
ne Organisation der DNA in Nukleosomenreduziert den räumlichen Anspruch desGenoms bereits um das Siebenfache [1]. Inder nächsthöheren Organisationsform wirddie sogenannte 30-nm-Faser gebildet, derenexakte Struktur bis heute unbekannt ist.Maximale Kondensation wird in mitotischenChromosomen erreicht, in denen die Ge nomeum das 10.000-fache kondensiert werden [1].Die Histone selbst lassen sich in globuläreDomänen, die das eigentliche Proteingerüstfür die DNA bilden, und unstrukturierte N-ter-minale Histon-tails unterteilen. Zumindestauf der Ebene einzelner Nukleosomen schei-nen die ca. 15 bis 37 Aminosäuren langenHiston-tails keine strukturelle Relevanz zuhaben, ihre Aminosäuresequenz ist jedochvon der Hefe bis zum Menschen hochkon-serviert, was auf eine zentrale biologischeFunktion hindeutet [2, 3]. Diese ist eng mitder zweiten essenziellen Funktion des Chro-matins verknüpft, der Regulation der Genak-tivität an der benachbarten DNA. Diese Wech-selwirkungen werden u. a. durch eine Viel-zahl von posttranslationalen Modifikationenvermittelt, die man primär an den Histon-tailsfindet. Eine große strukturelle Diversitätzeichnet diese Modifikationen aus. Bereitsseit den 1960er-Jahren kennt man Acetylie-rungen und Methylierungen von Lysinrestensowie die Phosphorylierung von Serin- undThreoninresten (Abb. 1B, [2, 3]). NeuereUntersuchungen erlaubten die Identifikationvon Methylierung und Desaminierung vonArginin, Ubiquitinierung von Lysin sowie wei-teren Modifikationen.
¯ Abb. 1: Das Nukleosom ist die strukturelleBasiseinheit des Chromatins. A, Struktur desNukleosoms mit den Histonen H3 (grün), H4(blau), H2A (rot), H2B (gelb) und der DNA (grau).Die Abbildung wurde basierend auf der PDB-Struktur 1AOI erstellt. B, Übersicht der bekann-ten Modifikationsstellen an den Histon-tails.Gelistet sind Acetylierungen, Methylierungen,Phosphorylierung von Serin- und Threoninres-ten und Arginin-Desaminierungen zu Citrullin.
248 WISSENSCHAFT
BIOspektrum | 03.12 | 18. Jahrgang
DIRK SCHWARZER
INTERFAKULTÄRES INSTITUT FÜR BIOCHEMIE, UNIVERSITÄT TÜBINGEN
Histones package DNA into chromatin and regulate genes through a multitude of histone modifications. A ‘histone code’ hypothesis has beenput forth that ascribes different patterns of histone modifications specificbiological functions. This article highlights recent chemical biologicalapproaches to uncover the complex cross-talk between histone marksand chromatin activity.
DOI: 10.1007/s12268-012-0169-9© Springer-Verlag 2012
Chromatin, Histone undGenregulationó Alle eukaryotischen Organismen müssenihre DNA effizient verpacken, um sie in dembegrenzten Volumen ihrer Zellkerne unter-zubringen. Die Verpackung der Genomeerfolgt in Form von Chromatin, einem Kom-
plex aus DNA und zugehörigen Verpa-ckungsproteinen, den Histonen [1]. Die ein-fachste Struktureinheit des Chromatins istdas Nukleosom, bestehend aus je einem Paarder Histone H2A, H2B, H3 und H4, die einProteingerüst bilden, um das ca. 150 Basen-paare DNA gewickelt sind (Abb. 1A). Die rei-
Histonmodifikation
Die Chemische Biologie beschreitet neue Wege in der Chromatinforschung
A
B
Die heute gängigste Theorie zum Zu sam -menspiel zwischen Histon-tails, posttransla-tionalen Modifikationen und Genaktivität istdie Histon-Code-Theorie [4]. Sie interpretiertdie Histon-tails als eine Art Programmier-plattform, die mit posttranslationalen Modi-fikationen beschrieben wird. Eine einzelneModifikation oder ein Modifikationsmusterdient hierbei als Erkennungsstelle für ein spe-zifisches Regulatorprotein, das diese Mar-kierung erkennt, an sie bindet und imAnschluss Prozessierungen der benachbar-ten DNA einleitet. Mithilfe der Histon-Code-Theorie lassen sich viele experimentelle Beob-achtungen erklären, und man kennt bereitseine Vielzahl von Proteindomänen, die His-tonmodifikationen binden. Allerdings stelltdie schier astronomische Zahl an möglichen„Histon-Codons“, die sich durch die freie Per-mutation der bekannten Histonmodifikatio-nen ergeben, ein Problem dar, und man mussdavon ausgehen, dass viele theoretisch mög-liche Modifikationsmuster keine physiologi-sche Bedeutung haben.
Der gezielte Einbau vonHistonmodifikationen durchProteinsemisyntheseUm zu klären, welche Modifikationen für dieRekrutierung eines Regulatorproteins not-wendig sind, bedarf es homogen modifizier-ter Histone, die in Nukleosomen inkorporiertwerden können. Dies stellt konventionellebiochemische Ansätze vor ein Problem, dadie Enzyme, die diese Modifikationen in vivoin Histone einfügen, entweder nicht rekom-binant hergestellt werden können oder in vitroweitestgehend unspezifisch sind [3]. Die Che-mische Biologie bietet hier durch die soge-
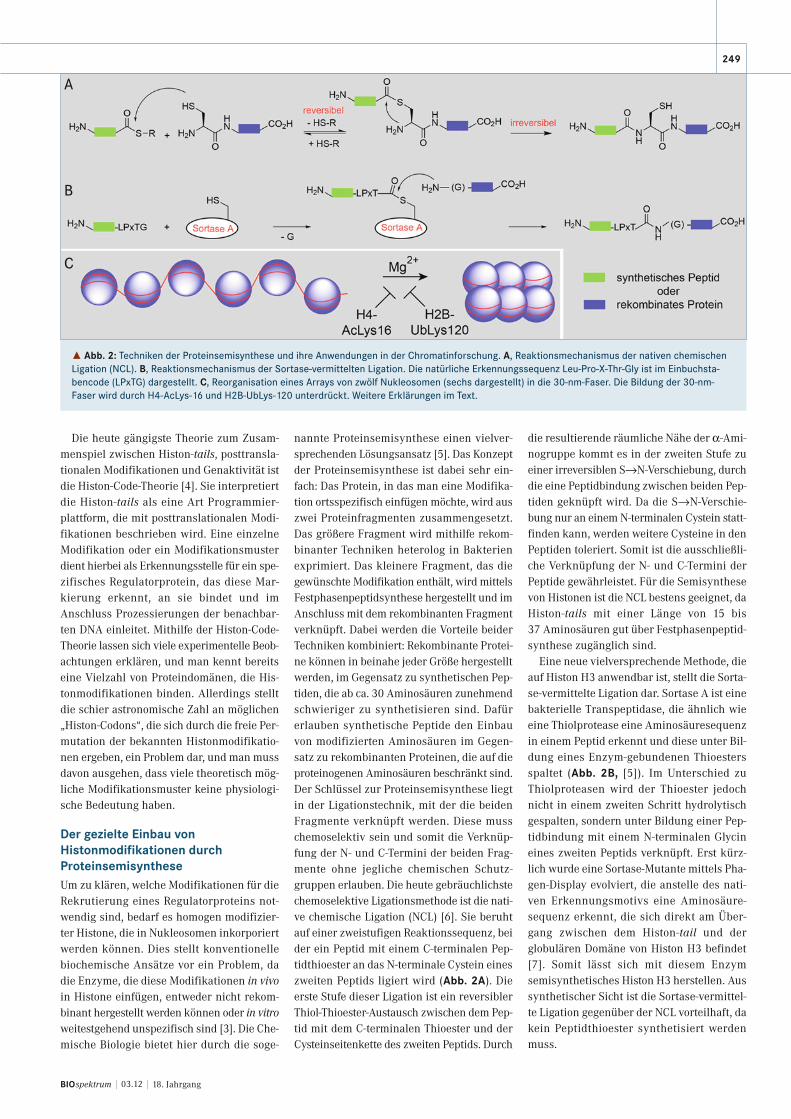
nannte Proteinsemisynthese einen vielver-sprechenden Lösungsansatz [5]. Das Konzeptder Proteinsemisynthese ist dabei sehr ein-fach: Das Protein, in das man eine Modifika-tion ortsspezifisch einfügen möchte, wird auszwei Proteinfragmenten zusammengesetzt.Das größere Fragment wird mithilfe rekom-binanter Techniken heterolog in Bakterienexprimiert. Das kleinere Fragment, das diegewünschte Modifikation enthält, wird mittelsFestphasenpeptidsynthese hergestellt und imAnschluss mit dem rekombinanten Fragmentverknüpft. Dabei werden die Vorteile beiderTechniken kombiniert: Rekombinante Protei-ne können in beinahe jeder Größe hergestelltwerden, im Gegensatz zu synthetischen Pep-tiden, die ab ca. 30 Aminosäuren zunehmendschwieriger zu synthetisieren sind. Dafürerlauben synthetische Peptide den Einbauvon modifizierten Aminosäuren im Gegen-satz zu rekombinanten Proteinen, die auf dieproteinogenen Aminosäuren beschränkt sind.Der Schlüssel zur Proteinsemisynthese liegtin der Ligationstechnik, mit der die beidenFragmente verknüpft werden. Diese musschemoselektiv sein und somit die Verknüp-fung der N- und C-Termini der beiden Frag-mente ohne jegliche chemischen Schutz-gruppen erlauben. Die heute gebräuchlichstechemoselektive Ligationsmethode ist die nati-ve chemische Ligation (NCL) [6]. Sie beruhtauf einer zweistufigen Reaktionssequenz, beider ein Peptid mit einem C-terminalen Pep-tidthioester an das N-terminale Cystein eineszweiten Peptids ligiert wird (Abb. 2A). Dieerste Stufe dieser Ligation ist ein reversiblerThiol-Thioester-Austausch zwischen dem Pep-tid mit dem C-terminalen Thioester und derCysteinseitenkette des zweiten Peptids. Durch
die resultierende räumliche Nähe der α-Ami-nogruppe kommt es in der zweiten Stufe zueiner irreversiblen S→N-Verschiebung, durchdie eine Peptidbindung zwischen beiden Pep-tiden geknüpft wird. Da die S→N-Verschie-bung nur an einem N-terminalen Cystein statt-finden kann, werden weitere Cysteine in denPeptiden toleriert. Somit ist die ausschließli-che Verknüpfung der N- und C-Termini derPeptide gewährleistet. Für die Semisynthesevon Histonen ist die NCL bestens geeignet, daHiston-tails mit einer Länge von 15 bis37 Aminosäuren gut über Festphasenpeptid-synthese zugänglich sind.
Eine neue vielversprechende Methode, dieauf Histon H3 anwendbar ist, stellt die Sorta-se-vermittelte Ligation dar. Sortase A ist einebakterielle Transpeptidase, die ähnlich wieeine Thiolprotease eine Aminosäuresequenzin einem Peptid erkennt und diese unter Bil-dung eines Enzym-gebundenen Thioestersspaltet (Abb. 2B, [5]). Im Unterschied zuThiolproteasen wird der Thioester jedochnicht in einem zweiten Schritt hydrolytischgespalten, sondern unter Bildung einer Pep-tidbindung mit einem N-terminalen Glycineines zweiten Peptids verknüpft. Erst kürz-lich wurde eine Sortase-Mutante mittels Pha-gen-Display evolviert, die anstelle des nati-ven Erkennungsmotivs eine Aminosäure -sequenz erkennt, die sich direkt am Über-gang zwischen dem Histon-tail und der globulären Domäne von Histon H3 befindet[7]. Somit lässt sich mit diesem Enzym semisynthetisches Histon H3 herstellen. Aussynthetischer Sicht ist die Sortase-vermittel-te Ligation gegenüber der NCL vorteilhaft, dakein Peptidthioester synthetisiert werdenmuss.
˚ Abb. 2: Techniken der Proteinsemisynthese und ihre Anwendungen in der Chromatinforschung. A, Reaktionsmechanismus der nativen chemischenLigation (NCL). B, Reaktionsmechanismus der Sortase-vermittelten Ligation. Die natürliche Erkennungssequenz Leu-Pro-X-Thr-Gly ist im Einbuchsta-bencode (LPxTG) dargestellt. C, Reorganisation eines Arrays von zwölf Nukleosomen (sechs dargestellt) in die 30-nm-Faser. Die Bildung der 30-nm-Faser wird durch H4-AcLys-16 und H2B-UbLys-120 unterdrückt. Weitere Erklärungen im Text.
A
B
C
249
BIOspektrum | 03.12 | 18. Jahrgang

250 WISSENSCHAFT
BIOspektrum | 03.12 | 18. Jahrgang
Die Chemische Biologie in derChromatinforschungZum gegenwärtigen Zeitpunkt sind chemisch-biologische Untersuchungsansätze in derChromatinforschung schon über das proof-of-concept-Stadium hinaus entwickelt wordenund konnten bereits wichtige Beiträge zurKlärung biologischer Beobachtungen leisten.Dies soll im Folgenden anhand ausgewählterBeispiele erläutert werden:
Eine zentrale Aussage der Histon-Code-The-orie ist die Existenz von Regulatorproteinen,die über bestimmte Modifikationen oderModifikationsmuster der Histon-tails an dasChromatin rekrutiert werden [4]. Die Anwen-dung der Proteinsemisynthese zur Erstellunghomogen modifizierter Histone, die in Nukle-osomen eingebaut als Köder für diese Regu-latoren dienen, ist äußerst vielversprechend.In zwei unabhängigen Studien wurden kürz-lich methylierungsspezifische Bindeproteinevon Histon H3 untersucht [8, 9]. Dazu wur-den zunächst synthetische H3-tails mit methy-lierten Lysinen an bekannten Modifikations-stellen als Peptidthioester hergestellt. NCL-vermittelte Ligation mit der rekombinantenglobulären Domäne lieferte semisynthetischeH3-Histone mit den gewünschten Methylie-rungen. Im Anschluss wurden zusammen mitunmodifiziertem H2A, H2B und H4 Nukleo-somen rekonstituiert, die über DNA-gebun-dene Biotin-Tags an Streptavidin-Agaroseimmobilisiert als Köder für Regulatorprotei-ne dienten. Mit diesen Ködern gelang es, zahl-reiche Chromatinfaktoren aus Zellkernex-trakten zu isolieren und über moderne Mas-senspektrometrie zu identifizieren. DieseUntersuchungen repräsentieren einen wich-tigen Schritt zur Aufklärung der Interak-tionsprofile nuklearer Proteine mit Chroma-tin, was einen wichtigen Beitrag zur experi-mentellen Überprüfung der Histon-Code-The-orie darstellt.
Interessanterweise können Histonmodifi-kationen nicht nur Regulatorproteine rekru-tieren, sondern auch die biophysikalischenEigenschaften des Chromatins direkt modu-lieren. Wie eingangs erwähnt falten sich län-gere Anordnungen von Nukleosomen in dienächsthöhere Organisationsstufe, die 30-nm-Faser. Es ist bekannt, dass sich Arrays, beste-hend aus zwölf Nukleosomen, in vitro inGegenwart von Mg2+ von einer linearenAnordnung in eine kompakte Struktur umor-ganisieren, die die biophysikalischen Eigen-schaften der 30-nm-Faser widerspiegeln(Abb. 2C, [10]). Für diese Reorganisation isteine Region des H4-tails, welche die Reste 14
bis 23 umfasst, essenziell. Da diese Regioneine Acetylierung an Lysin-16 aufweist, stell-te sich die Frage, ob diese Modifikation dieBildung der 30-nm-Faser beeinflussen kann.Zur Klärung dieser Vermutung wurde mittelsNCL an Lys-16 acetyliertes Histon H4 herge-stellt und anschließend in ein nukleosoma-les Array inkorporiert. Tatsächlich unter-drückte die Acetylierung von H4-Lys-16 dieBildung der 30-nm-Faser, wodurch gezeigtwurde, dass eine einzige Histonmodifikationdie Chromatinstruktur modulieren kann [10].
Unlängst konnte in einem weiteren che-misch-biologischen Ansatz gezeigt werden,dass auch eine andere Modifikation, die Ubi-quitinierung von Lys-120 von Histon H2B, dieBildung der 30-nm-Faser unterdrücken kann[11]. Dazu wurde das Ubiquitin-Protein che-misch über eine Disulfidbrücke mit H2B ver-knüpft. Die daraus rekonstituierten Nukleo-somen zeigten ebenfalls eine starke Beein-trächtigung in der Bildung der 30-nm-Faser.Interessanterweise stellte sich heraus, dassder Mechanismus, mit dem die Ubiquitinie-rung von H2B die Bildung der 30-nm-Faserbeeinflusst, sich deutlich von demjenigenunterscheidet, der durch die Acetylierung vonH4-Lys-16 vermittelt wird.
Diese ausgewählten Beispiele veranschau-lichen, wie elegant sich Fragestellungen zuder Funktion des Chromatins über moderneTechniken der Chemischen Biologie ent-schlüsseln lassen. Die Chemische Biologiehat sich somit als ein wichtiges Werkzeug zurUntersuchung von Chromatin etabliert. Nebender hier beschriebenen Proteinsemisynthesehält sie weitere Techniken bereit, um Modifi-kationen in Histone zu inkorporieren [2, 3].Das methodische Repertoire umfasst weiter-hin synthetische Moleküle und Naturstoffe,die die Aktivität Histon-modifizierender Enzy-me modulieren. Ein prominentes Beispiel, dasals ein Meilenstein der Chemischen Biologieverstanden werden kann, stellt die Isolierungund Identifizierung der ersten Histondeace-tylase mithilfe eines immobilisierten Histon-deacetylase-Inhibitors dar [12]. Vor diesemHintergrund erscheint es sehr wahrschein-
lich, dass auch zukünftig wichtige Impulsefür die Chromatinforschung und die Biologieim Allgemeinen von der Chemischen Biolo-gie kommen werden.
DanksagungUnsere Projekte werden von der DFG unter-stützt. Alexander Dose und Jan Oliver Jostdanke ich für die kritische Durchsicht desManuskripts. ó
Literatur[1] Turner BM (2001) Chromatin and Gene Regulation.Blackwell Science Ltd., Oxford[2] Allis CD, Muir TW (2011) Spreading chromatin into che-mical biology. Chembiochem 12:264–279[3] Schwarzer D (2010) Chemical tools in chromatin research.J Pept Sci 16:530–537[4] Strahl BD, Allis CD (2000) The language of covalent histo-ne modifications. Nature 403:41–45[5] Hackenberger CP, Schwarzer D (2008) Chemoselectiveligation and modification strategies for peptides and proteins.Angew Chem Int Ed Engl 47:10030–10074[6] Dawson PE, Muir TW, Clark-Lewis I et al. (1994) Synthesisof proteins by native chemical ligation. Science 266:776–779[7] Piotukh K, Geltinger B, Heinrich N et al. (2011) Directedevolution of sortase A mutants with altered substrate selecti-vity profiles. J Am Chem Soc 133:17536–17539[8] Nikolov M, Stützer A, Mosch K et al. (2011) Chromatinaffinity purification and quantitative mass spectrometry defi-ning the interactome of histone modification patterns. MolCell Proteomics 10:M110.005371[9] Bartke T, Vermeulen M, Xhemalce B et al. (2011)Nucleosome-interacting proteins regulated by DNA and histo-ne methylation. Cell 143:470–484[10] Shogren-Knaak M, Ishii H, Sun JM et al. (2006) HistoneH4-K16 acetylation controls chromatin structure and proteininteractions. Science 311:844–847[11] Fierz B, Chatterjee C, McGinty RK et al. (2011) HistoneH2B ubiquitylation disrupts local and higher-order chromatincompaction. Nat Chem Biol 7:113–119[12] Taunton J, Hassig CA, Schreiber SL (1996) A mammalianhistone deacetylase related to the yeast transcriptional regula-tor Rpd3p. Science 272:408–411
Korrespondenzadresse:Prof. Dr. Dirk SchwarzerInterfakultäres Institut für BiochemieEberhard Karls Universität TübingenHoppe-Seyler-Straße 4D-72076 TübingenTel.: 07071-29-73344Fax: [email protected]
AUTORDirk SchwarzerJahrgang 1972. 1993–2002 Chemiestudium und Promotion an der Universität Mar-burg. 2003–2006 Postdoc an der Johns Hopkins University, Baltimore, MD, USA.2006–2007 Postdoc an der Universität Dortmund. 2007–2011 Leiter einer Emmy-Noether-Gruppe am Leibniz-Institut für Molekulare Pharmakologie (FMP) in Berlin. Seit2011 Professor für Biochemie am Interfakultären Institut für Biochemie, UniversitätTübingen.
Recommended











![Chemische Verfahrenstechnik 2018 [Schreibgeschützt] · Universität Stuttgart Entwicklung der Chemische Verfahrenstechnik 5 Stoffumwandlung durch chemische Reaktion Bindeglied zwischen](https://img.pdfslide.org/doc/110x75/5d60386488c9930d758b7bc3/chemische-verfahrenstechnik-2018-schreibgeschuetzt-universitaet-stuttgart.jpg)




