Embed Size (px)
Citation preview

(Aus dem Zoologischen Institut der Universit~tt Leipzig und dem Kaiser Wilhelm- Institut fiir Biologic in Berlin-Dahlem, Abteilung HaxT~cutm~.)
I~BER DIE D I F F E R E N Z I E R U N G DER PANKREASZELLE IN VITRO 1.
Von ]LS]~ FlSC~EI~.
Mit 15 Textabbildungen (17 Einzelbildern).
(Eingegangen am 9. November 1937.)
Einleitung. Im Anschlu6 an die Arbeiten yon RIES (1935 a und b) fiber die Histo-
physiologie und die Differenzierung der Pankreaszelle und FlSCHEI~ und RIES (1936) fiber die Gewebekultur der Pankreaszelle war das Ziel der vorliegenden Untersuchungen, das Verhalten der Pankreaszelle w~hrend ihrer Differenzierungsperiode in vitro zu verfolgen. Es hatte sich ergeben, dal~ die differenzierte Pankreaszelle nach ihrer Explantation ihren nor- malen Arbeitsrhythmus aufgibt, keine Proenzyrngranula mehr bildet und durch Schwinden des Ergastoplasmas und der GoLGI-Substanz sowie durch eine gleichzeitig einsetzende Fettaufspeicherung zwar eine gewisse morphologische Ahnlichkeit mit embryonalen Zellen gewinnt, sich dabei aber keineswegs entdifferenziert, sondern ihre besonderen ,,organo- blastischen" Potenzen bewahrt, so dab sich unter UmstAnden aueh in ~lteren Kulturen, besonders im Mutterstfick, die Zellen wieder zu aeinus- s Gebilden anordnen kSnnen. Diese Befunde standen in Gegen- satz zu den Angaben yon CHLOPIN und PINKUS, nach denen sieh die Pankreaszelle in vitro entdifferenziert. Nicht nur die Pankreaszelle, sondern auch die meisten fibrigen Gewebe sollen sich in vitro zufolge der Auffassung verschiedener Autoren entdifferenzieren. Es kann hier im Augenblick davon abgesehen werden, dab man dem Begriff Entdifferen- zierung oft einen ganz verschiedenen Inhalt gegeben hat und einesteils dar- unter rein morphologische StrukturverAnderungen oder besser Struktur- verminderungen verstand, andernteils meinte, da6 die Zelle durch die Entdifferenzierung wirklich wieder den Charakter einer indifferenten pluripotenten Zelle annehmen kSnnte. Merkwfirdig erscheint auf jeden Fall, da6 den zahlreichen Angaben fiber eine Entdifferenzierung der Zelle in der Gewebekultur eine Reihe von Befunden gegenfiberstehen, nach denen sich indifferente embryonale Zellen in vitro differenzierten und mit der Ausfibung ihrer spezifischen Funktion begannen. Ich denke hier vor allem an die Arbeiten yon OLIvo (1928) fiber die Differenzierung der Herzanlage in vitro, yon STRANOEWAYS und FELL fiber die Differenzierung verschiedener Organanlagen des Hiihnchens, besonders des Auges in
Mit Unterstfitzung der Deutschen Forschungsgemeinschaft.

~ber die Differenzierung der Pankreaszelle in vitro. 641
Gewebekulturen und von FELL sowie FELL und ROBINSON fiber die Bil- dung des Femurs in vitro.
Wenn differenzierte Gewebezellen sieh nach der Explantation ent- differenzieren, warum differenzieren sich dann unter den gleichen Kultur- bedingungen auf friihen Entwicklungsstadien isolierte embryonale Zellen ? Es ersehien lohnend, diese Frage zu kl~ren, indem man einen Zelltyp auf verschiedenen Stadien der ontogenetischen Entwicklung explantierte. Ein geeignetes Objekt daffir war die Pankreaszelle des Axolotl, da sie wohl schon sehr frfihzeitig determiniert wird. Schon die ~lteren Arbeiten von SPEN[ANN ergaben, dab bei den Amphibien die meisten Organanlagen bereits w&hrend der Gastrulationsperiode determiniert werden. Durch Transplantationsversuche konnte HOLTrRETER (1925) nachweisen, dal~ die Anlagen yon Leber und Pankreas bei Anurenkeimen mindestens schon in der ~lteren Gastrula determiniert sein mfissen, da sie sich nach Implantat ion im Wirtskeim stets herkunfts-, nie o r t s g e m ~ weiterent- wickelten.
I m einzelnen habe ich bei den vorliegenden Untersuchungen folgende Fragen zu kl~ren versucht:
1. Differenziert sich die Pankreaszelle in der Gewebekultur aus undifferenzierten Elementen, und bis zu welchem frfihesten ontogene- tischen Entwicklungsstadium ist es mSglieh, aus Entodermzellen Pan- kreasgewebe zu zfichten ?
2. Wieweit gehen die Differenzierungserscheinungen ? Erreichen die Zellen nur das funktionsfs Stadium oder werden auch wirklich schon Proenzymgranula gebildet, und gesehieht dies gegebenenfalls in gleicher Weise wie in der differenzierten Zelle in vitro ?
3. Kommt es nur zu einer rein cytologischen oder auch zu einer organo- logischen Differenzierung ?
4. Is t eine wirkliche Gewebekultur mit unbegrenztem Wachstum der undifferenzierten entodermalen Zellen mSglieh, bei der diese nur auf Kosten der N~hrstoffe des Zfichtungsmediums, nicht der in den Dottersehollen gespeicherten Substanzen, leben ?
5. Welche morphologischen und vielleicht auch physiologischen Unter- sehiede bestehen zwischen den bereits determinierten aber noch nicht differenzierten und den differenzierten bzw. nach der Differenzierung explantierten Pankreaszellen ?
6. L~,l~t sich an den sich differenzierenden Pankreasexplantaten ein Gegensatz zwischen Wachstumsgesehwindigkeit und progressiver Diffe- renzierung nachweisen, wie er yon SCHM~U~H,tUSEN bei der Entwicklung versehiedener Tiere festgestellt werden konnte ?
Material und Teehnik. Als Objekt wurde die Pankreaszelle yon Amblystoma mexicanum deswegen
gew~hlt, weil sie ihrer GrSBe wegen fiir die be~bsichtigten Untersuchungen besonders geeignet ist.

642 Ilse Fischer:
E,'.plantiert wurden: Entoderm aus der Blastula. der vordere mediane Bezirk des Entoderms der Gastrula und der Neurula, aus dem nach HOLTFRETER bei Anuren die Anhangsorgane des Darmes sich absondern, der vordere Teil des Darmes yon Embryonen auf dem Schwanzknospenstadium, schlie~lich die hereits ab- gegliederte Leberpankreasanlage von etwa 10--12 mm langen Embryonen.
Als Zfichtungsmedium diente auf a/3 mit destilliertem, sterilem Wasser ver- diinntes Hiihnerplasma mit und ohne Zusatz yon Embryonalextrakt. Die Ex- plantate wurden nach der yon MAXIMOW angegebenen Methode auf einem be- sonderen kleinen GlimmerplAttchen angesetzt. Dabei ist es m6glich, die Kulturen mit diesem kleinen Glimmer zu waschen und umznsetzen, ohne sie aus dem alteu Ziichtungsmedium herauszuschneiden. Dies war wichtig, weil w~ihrend der Differen- zierungsvorg~nge st~irkere meehanische Reize vermieden werden sollten, auch durften Differenzierungsstrukturen im Gewebe nicht zerst6rt werden, schlieBlich sollten auch nicht durch das Setzen yon ,,~Vunden" l~egenerationserscheinungen ausgel6st werden, die das Bild der Entwicklungsvorg~nge nur gestOrt hatten. Die Kulturen wurden alle 6--8 Tage gewaschen und mit einem sehr kleinen Tropfen yon Plasma und Extrakt geffittert. Um das Wachstumsverm6gen der Explantate zu priifen, wurden gelegentlich auch einzelne Kulturen beim Umsetzen aus dem Medium herausgeschnitten und geteilt. Um den Zuwachs der Kulturen zu ermitteln, wurde eine Reihe t~glich gezeichnet und der Fl~icheninhalt der Umrifl- zeichnungen mit Hilfe eines Planimeters bestimmt.
Da der Embryonalextrakt nach HOL~rFRETER induzierend wirkt, und es nicht sieher ist, ob vielleicht nicht auch das Blutplasma Induktoreigenschaften hat, wurden zur Kontrolle einige Ektodermexplantate und Leber- und Par/kreasanlagen auch in der HOLTFRETERschen RI~O~R-L6sung geziichtet. Diese Explantate wurden in kleinen Glassch~lchen gehalten und etwa alle 10--14 Tage umgesetzt.
Vor der Explantation wurden die Eihiillen au$erlich sterilisiert, indem sie etwa 10Min. lang in eine 0,4%ige LSsung yon Chloramin in Rx~O:ER-IAisung gelegt wurden, hernaeh wurden sie in sterile RI~oE~-LSsung iibertragen, die so oft (etwa 9--12real) gewechselt wurde, bis die Waschfliissigkeit nicht mehr nach Chlor roeh. Dann wurden die ~[eime mit feinen Nadeln steril herauspr~tpariert und die betref- fenden Entodermbezirke bzw. die Organanlagen mit sehr feinen, angeschliffenen Nadeln herausgeschnitten. Die Kulturen wurden bei Zimmertemperatur gehalten. Es hat sich im Laufe der Untersuchungen herausgestellt, dab sie sich am besten im Winter in ungeheizten Zimmern, bei einer Temperatur yon etwa 10--120 ent- wickelten.
Die Kulturen wurden t~glich im Hell- und Dunkelfeld beobachtet, Ver~nderungen protokolliert und zum Tell gezeiehnet. Ffir Vitalf~rbungen verwandte ich haupt- siichlich Toluidinblau, Neutmlrot und Janusgriin in R~OER-LSsung (Farbstoff- konzentration etwa 1:75000); es wurden davon jeweils etwa 1--2 Tropfen dem Kulturmedium zugesetzt.
Ubersichtspr~tparate wurden nach BowN fixiert und mit Hgmalaun gef~rbt; Fet t wurde durch Sudan III-Fi~rbung an Praparaten nachgewiesen, die in Formol- RI~GER-LSsung fixiert worden waren. Zur Darstellung des GoLor-Apparates und der Mitochondrien wurden die Kulturen nach CKAMI~Y fixiert und dann bei 37 o in l%iger Osmiums~ture 3, 7 und 9 Tage nachosmiert.
Die Mikrophotographien wurden mit einer Aufsatzkamera (Makam yon Leitz) und die Zeichnungen mit einem Zeichenajaparat angefertigt.
l~bersicht der Ergebnisse an verschieden alten Explantaten. Die L e b e r b u c h t g l i eder t sieh bei A x o l o t l l a r v e n yon e twa 9 - - 1 0 m m
L~nge ab. Bei den f r f iheren E n t w i c k l u n g s s t a d i e n h a t m a n ke ine abso lu t e

(~ber die Differenzierung der PankreaszeUe in vitro. 643
Sicherheit, die Leber-Pankreasanlage und nut diese zu isolieren. Es entwickelten sich aber die friihembryonalen Entodermexplantate, die urspriinglich aus gro•en, dotterreichen, morphologisch gleichen Zellen bestehen, in der fiberwiegenden Mehrzahl zu histologisch differenzierten Gewebekomplexen. Neben Leber- und Pankreasanlagen bildeten sich vorwiegend Darmanlagen, h~ufig auch Muskulatur, die aus dem Rand- zonenmaterial des Entoderms entstanden sein diirfte, in wenigen F~llen differenzierten sich ferner Herzanlagen, besonders wenn Gastrula- und Neurulaentoderm explantiert worden war, da auf diesen friihen Ent- wicklungsstadien bei der Isolierung l~bergriffe in fremdes Anlagengebiet kaum zu vermeiden sind. Diese Mitnahme yon Zellen anderer prospek- tiver Bedeutung erschwert die Beurteilung der Selbstdifferenzierungs- fi~higkeit der Explantate. Es ist aber nicht mSglich, diesen Unsicherheits- faktor auszuschalten, da die zu Leber- und Pankreaszellen determinierten Elemente eben vor Abgliederung der Leber-Pankreasanlage in keiner Weise als solche zu identifizieren sind.
Niemals ist es mir gelungen, aus Explantaten aus der Blastula Pan- kreaszellen zu ziichten. Aus Blastulamaterial wurden bisher iiberhaupt keine typischen Gewebekulturen erhalten. Die ZeUen flossen bei der Ex- plantation und beim t~berffihren in das Ziichtungsmedium auseinander und starben spi~testens nach 14 Tagen ab, ohne dab sie irgendwelche Wachstums- und Differenzierungserscheinungen gezeigt hs Aller- dings wurden, um die auf diesem Stadium noch m5glichen Regulationen auszuschlieBen, immer relativ kleine Teile der Blastula explantiert. Mit diesen jiingsten Entwicklungsstadien konnte ich bisher jedoch nur ver- h~ltnisms wenig Versuche anstellen, deswegen m5chte ich nicht ent- scheiden, ob die Kulturen infolge irgendwelcher technischer Fehler bei der Explantat ion nicht angingen, oder ob bci ihnen eine Gewebekultur an sich nicht mSglich ist.
Fiir unsere spezielle Fragestellung genfigte es indessen, die Experi- mente mit der Gastrula zu beginnen.
Typische Pankreaszellen differenzierten sich aus Gastrulaentoderm ebenso wie aus defn Entoderm der Neurula und in Explanta ten aus ~lteren Embryonen. Tabelle 1 gibt eine Obersicht der Entwicklung
Gewebe
Gastrulaentoderm . . . . Neurulaentoderm . . . . S chwanzknospenstadium . Abgegliederte Leber-Pan-
kreasanlage . . . . .
Tabe l l e 1.
Zahl der
Exp lan- t a t e
30 36 12
33
Pankreas
1
18
Differenzier t zu
andere Gewebe, Lebe r besonders Da rm,
Muskula tur , Herz
i ~ 1 10 5
24 - -
Nlcht differen-
z ier t
10 6 4

644 Ilse Fischer:
von Pankreas- und Leberzellen aus verschieden alten Keimen. Kulturen, die infolge Infektion zugrunde gingen oder vor der Differenzierung zu Vitalf/~rbungen bzw. Fixierungszwecken verwandt wurden, sind nicht mit in die Tabelle aufgenommen worden.
Aus dieser Ubersicht folgt, da6 die Pankreaszelle sich in Entoderm- explantaten aus der Gastrula ebenso differenziert wie in sotchen von /~lteren Entwicklungsstadien. Wenn sich allerdings um so weniger Explan- tate zu Leber- oder Pankreasgewebe entwiekelten, je jiinger der Em- bryo war, aus dem das Material stammte, so ist dies einfach damit zu erkl/~ren, da6 die Isolation dieses Anlagenkomplexes auf den friihen Ent- wicklungsstadien mehr oder weniger vom Zufall abh/s Immerhin besteht etwa bis zum Neurulastadium eine gewisse Gew/~hr dafiir, da6 man die Leber- und Pankreasanlage mit explantiert, wenn man den vorderen medianen Bezirk des Entoderms isoliert. Indessen ist es unver- meidlich, dal3 gleiehzeitig Zellen aus anderen Anlagengebieten, vor allem dem des Darmes, mit entnommen werden. Daher differenzierten sieh sp/~ter in den Kulturen, aul3er Leber und Pankreas auch andere Gewebe, besonders h/iufig solehe, welche die typischen Strukturen der den Darm aufbauenden Zellen erkennen lie6en. Gelegentlieh wurden wohl auch Komplexe yon mesodermalen Zellen mit explantiert. Daftir sprieht, dab in einigen F/~llen besonders bei Gastrulaexplantaten auch eine Differen- zierung yon Muskelgewebe erfolgte. In zwei Explantaten bildeten sieh aueh typische Herzanlagen, die nach einiger Zeit zu pulsieren begannen und 4 bzw. 6 Wochen lang in einem gleiehbleibenden Rhythmus zuekten.
Leber- und Pankreaszellen differenzierten sieh meist gleichzeitig in den Explantaten.
Das h~ngt wohl damit zusammen, dab die Bezirke ihrer pr~sumptiven Anlagen unmittelbar benaehbart sind.
Ein Teil der Kulturen hat sich nicht differenziert. Diese Explant~te hatten eine relativ geringe Zellauswanderung gezeigt und die ausge- wanderten Zellen sich zu nicht immer liickenlosen Membranen zusammen- geschlossen. Das kSnnte vielleicht den Gedanken aufkommen lassen, da~ es sich bei den Explantaten, in denen sieh spezffische Gewebe diffe- renziert haben, um abhs Differenzierungsvorgs gehandelt haben k5nnte, und dab in den Kulturen, die sich nieht differenzierten, irgendwelche Induktoren, die vielleicht in besonderen Zellkomplexen lokalisiert sein kSnnten, nicht zur Wirkung gelangten, weil die betreffenden Zellen in den Explantaten fehlten. Allein es gibt eine viel zwanglosere und einfachere Erkl~rung fiir diese Erscheinung. Ein gro~er Teil der Entodermzellen gelangt in vivo w~hrend der Entwieklung in den zen- tralen Bezirk des Rumpfdarmes; diese Zellen werden, ohne sich irgendwie besonders zu differenzieren, resorbiert. Wahrscheinlich bestanden die Kulturen, die sich nieht differenziert haben, haupts~chlich aus solchen Nahrungsdotterzellen.

0ber dic Differenzierung der Pankreaszelle in vitro. 645
Nach der Holffreterschen Methode in Salzliisungen aufgezogene Entodermexplantate.
Nach ItOLTFRETER ist der Hiihnerembryonalextrakt besonders reich an induzierenden Stoffen, undes laitt sich yon vornherein nicht absehen, ob nicht vielleicht auch das Blutplasma Induktoreigenschaften hat. Um zu entscheiden, ob die beobachteten Differenzierungserscheinungen nicht vielleicht durch induzierende Eigenschaften des Mediums hervor- gerufen worden waren, habe ich einige Explantate auch in der von HO:LTF~ETE~ modifizierten RINGER- oder in entsprechend verdiinnter Tyrodel6sung kultiviert. Die Aufzucht von Entodermexplantaten in Salzl6sungen ist sehr schwer, worauf schon HOLTFRETER hingewiesen hat. Es gelang auch ihm unter zahlreichen Versuehen nur in wenigen Fs aus isoliertem Material in Salzl6sungen typisehes Darmgewebe aufzu- ziehen.
In Salzl6sungen kugelten sich die Entodermexplantate ss ab, nach 6--8 Tagen ,,zerliefen" sie aber meistens, und zwar merkwiirdiger- weise um so eher, je weiter fortgeschritten das Entwicklungsstadium war, auf dem sie entnommen wordcn waren. Nur wenige Explantate (Em- bryonen, kurz vor Abgliederung der Leberbucht) blieben bis zu ihrer Differenzierung am Leben. Sie formten sieh zu blasigen Gebilden, die in 6 F~,llen Darmzellen erkennen liel]en. Diese Explantate wiesen ss die eigenartige yon A. Fzsc~E~ 1930 und HOLTFRETER (1931) beschriebene Inversion der Darmzellen auf. In 3 Fitllen hatten sich leberartige Struk- turen gebildet; in zweien waren daneben auch typische, zu unregelm~Bigen Aeini angeordnete Pankreaszellen zu erkennen.
Diese, wenngleich verh~ltnism~l~ig wenigen positiven Experimente zeigen, dal3 die Differenzierung der Pankreas- und Leberexplantate nicht allein durch die induzierende Wirkung des Embryonalextraktes bedingt ist. Zwar verl/~uft der Differenzierungsprozel~ in vitro in reinem Plasma oder in einem Gemisch yon Plasma und Embryonalextrakt in der Regel etwas schneller. Vermutlich bietet ein solches Medium den Zellen giin- stigere allgemeine Lebensbedingungen als eine Salzl6sung. Das zeigt sich vor ahem auch darin, dal~ die in Plasma gezfichteten Explantate hie zerfielen und im allgemeinen l~nger am Leben erhalten werden konnten, als es in Salzl6sungen m6glich war.
Durch die Ziichtung in indifferenten Salzl6sungen ist jedoch noch nicht zu entscheiden, ob bei der Differenzierung der Leberpankreasanlagen das bUS anderen Anlagenkomplexen mit fibernommene Material eine induzie- rende Wirkung ausgefibt hat. Denn hier kann natiirlich ebenso durch einzelne, aus anderen Anlagen stammende Zellen eine Induktion erfolgen. Indessen erscheint das Selbstdifferenzierungsverm6gen der Leberpan- kreasanlage aber dureh die schon erw~hnten Transplantationsversuche HOLTFRETERS gesichert: Er implantierte diese Anlagenkomplexe auf ver- schiedenen Differenzierungsstufen in die ventrale Dotterzellenmasse von

646 Ilse Fischer:
Embryonen, die das Schwanzknospenstadium erreicht hatten (Anuren). Sowohl bei homSo- wie xenoplastischer Transplantation gingen aus der Leber- oder Pankreasanlage nie Teile der Darmwandung hervor, sondern (lie Implantate entwickelten sich immer herkunftgem/iB welter.
Die Entwicklung in vitro dauerte stets wesentlich liinger, sowohl ill Plasmakulturen als in SalzlSsungen als die Entwicklung der Organe bei den unter gleichen Temperaturbedingungen gehaltenen Kontrolltieren, und zwar scheint die Differenzierung der aus /ilteren Entwicklungs- stadien stammenden Explantate relativ am meisten vcrz5gert zu sein. Explantate, die auf dem Stadium der eben angedeuteten Leberbucht entnommen worden waren, erschienen meist noch gar nicht differenziert, wenn die Kontrolltiere schon 1/ingere Zeit Nahrung aufgenommen hatten, und bei diesen bereits vSllig differenzierte und dotterfreie Leber- und Pankreaszellen vorhanden waren. Genauere Zeitangaben lassen sich nach den vorliegenden Protokollen nicht machen, da die Entwicklungsprozesse anscheinend weitgehend yon der ~uBeren Temperatur abh/ingig sind und (tie Kulturen leider nicht unter konstanten Bedingungen gehalten werden konnten.
Das Verhalten des isolierten Entoderms nach der Explantation. Nach der Explantation zieht sich das Gewebe stark zusammen und
kugelt sich sowohl in SalzlSsung als auch in einem Plasmatropfen meist vSllig ab. Bei den jfingeren Stadien, yon der Gastrula bis zur Neurula, bemerkt man in den ersten drei Tagen nach der Explantation eine geringe Zellauswanderung. Die ausgewanderten Zellen schlieBen sich zu einer Membran zusammen. In SalzlSsungen kommt es zu solchen Membran- bildungen nur, wenn das Explantat auf den Boden des Zuchtsch/tlchens gesunken ist. Will man das Explantat aus der Schale entfernen, so reiBt die Membran regelm/iBig ab, ein Zeichen, dal~ die Zellen sehr lest am Glasboden haften. In den so gebildeten Wachstumszonen sind verh/ilt- nism/iBig wenig Zellteilungen zu beobachten. Durch t/iglich hergestellte UmriBzeichnungen ergibt sich auch, dab die F1/tchenvergr5Berung der Membranen relativ gering ist. Das Wachstum der Membranen erlischt aber schon nach einer Woche v611ig. Nun zeigen zwar nach der Methode des latenten Lebens in Carrelflaschen (R. PAI~KER 1936) oder nach der MAXI~Owschen Methode (I. FISCgER 1937) geziichtete Gewebekulturen stets nach 10--15 Tagen einen Wachstumsstillstand. Indessen wachsen solche Gewebekulturen dann immer wieder yon neuem aus, wenn man sie aus dem alten Plasmakoagulum herausschneidet und in einen frischen Mediumtropfen setzt. Bei den Versuchen yon P~KER war dies auch noch m5glich, nachdem die Kulturen 1 Jahr lang im Zustand vSlliger Wachstumruhe gehalten worden waren. Bei den Entodermexplantaten lieB sich eine erneute Wachstumsanregung nun niemals beobachten. Wenn man sie umsetzt und teilt, so wachsen die Zellen nicht wieder aus; die

0ber die Differenzierung der Pankreaszelle in vitro. 647
Subkulturen kugeln sich nur noch starker ab, wobei die an der Oberfl~che gelegenen Zellen ein flaches Epithel iibcr den Explantaten bilden.
Explantate aus i~lteren Em- b~Tonen und besonders die Ex- plantate der bereits abgeglieder- ten Leberbucht bilden meistens iiberhaupt keine Wachstumszonen aus. Erst sp~ter (nach 14 Tagen bis 3 Wochen), wenn die eigent- lichen Differenzierungsprozesse bereits einsetzen, wachsen diese Kulturen aus (Abb. 1). Die kurz Abb. 1. E n t o d e r m k u l t u r aus tier Gastrula.
Nach 5 Woehen h a t t e n sich in v i t ro Driisen- v o r ihrer Differenzierung explan- schl~tuche gebildet , haupts~chl ich im Mut te r -
stiick, links auch in der Wachs tumszone . tierten Leber- und Pankreaszellen Rechts Entodermzel len , die sich n ich t differen- machen in vitro also zun~chst eine zlert haben (Nahrungsdot te rze l len ?). Bouin,
H~malann . Verge. 17fach. gewisse Latenzzeit durch. W~hrcnd der Differenzierungsprozesse beginnen auch die Entodermexplantate, die von jiingeren Keimen stammen, einen neuen Zuwachs zu zeigen, der kaum auf erneute Zellenaus- wanderung, sondern wohl ausschlieBlich auf Zellver- mehrung zurtickzufiihren ist.
Die Differenzierungser- scheinungen beginnen und sind am auff~lligsten im Mut- terstiick der Kulturen. Sie erstrecken sich nicht nur auf die Differenzierung der Zellen an sich, sondern zugleich zeigen sich typische histologi- sche Formbildungsprozesse. In den Pankreasexplantaten formieren sich deutlich Acini (Abb. 2, 3 und 7); die Leber- zellen ordnen sich rosetten- fSrmig an und bilden schliei~- lich ganz wie in der Leber Tubuli. In Darmexplantaten Abb. 2. Polaris ier te , zu Acini angeordnete , berei ts
m i t P roenzymgranu l a beladene Pankreaszel len, die ents tand manchmal eine weir- sieh in e inem Neuru l aen tode rmexp lan t a t differen- gehend den Verh~ltnissen in ziert haben. Mut ters t i ick lebend photogeaphier t .
Vergr. e twa 500fach. vivo entsprechende Struktur der Darmwandung, nur lag das Epithel vielfach auBen (vgl. A. FISCHER 1930). Sparer zeigten diese Explantate sogar deutlich peristaltische Bewegungen. Wieweit die Differenzierungsprozesse gehen, das wurde
Z. f. Zel l forschung u. mikr . Anatomie . Bd. 27. 43

648 llse Fischer:
besonders eindeutig demonstriert, wenn - - unbeabsichtigt - - Herzan- lagematerial mit explantiert worden war. Es bildeten sich schlauch- oder blasenartige Komplexe, die Kui~erlich zwar vielfach eine recht unregelmKBige Gestalt annahmen, aber nach einiger Zeit zu pulsieren begannen. Dabei war eine regelm~i~iger Rhythmus der Bewegungen zu beobachten, obwohl diese Gebilde nicht mit einem Blutgef~tBnetz in Verbindung standen. In einigen F~llen pulsierten diese Herzkulturen 8 Woehen lang.
W/~hrend der Differenzierungsperiode kommt es in den Explantaten also auch zu typischen histologischen Formbil. dungsprozessen, wobei der Bau der Or- gane, zu denen sich die betreffenden
Abb. 3. Abb. 4.
Abb. 3. I n typ i scher Weise ac inus f6 rmig angeordne te Zellen aus dem Mut te rs t i i ck e iner En tode rmku l tu r . A m Zellapex f inden sich berei ts iiberall P roenzymgranu la , in den meis ten Zellen ist ein ne t z f6 rmige r GoLot-Appara t vorhanden , oder es sind wenigs tens noch einzelne S t range yon GOL~I-Substanz erhal ten. Die Mitochondr ien wurden ebenfalls impr~igniert. Die Lipoehondr ien f inden sich me i s t basal, un te rha lb des Kernes . F ix i e r t nach CIt~MP]=,
9 Tage osmiert . Vergr. e twa 500fach.
Abb. 4. GroI~e undif ferenzier te Entodermzel len aus der Gastru]a, m i t sehr viel Dot terschol len und einzelnen P igmen tk6 rnchen . Vergr, e twa 900fach.
Gewebe in vivo entwickelt haben wfirden, mehr oder weniger weitgehend nachgeahmt wird. MAxIMow hat solche Formbildungsprozesse in Gewebe- kulturen als ein organoides oder organotypisehes Wachstums bezeichnet. Es war nun in allen F~llen eine weitgehende Parallelit~t zwischen den Formbildungsprozessen und den Vorg~ngen der cytologischen Differen- zierung festzustellen. Die Zellen differenzieren sieh nicht erst und ordnen sich dann in gewebetypischer Weise, und lassen aueh nieht etwa umgekehrt erst eine bestimmte Zusammenordnung erkcnnen, um dann die Merk- male ihrer besonderen Struktur auszubilden. Die Vorg~,nge der Zelldiffe- renzierung und der geweblichen Formbildung laufen vielmehr gleichzeitig

Uber die Dffferenzierung der Pankreaszelle in vitro. 649
nebeneinander ab. Wghrend also die ersten Anzeichen der Zelldffferen- zierung sich geltend machen, erfolgt auch schon die ,,Ordnung" der Zellen im Gewebeverband.
Die Differenzierungs- und Formbildungsprozesse waren stets am deutliehsten im Mutterstfick der Kulturen. In der Wachstumszone waren sie, offenbar durch die Einordnung der Zellen in das Plasma- medium, gehemmt. Immerhin bildeten sich ausnahmsweise auch in der Wachstumszone zu dieser Zeit typische Driisenschlguche (Abb. 1).
Gegen Ende der Differenzierungsperiode zeigt sich immer deutlicher ein zunehmendes histiotypisches Wachstum der differenzierten Zellen
Abb. 5. Entodermzel len aus der Gas t ru la m i t Toluidinblau vi ta l gef~trbt. Keine Farbstoff- speicherung durch Cytoplasmaorganel le . u Kernfa rbung . Vergr. e twa 850fach.
(s. S. 655 und Abb. 11 und 15). Es ist yon nun an auch m6glich, Sub- kulturen yon den Geweben anzufertigen, die nach dem Umsetzen aus- wachsen und in mehreren Passagen weitergeziichtet werden k6nnen.
Die Zelldifferenzierung in den Leberexplantaten verli~uft recht kom- pliziert, da in diesen Zellen mehrerc funktionellc Prozesse nebeneinander hergehen. Auch ist es schwierig, diese Prozesse zu analysicren, da in den Leberzellen stets sehr viel Pigment gespeichert wird. Ich werde reich daher im folgenden darauf beschri~nken, nur die Differenzierungserschei- nungen an der Pankreaszelle und ihre spi~teren Vcr~nderungen beim Auswachsen ngher zu schildern.
Die Zelldifferenzierungsvorg~inge in vitro. Bis zum Beginn der Differenzicrungsperiode lassen die undffferenzier.
ten Entodermzellen der verschiedenen Entwicklungsstadien keine wesent- lichen Vergnderungen erkennen. Sie sind zun~chst noch ganz mit Dotter- schollen voUgepropft. AuBerdem enthalten sie viele Fettr6pfchen yon verschiedener Gr61~e, die sich yon den Dotterschollen auch in der
43*

650 llse Fischer:
lebenden Zelle durch ihr st/~rkeres LiehtbrechungsvermSgen unterseheiden lassen. Der Kern erscheint in ungesch/~digten Zellen nur als heller, strukturloser Hof. Seine Form wird oft durch die Lage benachbarter Dotterschollen, die ihn manchmal stark eindriicken, beeinflul3t (Abb. 4 und 6). Zwischen den Dotterschollen und den FettrSpfchen sind feine t ' igmentk6rnchen verteilt, die wohl noch direkt yon der Eizelle abstammen. Sie zeigen die BRow~sche Molekularbewegung, woraus man schliel]en kann, dal3 das Grundplasma der Zellen ziemlich diimffltissig ist. Die Mitoehondrien sind in der lebenden und ungef/~rbten Zelle nieht sichtbar; sie sind erst nach Anf/~rbung mit Janusgrtin oder in osmierten Pr/~pa- raten als lange feine F/~dchen zu erkennen. Ein GoLoI-Apparat ist weder in der lebenden Zelle nachzuweisen, noeh 1/~i~t er sieh in naeh ClUmPY fixierten Pr/~paraten mit Osmiums/~ure impr/ignieren. Auch Zusatz basi- seher Vitalfarbstoffe zu den Kulturen ergibt kein Anzeichen ftir das Vor- handensein eines Gol~oI-Apparates, der immer an einer intensiven Farb- stoffausfloekung in einer bestimmten Zone der Zelle zu erkennen ist. An osmierten Pr/~paraten ist noeh bemerkenswert, dal] das Grundplasma tiberhaupt nur sehr wenig osmiophil ist.
Besonders charakteristiseh ist das Verhalten der undifferenzierten entodermalen Zellen gegeniiber basischen Vitalfarbstoffen. Sowohl die Zellen des Mutterstiickes als aueh die der Wachstumszone haben nur ein /~uBerst geringes Speicherungsverm6gen fiir alle Vitalfarbstoffe. Nur die Pigmentk6rnehen f/~rben sich etwas an. Eigentliche Speicherorganelle (Lipoehondrien) fehlen. Ieh habe aueh niemals eine Anf/~rbung der Mito- chondrien mit anderen basischen Vitalfarbstoffen als Janusgriin beob- aehtet, die in den versehiedensten differenzierten Geweben sonst relativ oft zu bemerken ist. Aueh in vivo lassen sieh die Entodermzellen auf den friihembryonalen Entwicklungsstudien nieht vital anf/~rben, l~IES (1935) und GEI~SCI~ und RI~S (1936) stellten fest, dal3 die undifferenzierte Pan- kreaszelle des Axolotl und des Htihnchens ein versehwindend geringes Speieherungsverm6gen gegeniiber basisehen Vitalfarbstoffen besitzt. Die Dotterschollen bleiben meist auch nahezu ungef/~rbt (Abb. 5 und 6). Nur in absterbenden Zellen zeigen sie eine unregelm/~l~ige, mehr oder weniger starke Ald/~rbung. Manehmal nehmen die Reste zerfallender Dotter- schollen etwas mehr Farbstoff auf. Das geringe F/~rbungsvermSgen der undifferenzierten Entodermzellen beruht jedoeh nieht etwa darauf, dab der Farbstoff nieht in die Zelle eindringt, sondern tats/ichlieh auf dem Fehlen bzw. der Inaktivierung der Speieherungsorganellen. Dies wird besonders deutlieh dureh das Auftreten yon vitalen Kernf/~rbungen. Dabei fgrben sieh nieht nur die Ruhekerne sondern auch die Chromo- somen (Abb. 6), sehon in der Prophase kann man deutlieh den L/~ngs- spalt der Chromosomen erkennen. Zu solehen vitalen Kernf/~rbungen kommt es in diesen Kulturen relativ oft, und vor allem, wenn seit dom letzten Wasehen bereits einige Tage vergangen sind. Die Kernf~rbungen

g'ber die Differenzierung der Pankreaszellc in vitro. 651
m6gen dureh eine Sauerstoffverminderung (s. NASSONOW 1930 und ALEXA~DROW 1932) und vielleicht auch mit durch eine Ver~nderung der Kerneiweil~e durch Anh~ufung von Stoffwechselprodukten im Medium bedingt sein. Daffir spricht, dal~ diese Kernf~rbungen reversibel sind. Sie lassen sich in R~Gl~-L6sung wieder auswaschen. DaI~ nicht allein der Sauerstoffmangel die Kernfs bedingt, geht daraus hervor, dal~ das F~,rbungsbild etwas anders ausf~llt, wenn man die Kulturen in farb- stoffhaltige, ausgekoehte RI~G~-L6sung einlegt. Man bekommt dann
Abb. 6. Vitale Kernf~.rbung mi t Toluidinblau un te r Sauersto~fmangel in eincr Gewebc- ku l tu r en todermale r embryona le r Axolotlzellen, Die F~rbbarke i t t r i t t gleichzeit ig und g le ichar t ig in den freien Chromosomen eines P rophases t ad iums wie auch in den Chromat in - s t r u k t u r e n eines frt ihen Prophasekerns vo r Aufl6sung der Membran und eines Ruhekernes
ein. (Aus I~IES: Z, Zellforsch. 26, nach e inem P r ~ p a r a t Yon I . FISCHER.)
regelmi~13ig eine zarte Anfarbung der Kerne, aber ohne dai~ dabei die Kernstrukturen besonders deutlieh werden. NAssosow hat das Auf- treten der Kernfi~rbung mit dem Erl6schen von Oxydationsprozessen zusammengebracht, welche die Energie fiir die Granulaspeicherung liefern sollten. MAKAROV nimmt an, dal~ Glykolyse einsetzt, nachdem die Zellatmung ausgesehaltet ist, und dureh die frei werdende Milchs/~ure Gelbildungen im Kern hervorgerufen werden. Fiir eine eingehende Be- spreehung, wie diese Kernf/~rbungen m6glicherweise entstehen, sei auf die Untersuchungen von RIES (1937c) fiber die Fgrbbarkeit absterbender Zellen verwiesen. Nach den yon RIES ausgeftihrten Experimenten absor- bieren die ursprfinglieh hochdispersen und weitgehend hydratisierten Kerneiweil~e naeh Dispersit/itsverminderung und Dehydration in iihn- licher Weise Farbstoff wie andere ,,ausfallende" bzw. ,,entmischte" EiweiBsubstanzen in gesch/~digten Zellen oder auch in Modellversuchen. In sauerstofffreier RINOwR-L6sung fand iibrigens auch meist eine

652 Ilse Fischer:
zunehmende Anreicherung des Farbstoffes in den Dotterschollen und manchmal auch in den Fettr6pfchen statt.
Es ist besonders bemerkenswert, daft die undifferenzierten entoder- malen Zellen in den Explantaten wenig Veri~nderungen gegenfiber ihrem urspriinglichen Zustand in r ive aufweisen. Vor allem kommt es hier nicht zu der ffir Gewebekulturen sonst charakteristischen Verfettung der Zellen. Die Zahl der vorhandenen Fettr6pfehen in den Zellen der Wachstumszone nimmt auch nicht zu. Die einzige Ver/inderung dieser Zellen besteht darin, dab sie in der Kultur flach ausgebreitet werden. Ffir die Zellen im Muttersttick trifft nieht einmal dieses zu.
Als erstes Anzeichen der beginnenden Differenzierung bemerkt man meist eine gewisse ,,Aufhellung" der Explantate, was mit der zunehmenden Resorption der Dotterschollen zusammenh/ingen mag. Hand in Hand damit ist eine allm/ihliche Verkleinerung der Zellen festzustellen. AuBer- dem wird das Grundplasma der Zellen wohl wasser/~rmer, es erlischt die B~ow~sche Molekularbewegung der Pigmentk6rnchen, die sieh vielfach zusammenballen. In fixierten und osmierten Pr/~paraten zeigt sich mit dem Beginn der Zelldifferenzierung eine fast sprunghafte Zunahme der Osmiophilie des Grundplasmas. Dies trifft sowohl fiir die Pankreaszellen als auch fiir die Zellen anderer Gewebe zu.
Als Pankreaszelle ist eine sich differenzierende Entodermzelle ill den Kulturen aber erst an der Bildung der ersten Proenzymgranula mit Sicherheit zu erkennen. Sie linden sich meist zuerst in den aeinusf6rmigen Zellaggregaten (Abb. 2 und 3) am distalen Ende der Zellen. Die Preenzym- granula sind zun/tehst in der lebenden Zelle nut schwer yon den Fett- tr6pfchen zu unterscheiden, sie sind als solehe haupts&ehlich durch ihre regelm/iBige Gr6Be und durch ihre dichte Zusammenlagerung zu erkennen. Die ersten Proenzymgranula gehen offenbar direkt aus den Fettr6pfchen der embryonalen Zellen hervor; denn sic erscheinen in den Zellen, noch ehe vitalfitrbbare Lipochondrien nachzuweisen sind. Diese Beobachtung st immt mit den Befunden yon RIEs (1935) fiber die erste Bildung des Sekretes in der embryonalen Pankreaszelle des Axolotl in r ive fiberein. RIES nimmt jedoeh an, dab diese Vorgiinge weitgehend denen der Sekretbildung in der differenzierten Pankreaszelle entsprechen, da es sich bei jenen FettrSpf- chen wahrscheinlich um Lipochondrien handelt, die w/~hrend der Eient- wicklung (lurch Speicherung yon Neutralfetten angeschwollen waren und dadurch ihr Vitalf/~rbungsverm6gen verloren hatten (vgl. RIES und vA~ W~rz~ 1934}.
W/~hrend sich die Acini formieren und die ersten Proenzymgranula gebildet werden,/indert sich das Vitalf~rbungsbild der sich differenzieren- den Zellen. An einigen der apikal gelegenen Fettr6pfchen bemerkt man gelegentlieh vitalf/irbbare Hiillen oder Polkappen. Manchmal beobaehtet man aueh im Zentrum der Acini eine regelrechte Farbstoffausf|ockung

l~ber die Differenzierung der Pankreaszelle in vitro. 653
(Abb. 7 a und b), wie etwa im Pankreas der erwachsenen Maus auf den- jenigen Stadien des Arbeitsrhythmus, auf denen in den Zellen ein GOLGI- Apparat in Erscheinung tr i t t (vgl. RIVES 1935 a). Diese Zellen enthalten oft noch keine typischen vitalf/~rbbaren Lipochondrien. Aber mit fort- schreitender Differenzierung zeigen auch einige der basal gelegenen Fett- tr6pfchen ein sich allm/~hlich steigerndes Speicherungsverm6gen fiir basische Vitaliarbstoffe. Bemerkenswert ist jedoch, dab an diesen Lipochondrien zun~chst noch keine Tcilungserscheinungen auftreten. Einzelne Zellen differenzieren sich auch in der Wachstumszone bzw.
es gelangen infolge des nun einsetzenden Zellwachstums durch Mitosen differenzierte
~fuck X ~ , , J , ~
a b
A b b . 7a u n d b. S t a r k e F a r b s t o f f a u ~ f l o c k u n g i m Z e n t r u m e ines A c i n u s n a c h F ~ r b u n g m i t T o l u i d i n b l a u . a l e b e n d p h o t o g r a p h i e r t , b g e z e i c h n e t . V e r g r . ~ e t w a 750fach , b e t w a 400 fach .
oder sich differenzierende Zellen in grSl~erer Zahl aus dem Mutterstiick in die Wachstumszone. In der l~egel geht dabei sehr bald die acinus- fSrmige Anordnung und die polare Orientierung der Zellen wieder verloren. Einige dieser Zellen weisen aber noch die wesentlichen Merk- male sekretbildender Pankreaszellen auf. Manche sind mit Proenzym- granula ganz vollgepropft (Abb. 9) und andere zeigen in einer dem Kern benachbarten Zone bei Vitalf~rbungen eine starke Farbstoff- ausflockung, die auf das Vorhandensein eines GoLGx-Apparates hin- deutet (Abb. 8). Ganz entsprechend lassen sich an dieser Stelle in nach CHAMPY fixierten Pr~paraten auch mit Osmiums~ure Struk- turen impr~gnieren, die durchaus einem GoLGI-Apparat entsprechen (Abb. 3, 12a).
Die Mitochondrien lassen im Zusammenhang mit der Zelldifferenzie- rung keine charakteristischen Ver~nderungen erkennen. Sie erscheinen noch immer als dieselben gleichm~ig in der ganzen Zelle verteilten langen schmalen Fs wie in den embryonalen Zellen (Abb. 10).

654 llse Fischer:
Weder in den Zellen des Muttersttickes, noch in denen, die sich in der Wachstumszone differenzieren, habe ich jemals eine Ergastoplasmabildung beobachtet (Abb. 2, 7, 8). Es ist zwar eine zunehmende ,,Verdichtung" des
Abb. 8. Abb. 9. Abb. 8. Eine in der "Wachstumszone sich differcnzierende Zelle. S tarke Farbstoffau.,- f lockung im Be~eich des GoLGI-Apparates w/~hren4 der Bildung der ers ten P roenzymgranu la .
Gefhrbt m i t Toluidinblau, lebend photographier t . Vergr. e twa 1200fach. Abb. 9. Fer t ig differenzierte, m i t P roenzymgranu la s ta rk beladene Zelle in der Wachs- tllmszone. Keine polare Orientierung, keine Farbs toffausf lockung mehr , nu t noch sehr
wenige Dotterschonen. Vergr. e twa 1200fach.
Abb. 10. Mi~ochondrien, Lipochondrien und Fet t t rSpfchen, in Zellen aus der hist iotypiscb wachsenden Ep i the lmembran einer Pankreaskul tur , die sich in v i t ro differenziert ha t t e .
Fixier t nach GItAMPr, 8 Tage osmiert . Vergr. e twa 1200fach.
Plasmas zu bemerken; aber es konnte an Kulturen, die in dem BouIN- schen Gemisch fixiert worden waren, niemals eine fibrillenartige Koagula- tion im Orundplasma festgestellt werden.

Ober die Differenzierung der Pankreaszellc ia vitro. 655
Die Strukturen des Kernes wiesen w/ihrend der Zelldifferenzierung kaum besondere morphologische Ver/~nderungen auf. Manchmal war eine Vermehrung der Nukleolen zu bemerken.
Die Dotterschollen werden w/s der Differenzierungsperiode nahezu vollstitndig resorbiert.
Das weitere Schicksal der Explantate in der Kultur.
Nach dem Eintrlt t der Differenzierung nimmt dieWachstumsintensit~t der Kulturen fast sprunghaft zu (s. S. 658), und es bilden sich groBe, schDne Wachstumszonen. D~Herenzierte Pankreaszel len
Die Wachstumsmembranen bestehen bei den meisten Kul- turen vorwiegend aus Epithel- zellen (Abb. 1 und 11), nur sel- ten zeigt sich ein geringes binde- gewebiges Wachstum. In den Epithelzellen werden nun die letzten restlichen Dotterschol- len resorbiert. Die Proenzym- granulabildung h6rt damit v61- lig auf. Die Zahl der vorhan- denen Proenzymgranula in den einzelnen Zellen verringert sich naturgem/s bei jeder Zell-
Abb. 11. E n t o d e r m k u l t u r aus tier Neurula . Es teilung, auBerdem scheinen die h a t t e n sich P a n k r e a s - und Leberzel len differen-
eDen erst gebildeten Proenzym- ziert . N a c h der Di f fe renz ie rung h a t bere i t s das s t a rke re W a c h s t u m tier Epi the lze l len in der
granula aber mit der Zeit auch ,,~Vachstumszone" begonnen; alas Mutterstfick ist dahe r bere i t s yon e iner r e l a t iv groilen
resorbiert zu werden. Im Cyto- Epithelmembran umgeben. Vergr. 17fach. plasma der Zellen bilden sich vielfach kleinere oder grDBere Vakuolen, in denen die Proenzymgranula allmiihlich aufgel6st werden. Ebenso schwindet in den Zellen der Wachstumszone der GoLoI-Apparat. Zun/~chst linden sich in osmierten Pr/iparaten in den Zellen, meist in unmittelbarer Nachbarschaft des Kernes noch - - allerdings recht unregelm/~i~ige Str/inge von GOLGI- Substanz (Abb. 12 a), diese erfahren aber bald eine Auflockerung und zerfallen in einzelne unregelm/tl~ige kleine st/~bchen- oder keulen- fDrmige Gebilde und sind infolgedessen meist nicht mehr yon gleich- zeitig dargestellten Mitochondrien zu unterscheiden. In osmierten Pr/~paraten, die bald nach der Differenzierung fixiert worden waren, deutet ein dem Kern benachbarter Bezirk von etwas st~rkerer Osmio- philie bei manchen Zellen noch darauf hin, dal] hier wohl einmal ein GoLGx-Apparat vorhanden gewesen ist. Sp/iter ist auch diese besonders osmiophile Zone nicht mehr zu erkennen (Abb. 12a und b).

656 Ilse Fischer:
Nach Zusatz von basischen Vitalfarbstoffen f~rben sich regelmi~Big die Lipochondrien an (Abb. 13). Die Mitochondrien sind jetzt viel leichter als in undifferenzierten Zellen mit Janusgriin darzustellen, sie speichern auch oft andere basische Vitalfarbstoffe (Abb. 13).
a 1) A b b . 12a l ind b. a U n r e g e l l ~ i ~ i g c s GoL~[ -Ne t z in e i n e r d i f f e r e n z i c r t c n Zel le d e r W a c h s - t u m s z o n e , b Zel le a u s d e r s e l b e n K u l t u r , in d e r s i ch n u r n o c h L i p o e h o n d r i e n u n d F e t t - t r S p f c h e n i m p r f i g n i e r e n liei]en. F i x i e r t retch CHAMP~', 9 T a g e o s m i e r t . V e r g r . e t w a 1200fach .
Die Lipochondrien beginnen nun sich zu vermehren, indem sie Tochtergranula abschniiren. Besondere Abschnfirungsperioden sind aber nicht zu erkennen. Bei Zellteihmgen werden die Lipochondrien ziemlich
A b b . 13. V i t a l f ~ i r b u n g y o n P a n k r e a s z e l l e n a u s d e r W a c h s t u m s z o n e e i n e r K u l t u r n a c h d e r D l f f e r e n z i e r u n g . D u r e h w e g re ine L i p o c h o n d r i e n f f i r b u u g , k e i n e F a r b s t o f f a u s f l o c k u n g in
den Zel len. D ie M i t o c h o n d r i e n s i n d a u c h e t w a s angef , ' i rb t . V e r g r . e t w a l l 0 0 f a c h .
regelms auf die Tochterzellen verteilt, indem sie sich der Spindel dicht anlegen und sparer mit den Tochterplatten auseinanderweichen.
Ein Teil der Lipochondrien schwillt an, wobei vor allem Neutral- fette gespeichert werden. Dabei verlieren sie gleichzeitig ihr Speicherungs- vermSgen fiir Vitalfarbstoffe und werden zu typischen Fettvakuolen. Die Zellen erscheinen nun bald reich mit Fettvakuolen beladen, wie es im allgemeinen fiir in vitro geziichtete Zellen charakteristisch ist (Abb. 13

Uber die Differenzierung der 1%nkreaszelle in vitro. 657
und 15). I m Gegensatz zu CaLoPI~ (1925) kann ich nicht linden, dab die Zellen yon Kaltbliitlern weniger verfetten als die von Warmbltitlern. Es bleibt jedoch stets eine grSBere Zahl yon vitalf/~rbbaren Lipo- chondrien in allen Zellen erhalten.
Die Pankreaszelle des Axolotl verh/~lt sieh nach ihrer Differenzierung in vitro also genau so wie die differenzierte Pankreaszelle des Hfihnchens in der Gewebekultur: Es werden keine Proenzymgranula mehr gebildet, der GoLcI-Apparat sehwindet, dagegen findet eine lebhafte Vermehrung der Lipochondrien statt. Eine Speieherung yon Neutralfetten in den LipochondrienabkSmmlingen fiihrt zu der charakteristischen Verfettung der Kulturen (s. F~SCHE~ und R~]~s 1936).
Die Waehstumserscheinungen an den Explantaten. Die Wachstumsvorg~nge an den Kulturen lassen sich am besten an
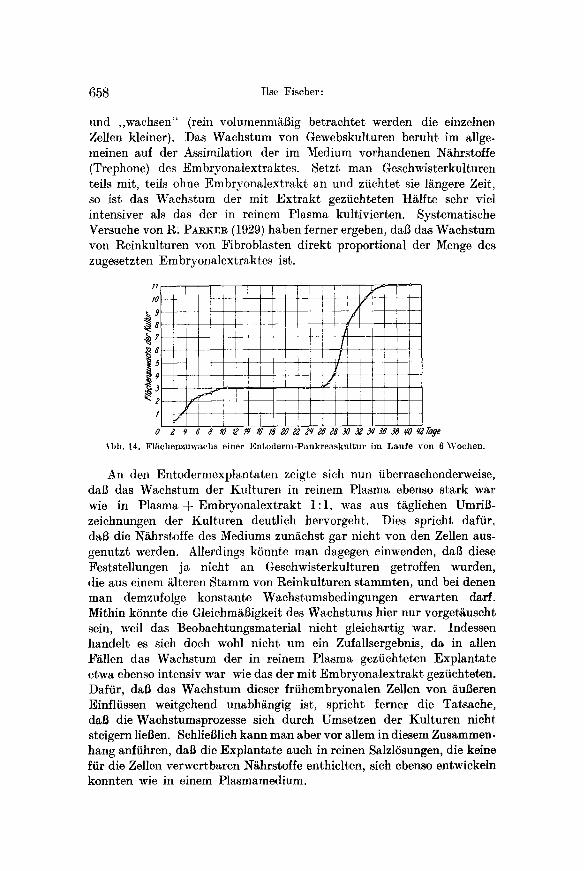
H a n d der Kurve in Abb. 14 erSrtern, die den Fl~chenzuwachs einer Kultur aus Neurulaentoderm im Laufe yon 6 Wochen darstellt. Wir finden zun~chst ein m~Biges, dann sich langsam verringerndes Wachstum und schlieBlich einen Wachstumsstillstand. In dieser Zeit des Wachstums- stillstandes beginnt die organoide und die Zelldifferenzierung (Nb. die Zellen haben in dieser Zeit nicht v511ig ihr Teilungsverm5gen ver- loren. Man findet ausnahmsweise auch in dieser Periode Mitosen; aber die Zellteilungen sind so selten, dab sie nicht zu einer wesentlichen VergrSBerung des Explantates fiihren. AuBerdem ist zu bedenken, daI~ in der Kurve ein Wachstumsstillstand auch schon zu einer Zeit, in der sich die Zellen noch teilen, zum Ausdruck kommen kann. Dureh die zunehmende Resorption des Dotters werden in dieser Periode die einzelnen Zellen n/imlich allm/~hlich kleiner.) Dafiir, dab zu Beginn der Differen- zierungsperiode aber tats/iehlich irgendwelehe wachstumshemmenden Faktoren eine Rolle spielen, spricht vor allem aber auch die Tatsaehe, dab es in dieser Zeit nicht m6glich war, (lie Kulturen zu teilen und Sub- kulturen zu erlangen, die wieder zu der alten Gr6Be heranwuchsen. Diese Erscheinung steht in Einklang mit mehreren Befunden yon SCHMAL- HAUSEN, der bei zahlreichen vielzelligen Organismen und an versehiedenen Geweben w/ihrend der Differenzierungsprozesse ein Herabsinken der Wachstumsgeschwindigkeit feststellen konnte. SCHMALHAUSEN hat daraus geschlossen, dab mit der progressiven Differenzierung eines Organismus die Wachstumsprozesse besehr~nkt werden. Der weitere Verlauf der Kurve in Abb. 14 scheint diesen Beziehungen zu widerspreehen. In- dessen ist anzunehmen, daf5 fiir die Steigerung der Wachstumsintensit~t der Pankreasexplantate ganz bestimmte Ursachen vorliegen, die in einer Umstellung der physiologischen Vorg/inge in den differenzierten Zellen begrtindet sind. Eine ganze Reihe yon Anhaltspunkten sprechen dafiir, dab die undifferenzierten Entodermzellen sich allein auf Kosten des in den Dotterschollen gespeieherten N/ihrmateriales differenzieren
658 Ilse Fischer:
und ,,wachsen" (rein volumenm~i~ig betrachtet werden die einzelnen Zellen kleiner). Das Wachstum yon Gewebskulturen beruht im allge- meinen auf der Assimilation der im Medium vorhandenen N/~hrstoffe (Trephone) des Embryonalextraktes. Setzt man Geschwisterkulturen tells mit, teils ohne Embryonalextrakt an und zfichtet sie 1/ingere Zeit, so ist das Wachstum der nfit Ext rakt geziichteten H~lfte sehr viel intensiver als das der in reinem Plasma kultivierten. Systematische Versuche von R: PARKER (1929) haben ferner ergeben, dab das Wachstum von Reinkulturen yon Fibroblasten direkt proportional der Menge des zugesetzten Embryonalextraktes ist.
0 2 q 8 8 ~ 12 1~ ,~ 18 2 0 2 2 2 q 2 6 Z 8 3 0 323z /3638 qog2Tage
Abb. 14. F l ~ c h e n z u w a c h s e ine r E n t o d e r m - P a n k r e a s k u l t u r i m L a u f e y o n 6 W o c h e n .
An den Entodermexplantaten zeigte sich nun fiberraschenderweise, dal3 das Wachstum der Kulturen in reinem Plasma ebenso stark war wie in Plasma + Embryonalextrakt 1:1, was aus t/iglichen UmriB- zeichnungen der Kulturen deutlich hervorgeht. Dies spricht dafiir, dab die N/~hrstoffe des Mediums zun/Lchst gar nicht yon den Zellen aus- genutzt werden. Allerdings kSnnte man dagegen einwenden, dal3 diese Feststellungen ja nicht an Geschwisterkulturen getroffen wurden, die aus einem ~lteren Stamm yon Reinkulturen stammten, und bei denen man demzufolge konstante Wachstumsbedingungen erwarten daft. Mithin kSnnte die Gleichm/~13igkeit des Wachstums hier nur vorget/~uscht sein, weft das Beobachtungsmaterial nicht gleichartig war. Indessen handelt es sich doch wohl nicht um ein Zufallsergebnis, da in allen F/illen das Wachstum der in reinem Plasma geziichteten Explantate etwa ebenso intensiv war wie das der mit Embryonalextrakt geziichteten. Daffir, dab das Wachstum dieser frfihembryonalen Zellen yon /iul3eren Einfliissen weitgehend unabh/~ngig ist, spricht ferner die Tatsache, dab die Wachstumsprozesse sich durch Umsetzen der Kulturen night steigern lieflen. Schliel31ich kann man abet vor allem in diesem Zusammen- hang anftihren, dab die Explantate aueh in reinen SalzlSsungen, die keine ffir die Zellen verwertbaren N/~hrstoffe enthielten, sich ebenso entwickeln konnten wie in einem Plasmamedium.

l~ber die Differenzierung der Pankreaszelle in vitro. 659
Wenn nun w/~hrend oder gegen Ende der Differenzierungsperiode eine sprunghafte Steigerung des Waehstums der Kulturen zu beob- achten ist, so h/~ngt dies wahrseheinlich damit zusammen, dai~ die Zellen jetzt die Ni~hrstoffe des Mediums aufnehmen, damit auch die spezifi- schen wachstumsf6rdernden Stoffe des Embryonalextraktes und sich infolgedessen hi~ufiger teilen. Es ist auch bezeiehnend, dait erst zu dieser Zeit die fiir Gewebekulturen eharakteristische Verfettung der Zellen (Abb. 13 und 15) beginnt und die Dotterschollen zu diesel" Zeit nahezu v611ig aufgebraucht sind.
Von nun an ist die Wachstums- energie der Kulturen so gesteigert, da$ es m6glich wird, die Kulturen zu teilen und die Subkulturen (Ab- bildung 14) in mehreren Passagen zu ziiehten. Bei den vorliegenden Versuchen wurden Kulturen, die sich in vitro di.fferenziert hatten, fiber 3--4 Passagen weitergeziichtet (24--26 Tage nach der Differen- zierung). Die Versuche mui~ten dann aus /tuBeren Grfinden abge- brochen werden. Man kann so ver- stehen, warum bei der embryonalen, undifferenzierten Pankreaszelle eine Gewebekultur im strengen Sinne Abb. 15. Ep i thc lku l tu r aus e inem Ento-
nicht m6glich ist. Die differenzierte de rmexp lan t a t , das sich in v i t ro differen- z ier t ha t t e . Die Zellen beginnen zu ver-
Entodermzelle spricht n i c h t a u f d i e f e t t en (2. Passage). Vergr . e twa 25fach.
wachstumsf6rdernden Stoffe des Em- bryonalextraktes an. Daraus ergibt sich weiterhin, dal~ fiir die M6glichkeit der Kultur eines Gewebes in vitro nicht allein das Teilungsverm6gen der Zellen aussehlaggebend ist, sondern dabei auch der besondere physiologi- sche Zustand, die Bereitschaft der Zellen, Stoffe aus der Umgebung aufzu- nehmen und zu verarbeiten, eine wesentliche Rolle spielt. In diesem Zu- sammenhange sei daran erinnert, daf~ das Pankreas yon Kiieken (bis zu einem Alter yon etwa 14 Tagen) in vitro nach Piloearpininjektionen, wenn die Zellen auf dem Stadium der friihen Sekretrestitution explantiert wurden, noch auswuchs, w~hrend sonst Pankreasexplantate etwa vom 3. Tage nach dem Schliipfen an nicht mehr in der Kultur auswachsen (FISCHER und l~I]~S 1936). Wi~hrend der Sekretrestitution aber weisen die Pankreaszellen ein erb6htes Aufnahme- und Speicherungsverm6gen ffir Stoffe aus der Umgebung auf, was sich besonders in ihrer starken Speicherung yon basischen Vitalfarbstoffen auspr/igt, die auf diesem Stadium des Arbeitsrhythmus haupts/ichlich durch den GoLGI-Apparat erfolgt.

660 llse Fischer:
l~brigens pr~gen sich ebenso die Unterschiede in der Bereitschaft, Stoffe aus der Umgebung aufzunehmen, zwischen undifferenzierten und differenzierten Pankreaszellen beim Axolotl in ihrem sehr verschiedenen SpeicherungsvermSgen ftir basische Vitalfarbstoffe aus, was deutlich aus einem Vergleieh der Abb. 5 und 6 und 7, 8 und 13 hervorgeht.
Allgemeine Folgerungen aus den Befunden. Mit groBer Wahrscheinlichkeit kann angenommen werden, da6 die
Pankreaszelle des Axolotls zum mindesten schon in der Gastrula deter- miniert und zur Selbstdifferenzierung befs ist, d. h. sich unabh/ingig yon /~ui3eren Einflfissen in bestimmt gerichteter Weise zu ihrer end- giiltigen Form entwickelt, und dab sie die Ursachen zu dieser Weiter- entwicklung in sich selbst tr/~gt. Wenn ich hier betone, dal3 es sich dabei um eine Wahrscheinlichkeitsannahme handelt, so mSchte ieh damit der Tatsache Rechnung tragen, dal3 nicht mit vSlliger Sicherheit auszuschlie/3en ist, dab mitexplantiertes Zellmaterial aus im Keim benachbarten Anlagenkomplexen vielleicht eine induzierende Wirkung ausgefibt hat. (Diese Fehlerquelle ist auch durch Aufzucht der Larven in der HoLTF~T~ttschen SalzlSsung nicht auszuschalten.) Indessen ist die Selbstdifferenzierungsf/thigkeit der Pankreaszelle wohl ziemlich sicher erwiesen durch die eingangs schon erw~hnten Transplantations- versuche HOLTF~ET~Rs an Anuren, bei denen sich die Anlagen yon Leber und Pankreas stets herkunftsgem/iB weiterentwickelten. Auch die Pankreaszelle des Hfihnchens kann sich unabh/~ngig differenzieren: DANTSCHAKOFF (1924) transplantierte Pankreasanlagen in die Allantois; die transplantierten Anlagen entwickelten sich ausnahmslos in v611ig normaler Weise. Allerdings diirfte diese Methode wohl weniger noch als die hier angewandte fiir Potenzprfifungen geeignet sein, da nach HOLT- FRET:E~ (1934) in der Chorio-Allantois mit der Anwesenheit induzierender Stoffe in besonders groi3er Menge zu rechnen ist.
Auf jeden Fall abet erscheint eine weitgehende Autonomie der Pan- kreaszelle in ihren friihesten Entwicklungsphasen gesichert. Dafiir sprechen folgende Tatsachen: Die Zellen differenzieren und entwickeln sich ebenso in anorganischen SalzlSsungen wie in einem Plasmamedium. Sie leben also zun~Lchst offenbar allein auf Kosten der bei der Re- sorption der Dotterschollen frei werdenden Energiequellen. Dal3 die undifferenzierten entodermalen Zellen die N/~hrstoffe des Mediums und die wachstumsfSrdernden Substanzen des Embryonalextraktes fiberhaupt nicht verwerten, geht daraus hervor, dal3 gleichalte und gleieh grol3e Entodermexplantate in reinem Plasma dieselbe Waehstums- intensit/it aufwiesen wie in Plasma und Embryona]extrakt. Wie gering das VermSgen dieser Zellen ist, Stoffe aus der Umgebung aufzunehmen und zu speichern, zeigte auch der Ausfall der Vitalfitrbungen; nut die PigmentkSrnchen nehmen geringe Mengen des Farbstoffes auf. (Die

~ber die Differenzierung der PankreaszelIe in vitro. 661
vitalen Kernf~rbungen dfirften nicht unter ganz normalen Bedingungen zustande kommen und kSnnen daher in diesem Zusammenhange auBer Betracht bleiben.)
Weiterhin vollzieht sich auch die Bildung der ersten Proenzym- granula auf Grund yon Material, das yon Anfang an in den Zellen vor- handen war. W~hrend in der Pankreaszelle des differenzierten und funktionierenden Organes die Lipochondrien Tochtergranula abschniiren, die dann unter Verlust ihres Lipoidgehaltes und unter Anreicherung yon EiweiBsubstanzen (s. RI]~s 1935) zu Proenzymgranula heranreifen, entstehen in den sich differenzierenden Zellen die ersten Proenzym- granula durch direkte Umwandlung yon FettrSpfchen, noch ehe die Lipo- chondrien der Zellen ein Speicherungs- und Teilungsverm6gen erkennen lassen. Erst nachdem bereits eine gr61~ere Menge yon Proenzymgranula in der Zelle vorhanden ist, und die Zellen sich polarisiert und zu Acini zusammengeordnet haben, gewinnen einzelne Lipochondrien ihr Speiche- rungsvermSgen fiir basische Farbstoffe zurfick und sind nun erst als solche zu identifizieren. Teilungserscheinungen sind an diesen Lipochon- drien zun~chst aber noch nicht in einem merkbaren Mal~e zu beob- achten. Es bedarf dazu scheinbar eines Anstol~es yon auBen (in vivo Sekretextrusion; in vitro kommt es unter den Bedingungen raschen Wachstums ebenfalls zu einer allerdings vielleicht geringeren Lipochon- drienvermehrung als in der funktionstfichtigen Drfisenzelle in vivo).
Es erscheint nun bedeutungsvoll, daf~ die Pankreaszelle bereits Proenzymgranula zu bilden vermag, noch ehe die Organellen yon denen in der ,,fertig differenzierten" Zelle die Sekretbildung ausgeht, die Lipochondrien, in den funktionsf~higen Zustand eingetreten sind. Es folgt daraus, dal~ nicht die Ausbildung der typischen morphologischen Strukturen die Differenzierung der Zelle ausmacht, sondern dab wesent- lich vor allem die Bef~higung der Zelle zu dem Funktionsvorgang ist. Im vorliegenden Falle geht die dynamische Differenzierung der Zelle ihrer endgiiltigen morphologischen Differenzierung voraus. Die Bildung der Proenzymgranula kann zu Beginn der Funktionsperiode auf ver- schiedenen Wegen erfolgen. Denn wenn man auch annimmt, dal~ die Fettr6pfchen, aus denen die ersten Proenzymgranul~ hervorgehen, ihrer- seits einstmals aus Lipochondrien entstanden (s. RIEs 1935b), so sind sie doch woh] verschieden - - zum mindesten was ihre quantitative stoff- liche Zusammensetzung anbelangt - - yon den eigentlichen Lipochondrien und ihren Abk6mmlingen; ihr verschiedenes Verhalten bei Vitalf~r- bungen deutet darauf hin. Die Bildung der ersten Proenzymgranula in der Pankreaszelle erscheint damit ,,doppelt gesichert". MSglicherweise hat das ,,Prinzip der doppelten Sicherung" fiberhaupt einen nicht nur auf entwicklungsmechanische Vorg~nge beschr~nkten Geltungsbereich, sondern gelangt auch bei zellphysiologischen Prozessen zur Anwendung. So ist z. B. auch in der Irisepithelzelle die Pigmentbildung doppelt

662 ]lse Fischer:
gesichert, indem sie sowohl yon den Lipoehondrien als auch yon den Mitochondrien ausgehen kann (I. FISCHER 1937). Es w~re wichtig, auch bei anderen Zelltypen darauf zu achten, wieweit in /ihnlicher Weise einzelne Arbeitsleistungen sich fiberschneiden bzw. harmonisch in- einandergreifen. Vielleicht k6nnten damit manche zur Zeit unverst/ind- lichen Widersprfiche fiber die Bedeutung der verschiedenen Zellstrukturen im Zellhaushalt ihre Erkl/irung finden. Allerdings w/~re es vielleicht besser, wenn man auf solche Erscheinungen nicht den aus der Technik fibernommenen finalen Begriff der ,,doppelten Sicherung" anwenden wfirde, sondern in Anlehnung an SrEEMA.WS (1931) synergetisches 1)rinzip der Entwicklung yon einem ,,synergetischen Prinzip der Zell- arbeit reden wfirdc".
Die Bedeutung der Zellorganelle als relativ selbst~ndiger Faktoren im Arbeitsrhythmus und Lebenszyklus der Zellen wird durch diese Erw/~gungen keineswegs eingeschri~nkt, sondern nur die M6glichkeit einer kombinierten Wirkung verschiedener Strukturen hervorgehoben.
W~hrend und vor allem gegen Ende der Differenzierungsperiode /indert sich indessen das Verhalten der Pankreaszelle zu ihrer Um- gebung. ])as Auftrcten einer organoiden Anordnung der Zellen zu dieser Zeit deutet ja schon auf Wechselbeziehungen der Zellen zu ihrer Umgebung hin. Sie beginnen nun auch immer mehr Stoffe aus dem Medium aufzunehmen, was vor allem durch die Speicherung yon Vitalfarbstoffen ersieht!ich wird. Besonders die Lipochondrien gewinnen nun auch wieder ein gro[tes Speicherungsverm6gen gegenfiber basischen Vitalfarbstoffen und wohl auch fiir die N~hrstoffe des Mediums, was aus der Umwandlung yon Lipochondrienabk6mmlingen in Fettvakuolen hervorgeht. Zu dieser Zeit zeigt sich auch eine sprunghafte Steigerung der Wachstumsintensit~t der Kulturen, ein Zeichen, dab sie jetzt auf die wachstumsf6rdernden Stoffe des Mediums ansprechen. In der Wachs- tumszone weisen die als undifferenzierte Entodermzellen explantierten Pankreaszellen nun immer mehr jene Merkmale der erst nach ihrer Differenzierung explantierten Pankreaszellen auf. Aber weder in morpho- logischer noch auch in physiologischer Hinsicht (Speicherungsverm6gen, Wachstumswert und -intensit~t) zeigen sie dabei jetzt die typischen Eigenschaften undifferenzierter entodermaler Zellen. Mithin erscheint in keiner Weise gerechtfertigt, die Ver~tnderungen, welche die Pankreas- zelle nach ihrer Explantation zeigt, als Entdifferenzierungsprozesse auf- zufassen, auch nicht in einem rein morphologischen Sinne.
Viele Autoren haben darauf hingewiesen, dab Differenzierung und Funktion das Wachstum der Gewebe, soweit es auf Zellteilung beruht, hemmen, so z. B. u. a. BE~INGHOrF, JACO~J, PETER. An Gewebekulturen wurde ein solcher Antagonismus vor allem yon A. FISCHER (1929), A. FiscHER und PARKER (1929) ffir Fibroblastenzellen aus Knorpel- gewebe und yon DOLJANSXI (1930) an Irisepithelkulturen nachgewiesen.

~ber die Differenzierung der Pankreaszelle in vitro. 663
Es hat sich nun herausgestellt, dab Irisepithelkulturen auch bei raschem Wachstum in vitro stets weiter Propigmentgranula produzieren, nur die spezifische Funktion der Melaninbildung erscheint bei gesteigerter Wachstumsintensit~t eingeschr~nkt (vgl. I. FISCHER 1937). Um die Be- ziehungen zwischen Wachstum, Differenzierung und Funktion weiter zu kli~ren, w/~re es daher wichtig, einmal quanti tat iv den Betriebsstoff- wechsel eines mit verschiedener Wachstumsgeschwindigkeit in vitro gezfichteten Gewebes zu untersuchen. Aus morphologischen Befunden ergeben sich vorl/~ufig keine Anhultspunkte, die dafiir sprechen, dal] Zellen rasch wachsender Kulturen ein besonders geringes MaB an Zell- arbeit leisten und sich funktionell mehr oder weniger in einem l~uhe- zustand befinden. Wenn die Pankreaszelle nach ihrer Differenzierung in vitro keine Proenzymgranula mehr bildet, so deutet doch die dauernde Vermehrung der Lipochondrien auf ihre funktionelle Aktivit~t im Gegensatz zur undifferenzierten Entodermzelle hin. Wir wissen noch nicht, warum die Proenzymgranulabildung in vitro unterbleibt; es kann sein, dab den Zellen dazu irgendwelche spezifischen Stoffe feh]en, die im Kulturmedium nicht vorhanden sind; es kann aber auch sein, dab die differenzierte Pankreaszelle infolge ihrer Bereitschaft, Stoffe aus der Umgebung aufzunehmen, mit Ni~hrstoffen iiberschwemmt wird und die speicherungsfi~higen Lipochondrien in /ihnlicher Weise wie in der heran- wachsenden Eizelle, in der ja auch ein ~berschuB yon N~hrstoffen an- gereichert wird, nunmehr zu typischen Fettvakuolen anschwellen (vgl. RIES und VAN WEEL 1934). Jedenfalls erf~hrt die differenzierte Pankreas- zelle in vitro - - im Gegensatz zu ihrem Verhalten in ihrer friihembryonalen Entwicklungsphase - - eine weitgehende Funktionsumstimmung, die wohl haupts~chlich durch die ver~nderten Verhaltnisse des Milieus bedingt sein diirfte.
Durch die Bedingungen des Milieus (Embryonalextrakt !) ist vermutlich auch die sprunghafte Steigerung der Wachstumsgeschwindigkeit in vitro nach effolgter Differenzierung veranlaBt. Nach der Differenzierung lassen die Kulturen dann ein relativ gleichm~Biges und intensives Wachstum er- kennen, so dab man sie teilen und in mehreren Passagen weiterziichten kann.
Indessen ist bei der Pankreaszelle nicht so wie bei der Fibroblasten- zelle eine unbegrenzte Dauerkultur m6glich. Die Arbeiten yon K ~ H L (1929), CHLOPIN (1930), AMOROSO (1931), PINKUS (1931), I. FISCHER und RIES (1936) ergeben iibereinstimmend, dab Sts von Pankreas- kulturen nie ls als etwa 6 Wochen geziichtet werden konnten. Pan- kreaskulturen wachsen um so schlechter, je alter sie sind. Schon nach 4 Wochen bleiben sie meistens so klein, dab man sie nicht mehr teilen kann, und diese Kulturen gehen schlieBlich unter zunehmender Ver- fettung zugrunde.
Das Verhalten der Pankreaszelle auf verschiedenen Entwieklungs- phasen in der Gewebekultur zeigt, daB hier ein streng determinierter
Z. f. Zel l fo l ,~chung n. m i k r . A n a t o m i e . B d . 27. 4 4

664 llse Fischer:
Zelltyp mit einem ganz bestimmten Lebenszyklus vorliegt. Dieser Lebenszyklus l~13t verscbiedene, deutlich voneinander abgcgrenzte Perioden erkennen : 1. die der determinierten, aber noch undifferenzierten teilungsfiihigen Entodermzelle (yon der Gastrulation bis zur Abgliederung der Leberbucht), 2. eine Latenzperiode bis zum Beginn der Differenzierung (sehr beschr~nktes WachstumsvermSgen der Kulturen), 3. die Differen- zierungsperiode, 4. die Funktionsperiode, 5. die Degenerationsperiode (.~) (vgl. hierzu auch das Schema yon RIES (1937 a) fiber die Entwicklungs- perioden und Ziichtbarkeit der Pankreaszelle). Die einzelnen Phasen des Lebenszyklus der Pankreaszelle pr~gen sich also auch in vitro aus, nur eben in wesentlich anderer Weise als in vivo in den Perioden, in denen die Pan- kreaszelle besonders auf Umweltseinfliisse anspricht. Es ergibt sich daraus weiterhin, dal~ Determination und Differenzierung bei der Pankreaszelle weitgehend irreversible Prozesse sind, die sich zum mindcsten durch Isolierung und Explantation nicht aufheben lassen.
Zusammenfassung. 1. Die Pankreaszelle ist bereits in der Gastrula determiniert, w~s
nicht nur durch die vorliegenden Explantationsversuche, sondern vor allem auch durch die Untersuchungen HOLTFR~T~RS gesichert erscheint.
2. Sic vermag sich in der Gewebekultur ebenso wie in physiologischen SalzlSsungen zu differenzieren.
3. Die dynamische Differenzierung der Zelle geht ihrer endgiiltigen morphologischen Differenzierung voraus.
4. Die Pankreaszellen polarisieren sich und bilden typische Pro- enzymgranula.
5. Gleichzeitig bildet sich im Muttersttick der Explantate eine organoide Gewebestruktur, w~hrend es in der ,,Wachstumszone" in der Regel nur zu einer rein cytologischen Differenzierung kommt.
6. Die Wachstumsintensit~t ~ndert sich periodisch im Lebens- zyklus der Pankreaszelle:
7. Die undifferenzierten entodermalen Zellen weisen ein m~l]iges und im Laufe der Zeit ganz allm~hlich abklingendes TeilungsvermSgen auf. Vor Beginn der Zelldifferenzierung zeigt sich hinsichtlich des Wachstums eine Latenzperiode. Die differenzierte Zelle l~,l]t wieder eine erhShte Wachstumsenergie erkennen. Dieses Wachstum ist bedingt durch die wachstumsf6rdernden Stoffe des Embryonalextraktes im Medium, die bei den differenzierten Zellen Mitosen hervorrufen, w~hrend die undifferenzierten Entodermzellen auf die waehstumsfSrdernden Stoffe nicht ansprechen. Nach einigen Wochen zeigt sich indessen eine Verminderung der Wachstumsprozesse der differenzierten Zellen, was schliel]lich zur Folge hat, dab die Kulturen nach einiger Zeit eingehen.
8. Die Beziehungen zwischen Waehstum und Differenzierung der Pankre~szelle in der Gewebekultur entsprechen der Wachstumstheorie yon SC~M~HAUSEN; mit ,,progressiver" Differenzierung erscheint das

Uber die Differenzierung der Pankreaszelle in vitro. 665
Wachs tum beschr~nkt, die differenzierte Zelle aber ist, unter Bedingungeu, die dasWachs tum begiinstigen, wieder zu einer intensiven Vermehrung f~hig.
9. Die entodermalen, undifferenzierten PankreaszeIlen lassen an- fangs eine weitgehende Autonomie ihrer Lebenserscheinungen erkennen: Sie entwickeln sich allein auf Kosten des in den Dotterschollen ge- speicherten N~hrmateriales. Die N~hrstoffe des Mediums werden gar nicht ausgeniitzt. Die Wachstumsintensi t~t der Zellen wird durch Em- bryonalextrakt nicht gesteigert. Die geringe Aufnahmeberei tschaft dieser Zellen fiir Stoffe aus ihrer Umgebung zeigt sich in fiberzeugender Weise auch nach vitaler Anf~rbung; im Gegensatz zur differenzierten Zelle vermSgen sie basische Vitalfarbstoffe nicht zu speichern.
10. Auch die Bildung der ersten Proenzymgranula geschieht auf Kosten yon Material, das bereits in den embryonalen Zelien vorhanden war. Noch bevor die Lipochondrien funktionsf~Lhig erscheinen, werden Proenzymgranula aus Fet trSpfchen gebildet. Die Bildung yon Pro- enzymgranula vor Beginn des Arbei ts rhythmus der Zellen erscheint damit ,,doppelt gesichert" (,,synergetisches Prinzip der Zellarbeit").
11. Der Lebenszyklus der Pankreaszelle erweist sich als streng determiniert und pr~gt sich auch im Verhalten der Zellen in vitro auf verschiedenen Entwicklungsstadien aus. Nu t wenn die Zelle nach der Differenzierung in besonderem MaBe bef~higt ist, Stoffe aus der Um- gebung aufzunehmen und zu verarbeiten, k o m m t es zu einer dutch das Milieu bedingten Funkt ionsumst immung. Im Zusammenhange damit ~ndern sich die morphologischen und physiologischen Merkmale der Zellen.
12. Determinat ion und Differenzierung erscheinen bei der Pankreas- zelle weitgehend irreversibel, sic k6nnen zum mindesten dutch Isolat ion und Explantat ion, nach HOLTFRETER aueh durch Transplantat ion, nicht aufgehoben werden.
L i t e r a t u r v e r z e i c h n i s .
Amoroso, E.: Die Ztichtung yon Epithel des embryonalen Hiihnerpankreas. Arch. exper. Zellforsch. 12 (1931). - - Champy, Ch.: Perte de S~cr~tion sp~cifique des cellules eultiv~s in vitro. C. r. Soc. Biol. Paris 83 (1920). - - Chlopin, G.: l~ber die Verwandlungen des Epithels der Bauchspeicheldriise in Gewebekulturen. Arch. exper. Zellforsch. 9 (1930). - - DantsehakoII, W.: Wachstum transplantierter embryonaler Gewebe in der Allantois. Z. Anat. 74 (1924). - - Doljanski, L.: Sur le rapport entre la prolif4ration et l'activit4 pigmentog~ne dans les cultures de l'iris. C. r. Soe. Biol. Paris 105 (1930). - - Fell, H. B.: The development in vitro of the isolated otoeyst of the embryonic fowl. Arch. exper. Zellforsch. 7 (1928). -- Fell, H. B. and R o b i n s o n : The development and phosphatase activity in vivo and in vitro of the mandibular skeletal tissue of the embryonic fowl. Biochemic. J. 24 (1930). - - F i scher , A. : Gewebeztichtung. Miinehen 1930. - - F i scher , A . u. R. C. P a r k e r : Dauerzfiehtung in vitro ohne Wachstumsbesehleunigung. Arch. exper. Zell- forsch. 8 (1929). - - Proliferation und Differenzierung. Arch. exper. Zellforsch. 8 (1929). - - F i scher , I.: Die Pigmentbfldung des Irisepithels in vitro, ein Beitrag zu den Beziehungen zwischen Differenzierung, Wachstum und Funktion. Arch. exper. Zell- forsch. 21 (1937) . - - F i scher , I. u. E. Ries : Das Verhalten der Pankreaszelle des Hiihnchens in der Gewebekultur. Arch. exper. Zellforsch. 18 (1936). - - Ho l t f re ter , J.: Defekt- und Transplantationsversuehe an der Anlage yon Leber und Pankreas
44*

666 llse Fischer.
j tmgster Amphibienkeimc. Roux' Arch. 105 (1925). (~'ber die Aufzucht isolierter Teile des Amphibienkeimes. I. Methode einer Gewebeztichtung in vivo. Roux' Arch. 117 (1929). - - Uber histologische Differenzierungen yon isoliertem Material jiingster Amphibienkeime. Verh. dtsch, zool. Ges. 1929. - - L'ber die Aufzueht isolierter Teile des Amphibienkeimes. II . Ziichtung yon Keimen uud Keimteilen in Salz- ]6sung. Roux' Arch. ]24 (1931). Formative Reize in der Embryonalentwicklung der Amphibicn, dargestellt an Explantationsversuchen. Arch. exper. Zellforsch. 15 (1934). - - Eigenschaften und Verbreitung induzierender St<)ffe. Naturwiss. 1933. Kapel~ 0. : Einige Untersuchungen fiber das Verhalten des Epithels in vitro. Arch. exper. Zellforsch. 8 (1929). - - 01ivo: tJber die frfihzeitige Determinierung der Herzanlage beimHfihnerembryo. Verh. dtsch, anat. Ges. 66 (1928). - - Parker, R.: The cultivation of tissues for prolonged periods in single flasks. J. of exper. Med. 75 (1936). - - Peter, K.: Zellteilung und Resorption, ]., 2. u. 3. Teil. Z. Anat. 72 (1924). --- Zur Histophysiologie der Amphibienuiere. Z. Anat. 73 (1924). - - Zell- teilung und Sekretion. Z. Anat. 75 (1925). - - Der EinfluB der Zel l~i lung auf die Zellt~tigkeit. Z. Zellforsch. 9 (1929). - - Pinkus, ]{.: Untersuchungen fiber das Verhalten des Hfihnerpankreas in der Gewebekultur. Arch. exper. Zellforsch. 10 (1931). - - Ries~ E.: Zur Histophysiologie des M~usepankreas nach Lebendbeobach- tung, Vitalf/irbung und Stufenuntersuchung. Z. Zetlforsch. 22 (1935a). --- Die Differenzierung der Pankreaszelle w~thrend der Embryonalentwicklung des Axolotl. Z. Zellforsch. 23 (1935 b). - - Untersuchungen fiber Differenzierung, Arbei tsrhythmus und Ztichtbarkeit der Pankreaszelle. Arch. exper. Zellforsch. 19 (1937a). - - (;rber Arbei tsrhythmen und Lebenszyklen yon Zellen. Verh. dtsch, zool. Ges. 1937 b. - - Untersuchungen fiber den Zelltod. I. Die Ver~nderung der F~trbbarkeit beim Absterben der Zellen, mit besonderer Berficksichtigung der vi ta len und post- vi talen Kernfiirbung. Z. Zellforsch. 26 (1937 c). - - Sehmalhausen, J. : Zur Wachstums- theorie. Roux' Arch. 116 (1929). --- Beitr~ge zur quant i ta t iven Analyse der Form- bildung. I. Uber die Gesetzm~fligkeiten des embryonalen Wachstums. Roux' Arch. 109 (1927). - - Das Wachstumsgesetz als Gesetz der progressiven Differenzierung. Roux' Arch. 123 (1930). - - Strangeways~ T.: Tissue culture in relation to growth an differentiation. Cambridge 1924. - - Strangeways~ T. and tI. ]L Fell: Experi- mental studies on the differentiation of embryonic tissue growing in vivo and in vitro. Proc. roy. Soc. Lond. B 99 (1926).
Nachtrag zum Literaturverzeiehnis. Alexandrov, W.: l~ber die Bedeutung der oxydoreduktiven Bedingungen fiir
die vitale Fi~rbung, mit besonderer Berficksichtigung der Kernf~rbung in lebenden Zellen. Protoplasma (Berl.) 17 (1932). - - Benninghoff, A.: Zur Kenntnis und Be- deutung der Amitose und amitose/~hnlicher Vorg~nge. Sitzgsber. Ges. Naturwiss. Marburg 1922. - - Chi0pin, S. u. A. Chlopin: Studien fiber Gewebskulturen in art- fremdem Blutplasma. Arch. exper. Zellforsch. 1 (1925). - - Gersch, M. u. E. Ries: Vergleichende Vitalf~rbungsstudien. Sonderungsprozesse und Differenzierungs- peri0den bei Eizellen und Entwicklungsstadien in verschiedenen Tiergruppen. Roux' Arch. 136 (I937). - - Jaeobj, W.: l~ber das rhythmische Wachstum der Zellen durch Verdoppelung ihres Volumens. Roux' Arch. 106 (1925). - - Makarov, P.: Analyse der Wirkung des Kohlenoxyds und der Cyanide mit Hilfe der Vitalf~rbung. Protoplasma (Berl.) 20 (1934). - - Nassonov, D.: ~ b e r den EinfluB der Oxydations- prozesse auf die Verteilung yon Vitalfarbstoffen in der Zelle. Z. Zellforsch. l l (1930). - - Parker, R.: Physiologische Eigenschaften mesenchymaler Zellen in vitro. Arch. exper. Zellforsch. 8. - - Ries, E. u. P. B. van Weel: Die Eibildung der Kleider- laus, untersucht an lebenden, vitalgef/irbten und fixierten Prb, paraten. Z. Zellforsch. 20 (1934). - - Spemann, H.: Ober den Anteil yon Implan ta t und Wirtskeim an der Orientierung und Bescbaffenheit der induzierten Embryonalanlage. Roux' Arch. 123, S. 508f. (1931).





























