Embed Size (px)
Citation preview

8 R. HA_~M:
Tabelle 6. Prii/ung der ~eproduzier- barkeit der Reduktionswerte von De- hydroascorbinsgure mittels Schwefel- wassersto// i~ber 18 Std bei pH 3,5. Konz.: Dehydroascorbins~iure etwa 2.10 -a tool/l, Puffer: 8,2 ml 0,1 n-Oxal- s~ure+2,5 ml 3% ige Na-acetatlSsung, Temperatur: 20 ° C; Volumen: i0 ml, Auftragungsvolumen: 10/A, Chromato-
graphiGpaPiGr: SS 2043bMG1.
Versuchs- Probe reihe
1 1 1 2 1 3 1 4 1 5
2 1 2 2 2 3 2 4 2 5 2 6
Verbrauch an Farbstoff*
nach der Reduktion
und papier- chromato-
graphischen Trennung
ml
2,98 3,00 2,99 2,97 3,01
2,75 2,74 2,71 2,73 2,75 2,73
* E~wa 2,25 • 10 -4 n-2,6-Dichlor- phenolindophGnollSsung.
Die Ergebnisse yon 2 Versuchsreihen zu 5 bzw. 6 P roben (Tab. 6), in denen jeweils gMche Mengen derselben Dehydroascorbins i iure lSsung mi~ Sehwefelwassers$off reduzier t wurden, be- weisen die Reproduz ie rbarke i¢ der Reduk t ions - wer~e unger den Bedingungen, wie Sie in der Arbe i t svorsehr i f t fesCgelegt sind.
B. A r b e i t s v o r s c h r i f t
In ein hSehstens 5 ml fassendes GefiiB werden nacheinander 0,129 g Na-acetat sowie 1 ml des Iiir die Ascorbins~urebestimmung hergestellten Extraktes (vgl. Mitt. I) Gingewogen, 15 min mit SchweMwasserstoff durchspfilt und das verschlossene Gef~B fiber Nacht bzw. wenigstens 12 Std vor Lieht geschfitzt aufbewahrt. ])ann werden davon wieder je nach Aseorbins£ure- gehM~ zwisehen 10 und 100/A auf das Chromatogramm aufgetragGn und welter WiG bei der Ascorbins~urebe- stimmung (I. Mitt.) verfahren. Die Differenz des As- corbins~urewertes zu dem hierbei erhaltenen Wert ent- sprieht dem Dehydroaseorbins~uregehalt.
Zusammen]assung
D a die Dehydroascorbinsi~ure nu r ind i rek t nach der R e d u k t i o n als Ascorbinsi~ure bes t imm- b a r is~, ha f t en ihr auch die gleichen Feh l e r wie der Ascorbinsgurebes~immung an, die sich aber durch Verwendung der in de r Mit t . I darge leg ten Verfahrensweise ve rmeiden lassen. Es wurde die
Verl/ iNichkei~ des gew/~hlten Reduk t ionsve r fah rens geprfif~ u n d fes~gestellt, daft d ie Oxals/~ure die Dehydroaseorb ins~ure u n t e r den gegebenen Bedingungen n ieh t reduzier t , wie dies yon SCH~IDT u n d STAU])~G~i~ mi tge te i l t wird.
tJber das Wasserbindungsverm~gen des S~iugetiermuskels X I . Mit te i lung
Ein f lu~ des M u s k e l - P l a s m a s u n d der in i h m e n t h a l t e n e n Mine ra l s to f f e au f das W a s s e r b i n d u n g s v e r m ( i g e n des R i n d e r m u s k e l s ~
Von R. HAMM
Mitieilung aus den* Institut /i~r Chemie und Physik der Bundesanstalt /iir Fleisch/orschung, Kulmbach**
Mit 9 Textabbfldungen
(Eingegangen am 2. Oktober 1961)
Fi i r alas Wasse rb indungsve rm6gen (WBV) des 1Yiuskels s ind zweifellos in der H a u p t s a c h e die s t ruk~urel len Pro~eine verantwor~l ich 1. Dies abe r bedeute~ niche, daft
* Abkiirzungen: WBV = WasserbindungsvermSgen; I.P. ---- isoelektriseher Punkt. ** Frgulein HEDWIG BREI-I~ danke ich ffir ihre geschickte Mitarbeit.
1 H A ~ , R. : Advanc. Food Res. 10, 355 (1960) ; hier sind auch die Definitionen ffir die Begriffe ,,WasserbindungsvermSgen" und ,,ttydratation" angegeben.

Uber das WasserbindungsvermSgen des S~ugetiermuskels. XI 9
die wasserlSsliehen, im Muskelsaf$ en~hal~enen Subs tanzen ffir das W B V bedeu- tungslos w~ren. I n Anbetraeh~ des s t a rken Einflusses yon Salzen ~uf das W B V des Fleisehes x is t im Gegen~eil zu erwarten, dal~ die Mineralstoffe des Sarkop lasmas ftir die Hydra ta~ ion der s~rukturel len Pro~eine reeh~ wesent l ieh sind. W a s die wasser- 15sliehen Muskelprote ine anbetr i f f t , so wurde die MSglichkei~ eines Zusammenhanges mi t dem Gehal$ des Muskels an diesen Stoffen und seinem W B V sehon in Erw/~gung gezogen ~. Bisher aber is t m a n der F rage naeh der Bedeu tung des Muskelp lasmas und seiner Bes tandtef le auf das W B V des Fleisehes exper imente l l noeh nich$ nachgegan- gen. Eine Bea rbe i tung dieses Prob lems ersehien mas u m so notwendiger , als m a n bei der Behand lung u n d Vera rbe i tung des Fleisehes wissen sollte, ob und in welehem AusmaI~ Saf tver lus te , wie sie e twa be im Auf t auen yon Gefrierfleisch oder aueh w/~hrend des Abh/~ngens auf t re ten, n icht nur zu einer Verminderung des Nghrs~off- gehal~es, sondern aueh zu einer Ver/~nderung des W B V und d a m i t der Fleisch- beschaffenheit ffihren.
Die vorl iegende Unte r suchung erfolgte in drei Schrit~en. Es wurden s tud ie r t : 1) Die Bedeu tung des ganzen Muskelsaf tes ffir das W B V ; 2) der Einfluf] der Pro te ine und n iedermolekula ren Bes tandte i le des Sarkop lasmas auf das W B V des Muskels ; 3) die W i r k u n g einzelner Minerals~offe des Muskelplasmas auf das W B V des Muskels. U b e r das spezielle Thema der W i r k u n g des Muskelsaf tes auf das W B V des Fleisches h inaus g ib t die vor l iegende Arbei* auch einige grunds/ t tzl iche In fo rma t ionen fiber die Weehse lwi rkung zwischen Muskeleiweil3 und anorganischen Ionen und ihre Bedeu- ~ung ftir H y d r a t a t i o n und Rigidi~/~ des Gewebes.
Methodik
Untersuchungsmaterial. Der yon Fe~f und Bindegewebe befreite M. longissimus dorsi 5 bis 7j/~hriger Ktihe der Handels~asse ]3, 5 Tage naeh dem Schlaehten.
ErschSpfende Extraktion des 3Iuskels. Rindermuskel wurde zweimal durch den ]~leischwolf gegeben und dann in der ~ehtfachen Menge seines Gewichts an eiskal~er 0,1 m-KCLLSsung kurz im Starmix homogenisiert. Das ttomogenat wurde 15 rain bei 0°C geriihr~ und 25 min bei 13000 U/min und 0°C zentrlfugiert. AnschlieBend wurde der Zentrifugenrficks~and dreimal - - bei zwei Versuchen fiinfmal - - mit en~ionisiertem Wasser in der gleichen Weise extrahiert. Der Gehal~ des drit~en Wasser-Ex~raktes an Trockeiisubstanz war in allen F/~llen gleich Null. Das in dieser Weise behandel~e, ~ast farblose Gewebe wird im folgenden als ,,extrahierter Muskel" bezeichnet. Es ist nicht IIur frei yon den niedermolekularen Bestandteflen des Muskelplasmas; dutch die anf/~ngliehe Ex~rak~ion mit KC1-LSsung wurden ihm auch die Plasmaproteine entzogen. Im extrahierten wie im frischen Muskel wurden Wassergehalt und pH-Wert bestimmt.
Ersetzen eines Teiles des Muskelplasmas dutch reines Wasser. 100 g l~indermuskel wurden zweimal dutch den Fleischwolf gegeben und 15 see im Starmix in 60 ml eiskaltem Wasser zer- kleinert. Zwei 40 g-Proben des Homogenats wurden bei 12000 U/min und 0°C zentrifugier~. Der Uberstand wurde abgegossen und sein Volumen gemesseii. Zum Zentrifugierrtiekstand der einen Probe wurde der abzentrifugierte Muskelsaft, zum Riicks~and der anderen Probe das gleiche Volumen an Wasser gegeben. ]~iickstand und Fliissigkeit wurden mit dem Spa~el grfindlich vermischt.
Dicdyse. 100 g des im Wolf zerkleiner~en Muske]s wurden in 60 ml eiskaltem Wasser 60 see im Starmix homogenisiert. E~wa 50 g des Homogenates wurden im Dialyseschlaueh unter lang- samer Umdrehung 20 S~d in der Kalte gegen entionisiertes Wasser dialysiert. Im Dialyse-Rfick- stand warden Wassergehalt und IO~-Wert bestimmt.
Ultrafiltration. Der dutch Pressen kleiner Muske]stiicke zwischen zwei Platten gewonnene Muskelprel~saft wurde bei einem Druck yon 8 kg/em 2 unter Sticks~off in einem Druckfiltrations- ger/£iG aus V2A-Stahl (Membranfil~er-Gesellschaft, GSt~ingen) durch ein eiweil]dich~es Ffl£er (,,Lsg. 60", Membranfiltergesellschaft, GSttingen) filtriert. Das Ultrafiltrat war klar und farblos; mit Triehloressigs/~ure entstand l~eine Trfibung.
HAMM, R. : ZR. S. 8, Anm. 1. 2 FuJIMAXI, M., u. Y. N~KAJI~A: J. agric, chem. Soe. Japan 32, 695 (1958).

10 R. I~L~¢:
Ge/riertrocknung. 2 g-Proben des normalen oder extrahierten 1, zerkleinerten ~uskels warden in Wggeglischen bei --20°C eingefroren und in der Anlage G 02 der Fa. Leybold (K..~ln) gefrier- getrocknet. Die Endtemperatur der Trocknung lag bei +25°C, das Endvakuum (Oldiffusions- pumpe) betrug 4× 10 -3 torr. Der Wassergehalt des lyophilisierten Gewebes lag zwischen 0,6 und 1,0%.
Die Trocknung Yon Muskelsaft und Ultrafiltraten effolgte unter den gleichen Bedingungen. Zur Herstellung yon doppelt konzentrierten Ultrafiltraten (und Muskelsaft) wurde das getroeknete )/[aterial mit der H~lfte der ihm entzogenen Wassermenge aufgenommen. Aus der klaren L6sung schieden sich bei den U1trafiltraten bei lingerem Stehen im Kfihlschrank schSne Kristalle ab, die sigh bei Erwirmen auf 40--50°C wieder vollst~ndig 15sten.
Vorbereitung der Proben zur Bestimmung des WJRV. 2 g dos zerkleinerten normalen oder extrahierten 35uskels wurden mit der angegebenen iVIenge an Fremdwasser versetzt. Im Falle des gusatzes yon Muskelsaft oder Ultrafiltrat enthielt das zugegebene Fremdwasser diejenige 3~enge an Muskelsa~t oder Ultrafiltrat, die der im normMen ~uskel enthaltenen und durch die Extraktion entzogenen Menge entsprach. Gegebenenfalls enthiel~ das Fremdwasser die zur p~-Einstellung notwendige Menge an ~aOH oder HC1 bzw. Milchs~ure und die zu priifenden Salze. Die Salze wurden stets in ge16ster Form zugesetzt, pH-Messung erfolg~e nach dem Vermisehen der Probe mit der Fliissigkeit vor und unmittelbar nach Bestimmung des WBV.
Die Anwendung lyophilisierter ~uskelproben gestattet es, dem Muskel LSsungen zuzusetzen (pH-Einstellung, Salze), olme seinen normMen Wassergehalt zu ~ndern. Die geffiergetroekneten Proben (vgl. oben) warden mit soviel Wasser vermiseht, wie dutch Trocknung entzogen worden war. Vor Messung des WBV lieB man die rehydratisierte Probe etwa 20 min bei +2°C stehen. ]3el EinsteHen eines bestimrnten p~-Wertes wurde zunichst ein Teil des Wassers zur Rehydratation zugemischt und erst dann die efforderliche Menge an S~ure oder Lauge zugefiigt. ])as gleiehe grit 1fir den Zusatz yon SalzlSsungen. Bei Zusatz yon Muskelsaft oder Ultrafiltrat unter gleichzeitiger pg-Einstellung mul]te, um nieht mehr Wasser zuzugeben als durch Trocknung entzogen war, der Salt bzw. das Ultrafiltrat in doppelt konzen~rierter ~orm (vgl. oben) angewendet werden.
Bestimmung des Wasserbindungsvermggens und der Ver/ormbar~eit (t~igiditSt). Die Bestimmung dos WBV ~ und der Ver~ormbarkeit (Rigiditit a) dos Muskels war schon friiher beschrieben worden. Bei der Bestimmung des WBV in den Zentrifugenrfieksti~nden (erschSpfend extrahiertes Fleisch) wurden zur Berectmung folgende Formeln angewendet:
1) Fremdwassermenge (X) ira Ext~raktionsrfickstand in % (bezogen au~ zusatzfreien Muskel) :
X = 100 (W~ - - W~) i o o - - w ~ '
wobei Wa WassergehMt des l~uskels und Wa Wassergehalt des Extraktionsriiekstandes bedeuten.
2) ~remdwassergehalt des Extraktionsriieks~andes (F~):
X . 100 - ~ I = - I O 0 + X "
(3) mg-Muskel (M~) in 300 mg des Extraktionsriiekst~ndes: ~1- 300
~g~ = 300 100
(4) ~remdwassergehMt des Extrak~ionsrfickstandes (F~) naeh Zumisehen yon Wasser (Wz) zum Rfickstand:
( ~ + W~)" 100 100 + Wz
(5) mg-Muskel (Mz) in 300 nag Mischung (Riickstand + Wz) :
~ " 300 ..3//2 = 300 100
Die weitere Auswertung effolgte in der frfiher angegebenen Weise ~.
Fiir das RehydratationsvermSgen des ]Suskeleiweil~es ist es ohne Belang, ob der Muskel in Gegenwart odor in Abwesenheit yon Sarkoplasma getrocknet wurde [l~M~¢, 1~. : Fleisehwirtsch. 14, 204 (1962)
GRAY, R., u. 1~. I-I~v~M: Diese Z. 105, 446 (1957). H A ~ , R.: Biochem. Z. 327, 149 (1956).
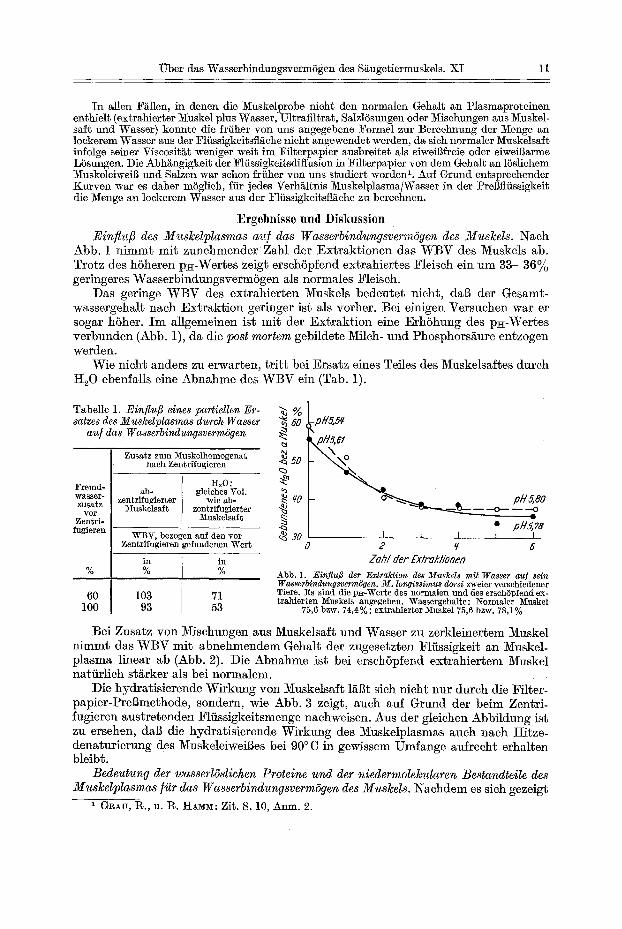
~Tber das WasserbindungsvermSgen des S~ugetiermuskels. XI 11
In allen F~llen, in denen die Muskelprobe nioht den normalen Gehalt an Plasmaproteinen enthielt (extr~hierter Muskel plus W~sser, Ultrafiltrat, Salzl6sungen oder Mischungen aus Muskel- salt und Wasser) konnte die ffiiher yon uns angegebene Formel zur Berechnung der Menge an lockerem Wasser aus der Fliissigkeitsfl~che nicht angewendet werden, d~ sich normaler Muskelsa~t infolge seiner Viscosit~t~ weniger weir im Fflterp~pier ausbreitet als eiweiBfreie oder eiweiBarme LSsungen. Die Abhi~ngigkeit der Fliissigkeitsdiffusion in Ffl~erpapier yon dem Gehalt an 15slichem MuskeleiweiB und S~lzen war schon ~riiher yon uns studiert wordenL Au~ Grund entsprechender Kurven w~r es daher mSglich, ffir jedes Verh~l~nis Muskelpl~sma/Wasser in der PreBfliissigkeit die Menge an lockerem Wasser ~us der Fliissigkeitsfl~che zu berechnen.
Ergebnisse und Diskussion Einflu[3 des Muslcelplasmas au] das WasserbindungsvermSgen des Muslcels. Nach
Abb. 1 nimmt mit zunehmender Zahl der Extraktionen das WBV des Muskels ab. Trotz des hSheren p~-Wel~es ze~gt ersch6pfend extrahiertes Fleisch ein um 33--36% geringeres WasserbindungsvermSgen als normales Fleisch.
Das geringe WBV des extrahierten Muskels bedeutet nicht, dab der Gesamt- wassergehalt nach Extraktion geringer ist als vorher. Bei einigen Versuchen war er sogar h6her. Im allgemeinen ist mi~ der Extraktion eine ErhShung des p~-Wertes verbvmden (Abb. 1), da die post mortem gebildete Milch- und Phosphors~ure entzogen werden.
Wie nicht anders zu erwarten, tritt bei Ersatz eines Teiles des Muskelsaftes durch H~O ebenfalls eine Abnahme des WBV ein (Tab. 1).
Tabelle 1. Einflufl eines partiellen Er- satzes des Muskelplasmas dutch Wasser
au] das Wasserbindungsverm6gen
Zusatz zum lV[uskelhomogenat nach Zentrffugieren
H20; Fremd- ab- gleiches Vol. wasser- zusatz zen~rifugierter wie ab-
~[uskelsaft zen~rifugierter vor Zentri- ]k~uskelsaft
fugieren WBV, bezo~ ~n auf den vor
Zen~rifugieren gefundenen Weft
%
6 0
100
in in % %
103 71 93 53
%
5O
~30 0
- .
8
• pH5,,78 I ] ~ I I I
2 q 6 Zahl der Extraktioflefl
Abb. 1. EinJluB der E~traktion des Muskels mit Wasser au] sein Wasserbindungsverm6gen. M. longissimus dorsl zweier verschiedener Tiere. Es sind die p~-Werte des normalen und des erschSpfend ex- trahierten ~uskels angegeben. Wassergehalte: Normaler Muskel
75,6 bzw. 74,4%; extrahierber ~aske175,6 bzw. 78,1%
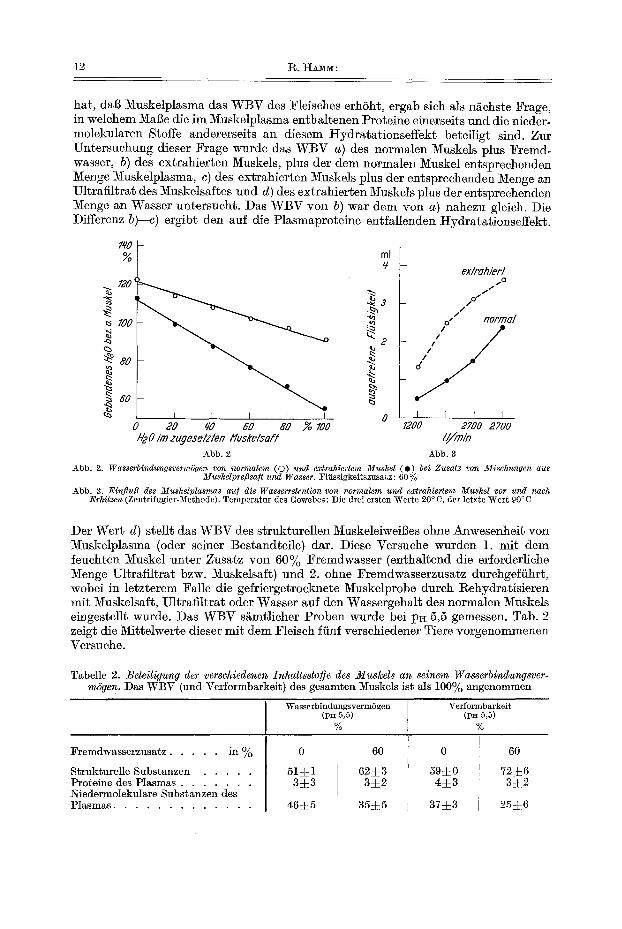
Bei Zusatz yon Mischungen aus Muskelsaft und Wasser zu zerkleinertem Muskel nimmt das WBV mit abnehmendem Gehalt der zugesetzten Fliissigkei~ an Muskel- plasma linear ab (Abb. 2). Die Abnahme ist bei erschSpfend extrahiertem Muskel natfirlich s*~rker als bei normalem.
Die hydratisierende Wirkung yon Mnskelsaft liiBt sich nicht nur durch die Filter- papier-Prel]methode, sondern, wie Abb. 3 zeigt, auch auf Grund der beim Zentri- fug4eren austretenden Fliissigkeitsmenge nachweisen. Aus der gleichen Abbildung ist zu ersehen, dal~ die hydratisierende Wirkung des Muskelplasmas auch nach Hitze- dena*urierung des Muskeleiweil3es bei 90°C in gewissem Umfange alffrecht erhalten bleibt.
Bedeutung der wasserI5slichen Proteine und der niedermolelcularen Bestandteile des Muslcelplasmas /i~r das Wasserbindungsverm6gen des Muskels. Nachdem es sich gezeigt
- - 1 GRAY, R., u. R. t t A ~ : Zi~. S. 10, Anm. 2.
12 R. H-~v~:
hat, dab Muskelplasma das WBV des Fleisches erh6ht, ergab sich als ngchste Frage, in welchem MaBe die im Muskelplasma enthaltenen Proteine einerseits und die nieder- molekularen Stoffe andererseits an diesem ttydratat ionseffekt beteiligt sind. Zur Untersuchung dieser Frage wurde das WBV a) des normalen Muskels plus Fremd- wasser, b) des extrahierten Muskels, plus der dem normalen Muskel en~sprechenden Menge Muskelplasma, c) des extrahierten Muskels plus der en~sprechenden Menge an Ultrafiltra~ des Muskelsaftes und d) des extrahierten Muskels plus der entsprechenden Menge an Wasser untersucht. Das WBV yon b) war dem yon a) nahezu gleich. Die Differenz b)--c) ergibt den auf die Plasmaproteine entfallenden tIydratationseffekt.
Iqo -
120 ( i
7OO
8O
8O
I 1 1 I I 0 go qo 60 80 ~ tOO H20 im zugesetzten Nuske/saft
Abb. 2
ml
3
2
extrahiert 1110
,o. / /
o" normal
J I I I I
1200 2?00 2700 Ll/m/n
Abb. 3
Abb. 2. WasserbindungsvermSgen von normalem (©) und ex~rahiertem Muskel ( e ) bei Zusatz yon Mischungen aus Mus/celpre[3sa]t und Wasser. Fliissigkei~szusatz: 60 %
Abb. 3. Einfiufl des Muskelplasmas au] die Wasserretention yon normalem und extrahiertem Muskel v o r u n d nach Erhitzen (Zentrifugier-Methode). Temperatur des Gewebes: Die drei ersten Werte 20°C, der letzte Wer~ 90°C
Der Wert d) stellt das WBV des strukturellen MuskeleiweiBes ohne Anwesenheit yon Muskelplasma (oder seiner Bestandteile) dar. Diese Versuche wurden 1. mi~ dem feuchten Muskel unter Zusatz yon 60% Fremdwasser (enthaltend die efforderliche Menge Ultrafiltrat bzw. Muskelsaft) und 2. ohne Fremdwasserzusatz durchgefiihr~, wobei in letzterem Falle die gefriergetrocknete Muskelprobe durch Rehydratisieren mit Muskelsaft, Ultrafiltrat oder Wasser auf den Wassergehal~ des normalen Muskels eingestellt wurde. ])as WBV s~mtlicher Proben wurde bei pE 5,5 gemessen. Tab. 2 zeig~ die Mittelwerte dieser mi~ dem Fleisch ffinf verschiedener Tiere vorgenommenen Versuche.
Tabelle 2. Beteiligung der verschiedenen Inhaltssto]e des Muskels an seinem Wasserbindungsver- m6gen. ])as WBV (and Vefformbarkeit) des gesamten Muskels is~ als 100% angenommen
~remdwasserzusatz . . . . . in ~o
Strukturelle Substanzen . . . . . Proteine des Plasmas . . . . . . . Niedermo]ekulare Substanzen des Plasmas . . . . . . . . . . . . .
Wasserbindungsverm6gen Verformbarkeit (PH 5,5) (pH 5,5)
% %
0 60
51=]=1 62±3 3±3 3=[_2
46±5 35~5
0
59±0 4±3
37~:3
6O
72~:6 3:~2
25:L6
Uber das WasserbindungsvermSgen des Sgugetiermuskels. XI 13
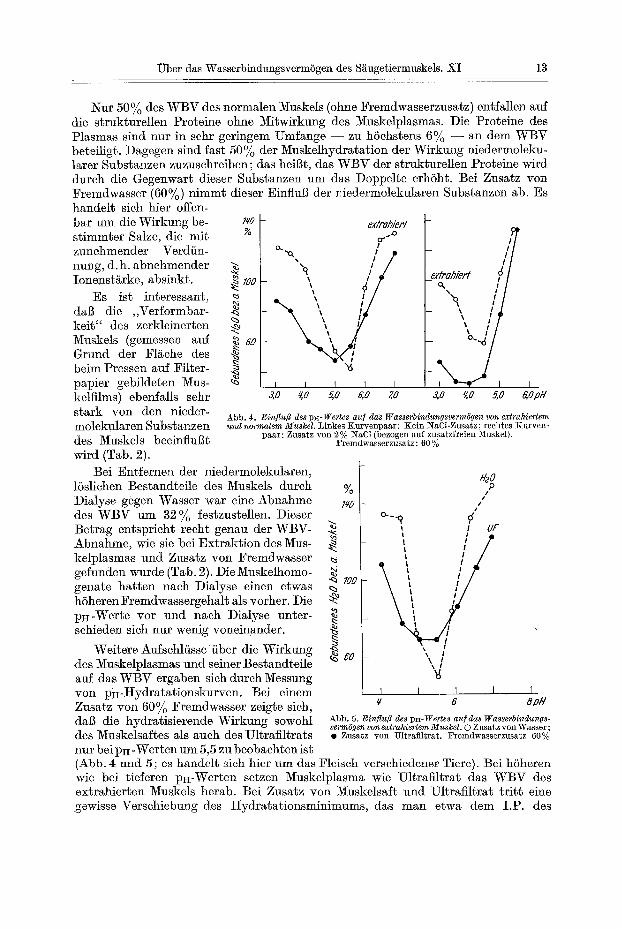
Nur 50% des WBV des normalen Muskels (ohne Fremdwasserzusatz) entfallen auf die strukturellen Proteine ohne Migwirkung des Muskelplasmas. Die Proteine des Plasmas sind nur in sehr geringem Umfange - - zu h6ehstens 6% - - an dem WBV beteiligt. Dagegen sind fast 50% der Muskelhydratation der Wirkung niedermoleku- later Substanzen zuzusehreiben; das heiBt, das WBV der strukgurellen Proteine wird dutch die Gegenwart dieser Subsganzen um das Doppelte erh6ht. Bei Zusatz yon Fremdwasser (60%) nimmt dieser Einflug der niedermolekularen Subsganzen ab. Es handelt sieh hier offen- bar um die Wirkung be- zo ex/rahieN st immter Salze, die mit g °"~ ~7 zunehmender Verdiin- c~.~ , // ~, I I ,~ nung, d.h. abnehmender X ,q / /
i # _extrahierl ~ l oneos,,rke, absio t, e,oo ",, itil
Es ist interessant, ~ °'"o -~1
~ ' o ~ _ ~o.
3,0 q.O 5,0 6,,0 7,0 3,0 ~0 5,0 G, OpH
dab die ,,Verformbar- kei6" des zerkleinerten Muskels (gemessen auf Grund d e r F1/~che des beim Pressen auf Filter- papier gebildeten Mus- kelfilms) ebenfalls sehr stark yon den nieder- molekularen Substanzen des Muskels beeinfluBt wird (Tab. 2).
Bei Entfernen der niedermolekularen, 16sliehen Bestandteile des Muskels dutch Dialyse gegen Wasser war eine Abnahme des WBV um 32 % festzustellen. Dieser Betrag entsprieht reeht genau der WBV- Abnahme, wie sie bei Extrakt ion des Mus- kelplasmas und Zusatz yon Fremdwasser gefunden wurde (Tab. 2). Die Muskelhomo- genate hat ten naeh Dialyse einen etwas h6heren Fremdwassergehal~ als vorher. Die p~-Werte vor und naeh Dialyse unter- sehieden sieh mlr wenig voneinander.
Weitere Aufsehl/issefiber die Wirkung des Muskelplasmas nnd seiner Bestandteile auf das WBV ergaben sich dureh Messnng yon pH-Hydratationskurven. Bei einem Zusatz yon 60% Fremdwasser zeigge sieh,
Abb. 4. EinfluB des pu-Wertes au] das Wasserbindungsverm6gen yon extrahiertem und normalem MuskeL Linkes Kurvenpaar: Kein NaC1-Zusatz; rechtes Kurven-
1)aar: Zusatz yon 2 % ~aC1 (bezogen a~f zusatzfreien 5~uskel). Fremdwasserzusatz: 60 %
% 140
"~ 100
60
i
l I
I UF / / I I
V, ? 1 I I I I
q 5 8pH
dab die hydratisierende Wirkung sowohl Abb. 5. Einflufl des pH-Wertes auf das Wasserbindungs- verm6gen yon extrahiertem Muskel. 0 Zlls~sz vort Wasser;
des Muskelsai~es als auch des Ultrafiltrats . Znsatz yon Ultrafiltrat. Fremdwasserzusatz 60% nur bei p~I-Werten um 5,5 zu beobachten is~ (Abb. 4 und 5 ; es handelt sich hier um das Fleisch verschiedener Tiere). Bei h6heren wie bei tieferen pi~-Werten setzen Muskelplasma wie Ultrafiltrat das WBV des extrahier~en Muskels herab. Bei Zusatz yon Muskelsaft and Ultraflltrat t r i t t eine gewisse Versehiebung des t Iydratat ionsminimums, das man etwa dem I.P. des
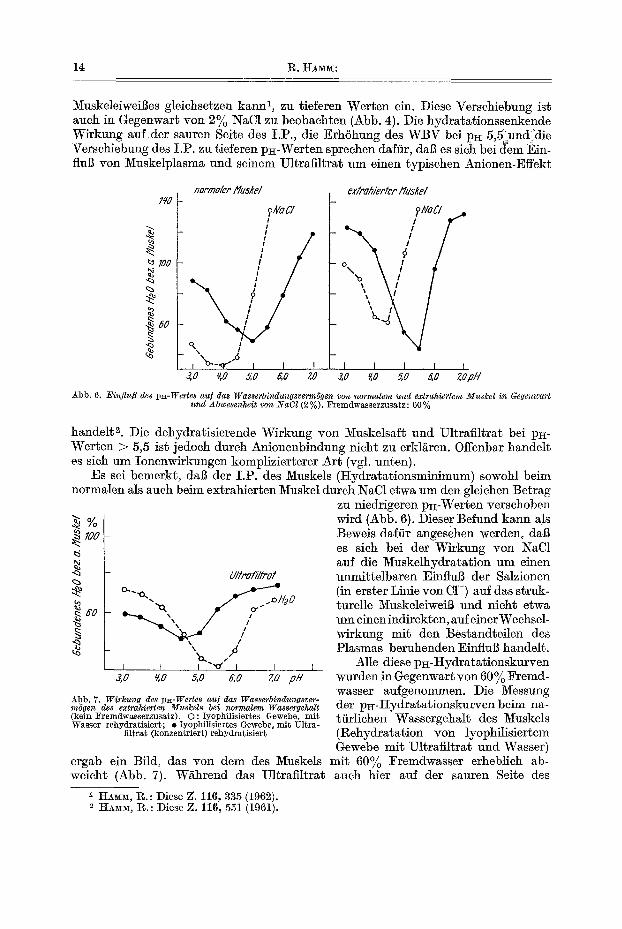
14 R. HAM~:
MuskeleiweiBes gleichsetzen kann ~, zu tieferen Werten ein. Diese ¥erschiebung ist auch in Gegenwart yon 2% NaC1 zu beobachten (Abb. 4). Die hydratationssenkende Wirkung auf der sauren SeRe des I.P., die Erh6hung des WBV bei lO~ 5,51~und die Verschiebung des I.P. zu tieferen p~-Werten sprechen daftir, dab es sich bei dem Ein- flul~ yon Muskelplasma und seinem Ultrafiltrat um einen typischen Anionen-Effekt
lqO
100
50
normgler Huskel
, Na CI
x t
3,0 ~,0 5,O ~,~ 7,0
exlfgbief ter llus ke l
/ / I I r f I
3,0 go 50 ~o ZopH
Abb. 6. Einflu~ des px-Wertes au[ das Wasserbindungsverm6gen yon normalem und extrahiertem MusSel i~z Gegenwart und Abwesenheit von ~YaC1 (2 % ). Fremdwasserzusa~z: 60 %
handelt 2. Die dehydratisierende Wirkung yon l~uskelsaft und Ul~rafiltrat bei p~- Werten > 5,5 ist jedoeh durch Anionenbindung nicht zu erkl/~ren. Offenbar handelt es sich am Ionenwirkungen komplizierterer Art (vgl. unten).
Es sei bemerkt, dab der I.P. des Muskels (tIydratationsminimum) sowohl beim normalen als auch beim extrahierten Muskel durch NaC1 etwa um den gleiehen Betrag
% ~ I00
.~ eo
Ultraf/Itrot
c~ " ~a" . o H2 0
, ,~ I I I
3,0 ~,0 5 , 0 G,O go f in
_A_bb. 7. Wirkung des p~-Wertes au] das Wasse~'bindungsver- m6gen des extrahierten Must:els be~ normalem Wassergehalt (kein ~'remdwasserzusatz). O: lyophilisiertes Gewebe, mi$ Wasser rehydratisiert; $ lyophilisiertes Gewebe, mit Ultra-
filtra$ (konzentriert) rehydratisier~
ergab ein Bild, das yon dem des 5Iuskels weicht (Abb. 7). W/~hrend das Ultrafiltrat
1 H ~ M , 1~.: D i e s e Z. 116, 335 (1962) . HA~, R. : Diese Z. 116, 551 (1961).
zu niedrigeren p~-Werten verschoben wird (Abb. 6). Diese r Befund kann als Beweis dafiir angesehen werden, dab es sich bei der Wirkung yon NaC1 auf die Muskelhydratation um einen unmittelbaren EinfluB der SMzionen (in erster Linie von C1-) auf das struk- turelle MuskeleiweiB und nicht etwa um einenindirekten, auf einer Wechsel- wirkung mit den Bestandteilen des Plasmas beruhenden EinfluB handelt.
Alle diese pit-Hydratationskurven wurden in Gegenwart yon 60% Fremd- wasser aufgenommen. Die Messung der p~-Hydratat ionskurven beim na- ttirlichen Wassergehalt des Muskels (Rehydratation yon lyophilisiertem Gewebe mit Ultrafiltrat und Wasser)
mit 60% l~remdwasser erheblich ab- auch bier auf der sauren Seite des

Uber das Wasserbindungsverm5gen des S~ugetiermuskels. XI 15
I.P. dehydratisierend wirkt, erhSht es bei pH-Werten zwischen 5,0 v_nd 7,0 das WasserbindungsvermSgen. Der I.P. des extrahierten Muskels wird dutch das Ultra- ill, ra t starker nach niedrigen p~-Werten verschoben als dies in Gegenwart yon 60% Fremdwasser der Fall is~. Dieser Befund li~l~t darauf schlieBen, dal] das Ultra- filtrat bei normalem Wassergehalt des Fleisches im gesamten gemessenen p~-Bereich vorwiegend einen Anionen-Effek~ ausfibt.
Aus den bisher erSrterten Versuchen lassen sich folgende Schlfisse ziehen: 1. Muskelplasma und Ultrafiltrat beeinflussen das WBV des Muskels prinzipiell in der gleichen Weise. Bei dem Einilul] des Plasmas auf die Muskelhydratation handelt es sich also um die Wirkung niedermolekularer Substanzen; 2. Bei normalem Wasser- gehalt des Gowebes iiben diese Substanzen einen ausgepri~g~en Anionen-Effekt aus. Mit zunehmendem Fremdwasscrgehalt wird dieser Anionen-Einflul~ durch andere Effekte fiberlagert; 3. Der I.P. des extrahierten Fleisches plus Ultrafiltra~ ist bei normalem Wassergehalt des Gewebes niedriger (p~ 4,5) als bei Gegenwart yon 60% Fremdwasser (p~ 5,0--5,5). Damit ist nachgewiesen, dal] es sich bei der frfiher yon uns beobachteten Verschiebung des I.P. zu hSheren pH-Werten bei Fremdwasser- zusatz 1 um einen Effekt handelt, der auf einer Verminderung der Wirkung der im Muskel enthaltenen Anionen dutch Verdfinnung (Erniedrigung der Ionenst£rke) beruht.
Wirlcung der anorganischen Ionen des Musl~elsa/tes auf das Wasserbindungsver- mSgen des Musl~els. Wir haben gezeigt, dal~ die Wirkung des Muskelsplasmas auf das WBV in erster Linie auf den Einflul~ niedermolekularer Substanzen zuriickzuffihren ist. Da kaum ein Zweifel bestand, da]3 es sich um die Wirkung anorganischer Ionen hundelt, wurde der Einflul~ der wich~igs~en, im Muskelsaft vorkommenden Ionen auf das WBV des Fleisches geprfift.
ErsehSpfend extrahier~es, lyophilisiertes Gewebe wurde mi~ Uitrafiltrat sowie mit verschiedenen SalzlSsungen rehydratisiert und das WBV beim normalen Wasser- gehalt des Muskels sowie bei Zusatz yon 60% Fremdwasser gemessen. Der mit Ultrafiltrat versetzte Muskel enthielt die niedermolekularen Substunzen des Fleisches in gleicher Konzentration wie der normale Muskel. In den Tab. 3 und 4 ist lediglich der Differenzwer~ A WBV angefiihr~, d .h . derjenige Betrag, um den das bei den betreffenden Zus~tzen gemessene WBV hSher oder niedriger ist als das WBV des mit reinem Wasser rehydratisierten, extrahierten Muskels bei dem gleiehen Gesamtwasser- gehalt.
Die bei Zusatz yon Ultrafiltrat gefundenen Werte stimmen mit denen der frfiheren Versuehe iiberein: Bei p~ 3,5 wird das WBV erniedrigt, bei p~ 5,5 erhSht ; bei pH 6,5 wirkt Ultrafiltrat in Gegenwart von Fremdwasser dehydra~isierend, bei normalem Wassergehalt dagegen hydratisierend (Tab. 3). Magnesium wurde als Acetat zuge- setzt, da das Acetat-Ion yore Muskel nut verhiiltnism~l]ig sehwach gebunden wird ~. In einer I~onzentration, die dem Gehal~ des Muskels an dissoziiertem Mg ++ entspricht, wirk~ dieses Salz auf den Muskel ohne Fremdwasserzusatz bei pH 3,5 etw~s dehydrati- sierend, bei pH 5,5 etwas hydratisierend. Beide Effekte sind geringer als die des Ultra- filtrats. Bei p~ 6,5 setzt Mg-Acetat im Gegensatz zum Ultrafiltrat das WBV herab. In Gegenwart yon 60% Fremdwasser wirkt Magnesiumaee~at bei p~ 3,5 und 5,5 hydrati- sierend, bei PH 6,5 hingegen stark dehydratisierend. Diese Erniedrigung is~ erheblich sti~rker als die durch Zusatz yon Ultrafiltrat (bei 60% Fremdwasserzusa~z) hervorge- rufeno. Ein Zus~tz yon K+-Ionen (als Aeetat) in einer Menge, welche dem Kaliumgehalt
G~A~, R., R. H A ~ u. A. B~V~A~- Diese Z. 32~, 1 (1953). - - HA~v~, R. : Diese Z. 109, 227 (1959).
HA~, R., u. R. G~v: Diese Z. 108, 280 (1958).
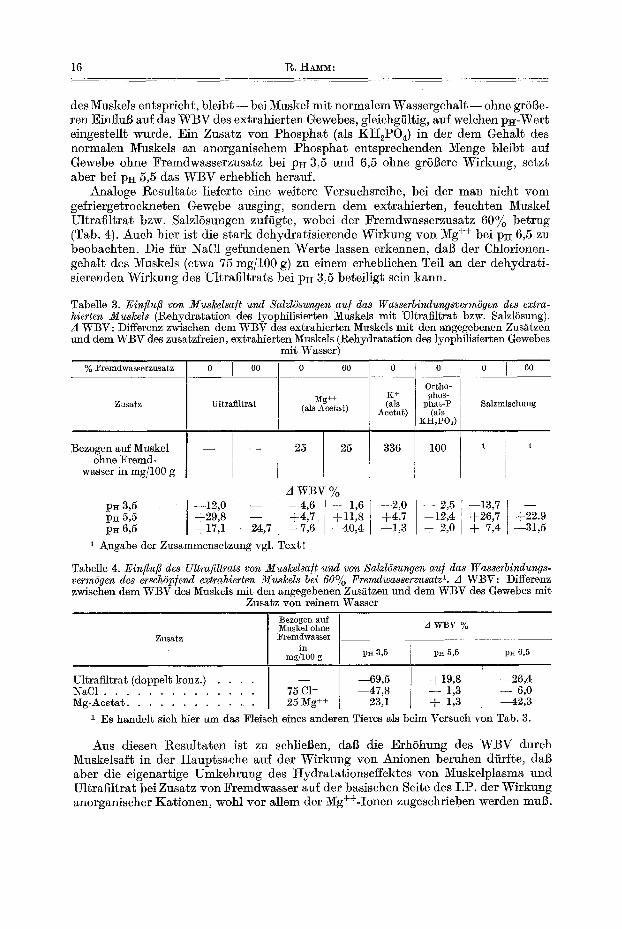
16 1%. H ~ :
des Muskels entspricht, bleibt - - bei 1V~uskel mit normalem Wassergehal t - - ohne gr6i3e- ten Einflu]3 auf das WBV des extrahierten Gewebes, gleichgfiltig, auf welchen p~-Wert eingestellt wurde. Ein Zusatz yon Phosphat (als KH2POa) in der dem Gehal~ des normalen Muskels an anorganischem Phosphat entsprechenden Menge bleib~ auf Gewebe ohne Fremdwasserzusatz bei pH 3,5 and 6,5 ohne grSBere Wirkung, se~zt aber bei loll 5,5 das WBV erheblich heratff.
Analoge Resultate lieferte eine weitere Versuchsreihe, bei der man nicht vom gefriergetrockncten Gewebe ausging, sondern dem ex~rahier~en, feuch~en Muskel Ultrafiltrat bzw. SalzlSsungen zuf/igte, wobei der Fremdwasserzusatz 60% betrug (Tab. 4). Auch hier ist die stark dehydratisierende Wirkung von Mg ++ bei PH 6,5 zu beobachten. Die ffir NaC1 gefundenen Werte lassen erkennen, da~ der Chlorionen- gehalt des Muskels (e~wa 75 rag/100 g) zu einem erheblichen Toil an der dehydrati- sierenden Wirkung des Ul~rafiltrats bei pH 3,5 beteiligt sein kann.
Tabelle 3. Einflufl yon Muskelsa/t und Salzl6sungen au/ das Wasserbindungsverm6gen des extra- hierten Muskels (Rehydrata~ion des lyophflisierten Muskels mit Ultrafil~ra~ bzw. Salz16sung). A WBV: Differenz zwischen dem WBV des extrahierten Muskels mit den angegebenen Zus~tzen und dem WBV des zusa~zfreien, ex~rabier~en Muskels (Rehydratation des lyophilisierten Gewebes
mi~ Wasser)
% Fremdwasserzusatz 0 I 60 0 I 60
Zusatz Ultrafil~rat
Bezogen au~ Muskel ohne Fremd-
wasser in rag/100 g
~g++ (als Acetat)
25 25
0
K+ (als
Acetat)
336
0 0 I 60
Ortho- phos-
Isha~-:P Salzmiscllung (als
KH2I'O,)
100 1
A w ~ v % p~3,5 i--12,O - - ~4,6 I + 1,6 --2,0 - - 2 , 5 --13,7 - - p~ 5,5 +29,8 +4,7 +11,8 +4,7 +12,4 +26,7 +22.9 pE6,5 +17,1 - - ~ , 7 --7,6 ~ 0 , 4 --1,3 + 2,0 + 7,4 --31,5
1 Axlgabe der Zusammensetzung vgl. Text!
Tabelle 4. Einflufl des Ultrafiltrats yon Muskelsa/t und von SalzlSsungen au] das Wasserbindungs- verm6gen des ersch6p/end extrahierten Muskels bei 60% Fremdwasserzusatz l. A WBV: Differenz zwischen dem W]3V des Muskels mit den angegebenen Zus~zen und dem W:BV des Gewebes mit
Zusatz yon reinem Wasser :Bezogen auf ~uskel ohne A WBV %
Zusatz Fremdwasser in
rag/100 g !0~ 3,5 1)g 5,5 1)H 6,5
Ultrafiltrat (doppe]~ konz.) . . . . I - - --69,5 + 19,8 --26,4 NaC1 . . . . . . . . . . . . . . ] 75 C1- 147,8 - - 1,3 - - 6,0 Mg-Ace~a~ . . . . . . . . . . . . 25 Mg ++ --23,1 + 1,3 -~2,3
1 Es handelt sick bier um das Fleisch eines anderen Tieres Ms beim Versuch yon Tab. 3.
Aus diesen Resttltaten ist zu schliel3en, dab die ErhShm~g des WBV durch Muskelsaft in der t tauptsache auf der Wirkung yon Anionen beruhen dfiffte, dab aber die eigenar~ige Umkehrung des l tydratationseffektes yon Muskelplasma und Ul~rafiltra~ bei Zusatz yon Fremdwasser auf der basischen Sei~e des I .P. der Wirkung anorganischer Kationen, wohl vor ahem der Mg++-Ionen zugeschrieben werden mu l l

~ber das WasserbindungsvermSgen des S£ugetiermuskels. XI 17
Es wurde ferner die Wirkung einer Salzmischung geprfif~, die s£mtliche der in hSherer Konzentration im Muskelplasma anwesenden anorganischen Ionen sowie Milchs~ure enthielt. Der Gehalt der Muskelproben an dieser Salzmischung entspraeh der Konzentration der im normalen Muskel in dissoziiertem Zustand anwesenden Ionen. Die Misehung setzte sieh zusammen aus KC1, KIt~P04, K2SO 4, Mg-Acetat, Ca-Acetat, K-Acetat, NaOH and Milehs~ure. Sie war in Wasser klar 15slich; erst bei l£ngerem Stehen in der K~lte bildete sich eine Triibung (wahrseheinlich Erdalkali- phosphate). Nach Zusatz der Salzmischang betrug der Gehalt der Niuskelproben bei normalem Wassergehalt des Gewebes in rag/100 g: 300 K +, 80 Na +, 25 Mg ++, 2,5Ca ++, 100 Orthophospbat, 80 Sulfat, 60 Chlorid und 50 Lactat. Ferner enthielt die Probe etwas Acetat. Wie Tab. 3 zeigt, ist die Wirkung dieser Salzmisehung auf das WBV des Muskels bei allen gemessenen p~-Wer~en derjenigen des Ultrafiltrates sehr £hnlieh, wiederum ein Zeichen, dab man die Wirkung des Muskelplasmas auf das WBV haupts£ehlich dem Effekt anorganischer Ionen zuschreiben muB.
Eine Zunahme der dehydratisierenden Wirkung des Ultrafiltrates yon MuskelpreB- salt bei pH 6,5 t r i t t nieht nut ein, wenn man dutch Fremdwasserzusabz verdiinnt, son- dern auch, werm man den Gehalt der Probe an Ultrafiltrat bei gleichbleibendemWasser- gehalt (vgl. Wer~e I mit I I I nnd I I mit IV auf Tab. 5) herabsetzt. Allerdings wirkt die Iterabsetzung der Ultrafiltrat-Konzentration im Muskel ohne Fremdwasser- zusatz wesentlieh starker dehydratisierend als eine Einstellung der gleichen Konzen- tration dureh Verdfinnen mit Fremdwasser. In der Probe mit der niedrigsten absolu- ten Menge an Ultrafil~rat (Wert III , Tab. 5) ist das WBV am niedrigsten. Dies zeigt, dab es sich bei dem Effekt der im Ultrafiltrat enthaltenen Ionen nicht einfach um den Einflul~ der IonensC4rke, sondern um eine spezifische Wechselwirkung der Ionen mit dem NIuskeleiweiB handeln dfirfte.
Tabelle 5. Einflufi der Ultrafiltrat~onzentration und des Fremdwasse~'zusatzes au/ das Wasser- bindungsvermSgen des Muskels. Erkl~rungen vgl. Legende yon Tab. 3
I II III IV
Fremdwasserznsatz % 0 60 0 60
Ultrafiltrat, bezogen auf Gesamt- wassergehalt der Probe . . in %
A WBV bei p~ 6,5 . . . . . . . . 100
~-17,1 100
--5,0 62,5
--41,8 62,5
--24,7
Wie wir aus den bisher angeffihrfen Versuchen entnehmen kSnnen, liil~t sich die dehydratisierende Wirkung des Muskelsaftes auf der sauren Seite des I.P. befrie- digend mit dem EinfluB der im Plasma enthaltenen anorganischen Anionen erkl£ren. Dieser Einflu]~ wird durch Zusa~z yon Fremdwasser abgeschwiieht. Die hydrati- sierende Wirkung des Muskelplasmas auf der basisehen Seite des I.P. bei normalem Wassergehalt des Gewebes l£Bt sich ebenfalls mit der Weehselwirkung der anorgani- schen Anionen des Muskels mit den strukturellen Proteinen erkl£ren. Bei Fremd- wasserzusatz aber treten auf der basischen Seite des I.P. Kationen-Effekte in Erschei- nung, die sich allein schon durch die Gegenwar~ der im Muskel yon t taus aus vor- handenen Mg++-Ionen erkl~ren lassen. Magnesium-Ionen allein setzen das WBV des Muskels (mit 60% Fremdwasser) st£rker herab als die gleiehe Magnesiumkonzen- tration im Ultrafiltrat oder in der ,,Salzmischung". Offenbar wird der Kationeneffekt dnreh die gleichzeitige Anwesenheit yon Anionen abgeschwiicht.
Es ergibt sich also die etwas iiberraschende Folgerung, dab es ganz yon der Kon- zentration des Salzes im N[uskel abhi~ngt, ob das Anion oder das Kation des Salzes
Z, Lebensmitt.-Un~ersuch., Band 117 2
18 g. tIAra:
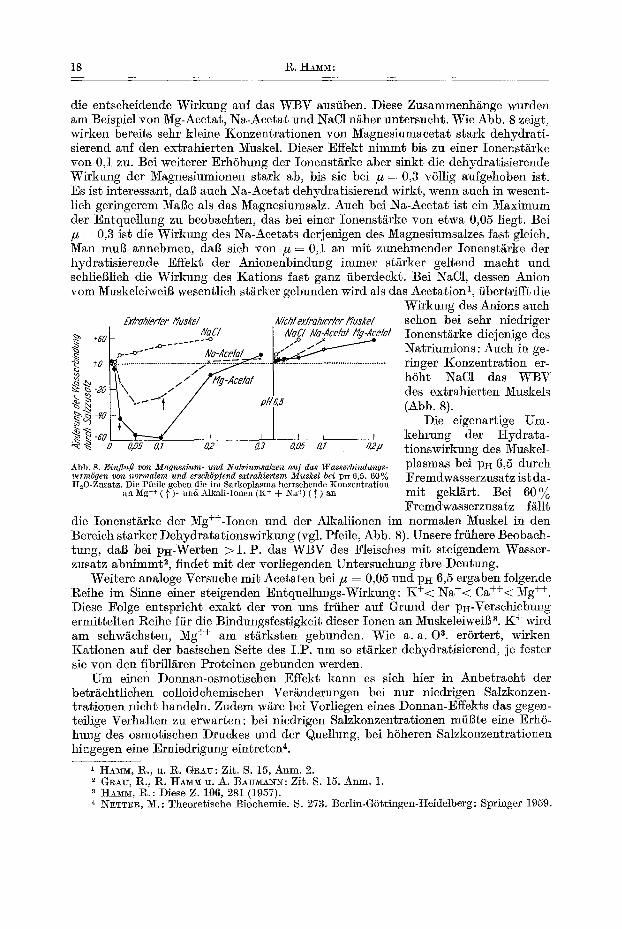
die entscheidende Wirkung auf das WBV ausfiben. Diese Zusammenh/inge wurden am Beispiel yon Mg-Acetat, Na-Acetat und NaC1 n/iher untersucht. Wie Abb. 8 zeigt, wirken bereits sehr kleine Konzentrat ionen yon Magnesinmacetat s tark dehydrati- sierend auf den extrahierten Muskel. Dieser Effekt n immt bis zu einer Ionenst/irke yon 0,1 zu. Bei weiterer ErhShung der Ionenst/irke abet sinkt die dehydratisierende Wirkung der Magnesiumionen stark ab, bis sic bei # = 0,3 v611ig anfgehoben ist. Es ist interessant, dab aueh Na-Acetat dehydratisierend wirkt, wenn aueh in wesent- lieh geringerem MaBe als das Magnesiumsalz. Auch bei Na-Acetat ist ein Maximum der Entquellung zu beobachten, das bei einer Ionenstgrke yon etwa 0,05 liegt. Bei # = 0,3 ist die Wirktmg des Na-Acetats derjenigen des Magnesiumsalzes fast gleich. Man mul? annehmen, dag sich yon /z = 0,1 an mit zunehmender Ionenstgrke der hydratisierende Effekt der Anionenbindung immer st/irker geltend maeht und schlieBlieh die Wirknng des Kations fast ganz fiberdeckt. Bei NaC1, dessen Anion yore Muskeleiweil~ wesentlieh stgrker gebnnden wird als das Aeetation 1,/ibertrifft die
Wirkung des Anions aueh £xlraNerler l f u ~ k e l N/?hlexlrah/erlerMuskel schon bei sehr niedriger
#a£/ I Nag/Na-Acetat/~g-Zce/a/ Ionenst£rke diejenige des ..~ +6~ - ___.-o I ..o / * _ . ~ ._~ ~.__~.- N~2Ac_e~/ ~ ~ Natriumions: Aueh in ge-
+-o ...................................... -~2/~ ........ T ......................... ringer Konzentrat ion er- ~ ,, / / /','¢g-Ace/a! ~ h6ht NaC1 das WBV ~-.o \~ ~<_!/ P ~ des extrahierten Muskels
~ //e,s (Abb. 8). ~ - s 0 Die eigenartige Um- :~ ~ -80 ~ I , I , I kehrung der Hydra ta-
o, o5 o,7 02 o,s o.o5 o,z a2# tionswirkung des Muskel-
Abb. 8. Einflufl yon Magnesium- und 2gatriumsalzen au] das Wasserbindungs- p l a s m a s bei pH 6,5 durch verm6gen yon normalem und ersch6p#nd extrahiertem Muskel bei pE 6,5. 60% Fremdwasserzusatz ist da- It20-ZuEatz. Die Pfefle geben die ira Sarkoplasma herrsehende ]~onzentration
an Mg ++ ( ]" )- mid Alkali-Ionen (K + ÷ Na +) ( ~ ) an m i t geklitrt. B e i 6 0 % Fremdwasserzusatz f~llt
die Ionenst/~rke der Mg++-Ionen nnd der Alka]iionen im normalen Muskel in den Bereieh starker Dehydratat ionswirkung (vgl. Pfeile, Abb. 8). Unsere frfihere Beobach- tnng, dag bei p~-Werten > I . P. das WBV des Fleisches mit steigendem Wasser- zusatz abnimmt 2, finder mit der vorliegenden Untersuchung ihre Deutung.
Weitere analoge Versuehe mit Acetaten bei/~ = 0,05 und pH 6,5 ergaben folgende geihe im Sinne einer steigenden Entqnellungs-Wirkung: K + < Na+< Ca++< Mg ++. Diese Folge entspricht exakt der yon uns frfiher auf Grund der p~-Versehiebu~g ermittelten l~eihe ffir die Bindnngsfestigkeit dieser Ionen an MuskeleiweiB 3. K + wird am sehw/~chsten, Mg ++ am st/irksten gebunden. Wie a. a. 0 8. erSrtert, wirken Kationen auf der basisehen Seite des I .P. um so stiirker dehydratisierend, je fester sie yon den fibrill/iren Proteinen gebunden werden.
U m einen Donnan-osmotisehen Effekt karm es sich hier in Anbetraeht der betr/iehtliehen colloidehemischen Ver/inderungen bei nur niedrigen Salzkonzen- trationen nieht handeln. Zudem w/~re bei Vorliegen eines Donnan-Effekts das gegen- teilige Verhalten zu erwarten: bei niedrigen Salzkonzentrationen mfiBte eine Erh6- hung des osmotisehen I)ruekes nnd der Quellung, bei h6heren Salzkonzentrationen hingegen eine Erniedrigung eintreten 4.
i tIAM~, 1%., u. R. G~Av: Zit. S. 15, Anm. 2. G~A~, I~., R. tIAM~ u. A. BAV~L~N~: Zi~. S. 15, Anm. 1. I-I~N, 1~. : Diese Z. 106, 281 (1957). N E ~ , M. : Theoretisehe ]3ioehemie. S. 273. Berlin-G6ttingen-tteidelberg: Springer 1959.
13ber das WasserbindungsvermSgen des Sgugetiermuskels. XI 19
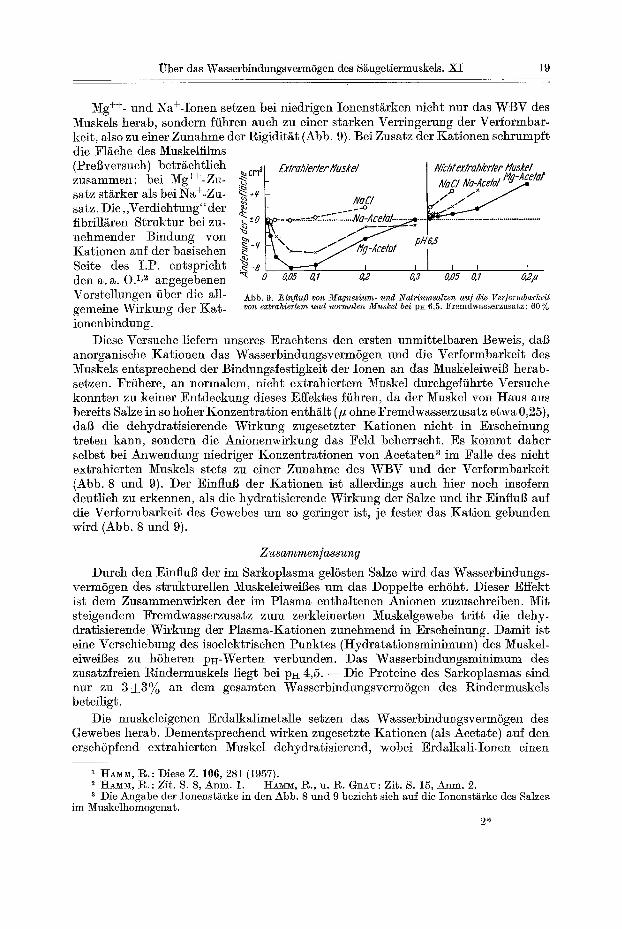
Mg ++- nnd Na+-Ionen setzen bei niedrigen Ionenst/irken nicht nur das WBV des Muskels herab, sondern fiihren auch zu einer starken Verringerung der Verformbar- keit, also zu einer Zunahme der Rigidit~t (Abb. 9). Bei Zusatz der Kationen sehrumpft die F1/~ehe des Muskelfilms (Prel~versuch) betr/£chtlich ~cm~ zus~mmen; bei Mg ++- Zu- ,~ satz st/irker als bei Na+-Zu- ~ *~ satz. Die, ,Verdichtung"der .~ fibrill~ren Struktur bei zu- ~-o nehmender Bindung yon ~_¢ Kationen auf der basischen Seite des I .P. entspricht ~ -a den a. ~. O. 1,2 a~gegebenen ~ o Vorstellungen fiber die all- gemeine Wirkung der Kat - ioncnbindung.
Exlrahierler lluskel N/chl exlrahierler lluakel NaCl Ne-Acelal Hg-~etot . / o f x
NaCI I. / ~ ..o FLj~..--~ ~--.o-~.-....~...=::.~.~.~Na-Acetel ........................................................
~ I I I I I I , 0,05 O,l g 2 0,3 8,05 0, I 021/
Abb. 9. JEinflu/3 yon M a g n e s i u m - u n d 2qatriumsalzen au] die Ver]ormbarkei t yon extrahiertem und normalen M u s k e l bei PE 6,5. Fremdwasserzusatz: 60 %
Diese Versuche liefern unseres Erachtens den ersten unmittelbaren Beweis, dab anorganische Kationen das WasserbindungsvermSgen und die Vefformbarkeit des ~uskels entsprechend der Bindungsfestigkeit der Ionen an das Muskeleiweil~ herab- setzen. Frfihere, an normalem, nicht extrahiertem Muskel durchgeffihrte Versuche konnten zu keiner Entdeckung dieses Effektes fiihren, da der Muskel yon Haus aus bereits Salze in so hoher Konzentrat ion enth/~lt (# ohne Fremdwasserzusatz etwa 0,25), daI3 die dehydratisierende Wirku~g zugesctzter Kationen nicht in Erscheinung treten kann, sondern die Anionenwirkung das Feld beherrscht. Es kommt daher selbst bei Anwendung niedriger Konzentrationen yon Acetatea 3 im Falle des nicht extrahierten Muskels ste~s zu einer Zunahme des WBV und der Verformbarkeit (Abb. 8 und 9). Der Einflul~ der Kationen ist allerdings auch bier noch insofern deutlich zu erkennen, als die hydratisierende Wirkung der Salze und ihr Einflul~ auf die Verformbarkeit des Gewebes nm so geringer ist, je fester das Kat ion gebunden wird (Abb. 8 und 9).
Zusammen]assung Durch den Einflul3 der im Sarkoplasma gelSsten Salze wird das Wasserbindungs-
verm6gen des strukturellen Muskeleiweil3es um das Doppelte erhSht. Dieser Effekt ist dem Zusammenwirken der im Plasma enthaltenen Anionen zuzuschreiben. Mit steigendem ~remdwasserzusatz zum zerkleinerten Muskelgewebe t r i t t die dehy- dratisierende Wirkung der Plasma-Kationen zunehmend in Erscheinung. Damit ist eine Verschiebung des isoelektrischen Punktes (Hydratationsminimum) des Muskel- eiweii3es zu h6heren pH-Werten verbunden. Das Wasserbindungsminimum des zusatzfreien Rindermuskels liegt bei pI~ 4,5. - - Die Proteine des Sarkoplasmas sind nur zu 3=~3~o an dem gesamten Wasserbindungsverm6gen des Rindermuskels beteiligt.
Die muskeleigencn Erdalkalimetalle setzen das WasserbindungsvermSgen des Gewebes herab. Dementsprechend wirken zugesetzte Kationen (als Acetate) auf den erschSpfend extrahierten Muskel dehydratisierend, wobei Erdalkali-Ionen einen
1 HAv!~r, t~. : Diese Z. 106, 281 (1957). HAMM, R.: Zit. S. 8, Amn. 1. - - HA~rM, R., u. R. GI~AU: Zit. S. 15, Anm. 2. Die Angabe der Ionenst~rke in den Abb. 8 und 9 bezieht sich auf die Ionenst/~rke des Salzes
im Muskelhomogenat. 2*

20 R. ttxM~ und H. IWATA:
wesent l ich sti~rkeren Ef fek t zeigen als Alka l i - Ionen . Die I t e r abse t zung des Wasser - b indungsvermSgens is t u m so sti~rker, je fes ter die K a t i o n e n v o m MuskeleiweiI3 gebunden werden. Bei hSherer Sa l z -Konzen t r a t i on her r sch t de r EinfluB des Anions (Aee ta t - Ion) des zugese tz ten Salzes vor. Se tz t m a n s t a t t de r A c e t a t e die Chloride de r Alkal i - u n d E r d a l k a l i m e t a l l e zu, so wi rd auch sehon bei n iedr igen Salzkonzen- t r a t i o n e n die I t y d r a t a t i o n des Gewebes durch die r e l a t i v s t a rke Bindung des Chtor- Ions be s t immt .
Chemische und physikalische Veriinderungen beim Erhitzen von Fleisch
I . Mitteilung
Wirkung des Erhitzens in Gegenwart yon 5~atriumchlorid au f W a s s e r b i n d u n g s . v e r m 6 g e n u n d p ~ - W e r t des Rindermuskels
~on
R. H~wM u n d H. IWATA*
Mitteilung aus dem Institut /iir Chemie und Physi~ der Bundesanstalt /iir Flelseh/orschung, Kulmbach
Mit 3 Tex~abbfldungen
(Eingegangen am 21. Oktober 1961)
Fr i ihere Un~ersuehnngen x haben gezeigt, dab sich Wasse rb indungsvermSgen 2 und p g - W e r t des zerk le iner ten R inde rmuske l s (post rigor) be im Erh i t zen in eharak te - r i s t i scher Weise i~ndern u n d dab sich aus diesen Ver~nderungen Rficksehlfisse auf den Chemismus der I - I i tzedenatur ierung der s t ruk tu re l l en Muskelpro te ine in s i tu ziehen lassen. I m Hinb l i ck auf die P rax i s de r F le i sehve ra rbe i tung war noch die F r a g e zu priffen, ob der Vorgang der I t i t z e d e n a t u r i e r u n g des MuskeleiweiBes in Gegenwar t yon N a t r i u m e h l o r i d (2 %) pr inzipie l l g le ichar t ig verli~uft wie im ungesalzenen Muskel. Hier - zu war es zun£chs t notwendig , die zei t l iehe Xnderung Yon Wasse rb indung und pH- W e r t des Muskels m i t n n d ohne Salzzusatz w~hrend des Erh i tzens bei verschiedenen T e m p e r a t u r e n zu verfolgen. Anschl ieBend wurde der EinfluB der T e m p e r a t u r auf H y d r a t a t i o n u n d pH-Wer t bei gleieher E rh i t zun gsda ue r un te r such t .
Hethodik Als Untersuchungsmaterial diente der 5 Tage nach dem Schlachten entnommene M. longis-
simus dorsi 5j~hriger Kiiho der Handelsklasse B. Je 10 g des im Fleischwolf zerkleinerten Rindermuskels wurden in den Glasgef~13en eines
Homogenisators (Get,t: Fa. Bfihler, Tfibingen) in diinner Schicht ausgebreitet und im Thermo- staten auf verschiedene Temperaturen erhitzt. Die Temperatur des Fleisches wurde mittels Thermoelementen im Innern der Schieht und in unmittelbarer N~he der Glaswandung gemessen. Der W~rmegang in der Probe erfolgte so raseh, dab prak~isch keine Temperaturdifferenz zwischen den beiden MeBstellen bestand. Die Zeit, die notwendig war, um die Probe yon Zimmertemperatur au~ die gewiinschte Temperatur zu bringen, betrug bei 30 ° und 50 ° C 4 rain, bei 70 ° und 90 ° C 5 rain.
* Frl. I-V~EDWIG BREttl~I danken wir fiir ihre wertvolle lYlitarbeit. 1 H A ~ , R., u. F. E. D~ATm~AG~: ~ood Res. 25, 587 (1960). 2 I)efinitionen fiir die Begriffe ,,WasserbindungsvermSgen" und ,,Hydratation" sind ange-
geben bei: R. HAM~: Advane. Food Res. 10, 355 (1960).























![[ÜBER DAS SONISCHE] ZUM BEGRIFF DES SONISCHEN · 2017. 11. 28. · ["ÜBER DAS SONISCHE"] ZUM BEGRIFF DES SONISCHEN Audiosignal Definitionen des Sonischen Klangerzeugung auf C64](https://img.pdfslide.org/doc/110x75/6132a8d8dfd10f4dd73a97eb/oeber-das-sonische-zum-begriff-des-sonischen-2017-11-28-oeber-das.jpg)





