Embed Size (px)
Citation preview

138 W. W ~ S ~ D T : Aktiver Transport durch Grenzfli~ehen Klinisehe Wochenschrift
besser, diesen Ausdruek fallen zu lassen und f/it den solvent drag allein die unmi t t e lba r gemessenen GrSgen anzngeben.
F a g t m a n zusammen, so ergibt sieh, dab sieh eine Ffille versehiedenart iger Beobaehtungen zwanglos und rela t iv einfaeh dureh die Annahme sehr enger, wasser- gefiillter Kan/ile, welehe die Membran durehziehen, erkl/tren lessen. Es mug abet offenbleiben, welehe Gestal t a n d welehe Abmessungen sie besitzen, ob sic alle gleiehartig sind odor untersehiedliehe Dimensionen aufweisen, a n d sehlieglieh, ob es sieh um permanente S t ruk tn ren odor Gebilde des Zufalls handel t .
Literatur. ~ As"Dn~s~, B., and tL H. USSL~'e: Aeta phy- siol. seand 39, 228 (1957). - - ~ ] 3 ~ , D. F., A. L. HoD~x~" and I. I. S~Aw: Nature (Land.) 190, 885 (1961). - - e~ DE G~oo~: The Thermodynamics of Irreversible Processes. Amsterdam 1951. - - ~ DAvso~, H., and J. F. I)ANIELLI: The permeability of natural membr~ns. Oxford 1943. - - ~ D~A- ~o~D, J.M.: J. Physiol. (Land.) 161, 503 (1962). - - s ED~- ~ ] ~ , R. : J. cell camp. Physiol. 41, 37 (1953). - - ~ GA]~DOS, G., u. F. B. SW~AU~ : Aeta physiol. Acad. Sci. hung. 12, 1 (1956).- 7 G ~ 5 ~ o ~ , E. : ])as Permeabilit/~tsproblem. Berlin 1929. - - s G~s~5, 0., u. It. PASSOW: Pfltigers Arch. ges. Physiol. 271, 378 (1960). - - ~ G ~ t s e ~ , G. : In Nierensymposion, GSttingen 1959, S. 49. Stuttgart 1960. --~0 G ~ , A., and K. Wm~z: Z. Naturforseh. 8a, 532 (1953). - - n GLASSTO~, S., K. J. LAIDLER and H. EYE~N(~: The Theory of Rate Processes. New York and London 19.4.1. ---- ~ HAYS, R. M., and A. LEA~: J. gen. Physiol. 45, 905 (1962). ---~ HAYs, R. M., and A. L~A~: J. gen. Physiol. 45, 933 (1962). - - ~ H~r~nve , H. G. : J. gen. Physiol. 44, 365 (1960). - - ~ HoD~x~, A.L., and R. D. K E ~ s : J. Physiol. (Land.) 1~8, 61 (1955). - - ~s H O ~ A ~ , J .F . : J. gen. Physiol. 4~, 9 (1959). - - ~s JAeo~s, M.H., H.N. GLASS~A~ and A. K. PA~]?A~: J. cell. camp. Physiol. 8, 403 (1936). - - ~ KATe~ALSK¥, A.: In Membrane Trans- port and Metabolism, p. 69. New York and London 1961. - - ~s K~D~5~, O.: In Membrane Transport and Metabolism,
p. 87. New York and London 1961. - - 19 K~D~, O., and A. KATCHALSKY: J. gem Physiol. 45, 143 ( 1 9 6 1 ) . - 20 t~LEINZELLER, shx,, and A. KoTYx: Membrane Trans- port and Metabolism. Proceedings of a symposion held in Prague 1960. London and New York 1961. - - 2t KO]~FO~D- JOENS~N, V., and H. H. UssINo: Aeta physiol, scand 28, 60 (1953). - - 22 LEAF, A., and R. M. HAYS: J. gen. Physiol. 45, 921 (1962) (drag). - - 2a LINDE~IANN, B., and A. K. SOLOMON: J. gem Physiol. 45, 821 (t962). - - 2t MOND, g., u. F. HOFF- mANN: Pfliigers Arch. ges. Physiol. -~19, 467 (1928). - - 2~ Mt~L- L~R, P., D. O. RUDIN, Ill. TI~IEN and W. C. WEseoT~': Nature (Land.) 191, 979 (1962). - - es PAGA~LL~, C.V., and A. K. SaLesmaN: J. gen. Physio141, 259 (1957). - - ~ PAP~E~'HE[~E~, J. R., E. M. l~m~rw and L. M. BOaREaO: Amer. J. Physiol. 167, 13 (1951). - - ~s P~ESeOT% D. M., and E. Z ~ T H ~ : Aeta physiol, stand. 28, 77 (I953). - - ~ R ~ ' K ~ , E. M., and J. t~. P A P f ~ : Ergebn. Physiol. 49, 59 (1957). - - s~ Ron- m~s, E., and A. MAU~o: J. gen. Physiol. 43, 523 (1960). - - s~ R,o~a~so~, J .D. : Progr. Biophys. 10, 343 (1960). - - ~" S~AV~m~A~, A. J.: Trans. Faraday Sac. 48, 176 (1952). as Se~JLTZ, S. G., and A.K. SoLomon: J. gen. Physiol. 44, 1189 (1960). - - ~a SID]~L, V. W., and A. K. SOLOMON': J. gen. Physiol. 41, 243 (1957). a.~ SoLo~ox, A. K. : In Membrane Transport and Metabolism, p 94. New York and London 1 9 6 1 . - a5 STO~eKE~:r'eS, W.: J. biophys, bioehem. Cytol. 5, 49I (1959). - - ~ S~rRAU~, F. B.: Acta physiol. Aead. Sci. hung. 4, 235 (1953). - - av TI~osc~, A. S. : In Membrane Transport and Metabolism, p. 45. New York and London 1961. - - as TRosem:x, A. S. : Des Problem der Zellpermea- bilit~t. Jena 1959. - - ~ VII~L~GAS, R., T.C. BARTO~ and A. K. SOLO.~oz¢: J, gen. Physiol. 42, 2, 355 (1958). - - a0 V~LL~- OAS, R., and G. M, V~LL~eAS" J. gen. Physiol. 43, Suppl. 73 (1960). - - ~ VILLEGAS, R., and F.V. BA~O~A: J. gen. Physiol. 44, 963 (1961). - - ~e WA~TIOVAA~A, V., u. R. COLLANDEI~: Permeabilit~tstheorien, in Protoplasmatologia, Handbuch der Protoplasmaforschung. Wien 1960. - - ~a W~L- ~A~DT, W. : Ergebn. Physiol. 40, 20¢ (1938). - - Symposion on Membrane Transport and Metabolism, Prague 1960. New York 1961. - - ~ WIS~D~A~, E. E., G. W~Z~TTES~J~¥, D.E. OK~-~-, I-I. J. SC~aTZ~A~ and A. K. SOLOS~O~: Amer. J. Physiol 197, 112I (1959). - - ~ W ~ D ~ A ~ . , E.E. , and G. G ~ s e ~ : Nature (Land.) 191, 1205 (1961).
Aktiver Transport dureh GrenzNiehen ~:~on
W. W'ILBI~ANDT, Bern
Unsere Kenntn i s se fiber die Bewegung gelSster Teilehen dutch biologisehe Grenzflgchen haben in den letzten Jah rzehn ten starke W a n d l u n g e n durehge- maeht . Es ha t sieh einmal gezeigt, dug epitheliale Zellsehichten in viol zahlreieheren F~llen als frfiher angenommen die Fghigkei t besitzen, gelSste Stoffe dutch besondere Mechanismen van einer Epithelsei te auf die andere zu bewegen a n d dug diese Bewegungen vielfaeh bergauf erfolgen, d.h. van niedrigerer zu hShe- rer Konzent ra t ion , so dab die daffir beni i tz ten Ein- r ieh tungen Arbei t leisten u n d kurz als P u m p e n be- zeiehnet werden k6nnen. Welter ha t sieh er.wiesen, da6 solehe P u m p e n nieht n u t transeellulgre Transpor te dureh Zellen h indureh bewerkstelligen, sondern dug Vorr ieh tungen ~hnlieher Ar t aueh eine bedeutende Rolle spielen f/ir den Austauseh zwisehen Zelt innerem n n d U m g e b u n g dutch die Ze l lmembran hindureh. Bevor auf des Wesen dieser P u m p e n eingegangen wird, sei eine kurze Dars te l lung dieser En twiek lung voraus- gesehiekt.
A. Beispiele and biologische Bedeutung Die frfihesten Beobachtungen, die Pumpvorr ich-
t ungen in Zellschichten erkennen lieBen, waren Selcre- tionen versehiedener Art, Bag der Magensaft Wasser- ~toffionen in einer Konzen t r a t i on besitzt, die etwa eine
Million mal hSher ist als im Blur u n d den inters t i t ie l len Fliissigkeiten, ist seit langem bekannt . E in anderes Beispiel ist die Sekretion va n Sauerstoff in die Sehwimmblase der Fisehe, deren I n h a l t fast aus reinem Sanerstoff besteht.
E in Gegenstfick zu den Sekret ionen bi lden konzen- trierende Exlcretionen, wie etwa die Ausseheidung der Stoffweehsel-Endprodukte Hippurs£ure, I Iarnstoff u n d Harns/~ure, im Harn , in mehrfach h6herer Kon- zen t ra t ion als im Blur.
Neueren Da tums ist die Erkenntn i s , dab die Resorp- tionstransporte im Darm a n d die viele Paral lelen auf- weisende Riicl~resorption in den Nierentubuli ebenfatls bergauf erfolgen k a n n und sieh offenbar eelluI~rer P u m p e n bedient . Der wichtigste H~nweis ist aueh bier die Tatsaehe, dug die Bewegung entgegen dem Kon- zentrationsgef/ille erIolgen kann . I m Darm ist des ffir Zueker erstmals dureh die Versuehe va n BARA~TY a n d S P ~ E R ~ gezeigt warden, in der Niere ist es evident , seitdera der kombinier te Fi l t ra t ions- a n d Riiekresorp- t ionsmeehanismus sichergestellt ist : alle Substanzen, die im Glomerulus fi l tr iert werden, aber bei nor- malen Konzen t ra t ionen nieht im E n d h a r n erseheinen ( , ,Sehwellensubstanzen"), mfissen den Tubulus durch Riickresorption verlassen und zwar mi t zunehmender Verarmung des Tubulusharns gegen einen steigenden

Jg. 41, ~Ie~t 3 W . WILBI%ANDT : A k t i v e r Transpor~ d u r c h G r e n z f l a c h e n 139 1. Februar 1963
Konzentrationsgradienten. Zu ihnen geh6ren einmM wiedermn wiehtige Nahrungsstoffe wie Zueker und Aminosauren, die dem K6rper erhalten werden miissen. Daneben spielen sieh hier abet auch I~esorptions- prozesse ab, die regulatorisehen Zweeken dienen, in erster Linie der 08moregulation und der KonstanterhaI- tung des inneren Milieus, so die Rfickresorption von Natrium und die Sekretion yon KMium, die beiden Zwecken dient und bei Saugetieren den wichtigsten Anteil an der Osmoregulation besitzen.
Bei anderen Tierarten, insbesondere bei wasser- lebenden (sowohl bei SiiBwasserformen aIs bei Meeres- formen) treten neben (tie Niere als osmoregulatorisehe Organe eine Anzahl weiterer transportfi~higer Epithel- strukturen43, 4a . Sfigwassertiere ben6tigen Einrieh- tungen, um den fortwahrenden Salzverlust in die salz- arme Umgebung zu kompensieren. Bei ihnen linden sich Einrichtungen zum Transport yon Salzen aus den hoehverdiinnten AuBenlSsungen in den Organismus, bei Amphibien dureh die Haut 42, bei Fisehen und Krebsen dureh Kiemen. Meeresfische, die ihr Innen- medimn auf einem niedrigeren osmotisehen Druck- niveau halten als demjenigen des umgebenden Meeres- wassers (Knoehenfisehe), ben6tigen Pumpen, die in der umgekehrten Riehtung arbeiten. Aueh bier spielen wiederum die Kiemen in erster Linie eine Rolle, die in diesem Falle Salz von innen naeh augen transpor- tieren. Bei S~ugetieren und VSgeln, die im oder auf dem Meere leben und, aueh wenn sie kein Meerwasser trinken, doch mit der Nahrung hohe Salzmengen auf- nehmen, deren sie sieh entledigen mfissen, sind beson- dere Drfisen gefunden worden, wie die Buecaldriisen der Walfisehe ~ und die Satzdriisen der MeeresvSgeI s~, die hoehkonzentrierte SalzlSsungen zur Ausscheidung bringen. Eine Reihe dieser osmoregulatorisch wirk- samen Organe sind in den letzten Jahren zum Gegen- stand experimenteller Untersuchungen gemaehg wor- den, so in weitem Ausmag die Amphibienhaut ~, ss, ~o, ferner die Kiemen bei Fisehen ~° und Krebsen ~ und die Salzdriise der MeeresvSgeI ss.
Unerwarteter war die Feststellung, dab Pumpen- transporte nicht nut transcellul/~r arbeiten, sondern im Austauseh zwischen Zelle und Umgebung eine wiehtige golle spielen. Eiu erster Hinweis in dieser Riehtung war wohl der Befund yon VAN SLYX]~ und M]sY~R s~ aus dem Jahre 1913, dab die lebenden Zellen der Sguge- tierorganismen erheblieh h6here Ar~inos/~urenkonzen- trationen in ihrem Inneren besitzen als in der Um- gebung. Das Studium dieser Erseheinung, insbeson-
s~ dere an Tumorzellen hat gezeigt, dab die Membranen Aminosgurenpumpen besitzen, die diesen Konzentra- tionsuntersehied aufrechterhalten. Es seheint, dab solehe Einriehtungen vor allem notwendig sind f f r die Bausteine der wichtffgen hoehmolekularen Bestandteile der Zelle, in ersger LiNe der EiweiBe und Nuclein- s~uren. Aueh f/Jr die Basen der Nueleinsiiuren sind /~hnliehe Konzentrierungsmechanismen in letzter Zeit gefunden worden ~. Dagegen bilden sie f/Jr Zuelcer- moleki~le, die in erster Linie Gegenstand des katabolen Stoffweehsels sind, eher die Ausnahme als die Regel. Zwar treten Zuckermo]ek/ile dutch Zellmembranen im allgemeinen sicher nieht auf dem Wege der einfachen Diffusion hindurch, sondern mit Hilie yon Prozessen, die heute im allgemeinen als Tri~germechanismen ge- deutet werden, zumeist besitzen sie aber keinen Pum- pencharakter. Das Wesen dieses Mechanismus besteht
darin, dab das Transportsubstrat sieh auf der einen Seite der Membran mit einem Trggermolekiil zu einem Komplex binder, der in irgendeiner Weise, z.B. dureh Diffusion imstande ist, die lVlembran zu durehqueren und auf der anderen Seite unter Freigabe des Snbstrats in die w/igrige L6sung wieder gespalten wird (s. Abb. 1).
Bei Zellen mit besonders hoher Stoffweehselinten- sitgt gibt es abet aueh flit Zuekermolekfile Membran- pumpen. Es sind die bei den Bakterien in den letzten Jahren 12, 36, 37 besehriebenen Systeme, die als Per- measen bezeiehnet worden sind, ein Ausdruek, der vor allem deshatb angreifbar erseheint, weil er die sicher irrefiihrende Vorstelhng dem Permeasesystem um eine einzelne, wohl- definierte ~,Iolek/itart und sie gehSre, worauf die Endung -ase hin- deutet, znr Gruppe der Enzyme.
Eine in der Zetl- membran gelegene Pum- penart,, deren biologL sehe Bedeutung auf den ersten Bliek weniger evident erseheinen mag,
erweckt, es handle sieh bei
Hembfan ,.rA
mSglieherweise aber ~ l mannigfaltiger ist als die S l~'\-, ,CS C/~'/I. der bisher genannten,------J " ~ \ / . / f / I H ist die Gruppe der Ionen- I ~ / ~ \ I
• . I / , \ S pumpen. Die unglelche I I • \ I '
Verteflung yon Natrium ~ ' / \ \ ] und Kalium z~dschen abb. 1. Schema eines Tragersystems, Ze]linnerem und Zell- umgebung ist, wie man heute aus den Erfahrun- gen mit Isotopen mit Sicherheit weiB, weder auf allgemeine Undurch- lgssigkeit der Membran fiir diese Ionen noch,
das nur Konzentrationsausgleich be- wirkt, aber nicht bergauf transpor- tiert. Das Subsirat S reagiert mi t dem Trigger C, forint den Komplex SC, der die Membran passiert und aui der anderen SeRe S freigibt. Die II.eak- tionen k6nnen d~lrch Enzyme kata- lysiert werden. In der :g~embran sind im stat ionaren Zustand zwei gegen-
t/iufige Gradienten fiir CS und C
entsprechend dem Vorsehlag yon CO,WAr 7, auf se]ek- t i re Undurchl/~ssigkeit f/Jr Natrium zurfickzuftihren, da die Isotopen dieser Ionen sich mit relativ hoher Ge- sehwindigkeit durch die Membran bewegen. Es ist daher ers~ma]s yon D~A~ - 19412° vorgeschlagen wet- den, daI} der Konzentrationsausgleich f/Jr diese Ionen verhindert wird durch die kontinuierliche Tgtigkeit von Pumpen, die in der entgegengesetzten Richtung arbeJten und die fortlaufend Natrium aus den Zellen heraus und Kalium in die Zellen hinein bef6rdern. Die gegenlgufigen Versehiebungen yon Natrium und Kalinm (s. Abb. 2) siud, mindestens an mehreren Zell- arten, mi~einander gekoppelt (s. Abb. 3), so dal~ man yon einer , ,Natrium-Kalium-Austauschpumpe" spre- ehen kann. Auch flit Calcium s4 und Magnesium ~7 ist die Annahme entsprechender Vorrichtungen notwendig geworden.
Die biologische Bedeutung der Ionenpumpen ist darin gesehen worden, dab gewisse intraeellulare En- zyme anf hoheKaliumkonzent, rM, ion angewiesen sind2S. 5~indestens ebenso wiehtig ist aber zweifellos die Tat- saehe, dab die Zelle sich dieser Einriehtungen ffir ver- schiedene biologische Zwecke bedient. Bekannt ist heute vor allem durch die gl/~nzenden Untersuchungen der Cambridger Gruppe yon A. L. t{ODGXI~ n. Mitarb.,
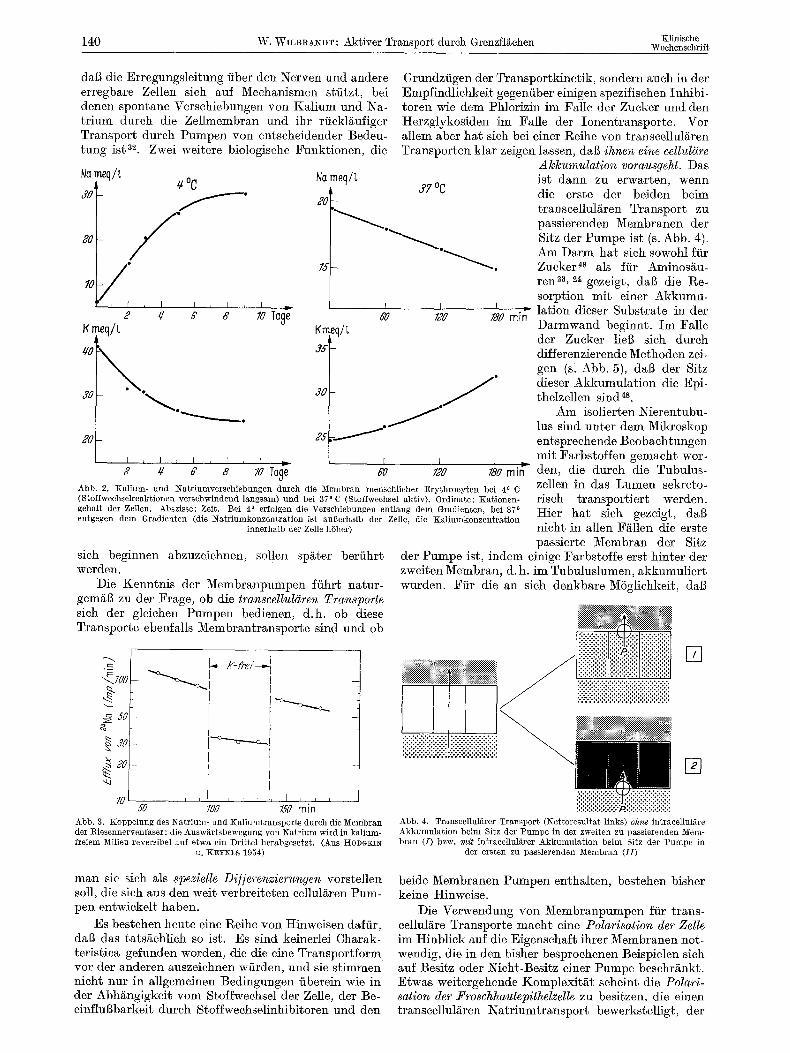
140 W. WILBRANDT: Aktiver Transporg durch Grenzflgehen Xlinische Woehensehrif~
dal3 die Erregungsleitung fiber den Nerven und andere erregbare Zellen sieh auf Meehanismen stfitzt, bei denen spontane Versehiebungen yon Kalium und Na- t r ium dureh die Zellmembran und ihr riickl/~ufiger Transport dureh Pumpen yon entseheidender Bedeu- tung ist ~. Zwei weitere biologisehe Funktionen, die
Na meq/~ °C Na meq/L sol g
r I , I , I , I ,
2 ¢ B 8 meq/t
X,X. 20-
I Io Ta~
Grundzfigen der Transportkinetik, sondern auch in der Empfindhehkeit gegenfiber einigen spezifischen Inhibi- toren wie dem Phlorizin im Falle der Zueker und den Herzglykosiden im Falle der Ionentransporte. Vor allem aber hat sich bei einer t~eihe yon transcellul/~ren Transporten Mar zeigen lassen, dag ihnen eine cellul~ire
Akkumulation vorausgeht. Das ist dann zu erwarten, wenn
' 37°C die erste der beiden beim , ~ transee]lu]/~ren Transport zu
• passierenden N{embranen der Sitz der Pumpe ist (s. Abb. 4).
" ~ Am Darm hat sieh sowohl ffir 75 • Zucker as als ffir Aminos£u-
renan, 24 gezeigt, dag die Re- sorption mit einer Akkumu-
I i I " lation dieser Substrate in der Kmec it ~0 120 180 rain Darmwand beginnt. I m Falle
der Zueker lieB sieh dureh 35 differenzierende Methoden zei-
gen (s; Abb. 5), da$ der Sibz
J " dieser Akkumulation die Epi- 30 thelzellen sind 4s.
J " Am isolierten Nierentubu- i ~ . lus sind unter dem Mikroskop
25 t entsprechende Beobaehtungen mit Farbstoffen gemaeht wor- den, die dutch die Tubulus- ze]len in das Lumen sekreto- riseh transportiert werden. Hier hat sieh gezeigt, dab nieht in allen t~/~llen die erste passierte Membran der Sitz
der Pumpe ist, indem einige Farbstoffe erst hinter der zweiten Membran, d.h. im Tubuluslumen, akkumuliert wurden. F/Jr die an sieh denkbare M6glichkeit, dab
2 4z 8 8 so T~ ~ Iz Iso rain
Abb. 2. Kalium- und Natrlumverschiebungen dutch die Membran menschlicher Erythrocyten bei 4 o 0 (Stoffwechselreaktionen verschwindend langsam) und bei 37 o C (Stoffwechsel aktiv). Ordinate: Kationen- gehalt der Zellen. Abszisse: Zeit. Bei 40 erfolgen die Verschiebungen entlang dem Gradienten, bei 370 entgegen dem Gradienten (die Natriumkonzentration ist augerhalb der Zelle, die Kalinmkonzentration
innerhalb der Zelle hSher)
sieh beginnen abzuzeiehnen, sollen sp/~ter berfihrt werden.
Die Kenntnis der Membranpumpen ffihrt natur- gem£$ zu der Frage, ob die transcellul@en Transporte sieh der gleichen Pumpen bedienen, d.h. ob diese Transporte ebenfalls Membrantransporte sind und ob
,=_
~. TO0
/0
[-~- /(-fre/-~
I $
I - I
I I
I . . . . . . I, I , , 50 700 750 m in
Abb. 3, Koppelung des Natrium- und ]~aliumtransports dutch die Membran der Riesennervenfaser: die Ausw~rtsbewegung yon Natrium wird in kalium- freiem Milieu reversibel auf etwa ein Drittel herabgesetzt. (Aus HODGKIN
u. J~EYNI~S 1954)
Abb. 4. Transcellul/~rer Transport (Nettoresultat links) ohne intraeellul/ire Akkumulation beim Sitz der Pumpe in der zweiten zu passierenden Mem- bran (I) bzw. mit lntracellul~rer Akkumulation beim Sitz der Pumpe in
der ersten zu passierenden Membran ( / / )
man sie sich als spezielle Di/]erenzierungen vorstellen soll, die sich aus den welt verbreiteten cellulgren Pure- pen entwickelt haben.
Es bestehen heute eine Reihe yon tIinweisen daf/ir, dag das tatsgchlich so ist. Es sind keinerlei Charak- teristiea gefunden worden, die die eine Transportform vor der anderen auszeichnen w/irden, und sie st immen nieht nur in Mlgemeinen Bedingungen iiberein wie in der Abh/~ngigkeit yore Stoflwechsel der Zelle, der Be- einflugbarkeit durch Stoffwechselinhibitoren und den
beide Membranen Pumpen enthalten, bestehen bisher keine Hinweise.
Die Verwendung yon Membranpumpen ffir trans- eellul/~re Transporte maeht eine Polarisation der Zelle im ttinbliek auf die Eigensehaft ihrer Membranen not- wendig, die in den bisher besproehenen Beispielen sieh anf Besitz oder Nicht-Besitz einer Pumpe besehrgnkt. Etwas weitergehende Komplexi tgt seheint die Polari- sation der Froschhautepithelzelle zu besitzen, die einen transcellul~ren Natr iumtransport bewerkstelligt, der
~g. tl, tte~t 3 W. W]:LB~_~NI)T : Aktiver Transport dutch Grenzfl~chen 141 1. Febraar 1963
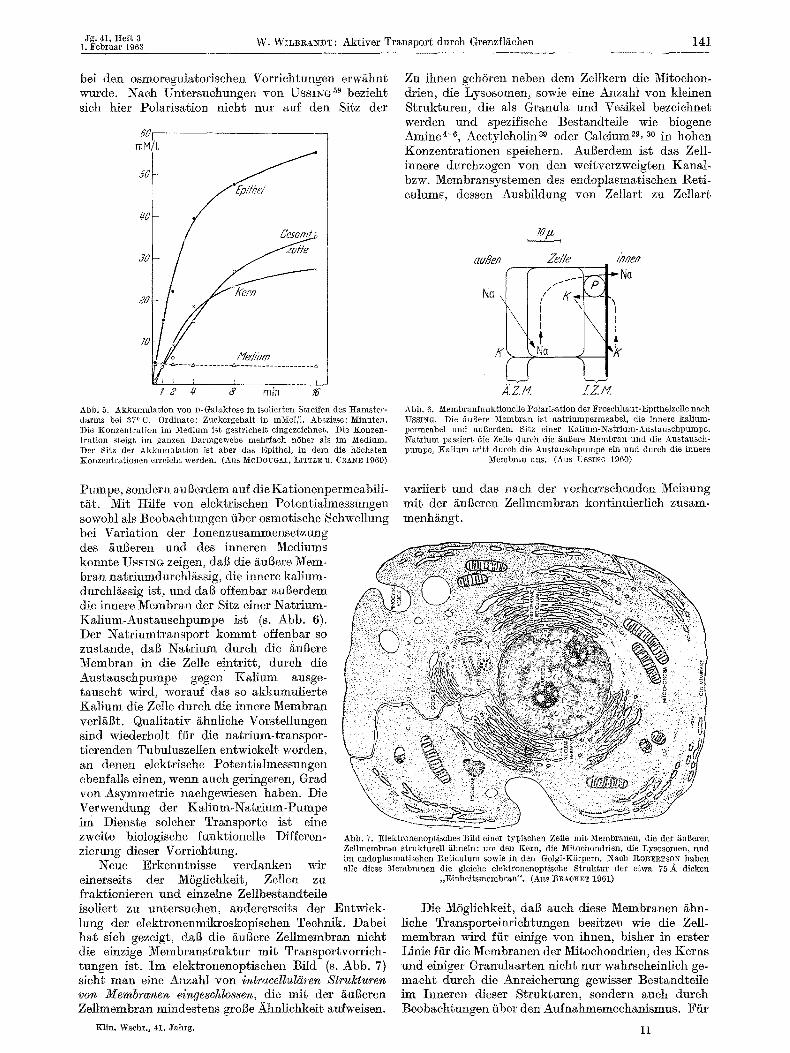
bei den osmoregulatorisehen Vorrichtungen erwghnt wurde. Naeh Untersuehungen yon U s s n ~ so bezieht sieh hier Polarisation nieht nut auf den Sitz der
mM/L
"~i/~c/
~ Med/~m
I 2 4" 6' rain 7~'
Abb. 5. Akkumula~ion yon D-Galaktose in isolierten Streifen des Hamster- darms bei 370 C. Ordinate: Zuekergehalt in mNol/1. Absziase: Minuten. Die Konzentration im Medium ist gestriehel~ eingezeiehneL Die Konzen- gration steigt im ganzen narmgewebe mehrfaeh h6her als im Medium. Der Sitz der Akkumulation ist abet das Epithel, in dem die h6ehsten Konzentrationen erreieht werden. (Aas MCDOUG:t~, LITTLE n. CRANE 1960)
Zu ihnen geh6ren neben dem Zellkern die 5fitoehon- drien, die Lysosomen, sowie eine Anzahl yon kleinen Strukturen, die als Granula und Vesikel bezeichnet werden und spezifisehe Bestandteile wie biogene Amine ~-6, Aeetyleholin a~ oder Calcium es, s0 in hohen Konzentrationen speiehern. Augerdem ist das Zell- innere durehzogen yon den weitverzweigten Kanal- bzw. Membransystemen des endoplasmatisehen get i- eulums, dessen Ausbildung yon Zellart zu Zellart
). i
auBez Zelle /nnen
Na / K -xl
I
A:ZM /~ZM Abb. 6. ~Iembranfunk~ione]le Polal%ation der Frosehhaat-Epithelzelle na.eh IJSSING. Die auBere 3iembran ist natriumpermeabel, die innere kalimn- permeabel und augerdem S:itz einer Kalinm-Natrium-Austausehpumpe. Natrium passiert die Zelle dnreh die ~ugere )Iembran und die Austauseh- pumpe, Kalium tritt dureh die Austausehpumpe ein und dureh die innere
Membran aus. (Aus lLssIi~ 1960)
Pumpe, sondern aul3erdem anf die Kationenpermeabili- t~t. Mit tIilfe yon elektrischen Potentialmessungen sowohl als Beobaehtungen fiber osmotisehe Sehwellung bei Variation der Ionenzusammensetzung des /~ugeren und des inneren Mediums konnte UssI~G zeigen, dab die ~uBere Mere- bran nat, rinmdtgehtg.ssig, die innere kalium- durehl/~ssig ist, und dab offenbar augerdem die innere Membran der Sigz einer Natrium- Kalium-Austausehpumpe ist (s. Abb. 6). Der Natriumtransport kommt offenbar so zustande, dab Natrium durch die £ul3ere Membran in die Zelle Nntritt, dutch die Austausehpumpe gegen Kalium ausge- tauseht wird, worauf das so akkumulierte Kalium die Zelte dutch die inhere Membran verl/~gt. Qualitativ ghnliehe Vorstellungen sind wiederholt ftir die natrium-transpor- tierenden Tubuluszellen entwiekelt worden, an denen elektrische Potentialmessungen ebenfalls einen, wenn aueh geringeren, Grad yon Asymmetrie naehgewiesen haben. Die Verwendung der Kalium-Natrium-Pumpe im Dienste soleher Transporte ist elne zweite biologisehe funk£ionelle Differen. zierung dieser Vorriehtung.
Neue Erkenntnisse verdanken vdr einerseits der MSgliehkeit, Zelten zu fraktionieren und einzelne Zellbestandtefle isoliert zu untersuehen, andererseits der Ent~4ek- lung der elektronenmikroskopisehen Teehnik. Dabei hat sieh gezeigt, dab die /~ul3ere Zellmembran nicht die einzige Membranstruktur mit Transportvorrieh- tungen is~. Im elektronenoptischen Bild (s. Abb. 7) sieht~ man eine Anzahl yon intracelluliiren Strukturen yon Membranen eingeschlossen, die mit der gugeren Zellmembran mindestens groBe ~hnlichkeit aufweisen.
varfiert nnd das naeh der vorherrsehenden Meinung mit der /Lul3eren Zellmembran kontinuierlieh zusam- menh~ngt.
Abb. 7. Elektronenoptisches Bild einer ~ypisehen Zelle mit Membranen, die der itugeren Zellmembran strukturell ~lmeln: um den Kern, die Mitoehondrien, die Lysosomen, und im endoplasmatisehen t~eticulum sowie in den Golgi-KSrpern. Naeh I~0~RTSOg haben alle diese Membranen die gleiche elektronenoptisehe Struktm" der etwa 75 ~ dieken
,:Einheitsmembran". (Aus ]~AOI~ET 1961)
Die M6gtichkeit, dal3 auch diese Membranen/~hn- liehe Transporteinrichtungen besitzen wie die Zell- membran wird f fir einige yon ihnen, bisher in erster Linie ffir die Membranen der Mitochondrien, des Kerns und einiger Granulaarten nieht nut wahrseheinlieh ge- maeht dureh die Anreieherung gewisser Bestandteile im hmeren dieser Struktttren, sondern aueh dureh Beobaehtungen fiber den Aufnahmemeehanismus. Ffir
Klin. Wschr., 41. Jahrg. 11
142 W. WILBI~ANDT : Aktiver Transport dutch Grenzfli~chen Klinische Wochenschri_tt
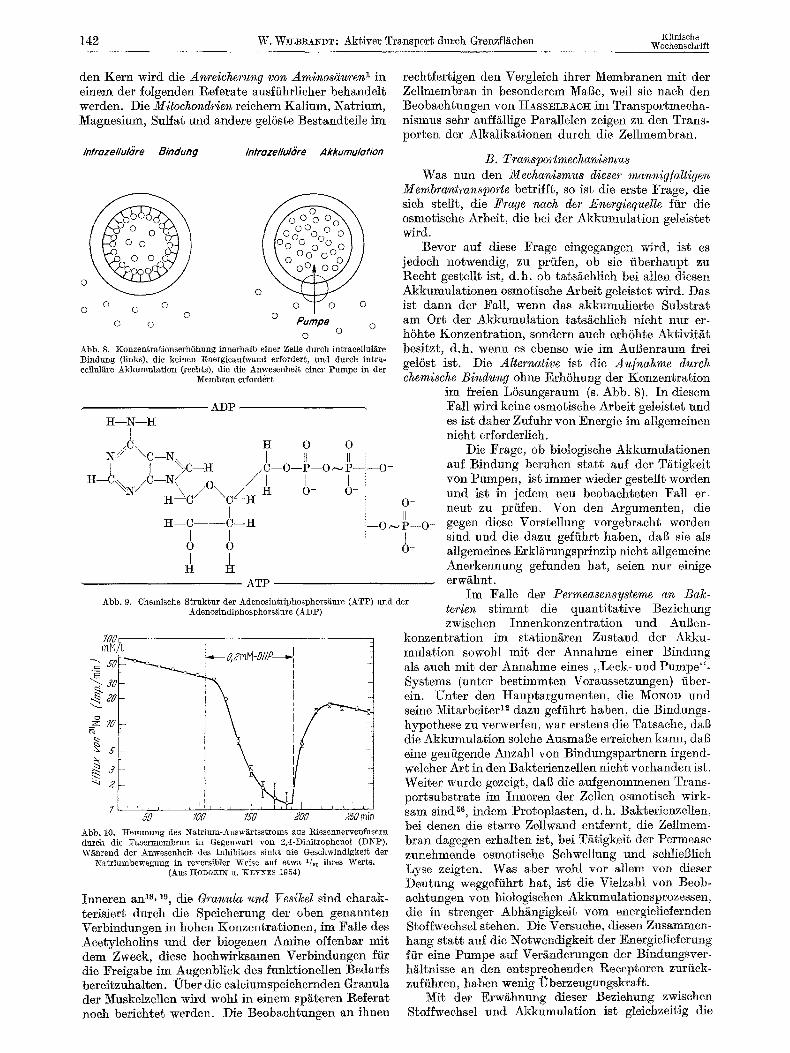
den Kern wird die Anreicherung von Aminosguren ~ in einem der folgenden Referate ausfiihrlieher behandelt werden. Die Mitochondrien reiehern Kalium, Natrium, Magnesium, Sulfat und andere gel6ste Bestandtefle im
Infrozellulore Bindung Intraze/lulb're Akkumu/otion
0 0 0 0 0 0 O
o o Pumpe o o o o o
Abb. 8. Xonzen%ra~ionserh6hung irmefl:alb einer ZelIe dttrch in~racellut~re Bindung (links), die keinen Energieaufwand e~ordert., und dutch in~ra- cellul~ire Akkumulation (rech~s), die die Jnwesenheit einer Pumpe in der
Nembran erfordert
ADP, H--N--H
1 /,C\ It 0 0
N .:/ "-C--N-- I1 II J I ~C--H .C--O--P--O ~ P--;:- -O-
H--C~._/C--~ .o, ./J { ! : ~-- H _ ~ c / \ c / ~ n o- o- :;
H--C----C--If
0 0
H I-I
ATP
Abb. 9. Chemische Struk~ur der Adenosin~rilohosphors~ure (ATP) and der Adenosindiphosphor s~ur e (ADP)
O -
N ~--o ~ P--O-
O-
700
s 1 I -
. . I . . . . , I , , i i ~ v T , I ~ ~ , i
:o 7oo /5o 200 25o rain Abb. 10. ]~emmung des Natrium-Auswg, rtssi~roms aus l~iesennerven£~sern dutch die Fasermembran in Gegenwaxt yon 2,4-Dinitrophenol (DNP). Wahrend der Anwesenheit des Inhibitors sinkt die Geschwindigkeit der
Natriumbewegung in reversibler Weise auf etwa ~!~s ihres Wefts. (Aus n:ODGKIN U. XEYNES 195~)
Inneren an is, 10, die Granuta und Vesilcet sind charak- terisiert durch die Speieherung der oben genannten Verbindungen in hohen Konzentrationen, im FMle des Acetylchotins and der biogenen Amine offenbar mit dem Zweck, diese hochwirksamen Verbindnngen fiir die Freigabe im Angenblick des funktionellen Bedarfs bereitztthalten, t~ber die calciumspeichernden Granula der Muskelzellen ~drd wohl in einem spBteren geferat noch berichtet werden. Die Beobachtungen an ihnen
reehtfertigen den Vergleieh ihrer Membranen mit der Zellmembran in besonderem Mage, weft sie nach den Beobaehtungen yon tIASS~LBAOH im Transportmeeha- nismus sehr auff~llige Parallelen zeigen zn den Trans- porten der Alkalikationen dutch die Zellmembran.
B. Transportmechan@mus Was nun den Mechanismus dieser mannig/altigen
Membrantransporte betrifft, so ist die erste Frage, die sich stellt, die Frage nach der Energiequelle ffir die osmotische Arbeit, die bei der Akkumulation geleistet ~drd.
Bevor auf diese Frage eingegangen wird, ist es jedoeh notwendig, zu priifen, ob sie iiberhaupt zu Reeht gestellt ist, d.h. ob tats~ehlieh bei allen diesen Akkumutationen osmotische Arbeit geleistet wird. Das ist dann der Fall, wenn das akkumulJerte Substrat am Ort der Akkumulation tats/iehIich nieht nnr er- hShte Konzentration, sondern aneh erhShte Aktivitgt besitzt, d.h. wenn es ebenso wie im AuBenraum frei gel6st ist. Die Alternative ist die Au/nahme dutch chemische Bindung ohr~e ErhShung der Konzentration
im freien LSsungsraum (s. Abb. 8). In diesem Fall wird keine osmotisehe Arbeit geleistet and es ist daher Zufuhr yon Energie im allgemeinen nieht erforderlieh.
Die Frage, ob biologisehe Akkumulationen auf Bindung beruhen start auf der Tgtigkeit yon Pumpen, ist immer wieder gestellt worden und ist in jedem neu beobaehteten Fall er- neut zu prfifen. Von den Argumenten, die gegen diese Vorstellung vorgebraeht worden sind and die dazu gefiihrt haben, dab sic Ms allgemeines ErklgrHngsprinzip nieht allgemeine Anerkennung gefunden hat, seien nur einige erwahnt.
Im Falle der Permeasensysteme an Bak- terien stimmt die quantitative Beziehung zwischen Innenkonzentration und Augen-
konzentration im station/iren Zustand der Akku- mulation sowohl mit der Annahme einer Bindung als aueh mit der Annahme eines ,,Leek-und Pumpe"- Systems (unter bestimmten Voraussetzungen) fiber- ein. Unter den Hauptargumenten, die Mo~oD and seine Mitarbeiter n dazu geffihrt haben, die Bindungs- hypothese zu verwerfen, war erstens die Tatsache, dab die Akkumulation solche Ausma.Be erreiehen kann, dag eine genfigende Anzahl yon Bindungspartnern irgend- weleher Art in den Bakterienzellen nieht vorhanden ist. Welter wurde gezeigt, dab die aufgenommenen Trans- portsubstrate im Inneren der Zelten osmotiseh wirk- sam sind 56, indem Protoplasten, d.h. Bakterienzelten, bei denen die starre Zellwand entfernt, die Zetlmem- bran dagegen erhalten ist, bei Tgtigkeit der Permease zunehmende osmotische Schwellung und sehlieBlieh Lyse zeigten. Was aber wohl vor allem yon dieser Dentung weggeffihrt hat, ist die Vielzahl yon Beob- achtungen yon biologisehen Akkumulationsprozessen, die in strenger Abh~ngigkeit yore energieliefernden Stoffweehsel stehen. Die Versuehe, diesen Zusammen- hang start auf die Notwendigkeit der Energielieferung far ekne Pumpe auf Ver£nderungen der Bindungsver- hgltnisse an den entspreehenden l~eceptoren zuriiek- zuffihren, haben wenig l~berzeugungskraf~.
Mit der Erw/~hnung dieser Beziehnng zwisehen Stoffwechsel und Akknmulation ist gleiehzeitig die
Jg. 41, Heft 3 W. WILBI~_~n~: Aktiver Transport durch Grenzfl~chen 143 5. Feb rua r 1963
Antwort auf die Frage nach der Ene~yiequelle (sofern sie nun als bereehtig~ anerkannt wh'd) gegeben: die Energiequelle ist chemische Energie aus dem Sto//- weehsel der Zelle. Zwar gibt es wahrseheinlieh eine Anzahl yon F/~llen, bei denen zwischen den akkumu- lierenden Transport und die chemisehe Energie noeh Zwisehenglieder eJngesehaltet sind, yon denen noeh die Rede sein wird, letztlich ist aber die Energiequelle aueh in diesen Fgllen chemisehe Stoffweehselenergie.
Allgemein ffihren alle Bedingungen, die die Stoff- wechselintensit~t vermindern, zur Verlangsamung bis Bloekiernng der Akkumulationsprozesse: niedrigeTem- peratur (s. Abb. 2), Sauerstoffmangel, Mangel an ener- gielieferndem Stoffweehselsubstrat, Anwesenheit yon Hemmstoffen ffir den oxydativen Stoffweehsel (Cya- nid, Acid) oder ffir den glykolytischen Stoffweehsel (Monojodessigsgure). Von besonderer Bedeutung ist die weitverbreitete Hemmung yon Inhibitoren, die die oxydative Phosphorylierung hemmen, d.h. die Bildung yon energiereiehen Phosphaten wie Adenosintriphos- phorsiiure (s. Abb. 9) aus der Energie des oxyda~iven Stoffwechsels. D~s meistverwendete Agens aus dieser Gruppe ist das 2,#Dinitrophenol. Die Tat.saehe, dab Dinitrophenol bei einer sehr groBen Zahl biologiseher Akkumulationen hemmend wirkt (ein Beispiel zeigt Abb. 10), sprieht daffir, dab auch hier wie bei anderen biologisehen Leis~ungen (Muskelkontraktion, chemi- sehe Synthese) das energiereiehe Phosphat die un- mittelbare Energieqnelle ist, die man in ansehaulieher Weise als die ,,bare Mfinze im Energieverkehr" be- zeichnet hat. Anch in F~llen, wo I)initrophenol nicht hemmend ~irkt , wie beim Kationentransport an roten BlutkSrperehen yon S/iugetieren, die ihre Energie vor- wiegend aus glykolytisehem Stoffweehsel beziehen, konnte die wichtige Bedeutung energiereichen Phos- phats erwiesen werden, indem y o n ~LECKENSTEIN n. Mitarb. ~5 gezeigt werden konnte, dab die t{emmwir- kung yon Monojodessigs~ure para?lel geht mit dem Versehwinden des energiereiehen Phosphats.
Zu den Beobaehtungen fiber die Hemmwirkung des Dinitrophenols gesellen sieh heute unmittelbarere Hin- weise auf die Bedeutung des energiereiehen Phosphats, vor allem aus Versuehen an l~iesennervenfasern, aber aueh an roten ]~lutk6rperehen und an den oben er- w£hnten eatciumspeiehernden Gralmla der Muskeln. An den Nervenfasern wurde yon der ttodgldnsehen Arbeitsgruppe in Cambridge gezeigt 9, dab naeh Bloekie- rung der Natriumpumpe dutch Cyanid der Natrium- transport wieder in Gang gebraeht werden kann (s. Abb. 11), wenn in das Innere der Faser bestimmte energiereiehe Phosphate injiziert werden, nnter ihnen Adenosintriphosphorsaure (anGerdem Argininphos- phorsgure und Phosphoenolbrenztraubens/inre). Die Znfuhr dieser energiereiehen Phosphatverbindungen wirkte reaktivierend nut im Inneren der Fasern, nieht dagegen an ihrer ~ul3eren Oberfigehe. Der dutch Di- nitrophenol oder Cyanid unterbrochene Prozefi kann also, wie diese Versuehe zeigen, tats/~chlich dutch die energie- reichen Verbindungen, deren Bildung dureh die Inhibi- toren verhindert wird, wieder in Gang gesetzt werden.
Xhnliche Untersuchungen lassen sieh mit spezieller Methodik am Erythroeyten durehffihren 2s, wobei wie- derum das ATP nur im Inneren der Zelle wirkt, nieht dagegen auBen. Der Eintr i t t in die Zelle wird hier durch die Straubsche Hamo]ysemethodik ermSglieht,
die auf der grogen Durehlgssigkeitszunahme der Mem- bran bei der H£molyse beruht.
Auf eine enge Verknfipfung zwisehen dem Stoff- weehsel des ATP und den Ionentransporten weisen aueh Beobachtungen aus einer anderen Riehtung, ngmlich aus Untersuehungen fiber das A TP-spaltende Enzym, die sog. ATPase in den Membranen einer Reihe yon Zellen.
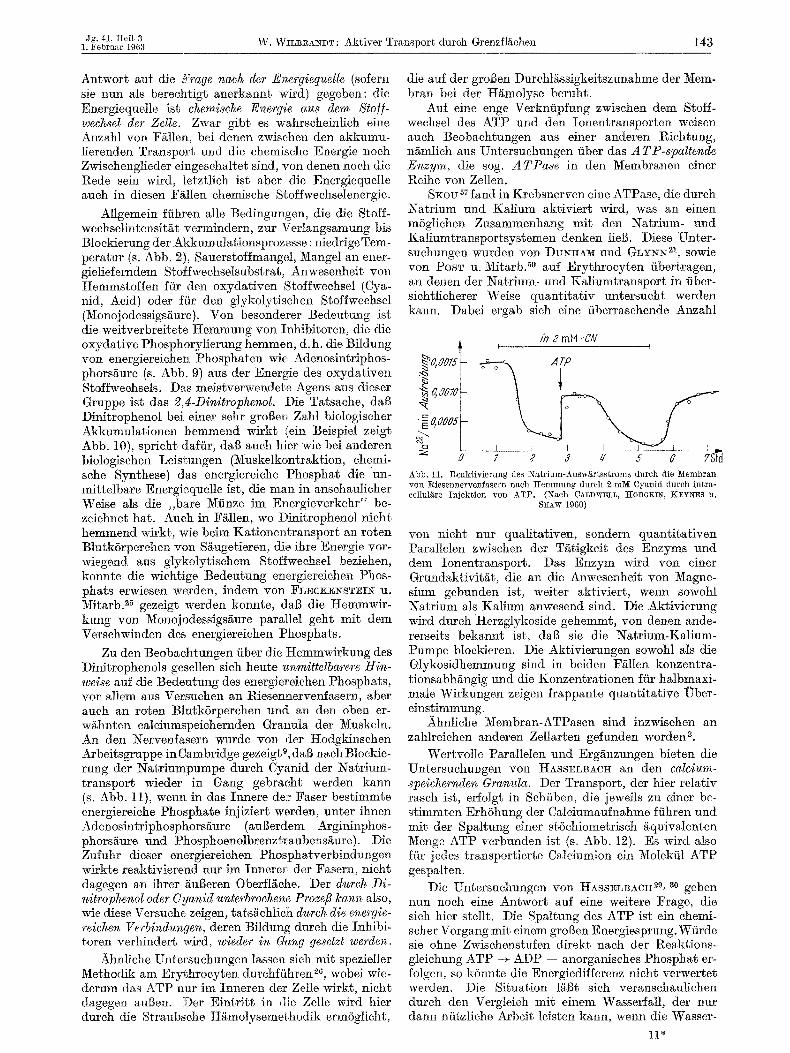
SKov 57 land in Krebsnerven eine ATPase, die dutch Natrium und Ka[ium aktiviert wird, was an einen mSglichen Zusammenhang mit den Natrium- und Kaliumtransportsystemen denken Iieg. Diese Unter- suehungen wurden yon DUNHASI and ~ L Y N N 21, sowie yon Posm u. M]tarb. a° anf Erythrocyten iibertragen, an denen der Natrium- und Kaliumtransport in iiber- siehtlieherer Weise quanti tat iv untersueht werden kann. Dabei ergab sieh eine fiberrasehende Anzahl
~O, OOZS~ Do, ool, . 40oo5
~zJ
0
/n 2 mM-C'N . . . . . . . . . [
7 2 3 z/ 6 G 7Sfd Abb. 11. I%eaktivierung des Natrium-Ausw~rtsstroms durch die Membran von Riesennervenfasern nach Jgemmung durch 2 mM Cyanid durch intra- cellul~re Injek~ion yon ATP. (Nach CALDWELL, lcJ[0DGKIN, KEYNES I1.
SHAW 1960)
yon nicht nur qualitativen, sondern quantitativen Parallelen zwischen der T/itigkeit des Enzyms uzad dera Ionentransport. Das Enzym ~ r d yon einer Grundaktivit/~t, die an die Anwesenheit yon Magne- sium gebunden ist, weiter aktiviert, wenn sowohl Natrium als Kalium anwesend sind. Die Aktivierung wird dureh }terzglykoside gehemmt, yon denen ande- rerseits bekannt ist, dag sie die Natrium-Kalium- Pumpe bloekieren. Die Aktivierungen sowohl als die Glykosidhemmung sind in beiden F/~llen konzentra- tionsabhgngig und die Konzentrationen f/it halbmaxi- male Wirkungen zeigen frappante quantitative LTber- einstimmung.
J~hnliehe Membran-ATPasen sind inzwisehen an zahlreichen anderen Zellarten gefunden worden a.
Wertvolle ParMlelen und Erg~nzungen bieten die Untersuchungen yon HASSELBAC~ an den calcium- speichernden Granula. Der Transport, der hier relativ raseh ist, erfolgt in Sehiiben, die jeweils zu einer be- stimmten ErhShung der Caleiumaufnahme f/ihren und mit der Spaltung einer st6ehiometriseh gquivalenten Menge ATP verbunden ist (s. Abb. 12). Es wird also f/Jr jedes transportierte Caleiumion ein Molekiil ATP gespalten.
Die Untersuehungen yon HASSELBAOII ~9, S0 geben nun noeh eine Antwort auf eine weitere Frage, die sieh hier stellt. Die Spaltung des ATP ist ein chemi- sober Vorgang mit einem groGen Energiesprung. Wfirde sie ohne Zwisehenstufen direkt naeh der Reaktions- gleiehung ATP -~ ADP + anorganisehes Phosphat er- folgen, so k6nnte die Energiedifferenz nieht verwertet werden. Die Situation 1/~Gt sich veranschauliehen durch den Vergleich mit einem Wasserfall, d e r n u r dann n/itzliehe Arbeit leisten kann, wenn die Wasser-
11"
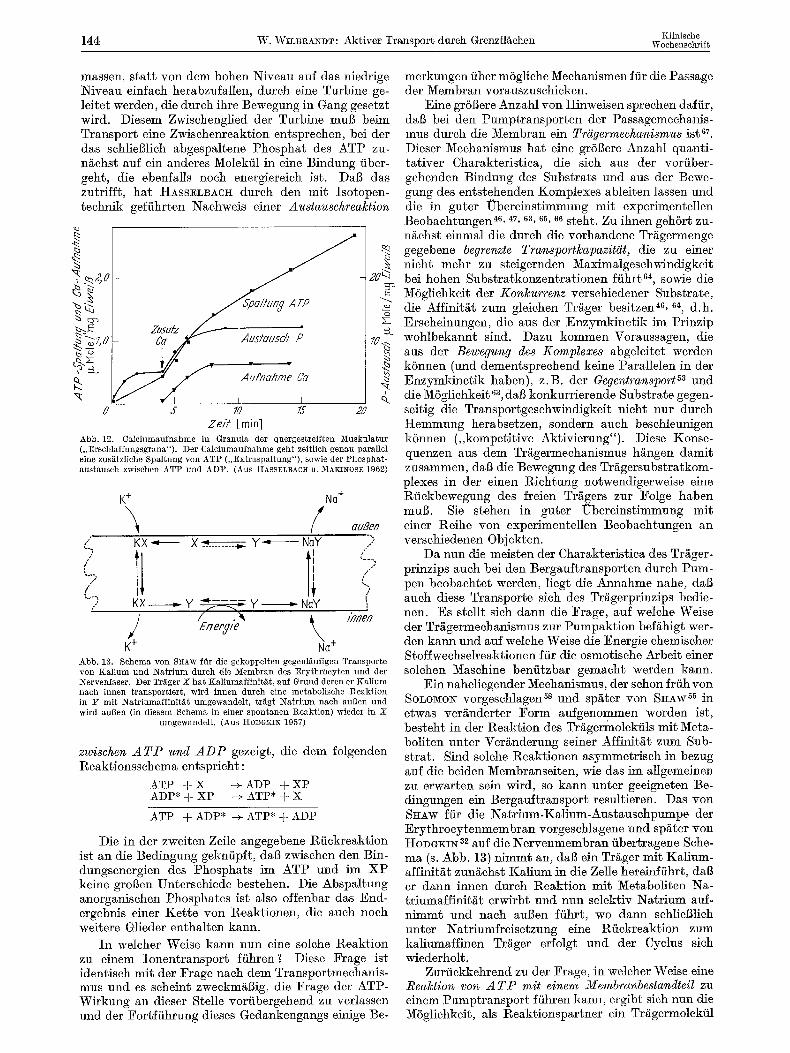
Kllnische 144 W. Wm~A~DT: Aktiver Transpor~ durch Grenzfl~chen Wochenschrift
msssen, start von dem hohen Nivesu auf das niedrige l~ivesu einfach herabznfsllen, durch eine Turbine ge- ]eitet werden, die durch ihre Bewegung in Gang gesetzt wird. Diesem Zwischenglied der Turbine mug beim Transport eine Zwischenreaktion entspreehen, bei der dss schlie$1ich abgespaltene Phosphat des ATP zu- nKchst auf ein snderes Molekfil in eine Bindung fiber- geht, die ebenfalls noeh energiereieh ist. DaB dss zutrffft, hat HASSELBACH dureh den mit Isotopen- technik geffihrten Nachweis einer Austauschreaktion
~-~ |
~ . . . ~ p a l t o z y A TP
% -8
2_
T %
0 £ 10 15 2# Zei/ [mini
Abb. 12. Calciumaufnahme in Granula der quergestreiften iVfuskulatur (,,Erschlaffungsgrana"). Der Calciumaufnahme geht zeitlich genau parallel eine zus~itzliche Spaltung yon ATP (,,Extraspaltung"), sowie der Phosphat- austausch zwischen ATP und ADP. (Aus HASSELBACtI U. MAKINOSE 1962)
K + Nn +
/ au~en ," KX = X .=-.. ........ p- Y = NaY .,-
/ ] ) "...,, ., / "') "..--i I
K× ~ Y ~ Y = NaY ¢"
K + Na + Abb. 13. Schema von S~AW ffir die gekoppelten gegenl~ufigen Transporte yon Xalium und Natrium dutch die Membran des Erythr0cyten und der Nervenfaser. Der Tr/iger X hat Kaliumaffiniti~t, auf Grund deren er Xalium nach innen transportiert, wird innen durch eine metabolische l~eaktion in Y mit NatriumaffinitKt umgewandelt, tr~igt Natrium nach an2en und wird aul3en (in diesem Schema in einer spontanen Reaktion) wieder in X
umgewandelt. (Aus I-IODGXIX 1957)
zwischen A T P und ADP gezeigt, die dem folgenden Reaktionsschema entspricht :
A T P d - X - ~ A D P d - X P ADP* d- XP --> ATP* + X
ATP d- ADP* -4 ATP* d~ ADP
Die in der zweiten Zeile angegebene Rfickresktion ist an die Bedingung geknfipft, dab zwisehen den Bin- dungsenergien des Phosphats im ATP und im XP keine gro~en Untersehiede bestehen. Die Abspaltung anorganischen Phosphates ist also offenbar das End- ergebnis einer Ket te yon Resktionen, die such noeh weitere Glieder enthalten kann.
In welcher Weise kann nun eine solehe Reaktion zu einem Ionentrsnsport ffihren ? Diese Frage ist identisch mit der Frage naeh dem Transportmechanis- mus und es scheint zweckmgl~ig, die Frage der ATP- Wirkung an dieser Stelle vorfibergehend zu verlsssen und der Fortffihrung dieses Gedsnkengangs einige Be-
merkungen fiber mSgliehe Mechsnismen ffir die Passage der Membran vorauszuschicken.
Eine grSl~ere Anzahl yon Hinweisen spreehen daffir, dab bei den Pumptrsnsporten der Psssagemeehsnis- mus durch die Membrsn ein Tri~germeJtanismus ist% Dieser Mechanismus hat eine grSl~ere Anzahl quanti- tativer Charakteristica, die sich aus der vorfiber- gehenden Bindung des Substrats und aus der Bewe- gung des entstehenden Komplexes ~bleiten lassen und die in guter ]~bereinstimmung mit experimentellen Beobaehtungen 46, 47, 63, s~, 66 s~eht. Zu ihnen geh6rt zu- n/~ehst einmal die dureh die vorhandene Tr~tgermenge gegebene begrenzte Transportkapazitdt, die zu einer nicht mehr zu steigernden Maximalgeschwindigkeit bei hohen Substrstkonzentrationen ffihrt% sowie die MSglichkeit der Konkurrenz versehiedener Substrate, die Affinitat zum gleichen Trgger besitzen 4s, % d.h. Erseheinungen, die aus tier Enzymkinetik im Prinzip wohlbekannt sind. Dszu kommen Voraussagen, die aus der Bewegung des Komplexes abgeleitet werden kSnnen (und dementspreehend keine Parallelen in der Enzymkinetik haben), z.B. der Gegentransport 53 und die MSglichkeit % da$ konkurrierende Substrate gegen- seitig die Transportgesehwindigkeit nieht nur dutch Hemmnng herabsetzen, sondern such beschleunigen kSnnen (,,kompetitive Aktivierung"). Diese Konse- quenzen sus dem Tr/~germechanismus h/ingen damit zusammen, dal~ die Bewegung des Tr~tgersubstratkom- plexes in der einen Richtung notwendigerweise eine t~iiekbewegung des freien Tr/~gers zur Folge haben muir. Sic stehen in guter Ubereinstimmung mit einer Reihe yon experimentellen Beobaehtungen an verschiedenen Objekten.
Da nun die meisten der Chsrskteristica des Tr/iger- prinzips such bei den Bergauftransporten durch Pure- pen beobaehtet werden, liegt die Annahme nahe, alas such diese Transporte sieh des Tr/~gerprinzips bedie- nen. Es stellt sich dann die Frage, auf welche Weise der Tr/tgermeehsnismus zur Pumpaktion bef~higt wet- den kann und auf welche Weise die Energie chemiseher Stoffwechselresktionen ffir die osmotisehe Arbeit einer solchen Maschine benfitzbsr gemacht werden kann.
Ein nsheliegender Meehsnismus, der schon frfih yon Sono~rox vorgeschlagen ss und sparer yon SgAw 55 in etwas ver/inderter Form aufgcnommen worden ist, besteht in der Reaktion des Tr/~germolekfils mit Meta- boliten unter Ver~nderung seiner Affinit£t zum Sub- strat. Sind solehe l~eaktionen asymmetriseh in bezug auf die beiden Membrsnseiten, wie dss im allgemeinen zu erwarten sein wird, so kann unter geeigneten Be- dingungen ein Bergauftransport resultieren. Das yon S~AW ffir die Natrium-Kalium-Austsuschpumpe der Erythrocytenmembran vorgesehlagene und sparer yon ttoDaxI~¢ s2 auf die Nervenmembran fibertragene Sche- ma (s. Abb. 13) nimmt an, da$ ein Tr/~ger mit Kalium- affinit£t zun/~chst Kalium in die Zelle hereinffihrt, da$ er dann innen durch 1%aktion mit Metaboliten Na- triumaffinit/it erwirbt und nun selektiv Natrium suf- nimmt und naeh auSen ffihrt, wo dann sehhe$1ich unter Nstriumfreisetzung eine Rfiekreaktion zum kaliumaffinen Tr/tger erfolgt und der Cyelus sich wiederholt.
Zurfiekkehrend zu der Frage, in weleher Weise eine Reaktion yon A T P mit einem Membranbestandteil zu einem Pumptransport ffihren kann, ergibt sich nun die MSgliehkeit, sls Reaktionspartner ein Tr/~germolektil

Jg. 4i, tIert s W. W~LBaiNDT: Aktiver Transpor~ durch Grenzflachen 145 t . Februar 1963
anzunehmen und das Wesen der 1~eaktion in einer Ver~nderung der Atfinit/tt dieses Trfi.gers zum Subs~rat zu erblicken. Auf dieser Basis hat P o s t ~ die yon ihm beobaehteten Parallelen zwisehen ATPaseaktivitat und Ionentranspor~ gedeutet (s. Abb. 1Q. Er nahm an, dab ATP einem zun/~ehs~ kaliumaffinen Trgger- molekiil (vermutlich dutch Phosphatfibertragung) Natriumaffinitgt verleiht, und dab sieh an diese Reak- tion dann der yon SHaw angenommene Cyelus yon Transporten und t~eaktionen anschliegt. Die Aktivie- rung der ATPase durch Na~rium und Kalium ware dann dadurch zu erklgren, dab das Enzymsubstrat , ngmlieh der Partner ffir die Reaktion mit ATP, dem ATPasemolekfil innen so lange kontinuierlieh immer wieder zugef/ihrt wird, als der Cyelus 1/~uft. Da welter-
/nn en Pl~m b n(/ n ~uBen
P/ ~) Na Na ~a
Abb. 1t, Tr~gerschema yon P0ST zur Deu~ung der l~Iembran-ATPase- Yersuehe am Erythrocyten nnd ihrer Beziehung zum gekoppelten Na~rium- und Kalimntransport d~xch die ]grythrocytenmembran. ATP aktiviert innen den Tr~ger (vermutlich dutch Phosphorylierung) und ~-ei'leiht ihm NatriumaHinitS~, w~hrend augen ein zweites nicht identifiziertes Enzym die Aktivierungsreaktien rfickg~ngig maehg and dem Tr'Xger Kalium- affinit~t verleiht. Die Aktivierung des Enzyms beraht auf der cyclischen Zufuhr des Substrats (des Trigers), die dutch Herzglykosid gehemmt wird.
(Naeh Pos t 1961)
hin der Cyclus mi~ Natrium- und I~aliumtranspor~ durch die Membran verbunden ist, ist die hemmende Wirkung der Herzglykoside auf die Aktivierung er- kl/~rbar aus der bekannten tIemmwirkung dieser Stoff- gruppe auf die Na~rium- und Kaliumtranspor~e.
In bezug auf die chemische h~at%r des Triiger~wle- kii ls geben diese gesul ta te keinen Aufschlug. Einige Beaehtung hat ein Vorsehlag yon I]:OKL~ as gefunden, nach dem das Trggermolekiil aus Phosphatidsgure be- stehen solt, die an der Innenseite der Membran dutch Reaktion mit ATP entstehen und an der Aul3enseite dutch eine Phosphatase gespalten werden soil (s. Abb. 16). Die experimentellen BeNnde, auf die sich diese Annahme st/itzt, sind in erster Linie an der Salz- dr/ise des Albatrosvogels (und an Erythrocyten) er- hoben worden. Sic besgehen darin, dab unter Bed_in- gungen, die der Driisentgtigkeit ent~sprechen (Zusa~z von Aeetyleholin zu Schnitten der eholinergischen Dr/ise) der Umsatz der Phosphatids~/ure, gemessen an der Geschwindigkeit des Einbaus radioaktiven Phos- phats, eine bedeutende Steigerung erf~hrt, dab diese Steigerung sieh bei anderen PhosphoHpiden nieht oder in geringerem AusmaB finder nnd dab sic (s. Abb. 17) unter den verschiedenen Zellfraktionen mit betr~eht- lieher Selektivit/~t nur hi der Mikrosomenfraktion auf- t r i t t (in die die Zellenmembran uncI das doplasmatische Refieulum eingehen).
Es scheint aus diesen Befunden tatsachlich rechb wahrseheinlieh, dal3 die Phosphatids£ure hi irgendeiner Weise mit dem Natriumtransport in Verbindung steht. Ob sich allerdings die Hokinschen Vorstellungen im einzelnen wer- den halgen k6nnen, erschein~ aus mehreren Gr/inden fraglich. Offene Fragen, die sich stellen, betreffen unter anderem die Spezifit~t, die Koppelung mit Kalium und das SchicksaI des an- organischen Phosphats (das nicht in der Aul3enlSsung erseheint).
CIt~--O--Fe~tsiure
CIt--O--Fettsiure I CIte--O--POaH e
Phosphatidsiure
Abbo 15. Chemisehe S~;ruktur der
Phosphatids~are
Man k6nnte daran denken, dug die Phosphatids/~ure ein Glied in einer l~ngeren Ket te ist. Die Annahme,
@/oplasma
_ A T P ~
2C~-
Dg -~
/0 ~-KI~LTEe
PA-
~L/mEn
Na2pA -
®t P/P-~? --~-2Na
=Nazn,4~i ,4'PO~ ?/Z, . ,
~2CL-
Abb. 16. ItOKINs Schema des Na~riumtransports mit t l i~e yon Phosphatid- s~ure. ATP phosphoryliert Diglyeerid atff tier Cytoplasmaseite der ~embran mit ttilfe der Diglycerid!dnase, die so gebildete :Phosphafids~ure trans- portiert Natrium linch aut~en (zum Lumen des Drt~sengangs), wo eine
Phosphatidphosphatase die ghosphatids£ure spaRe~ und gleiehzeitig Natrium freisetzt. (Aus ttOXIN u. HOXIN 1960)
-- . TO0 000
E
soooo
i
70000
32P-lncomorathn b Phospkatid-sb'ure
[ ~ qa /z c//'~'s e )
J Kern kilt. P//c. Lb's.
Abb. 17. Steigerung des Umsatzes alex Phospha~idsiure (gemessen an der Inkorporation ~on ~sp) in den Zetlfraktionen des Nucleus (Nut), der I~Iitoehondrien (Mit), der Mikrosomen (Mic) und der iibei~tehenden L6sung (Sol) im Gewebsschnitt der cholinergisehen Satzdrfise des Albatros, ,.stimuliert" dutch Zusatz yon Aeetylcholin. (Nach H0rdN u. HOg_IN 1960)
dal3 sic nicht dem Natriumtransport, sondern dem Phosphatrficktransport naeh innen dien~, wg.re mi~ den Hokinsehen Befunden rdcht in ~¥idersprueh und wiirde wenigstens eine der erw~hnten offenen Fragen, nam- lich das Schieksal des anorganisehen Phosphats, beant- worten k6nnen.
So bleibt die Frage der Natur des Tr/igers zun~chst often. Von anderen Vorsehl/~gen, die gemaeh~ worden

146 W. WILBRANDT: Aktiver Transport dareh Grenzflachen Klinisehe Woehenschrift
sind, sei derjenige yon Ct~ISTESSES ~° erwahnt, der f/Jr den Aminosauretransport in Tumorzellen als Trager Pyridoxsl vorschlug und annahm, dab der gebildete Komplex noch Alkalikationen enthalt, so dab er die Grundlage yon Beobachtnngen sein kSnnte, die auf eine Koppelung zwischen Alkalikationentransport und Aminosauretransport hinweisen.
Aueh ftir diese Vorstellung bestehen im Augenbliek einige Sehwierigkeiten n, auf die hier nieht eingegangen sei. Dagegen ist die Koppelung zw~schen Alkali- Kationen- Transporten und Transporten anderer Sub- strate nicht auf Aminos~uren besehr/~nkt, sondern eine Beobachtung, der offenbar allgemeinere Bedeutnng zukommt. So ist die Glueoseresorption im Darm yon der Amvesenheit des Natr iums a bh~ngig ~-~, el, das- selbe gilt fiir die Aminosaureresorption 14, ~s und f/it die
l //erz - Phlo/'/z/h --- - - --~Igc~s/c/e
g + Na
Abb. lS. gorstellung yon CI~A~E fiber den Meehanismus der Xoppelung zwisehen Olueosetransport und Natriumt,ransport an der ~pithelzelle des Dfinnd~rms (Abhingigkeit der Glueoscresorption yon der Anwesenheit yon Natrium). Ein gemeinsamer (durch einen Xreis bezclehneter Triger) f ~ r t Glucose (G) und Natrium (~a) durch die Membran in die Zelle (gehemmt dureh Phlorizin). Im Zellinneren wird Glucose and Natrium freigesetzt. Die En~fernung des Natriums dutch die herzglykosid-empfindliehe Pumpe erhSht den Gradienten des Xolnplexes und ifihrt so zu Glucoseakkumnla-
tion in der Zelle. (Naeh C~ANE, I~II~E~ u. BIH~ER 1961)
Jodidakkumulat ion in der Sehilddr/ise. In allen diesen Fallen hemmt nieht nur Natriumabwesenheit den Transport, sondern such die Anwesenheit yon tterz- glykosid, das Alkali-Kationen-Transporte hemmt.
Diese Beobachtungen maehen es wahrscheinlieh, dab die in Frage stehenden Transporte ihre Energie nicht unmit telbar aus dem Stoffwechsel beziehen, son- dern dab sie in irgendeiner Weise an den Natr iam- t ransport angekoppelt sind. Man hat solehe Trans- porte als se]cundi~re Transporte bezeichnetSL Die Art der Ankoppelung ist zunachst Gegenstand der Speku- lation. C ~ A ~ is hat ffir die Glucoseresoprtion ange- nommen, dab ein Natrium-Glucose-Trggertransport die Zellmembran passiert, innen Natr ium und Glucose freisetzt und dab dann die Entfernung des Natr iums dureh die Nat r iumpumpe dazu fiihrt, dab der Gradient Iiir den Komplex steit gehalten und aul diese Weise Glucose in der Zelle akkumuliert wird (s. Abb. 18).
Diese Vorstelhmg sollte zun/~ehst nut als Beispiel dafih- betraehtet werden, dab das Prinzip des Trager- transportes Koppelungen zwisehen zwei Transporten im Sinne einer Mitffihrung oder einer Gegenfiihrung leieht deutbar maehen kann. Die MSglichkeiten sind mannigfaltig, die sich aus der gemeinsamen Benfitzung eines Trggers dutch zwei Substrate oder des Trans- porges eines Substrates dutch zwei versehiedene Trgger oder dutch l~ea.ktionen z~dsehen verschiedenen Tr/~- gern oder Substraten ergeben. Die Natriumabh~ngig- keit der Glueoseresorption ware beispielsweise such
dann zur erwarten, wenn Natrium und Glucose ver-
schiedeneTrager beniitzen, die aber miteinanderreagie-
ren und einen Komplex bilden kSnnen. Eine erste Frage, die gestellt und experimentell
angegangen werden kann, ist die, ob das Erfordernis ffir den Ablauf des sekundaren Transportes darin liegt, dab die Pnmpe des pr imaren Transportes lauft oder nur darin, dab der Gradient, der durch die prim~tre Pumpe aufreehterhalten wird, besteht. Beides ware denkbar. Versuehe an Sehilddriisensehnitten ss spre- ehen im Falle der Jodidakkumulat ion dafiir, dab der sekundare Jodidtransport lauft, sofern der Natr ium- gradient an der Zelte erhalten bteibt, unabhangig da- yon, ob die Nat r iumpumpe bloekiert ist. Wenn an diesem Objekt der Zuffihrung yon Herzgtykosid eine natriumfreie Phase vorausgesehiekt wird, ~drd die intraeellul~re Anhanfung yon Nat r inm verhindert (weil anBen keines anwesend ist), obwoht die Na t r i ampumpe blockie~ ist. Stellt man dann in Anwesenheit des Herzglykosids den Natriumgr~dient dutch Zuffigung yon Nat r ium zum Augenmedium wieder her, so fangt die Jodidpumpe vorfibergehend wieder an zu laufen. Naeh vorlaufigen Versuehen scheint es mSglieh, dab die Situation bei der Glueoseresorption im Darm ahn- lich ist.
MSglicherweise sind sekundare Transporte, die in ahnlicher Weise ihre Energie unmit te lbar aus der Kat ionenpumpe nnd nur mit telbar aus dem Stoff- wechsel beziehen, weJter verbreitet, als bisher bekannt. Beispielsweise wurde gefunden, dab such dieAnfnahme yon Serotonin in Blutplat tchen durch Herzglykoside gehemmt wird s~. Die Bedeutung des Natr iums ist bier aber noch nicht im einzelnen untersucht. Aneh die Tatsache, dab HOKIN ghnHehe Umsatzsteigerungen der Phosphatidsaure wie beim Natr iumtranspor t such bei anderen Transportsystemen beobachtet hat, k6nnte auf einer Verknfipfung zwischen diesen Transporten und den Kationentransporten beruhen. Wieweit die Tragfahigkeit solcher Gedankengange reicht, kann nnr dutch weitere experimentelle Untersuehungen geklart werden.
Literatur. 1 ALsRmc, V. E., R. MENDT, J. W. HoPkins and A. E. 1VIIRSK:f : Proc. nat. Aead. Sci. (W~sh.) 47, 907 (1961). - - 2 BARA~¥, E., and E. SP]~Rm~R: Scand. Arch. Physiol. 81, 290 (1939). -.~ a BO~TI~G, S. L., K. A. SI~o~ and N. M. HAw~:I~s : Arch. Biochem. 95, 416 (1961). - - t Borer, G.V. 1%., and J. BR~CK~rJ~LL: J. Physiol. (Lond.) 147, 153 (1959).- s Bo~5¢, G. V. R., and ~. E. GrLLSOSr: J. Physiol. (Lond.) 146, 472 (1959). - - 6 Bo]~-, G. V. 1~., G. I. I~G~A~ and R. S. ST±OEY: Brit. J. Pharmacol. 13, 62 (1958). - - ~ BOYLE, P.J . , and E. J. Co,wAY: J. Physiol. (Lond.) 100, 1 (I941). __s BIgAc~]~, J. : Sci. Amer. 205, 51 (1961). - - ~ CALI)W]S~, P.C., A. L. HODGKIig, ]~. D. KEYIVES and T. I. SEAW: J. Physiol. (Lond.) I~2, 561 (1960). -- ~0 C~m~STE~SE~, H. N.: Science 122, 1087 (1955). -- n C~q~STE~SE~, H.N.: In A. K~sr~zs~ and A. KOTYK (edit.), Membrane transport and metabolism, p. 470. Prag 1961. --~a COHEn, G.N., and J. Mo~oD: Bact. Rev. 21, 169 (1957). -- zs C~A~E, R. K., D. M~L~ and i. B~gn~s~: In A. KL~XNZELnEX~ and A, KOTYK (edit.), l~embrane trans- port and metabolism, p. 439. Prag 1961. -- ~a CSAK¥, T. Z. : Biochem. Pharmacol. 8, 38 (1961). -- ~s CSAKY, T. Z. : Amer. J. Physiol. 201,999 (1961). --~s CSAK¥, T. Z, and MA~GIg~T~gE T~i[nnS: J. Physiol. (Lond.) 151, 59 (1960). -- ~¢ CSAK¥, T. Z., and L. ZOLL~CO~ER" Amer. J. Physiol. 198, 1056 (1960). -- ~s BArnES, ~. E.: Syrup. Soc. exp. ]~iol. 8, 476 (1954). -- ~s DAVIES, R. E./A. M. DELL~gVA, I. J. DEW'SUP and R.W. ~VL~TEIgS: In: Membrane transport and metabolism (edit. A. XLEISZELLEI~ and A. KOT~ZK), p. 285. Prag 1961. --
DEA~, I~,. B.: Biol. Symposia. ~, 331 (1941). -- ~ DUNHA~, E, T., and I. M. GLYS~: J. Physiol. (Lond.) 156, 274 (1961). -- ~ FETCH]ZIg, E.S.: Quart. Rev. Biol. 14, 451 (1939). - -

Jg. 41, tIeft 3 EUGENE M. ~=~ENKIN: Stofftransport durch die Wande der Blutcapillaren 147 1. Februar 1963
~a FI~cH, L. 1~., and F. J. 1~. HIRD: Biochim. biophys. Acta (Amst.) 43, 268 (1960). - - ea FINe~, L. R., and F. J. R. Hn~D : Biochim. biophys. Act~ (Amst.) 43, 278 (1960). - - ~ FLECKEN- STEIN, A., E. GERLAC]~ U. J . JANKE: Schweiz. med. Wschr. 86, 1041 (1956). - - ~ GARDOS, G., U. F. B. STRAIFB: Acta physiol. Aead. Sei. hung. 12, 1 (1957). - - ev G][LBERT, DANIEL L. and JOA~A MeGANN: J. geL. Physiol. 43, 1103 (1960). - - 2s GLYNN, I. M.: Progr. Biophys. 8, 241 (1957). - - e~ ttASSEL- ~AOH, W., u. M. MA]tI~OSE: Biochem. Z. 333, 518 (1961). - - 3o HASSELBACH, W., and M. ~V~AKINOSE: Biochem. biophys. Res. Comnmun. 7, 132 (1962). - - ~ HEINZ, E.: 12. Kolloq. GeE. physiol. Chemie 1961, S. 167. - - ~ HODGKIN, A.L.: Prec. roy. Soc. B 148, 1 (1957). - - aa HODGKIN, A. L., and R. D. KEYN~S: Syrup. Soc. exp. Biol. 8, 423 (1954). - - ~ HODGKI~, A.L., and R.D. KE~NES: J. Physiol. (Lend.) 138, 253 (1957). - - ~ HOKIN, L. E., and M. R. HoKI~: J. gen. Physiol. 44, 61 (1960). - - ~ HOREOKER, B. L., J. T~O~AS and J. MONOD : J. biol. Chem. 235, 1580 (1960). - - ~ HO~EeKER, B.L., J. T~O~AS and J. MONOD: J. biol. Chem. 23~,5, 1586 (1960). - - as IFF, H. W. : In Vorbereitung. - - ~ KATZ, B. : Sei. Amer. :]05, 209 (1961). - - ~0 K~Ys, A.: Prec. roy. Soc. B 112, 184 (1933). - -a~ KoctI, H . J . : In J. A. KITCttING (ed.), Recent developments in cell physiology, p. 15. London 1954. - - ~s KROGI~, A. :Scand. Arch. Physiol. 76, 60 (1937). - - ~a K ~ o ~ , A. : Osmotic regulation in aquatic animals. Cambridge: Uni- versity Press 1939. - - ~ K~oG~, A.: Prec. roy. Soc. B 133, 140 (1946). - - as LASSEN, ULRIX, V., ~nd K. OVERGAARD- HANSEN: Biochim. biophys. Acta (Amst.) 57, 111 (1962). - - as LE F~VRE, P. G., and R. I. DAVIES: J. gem Physiol. 34,
515 (1951). - - ~7 LE F~VRE, P. G., and M. E. LE F~vR]~: J. gen. Physiol. 8~, 891 (1952). - - as McDouGAL, D. B., K. D. LITTLE and R. K. CRAS~: Biochim. biophys. Acta (Amst.) 4.~, 483 (1960). - - 4s POST, R. L.: In: Twelfth Annual Conference on the Nephrotic Syndrome, ed. by Dr. J. METCOFF ~nd publ. by the National Kidney Disease Foundation, New York 1962. - - 50 POST, R. L., C. R. MERRIT, C. R. KINSOLVING and C. D. ALERIGttT: J. biol. Chem. 235, 1796 (1960). - - s~ Rn~Lis, E., and J. H. QUASTEn: Canad. J. Biochem. 36, 347 (1958). - - s* ROEE~WSON, M.D.: Progr. Biophys. 10, 343 (1960). - - s~ ROSENBEI~G, T., and W. W~LB~A~DT: J. gen. Physiol. 41, 289 (1957). - - s~ SOH~IDT-NIELSEN, K., C. B. JSRGENSEN and H. OsAxI: Amer. J . Physiol. 193, 101 (1958). - - s s S~Aw, T. I . : Ph. D. Thesis Cambridge University 1954. Zit. nach GLY~ ~. ss SIST~O~, W. R. : Biochim. biophys. Act~ (Amst.) '29, 579 (1958). - - s~ SKOU, JE~s CgR. : Biochim. biophys. Acta (Amst.) 23, 394 (1957). - - ss Sono~os, A.K.: J. gem Physiol. ~{;, 57 (1952/53). - - ss USSI~G, H.H. : J. fen. Physiol. 43, 135 (1960). - - so UssINa, H.H.: 12. Koll. Ges. physiol. Chemie, S. 1. 1961. - - 61 SLYKE, D. D. VAN, and G. IV[. MEYER: J . biol. Chem. 16, 197 (1913). - - se WEISSBACtt, H., B. J. RED- FIELD and E. TITvs: Nature (Lend.) 185, 99 (1960). - - 63 WIDDAS, F. W. F. : J . Physiol. (Lend.) 12~, 163 (1954). - - s~ WILB~ANDT, W.: Dtsch. med. Wschr. 28, 1153 (1957). - - ss WIL~A~DT, W.: 12. Koll. GeE. physiol. Chemie, S. 111. Mosbach (Baden) April 1961. - - ss WILBI~ANDT, W., S. FREI and T. ROSENBERG: Exp. Cell l~es. l l , 59 (1956). - - s~ W~L- ~aANDT, W., and T. ROSE~BEE~: Pharmacol. Rev. 13, 109 (1961).
Stofftransport durch die W~nde der Blutcapillaren Von
EUGenE M. RENKIN
Department of Physiology, The George Washington University, Washington, D.C.U.S.A.
Viele versehiedenart ige Subs tanzen werden yon der Blu tz i rkula t ion t ranspor t ie r t u n d dr ingen dureh die W~nde der Bluteapi l laren ein oder aus. Diese Sub- s tanzen reiehen yon der GrSl~e eines Wassermolekfils bis zur GrSBe der grSBten Plasmaeiweigmolektile. Kleine Molekfile, die vom Zellstoffwechsel aufgenom- men oder abgesehieden werden, werden in enormen Mengen t ransport ier t . Die grS~eren Molekfile: Hor- mone, ImmunkSrper , EiweiBkSrper usw., werden in kleineren, jedoch wiehtigen Mengen t ransport ier t . Die Blu tz i rkula t ion t ranspor t ie r t aueh die W~rrne, u n d ist verantwort l ich ffir das Gleiehgewieht des Wassers und der Elektrolyte tiberall im Organismus.
Die Capi l la rmembran er laubt den freien Durehgang all dieser Subs tanzen u n d verh inder t dabei einen Volumenver lus t des zirku]ierenden Blutes. Die stufen- weise En tdeekung der Meehanismen, wodureh dieses bewirkt wird, stellt ein s t imulierendes Kapi te l in der En twiek lung der modernen Physiologic dar. Unser Vers tandnis dieser Mechanismen ist noch weft von vollst~ndig. Heute werde ich eine ~[~bersieht fiber einige unserer Kenntn i sse des Stofftransportes dureh die Capi l la rmembranen prgsentieren. Ieh hoffe dabei Ihre Aufmerksamkei t auf manche ungelSste Probleme, die unsere Einb i ldungskraf t u n d unser Versuchs- gesehick herausfordern, zu ]enken.
Der Prozeg verantwort l ich ffir den Haup tan t e i l des Stofftransportes ist Diffusion. Dieses ist eine Folge der unaufh5r l iehen kinet isehen Bewegung der ein- zelnen Molekfile u n d Ionen, die ihre gleiehmggige Vertei lung in dem f fir sie zuggngliehen R a u m zu- s tande bringt . Obgleieh die einzelnen Molekiile sieh in allen R ieh tungen bewegen, Ne t to t ranspor t geht immer yon hohem zu niedrigem elektroehemisehen Potent ia l (NETTleR 1959, S. 67). Die Diffusion un-
geladener gelSster Molekfile durch eine dt inne Membran wird yon dem wohlbekannten Fickschen Gesetz be- schrieben :
M1-~2 = DAA x (ai--%).
Dieses stellt dar : Die Menge einer spezifischen Substanz t ranspor t ie r t yon Seite 1 zur Seite 2 pro Zeiteinheit (M 1-,2) ist gleich dem Produk t seiner freien Dif- fus ionskonstante (D), effektiver Diffusionsfl£che pro Einhei t der Wegl~nge in der Membrane (A/x] x) u n d der Aktivit~tsdifferenz der Substanz (al--%). I n sehr verdf innten LSsungen k a n n die Konzent ra t ions- differenz ans t a t t der Aktivi tatsdifferenz gebraucht werden. Da der Transpor t dureh Diffusion aus den individuel len Molekfilbewegungen resultiert , kSnnen versehiedenen Subs tanzen gleiehzeitig in entgegen- gesetzten Rieh tungen t ranspor t ie r t werden.
Charakterist isch fiir die Diffusion dureh die Capri- larwande sind die Geschwindigkeit ffir kleine Molekiile u n d die Selektivi t£t in Respekt zur MolektilgrS~e. Die Diffusionswege sind klein; die Capil larwand selbst ist weniger ale 1 # dick, u n d die En t f e rnungen zwisehen Capillaren reiehen yon 10--100/~. C~II~ARD u. Mitarb. (1955) zeigten mi t Hilfe yon Tracer-Iso- topen, dab 90 % oder mehr des Wassers und anderer kleiner Molektile des Blutes in einem Durchgang durch die Capillaren aus den Capillaren diffundieren kSnnen. Mit ansteigender MolekfilgrSl~e wird der Prozentsatz der d i f fundierenden Molektile framer geringer, u n d schlieBlieh ffir Molekfile yon der GrSf~e des Hamo- globins (Molekulargewieht 68000) und Serumalbu- rains (Melekulargewieht 70000) ist die diffundierende Menge ffir einen einzelnen Durehgang unmeBbar klein.
PAPPE~HEIME~ U. Mitarb. (1951) un te r such ten die Diffusionsgesehwindigkeit durch die Capi]larw~nde





























