Embed Size (px)
Citation preview

(Aus dem Histologisohen Institut der I. Universit~it Moskau. Prof. A. GURWITSC~.)
ZUR ENERGETIK DER MITOGENETISCHEN INDUKTION UND ZELLTEILUNGSREAKTION.
(26. MITTEILUN[G UBER MITOGENETISCHE STRAHLUNG UND INDUKTION.)
Von
ALEXANDER und LYDIA Gul~WlTSCm
Mit 3 Textabbildungen.
(Eingegangen am 7. Mal 1928.)
Wir bat ten bereits in einer vorangehenden Mitteilung i die l%age nach den unmittelbaren :Folgen der Perzeption des mitogenetischen Reizes durch die Zelle aufgeworfen und zum Teil auch beantwor~et : es liel~ sich n~mlich Iiir gewisse Zellen der Zwiebelwurzel der •achweis erbringen, dab die Bestrahlung mit mitogenetischen Strahlen dm'ch 8ekund;~re mito- genetische Ausstrahlung aus diesen Zellen beantwortet wird. Es wurde auch darauf hingewiesen, daI~ diese Tatsache yon grundlegender Bedeu- tung fiir unsere Auffassung der Energetik der Zellteihmg werden kSnnte. :Die nachfolgenden Zeilen diirften die Berechtigung dieser Vermutung erweisen und uns einen tieferen Einblick in das Getriebe der Zelle un- mittelbar vor und eventuell auch wghrend der Zellteilung gew~hren.
Die mannigfaltigen, bei der Analyse der Sekund~rstrahlung sich ver- flechtenden Probleme diirften am klarsten aus der erneuerten und ver- tieften Analyse des Grundproblems hervorgehen. Dasselbe lautet :
Wie is t Fremdindukt ion miiglich?
Es mag ohne weiteres zugegeben werden, daI3 diese, eigentlich grund- legende Frage Yon uns bisher hie ernstlich diskutiert wurde, indem wir einfach yon der allgemeinen Annahme ausgingen, dab ein Mehr an Reiz- intensit~Lt auch eine Steigerung des Effektes erzeugen dfirfte, dal3 mit andern Worten Zellen, die auf autochthonen Reiz, d. h. die aus dem Orga- nismus selbst stammende mitogenetische Strahlung nicht ansprechen, bei Steigertmg der Reizintensit~t durch hinzukommende Fremdinduktion in den fiir die Einleitung der Mitose adgquaten Erregungszustand ver- setzt werden kSrmen. Diese fiir unsere Untersuchungen ursprfinglich mai3gebende Annahme, der zufolge in einem Zellverbande (z. B. Wurzel-
1 19. Mitteilung: Dieses Archiv 109, Hef~ 3.

Zur Energetik der mitogenetisehen Induktion und Zellteilungsreaktion. 741
meristem) empfindliche und ,,torpide" Zellen gesetzt wurden, wurde in der bereits erw/~hnten vorangehenden )Yitteilung dutch eine andere, wle es uns scheint zutreffendere und wohl definitive Fassung ersetzt, die etw~ so lautet :
In einem allgemein gleichartigen, vermehrungsf/~higen embryonalen Gewebe, wie es z. B. das ~eristem ist, sind alle Zellen an der Vcrmehrung beteiligt, indem sie die Teilungen in einem bestimmten Zeitturnus durch- machen; dessen mittlere Gr51~e fiir jede Wurzel ann/ihernd konstant, jedenfalls typisch ist und als Konstitution derselben (oder offenbar der zugehSrigen Zwiebel) bezeichnet werden mag. Diese mittlere (also streng genommen fikiive, abstrakte) GrSi3e des Turnus kann aus zwei empiri- schen ])aten, der Zeitdauer der Mitose und dem Prozentsatze dcr sich simultan teilcnden Zellen bestimmt werden. Setz~ man die Dauer der Mitose in der Zwiebelwurzel unSer normalen Verh/~ltnissen auf etw~ 1 Stunde i und die Gesamtzahl der ]~eristemzellen in einem Median- schnitt dt~rch eine kr~ftige Wurzel auf 4000 (mittlere GrSge aus einigen Zi~hlungen), so ist die mittlere Gr/SBe des Teilungsturnus auch bei roger Vermehrtmg noch sehr bedeutend.
So ]~l~t sich dieselbe fiir eine sehr hohe Mitosenzahl yon 200 l~itosen im Mcdianschnitte unter obigcn Voraussetzungen auf ctwa 19--20 Stun- den berechnen, was aus folgenden Uberlegungcn hervorgeht :
Der Prozentsatz der bei Fixierung gerade ange~roffenen Zellteilungen (d. h. das Verh~ltnis der 1Viitosen zur Gesamtzahl der l~eristemzellen) ist gleich dem Wahrscheinlichkeitsbruche, der die Wahrscheinlichkeit aus der Gesamtdauer (N+I S~unden) des Turnus gerade die eine, yon der Mitose besetzte Stunde herauszugreifen, zum Ausdruck bringS. In diesem Bruche steh~ 1 (eine Stunde prasumierte )[itosendauer) natiirlich im Zahler, der Nenner is~ gleich 2V + 1, d. h. =der Zeitspanne (N Stunden) zwischen zwei Teilungen+Teilungsdauer. Es or- gibt sich die einfache Gleichung
1 -P% - = 1 ~ ' in unserem Zahlenbeispiel is~ P -- 400~200 = 0,05,
woraus 2V= 19 Stunden. Bei I00 ~itosen pro Medianschni~b (der h/iufigste Typus) be~r/ig~ N e~wa
39 Stunden usw.
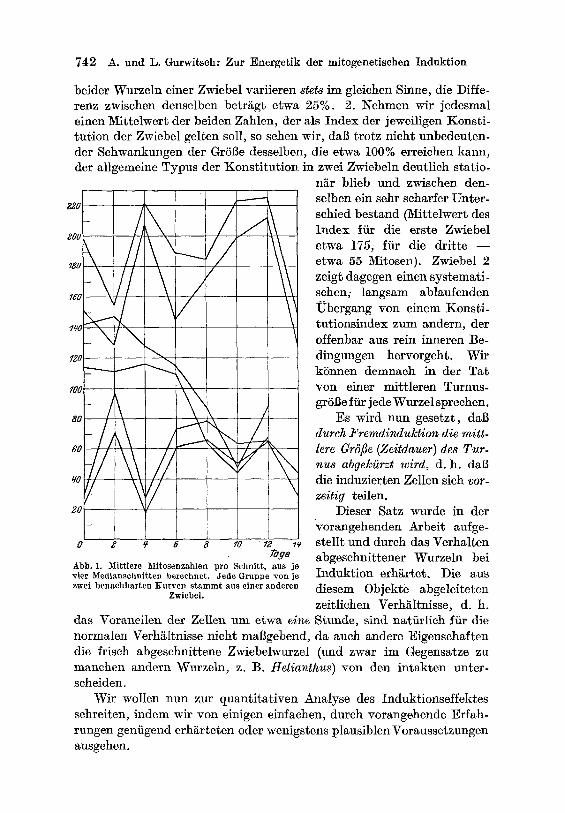
Dal~ eine individuelle , ,Konstitution" der Zwiebel tats~chlich be- steht und zi~he erhalten wird, diirfte aus Abb. i hervorgehen. Es wurden yon drei Zw~ebeln, die alle unter genau gleichen Bedingungen kultiviert wurden, ira Verlaufe yon 16 Tagen a11e zwei Tage (stets um 10 Uhr mor- gens) je zwei Wurzein (beliebiger L~ngen) genommen, in jeder je vier Medianschnitte auf die Mitosenzahl abgez~hlt und fiir jede Wurzel ein Mittelwert aus denselben gewonnen.
Die Ergebnisse der Z/~hlungen lehren uns folgendes: 1. Die Zahlen
1 Dieser (ann/~hernde) Wer$ ergibt Sich aus unseren Versuchen rnit Induktion abgeschnittener Zwiebelwurzeln (vgl. 19. Mitteilung, 109, Heft 3, S. 367).
742 A. und L. Gurwitsoh: Zur Energetik der mitogenetischen Induktion
220
180
160
beider Wurzeln einer Zwiebel variieren stets im gleichen Sinne, die Diffe- renz zwischen denselben betr~gt etwa 25%. 2. l~ehmen wir jedesmal einen ~Iittelwert der beiden Zahlen, der als Index der jeweiligen Konsti- fusion der Zwiebel gelten sell, so sehen wir, dal~ trotz nicht unbedeuten- der Schwankungen der Gr6Be desselben, die etwa 100% erreichen kann, der allgemeine Typus der Konst i tu t ion in zwei Zwiebeln deutlich statio-
n/~r blieb und zwischen den-
q0
Tage Abb. 1. Mittlere Mitosenzahlen pro Schnitt, aus je "~ier Med ianschn i t t eu berechnct , ffede Gruppr ,zon jo zwei b e n a c h b a r t e n K u r v e n s t a m m t aus einer anderen
Zwiebel .
das Voraneilen der ZeUen um etwa eine normalen Verh/s nicht ma$gebend,
selben ein sehr scharfer Unter- schied bestand (M_ittelwert des Index ffir die erste Zwiebel etwa 175, fiir die drit te - - etwa 55 M_itosen). Zwiebel 2 zeigt dagegen einen systemati- sehen; langsam ablaufenden Ubergang yon einem Konsti- tutionsindex zum andern, der offenbar aus rein inneren Be- dingungen hervorgeht. Wit kSnnen demnaeh in der Ta t yon einer mitt leren Turnus- gr6i~e ffir jede Wurzel spreehen.
Es wird nun gesetzt, dab durch 2"remdinduktion die mitt- lere GrSfle (Zeitdauer) des Tur- nws abgekiirzt wird, d .h . dab die induzierten Zellen sich vor- zeitig teilen.
Dieser Satz wurde in der vorangehenden Arbeit aufge- stellt und durch das Verhalten abgeschnittener Wurzeln bei ~ldukt ion erhKrtet. Die au~ diesem Objekte abgeleiteten zeitliehen Verh~ltnisse, d. h. Stunde, sind natorl ich fiir die da auch andere Eigensehaften
die friseh abgesehni~ene Zwiebelwttrzel (und zwar im Gegensatze zu manehen andern Wurzeln, z. B. Helianthus) yon den intakten unter- scheiden.
Wir wollen nun zur quant i ta t iven Analyse des Induktionseffektes schreiten; indem wir yon einigen einfachen, dureh vor~ngehende Erfah- rungen geniigend erh~rteten oder wenigstens plausiblenVoraussetzungen ausgehen.
und Zellteilungsreaktion. 743
1. Die mittlere Dauer der Mitosen ist ceteris paribus in den Wurzcln verschiedencr Konst i tut ion die gleiche.
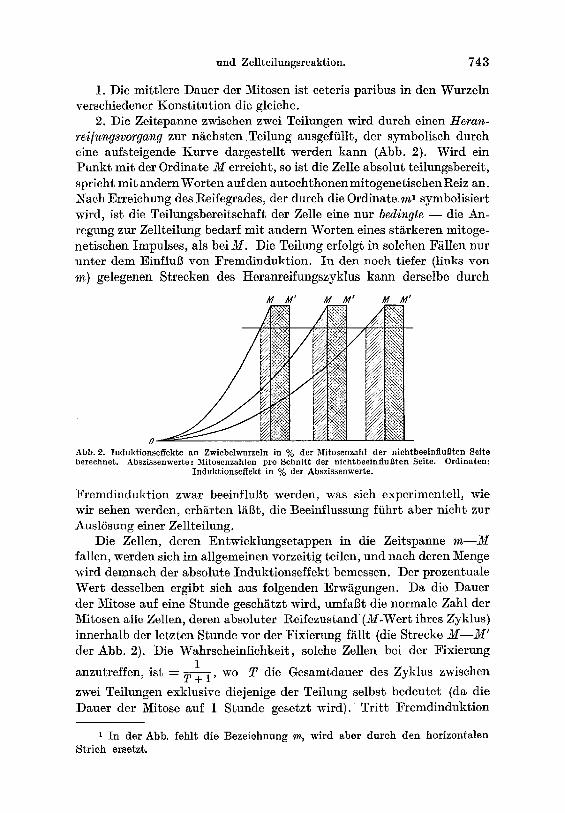
2. ;Die Zeitspanne zwisehen zwci Teilungen wird durch einen Heran- rei/ungsvorgang zur nachsten Tei lung ausgeffillt, der symbolisch durch eine aufsteigende Kurve dargestellt werden kann (Abb. 2). Wird ein Punkt mit der Ordinate M erreicht, so ist die Zelle absolut teilungsbereit, spricht mit andern Worten auf den autochthonen mitogenetischen Reiz an. Nach EiTeiehung des Reifegrades, der durch die Ordinate ml symbolisiert wird, ist die Teilungsbereitschaft der Zelle eine nur bedingte - - die An- regung zur Zellteilung bedarf mit andern Worten eines sts mitoge- netischen Impulses, als bei M. Die Teilung erfolgt in solchen F~llen nur nnter dem EinfluB yon :Fremdinduktion. In den noch tiefer (links yon m) gelegenen Strecken des Heranreifungszyklus kann derselbe durch
M M' M M' M M'
Abb. 2. Induk t ionse f fek te an Zwiebelwurzeln in % der Mitosenzahl de r n iehtbeeinf luBten Sei te berechnet . Absz i s senwer te : Mitoscnzahlen pro Schn i t t der n i ch tbee in i iuBten Seite. Ord ina ten :
Induk t ionse f f ek t in % der Abszissenwer te .
Fremdindukt ion zwar beeinfluBt werden, was sich experimentell, ~de wit sehen werden, erh~rten l~gt, die Beeinflussung ffihrt aber nicht zur Ausl6sung einer Ze]lteilung.
Die Zellen, deren Entwicklungsetappen in die Zeitspanne m - - M fallen, werden sich im allgemeinen vorzeitig teilen, und nach deren Menge wird demnach der absolute Induktionseffekt bemessen. Der prozentuale Wef t desselben ergibt sieh aus folgenden Erws Da die Dauer der Mitose auf eine Stunde geschi~tzt wird, umfaBt die normale Zahl der Mitosen alle Zellen, deren absoluter Reifezustand (M-Wert ihres Zyklus) innerhalb dcr letzten Stunde vor der Fixierung fallt (die Strecke M - - M ' der Abb. 2). Die WahrscheinHchkeit, solche Zellen bei der l~ixierung
1 anzutreffen, ist -- T + 1' wo T die Gesamtdauer des Zyklus zwischen
zwei Teilungen exklusive diejenige der Teilung selbst bedeutet (da die Dauer der l~iitose auf 1 Stunde gesetzt wi rd) . Tr i t t Fremdinduktion
1 In der Abb. fehlt die Bezeizhnung m, wird aber dureh den horizontalen Strich ersetzt.
744 A. und L. Gurwitsch: Zur Energetik der mitogenetischen Induktion
hinzu, so verl~ngert sich die Zeitspanne, die die sich teilenden Zellen umgreift und ist nunmehr gleich dem Bereich m - M'. Der Wahr- scheinlichkeitsbrueh erh~It nun an Stelle des Z~hlers 1 einen neuen Zi~hler a (a = der Streeke m - - ~ l der Abb. 3). Es entsteht nun das Problem den zeitlichen Rhythmus des dutch die Kurve veranschaulichten Heran- reifungsvorganges nach MSgliehkei~ zu analysieren, um daraus Schliisse fiber dessen Inha l t ziehen zu kSnnen.
Die grundlegende in erster Linie sich aufdr~ngende Frage li~Bt sich hier etwa folgendermal]en formulieren: bleibt die Kurve, die den Heran-
700g080 --*:
70 �9 ~ \ 8G ~, ~ ""
0, i . . ** *I
�9
za ~ �9 �9 I , �9
I 0
g
O e
0 10 20 30 qO 50 GO 20 80 ,r ~'00 ~'~IO 180
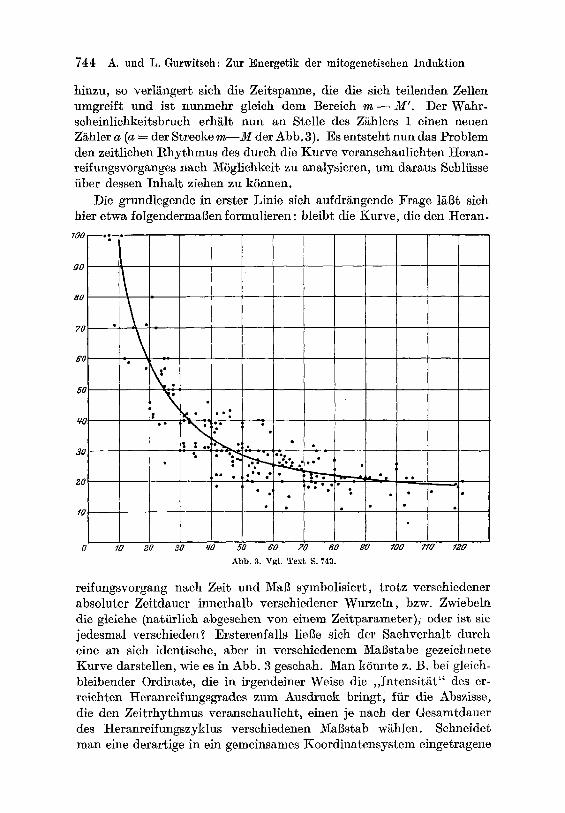
A b b . 3. u T e x t S. 743.
reifungsvorgang nach Zeit und Ma]~ symbolisiert , t rotz verschiedener absoluter Zeitdauer innerhalb verschiedener Wurzeln, bzw. Zwiebeln die gleiche (natiirlich ~bgesehen yon einem Zeitparameter) , oder ~st sie jedesmal verschieden? Ersterenfalls liel3e sich der Sachverhal~ dutch eine an sich identisehe, aber in versehiedenem Mal3stabe gezeichnete Kurve darstellen, wie es in Abb. 3 geschah. Man kSnnte z. B. bei gleich- bleibender Ordinate, die in irgendeiner Weise die , , Intensit~t" des er- reichten Heranreifungsgrades zum Ausdruek bringt, fiir die Abszisse, die den Zei t rhythmus veranschaulicht, einen je naeh der Gesamtdauer des Heranreifungszyklus verschiedenen )'IaBstab w~hlen. Schneider man eine derartige in ein gemeinsames Koordinatensystem eingetragene

und Zellteilungsreaktion. 745
Kurvenschar durch eine der Abszisse parallele GeradeL die dureh die Punkte m der Kurven gezogen wird, so werden durch die FuBplmkte der Ordinaten der Schnittpunkte, die Abszissen des jeweiligen Zyklus in zwei Strecken geteil~, deren VerhMtnis ffir alle Kurven konstant bleibt (vgl. Abb. 22). Die durch diese Operation aus dem Gesamtzyklus ausgesonderte , ,aktive" Strecke (m - - M) ist dabei mit andern Worten der Gesamtlgnge des Zyklus proportional. Man kann aber auch umgekehrt , allerdings nicht in obligatorischer, sondern nur in sehr plausibler Weise argumen- tieren. W e r d e n bei der i m vorangehenden darges te l l ten Opera t ion yon der
7 (urvenschar propor t iona le S t r e c k e n geschni t ten , so l iegt grofle W a h r s c h e i n -
l i chke i t ve t , daf t d ie K u r v e n z u m m i n d e s t e n i n i h rem aUgemeinen Ver lau fe
iden t i sch s ind . DaB eine derartige SchluBfo]gerung yon groBer Bedeu- tung ist, ~ i rd sich aus dem folgenden ergeben.
Es liiBt sieh nun aus unserm Versuchsmaterial eine derartige an- n~ihernde 1)roportionalitht der Strecken m - - M in der Ta t erkermen, was aus folgenden lJberlegungen und Befunden hervorgeht.
Wir sahen bereits, dab bei ) ' remdinduktion der Wahrscheinliehkeits- 1
bruch ~-~-T durch einen solchen mit einem neuen Zithler erset.zt werden
m u l l Sell die Voraussetzung, dai~ die ,,mltotmc e Zeitspanne m - - M um einen, dem Gesamtzyklus proportionalen Betrag zunimmt, zutreffen, so muB der neue Z/ihler a die Form 1 + k T (k - - eine konstante zuni~ehst unbest immte Gr6Be) haben. Wir schreiben demnach:
1 P = i-u . . . . . . . . . (I)
ffir autochtone Induktion, we p der Prozent der sich simultan teilenden ( absolute Mitosenzahl~
Zellen ist \ ~ ~ t ~ a h i / and l + k T
P + P - - I + T - - " . . . . . . . (2)
fiir l~remdinduktion, we P der Induktionseffekb in Prozenten der abso- luten Zellenzahl ist.
Dann haben wir l + / c T 1 kT
P = - I + T - - I + - - T = i + T . . . . . . . (3)
Indem wir aus der Gleiehung (3) das T eliminieren, erhalten wir folgen- den Ausdruck:
Aus Gleichung (1) folgt:
T= 1 1. p
Setzen wir diesen Wert in Gleichung (3), so erhalten wir:
p = k (Up - 1) _ k(1/p - 1) _ k - - k p . . . . . (4) 1 + 1/p - 1 1/p
An dcn Schnittpunkten fehlt die Bezeiehnung m. " 1). h. O M / m M = Konstant.
W. ROUX ~ Archly f. Entwiek lungsmechanik Bd. 113. 49

746 A. und L. Gurwitsch: Zur Energetik der mitogenetisehen Induktion
W i r wollen nun be ide H~lf ten der Gle iehung (4) P -~ k -- kp durch p d iv id ie ren und k o m m e n zum Ausdruck
P k - ~ . . . . . . . . . ( 5 )
p P ] )a die W e r t e p bzw. P alas p rozen tua le Verh~l tn is der n o r m a l e n (p) u n d der induz ie r t en (P) Mi tosen zur Gesamtzah l der Zel len ausdr i ickcn , wi r4 aus dem Bruch P / p l c t z te re r W e r t e l imin ie r t . P / p bedeu te t demnach den Induk t ionse f f ek t be rechne t in P rozen ten der no rma len Mi tosenzahl (bzw. der Mi tosenzahl der nichtbeeinfluBgen H/i l f te des Schni t tes) . Also z. B. Mi tosenzahl an der indu iz ie r t cn Seite 750, an der n i ch t induz ie r t en 500. P = 250. P / p = 50%.
Unsere Berechnung ging yon der Vorausse tzung der •ons t anz yon ]c aus. Diese Se tzung i s t g l e i ch lau tend m i t der A n n a h m e , dab die Ab- sz issenst recken m - - M und O - - M unscrer Abb . 2 c inander proportional sind. Gil t diesc Sctzung bzw. is t /c - kons t an t , so s te l l t die Gle ichung (5) eine gleichschenkel ige, auf ihrc A s y m p t o t e n als Achsen bezogene H y - pe rbe l dar , wo wir P / p als Yq-]c, p - - als X se tzen 1.
Es b l e ib t uns j c t z t demnach der l e tz te Schr i t t , i ndem wi t unser Ver- suchsmate r i a l auf e in K o o r d i n a t e n n e t z au f t r agen und die Ubere ins t im- mung der e rha l t enen Einze lwer te m i t der Gleichung (5) prfifcn. Die ab- so lu ten Mi tosenzahlen der n i ch t induz ie r t en Seite (die j a p ropo r t i ona l p, e twa ----- 2000 p sind), werden als Abszissenwerte , das Verh~l tnis P i p absoluter Induktionseffekt~ M ~ a ~ ~ ] als Ord ina tenwer t e aufge t ragen . E ine K u r v e
wird in m6gl ichs t ob j ek t ive r Weise den E inze lwer ten in t e rpo l i e r t und ihr AnschluB an die Gle ichung (5) geprfif t . Als eine solche In t e rpo l a t i ons - kurve is t in Abb . 3 eine glcichschenkel ige, nach der pos i t i ven 2 R i c h t u n g der Y-Achse verschobene glc ichschenkel ige H y p e r b e l e ingetragena. Dcr AnschluB des Versuchsmate r i a l s an diese I n t e r p o l a t i o n s k u r v e is t , wie ers icht l ich , ein sehr guter .
Die Diskuss ion dieser Gle ichung in bezug auf unsere Verh~l tn isse er- gibe fo lgendes :
1 Der Ausdruck - - P -- Y + k bedcutet, dab die Abszissc, an der das empi- P
rische l~aterial aufgetragen wird, um k nach der negativen Richtung der Y-Achse gegen die horizontale Asymptote verschoben ist.
2 Die horizontale Asymptote entsprieht dem I~orizontalstrieh 10. 3 Die Abh/~ngigkeit zwischen der Anzahl der Mitosen in den Wurzeln und
dem Induktionseffekt ist auch WAGing (Planta, Bd. 5) nicht entgangen (vgl. S. 82 und Abb. 3, S. 83). Ich kann aber seinem Satze, daB: ,,alle Versuehe mit Wurzeln, die eine groBe Mitosenzahl enthalten, immer ein negatives Resultat ergeben", absolut nicht beipflichten, da in unseren zahlreichen Versuchen, Wur- zeln mit sehr hohen Mitosenzahlen (fiber 160 bis 240 Mitosen im ]Icdianschnitt) stets einen absolut sehr bohen, prozentual aber geringen (Wert der interpolierten Kurve Abb. 3 = 21 bis 18%) Induktionseffekt ergaben.

und Zellteilungsreaktion. 747
1. 29 = dem Prozent der sich simultan ~eilenden Zellen auf die Gesamtzahl dcr ~Ieristemzellen bezogen, ist stets eirt kleiner Bruch (die sehr hohe :~Iitosenzahl = 200 ~[itosei1 pro Sehnitt, entspricht nur 5%). ])as erste Glied der rechten Seite =k/p ist daher stets grol~ im Vergleich zu k.
2. Der Brueh P erreicht nut hSchst selten den Wert 1 (= 100%), hat ihn bis- 79
her nicht iibersehritten. 3. Die Abszissenwcrte sind n~tfirlich nicht gleieh p, sondern demselben nur
proportional, kfnnen aus 79 mi~ genfigendem Genauigkeitsgrad dutch Multipli- kation mit 2000 berechnet werden (Gesamtzahl der ~ieristemzellen in einer t tglfte eines Medianschnittes).
4. Eine prinzipielle Abweichung der empirischen Kurve (Abb. 3) yon der Formel 5 ist schliel31ich im Vorzeichen des konstanten Gliedes K in der linken H~lfte der Gleichung gegcben, welches in letzterer ncgativ, im empirischen Ma- terial dagegen posi t ivis t (da die erapirisehe Kurve in positiver Y-Riehtung gegen die X-Achse verschoben ist). Der konkrete Sinn der durch die Abb. 3 veran- sehaulieht.en Abh~ngigkeit ist, wie leicht einzusehen, folgender. Der Induktions- effekt setzt sich aus zwei Gliedern zusammen: 1. Das erstc ist prozentual der Gesamtdauer eines Teilungsturnus proportional (oder dem 3'[itosenreichtum um- gekehr~ proportional). 2. Das zweite Glied ist konstant ( = k), d .h . yon den in 1. erw~thnten Umst~nden v61lig unabhfingig. Die Interpretation, die nmn diesen Tatsachen geben kann, ist etwa folgende: Der Induktionseffekt P, der
k T laut Formel (3) = ~ - T sein mfil~te, hat den tatshehlichen Ausdruek kT+l+TC
(c = konst.) (6). Es wird demnach dutch Induktion aus der ~eranreifungskurve tats~chlieh ein Bereich ,,herausgesehnitten", der der Gesamtdauer des jeweiligen Zyklus proportional isb, was de~ Wat~rscheinllchkeitssctdufl gestattet, daft die den Heranrei[ungszvrgan~ symbollsierenden K~zrven $rotz der Verschiedenheit der abso- lumen Zei$mafle sich ste~s gleich bleiben, oder bildlich ausgedriickt, zeltllcF~ ,,z~- samme~geclr?ingt" werde~ k6~nen, ohne sich zu de[ormierer~. Wean wir dieser rein formellen Dnrstellung elnen konkreten Inhal~ zugrunde legen, so ergibt sieh folgende VorsteUung yon weittragender Bedeutung.
Die , ,Heranre i fung" invo l ie r t u n b e s t r e i t b a r chemische Prozesse, und zwar hSchs t wahrscheinl ieh eine al lmiihl iche Speicherung energiel iefern- der Stoffe, die durch mi togene t i sehe S t r ah l en zersc tz t werden, wobei aus de r dabe i frei gewordenen Energie die Arbeib der Ze l l te i lung be- s t r i t t en wird . Der fiir d ie e inzelnen Wurze ln so verschiedene ze i t l iche R h y t h m u s der e inzelnen , ,Heranre i fungszyk len" miil3te demnach als ein verschiedenes Intensit~tsm~13 oder Geschwindigke i t der Aufspeiche- rung dieser 8toffe be t raehte~ werden Es l iegen nun zwei M6glichkeiter~ vor, wie m a n sich diese Abl~ufe zu denken h~t te . Es w~re denkbar , dal3 de r ganze Ab lau f aus e iner K e t t e yon verschiedenen chemisehen U m - s~tzen, even tue l l aueh s imu l t anen Prozessen bestehb, oder dal~ nu r ein grund legender Vorgang, z. B. Spe icherung eines be s t immte n Stoffes bis zu e iner m a x i m a l e n K o n z e n t r a t i o n im Spiele sei. Die W a h r u n g e ine r Ze i tp ropo r t i ona l i t~ t a l ler Gl ieder eines komplexen Ablaufes (erste MSg- l ichkei t ) i s t u n t e r b e s t i m m t e n Ums t~nden zwar nich~ unm6gUeh (w~re z. B. bei Abh~ngigke i t des R h y t h m u s des g~nzen Zyk lus yon der Tern-

748 A. und L. Gurwitseh: Zur Energetik der mitogenetischen Induktion
peratur zu erwarten), erscheint abet unter den gegebenen Verhaltnissen (d. h. bei der individuellen, relativ konstanten Veranlagung einer Zwiebel zur Reproduktion eines bestimmten Rhythmus der Zyklen) in allen Wur- zeln als sehr unwahrseheinlieh. Es waren hier jedenfalls komplizierte und schwer motivierbare Hilfsannahmen unerlaBlich. Um so plausibler erscheint dagegen die aufgedeekte Proportionalitat, bzw. die gesetzte ]~onstanz der Kurvenform, wenn wir uns den ganzen Ablauf der Heran- reifung als einen einheitlichen, monoton verlaufenden ProzeB, z. B. An- reicherung eines bestimmten Stories denken. Die durch die verschiedene Lange der Zyklen gekermzeiehnete individuelle, sich so za.he erhaltende Geschwindigkeit diese Vorganges ware natiirlich in ph~usibler Weise aus den konstitutionellen Verhaltnissen der Zwiebel, bzw. der Wurzeln ab- zuleiten. Denken wir uns z. B. individuelle Schwankungen in der Aus- bildung der Zwiebelsohle, der LeitgefaBbiindel~ des Gehaltes der Zwiebel an Speieherstoffen usw. so erscheint es ohne weiteres verstandlieh, dab die Versorgung der Meristemzellen mit dem bei Heranreifung in Betracht kommenden Grundstoffe naeh einem fiJr alle Individuen gleiehbleiben- den Gesetze, aber mit einem verschiedenen Zeitparameter geschieht.
Unsere definitive hypothetisehe l~ormulierung des Induktionseffcktes w/~re auf Grund des Vorangehenden etwa folgende:
Die Zellen rei/en zur Mitose allm~ihlich und stetig heran, indem sie einen energielie]ernden Sto/] anreichern. Die mitogenetische Indulction besteht in einer Zersetzung desselben und entsprechender Energieentbindung, die hauptsiichlich die Arbeit der Zellteilung leistet.
Dieser einfache Satz bedarf eines weiteren Zusatzes. Der angereicherte Stoff ist zwar, soweit ersichtlich, stets zersetzungsfahig. Es l~.13t sich aber durchaus nicht beh~upten, dult die dadurch entstchende Energie- entbindung in eindeutiger, zwangsm~iger Weise zur Mitose ifihre. Es gehSrt dazu in erster Linie eine bestimmte Schwellenkonzentration des euergieliefernden Stories (der Bereieh ~ t ~ - M unserer Graphik), d. h. ein Bestimmungsmoment offenbar rein quantitativer Art. Abet ab- gesehen davon w~re es natiirlieh noch verfriiht, den Komplex der ,,MSg- lichkeitsfaktoren" auf den einen, relativ elementaren Ablauf zuriick- zufiihren~.
1 Speziell das konstante Glied c (Formel 6) mahnt zur Vorsicht. Es entstehen bei Induktion.tats~chlich um e Mitosen mehr, als aus vorangehenden S/~tzen zu erwarten w/~re, i~Ian muB abet dabei den Umstand in Betraeht ziehen, dab die theoretisehe Ableitung unter Voraussetzung eines praktiseh momentanen Induk- ~ionsreizes entwiekelt wurde, wie er bei Versuchen mit Spektrallinien (1 Sekunde) tatsgchlich verwirklieht wird. Unser in Abb. 3 zusammengestelltes empirisches Material bezieht sieh ~ast ausschliei31ioh auf Versuehe mi~ biologischen Induk~ions- quellen mit langer Induktionsdauer. Es findet demnach eine ~berlagerung der einzelnen induktionsschritte statt, die sehr wohl fiir das Auftreten des ~ber- maBes an Mitosen verantwortlich gemacht werden kSnnen.

und Zellteilungsreaktion. 749
Unsere Hypothese mul3 und ]<ann auch durch eine Anzahl Tatsachen gestiitzt werden. Die Beweisfiihrung diirfte am besten unter ~Beriick- siehtigung der sich aus der Grundannahme ergebenden Konsequenzen durchfiihrbar sein. Sie lassen sich etwa ~ie folgt formulieren:
1. Wird die Stoffanreicherung als ein stetiger und allm/ihlicher u gang gesetzt, so miissen Korrelate dessen Zersetzung durch mitogeneti- sche Strahlen auch im Bereiche des Gesamtzyklus auflerhalb der Streeke m - - M naehweisbar sein.
2. Beruht der positive Induktionseffekt auf vorzeitlgem Verbrauehe des gespeicherten Stoffes, so mul~ demselben eine Art Depressionszustand nachfolgen, indem eine Anzahl sonst f/illiger Zellteilungen ganz weg- bleiben, bzw. mit Versp/itung auftreten.
3. Ist fiberhaupt Fremdinduktion miig]ich, so mul~ irgendeh]e quan- ti tative Beziehung zwischen Belichtungsintensit/it, bzw. Reizmenge und Reaktionseffekt naehweisbar sein.
4. Es w/ire denkbar, dab die Stoffanreicherung dutch entsprechende Beeinflussung besehleunigt oder begi~nstigt werde, was den Prozentsatz der Mitosen steigern miif~te.
Fiir alle vier Punkte lassen sich Belege anfi~hren. Was den ersten Punkt betrifft, so diirfen wir mi~ guten Griinden
setzen, dab das in den vorangehenden Arbeiten (dieses Archly, Bd. 109, S. 373 und Bioeh. Zeitschr. 128) ausfiihrlich geschilderte Ph/inomen tier Sekund/irstrahlung das gesuehte Korrelat ist. Von ganz anderen Erw/igun- gen ausgehend, haben wit nach ausfiihrlieher Diskussion des Problems die Ansieht ausgesprochen, daB: ,,als unmittelbare Reaktion reizrezeptiver Zellen auf mitogenetisehe Bestrahlung das Aussenden sekund/irer mito- genetiseher Strahlen erblickt werden mu$. Diese Reaktion diirfte nur in einen zeitlich ziemlich eng zirkumskripten Punkt der Latenzperiode der Meristemzellen fallen" (S. 377). Diese Vermutung erf/ihrt nun durch unsere jetzigen Ausfiihrungen eine tiefere Begriindung, wenn aueh ein strenger Beweis hier noch nicht erbracht wird i. Fiir die in den Punk- ten 2---4 angefiihrten Postulate lassen sieh dagegen ganz eindeutige Be- lege anfiihren: eine Depression (oder wie wir welter sagen werden - - Ersch6pfung) konnte in aller w/insehenswerten Klarheit nachgewiesen werden (vgl. die nachfolgende Arbeit von Frl. SUSS~r~OW~TSC~).
Es mag bier in allgemeinen Ziigen die Theorie der zu erwartenden Erseheinungen gegeben werden.
Es sollen alle Zellen des l%[eristems nach ihrem I~eifegrad in Gruppen naeh Zeitintervallen eingeteilt werden und zwar so, dab je n/iher der Reife die betreffende Gruppe, desto ,,empfindlicher" fiir die Strahlen, d . h . desto grOBer der Prozentsatz der vorzeitig , ,entladenen" Zellen. DaB aul]erdem der Prozentsatz der ]Sntladung auch als Funktion der
1 Er wird in einer demn&chsb erscheinenden Arbei~ gegeben.

750 A. und L. Gurwitsch: Zur Energetik der mitogenetischen Induktion
Intensi tEt und Dauer der Belichtung gesetzt wird, wurde bereits hervor- gehoben. Es seien z. B. Zeitin~ervalle yon 12 Minuten gesetzt. Die Dauer der Mitose auf 1 Stunde gesetzt, werden wir im fixierten Pr/~parat fiinf solche Intervalle als Mitosen vorfinden bzw. abz/~hlen. Wir wollen zunttchst den Fall nehmen, dal~ eine einmalige zum positiven Induk- tionseffekt ausreichende Reizmenge gegeben wird. Handel t es sich um eine Spektrallinie, so geniigt erfahrungsgcm~l~ 1 Sekunde. Bei einer der- artigen Reizgabe soll folgendes geschehen : Die absolut teilungsreife Zell- gruppe (1), die eben durch autochthone Indukt ion zur Teilung angeregt wird, wird natiirlich nicht wciter beeinflu2t. Es werden demnach, yon dem Augenblick der ersten l~eizung ab gerechnet, die Zellen der Gruppe 1 nach 2 Stunden zur Teilung schreiten, naeh 3 Stunden sich in Telophase befinden.
Von der Gruppe 2 sollen 50% durch Fremdinduktion vorzeitig ,,ent- laden" werden, was fiir diese Gruppe mit vorzeitiger Mitose gleichbe- deutend ist. Es werden demnaeh 2 Stunden naeh der ersten l~eizung, ab- gesehen yon allen Zellen der Gruppe 1 noch 50% der Gruppe 2 in Mitose angetroffen. Es sollcn aber abgesehen yon 50% der zweiten Gruppe, etwa 30 % der dri t ten Gruppe und 15% der vierten Gruppe durch den l~eiz zur vorzeitigen Ent ladung gebracht werden, was aber bier nicht zur vorzeitigen Mitose, sondern laut Voraussetzung, zur Verstrahlung der Energie und folglich zum Ausbleiben der rechtzeitigen Mitose dieser Zellen f fhren mug. Reift jetzt die Gruppe 2 zur Teilung heran, so sind es nur 50% der Normalzahl, da die ersten 50% sieh vorzeitig mit der ersten Gruppe teilten. K o m m t die dritte Gruppe an die Reihe, so wer- den sich 70% der Normalzahl teilen, bei der vierten Gruppe 85%, bei der ffinften Gruppe sehon 100%.
Das Ergebnis wird versehieden ausfallen, je nachdem wir die Wurzel 2 oder 3 Stunden nach der Induktion.f ixieren. Gesehieht ersteres, so miil3ten die Zellen der Gruppe 1 und 50% der Gruppe 2 in Mitose ange- troffen werden, die Defizite der iibrigen Gruppen kSnnten sieh aber noeh nicht geltend machen, da die Zellen derselben noch gar nicht zur Mitose schreiten. Erfolg~ die Fixierung nach 3 Stunden, so h/itten wir etwa folgendes: Neben 150% Telophasen (aus den zwei ersten Gruppen) nur 50% etwas jfingeren Stadiums (aus dem Rest der zweiten Gruppe), 70% der drit ten Gruppe, 85% der vierten Gruppe und 100% der fiinften Gruppe ~.
Es ergebe sich alles in allem ein Defizit aus der dri t ten Gruppe (30%) und der vier tenGruppe (15%), d.h. 45%, oder auf fiinf Gruppen bezogen 45 �9 - = 9 % . 5
1 Die Prozentzahlen jeder Gruppe jedesmal auf die entsprcchcndcn Gruppen der normalen Seitc bezogen.

und Zellteilungsreaktion. 751
Es versteht sich yon selbst, dab es sich um ein ganz rohes Schema handelt , aus dem nur herausgelesen werden kann, dab bei einmaliger kurzdauernder Indukt ion ein zeitlich relat iv eng umgrenzter positiver Induktionseffekt erwartet werden muB, dem eventuell sofort ein ebenso kurz w/ihrcnder schwacher ErschSpfungszustand nachfolgen k6nnte. Dal~ die Gegens/~tze zwischen positivem Effekt und Defizit in Wirk- lichkeit nicht so scharf sein werden und namentlich letzteres der Fest- stellung auch leicht entgehcn kann, braucht nicht erst betont zu werden.
Es wurde im Vorangehenden yon einer Reizmenge ausgegangen, die zur Beeinflussung yon 50% der zweiten Gruppe ausreicht, dabei aber so kurz bemessen ist, dab sie im Vergleich zu den Teilungsabl/~ufen, gewisser- mal]en als momentan bezeichnet werden kSnnte, l~ehmen wir nun jetzt die glciche Reizmenge mit geringer Intensit/~t, so f/iilt schon die Zeit- dauer der Reizwirkung sehr in Betracht. Es handle sich z. B. um 12 Mi- nuten Indukt ionsdauer (was ann/~hernd dem effektiven wirksamen Mi- nimalwert bei biologischer Indl~ktionsquelle entspricht), yon viel ge- ringerer Intensi t~t als im ersten Falle. Der Prozentsatz der beeinflui]ten Zellen jeder Gruppe miiBte hier viel niedriger gesetzt werden, z. B. 30% ffir die zweite, 15% ffir die drit te und 0% ffir die vierte Gruppe. Es h~tte sich aber bei Fixierung nach eLwa 2 Stunden folgendes ergeben: der Pluseffekt bestfinde aus 30% vorzeitiger Mitosen der zweiten Gruppe, die schon innerhalb der ersten Sekunden der Indukt ion zur Teilung an- geregt wfirden, und aus 30% der Zellen der drit ten Gruppe, die mittler- weile (d. h. gegen AbschluB der Induktion) in die zweite einrfickte. Da abet yon dieser Gruppe schon vorhin 15% vorzeitig entladen x~alrden, miissen wir die 30% yon 85 und nicht yon 100 berechnen, was etwa 25% ergeben wird. Mir ba t ten demnach auch bier einen Totaleffekt von eLwa 50%, wodurch dem Reizmengensatze vSllig Geniige getan wird. Die auf den ersten ]3lick absonderliehe Tatsache, dab die auBerordentlich sehwachen biologischen Quellen und die ganz unvergleichlich st/~rkeren Spektrallinien zum gleichen Effekt fi]hren, erfKhrt dadurch ihre volle Erkl/irung.
Wir wollen je~zt die Verh~ltnisse bei dauernder Belichtung des Me- ristems betrachten, wobei wir der gr61leren Anschaulichkeit wegen eben- falls yon den beiden eben besprochenen Quellen mi t ihren bzw. Zahlen ausgehen. Nehmen wir zun~chst die st~rkere vor, wo die ausreichende l~eizmenge = 1 Sekunde ist, so w/irde nach unserm Schema eine un- unterbrochene Belichtung yon 1 Stunde (oder was auf das gleiche hinaus- kornmt - - kurze Beliehtungen alle 3 - -5 Minuten w/~hrend einer Stunde odor mehr), zu folgenden l~esultaten fiihren. Es werden sich bei jeder Belichtung Zellen vorfinden; die in die zweite Gruppe einrficken, und daher zur vorzeitigen Teilung angeregt werden. Diese Verschiebung der zeitlichen Verh~ltnisse wird sich indessen nur in dem Falle als positiver

752 A.u.L. Gurwitseh: Energetik d. mitogen. Induktion u. Zellteilungsreakt.
Induktionseffekt widerspiegeln, weIm die betreffenden Zellen im Augen- blicke der Fixierung in Frfihphasen angetroffen werden, Jhre Partner an der andern Seite daher noch als Ruhezcllcn angesprochen werden. Sonst wird diese zeitliehe Verschiebung unbemerkt bleiben, da wir ja den Induktionseffekt nicht nach einzelnen Stadien, sondern global beur- teilen. Durch die wiederholte Belichtung werden aber jedesmal Zellen der verschiedenen Gruppen der hSheren Nummern betroffen, die den angereizherten Stoff ,,vers~rahlen" werden, und daher im Sinne recht- zeitiger Mitosen wegfallen. Setzt man die Latenzzeit der Teilungsreak- tion auf 2 Stunden und wird 3 Stunden nach Beginn der Belichtung fixiert, so wird der Gesamteffekt der ersten Belichtungsstunde sich schon als ein Defizit merkbar machen, die weitere Belichtung ist dagegen fiir den Ausgang des Versuches irrelevant. Sofern eine protrahierte Belich- tung nicht dirckt sch~dlich wirkt, z. B. nicht etwa den Ablauf der Mitose hemmt, und sich nur im Sinne der vorzeitigen , ,Entladung" der Zellen ~ul~ert, k6nnen wir aus dem Vorangehenden folgende allgemeine Si~tze ableiten: Bei protrahierter Belichtung ist im Sinne der positiven Beein- flussung, d. h. eines Pluseffektes nur eine relativ kurze Zeitspanne von Belang, deren AbschluI3 auf etwa 2 Stunden vor Abschlul~ des Ver- suches, bzw. Fixierung fs deren Dauer der Gr51]e der zeitlichen Ver- schiebung der Mitosen unter dem Einflusse der Induktion gleichkommt. Die ganzc angewendete Belichtungsdauer ist dagegen nur im Sinne einer Erschdpfung der Wurzel durch vorzeitige , ,Entladung" der Zellen, die noch nicht teilmlgsreif sind, wirksam.
Da wir hier yon unbegrenzter Belichttmgsdauer sprechen, haben wir nur die Intensit~ten der verschiedenen Quellen zu beriicksichtigen, und hier ergibt es sich aus vorangehenden Erws daG w~thrend die Gr6Be des 19ositiven Induktionseffektes v o n d e r Intensit~t einer Quelle unabh~ngig ist, der Zeitpunkt des ersten Auftretens des Defizits (d. h. der Ersch6pfung der Wurzel durch protruhierte Induktion) mit derselben zusammenh~ngt, da es durch vorzeitige Entladung der noch nicht tei- lungsreifen ZeUen entsteht, und letzterc in ihrer Ausdehnung, laut V o r - aussetzung yon der Intensits der Belichtung abh~ngt. Unsere Be- t rachtungen fiihren demnach zur Ableitung ganz bestimmter Erwar- tungen, die in der nachfolgenden Arbeit yon Frl. SCSSMA~OWITSCH cx- perimentell gepriift werden sollen.





























