Embed Size (px)
Citation preview

OBER DIE BIPOLARE DIFFERENZIERUNG DER EIZELLEN VON NEREIS LIMBATA UND CHAETOPTERUS PERGAMENTACEUS
Yon JOSEF SPEK (Heidelberg) Aus dem Marine Biological Laboratory Woods Hole, Mass. U.S.A.
Mit Tafel I I
Eingegangen am 3. M~trz 1934
Die Eizellen yon Nerds Dumerilii waren das erste Objekt, an welchem das Problem der bipolaren Differenzierung der Eizellen zum ersten Male yon physikaliseh-chemischen Gesichtspunkten beleuehtet worden ist (Spek 1930). l~aeh den Ergebnissen jener Untersuchung kommt die bipolare Differenzierung der Nereis-Eier dureh eine Erscheinung zustande, mit der man in der Zell- physiologie bis dahin gar nicht gereehnet hatte. Es galt daher zun~ichst, den Geltungsbereieh dieser Befunde aueh bei anderem, m6glichst verschiedenartigem Material zu prfifen und die physikalisch/ehemischen Grundlagen der Beob- aehtungen zu verbreitern und zu sichern. So entstand eine Serie yon Unter- suchungen, yon denen eine - - ausgef~ihrt an Eiern yon S i i 6 w a s s e r - T e l e o s t e e r n - - bereits publiziert ist ( S p e k 1933). Alle weiteren Untersuehungen dieser Serie wurden an der Marinen Biologischen Station in Woods Hole, Mass. U.S.A. ausgeffihrt und umfassen Versuche an Eiern yon 2~Tereis limbata, Chaetopterus pergamentaceus, Loligo vulgaris, Columbella avara, Ciona intestinalis und Asterias Forbesii. An den Eizellen von Columbella und Ciona konnten bis jetzt erst einige Grundversuche ausgeffihrt werden, und die Untersuchung an den Asterias- Eiern ist insofern noeh ein Fragment geblieben, als die Versuche hier mit anders- artigen Methoden fortgef/ihrt werden mfissen, was bisher noch nicht gesehehen konnte.
In der vorliegenden Arbeit berichte ich fiber die neuen Versuehe an Eiern yon 2Vereis limbata und Chaetopterus pergamentaceu8. Die Beriehte fiber die Untersuehungen an den andern Objekten werden folgen.
Zu einer Wiederholung der Versuche an den Eiern einer andern Nereis-Art ffihlte ich reich dadureh veranlal~t, dab sieh seit meiner ersten Untersuchung yon 1930 meine Methoden erweitert und verbessert batten. Dies sehien auch eine neue Chance zu bieten, einige Liieken meiner ersten Beweisffihrung aus- zufiillen.
Bei meinen ersten Studien an Nereis-Eiern hat te sich ergeben, dab die Versehiedenheit dos Protoplasmas am animalen und vegetativen Po lde r Eizellen ( ~ bipolare Differenzierung), welche nach der Ausseheidung der Riehtungsk6rper in Erseheinung tritt , im wesentliehen dadureh zustande kommt, da~ alkalisehe Plasmakolloide naeh dem animalen und saure Plasmakolloide naeh dem vegeta- tiven Pole wandern und sich bier immer starker anreichern. Erkannt wurdo

Bipolare Differenzierung der Eizellen yon Nereis limbata u. Chaetopterus pergamentaceus 395
dies durch Vitalf/~rbungen mi t I n d i k a t o r e n und an der Verschiebung des Fa rb - tons des na t i i r l ichen diffus gelSsten P igmen t s der E ie r yon Nereis Dumerilii. Dieser natf i r l iche I n d i k a t o r fehl t den E ie rn yon Nerds limbata. K l u m p e n von E ie rn erseheinen zwar blaBgriin. Dieses P igme n t sehls aber bei Zusa tz yon pene t r i e renden S~turen oder Alka l ien n ich t um. Bei mikroskopischer Bet rach- t ung sind die feinen br/~unlieh-grauen KSrnchen odor TrSpfehen un te r der Kor t ika l sch i ch t das einzige, was an den Eizel len gef/~rbt erscheint . W e n n ein diffus ver te i l tes P igmen t vo rhanden ist, i s t es jedenfal ls sehr blaB. - - F i i r Vi ta l f s s ind die E ie r von Nereis limbata ein he rvor ragend gutes Objek t .
Bei meiner e rs ten Nereis-Untersuehung s t anden mi r nur zwei I n d i k a t o r e n fiir d i eVi ta l f~ rbungen zurVer f i igung : N e u t r a l r o t und N i l b l a u s u l f a t B. Eine gewisse Kon t ro l l e zu den Umschl/~gen dieser Vi ta l fa rben l ieferte nur noch das erw. na t i i r l iche P igmen t der E ie r yon Nereis Dumerilii. Bei so wenigen In- d i k a t o r e n h a t m a n vie l le ieht i m m e r noch m i t der MSgliehkei t rechnen k5nnen, dab die verschiedenen F a r b t S n e dureh andere F a k t o r e n als das p H verursach t sein kSnnten , u n d e s nur Zufal l ist , dab ~lle drei im gleiehen Zel lbezi rk einen F a r b t o n zeigen, de r dem gleichen p H entspr ieh t . Al lerdings kennen wir fiir diese F a r b e n keinen dera r t igen F a k t o r , der eine para l le le Serie von F a r b - ~nderungen herbei f i ihren kSnnte wie das pH, und andererse i t s sehlagen Neu- t r a l r o t und der na t t i r l iche I n d i k a t o r in den Eiern , wenn m a n exper imente l l i rgendeine p t t -Versch iebung in der Zelle hervorruf t , im Augenb l iek im e rwar te t en Sinne urn. Auf alle F/~lle mul~te aber die Beweisf i ihrung sicherer werden, wenn m a n das gleiehe l~esul ta t mi t e iner gr6~eren Zahl yon I n d i k a t o r e n erhielt .
I ch pflege j e t z t zu Vi ta l fa rbungen zwecks p H - B e s t i m m u n g in de r lebenden Zelle auBer N e u t r a l r o t und N i l b l a u s u l f a t noch B r i l l a n t v i t a l r o t , B r i l - i a n t k r e s y l v i o l e t t , K r e s y l e c h t v i o l e t t und evtl . noeh M e t h y l r o t anzu- wenden. ~ b e r die Eigensehaf ten dieser Fa rbs to f fe habe ieh mich schon fri iher ge/iuBert ( S p e k 1933).
BriUantvitalrot kann man so anwenden wie Neutralrot; auch die Skala seiner Farb- tSne bei verschiedenem pH ist ~hnlieh: rosa auf der sauren, rostgelb auf der alkalischen Seite, dazwisehen zinnoberrot. Methylrot dringt meist nur aus Medien in die ]ebenden Zellen ein, welche die Permeabilit~t der Zellen erhOhen, wie etwa reine Jsoton. NaC1-LOsung. Besonders bei Zellen die selbst gelblich sind, sind blasse Anf/~rbungen im fahlgelben (alkalischen) Farbton von Neutralrot oder Brillantvitalrot mitunter nicht leicht mit Sicher- heir erkennbar. In solchen F/~llen sind Kontrollen mit blau~rosa Indikatoren sehr er- wiinseht. Bei vielen Objekten - - so auch bei unserm - - ]assen sich besonders sehOne F~rbungen dieser Art mit Brillantkresylviolett und Kresylechtviolett erhalten. Diese beiden Indikatoren und Nilblausulfat sind blau auf der sauren und rosa bzw. lila auf der alkalisehen Seite. Es sei auch bier hervorgehoben, da6 besonders bei den beiden ersten der genannten drei Indikatoren die Umsehl/~ge in reinen w/~sserigen LOsungen sehr langsam erfolgen. Sehr viel raseher erfolgen sie in organischen L6sungsmitteln wie 01en und Chloro- form und augenblieklieh erfolgen sie in manchen vitalgef~irbten Zellen (wie z. B. jungen Ooeyten yon Teleosteern). Is t mit irgendeinem Indikator in einer Zelle oder einem Zellbezirk ein bestimmtes p H gefunden worden, so f/~rbt sich diese Zelle oder Zellregion - - sofern sie sich iiberhaupt anf~rben l~f~t - - mit Brillantkresylviolett odor Kresylechtviolett s t e t s sofort in dem Farbton an, der diesem pi t entspricht, unabh/ingig davon, ob die Farb- 10sung der Zelle Yon aul]en her in blauer odor rosa Modifikation geboten wird, u n d auCh
26*

396 Spek
dann, wenn die Anfgrbung sehr rasch erfolgt. Hat man also z: B. mit irgendeiner Vital- fi~rbung mit einem Indikator oder sonstwie in einer Zellsubstanz ein pH fiber 7,6 vor- gefunden, kann man sicher sein, dab etwa Brillantkresylviolett, wenn es diese Zellsubstanz fiberhaupt fitrbt, sie in lila oder rosa Farbton und niemals im blauen anf/~rbt. Bei all den zahllosen F/~rbungen, welche ich mit diesen Farbstoffen ausgeffihrt habe, habe ieh von dieser Regel noch keine Ausnahme gefunden. DaB in dem angefiihrten Fall, auch wenn der Farbstoff in einer blauen L6sung geboten wird, nicht zuerst Blaufarbung erfolgt und dann der blaue Farbton allm/~hlich in Rosa fibergeht, ist jedenfalls so zu erkl&ren, dag die alkalischen Plasmateilchen sich yon vornherein nur mit den rosa Farbteilchen verbinden k6nnen (und saure PIasmateilchen in en~sprechender Weise nur mit b]auen). Setzt man zu dem AuBenmedium der mit Kresylechtviolett oder Brillantkresylviolett gefgrbten Zellen Milchs~ure oder Ammoniak zu, so erfo]gt meist eine rasche Entf/irbung der Zellen. Aus den Zellkolloiden herausgelSst sehlagen die Farbstoffe ebenso langsam um wie im Reagenz- glas. - - Brillantvitalrot und Neutralrot schlagen bei Zusatz penetrierender S/~uren oder Alkalien sofort um. Ihre Adsorption an den Plasmakolloiden wird aber etwa durch Am- moniak oder Milchs/~nre auch rfickg/~ngig gemacht.
Einen andern Faktor, der die gleiehen Farbumschlgge herbeiffihren k6nnte wie die Wasserstoffionenkonzentration, kennen wir auch bei den neueingeffihrten Indikatoren nicht. Wer etwa versucht, alle die gesetzm~Bigen F&rbungen durch ,,spezifische Adsorptionen" der Farbstoffe am Plasma zu erkl/~ren, st6gt sogleich auf die groBe Schwierigkeit, dab er ffir jeden Farbstoff so viele spezifische Adsorptionen annehmen mug, als FarbtOne bei der Vitalf/~rbung auftreten, und bei manehen Vitalf/~rbungen ist das die ganze Serie yon Farbt6nen, die der betr. Indikator iiberhaupt annehmen kann. ~ DaB nun diese vielen spezifisehen F/~rbungen alle so ausfallen, dab zuf/~llig alle Indikatoren den Farbton an- nehmen, der dem betr. pH entsprieht, ist /~uBerst unwahrseheinlieh.
Die Methode der Vitalf/s ist sehon friiher besehrieben worden (vgl. Spek 1933). Hauptsache ist, dab man stets nur ganz verdiinnte Farbl6sungen anwendet. Die L6sungen yon Kresyleehtviolett und Brillantkresylviolett sollen nieht 1/tnger als 3--4 Wochen ver- wendet werden, da sieh ihre physikalischen Eigensehaften allmahlieh /mdern.
Nereis limbata kann in Woods Hole an Sommerabenden beim Lampenlieht leicht in grogen Mengen gefangen werden. Setzt man M~nnchen und Weibehen zusammen, so erfolgt prompt Befruchtung. Technische Einzelheiten sind schon friiher besehrieben worden.
Eine Aufzfiehtung der Heteronereis-Form aus den Bodentieren, wie ieh sie in Neapel an Nereis Dumerilii stets ausfiihrte, habe ich in Woods Hole nicht versucht. Dement- sprechend konnte ich auch die allm~hliche Heranreifung des Ovarialeies und die Ent- stehung der konzentrischen Struktur des vollentwickelten Ovarialeies an diesem Objekt bisher noch nicht studieren.
Auf das Wesen der konzentr ischen S t ruk tu r des reifen, unbef ruch te ten Nereis-Eies gehe ich an sp/~terer Stelle (S. 401) genauer ein. Auch bei Nereis limbata verschwindet bekannt l ich bei der Befruchtung die/~uBere brei te Kort ikal - schicht dadurch, dab sie ganz oder zum gr6Bten Teil nach augen ausgeschieden wird. Hiernach weist, yon den oberfl/~chlich angereicherten Granu len abgesehen, der ganze EikSrper vori ibergehend eine gleichm/tgige Beschaffenheit auf und n i m m t bei Vitalf / i rbungen eine gleichm/iBige blasse Farbe an.
Die bipolare Differenzierung setzt auch hier erst nach der Ausscheidung der Richtungsk6rper ein, und k o m m t auch hier dadurch zustande, dag gewisse Subs tanzen nach dem Richtungsk6rperpol abwandern und sich dort immer

Bipolare Differenzierung der Eizellen yon Nereis limbata u. Chaetopterus pergamentaceus 397
sti~rker anreichern, w/~hrend andere von dem RichtungskBrperpol immer mehr abriicken and nach dem entgegengesetzten Pol wandern. Die (~ltropfen und DottertrBpfchen bewegen sich geradlinig nach dem vegetat iven Pol, bis sie dort dicht zusammengedr~ngt sind. Die Granulen, die sich am animalen Pol immer mehr anreichern, s tammen aus der Oberfli~chenschicht, welche bei dem unbe- ffuchteten Ei unter der breiten Kortikalschicht liegt.
Sowie die Sonderung der Substanzen einen gewissen Grad erreicht, kann man dann, wenn man die Eier vital f/~rbt, auch hier wieder die Beobaehtung machen, welche ich an vielen anderen Zellen schon beschrieben habe: In dem Substanzgemisch f~rbt sich vorwiegend nur eine Substanz an, und die ganze F/~rbung bleibt blaB. Sowie aber die verschiedenen Substanzen des Gemenges voneinander getrennt werden, wird jede Substanz fi~rbbar, und die F/~rbung wird immer intensiver, je vollst/~ndiger die Aussonderung der verschiedenen Substanzen vollzogen ist. Verwendet man aber zu diesen F/~rbungen Indikatoren, so erh~lt man ausnahmslos bei den am animalen Pol angereicherten Substanzen eine Anfiirbung in stark alkalischem Farbton, bei den Substanzen am anderen Pol hingegen eine solche im tiefsauren. Vor der Befruchtung f~rbt sich z. B. in Brillantkresylvio]ett die ganze l~asse des Protoplasmas innerhalb der Kor- tikalschicht gleichm/~Big blaBblau. Die gleiche Farbe hat auch der ganze Zell- kBrper des befruehteten Eies vor der bipolaren Differenzierung. Sowie aber nach der RichtungskSrperbildung der klare Hof des animalen Plasmas erscheint, ist auch schon sehr bald die erste Andeutung einer Verschiedenfarbigkeit der beiden Pole zu erkennen der Art, dab die animale Zellh/~lfte eine deutliche rosa F~rbung zeigt, withrend am andern Pol die blaue Fs vers ts erscheint. Diese Fi~rbungsunterschiede werden immer deutlicher. I m Vierzellenstadium bietet der mit Brillantkresylviolett gef/irbte Keim in der Seitenansicht etwa das Bild der Fig. 1 Tar. I I dar. (Das darunter liegende zweite Zellenpaar ist nicht gezeichnet.)
Wenn mari zu den Vitalf/~rbungen so minimale Zuss der Farbli~sungen zum Seewasser verwendet, wie ich das friiher beschrieben babe, dauert es eine Stunde oder mehr bis eine deutliche Fs erscheint. Auch wenn man also Eier gleich nach der Befruchtung in die FarblBsung setzt, t r i t t eine deutliche Anf/~rbung, da ja die Eier sich unterdessen weiter entwickeln, erst auf dem nach einer Stunde erreichten Stadium hervor. So kBnnte man meinen, dab man nur aus diesem Grunde eine intensivere F/~rbung der ersten Stadien nicht erreichen kann. Bei sorgf/~ltiger Vergleichung der Fi~rbung ganz junger und ~lterer Stadien mit schon bipolar differenzierten Blastomeren ergibt sich aber doch, daft letztere sich in der gleichen Zeit und in der gleichen L6sung wesentlich intensiver anf/~rben.
Die groge Masse des animalen Protoplasmas, welche sich nach der Sub- stanzsonderung in Brillantkresylviolett rosa fs ist natiirlich nicht neu ent- standen, sondern war vorher schon vorhanden, bloB gleichm/~Big zwischen dem Dotterplasma verteilt. Solange dies aber der Fall ist, erfolgt i iberhaupt keine Anf/~rbung derselben. Der rosa Farbton des animalen Plasmas wfirde mit dem hellen Blau des Dotterplasmas gemischt die Gesamtfarbe sicher nach Violett
398 Spek
verschieben. Ein solcher Mischton ist aber vor der bipolaren Differenzierung in dieser Farbe nie zu beobachten. Unbefruchtete Eier kann man ja darin beliebig lange f~rben, abet der zentrale ZellkDrper der Eier bleibt rein blau.
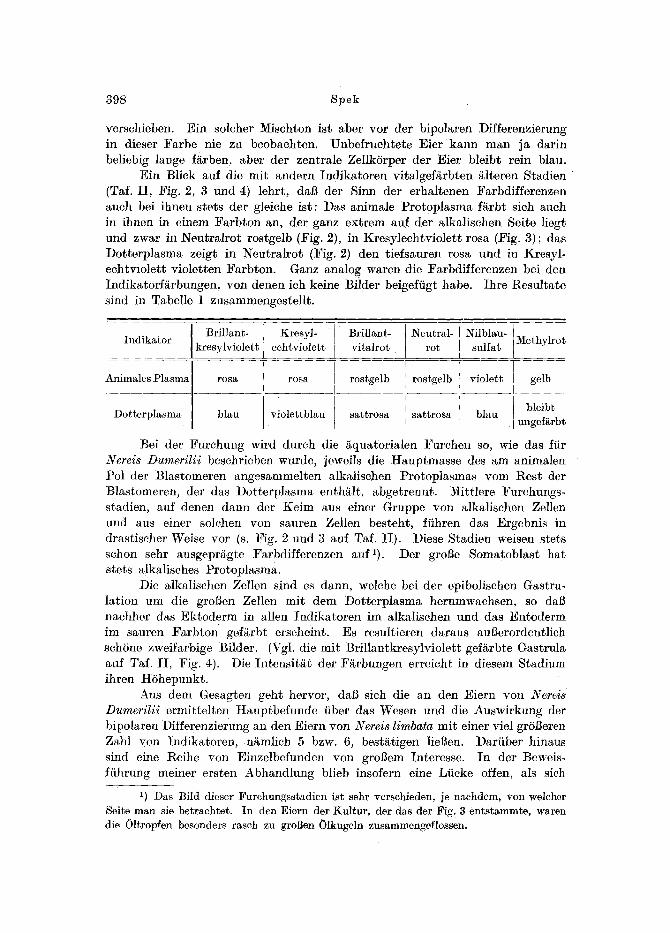
Ein Blick auf die mit andern Indikatoren vitalgef~rbten i~lteren Stadien (Taf. I I , Fig. 2, 3 und 4) lehrt, dad der Sinn der erhaltenen Farbdifferenzen auch bei ihnen stets der gleiche ist: Das animale Prot0plasma f~rbt sich auch in ihnen in einem Farbton an, der ganz extrem auf der alkalischen Seite liegt und zwar in Neutralrot rostgelb (Fig. 2), in Kresylechtviolett rosa (:Fig. 3) ; das Dotterplasma zeigt in NeutrMrot (Fig. 2) den tiefsauren rosa und in Kresyl- echtviolett violetten Farbton. Ganz analog waren die Farbdifferenzen bei den Indikatorfitrbungen, yon denen ich keine Bilder beigeffigt habe. Ihre Resultate sind in Tabelle 1 zusammengestellt.
Brillant- i Kresyl- Brillant- Neutral- Nilblau- Methylrot Indikator kresylviolett, echtviolett vita]rot rot sulfat
Animales Plasma
Dotterplasma
r o s a
blau
rosa
violettblau
rostgelb
sattrosa
rostgelb
sattrosa
violett
blau
gelb
bleibt ungei~rbt
Bei der Furchung wird durch die ~quatorialen Furchen so, wie das ffir Nerds Dumerilii beschrieben wurde, jeweils die Hauptmasse des am animalen P o l d e r Blastomeren angesammelten alkalischen Protoplasmas yore Rest der Blastomeren, der das Dotterplasma enth~lt, abgetrennt. Mittlere Furchungs- stadien, auf denen dann der Keim aus einer Gruppe yon alkalischen Zellen und aus einer solchen yon sauren Zellen besteht, fiihren das Ergebnis in drastiseher Weise vor (s. Fig. 2 und 3 auf Taf. II) . Diese Stadien weisen stets schon sehr ausgepr~gte Farbdifferenzen aufl). Der groBe Somatoblast hat stets alkalisches Protoplasma.
Die alkalischen Zellen sind es dann, welche bei der epibolischen Gastru- lation um die grol~en Zellen mit dem Dotterplasma herumwachsen, so dad nachher das Ektoderm in allen Indikatoren im alkalischen und das Entoderm in] sauren Farbton gef~rbt erscheint. Es resultieren daraus auDerordentlich schSne zweifarbige Bilder. (Vgl. die mit Brillantkresylviolett gef~rbte Gastrula auf Taf. I I , Fig. 4). Die Intensit~t der F~rbungen erreicht in diesem Stadium ihren HDhepunkt.
Aus dem Gesagten geht hervor, dad sich die an den Eiern yon Nerds Dumerilii ermittelten I tauptbefunde fiber das Wesen und die Auswirkung der bipolaren Differenzierung an den Eiern yon Nereis limbata mit einer viel grSl~eren Zahl yon Indikatoren, n~mlich 5 bzw. 6, best~tigen lieBen. Darfiber hinaus sind eine Reihe yon Einzelbefunden yon groBem Interesse. In der :Beweis- fiihrung meiner ersten Abhandlung blieb insofern eine Lticke often, als sich
1) Das Bild dieser Furchungsstadien ist sehr verschieden, je nachdem, yon welcher Seite man sie betrachtet. In den Eiern der Kultur, der das der Fig. 3 entstammte, waren die 01tropfen besonders rasch zu groi]en 01kugeln zusammengeflossen.

Bipolare Differenzierung der Eizellen von Nereis limbata u. Chaetopterus pergamentaceus 403
Es wurde auch bei diesem Objekt versucht auch durch Mikroinjektionen Indikatoren in die Zellen zu bringen. Hierzu ist nun aber das Ei yon Nereis limbata ein reeht wenig geeignetes Objekt, denn erstens sind die Membranen (zum Untersehied yon denen yon N. Dumerilii) auBerordentlich z~he, und zweitens tr i t t , wenn es schlieBlich doch gelingt, eine Pipet te in den EikSrper einzufiihren, sogleich cytolytische Verquellung der Eisubstanzen ein. Wie weiterhin Kontrollversuehe am ausflieBenden Plasma angestoehener oder zer- drfiekter Eier zeigten, werden die Clarkschen Indikatoren yon den Zellsub- stanzen nur sehr wenig oder gar nicht adsorbiert.
$ $ $
Zur Erg/inzung der Befunde an den Nereis'Eiern wurde noch eine Serie von parallellaufenden Vitalf/irbungen mit Indikatoren an den Eiern yon Chae- topterus pergamentaceus ausgeffihrt. Gesehleehtsreife Tiere sind in Woods Hole in den Sommermonaten leieht zu haben. K/instliehe Befruchtung ist ohne Schwierigkeit ausfiihrbar. Man braucht dazu den Tieren nur je ein mit reifen Geschlechtszellen prall gefiilltes Parapodium abzuzwicken, in Seewasser zu en~- leeren und den Eiern Sperma zuzusetzen. Eine Befruchtung von Eiern, die sehon vorher vitalgef~rbt worden sind, gelingt auch hier nicht, und die F/~rbung der ersten Entwieklungsstadien bleibt blaB. St/~rkere Konzentrat ionen der Farbstoffe als die oben angegebenen wirken, besonders kurz naeh der Be- fruchtung leicht sch/~digend und bringen die Entwicklung zum Stillstand.
Die Chaetopterus-Eier sind viel weniger durehsiehtig als die Nereis-Eier. Trotzdem sind aber die Farben deutlich genug erkennbar und in einigen Indi- katoren treten die verschiedenen Farben der beiden Eih~lften schon kurz nach der RichtungskSrperbildung aufs deutlichste hervor. Sowohl die Sonderung der Substanzen, als auch die Anf/~rbbarkeit derselbeu steigert sich dann noch w~hrend der weiteren Entwieklung ganz betr/~chtlich. Am st/irksten erscheinen die Farbunterschiede wieder im Gastrulastadium (zwischen den Zellen des Ektoderms und des Entoderms).
Die RichtungskSrperbildung ist in etwa 20 Min. vollendet. Die erste Furchungsteilung erfolgt eine Stunde nach der Befruehtung. Die biPolare Um- schiehtung des Eiinhaltes setzt, wie ja das schon die alten eytologischen Untersuchungen festgestellt haben, auch hier nach der RichtungskSrperbildung ein und erreicht sehon vor der ersten Teilung einen ziemlich hohen Grad. In diesem Stadium 1/~$t sich am leichtesten mit Kresylechtviolett eine Anf/irbung beider Plasmakomponenten, sowohl der alkalischen als auch der sauren erreichen. Der ganze hellere Plasmahof um den I~iehtungskSrperpol herum zeigt in diesem Indikator eine zartviolette Farbe, w~hrend das Dotterplasma b~al] grfinlichblau ist. (Zum blauen l~arbton des Indikators kommt die gelbliche Eigenfarbe der I)ot tertropfen hinzu.) Die Verschiedenfarbigkeit ist bei allen Eiern aufs deut- lichste ausgepr/~gt. In den sp~teren Stadien versehiebt sich der Farbton der alkalischen Hiilfte mehr naeh Rosa.
Die entwieklungsmeehanisch interessante Frage, was fiir Substanzen in den kleinen D o t t e r l a p p e n hineinkommen, der sich w~hrend der ersten

404 Spek
Furchungsteilung am vegetativen Pol der einen Blastomere vorfibergehend ab- schnfirt, l~13t sich nach der Kresyleehtf/~rbung dahin beantworten, da6 die Hauptmasse des Dotterlappens zwar so wie die ganze vegetative Blastomeren- hi~lfte aus sauren Kolloiden besteht; an der Oberfl~tche des Dotterlappens (nach vollzogener Abschnfirung besonders an der Kuploe ) ist aber deutlich noch ein Rest yon alkalischer Substal~z zu erkennen. Es machte mir den Eindruck, als ob der Abtransport der alkalischen Substanzen, die ursprfinglich fiber das ganze Ei ausgebreitet sind, yon dem Bezirk, der sich als Dotterlappen ab- schnfirt, verzSgert erfolgt, so da~ eine Diskrepanz gegenfiber der Umgebung entsteht. Die VerzSgerung in der Abwanderung der alkalischen Substanzen dieses Bezirkes nach dem animalen Pole k5nnte ja z. B. durch eine hShere Viskosit~t verursacht sein.
In Brillantkresylviolett ist die Lilaf~rbung des animalen Plasmas vor der ersten Teilung noch sehr blal~, aber immerhin erkennbar. Der Dotter ist deut- lich blau, mit einem Stich ins Grfine. Aul3erdem f~rbt sich in diesem Farb- stoff eine Sorte von grol3en Tropfen im Protoplasma intensiv blauviolett an. Auch sie wandern nach dem RichtungskSrl0erpol und ordnen sich bei der bipolaren Stoffsonderung stets auf das genaueste so zwischen die andern Plasma- komponenten ein, dal~ sie zwischen die lila und die blauen Substanzen zu liegen kommen. Sie liegen also zuerst in einer Querzone in einiger Entfernung vom RichtungskSrperpol. Bei den ~iquatorialen Teilungen kommen sowohl in die animalen, als auch in die vegetativen Zellen einige dieser Tropfen zu liegen. Bei der weiteren Entwicklung werden sie stets aufs neue in den alkalischen Zellen an den sauren und in den vegetativen an den alkalischen Pol geffihrt. An alten Furchungsstadien und Gastrulen ist die alkalische Seite der animalen bzw. ektodermalen Zellen auch in diesem Indikator aufs deutlichste rein rosa gef~irbt und sticht scharf ab gegen den tiefblauen Farbton der dunklen vege- ta t iven (entodermalen) Zellen.
In Brillantvitalrot und Neutralrot f~rbt sich eine im ungeschichteten Ei den ganzen ZellkSrper gleichm~l~ig dnrchsetzende saure Substanz intensiv rosa an. Ihre Abwanderung nach dem vegetat iven Pole erfolgt nut ganz langsam. Aber schliel~lich ist das Endresultat auch bei dieser F/irbung das gleiche: Die animalen H~lften der alten Furchungszellen bzw. die Aul~enh~lften der Ektoderm- zellen erscheinen rostgelb, wi~hrend die entodermalen Zellen rosa bleiben.
So kann nicht gezweifelt werden, dal] das au6erordentlich komplizierte Substanzgemisch, welches der Zellinhalt der Chaetopterus-Eier darstellt, bei der nach der RichtungskSrperbildung einsetzenden bipolaren Schichtung auch so ausgesondert wird, da[~ die alkalischen Substanzen nach dem einen und die sauren Substanzen nach dem andern Pol verlagert werden.
Kurze Zusammenfassung der Hauptresultate Die 1930 an den Eiern yon Nerei8 Dumerilii ausgeffihrte Untersuchung,
welche zur Entdeckung der , ,Kataphorese in lebenden Zellen" geftihrt hatte, wurde mit sechs Indikatoren und erweiterten Problemstellungen an den Eiern der amerikanischen Nereis limbata wiederholt. Die Methode waren wieder

Bipolare Differenzierung der Eizellen von Nereis limbata u. Chaetopterus pergamentaceus 405
Vitalf~rbungen mit Indikatoren und zwar mit Neutralrot, Brillantvitalrot, Kresyl- echtviolett, Brillantkresylviolett, Nilblausulfat B und Methylrot. Methylrot (in isoton. NaC1-LSsuug angewandt) f/~rbt nur die animalen Zellen bzw. Zellh/tlften an. Alle andern Farbstoffe (hier auch Nilblausulfat) fitrben den ganzen Keim leicht und intensiv an. Die F/~rbungen sind hier durch kein natiirliches Pig- ment kompliziert.
Die bipolare Differenzierung beginnt wieder nach der Richtungsk5rper- bildung. Bald nach ihrem Einsetzen ist auch schon eine Zweifarbigkeit des vitalgef~trbten Keimes andeutungsweise zu erkennen. Sie wird dann immer in- tensiver und fiihrt schlieBlich an/~lteren Furchungsstadien zu ganz extremen Farb- differenzen~ deren Sinn wiederum der ist, daI~ am animalen Pol des Eies bzw. der Blastomeren alkalische und am vegetativen Pol saure Kolloide angereichert werden. Der Sinn der Farbumschl/tge aller Indikatoren stimmt iiberein. Das pH der alkalischen Substanzen diirfte bei 7,8 oder noch hSher liegen, das der sauren konnte leider nicht ermittelt werden, da der einzige tiefumschlagende Indikator Methylrot die sauren Substanzen nicht anf/irbt. (Bei Nereis Dumerilii betrug es ungef/~hr 5,5.) Die alkalischen Blastomeren liefern wiederum das Ektoderm, die sauren das Entoderm. Die Farbdifferenzen der Keimschichten der Gastrula sind aui3erordentlich intensiv. In den Einzelzellen des Ektoderms kommt es zur Ausbildung einer neuen bipolaren Differenzierung und zwar so, dal3 in allen die alkalischsten Stoffe an die Aul]enseite, die sauren an die Innenseite zu liegen kommen.
An diesem Objekt gelang in Brillantkresylviolett auch eine Anf/~rbung aller Zonen des konzentrisch gebauten unbefruchteten Eies. In ihm entsteht die eigentfimliche Substanzanordnung so, da6 die alkalischsten Stoffe g l e i c h - m/~13ig an die g a n z e O b e r f l s wandern, die sauren die Zellmitte einnehmen und die Substanzen mit mittlerem pH dazwischenliegen.
Parallelversuche am Chaetopterus-Ei lehrten, dab die bipolare Differen- zierung desselben nach dem gleichen Prinzip erfoIgt.
L i te ra turverze ichnis siehe auf S. 425 in Bd. 9, (1930) dieser Zeitschrift.
Erkl~rung der Abbildungen der Tafel II
Abb. 1. Vierzellenstadium yon 2(ereis limbata, mit Brillantkresylviolett vital gefiirbt. Abb. 2. J~lteres Furchungsstadium yon 1Vereis limbata mit Neutralrot vital gcfi~rbt. Abb. 3. ~lteres Furchungsstadium yon Nereis limbata mit Kresylechtviolett vital gefi~rbt. Abb. 4. Gastrula yon Nereis limbata mit Brillantkresylviolett vital gef~rbt. Abb. 5. Unbefruchtetes Ei yon Nereis limbata mit Brillantkresylviolett vital gef/~rbt.





























