Embed Size (px)
Citation preview

Arch. exper. Path. u. Pharmakol., Bd. 226, S. 442--448 (1955).
Aus dem Institut fiir Pharmakologie und Toxikologie der Humboldt-Universit~t zu Berlin (Direktor: Professor Dr. F. Ju~G) und dem Institut fiir Medizin und Biologie
der I)eutschen Akademie der Wissenschaften, Berlin-Buch.
~ber Stoffweehselleistungen roter Blutkiirperehen. IV. Substratverarmung und Stoiiwechselaktivit~t.
Yon HANSJ~RGEN MATTHIES.
Mit 4 Textabbfldungen.
(Eingegangen am 16. Juni 1955.)
Bei Unte r suchuvgen fiber die Beziehungen zwischen der Meth~moglo- binrf ickbi ldung und dem Ka t ionen t r anspor t in den Ery th rocy ten 4 ha t t en wir die Fes ts te l luvg gemaeht, dab der Ze i tpunk t der Subst ra tzugabe yon groBer Bedeu tung fiir die Verwertung der Substra te durch die roten Blutzel len ist. Zugabe yon Glucose bewirkt nu r zu Beginn der I n k u b a t i o n der Zellen eine erhebliche Beschleunigung der Reduktionsgesehwindig- keit des Meth~moglobins [Hb(3)] und ermSglicht n u t d a n n die Aufreeht- e rha l tuvg des Ka t ionenbes tandes ; wurde das Subs t ra t erst ein oder zwei S tunden nach Beginn der Inkubat ionsper iode zugesetzt, so war die Hb(3)-Riickbfldung n u t geringffigig beschleunigt u n d der Ka l iumver lus t der Zellen n ieh t mehr aufzuhal ten. I m Hinbl iek auf die Notwendigkei t vergleichender Auswer tuvgen unserer Stoffwechselversuche war es not- wendig, diesen Vorgang einer systematischen Unte r suchuvg zu unter- ziehen, um seine Gesetzm~Bigkeiten aufzukli~ren.
Methodik. Kaninchenblut wurde durch Herzpunktion gewonnen, defibriniert und das
Plasma durch Zentrifugieren yon den Zellen abgetrennt. Das Zentrifugieren dauerte 10 rain (bei 3000 U/rain). AnschlieBend wurden die Erythrocyten mit dem gleiehen Volumen 2,4°/oiger NaNO2-LSsung gut durchmiseht und 5 min stehen gelassen. durch dreimaliges Waschen wurden die Zellen yon den Resten des Oxydations- mittels und den diffusiblen Stoffwechselsubstraten weitestgehend befreit. Die Waschung erfolgte mit einer isotonen SalzlSsung, die aus 4 Teilen 0,155 m NaCl und 1 Teil Phosphatpuffer (pH 7,4) bestand. Dann wurde aus den Zellen mit einer gleichen Salzl6sung eine 30~oige Suspension hergestellt. Die Hb(3)-Riiekbfldung wurde an der CO-Absorption durch das dabei entstehende H~moglobin [Hb(2)] bei Inkubation um 37 ° manometrisch gemossen. - - Zu verschiedenen Zeiten der Inku- bationsperiode erfolgte die Zugabe der Substrate : Glucose 10 -2 m, Glucose 10 -2 m -~ Methylenblau 5 • 10 -5 m, Fructose 10 -2 m, BTatriumlactat 5 • 10 -3 m Formalde- hyd 10 -3 m. Die Ausrechnung der Geschwindigkeit der Hb(3)-Riickbfldung erfolgte an Hand der Kurven. Da die Anfangskonzentration des Hb(3) bei etwa 90 bis 95~/o
l~ber Stoffwechselleistungen roter BlutkSrperchen. IV. 443
des Gesamthamoglobins lag, kam sic als Geschwindigkeits-bestimmender Faktor nicht in Betracht. Zur Kontrolle wurden, wie auch in unseren friiheren Unt~r- suchungen, in jeder Versuchsreihe colorimetrische Bestimmungen der Hb(3)-Riick- bildung durchgefiihrt, die in allen Fallen die manometrischen Messungen best~tigten.
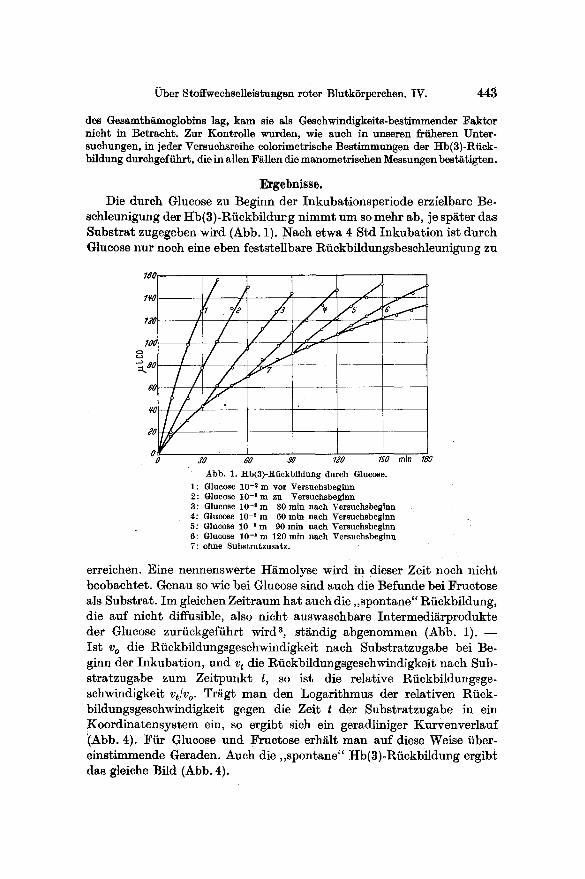
Ergebnisse. Die durch Glucose zu Beginn der Inkubationsperiode erzielbare Be-
schleunigung der Hb(3)-Riickbildurg n immt um so mehr ab, je sparer das Substrat zugegeben wird (Abb. 1). Nach etwa 4 Std Inkubat ion ist durch Glucose nur noch eine eben feststellbare R6ckbildungsbeschleunigung zu
706
~yo G6
0
I//- 30 EO 90 1gO
Abb. 1. Hb(3) -Rl ickbf ldung durch Glucose.
1: Glucose 10 -2 m vor Versuchsbeginn 2: Glucose 1 0 - ' m zu Versuchsbeginn 3: Glucose 10 -2 m 30 ra in nach Versuchsbcginn 4: Glucose 10 -2 m 60 ra in nach Vcrsuchsbeginn 5: Glucose 10 -2 m 90 ra in nach Versuehsbeginn 6: Glucose 10 -2 m 120 rain nach Versuchsbeginn 7: ohne Subs t ra tzusa tz .
150 rain 188
erreichen. Eine nennenswerte H~molyse wird in dieser Zeit noch nicht beobachtet. Genau so wie bei Glucose sind such die Befunde bei Fructose als Substrat. I m gleichen Zeitraum hat such die, ,spontane" Riickbildung, die auf nicht diffusible, also nicht auswasehbare Intermedi~rprodukte der Glucose zuriickgefiihrt wird a, st~ndig abgenommen (Abb. 1). - - Is t v o die Rfickbildungsgeschwindigkeit nach Substratzugabe bei Be- ginn der Inkubation, und v t die Riickbfldungsgeschwindigkeit nach Sub- stratzugabe zum Zeitpunkt t, so ist die relative Rfickbildungsge- schwindigkeit v t /v o. Trfigt man den Logarithmus tier relativen Riick- bildungsgeschwindigkeit gegen d i e Zeit t der Substratzugabe in ein Koord ina tensys tem ein, so ergibt sich ein geradliniger Kurvenver lauf (Abb. 4). Fiir Glucose und Fructose erh~lt man auf diese Weise fiber- einstimmende Geraden. Auch die , ,spontane" Hb(3)-Riickbildung ergib£ das gleiche Bild (Abb. 4).
444 ~L~NsJURGEN MATTHIES:
N u n meh r l iegt es nahe, auch die anderen Wege der Hb-(3)I~iickbildung auf diese Weise zu untersuchen.
Bekanntlich unterscheidet man haupts~chlich drei verschiedene MSglichkeiten der physiologiscben Hb(3)-Reduktion in den kernlosen roten Blutzellen a, s, ~,: Mit
M6
7E6
726 ~
700
£G
dO
/ / / [
~ L / j , !
1
0 20 ~0 GO 80 100 720 7~0 lEO 180 20Omen
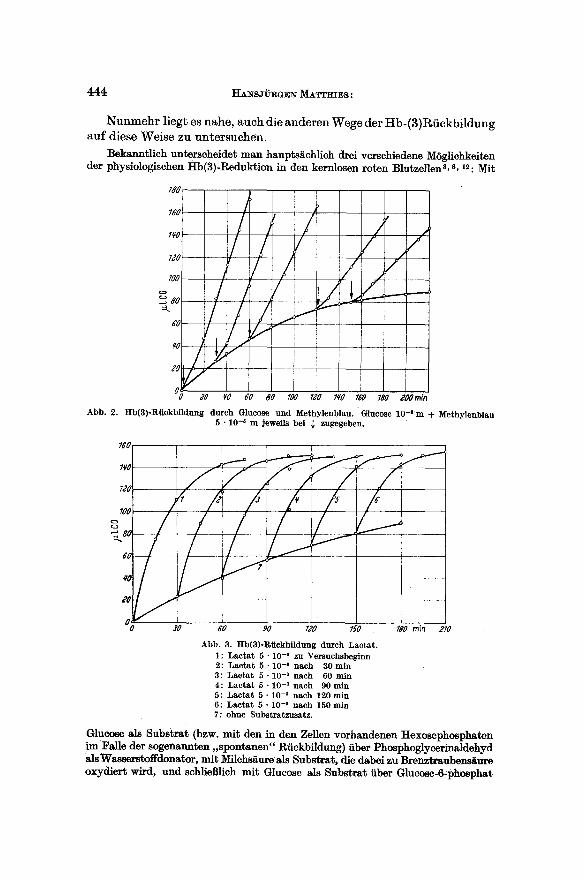
Abb. 2. Hb(3)-RiickbUdung durch Glucose und Methylenblau. Glucose 10-1m + Methylenblau 5 • 10 -5 m ]eweils bei ~ zugegeben.
~ 0
L J 90 720 750 180 rain dTO
Abb. 3. Hb(3)-Riickbildung durch Lactat. 1: Lactat 5 . 1 0 -8 zu Versuchsbeginn 2: Lactat 5 • 10 -a nach 30 min 3: Lactat 5 • 10 -a nach 60 rain 4 : Lactat 5 - 10 -3 nach 90 rain 5: Lactat 5 - 1 0 -s nach 120min 6: Lactat 5 . 1 0 -3 nach 150 rain 7: ohne Substratzusatz.
Glucose als Substrat (bzw. mit den in den ZeUen vorhandenen Hexosephospha~en i m F a U e d e r s o g e n a n n t e n , , s p o n t a n e n " R i i c k b i l d u n g ) f iber P h o s p h o g l y c e r i n a l d e h y d
a l s W a s s e r s C o f f d o n a t o r , m i t M i l c h s a u r e a l s S u b s t r a t , d ie d a b e i z u B r e n z t r a u b e n s ~ u r e o x y d i e r t w i r d , u n d schl ieBl ich m i t G lucose a ls S u b s t r a t f iber G l u c o s e - 6 , p h o s p h a t
~ber Stoffwechselleistungen roter BlutkSrperchen. IV. 445
und Phosphogluconsgure. IBese letzte MSglichkeit ist normalerweise fOr .die Hb(3):Riickbfldung yon untergeordneter Bedeutung, sie kann aber durch re- versible Farbstoffe, wie Thionin oder Methylenblau sehr stark beschleunigt werden und spielt dann eine gowisse Rolle in der Therapie der Methgmoglobingmie.
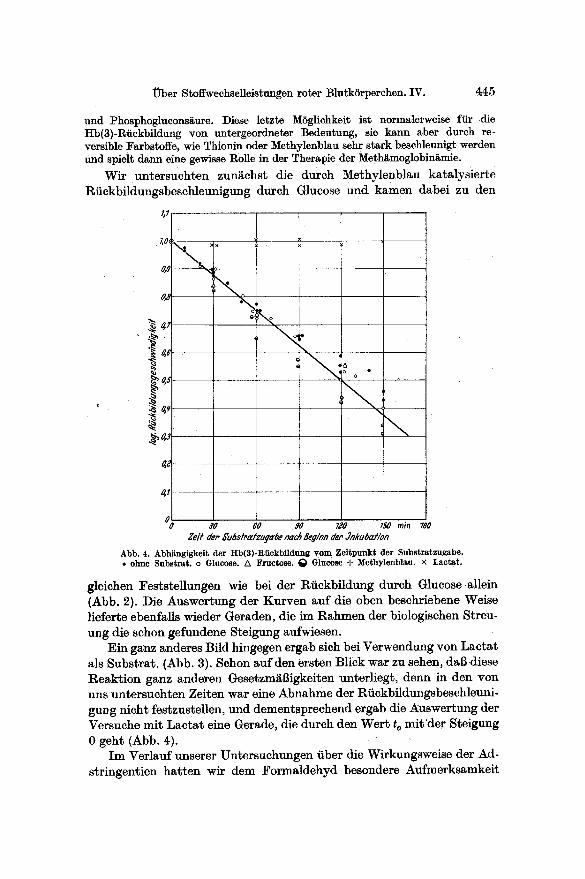
Wir untersuchten zungchst die dutch Methylenblau ka~alysierte Riickbildungsbeschleunigung durch Glucose und kamen dabei zu den
%,= ~ ~
\
I
60 dO 6"0 3o 720 15o rain 180 Zelt der ~'ubstpMzu~be noch 8eg/nn de," Jnku~ztlon
Abb. 4. Abhii~kelt der Hb(8)-REckbildung yore ZeRp~t der Substratzugabe. • ohne Substrat . o Glucose. /x Fructose. ~ Glucose -I- Methylenblau. × Lactat .
gleichen Feststellungen wie bei der Rfiekbildung durch Glucose allein (Abb. 2). Die Auswertung der Kurven auf die oben beschriebene Weise lieferte ebenfalls wieder Geraden, die im Rahmen der biologisehen Streu- ung die sehon gefundene Steigung aufwiesen.
Ein g~nz anderes Bild hingegen ergab sieh bei Verwendung yon Lacta t als Substrat. (Abb. 3). Schon auf den ersten Blick war zu sehen, dab diese Reakt ion ganz anderen Gesetzm~Bigkeiten unterliegt, denn in den yon uns untersuchten Zeiten war eine Abnahme der Rfickbildungsbeschleuni- gung nicht festzustellen, und dementsprechend ergab die Auswertung der Versuche mit Lacta t eine Gerade, die dutch den Wert t o mit 'der Steigung 0 geht (Abb. 4).
I m Verlauf unserer Untersuchungen fiber die Wirkungsweise der Ad- stringentien hat ten wir dem Formaldehyd besondere Aufmerksamkeit

446 HANSJURGENMATTHIES:
zugewandt ~, ~, 5, und es hatte sich ergeben, daI3 diese Substanz eine sehr starke Wirkung auf den Stoffwechsel der roten BlutkSrperchen ausfibt, die neben ihrer Wirkung auf die Zellstrukturen einhergeht 5. In der ttoff- nung, sowohl ffir die Aufkl~rung der hier besprochenen Befunde, als auch ffir die Vertiefung unserer Vorstellungen fiber den Mechanismus der Formaldehydwirkung welteres Material zu gewinnen, setzten wir diesen Stoff zu verschiedenen Zeiten den Erythrocytensuspensionen zu. Auch die Beschleunigun g der Hb(3)-Rfickbildung durch Formaldehyd war in den yon uns untersuchten Zeiten unabh~ngig vom Zeitpunkt der Zugabe des Aldehyds; sein Verhalten entspricht also dem des Lactats. In Abb. 4 sind die Ergebnisse dieser Arbeit in ein Koordinatensystem eingetragen, wodureh die Unterschiede und ~bereinstimmungen deutlich gemacht werdeu.
Diskussion. Ffir die I)eutung dieser Versuche muB man einen fermentativen
ProzeB oder einen Faktor suchen, der den zeitabh~ngigen Reaktionen gemeinsam ist, w~hrend er ffir die Rfickbildung durch Lactat der Form- aldehyd keine Bedeutung haben dfirfte. Die eigentlichen Dehydrierungs- fermente shad auszuschlieBen, da sic im Fall der Rfickbfldung dutch Glucose allein und der durch Methylenblau katalysierten Rfickbildung durch Glucose verschieden sind: Im ersten Fail ist es die Triosephosphat- dehydrogenase, im zweiten Fall die Glucons~urephosphatdehydrogenase.
Aus dem gleichen Grund scheiden auch die Co-Fermente aus der weiteren Betrachtu~g aus: Bei der durch Methylenblau katalysierten Reaktion dient Triphosphopyridinnucleotid, bei der Dehydrierung des Triosephosphats Diphosphopyridinnucleotid als Wasserstoffacceptor. Eine Reaktion ist den zeitabh~ngigen Rfickbfldungsarten gemeinsam, und das ist die Phosphorylierung. Da abet ffir diesen Vorgang bei den ehlzelnen Substraten gleichfalls verschiedene Enzyme wirksam sind - - ffir die Glucose die Glucokinase und Phosphohexokinase, ffir Fructose die Fructokinase und' Phosphohexokinase, ffir die durch Methylenblau katatysierte Reaktion nut die Glucokinase, ffir das nieht auswaschbare Substrat, sower es sich um Glucose-6-phosphat oder ~Yuctose-6-phos- phat handelt, nur die Phosphohexokinase - - so bleibt als einziger ge- meinsamer Faktor der Phosphatdonator, das Adenosintriphosphat (ATP) fibrig. Man kann also annehmen, dab ein zunehmender Mangel an ATP die Ursache der abnehmenden Geschwindigkeit der zeitabh~ngigen Rfiek- bildurgsprozesse ist. Die Ursache ffir die ATP-Abnahme ist wohl darin zu suchen, dab auf der einen Seite ]aufend eine Verwertung der energie- reichen Phos'phatbindurg des ATP ffir die allgemeinen Zelleistu~gen, wie etwa ffir den Kationentransport erfolgt 1, 9, daB aber andererseits infolge der Abnahme des nieht auswaschbaren Substrates diese Verluste durch Re- synthese niche mehr ausgeglichen werden kSnnen. Wenn man noeh einmal

(~ber Stoffwechselleis~angen toter Blutk6rperchen. IV. 447
die einze]nen Sehritte unserer ~berlegungen iiberbliekt, ergibt sich also folgendes Bild: Schwund des ,,nicht auswaschbaren Substrats" , ab- nehmende Resynthese yon ATP bei gleichzeitiger Spaltung, abnehmendes PhosphorylierungsvermiSgen, verminderte Ausnutzung des Substrats im Falle einer sp~teren Zugabe, damit wiederum verbundene UnmSglich- keit einer neuerlichen Resynthese yon ATP. Es ist also anzunehmen, dal~ unterhalb eines gewissen ATP-Gehaltes der Zelle der Zusammenbruch des Stoffwechsels nicht mehr aufzuhalten ist. Die Hb(3)-Riickbildung ist gewissermal~en als Indikator fiir den Substratumsatz aufzufassen.
PAYS~-T und WOL~ 6 machten ~hnliche Beobachtungen bei der Untersuchung der Glucoseutilisation dutch Hb(2)-Zellen in Abhangigkeit yon der Glucosezugabe. Sie stellten lest, dab yon einem kritischen Zeitpunkt an auch dutch Zusatz sehr hoher Substratmengen der Abbau des ATP nicht mehr aufzuhalten ist.
Unsere Auswertung der Versuche erlaubt uns auch, noch einige weitere Angaben zu machen. Offensichtlich liegt den zeitabh~ngigen Reaktionen ein gemeinsamer Vorgang zugrunde; wenn man unseren l~berlegungen fo]gen will, w~re dies die geschwindigkeitsbestimmende Reaktion beim Abbau des nicht auswaschbaren Substrates. Aus Abb. 4 l~flt sich ~ un- mit telbar die Halbwertazeit en~nehmen; sic betr~gt 72 rain. LieBe sich fiir den Abbau des ATP unter den Bedingungen des Substratmangels eine gleiche Halbwertszeit feststellen, so w~re unsere Vorstellung, dab die Abnahme des ATP in der Zelle die Ursache ffir die mangelnde Sub- stratverwertung in unserer Versuchsanordnung darstellt, richtig. In der Ta t haben orientierende Versuche eine solche Beziehung aufgedeckt.
Die Ergebnisse der vorliegenden Untersuchungen haben fiber die ein- gangs erw~hnte Bedeutung fiir die Aufkl~rung einiger Differenzen un- serer Versuchsergebnisse und die kiinftige Berficksichtigung bei der Entwicklung methodischer Einzelheiten hinaus Bedeutung fiir das Pro- blem der Blutkonservierung, bei dem die Erhal tung der Vitalit~t der Zellen einen entseheidenden Faktor darstellt. Wie wir bereits an anderer Stelle berichteten 4, zeig~ sich bei derartigen substratverarmenden, das heis t also mit anderen Worten auch ATP-verarmenden Zellen, ein be- schleunigter Kaliumverlust als Vorl~ufer der H~molyse. In unseren damaligen Versuchen war dieser Vorgavg info]ge der gleichzeitig ablaufen- den Hb(3)-Riickbfldung ebenfalls s tark beschleunigt, bei I~ichtberiick- sichtigung der hier erhobenen Befunde mag er aber auch bei nicht so hohem Hb(3)-Gehalt der Zellen eine Rolle spielen.
Zusammenfassung.
1. Die Beschleunigun g der Hb(3)-Rfickbildung durch Glucose, Fructose oder durch Glucose und Methylenblau ist vom Zeitpunkt der Zugabeder Substrate nach Einbringen der Zellsuspensionen in den Thermostaten abh~ngig.

448 HA~SJi~RGEN MATTHIES : l~ber Stoffweehselleistungen roter BlutkSrperchen. IV.
2. D i e B e s c h l e u n i g u n g de r H b ( 3 ) - R i i c k b i l d u n g d u r e h L a c t a t u n d
F o r m a l d e h y d ze ig t ke ine solche A b h ~ n g i g k e i t w ~ h r e n d de r g le i chen Ze i t . 3. A u s den B e f u n d e n w i r d geschlossen , daft es sieh be i d e m sogenann-
t e n , , n i ch t a u s w a s c h b a r e n S u b s t r a t " u m die H e x o s e p h o s p h a t e hande l t . 4. Als U r s a c h e f i i r die a b n e h m e n d e A u s n u t z u n g de r z u g e s e t z t e n Sub-
s t r a f e w i r d de r A T P - A b b a u im Zuge a l l geme ine r Ze l le i s tungen , insbe- sonde re des K a l i u m t r a n s p o r t s , be i g le ichze i t ige r A b n a h m e der R e s y n -
t he se y o n A T P angesehen .
Literatur. 1GA~DOS, G.: Aeta physiol. (Budapest) 6, 191 (1954). - - 2 Juno, F., u. K.
O~NE~: Arch. exper. Path. u. Pharmakol. 224, 179 (1955). - - 3 KIESE, i~. : Biochem. Z. 816, 264 (1944). - - 4 MATTHIES, H. : Arch. exper. Path. u. Pharmakol. 921, 497 (1954). - - 5 MATTHIES, ~-I. : Symposion fiber Struktur und Funktion der roten Blut- kSrperchen. Berlin 1955. - - e PAYSA~T U. WOLFF: C. r. Soc. Biol. (Paris) 146, 1250, 1253 (1952). - - 7 SCHELER, W . , K. SCHELER U. F. JUNO -" Bioehem. Z. 826, 288 (1955). __8 SPICER, S. S., C. H . HANNA U. A. M. CLARK: J . of Biol. Chem. 177,217 (1949). - - 9 STRAUB, F. B. : Acta physiol. (Budapest) 4, 235 (1953). - - 10 W~BURG, 0., F. Ku- BOWITZ U. W. CHriSTIAN: Bioehem. Z. 997, 245 (1931).
Dr. HANSJURGEN MATTHIES, Iusti tut fiir Pharmakologie u. Toxikologie der Humboldt-Universit~t Berlin NW 7 Clara-Zetkin-Str. 94.





























