Embed Size (px)
Citation preview

Aus dem Institut für Anatomie und Zellbiologie
(Direktor: Prof. Dr. med. Karlhans Endlich)
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
Zur Pneumatisation des Processus alveolaris der Anthropoidea
– eine biometrische Studie –
Inauguraldissertation
zur
Erlangung des akademischen Grades
Doktor der Zahnmedizin
(Dr. med. dent.)
der Universitätsmedizin der Ernst-Moritz-Arndt-Universität Greifswald
2014
vorgelegt von
Romana Marion Krapf
geb. 10.01.1985
in Neu-Ulm

Dekan: Prof. Dr. med. dent. Reiner Biffar
1. Gutachter: Prof. Dr. Thomas Koppe
2. Gutachter: Prof. Dr. Jens Christoph Türp
Ort, Raum: Greifswald, Hörsaal der HNO-Klinik, W.-Rathenau-Str. 43 - 45
Tag der Disputation: 27. April 2015

Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung und Fragestellung ........................................................................1
2 Literaturübersicht ..........................................................................................3
2.1 Taxonomie der Primaten ....................................................................3
2.2 Vorkommen und Verteilung der Nasennebenhöhlen bei
Primaten ............................................................................................8
2.3 Anatomie der Kieferhöhle ..................................................................9
2.4 Funktionen der Nasennebenhöhlen ................................................. 13
2.5 Odontogene Sinusitis maxillaris ....................................................... 16
2.6 Klinische Bedeutung der Kieferhöhlen in der dentalen
Implantologie.................................................................................... 19
3 Material und Methoden ............................................................................... 23
3.1 Material ............................................................................................ 23
3.2 Methoden ......................................................................................... 24
3.2.1 Metrische Schädelvermessung ............................................. 24
3.2.2 Computertomographische Vermessung des
Recessus alveolaris .............................................................. 28
3.2.3 Biostatistische Auswertung .................................................... 32
4 Ergebnisse .................................................................................................. 36
4.1 Morphologie des Recessus alveolaris .............................................. 36
4.2 Ergebnisse der Korrelationsanalyse ................................................ 54
4.4 Ergebnisse der Regressionsanalyse ................................................ 60
5 Diskussion .................................................................................................. 70
6 Zusammenfassung ..................................................................................... 86
7 Literaturverzeichnis ..................................................................................... 88
8 Anhang ...........................................................................................................

Einleitung und Fragestellung
1
1 Einleitung und Fragestellung
Der Recessus alveolaris ist einer von vier Recessus, welcher den Sinus
maxillaris in seiner Größe erweitern kann (Fanghänel et al. 2003). Er
pneumatisiert den Processus alveolaris der Maxilla und hat dadurch häufig
engen Kontakt zu den Oberkieferseitenzähnen (Schumacher 1971). Dies ist für
verschiedene Disziplinen der Zahnmedizin von klinischer Bedeutung, da unter
anderem bei der Extraktion, einer endodontischen Behandlung oder bei der
Insertion eines enossalen Implantates das Risiko einer iatrogenen Verbindung
zur Kieferhöhle besteht (Reichert 2009). Interdisziplinär spielt dieses Wissen um
die anatomische Nähe des Recessus alveolaris und der Oberkieferseitenzähne
für die Hals-Nasen-Ohren-Heilkunde auch eine bedeutende Rolle, da in 40,6%
der Sinusitiden eine dentogene Ursache zugrunde liegt (Melén et al. 1986). In
den letzten Jahren haben sich verschiedene Arbeiten mit Hilfe von
Knochendickemessungen zwischen dem Recessus alveolaris und den
Wurzelspitzen der Oberkieferseitenzähne dieser Problematik zugewandt
(Eberhardt et al. 1992; Nogi and Kanazawa 2001; Kwak et al. 2004).
Der Recessus alveolaris wurde nicht nur beim Menschen, sondern auch bei
Neuweltaffen und verschiedenen Altweltaffen beschrieben (Cave and Haines
1940; Ward and Pilbeam 1983; Koppe and Ohkawa 1999; Koppe and Rae 2001;
Nishimura et al. 2005; Rossie 2006). Intra- und interspezifische Unterschiede
zeigen sich sowohl beim Ausmaß als auch in der Morphologie des Recessus
alveolaris maxillae.
Die Faktoren und die Gesetzmäßigkeiten, welche die Pneumatisation des
Processus alveolaris beeinflussen, sind jedoch weitgehend unbekannt. Obgleich
Studien zur Tiefe des Recessus alveolaris darauf hindeuten, dass bei Makaken
(Koppe and Rae 2001) und Homo sapiens (Koppe et al. 2005b) ein
Zusammenhang zwischen der Gaumenmorphologie und der Größe des
Recessus alveolaris besteht, ist die Datenlage insbesondere anhand
dreidimensionaler Volumenvermessungen rar. Mit Ausnahme einer Studie von

Einleitung und Fragestellung
2
Oesterreich et al. (2005), die das Volumen des Recessus alveolaris an Schädel
von Neuweltaffen analysierten, existieren keine weiteren Daten über das
Volumen des Recessus alveolaris.
Solche Daten über rezente Primaten und den Menschen können von großer
Bedeutung für das Verständnis des Recessus alveolaris und solcher Faktoren
sein, die seine Größe und sein Ausmaß beeinflussen. Vor diesem Hintergrund
befasst sich die vorliegende Arbeit mit der Vermessung des Volumens des
Recessus alveolaris anhand dreidimensionaler CT-Bilder verschiedener Spezies
von Alt- und Neuweltaffen.
Im Einzelnen verfolgt die vorliegende Arbeit folgende Zielstellungen:
1. Vermessung des Volumens des Recessus alveolaris und seine
morphologische Beschreibung bei verschiedenen Spezies der Altweltaffen
und Neuweltaffen anhand von computertomographischen
Untersuchungen
2. Vermessung der tiefsten Stelle des Recessus alveolaris auf Höhe
verschiedener Zahnregionen
3. Untersuchung der Asymmetrie von rechtem und linkem Recessus
alveolaris
4. Prüfung statistischer Beziehungen zwischen Recessus alveolaris, Sinus
maxillaris und ausgewählten Schädelmaßen bei Neuweltaffen und
Altweltaffen
5. Darstellung der Bedeutung der vorliegenden Studie

Literaturübersicht
3
2 Literaturübersicht
2.1 Taxonomie der Primaten
Primaten lassen sich unter anderem über anatomisch-morphologische Merkmale
definieren. Einige dieser Merkmale sind Geissmann (2003) zufolge greiffähige
Extremitäten, flache Nägel an Fingern und Zehen und ein ausgeprägter
Gesichtssinn. Die rezenten Primaten werden in die Unterordnung der Haplorrhini
(Trockennasenaffen) und der Strepsirrhini (Feuchtnasenaffen) unterteilt (Abb. 1).
Neben weiteren Unterscheidungsmerkmalen der beiden Gruppen finden sich
zwei solcher bei der Betrachtung der Nase und des Auges (Geissmann 2003).
Strepsirrhini besitzen ein Philtrum und einen Nasenspiegel (Rhinarium). Dabei
handelt es sich um einen feuchten und drüsenreichen Bereich um die
Nasenlöcher, welcher bei den Trockennasenprimaten fehlt (Geissmann 2003;
Fleagle 2013). Ein weiteres Unterscheidungsmerkmal zwischen Strepsirrhini und
Haplorrhini ist die knöcherne Orbita. Während die Orbita der Strepsirrhini bis auf
einen Orbitaring nicht geschlossen ist, liegt bei haplorrhinen Primaten ein
Orbitatrichter vor, welcher durch ein zusätzliches postorbitales Septum das Auge
nach dorsal einfasst (Geissmann 2003; Fleagle 2013). Dadurch wird vermutlich
gewährleistet, dass bei Kontraktion der Kaumuskulatur das Auge nicht
beeinträchtigt wird (Geissmann 2003).
Die Haplorrhini werden in die Gruppen der Tarsiforme (Koboldmakis) und der
Antropoideae (eigentliche Affen) gegliedert. Letztgenannte teilten sich vermutlich
vor ca. 40 Millionen Jahren in die Platyrrhini (Neuweltaffen) und die Catarrhini
(Altweltaffen) (Goodman et al. 1998) (Abb. 1). Catarrhini unterscheiden sich von
den Platyrrhini unter anderem durch einen knöchernen äußeren Gehörgang,
eine schmale Nase mit nach vorn unten geöffneten Nasenlöchern, opponierbare
Daumen und eine spezifische Zahnformel (Geissmann 2003). Im Vergleich zur
ursprünglichen Zahnformel der Säugetiere von 3143/3143 ist bei allen Primaten
ein Inzisivus verloren gegangen. Darüber hinaus hat sich bei den Neuweltaffen
die Zahl der Prämolaren um einen Prämolar und bei den Catarrhini um zwei

Literaturübersicht
4
Prämolaren reduziert. Demzufolge lautet die Zahnformel der Neuweltaffen
2133/2133 und die der Catarrhini 2123/2123 (Geissmann 2003). Eine
vereinfachte Übersicht der Hauptverzweigungslinien der Primaten zeigt
Abbildung 1.
Abb. 1: Schematische Darstellung der Hauptverzweigungslinien der rezenten
Primaten.
Bei den Catarrhini kam es vor ca. 25 Mio. Jahren zu einer Teilung in die
Cercopithecoidea und Hominoidea (Goodman et al. 1998). Hominoidea werden
wiederum in Hylobatidae (Gibbons) und die Hominidae eingeteilt (Geissmann
2003).
Über die Unterteilung der Hominidae in Familien und Subfamilien gibt es
unterschiedliche Ansichten, welche sich aufgrund verschiedener
Klassifikationskonzepte ergeben. Dabei wird die traditionelle von der
phylogenetischen Klassifikation unterschieden (Abb. 2) (Ax 1984). Die
traditionelle Einteilung beruht auf anatomisch-morphologischen Merkmalen, nach
denen der Mensch als einziger Vertreter der Subfamilie der Hominidae
zugeordnet wird (Abb. 2a). Die großen Menschenaffen Gorilla, Schimpanse und
Orang-Utan sind Teil der Subfamilie Pongidae (Abb. 2a) (u.a. Simpson 1961;
Ankel 1970). Nach der phylogenetischen Systematik erfolgt die Einteilung

Literaturübersicht
5
aufgrund molekularer Untersuchungen. Dabei werden alle rezenten
Menschenaffen der Familie Hominidae zugeordnet, welche sich in die
Unterfamilien der Ponginae und Homininae teilt. Letztere beinhaltet danach die
afrikanischen Menschenaffen und die Gattung Homo; zu den Ponginae wird als
einziger Vertreter der Orang-Utan gezählt (Goodman et al. 1998; Wildmann et al.
2003). Dieser hat am wenigsten genetische Gemeinsamkeiten zur Gruppe der
Homininae (Abb. 2b) (Miyamoto et al. 1988; Wildmann et al. 2003).
Abb. 2: Zwei mögliche Klassifikationen von Menschenaffen und Mensch nach
der traditionellen (a) und phylogenetischen (b) Einteilung (aus Ax (1984) mit
freundlicher Genehmigung des © Gustav Fischer Verlags).
Goodman et al. (1998) und Wildmann et al. (2003) sprechen sich sogar dafür
aus, die Gattung Homo in drei Spezies zu unterteilen, um die
Verwandtschaftsverhältnisse noch genauer zu betonen: den Mensch als Homo
(Homo) sapiens, den Schimpansen als Homo (Pan) troglodytes und den Bonobo
als Homo (Pan) paniscus.
Die Einteilung der Neuweltaffen ist bis heute umstritten. Unterschiedliche
analytische Methoden wurden vorgenommen, um die

Literaturübersicht
6
Verwandtschaftsverhältnisse innerhalb der Neuweltaffen zu analysieren. Dazu
dienten zum einen makroskopisch-anatomische Merkmale, zum anderen
molekular-genetische, biochemische und zytogenetische Charakteristika.
Eine Einteilung der Neuweltaffen nach makroskopisch-anatomischen Merkmalen
wurde unter anderem von Ford (1986) und Rosenberger et al. (1990)
vorgenommen, welche auf Basis von Zahnmorphologien, kranialen und
postkranialen Parametern beruhen. Während Rosenberger et al. (1990) die
Platyrrhini in zwei Familien (Cebidae und Atelidae) unterteilen, nimmt Ford
(1986) eine Einteilung in drei Familien (Callitrichidae, Cebidae und Atelidae) vor.
Neuere molekulare und biochemische Untersuchungen führten zur
Unterscheidung von drei monophyletischen Gruppen der Neuweltaffen: Cebidae,
Pithecidae und Atelidae (Schneider et al. 1993; Goodman et al. 1998; Dornum
and Ruvolo 1999; Schneider et al. 2001; Steiper and Ruvolo 2003; Wildmann et
al. 2009). Schneider et al. (1996) und Goodman et al. (1998) bezeichnen
Cebidae, Pithecidae und Atelidae nicht als Gruppen, sondern als Familien. Dabei
repräsentiert die Familie Cebidae die Callitrichinae sowie Cebus, Saimiri und
Aoutus. Zur Familie der Pithecidae zählen Callicebus, Pithecia, Cacajao und
Chiropotes. Der Familie der Atelidae gehören Brachyteles, Lagothrix, Ateles und
Alouatta an. Über die genauen Verwandtschaftsverhältnisse zwischen und
innerhalb der drei Familien besteht in der Literatur keine Übereinstimmung.
Insbesondere die Beziehungen innerhalb der Atelinae, der Callitrichinae und der
Gattungen von Saimiri, Cebus, Aoutus sind nach wie vor ungeklärt (z.B.
Schneider et al. 2001). Schneider et al. (2001) führte eine vergleichende Analyse
zu vier verschiedenen DNA-Datensätzen für Neuweltaffen durch. Abbildung 3
informiert über die Phylogenie der Neuweltaffen basierend auf den Ergebnissen
der von Schneider et al. (2001) durchgeführten Analyse.

Literaturübersicht
7
Abb. 3: Stammbaum der Neuweltaffen basierend auf DNA-Datenanalysen.
(verändert nach Schneider et al. (2001) mit freundlicher Genehmigung des
Autors und des Springer Verlags ©).

Literaturübersicht
8
2.2 Vorkommen und Verteilung der Nasennebenhöhlen bei Primaten
Nasennebenhöhlen sind bei fast allen Primaten nachweisbar. Die beim
Menschen beschriebenen Nasennebenhöhlen (Sinus ethmoidales, Sinus
frontalis, Sinus sphenoidalis und Sinus maxillaris) treten bei anderen Primaten in
unterschiedlicher Häufigkeit auf. Darüber hinaus bestehen bezüglich der
Homologie dieser Nasennebenhöhlen bei den Primaten unterschiedliche
Auffassungen. Lediglich der Sinus maxillaris wird übereinstimmend als
homologes Merkmal angesehen (Moore 1981). Ein kurzer Überblick über das
Vorkommen und die Verteilung von Nasennebenhöhlen bei Altweltaffen und
Neuweltaffen soll an dieser Stelle gegeben werden.
Unter den Altweltaffen besitzen sowohl die afrikanischen Menschenaffen als
auch der Mensch vier paarige Nasennebenhöhlen (Sinus frontalis, Cellulae
ethmoidales, Sinus sphenoidalis und Sinus maxillaris). Sinus frontalis und
Cellulae ethmoidales fehlen dagegen bei Gibbons und Orang-Utans (Cave and
Haines 1940). Von einigen Autoren werden vereinzelte pneumatisierte Räume
innerhalb des Siebbeins der Gibbons beschrieben (Koppe and Ohkawa 1999;
Rae and Koppe 2004). Unter den Nasennebenhöhlen der Altweltaffen nehmen
die Kieferhöhlen den größten Volumenanteil ein (Koppe und Schumacher 1992)
und zeigen bei männlichen Tieren ein größeres Volumen als bei weiblichen
(Koppe and Ohkawa 1999). Eine Ausnahme stellen die geschwänzten Affen
(Cercopithecoidea) dar, welche keine pneumatisierten Räume aufzeigen.
Innerhalb der Cercopithecoidea ist lediglich bei Makaken eine Kieferhöhle
nachweisbar (Lund 1988; Koppe and Ohkawa 1999).
Über die Existenz von Nasennebenhöhlen bei Neuweltaffen herrscht in der
Literatur kein Konsens. Innerhalb der Gattungen und auch innerhalb der Spezies
ist das Vorkommen von Nasennebenhöhlen sehr variabel, weshalb in der
Literatur kontroverse Angaben zu finden sind. Insbesondere die Frage, ob ein
pneumatisierter Raum eine eigenständige Nasennebenhöhle darstellt oder
lediglich eine Ausdehnung einer Nasennebenhöhle in einen anderen Knochen

Literaturübersicht
9
ist, in dem eine Nasennebenhöhle zu erwarten ist, wird unterschiedlich
beantwortet.
Der Sinus sphenoidalis ist nach Hershkovitz (1977) bei Cebus, Lagothrix,
Brachyteles, Alouatta, Callimico und Leontopithecus eine eigenständige Höhle.
Cave (1967) hingegen sieht diesen pneumatisierten Raum bei Alouatta als eine
Ausdehnung des Sinus maxillaris in das Os sphenoidale. Dem stimmen auch
Koppe et al. (2005a) zu.
Bei allen Platyrrhini ist nach Hershkovitz (1977) ein Sinus frontalis vorhanden.
Schiwal (2005) kann bei Callithrix jacchus weder Sinus frontalis noch
Siebbeinzellen nachweisen. Im Gegensatz dazu beschreibt Rossie (2006) das
Vorkommen von Siebbeinzellen bei Cebus, Callimico, Lagothrix und Alouatta,
Aoutus und Sanguinus.
Eine Kieferhöhle findet sich bei den meisten Neuweltaffen (Cave 1967;
Oesterreich et al. 2005). Lund (1988) sieht allerdings bei einzelnen Spezies der
Gattung Sanguinus, Callithrix, Saimiri, Chiropotes, Cacajo und Ateles keine
eindeutige pneumatisierte Höhle, sondern lediglich weite Spongiosaräume
zwischen Orbita und Zahnwurzeln des Oberkiefers.
2.3 Anatomie der Kieferhöhle
Anatomie der menschlichen Kieferhöhle
Der Sinus maxillaris ist eine typische Bildung des Schädels der Säugetiere und
der meisten Primaten (Moore 1981). Bei den Hominoiden, einschließlich des
Menschen, weist er das größte Volumen aller Nasennebenhöhlen auf (Koppe
and Ohkawa 1999). Das Volumen des Sinus maxillaris des Menschen wird beim
Erwachsenen mit 14,71cm³ (Ariji et al. 1994) bzw. 18cm³ (Emirzeoglu et al.
2007) angegeben. Die Kieferhöhle ist bereits beim Neugeborenen in einer
ovalen Form vorhanden und nimmt von der Geburt an bis zum vollständigen
Durchbruch der zweiten Dentition stetig an Größe zu (Fanghänel et al. 2003).

Literaturübersicht
10
Ein beschleunigtes Wachstum tritt während der Pubertät ein und bereits
zwischen 12 und 15 Jahren erreicht die Kieferhöhle ihre maximale Größe
(Schumacher 1971). Eine weitere Ausformung dauert bis zum 18. Lebensjahr an
(Schumacher 1971). Ab dem 20. Lebensjahr soll die Kieferhöhle wieder an
Größe verlieren (Ariji et al. 1994).
Die Anatomie der Kieferhöhle wird ausführlich von Fanghänel et al. (2003)
beschrieben, welche im Folgenden genauer dargestellt wird. Die Form des Sinus
maxillaris entwickelt sich von einer ovalen Form bei der Geburt zu einer drei-
oder vierseitigen Pyramide im Erwachsenenalter. Die Spitze der Pyramide zeigt
zum Processus zygomaticus der Maxilla, die Basis der Pyramide ist in Richtung
laterale Nasenwand gerichtet. Letztere bildet zugleich die mediale Begrenzung
der Kieferhöhle. Eine Verbindung zum mittleren Nasengang besteht über das
Infundibulum ethmoidale, einer schmalen Rille, in welches die Sinus maxillaris
und frontalis und die Cellulae ethmoidales anteriores münden. Diese
halbmondförmige Öffnung des Sinus maxillaris, der Hiatus semilunaris, liegt über
dem Boden der Kieferhöhle. Dies spielt bei der Entstehung von pathologischen
Prozessen eine wichtige Rolle, denn durch seine anatomisch ungünstige Lage
kann er der Funktion des Sekretabflusses bei pathologischen Prozessen oftmals
nicht gerecht werden (Fanghänel et al. 2003).
Fanghänel et al. (2003) beschreiben weiter, dass das Dach des Sinus maxillaris
dem Orbitaboden entspricht, in welchem sich der Canalis infraorbitalis befindet.
Letzterer mündet mit dem Foramen infraorbitale an der fazialen Wand der
Maxilla. Die Hinterwand der Kieferhöhle wird vom Tuber maxillae gebildet und
grenzt an die Fossa pterygopalatina (Schumacher 1971). Die Vorderwand der
Kieferhöhle weist meist eine charakteristische Einbuchtung, die Fossa canina auf
und wird von der Facies anterior der Maxilla begrenzt (Fanghänel et al. 2003).
Der Boden des Sinus maxillaris reicht unterschiedlich weit in den Processus
alveolaris der Maxilla (Fanghänel et al. 2003).

Literaturübersicht
11
Ausgehend vom Boden oder der lateralen Wand der Kieferhöhle können
knöcherne Septen, auch Underwood-Septen genannt, in die Kieferhöhle ragen
und diese partiell oder komplett unterteilen (Underwood 1910). Die Prävalenz
von Septen in Bezug auf die Anzahl der Kieferhöhlen wird mit 13–35,9%
angegeben (Maestre-Férrin et al. 2010). Krennmair et al. (1995) und Kim et al.
(2006) berichten über eine größere Prävalenz von Septen bei zahnlosen
Patienten mit atrophischem Kiefer im Gegensatz zu Patienten mit teilbezahntem,
nicht-atrophischem Oberkiefer. Des Weiteren kann sich der Sinus maxillaris
durch die Ausbuchtungen Recessus frontalis, zygomaticus, palatinus und
alveolaris in die umliegenden Knochen ausdehnen (Schumacher 1971).
Der Kieferhöhlenboden entspricht meist dem Recessus alveolaris. Er kann im
Bezug zum Nasenboden auf unterschiedlichem Niveau liegen. Während
Schumacher (1971) den Boden der Kieferhöhle in der Regel auf einer Höhe mit
dem Nasenboden und bei Menschen mit niedrigem Mittelgesicht sogar oberhalb
des Nasenbodens beschreibt, berichten Koppe et al. (2005b), dass sich der
Recessus alveolaris bei den von ihnen untersuchten japanischen Schädeln und
der Jomon-Population, einer archäologischen Population Japans, in der Regel
unterhalb des Nasenhöhlenbodens befindet. Dadurch hat der Recessus
alveolaris häufig enge topographische Beziehung zu den Wurzelspitzen der
Oberkieferseitenzähne. Nimigean et al. (2008) berichten, dass die Knochendicke
von der Eckzahnregion (6,9mm) zum zweiten Molaren bis zu 1,7mm abnimmt
und zum dritten Molaren hin wieder etwas dicker wird. Dass die geringste
Distanz zwischen Kieferhöhlenboden und zweitem Molaren besteht, beschreiben
auch Eberhardt et al. (1992) und Kwak et al. (2004). Durch die enge Beziehung
zwischen Recessus alveolaris und den ersten und zweiten Molaren sind
Unebenheiten im Kieferhöhlenboden keine Seltenheit (u.a. Koppe et al. 2005b;
Nimigean et al. 2008).

Literaturübersicht
12
Anatomie der Kieferhöhle bei Altweltaffen
Die Morphologie der Kieferhöhle und deren Kommunikation zu den anderen
Nasennebenhöhlen weist bei Gorillas und Schimpansen große Ähnlichkeiten mit
dem Menschen auf (Lund 1988). Ein Unterschied zwischen Mensch und Gorilla
ist die Bulla nasolacrimalis, ein stark erweiterter Canalis nasolacrimalis, welche
die Kieferhöhle beim Gorilla nach medial begrenzt (Cave and Haines 1940;
Koppe and Ohkawa 1999). Nach distal wölbt sich die Kieferhöhle des Gorillas
weit in das Os sphenoidale und reicht bis an die anteriore Wand des Sinus
sphenoidalis. Zudem pneumatisiert der Sinus maxillaris des Gorillas auch Teile
des Gaumens und des Ethmoids (Cave and Haines 1940). Für den
Schimpansen ist ein Recessus palatinus charakteristisch, welcher so weit in den
harten Gaumen extendieren kann, dass die beiden benachbarten Recessus
palatini nur noch durch eine dünne sagittale Knochenwand voneinander getrennt
werden (Cave and Haines 1940; Lund 1988; Koppe and Nagai 1998).
Die Kieferhöhlen des Orang-Utans extendieren massiv in alle umliegenden
Knochen, ausgenommen der Praemaxilla (Cave and Haines 1940). Eine direkte
Verbindung von Sinus maxillaris in den mittleren Nasengang ohne Hiatus
semilunaris ist beim Orang-Utan typisch (Cave and Haines 1940).
Die Kieferhöhle des Gibbons breitet sich lediglich innerhalb des Oberkiefers aus
und erstreckt sich vom zweiten Prämolaren bis zum Tuber maxillae (Klug 2011).
Das Volumen der Kieferhöhle zeigt unter den Altweltaffen große Variationen.
Während der Gibbon das kleinste Kieferhöhlenvolumen mit durchschnittlich
2,85cm³ hat, weist der Gorilla mit 38,45cm³ das größte Volumen auf (Koppe et
al. 1999b).
Anatomie der Kieferhöhle bei Neuweltaffen
Bei Alouatta, Callicebus, Cebus und Lagothrix erstreckt sich die Kieferhöhle
zwischen Prämolaren und Molaren und der Orbita. Die seitliche Begrenzung der
Kieferhöhle liegt lateral der Zahnwurzeln bei Callicebus (Schiwal 2005) und

Literaturübersicht
13
Alouatta (Koppe et al. 2005a). Nach dorsal reicht die Kieferhöhle bis an den
Tuber maxillae, ausgenommen bei Sanguinus, Callimico und Aoutus. Bei diesen
Gattungen liegt die dorsale Begrenzung mesial des letzten Molaren (Rossie
2006). Der Boden der Kieferhöhle liegt unter dem Niveau des Nasenbodens bei
Alouatta, Cebus, Lagothrix, Ateles, Brachyteles und Callicebus und wird somit
als Recessus alveolaris bezeichnet (Oesterreich et al. 2005; Nishimura et al.
2005). Der Boden der Kieferhöhle ist meist durch die Zahnwurzeln aufgewölbt
(Nishimura et al. 2005; Rossie 2006).
Die Gattung Alouatta zeigt im Vergleich zu anderen Spezies der Plattyrhini das
größte Kieferhöhlenvolumen (Koppe et al. 1999b; Oesterreich et al. 2005;
Nishimura et al. 2005). Koppe et al. (2005a) berichten über einen signifikanten
Sexualdimorphismus bei Alouatta. So haben männliche Brüllaffen mit
durchschnittlich 4,08cm³ ein doppelt so großes Kieferhöhlenvolumen wie die
weiblichen Tiere mit 2,00cm³. Eine Erweiterung des Sinus maxillaris in das Os
sphenoidale bei Alouatta beschreiben Cave (1967) und Koppe et al. (2005a). Im
Gegensatz dazu betrachtet Hershkovitz (1977) diesen pneumatisierten Raum als
eigenständigen Sinus sphenoidales unter anderem bei den Gattungen von
Cebus, Lagothrix, Brachyteles, Alouatta, Callimico und Leontopithecus.
2.4 Funktionen der Nasennebenhöhlen
In der Literatur finden sich zahlreiche und zum Teil widersprüchliche
Informationen über die Funktion der Kieferhöhle (Blanton and Biggs 1969;
Blaney 1990; Rae and Koppe 2004; Keir 2008; Márquez 2008). In einer
Übersichtsarbeit diskutieren Blanton and Biggs (1969) folgende Funktionen der
Nasennebenhöhlen:

Literaturübersicht
14
1. Stimmresonanzkörper
2. Öberflächenvergrößerung der Riechschleimhaut zur Verbesserung des
Riechvermögens
3. Schleimsekretion zum Feuchthalten der Nasenhöhle
4. Stoßdämpfer für intrakranielle Organe bei Verletzungen und Traumata
5. Thermische Isolation des zentralen Nervensystems
6. Unterstützung bei Gesichtsschädelwachstum und -architektur
7. Gewichtsreduktion des Schädels
8. Überrest aus evolutionärer Sicht bzw. ein unerwünschter luftgefüllter
Raum
Blaney (1990) ordnet die zahlreichen Theorien drei großen Hauptgruppen zu:
(1) Nasennebenhöhlen haben eine strukturelle Funktion zur Verbesserung der
Schädelarchitektur, (2) Nasennebenhöhlen existieren aus physiologischen
Gründen und (3) Nasennebenhöhlen sind funktionslose Räume bzw.
evolutionäre Überreste.
Strukturelle Funktion
Hierbei werden Nasennebenhöhlen als unterstützende Strukturen des
Gesichtsschädelwachstums und der -architektur angesehen. Sie sollen das
Schädelgewicht reduzieren und als Stoßdämpfer die intrakraniellen
Weichteilstrukturen schützen (Blanton and Biggs 1969). Bereits Weidenreich
(1924) nahm an, dass Nasennebenhöhlen an denjenigen Regionen des
Schädels entstehen, die von geringen statischen und mechanischen
Belastungen beansprucht werden. Preuschoft et al. (2002) beschreiben die
Nebenhöhlen des Schädels als dünnwandige Schalenkonstruktionen, die trotz
geringen Knochenaufwands eine gute mechanische Belastung und Tragekraft
ermöglichen. Die honigwabenartige Struktur der pneumatisierten Knochen und

Literaturübersicht
15
der Maxilla können mechanische Kräfte abfangen und auf andere Knochen leiten
(Preuschoft et al. 2002). Dass die Schädelstruktur durch die Nasennebenhöhlen
optimiert wird, lassen Untersuchungen an Altweltaffen und Neuweltaffen
vermuten, die auf eine allometrische Beziehung zwischen Schädelgröße und
Kieferhöhlengröße hinweisen (Lund 1988; Koppe et al. 1999b, 2005a). Einen
Beweis für die alleinige strukturelle bzw. gewichtssparende Funktion der
Nasennebenhöhlen sehen Koppe et al. (1999b) darin nicht. Außerdem soll der
Gewichtsverlust durch die pneumatisierten Räume bei gerade mal 1% liegen
(Braune und Clasen 1877).
Physiologische Funktion
Dieser Auffassung zufolge werden den Nasennebenhöhlen physiologische und
funktionelle Aufgaben zugeschrieben. Dazu gehören z.B. die Konditionierung der
Atemluft und die Funktion als Resonanzkörper für die Stimme (Blaney 1990).
Bereits Wegner (1955/56) begründete letztgenannte Funktion damit, dass
Brüllaffen verhältnismäßig große pneumatisierte Räume besitzen und damit
befähigt sind ausgesprochen laute Töne von sich zu geben. Negus (1954)
beobachtete allerdings, dass Giraffen mit großen Stirnhöhlen nur leise Töne
erzeugen. Blanton and Biggs (1969) unterstützen die Theorie der
Nasennebenhöhle als Resonanzkörpers ebenfalls nicht. Die Beteiligung der
Nasennebenhöhlen am Erwärmen und Anfeuchten der Atemluft scheint ebenso
umstritten, da circa fünf Minuten benötigt werden, um 90% des Luftvolumens der
Kieferhöhle bei normaler Nasenatmung auszutauschen (Drettner 1980).
Des Weiteren wird den Nasennebenhöhlen eine Abwehrfunktion zugeschrieben
(Stierna 2001). Die in den Nasennebenhöhlen vorkommenden Becherzellen
sowie muköse und seröse Drüsen sind dabei für die Herstellung von
Plasmaproteinen wie Albuminen, Immunoglobulinen und Komplementfaktoren
sowie Lactoferrin, Lysozymen und Glycoproteinen verantwortlich (Stierna 2001).
Zudem wird von den epithelialen Zellen der Kieferhöhlen Stickstoffmonoxid (NO)
produziert (Lundberg et al. 1995; Stierna 2001; Keir 2008). Stickstoffmonoxid

Literaturübersicht
16
reguliert unter anderem die Schlagfrequenz der Kinozilien der Epithelzellen in
den Nasennebenhöhlen (Lundberg et al. 1995) und stellt somit einen
entscheidenden Faktor für den mukoziliaren Transport und für die mechanische
Reinigungsfunktion eingeatmeter Partikel oder Bakterien dar. Weiterhin hat
Stickstoffmonoxid bakteriostatische (Lundberg et al. 1995) und antivirale
Wirkung (Croen 1993).
Evolutionäre Überreste
Einer weiteren Gruppe von Autoren zufolge haben die Nebenhöhlen keine
Funktion und sind lediglich als evolutionäre Relikte zu betrachten (Negus 1954;
Blanton and Biggs 1969). Weidenreich (1924) bezeichnete sie als „tote Räume“,
die keine weitere Funktion besitzen. Negus (1954) vertritt die Meinung, dass die
Kieferhöhle ein übrig gebliebener Raum ist, der sich aufgrund der Rückbildung
der maxilloturbinalen Region ausgebildet hat.
2.5 Odontogene Sinusitis maxillaris
Die chronische Sinusitis ist in den USA die häufigste diagnostizierte chronische
Erkrankung nach orthopädischen Fehlstellungen (Collins 1997). Obgleich sich
Sinusitiden häufig auf rhinogenen Ursprung zurückführen lassen, ist eine
dentogene Ursache nicht auszuschließen. Der Anteil einer dentogenen Ursache
wird mit 10–12% (Brook 2009) angegeben. Melén et al. (1986) sind sogar der
Auffassung, dass bis 40,6% aller Sinusitiden dentogenen Ursprungs sind.
Ursachen und Pathophysiologie der dentogenen Sinusitis
Die Kieferhöhle kann durch eine Vielzahl dentogener Pathologien infiziert
werden. Zu nennen sind insbesondere Zysten, Tumoren, Traumata,
Fremdkörper, retinierte Zähne sowie Endo- und Parodontopathien (Schmelzle
und Schwenzer 1982; Reichert 2009). Kieferhöhlenentzündungen können
genauso durch Zahnbehandlungen wie implantologische Eingriffe, insbesondere
in Zusammenhang mit Sinusbodenaugmentation, Wurzelkanalbehandlungen mit

Literaturübersicht
17
Überinstrumentierung, überpresstes Wurzelfüllmaterial mit der Folge einer
Aspergillusmykose und Mund-Antrum-Verbindungen nach zahnärztlichen
Extraktionen entstehen (Reinert und Machtens 2002; Reichert 2009). Das
höchste Risiko für eine Mund-Antrum-Verbindung stellt die Extraktion eines
parodontal oder periapikal geschädigten ersten Molaren bei Zahnentfernungen
dar (Kishi et al. 1995).
Als häufigste Ursache einer dentogenen Sinusitis wird eine chronische apikale
Parodontitis des ersten Molaren angesehen (Schmelzle und Schwenzer 1982).
Bedingt durch die Nähe der Wurzeln der Oberkiefermolaren zum
Kieferhöhlenboden beginnt die dentogene Sinusitis meist mit einer lokalen, auf
den Kieferhöhlenboden beschränkten Entzündungsreaktion (Bauer 1943; Melén
1996). Wird die auslösende Ursache und die dentale Infektion in diesem Stadium
behandelt, ist eine schnelle Regeneration der Kieferhöhlenschleimhaut möglich
(Melén 1996). Studien zeigen, dass bereits eine entsprechende konservative
Maßnahme wie unter anderem eine endodontische Therapie (Nurbakhsh et al.
2011) oder Parodontaltherapie (Engström et al. 1988) zu einer Ausheilung der
Kieferhöhlenschleimhaut führen kann. Häufig bleibt jedoch durch den langsamen
Entstehungsprozess einer apikalen oder marginalen Parodontitis die Sinusitis
unbemerkt und damit unbehandelt (Melén 1996). Eine bakterielle Besiedlung
kann zur Exazerbation der Symptome oder gar über viele Jahre zu einem
Kieferhöhlenempyem führen (Melén 1996).
Dentogene Sinusitis bei nicht-menschlichen Primaten
Angaben darüber, ob auch freilebende Primaten an Sinusitiden leiden, gibt es in
der Literatur kaum. Bluestone et al. (2012) vertreten die Auffassung, dass
Erkrankungen wie die Rhinosinusitis bzw. die Otitis media vorwiegend den
Menschen betreffen. Die Einschränkung von Hör- bzw. Riechsinn durch diese
Erkrankungen hätten das Überleben von Primaten in der freien Wildbahn nicht
möglich gemacht, wohingegen der Mensch mit seinem gut ausgeprägten
Sehvermögen, seinem ausgebildeten Gehirn und seinen sozialen Strukturen den

Literaturübersicht
18
Verlust von Hör- bzw. Riechsinn kompensieren kann (Bluestone et al. 2012).
Bluestone et al. (2012) gehen somit davon aus, dass derartige Erkrankungen
eine Konsequenz von menschlicher Evolution, immunologischen Faktoren und
Umweltfaktoren sind.
Dennoch ist anzumerken, dass nicht nur beim Menschen, sondern auch bei
freilebenden Primaten Erkrankungen der Zähne wie Karies, Parodontitiden sowie
periapikale Abszesse auftreten können (Wegner 1962; Kakehashi et al. 1963;
Hall et al. 1967; Kilgore 1989; Lovell 1990; Stoner 1995). Während Karies bei
Gorillas, Schimpansen und Orang-Utans selten auftreten, sind parodontale
Läsionen und Abszesse häufiger zu finden. Kakehashi et al. (1963) beobachten,
dass bei 76,4% der von ihnen untersuchten 292 Gorillaschädel parodontale
Läsionen aufzeigen. Kilgore (1989) stellte beim Schimpansen fest, dass
parodontale Läsionen, Abszesse und vorzeitiger Zahnverlust im höheren
Lebensalter ab Mitte 30 auftreten. Prämolaren und Molaren waren am häufigsten
von Parodontitiden betroffen, während Abszesse vor allem an den Eckzähnen
auftraten (Kilgore 1989).
Klinik und Diagnose der odontogenen Sinusitis maxillaris
Bei Verdacht einer Sinusitis dentogenen Ursprungs lassen die genaue
anamnestische Befragung und die klinische Untersuchung des Patienten
insbesondere bei der akuten Form der dentogenen Sinusitis maxillaris einen
Rückschluss auf die Erkrankung zu. Häufige Symptome, welche individuell
unterschiedlich ausgeprägt sein können, sind unter anderem diffuse
Zahnschmerzen mit Aufbiss- und Perkussionsempfindlichkeit, einseitiger
Schnupfen, eine druckdolente Schwellung der entsprechenden Wange, diffuse
Gesichtsschmerzen in der Region des betroffenen Oberkiefers mit Ausstrahlung
in Richtung Stirn oder Zähne mit einer möglichen Verstärkung beim Bücken
sowie allgemeine Abgeschlagenheit und Fieber mit erhöhten
Entzündungsparametern im Blutbild (u.a. Reinert und Machtens 2002; Reichert
2009). Für die chronische Sinusitis sind die klinischen Symptome eher

Literaturübersicht
19
unspezifisch und können dabei als dumpfe Kopfschmerzen, einseitige Rhinitis
und Hyposmie auftreten (Reinert und Machtens 2002).
Neben einer ausführlichen Anamnese ist eine ausgiebige klinische
Untersuchung zur Diagnose einer Sinusitis maxillaris unabdingbar. Die
Beurteilung, Inspektion und Palpation von Gesichtsasymmetrien, Schwellungen,
Schmerzen und Rötung der Regio maxillaris, intra- und extraoraler Schleimhaut,
Zähnen, Alveolarknochen und Parodont sind durchzuführen (Reichert 2009). Die
Erhebung eines Zahnstatus inklusive Veränderungen der Zahnfarbe, Prüfung der
Sensibilität, Taschentiefenmessung sowie Perkussions- und Palpationstests ist
nicht weniger wichtig als ein Nasen-Blas-Versuch bzw. ein stumpfer Sondentest
mit dem Hintergrund einer möglichen Mund-Antrum-Verbindung (Schmelzle und
Schwenzer 1982; Reichert 2009). Röntgenbilder in Form von
Panoramaschichtaufnahmen sind als Basisdiagnostik geeignet, wobei
Übersichtsaufnahmen mit okzipitofrontalem (Caldwell) und okzipitomentalem
(Waters) Strahlengang zusätzlich notwendig sind (Reichert 2009). Weitere
Maßnahmen, um eine Diagnose zu stellen, sind unter anderem die anteriore
Rhinoskopie, eine Endoskopie und/oder mikrobiologische und zytologische Tests
(Reinert und Machtens 2002; Reichert 2009). Für operative Eingriffe in die
Nasennebenhöhlen ist insbesondere zur Abschätzung von Komplikationen die
Anfertigung eines präoperativen CT-Bildes unabdingbar (Stuck et al. 2007).
2.6 Klinische Bedeutung der Kieferhöhlen in der dentalen Implantologie
Die Morphologie der Kieferhöhle ist für die enossale Implantologie von großer
Bedeutung. Insbesondere im Oberkieferseitenzahnbereich stellt häufig eine
unzureichende Knochendicke des Processus alveolaris zwischen
Kieferhöhlenboden und der Mundhöhle eine Limitation für das Einsetzen von
schraubenförmigen Implantaten dar.
Nach Zahnverlust ist eine Inaktivitätsatrophie des Alveolarkammknochens
sowohl in der Horizontalen als auch in der Vertikalen die Folge (Cawood and

Literaturübersicht
20
Howell 1988). Die im Rahmen der Atrophie zunehmende Pneumatisation der
Kieferhöhlen reduziert das Knochenangebot im Oberkiefer in zunehmendem
Maße (Cawood and Howell 1988). Allerdings ist eine genügende Schichtstärke
des Alveolarfortsatzes ein maßgeblicher Faktor für eine ausreichende
Primärstabilität des Implantates. Im Oberkiefer ist eine Knochenhöhe des
Processus alveolaris von 10mm und eine Knochenbreite von 6mm erforderlich,
um ein Implantat ohne weitere Knochenaufbaumaßnahmen zu inserieren
(Vogeler et al. 2010). Sind diese Voraussetzungen nicht gegeben, kann bei
geringerem Knochenangebot folglich nur ein kürzeres und dünneres Implantat
eingebracht werden. Dies ist von erheblichem Nachteil, denn die Länge und der
Durchmesser des Implantates haben einen maßgeblichen Einfluss auf die
Erfolgsrate eines Implantates (Bahat 2000; Winkler et al. 2000). Winkler et al.
(2000) konnten in ihrer Studie eine Erfolgsrate von 96,4% bei einem 16mm
langen Implantat im Gegensatz zu einer Erfolgsrate von 66,7% bei einem 7mm
langen Implantat nachweisen .
Allerdings haben nicht nur Restknochenhöhe und Implantatgröße einen
maßgeblichen Einfluss auf Stabilität und Widerstandsfähigkeit des Implantates,
sondern neben weiteren Faktoren auch die Qualität des Knochens. Nach
Lekholm and Zarb (1985) wird der Knochen in vier Typen unterteilt, wobei Typ I
einen vorwiegend kompakten Knochen mit wenig spongiösem Anteil beschreibt
und Typ IV einen hohen Anteil an weitmaschiger Spongiosa und einer dünnen
Kompakta enthält. Typ II und III sind Mischtypen. So stellen Jaffin and Bermann
(1991) eine Verlustrate von 44% von Oberkieferimplantaten bei Knochen Typ IV
heraus. Lediglich 3,6% der Implantate, die im Oberkiefer inseriert wurden,
gingen bei Typ I–III verloren.
Die genaue Kenntnis der Pneumatisation des Processus alveolaris ist somit von
entscheidender Bedeutung, denn sie legt mitunter fest, ob bzw. welches
Implantat gesetzt werden kann.

Literaturübersicht
21
Sinusbodenelevation als erweiterte Indikation für Zahnimplantate
Eine unzureichende Knochensubstanz ist heute dennoch keine Kontraindikation
für eine Implantatinsertion in den posterioren, atrophierten Regionen des
Oberkiefers. Bereits Tatum (1977) und Boyne and James (1980) präsentierten
mit der externen Sinusbodenelevation ein operatives Verfahren, welches es
ermöglicht, ein ausreichendes Implantatlager bei mangelndem Knochenangebot
zu schaffen. Durch diese Methode des externen Sinusliftes hat sich die
Indikation für das Einsetzen von Zahnimplantaten im Oberkieferseitenzahngebiet
erheblich verbreitert. Dabei wird nach Anlegen eines Knochenfensters in die
laterale Kieferhöhlenwand die Kieferhöhlenschleimhaut, auch Schneider’sche
Membran genannt, vom Boden der Kieferhöhle vorsichtig abgelöst und nach
kranial und medial verlagert. Der entstandene Hohlraum zwischen
Kieferhöhlenboden und Kieferhöhlenschleimhaut wird nun mit autogenem
Knochen, synthetisch hergestellten Knochenersatzmaterialien oder einer
Kombination aus beiden aufgefüllt. Dadurch wird gewissermaßen die räumliche
Ausdehnung der Kieferhöhle verkleinert und durch die Einengung des
Augmentats die Höhe und Breite des Processus alveolaris zur Aufnahme eines
Implantates entscheidend verbessert.
Beträgt die Restknochenhöhe des Processus alveolaris maxillae vor dem
Sinuslift mindestens 4mm kann das Implantat nach Vogeler et al. (2010) in
einem einzeitigen Verfahren inseriert werden. Bei einer geringeren Restsubstanz
muss eine Einheilphase von 4–6 Monaten abgewartet werden, sodass das in die
Kieferhöhle eingebrachte Material knöchern um- und eingebaut werden kann
(Vogeler et al. 2010). Erst dann erfolgt die eigentliche Implantation. In diesem
Fall wird von einem zweizeitigen Verfahren gesprochen (Vogeler et al. 2010).
Neben dem externen Sinuslift existiert der interne Sinuslift. Diese weniger
invasive Methode der Osteotom-Technik wurde von Summers (1994) vorgestellt.
Nach der Technik von Summers (1994) wird zunächst das vorhandene
Implantatbett bis zum knöchernen Kieferhöhlenboden mit entsprechenden

Literaturübersicht
22
Instrumenten (Osteotomen) aufbereitet. Anschließend werden transalveolär
Kieferhöhlenboden und Kieferhöhlenschleimhaut nach kranial angehoben. Das
so gewonnene Lumen wird mit Augmentationsmaterial aufgefüllt und das
Implantat eingeschraubt (Summers 1994). Eine simultane Implantatinsertion ist
nur bei einer ausreichenden Restknochenhöhe möglich. Pjetersson et al. (2009)
stellen heraus, dass die transalveolare Technik eine verlässliche Methode bei
mindestens 5mm Restknochenhöhe ist. In einer Untersuchung von Rosen et al.
(1999) zeigt sich eine Implantatüberlebensrate von 96% bei einer
Restknochenhöhe von mindestens 5mm und sinkt auf 85,7% bei lediglich 4mm
oder weniger. Nach Toffler (2004) verringert sich die Überlebensrate von
Implantaten gar auf 73,3% bei 4mm oder einer geringeren Restknochenhöhe.
Material und Methoden
23
3 Material und Methoden
3.1 Material
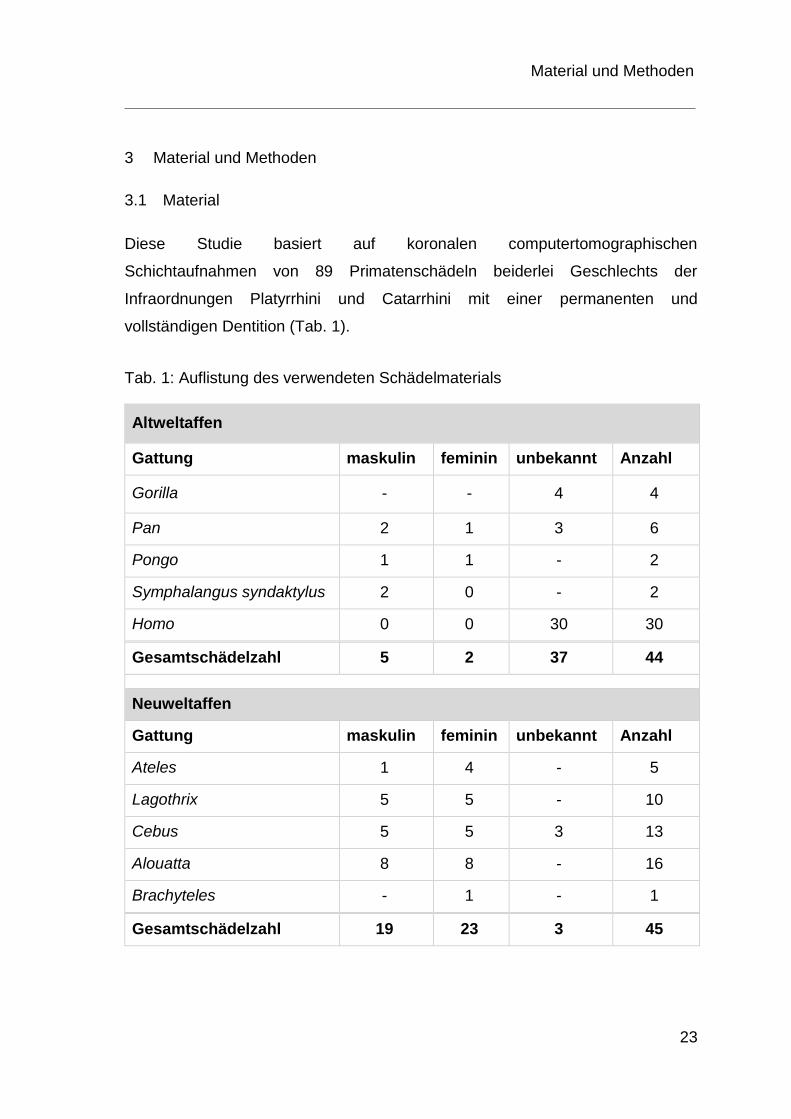
Diese Studie basiert auf koronalen computertomographischen
Schichtaufnahmen von 89 Primatenschädeln beiderlei Geschlechts der
Infraordnungen Platyrrhini und Catarrhini mit einer permanenten und
vollständigen Dentition (Tab. 1).
Tab. 1: Auflistung des verwendeten Schädelmaterials
Altweltaffen
Gattung maskulin feminin unbekannt Anzahl
Gorilla - - 4 4
Pan 2 1 3 6
Pongo 1 1 - 2
Symphalangus syndaktylus 2 0 - 2
Homo 0 0 30 30
Gesamtschädelzahl 5 2 37 44
Neuweltaffen
Gattung maskulin feminin unbekannt Anzahl
Ateles 1 4 - 5
Lagothrix 5 5 - 10
Cebus 5 5 3 13
Alouatta 8 8 - 16
Brachyteles - 1 - 1
Gesamtschädelzahl 19 23 3 45

Material und Methoden
24
Das Material entstammt der Zoologischen Staatssammlung der Ludwig-
Maximilian-Universität München, dem Institiut für Anatomie und Zellbiologie der
Ernst-Moritz-Arndt-Universität Greifswald, dem Primate Research Institute der
Kyoto University, Japan und dem Department of Anatomy, Histology and
Anthropology der Vilnius University, Litauen.
3.2 Methoden
3.2.1 Metrische Schädelvermessung
Messpunkte am Schädel
Um einen möglichen Zusammenhang zwischen dem Ausmaß des
Pneumatisationsgrades des Recessus alveolaris und der Schädelmorphologie
zwischen den Gattungen festzustellen, wurden zunächst in Anlehnung an Martin
(1928) und Harzer (1999) die folgenden Messpunkte definiert:
Opisthocranion Dorsalster Punkt des Schädels in der Mediansagittalebene
Basion Anteriorster Punkt des Foramen magnums
Nasion Schnittpunkt von Os nasale und Os frontale in der
Mediansagittalebene
Prosthion Anteriorster Punkt des Oberkiefer-Alveolarrandes zwischen
den mittleren Inzisivi in der Mediansagittalebene
Zygomaxillare Unterster Punkt der Sutura zygomticoaxillaris
Endomolare Palatinaler Rand des Limbus alveolaris auf Höhe des 2.
Molaren
Orale Schnittpunkt der Tangente durch die palatinalen Flächen
der mittleren oberen Inzisivi mit der Mediansagittalebene
Staphylion Schnittpunkt der Sutura palatina mediana und dem
hintersten Rand des harten Gaumens

Material und Methoden
25
Schädelmaße
Nach der Festlegung der Messpunkte wurden kraniometrische
Schädelvermessungen durchgeführt, die auf Martin (1928) zurückgehen
(Abb. 4–6). Ein Teil der Daten der Schädelvermessungen und
Volumenvermessungen entstammt den Untersuchungen von Koppe und
Schumacher (1990), Oesterreich et al. (2005), Röhrer-Ertl (persönliche
Mitteilung) und Frenkert (2008). Alle übrigen Messungen erfolgten an den unter
3.2.2. beschriebenen CT-Schichtaufnahmen mit dem Programm
OsiriX® Version 2.7.1. (Antoine Rosset & Osman Ratib) bzw.
ImageJ® Version 1.45s (National Institutes of Health, USA).
Folgende Schädelmaße wurden erhoben:
Längenmaße
SL: Gesamtschädellänge Prosthion – Opistokranion
GSL: Gesichtsschädellänge Nasion – Basion
SBL: Schädelbasislänge Basion – Prosthion
GL: Gaumenlänge Orale – Staphylion
Breitenmaße
MGB: Mittelgesichtsbreite Zygomaxillare – Zygomaxillare
MAB: Maxilloalveolarbreite Ektomolare – Ektomolare
GB: Gaumenbreite Endomolare – Endomolare
Höhenmaße
OGH: Obergesichtshöhe Nasion – Prosthion
GH: Gaumenhöhe Lot von höchsten Punkt des
Gaumengewölbes auf die Gaumenbreite in
Region der ersten Molaren
Material und Methoden
26
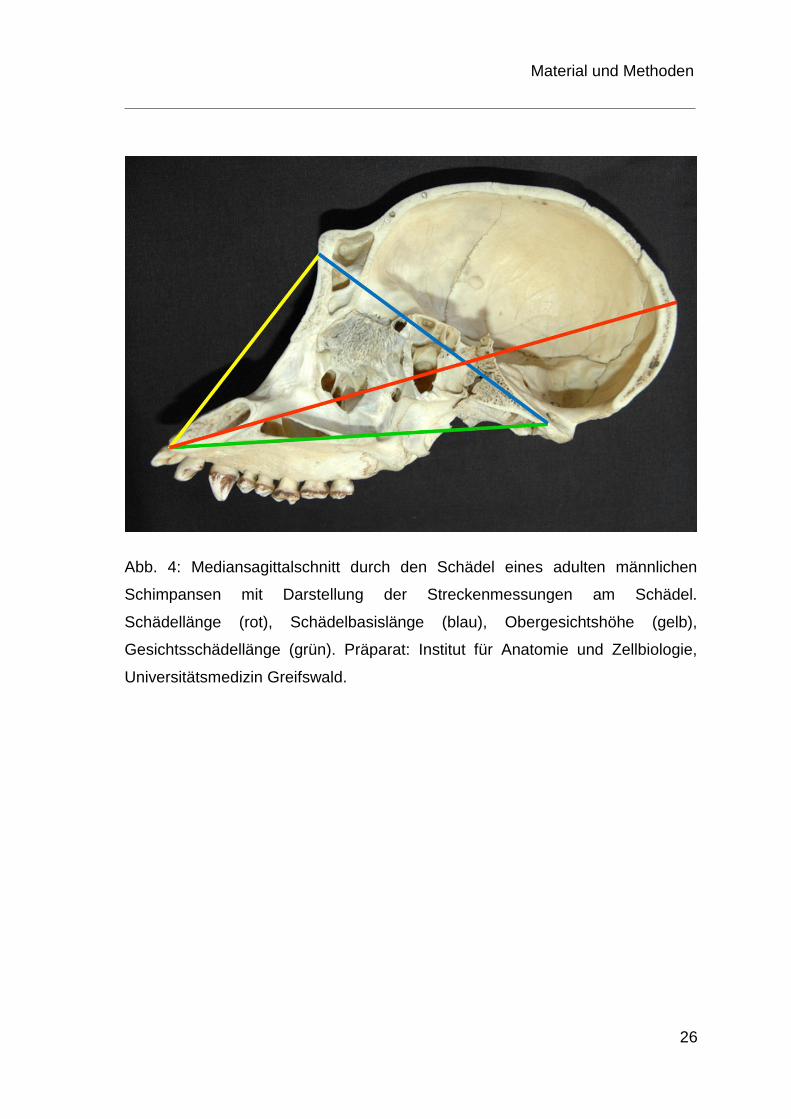
Abb. 4: Mediansagittalschnitt durch den Schädel eines adulten männlichen
Schimpansen mit Darstellung der Streckenmessungen am Schädel.
Schädellänge (rot), Schädelbasislänge (blau), Obergesichtshöhe (gelb),
Gesichtsschädellänge (grün). Präparat: Institut für Anatomie und Zellbiologie,
Universitätsmedizin Greifswald.
Material und Methoden
27
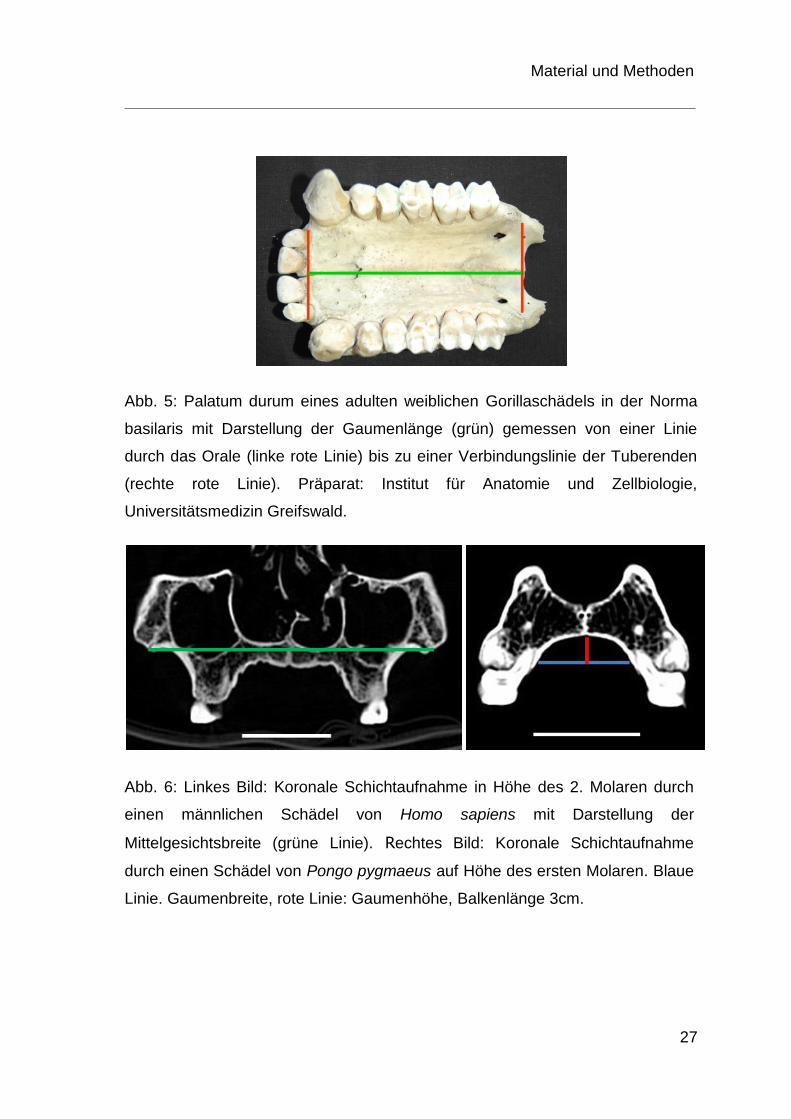
Abb. 5: Palatum durum eines adulten weiblichen Gorillaschädels in der Norma
basilaris mit Darstellung der Gaumenlänge (grün) gemessen von einer Linie
durch das Orale (linke rote Linie) bis zu einer Verbindungslinie der Tuberenden
(rechte rote Linie). Präparat: Institut für Anatomie und Zellbiologie,
Universitätsmedizin Greifswald.
Abb. 6: Linkes Bild: Koronale Schichtaufnahme in Höhe des 2. Molaren durch
einen männlichen Schädel von Homo sapiens mit Darstellung der
Mittelgesichtsbreite (grüne Linie). Rechtes Bild: Koronale Schichtaufnahme
durch einen Schädel von Pongo pygmaeus auf Höhe des ersten Molaren. Blaue
Linie. Gaumenbreite, rote Linie: Gaumenhöhe, Balkenlänge 3cm.

Material und Methoden
28
3.2.2 Computertomographische Vermessung des Recessus alveolaris
Um das Ausmaß des Pneumatisationsgrades des Recessus alveolaris näher
untersuchen zu können, wurden Volumen- und Streckenmessungen erhoben.
Die Anfertigung der CT-Bilder der Schädel aus der Zoologischen
Staatssammlung München erfolgte am Institut für Radiologische Diagnostik der
Ludwig-Maximilian-Universität München mithilfe eines Picker Spiral-CT PQ 5000
bei 120kV und 30mA. Das Material aus dem Primate Research Institute der
Kyoto University, Japan wurde am Department of Radiology, Okayama
University Medical School, Japan bei 120kV und 150mA mittels eines HiSpeed
Advantage RP CT (General Electric Medical Systems, Milwaukee, USA)
geröntgt. Die Schädel aus dem Institut für Anatomie und Zellbiologie der
Universitätsmedizin Greifswald und die Schädel vom Department of Anatomy,
Histology and Anthropology der Vilnius University Litauen wurden im Institut für
Radiologie der Universitätsmedizin Greifswald bei 120 kV und 75 mA mit dem
Siemens Sensation 16CT (Siemens AG, Berlin, Deutschland) geröntgt. Die
Aufnahmen erfolgten nach Ausrichtung der Schädel nach der Frankfurter
Horizontalen von rostral nach dorsal. Die Schichtdicke der Aufnahmen betrug
1–3mm.
Volumenmessungen des Recessus alveolaris
Die frontalen CT-Schichtaufnahmen lagen in Form von DICOM-Dateien vor.
Bevor diese mithilfe der 3D-Rekonstruktionssoftware „WinSurf® 4-1-0“ (Golden
Software Inc., USA) bearbeitet werden konnten, wurden die entsprechenden
Bildabschnitte des Recessus alveolaris unter Verwendung der Software eFilm
Workstation™ 1.5.3 (eFilm Medical Inc. Toronto, Ontario, Canada) in JPEG-
Dateien konvertiert und gespeichert. Die 3D-Rekonstruktionssoftware berechnet
nach Kennzeichnung der zu messenden Fläche in jeder CT-Schicht das
entsprechende Volumen und konstruiert ein dreidimensionales Bild. Dabei
wurde, um das Volumen des Recessus alveolaris zu berechnen, eine
horizontale Linie vom tiefsten Punkt der Nasenhöhle als Bezugsebene gezogen
Material und Methoden
29
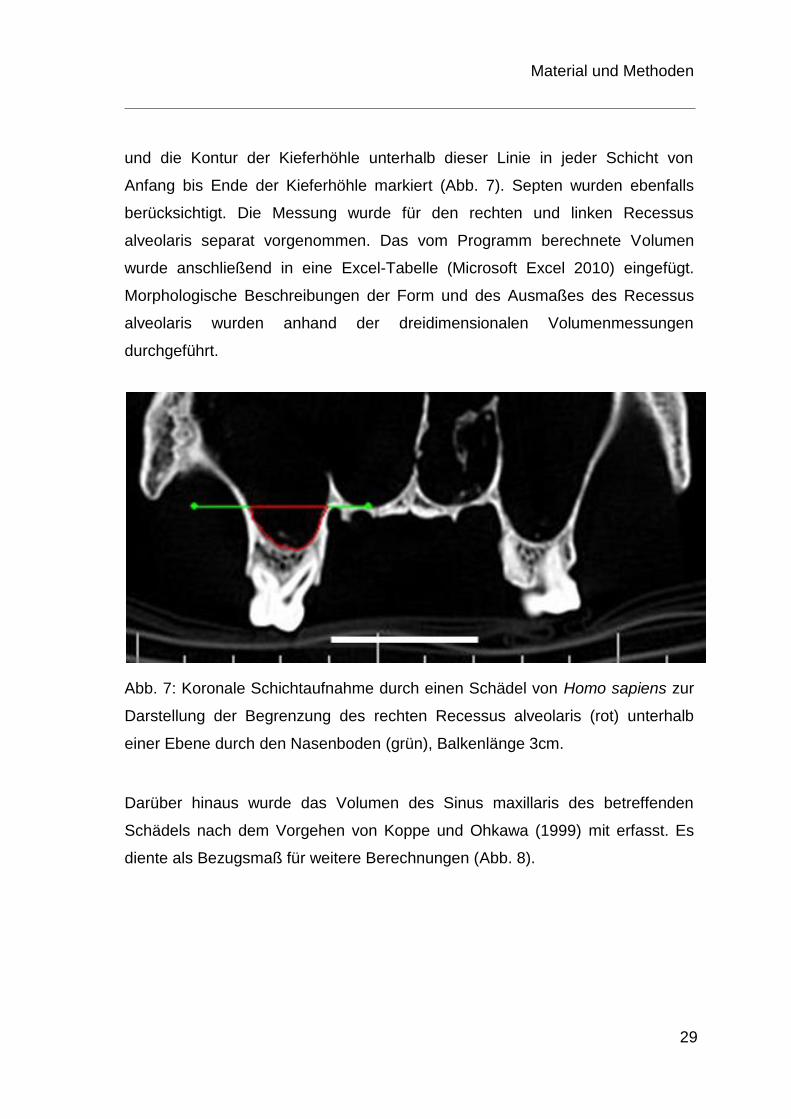
und die Kontur der Kieferhöhle unterhalb dieser Linie in jeder Schicht von
Anfang bis Ende der Kieferhöhle markiert (Abb. 7). Septen wurden ebenfalls
berücksichtigt. Die Messung wurde für den rechten und linken Recessus
alveolaris separat vorgenommen. Das vom Programm berechnete Volumen
wurde anschließend in eine Excel-Tabelle (Microsoft Excel 2010) eingefügt.
Morphologische Beschreibungen der Form und des Ausmaßes des Recessus
alveolaris wurden anhand der dreidimensionalen Volumenmessungen
durchgeführt.
Abb. 7: Koronale Schichtaufnahme durch einen Schädel von Homo sapiens zur
Darstellung der Begrenzung des rechten Recessus alveolaris (rot) unterhalb
einer Ebene durch den Nasenboden (grün), Balkenlänge 3cm.
Darüber hinaus wurde das Volumen des Sinus maxillaris des betreffenden
Schädels nach dem Vorgehen von Koppe und Ohkawa (1999) mit erfasst. Es
diente als Bezugsmaß für weitere Berechnungen (Abb. 8).

Material und Methoden
30
Abb. 8: Koronale CT-Schichtaufnahme eines Schimpansenschädels mit
Darstellung der Benutzeroberfläche von WinSurf® 4.0 und Konturierung des
vermessenen rechten Sinus maxillaris (grün) auf Höhe des 3. Molaren. Inset:
vergrößerter Ausschnitt des Bildes mit Darstellung der rechten Kieferhöhle
(grün), dem Canalis infraorbitalis (roter Pfeil) und dem Nasenboden (gelbe
Pfeile), Balkenlänge 2cm.
Material und Methoden
31
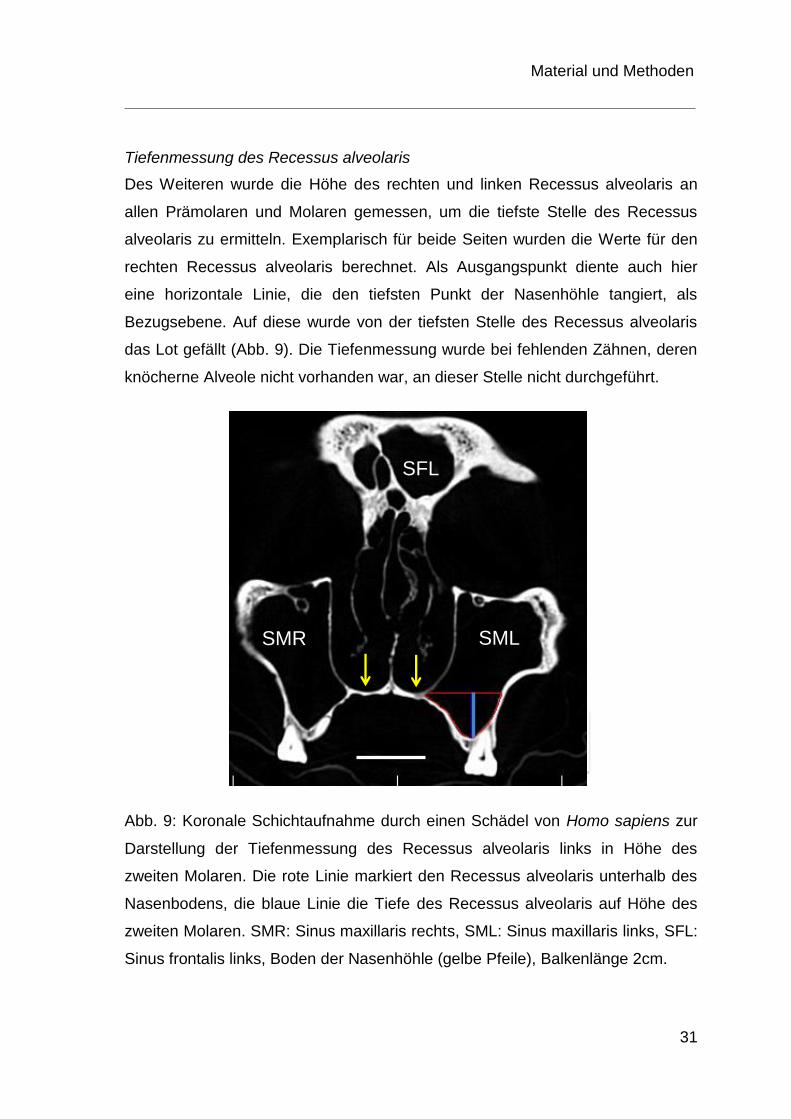
Tiefenmessung des Recessus alveolaris
Des Weiteren wurde die Höhe des rechten und linken Recessus alveolaris an
allen Prämolaren und Molaren gemessen, um die tiefste Stelle des Recessus
alveolaris zu ermitteln. Exemplarisch für beide Seiten wurden die Werte für den
rechten Recessus alveolaris berechnet. Als Ausgangspunkt diente auch hier
eine horizontale Linie, die den tiefsten Punkt der Nasenhöhle tangiert, als
Bezugsebene. Auf diese wurde von der tiefsten Stelle des Recessus alveolaris
das Lot gefällt (Abb. 9). Die Tiefenmessung wurde bei fehlenden Zähnen, deren
knöcherne Alveole nicht vorhanden war, an dieser Stelle nicht durchgeführt.
Abb. 9: Koronale Schichtaufnahme durch einen Schädel von Homo sapiens zur
Darstellung der Tiefenmessung des Recessus alveolaris links in Höhe des
zweiten Molaren. Die rote Linie markiert den Recessus alveolaris unterhalb des
Nasenbodens, die blaue Linie die Tiefe des Recessus alveolaris auf Höhe des
zweiten Molaren. SMR: Sinus maxillaris rechts, SML: Sinus maxillaris links, SFL:
Sinus frontalis links, Boden der Nasenhöhle (gelbe Pfeile), Balkenlänge 2cm.
SMLSMRSMLSMR
SFL

Material und Methoden
32
Im Einzelnen wurden folgende Messungen an den CT-Schichtaufnahmen
vorgenommen:
SMR: Volumen des rechten Sinus maxillaris
SML: Volumen des linken Sinus maxillaris
RAR: Volumen des rechten Recessus alveolaris
RAL: Volumen des linken Recessus alveolaris
TRAR: Tiefe des rechten Recessus alveolaris
TRAL Tiefe des linken Recessus alveolaris
3.2.3 Biostatistische Auswertung
Der Fokus dieser Arbeit liegt auf der Untersuchung der möglichen
Zusammenhänge zwischen der Größe des Recessus alveolaris und der
Schädelmorphologie von Neuweltaffen und Altweltaffen. Da für diese Studie
unterschiedliche Spezies vermessen wurden, erfolgte zunächst eine deskriptive
Statistik für die Werte des Recessus alveolaris der Schädel der einzelnen
Spezies von den entsprechenden Mittelwerten und Standardabweichungen. Die
Ergebnisse sind in Form von Boxplots graphisch dargestellt (Abb. 10).
Für den Vergleich der Gruppen wurden die Spezies entsprechend den
Altweltaffen bzw. Neuweltaffen zugeordnet. Die Prüfung auf Normalverteilung
erfolgte mit Hilfe des Kolmogorov-Smirnov-Anpassungstests. Berechnungen von
Mittelwerten und Standardabweichungen erfolgten auch hier. Die Mittelwerte der
zwei Gruppen wurden ferner einem T-Test unterzogen und dessen Signifikanz
auf einem Signifikanzniveau von p < 0,05 geprüft (Sokal and Rohlf 1995). Für
den T-Test wurden die Werte in den natürlichen Logarithmus transformiert.

Material und Methoden
33
Asymmetrieindex
Aufgrund der hohen Variabilität der Nasennebenhöhlen (Cave and Haines 1940,
Shankar et al. 1994) wurde weiterhin die Asymmetrie von rechtem und linkem
Recessus alveolaris für Altweltaffen und Neuweltaffen anhand des
Asymmetrieindex AI in Anlehnung an Lovich and Gibbons (1992) und Koppe and
Nagai (1997) berechnet. Dabei beschreibt GRA den größeren Recessus
alveolaris und KRA den kleineren Recessus alveolaris. Die Berechnung von
Mittelwerten und Standardabweichungen wurde dazu getrennt für Neuweltaffen
und Altweltaffen vorgenommen.
𝑨𝑰 = [− (𝑮𝑹𝑨
𝑲𝑹𝑨) ] + 𝟏𝒂)
𝑨𝑰 = [+ (𝑮𝑹𝑨
𝑲𝑹𝑨) ] − 𝟏𝒃)
a) Formel, wenn der Recessus alveolaris rechts größer ist
b) Formel, wenn der Recessus alveolaris links größer ist
Korrelation und partielle Korrelation
Um mögliche Beziehungen zwischen den Dimensionen des Recessus alveolaris
und den erhobenen Schädelmaßen herauszufinden, wurden die Parameter einer
Korrelationsanalyse unterzogen. Dabei wurden alle Paarungskombinationen
zwischen Recessus alveolaris und Schädelmaßen für Altweltaffen und
Neuweltaffen getrennt voneinander in Zusammenhang gebracht und die
Signifikanz der Korrelationskoeffizienten auf einem Signifikanzniveau von
p < 0,05 nach Pearson überprüft. Die Korrelationsanalyse wurde in einer
Korrelationsmatrix tabellarisch zusammengefasst.
Mögliche Scheinkorrelationen zwischen dem Recessus alveolaris und einem
entsprechenden anderen Schädelmaß wurden durch eine partielle
Korrelationsanalyse unter Berücksichtigung einer Kontrollvariablen geprüft. Als

Material und Methoden
34
Kontrollvariable wurde zum einen die Gesichtslänge, zum anderen die
Gesamtschädellänge einbezogen.
Um die Stärke der Zusammenhänge zwischen den untersuchten
Merkmalspaaren der Primatengruppen zu prüfen, wurden darüber hinaus die
Korrelationskoeffizienten nach Pearson mithilfe der interaktiven Internetseite
http://home.ubalt.edu/ntsbarsh/Business-stat/otherapplets/MultiCorr.htm (Zugriff
am 28.01.2013) verglichen. Die Berechnung ist auf der von Hays (1988)
beschriebenen Methode des Vergleichs von Korrelationskoeffizienten gestützt.
Regressionsanalyse
Für die Variablenpaare, bei denen eine signifikante Korrelation besteht, folgte
eine lineare Regressionsanalyse getrennt für Neuweltaffen und Altweltaffen mit
einer Irrtumswahrscheinlichkeit von p < 0,05, um Ausmaß und Stärke zwischen
zwei Merkmalspaaren zu errechnen. Insbesondere interessiert dabei das
Volumen des Recessus alveolaris in Bezug zu den anderen Schädelmaßen.
Graphisch dargestellt zeigte sich für die entsprechenden Merkmalspaare eine
Punkteverteilung entlang einer Regressionsgeraden, welche sich durch
folgenden Gleichungssatz mathematisch beschreiben lässt:
y = b x + a
mit:
x, y: korrelierende Merkmalspaare, wobei x (unabhängige
Variable) für den Recessus alveolaris und y (abhängige
Variable) für das korrelierende Schädelmaß steht
b: Anstieg der Regressionsgeraden
a: Schnittpunkt der Regressionsgeraden mit der y-Achse

Material und Methoden
35
Zusätzlich wurden, um die Beziehungen des Recessus alveolaris und anderen
Schädelmaßen zwischen Altweltaffen und Neuweltaffen zu untersuchen, die
Geradenanstiege mit der linearen Regressionsanalyse innerhalb einer
Merkmalspaarung verglichen. Das Signifikanzniveau wurde auch hier auf
p < 0,05 festgelegt. Um mögliche Ausreißer angemessen zu schätzen, wurden
im Berechnungsmodell die Huber-/White-/Sandwich-Schätzer als robuste
Schätzer bei der statistischen Analyse gewählt, damit der Einfluss der
Ausreißerwerte angemessen geschätzt wurde (Hardin and Hilbe 2012).
In Anlehnung an Ravosa (1991) wurden für die Korrelations- und
Regressionsanalyse die Messdaten in den natürlichen Logarithmus
transformiert, sodass eine Vergleichbarkeit innerhalb einer Gruppe möglich ist
(Jungers et al. 1995). Männliche und weibliche Tiere wurden bei der statistischen
Analyse zusammengefasst. Die biostatistische Auswertung erfolgte mit dem
Statistikprogramm SPSS® Version 19.0 (SPSS Inc.) und Microsoft Excel® 2010.

Ergebnisse
36
4 Ergebnisse
4.1 Morphologie des Recessus alveolaris
Volumenmessungen
Zunächst wurden die Volumina der rechten und linken Seite für die Kieferhöhle
und den Recessus alveolaris der entsprechenden Spezies getrennt für
Neuweltaffen und Altweltaffen berechnet. Ein Schädel der Spezies Homo
sapiens konnte nicht mit in die Berechnung einbezogen werden, bei einem
weiteren Schädel dieser Spezies lag der Boden der Kieferhöhle auf der linken
Seite auf einer Bezugslinie mit der Nasenebene. Insofern lag hier auf der linken
Seite nach unserer Definition kein Recessus alveolaris vor. Das
durchschnittliche Volumen des rechten Recessus alveolaris ergab bei den
Altweltaffen einen Wert von 2,2cm³ und auf der linken Seite von 2,47cm³. Bei
den Neuweltaffen betrug das Volumen lediglich 0,46cm³ auf der rechten Seite
und 0,41cm³ auf der linken Seite. Unter den Neuweltaffen zeigte Alouatta das
größte Volumen mit 4,31cm³ für den rechten Sinus maxillaris und mit 0,88cm³
für den rechten Recessus alveolaris gegenüber Cebus mit dem geringeren
Volumen von 1,24cm³ für die rechte Kieferhöhle und von 0,22cm³ für den
rechten Recessus alveolaris. Unter Einbeziehung der Volumenmessungen von
Oestereich et al. (2005) zeigt jedoch Brachyteles unter den Neuweltaffen das
kleinste Volumen des Recessus alveolaris auf. Demgegenüber wies unter den
Altweltaffen der Gorilla sowohl für den rechten Sinus maxillaris mit 27,00cm³ als
auch für den rechten Recessus alveolaris mit 5,00cm³ das größte Volumen auf.
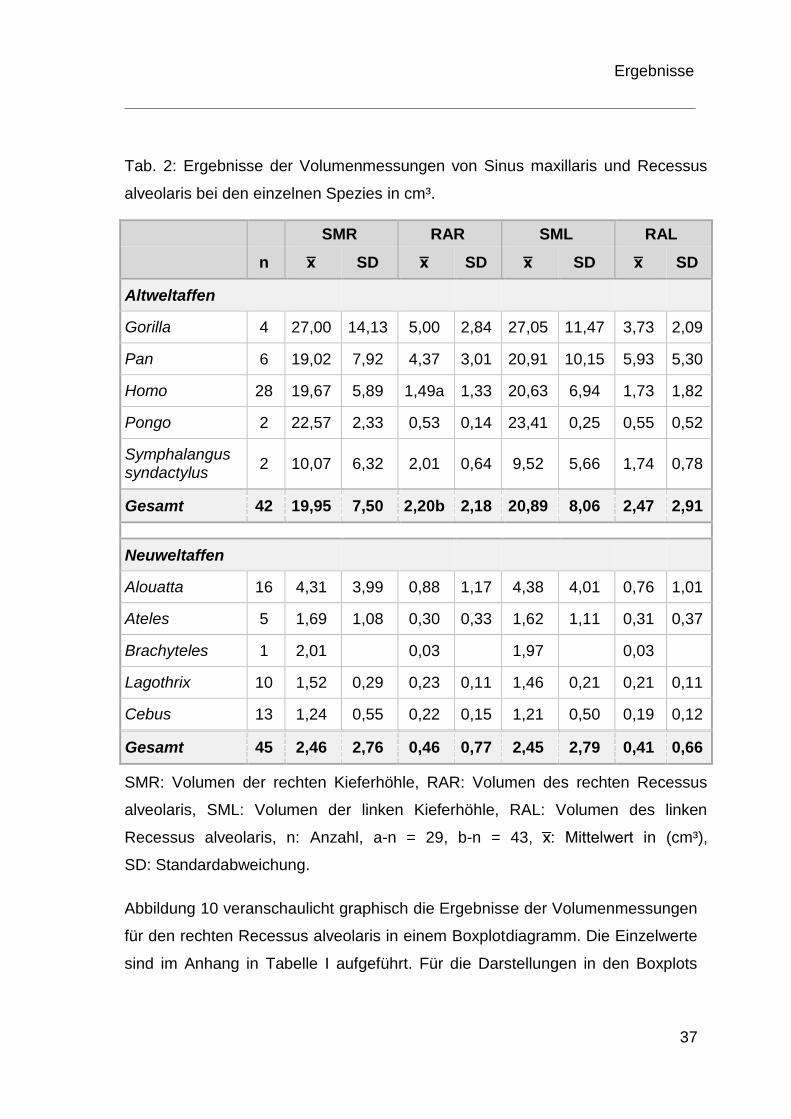
Die entsprechenden Ergebnisse sind Tabelle 2 zu entnehmen.
Ergebnisse
37
Tab. 2: Ergebnisse der Volumenmessungen von Sinus maxillaris und Recessus
alveolaris bei den einzelnen Spezies in cm³.
SMR RAR SML RAL
n x̅ SD x̅ SD x̅ SD x̅ SD
Altweltaffen
Gorilla 4 27,00 14,13 5,00 2,84 27,05 11,47 3,73 2,09
Pan 6 19,02 7,92 4,37 3,01 20,91 10,15 5,93 5,30
Homo 28 19,67 5,89 1,49a 1,33 20,63 6,94 1,73 1,82
Pongo 2 22,57 2,33 0,53 0,14 23,41 0,25 0,55 0,52
Symphalangus syndactylus
2 10,07 6,32 2,01 0,64 9,52 5,66 1,74 0,78
Gesamt 42 19,95 7,50 2,20b 2,18 20,89 8,06 2,47 2,91
Neuweltaffen
Alouatta 16 4,31 3,99 0,88 1,17 4,38 4,01 0,76 1,01
Ateles 5 1,69 1,08 0,30 0,33 1,62 1,11 0,31 0,37
Brachyteles 1 2,01
0,03
1,97
0,03
Lagothrix 10 1,52 0,29 0,23 0,11 1,46 0,21 0,21 0,11
Cebus 13 1,24 0,55 0,22 0,15 1,21 0,50 0,19 0,12
Gesamt 45 2,46 2,76 0,46 0,77 2,45 2,79 0,41 0,66
SMR: Volumen der rechten Kieferhöhle, RAR: Volumen des rechten Recessus
alveolaris, SML: Volumen der linken Kieferhöhle, RAL: Volumen des linken
Recessus alveolaris, n: Anzahl, a-n = 29, b-n = 43, x̅: Mittelwert in (cm³),
SD: Standardabweichung.
Abbildung 10 veranschaulicht graphisch die Ergebnisse der Volumenmessungen
für den rechten Recessus alveolaris in einem Boxplotdiagramm. Die Einzelwerte
sind im Anhang in Tabelle I aufgeführt. Für die Darstellungen in den Boxplots
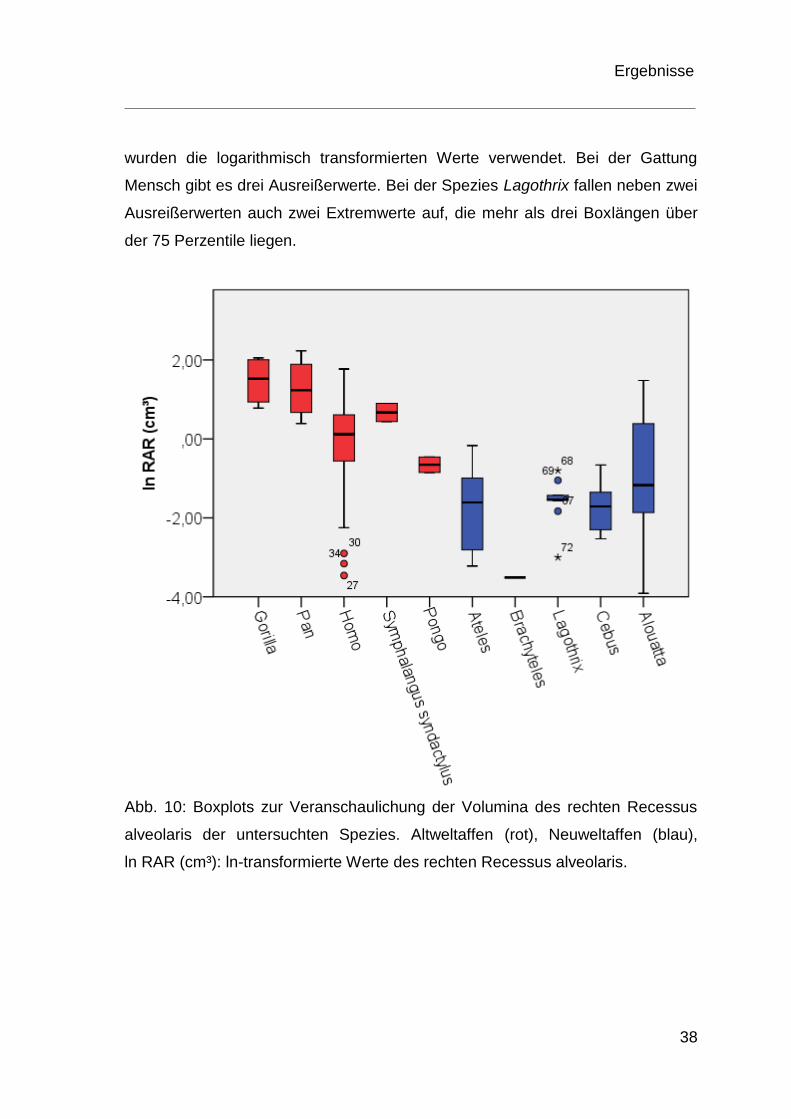
Ergebnisse
38
wurden die logarithmisch transformierten Werte verwendet. Bei der Gattung
Mensch gibt es drei Ausreißerwerte. Bei der Spezies Lagothrix fallen neben zwei
Ausreißerwerten auch zwei Extremwerte auf, die mehr als drei Boxlängen über
der 75 Perzentile liegen.
Abb. 10: Boxplots zur Veranschaulichung der Volumina des rechten Recessus
alveolaris der untersuchten Spezies. Altweltaffen (rot), Neuweltaffen (blau),
ln RAR (cm³): ln-transformierte Werte des rechten Recessus alveolaris.
Ergebnisse
39
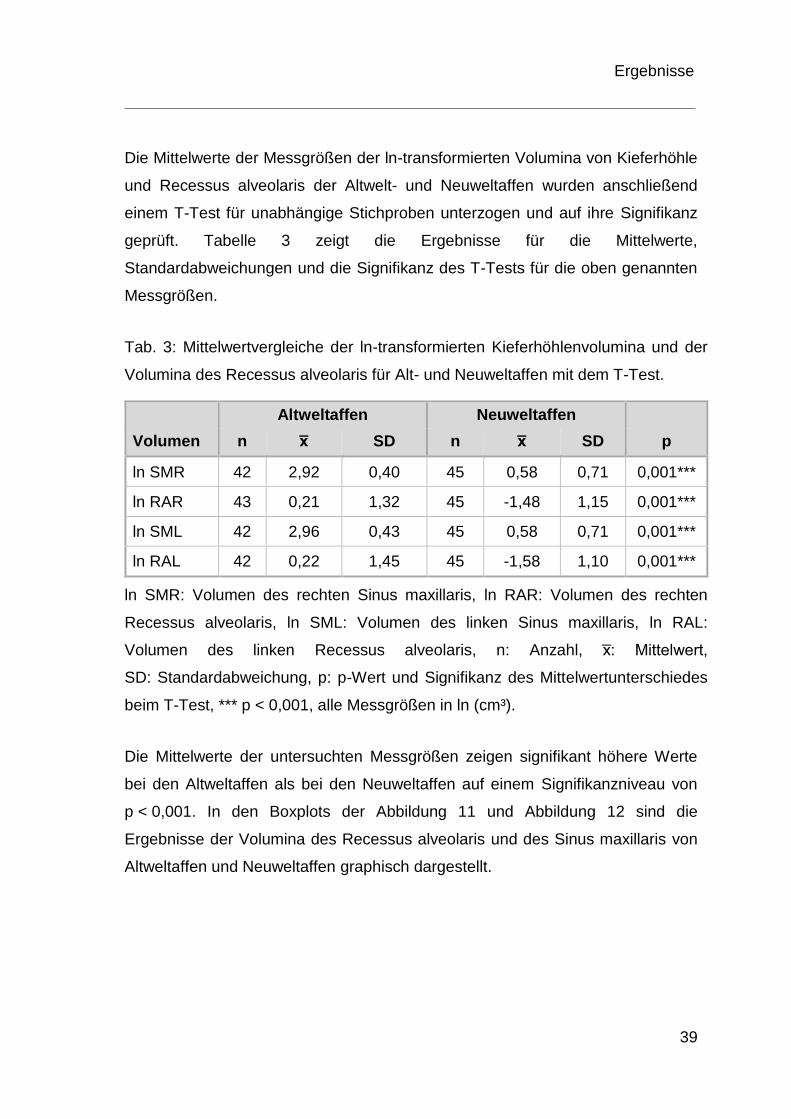
Die Mittelwerte der Messgrößen der ln-transformierten Volumina von Kieferhöhle
und Recessus alveolaris der Altwelt- und Neuweltaffen wurden anschließend
einem T-Test für unabhängige Stichproben unterzogen und auf ihre Signifikanz
geprüft. Tabelle 3 zeigt die Ergebnisse für die Mittelwerte,
Standardabweichungen und die Signifikanz des T-Tests für die oben genannten
Messgrößen.
Tab. 3: Mittelwertvergleiche der ln-transformierten Kieferhöhlenvolumina und der
Volumina des Recessus alveolaris für Alt- und Neuweltaffen mit dem T-Test.
Altweltaffen Neuweltaffen
Volumen n x̅ SD n x̅ SD p
ln SMR 42 2,92 0,40 45 0,58 0,71 0,001***
ln RAR 43 0,21 1,32 45 -1,48 1,15 0,001***
ln SML 42 2,96 0,43 45 0,58 0,71 0,001***
ln RAL 42 0,22 1,45 45 -1,58 1,10 0,001***
ln SMR: Volumen des rechten Sinus maxillaris, ln RAR: Volumen des rechten
Recessus alveolaris, ln SML: Volumen des linken Sinus maxillaris, ln RAL:
Volumen des linken Recessus alveolaris, n: Anzahl, x̅: Mittelwert,
SD: Standardabweichung, p: p-Wert und Signifikanz des Mittelwertunterschiedes
beim T-Test, *** p < 0,001, alle Messgrößen in ln (cm³).
Die Mittelwerte der untersuchten Messgrößen zeigen signifikant höhere Werte
bei den Altweltaffen als bei den Neuweltaffen auf einem Signifikanzniveau von
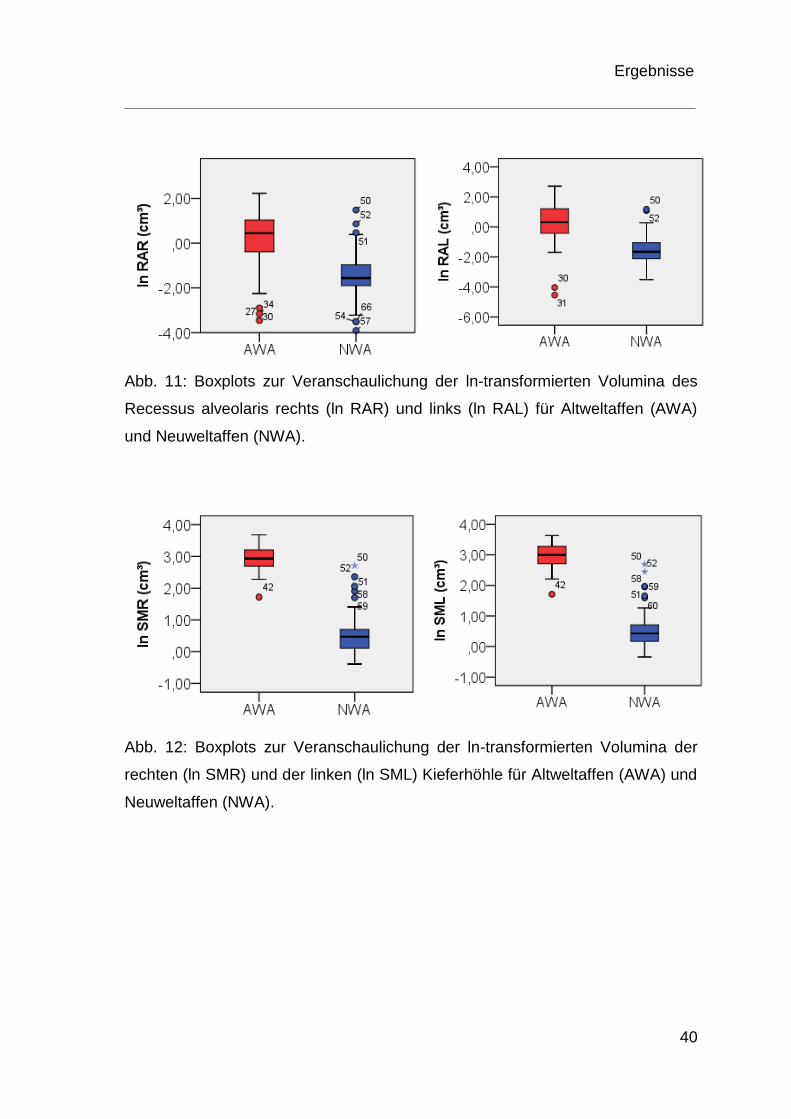
p < 0,001. In den Boxplots der Abbildung 11 und Abbildung 12 sind die
Ergebnisse der Volumina des Recessus alveolaris und des Sinus maxillaris von
Altweltaffen und Neuweltaffen graphisch dargestellt.
Ergebnisse
40
Abb. 11: Boxplots zur Veranschaulichung der ln-transformierten Volumina des
Recessus alveolaris rechts (ln RAR) und links (ln RAL) für Altweltaffen (AWA)
und Neuweltaffen (NWA).
Abb. 12: Boxplots zur Veranschaulichung der ln-transformierten Volumina der
rechten (ln SMR) und der linken (ln SML) Kieferhöhle für Altweltaffen (AWA) und
Neuweltaffen (NWA).

Ergebnisse
41
Morphologische Beschreibung des Recessus alveolaris
Bei den Altweltaffen lag bei insgesamt drei Fällen (drei Schimpansenschädel)
nach unserer Definition ein Recessus alveolaris bereits auf Höhe des ersten
Prämolaren vor. Bei 12 Schädeln begann der Recessus alveolaris auf Höhe des
zweiten Prämolaren. Dies traf bei zehn Schädeln von Homo sapiens, einem
Schädel eines Gorillas und einem Schimpansenschädel zu. In 23 Fällen
(17 Menschenschädel, drei Gorillaschädel, ein Schimpansenschädel, zwei
Schädel von Symphalangus syndactylus) lag der Recessus alveolaris rechts in
der Region des ersten Molaren vor. Ein weiterer Schädel der Spezies Homo
sapiens konnte in der Region des ersten Molaren nicht vermessen werden, da
der erste Molar fehlte und keine eindeutige Zuordnung getroffen werden konnte.
Bei beiden untersuchten Schädeln des Orang-Utans und in einem Fall von Homo
sapiens befand sich der Kieferhöhlenboden erst auf Höhe des zweiten Molaren
unterhalb einer Bezugsebene durch den Nasenboden. Der Recessus alveolaris
wurde nach dorsal hin flacher. Dennoch lag in 31 Fällen der Altweltaffen der
Kieferhöhlenboden auch noch hinter den dritten Molaren unterhalb einer
Bezugsebene durch den Nasenboden. In acht Fällen der Spezies Homo sapiens
endete der Recessus alveolaris distal des zweiten Molaren, in drei weiteren
Fällen derselben Spezies bereits mittig des zweiten Molaren. Anzumerken ist,
dass von den eben genannten elf menschlichen Schädeln in acht Fällen uni-
bzw. bilateral kein dritter Molar vorhanden war.
Die Morphologie des Bodens des Recessus alveolaris stellte sich bei jeder
Spezies sehr variabel dar. Er variierte von schmal bis breitgezogen, von flach bis
tief und von einer zerklüfteten Form bis hin zu einer wannenförmigen Struktur.
Die Wurzeln der Molaren beim Schimpansen hatten engen Kontakt zum
Recessus alveolaris und waren meist nur durch eine dünne Knochenlamelle
getrennt. Durch diese anatomische Nähe von Kieferhöhlenboden und Zähnen
wölbte sich der Boden der Kieferhöhle regelrecht auf. Somit stellte sich der
Kieferhöhlenboden unregelmäßig und zerklüftet dar (Abb. 13).

Ergebnisse
42
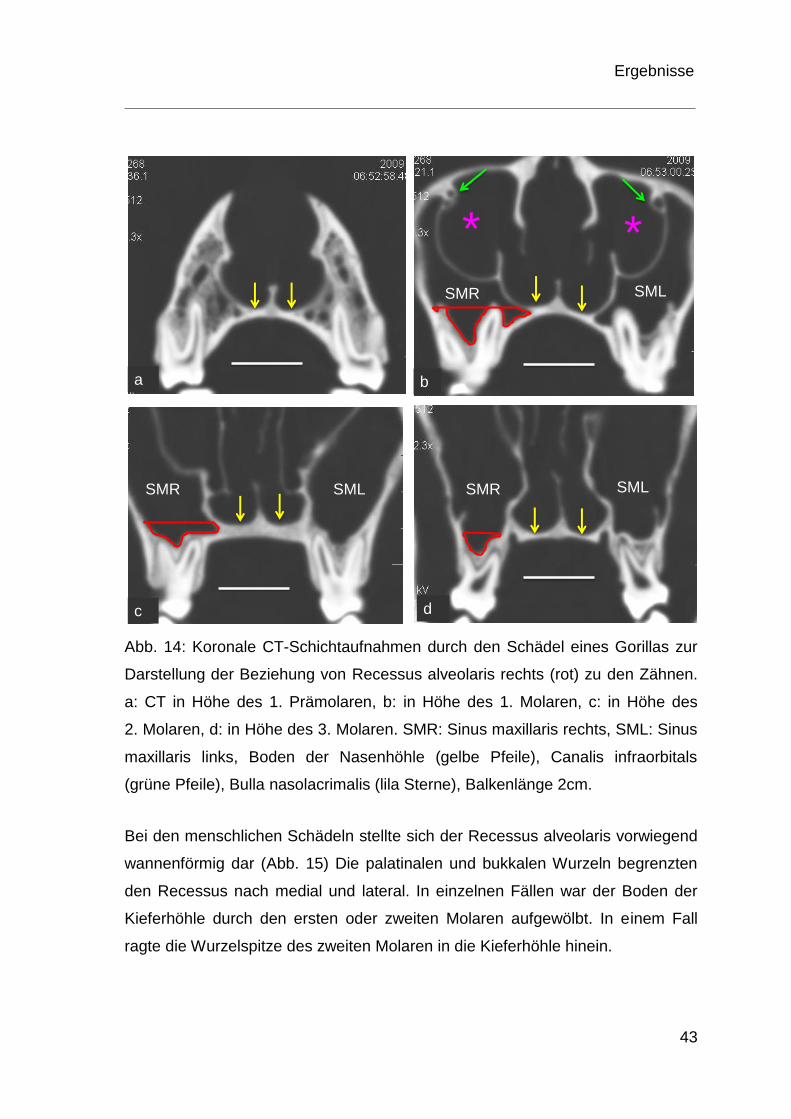
Beim Gorilla zeigte sich ein anderes Bild. Bei drei von vier Schädeln zeigt sich
eine Wannenform des Recessus alveolaris, der sich in die Furkation der ersten,
zweiten und dritten Molaren ausdehnte (vgl. Abb. 14c, d). Die palatinale Wurzel
des ersten Molaren stellte die mediale Begrenzung des Recessus alveolaris dar.
In einem Fall ragte die palatinale Wurzel des ersten Molaren direkt in die
Kieferhöhle (Abb. 14b), wodurch sich der Kieferhöhlenboden ebenfalls uneben
darstellte.
Abb. 13: 3D-Rekonstruktion des Bodens des rechten Recessus alveolaris eines
adulten Schimpansen a: Ansicht von basal. Beachte die durch Zahnwurzeln
hervorgerufenen Impressionen des Kieferhöhlenbodens (rote Pfeile). b: Ansicht
von dorsal. Die 3D-Figur ist in der Frontalebene um 45 Grad nach kranial
gedreht. Beachte den stark zerklüfteten Boden der Kieferhöhle.
Ergebnisse
43
Abb. 14: Koronale CT-Schichtaufnahmen durch den Schädel eines Gorillas zur
Darstellung der Beziehung von Recessus alveolaris rechts (rot) zu den Zähnen.
a: CT in Höhe des 1. Prämolaren, b: in Höhe des 1. Molaren, c: in Höhe des
2. Molaren, d: in Höhe des 3. Molaren. SMR: Sinus maxillaris rechts, SML: Sinus
maxillaris links, Boden der Nasenhöhle (gelbe Pfeile), Canalis infraorbitals
(grüne Pfeile), Bulla nasolacrimalis (lila Sterne), Balkenlänge 2cm.
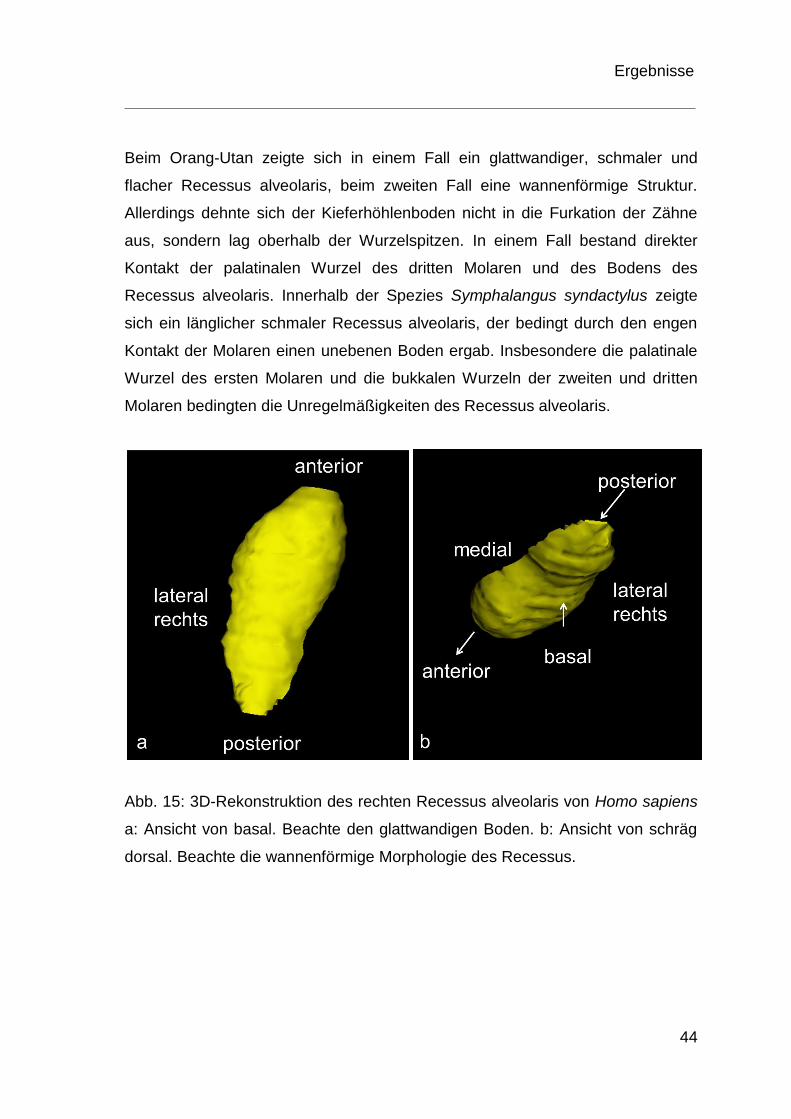
Bei den menschlichen Schädeln stellte sich der Recessus alveolaris vorwiegend
wannenförmig dar (Abb. 15) Die palatinalen und bukkalen Wurzeln begrenzten
den Recessus nach medial und lateral. In einzelnen Fällen war der Boden der
Kieferhöhle durch den ersten oder zweiten Molaren aufgewölbt. In einem Fall
ragte die Wurzelspitze des zweiten Molaren in die Kieferhöhle hinein.
* *SMLSMR
SMLSMR SMLSMR
a
dc
b
Ergebnisse
44
Beim Orang-Utan zeigte sich in einem Fall ein glattwandiger, schmaler und
flacher Recessus alveolaris, beim zweiten Fall eine wannenförmige Struktur.
Allerdings dehnte sich der Kieferhöhlenboden nicht in die Furkation der Zähne
aus, sondern lag oberhalb der Wurzelspitzen. In einem Fall bestand direkter
Kontakt der palatinalen Wurzel des dritten Molaren und des Bodens des
Recessus alveolaris. Innerhalb der Spezies Symphalangus syndactylus zeigte
sich ein länglicher schmaler Recessus alveolaris, der bedingt durch den engen
Kontakt der Molaren einen unebenen Boden ergab. Insbesondere die palatinale
Wurzel des ersten Molaren und die bukkalen Wurzeln der zweiten und dritten
Molaren bedingten die Unregelmäßigkeiten des Recessus alveolaris.
Abb. 15: 3D-Rekonstruktion des rechten Recessus alveolaris von Homo sapiens
a: Ansicht von basal. Beachte den glattwandigen Boden. b: Ansicht von schräg
dorsal. Beachte die wannenförmige Morphologie des Recessus.

Ergebnisse
45
Bei den Neuweltaffen wies Alouatta einen breitflächigen Recessus alveolaris auf.
Dieser dehnte sich nicht nur in die Furkation der Molaren aus, sondern auch
nach lateral über die Wurzeln der Molaren hinaus. Die Wurzeln der ersten und
zweiten Molaren ragten in die Kieferhöhle hinein (Abb. 16). Bei Cebus zeigte
sich eine ähnliche Morphologie. Allerdings berührten bereits die Wurzeln des
dritten Prämolaren den Kieferhöhlenboden bzw. ragten in diesen hinein
(Abb. 18). Von basal stellte sich eine schmale, längliche Form des Recessus
alveolaris dar (Abb. 17).
Abb. 16: 3D-Rekonstruktion des rechten Recessus alveolaris von Alouatta
seniculus. Linkes Bild: Ansicht von basal. Beachte die breit gezogene Form des
Recessus alveolaris. Rechtes Bild: Ansicht von schräg dorsal. Beachte den
unregelmäßigen Boden des Recessus alveolaris.
Ergebnisse
46
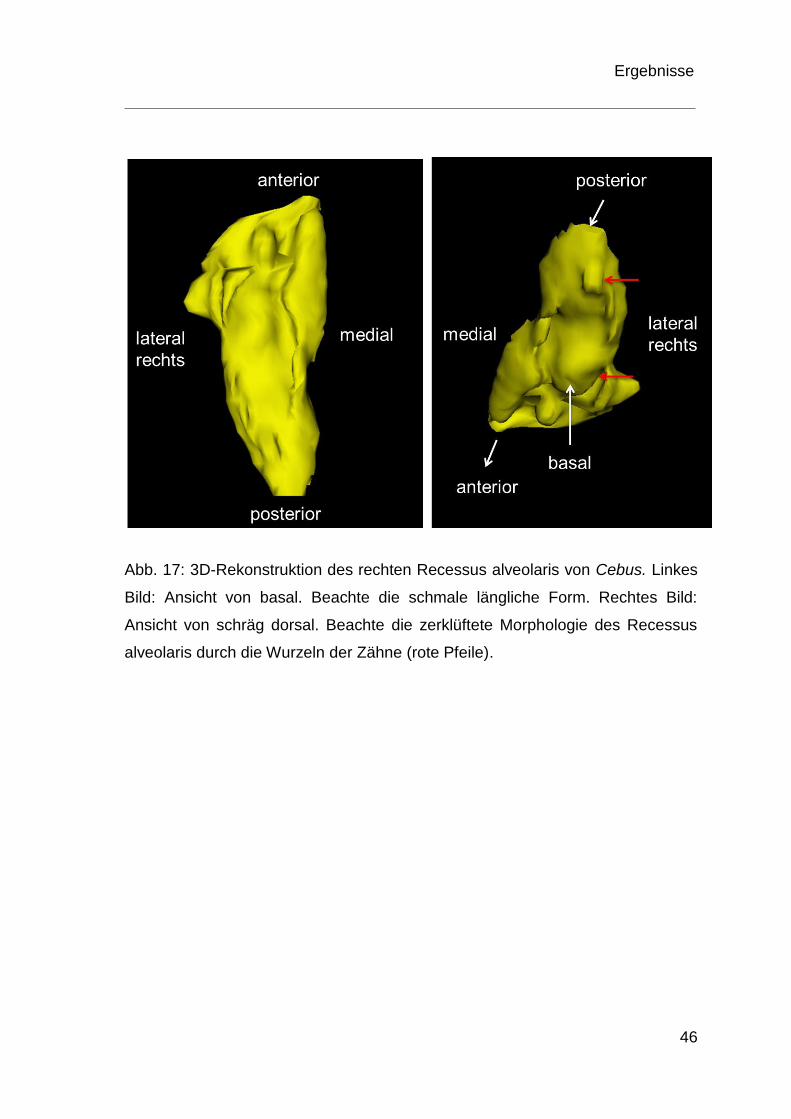
Abb. 17: 3D-Rekonstruktion des rechten Recessus alveolaris von Cebus. Linkes
Bild: Ansicht von basal. Beachte die schmale längliche Form. Rechtes Bild:
Ansicht von schräg dorsal. Beachte die zerklüftete Morphologie des Recessus
alveolaris durch die Wurzeln der Zähne (rote Pfeile).
Ergebnisse
47
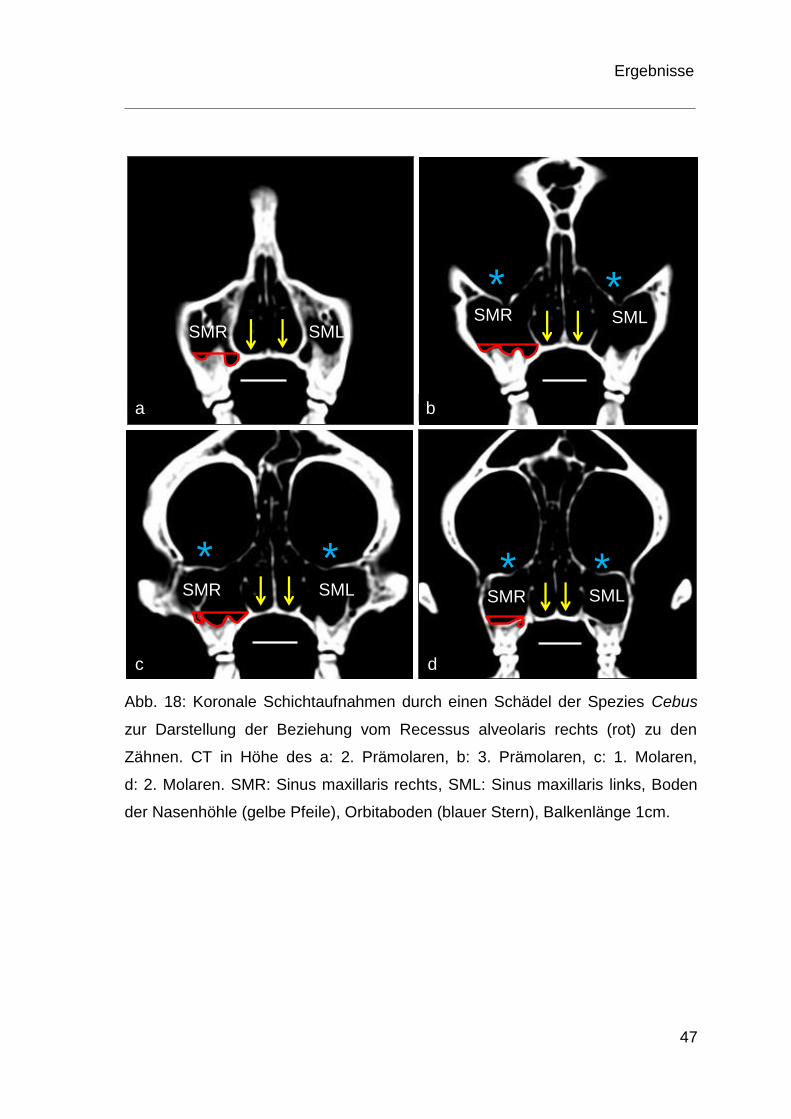
Abb. 18: Koronale Schichtaufnahmen durch einen Schädel der Spezies Cebus
zur Darstellung der Beziehung vom Recessus alveolaris rechts (rot) zu den
Zähnen. CT in Höhe des a: 2. Prämolaren, b: 3. Prämolaren, c: 1. Molaren,
d: 2. Molaren. SMR: Sinus maxillaris rechts, SML: Sinus maxillaris links, Boden
der Nasenhöhle (gelbe Pfeile), Orbitaboden (blauer Stern), Balkenlänge 1cm.
* *SMR SML
SMR SML
* *SMR SML
* *SMR SML
c
b
d
a

Ergebnisse
48
Tiefenmessung des Recessus alveolaris
Die Tiefen des rechten und linken Recessus alveolaris wurden auf Höhe der
Prämolaren und Molaren vermessen. Die Berechnungen erfolgten für den
rechten Recessus alveolaris. Die genauen Werte sind dem Anhang zu
entnehmen (Tab. II). Je ein Schädel der Spezies Homo sapiens und ein Schädel
von Pan troglodytes konnten hinsichtlich der Tiefenmessung keine
Berücksichtigung finden und wurden ausgeschlossen. Bei einem weiteren
menschlichen Schädel fehlte der erste Molar. Die Tiefenvermessung wurde für
diesen Schädel auf Höhe des ersten Molaren somit nicht durchgeführt.
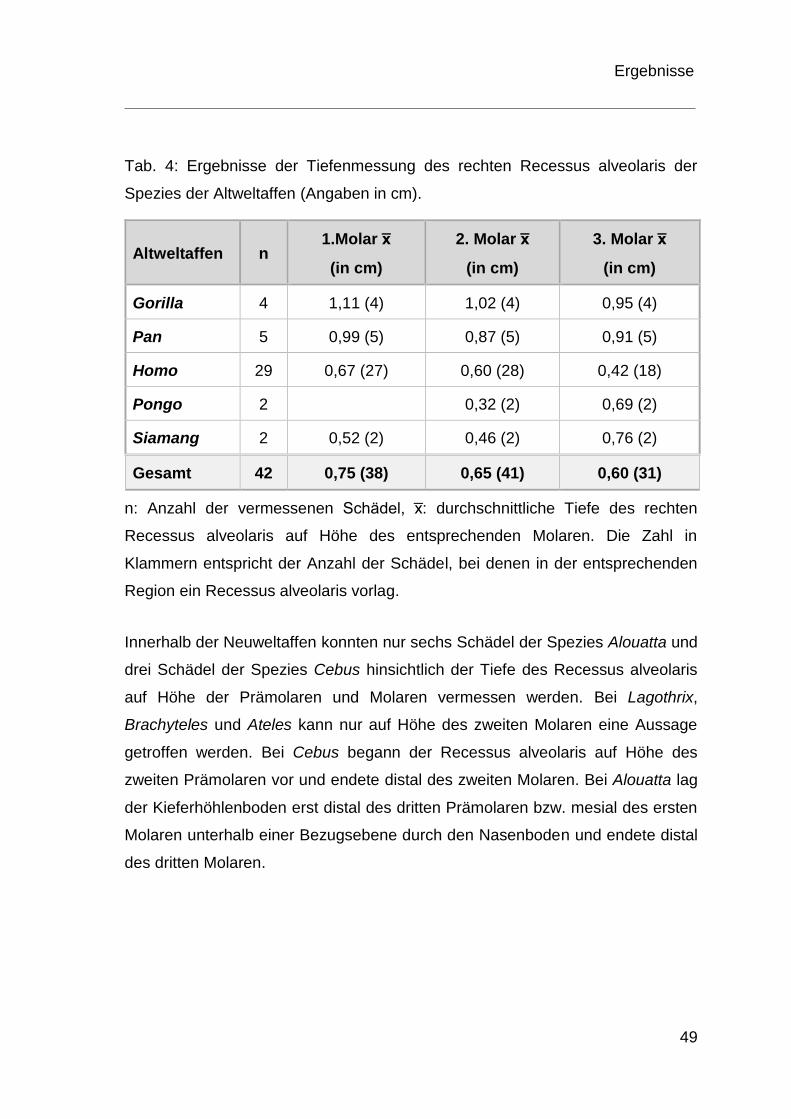
Die durchschnittlichen Tiefen auf Höhe der Molaren des rechten Recessus
alveolaris sind in Tabelle 4 für Altweltaffen und Tabelle 5 für Neuweltaffen
aufgelistet. Bei den Schädeln von Gorilla, Schimpansen und Homo sapiens
zeigte sich, dass die tiefste Stelle in der Region des ersten Molaren lag. Beim
Orang-Utan und beim Siamang befand sich die tiefste Stelle auf Höhe des dritten
Molaren. Werden alle Werte der vermessenen Altweltaffen betrachtet, liegt der
durchschnittliche Wert für die tiefste Stelle des rechten Recessus alveolaris mit
0,75cm auf Höhe des ersten Molaren.
Ergebnisse
49
Tab. 4: Ergebnisse der Tiefenmessung des rechten Recessus alveolaris der
Spezies der Altweltaffen (Angaben in cm).
Altweltaffen n 1.Molar x̅
(in cm)
2. Molar x̅
(in cm)
3. Molar x̅
(in cm)
Gorilla 4 1,11 (4) 1,02 (4) 0,95 (4)
Pan 5 0,99 (5) 0,87 (5) 0,91 (5)
Homo 29 0,67 (27) 0,60 (28) 0,42 (18)
Pongo 2 0,32 (2) 0,69 (2)
Siamang 2 0,52 (2) 0,46 (2) 0,76 (2)
Gesamt 42 0,75 (38) 0,65 (41) 0,60 (31)
n: Anzahl der vermessenen Schädel, x̅: durchschnittliche Tiefe des rechten
Recessus alveolaris auf Höhe des entsprechenden Molaren. Die Zahl in
Klammern entspricht der Anzahl der Schädel, bei denen in der entsprechenden
Region ein Recessus alveolaris vorlag.
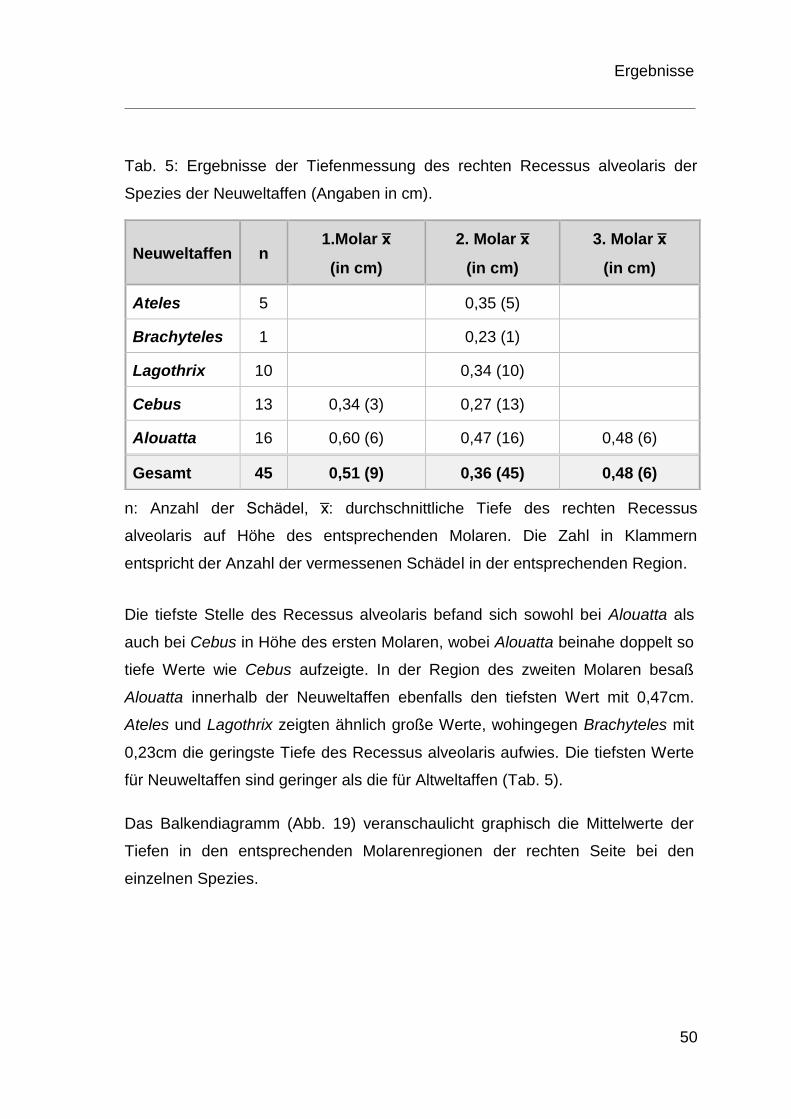
Innerhalb der Neuweltaffen konnten nur sechs Schädel der Spezies Alouatta und
drei Schädel der Spezies Cebus hinsichtlich der Tiefe des Recessus alveolaris
auf Höhe der Prämolaren und Molaren vermessen werden. Bei Lagothrix,
Brachyteles und Ateles kann nur auf Höhe des zweiten Molaren eine Aussage
getroffen werden. Bei Cebus begann der Recessus alveolaris auf Höhe des
zweiten Prämolaren vor und endete distal des zweiten Molaren. Bei Alouatta lag
der Kieferhöhlenboden erst distal des dritten Prämolaren bzw. mesial des ersten
Molaren unterhalb einer Bezugsebene durch den Nasenboden und endete distal
des dritten Molaren.
Ergebnisse
50
Tab. 5: Ergebnisse der Tiefenmessung des rechten Recessus alveolaris der
Spezies der Neuweltaffen (Angaben in cm).
Neuweltaffen n 1.Molar x̅
(in cm)
2. Molar x̅
(in cm)
3. Molar x̅
(in cm)
Ateles 5 0,35 (5)
Brachyteles 1 0,23 (1)
Lagothrix 10 0,34 (10)
Cebus 13 0,34 (3) 0,27 (13)
Alouatta 16 0,60 (6) 0,47 (16) 0,48 (6)
Gesamt 45 0,51 (9) 0,36 (45) 0,48 (6)
n: Anzahl der Schädel, x̅: durchschnittliche Tiefe des rechten Recessus
alveolaris auf Höhe des entsprechenden Molaren. Die Zahl in Klammern
entspricht der Anzahl der vermessenen Schädel in der entsprechenden Region.
Die tiefste Stelle des Recessus alveolaris befand sich sowohl bei Alouatta als
auch bei Cebus in Höhe des ersten Molaren, wobei Alouatta beinahe doppelt so
tiefe Werte wie Cebus aufzeigte. In der Region des zweiten Molaren besaß
Alouatta innerhalb der Neuweltaffen ebenfalls den tiefsten Wert mit 0,47cm.
Ateles und Lagothrix zeigten ähnlich große Werte, wohingegen Brachyteles mit
0,23cm die geringste Tiefe des Recessus alveolaris aufwies. Die tiefsten Werte
für Neuweltaffen sind geringer als die für Altweltaffen (Tab. 5).
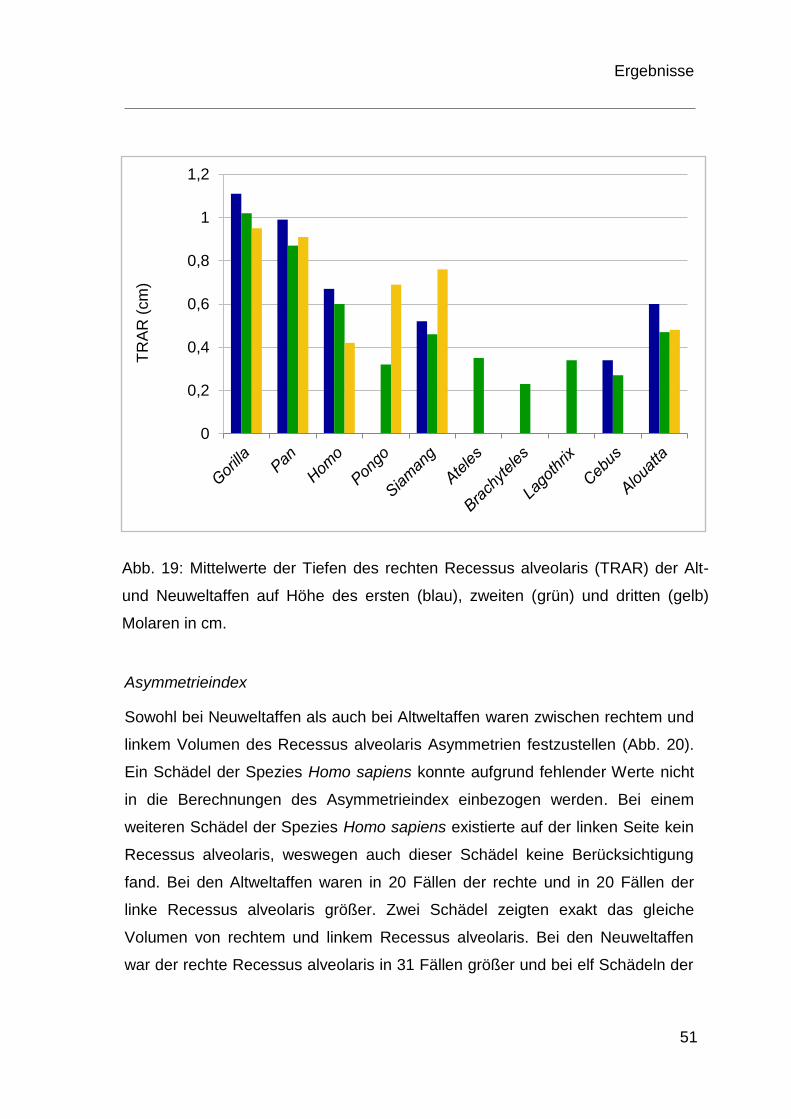
Das Balkendiagramm (Abb. 19) veranschaulicht graphisch die Mittelwerte der
Tiefen in den entsprechenden Molarenregionen der rechten Seite bei den
einzelnen Spezies.
Ergebnisse
51
Abb. 19: Mittelwerte der Tiefen des rechten Recessus alveolaris (TRAR) der Alt-
und Neuweltaffen auf Höhe des ersten (blau), zweiten (grün) und dritten (gelb)
Molaren in cm.
Asymmetrieindex
Sowohl bei Neuweltaffen als auch bei Altweltaffen waren zwischen rechtem und
linkem Volumen des Recessus alveolaris Asymmetrien festzustellen (Abb. 20).
Ein Schädel der Spezies Homo sapiens konnte aufgrund fehlender Werte nicht
in die Berechnungen des Asymmetrieindex einbezogen werden. Bei einem
weiteren Schädel der Spezies Homo sapiens existierte auf der linken Seite kein
Recessus alveolaris, weswegen auch dieser Schädel keine Berücksichtigung
fand. Bei den Altweltaffen waren in 20 Fällen der rechte und in 20 Fällen der
linke Recessus alveolaris größer. Zwei Schädel zeigten exakt das gleiche
Volumen von rechtem und linkem Recessus alveolaris. Bei den Neuweltaffen
war der rechte Recessus alveolaris in 31 Fällen größer und bei elf Schädeln der
0
0,2
0,4
0,6
0,8
1
1,2
TR
AR
(cm
)

Ergebnisse
52
linke größer. Hier zeigten drei Schädel exakt das gleiche Volumen der rechten
und linken Seite. Bei den Neuweltaffen zeigte ein Schädel von Alouatta einen
Extremwert der linken Seite mit 1,00 auf. Bei den Altweltaffen stellten sich zwei
Extremwerte des Recessus alveolaris heraus. Dies war bei Homo sapiens mit
-71,00 zugunsten der rechten Seite und mit 5,67 zugunsten der linken Seite der
Fall.
In der Abbildung 20 ist der besseren Übersicht halber der Extremwert eines
Homo sapiens von -71,00 nicht dargestellt. Für Altweltaffen ergab der mittlere
Wert für den Asymmetrieindex -1,58 bei einer Standardabweichung 11,05, für
Neuweltaffen lag der Mittelwert des Asymmetrieindex bei -0,11 bei einer
Standardabweichung von 0,33 (Tab. 6). Die genauen Werte für den
Asymmetrieindex sind dem Anhang zu entnehmen (Tab. I)
Tab. 6: Ergebnisse des Asymmetrieindex von Altweltaffen und Neuweltaffen.
Asymmetrieindex
n x̅ SD Min Max n (re) n (li) n (=)
Altweltaffen
AI (cm) 42 -1,58 11,05 -71,00 5,67 20 20 2
Neuweltaffen
AI (cm) 45 -0,11 0,33 -0,90 1,00 31 11 3
AI (cm): Asymmetrieindex des Recessus alveolaris in cm, n: Anzahl, x̅:
Mittelwert, SD: Standardabweichung, Min: kleinster Wert des Asymmetrieindex,
Max: größter Wert des Asymmetrieindex, n (re): Anzahl der Schädel, deren
rechter Recessus alveolaris größer ist, n (li): Anzahl der Schädel, deren linker
Recessus alveolaris größer ist, n (=): Anzahl der Schädel, deren rechter und
linker Recessus alveolaris gleich groß sind.
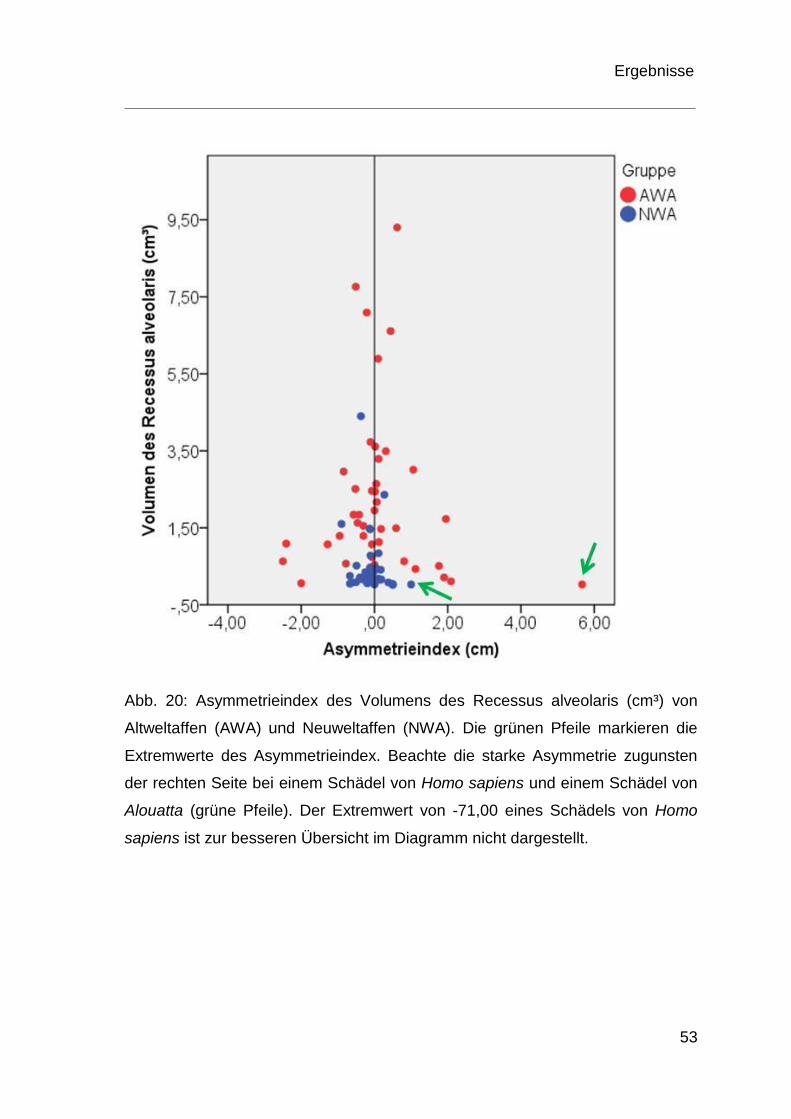
Ergebnisse
53
Abb. 20: Asymmetrieindex des Volumens des Recessus alveolaris (cm³) von
Altweltaffen (AWA) und Neuweltaffen (NWA). Die grünen Pfeile markieren die
Extremwerte des Asymmetrieindex. Beachte die starke Asymmetrie zugunsten
der rechten Seite bei einem Schädel von Homo sapiens und einem Schädel von
Alouatta (grüne Pfeile). Der Extremwert von -71,00 eines Schädels von Homo
sapiens ist zur besseren Übersicht im Diagramm nicht dargestellt.

Ergebnisse
54
4.2 Ergebnisse der Korrelationsanalyse
Die Zusammenhänge zwischen den erfassten Längen- und Breitenmaßen und
den vermessenen Volumina wurden in einer Korrelationsanalyse nach Pearson
getrennt für Altweltaffen und Neuweltaffen ermittelt. Die genauen Ergebnisse der
Korrelationsanalyse sind für alle Messgrößen im Anhang für Altweltaffen
(Tab. III) und Neuweltaffen (Tab. IV) aufgelistet. Anhand dieser Ergebnisse
wurde eine Korrelationsmatrix erstellt, die die signifikanten Korrelationen
innerhalb der jeweiligen Gruppe der Affen zusammenfasst (Tab. 7).
Innerhalb der Neuweltaffen korrelierten sowohl rechter als auch linker Recessus
alveolaris mit jeder anderen Messgröße ausgenommen mit der
Mittelgesichtsbreite und der Maxilloalveolarbreite. Für die Altweltaffen ergab sich
kein Zusammenhang zwischen dem rechten Recessus alveolaris und der
Gesamtschädellänge, der Schädelbasislänge, der Mittelgesichtsbreite,
Maxilloalveolarbreite und der Gaumenbreite. Für die linke Seite des Recessus
alveolaris ergab sich der gleiche Sachverhalt. Aufgrund dieser Ergebnisse
wurden die folgenden Berechnungen exemplarisch für beide Seiten anhand der
rechten Seite durchgeführt.

Ergebnisse
55
Tab. 7: Übersicht der signifikanten korrelierenden Merkmalspaare innerhalb der
Altwelt- und Neuweltaffen.
SL
SB
L
GS
L
MG
B
MA
B
GL
GB
GH
OG
H
SM
R
RA
R
SM
L
RA
L
SL 12 2 12 12 2 12 12 12 12 2 12 2
SBL 12 2 12 12 2 12 12 12 12 2 12 2
GSL 2 2 2 2 12 12 12 1 12 12 12 12
MGB 12 12 2 1 1 1 1 1 1
MAB 12 12 2 1 12 1 1 12 12 2
GL 2 2 12 12 12 1 12 12 12 12
GB 12 12 12 1 12 12 2 12 2 12 2
GH 12 12 12 1 1 12 2 1 12 12 12 12
OGH 12 12 1 1 1 1 1 1 12 12 12
SMR 12 12 12 1 1 2 12 12 1 12 12 12
RAR 2 2 12 12 2 12 12 12 12 12
SML 12 12 12 1 1 12 12 12 12 12 12 12
RAL 2 2 12 12 2 12 12 12 12 12
SL: Gesamtschädellänge, SBL: Schädelbasislänge, GSL: Gesichtslänge, MGB:
Mittelgesichtsbreite, MAB: Maxilloalveolarbreite, GL: Gaumenlänge, GB:
Gaumenbreite, GH: Gaumenhöhe, OGH: Obergesichtshöhe, SMR: Volumen des
rechten Sinus maxillaris, RAR: Volumen des rechten Recessus alveolaris, SML:
Volumen des linken Sinus maxillaris, RAL: Volumen des linken Recessus
alveolaris.
1: Korrelation des Variablenpaares innerhalb der Altweltaffen
2: Korrelation des Variablenpaares innerhalb der Neuweltaffen
Für die korrelierenden Merkmalspaare der entsprechenden Affengruppe wurde
zudem eine partielle Korrelationsanalyse durchgeführt, um den Einfluss einer
dritten Messgröße auf das Wertepaar zu prüfen. Die partielle
Korrelationsanalyse wurde zum einen bei konstanter Gesamtschädellänge und

Ergebnisse
56
zum anderen bei konstanter Gesichtslänge getestet. In den Tabellen 8 und 9
sind die entsprechenden Ergebnisse für Altweltaffen und Neuweltaffen
dargestellt. Bei den Altweltaffen stellte sich kein Einfluss der
Gesamtschädellänge auf das Abhängigkeitsmerkmal der Merkmalspaare von
Recessus alveolaris rechts und Sinus maxillaris rechts, Recessus alveolaris
rechts und Obergesichtshöhe sowie Recessus alveolaris und Gaumenhöhe
heraus. Auf alle weiteren Merkmalspaare konnte ein Einfluss der
Gesamtschädellänge nachgewiesen werden. Die partiellen
Korrelationskoeffizienten fallen kleiner aus als die Korrelationskoeffizienten nach
Pearson. Durchaus Auswirkung zeigte die Gesichtslänge auf die untersuchten
Merkmalspaare bei der partiellen Korrelationsanalyse. Die Korrelationsindizes für
die partielle Korrelation fielen für alle Merkmalspaare geringer aus (Tab. 8).
Bei der partiellen Korrelationsanalyse des rechten Recessus alveolaris der
Neuweltaffen zeigte sich, dass die Gesamtschädellänge keinen Einfluss auf das
Abhängigkeitsverhältnis von Recessus alveolaris rechts und Sinus maxillaris
rechts, Recessus alveolaris rechts und Sinus maxillaris links sowie Recessus
alveolaris rechts und Obergesichtshöhe hat. Die Merkmalspaare von Recessus
alveolaris und Sinus maxillaris rechts sowie Recessus alveolaris rechts und
Obergesichtshöhe wurden genauso wenig von der Gesichtslänge beeinflusst.
Hinsichtlich der anderen in Tabelle 9 aufgeführten Merkmalspaare zeigte sich ein
Einfluss von Gesichtslänge und Gesamtschädellänge auf das
Abhängigkeitsverhältnis. Die entsprechenden Signifikanzniveaus sind Tabelle 9
zu entnehmen.
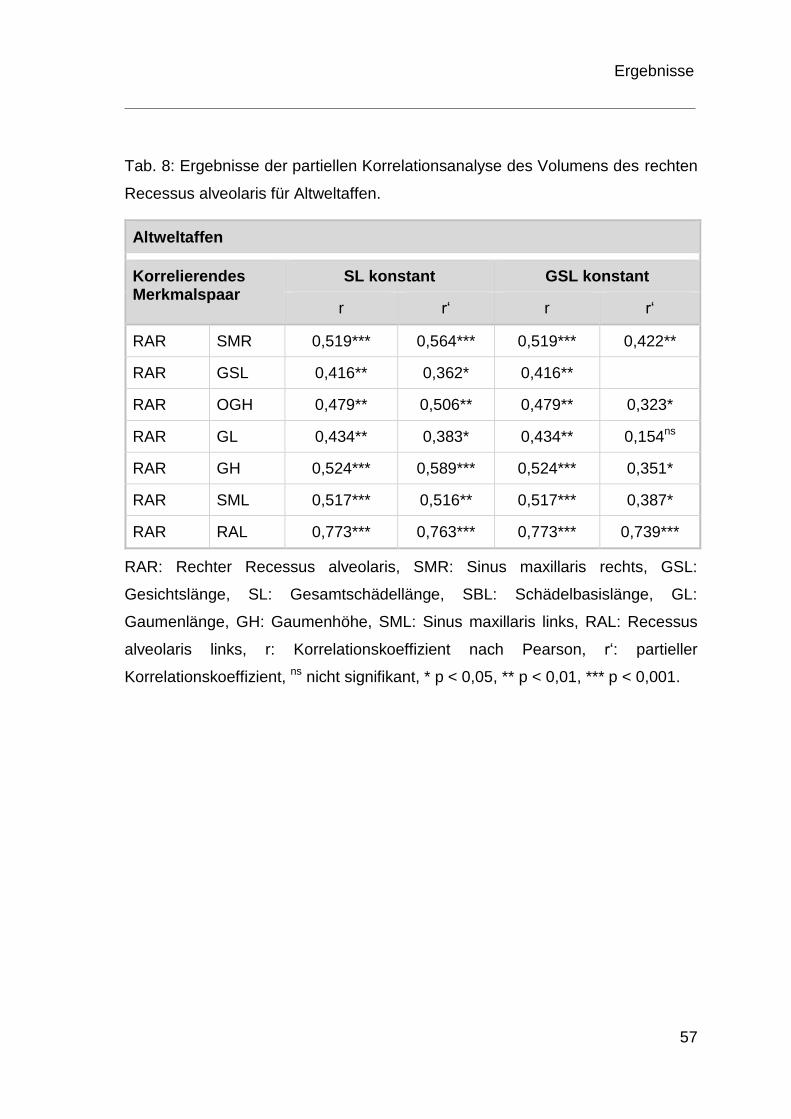
Ergebnisse
57
Tab. 8: Ergebnisse der partiellen Korrelationsanalyse des Volumens des rechten
Recessus alveolaris für Altweltaffen.
Altweltaffen
Korrelierendes Merkmalspaar
SL konstant GSL konstant
r r‘ r r‘
RAR SMR 0,519*** 0,564*** 0,519*** 0,422**
RAR GSL 0,416** 0,362* 0,416**
RAR OGH 0,479** 0,506** 0,479** 0,323*
RAR GL 0,434** 0,383* 0,434** 0,154ns
RAR GH 0,524*** 0,589*** 0,524*** 0,351*
RAR SML 0,517*** 0,516** 0,517*** 0,387*
RAR RAL 0,773*** 0,763*** 0,773*** 0,739***
RAR: Rechter Recessus alveolaris, SMR: Sinus maxillaris rechts, GSL:
Gesichtslänge, SL: Gesamtschädellänge, SBL: Schädelbasislänge, GL:
Gaumenlänge, GH: Gaumenhöhe, SML: Sinus maxillaris links, RAL: Recessus
alveolaris links, r: Korrelationskoeffizient nach Pearson, r‘: partieller
Korrelationskoeffizient, ns nicht signifikant, * p < 0,05, ** p < 0,01, *** p < 0,001.
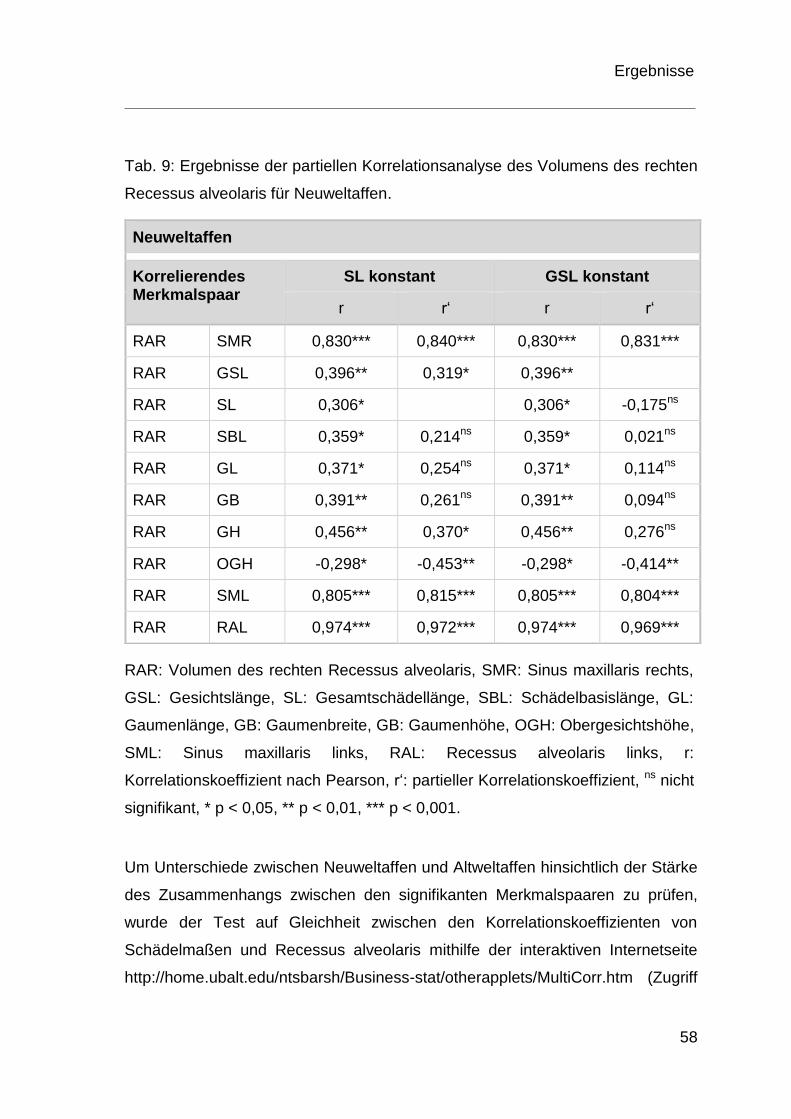
Ergebnisse
58
Tab. 9: Ergebnisse der partiellen Korrelationsanalyse des Volumens des rechten
Recessus alveolaris für Neuweltaffen.
Neuweltaffen
Korrelierendes Merkmalspaar
SL konstant GSL konstant
r r‘ r r‘
RAR SMR 0,830*** 0,840*** 0,830*** 0,831***
RAR GSL 0,396** 0,319* 0,396**
RAR SL 0,306* 0,306* -0,175ns
RAR SBL 0,359* 0,214ns 0,359* 0,021ns
RAR GL 0,371* 0,254ns 0,371* 0,114ns
RAR GB 0,391** 0,261ns 0,391** 0,094ns
RAR GH 0,456** 0,370* 0,456** 0,276ns
RAR OGH -0,298* -0,453** -0,298* -0,414**
RAR SML 0,805*** 0,815*** 0,805*** 0,804***
RAR RAL 0,974*** 0,972*** 0,974*** 0,969***
RAR: Volumen des rechten Recessus alveolaris, SMR: Sinus maxillaris rechts,
GSL: Gesichtslänge, SL: Gesamtschädellänge, SBL: Schädelbasislänge, GL:
Gaumenlänge, GB: Gaumenbreite, GB: Gaumenhöhe, OGH: Obergesichtshöhe,
SML: Sinus maxillaris links, RAL: Recessus alveolaris links, r:
Korrelationskoeffizient nach Pearson, r‘: partieller Korrelationskoeffizient, ns nicht
signifikant, * p < 0,05, ** p < 0,01, *** p < 0,001.
Um Unterschiede zwischen Neuweltaffen und Altweltaffen hinsichtlich der Stärke
des Zusammenhangs zwischen den signifikanten Merkmalspaaren zu prüfen,
wurde der Test auf Gleichheit zwischen den Korrelationskoeffizienten von
Schädelmaßen und Recessus alveolaris mithilfe der interaktiven Internetseite
http://home.ubalt.edu/ntsbarsh/Business-stat/otherapplets/MultiCorr.htm (Zugriff
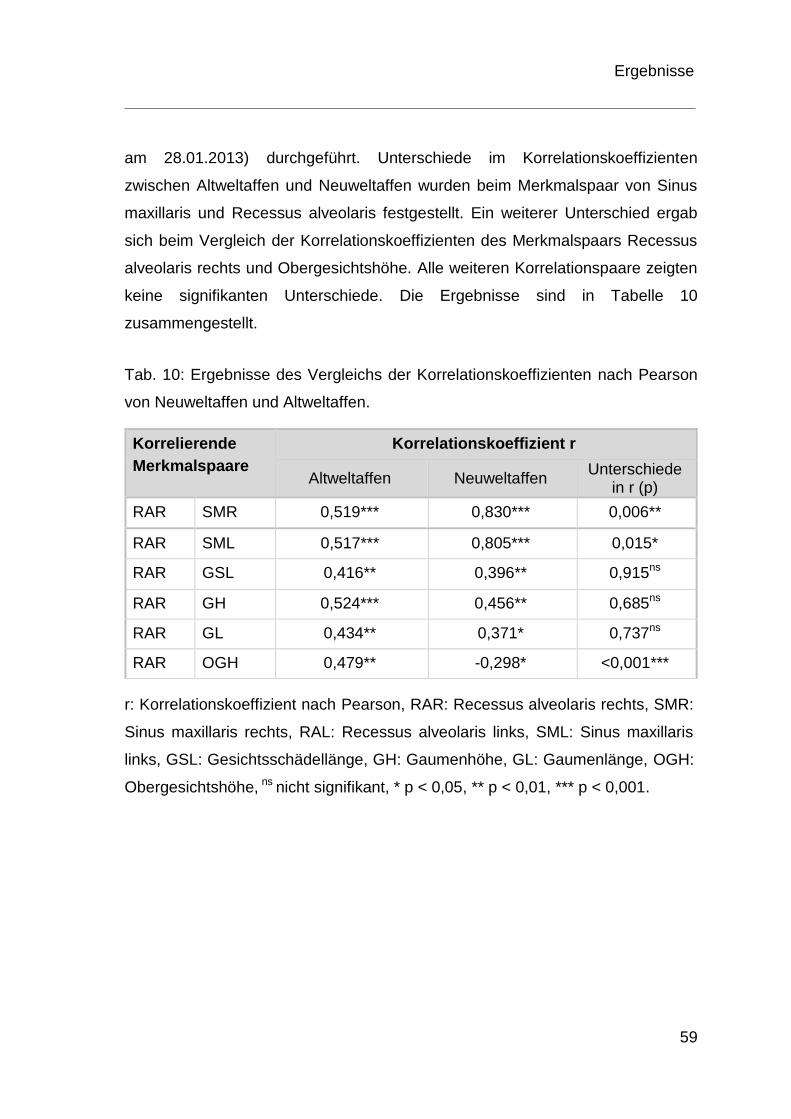
Ergebnisse
59
am 28.01.2013) durchgeführt. Unterschiede im Korrelationskoeffizienten
zwischen Altweltaffen und Neuweltaffen wurden beim Merkmalspaar von Sinus
maxillaris und Recessus alveolaris festgestellt. Ein weiterer Unterschied ergab
sich beim Vergleich der Korrelationskoeffizienten des Merkmalspaars Recessus
alveolaris rechts und Obergesichtshöhe. Alle weiteren Korrelationspaare zeigten
keine signifikanten Unterschiede. Die Ergebnisse sind in Tabelle 10
zusammengestellt.
Tab. 10: Ergebnisse des Vergleichs der Korrelationskoeffizienten nach Pearson
von Neuweltaffen und Altweltaffen.
Korrelierende
Merkmalspaare
Korrelationskoeffizient r
Altweltaffen Neuweltaffen Unterschiede
in r (p)
RAR SMR 0,519*** 0,830*** 0,006**
RAR SML 0,517*** 0,805*** 0,015*
RAR GSL 0,416** 0,396** 0,915ns
RAR GH 0,524*** 0,456** 0,685ns
RAR GL 0,434** 0,371* 0,737ns
RAR OGH 0,479** -0,298* <0,001***
r: Korrelationskoeffizient nach Pearson, RAR: Recessus alveolaris rechts, SMR:
Sinus maxillaris rechts, RAL: Recessus alveolaris links, SML: Sinus maxillaris
links, GSL: Gesichtsschädellänge, GH: Gaumenhöhe, GL: Gaumenlänge, OGH:
Obergesichtshöhe, ns nicht signifikant, * p < 0,05, ** p < 0,01, *** p < 0,001.
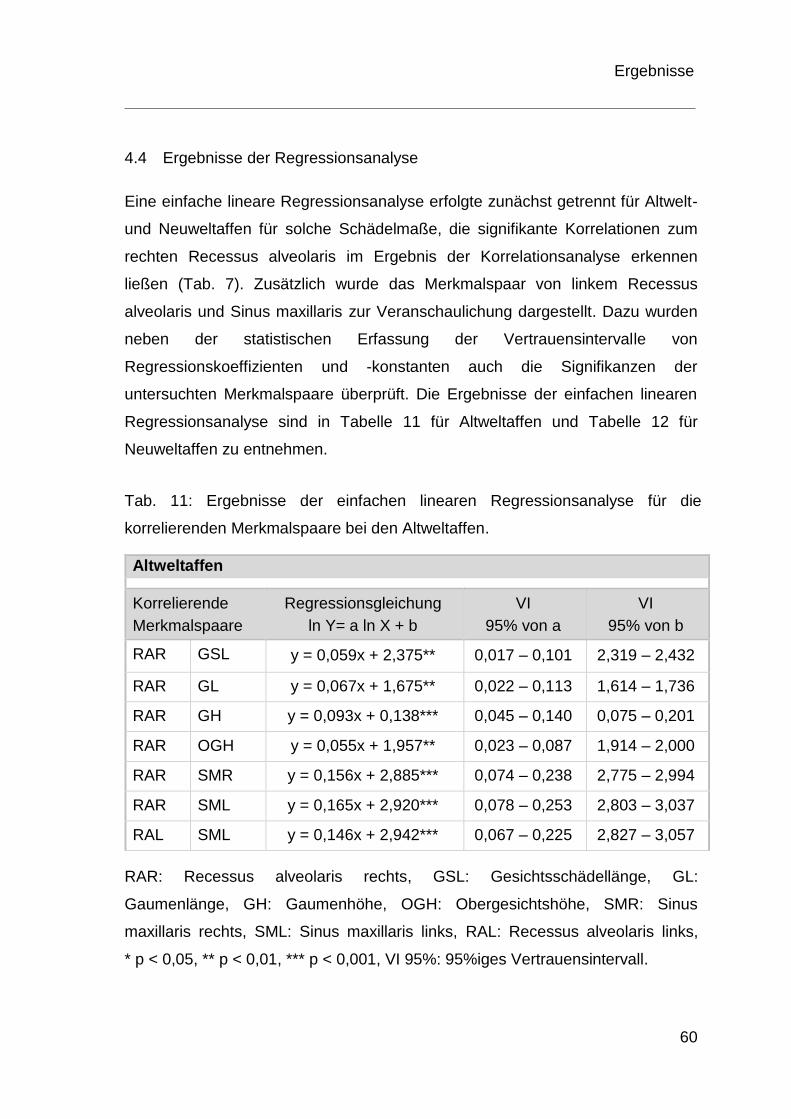
Ergebnisse
60
4.4 Ergebnisse der Regressionsanalyse
Eine einfache lineare Regressionsanalyse erfolgte zunächst getrennt für Altwelt-
und Neuweltaffen für solche Schädelmaße, die signifikante Korrelationen zum
rechten Recessus alveolaris im Ergebnis der Korrelationsanalyse erkennen
ließen (Tab. 7). Zusätzlich wurde das Merkmalspaar von linkem Recessus
alveolaris und Sinus maxillaris zur Veranschaulichung dargestellt. Dazu wurden
neben der statistischen Erfassung der Vertrauensintervalle von
Regressionskoeffizienten und -konstanten auch die Signifikanzen der
untersuchten Merkmalspaare überprüft. Die Ergebnisse der einfachen linearen
Regressionsanalyse sind in Tabelle 11 für Altweltaffen und Tabelle 12 für
Neuweltaffen zu entnehmen.
Tab. 11: Ergebnisse der einfachen linearen Regressionsanalyse für die
korrelierenden Merkmalspaare bei den Altweltaffen.
Altweltaffen
Korrelierende
Merkmalspaare
Regressionsgleichung
ln Y= a ln X + b
VI
95% von a
VI
95% von b
RAR GSL y = 0,059x + 2,375** 0,017 – 0,101 2,319 – 2,432
RAR GL y = 0,067x + 1,675** 0,022 – 0,113 1,614 – 1,736
RAR GH y = 0,093x + 0,138*** 0,045 – 0,140 0,075 – 0,201
RAR OGH y = 0,055x + 1,957** 0,023 – 0,087 1,914 – 2,000
RAR SMR y = 0,156x + 2,885*** 0,074 – 0,238 2,775 – 2,994
RAR SML y = 0,165x + 2,920*** 0,078 – 0,253 2,803 – 3,037
RAL SML y = 0,146x + 2,942*** 0,067 – 0,225 2,827 – 3,057
RAR: Recessus alveolaris rechts, GSL: Gesichtsschädellänge, GL:
Gaumenlänge, GH: Gaumenhöhe, OGH: Obergesichtshöhe, SMR: Sinus
maxillaris rechts, SML: Sinus maxillaris links, RAL: Recessus alveolaris links,
* p < 0,05, ** p < 0,01, *** p < 0,001, VI 95%: 95%iges Vertrauensintervall.
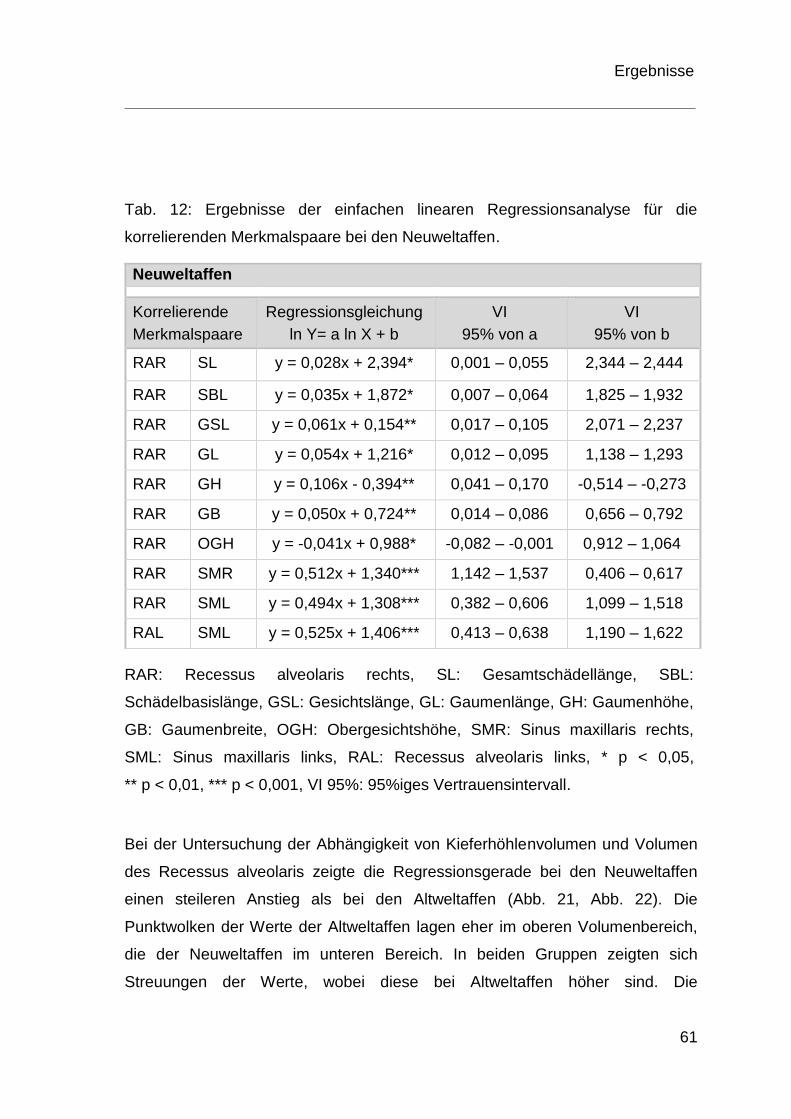
Ergebnisse
61
Tab. 12: Ergebnisse der einfachen linearen Regressionsanalyse für die
korrelierenden Merkmalspaare bei den Neuweltaffen.
Neuweltaffen
Korrelierende
Merkmalspaare
Regressionsgleichung
ln Y= a ln X + b
VI
95% von a
VI
95% von b
RAR SL y = 0,028x + 2,394* 0,001 – 0,055 2,344 – 2,444
RAR SBL y = 0,035x + 1,872* 0,007 – 0,064 1,825 – 1,932
RAR GSL y = 0,061x + 0,154** 0,017 – 0,105 2,071 – 2,237
RAR GL y = 0,054x + 1,216* 0,012 – 0,095 1,138 – 1,293
RAR GH y = 0,106x - 0,394** 0,041 – 0,170 -0,514 – -0,273
RAR GB y = 0,050x + 0,724** 0,014 – 0,086 0,656 – 0,792
RAR OGH y = -0,041x + 0,988* -0,082 – -0,001 0,912 – 1,064
RAR SMR y = 0,512x + 1,340*** 1,142 – 1,537 0,406 – 0,617
RAR SML y = 0,494x + 1,308*** 0,382 – 0,606 1,099 – 1,518
RAL SML y = 0,525x + 1,406*** 0,413 – 0,638 1,190 – 1,622
RAR: Recessus alveolaris rechts, SL: Gesamtschädellänge, SBL:
Schädelbasislänge, GSL: Gesichtslänge, GL: Gaumenlänge, GH: Gaumenhöhe,
GB: Gaumenbreite, OGH: Obergesichtshöhe, SMR: Sinus maxillaris rechts,
SML: Sinus maxillaris links, RAL: Recessus alveolaris links, * p < 0,05,
** p < 0,01, *** p < 0,001, VI 95%: 95%iges Vertrauensintervall.
Bei der Untersuchung der Abhängigkeit von Kieferhöhlenvolumen und Volumen
des Recessus alveolaris zeigte die Regressionsgerade bei den Neuweltaffen
einen steileren Anstieg als bei den Altweltaffen (Abb. 21, Abb. 22). Die
Punktwolken der Werte der Altweltaffen lagen eher im oberen Volumenbereich,
die der Neuweltaffen im unteren Bereich. In beiden Gruppen zeigten sich
Streuungen der Werte, wobei diese bei Altweltaffen höher sind. Die

Ergebnisse
62
Regressionsgerade von Recessus alveolaris rechts und Obergesichtshöhe
zeigte bei Neuweltaffen und Altweltaffen deutliche Unterschiede. Die
Regressionsgeraden verlaufen scherenartig (Abb. 26). Dies zeigte sich bereits in
der Korrelationsanalyse durch die negative Korrelation bei Neuweltaffen und die
positive Korrelation bei Altweltaffen. Für die Merkmalspaare Recessus alveolaris
und Gesichtslänge, Gaumenlänge sowie Gaumenhöhe stellte sich ein ähnlicher
Zusammenhang von Neuweltaffen und Altweltaffen dar, was durch den
parallelen Anstieg der Regressionsgeraden zu sehen ist (Abb. 23, Abb. 24,
Abb. 25). Die Punktwolken der Altweltaffen befanden sich auch bei diesen
Paaren eher im oberen Bereich. Eine breitere Streuung wiesen die Neuweltaffen
für diese drei Merkmalspaare auf.
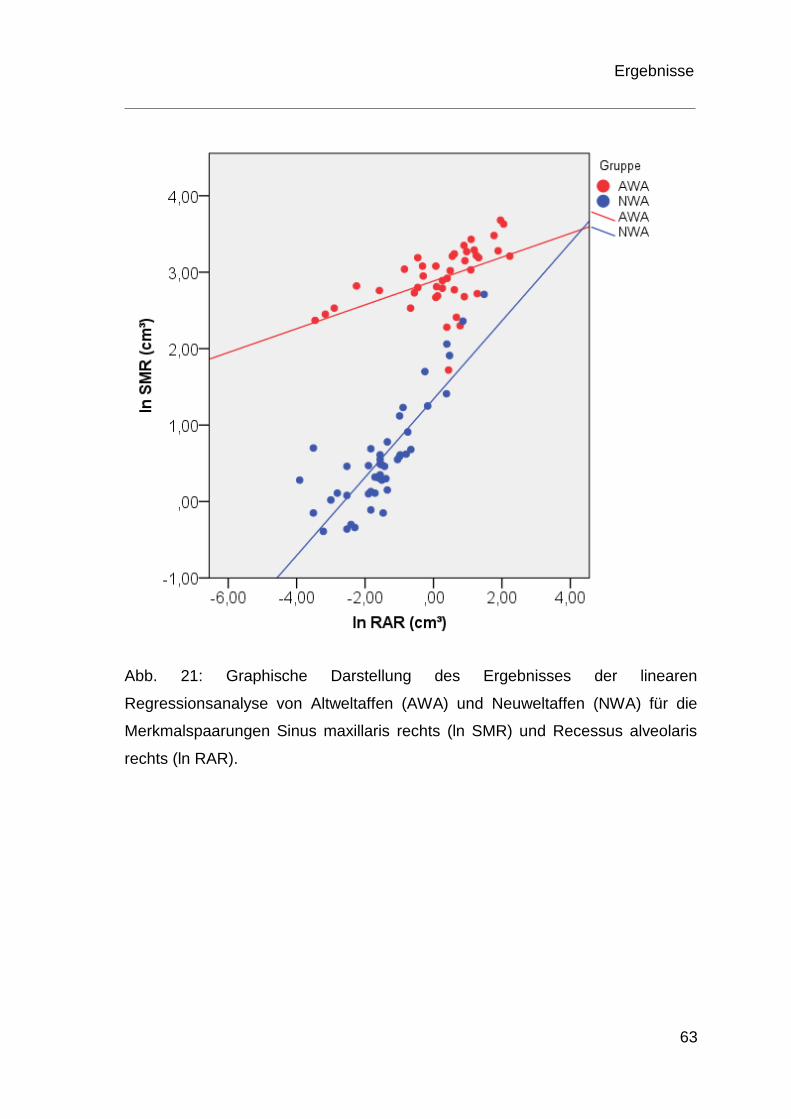
Ergebnisse
63
Abb. 21: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarungen Sinus maxillaris rechts (ln SMR) und Recessus alveolaris
rechts (ln RAR).
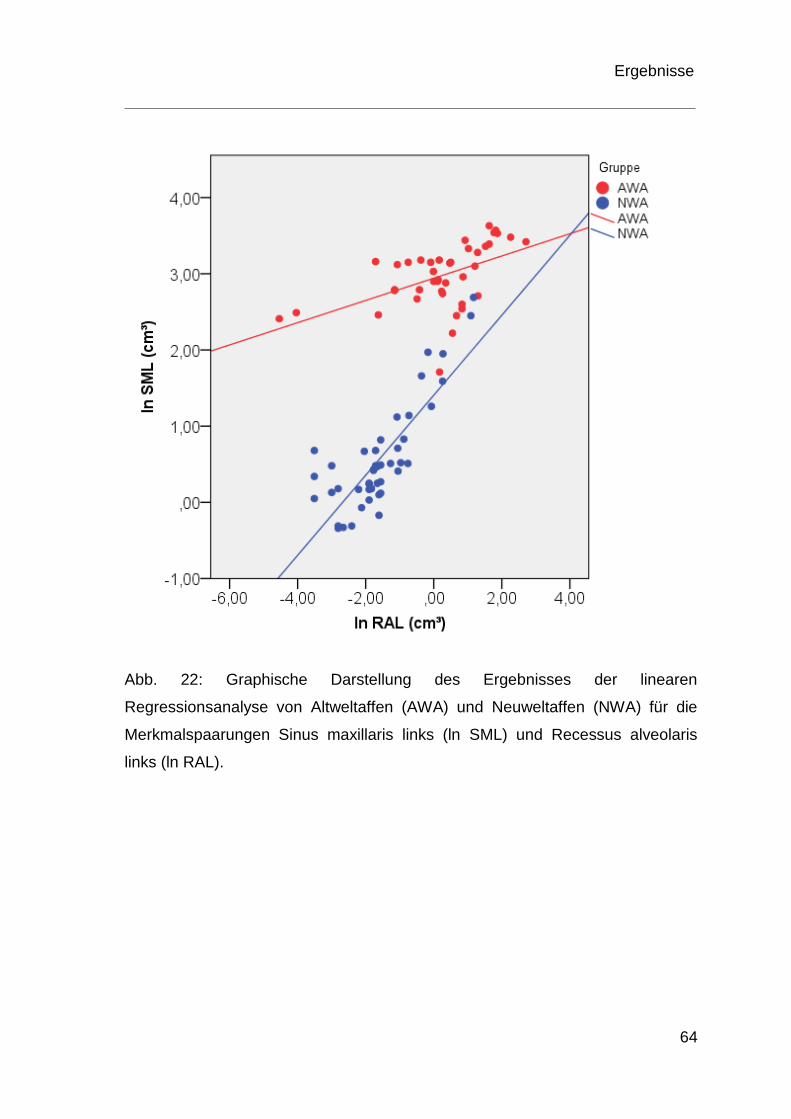
Ergebnisse
64
Abb. 22: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarungen Sinus maxillaris links (ln SML) und Recessus alveolaris
links (ln RAL).
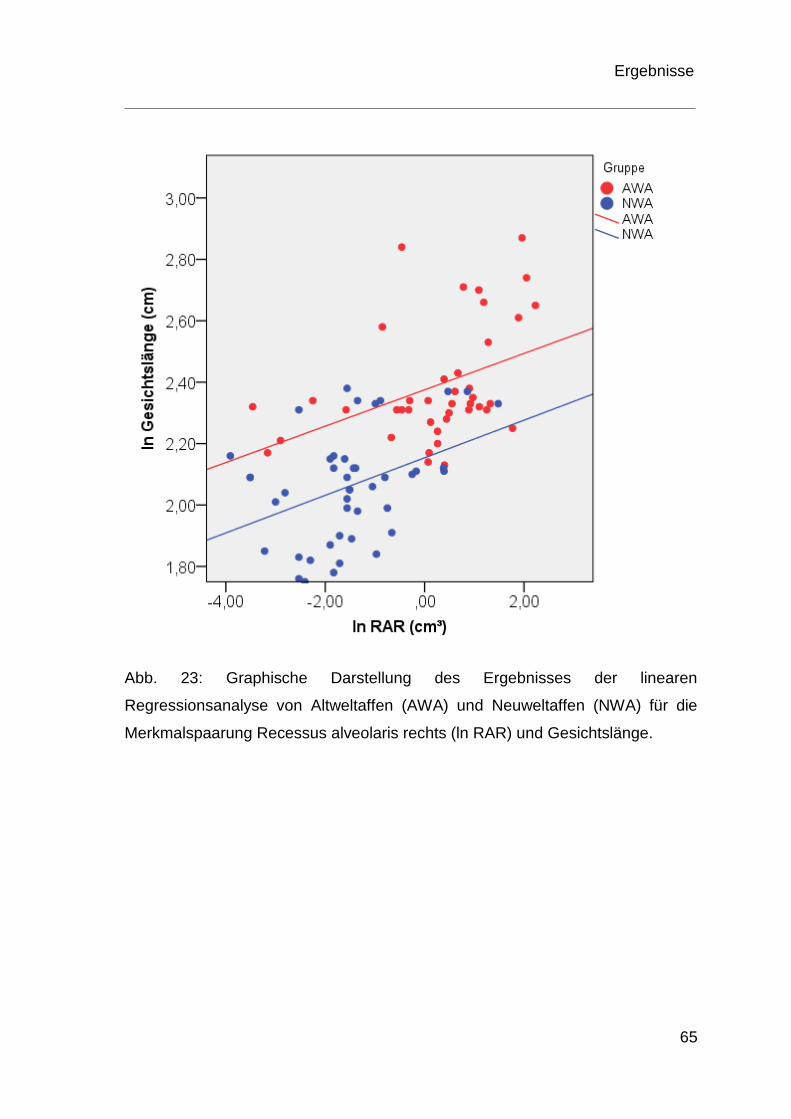
Ergebnisse
65
Abb. 23: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarung Recessus alveolaris rechts (ln RAR) und Gesichtslänge.
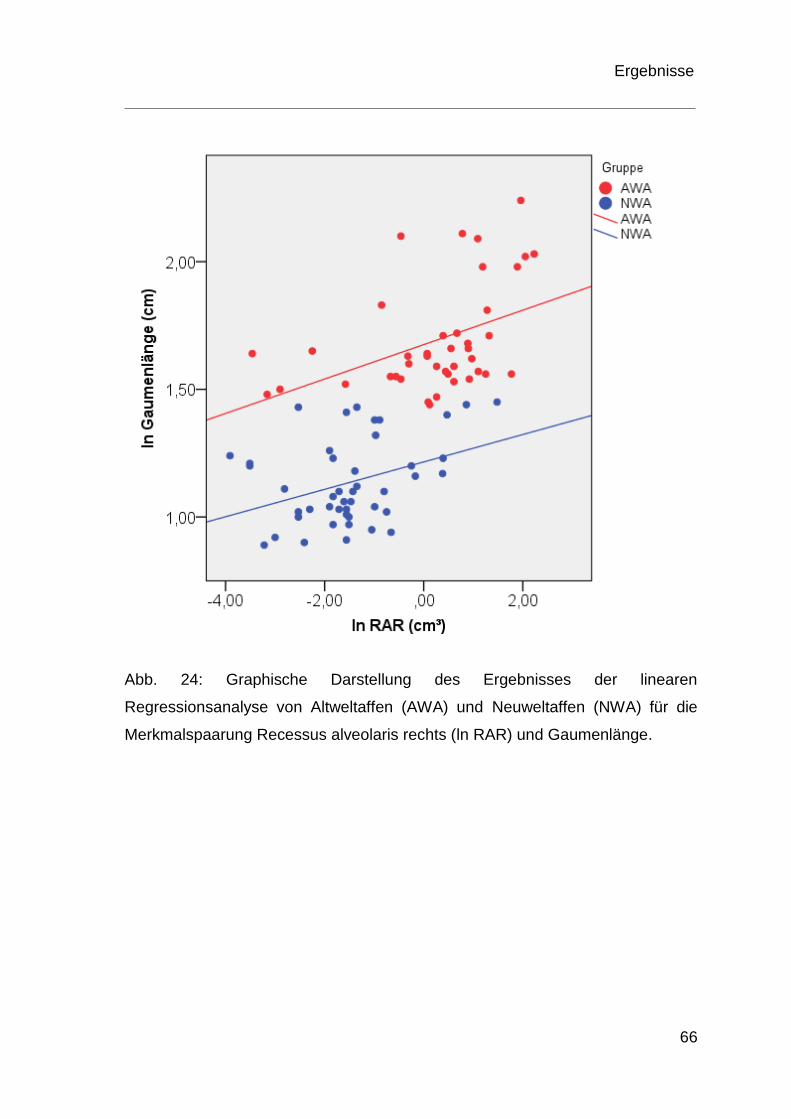
Ergebnisse
66
Abb. 24: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarung Recessus alveolaris rechts (ln RAR) und Gaumenlänge.
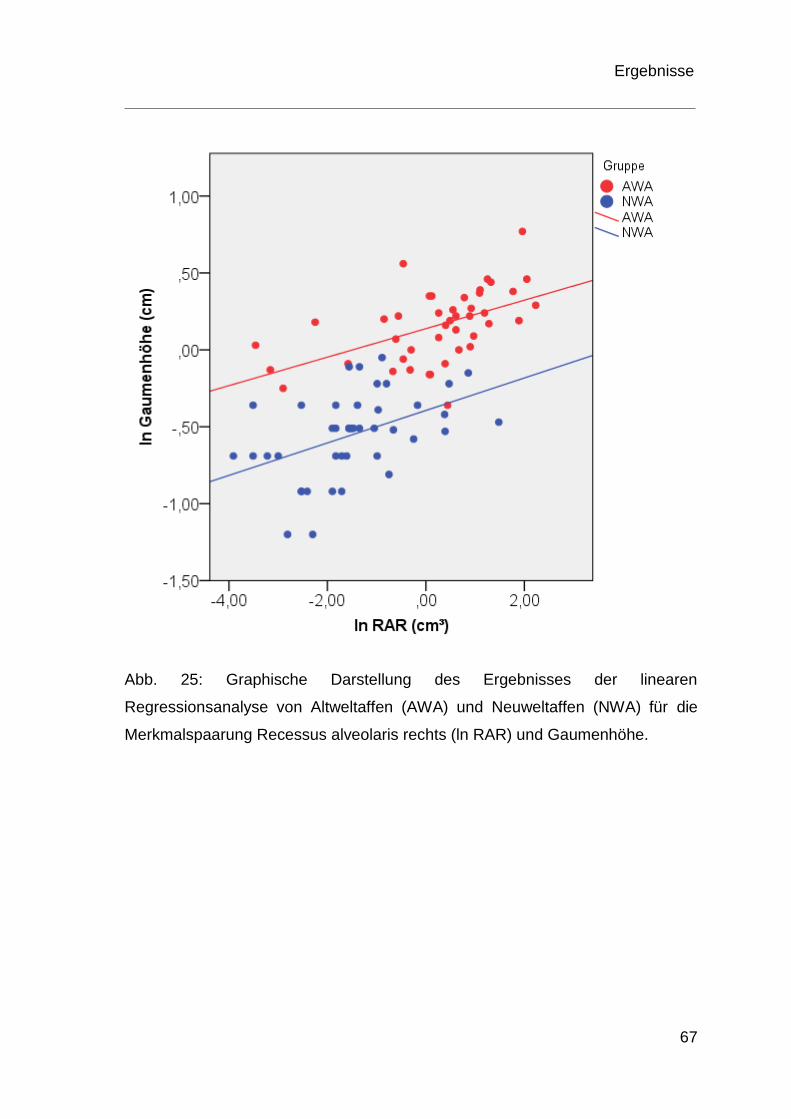
Ergebnisse
67
Abb. 25: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarung Recessus alveolaris rechts (ln RAR) und Gaumenhöhe.
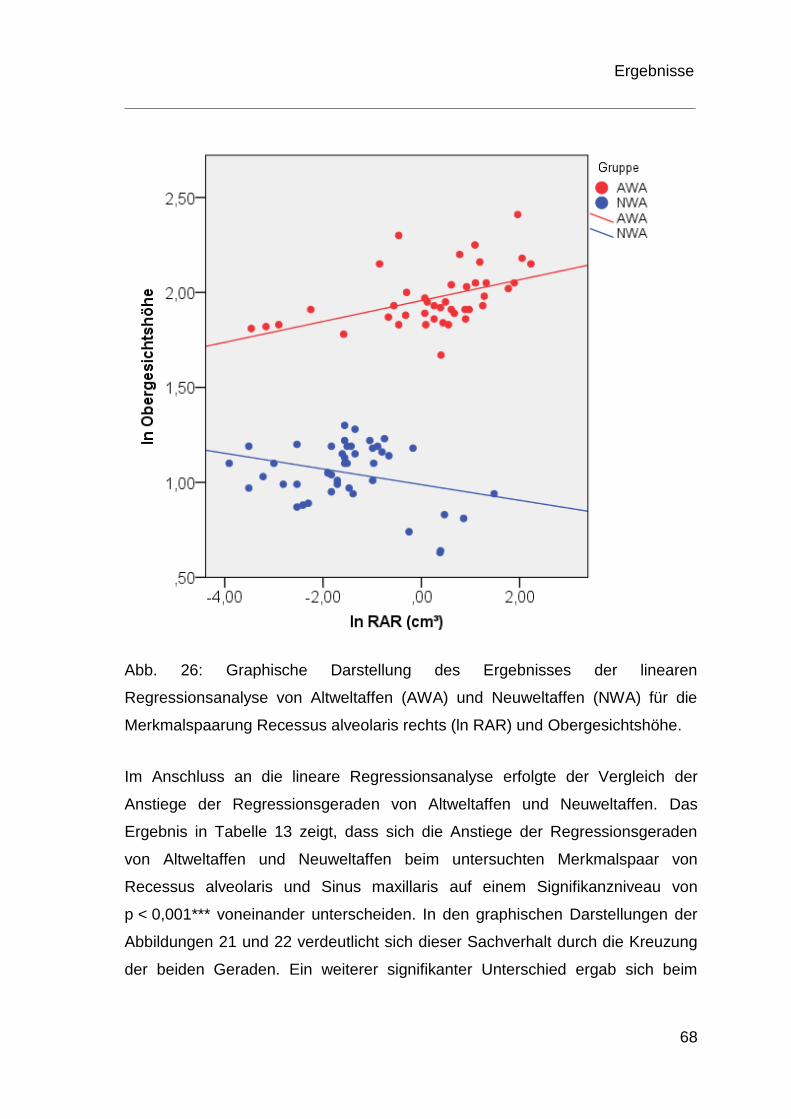
Ergebnisse
68
Abb. 26: Graphische Darstellung des Ergebnisses der linearen
Regressionsanalyse von Altweltaffen (AWA) und Neuweltaffen (NWA) für die
Merkmalspaarung Recessus alveolaris rechts (ln RAR) und Obergesichtshöhe.
Im Anschluss an die lineare Regressionsanalyse erfolgte der Vergleich der
Anstiege der Regressionsgeraden von Altweltaffen und Neuweltaffen. Das
Ergebnis in Tabelle 13 zeigt, dass sich die Anstiege der Regressionsgeraden
von Altweltaffen und Neuweltaffen beim untersuchten Merkmalspaar von
Recessus alveolaris und Sinus maxillaris auf einem Signifikanzniveau von
p < 0,001*** voneinander unterscheiden. In den graphischen Darstellungen der
Abbildungen 21 und 22 verdeutlicht sich dieser Sachverhalt durch die Kreuzung
der beiden Geraden. Ein weiterer signifikanter Unterschied ergab sich beim

Ergebnisse
69
Vergleich der Regressionsgeraden von Obergesichtshöhe und Recessus
alveolaris bei Neuweltaffen und Altweltaffen.
Ein anderer Sachverhalt zeigt sich hingegen bei der Analyse der in Tabelle 13
aufgeführten weiteren Merkmalspaare. Die Punkt-/Streudiagramme in den
Abbildungen 23, 24 und 25 lassen einen parallelen Verlauf der
Regressionsgeraden erkennen. Es stellten sich keine signifikanten Unterschiede
beim Vergleich der Anstiege heraus. Die Zusammenhänge von Recessus
alveolaris und Gesichtslänge (Abb. 23), Gaumenlänge (Abb. 24) und
Gaumenhöhe (Abb. 25) sind bei Neuweltaffen und Altweltaffen somit ähnlich.
Tab. 13: Ergebnisse des Vergleichs der linearen Regressionsgeraden von
Neuweltaffen und Altweltaffen
Korrelierende
Merkmalspaare
Regressionsgleichung
Altweltaffen Neuweltaffen p
RAR SMR y = 0,156x - 2,885 y = 0,512x + 1,340 <0,001***
RAL SML y = 0,146x + 2,942 y = 0,525x + 1,406 <0,001***
RAR GSL y = 0,059x + 2,375 y = 0,061x + 0,154 0,940ns
RAR GH y = 0,093x + 0,138 y = 0,106x - 0,394 0,715ns
RAR GL y = 0,067x + 1,675 y = 0,054x + 1,216 0,639ns
RAR OGH y = 0,055x + 1,957 y = -0,041x + 0,988 <0,001***
RAR: Recessus alveolaris rechts, SMR: Sinus maxillaris rechts, RAL: Recessus
alveolaris links, SML: Sinus maxillaris links, GSL: Gesichtsschädellänge, GH:
Gaumenhöhe, GL: Gaumenlänge, OGH: Obergesichtshöhe, ns nicht signifikant,
* p < 0,05, ** p < 0,01, *** p < 0,001.

Diskussion
70
5 Diskussion
Unsere Arbeit hatte zum Ziel, die Pneumatisation des Processus alveolaris der
Maxilla an Schädeln von Neuweltaffen und Altweltaffen zu vermessen und zu
beschreiben. Darüber hinaus interessierten uns mögliche Zusammenhänge
zwischen der Größe des Recessus alveolaris und der Schädelmorphologie.
Wir definierten den Recessus alveolaris als den Teil der Kieferhöhle, der
unterhalb einer definierten Bezugsebene durch den tiefsten Punkt des
Nasenbodens liegt. Dieses Vorgehen verwendeten bereits Koppe and Rae
(2001), Oesterreich et al. (2005) und Koppe et al. (2005b). Unserer Meinung
nach eignete sich der tiefste Punkt des Nasenbodens für unsere Studie gut, um
in jeder CT-Schicht einen eindeutigen Ausgangspunkt für die
Volumenvermessungen zu haben. Den Vorteil dieser Vorgehensweise sahen wir
darin, dass die Vergleichbarkeit und die genaue Rekonstruktion dieser
Bezugslinie auch bei unterschiedlichen Spezies möglich sind.
In der uns zur Verfügung stehenden Literatur fanden wir nur eine Studie, in der
das Volumen des Recessus alveolaris an Schädeln von Neuweltaffen
vermessen wurde (Oesterreich et al. 2005). Zum Volumen des Recessus
alveolaris bei Altweltaffen gibt es in der Literatur keine vergleichbaren
quantitativen Daten. Andere Studien untersuchten die Tiefe des Recessus
alveolaris an Schädeln von Menschen (Koppe et al. 2005b) und Makaken
(Koppe and Rae 2001). Diese Studien erfassten den Recessus alveolaris als
zweidimensionale Größe. Unserer Meinung nach ist die dreidimensionale
Vermessung, wie in der vorliegenden Studie, unabdinglich, um die Ausdehnung
der Pneumatisation der Sinus maxillaris in den Processus alveolaris maxillae
und den Recessus alveolaris als Teilvolumen der Kieferhöhle verstehen zu
können.
Zwei Auswahlkriterien definierten wir vor Beginn der Studie. Unser primäres
Auswahlkriterium war das Vorhandensein einer permanenten Dentition. Da der
Zahndurchbruch und der Stand der Dentition maßgeblich die Morphologie und

Diskussion
71
die Ausdehnung der Kieferhöhlen beeinflussen (Schaeffer 1920), bezogen wir
nur solche Schädel in die Untersuchung ein, die ein vollständiges permanentes
Gebiss aufwiesen. Mit der permanenten Dentition erreicht die Kieferhöhle beim
Menschen im Alter von 12 bis 15 Jahren ihre maximale Größe (Schumacher
1971). Durch dieses Kriterium konnten wir sicherstellen, dass trotz des
unbekannten Alters der Schädel, nur Schädel mit einer ausgewachsenen
Kieferhöhle vermessen wurden. Ein sekundäres Auswahlkriterium war das
Vorhandensein einer vollständigen Zahnreihe.
Die Morphologie der Kieferhöhle unterliegt erheblichen Altersschwankungen, die
sich insbesondere auch nach Zahnverlust in einer Zunahme der Pneumatisation
des Recessus alveolaris manifestiert (Sharan and Madjar 2008). Dies entspricht
auch klinischen Erfahrungen. Dennoch zeigen Untersuchungen, dass der
Einfluss des Zahnverlustes auf die Morphologie der Kieferhöhle unterschiedlich
bewertet wird. Während Ariji et al. (1994) keinen signifikanten Unterschied
bezüglich des Kieferhöhlenvolumens bei Patienten mit vorhandenen oder
fehlenden Prämolaren oder Molaren feststellen, nimmt die basale
Pneumatisation der Kieferhöhle nach Sharan and Madjar (2008) insbesondere
nach Extraktion der ersten und/oder zweiten Molaren zu. Die letztgenannte
Studie untersuchte die Dimension der Kieferhöhle allerdings anhand von
Panoramaschichtaufnahmen. Der Nachteil dieses Verfahrens liegt unter
anderem darin, dass nur eine Schicht dargestellt wird und dass sich bedingt
durch den unterschiedlichen Abstand von Fokus, Objekt und Film bei der
Rotation während der Röntgenaufnahme abweichende Vergrößerungsfaktoren
ergeben (Zäschke und Schopf 1975; Nötzel und Schultz 2009). Somit sind
exakte Vermessungen nicht verlässlich durchzuführen (Zäschke und Schopf
1975; Al Bourney 1999; Nohadani and Ruf 2008). Die Studie von Ariji et al.
(1994) hingegen beruht auf der Vermessung des Kieferhöhlenvolumens anhand
von CT-Bildern, obgleich nicht die Pneumatisation des Processus alveolaris
speziell untersucht worden ist.

Diskussion
72
Um die mögliche Volumenzunahme des Recessus alveolaris durch Zahnverlust
als Störfaktor in unserer Studie auszuschließen, verwendeten wir nahezu nur
Schädel mit einer geschlossenen, vollständigen Zahnreihe. Lediglich bei einem
menschlichen Schädel fehlte der erste Molar. Bei acht weiteren fehlte der dritte
Molar uni- bzw. bilateral. Dies begründen wir damit, dass fehlende
Weisheitszahnanlagen beim Menschen keine Seltenheit sind und in
verschiedenen Bevölkerungsgruppen mit unterschiedlicher Prävalenz auftreten
(Keene 1965; Rozkovcová et al. 2004). Unsere Untersuchungen ergaben, dass
die Morphologie des Kieferhöhlenbodens intra- und interspezifisch eine große
Variabilität aufzeigt. Diese Variabilität des Recessus alveolaris wurde bereits in
der Literatur unter anderem an Schädeln von Neuweltaffen (u.a. Nishimura et al.
2005; Oesterreich et al. 2005), von rezenten Hominoiden (Koppe and Ohkawa
1999; Koppe et al. 2005b) und von miozänen Hominoiden beschrieben (Ward
and Pilbeam 1983).
Beim Schimpansen und bei Syndactylus symphalangus zeigte der Boden des
Recessus alveolaris eine unregelmäßige Form. Dies entspricht auch den
Beobachtungen von Koppe and Ohkawa (1999). Beim Gorilla beschreiben
letztgenannte Autoren den Recessus alveolaris ebenfalls als unregelmäßig.
Unseren Untersuchungen zufolge ist der Recessus alveolaris eher
wannenförmig, da er sich zwischen die bukkalen und palatinalen Wurzeln der
Molaren erstreckte.
Bei den von uns untersuchten Orang-Utans beobachteten wir sowohl einen
flachen als auch einen wannenförmigen Recessus alveolaris. Im Gegensatz zum
Gorilla dehnte sich der Recessus alveolaris nicht in die Furkation der Molaren
aus. Wannenförmige Recessus alveolaris fanden wir auch bei Homo sapiens,
wobei allerdings auch unregelmäßige Formen vorkamen. Letztgenannte Formen
zeichneten sich dadurch aus, dass sich insbesondere die ersten und zweiten
Molaren in den Boden vorwölbten. Dies deckt sich mit den Ergebnissen von
Koppe et al. (2005b). Deren Studie hat ergeben, dass bei ca. 50% der von ihnen

Diskussion
73
vermessenen Schädel einer Jomon und Kyoto-Population der
Kieferhöhlenboden durch die Wurzeln der ersten und zweiten Molaren
aufgewölbt war. Im Gegensatz dazu berichten Eberhardt et al. (1992) in lediglich
5% der untersuchten Fälle über Zahnwurzeln, die in die Kieferhöhle hineinragen.
Nogi and Kanazawa (2001) führen die unterschiedlichen Morphologien des
Kieferhöhlenbodens auf die Abstände der Zahnwurzeln vom Kieferhöhlenboden
zurück. Hierbei geht ein unregelmäßiger und flacher Boden des Recessus
alveolaris mit einer kürzeren Distanz zu den Zahnwurzeln einher. Koppe and
Ohkawa (1999) beschreiben einen unregelmäßigen Boden bei Primatenspezies,
deren Wurzeln der Oberkieferseitenzähne in engem Kontakt zum
Kieferhöhlenboden stehen.
Obgleich eine enge anatomische Beziehung zwischen Zahnwurzeln und
Recessus alveolaris besteht, lassen sich unseren Untersuchungen zufolge die
unterschiedlichen Morphologien des Kieferhöhlenbodens, insbesondere
innerhalb einer Spezies, nicht ausschließlich auf den Stand der Dentition
zurückführen. Diese Vermutung wird gestützt durch Untersuchungen von
Swindler (1999), der berichtet, dass selbst das Fehlen der gesamten
Kieferhöhle, wie bei einigen Vertretern der Cercopithecoidea, nicht allein auf die
Zahnmorphologie und damit auch auf die Dentition zurückzuführen ist.
In einer Studie von Rossie (2006) zu den Nasennebenhöhlen von Neuweltaffen
findet sich eine mögliche Erklärung im Zusammenhang mit der Morphologie des
Kieferhöhlenbodens. Rossie (2006) vermutet, dass die Ausdehnung der
Kieferhöhle nach lateral über die bukkalen Wurzeln der Seitenzähne maßgeblich
davon abhängt, wie viel Platz zwischen Orbita und Zahnwurzeln vorhanden ist.
Während bei Aoutus dieser Raum nicht vorhanden ist, existiert er bei Alouatta
und Callicebus (Rossie 2006). Der Recessus alveolaris stellt sich bei diesen
Spezies demzufolge breitflächig und zerklüftet dar und die Wurzeln der Molaren
projizieren sich in die Kieferhöhle hinein (Moormann 2004; Oesterreich et al.
2005; Nishimura et al. 2005; Rae and Koppe 2008). Dies konnten wir in unseren

Diskussion
74
Untersuchungen für Alouatta und Cebus bestätigen. Bei kleineren Neuweltaffen
wie Sanguinus, Callimico und Aoutus liegt zwar der Boden der Kieferhöhle unter
dem Niveau der Wurzelspitzen von Prämolaren und Molaren, aber lediglich
deren palatinale Wurzeln ragen in die Kieferhöhle hinein (Rossie 2006). Schiwal
(2005) vermutet gleichermaßen nach ihren Untersuchungen an Callithrix dass
ein tiefer Orbitaboden und lange Zahnwurzeln nur einen kleinen Raum für die
Pneumatisation zulassen.
Auch wenn dieser Sachverhalt eine mögliche Erklärung für die Pneumatisation
der Maxilla nach lateral und den unregelmäßigen Recessus alveolaris von
Neuweltaffen ist, lässt sich diese nach unseren Ergebnissen nicht auf die
Hominoiden übertragen. Wir stellten fest, dass sich die laterale Ausdehnung der
Sinus maxillaris nicht immer über die bukkalen Wurzeln ausdehnt, obwohl bei
den großen Menschenaffen und beim Menschen ausreichend Platz zwischen
Orbita und Kieferhöhle besteht. Weitere Untersuchungen hierzu können
möglicherweise Aufschluss geben.
Verschiedene Autoren nutzten die unterschiedlichen Morphologien des
Kieferhöhlenbodens, um eine Klassifikation für die gesamte Kieferhöhle
vorzunehmen. Demnach kann die Kieferhöhle beim Menschen in bis zu sechs
verschiedene Typen eingeteilt werden (Anagnostopoulou et al. 1991; Kim et al.
2002) Im Gegensatz dazu unterscheiden Nogi and Kanazawa (2001) nur drei
Typen: den runden, den flachen und den irregulären Typ. Diese von Nogi and
Kanazawa (2001) beschriebenen drei Typen konnten wir in ähnlicher Weise
auch bei Neuweltaffen und Altweltaffen feststellen. Eine weitere Unterteilung
scheint uns nicht sinnvoll.
Einen Nachteil dieser Klassifikation sehen wir darin, dass sie in erster Linie
anhand einer visuellen Beurteilung vorgenommen wird und sich keiner
kalibrierten Einteilung bedient. So stellen Nogi and Kanazawa (2001) den runden
Typ beim Menschen vor allem bei solchen Schädeln fest, bei denen die Zähne
einen größeren Abstand zum Kieferhöhlenboden haben im Gegensatz zu den

Diskussion
75
anderen beiden Typen. Wir fanden einen runden Typ vorwiegend bei Fällen, bei
denen sich der Recessus alveolaris in die Furkation der Oberkiefermolaren
ausbreitet, wie z.B. bei den meisten Schädeln des Menschen und des Gorillas.
Diese Form war jedoch in unseren Fällen eher wannenförmig als rund.
Einen weiteren Nachteil der Klassifikation von Nogi and Kanazawa (2001) sehen
wir darin, dass die Kieferhöhle keine konstante Morphologie besitzt. Beim
Menschen wird die maximale Ausdehnung der Kieferhöhle mit dem
Vorhandensein der vollständigen zweiten Dentition erreicht (Schumacher 1971).
Bereits ab dem 20. Lebensjahr soll sich das Volumen wieder verkleinern (Ariji et
al. 1994). Es ist also anzunehmen, dass nicht nur das Volumen der Kieferhöhle,
sondern auch die Morphologie des Recessus alveolaris und damit die
entsprechende Einteilung nach der Klassifikation im Laufe des Lebens variiert.
Inwieweit eine solche Klassifikation ein Ausdruck für das Ausmaß und die Größe
des Recessus alveolaris ist, können wir nicht beurteilen. Weitere Studien an
unterschiedlichen Spezies zu diesem Sachverhalt dazu können möglicherweise
Aufschluss geben.
Unsere Ergebnisse zeigten, dass das Volumen des Recessus alveolaris bei
Altweltaffen größer ist als bei Neuweltaffen. Unter den Neuweltaffen besaß
Alouatta das größte und Brachyteles das kleinste Volumen des Recessus
alveolaris. Unter den Altweltaffen zeigte der Gorilla das größte Volumen des
Recessus alveolaris rechts auf. Das kleinste Volumen besitzt der Orang-Utan.
Die Spezies Alouatta und der Orang-Utan fielen bei der Gesamtübersicht
besonders auf. Während Alouatta den größten Recessus alveolaris unter den
Neuweltaffen aufzeigte, wies der Orang-Utan den kleinsten Recessus alveolaris
unter den Altweltaffen auf.
Zunächst sollen zwei mögliche Aspekte für die Spezies Alouatta diskutiert
werden, welche den vergleichsweise großen Recessus alveolaris bedingen. Der
erste mögliche Grund für den ausgeprägten Recessus alveolaris bei Alouatta
besteht darin, dass bei dieser Spezies ein Recessus palatinus existiert, welcher

Diskussion
76
den harten Gaumen über die gesamte Länge der Kieferhöhle pneumatisiert
(Moormann 2004). Dieser Teil der Kieferhöhle erweitert die Kieferhöhle nach
anterior und liegt wie auch der Recessus alveolaris unter einer Bezugsebene
durch den Nasenboden. Somit war eine klare Abgrenzung zwischen diesen
Recessus nicht möglich, weswegen wir in unserer Studie den Recessus
palatinus gemeinsam mit dem Recessus alveolaris vermessen haben. Dieser
Sachverhalt trifft auch für den Schimpansen zu. Inwieweit das Volumen des
Recessus alveolaris durch den Anteil des Recessus palatinus vergrößert ist,
muss in weiteren Studien geprüft werden.
Eine zweite mögliche Erklärung für das große Volumen des Recessus alveolaris
von Alouatta lässt sich bei der Betrachtung der Stellung von Gesichts- und
Gehirnschädel dieser Spezies finden. Die Stellung der beiden Schädelanteile
kann über den Winkel zwischen Gaumenebene und der Klivusebene bestimmt
werden. Hofer (1952) prägte in diesem Zusammenhang die Begriffe der
Klinorhynchie und Airorhynchie. Bei einem kleinen Winkel wird von Klinorhnychie
gesprochen, bei einem großen Winkel von Airorhynchie. Die Klinorhynchie bei
Alouatta ist im Vergleich zu anderen Primaten gering ausgeprägt (Schumacher
et al. 1994). Untersuchungen von Moormann (2004) an Alouatta deuten darauf
hin, dass ein gering klinorhyncher Schädel mit einem vergrößerten Sinus
maxillaris in Verbindung gebracht werden kann. Übertragen wir diese Erklärung
auf den Recessus alveolaris, können wir für Alouatta den Rückschluss ziehen,
dass ein gering klinorhyncher, stark pneumatisierter Schädel zugleich mit einem
verhältnismäßig großen Recessus alveolaris einhergeht. Obgleich diese
Erklärung für Alouatta zutrifft, können wir sie nach unseren Ergebnissen nicht als
allgemeingültig betrachten.
Unter diesem Aspekt ist der Orang-Utan zu erwähnen, welcher unter den
Altweltaffen den kleinsten Recessus alveolaris aufzeigte. Der Orang-Utan besitzt
wie Alouatta einen gering klinorhynchen Schädel (Schumacher et al. 1994). Zwar
deutet eine Studie von Koppe et al. (1999a) darauf hin, dass ein gering

Diskussion
77
klinorhyncher Schädel beim Orang-Utan mit einem vergrößerten Sinus maxillaris
in Zusammenhang gebracht werden kann, aber die Ergebnisse unserer Studie
zeigten, dass der Orang-Utan unter den Altweltaffen ein kleines Volumen des
Recessus alveolaris besitzt. Somit können wir für den Orang-Utan nicht
bestätigen, dass ein gering klinorhyncher Schädel mit einem großen Recessus
alveolaris in Zusammenhang steht.
Eine andere mögliche Ursache für den kleinen Recessus alveolaris des Orang-
Utan ergibt sich aus der anterioren Ausdehnung der Sinus maxillaris. So
vermuten Ward and Pilbeam (1983), dass eine tiefe Fossa canina, wie sie beim
Orang-Utan zu finden ist, die Pneumatisation der Kieferhöhle nach anterior über
die Molaren hinaus hindert. Hingegen pneumatisiert die Kieferhöhle des Orang-
Utans über den Tuber maxillae hinaus und reicht nach posterior bis an die Sinus
sphenoidales (Koppe and Ohkawa 1999). Wir vermuten, dass die posteriore
Pneumatisation der Kieferhöhle in Zusammenhang mit dem kleinen Volumen
des Recessus alveolaris steht. In unseren Untersuchungen lag der Recessus
alveolaris beim Orang-Utan erst auf Höhe des zweiten Molaren unterhalb des
Nasenbodens und begründet somit möglicherweise sein kleines Volumen.
Aufgrund der geringen Speziesanzahl ist dieser Sachverhalt jedoch an weiteren
Schädeln zu prüfen.
Obgleich unsere Untersuchungsergebnisse zeigten, dass die Ausbildung eines
Recessus alveolaris ein typisches Merkmal des Sinus maxillaris der Primaten ist,
stellte sich heraus, dass es auch einzelne Schädel gibt, bei denen der
Nasenboden auf einer Ebene mit dem Kieferhöhlenboden liegt. Dies war in
unseren Untersuchungen bei einem Schädel von Homo sapiens der Fall, bei
dem auf der linken Seite der Boden der Kieferhöhle auf einer Ebene mit dem
Nasenboden lag und somit nach unserer Definition kein Recessus alveolaris
existierte. Mit Blick auf die bestehende Literatur ist diese Beobachtung nicht
verwunderlich. Schumacher (1971) beschreibt, dass der Kieferhöhlenboden
beim Menschen in der Regel auf einer Ebene mit dem Nasenboden liegt, bei

Diskussion
78
Menschen mit niedrigem Mittelgesicht sogar oberhalb des Nasenbodens. Auch
in der Studie von Oesterreich et al. (2005) zeigt sich, dass der Recessus
alveolaris unterhalb einer gedachten Bezugslinie durch den Nasenboden bei
Neuweltaffen in einigen Fällen nicht immer vorhanden ist (Oesterreich et al.
2005).
Unter diesem Gesichtspunkt muss erwähnt werden, dass auch bei Macaca
fuscata zu beobachten ist, dass der Boden der Kieferhöhle bei vielen Schädeln
oberhalb einer Bezugsebene durch den Nasenboden liegt (Koppe and Rae
2001). Letztgenannte Autoren zeigen in ihrer Studie, dass Macaca fuscata ein
kleines Kieferhöhlenvolumen im Vergleich zu anderen Makakenarten besitzt.
Rae et al. (1997) diskutieren, dass ein kleines Kieferhöhlenvolumen von Macaca
fuscata mit kalten Temperaturen in Zusammenhang steht. Je kälter die
Temperaturen sind, denen die Primaten ausgesetzt sind, umso kleiner ist das
Kieferhöhlenvolumen (Rae et al. 1997). Gleichermaßen beschreibt Shea (1977)
diesen Zusammenhang auch bei Eskimoschädeln. Ob und inwiefern jedoch ein
kleines Kieferhöhlenvolumen, bedingt durch kalte Temperaturen, eine Erklärung
für die nicht vorhandene Pneumatisation des Processus alveolaris ist, muss in
weiteren Studien für verschiedene Spezies geprüft werden.
Tiefe des Recessus alveolaris
Wir beobachteten, dass die tiefste Stelle bei den Altweltaffen vorwiegend in
Region des ersten Molaren liegt. Bei Cebus und Alouatta ist dies gleichermaßen
der Fall. Sowohl bei Neuweltaffen als auch bei Altweltaffen hatten die
Zahnwurzeln der Oberkiefermolaren meist engen Kontakt zum Boden des
Recessus alveolaris. Dies wurde auch von Koppe and Ohkawa (1999), Koppe
(2005b) und Oesterreich et al. (2005) berichtet.
Dies lässt die Annahme zu, dass die tiefste Stelle des Recessus alveolaris
zugleich in der Region liegt, in der die geringste Knochendicke zwischen
Zahnwurzeln und Kieferhöhlenboden liegt. In der vorliegenden Studie wurde die

Diskussion
79
Knochendicke zwar nicht vermessen, aber einige Untersuchungen am
Menschen liegen diesbezüglich in der bestehenden Literatur vor (u.a. Eberhardt
et al. 1992; Nogi and Kanazawa 2001; Kwak et al. 2004). Diese zeigen jedoch,
entgegengesetzt unseren Ergebnissen, dass die geringste Distanz nicht auf
Höhe des ersten Molaren, sondern zwischen zweitem Molaren und
Kieferhöhlenboden besteht. Die Knochendicke oberhalb des zweiten Molaren
variiert dabei zwischen 0,83mm (Eberhardt et al. 1992) und 2,9mm (Nogi and
Kanazawa 2001). Letztgenannte Autoren vermuten, dass der größere Abstand
zwischen Zahnwurzel und Kieferhöhlenboden auf die kürzeren Zahnwurzeln bei
der japanischen Bevölkerung im Gegensatz zur kaukasischen
Bevölkerungsgruppe zurückzuführen ist, die Eberhardt et al. (1992)
untersuchten. Diese Erklärung ist unserer Auffassung nach nicht stichhaltig,
denn bei der Betrachtung von Macaca fuscata zeigt sich ein deutlicher Abstand
von Zahnwurzeln und Kieferhöhlenboden, obgleich diese Spezies sehr lange
Zahnwurzeln besitzt (Koppe and Rae 2001). Letztgenannte Autoren weisen
darauf hin, dass möglicherweise lange Wurzeln die Pneumatisation des
Recessus alveolaris bei Macaca fuscata hemmen. Denn bei Macaca fuscata
liegen nicht nur die Zahnwurzeln oberhalb einer Bezugsebene durch den
Nasenboden, sondern auch der Boden der Kieferhöhle oberhalb einer
Bezugsebene durch den Nasenboden (Koppe and Rae 2001).
Asymmetrien des Recessus alveolaris
Unsere Untersuchungsergebnisse zeigten, dass Asymmetrien von rechtem und
linkem Recessus alveolaris sowohl bei Altweltaffen als auch bei Neuweltaffen
auftreten. Zwei Schädel der Spezies Homo sapiens und ein Schädel der Spezies
Alouatta wiesen deutliche Seitenabweichungen auf. Bei einem dieser Schädel
von Homo sapiens lag der linke Boden der Kieferhöhle mit dem Nasenboden
nahezu auf einer Ebene, wohingegen auf der rechten Seite ein Recessus
alveolaris vorhanden war; bei dem anderen Schädel zeigte sich dieser
Sachverhalt seitenverkehrt. Bei dem Schädel der Spezies Alouatta war der

Diskussion
80
rechte Recessus alveolaris beinahe doppelt so groß wie die linke Seite.
Bisher sind uns keine Studien bekannt, die die Asymmetrie des Volumens des
Recessus alveolaris prüften. Untersuchungen zu Seitenabweichungen von
Kieferhöhlen liegen jedoch vor. Diese veranschaulichen, dass Asymmetrien von
rechter und linker Kieferhöhle beim Menschen keine Seltenheit sind (u.a.
Schumacher et al. 1972a; Anagnostopoulou et al. 1991). Fehlende Zähne haben
dabei offenbar keinen Einfluss auf Seitendifferenzen von rechtem und linkem
Kieferhöhlenvolumen (Ariji et al. 1994). Ausgesprochen große Abweichungen
sind selten (Schumacher et al. 1972b). Dies konnten wir durch unsere
Untersuchung für den Recessus alveolaris gleichermaßen bestätigen.
Eine mögliche Erklärung für die Seitendifferenzen liefern Schumacher et al.
(1972b). Sie sehen in den ungleichen Kieferhöhlenmorphologien von rechter und
linker Seite eine natürliche biologische Varianz. Einen weiteren Ansatz liefern
Manning and Chamberlain (1993), die von einer fluktuierenden Asymmetrie
sprechen, welche als zufällige genetische oder umweltbedingte Störung in der
Ontogenese betrachtet werden kann. Dieser Ansicht sind auch Moormann
(2004) und Koppe et al. (1997) nach ihren Untersuchungen zum
Kieferhöhlenvolumen an höheren Primaten. Vor dem Hintergrund dieser Studien
ist anzunehmen, dass diese Erklärung auch auf unsere Studie für den Recessus
alveolaris bei Neuweltaffen und Altweltaffen übertragen werden kann.
Recessus alveolaris und Schädelmorphologie
Unsere Untersuchungsergebnisse zeigten, dass signifikante Korrelationen
zwischen den Volumina von Recessus alveolaris und Sinus maxillaris sowohl für
Neuweltaffen als auch Altweltaffen bestehen. Je größer die Kieferhöhle ist, umso
mehr tendiert sie offenbar dazu, den Processus alveolaris zu pneumatisieren.
Dieser Zusammenhang wurde auch in der partiellen Korrelationsanalyse bei
Neuweltaffen bestätigt. Dieses Ergebnis deckt sich mit den Untersuchungen von
Oesterreich et al. (2005). Bei den Altweltaffen hingegen waren die

Diskussion
81
Zusammenhänge schwächer, was sich in der partiellen Korrelationsanalyse und
den unterschiedlichen Steigungen der Regressionsgeraden zeigte. Somit kann
der Recessus alveolaris bei den Altweltaffen nicht allein durch das Ausmaß der
Kieferhöhle selbst erklärt werden. Dies begründet sich möglicherweise in den
unterschiedlichen Pneumatisationsrichtungen der Spezies. So extendierte
beispielsweise die Kieferhöhle des Orang-Utans nach posterior in Richtung
Sinus sphenoidale und weniger nach kaudal in den Processus alveolaris.
Vergleichbare Daten zum Volumen des Recessus alveolaris an einer breiten
Variation unterschiedlicher Spezies von Altweltaffen liegen bisher nicht vor.
Weitere Studien können möglicherweise Aufschluss über die genauen
Zusammenhänge geben.
Unsere Korrelationsanalyse von Recessus alveolaris und Schädelmaßen zeigte,
dass das Merkmalspaar von Obergesichtshöhe und Recessus alveolaris bei
Neuweltaffen eine negative Korrelation und bei Altweltaffen eine positive
Korrelation aufzeigt. Für die anderen Merkmalspaare besteht ledigich ein
schwacher Zusammenhang. Dies wird vor allem durch die partielle
Korrelationsanalyse bestätigt, welche zeigte, dass sowohl Gesichtslänge als
auch Schädellänge einen Einfluss auf die entsprechenden Merkmalspaare
haben. Insbesondere der schwache Zusammenhang zwischen Recessus
alveolaris und Gaumenmaßen ist bemerkenswert, denn in der Studie von
Oesterreich et al. (2005) zeigte sich durchaus ein solcher Zusammenhang.
Lediglich bei den Altweltaffen ergab sich nach unseren Ergebnissen der
partiellen Korrelationsanalyse eine signifikante Korrelation für die Gaumenhöhe
und den Recessus alveolaris bei konstanter Gesichtslänge.
Obwohl der Einfluss des Gaumens auf die Kieferhöhlenmorphologie in einigen
Studien untersucht wurde, ist nach wie vor nicht geklärt, welche Rolle der
knöcherne Gaumen in Bezug auf die Morphologie der Kieferhöhlen spielt. In
diesem Zusammenhang vermuten Koppe und Schumacher (1992), dass die
Reduktion des Kiefers in der Anthropogenese mit einer Verminderung des

Diskussion
82
Pneumatisationsgrades des Gesichtsschädels einhergegangen ist. Frühere
Untersuchungen zu dieser Problematik liefern kein einheiltiches Bild. So ist beim
Menschen der Einfluss des Gaumens auf die Kieferhöhlengröße vorhanden
(Eller 1932), wohingegen dieser bei Macaca fuscata (Geißler 2007) und beim
Gibbon (Klug 2011) offensichtlich gering ist. Beim Orang-Utan wird das
Abhängigkeitsverhältnis von Sinus maxillaris und Gaumen von der Schädellänge
beeinflusst (Koppe et al. 1996). Übertragen wir dieses Ergebnis auf den
Recessus alveolaris, stellen wir fest, dass die Schädellänge gleichermaßen
einen Einfluss auf das Abhängigkeitsverhältnis von Gaumenhöhe und Recessus
alveolaris hat. Studien von Koppe et al. (2005b) und Koppe and Rae (2001)
wiederum deuten darauf hin, dass beim Menschen und bei Makaken ein hoher
Gaumen mit einem tiefen Recessus alveolaris in Zusammenhang steht.
Aufgrund unserer Ergebnisse und der Ergebnisse der bisherigen Studien
schließen wir einen Zusammenhang zwischen Gaumen und Recessus alveolaris
nicht aus. Dennoch kommen wir zu dem Schluss, dass weitere Faktoren für die
Pneumatisation des Processus alveolaris verantwortlich sind.
Die Untersuchungen von Moss and Greenberg (1967) zur Hypothese der
funktionellen Matrix sind in diesem Zusammenhang von besonderem Interesse.
Moss and Greenberg (1967) sehen die Entwicklung des Schädels als ein
Resultat aus dem Zusammenspiel von funktioneller Matrix
(Weichgewebskomponente) und skelettaler Einheit (Hartgewebskomponente).
Dabei wird das ontogenetische Wachstum primär durch genetische Faktoren
determiniert. Für das postnatale Wachstum und die Ausformung der skelettalen
Komponenten sind letztendlich die Weichgewebsanteile mit deren
physiologischen Funktionen (funktionelle Matrix) verantwortlich. Somit sind
Größe und Form der knöchernen Komponenten Folge und Antwort auf die
Wachstumsänderungen der funktionellen Matrix (Moss and Greenberg 1967).
Insofern muss jede dieser skelettalen Einheiten eigenständig betrachtet werden
und ist von anderen Komplexen abzugrenzen. Folglich kann der Recessus
alveolaris als Teil der Kieferhöhle nicht allein durch die Dimension des Gaumens

Diskussion
83
oder andere Schädelmaße erklärt werden. Des Weiteren haben Faktoren
extrinsischer Herkunft Einfluss auf die funktionelle Matrix. An dieser Stelle sei
nochmals auf die Studien von Shea (1977) und Rae et al. (2003) hingewiesen,
die darauf hindeuten, dass in Regionen mit kälteren Temperaturen die Volumina
der Kieferhöhlen kleiner sind.

Diskussion
84
Schlussfolgerung
Unter Bezugnahme auf die in der Einleitung formulierten Ziele dieser Arbeit
kommen wir zu folgenden Ergebnissen:
1. Der Recessus alveolaris, der Teil der Kieferhöhle, der unterhalb einer
Bezugslinie durch den Nasenboden liegt, ist bei allen Schädeln rechts
vorhanden. Das größte Volumen unter den Altweltaffen zeigt der Gorilla
mit 5,00cm³, das kleinste der Orang-Utan mit 0,53cm³. Unter den
Neuweltaffen hat Alouatta mit 0,88cm³ das größte, Brachyteles mit
0,03cm³ das kleinste Volumen. Der Recessus alveolaris beginnt bei den
Altweltaffen anterior in den meisten Fällen auf Höhe des zweiten
Prämolaren bzw. des ersten Molaren unterhalb des Nasenbodens und
endet posterior hinter den letzten vorhandenen Molaren. Bei den
Neuweltaffen liegt der anteriore Beginn des Recessus alveolaris auf Höhe
des zweiten bzw. dritten Prämolaren unterhalb des Nasenbodens und
endet distal des zweiten bzw. dritten Molaren. Die Zähne haben engen
Kontakt zum Kieferhöhlenboden oder projizieren sich in diesen hinein.
Somit zeigt sich der Kieferhöhlenboden häufig uneben, wie unter anderem
bei den Spezies Alouatta, Cebus, Pan und Symphalangus syndactylus.
Bei Homo sapiens und beim Gorilla stellt sich der Kieferhöhlenboden
zerklüftet oder wannenförmig dar und breitet sich in die Furkation der
ersten und zweiten Molaren aus. Lediglich bei einem Schädel des Orang-
Utans zeigt sich der Recessus alveolaris schmal und flach.
2. Die tiefste Stelle des Recessus alveolaris liegt sowohl bei den
Neuweltaffen als auch Altweltaffen in der Regel in Höhe des ersten
Molaren. Bei Neuweltaffen weist die tiefste Stelle des Recessus alveolaris
in der Region des ersten Molaren eine Größe von 0,51cm und bei den
Altweltaffen von 0,75cm auf.
3. Asymmetrien des Recessus alveolaris existieren sowohl bei Neuweltaffen
als auch bei Altweltaffen. Diese lassen sich durch den Asymmetrieindex

Diskussion
85
quantifizieren. Extreme Seitenabweichungen sind selten. Lediglich zwei
Schädel der Spezies Homo sapiens und ein Schädel der Spezies Alouatta
zeigen Extremwerte beim Asymmetrieindex auf. Wir ordnen die
Asymmetrien als fluktuierend ein.
4. Sowohl bei den Neuweltaffen als auch bei den Altweltaffen besteht eine
signifikante Korrelation zwischen den Volumina der Kieferhöhlen und
seines Recessus alveolaris. Der Processus alveolaris ist umso mehr
pneumatisiert, je größer die Kieferhöhle ist. Der Zusammenhang von
Recessus alveolaris und Sinus maxillaris ist bei Neuweltaffen jedoch
stärker als bei Altweltaffen, was durch die unterschiedlichen Steigungen
der Regressionsgeraden bestätigt wird. Bei den Altweltaffen kann ein
Einfluss der Gesichtslänge auf das Merkmalspaar Recessus alveolaris
und Sinus maxillaris nachgewiesen werden. Hinsichtlich der Beziehung
der Merkmalspaare von Recessus alveolaris und weiteren Schädelmaßen
zeigt sich, dass nur schwache Korrelationen bestehen und diese in
unterschiedlichem Ausmaß von der Gesichtslänge und der Schädellänge
beeinflusst werden. Lediglich das Merkmalspaar von Obergesichtshöhe
und Recessus alveolaris zeigt auch in der partiellen Korrelationsanalyse
signifikante Korrelationen bei Neuweltaffen und Altweltaffen.
5. Die vorliegende Arbeit zeigt erstmals eine vergleichende Analyse der
Volumina des Recessus alveolaris von Neuweltaffen und Altweltaffen. Die
Gesetzmäßigkeiten, mit welchen sich die Kieferhöhle nach basal
ausdehnt, sind bei Neuwelt- und Altweltaffen offenbar unterschiedlich. Für
die Anthropologie und Paläoanthropologie kann die vorliegende Studie
von Bedeutung sein, da sich Fossilien und archäologische Schädelfunde
häufig nur auf unvollständige Schädelfragmente beschränken. Mitunter
sind lediglich Bruchstücke der Maxilla zu finden. Durch das Wissen um die
Pneumatisation des Processus alveolaris können Rückschlüsse auf das
Kieferhöhlenvolumen und damit möglicherweise auch auf das
Mittelgesicht gezogen werden. Weitere Studien sollten zu diesen
Ergebnissen der vorliegenden Studie durchgeführt werden.

Zusammenfassung
86
6 Zusammenfassung
In der vorliegenden Arbeit wurde die Pneumatisation des Processus alveolaris
maxillae unterschiedlicher Spezies von Catarrhini und Platyrrhini untersucht.
Dazu standen 89 computertomographische Datensätze adulter Schädel mit einer
permanenten Dentition beiderlei Geschlechts zur Verfügung. Der Recessus
alveolaris wurde als der Teil der Kieferhöhle definiert, der unter einer gedachten
Bezugslinie zum Nasenboden liegt. Anhand der CT-Datensätze wurde das
Volumen des Recessus alveolaris mit der 3D-Rekonstruktionssoftware
„WinSurf® 4-1-0“ (Golden Software Inc., USA) berechnet sowie 3D-
Rekonstruktionen zur Beurteilung der Form des Kieferhöhlenbodens angefertigt.
Darüber hinaus wurden die Schädel unter vergleichend-anatomischen
Gesichtspunkten vermessen. Die Beziehungen der erhobenen Volumina und
Messtrecken wurden mit Hilfe von Korrelations- und Regressionsanalysen
geprüft.
Das Volumen des Recessus alveolaris war bei Altweltaffen größer als bei
Neuweltaffen. Der Gorilla zeigte unter den Altweltaffen mit 5,00cm³ bzw. Alouatta
unter den Neuweltaffen mit 0,88cm³ das größte Volumen der jeweiligen Gruppe.
Das kleinste Volumen des Recessus alveolaris besaß unter den Neuweltaffen
Brachyteles mit 0,03cm³ und unter den Altweltaffen der Orang-Utan mit 0,53cm³.
Die tiefste Stelle des Recessus alveolaris lag sowohl bei Altweltaffen als auch
bei den Spezies Alouatta und Cebus in der Regel auf Höhe des ersten Molaren.
Die Morphologie des Recessus alveolaris zeigte inter- und intraspezifisch eine
große Variationsbreite. So stellte sich der Recessus alveolaris beim Menschen
und Gorilla als wannenförmig oder unregelmäßig dar; bei den Schimpansen,
Gibbons und den Neuweltaffen zeigte er eine unregelmäßige Form. Die
Zahnwurzeln hatten häufig direkten Kontakt zum Boden der Kieferhöhle und
projizieren sich zum Teil in die Kieferhöhle hinein. In einem Fall des Orang-Utans
zeigte sich eine Wannenform, im anderen Fall ein flacher, schmaler Recessus
alveolaris, der keinen direkten Kontakt zu den Zahnwurzeln aufwies.

Zusammenfassung
87
Asymmetrien von rechtem und linkem Recessus alveolaris traten sowohl bei
Neuweltaffen als auch bei Altweltaffen auf. Diese Asymmetrien wurden als
fluktuierend eingestuft.
Mit der Korrelationsanalyse konnte ein signifikanter Zusammenhang zwischen
den Volumina von Recessus alveolaris und Sinus maxillaris sowohl bei Alt- als
auch Neuweltaffen nachgewiesen werden. Je größer die Kieferhöhle, umso mehr
tendierte sie dazu den Processus alveolaris zu pneumatisieren. Die Ausdehnung
der Sinus maxillaris in den Processus alveolaris war jedoch bei den
Affengruppen unterschiedlich. Dies bestätigte sich in den unterschiedlichen
Anstiegen der Regressionsgeraden. Bei den Altweltaffen wurde das
Merkmalspaar von Recessus alveolaris und Sinus maxillaris von der
Gesichtsschädellänge beeinflusst.
Obwohl unsere Ergebnisse darauf hindeuten, dass ein großes
Kieferhöhlenvolumen mit einem großen Volumen des Recessus alveolaris in
Zusammenhang steht, vermuten wir, dass eine alleinige strukturelle Erklärung
unzureichend ist. Dies begründen wir durch die unterschiedlichen Ergebnisse
zwischen den Affengruppen und durch den schwachen Zusammenhang
zwischen Recessus alveolaris und Schädelmaßen. Wir nehmen an, dass eine
komplexe Interaktion mehrerer Faktoren intrinsischer und extrinsischer Natur für
die Morphologie und die Pneumatisation des Recessus alveolaris verantwortlich
sind. In welchem Maß extrinsische Faktoren Einfluss auf die Gestalt des
Recessus alveolaris nehmen, sollte anhand weiterer Studien geprüft werden.

Literaturverzeichnis
88
7 Literaturverzeichnis
Al Bourney, M (1999) Verwendung des Fernröntgenseitenbildes des Kopfes und des Orthopantomogramms zur metrischen Analyse des Schädels in der Kieferorthopädie. Inaug Diss, Universität Giessen
Anagnostopoulou S, Venieratos D, Spyropoulos N (1991) Classification of human maxillar sinuses according to their geometric features. Anat Anz 173: 121–130
Ariji Y, Kuroki T, Moriguchi S, Ariji E, Kanda S (1994) Age changes in the volume of the human maxillary sinus: a study using computed tomography. Dentomaxillofac Radiol 23: 163–168
Ax P (1984) Das phylogenetische System. Systematisierung der lebenden Natur aufgrund ihrer Phylogenese. Stuttgart, Gustav Fischer
Bahat O (2000) Brånemark system implants in the posterior maxilla: clinical study of 660 implants followed for 5 to 12 years. Int J Oral Maxillofac Implants 15: 646–653
Bauer HB (1943) Maxillary sinusitis of dental origin. Am J Orthodo Oral Surg 29: B133–B151
Blaney SP (1990) Why paranasal sinuses? J Laryngol Otol 104: 690–693
Blanton PI, Biggs NL (1969) Eighteen hundred years of controversy: The paranasal sinuses. Am J Anat 124: 135–148
Bluestone CD, Pagano AS, Swarts JD, Laitman JT (2012) Consequences of evolution: Is rhinosinusitis, like otitis media, a unique disease of humans? Otolaryng Head Neck 147: 986–991
Boyne PJ, James RA (1980) Grafting of the maxillary sinus floor with autogenous marrow and bone. J Oral Surg 38: 613–616
Braune W, Clasen FE (1877) Die Nebenhöhlen der menschlichen Nase in ihrer Bedeutung für den Mechanismus des Riechens. Z Anat Entwicklungsgesch 2: 1–28
Brook I (2009) Sinusitis. Periodontol 2000 49: 126–139
Cave AJ (1967) Observations on the platyrrhine nasal fossa. Am J Phys Anthropol 26: 277–288
Cave AJ, Haines W (1940) The paranasal sinuses of the anthropoid apes. J Anat 74: 493–523

Literaturverzeichnis
89
Cawood JD, Howell RA (1988) A classification of the edentulous jaws. Int J Oral Max Surg 17: 232–236
Collins JG (1997) Prevalence of selected chronic conditions: United States 1990-1992. Vital Health Stat 10: 1–89
Croen KD (1993) Evidence for an antiviral effect of nitrit oxide. Inhibition of herpes simplex virus type 1 replication. J Clin Invest 91: 2446–2452
Dornum M von, Ruvolo M (1999) Phylogenetic relationships of the new world monkeys (Primates, Platyrrhini) based on nuclear G6PD DNA sequences. Mol Phylogenet Evol 11: 459–476
Drettner B (1980) Pathophysiology of paranasal sinuses with clinical impliations. Clin Otolaryngol Allied Sci 5: 277–284
Eberhardt JA, Torabinejad M, Christiansen EL (1992) A computed tomographic study of the distances between the maxillary sinus floor and the apices of the maxillary posterior teeth. Oral Surg Oral Med Oral Pathol 73: 345–346
Eller H (1932) Der Sinus maxillaris und seine Nachbarorgane bei verschiedenen Affen und beim Hunde. Z Anat Entwicklungsgesch 97: 725–756
Emirzeoglu M, Sahin B, Bilgic S, Celebi M, Uzun A (2007) Volumetric evaluation of the paranasal sinuses in normal subjects using computer tomography images: A stereological study. Auris Nasus Larynx 34: 191–195
Engström H, Chamberlain D, Kiger R, Egelberg J (1988) Radiographic evaluation of the effect of initial periodontal therapy on thickness of the maxillary sinus mucosa. J Periodontol 59: 604–608
Fanghänel J, Giebel J, Koppe T, Miehe B, Splieth C, Kocher T, Weingärtner J, Kubein-Meesenburg D (2003) Kopf, Cranium und Hals, Collum. In: Fanghänel J, Pera F, Anderhuber F, Nitch R (Hrsg.) Waldeyer – Anatomie des Menschen. 17. Aufl. Berlin, de Gruyter: 177–353
Fleagle JG (2013) Primate Adaptation and Evolution. 3rd Ed, San Diego, Academic Press
Ford SM (1986) Systematics of the New World Monkeys. New York, A. R. Liss
Frenkert K (2008) Zur postnatalen Entwicklung des Gaumengewölbes des Schimpansen (Pan troglodytes). Inaug Diss, Universität Greifswald
Geißler F (2007) Biometrische Untersuchungen zum Wachstum des harten Gaumens von Macaca fuscata. Inaug Diss, Universität Greifswald
Geissmann T (2003) Vergleichende Primatologie. Berlin, Springer

Literaturverzeichnis
90
Goodman M, Porter CA, Czelusniak J, Page AL, Schneider H (1998) Toward a phylogenetic classification of primates based on DNA evidence complemented by fossil evidence. Mol Phylogenet Evol 9: 585–598
Hall WB, Grupe HE, Claycomb CK (1967) The periodontium and periodontal pathology in the howler monkey. Arch Oral Biol 12: 359–365
Hardin JW, Hilbe JM (2012) Generalized linear models and extensions. College Station, Texas, Stata Press
Harzer W (1999) Lehrbuch der Kieferorthopädie. München, Hanser
Hays WL (1988) Statistics. Fort Worth, Holt, Rinehart and Winston
Hershkovitz P (1977) Living New World Monkeys (Platyrrhini). With an
Introduction to Primates. Vol 1. Chicago, Chicago University Press
Hofer H (1952) Der Gestaltwandel des Schädels der Säugetiere und Vögel mit
besonderer Berücksichtigung der Knickungstypen der Schädelbasis.
Verhandlungen der Anatomische Gesellschaft 50: 102–113
Jaffin RA, Berman CL (1991) The excessive loss of Branemark fixtures in type IV Bone: a 5-Year analysis. J Periodontol 62: 2–4
Jungers WL, Falsetti AB, Wall CE (1995) Shape, relative size, and size-adjustments in morphometrics. Yearb Phys Anthropol 38: 137–161
Kakehashi S, Bear PN, White CL (1963) Comparative pathology of periodontal disease: I. Gorilla. Oral Surg Oral Med Oral Pathol 16: 397–406
Keene HJ (1965) The relationsship between third molar agenesis and the morphologic variability of the molar teeth. Angle Orthod 35: 289–298
Keir J (2008) Why do we have paranasal sinuses? J Laryngol Otol 123: 4–8
Kilgore L (1989) Dental pathologies in ten free-ranging chimpanzees from Gombe National Park, Tanzania. Am J Phys Anthropol 80: 219–227
Kim HJ, Yoon HR, Kim K, Kang MK, Kwak HH, Park HD, Han SD, Park CS (2002) Personal-computer-based three-dimensional reconstruction and simulation of maxillary sinus. Surg Radiol Anat 24: 393–399
Kim M, Jung U, Kim C, Kim K, Choi S, Kim C, Cho K (2006) Maxillary sinus septa: prevalence, height, location and morphology. A reformatted computed tomography scan analysis. J Periodontol 77: 903–908

Literaturverzeichnis
91
Kishi K, Nakatsu T, Wakasa T, Hasegawa I, Uemura M, Hazawa K, Shimizu K, Doi Y (1995) Clinical and radiographic study of antro-oral fistula. Oral Radiol 11: 17–22
Klug S (2011) Zur funktionellen Morphologie der Nasennebenhöhlen der Hylobatiden (Primates, Hylobatidae). Inaug Diss, Universität Greifswald
Koppe T, Moormann T, Wallner C, Röhrer-Ertl O (2005a) Extensive enlargement of the maxillary sinus in Alouatta caraya (Mammalia, Primates, Cebidae): An allometric approach to skull pneumatization in Atelinae. J Morphol 263: 238–246
Koppe T, Nagai H (1997) Growth pattern of the maxillary sinus in the Japanese macaque (Macaca fuscata): reflections on the structural role of the paranasal sinuses. J Anat 190: 533–544
Koppe T, Nagai H (1998) The maxillary sinus of extant catarrhine primates. In: Alt K, Rösing F, Teschler-Nicola, N (Eds.) Dental Anthropology: Fundamentals, Limits and Prospects. Wien, Springer Verlag: 167–177
Koppe T, Nagai H, Rae TC (1999a) Factors in the evolution of the primate pneumatic cavities: possible roles of the paranasal sinuses. In: Koppe T, Nagai H, Alt K (Eds.) The Paranasal Sinuses of Higher Primates: Development, Function and Evolution. Chicago, Quintessence: 151–174
Koppe T, Nakatsukasa M, Yamanaka A (2005b) Implication of craniofacial morphology for the pneumatization pattern of the human alveolar process. Acta Med Lithuania 12: 43–45
Koppe T, Ohkawa Y (1999) Pneumatization of the facial skeleton in catarrhine primates. In: Koppe T, Nagai H und Alt K (Eds.) The Paranasal Sinuses of Higher Primates: Development, Function and Evolution. Chicago, Quintessence: 77–119
Koppe T, Rae TC (2001) Factors influencing the pneumatization of the alveolar process in macaques. In: Brook A (Ed.) Dental Morphology 2001. Sheffield, Sheffield Academic Press: 267–276
Koppe T, Rae TC, Swindler DR (1999b) Influence of craniofacial morphology on primate paranasal pneumatization. Ann Anat 181: 77–80
Koppe T, Röhrer-Ertl O, Hahn D, Reike R, Nagai H (1996) The relationship between the palatal form and the maxillary sinus in orang-utan. Okajimas Folia Anat Jpn 72: 31–41
Koppe T, Schumacher GH, Nagai H (1997) Asymmetry in maxillary sinus size of higher primates. J Morphol 232: 278

Literaturverzeichnis
92
Koppe T, Schumacher KU (1990) Vergleichend-anatomische Untersuchungen zur funktionellen Morphologie der Nasennebenhöhlen des Menschen und der Pongoiden. Eine quantitative Studie auf der Grundlage der Computertomographie. Med Diss B, Universität Rostock
Koppe T, Schumacher KU (1992) Untersuchungen zum Pneumatisationsgrad des Viscerocranium beim Menschen und bei den Pongiden. Acta Anat Nippon 67: 725–734
Krennmair G, Ulm C, Lugmayr H (1997) Maxillary sinus septa: incidence, morphology and clinical implications. J Cran Maxill Surg 25: 261–265
Kwak HH, Park HD, Yoon HR, Kang MK, Koh KS, Kim HJ (2004) Topographic anatomy of the inferior wall of the maxillary sinus in Koreans. Int J Oral Max Surg 33: 382–388
Lekholm U, Zarb GA (1985) Patient selection and preparation. In: Brånemark PI, Zarb GA, Albrektsson T (Eds.) Tissue integrated prostheses: osseointegration in clinical dentistry. Chicago, Quintessence: 199–209
Lovell NC (1990) Skeletal and dental pathology of free-ranging mountain gorillas. Am J Phys Anthropol 81: 399–412
Lovich E, Gibbons J (1992) A review of techniques for quantifying sexual size dimorphismus. Growth Develop Aging 56: 269–281
Lund VJ (1988) The maxillary sinus in the higher primates. Acta Otolaryngol 105: 163–171
Lundberg J, Farkas-Szallasi T, Weitzberg E, Rinder J, Lidholm J, Änggåard A, Hökfelt T, Lundberg JM, Alving K (1995) High nitric oxide production in human paranasal sinuses. Nat Med 1: 370–373
Maestre-Ferrin L, Galan-Gil S, Rubio-Serrano M, Penarrocha-Diago M, Penarrocha-Oltra D (2009) Maxillary sinus septa: A systematic review. Med Oral Patol Oral Cir Bucal 15: 383–386
Manning J, Chamberlain AT (1993) Fluctuating asymmetry, sexual selection and canine teeth in primates. Proc R Soc Lond, B 251: 83–87
Márquez S (2008) The paranasal sinuses: the last frontier in craniofacial biology. Anat Rec 261: 1350–1361
Martin R (1928) Lehrbuch der Anthropologie. Jena, Gustav Fischer

Literaturverzeichnis
93
Melén I (1996) Maxillary inflammation and infection in relation to dental infection. In: Stierna P, Karlsson G, Melén I, Jannert M (Eds.) Aspects on Sinusitis. Diagnosis and Treatment in Adults. HNS. Stockholm, Eli Lilly Sweden AB: 51–56
Melén I, Lindahl L, Adréasson L, Rundcrantz H (1986) Chronic maxillary sinusitis. Definition, Diagnosis and Relation to Dental Infections and Nasal Polyposis. Acta Otolaryngol 101: 320–327
Miyamoto MM, Koop BF, Slightom JL, Goodmann M, Tennant MR (1988) Molecular systematics of higher primates: genealogical relations and classification. Proc Natl Acad Sci USA 85: 7627–7631
Moore W (1981) The Mammalian Skull. Cambridge, Cambridge Universitiy Press
Moormann T (2004) Funktionelle Morphologie der Nasennebenhöhlen des Brüllaffen Alouatta caraya (Primates, Platyrrhini). Inaug Diss, Universität Greifswald
Moss ML, Greenberg SN (1967) Functional cranial analysis of the human maxillary bone: I, Basal bone. Angle Orthod 37: 151–164
Negus VE (1954) The function of the paranasal sinuses. Acta Otolaryngol 44: 408–426
Nimigean V, Nimigean VR, Maru N, Salavastru DI, Badita D (2008) The maxillary sinus floor in the oral implantology. Rom J Morphol Embryol 49: 485–489
Nishimura TD, Takai M, Tsubamoto T, Egi N, Shigehara N (2005) Variation in maxillary sinus anatomy among platyrrhine monkeys. J Hum Evol 49: 370–389
Nogi T, Kanazawa E (2001) The relationship between tooth roots and the floor of maxillary sinus assessed using CT images. In: Brook A (Ed) Dental Morphology 2001. Sheffield, Sheffield Academic Press: 257–266
Nohadani N, Ruf S (2008) Assessment of vertical facial and dentoalveolar changes using panoramic radiography. Eur J Orthod 3: 262–268
Nötzel F, Schultz C (2009) Leitfaden der kieferorthpädischen Diagnostik. 2. Aufl. Köln, Deutscher Zahnärzte Verlag
Nurbakhsh B, Friedman S, Kulkarni GH, Basrani B, Lam E (2011) Resolution of maxillary sinus mucositis after endodontic treatment of maxillary teeth with apical periodontitis: A cone-beam computed tomography pilot study. J Endodont 37: 1504–1511

Literaturverzeichnis
94
Oesterreich A, Koppe T, Wallner C, Röhrer-Ertl O (2005) New insights into the pneumatization pattern of the alveolar process in cebidae. In: Zadzinska E (Ed) Current Trends in Dental Morphology Research. Lodz, Univerity of Press: 269–277
Pjetursson BE, Rast C, Brägger C, Schmidlin K, Zwahlen M, Lang NP (2009) Maxillary sinus floor elevation using the (transalveolar) osteotome technique with or without grafting material. Part I: implant survival and patients' perception. Clin Oral Impl Res 20: 667–676
Preuschoft H, Witte H, Witzel U (2002) Pneumatized spaces, sinuses and spongy bones in the skulls of primates. Anthropol Anz 60: 67–79
Rae TC, Hill RA, Hamada Y, Koppe T (2003) Clinal variation of maxillary sinus volume in Japanese macaques (Macaca fuscata). Am J Primatol 59: 153–158
Rae TC, Koppe T (2004) Holes in the head: evolutionary interpretations of the paranasal sinuses in catarrhines. Evol Anthropol 12: 211–223
Rae TC, Koppe T (2008) Independence of biomechanical forces and craniofacial pneumatization in Cebus. Anat Rec 291: 1414–1419
Rae TC, Koppe T, Nagai H (1997) Maxillary sinus reduction due to cold stress in the Macaca fascicularis species group. Am J Phys Anthropol 24: 190–191
Ravosa MJ (1991) Ontogenetic perspective on mechanical and nonmechanical models of primate circumorbital morphology. Am J Phys Anthropol 85: 95–112
Reichert TE (2009) Odontogene Kieferhöhlenerkrankungen. Pathophysiologie, Diagnostik und Therapie aus chirurgischer Sicht. MKG-Chirurg 2: 293–310
Reinert S, Machtens E (2002) Erkrankungen der Nasennebenhöhlen. In: Schwenzer N und Ehrenfeld M (Hrsg.) Zahn-, Mund- und Kieferheilkunde, Band 2: Spezielle Chirurgie. 3. Aufl. Stuttgart, Georg Thieme: 1–22
Röhrer-Ertl O. Persönliche Mitteilung
Rosen PS, Summers R, Mellado JR, Salkin LM, Shanaman RH, Marks H (1999) The bone-added osteotome sinus floor elevation technique: multicenter retrospective report of consecutively treated patients. Int J Oral Max Impl 14: 853–858
Rosenberger AL, Setoguchi T, Shigehara N (1990) The fossil record of callitrichine primates. J Hum Evol 19: 209–236
Rossie JB (2006) Ontogeny and Homology of the paranasal sinuses in Platyrrhini (Mammalia: Primates). J Morphol 267: 1–40

Literaturverzeichnis
95
Rozkovcová E, Marková M, Lánik J, Zvárová J (2004) Agenesis of third molars in young Czech population. Prague Medical Report 105: 35–52
Schaeffer J (1920) The Nose, Paranasal Sinuses, Nasolacrymal Passageways, and Olfactory Organ in Man. Philadelphia, Blakiston's Son
Schiwal OS (2005) Zur funktionellen Morphologie der Sinus paranasales von Callithrix jacchus (Primates, Platyrrhini). Inaug Diss, Universität Greifswald
Schmelzle R, Schwenzer N (1982) Die odontogenen Erkrankungen der Kieferhöhle. Acta Otorhinolaryngol 235: 379–386
Schneider H, Canavez FC, Sampaio I, Moreira MA, Tagliaro CH, Seuánez HN (2001) Can molecular data place each neotropical monkey in its own branch? Chromosoma 109: 515–523
Schneider H, Sampaio I, Harada ML, Barroso CM, Schneider MP, Czelusniak J, Goodman M (1996) Molecular phylogeny of the New World monkeys (Platyrrhini, primates) based on two unlinked nuclear genes: IRBP intron 1 and ϵ-globin sequences. Am J Phys Anthropol 100: 153–179
Schneider H, Schneider MP, Sampaio I, Harada ML, Stanhope M, Czelusniak J, Goodmann M (1993) Molecular phylogeny of the new world monkeys (Platyrrhini, Primates). Mol Phylogenet Evol 2: 225–242
Schumacher GH (1971) Topographische Anatomie der Kieferhöhle. Dtsch Stomatol 21: 231–236
Schumacher GH, Fanghänel J, Woithe G, Pawlak K (1972b) Quantitative Untersuchung zur Erfassung von Asymmetrien des Kopfes. 3. Mitteilung: Nasenhöhlen und Nasennebenhöhlen. Anat Anz 132: 197–210
Schumacher KU, Koppe T, Fanghänel J, Schumacher GH, Nagai H (1994) Morphometric studies on the facial skeleton of humans and pongoids based on CT-scans. Acta Anat Nippon 69: 636–644
Schumacher GH, Heyne HJ, Fanghänel R (1972a) Zur Anatomie der menschlichen Nasennebenhöhlen. 2. Mitteilung. Volumetrie. Anat Anz 130: 143–157
Shankar L, Evans K, Hawke M, Stammberger H (1994) Atlas der Nasennebenhöhlen. Weinheim, Chapman & Hall
Sharan A, Madjar D (2008) Maxillary sinus pneumatization following extractions: a radiographic study. Int J Oral Max Impl 23: 48–56

Literaturverzeichnis
96
Shea BT (1977) Eskimo craniofacial morphology, cold stress and the maxillary sinus. Am J Phys Anthropol 47: 289–300
Simpson GG (1961) Principles of animal taxonomy. New York, Columbia University Press (20)
Sokal RR, Rohlf FJ (1995) Biometry: the principles and practice of statistics in biological research. New York, W. H. Freeman and Co
Steiper ME, Ruvolo M (2003) New world monkey phylogeny based on X-linked G6PD DNA sequences. Mol Phylogenet Evol 27: 121–130
Stierna P (2001) Physiology, mucociliary clearance and neutral control. In: Kennedy DW, Bolger W, Zinnreich S (Eds.): Disease of the Sinuses: Diagnosis and Management, Hamilton, B. C. Decker: 35–45
Stoner KE (1995) Dental pathology in Pongo satyrus borneensis. Am J Phys Anthropol 98: 307–321
Stuck BA, Bachert C, Federspil P, Hosemann W, Klimek L, Mösges R, Pfaar O, Rudack C, Sitter H, Wagenmann M, Hörmann K (2007) Leitlinie „Rhinosinusitis“ der Deutschen Gesellschaft für Hals-Nasen-Ohren-Heilkunde, Kopf- und Hals-Chirurgie. HNO 58: 272–278
Summers RB (1994) A new concept in maxillary implant surgery: the osteotome technique. Compendium (Newtown, Pa) 15: 152–154
Swindler DR (1999) Maxillary sinuses, dentition, diet, and arch form in some anthropoid primates. In: Koppe T, Nagai H, Alt K (Eds.) The Paranasal Sinuses of Higher Primates: Development, Function and Evolution. Chicago, Quintessence: 191–206
Tatum H (1986) Maxillary and sinus implant reconstructions. Dentistry Clinical North America 30: 207–229
Toffler M (2004) Osteotome-mediated sinus floor elevation: a clinical report. Int J Oral Max Impl 19: 266–273
Underwood AS (1910) An inquiry into the anatomy and pathology of the maxillary sinus. J Anat 44: 354–369
Vogeler M, Bucher R, Schmelzeisen R (2010) Implantologie. In: Gutwald R, Gellrich NC, Schmelzeisen R (Hrsg.) Einführung in die zahnärztliche Chirurgie und Implantologie. Für Studium und Beruf. 2. Aufl. Köln, Deutscher Zahnärzte Verlag: 383–432

Literaturverzeichnis
97
Ward S, Pilbeam D (1983) Maxillofacial morphology of miocene hominoids from Africa and Indo-Pakistan. In: Ciochion R, Corruccini RS (Eds.) New Interpretations of Ape and Human Ancestry. New York, Plenum Press: 211–238
Wegner RN (1962) Abszesse an dern Eckzähnen bei den Anthropoiden. Dtsch Zahn-Mund-Kieferheilkd 37: 433–444
Wegner RN (1955/56) Studien über Nebenhöhlen des Schädels: Verschmelzungen der Nebenhöhle der Nase und Abänderungen ihrer Öffnungen beim Primaten. Wissenschaftliche Zeitschrift der Ernst-Moritz-Arndt-Universität Greifswald, Mathematisch-naturwissenschaftliche Reihe 5: 1–127
Weidenreich F (1924) Über die pneumatischen Nebenräume des Kopfes. Ein Beitrag zur Kenntnis des Bauprinzips der Knochen, des Schädels und des Körpers. Anat Embryol 72: 55–93
Wildmann DE, Uddin M, Liu G, Grossman LI, Goodmann M (2003) Implications of natural selection in shaping 99,4% nonsynonymous DNA identity between humans and chimpanzees: enlarging genus Homo. P Natl Acad Sci USA 100: 7181–7188
Winkler S, Morris HF, Ochi S (2000) Implant survival to 36 months as related to length and diameter. Ann Periodontol 5: 22–31
Zäschke C, Schopf PM (1975) Zur metrischen Analyse von Panorama-Röntgenbildern. Fortschr Kieferorthop 36: 231–243

Anhang
8 Anhang

Anhang
Ta
b.
I: S
chä
de
lma
ße
de
r A
ltw
elta
ffe
n u
nd
Neu
welta
ffe
n
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
1A
WA
Gorilla
G1
020,9
810,9
415,4
410,2
86,3
87,5
23,4
51,5
88,8
537,6
77,7
637,8
85,1
3-0
,51
2A
WA
Gorilla
G2
021,5
210,4
314,9
510,4
56,5
18,0
83,2
81,4
59,4
920,7
32,9
623,1
11,6
1-0
,84
3A
WA
Gorilla
G3
020,9
310,7
815,0
510,2
06,6
98,2
53,1
81,4
09,0
010,0
22,1
712,6
92,3
00,0
6
4A
WA
Gorilla
G4
023,8
611,7
517,6
212,0
57,2
79,4
33,6
32,1
611,0
939,5
67,0
934,5
15,8
7-0
,21
5A
WA
Pan
Litt
10
9,8
613,5
57,2
76,1
97,2
63,0
31,2
17,7
426,6
16,6
132,4
29,5
40,4
4
6A
WA
Pan
W3
016,6
88,6
111,3
27,3
94,8
65,6
12,9
61,0
06,6
011,1
41,9
511,5
81,9
50,0
00,0
0
7A
WA
Pan
W4
017,5
39,1
912,5
78,6
15,9
36,1
02,9
41,1
97,2
315,1
13,6
115,0
93,6
70,0
2
8A
WA
Pan
15.5
.13
m19,4
39,6
914,2
98,5
66,3
47,2
33,1
61,2
78,7
026,7
73,2
926,6
73,6
40,1
1
9A
WA
Pan
Litt
2m
20,2
810,4
214,1
98,6
46,1
27,6
33,1
01,3
48,6
124,7
49,3
030,4
915,0
70,6
2
10
AW
AP
an
15.5
.08
w16,5
48,8
911,1
47,6
85,0
35,5
42,8
40,9
26,8
19,7
51,4
79,2
21,7
30,1
8
11
AW
AP
linka
igalis
957
018,3
19,7
38,3
89,9
56,2
84,0
91,1
75,3
318,6
01,4
919,3
12,3
70,5
9
12
AW
AP
linka
igalis
958
020,1
210,4
810,0
38,9
75,9
44,6
64,0
70,9
46,2
116,3
70,6
318,5
81,1
40,8
1
13
AW
AP
linka
igalis
960
019,5
89,8
29,0
39,4
16,2
84,3
74,2
31,2
76,4
318,0
61,2
920,7
10,9
9-0
,30
14
AW
AP
linka
igalis
963
019,4
010,9
210,1
19,2
55,9
74,5
73,8
90,9
15,9
515,7
50,2
114,4
80,6
11,9
0
15
AW
AP
linka
igalis
1083
020,8
510,5
610,3
19,5
76,6
64,6
84,1
81,3
17,6
023,4
22,5
123,2
31,6
5-0
,52

Anhang
Fort
se
tzun
g T
ab
. I
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
16
AW
AP
linka
igalis
1086
01,0
80,5
40,5
40,0
00,0
0
17
AW
AP
linka
igalis
1077
021,1
010,8
410,4
19,8
16,3
15,0
84,0
01,4
27,1
814,4
11,0
718,1
11,0
0-0
,07
18
AW
AP
linka
igalis
1079
020,3
411,0
110,5
09,8
96,8
15,0
34,4
61,1
06,7
526,3
02,6
427,9
32,7
70,0
5
19
AW
AP
linka
igalis
1080
020,1
910,3
910,3
49,8
96,1
94,9
73,5
91,0
07,4
019,1
00,7
423,9
30,6
9-0
,07
20
AW
AP
linka
igalis
1081
020,2
810,3
49,6
49,4
06,4
64,2
13,9
01,4
27,0
014,7
71,1
315,9
51,2
60,1
2
21
AW
AP
linka
igalis
1082
020,3
910,4
210,0
89,8
46,7
34,7
84,3
81,5
96,9
125,1
13,4
928,9
34,5
60,3
1
22
AW
AP
linka
igalis
1094
020,8
29,9
210,2
69,6
96,6
25,2
54,5
61,2
96,2
624,6
81,7
329,7
15,1
01,9
5
23
AW
AP
linka
igalis
1095
020,7
910,3
410,4
19,5
56,2
85,2
14,2
81,2
06,7
216,8
60,1
122,5
80,3
42,0
9
24
AW
AP
linka
igalis
1096
019,7
610,2
710,0
29,6
76,7
14,7
54,2
31,2
17,0
620,4
41,6
318,1
51,1
2-0
,46
25
AW
AP
linka
igalis
1226
021,2
011,0
410,7
39,0
46,2
04,9
13,8
41,1
46,7
815,9
51,8
415,5
51,3
0-0
,42
26
AW
AP
linka
igalis
1228
020,8
810,5
210,0
68,5
25,8
64,7
14,0
21,2
56,9
215,3
10,5
716,2
60,3
2-0
,78
27
AW
AA
lytu
s3574
018,7
99,6
110,1
89,2
05,6
85,1
43,7
51,0
36,1
210,6
90,0
311,7
20,2
05,6
7
28
AW
AA
lytu
s3577
019,0
110,0
18,5
29,8
06,2
95,1
44,2
00,8
56,6
521,7
71,0
723,3
30,4
7-1
,28
29
AW
AA
lytu
s3593
020,8
610,3
410,0
410,0
66,7
35,3
84,2
81,2
56,7
428,3
92,4
431,1
22,5
00,0
2
30
AW
AA
lytu
s3595
018,8
39,3
49,1
29,6
25,5
54,4
83,9
20,7
86,2
112,5
30,0
612,1
00,0
2-2
,00

Anhang
Fort
se
tzun
g T
ab
. I
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
31
AW
AA
lytu
s3601
020,5
410,0
710,0
39,1
36,6
55,0
84,4
70,8
86,5
421,8
10,7
211,1
70,0
1-7
1,0
0
32
AW
AA
lytu
s3605
018,6
59,4
08,7
88,8
25,5
54,2
53,5
40,8
56,2
516,6
41,0
916,1
20,3
2-2
,41
33
AW
AA
lytu
s3607
019,7
39,4
19,2
49,1
75,9
74,7
33,9
20,8
76,5
012,5
30,5
117,7
41,4
11,7
6
34
AW
AA
lytu
s3610
019,0
39,7
18,7
38,3
35,5
94,4
13,7
20,8
86,1
711,6
00,0
49,1
3
35
AW
AA
lytu
s3613
020,3
09,3
89,4
49,1
56,3
84,8
94,3
91,0
96,9
016,3
01,2
916,3
20,6
6-0
,95
36
AW
AA
lytu
s3617
020,9
510,1
810,2
99,9
66,9
15,5
24,1
21,5
57,7
724,4
13,7
322,1
23,3
6-0
,11
37
AW
AP
linka
igalis
813
m20,8
110,9
310,2
09,5
46,6
04,8
04,5
41,4
87,7
730,8
93,0
135,4
66,1
91,0
6
38
AW
AP
linka
igalis
814
m20,5
210,0
29,4
59,2
86,2
84,7
54,0
11,4
77,5
232,5
25,8
933,9
86,4
80,1
0
39
AW
AP
linka
igalis
817
m20,1
110,1
16,4
14,6
34,4
61,2
47,7
125,4
51,8
424,0
51,1
7-0
,57
40
AW
AP
linka
igalis
959
020,1
19,7
99,4
79,3
16,2
14,5
33,9
86,1
1
41
AW
AS
ymphala
ngus s
yndacty
lus
M4099307
m13,7
08,1
010,8
17,0
24,4
85,2
52,5
41,0
26,4
014,5
42,4
613,5
22,2
9-0
,07
42
AW
AS
ymphala
ngus s
yndacty
lus
M4099309
m12,7
07,9
09,7
36,6
44,2
04,8
12,4
20,7
06,3
05,6
01,5
55,5
11,1
9-0
,30
43
AW
AP
ongo s
aty
rus b
orn
eensis
1981/1
86
m23,3
010,0
017,2
011,7
27,8
08,2
04,5
01,7
510,0
024,2
20,6
323,5
90,1
8-2
,50
44
AW
AP
ongo s
aty
rus b
orn
eensis
1981/2
01
w18,5
08,7
013,2
010,4
06,2
06,2
53,6
21,2
28,6
020,9
20,4
323,2
30,9
11,1
2
45
NW
AA
louatta c
ara
yaA
M 0
m12,5
27,5
810,3
35,7
74,2
54,1
62,4
70,9
03,5
92,1
80,2
62,2
60,2
1-0
,24

Anhang
Fort
se
tzun
g T
ab
. I
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
46
NW
AA
louatta c
ara
yaA
M 1
342
m12,6
37,6
410,8
06,2
54,6
44,1
12,4
00,9
03,6
81,7
40,2
11,9
70,1
8-0
,17
47
NW
AA
louatta c
ara
yaA
M 1
359
m11,8
37,1
210,1
25,9
74,2
84,1
62,2
90,7
03,3
21,5
90,0
81,6
10,0
5-0
,60
48
NW
AA
louatta c
ara
ya1925/4
21
m12,5
07,4
410,3
76,0
24,6
63,9
72,5
00,9
53,3
03,4
10,4
13,1
30,4
80,1
7
49
NW
AA
louatta c
ara
ya1925/2
43
m11,7
77,2
710,2
76,2
64,5
53,9
62,3
80,8
03,2
63,0
80,3
73,0
70,3
4-0
,09
50
NW
AA
louatta b
elz
ebul
363
m12,0
16,9
810,2
46,1
24,6
14,2
52,4
30,6
32,5
515,0
74,4
014,8
03,2
2-0
,37
51
NW
AA
louatta fusca
720
m12,0
07,6
810,7
06,5
04,3
14,0
52,4
50,8
02,3
06,7
41,6
07,1
90,8
4-0
,90
52
NW
AA
louatta s
enic
ulu
s365
m12,6
47,4
210,7
27,2
84,2
64,2
02,4
50,8
62,2
510,6
42,3
611,6
22,9
90,2
7
53
NW
AA
louatta c
ara
ya1925/2
44
w10,1
46,0
98,3
15,2
93,7
73,2
41,9
20,7
02,5
71,3
50,2
51,1
90,1
5-0
,67
54
NW
AA
louatta c
ara
ya1925/2
48
w10,0
65,9
88,0
85,0
03,8
33,3
42,1
30,7
02,6
30,8
60,0
31,2
00,0
61,0
0
55
NW
AA
louatta c
ara
ya1925/3
09
w10,5
56,2
68,6
45,3
73,8
13,4
12,1
10,7
02,8
42,0
00,1
61,9
60,1
3-0
,23
56
NW
AA
louatta c
ara
ya1925/3
23
w10,2
75,0
18,5
65,1
73,8
83,5
22,0
70,6
02,8
71,6
00,1
51,5
30,1
70,1
3
57
NW
AA
louatta c
ara
ya1925/3
25
w10,6
86,4
08,6
75,3
83,9
63,4
42,0
40,5
03,0
11,3
20,0
21,4
10,0
30,5
0
58
NW
AA
louatta b
elz
ebul
364
w10,1
16,0
98,2
25,7
73,9
53,4
22,2
50,5
91,8
97,8
71,4
87,0
41,3
1-0
,13
59
NW
AA
louatta fusca
721
w10,2
16,0
28,2
05,5
63,6
13,3
11,9
70,5
62,1
05,4
60,7
85,2
40,7
0-0
,11
60
NW
AA
louatta s
enic
ulu
s367
w10,3
06,0
68,3
05,3
13,6
53,2
21,8
60,6
61,8
84,0
81,4
64,8
81,3
0-0
,12

Anhang
Fort
se
tzun
g T
ab
. I
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
61
NW
AA
tele
s g
eoffro
yi1972/1
93
m11,6
46,2
78,2
65,1
63,7
93,1
82,2
90,7
03,2
73,4
80,8
43,5
30,9
30,1
1
62
NW
AA
tele
s fuscic
eps
1960/1
72
w11,7
14,6
13,3
42,8
42,0
20,5
02,7
51,8
10,3
71,5
10,3
5-0
,06
63
NW
AA
tele
s g
eoffro
yi1953/8
2w
10,3
35,3
76,3
83,9
43,3
02,4
41,7
50,5
02,8
10,6
80,0
40,7
10,0
60,5
0
64
NW
AA
tele
s p
anis
cus
1962/2
54
w11,4
96,1
27,6
84,3
83,2
63,0
21,9
10,3
02,6
81,1
20,0
61,1
40,0
5-0
,20
65
NW
AA
tele
s p
anis
cus
1962/2
84
w11,8
16,8
38,6
245,7
03,1
02,8
91,8
70,5
03,1
61,3
60,2
01,2
00,1
6-0
,25
66
NW
AB
rachyt
ele
s a
rachnoid
es
1944/3
005
w11,5
26,6
58,0
94,1
13,3
21,9
20,5
03,2
82,0
10,0
31,9
70,0
30,0
00,0
0
67
NW
ALagoth
rix
lagotr
icha
AM
/1319
m11,1
46,7
58,3
15,2
535,2
02,9
41,8
50,6
03,3
01,1
40,1
61,2
90,1
90,1
9
68
NW
ALagoth
rix
lagotr
icha
AM
/1322
m10,7
46,5
38,1
05,1
63,3
73,0
11,8
20,8
03,2
01,8
60,4
51,6
70,4
70,0
4
69
NW
ALagoth
rix
lagotr
icha
AM
/1325
m10,9
36,3
47,8
44,8
93,3
62,5
81,7
50,6
03,4
01,7
30,3
51,6
60,2
8-0
,25
70
NW
ALagoth
rix
lagotr
icha
AM
/1327
m10,4
26,5
97,7
45,1
53,2
52,7
31,8
10,6
03,0
01,3
20,2
21,3
10,2
1-0
,05
71
NW
ALagoth
rix
lagotr
icha
AM
/1330
m10,8
56,6
28,3
75,0
43,3
23,0
11,8
73,3
01,5
90,2
41,6
30,2
1-0
,14
72
NW
ALagoth
rix
lagotr
icha
AM
/1321
w9,9
96,1
27,4
34,6
03,2
52,5
11,7
80,5
03,0
01,0
20,0
51,0
50,0
3-0
,67
73
NW
ALagoth
rix
lagotr
icha
AM
/1326
w10,6
36,5
78,0
54,7
13,1
22,7
51,6
40,6
03,4
01,8
40,2
11,6
00,1
9-0
,11
74
NW
ALagoth
rix
lagotr
icha
AM
/1329
w10,2
46,3
27,5
54,7
23,2
02,8
01,7
20,6
03,1
01,4
20,2
11,2
80,1
5-0
,40
75
NW
ALagoth
rix
lagotr
icha
AM
/1332
w10,5
06,2
17,7
34,7
43,3
42,6
51,8
00,6
03,3
01,6
10,2
21,5
20,1
7-0
,29

Anhang
Fort
se
tzun
g T
ab
. I
Nr:
N
um
me
r, A
WA
: A
ltw
elta
ffen
, N
WA
: N
euw
elta
ffen
, ID
: Id
en
tifika
tio
nsn
um
me
r, S
ex:
Ge
sch
lech
t, m
: m
än
nlic
h,
w:
weib
lich
, 0:
unb
ekan
nt,
SL:
Ge
sam
tsch
äd
ellä
nge
, S
BL
: S
ch
äde
lba
sis
län
ge
, G
SL
: G
esic
hts
län
ge
,
MG
B:
Mitte
lge
sic
hts
bre
ite
, M
AB
: M
axill
oa
lveo
larb
reite
, G
L:
Ga
um
en
län
ge
, G
B:
Ga
um
enb
reite
, G
H:
Gau
me
nhö
he,
OG
H:
Obe
rge
sic
hts
höh
e,
SM
R:
Vo
lum
en
de
s
rech
ten
S
inu
s
maxill
aris,
RA
R:
Vo
lum
en
d
es
rech
ten
R
ece
ssu
s
alv
eo
laris,
SM
L:
Vo
lum
en
de
s
linken
S
inu
s
ma
xill
aris,
RA
L:
Vo
lum
en
de
s
linken
R
ece
ssu
s
alv
eo
laris,
AI
RE
: A
sym
me
trie
inde
x,
wenn
de
r re
ch
te R
ece
ssu
s a
lveo
laris r
ech
ts g
röß
er
ist, A
I LI:
Asym
me
trie
ind
ex,
wen
n d
er
linke
Re
ce
ssu
s a
lveo
laris g
röß
er
ist.
Die
bla
u m
ark
iert
en
Wert
e e
nts
pre
ch
en
de
r eig
en
en
Da
tene
rheb
ung
.
Nr
Gru
ppe
Gattung
IDS
ex
SL
SB
LG
SL
MG
BM
AB
GL
GB
GH
OG
HS
MR
RA
RS
ML
RA
LA
I RE
AI L
I
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m³)
(cm
³)(c
m³)
(cm
³)(c
m)
(cm
)
76
NW
ALagoth
rix
lagotr
icha
AM
/1334
w10,2
46,0
47,3
44,6
53,3
22,4
81,7
30,6
03,0
01,6
30,2
11,6
20,1
8-0
,17
77
NW
AC
ebus
13
09,1
25,3
96,3
14,1
23,0
93,7
41,9
70,6
73,0
21,8
50,3
81,6
80,3
80,0
00,0
0
78
NW
AC
ebus
14
09,6
36,0
46,7
24,3
53,1
72,5
61,7
90,6
03,1
11,9
80,5
22,0
30,3
5-0
,49
79
NW
AC
ebus
15
010,2
26,2
67,3
24,8
83,4
42,7
71,5
60,4
53,4
12,4
80,4
72,2
90,4
2-0
,12
80
NW
AC
ebus a
pella
1368
m9,9
96,0
77,2
35,9
23,4
43,0
51,8
20,6
03,1
61,1
60,2
61,1
30,2
1-0
,24
81
NW
AC
ebus a
pella
1903/8
2m
9,7
25,4
86,5
05,5
63,2
32,8
21,8
10,4
02,8
71,1
00,1
51,0
30,1
50,0
00,0
0
82
NW
AC
ebus a
pella
1904/1
882
m9,1
45,4
96,2
65,1
93,2
32,7
81,6
60,4
02,7
01,0
80,0
81,1
80,1
10,3
8
83
NW
AC
ebus a
pella
1953/3
9m
9,7
15,6
96,6
85,6
03,1
73,0
11,8
00,5
02,7
01,3
80,1
81,2
70,1
5-0
,20
84
NW
AC
ebus a
pella
1982/1
19
m9,7
05,8
46,5
95,4
23,1
72,8
91,6
70,6
02,6
30,8
60,2
30,8
40,2
0-0
,15
85
NW
AC
ebus a
pella
AM
1491
w8,9
75,0
95,7
44,7
22,9
82,4
51,4
80,4
02,4
20,7
40,0
90,7
30,0
6-0
,50
86
NW
AC
ebus a
pella
1903/9
3w
8,6
95,2
45,7
94,6
93,0
42,7
11,5
90,4
02,3
80,7
00,0
80,7
20,0
7-0
,14
87
NW
AC
ebus a
pella
1957/1
23
w8,8
45,3
16,0
84,9
72,9
22,8
11,5
20,4
02,7
51,1
20,1
81,1
10,2
00,1
1
88
NW
AC
ebus a
pella
1982/1
16
w8,9
65,2
75,9
14,8
52,9
52,6
51,5
70,5
02,5
90,9
00,1
60,9
30,1
2-0
,33
89
NW
AC
ebus a
pella
1982/1
18
w8,9
25,4
96,1
84,8
33,0
72,8
01,5
90,3
02,4
30,7
10,1
00,7
30,0
9-0
,11
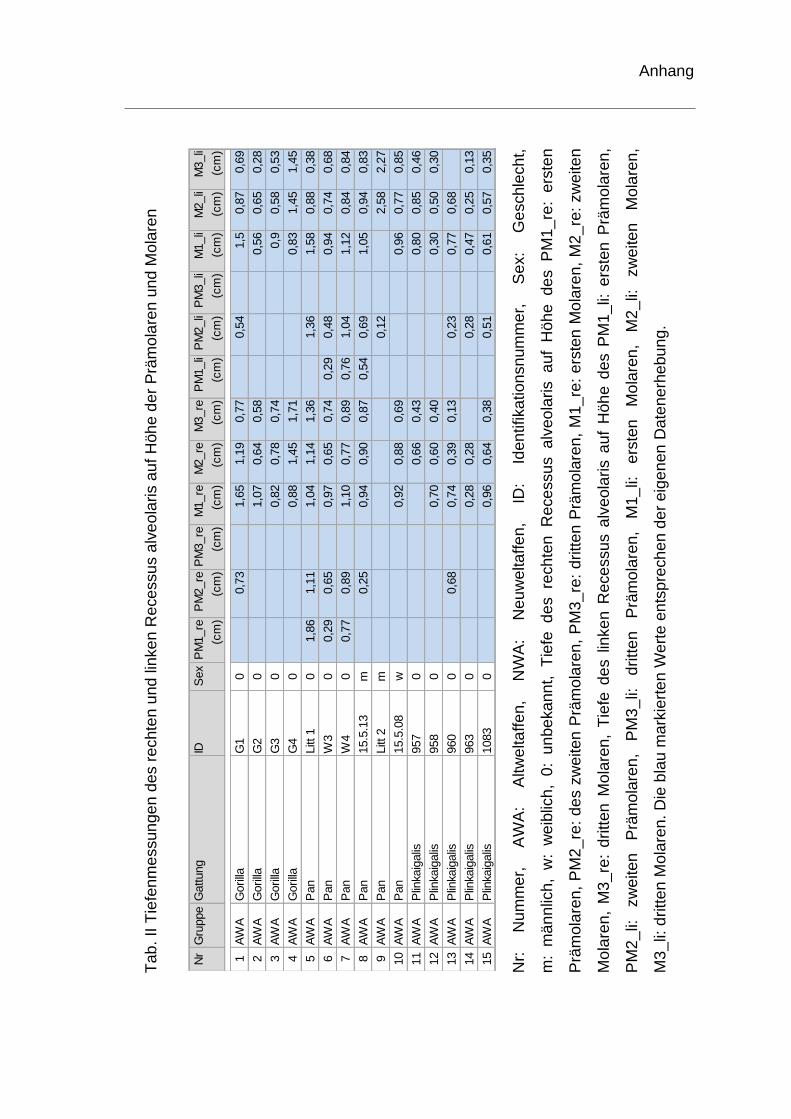
Anhang
Tab
. II T
iefe
nm
essun
ge
n d
es r
echte
n u
nd
lin
ke
n R
ece
ssu
s a
lve
ola
ris a
uf
Höh
e d
er
Prä
mo
lare
n u
nd
Mola
ren
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dritte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Date
ne
rhe
bu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1_re
PM
2_re
PM
3_re
M1_re
M2_re
M3_re
PM
1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
1A
WA
Gorilla
G1
00,7
31,6
51,1
90,7
70,5
41,5
0,8
70,6
9
2A
WA
Gorilla
G2
01,0
70,6
40,5
80,5
60,6
50,2
8
3A
WA
Gorilla
G3
00,8
20,7
80,7
40,9
0,5
80,5
3
4A
WA
Gorilla
G4
00,8
81,4
51,7
10,8
31,4
51,4
5
5A
WA
Pan
Litt
10
1,8
61,1
11,0
41,1
41,3
61,3
61,5
80,8
80,3
8
6A
WA
Pan
W3
00,2
90,6
50,9
70,6
50,7
40,2
90,4
80,9
40,7
40,6
8
7A
WA
Pan
W4
00,7
70,8
91,1
00,7
70,8
90,7
61,0
41,1
20,8
40,8
4
8A
WA
Pan
15.5
.13
m0,2
50,9
40,9
00,8
70,5
40,6
91,0
50,9
40,8
3
9A
WA
Pan
Litt
2m
0,1
22,5
82,2
7
10
AW
AP
an
15.5
.08
w0,9
20,8
80,6
90,9
60,7
70,8
5
11
AW
AP
linka
igalis
957
00,6
60,4
30,8
00,8
50,4
6
12
AW
AP
linka
igalis
958
00,7
00,6
00,4
00,3
00,5
00,3
0
13
AW
AP
linka
igalis
960
00,6
80,7
40,3
90,1
30,2
30,7
70,6
8
14
AW
AP
linka
igalis
963
00,2
80,2
80,2
80,4
70,2
50,1
3
15
AW
AP
linka
igalis
1083
00,9
60,6
40,3
80,5
10,6
10,5
70,3
5
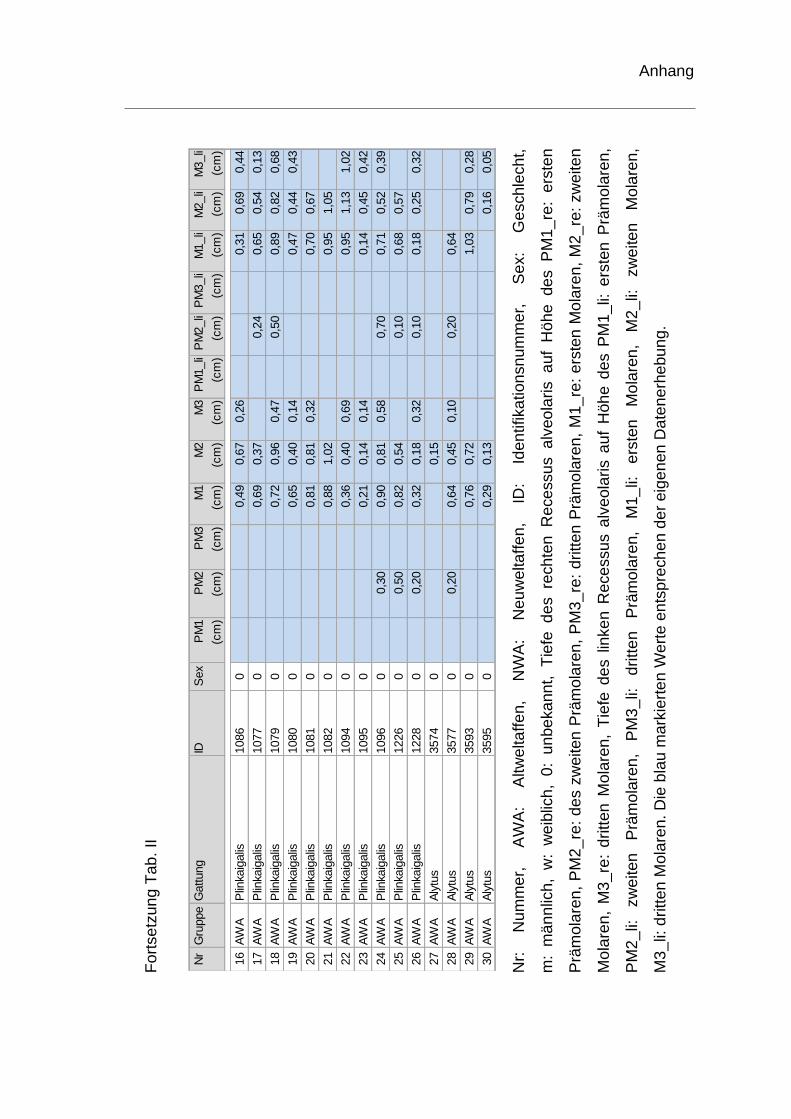
Anhang
Fort
se
tzun
g T
ab
. II
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dritte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Da
ten
erh
ebu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1P
M2
PM
3M
1M
2M
3P
M1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
16
AW
AP
linka
igalis
1086
00,4
90,6
70,2
60,3
10,6
90,4
4
17
AW
AP
linka
igalis
1077
00,6
90,3
70,2
40,6
50,5
40,1
3
18
AW
AP
linka
igalis
1079
00,7
20,9
60,4
70,5
00,8
90,8
20,6
8
19
AW
AP
linka
igalis
1080
00,6
50,4
00,1
40,4
70,4
40,4
3
20
AW
AP
linka
igalis
1081
00,8
10,8
10,3
20,7
00,6
7
21
AW
AP
linka
igalis
1082
00,8
81,0
20,9
51,0
5
22
AW
AP
linka
igalis
1094
00,3
60,4
00,6
90,9
51,1
31,0
2
23
AW
AP
linka
igalis
1095
00,2
10,1
40,1
40,1
40,4
50,4
2
24
AW
AP
linka
igalis
1096
00,3
00,9
00,8
10,5
80,7
00,7
10,5
20,3
9
25
AW
AP
linka
igalis
1226
00,5
00,8
20,5
40,1
00,6
80,5
7
26
AW
AP
linka
igalis
1228
00,2
00,3
20,1
80,3
20,1
00,1
80,2
50,3
2
27
AW
AA
lytu
s3574
00,1
5
28
AW
AA
lytu
s3577
00,2
00,6
40,4
50,1
00,2
00,6
4
29
AW
AA
lytu
s3593
00,7
60,7
21,0
30,7
90,2
8
30
AW
AA
lytu
s3595
00,2
90,1
30,1
60,0
5
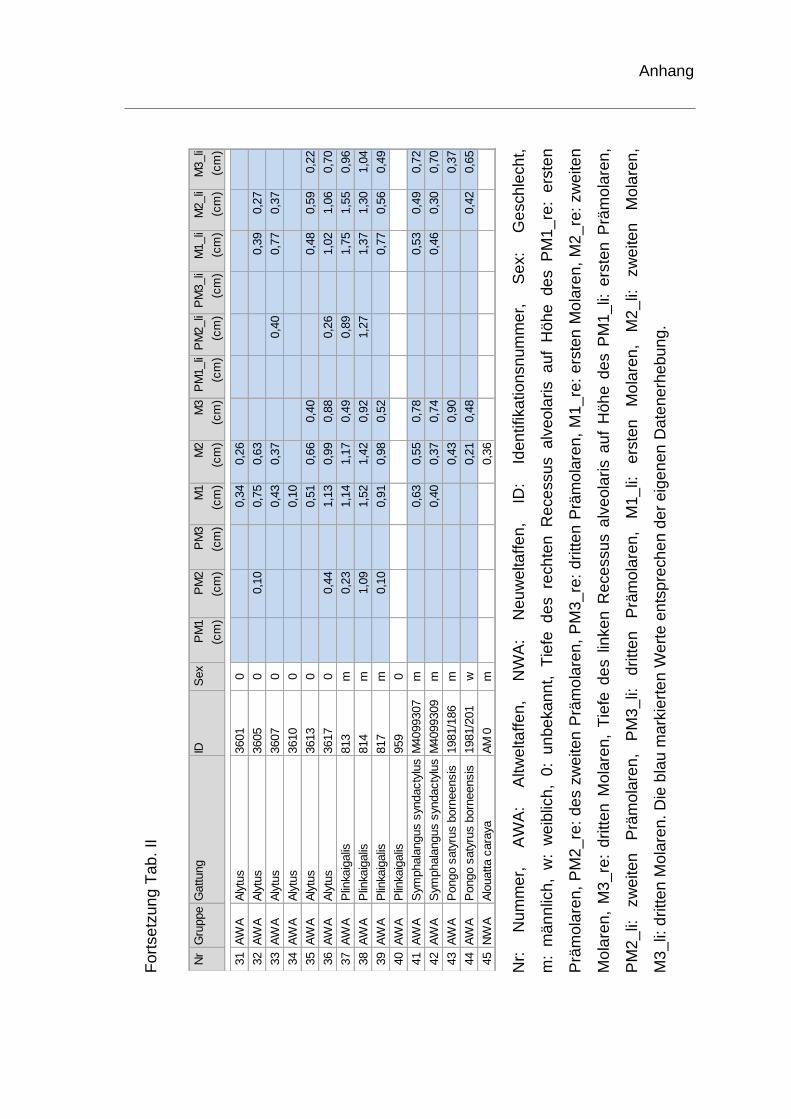
Anhang
Fort
se
tzun
g T
ab
. II
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dritte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Date
ne
rhe
bu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1P
M2
PM
3M
1M
2M
3P
M1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
31
AW
AA
lytu
s3601
00,3
40,2
6
32
AW
AA
lytu
s3605
00,1
00,7
50,6
30,3
90,2
7
33
AW
AA
lytu
s3607
00,4
30,3
70,4
00,7
70,3
7
34
AW
AA
lytu
s3610
00,1
0
35
AW
AA
lytu
s3613
00,5
10,6
60,4
00,4
80,5
90,2
2
36
AW
AA
lytu
s3617
00,4
41,1
30,9
90,8
80,2
61,0
21,0
60,7
0
37
AW
AP
linka
igalis
813
m0,2
31,1
41,1
70,4
90,8
91,7
51,5
50,9
6
38
AW
AP
linka
igalis
814
m1,0
91,5
21,4
20,9
21,2
71,3
71,3
01,0
4
39
AW
AP
linka
igalis
817
m0,1
00,9
10,9
80,5
20,7
70,5
60,4
9
40
AW
AP
linka
igalis
959
0
41
AW
AS
ymphala
ngus s
yndacty
lus
M4099307
m0,6
30,5
50,7
80,5
30,4
90,7
2
42
AW
AS
ymphala
ngus s
yndacty
lus
M4099309
m0,4
00,3
70,7
40,4
60,3
00,7
0
43
AW
AP
ongo s
aty
rus b
orn
eensis
1981/1
86
m0,4
30,9
00,3
7
44
AW
AP
ongo s
aty
rus b
orn
eensis
1981/2
01
w0,2
10,4
80,4
20,6
5
45
NW
AA
louatta c
ara
yaA
M 0
m0,3
6
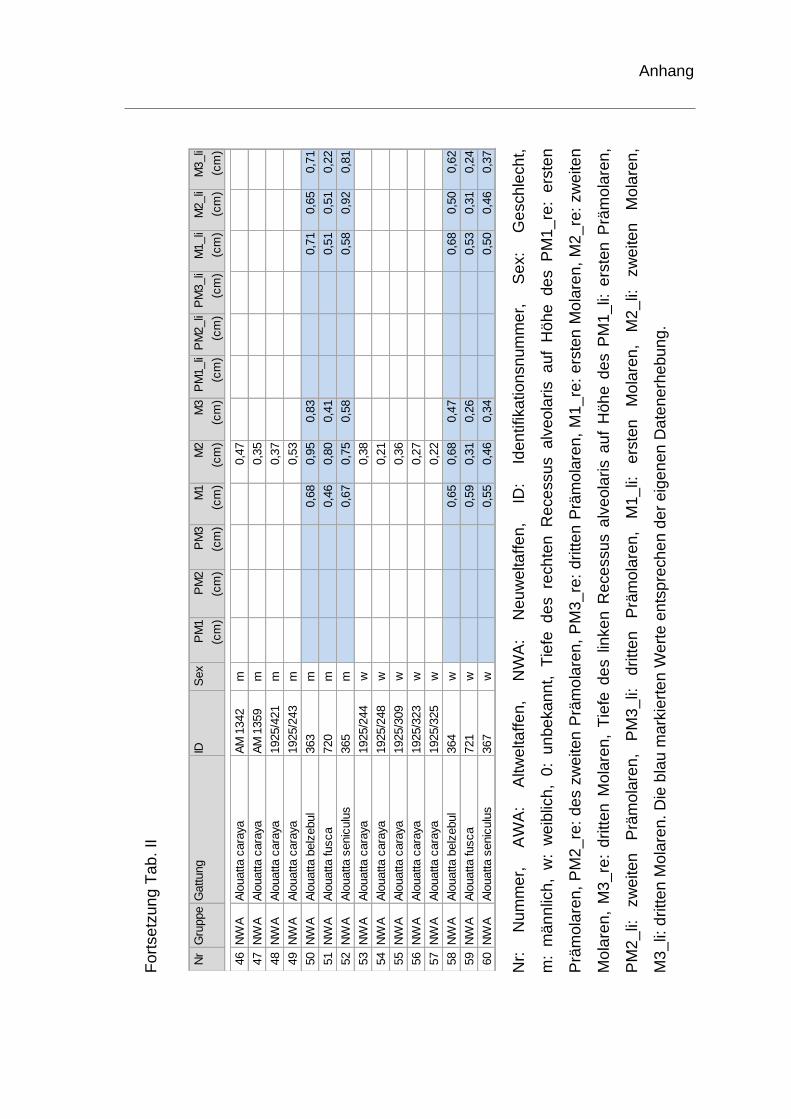
Anhang
Fort
se
tzun
g T
ab
. II
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dritte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Da
ten
erh
ebu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1P
M2
PM
3M
1M
2M
3P
M1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
46
NW
AA
louatta c
ara
yaA
M 1
342
m0,4
7
47
NW
AA
louatta c
ara
yaA
M 1
359
m0,3
5
48
NW
AA
louatta c
ara
ya1925/4
21
m0,3
7
49
NW
AA
louatta c
ara
ya1925/2
43
m0,5
3
50
NW
AA
louatta b
elz
ebul
363
m0,6
80,9
50,8
30,7
10,6
50,7
1
51
NW
AA
louatta fusca
720
m0,4
60,8
00,4
10,5
10,5
10,2
2
52
NW
AA
louatta s
enic
ulu
s365
m0,6
70,7
50,5
80,5
80,9
20,8
1
53
NW
AA
louatta c
ara
ya1925/2
44
w0,3
8
54
NW
AA
louatta c
ara
ya1925/2
48
w0,2
1
55
NW
AA
louatta c
ara
ya1925/3
09
w0,3
6
56
NW
AA
louatta c
ara
ya1925/3
23
w0,2
7
57
NW
AA
louatta c
ara
ya1925/3
25
w0,2
2
58
NW
AA
louatta b
elz
ebul
364
w0,6
50,6
80,4
70,6
80,5
00,6
2
59
NW
AA
louatta fusca
721
w0,5
90,3
10,2
60,5
30,3
10,2
4
60
NW
AA
louatta s
enic
ulu
s367
w0,5
50,4
60,3
40,5
00,4
60,3
7
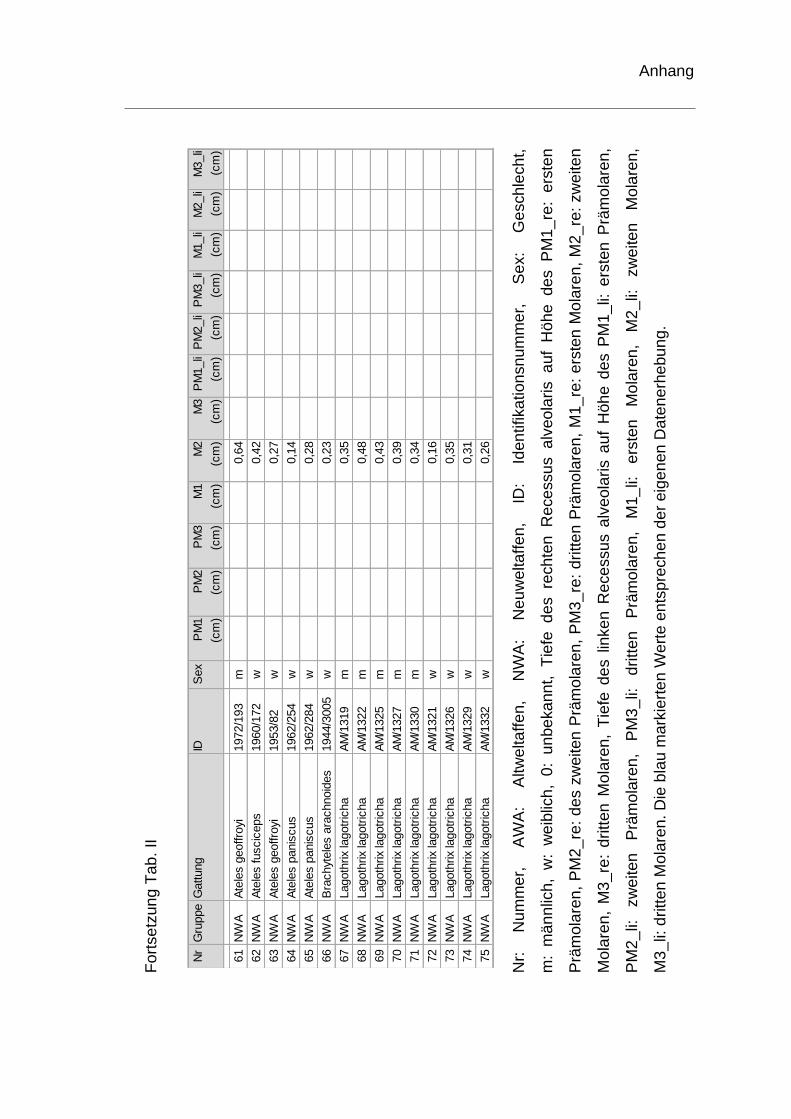
Anhang
Fort
se
tzun
g T
ab
. II
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dri
tte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Da
ten
erh
ebu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1P
M2
PM
3M
1M
2M
3P
M1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
61
NW
AA
tele
s g
eoffro
yi1972/1
93
m0,6
4
62
NW
AA
tele
s fuscic
eps
1960/1
72
w0,4
2
63
NW
AA
tele
s g
eoffro
yi1953/8
2w
0,2
7
64
NW
AA
tele
s p
anis
cus
1962/2
54
w0,1
4
65
NW
AA
tele
s p
anis
cus
1962/2
84
w0,2
8
66
NW
AB
rachyt
ele
s a
rachnoid
es
1944/3
005
w0,2
3
67
NW
ALagoth
rix
lagotr
icha
AM
/1319
m0,3
5
68
NW
ALagoth
rix
lagotr
icha
AM
/1322
m0,4
8
69
NW
ALagoth
rix
lagotr
icha
AM
/1325
m0,4
3
70
NW
ALagoth
rix
lagotr
icha
AM
/1327
m0,3
9
71
NW
ALagoth
rix
lagotr
icha
AM
/1330
m0,3
4
72
NW
ALagoth
rix
lagotr
icha
AM
/1321
w0,1
6
73
NW
ALagoth
rix
lagotr
icha
AM
/1326
w0,3
5
74
NW
ALagoth
rix
lagotr
icha
AM
/1329
w0,3
1
75
NW
ALagoth
rix
lagotr
icha
AM
/1332
w0,2
6
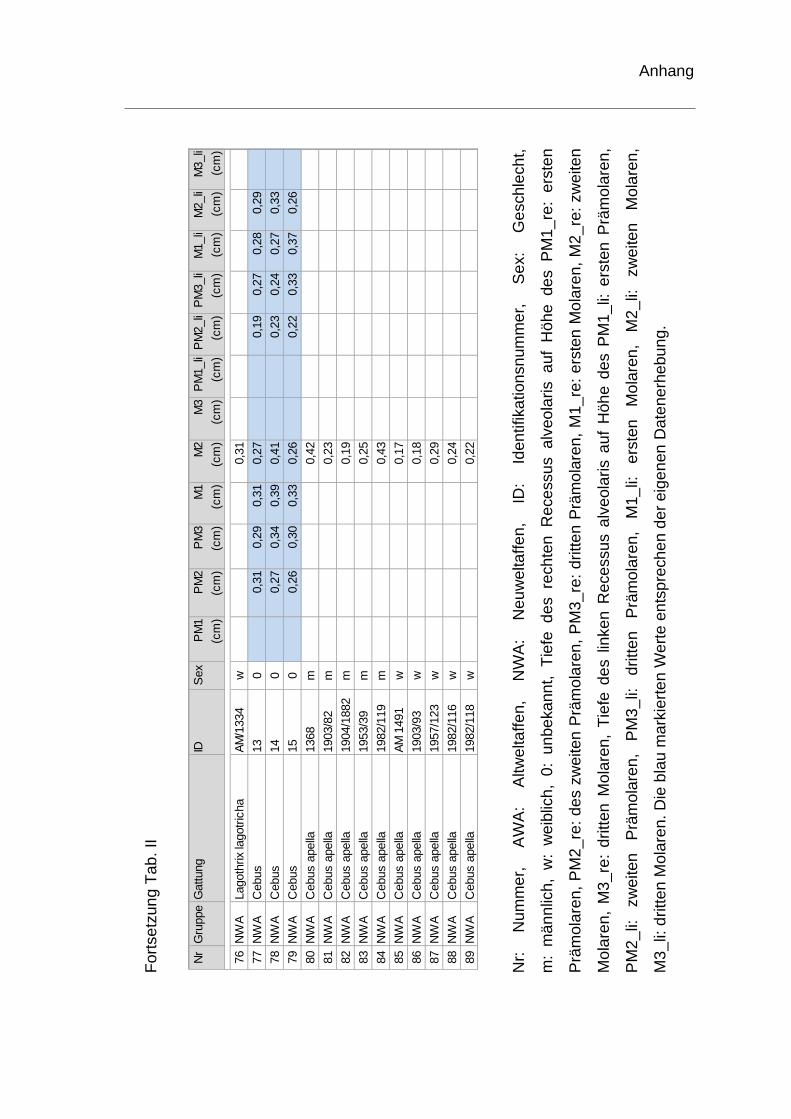
Anhang
Fort
se
tzun
g T
ab
. II
Nr:
N
um
me
r,
AW
A:
Altw
elta
ffe
n,
NW
A:
Neu
welta
ffe
n,
ID:
Iden
tifika
tion
sn
um
me
r,
Se
x:
Ge
schle
ch
t,
m:
mä
nn
lich
, w
: w
eib
lich
, 0
: u
nbe
ka
nn
t, T
iefe
d
es re
ch
ten
R
ece
ssu
s a
lveo
laris a
uf
Höh
e de
s P
M1_
re:
ers
ten
Prä
mola
ren
, P
M2
_re
: d
es z
weiten
Prä
mola
ren
, P
M3_
re:
dritte
n P
räm
ola
ren
, M
1_
re:
ers
ten
Mo
lare
n,
M2
_re
: zw
eite
n
Mo
lare
n,
M3_
re:
dritt
en
Mo
lare
n,
Tie
fe d
es l
inke
n R
ece
ssu
s a
lveola
ris a
uf
Höh
e d
es P
M1
_li:
ers
ten
Prä
mola
ren
,
PM
2_
li:
zw
eiten
P
räm
ola
ren
, P
M3
_li:
d
ritt
en
P
räm
ola
ren,
M1_
li:
ers
ten
M
ola
ren
, M
2_
li:
zw
eite
n
Mo
lare
n,
M3
_li:
dritt
en M
ola
ren
. D
ie b
lau
mark
iert
en
We
rte e
nts
pre
ch
en d
er
eig
en
en
Date
ne
rhe
bu
ng.
Nr
Gru
ppe
Gattung
IDS
ex
PM
1P
M2
PM
3M
1M
2M
3P
M1_li
PM
2_li
PM
3_li
M1_li
M2_li
M3_li
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
(cm
)(c
m)
76
NW
ALagoth
rix
lagotr
icha
AM
/1334
w0,3
1
77
NW
AC
ebus
13
00,3
10,2
90,3
10,2
70,1
90,2
70,2
80,2
9
78
NW
AC
ebus
14
00,2
70,3
40,3
90,4
10,2
30,2
40,2
70,3
3
79
NW
AC
ebus
15
00,2
60,3
00,3
30,2
60,2
20,3
30,3
70,2
6
80
NW
AC
ebus a
pella
1368
m0,4
2
81
NW
AC
ebus a
pella
1903/8
2m
0,2
3
82
NW
AC
ebus a
pella
1904/1
882
m0,1
9
83
NW
AC
ebus a
pella
1953/3
9m
0,2
5
84
NW
AC
ebus a
pella
1982/1
19
m0,4
3
85
NW
AC
ebus a
pella
AM
1491
w0,1
7
86
NW
AC
ebus a
pella
1903/9
3w
0,1
8
87
NW
AC
ebus a
pella
1957/1
23
w0,2
9
88
NW
AC
ebus a
pella
1982/1
16
w0,2
4
89
NW
AC
ebus a
pella
1982/1
18
w0,2
2

Anhang
Tab. III: Pearson-Korrelation der Altweltaffen
SL: Gesamtschädellänge, SBL: Schädelbasislänge, GSL: Gesichtslänge,
MGB: Mittelgesichtsbreite, MAB: Maxilloalveolarbreite, GL: Gaumenlänge,
GB: Gaumenbreite, GH: Gaumenhöhe, OGH: Obergesichtshöhe,
SMR: Volumen des rechten Sinus maxillaris, RAR: Volumen des rechten
Recessus alveolaris, SML: Volumen des linken Sinus maxillaris, RAL: Volumen
des linken Recessus alveolaris.
SL SBL GSL MGB MAB GL GB GH OGH SMR RAR SML RAL
r 1 0,848*** 0,287 0,839*** 0,904*** 0,264 0,707*** 0,639*** 0,462** 0,646*** 0,089 0,652*** 0,017
p 0,000 0,068 0,000 0,000 0,095 0,000 0,000 0,002 0,000 0,579 0,000 0,917
r 0,848*** 1 0,214 0,658*** 0,763*** 0,183 0,564*** 0,560*** 0,344* 0,570*** 0,18 0,569*** 0,151
p 0,000 0,174 0,000 0,000 0,251 0,000 0,000 0,026 0,000 0,260 0,000 0,354
r 0,287 0,214 1 0,266 0,292 0,956*** -0,348* 0,596*** 0,880*** 0,337* 0,416** 0,334* 0,318*
p 0,068 0,174 0,088 0,061 0,000 0,024 0,000 0,000 0,031 0,007 0,033 0,046
r 0,839*** 0,658*** 0,266 1 0,845*** 0,274 0,663*** 0,601*** 0,454** 0,566*** 0,011 0,559*** -0,079
p 0,000 0,000 0,088 0,000 0,079 0,000 0,000 0,002 0,000 0,945 0,000 0,622
r 0,904*** 0,763*** 0,292 0,845*** 1 0,297 0,717*** 0,694*** 0,476** 0,717*** 0,218 0,693*** 0,089
p 0,000 0,000 0,061 0,000 0,056 0,000 0,000 0,001 0,000 0,165 0,000 0,582
r 0,264 0,183 0,956*** 0,274 0,297 1 -0,353* 0,546*** 0,847*** 0,329* 0,434** 0,339* 0,344*
p 0,095 0,251 0,000 0,079 0,056 0,022 0,000 0,000 0,036 0,005 0,030 0,030
r 0,707*** 0,564*** -0,348* 0,663*** 0,717*** -0,353* 1 0,247 -0,124 0,458** -0,21 0,418** -0,268
p 0,000 0,000 0,024 0,000 0,000 0,022 0,114 0,429 0,002 0,182 0,006 0,090
r 0,639*** 0,560*** 0,596*** 0,601*** 0,694*** 0,546*** 0,247 1 0,722*** 0,660*** 0,524*** 0,690*** 0,500**
p 0,000 0,000 0,000 0,000 0,000 0,000 0,114 0,000 0,000 0,000 0,000 0,001
r 0,462** 0,344* 0,880*** 0,454** 0,476** 0,847*** -0,124 0,722*** 1 0,494** 0,479** 0,490** 0,321*
p 0,002 0,026 0,000 0,002 0,001 0,000 0,429 0,000 0,001 0,001 0,001 0,041
r 0,646*** 0,570*** 0,337* 0,566*** 0,717*** 0,329* 0,458** 0,660*** 0,494** 1 0,519*** 0,920*** 0,356*
p 0,000 0,000 0,031 0,000 0,000 0,036 0,002 0,000 0,001 0,000 0,000 0,022
r 0,089 0,18 0,416** 0,011 0,218 0,434** -0,21 0,524*** 0,479** 0,519*** 1 0,517*** 0,773***
p 0,579 0,260 0,007 0,945 0,165 0,005 0,182 0,000 0,001 0,000 0,000 0,000
r 0,652*** 0,569*** 0,334* 0,559*** 0,693*** 0,339* 0,418** 0,690*** 0,490** 0,920*** 0,517*** 1 0,514**
p 0,000 0,000 0,033 0,000 0,000 0,030 0,006 0,000 0,001 0,000 0,000 0,001
r 0,017 0,151 0,318* -0,079 0,089 0,344* -0,268 0,500** 0,321* 0,356* 0,773*** 0,514** 1
p 0,917 0,354 0,046 0,622 0,582 0,030 0,090 0,001 0,041 0,022 0,000 0,001
SML
RAL
GL
GB
GH
OGH
SMR
RAR
MAB
SL
SBL
GSL
MGB
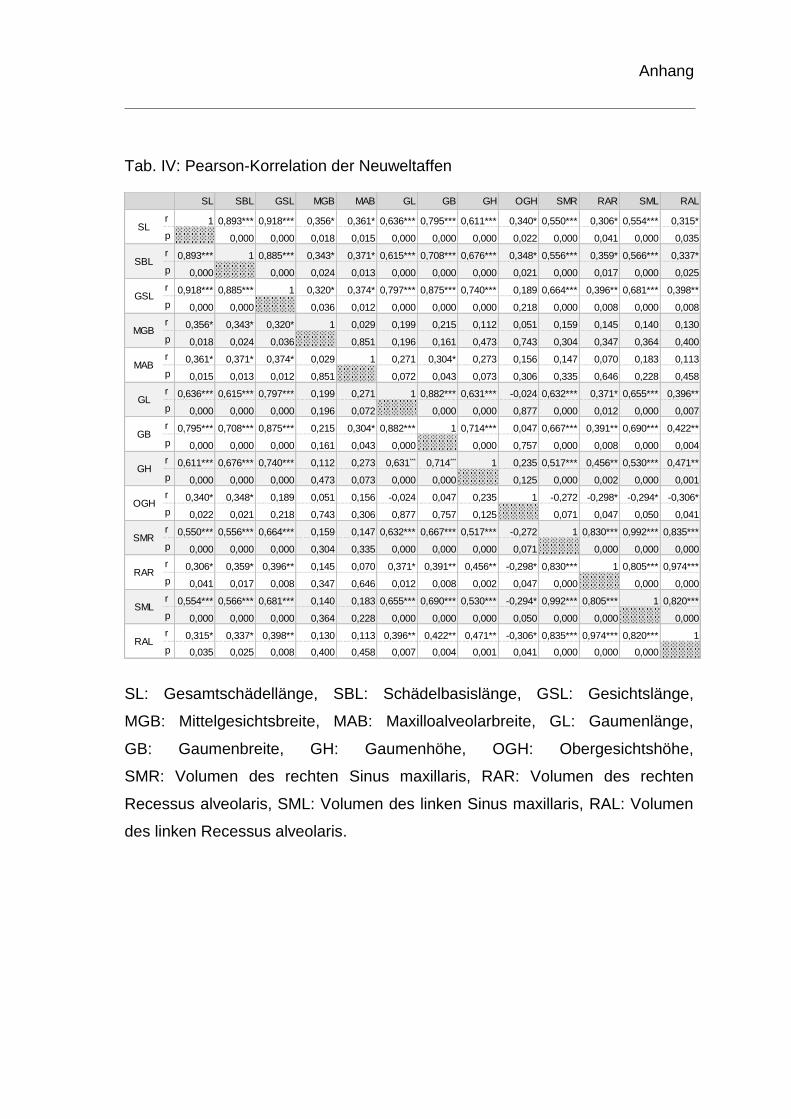
Anhang
Tab. IV: Pearson-Korrelation der Neuweltaffen
SL: Gesamtschädellänge, SBL: Schädelbasislänge, GSL: Gesichtslänge,
MGB: Mittelgesichtsbreite, MAB: Maxilloalveolarbreite, GL: Gaumenlänge,
GB: Gaumenbreite, GH: Gaumenhöhe, OGH: Obergesichtshöhe,
SMR: Volumen des rechten Sinus maxillaris, RAR: Volumen des rechten
Recessus alveolaris, SML: Volumen des linken Sinus maxillaris, RAL: Volumen
des linken Recessus alveolaris.
SL SBL GSL MGB MAB GL GB GH OGH SMR RAR SML RAL
r 1 0,893*** 0,918*** 0,356* 0,361* 0,636*** 0,795*** 0,611*** 0,340* 0,550*** 0,306* 0,554*** 0,315*
p 0,000 0,000 0,018 0,015 0,000 0,000 0,000 0,022 0,000 0,041 0,000 0,035
r 0,893*** 1 0,885*** 0,343* 0,371* 0,615*** 0,708*** 0,676*** 0,348* 0,556*** 0,359* 0,566*** 0,337*
p 0,000 0,000 0,024 0,013 0,000 0,000 0,000 0,021 0,000 0,017 0,000 0,025
r 0,918*** 0,885*** 1 0,320* 0,374* 0,797*** 0,875*** 0,740*** 0,189 0,664*** 0,396** 0,681*** 0,398**
p 0,000 0,000 0,036 0,012 0,000 0,000 0,000 0,218 0,000 0,008 0,000 0,008
r 0,356* 0,343* 0,320* 1 0,029 0,199 0,215 0,112 0,051 0,159 0,145 0,140 0,130
p 0,018 0,024 0,036 0,851 0,196 0,161 0,473 0,743 0,304 0,347 0,364 0,400
r 0,361* 0,371* 0,374* 0,029 1 0,271 0,304* 0,273 0,156 0,147 0,070 0,183 0,113
p 0,015 0,013 0,012 0,851 0,072 0,043 0,073 0,306 0,335 0,646 0,228 0,458
r 0,636*** 0,615*** 0,797*** 0,199 0,271 1 0,882*** 0,631*** -0,024 0,632*** 0,371* 0,655*** 0,396**
p 0,000 0,000 0,000 0,196 0,072 0,000 0,000 0,877 0,000 0,012 0,000 0,007
r 0,795*** 0,708*** 0,875*** 0,215 0,304* 0,882*** 1 0,714*** 0,047 0,667*** 0,391** 0,690*** 0,422**
p 0,000 0,000 0,000 0,161 0,043 0,000 0,000 0,757 0,000 0,008 0,000 0,004
r 0,611*** 0,676*** 0,740*** 0,112 0,273 0,631*** 0,714*** 1 0,235 0,517*** 0,456** 0,530*** 0,471**
p 0,000 0,000 0,000 0,473 0,073 0,000 0,000 0,125 0,000 0,002 0,000 0,001
r 0,340* 0,348* 0,189 0,051 0,156 -0,024 0,047 0,235 1 -0,272 -0,298* -0,294* -0,306*
p 0,022 0,021 0,218 0,743 0,306 0,877 0,757 0,125 0,071 0,047 0,050 0,041
r 0,550*** 0,556*** 0,664*** 0,159 0,147 0,632*** 0,667*** 0,517*** -0,272 1 0,830*** 0,992*** 0,835***
p 0,000 0,000 0,000 0,304 0,335 0,000 0,000 0,000 0,071 0,000 0,000 0,000
r 0,306* 0,359* 0,396** 0,145 0,070 0,371* 0,391** 0,456** -0,298* 0,830*** 1 0,805*** 0,974***
p 0,041 0,017 0,008 0,347 0,646 0,012 0,008 0,002 0,047 0,000 0,000 0,000
r 0,554*** 0,566*** 0,681*** 0,140 0,183 0,655*** 0,690*** 0,530*** -0,294* 0,992*** 0,805*** 1 0,820***
p 0,000 0,000 0,000 0,364 0,228 0,000 0,000 0,000 0,050 0,000 0,000 0,000
r 0,315* 0,337* 0,398** 0,130 0,113 0,396** 0,422** 0,471** -0,306* 0,835*** 0,974*** 0,820*** 1
p 0,035 0,025 0,008 0,400 0,458 0,007 0,004 0,001 0,041 0,000 0,000 0,000
RAR
SML
RAL
MAB
GL
GB
GH
OGH
SMR
SL
SBL
GSL
MGB

Anhang
Danksagung
An erster Stelle gilt mein besonderer Dank Herrn Prof. Dr. Thomas Koppe für die
freundliche Überlassung des Themas dieser Dissertation und die umfassende
und geduldige Betreuung dabei. Sein wertvoller Rat, die konstruktiven
Gespräche sowie die motivierenden Ratschläge waren unersetzlich bis zur
Fertigstellung der Arbeit. Des Weiteren möchte ich mich bei ihm für die
Bereitstellung der CT-Aufnahmen und eines Großteils der Schädeldaten
bedanken.
Weiterhin möchte ich mich bei Prof. Dr. Karlhans Endlich, Direktor des Instituts
für Anatomie und Zellbiologie der Universitätsmedizin Greifswald bedanken, der
mir die Arbeit am Institut ermöglicht hat.
Mein Dank gilt weiterhin Herrn Dr. Christian Schwahn, der mir bei Fragen in
statistischer und mathematischer Hinsicht immer zur Seite stand.
Danke auch an meine Freundin Dr. Julia Skrobek für ihre zahlreichen
Anregungen und Ratschläge.
Eine besondere Stellung nehmen meine Eltern, meine Geschwister Benjamin
und Michael und mein Freund Raphael Jacobs ein. Ihnen möchte ich herzlichst
für die liebevolle Fürsorge mit immer wieder aufbauenden und motivierenden
Worten bis zur Vollendung dieser Arbeit danken.