Embed Size (px)
Citation preview
12.2 Mundhöhle, Pharynxund Ösophagus
In der Mundhöhle wird die aufgenommene feste Nah-rung durch Kauen und Einspeicheln in einen gleit-fähigen Zustand überführt. Der Speichel enthält vorallem Elektrolyte, ·-Amylase und Muzine.
12.2.1 Anatomie von Mundhöhle,Pharynx, Ösophagus undSpeicheldrüsen
Die Mundhöhle stellt den Anfangsteil des Verdau-ungskanals dar. Sie wird nach vorn von den Lippen mitder Mundspalte, seitlich durch die Wangen, untendurch den Mundboden und oben durch den harten undweichen Gaumen begrenzt. Nach hinten geht dieMundhöhle in den mittleren Abschnitt des Pharynx(Rachens) über. An diesem Übergang liegen die Gau-menbögen, welche die Gaumentonsillen (Gaumen-mandeln) einschließen. Der Raum zwischen den Zäh-nen mit den Alveolarfortsätzen der Kiefer und denWangen bzw. Lippen wird Mundvorhof genannt. DasInnere der Mundhöhle ist, mit Ausnahme der Zähne,von Schleimhaut mit zahlreichen kleinen Speichel-drüsen ausgekleidet.
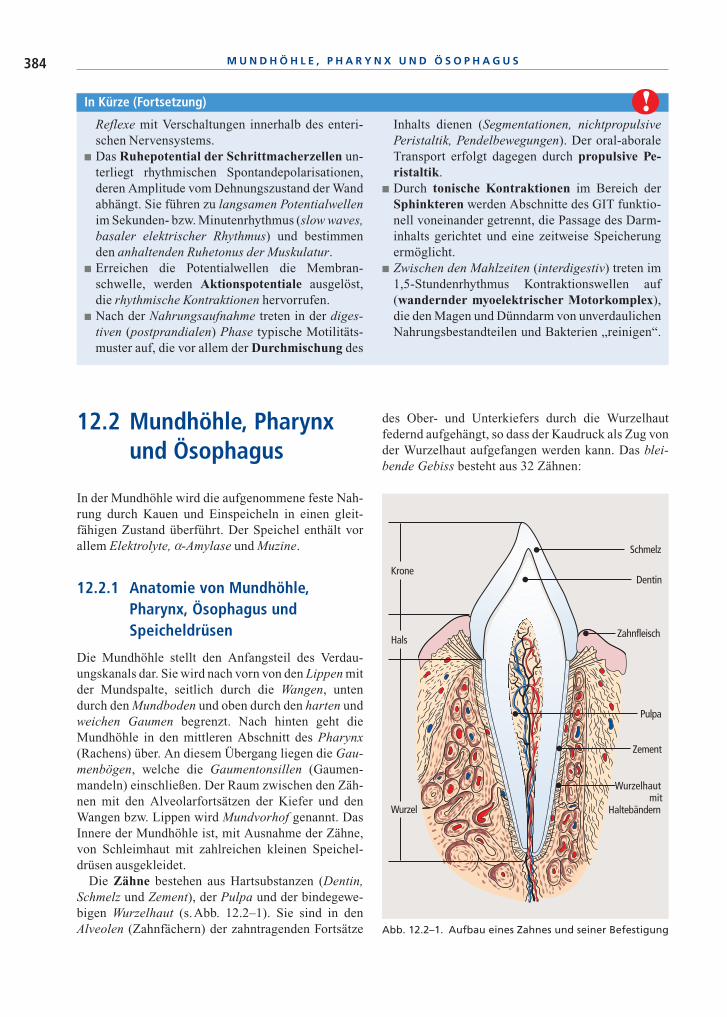
Die Zähne bestehen aus Hartsubstanzen (Dentin,Schmelz und Zement), der Pulpa und der bindegewe-bigen Wurzelhaut (s.Abb. 12.2–1). Sie sind in denAlveolen (Zahnfächern) der zahntragenden Fortsätze
des Ober- und Unterkiefers durch die Wurzelhautfedernd aufgehängt, so dass der Kaudruck als Zug vonder Wurzelhaut aufgefangen werden kann. Das blei-bende Gebiss besteht aus 32 Zähnen:
M U N D H Ö H L E , P H A R Y N X U N D Ö S O P H A G U S384
Zahnfleisch
Schmelz
Dentin
Wurzelhautmit
Haltebändern
Zement
Pulpa
Wurzel
Hals
Krone
Abb. 12.2–1. Aufbau eines Zahnes und seiner Befestigung
In Kürze (Fortsetzung)
Reflexe mit Verschaltungen innerhalb des enteri-schen Nervensystems.
� Das Ruhepotential der Schrittmacherzellen un-terliegt rhythmischen Spontandepolarisationen,deren Amplitude vom Dehnungszustand der Wandabhängt. Sie führen zu langsamen Potentialwellenim Sekunden- bzw. Minutenrhythmus (slow waves,basaler elektrischer Rhythmus) und bestimmenden anhaltenden Ruhetonus der Muskulatur.
� Erreichen die Potentialwellen die Membran-schwelle, werden Aktionspotentiale ausgelöst,die rhythmische Kontraktionen hervorrufen.
� Nach der Nahrungsaufnahme treten in der diges-tiven (postprandialen) Phase typische Motilitäts-muster auf, die vor allem der Durchmischung des
Inhalts dienen (Segmentationen, nichtpropulsivePeristaltik, Pendelbewegungen). Der oral-aboraleTransport erfolgt dagegen durch propulsive Pe-ristaltik.
� Durch tonische Kontraktionen im Bereich derSphinkteren werden Abschnitte des GIT funktio-nell voneinander getrennt, die Passage des Darm-inhalts gerichtet und eine zeitweise Speicherungermöglicht.
� Zwischen den Mahlzeiten (interdigestiv) treten im1,5-Stundenrhythmus Kontraktionswellen auf(wandernder myoelektrischer Motorkomplex),die den Magen und Dünndarm von unverdaulichenNahrungsbestandteilen und Bakterien „reinigen“.
!
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 384
� 8 meißelförmigen Schneidezähnen (Dentes incisi-vi),
� 4 Eckzähnen (Dentes canini) mit einer Kauspitze,
� 8 Backenzähnen (Dentes praemolares) mit zweiKauspitzen und
� 12 Mahlzähnen (Dentes molares) mit 4–5 Kau-spitzen, von denen der jeweils hinterste Mahlzahn(Weisheitszahn) oft Rückbildungserscheinungenzeigt.
Das Milchgebiss umfasst lediglich 20 Zähne (8Schneide-, 4 Eck- und 8 Mahlzähne).
Die Öffnungs- und Schließbewegungen, die Vor-und Rückschiebe- und seitlichen Bewegungen derZahnreihen beim Kauen werden durch vier paarigangelegte Kaumuskeln (M. masseter, M. temporalisund zwei weitere Muskelpaare, die Mm. pterygoidei,die an der Schädelbasis entspringen und zur Unter-seite des Unterkiefers ziehen) ermöglicht.
Die Zunge ist ein von Schleimhaut umkleideterMuskelkörper. Man unterscheidet den frei bewegli-chen Zungenrücken und den Zungengrund, der dashintere Drittel der Zunge einnimmt und den Übergangzum Pharynx bildet. Der Zungenrücken trägt Papillenfür die Tast- und Geschmacksempfindung (s.S. 721f.). Die quer gestreifte Zungenbinnenmuskulatur ver-läuft in longitudinalen, transversalen und vertikalenFaserzügen; in die Zunge einstrahlende Außenmus-keln haben ihren Ursprung am Unterkiefer, am Zun-genbein und am Schläfenbein. An der Oberfläche desZungengrundes liegt eine Ansammlung lymphati-schen Gewebes, die Tonsilla lingualis, die zusammenmit den Gaumentonsillen dem sog. lymphatischenRachenring zugerechnet wird.
Der Gaumen bildet das Dach der Mundhöhle. Manunterscheidet zwei Abschnitte, den harten Gaumen,der durch eine Knochenplatte versteift ist, und denweichen Gaumen mit der Uvula („Zäpfchen“).
Der Pharynx (Rachen) ist ein von Schleimhaut aus-gekleideter Muskelschlauch, dessen oberes Ende ander Schädelbasis aufgehängt ist, während das untereEnde in den Ösophagus übergeht. An seiner Vorder-wand finden sich drei Öffnungen, die zur paarigenNasenhöhle, zur Mundhöhle und zum Kehlkopfein-gang führen. Entsprechend wird der Pharynx in dreiAbschnitte unterteilt: Einen oberen (Epipharynx, Na-sen-Rachen-Raum), einen mittleren (Mesopharynx),der hinter der Mundhöhle liegt, und einen unterenAbschnitt (Hypopharynx) mit Zugang zum Kehlkopf.Im Pharynx kreuzen Luft- und Speisewege.
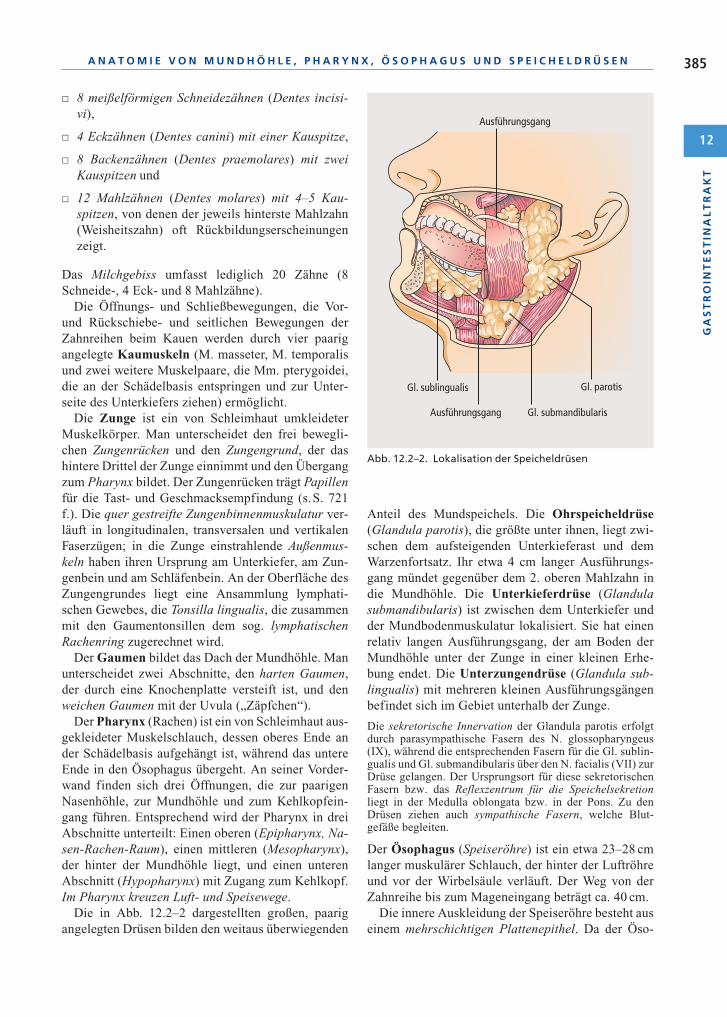
Die in Abb. 12.2–2 dargestellten großen, paarigangelegten Drüsen bilden den weitaus überwiegenden
Anteil des Mundspeichels. Die Ohrspeicheldrüse(Glandula parotis), die größte unter ihnen, liegt zwi-schen dem aufsteigenden Unterkieferast und demWarzenfortsatz. Ihr etwa 4 cm langer Ausführungs-gang mündet gegenüber dem 2. oberen Mahlzahn indie Mundhöhle. Die Unterkieferdrüse (Glandulasubmandibularis) ist zwischen dem Unterkiefer undder Mundbodenmuskulatur lokalisiert. Sie hat einenrelativ langen Ausführungsgang, der am Boden derMundhöhle unter der Zunge in einer kleinen Erhe-bung endet. Die Unterzungendrüse (Glandula sub-lingualis) mit mehreren kleinen Ausführungsgängenbefindet sich im Gebiet unterhalb der Zunge.
Die sekretorische Innervation der Glandula parotis erfolgtdurch parasympathische Fasern des N. glossopharyngeus(IX), während die entsprechenden Fasern für die Gl. sublin-gualis und Gl. submandibularis über den N. facialis (VII) zurDrüse gelangen. Der Ursprungsort für diese sekretorischenFasern bzw. das Reflexzentrum für die Speichelsekretionliegt in der Medulla oblongata bzw. in der Pons. Zu denDrüsen ziehen auch sympathische Fasern, welche Blut-gefäße begleiten.
Der Ösophagus (Speiseröhre) ist ein etwa 23–28 cmlanger muskulärer Schlauch, der hinter der Luftröhreund vor der Wirbelsäule verläuft. Der Weg von derZahnreihe bis zum Mageneingang beträgt ca. 40 cm.
Die innere Auskleidung der Speiseröhre besteht auseinem mehrschichtigen Plattenepithel. Da der Öso-
385A N A T O M I E V O N M U N D H Ö H L E , P H A R Y N X , Ö S O P H A G U S U N D S P E I C H E L D R Ü S E N
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12Ausführungsgang
Gl. parotis
Gl. submandibularisAusführungsgang
Gl. sublingualis
Abb. 12.2–2. Lokalisation der Speicheldrüsen
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 385

phagus ausschließlich der Weiterbeförderung desSpeisebreis dient, findet man in der Schleimhaut le-diglich Drüsen, die zum Schutz des Epithels Schleimsezernieren. Die Wand enthält im oberen Drittel quergestreifte, im unteren Drittel glatte Muskulatur. Dasmittlere Drittel weist beide Muskelarten auf.
Der Ösophagus hat drei anatomische „Engstellen“:Die erste liegt in Höhe des Ringknorpels (Kehlkopf-bereich), die mittlere in Höhe der Luftröhrengabe-lung, die untere in Höhe des Zwerchfelldurchtritts.
12.2.2 Kauen
Beim Kauen wird die feste Nahrung zerschnitten, zer-rissen und zermahlen. Obwohl diese Zerkleinerungkeine zwingende Voraussetzung für die Verdauungund Absorption ist, erleichtert sie diese Vorgänge er-heblich (z.B. durch Verbesserung des enzymatischenAufschlusses infolge Oberflächenvergrößerung). AmKauvorgang beteiligt sind Ober- und Unterkiefer mitden Zähnen, Kaumuskulatur, Zunge und Wangen so-wie Mundboden und Gaumen.
Die rhythmische Aktion des Kauvorgangs erfolgtprimär willkürlich, dann auch weitgehend unbewusst.Der Berührungsreiz der Speisepartikel an Gaumenund Zähnen steuert reflektorisch die Kaubewegung.Die Kräfte, die dabei aufgewandt werden, betragen imBereich der Schneidezähne 100–250 N, im Bereichder Molaren 300–700 N.
Zunge und Wangen schieben den Bissen immerwieder zwischen die Kauflächen, so dass feste Nah-rung zu Partikeln bis zu einer Größe von wenigenmm3 zermahlen wird. Der durch den Kauvorgang sti-mulierte Speichelfluss bereitet die Konsistenz desBissens (Bolus) zum Schlucken vor. Beim Kauen wirddurch Freisetzung flüchtiger Komponenten aus derNahrung sowie durch Auflösung oder Aufschwem-mung fester Bestandteile im Speichel die Ge-schmackswahrnehmung gefördert. Dies führt reflek-torisch zur weiteren Anregung des Speichelflussesund der Magensekretion (s.S. 395 ff.).
Saugreflex. Dieser nutritive Reflex wird durch Berüh-rungsreize von den Lippen oder von der Mundschleimhautdes Säuglings her ausgelöst. Bei luftdichtem Abschluss zwi-schen Lippen und Warzenhof der mütterlichen Brust sowienach Abdichtung der nasalen und trachealen Luftwegeerfolgt zunächst eine Senkung des Mundbodens. Der da-durch im Mundraum entstehende Unterdruck saugt dieMuttermilch an. Anschließend werden die Kiefer zusam-mengedrückt und damit die Milchgänge der Brustdrüseausgepresst. Der gesamte komplexe Vorgang, der mit einerrhythmischen Freigabe der Nasenatmung koordiniert ist,steht unter der Kontrolle von Neuronen in der Medullaoblongata.
12.2.3 Speichelsekretion
Die Glandula parotis ist eine seröse Drüse, die nebenWasser und Elektrolyten Glykoproteine sezerniert.Die Glandula submandibularis und die Glandula sub-lingualis sind gemischte Drüsen, die zusätzlichSaccharid-reiche Glykoproteine (Muzine) produzie-ren.
Regulation der Speichelsekretion. Täglich werden0,6–1,5 l Mundspeichel gebildet. Dieser hält denMund feucht und erleichtert das Sprechen, macht diegekaute Nahrung gleitfähig und fördert die Ge-schmacksentwicklung. Er ist essentiell für die Ge-sundheit der Zähne, die ohne Speichel kariös werden.Der Speichel hat eine reinigende und durch seinenGehalt an Lysozym, sekretorischem IgA, Lactoferrinund verschiedenen Prolin-reichen antibakteriellenProteinen eine antibakterielle bzw. antivirale Wir-kung.
Auch ohne Nahrungsaufnahme findet immer einegeringe Basalsekretion (Ruhesekretion) von Mund-speichel (ca. 0,5 l/Tag) statt. Kommt es zu einerBerührung der Mundschleimhaut mit aufgenomme-nen Speisen und/oder zu Geschmacksempfindungen,so wird die Speichelsekretion reflektorisch gesteigert.Aber auch der Anblick, der Geruch oder die bloßeVorstellung von Speisen „lassen das Wasser im Mun-de zusammenlaufen“ („bedingte Reflexe“, kephaleSekretionsphase, s.S. 395). Dabei wird die Zusam-mensetzung des Speichels durch den Einfluss desvegetativen Nervensystems variiert.
Eine Aktivierung des Parasympathikus bewirkt inallen Drüsen eine erhebliche Steigerung der Sekretioneines dünnflüssigen, glykoproteinarmen Speichels,die mit einer Durchblutungszunahme der Drüsen ein-hergeht. Letztere wird durch die gefäßerweiterndeWirkung von VIP vermittelt.
Eine Erregung des Sympathikus liefert dagegendurch Stimulation der Unterkieferdrüse geringe Men-gen eines viskösen, Glykoprotein-, K+- und HCO3
–-reichen Speichels.
Während der Basalsekretion haben die einzelnenDrüsen an der Gesamtspeichelproduktion folgendeAnteile: Gl. submandibularis 70%, Gl. parotis 25%und Gl. sublingualis 5%; nach Stimulation: 63%,34% und 3%.
Zusammensetzung des Speichels. Der Speichel be-steht zu 99% aus Wasser. Die wichtigsten darin ent-haltenen Elektrolyte sind Na+, K+, Cl– und HCO3
–.Der Primärspeichel, der von den Azini sezerniertwird, ist plasmaisoton.
M U N D H Ö H L E , P H A R Y N X U N D Ö S O P H A G U S386
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 386

In den Azini der meisten Speicheldrüsen wird Cl–
durch einen Na+/2 Cl–/K+-Symporter der basolate-ralen Membran in die Zelle aufgenommen, wofür dieNa+/K+-ATPase den Antrieb liefert (s.Abb. 12.2–3).Die apikale Cl–-Sekretion erfolgt dann über einenCl–-Kanal, während Na+ und Wasser auf parazellulä-rem Weg folgen.
In den Ausführungsgängen werden, bei relativgeringer Wasserpermeabilität, Na+ (aldosteronabhän-gig) und Cl– aus dem Lumen resorbiert und kleinereMengen an K+ und HCO3
– sezerniert, wodurch derMundspeichel hypoton wird (s.Abb. 12.2–4).
Die Elektrolytzusammensetzung des Speichels än-dert sich mit der Sekretionsrate: Mit zunehmendemSekretionsvolumen steigen die Na+- und Cl–-Konzen-trationen an, während die K+- und HCO3
–-Konzen-
trationen leicht abfallen (s.Abb. 12.2–4), da die zurVerfügung stehende Zeit zur Resorption von Na+ bzw.Sekretion von K+ mit steigender Durchflussrate ver-kürzt bzw. die maximale Kapazität der Transport-systeme erreicht ist. Der pH-Wert des Mundspei-chels liegt bei Ruhesekretion zwischen 6,5 und 6,9und steigt nach Stimulation auf 7,2 an.
Zusätzlich zu Elektrolyten und Wasser sezernierendie Speicheldrüsen verschiedene Makromoleküle: ·-Amylase, Glykoproteine, Muzine, Lysozym, Lacto-ferrin, Immunglobulin A, Haptocorrine (s.S. 393)und häufig auch Blutgruppen-Antigene (s.S. 186 f.)sowie Wachstumsfaktoren, welche die wundheilendeWirkung des Speichels erklären. Die funktionellwichtigsten sind die ·-Amylase, die vorwiegend vonder Gl. parotis ausgeschieden wird, und die Schleim-substanzen (aus Gl. submandibularis und Gl. sublin-gualis). Die ·-Amylase (Ptyalin) ist zwischen pH-Werten von 4 bis 11 stabil und hat ihr Wirkungs-optimum bei pH 6,7–6,9. Dieses Enzym leitet dieVerdauung der Stärke ein.
12.2.4 Schluckakt
Der Schluckakt gliedert sich in eine willkürliche oralesowie eine reflektorisch ablaufende pharyngeale und
387S C H L U C K A K T
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
Azinuszelle
Lumen Gangepithel Interstitium
Na+
H2O2 CI– Na+
H2O
2 CI–
K+ Na+ 3 Na+
HCO3–
H+
K+
CI–
Na+
H+
2 K+
3 Na+
CI–
2 K+
A
B
Abb. 12.2–3. Modell der wichtigsten Elektrolyttransportein den Azinuszellen der Gl. submandibularis und Gl. parotis(Primärsekretbildung A) sowie in den Ausführungsgängen(B). In den Azinuszellen wird Cl– über einen apikal gelege-nen Chloridkanal sezerniert; Na+ und Wasser folgen passivauf parazellulärem Weg. In den Ausführungsgängen wer-den Na+ und Cl– aus dem Lumen resorbiert und kleinereMengen an K+ und HCO3
– sezerniert
0
100
200
300
0 1 2 3 4
Osmolalität (mosmol/kg H2O)
Speichelfluss (ml/min)
00 1 2 3 4
50
100
150
Na+
Cl–
K+
HCO3–
Elektrolytkonzentration (mmol/l)
Speichelfluss (ml/min)
Abb. 12.2–4. Osmolalität (oben) und Elektrolytzusammen-setzung (unten) des Mundspeichels als Funktion der Sekre-tionsrate
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 387
M U N D H Ö H L E , P H A R Y N X U N D Ö S O P H A G U S388
ösophageale Phase, in welcher der Bissen durch peri-staltische Wellen in den Magen befördert wird.
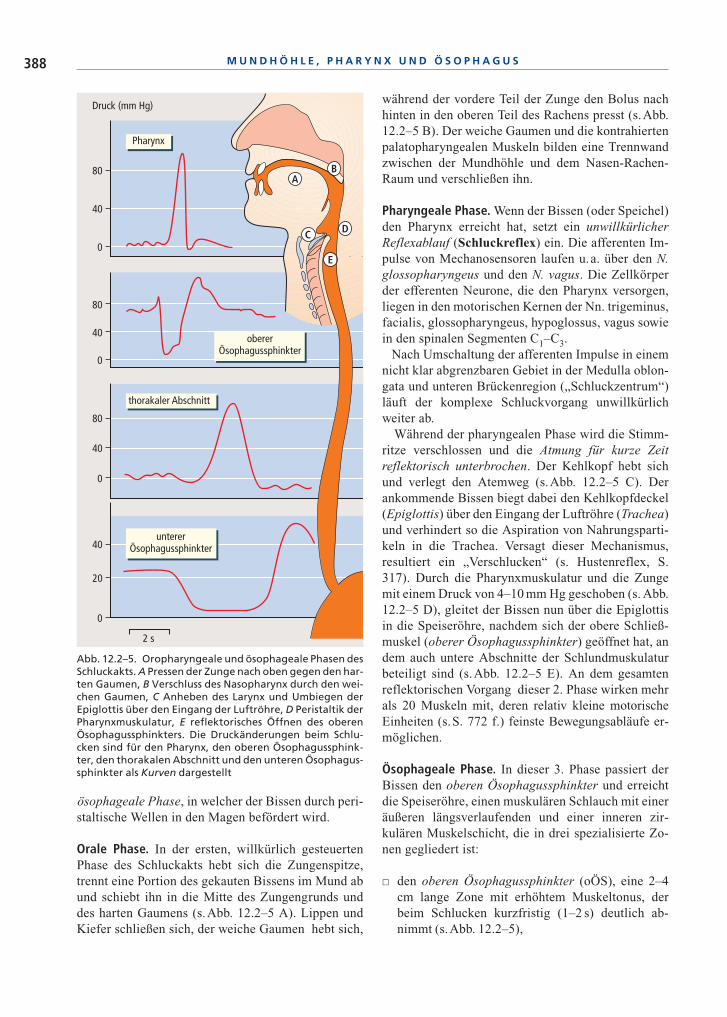
Orale Phase. In der ersten, willkürlich gesteuertenPhase des Schluckakts hebt sich die Zungenspitze,trennt eine Portion des gekauten Bissens im Mund abund schiebt ihn in die Mitte des Zungengrunds unddes harten Gaumens (s.Abb. 12.2–5 A). Lippen undKiefer schließen sich, der weiche Gaumen hebt sich,
während der vordere Teil der Zunge den Bolus nachhinten in den oberen Teil des Rachens presst (s.Abb.12.2–5 B). Der weiche Gaumen und die kontrahiertenpalatopharyngealen Muskeln bilden eine Trennwandzwischen der Mundhöhle und dem Nasen-Rachen-Raum und verschließen ihn.
Pharyngeale Phase. Wenn der Bissen (oder Speichel)den Pharynx erreicht hat, setzt ein unwillkürlicherReflexablauf (Schluckreflex) ein. Die afferenten Im-pulse von Mechanosensoren laufen u.a. über den N.glossopharyngeus und den N. vagus. Die Zellkörperder efferenten Neurone, die den Pharynx versorgen,liegen in den motorischen Kernen der Nn. trigeminus,facialis, glossopharyngeus, hypoglossus, vagus sowiein den spinalen Segmenten C1–C3.
Nach Umschaltung der afferenten Impulse in einemnicht klar abgrenzbaren Gebiet in der Medulla oblon-gata und unteren Brückenregion („Schluckzentrum“)läuft der komplexe Schluckvorgang unwillkürlichweiter ab.
Während der pharyngealen Phase wird die Stimm-ritze verschlossen und die Atmung für kurze Zeitreflektorisch unterbrochen. Der Kehlkopf hebt sichund verlegt den Atemweg (s.Abb. 12.2–5 C). Derankommende Bissen biegt dabei den Kehlkopfdeckel(Epiglottis) über den Eingang der Luftröhre (Trachea)und verhindert so die Aspiration von Nahrungsparti-keln in die Trachea. Versagt dieser Mechanismus,resultiert ein „Verschlucken“ (s. Hustenreflex, S.317). Durch die Pharynxmuskulatur und die Zungemit einem Druck von 4–10 mm Hg geschoben (s. Abb.12.2–5 D), gleitet der Bissen nun über die Epiglottisin die Speiseröhre, nachdem sich der obere Schließ-muskel (oberer Ösophagussphinkter) geöffnet hat, andem auch untere Abschnitte der Schlundmuskulaturbeteiligt sind (s.Abb. 12.2–5 E). An dem gesamtenreflektorischen Vorgang dieser 2. Phase wirken mehrals 20 Muskeln mit, deren relativ kleine motorischeEinheiten (s.S. 772 f.) feinste Bewegungsabläufe er-möglichen.
Ösophageale Phase. In dieser 3. Phase passiert derBissen den oberen Ösophagussphinkter und erreichtdie Speiseröhre, einen muskulären Schlauch mit eineräußeren längsverlaufenden und einer inneren zir-kulären Muskelschicht, die in drei spezialisierte Zo-nen gegliedert ist:
� den oberen Ösophagussphinkter (oÖS), eine 2–4cm lange Zone mit erhöhtem Muskeltonus, derbeim Schlucken kurzfristig (1–2 s) deutlich ab-nimmt (s.Abb. 12.2–5),
Abb. 12.2–5. Oropharyngeale und ösophageale Phasen desSchluckakts. A Pressen der Zunge nach oben gegen den har-ten Gaumen, B Verschluss des Nasopharynx durch den wei-chen Gaumen, C Anheben des Larynx und Umbiegen derEpiglottis über den Eingang der Luftröhre, D Peristaltik derPharynxmuskulatur, E reflektorisches Öffnen des oberenÖsophagussphinkters. Die Druckänderungen beim Schlu-cken sind für den Pharynx, den oberen Ösophagussphink-ter, den thorakalen Abschnitt und den unteren Ösophagus-sphinkter als Kurven dargestellt
0
40
80
0
40
80
0
40
80
0
20
40
BA
C D
E
obererÖsophagussphinkter
Druck (mm Hg)
2 s
thorakaler Abschnitt
untererÖsophagussphinkter
Pharynx
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 388

� den thorakalen Abschnitt und
� den unteren Ösophagussphinkter (uÖS), eine wei-tere Zone mit anhaltend erhöhtem Muskeltonus, derden Verschluss zum Magen gewährleistet.
Da der größte Teil des Ösophagus im Brustraum verläuft,liegen die Binnendrücke bei Ruheatmung um 4–6 mm Hgunter dem Atmosphärendruck (s.S. 306). Der Abschlussnach oben durch den oÖS mit einem Verschlussdruck von50–100 mm Hg verhindert ein ständiges Eindringen von Luftin den Ösophagus, während der uÖS mit einer Druckdiffe-renz von 15–25 mm Hg gegenüber dem Magenfundus denRückfluss (Reflux) von Mageninhalt in den Ösophagus ver-hindert.
Als primäre Peristaltik wird der durch den N. vagusnerval gesteuerte Bewegungsablauf bezeichnet, derdie Fortsetzung des begonnenen Schluckakts darstellt.Eine sekundäre Peristaltik entsteht durch afferenteImpulse vom Ösophagus selbst (z.B. durch mechani-sche Reizung). Sie wird durch Reste eines Bissens
verursacht, die durch die primäre Peristaltik nicht denMagen erreicht haben, und durch das enterische Ner-vensystem koordiniert.
Die peristaltische Welle im Ösophagus erfasst jeweils einKontraktionsareal von 2–4 cm Länge, schreitet mit einerGeschwindigkeit von 3–5 cm/s nach unten fort und erreichtden uÖS nach ca. 9 s (s.Abb. 12.2–5). Die Passagege-schwindigkeit hängt allerdings wesentlich von der Konsis-tenz des Bissens und der Körperlage ab. In aufrechter Kör-perhaltung erreichen Flüssigkeiten den Magen nach 1 s, brei-ige Nahrung nach 5 s und feste Partikel nach ca. 10 s. DerDruck der peristaltischen Welle steigt nach distal an underreicht im unteren Ösophagus 30–120 mm Hg. Die Druck-amplitude nimmt mit der Größe des Bissens zu. Der uÖS öff-net sich für 6–9 s, bevor der Bissen in den Magen eintritt undschließt sich danach wieder. Dabei nimmt er nach einer kur-zen Phase erhöhten Drucks erneut den Ruhetonus an. DieRelaxation des uÖS erfolgt reflektorisch unter dem Einflussvon NANC-Neuronen (s.S. 379) des N. vagus; als Neuro-transmitter werden das vasoaktive intestinale Polypeptid(VIP) und/oder Stickoxid (NO) angenommen.
389S C H L U C K A K T
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
!In Kürze !� Die Mundhöhle wird von den Lippen, den Wan-
gen, dem Gaumen und dem Mundboden begrenzt.Nach hinten geht sie in den Rachen über.
� Das Gebiss des Erwachsenen umfasst 32 Zähne,die in den Alveolen federnd aufgehängt sind. Manunterscheidet Zahnwurzel, -hals und -krone. Den-tin, Schmelz und Zement sind die Hartsubstanzender Zähne.
� Die Zahnreihen des Ober- und Unterkiefers wer-den durch die Kaumuskeln bewegt.
� Die Zunge ist ein von Schleimhaut umkleideterMuskelkörper. Die Schleimhaut des Zungen-rückens trägt Papillen für die Tast- und Ge-schmacksempfindung sowie Drüsen (Lipase-Se-kretion) und lymphatisches Gewebe („Zungen-mandel“). Der Muskelkörper ermöglicht die fürKauen, Bissenbildung, Schlucken und Sprechenerforderlichen Bewegungen.
� In die Mundhöhle münden zahlreiche kleine, in derMundschleimhaut gelegene sowie drei große, paa-rige Speicheldrüsen, die Glandulae parotis, sub-mandibularis und sublingualis.
� Die großen Speicheldrüsen besitzen sekretorischeEndstücke für die Bildung des Primärsekrets undein Ausführungsgangsystem, in dem durch Re-sorptions- und Sekretionsvorgänge der sog. Sekun-därspeichel entsteht.
� Die Ohrspeicheldrüse ist eine seröse Drüse, diebeiden anderen sind gemischte Drüsen, die Wasser,Elektrolyte und Glykoproteine (einschließlich Mu-zin) sezernieren.
� Durch Kauen und Einspeicheln wird in der Mund-höhle die aufgenommene feste Nahrung in einengleitfähigen Zustand überführt (Bolusbildung).
� Durch den Speichel werden Verdauungsenzyme(·-Amylase) und Abwehrstoffe bereitgestellt so-wie die Zähne vor Karies geschützt, die Ge-schmacksempfindung wird gefördert und dasSprechen erleichtert.
� Mundspeichel ist alkalisch. Geschluckter Speichelkann daher in die Speiseröhre gelangte Magen-salzsäure neutralisieren (sog. pH-Clearance).
� Die tägliche Speichelsekretion beträgt ca. 1 l. Siewird vorrangig durch Aktivierung des Parasym-pathikus reflektorisch gesteigert.
� Der Rachenraum (Pharynx) ist der gemeinsameAbschnitt des Atmungs- und Verdauungstrakts,die sich hier überkreuzen.
� Der Ösophagus (Speiseröhre), ein 23–28 cm lan-ger Muskelschlauch, ist lumenseitig mit einemmehrschichtigen, unverhornten Plattenepithel aus-gekleidet, das durch Muzin der Ösophagusdrüsengeschützt wird. Das obere Drittel der Speiseröhrebesitzt quer gestreifte, das untere Drittel glatte Mus-kulatur, das mittlere Drittel beide Muskeltypen.
� Der Ösophagus weist zwei Zonen mit erhöhtemMuskeltonus auf, den oberen Ösophagussphink-ter am Beginn und den unteren Ösophagus-sphinkter am Übergang zum Magen. Ihr hoherTonus verhindert das Eindringen von Luft bzw.Rückfluss von saurem Mageninhalt in den Öso-phagus.
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 389
12.3 Magen
Im Magen werden die geschluckten Speisen gespei-chert, zerkleinert und homogenisiert. Nach einerVerweildauer von 1–5 Stunden erfolgt die portions-weise Entleerung des Speisebreis (Chymus) in dasDuodenum.
12.3.1 Anatomie des Magens
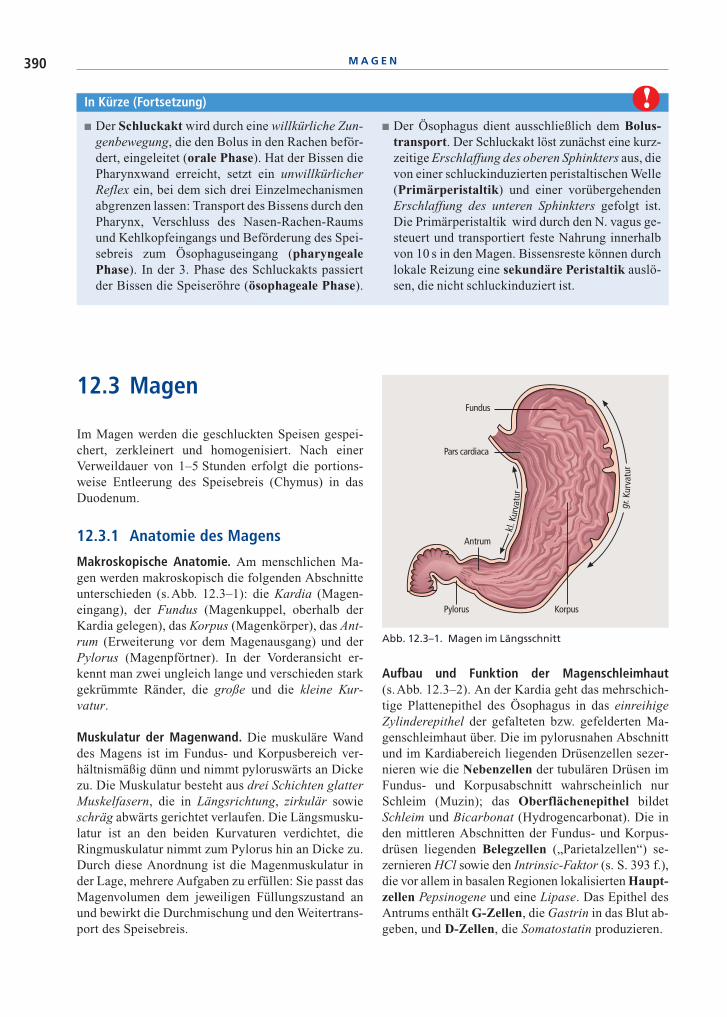
Makroskopische Anatomie. Am menschlichen Ma-gen werden makroskopisch die folgenden Abschnitteunterschieden (s.Abb. 12.3–1): die Kardia (Magen-eingang), der Fundus (Magenkuppel, oberhalb derKardia gelegen), das Korpus (Magenkörper), das Ant-rum (Erweiterung vor dem Magenausgang) und derPylorus (Magenpförtner). In der Vorderansicht er-kennt man zwei ungleich lange und verschieden starkgekrümmte Ränder, die große und die kleine Kur-vatur.
Muskulatur der Magenwand. Die muskuläre Wanddes Magens ist im Fundus- und Korpusbereich ver-hältnismäßig dünn und nimmt pyloruswärts an Dickezu. Die Muskulatur besteht aus drei Schichten glatterMuskelfasern, die in Längsrichtung, zirkulär sowieschräg abwärts gerichtet verlaufen. Die Längsmusku-latur ist an den beiden Kurvaturen verdichtet, dieRingmuskulatur nimmt zum Pylorus hin an Dicke zu.Durch diese Anordnung ist die Magenmuskulatur inder Lage, mehrere Aufgaben zu erfüllen: Sie passt dasMagenvolumen dem jeweiligen Füllungszustand anund bewirkt die Durchmischung und den Weitertrans-port des Speisebreis.
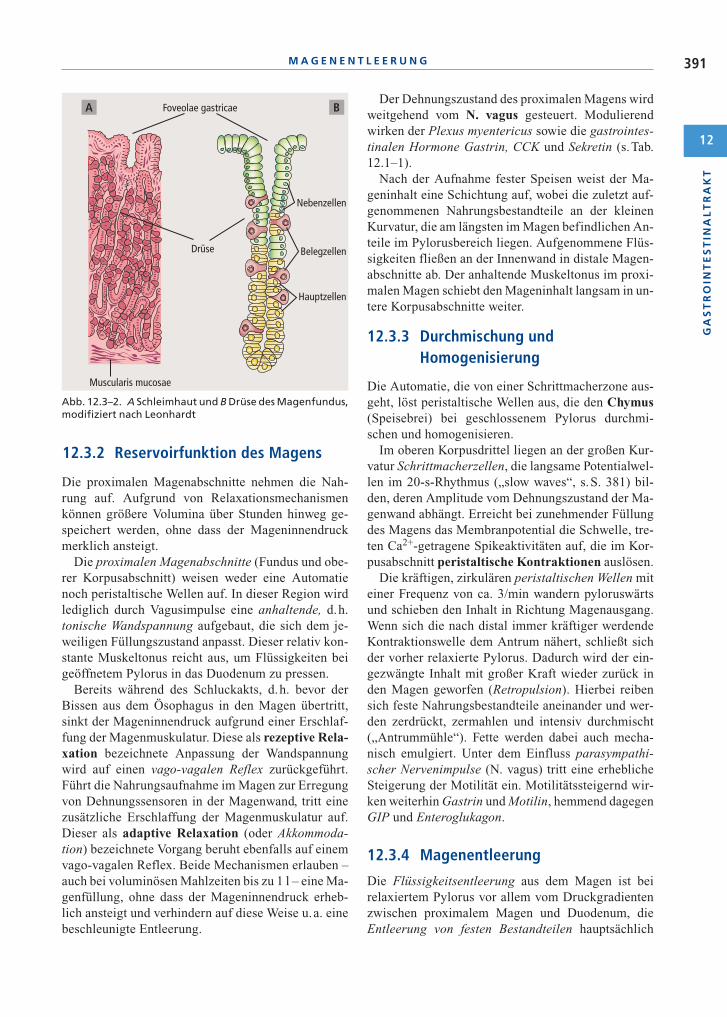
Aufbau und Funktion der Magenschleimhaut(s.Abb. 12.3–2). An der Kardia geht das mehrschich-tige Plattenepithel des Ösophagus in das einreihigeZylinderepithel der gefalteten bzw. gefelderten Ma-genschleimhaut über. Die im pylorusnahen Abschnittund im Kardiabereich liegenden Drüsenzellen sezer-nieren wie die Nebenzellen der tubulären Drüsen imFundus- und Korpusabschnitt wahrscheinlich nurSchleim (Muzin); das Oberflächenepithel bildetSchleim und Bicarbonat (Hydrogencarbonat). Die inden mittleren Abschnitten der Fundus- und Korpus-drüsen liegenden Belegzellen („Parietalzellen“) se-zernieren HCl sowie den Intrinsic-Faktor (s. S. 393 f.),die vor allem in basalen Regionen lokalisierten Haupt-zellen Pepsinogene und eine Lipase. Das Epithel desAntrums enthält G-Zellen, die Gastrin in das Blut ab-geben, und D-Zellen, die Somatostatin produzieren.
M A G E N390
In Kürze (Fortsetzung)
� Der Schluckakt wird durch eine willkürliche Zun-genbewegung, die den Bolus in den Rachen beför-dert, eingeleitet (orale Phase). Hat der Bissen diePharynxwand erreicht, setzt ein unwillkürlicherReflex ein, bei dem sich drei Einzelmechanismenabgrenzen lassen: Transport des Bissens durch denPharynx, Verschluss des Nasen-Rachen-Raumsund Kehlkopfeingangs und Beförderung des Spei-sebreis zum Ösophaguseingang (pharyngealePhase). In der 3. Phase des Schluckakts passiertder Bissen die Speiseröhre (ösophageale Phase).
� Der Ösophagus dient ausschließlich dem Bolus-transport. Der Schluckakt löst zunächst eine kurz-zeitige Erschlaffung des oberen Sphinkters aus, dievon einer schluckinduzierten peristaltischen Welle(Primärperistaltik) und einer vorübergehendenErschlaffung des unteren Sphinkters gefolgt ist.Die Primärperistaltik wird durch den N. vagus ge-steuert und transportiert feste Nahrung innerhalbvon 10 s in den Magen. Bissensreste können durchlokale Reizung eine sekundäre Peristaltik auslö-sen, die nicht schluckinduziert ist.
Fundus
Pars cardiaca
Antrum
Korpus
kl. K
urva
tur
gr. K
urva
tur
Fundus
Pars cardiaca
Antrum
Korpus
kl. K
urva
tur
Pylorus
Abb. 12.3–1. Magen im Längsschnitt
!
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 390
391M A G E N E N T L E E R U N G
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
12.3.2 Reservoirfunktion des Magens
Die proximalen Magenabschnitte nehmen die Nah-rung auf. Aufgrund von Relaxationsmechanismenkönnen größere Volumina über Stunden hinweg ge-speichert werden, ohne dass der Mageninnendruckmerklich ansteigt.
Die proximalen Magenabschnitte (Fundus und obe-rer Korpusabschnitt) weisen weder eine Automatienoch peristaltische Wellen auf. In dieser Region wirdlediglich durch Vagusimpulse eine anhaltende, d.h.tonische Wandspannung aufgebaut, die sich dem je-weiligen Füllungszustand anpasst. Dieser relativ kon-stante Muskeltonus reicht aus, um Flüssigkeiten beigeöffnetem Pylorus in das Duodenum zu pressen.
Bereits während des Schluckakts, d.h. bevor derBissen aus dem Ösophagus in den Magen übertritt,sinkt der Mageninnendruck aufgrund einer Erschlaf-fung der Magenmuskulatur. Diese als rezeptive Rela-xation bezeichnete Anpassung der Wandspannungwird auf einen vago-vagalen Reflex zurückgeführt.Führt die Nahrungsaufnahme im Magen zur Erregungvon Dehnungssensoren in der Magenwand, tritt einezusätzliche Erschlaffung der Magenmuskulatur auf.Dieser als adaptive Relaxation (oder Akkommoda-tion) bezeichnete Vorgang beruht ebenfalls auf einemvago-vagalen Reflex. Beide Mechanismen erlauben –auch bei voluminösen Mahlzeiten bis zu 1 l – eine Ma-genfüllung, ohne dass der Mageninnendruck erheb-lich ansteigt und verhindern auf diese Weise u.a. einebeschleunigte Entleerung.
Der Dehnungszustand des proximalen Magens wirdweitgehend vom N. vagus gesteuert. Modulierendwirken der Plexus myentericus sowie die gastrointes-tinalen Hormone Gastrin, CCK und Sekretin (s.Tab.12.1–1).
Nach der Aufnahme fester Speisen weist der Ma-geninhalt eine Schichtung auf, wobei die zuletzt auf-genommenen Nahrungsbestandteile an der kleinenKurvatur, die am längsten im Magen befindlichen An-teile im Pylorusbereich liegen. Aufgenommene Flüs-sigkeiten fließen an der Innenwand in distale Magen-abschnitte ab. Der anhaltende Muskeltonus im proxi-malen Magen schiebt den Mageninhalt langsam in un-tere Korpusabschnitte weiter.
12.3.3 Durchmischung und Homogenisierung
Die Automatie, die von einer Schrittmacherzone aus-geht, löst peristaltische Wellen aus, die den Chymus(Speisebrei) bei geschlossenem Pylorus durchmi-schen und homogenisieren.
Im oberen Korpusdrittel liegen an der großen Kur-vatur Schrittmacherzellen, die langsame Potentialwel-len im 20-s-Rhythmus („slow waves“, s.S. 381) bil-den, deren Amplitude vom Dehnungszustand der Ma-genwand abhängt. Erreicht bei zunehmender Füllungdes Magens das Membranpotential die Schwelle, tre-ten Ca2+-getragene Spikeaktivitäten auf, die im Kor-pusabschnitt peristaltische Kontraktionen auslösen.
Die kräftigen, zirkulären peristaltischen Wellen miteiner Frequenz von ca. 3/min wandern pyloruswärtsund schieben den Inhalt in Richtung Magenausgang.Wenn sich die nach distal immer kräftiger werdendeKontraktionswelle dem Antrum nähert, schließt sichder vorher relaxierte Pylorus. Dadurch wird der ein-gezwängte Inhalt mit großer Kraft wieder zurück inden Magen geworfen (Retropulsion). Hierbei reibensich feste Nahrungsbestandteile aneinander und wer-den zerdrückt, zermahlen und intensiv durchmischt(„Antrummühle“). Fette werden dabei auch mecha-nisch emulgiert. Unter dem Einfluss parasympathi-scher Nervenimpulse (N. vagus) tritt eine erheblicheSteigerung der Motilität ein. Motilitätssteigernd wir-ken weiterhin Gastrin und Motilin, hemmend dagegenGIP und Enteroglukagon.
12.3.4 Magenentleerung
Die Flüssigkeitsentleerung aus dem Magen ist beirelaxiertem Pylorus vor allem vom Druckgradientenzwischen proximalem Magen und Duodenum, dieEntleerung von festen Bestandteilen hauptsächlich
Abb. 12.3–2. A Schleimhaut und B Drüse des Magenfundus,modifiziert nach Leonhardt
Drüse
Foveolae gastricae
Muscularis mucosae
Nebenzellen
Belegzellen
Hauptzellen
A B
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 391

vom Pyloruswiderstand und damit letztlich auch vonder Größe der Partikel abhängig. Flüssigkeiten ver-lassen den Magen relativ schnell, feste Bestandteiledagegen erst, wenn sie auf eine Partikelgröße < 2 mmzerkleinert sind.
Die Entleerung des Magens erfolgt, vermitteltdurch den N. vagus, reflektorisch und zwar durch syn-chrone Erschlaffung der Pylorusmuskulatur beimEintreffen peristaltischer Wellen im Antrum. Aller-dings wird der zeitliche Ablauf des Entleerungsvor-gangs von einer Vielzahl weiterer Faktoren beein-flusst. Auch gastrointestinale Hormone sind an derRegulation der Magenentleerung beteiligt (s.Tab.12.1–1). Eine Hemmung der Magenentleerung erfolgtvor allem durch Sekretin, eine Beschleunigung durchMotilin. Eine Unterbrechung des N. vagus, wie z.B.bei einer operativen Vagotomie, führt zu Entleerungs-störungen des Magens mit Retention des Inhalts.
Die Entleerungsrate hängt zusätzlich von der Be-schaffenheit des Mageninhalts (Chymus) ab. Unterreflektorischem Einfluss, vermittelt durch Chemo-sensoren im Duodenum, ist die Verweildauer im Ma-gen länger
� bei saurem als bei neutralem Chymus,
� bei hyperosmolarem als bei hypoosmolarem Chy-mus,
� bei fettreichem als bei eiweißreichem Chymus(wobei allerdings Tryptophan als CCK-Antagonisteine motilitätshemmende Wirkung ausübt) sowie
� bei eiweißreichem als bei kohlenhydratreichemChymus.
Daher variiert die Verweildauer – je nach Zusammen-setzung der Speisen – zwischen 1 und 5 Stunden.
Große, feste Bestandteile können den Magen während dieserEntleerungsphase nicht verlassen. Solche Partikel werdenaber zwischen den Verdauungszeiten, d.h. in der Verdau-ungsruhe, durch den Mechanismus des interdigestiven wan-dernden myoelektrischen Motorkomplexes (s.S. 383 f.) ent-leert. In dessen Phase III kommt es zu kräftigen Antrumkon-traktionen, so dass jetzt auch große unverdaute Nahrungs-partikel oder z.B auch unaufgelöste feste Arzneiformen (u.a.Magensaft-resistente Tabletten) zusammen mit Magensaftdurch den Pylorus in das Duodenum getrieben werden.
Erbrechen. Das Erbrechen (Vomitus, Emesis) stellt einenkomplexen Schutzreflex dar, der von Neuronenverbänden(„Brechzentrum“) im Nucl. tractus solitarii bzw. von einerchemosensiblen Triggerzone im Bereich der Area postremaam Boden des 4. Ventrikels gesteuert wird. Er ist von vege-tativen Begleitsymptomen (Übelkeit, Blässe, Schweiß- undSpeichelsekretion, Blutdruckabfall und Tachykardie) be-gleitet. Das Erbrechen wird durch eine tiefe Inspiration mitnachfolgendem Verschluss der Glottis und des Nasopharynxeingeleitet. Anschließend erschlaffen die Magenmuskulaturund die Ösophagussphinkter, während sich das Zwerchfell
und die Bauchdeckenmuskulatur ruckartig kontrahieren.Letzteres bewirkt eine Erhöhung des intraabdominalenDrucks, und der Mageninhalt wird (teilweise) retrograd ent-leert. Aufgrund einer Tonussteigerung im Duodenum und/oder Pyloruserschlaffung kann es dabei auch zum Erbrechenvon Galle und Duodenalinhalt kommen.
Erbrechen wird durch eine Vielzahl von Ursachen aus-gelöst: durch mechanische Reizung des Oropharynx, mecha-nische und chemische Alteration von Magen und Darm,Entzündungen im Bauchraum, starke Schmerzzustände(Koliken, Herzinfarkt), hormonelle Umstellungen in derSchwangerschaft, Stoffwechselkrankheiten (z.B. nichtrespi-ratorische Azidose bei entgleistem Diabetes mellitus),Reisekrankheit und Schwerelosigkeit im All, Hirndruck-steigerung, Gabe von bestimmten Arzneimitteln (z.B.Apomorphin, Zytostatika), Intoxikationen, Bestrahlung undnicht zuletzt psychische Einflüsse (z.B. ekelerregenden Ge-ruch oder Anblick).
Chronisches Erbrechen führt zum Verlust von H+-, K+-und Cl–-Ionen sowie von Wasser, gefolgt von einer Hypo-volämie und einer nichtrespiratorischen Alkalose (s. S. 513 f.)
12.3.5 Magensaftsekretion
Die Magenmukosa sezerniert täglich 2–3 l Magen-saft, dessen wesentliche Bestandteile Salzsäure, In-trinsic-Faktor, Pepsinogene, Muzine und Bicarbonatsind.
Die Bicarbonat- und Muzinsekretion im Magenerfolgen kontinuierlich. Die HCl- und Pepsinogen-abgabe unterliegen dagegen einer Regulation imZusammenhang mit der Verdauung. Im Nüchtern-zustand (in der interdigestiven Phase) werden nurgeringe Mengen (50–70 ml/h) eines zähflüssigen,neutralen bis leicht alkalischen Sekrets abgegeben.Dagegen kommt es im Zusammenhang mit derNahrungsaufnahme zur Bildung eines stark sauren(pH = 0,8–1,5), nahezu blutisotonen, enzymreichenSekrets.
HCl-Sekretion. Die von den Belegzellen („Parietal-zellen“) unter Mitwirkung der Carboanhydratase ge-bildeten H+-Ionen werden mit Hilfe einer H+/K+-ATPase in intrazelluläre Kanalikuli (s.u.) gepumpt.Die Salzsäure des Magensafts aktiviert die Pepsino-gene, tötet Mikroorganismen ab, setzt Eisen, Calciumund Vitamin B12 aus Nahrungsproteinen frei und de-naturiert noch native Nahrungsproteine, die dann vonProteasen leichter gespalten werden können.
Die Belegzellen sind einzigartig in ihrer Fähigkeit,HCl in hoher Konzentration zu produzieren, wobeieine H+-Konzentrierung etwa um den Faktor 106
gegenüber dem Blut erzielt wird. Sie besitzen Tubu-lovesikel (s.Abb. 12.3–3), deren Membran die pro-tonentransportierende H+/K+-ATPase („Protonen-pumpe“) enthält, und intrazelluläre Kanalikuli, die ander apikalen Seite der Zelle in das Magenlumen ein-
M A G E N392
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 392
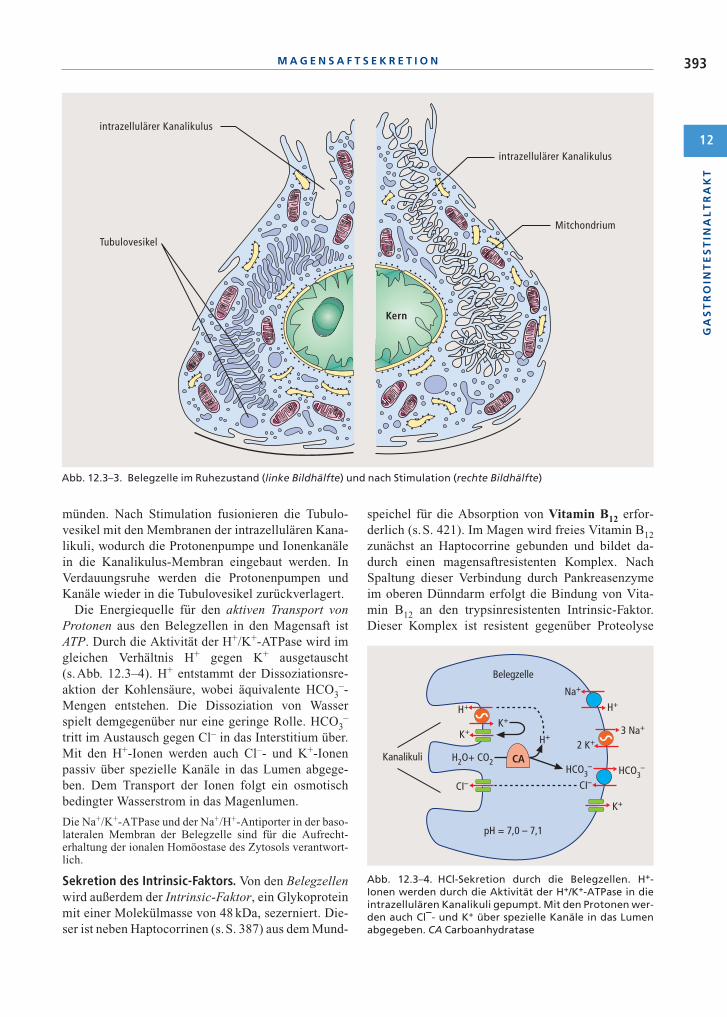
münden. Nach Stimulation fusionieren die Tubulo-vesikel mit den Membranen der intrazellulären Kana-likuli, wodurch die Protonenpumpe und Ionenkanälein die Kanalikulus-Membran eingebaut werden. InVerdauungsruhe werden die Protonenpumpen undKanäle wieder in die Tubulovesikel zurückverlagert.
Die Energiequelle für den aktiven Transport vonProtonen aus den Belegzellen in den Magensaft istATP. Durch die Aktivität der H+/K+-ATPase wird imgleichen Verhältnis H+ gegen K+ ausgetauscht(s.Abb. 12.3–4). H+ entstammt der Dissoziationsre-aktion der Kohlensäure, wobei äquivalente HCO3
–-Mengen entstehen. Die Dissoziation von Wasserspielt demgegenüber nur eine geringe Rolle. HCO3
–
tritt im Austausch gegen Cl– in das Interstitium über.Mit den H+-Ionen werden auch Cl–- und K+-Ionenpassiv über spezielle Kanäle in das Lumen abgege-ben. Dem Transport der Ionen folgt ein osmotischbedingter Wasserstrom in das Magenlumen.
Die Na+/K+-ATPase und der Na+/H+-Antiporter in der baso-lateralen Membran der Belegzelle sind für die Aufrecht-erhaltung der ionalen Homöostase des Zytosols verantwort-lich.
Sekretion des Intrinsic-Faktors. Von den Belegzellenwird außerdem der Intrinsic-Faktor, ein Glykoproteinmit einer Molekülmasse von 48 kDa, sezerniert. Die-ser ist neben Haptocorrinen (s.S. 387) aus dem Mund-
speichel für die Absorption von Vitamin B12 erfor-derlich (s.S. 421). Im Magen wird freies Vitamin B12zunächst an Haptocorrine gebunden und bildet da-durch einen magensaftresistenten Komplex. NachSpaltung dieser Verbindung durch Pankreasenzymeim oberen Dünndarm erfolgt die Bindung von Vita-min B12 an den trypsinresistenten Intrinsic-Faktor.Dieser Komplex ist resistent gegenüber Proteolyse
393M A G E N S A F T S E K R E T I O N
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
•••
• • ••
•
•
•
••
•••
• • ••
• ••
••
•••
•
••••••
•
•••
•
• • • • •
•••••
•
• • • • •
••
•••
• • ••
••
••
•• •
••
••
•
•
•
••
••
••
••
•
•
•
••
••
•
•
••
••
••
•
• ••
•
••
•
•
•
•
•••
••
•
•
••
•••
•• •
••
••
•• •
•
•
•
•
•
••
•
•
••
••
• ••
•••
••
••
••
••
••
••
••
•••
••••••••
••••
••
•
••
••
••
•••
••
•
••
• • • •• • • • •
• • • • • • • ••
•
intrazellulärer Kanalikulus
Kern
Mitchondrium
intrazellulärer Kanalikulus
Tubulovesikel
Abb. 12.3–3. Belegzelle im Ruhezustand (linke Bildhälfte) und nach Stimulation (rechte Bildhälfte)
H2O+ CO2
H+
K+
CI–
Na+
H+
HCO3– HCO3
–
K+
2 K+H+
K+
CI–
3 Na+
Belegzelle
pH = 7,0 – 7,1
Kanalikuli CA
Abb. 12.3–4. HCl-Sekretion durch die Belegzellen. H+-Ionen werden durch die Aktivität der H+/K+-ATPase in dieintrazellulären Kanalikuli gepumpt. Mit den Protonen wer-den auch Cl–- und K+ über spezielle Kanäle in das Lumenabgegeben. CA Carboanhydratase
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 393

und Absorption im oberen Dünndarm. Er wird schließ-lich durch rezeptorvermittelte Endozytose in die Mu-kosa des Ileums aufgenommen. Funktionsbestandteiledieses spezifischen Rezeptors sind die Proteine Cubi-lin und Megalin.
Sekretion von Pepsinogenen. Die Hauptzellen derMagenmukosa geben ein Gemisch von Proteasenvor-stufen (Pepsinogenen) ab. Die Stimulation der Pepsi-nogensekretion erfolgt auf dieselbe Weise wie die derSalzsäureproduktion. Es lassen sich acht verschie-dene Vorstufen dieser proteolytischen Isoenzyme(Endopeptidasen) elektrophoretisch nachweisen. Siewerden durch die Magensalzsäure zu den wirksameneiweißspaltenden Enzymen, den Pepsinen, durch hyd-rolytische Abspaltung eines blockierenden Peptidsaktiviert, ein Vorgang, der sich anschließend autoka-talytisch fortsetzt. Die Pepsine wirken nur bei saurenpH-Werten mit Optima zwischen 1,8 und 3,5; im alka-lischen Milieu werden sie irreversibel geschädigt.
Sekretion von Schleim und Bicarbonat. In den Ober-flächen- und Nebenzellen der Korpusdrüsen sowie inden Kardia- und Pylorusdrüsen wird Schleim (Mu-zin) produziert, der den gesamten Magen mit einer bis0,6 mm dicken Schicht als visköses Gel überzieht. Ererzeugt einen Gleitfilm und schützt die Schleimhautvor mechanischen und chemischen Schäden. DieSchleimschicht muss ständig intakt gehalten bzw.erneuert werden, da sie sonst anhaltenden mechani-schen und enzymatischen Angriffen ausgesetzt wäre.Hauptbestandteile des Schleims sind unterschiedlicheSaccharid-reiche Glykoproteine (Muzine).
Salzsäure und Pepsinogene gelangen durch 5 µmmessende feine Kanäle in der Schleimschicht von derapikalen Zellmembran ins Magenlumen.
Neben Schleim wird vom Oberflächenepithel desMagens Bicarbonat gebildet, das durch einenHCO3
_-Kanal sezerniert wird.
Bicarbonat hat zusammen mit dem Magenschleimeine wichtige Schutzfunktion gegenüber dem aggres-siven Magensaft. Das gebildete HCO3
– wird in derdem Magenepithel aufliegenden, strömungsfreienFlüssigkeits- bzw. Schleimschicht (unstirred layer)festgehalten und erzeugt dadurch einen pH-Gradien-ten von pH ≈ 7 in der Zelle auf pH ≈ 2 im Magen-lumen. Damit findet man den durch die Salzsäure be-dingten niedrigen pH-Wert nicht schon an der Epi-theloberfläche, sondern erst im Magenlumen. Da-rüber hinaus gelangt Bicarbonat, das in den Belegzel-len während der Sekretionsphase vermehrt gebildetund in das Blut abgegeben wird (s.Abb. 12.3–4),durch senkrecht in der Schleimhaut verlaufende Ka-
pillarschlingen zur Epitheloberfläche. Die Durchblu-tung dieser Kapillaren wird wesentlich durch Prosta-glandin E2 (PGE2) gesteuert, dem somit im Zusam-menspiel mit Bicarbonat und der strömungsfreienSchicht eine wichtige protektive Funktion für die Ma-genschleimhaut zukommt.
Mukosabarriere. Zu den protektiven Mechanismen der sog.Mukosabarriere zählen neben der bicarbonathaltigen, strö-mungsfreien Muzinschicht die Unversehrtheit der Membra-nen aller Oberflächenzellen. Diese wird durch eine guteSchleimhautdurchblutung, eine ungestörte PGE2-Wirkung(Steigerung der Durchblutung, der Schleim- und HCO–
3-Sekretion), die Intaktheit der interzellulären Schlussleistenund die Fähigkeit zur Epithelregeneration gewährleistet.Letztere wird durch eine Reihe von Wachstumsfaktorenkontrolliert.
Zu den aggressiven Faktoren, die den Schutz der Magen-schleimhaut gegen die von ihren Drüsen produzierten Pep-sine und HCl vermindern („Barrierenbrecher“), rechnet manbiologische Detergenzien (Gallensalze und Lysolecithin beiGallereflux), Glucocorticoide, nichtsteroidale entzündungs-hemmende Arzneimittel wie Acetylsalicylsäure (Hemmerder Prostaglandinsynthese), Minderdurchblutung der Schleim-haut sowie eine bakterielle Infektion mit Helicobacter pylo-ri (s.S. 430 f.).
M A G E N394
kephalePhase
gastralePhase
intes-tinalePhase
ACh
GRP
Somatostatin
Sekretin,GIP,Neurotensin
Histamin
Gastrin-sekretion
lokaleReflexe
HCI-Sekretion
GeruchGeschmackAnblickVorstellung
Magen-dehnung
Magen– Eiweiß- abbau- produkte
Antrum– pH < 3
Dünndarm– Dehnung– Eiweiß- abbau- produkte
Dünndarm– pH < 4– Fette– Hyper- osmolarität
ECL-Zellen
N. vagus
Abb. 12.3–5. Schematische Darstellung der an der HCl-Sekretion beteiligten fördernden und hemmenden Mecha-nismen. ACh Acetylcholin, GRP gastrin releasing peptide,GIP gastrin inhibitory peptide; ––ı hemmende Einflüsse, �fördernde Einflüsse
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 394
395M A G E N S A F T S E K R E T I O N
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
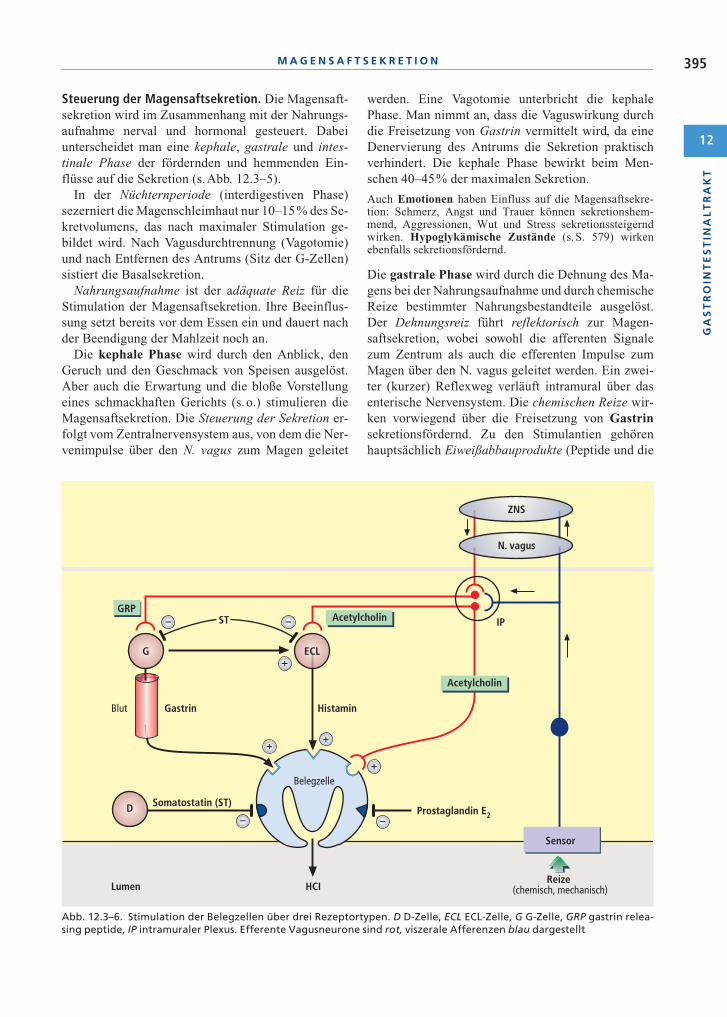
Steuerung der Magensaftsekretion. Die Magensaft-sekretion wird im Zusammenhang mit der Nahrungs-aufnahme nerval und hormonal gesteuert. Dabeiunterscheidet man eine kephale, gastrale und intes-tinale Phase der fördernden und hemmenden Ein-flüsse auf die Sekretion (s.Abb. 12.3–5).
In der Nüchternperiode (interdigestiven Phase)sezerniert die Magenschleimhaut nur 10–15% des Se-kretvolumens, das nach maximaler Stimulation ge-bildet wird. Nach Vagusdurchtrennung (Vagotomie)und nach Entfernen des Antrums (Sitz der G-Zellen)sistiert die Basalsekretion.
Nahrungsaufnahme ist der adäquate Reiz für dieStimulation der Magensaftsekretion. Ihre Beeinflus-sung setzt bereits vor dem Essen ein und dauert nachder Beendigung der Mahlzeit noch an.
Die kephale Phase wird durch den Anblick, denGeruch und den Geschmack von Speisen ausgelöst.Aber auch die Erwartung und die bloße Vorstellungeines schmackhaften Gerichts (s.o.) stimulieren dieMagensaftsekretion. Die Steuerung der Sekretion er-folgt vom Zentralnervensystem aus, von dem die Ner-venimpulse über den N. vagus zum Magen geleitet
werden. Eine Vagotomie unterbricht die kephalePhase. Man nimmt an, dass die Vaguswirkung durchdie Freisetzung von Gastrin vermittelt wird, da eineDenervierung des Antrums die Sekretion praktischverhindert. Die kephale Phase bewirkt beim Men-schen 40–45% der maximalen Sekretion.
Auch Emotionen haben Einfluss auf die Magensaftsekre-tion: Schmerz, Angst und Trauer können sekretionshem-mend, Aggressionen, Wut und Stress sekretionssteigerndwirken. Hypoglykämische Zustände (s.S. 579) wirkenebenfalls sekretionsfördernd.
Die gastrale Phase wird durch die Dehnung des Ma-gens bei der Nahrungsaufnahme und durch chemischeReize bestimmter Nahrungsbestandteile ausgelöst.Der Dehnungsreiz führt reflektorisch zur Magen-saftsekretion, wobei sowohl die afferenten Signalezum Zentrum als auch die efferenten Impulse zumMagen über den N. vagus geleitet werden. Ein zwei-ter (kurzer) Reflexweg verläuft intramural über dasenterische Nervensystem. Die chemischen Reize wir-ken vorwiegend über die Freisetzung von Gastrinsekretionsfördernd. Zu den Stimulantien gehörenhauptsächlich Eiweißabbauprodukte (Peptide und die
+
–
+
+
–
G
Blut Gastrin Histamin
Belegzelle
–
+ECL
– ST
Somatostatin (ST)Prostaglandin E2
HCI
Sensor
ReizeLumen (chemisch, mechanisch)
Acetylcholin
Acetylcholin IP
D
ZNS
N. vagus
GRP
Abb. 12.3–6. Stimulation der Belegzellen über drei Rezeptortypen. D D-Zelle, ECL ECL-Zelle, G G-Zelle, GRP gastrin relea-sing peptide, IP intramuraler Plexus. Efferente Vagusneurone sind rot, viszerale Afferenzen blau dargestellt
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 395

aromatischen Aminosäuren Phenylalanin und Tryp-tophan), ferner Ca2+-Ionen sowie Alkohol (Aperitif-Effekt) und Kaffee (Coffein, Röststoffe). Die gastralePhase trägt 50–55% zur maximalen Sekretion bei.
Bei pH < 3 im Antrum wird Somatostatin freige-setzt, das parakrin die Gastrinproduktion und endo-krin auch die Magensaftsekretion hemmt (negativeRückkopplung).
In der intestinalen Phase kann die Magensaftse-kretion vom Duodenum her sowohl fördernd als auchhemmend beeinflusst werden. Die Dehnung derDarmwand und der Übertritt von Eiweiß oder dessenSpaltprodukten ins Duodenum fördert über (nochnicht identifizierte) humorale Faktoren die Magen-saftsekretion, allerdings nur in geringem Maß (5%der Maximalsekretion).
Wichtiger ist in dieser Phase jedoch der hemmendeEinfluss. Tritt saurer (pH < 4), stark fetthaltiger oderhyperosmolarer Chymus in das Duodenum über, er-folgt dort eine Freisetzung von Sekretin, das die HCl-Sekretion hemmt und damit eine weitere Säure-belastung verhindert, die Pepsinogensekretion dage-gen stimuliert. Bei stark fetthaltigem Darminhalt wirddie Säuresekretion zusätzlich durch die Peptide Neu-rotensin, Peptid YY und GIP gehemmt.
Aktivierung der Belegzellen. Acetylcholin, Gastrinund Histamin reagieren mit jeweils spezifischenRezeptoren in der basolateralen Membran der Be-legzellen und aktivieren damit die HCl-Sekretion(s.Abb. 12.3–6).
Acetylcholin reagiert mit m3-Rezeptoren und Gast-rin mit Gastrinrezeptoren (CCK-2), wobei in beidenFällen die Sekretion über den second messenger IP3angeregt wird. Histamin, das aus den ECL-Zellen(enterochromaffin-like cells) der Magendrüsen undaus Mastzellen freigesetzt wird, reagiert mit H2-Re-zeptoren. Dadurch kommt es, vermittelt durch den se-cond messenger cAMP, zur Aktivierung der HCl-Sekretion.
Histamin spielt eine zentrale Rolle bei der Regula-tion der HCl-Sekretion. Da die ECL-Zellen sowohldurch Gastrin als auch durch Vagusaktivierung stimu-liert werden, lässt sich durch Blockade der H2-Rezep-toren mit H2-Antagonisten sowohl die durch Gastrinals auch die durch Acetylcholin vermittelte Sekretionherabsetzen. Eine Hemmung der HCl-Sekretion be-wirken auch Prostaglandin E2 und Somatostatin.
Neben der direkten Aktivierung der Belegzellen über m3-Cholinozeptoren wirkt der N. vagus auch indirekt stimulie-rend auf die Belegzellen, indem er die Gastrinfreisetzung ausden G-Zellen fördert. Als postganglionäre Überträgersub-stanz wird hierbei gastrin releasing peptide (GRP, Bom-besin) diskutiert.
M A G E N396
!In Kürze !� Im Magen werden die festen Nahrungsbestand-
teile gespeichert, zerkleinert und homogenisiert.Nach einer Verweildauer von 1–5 h erfolgt die por-tionsweise Entleerung des Speisebreis (Chymus)ins Duodenum.
� Am Magen werden folgende Abschnitte unter-schieden: Kardia, Fundus, Korpus, Antrum undPylorus.
� Die Magenwand besteht im Wesentlichen aus dreiSchichten glatter Muskulatur, die pyloruswärtsan Dicke zunehmen, und der gefalteten Magen-schleimhaut mit den für die verschiedenen Re-gionen typischen Drüsen.
� Das maximale Füllungsvolumen des Magensbeträgt 1,5–2 l.
� Die proximalen Magenabschnitte nehmen dieNahrung auf. Durch den N. vagus gesteuerte Rela-xationsmechanismen (rezeptive und adaptiveRelaxation) ermöglichen die Speicherung größe-rer Volumina über Stunden, ohne dass der Magen-innendruck merklich ansteigt.
� Von einer Schrittmacherzone im oberen Korpus-bereich gehen bei Wanddehnung peristaltischeWellen aus, die den Chymus bei geschlossenemPylorus durchmischen und homogenisieren.
� Die Entleerung des Magens erfolgt portionsweisebeim Eintreffen peristaltischer Wellen im Antrumdurch synchrone Erschlaffung des Pylorus. Siewird reflektorisch über den N. vagus gesteuert,aber auch durch gastrointestinale Hormone sowiedie Zusammensetzung und Beschaffenheit derNahrung bzw. des Chymus im Magen und im Duo-denum beeinflusst.
� Die Magenmukosa sezerniert täglich 2–3 l Ma-gensaft, dessen wesentliche Bestandteile Salz-säure, Intrinsic-Faktor, Pepsinogene, Muzin(e)und Bicarbonat sind.
� Die Bicarbonat- und Muzinsekretion durch das Oberflächenepithel erfolgt kontinuierlich.Schleim wird zusätzlich von den Nebenzellen derKorpusdrüsen sowie von Kardia- und Pylorus-drüsen gebildet.
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 396

12.4 Dünndarm
Der Dünndarm, in dem die Verdauungsvorgänge fort-gesetzt und die dabei anfallenden kleinmolekularenBruchstücke zum überwiegenden Teil absorbiert wer-den, erstreckt sich vom Pylorus des Magens bis inHöhe der rechten Darmbeinschaufel, wo er in denDickdarm einmündet.
Er gliedert sich in 3 Abschnitte:
� das Duodenum (Zwölffingerdarm),
� das am Treitz-Band beginnende Jejunum (Leer-darm) und
� das Ileum (Krummdarm), das sich ohne definierteGrenze anschließt.
Die Gesamtlänge des Dünndarms beträgt im tonisier-ten Zustand (in vivo) etwa 3,75 m, im relaxierten (postmortem) etwa 6 m.
12.4.1 Anatomie des Dünndarms
Makroskopische Anatomie. Der erste Dünndarmab-schnitt, das 25–30 cm lange Duodenum, hat die Formeines C, in dessen Konkavität der Kopf der Bauch-speicheldrüse eingebettet ist (s.Abb. 12.6–2, S. 405).Der obere Abschnitt ist zum Bulbus duodeni erwei-tert. In den absteigenden Schenkel münden der Aus-führungsgang der Bauchspeicheldrüse und der Gal-lengang in einem gemeinsamen Endstück (s.S. 414).
In Höhe des 2. Lendenwirbels geht das Duodenum indas etwa 1,5 m lange Jejunum über. An diesesschließt sich das etwa 2 m lange Ileum an, dessenBezeichnung auf den vielfach gewundenen Verlaufhinweist. Die Schlingen von Jejunum und Ileum sindam Mesenterium (Gekröse) aufgehängt.
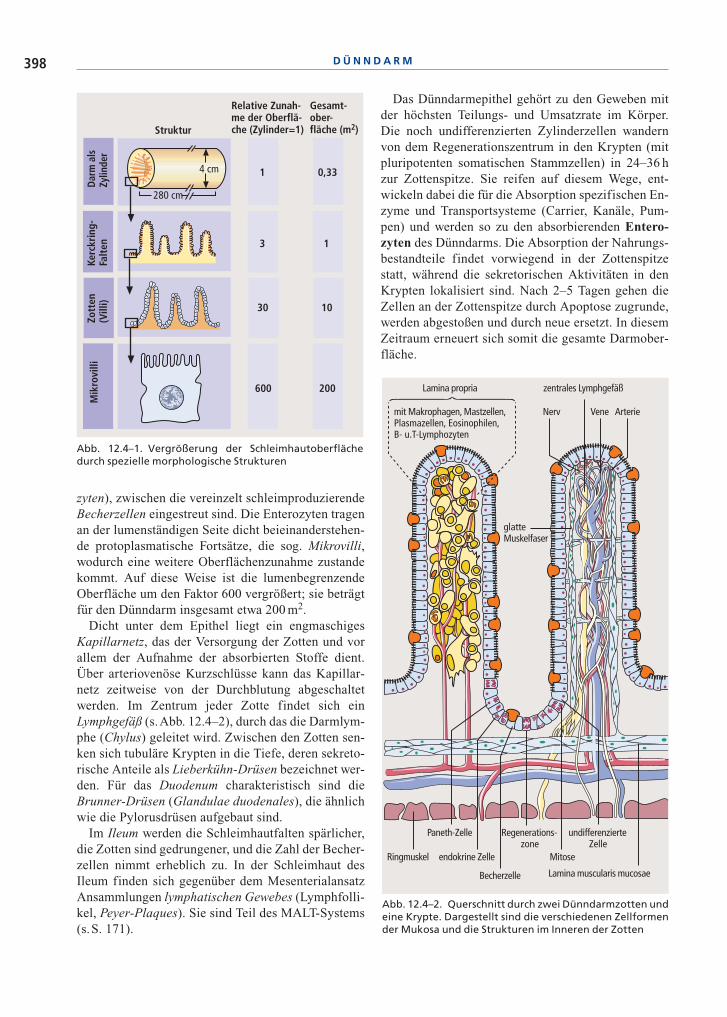
Wandaufbau. Der Wandaufbau des Dünndarms wur-de bereits in Abb. 12.1–2 (s.S. 378) schematisch dar-gestellt. Die Schleimhaut, die neben der einschichti-gen Epithelschicht und dem Schleimhautbindegewe-be eine eigene feine Muskulatur enthält, ist von einerRingmuskelschicht umgeben, an die sich nach außenhin eine Längsmuskelschicht anschließt. Dazwischenliegen zwei Nervengeflechte, der Plexus submucosus(Meißner-Plexus), der die Schleimhaut innerviert,und der Plexus myentericus (Auerbach-Plexus), derdie Muskulatur versorgt (s.S. 378). Das Mesenterium,eine Bauchfellduplikatur, nimmt die zuführendenbzw. abführenden Arterien, Venen, Lymphgefäße undNerven auf.
Schleimhautaufbau. Die Schleimhaut des Dünn-darms ist so strukturiert, dass eine starke Vergröße-rung der Oberfläche erzielt wird (s.Abb. 12.4–1). DieOberflächenvergrößerung entsteht in der ersten Stufedurch zirkuläre Schleimhautfalten (Ring- oder Kerck-ring-Falten). Auf diesen Falten befinden sich finger-förmige etwa 1 mm hohe Zotten, die den zweitenVergrößerungsfaktor darstellen. Das Epithel der Zot-ten besteht vorwiegend aus sog. Saumzellen (Entero-
397A N A T O M I E D E S D Ü N N D A R M S
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
In Kürze (Fortsetzung)
� Die Muzine machen den Chymus gleitfähig undhaben – zusammen mit Bicarbonat in der strö-mungsfreien Schleimschicht – protektive Eigen-schaften für die Magenschleimhaut.
� Die Hauptzellen geben ein Gemisch von Pepsino-genen ab, deren Aktivierung zu Pepsinen durchHCl eingeleitet und autokatalytisch fortgesetztwird.
� Die von den Belegzellen gebildeten H+-Ionenwerden mit Hilfe einer H+/K+-ATPase (Protonen-pumpe) sezerniert, wodurch im Magenlumen eine106 höhere H+-Konzentration im Vergleich mitdem Zytosol erreicht wird (pH-Wert ≈ 1). Zusam-men mit den Protonen werden Cl–-und K+-Ionenüber entsprechende Kanäle ins Lumen abgegeben.
� Die Belegzellen besitzen Rezeptoren für Acetyl-
cholin, Histamin und Gastrin, über welche dieHCl-Sekretion stimuliert wird.
� Der ebenfalls von den Belegzellen sezernierte In-trinsic-Faktor ist essentiell für die Bindung undspätere Absorption von Vitamin B12 im Ileum.
� Die mit der Nahrungsaufnahme gekoppelte Ma-gensaftsekretion wird nerval und hormonal ge-steuert. Ausgelöst wird die Sekretion durch Sin-neseindrücke, Vorstellungen und beim Kontakt derNahrung mit der Mundschleimhaut (kephale Pha-se). Unterhalten wird sie durch eine Dehnung vonMagen und Duodenum sowie durch Eiweißspalt-produkte (gastrale Phase). Während der intesti-nalen Phase wird die Sekretion durch Peptide imDuodenum stimuliert, durch Fette und saurenChymus gehemmt.
!
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 397
D Ü N N D A R M398
zyten), zwischen die vereinzelt schleimproduzierendeBecherzellen eingestreut sind. Die Enterozyten tragenan der lumenständigen Seite dicht beieinanderstehen-de protoplasmatische Fortsätze, die sog. Mikrovilli,wodurch eine weitere Oberflächenzunahme zustandekommt. Auf diese Weise ist die lumenbegrenzendeOberfläche um den Faktor 600 vergrößert; sie beträgtfür den Dünndarm insgesamt etwa 200 m2.
Dicht unter dem Epithel liegt ein engmaschigesKapillarnetz, das der Versorgung der Zotten und vorallem der Aufnahme der absorbierten Stoffe dient.Über arteriovenöse Kurzschlüsse kann das Kapillar-netz zeitweise von der Durchblutung abgeschaltetwerden. Im Zentrum jeder Zotte findet sich einLymphgefäß (s.Abb. 12.4–2), durch das die Darmlym-phe (Chylus) geleitet wird. Zwischen den Zotten sen-ken sich tubuläre Krypten in die Tiefe, deren sekreto-rische Anteile als Lieberkühn-Drüsen bezeichnet wer-den. Für das Duodenum charakteristisch sind dieBrunner-Drüsen (Glandulae duodenales), die ähnlichwie die Pylorusdrüsen aufgebaut sind.
Im Ileum werden die Schleimhautfalten spärlicher,die Zotten sind gedrungener, und die Zahl der Becher-zellen nimmt erheblich zu. In der Schleimhaut desIleum finden sich gegenüber dem MesenterialansatzAnsammlungen lymphatischen Gewebes (Lymphfolli-kel, Peyer-Plaques). Sie sind Teil des MALT-Systems(s.S. 171).
Das Dünndarmepithel gehört zu den Geweben mitder höchsten Teilungs- und Umsatzrate im Körper.Die noch undifferenzierten Zylinderzellen wandernvon dem Regenerationszentrum in den Krypten (mitpluripotenten somatischen Stammzellen) in 24–36 hzur Zottenspitze. Sie reifen auf diesem Wege, ent-wickeln dabei die für die Absorption spezifischen En-zyme und Transportsysteme (Carrier, Kanäle, Pum-pen) und werden so zu den absorbierenden Entero-zyten des Dünndarms. Die Absorption der Nahrungs-bestandteile findet vorwiegend in der Zottenspitzestatt, während die sekretorischen Aktivitäten in denKrypten lokalisiert sind. Nach 2–5 Tagen gehen dieZellen an der Zottenspitze durch Apoptose zugrunde,werden abgestoßen und durch neue ersetzt. In diesemZeitraum erneuert sich somit die gesamte Darmober-fläche.
Abb. 12.4–1. Vergrößerung der Schleimhautoberflächedurch spezielle morphologische Strukturen
Struktur
Relative Zunah-me der Oberflä-che (Zylinder=1)
Gesamt-ober-fläche (m2)
Mik
rovi
lliZo
tten
(Vill
i)Ke
rckr
ing-
Falt
enD
arm
als
Zylin
der
280 cm
4 cm
600
30
3
1
200
10
1
0,33
glatteMuskelfaser
Lamina propria
mit Makrophagen, Mastzellen,Plasmazellen, Eosinophilen,B- u.T-Lymphozyten
zentrales Lymphgefäß
Nerv Vene Arterie
Ringmuskel
Paneth-Zelle
endokrine Zelle
Becherzelle
Regenerations-zone
Mitose
undifferenzierteZelle
Lamina muscularis mucosae
Abb. 12.4–2. Querschnitt durch zwei Dünndarmzotten undeine Krypte. Dargestellt sind die verschiedenen Zellformender Mukosa und die Strukturen im Inneren der Zotten
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 398

Außer den Enterozyten enthält die Dünndarm-schleimhaut noch schleimbildende Becherzellen undverschiedene endokrine Zellen (z.B. amine content,precursor uptake, ability to decarboxylate cells, sog.APUD-Zellen, die u.a. Serotonin bilden). Die M-Zellen gehören zum Darm-assoziierten Lymphgewe-be (s.S. 398). An der Abwehr sind auch die Paneth-Zellen an der Kryptenbasis beteiligt, die Lysozym undDefensine bilden.
12.4.2 Dünndarmmotilität
Darmwandbewegungen. Durch die Bewegungen desDünndarms wird der Nahrungsbrei in der digestivenPhase mit den Verdauungssäften, insbesondere mitdem Pankreassekret und der Galle, intensiv durch-mischt. Dabei unterscheidet man verschiedene Be-wegungsformen, die durch Kontraktionen der Ring-und Längsmuskulatur zustande kommen: die rhythmi-schen Segmentationen und Pendelbewegungen (s.S.382).
Die Durchmischungsbewegungen im Dünndarmwerden in erster Linie durch die Schrittmacher-Au-tomatie gesteuert, die langsame Depolarisationswel-len (slow waves, s.S. 380) mit überlagerten Aktions-potentialen auslöst. Die Schrittmacher der langsamenWellen haben im Duodenum eine intrinsische Fre-quenz von ca. 12/min; diese nimmt stufenweise auf8/min im Ileum ab. Durch diesen Frequenzgradientenvon oben nach unten wird auch bei den nichtpropulsi-ven Segmentationen eine langsame Verschiebung desDarminhalts nach distal gewährleistet, da mit demFrequenzgefälle ein gleichgerichteter Druckgradientim Darm verbunden ist.
Für das Auftreten peristaltischer Wellen (s.S. 382 f.),welche die Durchmischungsvorgänge überlagern undden Inhalt (in Abhängigkeit von der Nahrungszusam-mensetzung) in 2–4 h zum Caecum befördern, sindvor allem motorische Aktivitäten des enterischenNervensystems verantwortlich. Sie werden insbeson-dere durch Dehnung der Darmwand ausgelöst undsind an die Erregungsimpulse aus dem Plexus myen-tericus gebunden. Parasympathikus und Sympathikushaben lediglich einen modulierenden Einfluss auf diePeristaltik (s.Abb. 12.1–4, S. 382).
Der Plexus submucosus erhält Signale von Mecha-no- und Chemosensoren, die über viszerale Afferen-zen entweder die Medulla oblongata oder das Rücken-mark erreichen. Die sympathischen Efferenzen hem-men vor allem erregende Darmneurone, wodurch derTonus der Darmmuskulatur herabgesetzt wird. Dieglatte Sphinktermuskulatur wird dagegen stimuliert.Eine Aktivierung der parasympathischen Efferenzen
(N. vagus) hat in der Regel eine Tonussteigerung zurFolge.
Der Einfluss gastrointestinaler Hormone und Pep-tide auf die Dünndarmmotilität ist gering bzw. unklar.Gesichert ist lediglich die motilitätssteigernde Wir-kung von Cholecystokinin.
Zottenbewegungen. Die stempelartigen Bewegun-gen der Zotten dienen der besseren Durchmischungdes Darminhalts und wirbeln die ruhende, den Entero-zyten aufliegende Schleimschicht (unstirred layer)auf. Die Kontraktion fördert auch die Entleerung derzentral in der Zotte verlaufenden Lymphkapillare(Chylusgefäß) in größere Lymphgefäße tiefererDarmwandschichten. Die Zottenbewegungen werdendurch das in der Dünndarmmukosa lokalisierte PeptidVillikinin aktiviert.
Ileozäkaler Übergang. Am Ende des Dünndarms kontrol-liert ein ca. 4 cm langes Segment den Übertritt von Darm-inhalt in den Dickdarm. Dieser Sphinkter ist andauernd kon-trahiert und erzeugt eine Zone erhöhten Drucks von ca. 20 mm Hg. Bei Dehnung des terminalen Ileums erschlafftder Sphinkter, bei Druckerhöhung im Caecum steigt seinTonus. Darüber hinaus bildet der als Bauhin-Klappe (Valvaileocaecalis, s.Abb. 12.5–1) in das Caecum hineinragendeEndteil des Ileums ein Ventil, das Drücken im Caecum biszu 40 mm Hg widersteht. Aufgrund dieser anatomischenBarriere ist die Bakterienbesiedlung im Ileum um den Faktor105 niedriger als im Caecum.
12.4.3 Dünndarmsekretion
Zusammensetzung des Dünndarmsekrets. DieDünndarmmukosa produziert täglich 2,5–3,0 l Darm-saft. Dessen Hauptbestandteile sind darmwandschüt-zende Muzine, die von den Brunner-Drüsen (im Duo-denum) sowie von den Becherzellen der Zotten undLieberkühn-Krypten sezerniert werden. Aufgrund derhohen HCO3
–-Konzentration des Sekrets der Brun-ner-Drüsen weist das Duodenalsekret einen pH-Wertvon 8–9 auf.
Muzine überziehen die Darmwand in einer gelarti-gen Schicht (unstirred layer, s.S. 394) und schützendas Epithel des Duodenums vor der Einwirkung vonsaurem Chymus sowie das gesamte Dünndarmepithelvor der Zerstörung durch Proteasen. Außerdem ermög-lichen sie ein reibungsfreies Gleiten des Darminhalts.
Die Hauptzellen der Krypten sezernieren eine plas-maisotone NaCl-Lösung. Cl– wird auf der basolatera-len Seite sekundär-aktiv in die Zellen aufgenommenund über Cl–-Kanäle an das Darmlumen abgegeben.Die Kanäle können durch VIP und Acetylcholin unterVermittlung von Botenstoffen (cAMP bzw. IP3, DAG)aktiviert werden. Wasser und Na+ folgen dem trans-zellulären Cl–-Transport auf parazellulärem Weg.
399D Ü N N D A R M S E K R E T I O N
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 399
Der Darmsaft enthält praktisch keine Enzyme.Durch Abschilferung von Mukosazellen können aller-dings sekundär Enzyme, die im Bürstensaum dieserZellen lokalisiert sind, in das Darmlumen gelangen.
Regulation der Dünndarmsekretion. Die Sekretions-vorgänge im Dünndarm werden sowohl neuronal alsauch humoral reguliert. Die Submukosa enthält reich-lich Chemo- und Mechanosensoren, die auf Ände-rungen der Zusammensetzung des Darminhalts (u.a.pH-Wert, Aminosäurenkonzentration) und auf Be-rührung reagieren. Über lokale (intramurale) Reflex-wege werden die Drüsenzellen aktiviert (s.Abb.
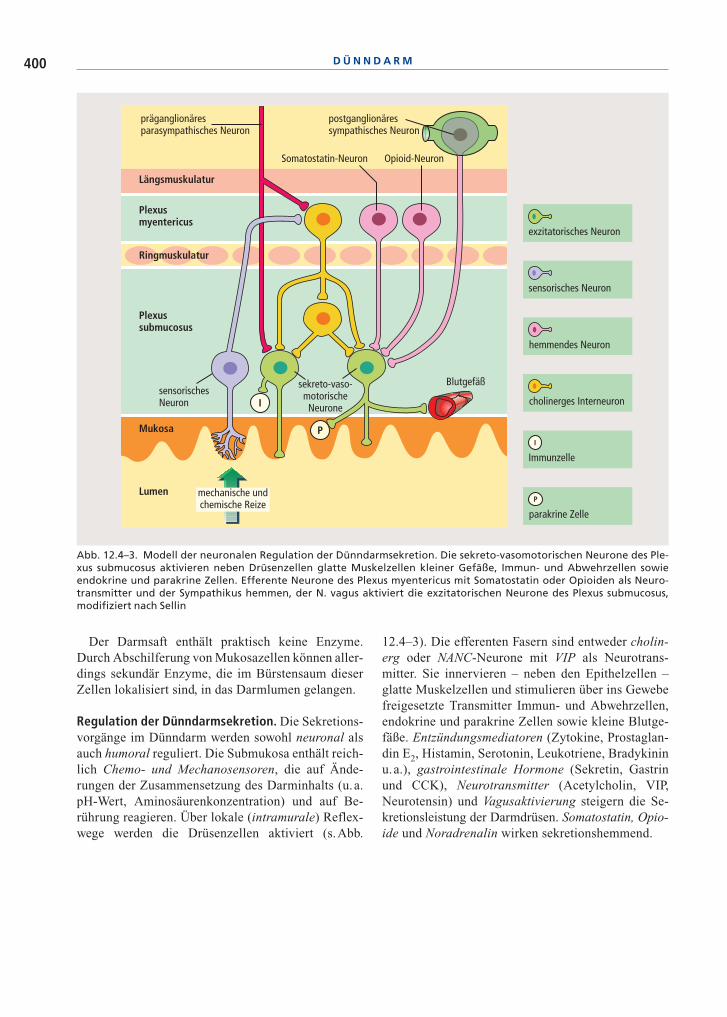
12.4–3). Die efferenten Fasern sind entweder cholin-erg oder NANC-Neurone mit VIP als Neurotrans-mitter. Sie innervieren – neben den Epithelzellen –glatte Muskelzellen und stimulieren über ins Gewebefreigesetzte Transmitter Immun- und Abwehrzellen,endokrine und parakrine Zellen sowie kleine Blutge-fäße. Entzündungsmediatoren (Zytokine, Prostaglan-din E2, Histamin, Serotonin, Leukotriene, Bradykininu.a.), gastrointestinale Hormone (Sekretin, Gastrinund CCK), Neurotransmitter (Acetylcholin, VIP,Neurotensin) und Vagusaktivierung steigern die Se-kretionsleistung der Darmdrüsen. Somatostatin, Opio-ide und Noradrenalin wirken sekretionshemmend.
D Ü N N D A R M400
Ringmuskulatur
Plexussubmucosus
Lumen
Längsmuskulatur
Plexusmyentericus
I
P
sensorischesNeuron
postganglionäressympathisches Neuron
Somatostatin-Neuron Opioid-Neuron
sekreto-vaso-motorischeNeurone
präganglionäresparasympathisches Neuron
Mukosa
Blutgefäß
mechanische undchemische Reize
sensorisches Neuron
I
Immunzelle
exzitatorisches Neuron
P
parakrine Zelle
hemmendes Neuron
cholinerges Interneuron
Abb. 12.4–3. Modell der neuronalen Regulation der Dünndarmsekretion. Die sekreto-vasomotorischen Neurone des Ple-xus submucosus aktivieren neben Drüsenzellen glatte Muskelzellen kleiner Gefäße, Immun- und Abwehrzellen sowieendokrine und parakrine Zellen. Efferente Neurone des Plexus myentericus mit Somatostatin oder Opioiden als Neuro-transmitter und der Sympathikus hemmen, der N. vagus aktiviert die exzitatorischen Neurone des Plexus submucosus,modifiziert nach Sellin
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 400

12.5 Dickdarm
Den letzten Abschnitt des Intestinaltrakts bildet derDickdarm. Man unterscheidet folgende Abschnitte:
� Caecum (Blinddarm) mit der Appendix vermifor-mis (Wurmfortsatz),
� Kolon (Grimmdarm) und
� Rektum (Mastdarm oder Enddarm).
In den einzelnen Abschnitten des Dickdarms werdendurch Eindickung des Darminhalts und durch Bei-mengung von Schleim die Fäzes (Kot, „Stuhl“) gebil-det, doch entsteht „Stuhl“ auch ohne Nahrungszufuhraus Schleim, abgeschilferten Darmzellen und Bak-terien.
12.5.1 Anatomie des Dickdarms
Makroskopische Anatomie. Der Dickdarm beginntim Bereich der rechten Darmbeinschaufel mit derbereits erwähnten Valva ileocaecalis (Bauhin-Klap-pe). An dieser Stelle mündet das Ileum seitlich in denaufsteigenden Dickdarmabschnitt, so dass unterhalbder Mündungsstelle ein blindes Ende, der Blinddarm(Caecum) abgegrenzt wird (s.Abb. 12.5–1).
Der Wurmfortsatz des Caecums, die Appendix ver-miformis (der Blinddarm des Laien), geht von der zurKörpermitte gerichteten Seite des Caecums ab. SeineLänge variiert zwischen 1 und 20 cm, sein Durch-messer zwischen 0,5 und 1 cm.
Am Kolon, dem Hauptteil des Dickdarms (s.Abb.12.5–2), unterscheidet man einen aufsteigenden,einen querverlaufenden, einen absteigenden undeinen S-förmigen Abschnitt (Colon ascendens, trans-versum, descendens, sigmoideum). Das Kolon hateine Länge von insgesamt etwa 1,2 m und eine lichte
401A N A T O M I E D E S D I C K D A R M S
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
!In Kürze !� Der Dünndarm gliedert sich in drei Abschnitte,
Duodenum, Jejunum und Ileum. Dieser längsteAbschnitt des GIT dient vor allem der Verdauungder Nahrungsbestandteile und der Absorptionkleinmolekularer Spaltprodukte.
� Die Absorption der Nahrungsbestandteile findet inden Zotten statt, die Sekretion von Darmsaft in denKrypten.
� Die Muskulatur der Darmwand entspricht in An-ordnung und Verlauf dem allgemeinen Wand-aufbau des GIT (s.o.). Die innere Ringmuskel-schicht ist wesentlich stärker als die äußere Längs-muskelschicht.
� Im Dünndarm erfolgt eine Durchmischung desInhalts mit den Verdauungssekreten durch rhyth-mische Segmentationen, Pendelbewegungen undstempelartige Zottenkontraktionen.
� Propulsive Peristaltik verlagert den Darminhaltüber größere Strecken in aboraler Richtung.
� Gesteuert werden die motorischen Aktivitäten vor
allem durch die Schrittmacher-Automatie und dasenterische Nervensystem (ENS). Parasympathikusund Sympathikus greifen lediglich modifizierendein.
� Die Dünndarmschleimhaut produziert täglich 2,5–3 l Sekret, dessen Abgabe durch lokale (intra-murale) Reflexe über Efferenzen des ENS zu denDrüsenzellen aktiviert wird. An der Sekretions-steuerung sind gastrointestinale Hormone, Neuro-transmitter und das vegetative Nervensystem be-teiligt.
� Die von den Becherzellen und Brunner-Drüsengebildeten Muzine haben vor allem Schutzfunk-tionen.
� Kryptenzellen und Brunner-Drüsen im Duode-num produzieren ein Bicarbonat-reiches, alkali-sches Sekret.
� Hauptzellen der Krypten sezernieren eine isotoneNaCl-Lösung.
Dickdarm
Ileum
Valvaileocaecalis
Wurmfortsatz(Appendix vermiformis)
Blinddarm(Caecum)
Abb. 12.5–1. Schnitt durch die Einmündung des Dünn-darms in den Dickdarm mit Blinddarm und Wurmfortsatz
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 401

Weite von 6–8 cm. Das besondere Charakteristikumdieses Darmabschnitts sind die Taenien und Haustren.Die drei Taenien stellen oberflächlich gelegeneStreifen der äußeren gebündelten Längsmuskulaturdar. Der Tonus der Taenien und lokale Kontraktionender Ringmuskulatur (Segmentationen, s.u.) lassenEinschnürungen entstehen, die als Falten (Plicae se-milunares) ins Darmlumen vorspringen. Zwischendiesen treten als halbkugelförmige Ausbuchtungen,die Haustren, hervor. Weiterhin trägt das Kolon cha-rakteristische zipfelförmige Fettanhängsel, die sog.Appendices epiploicae, deren Ausprägung vom Er-nährungszustand abhängig ist.
Das 15–20 cm lange Rektum beginnt dort, wo dasKolon sein Mesenterium verliert. In diesem letztenDarmabschnitt findet man wieder eine geschlosseneLängsmuskelschicht. Am Anus bildet die Ringmus-kulatur einen inneren Schließmuskel (Sphinkter) ausglatten Muskelfasern, der von einem äußeren Schließ-muskel aus quer gestreifter Muskulatur des Becken-bodens bedeckt ist. Unter der Schleimhaut befindetsich in der sog. Hämorrhoidalzone ein Geflecht vonarteriellen Blutgefäßen, die den muskulären Ver-schluss verstärken (s.S. 284). Oberhalb der beidenSchließmuskeln weist das Rektum eine Erweiterung(Ampulla recti) auf.
Aufbau der Dickdarmschleimhaut. Die Schleimhautdes gesamten Dickdarms ist zottenlos. Die Kryptender Dickdarmschleimhaut sind besonders tief und ste-hen dicht nebeneinander. Das Epithel der Krypten undder Oberfläche besteht vorwiegend aus Becherzellen,die Schleim produzieren. Ein Teil der oberflächlichen
Zellen ist mit einem Bürstensaum versehen und dientder Absorption.
Die Schleimhaut des Wurmfortsatzes zeigt den glei-chen Aufbau wie die des Dickdarms, jedoch sind ihreKrypten kleiner. Der Wurmfortsatz ist beim Men-schen Teil des Immunsystems (MALT, s.S. 171), undweist eine Vielzahl von Lymphfollikeln auf, die in dieSchleimhaut eingebettet sind.
12.5.2 Kolonmotilität
Die Hauptkomponenten der Kolonmotilität sindnichtpropulsive Segmentationen und werden vorwie-gend durch Schrittmacher-Automatie gesteuert. Hie-raus ergeben sich relativ lange Passagezeiten. Je nachNahrungszusammensetzung oder psychischem Zu-stand beträgt die durchschnittliche Passagezeit beiErwachsenen etwa 20–30 h (mit Schwankungen zwi-schen 5 und 70 h, s.S. 377).
Im Gegensatz zum Dünndarm haben die Seg-mentationen ihre niedrigste Frequenz am Beginn desKolon (ca. 7/min) und erreichen ihr Maximum imdistalen Kolon (ca. 15/min). Die Schrittmacherzoneliegt demnach im Colon descendes, von dem aus Kon-traktionswellen der Ringmuskulatur sowohl rück-wärts (Antiperistaltik) als auch in aboraler Richtungverlaufen. Hierdurch wird der Darminhalt im Caecumund im Colon ascendens längere Zeit zurückgehalten(Reservoirfunktion) und eingedickt. Motilitätsstei-gernd wirken cholinerge parasympathische Efferen-zen, hemmend dagegen NANC-Neurone mit denTransmittern VIP, NO und ATP.
In den Haustren bleibt der Dickdarminhalt über einen län-geren Zeitraum liegen, wodurch eine ausreichende Absorp-tion von Elektrolyten, Wasser und kurzkettigen Fettsäurenaus bakteriellen Abbauvorgängen (s.S. 404) sowie ein bak-terieller Aufschluss nicht-absorbierbarer oder nicht absor-bierter Nahrungsbestandteile gewährleistet wird. Die Ring-muskelkontraktionen bleiben lange Zeit an derselben Stellebestehen, so dass sie dem Koloninhalt einen Widerstand ent-gegensetzen, der eine zu schnelle Passage in das Rektumverhindert.
Bei herabgesetzter segmentaler Kontraktion, d.h. beimFehlen des Widerstandes, läuft der flüssige Inhalt schnell insRektum und verursacht vegetativ-funktionelle Durchfälle (z.B. durch gesteigerten Sympathikustonus bei Angst, Furchtoder Stress).
Peristaltische Wellen sind im Kolon selten. Dafür tre-ten, insbesondere nach den Mahlzeiten, propulsiveMassenbewegungen auf, die für den Transport desDarminhalts vom Colon transversum bis in das Rekto-sigmoid verantwortlich sind.
Die Massenbewegungen beginnen mit dem Sis-tieren der Segmentationen und einer Taenien-Er-schlaffung. Anschließend startet die Kontraktions-
D I C K D A R M402
HaustraTaenia
Colontransversum
Colondescendens
Colonsigmoideum
Anus
RektumAppendixvermiformis
Blinddarm(Caecum)
Colonascendens
Ileum
Abb. 12.5–2. Kolon und Rektum in Vorderansicht
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 402
welle proximal und setzt sich analwärts fort, wobeibeträchtliche Stuhlmengen durch die aboral relaxier-ten Abschnitte verschoben werden. Solche Bewegun-gen treten durchschnittlich 3–4 mal täglich auf undkönnen mit Stuhldrang und ggf. nachfolgender Stuhl-entleerung verbunden sein.
Die propulsiven Massenbewegungen stehen wahrscheinlichunter der Kontrolle des autonomen Nervensystems. Cho-linerge parasympathische Efferenzen des N. vagus bzw. ausdem Plexus sacralis sind vermutlich für das Auslösen derMassenbewegungen verantwortlich.
Im Gegensatz zum Dünndarm gibt es im Kolon keinenwandernden myoelektrischen Motorkomplex in der interdi-gestiven Phase (s.S. 383).
12.5.3 Darmkontinenz und Defäkation
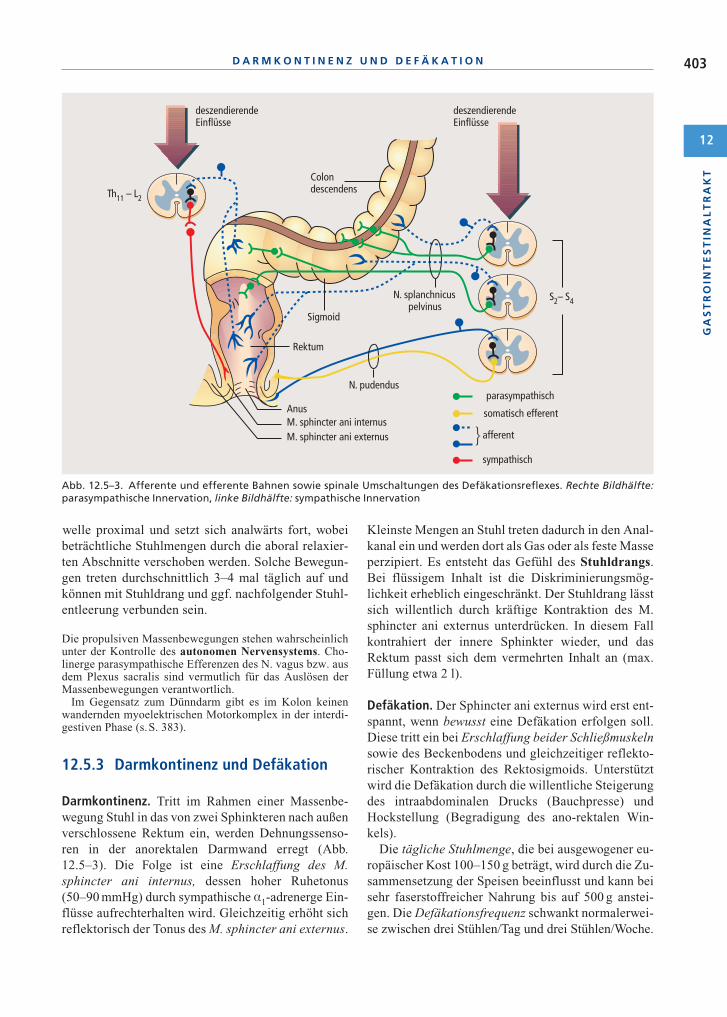
Darmkontinenz. Tritt im Rahmen einer Massenbe-wegung Stuhl in das von zwei Sphinkteren nach außenverschlossene Rektum ein, werden Dehnungssenso-ren in der anorektalen Darmwand erregt (Abb.12.5–3). Die Folge ist eine Erschlaffung des M.sphincter ani internus, dessen hoher Ruhetonus(50–90 mmHg) durch sympathische ·1-adrenerge Ein-flüsse aufrechterhalten wird. Gleichzeitig erhöht sichreflektorisch der Tonus des M. sphincter ani externus.
Kleinste Mengen an Stuhl treten dadurch in den Anal-kanal ein und werden dort als Gas oder als feste Masseperzipiert. Es entsteht das Gefühl des Stuhldrangs.Bei flüssigem Inhalt ist die Diskriminierungsmög-lichkeit erheblich eingeschränkt. Der Stuhldrang lässtsich willentlich durch kräftige Kontraktion des M.sphincter ani externus unterdrücken. In diesem Fallkontrahiert der innere Sphinkter wieder, und dasRektum passt sich dem vermehrten Inhalt an (max.Füllung etwa 2 l).
Defäkation. Der Sphincter ani externus wird erst ent-spannt, wenn bewusst eine Defäkation erfolgen soll.Diese tritt ein bei Erschlaffung beider Schließmuskelnsowie des Beckenbodens und gleichzeitiger reflekto-rischer Kontraktion des Rektosigmoids. Unterstütztwird die Defäkation durch die willentliche Steigerungdes intraabdominalen Drucks (Bauchpresse) undHockstellung (Begradigung des ano-rektalen Win-kels).
Die tägliche Stuhlmenge, die bei ausgewogener eu-ropäischer Kost 100–150 g beträgt, wird durch die Zu-sammensetzung der Speisen beeinflusst und kann beisehr faserstoffreicher Nahrung bis auf 500 g anstei-gen. Die Defäkationsfrequenz schwankt normalerwei-se zwischen drei Stühlen/Tag und drei Stühlen/Woche.
403D A R M K O N T I N E N Z U N D D E F Ä K A T I O N
GA
ST
RO
INT
ES
TIN
AL
TR
AK
T
12
Abb. 12.5–3. Afferente und efferente Bahnen sowie spinale Umschaltungen des Defäkationsreflexes. Rechte Bildhälfte:parasympathische Innervation, linke Bildhälfte: sympathische Innervation
parasympathisch
somatisch efferent
afferent
sympathisch
Sigmoid
Colondescendens
deszendierendeEinflüsse
N. splanchnicuspelvinus
deszendierendeEinflüsse
Rektum
N. pudendus
AnusM. sphincter ani internusM. sphincter ani externus
S2– S4
Th11 – L2
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 403

12.5.4 Sekretion und bakterielleBesiedlung des Dickdarms
Dickdarmsekretion. Die Kolonmukosa produziertnormalerweise nur kleinere Volumina einer plasma-isotonen, Muzin-, HCO3
–- und K+-reichen, alkali-schen Flüssigkeit. Sekretionssteigernd wirken Dihyd-roxygallensäuren, die aus dem Dünndarm in das Ko-lon gelangen, sowie VIP. Die HCO3
–-Sekretion in dasLumen erfolgt über einen HCO3
–/Cl–-Antiporter. Dasvon den Epithelzellen der Krypten sezernierte K+
wird bevorzugt auf parazellulärem Weg, teilweiseauch durch luminale K+-Kanäle, in das Dickdarmlu-men sezerniert. Die im Oberflächenepithel stattfin-dende Absorption übersteigt die Sekretion in denKrypten bei weitem.
Bakterielle Besiedlung des Dickdarms. Während derMagen und der obere Dünndarm keimarm sind,nimmt die Bakterienzahl nach distal hin zu. Sie steigtvon 106/ml Darminhalt im Ileum an der Bauhin-Klappe sprunghaft auf 1011–1012/ml im Kolon an.
Die Mehrzahl der Kolonbakterien sind obligate An-aerobier, in erster Linie Bacteroides (gramnegative,nichtsporenbildende Stäbchen). Die Anaerobier spal-ten unverdauliche Nahrungsbestandteile (z.B. Zellu-lose) teilweise auf, wobei u.a. kurzkettige Fettsäurenentstehen. Diese werden von der Kolonschleimhautabsorbiert und verwertet, wobei sie etwa 70% des lo-kalen Energiebedarfs decken. Aerobe Stämme wieEscherichia coli, Enterokokken und Lactobakterienmachen nur 1% der Kolonbakterien aus. Die über 400Bakterienarten im Kolon sind zu etwa 30–50%, bis-weilen sogar zu 75%, an der Stuhltrockenmasse betei-ligt.
L E B E R U N D G A L L E N W E G E404
!In Kürze
� Der etwa 120 cm lange Dickdarm besteht ausCaecum (mit Wurmfortsatz), Kolon und Rektum.
� Im Dickdarm wird der Darminhalt durchmischt,eingedickt und gespeichert.
� Die Schleimhaut des Dickdarms weist lediglichtiefe Krypten auf, Falten und Zotten fehlen.
� Die innere Ringmuskelschicht ist stärker ausge-bildet als die äußere Längsmuskelschicht, diein drei Bändern (Tanien) zusammengefasst ist.
� Die Hauptkomponenten der Kolonmotilität sindnicht-propulsiv. Hieraus ergeben sich relativlange Passagezeiten (20–30 h).
� Segmentationen beruhen auf der Schrittmacher-Automatie. Sie sind für die Durchmischung undindirekt auch für die Eindickung des Darminhaltsverantwortlich.
� Propulsive Massenbewegungen, die 3–4-maltäglich auftreten, verlagern den Darminhalt insRektosigmoid, wodurch Stuhldrang hervorgeru-fen werden kann, der dann zur willentlich aus-gelösten Stuhlentleerung (Defäkation) führt.
� Kontinenz, Stuhldrang und Defäkation werdenüber Aktivitäten des enterischen Nervensystems,vegetative und somatische Efferenzen gesteuert.
� Die Dickdarmmukosa produziert kleine Volu-mina eines Muzin-reichen, alkalischen Sekrets,das zur Neutralisation der bakteriell gebildetenSäuren dient.
� Das Kolon ist mit Bakterien, hauptsächlich An-aerobiern besiedelt, die unverdaute Faserstoffeaufspalten und u.a. kurzkettige Fettsäuren (s.o.),Vitamin K, Biotin, Schwefelwasserstoff, Methanund Wasserstoff produzieren (s. S. 428).
!
12.6 Leber und Gallenwege
Die Leber ist das zentrale Organ des Intermediär-stoffwechsels im Organismus (s.S. 15 ff.) sowie Spei-cher- und Syntheseort wichtiger Verbindungen (vgl.Tab. 12.6–1, s. S. 406). Weiterhin übernimmt siewichtige Funktionen bei der Metabolisierung (Bio-transformation) und Ausscheidung körpereigener undkörperfremder Substanzen. Im Hinblick auf die Bil-dung und Ausscheidung der Galle stellt die Leber diegrößte exokrine Drüse dar. Als Teilorgan des mono-nukleären Phagozytensystems (s.S. 44) nimmt sieweiterhin an den Abwehr- und Filterfunktionen des
Organismus teil. Vom Ende des zweiten Schwanger-schaftsmonats bis zur Geburt ist die Leber an der feta-len Blutbildung maßgeblich beteiligt (s.S. 137).
Im Rahmen dieser Darstellung soll nur auf die Ver-dauungsfunktion der Leber (Gallensekretion) einge-gangen werden. Bezüglich der weiteren Funktionenwird auf andere Kapitel dieses Lehrbuchs bzw. aufLehrbücher der Biochemie und Pharmakologie ver-wiesen.
12.6.1 Makroskopische Anatomie derLeber und der Gallenwege
Makroskopische Anatomie der Leber. Der größteTeil der beim Erwachsenen etwa 1,5 kg schweren
Kap 12 U_fe 30.03.2007 19:29 Uhr Seite 404













![Anhang-Bemessungshilfen (DIN 1052 neu)978-3-540-95899...Anhang-Bemessungshilfen (DIN 1052 neu) 395Tafel A.4. Beiwert ka,c a (BSH-Pultdachträger) a[ ] GL 24c GL 28c GL 32c GL 36c 3](https://img.pdfslide.org/doc/110x75/5f0544e97e708231d4121ff5/anhang-bemessungshilfen-din-1052-neu-978-3-540-95899-anhang-bemessungshilfen.jpg)












![Curriculum GSSM GL 10[1]](https://img.pdfslide.org/doc/110x75/616f6fcfb93cef78486fe63b/curriculum-gssm-gl-101.jpg)


