernknospen aus- gezeichnet sein.
Nun haben Sach t l ebens (55) Untersuchungen bereits ergeben, daB die Jnngraupen die Kiefernknospen auf einem weit fruheren Entwicklungs- zustand zu befressen imstande sind, als dies von Wolff und KrauBe an- genommen worden ist, und so folgert auch dieser Autor, ,,da13 io Jahren mit normalem Witterungsverlauf die Forleulenraupe - zum mindesten

Temperatur 4" 8u
s s s % & m Relat. Luftfeuchtigkeit o
Minimum . . . . . 4 - - 3 4 2
Maximum . . . . . 131-1- 1119 16 Mittel . . . . . .
2 E I n E Z I n - 8,2 - - 7, l 4,9 3,O
14O 180 220 280
.o s z s s s s z s s a &.O s;: " I n 2 L* In 3 E I n 3 Z I n 2
2 1,5 I 1,5 l ,50,5 I 1,00.5 0,5 0,5 0,5 0,5 4,5 3,s P,1 2,s 2,2 0,'i 2.4 1,4 1,2 0.9 0,6 0,5 8 14 14 5 1 4 l1,5 I 3 12,512 1,511,511.5
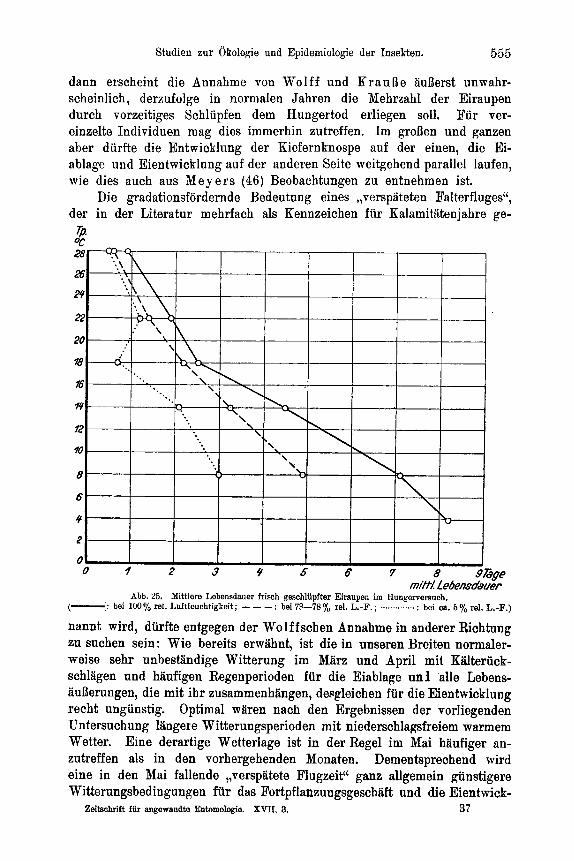
Studien zur Okologie und Epidemiologie der Insekten. 555
dann erscheint die Annahme von Wolff und KrauBe auBerst unwahr- scheinlich, derzufolge in normalen Jahren die Mehrzahl der Eiraupen durch vorzeitiges Schliipfen dem Hungertod erliegen soll. Piir ver- einzelte Individuen mag dies immerhin zutreffen. I m groBen und ganzen aber diirfte die Entwicklung der Kiefernknospe auf der einen, die Ei- ablage und Eientwicklung auf der anderen Seite weitgehend parallel laufen, wie dies auch aus Meyer s (46) Beobachtungen zu entnehmen ist.
Die gradationsfordernde Bedeutung eines ,,verspateten Falterfluges", der in der Literatur mehrfach als Kennzeichen fur Kalamitatenjahre ge-
@.
nannt wird, diirfte entgegen der Wolffschen Annahme in anderer Richtung zu suchen sein: Wie bereits erwahnt, ist die in unseren Breiten normaler- weise sehr unbestandige Witterung im Marz und April rnit Kalteruck- schlagen und haufigen Regenperioden fur die Eiablage uni alle Lebens- auBerungen, die mit ihr zusammenhangen, desgleichen fur die Eientwicklung recht ungunstig. Optimal waren nach den Ergebnissen der vorliegenden Untersuchung langere Witterungsperioden mit niederschlagsfreiem warmem Wetter. Eine derartige Wetterlage ist in der Regel im Mai haufiger an- zntreffen als in den vorhergehenden Monaten. Dementsprechend wird eine in den Mai fallende ,,verspatete Flugzeit" ganz allgemein giinstigere Witterungsbedingungen fur das Fortpflanzungsgeschaft und die Eientwick-
Zeitschrift fiir angewandte Entomologie. XVII, 3. 37

556 Z m o I f e r:
lung antreffen als eine solche, die im April oder gar im Marz stattfindet. Hierbei liegt somit ein d i r ek te r EinfluB der Witterung vor, und es ist keineswegs notwendig, eine indirekte Wirkung der Witterung zur Er- klarung des gradationsfordernden Einflusses verspateten Falterfluges heran- zuziehen, welch letztere der Wo If fschen Annahme zugrunde liegt. -
Es ist hinlanglich bekannt und oben schon erwahnt, daB das I. Larven- Stadium der Kieferneule in seiner Ernahrung auf Maitriebe angewiesen ist. Gelegentlich der Aufzucht des Versuchsmaterials lieBen sich die hier-
I Abb. 28. Eiraupe in Kaltestsrre.
uber bereits vorliegenden Beobachtungen bestatigen.
Bei den Zuchtversuchen mit kon- stanten Temeratnr- und Luftfeuchtigkeits- verhaltnissen wurden nun wiederholt Er- fahrungen gemacht, die darauf hindeuten, daB die Maitriebe, wenn sie als Nahrung fur Eiraupen noch in Prage kommen sollen, einen gewissen Entwicklungs- zustand nicht uberschritten haben durfen.
Wenn die Achse der Maitriebe zu verholzen beginnt und die Nadeln von der Basis an gemessea, eine Lange von ungefahr 18-20 mm erreicht haben, kommen sie - zum mindesten fur die Eiraupe als Putter kaum noch in Frage. So konnten in Zuchtversuchen bei kon- stanten Temperatur- und Feuchtigkeits- verhaltnissen im Mai mit jungen Trieben bezw. geschalten Knospen 60-700/0 und daruber von den schlupfenden Raupen
bis zur ersten Hautung gebracht werden, wahrend unter denselben gunstigen Temperatur-Peuchtigkeitsbedingungen im Juoi mit einem Putter, das aus Maitrieben in einem fortgeschrittenen Entwicklungszustand bestand, die Sterblichkeit der Eiraupen 80-100 O/,, erreichte.
Auf Grund dieser Beobachtungen ist zu vermuten, daB in der Natiir bei rascher Entwicklung der Maitriebe ein Teil derselben den Eiraupen ,,entwachst" und diese, soweit es sich um spatgeborene Individuen handelt, welche von Faltern abstammen, die Ende Msi bis Anfang Juni schlupften, schwerlich ihr erstes Hautungsstadium erreichen werden.
Es ist sehr wohl mogliah, daB das unterschiedliche Verhalten der Eiefernstangen- und Altholzer, von denen die ersteren zu Beginn einer Kalamitat meist erheblich stiirkeren Befall aufweisen, zum Teil mit dieser Erscheinung h Zusammenhang steht. Allerdings muBte hierzu noch der Nachweis erbrayht werden, daB die Maitriebe der Altholzer normalerweise rascher ,,ausreifen" als jene der Stangenholzer.

Studien zur Oliologie und Epidemiologie der Insekten. 557
Zussmmen fassung.
I. Der Begriff der , ,absoluten" und , , re la t iven" Zeugungskraft sowie
jener des Gle ichgewichtswiders tandes wird erortert und geaeigt, daB ,,absolute" Zeugungskraft und ,,Gleichgewichtswiderstand" als artspezifische GroBen aufgefal3t werden konnen.
Der Gesamtwiderstand W,, der auf die Generation einer Insektenart einwirkt, la& sich in Prozeut der Eizahlreduktion und vernichteter In- dividuen berechnen nach der Pormel:
100 (PI -Pz) + W,,P, PI
w - _ _ - .. - A -
wenn bekannt sind Ausgangspopulationsdichte PI und Endpopulations- dichte Pz, sowie der Gleichgewichtswiderstand W, fur die betreffende Art.
Die Populationsdichte P, zu Ende einer Generation ist bestimmbar alach der Gleichung (Populationsgleichung)
in der PI die Ausgangspopulationsdichte bedeutet, m : f das Geschlechter- verhaltnis, e die absolute Zeugungskraft der Art, wit w2, w,, . . . . . . Wn 7
endlich die Widerstandskomponenten, die auf die betreffende Generation einwirkten, gemessen in Prozent der Eireduktion und in Prozent des jeweils vernichteten Individuenanteiles der Population. - Theoretische Polgerungen und einige praktische Anwendunpmoglichkeiten der Gleichung werden besprochen.
11. Die absolute Eizahl der Kieferneule wird mit 190 ermittelt. Im Ver-
gleich mit niederen Temperaturen uben hohere lediglich einen be- schleunigenden EinfluB aul die Bildung legereifer Eier in den Ovarien der Weibchen aus. Die Gesamtproduktion iegereifer Eier wird hierdurch nicht beeinflult. Die untere Grenee wirksamer Temperaturen zur Er- langung der Schlupfreife bei der uberwinternden Puppe liegt zwischen 4- 80 C. Theoretische Erwagungen fiihren auf einen Grenawert von rund 6 0 C. - Der optimale Bereich der geschlechtlichen Aktivitat lie@ zwischen 12-160 C und goo/, bis schatzungsweise 6OO/, rel. Luftfeuchtigkeit, der entsprechende fur die Eiablage zwischen 14-190 C und ebensolchen Luftfeuchtigkeitsgraden. - Anhaltend extrem hohe Luftfeuchtiglteit von 100 o/,, gleichviel mit welcher Temperatur sie kombiniert wird, beein- trachtigt die ovariale Eientwicklung, die geschlechtliche Aktivitat, die Ge- samteiablage und die Lebensdauer der Imago erheblich.
37 *

Zwolfer : 558
111. Das d a l e Optimum fur das Ei ist relativ weit begrenzt und liegt
innerhalb eines Temperaturbereiches von 12 -22 0 C und Luftfeuchtigkeits- graden von 65-85 O / o . Gegen konstant feuchtigkeitsgesattigte Atmosphare, gleichviel mit welchen Temperaturen verbunden, erweisen sich die Eier auflerordentlich empfindlich. - Das vitale Optimum der Larve I ist wesent- lich enger begrenzt als jenes des Eies und liegt bei 17-180 C und 80 bis 90 O rel. Luftfeuchtigkeit. Das vitale Optimum der uberwinternden Puppe liegt bei 1000/o rel. Luftfeuchtigkeit und Temperaturen unterhalb 6 O C.
IV. An Hand der Versuchsergebnisse iiber die Abhangigkeit der Vitalitat
der praimaginalen Entwicklungsstufen von Temperatur und Luftfeuchtig- keit wird wahrscheinlich gemacht, da13 Larve I der Kieferneule dasjenige ihrer Entwicklungsstadien ist, welches die geringste okologische Valenz gegenuber Klimaeinflussen aufweist, und damit als das kritische Stadium im epidemiologischen Sinne anzusprechen ist. Unter Berucksichtigung von Meyers phanologischen Daten wird fur das mittelfrankische Gebiet im besonderen eine in die Zeit von Ende Mai bis Mitte Juni, meist aber in das erste Junidrittel fnllende Dekade als ,,kritische Zeit" erkannt und der Nachweis zu fuhren versucht, da13 hier wahrend der kritischen Zeit Niederschlagsmengen von rund 6 mm gradationsfordernd, hohere, von ca. 15-20 mm an aufwarts gradationshemrnend wirken.
V. Die Entwicklungsdauer der praimaginalen Stadien der Kieferneule
ist weitgehend von der jeweiligen Temperatur abhangig. Die Luftfeuchtig- keit spielt hierbei eine untergeordnete R o b . Die Temperaturabhangigkeit von der Ablage des Eies bis zur verpuppungsreifen Larve und jene der dazwischenliegenden Entwicklungsstufen wird in Form einer Kurven- schar eur Darstellung gebracht, die ermoglicht, durch graphische Inter- polation fiir jede konstante Temperatur die zugehorige Entwicklungsdauer naherungsweise zu bestimmen.
VI. Hungerversuche mit frisch geschliipften Eiraupen ergeben, da13 deren
Lebenadauer wesentlich von Temperatur und Luftfeuchtigkeit abhangig ist. Der maximale Wert wurde bei 4O C und 100°/o rel. Luftfeuchtigkeit mit 13 Tagen erreicht. - Larve I der Kieferneule ist in ihrer Ernahrung auf Maitriebe angewiesen, die einen bestimmten Entwicklungszustand nicht iiberschritten haben durfen.

Stndien zur Okologie und Epidemiologie der Insekten. 559
Literatnr. 1. Andersen , K. Th., Der EinfluB der Temperatur und der Luftfeuchtigkeit auf die
Dauer der Eizeit. (I. Beitrag zu einer exakten Biologie des linierten GrauriiBlers Sitonia Zineatn L.) Zeitschr. Morph. u. Okol. d. Tieve 17, 1930, S. 650-676.
2. Bachmetjew, P., Experimentelle entomologische Studien. Bd. 2. Sophia 1907. 3. Bermig, W., Die Forleule in Bayern (Histoiisch - klimatologische Betraohtung).
Forstwirtsch. Zentrlbl. 70, 1926, S. 165-181, 205-217, 259-265, 294-4297, 318 bis 328.
4. - - Laboratoriumsversuche zur Bionomie und Bekampfung der Forleule. Zeitschr. ang. Entom. 17, 1931, S. 563-586.
5. Berwig, W., und L e h n e r , Der EulenfraId 1923/24 und seine Folgen in der fiirst- lich-Hohenzollernschen Oberforsterei Griesel (Ncumark). Allg. Forst- u. Jagdztg. 105,
6. Blunck, H., Die Entwicklung des Dytiscus marginalis L. vom Ei bis Imago. 2. Die Metamorphose. Zeitschr. wiss. Zool. 121, 1923, S. 171-391.
7. - - Der Massenwechsel der lnsekten und seine Ursachen. 4. Wanderversamm- lung deutscher Entomologen in Kiel 1930, S. 19-41. Berlin-Dahlem 1930.
3, Bodenheimer , F. S., fjber die Voraussage der Generationenzahl von Insekten. 111. Die Bedeutung des Klimas fiir die landwirtsohaftliche Entomologie. Zeitschr. ang. Entom. 12, 1926, S. 91-122.
9. - - Welche Faktoren regnlieren die Individuenzahl einer Insektenart in der Natur. Biol. Zentrlbl. 48, 1928, 5. 714-739.
10. - - und Mi t a r b e i t e r , Epidemiologische, okologische und physiologische Studien iiber die afrikanische Wanderheuschrecke Schistocerca gregaria Forsk. Zeitschr. ang. Entom. 15, 1929, S. 435-557.
11. - - nber die Grundlagen einer allgemeinen Epidemiologie der Insektenkdamitiaten. Zeitschr. ang. Entom. 16, 1930, S. 433-450.
12. - - Die Schadlingsfauna Paliistinas. Berlin 1930. 13. - - Der Massenwechsel im Tierreich. Fomchungen und Fortschritte. Berlin 1931.
Bd. 7, S. 10. 14. B r e m er, H., Grundsatzliches iiber den Massenwechsel von Insekten. Zeitsch. ang.
Entom. 14, 1929, S. 254-272. 15. Chapman, R. K., Biotic potential, environmental resistance and insect abundance.
Cong. intern. Zool. X, Budapest 1927, S. 1209-1218. 16. - - The quantitative analysis of environmental factors. Ecology 9, 1928, S. 111
bis 122. 17. - - Quantitative results in the prediction of insects abundance on the basis of
biotic potential and environmental resistance. Journ. Ec. Entom. 21, 1928, S. 349 bis 352.
18. Dolles, Streifzug im Gebiet ron Feinden unserer schadlichen Waldinsekten. Forstl. naturw. Zeitschr. 6, 1897, 6. 257-270.
19. Ecks te in , F., Zoologisch-meteorologische Studien. 1. Mitteilung: fjber den EinfluB von Standort und Klima auf die Gradation des Kiefernspanners (Bupalus p~nkrius L.). Zoitschr. ang. Entom. 1923, S. 247-305.
20. Ecks t e i n , K., Bausteine zur Lebensgeschichte der Forleule. Zeitschr. ang. Entom. 10,
21. Eidmann, EL, Morphologische und physiologische Untersuchungen am weiblichen Genitalapparat der Lepidopteren. Zeitschr. ang. Entom. 15. 1929, S. 1-66.
1929, S. 416-428.
1924, S. 313-326.

560 Zwolfer:
22. E s c h e r i c h , K., und Baer , X., Tharandter zoologische Miszellen. Naturw. Zeitschr. Forst- u. Landw. 8, 1910, S. 164--168.
23. Escher ich , K., Forstinsekten Mitteleuropas Bd. I u. 11. 24. - - Kieferneulenkatastrophen und Fmstentomologie. Ber. uber die 21. Hauptvers.
Deutschen Forstver. Bamberg 1924. 25. - - Das neue Gesicht der Forstentomologie. Forstw. Zentrlbl. 52, 1930, S. 525
bis 546. 26. E s c h e r ich , G. U., Ein multipler Therniohygrostat mit standiger Lufterneuerung.
Anz. f. Schadlingsk. 6, 1930, S. 13-14. 27. F r i e d e r i c h s , K., Die Grundfragen und GesetzmaWigkeiten der Land- und forst-
wirtschaftlichen Zoologie Bd. I u. 11. Berlin 1930. 28. - - Zur Epidemiologie des Kiefernspanners. Ein Programm. Zeitschr. ang. Entom.
29. - - und S t e i n e r , P., Licht und Insektenentwicklung. Zeitschr. f. Bakt.. Parasitenk. usw. 80, 1930, S. 71-77.
30. Headlee , T. J., Some data on the effect of temperature and moisture on the rate of insect metabolism. Journ. Econ. Ent. 7, 1914, S. 413-417.
31. Hesse, R., Tiergeographie auf okologischer Grundlage. Jena 1924. 32. Horn, W., Einige Bedenken iiber neuere Anschauungen iiber Massenvermehrung
von Insekten. 4. Wanderversammlung Deutscher Entomologen in Kiel 1930, S. 43 bis 50. Berlin-Dahlem 1930.
33. J an isc h , E., Das Exponentialgesetz als Grundlage einer vergleichenden Biologie Berlin. Berlin 1927.
34. - - fiber die WirkungsgroSe der Umweltsfaktoren bei der Massenvermehrung von Insekten. Deutsche Forschung. Aus d. Arb. d. Notgem. d. Deutsch. Wissensch. Heft 9 : Landwirtschaftswissensch. 1929, S. 1--41.
35. - - Experimentelle Untersuchungen uber die Wirkung der Umwcltsfaktoren auf Insekten. I. Die Massenvermehrung der Baumwolleule Prodenia littoralis in Agypten. Zeitschr. Morph. n. Okol. d. Tiere 17, 1930, S. 339-416.
36. - - Einige Grundfragen der Insektenepidemiologie. Ber. VIII. Mitgliedervers. d. Deutschen Ges. f. ang. Entom.
37. J u d e i c h , J. F., und N i t s c h e , H., Lehrbuch der mitteleuropaischen Forstinsekten- kunde Bd. I u. 11. Wien 1895.
38. Mar t in i , E., Beobachtungen und Berechnungen zur Epidemiologie und Bekampfung der Malaria. Hamburg 1921.
39. - - Gedanken und Berechnungen iiber Vermehrung und Verbreitung der Lebe- wesen. Mit Anwendung auf die Stechmiickenfragen. Arch. f. Eydrobiol. 14, 1923,
40. - - Theoretisches zur Bestimmung der Lebensdauer von Schalingen auf Grund der Anopheles-Untersuchungen von Schuffner und von Grassi. Zeitschr. ang. Ent. 9, 1923. S. 133-146.
41. - - fiber die Warmesummenregel. 42. - - Beiti%ge zur medizinischen Entomologie nnd zur Malaria- Epidemiologie des
unteren Wolgagebietes. Hamburger Univ.- Abh. aus d. Geb. d. Auslandsk. Bd. 29, Reihe D, Bd. 3. Hamburg 1928.
43. - - uber Klima und Seuchen (vom Standpunkt des Entomologen). Anz. f. Schiid- lingsk. 5, 1929, S. 45-48.
44. - - Zur Gradationslehre. Ber. VIII. Mitgliedervers. d. Deutsch. Ges. f. ang. Entom. Anz. f. Schadlingsk. 6, 1930, S. 112-113.
45. Meteoro logische B e o b a c h t u n g e n in Bayern. In: Deutsches Met. Jahrbuch (1893-1913). Miinchen 1896-1914.
Berlin 1914/1923.
16, 1930, S. 197-205.
Anz. f. Schadlingsk. 6, 1930, S. 116-117.
S. 726-740.
Zeitschr. ang. Ent. 11, 1925, S. 301-305.

Studien zur dkologie und Epidemiologie der Insekten. 661
46. Me y e r , E., Untersuchungen zur Biologie und Bekampfung der Kieferneule Panolis flamnzen Schiff. Im Druck. Erscheint: Zeitschr. ang. Entom. 18, 1931.
47. NuBlin, O., und Rhumbler , L., Porstinsektenkunde 4. hufl. 48. P e a r l , R., The Biology of population growth. London 1926. 49. P i c t e t , A , Influence de la pression atmosphkrique sur le developpement des LQpi-
dopteres. 50. P i e r c e , W. D., A new interpretation of the relationship of temperature and humi-
dity to insect development. Journ. agr. Res. 5, 1916. 51. Ratzeburg , J. Th. Ch., Die Forstinsekten Bd. I (NachtrQe). 52. - - Die Porstinsekten Bd. 11. Berlin 1840. 53. - - Die Waldverderber. Bd. I. Berlin 1866. 54. ROB, R., The prevention of malaria. London 1910. 55. Sacht leben , H.. Beitriige zur Naturgeschichte der Forleule Panolis flammeu Schiff.
und ihrer Parasiten. Arb. Biol. Reichsanst. Land- u. Forstwirtsch. 15, 1927, S. 437 bis 536.
56. - - Die Forleule Pavzolis flammea Schiff. Monogr. z. Pflanzenschutz Bd. 3. Berlin 1929.
57. Sanderson , D. E., The relation of temperature to the growth of insects. Journ. Ec. Ent. 3, 1910, S. 113-140.
58. Schi lder , F. A., und Schi lder , M., Die Nahrung der Coccinelliden und ihre Be- ziehung zur Verwandtschaft der Arten. Arb. Biol. Reichsanst. f. Land- u. Forstwirtsch.
59. Schwerdt feger , F., Beobachtungen undUntersuchungen zur Biologie undBekampfung des Kiefernspanners wahrend des FraBjahres 1929 in der Letzlinger Heide. Zeitschr. f. Porst- u. Jagdw. 62, 1930, S. 65-166.
60. Sella , M . , in: Grass i , B., und Sel la , M., Seconda relazione della 10th antimalarica a Piumicino (Roma). Roma 1920.
61. Shel ford , V. E., Methods for the experimental study of the relations of insects to weather. Journ. Ec. Entom. 19, 1926, S. 251-261.
62. - - The relation of abundance of parasites to weather conditions. Journ. Ec. Entom. 19, 1926, 8. 283-289.
63. - - An experimental investigation of the relations of the codling moth to weather and climate. Bull. Ill. St. Nat. Eist. Surv. 16, 1927, S. 311-440.
64. - - Laboratory and field ecology. London 1929. 65. Sprengel , L., Uber Zustand und Entwicklnng der Eier in den Ovarien geschliipfter
Lepidopteren. 66. Thompson, W. R., La theorie mathbmatique de l’action des parasites entomophwes
et le facteur du hasard. Ann. FacultQ Sci. Marseille 2 nd. Ser, 11. pt., 8. 69-89. Marseille 1924.
67. - - A contribution to study of biological control and parasite introduction in con- tinental areas. Parasitology 20, 1928, S. 90--112.
68. Uvarov, B. P., Weather and climate in their relation to insects. Conf. of Empire meteorologists 1929. Agr. sect. London 1929.
69. -Volkel, E., Vorlaufige Mitteilung iiber den gesetzm5113igen Ablauf der Yassenver- mehrungen von Insekten. Nachrichtenbl. f. d. Deutschen Pllanzensch. 1930, 8. 44.
70. Volterra , V., Variations and fluctuations of the number of individuals in animal species living together, Journ. Cons. Intern. pour I’Explor. Xer., Val. 111, 1, 8. 3 bis 51, 1928.
71. - - Leqons sur la thQorie mathQmatique de la lutte pour la vie. Cahiers Sci. Fasc. 7. Paris 1931.
72. Weber, H., Biologie der Hemipteren. Berlin 1930.
Berlin 1927.
Arch. Sci. Phys. Eist. Wat. Genkve 44, 1917, S. 413-454.
S. 1183-1191. Berlin 1839.
16, 1928, S. 213-281.
Anz. f. Schadlingsk. Bd. 4, 1928, S. 25-30.

562 Z wolf er: Studien zur Okologie und Epidemiologie der Insekten.
73. W o l f f , M., und KrauBe, A,, Die forstlichen Lepidopteren. Jena 1922. 74. - - Die Erankheiten der Forleule und ihre prognostische Bedeutung fur die Praxis.
Breslau 1925. 7.5. Z w olfer, W., Untersuchungen zur Biologie und Bekampfung des Maiszunslers
(Pyruu-stu nubilulis Eb.) in Suddeutschland. 11. Teil. Arb. Biol. Reichsanst. f. Land- u. Forstuirtsch. 17, 1929, S. 460-498.
76. - - Beitrage zur Kenntnis der Schadlingsfauna Kleinasiens. I. Untersuchungen zur Epi6emiologie der ' Getreidewanze aryguster iategriwps Put. (Hem. Het.). Zeitschr. ang. Erztom. 17, 1930, S. 227-251.
77. - - Zur Theorie der Insektenepidemien. Biol. Zentrlbl. 50, 1930, S. 724-759.





























