Embed Size (px)
Citation preview

Zeitschrift fiir die gesamte experimentelle Medizin 144, 210--216 (1967)
Zum Ammoniakstoffwechsel der Leber*
PETER GLOGNER
Medizinische UniversitEtsklinik Tfibingen (Direktor: Prof. Dr. H. E. BOCK) und Medizinische Poliklinik der Universit~t Marburg/Lahn (Direktor: Prof. Dr. reed.
G. W. L6H~)
Eingegangen am 30. Juni 1966
Summary. 1. The effects of ammonia, on carbohydrate metabolites and urea in the perfusion medium of isolated rat livers and the mechanism of ammonia extrac- tion were studied. The influence of ammonia on oxygen consumption and carbon dioxide formation from glucose-6-14C of rat liver slices was also investigated.
2. After ammonia addition the levels of a-ketoglutarate and pyruvate fall in the medium. Ammonia disappears rapidly and the urea concentration rises with a slight retardation. The results are discussed.
3. Reduced blood flow and/or elevated portal blood levels result in higher ammonia extraction, suggesting non ionic diffusion of ammonia into the liver cell.
4. In liver slices ammonia inhibits g]ucose-6-14C decarboxylation in the citric acid cycle considerally more, than O 3 consumption. I t is concluded that glucose is substituted by other substrates maintaining tissue respiration.
Zusammen/assung. 1. An isoliert perfundierten Rattenlebern und Leberschnitten wurden der Mechanismus der Ammoniakextraktion und der EinfluB yon Ammoniak auf den Kohlenhydratstoffwechsel untersucht.
2. Nach Ammoniakbelastung fallen Pyruvat und a-Ketoglutaratspiegel im Perfusionsmedium ab. Ammoniak wird sehr raseh aufgenommen, und der Harnstoff- spiegel steigt verz6gert an.
3. Gedrosselte Durchblutung und/oder steigende NHs-Konzentrationen in der vena portae steigern die Ammoniakextraktion.
4. In Leberschnitten hemmt Ammoniak die Glucose-6-14C-Deearboxylierung erheblich, w~hrend der 02-Verbrauch kaum beeinflul~t wird.
5. Die erhobenen Befunde werden diskutiert.
R u n d 25 % des t/~glieh im mensehl iehen Organismus gebi lde ten t t a r n - stoffs d i f fundier t in den 1Vfagen-Darmtrakt, wird durch ureaseha l t ige Bak- te r ien gespa l ten und ge langt als A m m o n i a k fiber die P fo r t ade r in die Leber [13]. Dor t wird A m m o n i a k in Harns to f f fibergeffihrt , so dal3 im per ipheren Blur ein sehr n iedr iger Spiegel yon 0 ,5 - -1 ,0 7/ml A m m o n i a k - St ickstoff mel~bar ist. Bekann t l i ch finder sieh eine Hype rammon i i im ie vorwiegend bei schweren Lebere rk rankungen . Sic is t wahrscheinl ich ffir die Bewul~tseinsst6rung Lebe rk ranke r ve ran twor t l i eh . Ff i r die Neuro-
* Der Deutsehen Forschungsgemeinschaft sei ffir die Unterstiitzung der Arbeit gedankt.

Zum Ammoniakstoffweehsel der Leber 211
toxiciti~t des Ammoniak sprechen auch in letzter Zeit gefundene Enzym- defekte im Harnstoffcyclus [2, 6], die mit Erh6hung des Blutammoniak- spiegels einhergehen.
Veranlassung fiir die folgenden tierexperimentellen Untersuchungen war die Frage, welche physikochemischen Mechanismen fiir die Ammo- niakaufn~hme in der Leberzelle verantwortlich sind, urtd wie Ammoniak don Kohlenhydratstoffwechsel der Leber beeinflugt. Da fiber a-Keto- glutars/~ure Tricarbonsi~ureeyelus und Harnstoffsynthese verknfipft sind (reduktive Aminierung dutch Ammoniak mit NADI-I zu Glutamins~ure bzw. oxydative Decarboxylierung zu Suecinyl-Coenzym A), k6nnen StS- rungen im Gleichgewieht des Kohlenhydratstoffweehsels dureh Am- moniak erwartet werden.
Methoden Angaben fiber die Technik der Perfusion iso]ierter Rattenlebern und Substrat-
bestimmungen finden sich in der vorangegangenen Arbeit [3]. Als Perfusionsmedium wurde modifiziertes Rinderblut verwendet.
Versuehe mit Lebersehnitten: Jeweils 100 mg Leberschnitte wurden in Warburg- GefEflen mit Tyrodel5sung, enthMtend 0,2% Glucose, m/100 Phosphatpuffer pH 7,4 und 1 ~z Ci Glucose-6-14C (RadiochemicM Centre, Amersham, England) 1 Std unter 02 bei 370 geschiittelt. Stoppen der Reaktion dureh Einkippen yon 3% Pereh]orsEure (Endkonzentration) und Auffangen des retinierten C02 in kleinen Plastikbehiiltern, mit 0,1 ml Hyamine, Packard. Diese EinsEtze wurden direkt in PlastikmeBgefEBe der Fa. Medipha, Geislingen/Steige, iibergeffihrt und in iiblicher Weise mit 15 ml- Scintillator (4 g PPO und 100 mg Dimethyl-POPOP, Packard/1 Toluol) ira Flfissig- keitsseintillationszi~hler der Fa. Nuclear Corp., Chicago, gemessen.
Ergebnisse
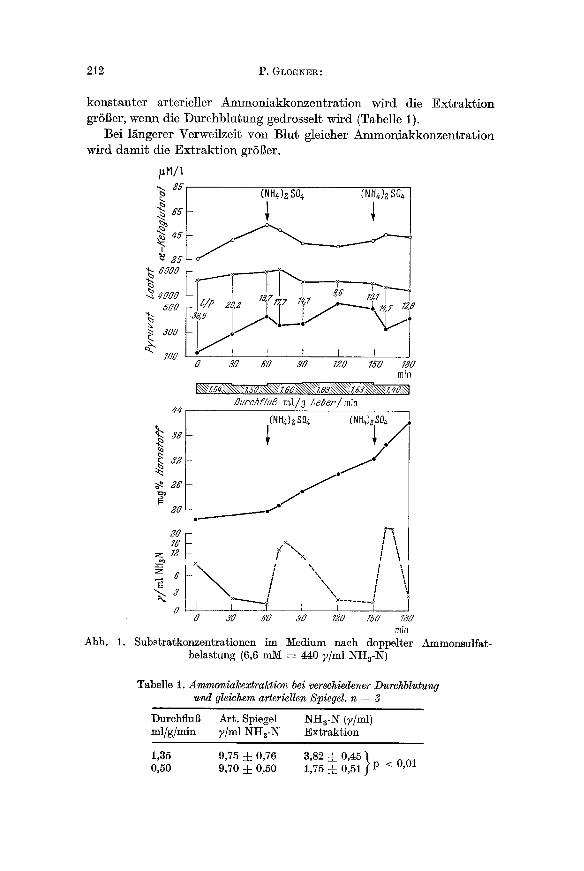
Wie frfiher gezeigt [3], vermag die isoliert perfundierte Rattenleber Ammoniak dem Medium zu entnehmen und den Blutspiegel w~hrend der Perfusion konstant niedrig zu hM~en. Nach Belastung mit hohen Ammoniumkonzentrationen lassen sich im Perfunda~ mehrere Ver~nde- rungen nachweisen (Abb. 1). Ammoniak wird mit groBer Geschwindig- keit yon der Leber extrahiert, so dab nach 10 rain weniger Ms 10% der vorgegebenen Menge nachweisbar ist. I)er in der Vorperiode anstei- gende a-Ke~oglutaratspiegel fi~llt erheblich ab. Die Ammoniakbelastung bewirkt auBerdem einen kurzfristigen Abfall der Pyruvatkonzentration, der sich rasch wieder ausgleicht (s. Diskussion). Zwischen Ammoniak- abfall und Harnstoffanstieg liegt eine zeitliche Verz6gerung.
Um den 1YIechanismus der Ammoniakextraktion zu studieren, wurden Belastungsversuche unter verschiedenen Durehblutungsgr5Ben und Blut- spiegeha durchgef~ihrt.
Wie aus Abb. 2 hervorgeht, nimmt bei kontinuierlicher ErhShung des arteriellen (portalen) Ammoniakspiegels die Elimination zu. Bei
212 P. GLOG~E~:
kons tan te r arterieller Ammoniakkonzen t ra t ion wird die E x t r a k t i o n gr6Ber, wenn die Durehb lu tung gedrosselt wird (Tabelle 1).
Bei 1/~ngerer Verweilzeit von Blur gleioher Ammoniakkonzen t ra t ion wird damit die Extraktion gr6Ber.
Abb. 1.
' f & ' | (NH4) ~ 804 (NH4)~. S04
N 35
L I T z/z z l~ZlZZ 1~,I :~__ 1~1 T
700 �9 I
##
~ ,qS -
~ 8Z,-
2 8 -
0 30 80 ,90 TZO 15-0 f80 rain
gunchflug ~l,/g lebeP/ rain
2 0 - ~
" X l g ~ •
~ zz I "~, I \ : ~ k ' N ' " / z # • I , , l -~ ~ / \', !
0 ]0 80 80 IZO 750
i NHz+)~ S04 (NH4)zS04 , l . /
_~..~.~'~'/
Substratkonzentrationen im Medium belastung (6,6 m M = 440 7/ml NHa-N )
; b'O rain
nach doppelter Ammonsulfet-
Tabelle 1. Ammonia~extraktion bei verschiedener Durchblutung und gleichem arteriellen Spiegel. n = 3
Durehflug Art. Spiegel NHa-N (y/ml) ml/g/min 7/ml NH3-N Extraktion
1,35 9,75 4- 0,76 3,82 4- 0,45 "~ 0,50 9,70 4- 0,50 1,75 4- 0,51 .~ p < 0,01
Zum Ammoniakstoffwechsel der Leber 213
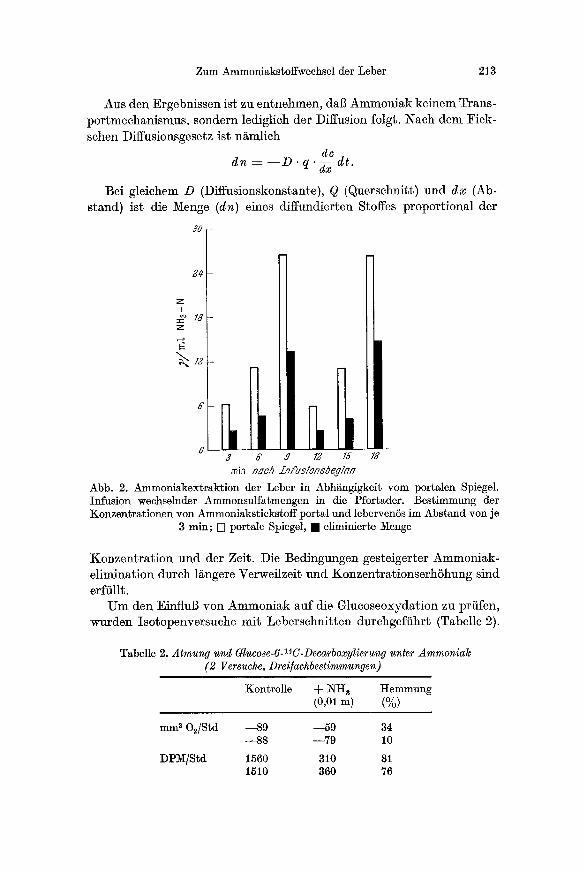
Aus den Ergebnissen ist zu entnehmen, dab Ammoniak keinem Trans- portmechanismus, sondern lediglich der Diffusion folgt. Nach dem Fick- schen Diffusionsgesetz ist ni~mlich
ds d n = - - D . q . dx dr .
Bei gleichem D (Diffusionskonstante), Q (Querschnitt) und d x (Ab- stand) ist die Menge (dn) eines diffundierten Stoffes proportional der
3 # -
2#
Z I
Z
3 6 # ~q 72 75 18
rain nach Infus/onsbeg/n#
Abb. 2. Ammoniakextraktion der Leber in Abh~ngigkeR vom port~len Spiegel. Infusion wechselnder Ammonsulfatmengen in die Pfortader. Bestimmung der Konzentrationen yon Ammoniakstiekstoff portal und lebervenSs im Abstand yon je
3 min; [] portale Spiegel, �9 eliminierte Menge
Konzentrat ion und der Zeit. Die Bedingungen gesteigerter Ammoniak- elimination durch l&ngere VerweilzeR und Konzentr~tionserhShung sind erffilR.
U m den EinfluB von Ammoniak auf die Glucoseoxydation zu prfifen, wurden Isotopenversuche mit Leberschnitten durchgeffihr~ (Tabelle 2).
14 Tabelle 2. Atmung und Glucose-6- C-Decarboxylierung unter Ammoniak (2 Versuche, Drei/achbestimmungen)
Kontrolle d- NHa Hemmung (0,0, m) (%)
mm a O~/Std --89 --59 34 --88 --79 10
DP)/I/Std 1560 310 81 1510 360 76

214 P. GLOGN~R:
W~hrend die Atmung dureh sehr hohe Ammoniakkonzentrationen (entsprechend 1,6 - 105 y/ml 1NHa-N) nur geringffigig beeintr~chtigt wird, kommt es zu einer starken Hemmung der Glueose-6-C-Decarboxylierung im Tricarbons/~urecyclus.
Diskussion
Die Untersuchungen zeigen, dab der Ketoglutaratspiegel im Perfu- sionsmedium nach Ammoniakbelastung abf~llt. Das Verhalten spricht ffir die besondere Bedeutung der ]~eaktion :
Ketoglutarat Jr NADH -~ H + ~- NtI4+ ---- Glutaminat -~ NAD [4]
als ersten Schritt der Ammoniakmetabolisierung. a-KGS ist im Gegen- satz zu Lactat and Pyruwt [10] wahrscheinlich night frei diffusibel. Schwankungen in der Mediumkonzentration kSnnen daher nieht die intracellularen Verhaltnisse, sondern nur die fallende Tendenz unter Am- moniak wiedergeben.
Der kurzfristige und reversible Abfall der Pyruvatkonzentration nach Ammoniakbelastung l~l~t mehrere Deutungen zu. Zu diskutieren ist eine besehleunigte NAD-Reduzierung, wodurch Pyruvat rascher oxydiert werden k~nn. Denkbar ist much eine gesteigerte l~aehbildung yon a-KGS aus Pyruvat, wobei NAD nieht den limitierenden Faktor darstellt. We- niger wahrscheinlich ist eine erhShte Glutamat-Pyruvat-Transaminie- rung, da diese Oxalacetat liefern wfirde, dieses jedoch unter Ammoniak intr~eellul~r erheblich absinkt [9]. Die VerzSgerung zwischen Ammoniak- abfall und tIarnstoffanstieg laBt darauf schlieBen, dab zun~ehst inter- mediare, stickstoffhaltige Produkte, wie Glutamins~ure, Glutamin and Asparagins~ure,intracellnls angeh~uft werden, bevor Harnstoff entsteht.
Ammoniak liegt bei 37 o und p g 7,4 ganz iiberwiegend ~ls NH4+-Ion vor (pK ~ 9). Dieses kann die Zellmembran night passieren [7], so dab aus der Elektronenreaktion NH4+ = NH 3 -~ I-I+ entsprechend den Gleichgewiehtsbedingungen permeables Ammoniak nachgebildet werden muB. In der Zelle wird es reionisiert und im Stoffwechsel aufgenommen.
Aus den Untersuchungen geht ferner hervor, dab der Ammoniak- anstieg im Blur durch l~tngere Verweilzeit und somit grSBere Extraktion verhindert werden kann. Diese Bedingungen sind bei einer Oesophagus- varicenblutung mit plStzlicher Drucksenkung im Pfortadersystem anzu- nehmen. AuI~erdem mug die Ammoniakextraktion naeh eiwei~reiehen Mahlzeiten, bei denen portale Spiegel von 10--15 N 1NHs-N/ml beob- aehtet warden [5], erheblich ansteigen. Dadurch bleibt die periphere Ammoniakkonzentration im 1Normbereich.
Nach unseren Ergebnissen wird Ammoniak durch Diffusion in die Leberzelle aufgenommen. Diffusion mul~ gleichzeitig der gesehwindig- keitsbegrenzende Schritt der Ammoniakextraktion sein: Der Konzen-

Zum Ammoniakstoffwechsel der Leber 215
trationsgradient kann nur dann aufreehterhalten werden, wenn jedes in die Zelle eintretende Molekfil im Sboffweehsel assimiliert wird. Dies ist naeh den vorliegenden Untersuchungen der Fall. Der gleiche Meehanis- mus gilt naeh Ammoniakverteilungsstudien (Blur/Liquor) bei Leber- kranken [8] auch fiir das Zentralnervensystem. In beiden Organsyste- men erfolgt demnach eine rasehe Assimilierung yon Ammoniak, wobei der Leber durch die groBe Kapazi tgt zur t tarnstoffsynthese die Haupt- rolle zuf/~llt, endogenes und exogenes Ammoniak zu entgfften. Ftir die Ammoniakaufnahme im Gehirn kommt zus~tzlieh der Aufnahmeweg a-Ketoglutarat--> Glutaminat--> y-Aminobutters/iure in Betraeht [11]. Seine Bedeutung ist noeh nieht vSllig aufgekl/~rt. Wie aus den vorliegen- den Befunden hervorgeht, kann die ammoniakbedingte Herabsetzung der Glueose-6-Deearboxylierung im Tricarbons/~ureeyelus in der Leber kompensiert werden: Die Zellatmung wird kaum beeintr/~chtigt, sic diirfte damit dureh Einsehleusen niehtglyeolytischer Zwisehenprodukte aufreehterhalten werden.
I m Gegensatz zur Leber ist das Gehirn ganz tiberwiegend auf die Glu- coseoxydation angewiesen. Ein Entzug yon a-Ketoglutarat aus dem Tri- earbons/~ureeyelus dureh Ammoniak, wie er yon RECKNAGEL U. POTTER [9] ffir die Leber gezeigt wurde, muB fiir das Zentralnervensystem sehwer- wiegende Folgen haben, da die Veratmung niehtglyeolytischer Stoff- weehselprodukte nieht ins Gewieh~ f/tilt [14].
Somit ergibt sieh ein weiterer Gesiehtspunkt fiir die yon B~ssMA~ u. B~SSMA~ [1] vertretene tIypothese, dab BewugtseinsstSrungen Leber- kranker mit I typerammonigmie darauf zuriiekzuf/ihren seien, dab Am- moniak-Ketoglutars~ure aus dem Tricarbons~urecyelus entfernt und die Zellatmnng gehemmt wird.
Der Beweis, dab Ammoniakkonzentrationen, wie sie bei komat6sen Leberkranken im Blur beobaehtet werden, die Atmung der Ganglien- zellen hemmen, steht ]edoch noch aus.
Literatur 1. B~SS~AN, S. P. : Ammonia metabolism in animals. Symposion on inorganic
nitrogen metabolism; p. 408. Baltimore: John Hopkins Press 1956. 2. COLOMBO, J. P., R. RIC~T~RLIC~, A. DONATe, A. SPerm% and E. RossI: Con-
genital lysine intolerance with periodic ammonia intoxication. Lancet 1964/I, 1014.
3. GLOGNER, P., I-I. WAGNER, K. GLOED:E U. IX. NIETH: Perfusion der isolierten Rattenleber mit Rinderblut. Klin. Wschr. 44, 1040 (1966).
4. KREBS, H. A., J. B. SUMNER, and K. lV[u The Enzymes. New York: Academic Press 1, 499; 2, 866 (1951).
5. MARTINI, G. A.: Die Bedeutung des Blutammoniaks fiir die Entstehung des Leberkomas. Dtsch. med. Wschr. 86, 1351 (1961).
6. Mcl~t'RR/~Y, W. C., J. C. RAT~Bm% F. ~o]tYm)nlN, and S. J. KOEGLER: Congenital citrullinuria. Pediatrics 32, 347 (1963).

216 P. GLOGNER: Zum Ammoniakstoffwechsel der Leber
7. MmNE, M. D., B. H. SChillER, and M. A. CRAWFORD: Non-ionic diffusion and the excretion of weak acids and bases. Amer. J. Med. 24, 709 (1958).
8. MOORE, E. W., G. W. ST~Om~EYER, and T. C. CKALMERS: Distribution of ammonia across the blood-cerebrospinal fluid barrier in patients with hepatic failure. Amer. J. Med. 85, 350 (1963).
9. R~CX~AGv, L, R. 0., and R. POTTER: Ketogenic effect of ammonium chloride. J. biol. Chem. 191, 236 (1951). "
10. SOHIMASSEK, H.: Der EinfluB der Leber auf den extracellul~iren Redox- Quotienten Lactat/Pyruvat. Biochem. Z. 886, 468 (1963).
11. SCnWA~E, U. : Vorkommen und Bedeutung der y-Aminobuttcrs~ure im Zentral- nervensystem. Dtsch. reed. Wschr. 86, 2235 (1961).
12. S~ERLOCK, S. : Krankheiten der Leber und der Gallenwege. 3. Aufl. Mfinchen: Lehmann 1965.
13. WALSER, M., and L. J. BODE~LOS : Urea metabolism in man. J. clin. Invest. 88, 1617 (1959).
14. WEIL-MALaEEB:~, H. : Der Energiestoffweehsel des Gehirns. Miinchn. med.Wschr. 104 (1962).
15. W ~ I ~ , L. P., P. A. PHEA~, and W. H. J. HVSTON: The oral ammonium test as aid in the investigation of suspected esophagogastric varices. J. din. Invest. 84, 15s (1955).
Dr. PETER GLOGNER Medizinische Universit~ts-Poliklinik 3550 Marburg/Lahn






![Morbidität und Mortalität nach Eingriffen an der Leber · PDF fileDetailreichere Aufschlüsse über die Anatomie der Leber gaben Capri (1470-1530) [14],](https://img.pdfslide.org/doc/110x75/5a7a216f7f8b9adf778d09f3/morbiditt-und-mortalitt-nach-eingriffen-an-der-leber-aufschlsse-ber-die-anatomie.jpg)