Embed Size (px)
Citation preview

Z Lebensm Unters Forsch (t982) 175:321-326 Zeitschrift fur
Lebensmittel- Untersuchung
und-Forschung QJ. F. Bergmann Verlag 1982
Toxicity of Different Wheat Gliadins in Coeliac Disease*
Herbert Wieser 1, Gudrun Springer 1, Hans-Dieter Belitz 1, Azaria Ashkenazi 2, and Dalia Idar 2
1 Deutsche Forschungsanstalt ffir Lebensmittelchemie und Institut ffir Lebensmittelchemie der Technischen Universit/it M/inchen, LichtenbergstraBe 4, D-8046 Garching, Federal Republic of Germany
z Department of Pediatrics and Pediatric Research, Kaplan Hospital, Rehovot, affiliated to the Hadassah and Hebrew University Medical School, Jerusalem, Israel
Untersuchungen von Gliadinfraktionen des Weizens auf Ciiliakieaktivifiit
Zusammenfassung. Zur Oberprfifung der teilweise wi- dersprfichlichen Angaben in der Literatur fiber die c61iakieausl6sende Wirkung einzelner Gliadinfrak- tionen wird Weizenprolamin (Gliadin) durch Ionen- austauschchromatographie an Sulfopropyl-Sephadex C-50 in die Hauptfraktionen u-, fl-, 7- und co-Gliadin aufgetrennt. Die Gliadine werden durch PAG-Elektro- phorese, Aminosfiureanalyse, Amid-N-Bestimmung und fiber die nach partieller enzymatischer Hydrolyse erhaltenen Peptidrnuster in ihrer chemischen Zusam- mensetzung charakterisiert. Durch peptisch-tryptische Partialhydrolyse, Ultrafiltration und Gelchromatogra- phie an Sephadex G-50 werden aus den einzelnen Glia- dinen die Peptidfraktionen B2 und B3 gewonnen. Die Protein- und Peptidfraktionen werden in einem immu- nologischen Test (LIF-Test) bzw. in einem Organkul- fur-Test auf C61iakieaktivit/it untersucht.
Die Ergebnisse zeigen, dab die Peptidfraktionen durchweg gr6Bere Aktivit/it haben als die entsprechen- den Proteine, und dab yon den Peptidfraktionen aller Gliadine eine c61iakiespezifische Wirkung ausgeht, wo- bei die Aktivitfit mit der chemischen Zusammenset- zung der Gliadine korreliert ist.
Summary. To determine the toxic effect of different gliadins on coeliac patients, which has been variably as- sessed in the literature, wheat prolamines (gliadin) were separated into the main fractions a-, fi-, 7-, and co-glia- dins by chromatography on Sulfopropyl Sephadex C-50. The chemical compositions of the gliands were characterized by polyacrylamide gel electrophoresis, amino-acid analysis, determination of amide nitrogen and peptide maps.
* Supported by a grant from Stiftung Volkswagenwerk
The peptide fractions B2 and B3 were isolated from the gliadins by a peptic tryptic digestion, ultrafiltration and gel filtration on Sephadex G-50.
The gliadins and the peptide fractions were exam- ined for coeliac activity by immunological tests (LIF tests) and by organ-culture tests.
The results show that the peptide fractions are gen- erally more active than their respective gliadins, The peptide fractions of all gliadins have a coeliac-specific toxic effect; their activities correlate with the chemical composition of the gliadins.
Introduction
Coeliac disease, or gluten-sensitive enteropathy, is known to be caused by the prolamines of wheat, rye and barley, but the toxic effect of oat prolamines re- mains controversial [1]. Wheat prolamines (gliadin) consist of many components [2], which can be grouped into four main fractions (~-, fl-, 7-, o)-gliadins) accord- ing to their electrophoretic mobility [3].
In the literature the toxic effect of the different glia- dins to coeliac patients has been variably assessed. Some authors [4-6] have suggested that c~-gliadin bears the toxic factor. Jos et al. [7] found that 7-gliadin and its peptide fractions were as toxic as ~-gliadin, 7-gliadin was less toxic, and co-gliadin was not toxic at all.
To clarify these contradictions the different gliadins and their respective peptide fractions were examined for coeliac activity with the aid of an immunological test, the LIF test (LIF = leucocyte migration inhibition factor). In previous work [8-10] the LIF test has been proved as a specific test not only for the diagnosis of coeliac disease, but also for the examination of the anti- genic effect of peptide fractions from gluten. Coeliac patients on a normal diet, on a gluten free diet and con- trols were included in the tests. Additionally some tests with organ cultures of intestinal mucosa were per- formed.
0044-3026/82/0175/0321/$1.20
322 H. Wieser et al.: Toxicity of Different Wheat Gliadins
Methods
Preparation of Gliadin
200 g wheat flour (type 630) from the pure strain "Kolibri" was twice dispersed in 600 ml of 40°-60 ° light petroleum and stirred for 30 min. The solvent was removed and the residue was dried on a filter paper. 50 g of the defatted flour were stepwise extracted with 3 x 200 ml of 0.4 M sodium chloride and 3 x 200 ml of 70% aqueous ethanol by homogenizing with an "Ultra-Turrax' , in a centrifuge bottle, for 5 min. Each suspension was centrifuged for 20 rain at 4,000 rpm and 4 °C and the supernatant liquid was decanted. The three ethanol ex- tracts were combined, dialyzed against tap water and 0.01 M acetic acid and freeze-dried.
Ion-exchange Chromatography of Gliadin
The chromatography of gliadin by cation exchange was performed according to the technique used by Charbonnier and Moss6 [10].
Conditions
Column: 5 c m x 90 cm - temperature: 22 °C - cation exchanger: Sul- fopropyl Sephadex C-50 (Pharmacia) - sample: 3 g of gliadin, dis- soved in buffer A elution solvents: 1,200 ml of buffer A: 2 M urea, 0.04 M ethylenediamine, 0.08 M HCI, acetic acid pH 3.1.1,200 ml of buffer B: 0.5 M CH3COONa solution in buffer A without acetic acid
- flow-rate: 50 ml x h - ~-fraction size: 12 ml - detection: trans- mission at 280 nm.
The fractions were combined as shown in Fig. 1, dialyzed against tap water and 0.01 M acetic acid and freeze-dried.
Preparation of the Protein Digests
The gliadins were successively digested by pepsin and trypsin as de- scribed by Frazer et al. [11]. After adjustment to pH 4.5, the digests were centrifuged and the supernatants liquids were freeze-dried.
Ultrafiltration
The soluble portions of the partial hydrolysates were fractionated by ultrafiltration with an U M 10 membrane (Amicon Ltd.) at 4 °C ac- cording to the procedure used by Dissanayake et al. [12] and the re- tentates (fractions B) were freeze-dried.
Gel Filtration
Gel filtration of the fractions B was performed under the following conditions: Column. 2.6 cm x 95 cm - temperature: 4 °C - gel: Sephadex G-50 fine (Pharmacia Ltd.) - sample: ~- 50 mg of the retentates from the ultrafiltration, dissolved in 2.0 mt of 0.1 M acetic acid - elution sol- vent: 0.1 M acetic acid - flow-rate: 20 ml x h - 1 _ fraction size: 5 ml
- detection: development of 0.1 ml aliquot with ninhydrin reagent af- ter alkaline hydrolysis (2 M NaOH, 15 min, 100 °C) and measure- ment of the absorbance at 575 nm. The eluates were combined into the fractions B4-B1 and freeze-dried.
Amino-acid Analysis
200 ~tg of the freeze-dried gliadins and peptide fractions were hy- drolyzed with 3 ml of 6 M HC1 at 110 °C under vacuum. The samples were dried over KOH pellets in a vacuum desiccator and were then redissolved in 1.00 ml of 0.2 M sodium citrate buffer (pH 2.2). From these solutions 0.4 ml were analysed using a Beckman "Multichrom" amino-acid analyser.
Determination of Amide Nitrogen
The amide nitrogen content of the gliadins was determined according to [13].
PA G-electrophoresis
The ultra-thin-layer electrophoresis of the gliadins on polyacryl- amide gel was performed as described by Gt rg et al. [14] with the usual conditions for the electrophoresis of gliadins [15].
LIF-test and Organ-culture Test
The examinations of the gliadins and their peptide fractions for coeliac activity were performed according to [16].
Results
In previous work [7, 8] two peptide fractions (B2, B3), which were obtained from gluten by peptic tryptic di- gestion, ultrafiltration and gel filtration, showed a coeliac specific effect in the LIF test. Therefore the comparative studies on the toxic effect of the individual gliadins (~-,/~-, 7-, ~o-gliadins) were similarly performed.
Separation and Character&ation of the Gliadins
Wheat prolamines (gliadin) were prepared from the de- fatted flour of the pure strain "Kolibri" by extraction with 70% ethanol after the albumins and globulins had been separated. The yield of dialyzed and freeze-dried gliadin was 5.6 g per 100 g of flour.
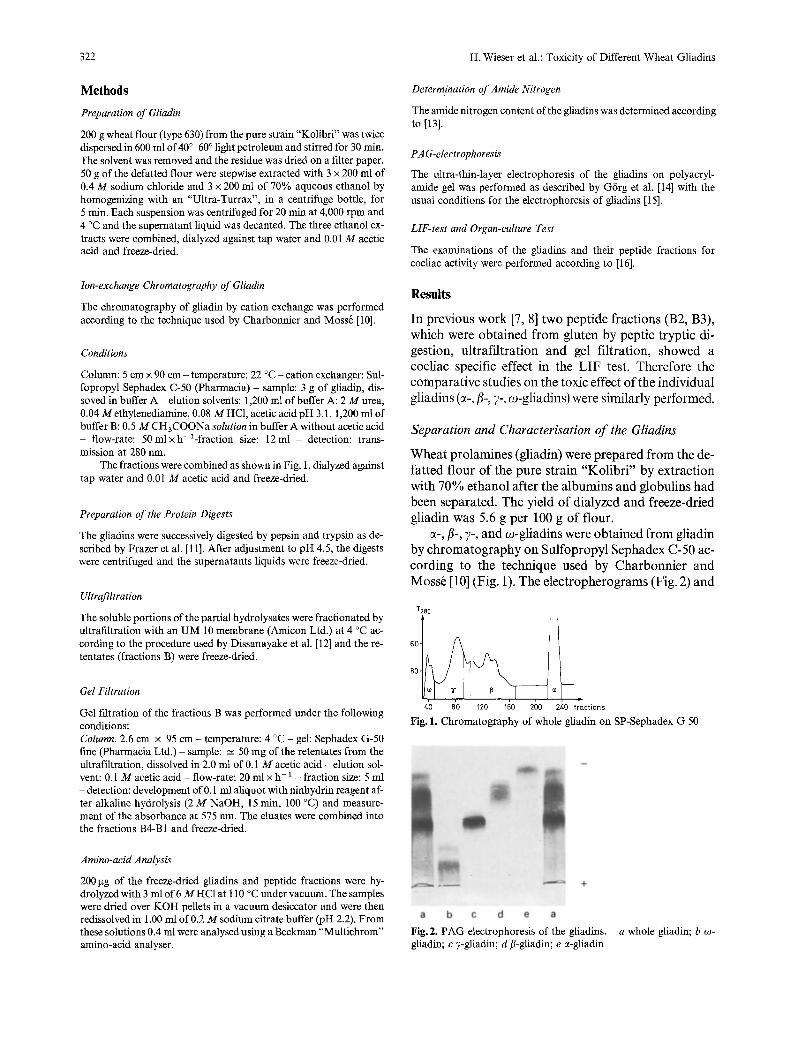
~-,/3-, 7-, and co-gliadins were obtained from gliadin by chromatography on Sulfopropyl Sephadex C-50 ac- cording to the technique used by Charbonnier and Moss6 [10] (Fig. 1). The electropherograms (Fig. 2) and
T280
g0-
~t
40 80 120 160 200 240 fractions
Fig. 1. Chromatography of whole gliadin on SP-Sephadex G 50
Fig. 2. PAG electrophoresis of the gliadins. - a whole gliadin; b co- gliadin; c 7-gliadin; d fl-gliadin; e ~-gliadin
H. Wieser et al.: Toxicity of Different Wheat Gliadins
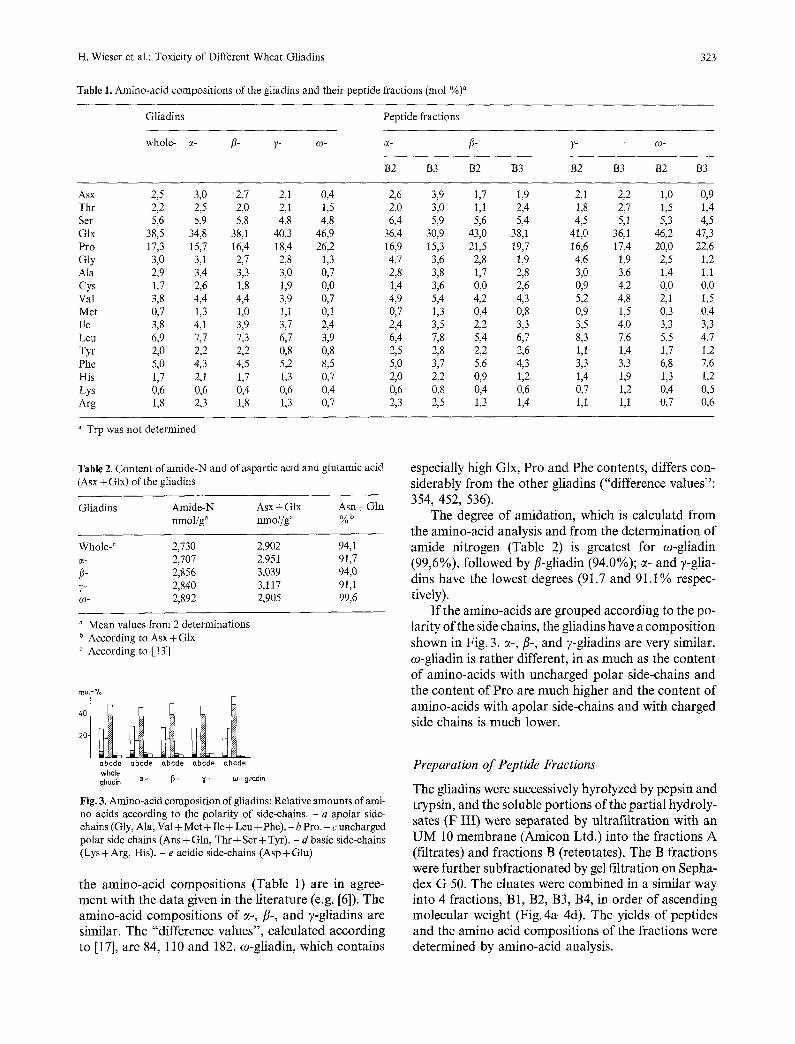
Table 1. Amino-acid compositions of the gliadins and their peptide fractions (tool %)"
323
Gliadins Peptide fractions
whole- ~- fl- 7- ~o- g- fi- 7" co-
B2 B3 B2 B3 B2 B3 B2 B3
Asx 2,5 3,0 2,7 2,1 0,4 2,6 3,9 Thr 2,2 2,5 2,0 2,1 1,5 2,0 3,0 Ser 5,6 5,9 5,8 4,8 4,8 6,4 5,9 Glx 38,5 34,8 38,1 40,3 46,9 36,4 30,9 Pro 17,3 15,7 16,4 18,4 26,2 16,9 15,3 Gly 3,0 3,1 2,7 2,8 1,3 4,7 3,6 Ala 2,9 3,4 3,3 3,0 0,7 2,8 3,8 Cys 1,7 2,6 1,8 1,9 0,0 1,4 3,6 Val 3,8 4,4 4,4 3,9 0,7 4,9 5,4 Met 0,7 1,3 1,0 1,1 0,1 0,7 1,3 Ile 3,8 4,1 3,9 3,7 2,4 2,4 3,5 Leu 6,9 7,7 7,3 6,7 3,9 6,4 7,8 Tyr 2,0 2,2 2,2 0,8 0,8 2,5 2,8 Phe 5,0 4,3 4,5 5,2 8,5 5,0 3,7 His 1,7 2,1 1,7 1,3 0,7 2,0 2,2 Lys 0,6 0,6 0,4 0,6 0,4 0,6 0,8 Arg 1,8 2,3 1,8 1,3 0,7 2,3 2,5
1,7 1,9 2,1 2,2 1,0 0,9 1,1 2,4 1,8 2,7 1,5 1,4 5,6 5,4 4,5 5,1 5,3 4,5
43,0 38,1 41,0 36,1 46,2 47,3 21,5 19,7 16,6 17,4 20,0 22,6
2,8 1,9 4,6 1,9 2,5 1,2 1,7 2,8 3,0 3,6 1,4 1,1 0,0 2,6 0,9 4,2 0,0 0,0 4,2 4,3 5,2 4,8 2,1 1,5 0,4 0,8 0,9 1,5 0,3 0,4 2,2 3,3 3,5 4,0 3,3 3,3 5,4 6,7 8,3 7,6 5,5 4,7 2,2 2,6 1,1 1,4 1,7 1,2 5,6 4,3 3,3 3,3 6,8 7,6 0,9 1,2 1,4 1,9 1,3 1,2 0,4 0,6 0,7 1,2 0,4 0,5 1,3 1,4 1,1 1,1 0,7 0,6
Trp was not determined
Table 2. Content of amide-N and of aspartic acid and glutamic acid (Asx + Glx) of the gliadins
Gliadins Amide-N Asx + Glx Asn+ Gln nmol/g" nmol/ga % b
Whole -c 2,730 2,902 94,1 c~- 2,707 2,951 91,7 fl- 2,856 3,039 94,0 7- 2,840 3,117 91,1 co- 2,892 2,905 99,6
Mean values from 2 determinations b According to Asx + Glx c According to [13]
mot-%
abcde abcde abcde abcde abode whore g[iodin 0¢- 13 - "I"- w- gtiodin
Fig. 3. Amino-acid composition of gliadins: Relative amounts of ami- no acids according to the polarity of side-chains. - a apolar side- chains (Gly, Ala, Val + Met + Ile + Leu + Phe). - b Pro. - c uncharged polar side chains (Ans+ Gln, Thr + Ser + Tyr). - d basic side-chains (Lys + Arg, His). - e acidic side-chains (Asp + Glu)
the amino-acid compositions (Table 1) are in agree- ment with the data given in the literature (e.g. [6]). The amino-acid compositions of c~-, fl-, and v-gliadins are similar. The "difference values", calculated according to [17], are 84, 110 and 182. co-gliadin, which contains
especially high Glx, Pro and Phe contents, differs con- siderably from the other gliadins ("difference values": 354, 452, 536).
The degree of amidation, which is calculatd from the amino-acid analysis and from the determination of amide nitrogen (Table 2) is greatest for o)-gliadin (99,6%), followed by fl-gliadin (94.0%); c~- and ?-glia- dins have the lowest degrees (91.7 and 91.1% respec- tively).
If the amino-acids are grouped according to the po- larity of the side chains, the gliadins have a composition shown in Fig. 3. ~-, fl-, and v-gliadins are very similar. co-gliadin is rather different, in as much as the content of amino-acids with uncharged polar side-chains and the content of Pro are much higher and the content of amino-acids with apolar side-chains and with charged side chains is much lower.
Preparation of Peptide Fractions
The gliadins were successively hyrolyzed by pepsin and trypsin, and the soluble portions of the partial hydroly- sates (F III) were separated by ultrafiltration with an UM 10 membrane (Amicon Ltd.) into the fractions A (filtrates) and fractions B (retentates). The B fractions were further subfractionated by gel filtration on Sepha- dex G 50. The eluates were combined in a similar way into 4 fractions, B1, B2, B3, B4, in order of ascending molecular weight (Fig. 4a-4d). The yields of peptides and the amino acid compositions of the fractions were determined by amino-acid analysis.

324
A575
1 '2 t
0 , 4
,7
a)
40 80 fractions
[ I ~ d)
Fig. 4 a~l. Chromatography of the fractions B from the ultrafiltrati- on on Sephadex G 50. - a = c~-gliadin; b = fi-gliadin; c = ?-gliadin; d = co-gliadin
The recoveries from the gel filtrations range from 89.8 to 105.9%. The distributions of the peptide yields among the fractions B4-B1 are similar (Table 3). The yields of the B4 fractions were generally the highest and those of B 1, with the exception of ?-gliadin, the lowest. With the exception of y-gliadin the yields of fractions B2 and B3 were of the order of 20%.
The distributions of the individual amino-acids among the fractions B4-B1 are given in Fig. 5. The di- stributions of several amino-acids are either similar (Lys, Met, His, Thr) or almost parallel (Tyr, Tle, Arg). Particularly c~- and fl-gliadins show a parallel distribu- tion of many amino-acids (Thr, Glx, Pro, Val, Ile, Tyr, Phe). co-Gliadin differs considerably from the other gliadins in the distribution of Ala, Cys, Val and Phe.
Within the gliadin components the fractions B2 and B3 have similar amino-acid compositions (Table 1; Fig. 5), in most cases the fractions B4 and B1 are diffe- rent (Fig. 5).
H. Wieser et al.: Toxicity of Different Wheat Gliadins
Table 3. Gel filtration: yield of peptides (%)a
Gliadins Peptide fractions
B4 B3 B2 B1
Recovery
e- 55,6 17,6 18,0 8,9 105,9 /3- 41,2 23,5 21,9 13,4 98,1 7- 57,0 16,0 9,6 17,4 104,1 ~o- 51,9 20,6 18,3 9,2 89,8
a According to recovered peptides = 100
Examination of Coeliac Activity
The toxic effect of the gliadin components and their peptide fractions B2 and B3 were examined in an im- munological test (LIF test) and in an organ-culture test.
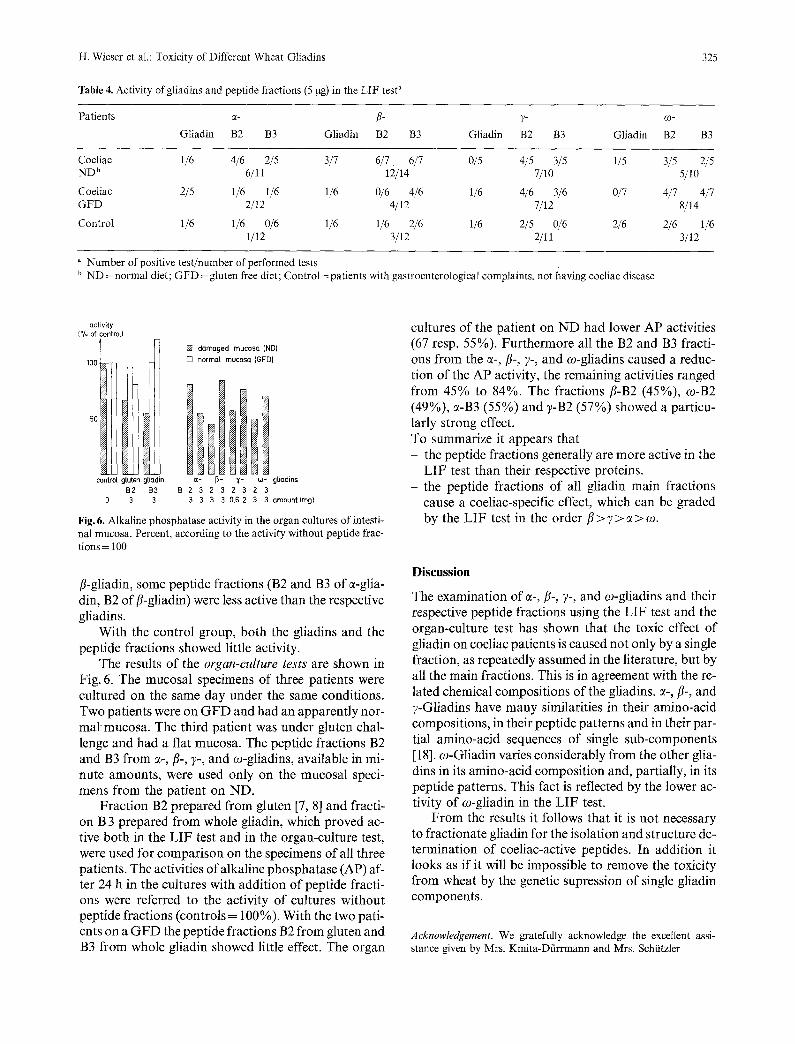
The results for the LIF tests, which are performed with patients on a normal diet (ND), patients on a glu- ten-free diet (GFD) and controls, are summarized in Table 4. Among the gliadins fl-gliadin was active for the patients on N D (3/7) 1 and c~-gliadin for the patients on GFD (2/5); the order gliadins did not show any coe- liac specific effect.
The peptide fractions B2 and B3 were generally mo- re active than the gliadins for patients on N D in the fol- lowing succession: fl > 7 > c~ > c0. With one exception (fl-gliadin) the B3 fractions showed somewhat less acti- vity in comparison to the B2 fractions.
As expected from previous studies [9], the patients on GFD responded less in most cases. The best respon- ses were obtained with B2 of ?-gliadin and with B3 of
1 Number of positive tests/number of performed tests
mo[-°/o Asx Thr Ser Met lie Leu
, , , , i , , , , . . . . , , , ,
G[x Pro Gty Tyr Phe His
. . . . . . .
AIo
I § i 3 2 i B
Cys Va[ Lys Arg
, , , , -~ , , , 3 2 1 B 4 3 2 1 1 3 4 3 2 1 g 4 : 3 2 i fro.ctions
Fig. 5. Distribution of amino-acids among the fractions B1-B4
H. Wieser et al.: Toxicity of Different Wheat Gliadins
Table 4. Activity of gliadins and peptide fractions (5 ~tg) in the LIF test a
325
Patients ~- /~- 7- co-
Gliadin B2 B3 Gliadin B2 B3 Gliadin B2 B3 Gliadin B2 B3
Coeliac 1/6 4/6 2/5 3/7 6/7 6/7 0/5 4/5 3/5 1/5 3/5 2/5 ND b 6/11 12/14 7/10 5/10
Coeliac 2/5 1/6 1/6 1/6 0/6 4/6 1/6 4/6 3/6 0/7 4/7 4/7 GFD 2/12 4/12 7/12 8/14
Control 1/6 1/6 0/6 1/6 1/6 2/6 1/6 2/5 0/6 2/6 2/6 1/6 1/12 3/12 2/11 3/12
a Number of positive test/number of performed tests b ND = normal diet; GFD = gluten free diet; Control = patients with gastroenterological complaints, not having coeliac disease
ectivity (% of controt)
control gluten g[iadin B2 B3
0 3 3
domaged mucosa (ND) D normal rnucosa (GFD)
t~- g[iadins B 2 3 2 3 2 3 2 3
3 3 3 3 0 . 6 2 3 3 ornount(rng)
Fig. 6. Alkaline phosphatase activity in the organ cultures of intesti- nal mucosa. Percent, according to the activity without peptide frac- tions = 100
/~-gliadin, some peptide fractions (B2 and B3 of c~-glia- din, B2 of/%gliadin) were less active than the respective gliadins.
With the control group, both the gliadins and the peptide fractions showed little activity.
The results of the organ-culture tests are shown in Fig. 6. The mucosal specimens of three patients were cultured on the same day under the same conditions. Two patients were on GFD and had an apparently nor- mal' mucosa. The third patient was under gluten chal- lenge and had a flat mucosa. The peptide fractions B2 and B3 from c~-,/3-, y-, and co-gliadins, available in mi- nute amounts, were used only on the mucosal speci- mens from the patient on ND.
Fraction B2 prepared from gluten [7, 8] and fracti- on B 3 prepared from whole gliadin, which proved ac- tive both in the LIF test and in the organ-culture test, were used for comparison on the specimens of all three patients. The activities of alkaline phosphatase (AP) af- ter 24 h in the cultures with addition of peptide fracti- ons were referred to the activity of cultures without peptide fractions (controls = 100%). With the two pati- ents on a GF D the peptide fractions B2 from gluten and B3 from whole gliadin showed little effect. The organ
cultures of the patient on ND had lower AP activities (67 resp. 55%). Furthermore all the B2 and B3 fracti- ons from the e-,/~-, 7-, and c0-gliadins caused a reduc- tion of the AP activity, the remaining activities ranged from 45% to 84%. The fractions fl-B2 (45%), co-B2 (49%), e-B3 (55%) and 7-B2 (57%) showed a particu- larly strong effect. To summarize it appears that - the peptide fractions generally are more active in the
LIF test than their respective proteins. the peptide fractions of all gliadin main fractions cause a coeliac-specific effect, which can be graded by the LIF test in the order/7 > 7 > c~ > co.
Discussion
The examination of e-, fl-, 7-, and co-gliadins and their respective peptide fractions using the LIF test and the organ-culture test has shown that the toxic effect of gliadin on coeliac patients is caused not only by a single fraction, as repeatedly assumed in the literature, but by all the main fractions. This is in agreement with the re- lated chemical compositions of the gliadins. ~-, fl-, and 7-Gliadins have many similarities in their amino-acid compositions, in their peptide patterns and in their par- tial amino-acid sequences of single sub-components [18]. co-Gliadin varies considerably from the other glia- dins in its amino-acid composition and, partially, in its peptide patterns. This fact is reflected by the lower ac- tivity of co-gliadin in the LIF test.
From the results it follows that it is not necessary to fractionate gliadin for the isolation and structure de- termination of coeliac-active peptides. In addition it looks as if it will be impossible to remove the toxicity from wheat by the genetic supression of single gliadin components.
Acknowledgement. We gratefully acknowledge the excellent assi- stance given by Mrs. Kmita-Dfirrmann and Mrs. Schfitzler

326 H. Wieser et al.: Toxicity of Different Wheat Gliadins
References
l. Kasarda DD (1975) Protein nutritional quality of foods and feeds. Friedman M (ed.) Vol. 1, Part 2, Dekker, New York p. 565
2. Wrigley CW, Shepherd KW (1973) Ann N Y Acad Sci 209:154 3. Hekkens WTJM, Haex AJCh, Willihagen RGH (1970) In: Coe-
liac disease. Proc. Int. Coeliac Symposium. Booth CC, DoMing RM (eds.) p, 11. Churchill Livingstone, Edinburgh
4. Kendall MJ, Schneider R, Cox PS, Hawkins CF (1972) Lancet ii: 1065
5. Kasarda DD, Qualset CO, Mecham DK, Goodenberger DM, Struder W (1978) In: Perspectives in eoeliac disease. McNicholl B, McCarthy CF, Fottrell PF (eds.) p. 55, Lancaster MTP Press Ltd
6. Jos J, Charbonnier L, Mougenot JF, Moss6 J, Rey J (1978) In: Perspectives in coeliac disease. McNicholl B, McCarthy CF, Fot- trell PF (eds.) p. 75, Lancaster MTP Press Ltd
7. Ashkenazi A, ldar D, Handzel TZ, Ofarim M, Levin S (1978) Lancet 1:627
8. Ashkenazi A, Levin S, Idar D, Rosenberg J, Handzel TZ (1980) Pediatr Res 14:776
9. Ashkenazi A, Levin S, Idar D, Or A, Barzilai N, Handzel ZT (1981) Lancet 1:914
10. Charbonnier L, Moss6 J (1980) J Sci Food Agric 31:54 11. Frazer AC, Fletcher RF, Ross CAC, Shaw B, Sammons HG,
Schneider R (1959) Lancet ii:252 12. Dissanayake AS, Jerrome DW, Offord RE, Truelove SC, White-
head R (1974) Gut 15:931 13. Wieser H, Seilmeier W, Belitz H-D (1981) Z Lebensm Unters
Forsch 173:90 14. G6rg A, Postel W, Westermeier R (1979) GIT Labormedizin
1:32 15. Platt SG, Kasarda DD (1971) Biochim Biophys Acta 243:407 16. Ashkenazi A, Idar D, Maimon M, Levin S, Wieser H, Springer
G, Belitz H-D: J Pediatr Gastroenterol Nutr 17. Wieser H, Seilmeier W, Belitz H-D (1980) Z Lebensm Unters
Forsch 170:17 18. Bietz JA, Hnebner FR, Sanderson JE, Wall JS (1977) Cer Chem
54:1070
Received May 12, 1982