Embed Size (px)
Citation preview

Aus der Klinik für Kinder- und Jugendmedizin
des St. Josef-Hospitals Bochum - Universitätsklinik -
der Ruhr-Universität Bochum Direktor: Prof. Dr. med. Eckard Hamelmann
Ausdifferenzierung von Memory-Zellen unter Einfluss Respiratory Syncytial Virus-infizierter
Dendritischer Zellen in vitro
Inaugural-Dissertation zur
Erlangung des Doktorgrades der Medizin einer
Hohen Medizinischen Fakultät der Ruhr-Universität Bochum
vorgelegt von Sandra Zawatzki
aus Gelsenkirchen
2012

Dekan: Prof. Dr. med. Klaus Überla Referent: Prof. Dr. med. Uwe Schauer Korreferent: Prof. Dr. med. Albrecht Bufe Tag der Mündlichen Prüfung: 14. Mai 2013

Meinen Eltern
und
Großeltern
„Phantasie
ist wichtiger als Wissen,
denn Wissen ist begrenzt.“
Albert Einstein

1
INHALTSVERZEICHNIS
1. EINLEITUNG .........................................................................10
1.1 DAS RESPIRATORY SYNCYTIAL VIRUS .......................................10
1.1.1 Geschichte und Virologie ...........................................................10
1.1.2 Klinisches Erscheinungsbild und Pathophysiologie ...................11
1.1.3 Diagnostik ..................................................................................13
1.1.4 Therapie und Impfung ...............................................................14
1.2 DIE IMMUNANTWORT .......................................................................15
1.2.1 Die primäre Immunantwort: T-Zellen und Dendritische Zellen ...15
1.2.2 Das immunologische Gedächtnis: Zentrale Memory- und
Effektor-Memory-T-Zellen ..........................................................17
1.3 OBERFLÄCHENMOLEKÜLE ANTIGENPRÄSENTIERENDER ZELLEN ...............................................................................................19
1.3.1 Antigenpräsentation durch Humane Leukozytenantigene .........19
1.3.2 CD80 und CD86 als Co-Stimulatoren Antigenpräsentierender
Zellen .........................................................................................21
1.3.3 CD83 als Marker reifer Dendritischer Zellen ..............................23
1.4 POLY-IC: EIN SIMULATOR VIRALER INFEKTIONEN IN VITRO
UND SEINE BINDUNG AN DEN TOLL-LIKE-REZEPTOR 3 .........25
1.5 SUPERANTIGENE ...............................................................................27
1.5.1 Der Aufbau des T-Zell-Rezeptors ..............................................27
1.5.2 Superantigene und ihre besondere Bindung an den TCR .........27
1.5.3 Aufbau und Bindungsspezifität des Toxic Shock Syndrome
Toxins-1 .....................................................................................29

2
2. FRAGESTELLUNG ..............................................................30
3. MATERIAL UND METHODEN ...........................................31
3.1 GEWINNUNG DENDRITISCHER ZELLEN AUS NABEL- SCHNURBLUT .....................................................................................31
3.1.1 Gewinnung von Nabelschnurblut ...............................................31
3.1.2 Gewinnung mononukleärer Zellen mittels Dichtegradienten-
zentrifugation .............................................................................32
3.1.3 Gewinnung CD34+ Stammzellen mittels magnetischer
Zellseparation ...........................................................................32
3.1.4 Einfrieren der CD34--Fraktion ....................................................33
3.1.5 Anzucht Dendritischer Zellen .....................................................33
3.1.6 Auftauen Dendritischer Zellen ...................................................34
3.1.7 Restimulation Dendritischer Zellen an Tag 7 .............................35
3.2 ISOLIERUNG NAIVER T-ZELLEN UND CO-KULTUR MIT DENDRITISCHEN ZELLEN ................................................................35
3.2.1 Gewinnung naiver CD4+ T-Zellen ..............................................35
3.2.2 Co-Kulturen naiver T-Zellen mit Dendritischen Zellen als
in vitro-Modell der primären Immunantwort ...............................36
3.3 PRÄPARATION VON UND INFEKTION MIT RSV .........................37
3.4 ZELLBESTIMMUNG MITTELS DURCHFLUSSZYTOMETRIE .........38
3.4.1 Das Prinzip der Durchflusszytometrie ........................................38
3.4.2 Fluoreszenzfärbungen und durchflusszytometrische
Messungen ................................................................................38
3.5 DIE SYNGENE LANGZEIT-CO-KULTUR IM ZEITLICHEN ABLAUF ...............................................................................................39

3
3.6 DATENAUFNAHME UND -ANALYSE ...............................................42
3.7 STATISTISCHE ANALYSE .................................................................42 3.8 EINGESETZTE MATERIALIEN UND HERSTELLERNACHWEIS ....43
4. ERGEBNISSE .......................................................................46 4.1 DIE RSV-INFEKTION FÜHRT LEDIGLICH ZU EINER SCHWACHEN REIFUNG DENDRITISCHER ZELLEN IN VITRO ..47
4.2 DIE RSV-INFEKTION HAT KEINEN NEGATIVEN EINFLUSS AUF DIE AUSBILDUNG VON MEMORY-T-ZELLEN BEI DER IMMUNOLOGISCHEN SEKUNDÄRANTWORT .................................51
5. DISKUSSION ........................................................................57
5.1 DIE RSV-INFEKTION FÜHRT ZU EINER IMMUNOLOGISCHEN
SEKUNDÄRANTWORT MIT AUSBILDUNG VON MEMORY- T-ZELLEN ............................................................................................57
5.2 DIE RSV-INFEKTION INDUZIERT DIE REIFUNG DENDRI- TISCHER ZELLEN WENIGER STARK ALS POLY-IC ...................61
5.3 DIE REAKTION DER T-ZELLEN AUF TSST-1 IST EHER
SUPERANTIGEN- ALS ANTIGEN-SPEZIFISCH ..............................64

4
6. ZUSAMMENFASSUNG ........................................................65
7. LITERATURVERZEICHNIS ..................................................67
DANKSAGUNG LEBENSLAUF

5
VERZEICHNIS DER ABKÜRZUNGEN
+ positiv
- negativ
α alpha
β beta
γ gamma
Δ Delta
ĸ kappa
µ mikro
Abb. Abbildung
al. andere (alii/aliae)
APC Antigenpräsentierende Zelle (antigen presenting cell)
BAL Bronchoalveoläre Lavage
BPD Bronchopulmonale Dysplasie
CCR Chemokinrezeptor (chemokine receptor)
CD Cluster of Differentiation
CMV Cytomegalie-Virus
CPAP Continuous Positive Airway Pressure
Da Dalton
DC Dendritische Zelle (dendritic cell)
DNA Desoxyribonukleinsäure (desoxyribonucleic acid)
ds doppelsträngig (double-stranded)
ELISA Enzyme-linked immunosorbent Assay
ER Endoplasmatisches Retikulum
ExT Exfoliating toxin
FSC Vorwärtsstreulicht (forward scatter channel)
g Gramm
g Gravitationskonstante
h human
HEp Humane Epithelzelle
HIV Human Immunodeficiency Virus

6
HLA Humanes Leukozyten-Antigen
HSV Herpes Simplex Virus
IE Internationale Einheit
IFN Interferon
Ig Immunglobulin
IL Interleukin
IU International Unit
k Kilo
L Levo (links)
l Liter
LPS Lipopolysaccharid
LRR domain leucinreiche Wiederholungsdomäne (leucine-rich repeat
domain)
m Bote (messenger)
m mega
m Meter
m milli
m murin
M Molare Masse
MAP Mitogen-aktiviertes Protein (mitogen-activated protein)
MFI mittlere Fluoreszenzintensität (mean fluorescence
intensity)
MHC Majorhistokompatibilitätskomplex (Major
Histocompatibility Complex)
MOI Infektionsmultiplizität (Multiplicity of Infection)
MyD88 Adapterprotein (Myeloid differentiation primary response
gene 88)
n nano
NFĸB Transkriptionsfaktor (nuclear factor of kappa-light chain
inducer in mature B cells)
NK Natürliche Killerzellen
n. s. nicht signifikant
PAMP Pathogen-assoziiertes Molekülmuster (pathogen-
associated molecular pattern)

7
PCR Polymerase-Kettenreaktion (Polymerase chain reaction)
PGE Prostaglandin E
PKR Proteinkinase R
Poly-IC Polyinosinic-polycytidylic acid
PRR Mustererkennungsrezeptor (pattern recognition receptor)
RNA Ribonukleinsäure (ribonucleic acid)
RSV Respiratory Syncytial Virus
s. siehe
SEB Staphylococcus aureus Enterotoxin B
SEC2 Staphylococcus aureus Enterotoxin C2
SEE Staphylococcus aureus Enterotoxin E
ss einzelsträngig (single-stranded)
SSC Seitwärtsstreulicht (side scatter channel)
SSW Schwangerschaftswoche
TCR T-Zell-Rezeptor (T-Cell Receptor)
TIR domain Toll-IL-1-Rezeptordomäne
TLR Toll-like Rezeptor
TSST Toxic Shock Syndrome Toxin
vs. versus
W Watt

8
VERZEICHNIS DER ABBILDUNGEN
Abbildung 1: Aufbau des Respiratory Syncytial Virus ........................11
Abbildung 2: Anteil RSV-F-Protein-positiver Dendritischer Zellen
nach Infektion mit RSV, Stimulation mit Poly-IC und
ohne Behandlung (Kontrolle) ........................................46
Abbildung 3: MFI (mean fluorescence intensity) HLA-DR-positiver
Dendritischer Zellen nach Infektion mit RSV,
Stimulation mit Poly-IC und ohne Behandlung
(Kontrolle) .....................................................................48
Abbildung 4: Anteil CD86-positiver Dendritischer Zellen nach
Infektion mit RSV, Stimulation mit Poly-IC und
ohne Behandlung (Kontrolle) ........................................49
Abbildung 5: MFI (mean fluorescence intensity) CD83-positiver
Dendritischer Zellen nach Infektion mit RSV,
Stimulation mit Poly-IC und ohne Behandlung
(Kontrolle) .....................................................................50
Abbildung 6: Präsentation des naiven T-Zell-Oberflächenmarkers
CD45RA an den Tagen 1, 3, 7 und 22 nach Infektion
mit RSV, Stimulation mit Poly-IC und ohne
Behandlung (Kontrolle) .................................................53
Abbildung 7: Präsentation des Memory-T-Zell-Oberflächenmarkers
CD45R0 an den Tagen 1, 3, 7 und 22 nach Infektion
mit RSV, Stimulation mit Poly-IC und ohne
Behandlung (Kontrolle) .................................................54

9
Abbildung 8: Präsentation des Memory-T-Zell-Oberflächenmarkers
CD49a an den Tagen 1, 3, 7 und 22 nach Infektion
mit RSV, Stimulation mit Poly-IC und ohne
Behandlung (Kontrolle) .................................................55
Abbildung 9: Präsentation des T-Zell-Rezeptor-Oberflächenmarkers
Vβ2 an den Tagen 1, 3, 7 und 22 nach Infektion
mit RSV, Stimulation mit Poly-IC und ohne
Behandlung (Kontrolle) .................................................56

10
1. EINLEITUNG
1.1 DAS RESPIRATORY SYNCYTIAL VIRUS 1.1.1 Geschichte und Virologie
Das RNA-Virus Respiratory Syncytial Virus (RSV) gehört zu einer Subfamilie
der Paramyxoviren, den Pneumoviren. Es wurde erstmals im Jahr 1956 von
Morris als Erreger eines Schimpansen-Schnupfens beschrieben (Morris et al.,
1956). Ein Jahr später konnte Chanock denselben Erreger bei Kindern mit
Atemwegsinfektionen nachweisen (Chanock et al., 1957).
Der Name Respiratory Syncytial Virus beschreibt die Fähigkeit des Erregers,
Atemwegsinfektionen hervorzurufen und in der Zellkultur große Synzytien zu
bilden. Es ist ein einzelsträngiges RNA-Virus negativer Polarität [(-)ssRNA] und
umfasst 15.222 Basenpaare, welche die zehn Virusproteine kodieren
(Sullender, 2000). Umschlossen wird das Genom von einem Nukleokapsid von
etwa 13,5 nm Durchmesser, das nochmals von einer Lipiddoppelschicht, dem
so genannten envelope, umgeben ist. Die envelope-Proteine der Pneumoviren
besitzen keine Neuraminidase- und Hämagglutinin-Aktivität (Eigenschaften, die
das Anheften an die Wirtszelle und das Eindringen ermöglichen), was sie von
den übrigen Paramyxoviren unterscheidet. Die Fähigkeit, sich an die Wirtszelle
zu heften und in sie einzudringen, erhält das RS-Virus durch zwei seiner drei
Oberflächenproteine: Das Glycoprotein G stellt den Kontakt zur Wirtszelle her
und ermöglicht über spikes die Anheftung. Das Fusionsprotein F ermöglicht die
Penetration in die Wirtszelle und die Virenverbreitung. Die Funktion des dritten,
hydrophoben Glycoproteins, des SH-Proteins (small hydrophobic), ist bisher
noch unklar, es scheint jedoch - gemeinsam mit dem G-Protein - eine Rolle bei
der angeborenen Immunantwort auf die RSV-Infektion zu spielen (Tripp et al.,
1999).
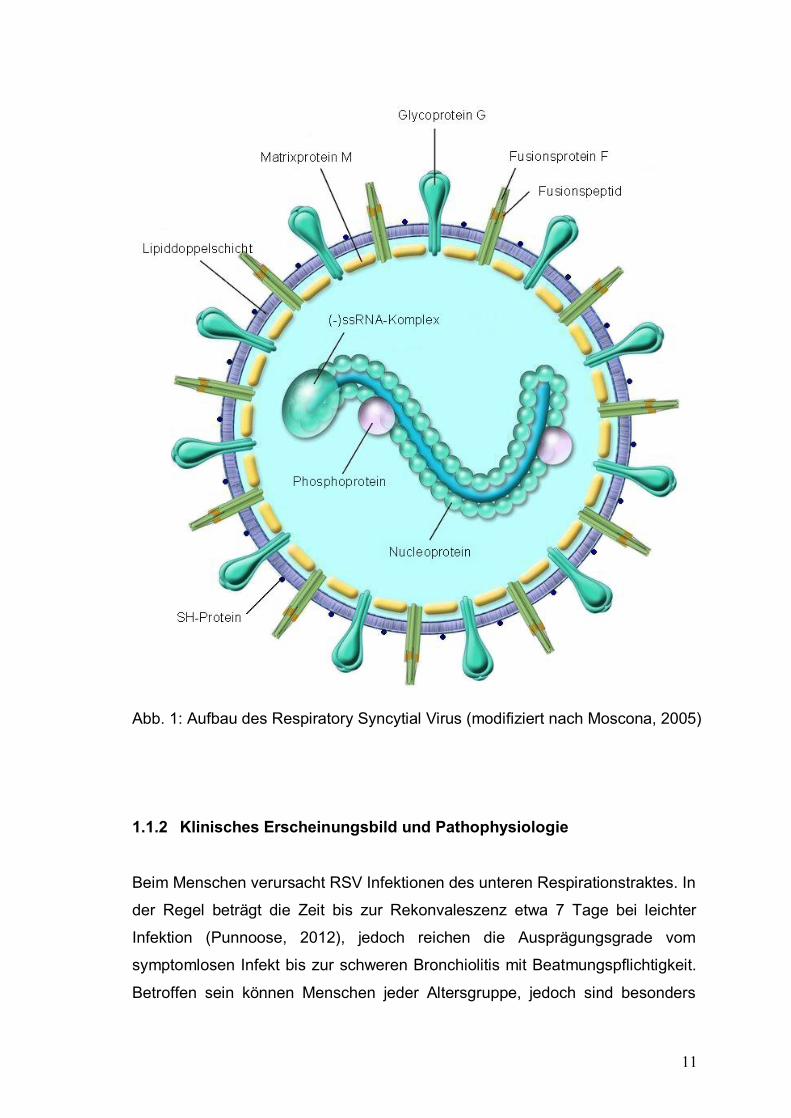
11
Abb. 1: Aufbau des Respiratory Syncytial Virus (modifiziert nach Moscona, 2005)
1.1.2 Klinisches Erscheinungsbild und Pathophysiologie
Beim Menschen verursacht RSV Infektionen des unteren Respirationstraktes. In
der Regel beträgt die Zeit bis zur Rekonvaleszenz etwa 7 Tage bei leichter
Infektion (Punnoose, 2012), jedoch reichen die Ausprägungsgrade vom
symptomlosen Infekt bis zur schweren Bronchiolitis mit Beatmungspflichtigkeit.
Betroffen sein können Menschen jeder Altersgruppe, jedoch sind besonders

12
junge Säuglinge im Alter von unter sechs Monaten gefährdet, an einer
schweren RSV-Infektion zu erkranken.
Während es bei größeren Kindern und Erwachsenen meist lediglich zu milden
Erkältungsbeschwerden wie Rhinitis oder leichter Pharyngitis kommt, können
junge Säuglinge schwere klinische Verläufe mit Atemwegsobstruktion, pneumo-
nieähnlichen Symptomen der Bronchiolitis bis hin zu Sauerstoffbedarf oder
Intubationspflichtigkeit aufgrund von Hypoxämie und Hyperkapnie zeigen.
Besondere Risikogruppen stellen Frühgeborene mit vorgeschädigter Lunge
(Bronchopulmonale Dysplasie), Kinder mit Shuntvitien (insbesondere mit
Belastung des Lungenkreislaufs) und immunsupprimierte oder immundefiziente
Patienten dar. Innerhalb der immunsupprimierten Patienten sind Patienten nach
allogener hämatopoetischer Stammzelltransplantation besonders gefährdet,
komplizierte Krankheitsverläufe zu erleiden, da diese häufig schwere
Pneumonien mit einer hohen Letalität entwickeln (Ullmann und Maschmeyer,
2012).
Die Ansteckung erfolgt über Tröpfcheninfektion. Als Eintrittspforte dient das
respiratorische Epithel insbesondere der Nasenschleimhaut. Das RS-Virus
vermehrt sich auf den zilientragenden Schleimhäuten der Atemwege, von dort
breitet es sich direkt oder hämatogen auf den unteren Respirationstrakt aus.
Die Inkubationszeit beträgt zwei bis acht Tage. Besonders in den
Wintermonaten (November bis April) treten RSV-Infektionen gehäuft auf, man
spricht daher auch von der RSV-Saison.
Durch starkes Anschwellen der Atemwegsschleimhaut kommt es zur Zunahme
des Atemwegswiderstandes in den kleinen Bronchien und Bronchiolen, was
wiederum zu einem Missverhältnis zwischen Ventilation und Perfusion im
Lungenkreislauf führt. Die daraus resultierende Hypoxämie kann bei
respiratorischer Erschöpfung zu einer lebensbedrohlichen Hyperkapnie führen.
Bei Säuglingen können neben typischen Symptomen der Bronchiolitis wie
Tachydyspnoe, Einziehungen und Giemen auch Apnoen (mitunter auch als
Erstsymptom) auftreten. Sie stellen somit eine besondere Risikogruppe dar.
Etwa 5% der erkrankten Kinder werden ateminsuffizient und bedürfen einer
maschinellen Beatmung, die Letalität beträgt unter heutigen intensiv-
medizinischen Bedingungen 1% (Therapiehinweis zu Palivizumab, Bundes-
ausschuss für Arzneimittel, 2008).

13
Doch nicht nur während der akuten Erkrankung, auch in Bezug auf die Allergie-
entwicklung hat die RSV-Infektion einen Einfluss: Insbesondere innerhalb des
ersten Jahres nach schwerer RSV-Infektion ist die Wahrscheinlichkeit einer
Sensibilisierung für Nahrungsmittel deutlich erhöht im Vergleich zu milder oder
nicht durchlebter RSV-Erkrankung (Schauer et al., 2002). Zudem wurde die
RSV-Bronchiolitis als wichtigster und unabhängiger Risikofaktor für wieder-
kehrendes Giemen identifiziert. Möglicherweise besteht auch ein Zusammen-
hang mit der Sensibilisierung für inhalative Allergene und der Entwicklung einer
Atopischen Dermatitis.
Innerhalb der ersten Lebenswochen besteht ein Nestschutz durch diaplazentar
übertragene mütterliche Antikörper. Eine einmalig durchgemachte RSV-
Infektion schützt jedoch nicht vor der Reinfektion; beschrieben wurden sogar
Reinfektionen kurz nach der primären Infektion mit dem genetisch identischen
Virus (Scott et al., 2006).
1.1.3 Diagnostik
Die Viruskultur ist nach wie vor Goldstandard in der Labordiagnostik zum
Nachweis von RSV (Respiratory Syncytial Viren, Robert-Koch-Institut, 2011).
Da diese sehr zeitaufwändig ist (zytopathische Effekte treten erst nach vier bis
sieben Tagen auf), haben sich in der klinischen Praxis sensitive und spezifische
immunologische Nachweistechniken wie der Enzyme-linked-immunosorbent
Assay (ELISA) oder Immunfluoreszenzverfahren durchgesetzt, die innerhalb
weniger Stunden virale Antigene mithilfe monoklonaler Antikörper in Nasen-
rachenspülwasser nachweisen. Breiten Einsatz finden auch Antigen-
schnelltests, die in etwa 20 Minuten das Ergebnis liefern und so für die
patientennahe Diagnostik geeignet sind. Auch ein Nachweis von RSV-
Genomfragmenten mittels Polymerase-Kettenreaktion (PCR) ist möglich. Die
PCR-Methode ist sehr spezifisch, schnell und hochsensitiv.

14
1.1.4 Therapie und Impfung
Bisher gibt es keine Kausaltherapie, so dass sich die Behandlung schwer RSV-
erkrankter Kinder auf die symptomatische Therapie beschränkt. Der Einsatz
inhalativer Betasympathomimetika wird kontrovers diskutiert, ein Therapie-
versuch sollte daher unter stationären Bedingungen durchgeführt werden,
ebenso wie die Inhalation mit Epinephrin. Neben einer Flüssigkeitssubstitution
sollte bei transcutan gemessener Sauerstoffsättigung unter 94% eine Sauer-
stoffgabe erfolgen, bei drohender respiratorischer Insuffizienz muss eine CPAP-
oder invasive Beatmung erwogen werden.
Eine routinemäßig durchgeführte antibiotische Therapie erbrachte keinen
Vorteil, ebenso wenig die Gabe inhalativer oder systemischer Glucocorticoide.
Die antivirale Therapie mit Ribavirin kann in Einzelfällen sinnvoll sein
(Respiratory Syncytial Viren, Robert-Koch-Institut, 2011), da sie sich als
wirksam bezüglich eines geringeren Sauerstoffbedarfs betroffener Säuglinge
und einer geringeren Dauer der invasiven Beatmung zeigte. Auf das Outcome
hatte die Behandlung jedoch keinen nachweisbaren Nutzen (Ventre and
Randolph, 2004).
Ein aktiver Impfstoff wird derzeit entwickelt. Als am besten geeignet erscheint
der rekombinante attenuierte Lebendimpfstoff rA2cp248/404/1030ΔSH, welcher
bereits in Phase IIa-Studien getestet wurde (Karron et al., 2005).
Zur allgemeinen Verfügung steht bislang jedoch lediglich der monoklonale
Antikörper Palivizumab gegen das F-Protein zur passiven Immunisierung. Der
guten Wirksamkeit [nach prophylaktischer Gabe von Palivizumab auf einer
neonatologischen Intensivstation während eines RSV-Ausbruchs wurden keine
weiteren Neuinfektionen mit dem RS-Virus beobachtet (O´Connell et al., 2011)]
stehen eine hohe Belastung des Kindes und ein enormer, jedoch auch
gerechtfertigter Kostenfaktor gegenüber (Resch et al., 2012), da während der
RSV-Saison monatlich eine gewichtsadaptierte Menge Palivizumab intra-
muskulär verabreicht wird.
Die derzeitigen Impfempfehlungen des Robert-Koch-Institutes erstrecken sich
auf ehemals Frühgeborene mit chronischer Lungenerkrankung infolge

15
Bronchopulmonaler Dysplasie (BPD) bis zum Alter von 24 Monaten, wenn sie in
den letzten sechs Monaten behandlungsbedürftig waren. Frühgeborene der 32.
bis 35. SSW ohne BPD sollten eine Prophylaxe erhalten, wenn zusätzliche
Risikofaktoren vorliegen (Respiratory Syncytial Viren, Robert-Koch-Institut,
2011).
Um die Pathogenese der Erkrankung noch besser verstehen zu können,
beschäftigt sich diese Arbeit mit der Frage, welche Mechanismen zu den
häufigen und klinisch symptomatischen Reinfektionen führen können und mit
welchen Mechanismen das RS-Virus die Immunabwehr des Wirtes unterläuft.
1.2 DIE IMMUNANTWORT
1.2.1 Die primäre Immunantwort: T-Zellen und Dendritische Zellen
Es zeigt sich, dass das RS-Virus Strategien entwickelt hat, die antiviralen
Mechanismen seines Wirtes zu durchbrechen. Um diese verstehen zu können,
muss man sich zunächst mit den Zellen der spezifischen Immunantwort
befassen: Die T-Lymphozyten nehmen eine Schlüsselstellung bei der
erworbenen, zellvermittelten Immunantwort ein.
Sie entstehen aus der pluripotenten Stammzelle des Knochenmarks, wandern
in den Thymus ein und durchwandern diesen von der Rinde her ins Mark. Dabei
differenzieren sie sich zu reifen T-Lymphozyten, von denen der größte Teil
(etwa 90%) abstirbt und nur ein Teil von etwa 10% den Thymus als reife und
immunologisch kompetente T-Zelle verlässt.
Solange diese reifen T-Zellen noch keinen Antigenkontakt hatten, gelten sie als
immunologisch naive T-Zellen. Erst der Kontakt mit einem Antigen führt zur
endgültigen Reifung und ermöglicht es der T-Zelle, an der Immunantwort
teilzunehmen.
Als professionellen, also hochspezialisierten, Antigenpräsentierenden Zellen
(APC) kommt den Dendritischen Zellen die Aufgabe zu, mit naiven T-Zellen in
Kontakt zu treten und deren Differenzierung auszulösen. Dendritische Zellen

16
sind die stärksten Stimulatoren naiver T-Zellen, und die Aktivierung naiver T-
Zellen durch Dendritische Zellen ist der erste entscheidende Schritt zur
Einleitung der spezifischen Immunantwort: Durch Antigenaufnahme und
-präsentation über den Major Histocompatibility Complex (MHC) interagieren
sie mit den T-Zellen über deren T-Zell-Rezeptor (TCR).
Bevor die Dendritische Zelle jedoch in der Lage ist, eine T-Zelle zu aktivieren,
muss sie einen Reifungsprozess durchlaufen. Die unreife Dendritische Zelle
besitzt nur eine geringe Anzahl antigenpräsentierender MHC-Moleküle und
keine co-stimulierenden Oberflächenmoleküle der B7-Immunglobulinfamilie.
Über Phagozytose und Pinozytose nehmen unreife Dendritische Zellen im
Verlauf ihrer Entwicklung Antigene in peripheren Geweben auf und wandern zu
benachbarten Lymphknoten. Dabei verlieren sie die Fähigkeit zur Antigen-
aufnahme und synthetisieren stattdessen MHC-Moleküle, mit denen sie Peptide
des Antigens an ihrer Oberfläche präsentieren. Außerdem exprimieren sie jetzt
co-stimulierende B7- und Adhäsionsmoleküle, was sie zu optimalen
Stimulatoren naiver T-Zellen macht.
Die RSV-Infektion führt zu einer vermehrten Anzahl von reifen Dendritischen
Zellen im Lungengewebe, welche die Primär-T-Zell-Antwort induzieren. Die
Beeinträchtigung der Funktion dieser Zellen ist ein starker viraler Mechanismus,
die Immunantwort zu umgehen oder abzuschwächen. Insbesondere scheint
hier die Fähigkeit von Bedeutung zu sein, die Interferon-γ-Produktion
herabzusetzen. IFN-γ hat zum einen einen direkt antiviralen Effekt, zum
anderen fördert es indirekt die Viruselimination durch Aktivierung natürlicher
Killerzellen, Makrophagen und zytotoxischer T-Zellen. So zeigen Lymphozyten
während der RSV-Bronchiolitis eine deutlich herabgesetzte IFN-γ-Produktion im
Vergleich zu Zellen der nichtinfizierten Kontrollgruppe. Dendritische Zellen
modulieren demnach T-Zell-Antworten wie IFN-γ-Ausschüttung. Weiterhin
konnten Schauer et al. zeigen, dass die erniedrigte IFN-γ-Produktion noch drei
Jahre später nachgewiesen werden konnte. Eine milde RSV-Infektion scheint
im Vergleich zur RSV-Bronchiolitis diese Effekte nicht in einem solchen Maße
zu erzielen (Schauer et al., 2004).
IFN-γ ist als ein die Virusreplikation limitierender Mediator essenziell bei der
Beeinflussung einer RSV-Infektion, und es konnte anhand eines in vitro-Modells

17
der Primärimmunantwort gezeigt werden, dass eine RSV-Infektion die Anzahl
IFN-γ-produzierender T-Zellen vermindert, wohingegen eine hohe Produktion
von IFN-γ vor RSV-vermittelter Erkrankung schützt.
Die verminderte IFN-γ-Produktion steht in Zusammenhang mit der Induktion
einer Apoptose in unreifen DCs, einem Anstieg der CD86-Expression und einer
verminderten Ausschüttung proinflammatorischer Zytokine (Bartz et al., 2003).
Ein lediglich leichter Anstieg der Expression von CD83 und HLA-DR, die als
Marker für reife DCs gelten, konnte nachgewiesen werden. Die Induktion der
Apoptose unreifer DCs könnte die effiziente T-Zell-Antwort verhindern und
somit ein Grund für die herabgesetzte zellvermittelte Immunität sein.
Diese Arbeit beschäftigt sich mit der Frage, ob RSV-infizierte, unreife
Dendritische Zellen in vitro vollständig ausreifen oder ob es Mechanismen gibt,
die die Ausreifung beeinträchtigen.
1.2.2 Das immunologische Gedächtnis: Zentrale Memory- und Effektor-
Memory-T-Zellen
Treten bisher unbekannte Pathogene in den Körper ein, reagiert das
Immunsystem zunächst verzögert wegen des Fehlens antigenspezifischer
Zellen. Unbekannte Antigene müssen zunächst ins lymphatische Gewebe
transportiert werden, wo sie eine primäre Immunantwort durch Interaktion
naiver CD4+ T-Lymphozyten und APCs auslösen. Dieser Primärantwort ent-
stammen Memory-Zellen, die sich wiederum in zwei unterschiedliche Zell-
formen unterteilen: zentrale Memory-T-Zellen, die vor nachfolgenden
Infektionen durch diesen Erreger schützen, und Effektor-Memory-T-Zellen,
welche die Ausbreitung des Pathogens verhindern. Etwa 5% aller naiven T-
Zellen differenzieren zu Memory-T-Zellen aus.
Mithilfe der unterschiedlichen Expression von CD45-Isoformen lassen sich
naive T-Zellen von Memory-T-Zellen unterscheiden: Naive T-Zellen exprimieren
die Isoform CD45RA, Memory-T-Zellen die Isoform CD45R0, die einer
Spleißvariante des CD45RA entspricht.

18
Das Glykoprotein CD45, auch Leukocyte Common Antigen genannt, findet man
auf allen kernhaltigen, blutbildenden Zellen; es interagiert mit dem T-Zell-
Rezeptor (Trowbridge, 1991). Seine drei Domänen sind die extrazelluläre, die
transmembranöse und die intrazelluläre bzw. zytoplasmatische Domäne
(Fischer et al., 1991, Trowbridge et al., 1991). Die extrazelluläre Domäne kann
durch alternatives Spleißen, Glycosylierung oder Sulfatierung vielfältig
prozessiert werden, so dass eine große Bandbreite an möglichen
Ligandenrezeptoren entsteht (Cyster et al, 1994, Trowbridge et al, 1992).
CD45RA gilt als Marker für ruhende, naive T-Zellen, wohingegen CD45R0 die
Memory-T-Zellen charakterisiert. Die Assoziation zum T-Zell-Rezeptor ist beim
RA-Typ eher gering und beim R0-Typ stark. Es wurden bisher einige CD45R-
Isoformen identifiziert. Diese Isoformen entstehen durch alternatives Spleißen
von Exons der Messenger-RNA (mRNA).
Unter Einbeziehen des Chemokinrezeptors CCR7 und der Oberflächenmarker
CD45RA bzw. CD45R0 lassen sich nicht nur naive von Memory-T-Zellen
unterscheiden, es lassen sich ebenfalls noch die Effektor-Memory-T-Zellen von
den zentralen Memory-T-Zellen abgrenzen (Sallusto et al., 1999).
Somit ergeben sich drei Typen von T-Zellen: Naive T-Zellen (CD45RA+,
CCR7+), zentrale Memory-T-Zellen (CD45R0+, CCR7+) und Effektor-Memory-T-
Zellen (CD45R0+, CCR7-). Alle drei Untergruppen übernehmen unterschiedliche
Aufgaben bei der zellulären Immunabwehr.
Insbesondere auf epithelialen Oberflächen findet man Effektor-Memory-T-
Zellen, welche die Ausbreitung des Pathogens verhindern, da hier die Haupt-
eintrittspforte pathogener Erreger ist. Sie produzieren sehr rasch Effektor-
zytokine wie Interferon γ (IFN-γ), Interleukin 4 (IL-4) und Interleukin 5 (IL-5).
Zentrale Memory-T-Zellen exprimieren kein IFN-γ und kein IL-4, dafür aber
hohe Mengen an Interleukin 2 (IL-2) (Mackay, 1999).
CCR7+ zentrale Memory-T-Zellen können sich in die CCR7- Effektor-Memory-T-
Zellen differenzieren, wenn sie durch ein Antigen stimuliert werden. Beide
Zellgruppen unterscheiden sich in ihrer Fähigkeit, L-Selektin zu exprimieren,
was - neben den ebenfalls CCR7+ naiven T-Zellen - nur den zentralen Memory-
Zellen möglich ist. L-Selektin befähigt die Zellen, in Lymphknoten einzutreten.

19
Nur das Zusammenspiel zentraler mit Effektor-Memory-T-Zellen ermöglicht eine
kompetente und schnelle Immunantwort auf bereits bekannte Antigene
(Mackay, 1993). Während Effektor-Memory-T-Zellen eine Abwehr bereits an
oberflächlichen epithelialen Geweben, also der Haupteintrittspforte für Erreger,
darstellen, können zentrale Memory-T-Zellen nach Kontakt mit dem Antigen in
den Lymphknoten zu Effektor-Memory-T-Zellen differenzieren. Nur so kann eine
optimale Immunantwort generiert werden.
Da bekannt ist, dass bereits kurze Zeit nach durchlebter RSV-Infektion eine Re-
infektion mit dem genetisch identischen Virus auftreten kann, stellte sich uns die
Frage, ob sich dies auf die fehlende Ausbildung eines immunologischen
Gedächtnisses zurückführen lässt und somit eine unzureichende immuno-
logische Sekundärantwort der frühzeitigen Reinfektion zugrunde liegt. In
unserer Arbeit unterscheiden wir zwischen den Isoformen CD45RA und
CD45R0, also zwischen naivem und Memory-T-Zell-Typ, wobei nicht weiter
zwischen zentralen Memory- und Effektor-Memory-T-Zellen differenziert wurde.
1.3 OBERFLÄCHENMOLEKÜLE ANTIGENPRÄSENTIERENDER ZELLEN
1.3.1 Antigenpräsentation durch Humane Leukozytenantigene
Humane Leukozytenantigene (HLA) sind Oberflächenmoleküle, die auf dem
kurzen Arm von Chromosom 6 codiert werden. Diesen Genort bezeichnet man
als Major Histocompatibility Complex (MHC). Man unterscheidet die MHC-
Klassen Ia (HLA-A, -B, -C), Ib (HLA-G, -E, -H, -CD1 u. a.) und II (HLA-DR, -DP,
-DQ), die sich in Struktur, Funktion und Zellverteilungsmuster unterscheiden
(Pichler und Peter, 2001).
HLA-Klasse I-Moleküle findet man auf allen kernhaltigen Zellen in unterschied-
licher Dichte. Fehlt die HLA-Klasse I-Expression oder ist sie deutlich reduziert,
kommt es zur Aktivierung Natürlicher Killerzellen (NK).
HLA-Klasse II-Moleküle werden insbesondere auf professionellen Antigen-
präsentierenden Zellen exprimiert. Diese nehmen Antigene aus der Umgebung

20
auf und präsentieren sie nach Prozessierung auf ihrer Oberfläche. Daher
werden mittels MHC-Klasse II-Molekülen hauptsächlich Peptide aus Proteinen
präsentiert, die mittels Phagozytose, rezeptorvermittelter Endozytose oder
Pinozytose aufgenommen wurden.
HLA-DR (Genlocus 6p21.31) findet man auf der Oberfläche von Makrophagen,
B-Zellen und Dendritischen Zellen. T-Zellen binden mit ihrem T-Zell-Rezeptor
an von HLA-Oberflächenmolekülen präsentierten Antigenen.
Strukturell handelt es sich bei HLA-DR um ein Heterodimer, bestehend aus
einer α- und einer β-Kette, die in der Plasmamembran verankert sind. Jede
Untereinheit besteht wiederum aus zwei extrazellulären Domänen, einer
membranverankerten Domäne und einem zytoplasmatischen Teil.
Bei der Immunantwort spielt das HLA-DR eine wichtige Rolle in der Antigen-
präsentation. Des Weiteren kann HLA-DR als Marker einer Immunstimulation
dienen, da häufig die gesteigerte Anwesenheit von HLA-DR auf der
Zelloberfläche die Antwort auf eine Stimulation darstellt.
MHC-Klasse II-Moleküle werden von Antigenpräsentierenden Zellen an deren
Oberfläche präsentiert und von CD4+ T-Helferzellen erkannt. MHC II-Moleküle
nehmen Peptide mittels Endozytose auf und bilden aus diesen T-Zell-Rezeptor-
Liganden. In den Lysosomen unreifer DCs werden Antigene prozessiert und in
Peptid-MHC II-Komplexe umgewandelt, wobei die Reifung Dendritischer Zellen
induziert wird. Diese Komplexe migrieren dann an die Zelloberfläche. Hier
spielen sie eine wichtige Rolle als Liganden für den T-Zell-Rezeptor.
Ein entscheidender Schritt für die T-Zell-Immunität ist die Antigenpräsentation
durch MHC-Produkte auf DCs.
In unreifen DCs werden jedoch weder Peptide noch Proteine in Peptid-MHC II-
Komplexe umgebaut, solange kein Reifungsstimulus wie beispielsweise
Lipopolysaccharid (LPS) anwesend ist (Turley et al., 2000). Sobald dieser
Stimulus hinzugegeben wird, wandern die Komplexe zunächst ins lysosomale
Kompartiment und von dort aus über das Endoplasmatische Retikulum (ER) an
die Plasmamembran. Hierbei durchlaufen die DCs ihren Reifungsprozess über
eine intermediäre Form hin zu reifen DCs. Die DC-Reifung ist demnach
charakterisiert durch eine deutliche Hochregulation der Oberflächenexpression
von MHC II und co-stimulierenden Molekülen.

21
Dendritische Zellen gehören zu den potentesten APCs. Ihre immunologische
Fähigkeit, naive T-Zellen zu stimulieren, ist unübertroffen. Hierzu bedienen sie
sich der Fähigkeit, Fremdprotein in Peptid-MHC II-Komplexe umzubauen und
an ihrer Zelloberfläche zu präsentieren. Als Reifemarker kann HLA-DR neben
CD83 für reife Dendritische Zellen genutzt werden.
Wir nutzten das Oberflächenmolekül HLA-DR zum einen als Marker der
Immunstimulation und zum anderen als Marker reifer Dendritischer Zellen.
1.3.2 CD80 und CD86 als Co-Stimulatoren Antigenpräsentierender Zellen
Um Lymphozyten zu aktivieren, muss das Antigen über MHC-Klasse II-
Moleküle präsentiert werden. Für die Aktivierung ist jedoch noch ein weiteres
co-stimulierendes Signal notwendig. Wird T-Zellen ein MHC-Antigen-Komplex
ohne co-stimulierende Moleküle präsentiert, können sie eine Toleranz-
entwicklung für das Antigen zeigen. CD80 und CD86 gehören zu den
wichtigsten co-stimulierenden Molekülen.
Die Abkürzung CD steht für cluster of differentiation. Es handelt sich um
Oberflächenmoleküle verschiedener Zellen, die zum Teil zellspezifisch sind.
CD80 und CD86 gehören einer Immunglobulin-Familie an, der so genannten
B7-Familie (June et al., 1994). Beide spielen eine Rolle bei der Ausbildung von
Immuntoleranz oder der Abstoßung eigener (Autoimmunreaktion) und fremder
Gewebe als co-stimulierende Faktoren bestimmter Antigenpräsentierender
Zellen.
Als „professionelle“ Antigenpräsentierende Zellen werden diejenigen Zellen
bezeichnet, die hohe Mengen CD80 und CD86 auf ihrer Zelloberfläche
präsentieren. Sie sind starke Stimulatoren der T-Zell-Aktivierung. Zu den
professionellen APCs gehören aktivierte B-Zellen, Makrophagen und reife
Dendritische Zellen.
„Nicht-professionelle“ Antigenpräsentierende Zellen tragen deutlich weniger
CD80- und CD86-Moleküle, sie exprimieren hingegen bevorzugt MHC-
Moleküle. Zu ihnen gehören die ruhenden B-Zellen und unreife Dendritische

22
Zellen. Professionelle und nicht-professionelle APCs unterscheiden sich
demnach in der Stärke ihrer Antigen-Präsentation (Lu et al., 1997).
CD86 findet man auf reifen Dendritischen Zellen, Monozyten und Makrophagen.
Bei Aktivierung wird es schnell (innerhalb von Stunden) exprimiert und
hochreguliert. Daher spielt es in der Initialphase der Immunantwort die
entscheidende Rolle.
CD80 findet man ebenfalls auf reifen Dendritischen Zellen, Makrophagen und
Monozyten. Es wird jedoch während der Immunantwort deutlich langsamer
(innerhalb von Tagen) exprimiert und auch langsamer hochreguliert. Demnach
kommt diesem Molekül die herausragende Rolle bei der Aufrechterhaltung einer
Immunantwort, der chronischen Phase, zu (Inaba et al., 1995, Hathcock et al.,
1994).
Beide Moleküle sind als Co-Stimulatoren gemeinsam bei der Toleranz-
entwicklung Eigen- und Fremdgeweben gegenüber beteiligt. Anhand von
Autoimmun-Krankheitsmodellen oder der Transplantatforschung konnte man
ihre Rolle besser verstehen (Khoury et al., 1995, Charlton et al., 1994). Die Co-
Stimulation von T-Zellen durch CD80/CD86 stellt einen entscheidenden Schritt
bei der Immunantwort dar. Blockiert man die CD80/CD86-Co-Stimulation, so
führt dies zu einer Blockade der T-Zell-Aktivierung und damit zur
Immuntoleranz, da Antigene, die ohne co-stimulierende Signale präsentiert
werden, vom Immunsystem als harmlos registriert werden. Lu et al. zeigten,
dass die CD80/CD86-Blockade bei Autoimmunerkrankungen zu einer Langzeit-
T-Zell-Anergie führte, die das Nichtreagieren auf ein Antigen bei herabgesetzter
Immunlage beschreibt und als eine Form der T-Zell-Toleranz angesehen wird
(Lu et al., 1997).
Das von Matzinger entworfene „danger-Modell“ erklärt den Zusammenhang von
T-Zell-Aktivierung und Toleranzentwicklung folgendermaßen: Nur wenn ein
Antigen durch professionelle, co-stimulierte APCs präsentiert wird, wird dieses
Antigen als Gefahrsignal vom Immunsystem klar erkannt, und die T-Zell-
Aktivierung wird initiiert. Im Gegensatz hierzu können Antigene, die ohne co-
stimulierende Faktoren präsentiert werden, keine ausreichende Immunantwort
auslösen und führen so zur Toleranzentwicklung (Matzinger, 1994).

23
Wir nutzten den Nachweis des Moleküls CD86 als Marker der primären
Immunantwort, da dieser Co-Stimulator nach Aktivierung auf der Zelloberfläche
Dendritischer Zellen hochreguliert wird und zur T-Zell-Aktivierung führt.
1.3.3 CD83 als Marker reifer Dendritischer Zellen
CD83 ist ein 45 kDa-Glykoprotein und gehört zur Ig-Superfamilie. Es wird unter
anderem von reifen Dendritischen Zellen, Langerhans-Zellen, B- und T-Zellen
exprimiert. Als Marker wird es für reife Dendritische Zellen genutzt.
Zudem scheint es auch eine wichtige Rolle bei der Immunantwort zu spielen.
So konnte gezeigt werden, dass die lösliche Form des humanen CD83
(hCD83ext) die von Dendritischen Zellen vermittelte T-Zell-Proliferation
dosisabhängig inhibiert. Diese lösliche Form konnte auch in menschlichen
Seren nachgewiesen werden; sie wird von aktivierten Dendritischen Zellen und
B-Zellen freigesetzt (Hock et al., 2001).
Ebenfalls inhibiert hCD83ext die Reifung Dendritischer Zellen. Lechmann et al.
zeigten, dass mit hCD83ext inkubierte unreife Dendritische Zellen nicht
vollständig ausreiften; es kam zu einer Herabregulation von CD80, einem co-
stimulierenden Molekül, und von CD83 selbst (Lechmann et al., 2001). Diese
Reifungsblockade könnte einen Mechanismus darstellen, Immunantworten zu
beeinflussen, indem unter Stimulationsbedingungen das membrangebundene
CD83 an seinen Liganden auf Dendritischen Zellen bindet und somit ein Signal
auslöst, welches zur Aktivierung der Reifung Dendritischer Zellen führt. Bindet
jedoch lösliches CD83 an diesen Rezeptor, wird er blockiert, so dass das
Signal, das zur Ausreifung Dendritischer Zellen führt, nicht weiter transmittiert
werden kann.
Obwohl nur reife Dendritische Zellen CD83 exprimieren und CD83 somit als
Marker für diese ausgereiften Dendritischen Zellen gilt, binden auch unreife
Dendritische Zellen CD83 (Lechmann et al., 2001).
Details, die die Existenz, Expression und Struktur des CD83-Liganden näher
definieren, blieben bisher noch ungeklärt. Erstmals beschrieben wurde ein
möglicher CD83-Ligand von Cramer et al. im Jahr 2000. Die Arbeitsgruppe
nutzte das rekombinante Fusionsprotein mCD83-IgG1, erzeugt aus der

24
extrazellulären Domäne von murinem CD83 (mCD83) und der Fc-Domäne von
humanem Immunglobulin G1 (IgG1). Dieses dimere Molekül war in der Lage,
eine Subpopulation muriner B-Zellen zu binden, nicht jedoch CD4-positive
Zellen (Cramer et al., 2000).
Lechmann et al. identifizierten 2001 einen möglichen CD83-Liganden auf reifen
und unreifen menschlichen DCs. Ähnlich wie Cramer et al. nutzten sie ein
Fusionsmolekül aus humanem CD83 und IgG1 (hCD83-IgG1) und konnten
zeigen, dass dieses Molekül dosisabhängig eine DC-Bindung vermittelt.
Scholler et al. nutzten das gleiche Fusionsprotein und konnten eine schwache
Bindung von hCD83-IgG1 an CD8+ T-Zellen sowie an menschliche Monozyten
nachweisen, wohingegen B-Zellen, CD56+ Natürliche Killerzellen oder Granu-
lozyten keine Bindung eingingen (Scholler et al., 2001).
Obwohl es deutliche Hinweise auf die Existenz und Funktion eines CD83-
Liganden gibt, bleibt eine genauere Beschreibung dieses Liganden weiterhin
abzuwarten.
Einen weiteren Hinweis auf eine funktionelle Rolle von CD83 lieferte die
Beobachtung, dass in viral infizierten Zellen die CD83-Expression deutlich
herunterreguliert wurde; dies wiederum führte zu einer Inhibition der Reifung
Dendritischer Zellen. Kruse et al. infizierten DCs mit Herpes simplex Virus Typ 1
und analysierten sie phänotypisch und strukturell: Zehn Stunden nach Infektion
kam es zu einer starken Herabregulation von CD83 an der Zelloberfläche reifer
DCs. Da die fehlende CD83-Oberflächenexpression nicht durch die Inhibition
der CD83-mRNA-Synthese hervorgerufen wurde (es konnte kein Rückgang der
CD83-mRNA-Level beobachtet werden), vermutete man, dass ein post-
transkriptionelles Ereignis für die Herabmodulation von CD83 und damit auch
für die Reifung Dendritischer Zellen verantwortlich sein müsse: Es konnte
gezeigt werden, dass CD83 in Lysosomen abgebaut wird (Kruse et al., 2000).
Zusammengefasst lässt sich sagen, dass CD83 als Oberflächenmolekül reifer
Dendritischer Zellen eine wichtige Rolle in der DC-vermittelten Immunantwort
spielt. Lösliches CD83 wirkt immunsuppressiv (Scholler et al., 2002). Zudem
wird CD83 für die CD4+ T-Zell-Generierung benötigt (Fujimoto et al., 2002).

25
Uns stellte sich die Frage, wie sich die RSV-Infektion Dendritischer Zellen auf
die CD83-Generierung auswirkt.
1.4 POLY-IC: EIN SIMULATOR VIRALER INFEKTIONEN IN VITRO
UND SEINE BINDUNG AN DEN TOLL-LIKE-REZEPTOR 3
Poly-IC (Polyinosinic-polycytidylic acid) ist ein Molekül, das mit dem Toll-like-
Rezeptor 3 (TLR3) interagiert. Dieser Rezeptor wird in B-Zellen und Dendri-
tischen Zellen exprimiert und geht eine Bindung mit doppelsträngiger RNA ein,
welche unter anderem von den meisten Viren während ihres Infektionszyklus
gebildet wird. Strukturell ähnelt Poly-IC doppelsträngiger RNA; diese synthe-
tische dsRNA wird als Immunmodulator angewandt, um virale Infektionen in
vitro zu simulieren.
Toll-like-Rezeptoren spielen als Teil des angeborenen Immunsystems eine
Rolle bei der Erkennung mikrobieller Erreger, der Induktion antimikrobieller
Gene und der Kontrolle der adaptiven Immunantwort. Sie unterstützen ebenfalls
die Kontrolle der DC-Reifung und der Differenzierung von T-Helferzellen
(Medzhitov, 2001).
Strukturell handelt es sich um transmembrane Rezeptoren, die charakterisiert
sind durch eine extrazelluläre, leucinreiche Wiederholungsdomäne [leucine-rich
repeat (LRR) domain] und eine intrazelluläre Toll-IL-1-Rezeptordomäne (TIR
domain). Sie sind eine Art „Mustererkennungsrezeptor” (pattern recognition
receptor, PRR) und erkennen Moleküle, die verschiedenen Pathogenen gemein
sind (pathogen-associated molecular patterns, PAMPs), jedoch unterscheidbar
von Eigenmolekülen. Mit dem IL-1-Rezeptor bilden TLRs eine Superfamilie, die
„IL-1-Receptor/Toll-like Receptor Superfamily“, deren Mitglieder alle eine so
genannte Toll-IL-1-Rezeptor(TIR)-Domäne besitzen. Die TIR-Domäne gilt als
Protein-Protein-Interaktions-Modul und umfasst drei Untergruppen:
Proteine der ersten Subgruppe sind Interleukinrezeptoren, die von Makro-
phagen, Monozyten und Dendritischen Zellen gebildet werden. Alle tragen
extrazelluläre Immunglobulindomänen.

26
Proteine der zweiten Untergruppe sind klassische TLRs, die direkt oder indirekt
mikrobielle Moleküle binden.
Proteine der dritten Untergruppe besitzen zytosolische Adapterproteine und
vermitteln Signalwege von Proteinen der ersten und zweiten Subgruppe.
TLR-Liganden sind durchaus verschieden in Struktur und Ursprung, jedoch gibt
es auch Gemeinsamkeiten: Die meisten TLR-Liganden sind mikrobielle
Produkte, PAMPs, welche die Anwesenheit einer Infektion signalisieren. Zudem
können TLRs verschiedene, strukturell nicht verwandte Liganden erkennen.
Einige TLRs nutzen akzessorische Proteine, um Liganden zu identifizieren. Es
gibt Hinweise, dass Säugetier-TLRs ihre Liganden direkt binden und als PRRs
funktionieren.
TLR3 erkennt virale Doppelstrang-RNA und das synthetische Analogon Poly-IC.
Nach Bindung wird die Interferon-Ausschüttung induziert und mit ihr die
inflammatorische Zytokin- und Chemokin-Produktion sowie die DC-Reifung
(Matsumoto et al., 2008).
Im Gegensatz zu anderen Säugetier-TLRs besitzt der TLR3 eine strukturelle
Besonderheit: Er hat keinen Prolin-Rest an der sonst typischen Position. Dies
bedeutet, dass der TLR3-Signalweg sich möglicherweise ebenfalls von dem
anderer TLRs unterscheidet. Der TLR3 wird bevorzugt, jedoch nicht
ausschließlich, in Dendritischen Zellen exprimiert (Medzhitov, 2001).
Bindet dsRNA an den TLR3, wird eine dsRNA-abhängige Proteinkinase
aktiviert, die PKR. Der gleiche Effekt wird ausgelöst, wenn Poly-IC an den TLR3
bindet. Es können dann zwei verschiedene Signaltransduktionswege ein-
geschlagen werden: ein vom Adapterprotein MyD88-abhängiger Signalweg,
den alle TLRs nutzen (shared pathway), und ein MyD88-unabhängiger Signal-
weg, der von TLR4 und TLR3 getriggert wird (specific pathway). Dieser MyD88-
unabhängige Signalweg kann die DC-Reifung induzieren.
Poly-IC ermöglicht die TLR3-vermittelte Aktivierung von NFκB und einer MAP
(mitogen activated protein)-Kinase und induziert zudem die Ausschüttung
zytotoxischer Zytokine und die Interferon-Produktion MyD88-abhängig (Fortier
et al., 2004).

27
Außerdem kann Poly-IC die Aktivierung von NFκB und der MAP-Kinase
MyD88-unabhängig auslösen, womit die Reifung Dendritischer Zellen induziert
wird (Alexopoulou et al., 2001).
Verdijk at al. zeigten, dass die Behandlung unreifer Dendritischer Zellen mit
Poly-IC deren Reifung induziert und dieser reife Phänotyp hohe Level von HLA-
DR, CD86 und CD83 exprimiert (Verdijk et al., 1999).
Daher wählten wir in unseren Experimenten Poly-IC, einen Simulator viraler
Infektionen in vitro, als Positiv-Kontrolle im Vergleich zur Infektion unreifer
Dendritischer Zellen mit RSV aus.
1.5 SUPERANTIGENE
1.5.1 Der Aufbau des T-Zell-Rezeptors
Der T-Zell-Rezeptor (TCR) ist bei 95% aller T-Zellen ein Heterodimer aus einer
α- und einer β-Kette. Zur Rezeptorseite gehören neben nicht keimbahn-
codierten Aminosäuren auch fünf genetisch codierte, variable Elemente (Vα, Jα,
Vβ, Dβ, Jβ). Bei allen T-Zellen bildet der Oberflächenmarker CD3 einen
Komplex mit dem TCR, so dass am Rezeptor empfangene Signale durch
Aktivierung einer intrazellulären Proteinkinase verstärkt weitergeleitet werden
können.
Liganden der α- oder β-Untereinheit des TCRs sind an MHC gebundene
Peptide. Normalerweise wird die Spezifität der TCRs für an MHC gebundene
Antigene bestimmt durch alle variablen Elemente der α- und β-Kette. Die
Region Vβ, die als Bindungsstelle für Superantigene identifiziert werden konnte,
liegt an einem Teil der β-Kette.
1.5.2 Superantigene und ihre besondere Bindung an den TCR
„Superantigene“ haben sich experimentell als Simulatoren der Virusantigen-
Reaktion bewährt. Es sind Rezeptorliganden, die - unabhängig von den

28
anderen variablen Elementen beider Ketten - mit ihrer MHC-Kombination die-
jenigen T-Zellen stimulieren, die ein bestimmtes Vβ tragen. Es gibt sowohl
exogene als auch endogene Superantigene. Hierbei handelt es sich um hitze-
stabile, hydrophile Moleküle mit einem Molekulargewicht von 22 bis 30 kDa,
welche zudem auch relativ stabil gegenüber proteolytischer Spaltung sind.
Im Unterschied zu gewöhnlichen Antigenen werden Superantigene direkt von
T-Zellen erkannt, ohne zuvor über MHC-Moleküle präsentiert worden zu sein.
Gascoigne und Ames zeigten, dass Superantigene den T-Zell-Rezeptor und
MHC II-Moleküle praktisch miteinander „verschmelzen“ können und somit ein
Dauersignal in der T-Zelle auslösen (Gascoigne und Ames, 1991). Es kommt zu
einer starken, polyklonalen und antigenunspezifischen T-Zell-Aktivierung, die
jedoch Vβ-selektiv ist (Dellabona et al., 1990). Hierbei findet die Bindung des
TCRs mit dem Superantigen nicht an der typischen antigenspezifischen
Bindungsstelle im Innern des Moleküls statt, sondern außen am MHC II-
Komplex.
Die Bindungsstelle am TCR für Superantigene ist die Region Vβ.
Normalerweise ist eine Aktivierung des TCRs nur möglich durch Bindung an
den MHC I- oder II-Komplex der Antigenpräsentierenden Zelle, womit die
Superantigen-Bindung hier eine Sonderstellung einnimmt: Superantigene
müssen nicht erst durch APCs prozessiert und dann den T-Zellen mit dem
MHC-Komplex als Peptidfragmente präsentiert werden, sondern sie binden
direkt an den TCR der T-Zelle einerseits und den MHC II-Komplex der APC
andererseits (Fleischer und Schrezenmeier, 1988, White et al., 1989). Dies
erklärt die außergewöhnliche stimulative Potenz der Superantigene.
Es sind bisher etliche Vβ-Elemente beschrieben worden. Weil die Vβ-Region
des TCRs nicht die Variabilität des gesamten Rezeptors erreicht, werden durch
ein Vβ-spezifisches Superantigen jeweils mehrere Prozent (5% bis 20%) des
gesamten T-Zell-Pools aktiviert, wohingegen bei der antigenabhängigen T-Zell-
Aktivierung nur 0,0001% bis 0,01% der T-Zellen aktiviert werden (Choi et al.,
1989, Choi et al., 1990). Es resultieren somit eine starke T-Zell-Aktivierung mit
einer deutlichen Zytokinproduktion und eine Expansion von T-Zell-Klonen mit
spezifischen Vβ-Strukturen.
Nach zunächst extremer T-Zell-Proliferation kommt es zur Apoptose und T-Zell-
Anergie, was in vivo eine systemische Inflammation mit Symptomen wie Fieber,

29
Hypotension und Kreislaufdysregulation bis hin zum Schock auslösen kann
(Krakauer, 1999).
In unseren Versuchen setzten wir das Superantigen Toxic Shock Syndrome
Toxin-1 als Simulator der Virusantigen-Reaktion in vitro ein.
1.5.3 Aufbau und Bindungsspezifität des Toxic Shock Syndrome Toxins-1
TSST-1, das als Auslöser des Toxic Shock Syndromes bekannt ist, gehört zur
Gruppe der Superantigene und umfasst 194 Aminosäuren bei einem
Molekulargewicht von 22.049 Da. In seiner Sekundärstruktur weist es - wie
andere Superantigene auch - hauptsächlich β-Faltblattstrukturen sowie nur
wenige α-Helices und aperiodische Bereiche auf, was der Tertiärstruktur mit
einer zylindrischen oder Fassform eine große Kontaktfläche nach außen
ermöglicht (Singh et al., 1988). TSST-1 bindet einerseits an MHC-Klasse II-
Moleküle der Dendritischen Zellen, andererseits an die Vβ2-Kette des T-Zell-
Rezeptors.
Choi et al. zeigten, dass das Toxic Shock Syndrome Toxin in besonderem
Maße (45%) die Vβ2+ Untergruppe der T-Zellen stimuliert und zudem - in jedoch
nur geringem Maße (anteilig zwischen 8,2% und <1%) - Vβ1+ und Vβ3+ bis
Vβ20+ T-Zellen (Choi et al., 1989).
Auch die anderen untersuchten Toxine scheinen bevorzugte Vβ T-Zellen zu
stimulieren [Staphylococcus aureus Enterotoxin E (SEE) Vβ8; Staphylococcus
aureus Enterotoxin B (SEB) Vβ3 und Vβ17; Staphylococcus aureus
Enterotoxin C2 (SEC2) Vβ13.1 und Vβ17; Exfoliating toxin (ExT) Vβ2 (<15%)].
Jedoch fand sich neben TSST kein anderes Staphylokokkentoxin, das in
diesem hohen Maße von 45% ein Vβ so spezifisch stimulierte wie TSST das
Vβ2.
In unseren Versuchen verwendeten wir TSST-1 als TCR-Ligand, da die
Stimulation hochspezifisch ist und der TSST-1-bindende T-Zell-Rezeptor Vβ2
schnell mittels Durchflusszytometrie erkannt werden kann.

30
2. FRAGESTELLUNG
Nach wie vor stellt die RSV-Infektion eine häufige und zum Teil schwer
verlaufende Erkrankung insbesondere junger Säuglinge dar. Eine besondere
Risikogruppe bilden ehemalige Frühgeborene und Kinder mit Shuntvitien. Auf
diese beziehen sich die derzeitigen Impfempfehlungen des Robert-Koch-
Instituts. Die üblichen Immunmechanismen nach durchlaufener Infektion
scheinen für das RS-Virus nicht zu gelten: Es konnten nach überstandener
Erkrankung Reinfektionen mit dem genetisch identischen Virus nachgewiesen
werden.
Da sich in den letzten Jahrzehnten große Fortschritte in der Behandlung
Frühgeborener erzielen ließen, stellt sich immer mehr die Frage nach einer
suffizienten, kostengünstigen und patientenschonenden Immunisierung. Zur
Verfügung steht derzeit lediglich ein Passivimpfstoff, der in der RSV-Saison
monatlich injiziert werden muss. Dies stellt eine erhebliche Belastung für die
kleinen Patienten dar und ist zudem mit hohen Kosten verbunden.
Diese Arbeit beschäftigt sich daher mit der Frage, welche Mechanismen die
Immunantwort nach durchlaufener RSV-Infektion beeinflussen und das
immunologische Gedächtnis beeinträchtigen. Als immunologisches Modell
dienten RSV-infizierte Dendritische Zellen.
Zusammengefasst ergaben sich folgende Fragestellungen:
1. Führt die RSV-Infektion unreifer Dendritischer Zellen im in vitro-Modell zu
einer Reifung mit Ausbildung der Oberflächenmarker CD83 und HLA-DR
in der immunologischen Primärantwort und somit zu einer potenten und
zielgerichteten Immunantwort?
2. Kommt es zu einer immunologischen Sekundärantwort mit Ausbildung
von Memory-T-Zellen und somit zur Generierung eines immunologischen
Gedächtnisses, oder ist eine ausbleibende Bildung von Memory-T-Zellen
begünstigend für die frühzeitige Reinfektion?

31
3. MATERIAL UND METHODEN Diese Arbeit entstand im Rahmen des DFG Projekts „Modulation von
Dendritischen Zellen und T-Zellen durch das Respiratory Syncytial Virus in
vitro", an dem eine Gruppe von Doktoranden beteiligt war. Ausgangsmaterial
für die Untersuchungen waren aus Nabelschurblut angezüchtete Dendritische
Zellen und isolierte naive T-Zellen. Am Aufbau eines Pools an Zellen waren alle
Doktoranden gleichermaßen beteiligt.
Die Untersuchung der Expression von Markern auf Dendritischen Zellen und
die Ausreifung von Memory-T-Zellen ist alleiniger Gegenstand dieser Arbeit.
In dem Artikel „Differential response of human naive and memory/effector T
cells to dendritic cells infected by respiratory syncytial virus“ wurden die
Ergebnisse des Gesamtprojektes veröffentlicht (Rothoeft et al., 2007). Im
Folgenden stelle ich den Teil der Untersuchungen dar, den ich experimentell
betreut habe.
3.1 GEWINNUNG DENDRITISCHER ZELLEN AUS NABELSCHNUR-
BLUT 3.1.1 Gewinnung von Nabelschnurblut
Nabelschnurblut klinisch gesunder Neugeborener der geburtshilflichen
Abteilung der Augusta Krankenanstalt Bochum wurde mittels Punktion der
Umbilicalvene steril entnommen. Um eine mögliche präpartale Aktivierung oder
Unreife des Immunsystems zu vermeiden, waren Frühgeburtlichkeit, eine
bekannte mütterliche Hepatitis- oder HIV-Infektion sowie Schwangerschafts-
komplikationen Ausschlusskriterien. Zuvor waren die Eltern über Entnahme und
Verwendung des Blutes aufgeklärt worden und gaben ihr schriftliches
Einverständnis. Da es sich um Untersuchungen an von Menschen gewonnenen
Proben handelte, durchlief die Studie zudem die Ethikkommission der Ruhr-
Universität Bochum.

32
Verwendet wurden Proben, die bei Beginn der Weiterverarbeitung nicht älter als
zwölf Stunden waren und deren Blutmenge mindestens 30 ml betrug, um eine
ausreichende Menge an Zellen zu gewinnen.
3.1.2 Gewinnung mononukleärer Zellen mittels Dichtegradientenzentri-fugation
Das Nabelschnurblut wurde steril in einen 50 ml Falcon Tube gegeben. Diesem
waren bereits 7 ml einer gerinnungs- und keimhemmenden Pufferlösung aus
Citrat-Phosphat-Dextrose-Lösung, Hepes 0,02 M, Kanamycin 0,1 mg/ml und
Partricin 0,5 µg/ml zugesetzt worden.
Mittels Ficoll-Dichtegradientenzentrifugation wurden die Zellen des Gesamt-
blutes aufgetrennt und die mononukleären Zellen - neben Lymphozyten und
Monozyten gehören auch die hämatopoetischen Stammzellen zu dieser
Zellgruppe - aus dem Nabelschnurblut isoliert. Aufgrund ihrer größeren Dichte
trennen sich bei der Dichtegradientenzentrifugation die Erythrozyten sowie die
polymorphkernigen Leukozyten von den mononukleären Zellen ab. An der
Phasengrenze zwischen Plasma und Ficoll reichern sich die mononukleären
Zellen an und können dort abpipettiert werden. Das Nabelschnurblut wurde
hierzu mit PBS im Verhältnis 1:4 verdünnt und mit 15 ml Ficoll-Lösung
unterschichtet. Nach Zentrifugation bei 400 x g für 20 Minuten wurde der
Interphasenring abpipettiert und zweimal mit PBS bei 300 x g für 10 Minuten
gewaschen. Die Zellzahl wurde nach Anfärben mit Türkscher Lösung in der
Neubauer-Zählkammer ermittelt.
3.1.3 Gewinnung CD34+ Stammzellen mittels magnetischer Zellseparation
Von diesen mononukleären Zellen separierten wir die CD34+ Stammzellen
mittels einer Positivselektion durch anti-CD34-beladene MicroBeads und
MiniMACS-Separationssäulen nach Protokoll des Herstellers Miltenyi:
Die Zellen wurden hierzu in 300 µl Puffer je 108 Zellen aufgenommen und
- ebenfalls je 108 Zellen - mit 100 µl humanem IgG und 100 µl Hapten-

33
konjugierten CD34-Antikörpern für 15 Minuten bei 4°C bis 6°C inkubiert.
Danach wurden die Zellen bei 300 x g für 10 Minuten gewaschen, in 400 µl
Puffer je 108 Zellen aufgenommen und mit 100 µl je 108 Zellen mit gegen das
Hapten des CD34-Antikörpers gerichteten MicroBeads markiert, für 15 Minuten
bei 4°C bis 6°C inkubiert und erneut für 10 Minuten mit 300 x g gewaschen.
Nach Resuspension in 500 µl Puffer je 108 Zellen wurden die Zellen über eine
MiniMACS-Separationssäule nach Herstellervorschrift sortiert. Um eine
möglichst hohe Reinheit der Sortierung zu erzielen, wurde die CD34+-Fraktion
der ersten Sortierung ein zweites Mal über eine MiniMACS-Separationssäule
sortiert.
3.1.4 Einfrieren der CD34--Fraktion
Das Eluat der MiniMacs-Säulen enthielt alle anderen mononukleären Zellen
inklusive naiver T-Zellen. Bis zur weiteren Verwendung wurden diese
eingefroren.
Hierzu wurden die Zellkulturplatte für 10 Minuten auf Eis gestellt, die Zellen mit
einer Pipette gut gemischt und in ein 50 ml Falcon Tube geerntet. Danach
wurden die Zellen bei 300 x g für 10 Minuten zentrifugiert, der Überstand wurde
verworfen, das Pellet zur Stabilisierung in eine Gefrierpufferlösung (bestehend
aus 45% FCS, 44% RPMI-1640, 10% Dimethylsulfoxid und 1% Penicillin/
Streptomycin) resuspendiert und bei -80°C in einem KryoTube eingefroren.
Nach 24 Stunden wurden die Zellen bei -196°C in Flüssigstickstoff
kryokonserviert.
3.1.5 Anzucht Dendritischer Zellen
Die Anzucht Dendritischer Zellen erfolgte modifiziert nach der von Caux
beschriebenen Methode (Caux et al., 1992).
Aus den durch Positivselektion gewonnenen, CD34+ hämatopoetischen
Progenitorzellen wurde eine Probe entnommen, die Zellzahl nach Anfärben mit
Türkscher Lösung in der Fuchs-Rosenthal-Zählkammer ermittelt und in

34
endotoxinfreiem Kulturmedium eingestellt. Das Kulturmedium setzte sich
zusammen aus RPMI-1640, 10% hitzeinaktiviertem fetalem Kälberserum (FCS)
und 4% nährstoffreicher und keimhemmender Lösung. Die Kultivierung erfolgte
in 24-Loch-Platten in einer Dichte von 1,5 x 105 Zellen/ml.
Den Kulturen wurden 100 ng/ml GM-CSF, 2,5 ng/ml TNF-α und 100 ng/ml SCF
als Stimuli zugesetzt. Bei 37°C und 8% Kohlendioxid wurden die Zellen für 7
Tage kultiviert, wenn nötig die Kulturen geteilt und mit frischem Kulturmedium
versehen. Die Proliferation wurde mikroskopisch kontrolliert.
An Tag 7 wurden die Dendritischen Zellen geerntet und bis zur weiteren
Verwendung eingefroren.
Hierzu wurden die Zellkulturplatte für 10 Minuten auf Eis gestellt, die Zellen mit
einer Pipette gut gemischt und in ein 50 ml Falcon Tube geerntet. Danach
wurden die Zellen bei 300 x g für 10 Minuten zentrifugiert, der Überstand wurde
verworfen, das Pellet zur Stabilisierung in eine Gefrierpufferlösung (bestehend
aus 45% FCS, 44% RPMI-1640, 10% Dimethylsulfoxid und 1% Penicillin/
Streptomycin) resuspendiert und bei -80°C in einem KryoTube eingefroren.
Nach 24 Stunden wurden die Zellen bei -196°C in Flüssigstickstoff
kryokonserviert.
3.1.6 Auftauen Dendritischer Zellen
Zum Auftauen der Dendritischen Zellen am benötigten Zeitpunkt wurden die
KryoTubes in einem 37°C warmen Wasserbad geschwenkt, bis sich im
Randbereich eine Flüssigkeitsschicht bildete. Danach wurden die Zellen in
einen 50 ml Falcon Tube, der zuvor mit 40 ml PBS befüllt worden war, überführt
und bei 300 x g für 10 Minuten zentrifugiert. Der Überstand wurde verworfen,
danach wurden die Zellen noch zweimal mit PBS gewaschen und erneut für
jeweils 10 Minuten bei 300 x g zentrifugiert. Die Zellen wurden danach in 10 ml
Kulturmedium, bestehend aus RPMI, 10% FCS und 4% nährstoffreicher und
keimhemmender Lösung, resuspendiert. Die Bestimmung der Zellzahl erfolgte
in der Neubauer-Zählkammer nach Anfärbung mit Türkscher Lösung.

35
3.1.7 Restimulation Dendritischer Zellen an Tag 7
Nach dem Auftauen und Waschen wurden die Zellen auf eine Konzentration
von 4 x 105 je ml Kulturmedium aus RPMI, 10% FCS und 4% nährstoffreicher
und keimhemmender Lösung eingestellt und in eine 24-Loch-Platte gesetzt. Zur
Stimulation wurden 125 IU/ml IL-4, 2,5 ng/ml TNF-α und 100 ng/ml GM-CSF
hinzugegeben. Bei 37°C und 8% Kohlendioxid reiften die Zellen weiter aus,
wurden an Tag 8 mit GM-CSF und TNF-α nachstimuliert und bis zu Tag 12
nicht weiter geteilt.
An Tag 12 wurden die Dendritischen Zellen dann entweder mit RSV infiziert
oder mit Poly-IC stimuliert und 24 Stunden später in der Co-Kultur eingesetzt.
An Tag 14 wurden die Dendritischen Zellen - wie bereits in 3.1.4 und 3.1.5
beschrieben - geerntet und nach Anfärbung mit Türkscher Lösung und
Bestimmung der Zellzahl in der Neubauer-Zählkammer zunächst für 24 Stun-
den bei -80°C in KryoTubes eingefroren, danach erfolgte eine Langzeitlagerung
in Flüssigstickstoff bei -196°C.
3.2 ISOLIERUNG NAIVER T-ZELLEN UND CO-KULTUR MIT DENDRI-TISCHEN ZELLEN
3.2.1 Gewinnung naiver CD4+ T-Zellen
Zur Gewinnung naiver T-Zellen wurde erneut die magnetische Zellseparation
eingesetzt.
Diesmal wurden die Zellen mittels Negativselektion sortiert, das heißt, es
wurden alle Zellen mit Ausnahme der naiven CD4+ T-Zellen depletiert.
Die Zellen, die entfernt werden sollten, wurden mit paramagnetischen beads
der Firma Miltenyi wie im Herstellerprotokoll beschrieben markiert und über eine
AutoMACS Separationssäule über das Programm depleteS entfernt. Die
Negativfraktion beinhaltete dann die unmarkierten CD4+ T-Zellen.
Folgende Zellen mussten aus dem Eluat der Stammzellseparation depletiert
werden: CD14+ Monozyten, B-Lymphozyten, CD56+ NK-Zellen, CD8+ zyto-

36
toxische T-Zellen, CD45R0+ Memory-T-Zellen und eventuell noch vorhandene
Granulozytenverunreinigungen. Zudem wurden Glycophorin A (CD235a)-
positive Zellen (Erythrozyten und Erythrozytenvorläufer) depletiert, um eine
Störung der durchflusszytometrischen Messung durch Normoblasten zu
verhindern.
Das am Tag der CD34+-Stammzellsortierung gewonnene und kryokonservierte
CD34- Eluat (s. 3.1.4) enthielt alle Leukozyten des Nabelschnurbluts (inklusive
naiver T-Zellen) und Normoblasten.
Zur Gewinnung naiver CD4+ T-Zellen wurde das Eluat in einem 37°C warmen
Wasserbad vorsichtig aufgetaut und wie oben bereits beschrieben dreimal
gewaschen. Nach dem dritten Waschen wurde die Zellzahl nach Anfärben mit
Türkscher Lösung in der Neubauer-Zählkammer bestimmt.
Danach wurden die Zellen in einem Kleinstvolumen von 100 µl resuspendiert.
Je 107 Zellen wurden 20 µl der folgenden beads hinzugegeben: CD8, CD14,
CD56, HLA-DR, CD45R0, Glycophorin A und 10 µl je 107 Zellen CD16. Dieser
Ansatz wurde für 15 Minuten bei 4°C bis 6°C im Kühlschrank inkubiert, danach
mit 15 ml PBS für 10 Minuten bei 300 x g gewaschen. Nach dem Waschen
wurde der Überstand abgegossen, das Pellet in 1 ml PBS resuspendiert und im
AutoMACS (depleteS) nach Herstellerprotokoll sortiert.
Die CD4+ naiven T-Zellen (also die Negativfraktion) wurden nach der Sortierung
abzentrifugiert und in 1 ml Kulturmedium aufgenommen. Nach Anfärben mit
Türkscher Lösung wurde die Zellzahl in der Neubauer-Zählkammer bestimmt.
3.2.2 Co-Kulturen naiver T-Zellen mit Dendritischen Zellen als in vitro-
Modell der primären Immunantwort
Die Co-Kulturen naiver T-Zellen und Dendritischer Zellen wurden in 24-Loch-
Platten angesetzt. Hierzu wurden je ml Medium 106 CD4+ T-Zellen mit 105
Dendritischen Zellen co-kultiviert. Je well wurde 0,1 ng/ml TSST-1
hinzugegeben.

37
TSST-1 wurde verwendet, da der TSST-1-bindende T-Zell-Rezeptor Vβ2
schnell mittels Durchflusszytometrie erkannt werden kann. In T-Zell-
Monokulturen induziert TSST-1 keine T-Zell-Proliferation.
3.3 PRÄPARATION VON UND INFEKTION MIT RSV
Die Dendritischen Zellen wurden entweder unbehandelt belassen, mit RSV
infiziert oder mit 10 µg/ml Poly-IC für 24 Stunden stimuliert.
Die Präparation von und Infektion mit RSV erfolgte wie von Thurau beschrieben
(Thurau et al., 1998):
HEp-2-Wirtszellen wurden mit einem RS-Virenstamm (long strain) mit geringer
Infektionsmultiplizität (MOI 3) infiziert. Sobald die Infektion gelungen war,
wurden die Zellüberstände abgeerntet und mittels Ultraschall die zum Teil noch
intakten Zellen zerstört, um das Virus freizusetzen. Zellen und Debris wurden
abzentrifugiert, und der virushaltige Überstand wurde weiter gereinigt über
ultrazentrifugierte Sucrose-Gradienten. Die dadurch gewonnenen Virus-
präparationen wurden bei -80°C eingefroren.
Infektionstiter des Virenstammes wurden durch die Beimpfung von HEp-2-
Zellen in einer Verdünnungsreihe bestimmt. Das Virenwachstum wurde anhand
typischer zytopathischer Effekte und darauf folgender immunzytochemischer
Färbung der infizierten Zellen beurteilt.
Eine Testung auf Mycoplasmen mittels Polymerasekettenreaktion erfolgte
mithilfe eines Mycoplasmenkits (Venor GeM®) wie im Herstellerhandbuch
beschrieben.

38
3.4 ZELLBESTIMMUNG MITTELS DURCHFLUSSZYTOMETRIE
3.4.1 Das Prinzip der Durchflusszytometrie
Die Durchflusszytometrie erlaubt, verschiedene chemische und physikalische
Eigenschaften einzelner Zellen oder Zellpartikel simultan in einem Flüssigkeits-
strom zu messen. Die Analyse erfolgt dabei über unterschiedliche Fluoreszenz-
intensitäten und Streulichtsignale. Es können bis zu fünf Parameter gleichzeitig
gemessen werden: Vorwärtsstreulicht (FSC) als Maß für die Zellgröße,
Rechtwinkel- oder Seitwärtsstreulicht (SSC) als Maß für die Granularität und
Zelldichte und drei verschiedene Fluoreszenzfärbungen. Hierbei müssen die
Fluoreszenzfarbstoffe so gewählt werden, dass deren Emissionsspektrum dem
Photodetektionsbereich des Durchflusszytometers entspricht. Das verwendete
Gerät (FACScan, Firma BD) enthält einen 15 mW Argon-Ionen-Laser der
Wellenlänge 488 nm und drei Photodetektoren, die Licht der Wellenlänge
530 nm, 585 nm und >650 nm aufnehmen können. Dementsprechend sind
folgende Fluoreszenzfarbstoffe geeignet: Fluoresceinisothiocyanat FITC
[530 nm (Fl-1)], Phycoerythrin PE [585 nm (Fl-2)] und Tricolor [>650 nm (Fl-3)].
Die Daten wurden im „List-Mode“ anhand der fünf beschriebenen Parameter
aufgenommen und mittels CellQuest® Software analysiert.
3.4.2 Fluoreszenzfärbungen und durchflusszytometrische Messungen
Um die Expression der co-stimulierenden Moleküle HLA-DR, CD83 und CD86
der Dendritischen Zellen zu messen, wurden die kultivierten Zellen zweimal
gewaschen und die Konzentration auf 2 x 105 Zellen je ml in PBS eingestellt.
Bei 4°C bis 6°C wurden sie mit Phycoerythrin-gekennzeichneten Antikörpern
gegen die Oberflächenproteine (HLA-DR, CD83 und CD86) für 20 Minuten
inkubiert; im Anschluss wurde die Expression der co-stimulierenden Moleküle
durchflusszytometrisch gemessen.

39
Die Expression der T-Zell-Oberflächenmarker wurde nach Anfärben mit
anti-CD45RA-Fluoresceinisothiocyanat, anti-CD45R0-Phycoerythrin oder anti-
CD49a-Phycoerythrin und parallel mit anti-CD3-Tricolor oder TCR-Vβ2-
Phycoerythrin durchflusszytometrisch gemessen.
Um die Rate RSV-infizierter Dendritischer Zellen zu ermitteln, mussten die
Zellen zunächst mit 4% Paraformaldehyd fixiert und anschließend
permeabilisiert werden. Hierfür resuspendierten wir die Zellen in Saponinpuffer,
bestehend aus HBSS, 0,1% Saponin und 0,01 M HEPES-Puffer. Diese Zellen
mit der nun permeablen Zellmembran wurden mit biotinyliertem goat-anti-RSV-
Antikörper für 20 Minuten bei 4°C bis 6°C inkubiert, mit Saponinpuffer
gewaschen und dann mit FITC-markiertem Streptavidin für 20 Minuten bei 4°C
bis 6°C im Kühlschrank inkubiert. Nach erneutem Waschen mit Saponinpuffer
wurden die Zellen in 200 µl PBS für die durchflusszytometrische Analyse
resuspendiert.
3.5 DIE SYNGENE LANGZEIT-CO-KULTUR IM ZEITLICHEN ABLAUF
Co-kultiviert wurden naive CD4+ T-Zellen (aus 3.2.1) mit Dendritischen Zellen
(aus 3.1.7) desselben Spenders.
Die Co-Kultur lief von Tag -1 über Tag 0 bis Tag 22.
An Tag -1 wurden die Dendritischen Zellen aus 3.1.7 nach zwölftägiger Kultur
geerntet und bei 300 x g für 10 Minuten gewaschen. Der Überstand wurde
abgegossen und die Zellen in 3 ml DMEM ohne Zusätze resuspendiert.
Nach Anfärben mit Türkscher Lösung wurde die Zellzahl in der Neubauer-
Zählkammer bestimmt und eine Zellkonzentration von 5 x 104 Zellen je
300 µl DMEM eingestellt.
Um die Dendritischen Zellen mit RSV zu infizieren, inkubierten wir 300 µl der
Zellsuspension für drei Stunden mit gereinigtem Virus der Infektionsmultiplizität
von 20 (MOI 20). Bei 5 x 104 Dendritischen Zellen wurden also 1 x 106 Viren
zugegeben. Im Anschluss wurde das Virus entfernt und durch Kulturmedium

40
RPMI 1640 mit 10% FCS und 4% nährstoffreicher und keimhemmender Lösung
ersetzt. Co-Kulturen wurden 24 Stunden nach Infektion angesetzt. Vorab-
untersuchungen haben gezeigt, dass 85% der Dendritischen Zellen noch
lebensfähig waren.
Folgende drei Ansätze wurden bei 37°C und 8% Kohlendioxid für drei Stunden
im Brutschrank in einer 24-Loch-Platte kultiviert:
• 8 wells nicht-infizierte Kontrollansätze,
• 8 wells RSV long strain-infizierte Ansätze (MOI 20) und
• 8 wells Poly-IC-stimulierte Ansätze (10 µg/ml).
Nach drei Stunden wurden 200 µl Medium bzw. virushaltiger Überstand
vorsichtig abpipettiert und 500 µl Kulturmedium aus RPMI-1640, 10% FCS und
4% nährstoffreicher und keimhemmender Lösung hinzugegeben. Über Nacht
wurden die Ansätze im Brutschrank inkubiert. Alle Zellen, die an Tag -1 nicht
eingesetzt wurden, wurden in drei gleichgroße Portionen aufgeteilt und
eingefroren.
An Tag 0 erfolgte die oben beschriebene Sortierung der CD4+ T-Zellen aus
dem Eluat (s. 3.2.1). Die Zellkonzentration wurde auf 5 x 105 je 500 µl
eingestellt, und die T-Zellen wurden zu den an Tag -1 vorbereiteten
Dendritischen Zellen gegeben. Danach wurde in jedes well 0,1 ng/ml TSST-1
gegeben.
An Tag 1 wurde je ein well der drei Ansätze geerntet, und die Oberflächen-
marker CD45RA, CD45R0, CD49a und Vβ2 wurden mittels Durchfluss-
zytometrie - wie in 3.4.2 beschrieben - analysiert.
An Tag 3 wurde erneut je ein well der drei Ansätze geerntet, und die
Oberflächenmarker CD45RA, CD45R0, CD49a und Vβ2 wurden mittels
Durchflusszytometrie - wie in 3.4.2 beschrieben - analysiert.
An Tag 6 wurde in alle wells 50 IU/ml IL-2 (auch: TCGF, T-Cell Growth Factor)
hinzugegeben.

41
Zudem wurde eine Portion der an Tag -1 eingefrorenen Dendritischen Zellen
dreimal gewaschen. Nach Anfärben mit Türkscher Lösung wurde die Zellzahl in
der Neubauer-Zählkammer bestimmt und die Zellkonzentration auf 1 x 105 in
300 µl DMEM eingestellt.
Folgende drei Ansätze wurden bei 37°C und 8% Kohlendioxid für drei Stunden
im Brutschrank in einer 24-Loch-Platte kultiviert:
• 4 wells nicht-infizierte Kontrollansätze,
• 4 wells RSV long strain-infizierte Ansätze (MOI 20) und
• 4 wells Poly-IC-stimulierte Ansätze (10 µg/ml).
Nach drei Stunden wurden Medium beziehungsweise virushaltiger Überstand
vorsichtig abpipettiert und 1 ml Kulturmedium aus RPMI-1640, 10% FCS und
4% nährstoffreicher und keimhemmender Lösung hinzugegeben. Über Nacht
wurden die Ansätze im Brutschrank inkubiert.
An Tag 7 wurde erneut je ein well der drei Ansätze geerntet, und die
Oberflächenmarker CD45RA, CD45R0, CD49a und Vβ2 wurden mittels
Durchflusszytometrie - wie in 3.4.2 beschrieben - analysiert.
Zudem wurden die restlichen wells der Co-Kultur durch Zugabe Dendritischer
Zellen restimuliert. Hierzu wurden die an Tag 6 vorbereiteten Dendritischen
Zellen für 10 Minuten auf Eis gestellt. Währenddessen wurde das Zellwachstum
in der Co-Kultur mikroskopisch beurteilt. In der Regel hatten sich die Zellen
stark vermehrt, so dass die Co-Kultur zunächst gründlich mit einer Eppendorf-
Pipette gemischt wurde und 500 µl des Ansatzes verworfen wurden.
Nun konnten - nach gründlichem Durchmischen der auf Eis stehenden
Dendritischen Zellkultur - 500 µl Dendritische Zellen je well zur Co-Kultur
hinzugegeben werden. Zusätzlich wurde 0,1 ng/ml TSST-1 je well
hinzugegeben.
An Tag 10 wurde in alle wells 50 IU/ml IL-2 hinzugegeben.
An Tag 13 wurde erneut eine Portion der an Tag -1 eingefrorenen Dendri-
tischen Zellen aufgetaut und - wie an Tag 6 beschrieben - in je 4 wells
unbehandelt, RSV-infiziert oder Poly-IC-stimuliert kultiviert.

42
An Tag 14 wurde die Co-Kultur - wie an Tag 7 beschrieben - restimuliert.
An Tag 17 wurde in alle wells 50 IU/ml IL-2 hinzugegeben.
An Tag 20 wurde erneut eine Portion der an Tag -1 eingefrorenen Dendri-
tischen Zellen aufgetaut und - wie an Tag 6 beschrieben - in je 4 wells
unbehandelt, RSV-infiziert oder Poly-IC-stimuliert kultiviert.
An Tag 21 wurde die Co-Kultur - wie an Tag 7 beschrieben - restimuliert.
An Tag 22 wurde erneut je ein well der drei Ansätze geerntet, und die
Oberflächenmarker CD45RA, CD45R0, CD49a und Vβ2 wurden mittels
Durchflusszytometrie - wie in 3.4.2 beschrieben - analysiert.
3.6 DATENAUFNAHME UND -ANALYSE
Es wurde ein FACScan® Durchflusszytometer benutzt mit Filtereinrichtungen
für FITC-, PE- und Tricolor-Emissionen. 10.000 bis 50.000 Ereignisse wurden
im „List-Mode“ aufgenommen und mittels CellQuest® Software analysiert.
3.7 STATISTISCHE ANALYSE
Die Daten wurden mittels Varianzanalyse (ANOVA) und dem Bonferroni Test
für Nachtestung selektiver Daten-Sets analysiert.

43
3.8 EINGESETZTE MATERIALIEN UND HERSTELLERNACHWEIS
24-Loch-Platte Nunc, Wiesbaden, Deutschland
96-Loch-Platte Nunc, Wiesbaden, Deutschland
4% nährstoffreiche und keimhemmende
Lösung, bestehend aus
1 % Natriumpyruvat 100 mM, Biochrom AG, Berlin, Deutschland
1 % L-Glutamin 200 mM, Biochrom AG, Berlin, Deutschland
1 % nicht-essentiellen Aminosäuren Biochrom AG, Berlin, Deutschland
(L-Alanin, L-Asparagin, Glycin,
L-Glutamat, L-Prolin, L-Serin),
1% Penicillin 10.000 IE/ml und Biochrom AG, Berlin, Deutschland
1% Streptomycin 10.000 IE/ml Biochrom AG, Berlin, Deutschland
im Mischverhältnis 1 : 1 : 1 : ½ : ½
50 ml Falcon Tube Falcon, Heidelberg, Deutschland
anti-CD3-Tricolor Caltag, Hamburg, Deutschland
anti-CD45RA-Fluoresceinisothiocyanat BD Biosciences, Californien, USA
anti-CD45R0-Phycoerythrin BD Biosciences, Californien, USA
anti-CD49a-Phycoerythrin BD Biosciences, Californien, USA
AutoMACS Separationssäule Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD14 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD16 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD34 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD45R0 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD56 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
CD8 MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland

44
CD83-Phycoerythrin-Antikörper Beckman Coulter, Krefeld,
Deutschland
CD86-Phycoerythrin-Antikörper BD Biosciences, Heidelberg,
Deutschland
CellQuest® Software BD Biosciences, Californien, USA
Citrat-Phosphat-Dextrose-Lösung Sigma Aldrich, Taufkirchen,
Deutschland
Dimethylsulfoxid Sigma Aldrich, Taufkirchen,
Deutschland
DMEM (Dulbecco's Modified Eagle
Medium) Biochrom AG, Berlin, Deutschland
FACScan (Fluorescence Activated
Cell Sorting) BD Biosciences, Californien, USA
FCS (fetal calf serum) Biochrom AG, Berlin, Deutschland
Ficoll Seperating Solution Biochrom AG, Berlin, Deutschland
FITC-markiertes Streptavidin Caltag Laboratories, Burlingame,
Californien, USA
Glutamin 2 mM Biochrom AG, Berlin, Deutschland
Glycophorin A MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
GM-CSF (Granulocyte macrophage
colony-stimulating factor) Peprotech, Rocky Hill, New Jersey,
USA
goat-anti-RSV Antikörper Biodesign International, Saco, Maine,
USA
HBSS (Hank´s balanced salt solution) Biochrom AG, Berlin, Deutschland
HEp-2-Zellen ATCC-LGC Standards, Wesel,
Deutschland
HEPES-Puffer (Hydroxyethylpiperazinyl-
Ethansulfonsäure-Puffer) Biochrom AG, Berlin, Deutschland
HLA-DR MicroBeads Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
HLA-DR-Phycoerythrin-Antikörper BD Biosciences, Heidelberg,
Deutschland

45
IL-2 (Interleukin 2) Peprotech, Rocky Hill, New Jersey,
USA
IL-4 (Interleukin 4) Peprotech, Rocky Hill, New Jersey,
USA
Kanamycin 0,1 mg/ml Biochrom AG, Berlin, Deutschland
KryoTube Nunc, Wiesbaden, Deutschland
MiniMACS Separationssäulen Miltenyi Biotech GmbH, Bergisch
Gladbach, Deutschland
Natriumpyruvat 1 mM Biochrom AG, Berlin, Deutschland
Paraformaldehyd, Pulver Riedel-de Haën AG, Seelze,
Deutschland
Partricin 0,5 µg/ml Biochrom AG, Berlin, Deutschland
PBS (Phosphate Buffered Saline) Biochrom AG, Berlin, Deutschland
Penicillin 100 µU/ml Biochrom AG, Berlin, Deutschland
Poly-IC 10 µg/ml (polyinosinic:
polycytidylic acid) Sigma Aldrich, Taufkirchen,
Deutschland
RPMI-1640 Kulturmedium Biochrom AG, Berlin, Deutschland
RS-Virenstamm „long strain“ American Type Culture Collection,
Maryland, USA
Saponin Riedel-de Haën AG, Seelze,
Deutschland
SCF (stem cell factor) Peprotech, Rocky Hill, New Jersey,
USA
Streptomycin 100 µg/ml Biochrom AG, Berlin, Deutschland
TCR-Vβ2-Phycoerythrin Beckman Coulter, Krefeld,
Deutschland
TNF-α (Tumor-Nekrose-Faktor-Alpha) Peprotech, Rocky Hill, New Jersey,
USA
TSST-1 (toxic shock syndrome toxin-1) Toxin Technology, Sarasota, Florida,
USA
Türksche Lösung Fluka Chemika, Buchs, Schweiz
Venor GeM® Mycoplasmenkit Minerva Biolabs, Berlin, Deutschland
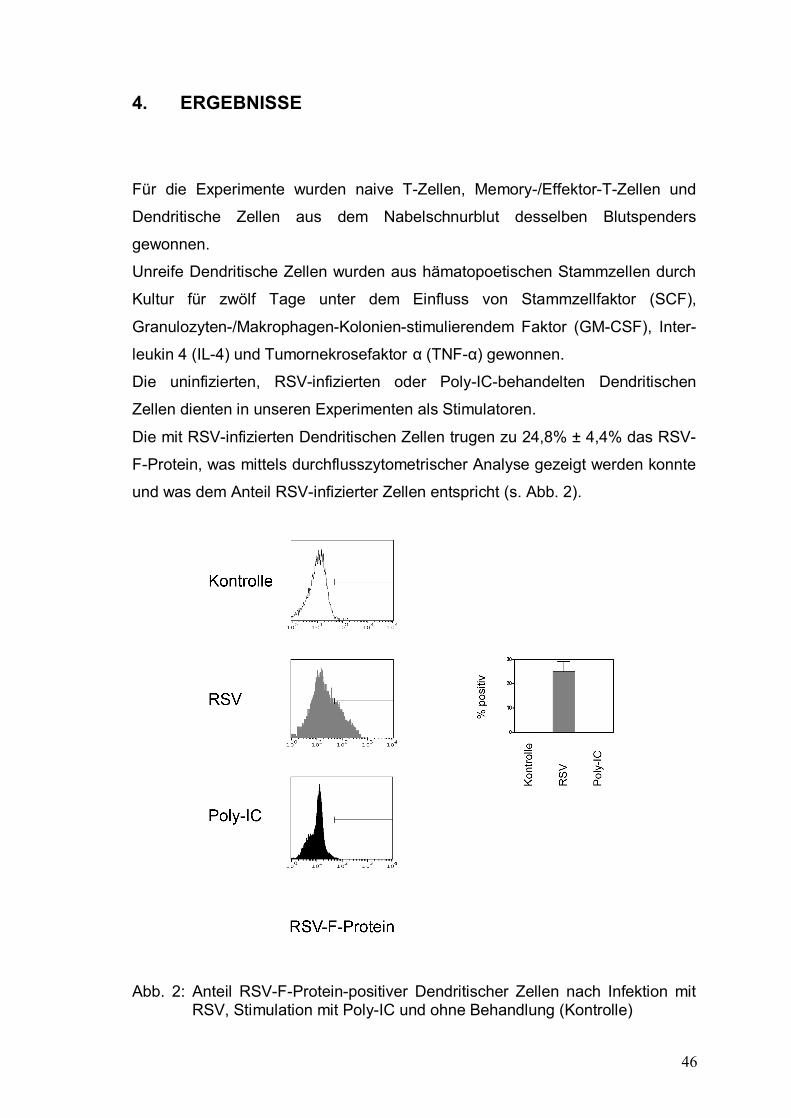
46
4. ERGEBNISSE
Für die Experimente wurden naive T-Zellen, Memory-/Effektor-T-Zellen und
Dendritische Zellen aus dem Nabelschnurblut desselben Blutspenders
gewonnen.
Unreife Dendritische Zellen wurden aus hämatopoetischen Stammzellen durch
Kultur für zwölf Tage unter dem Einfluss von Stammzellfaktor (SCF),
Granulozyten-/Makrophagen-Kolonien-stimulierendem Faktor (GM-CSF), Inter-
leukin 4 (IL-4) und Tumornekrosefaktor α (TNF-α) gewonnen.
Die uninfizierten, RSV-infizierten oder Poly-IC-behandelten Dendritischen
Zellen dienten in unseren Experimenten als Stimulatoren.
Die mit RSV-infizierten Dendritischen Zellen trugen zu 24,8% ± 4,4% das RSV-
F-Protein, was mittels durchflusszytometrischer Analyse gezeigt werden konnte
und was dem Anteil RSV-infizierter Zellen entspricht (s. Abb. 2).
Abb. 2: Anteil RSV-F-Protein-positiver Dendritischer Zellen nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle)

47
4.1 DIE RSV-INFEKTION FÜHRT LEDIGLICH ZU EINER SCHWACHEN
REIFUNG DENDRITISCHER ZELLEN IN VITRO
Zunächst untersuchten wir den Oberflächenphänotyp der Dendritischen Zellen.
Hierfür wurden unreife Dendritische Zellen entweder unbehandelt gelassen, mit
RSV (MOI 20) infiziert oder mit Poly-IC (10 µg/ml) stimuliert.
Danach wurde die Expression co-stimulierender Moleküle untersucht, indem die
Dendritischen Zellen mit spezifischen Antikörpern für HLA-DR, CD86 und CD83
inkubiert wurden.
Die durchflusszytometrische Analyse nach Fluoreszenzfärbung zeigte, dass die
Poly-IC-Behandlung zu einer starken Ausbildung aller drei untersuchten co-
stimulierenden Moleküle führte. Die mean fluorescence intensity (MFI) für HLA-
DR lag bei 1061 ± 223. 69,2% ± 10,2% der Zellen trugen den Oberflächen-
marker CD86, die MFI der CD83+ Zellen lag bei 423 ± 36. Dies bedeutet, dass
Poly-IC als „in vitro-Simulator“ einer Virusinfektion zu einer Expression des co-
stimulierenden Moleküls CD86 führt und eine Reifung Dendritischer Zellen
induziert, was der Anstieg von CD83 und HLA-DR zeigt.
In der Negativ-Kontrollgruppe der unbehandelten Dendritischen Zellen zeigte
HLA-DR eine MFI von 727 ± 211. 33,8% ± 9,6% der Zellen trugen den
Oberflächenmarker CD86, CD83 wies eine MFI von 221 ± 117 auf; verglichen
mit der Poly-IC-Gruppe kam es erwartungsgemäß zu einem deutlich geringeren
Effekt.
Die RSV-Infektion zeigte lediglich einen leichten Anstieg des co-stimulierenden
Moleküls CD86 (47,7% ± 2,8% CD86-positive Zellen). Auch wurde die Reifung
Dendritischer Zellen nur mäßig induziert: CD83 zeigte eine MFI von 283 ± 144
im Vergleich zu 423 ± 36 bei der Stimulation mit Poly-IC, es zeigte sich kein
Effekt beim HLA-DR (MFI 705 ± 106) im Vergleich zur Kontrollgruppe mit einer
MFI von 727 ± 211 (s. Abb. 3 bis 5).
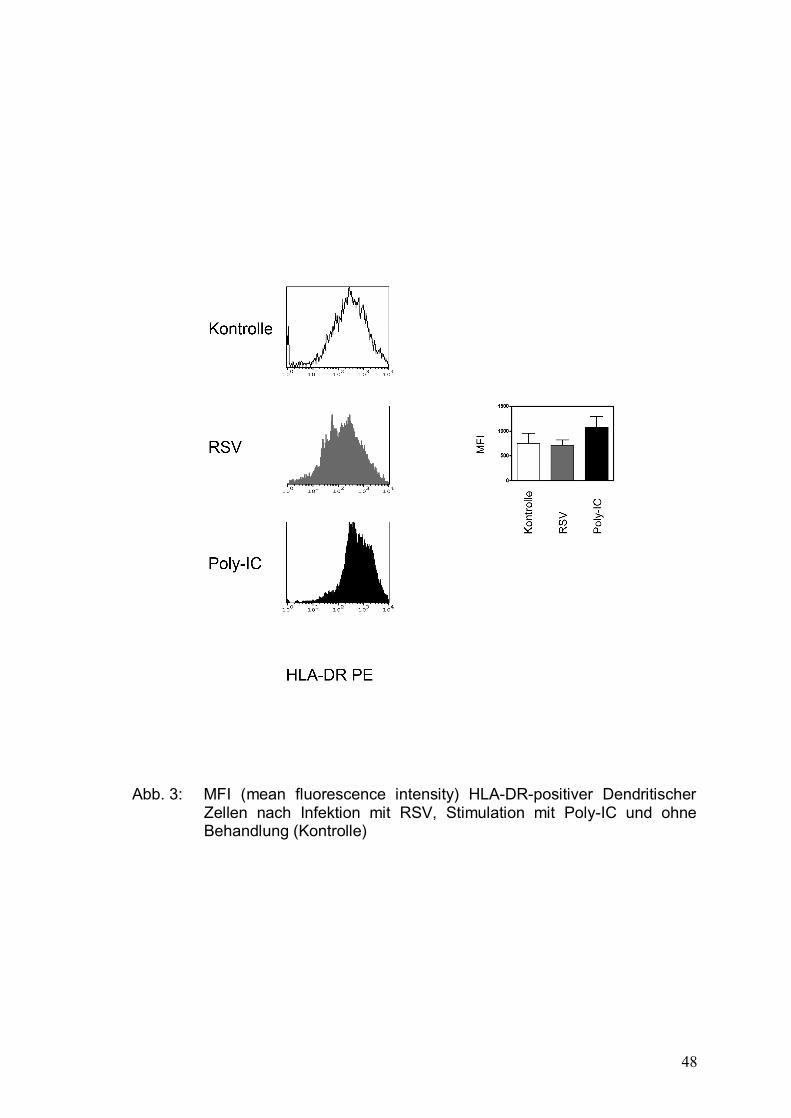
48
Abb. 3: MFI (mean fluorescence intensity) HLA-DR-positiver Dendritischer Zellen nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle)
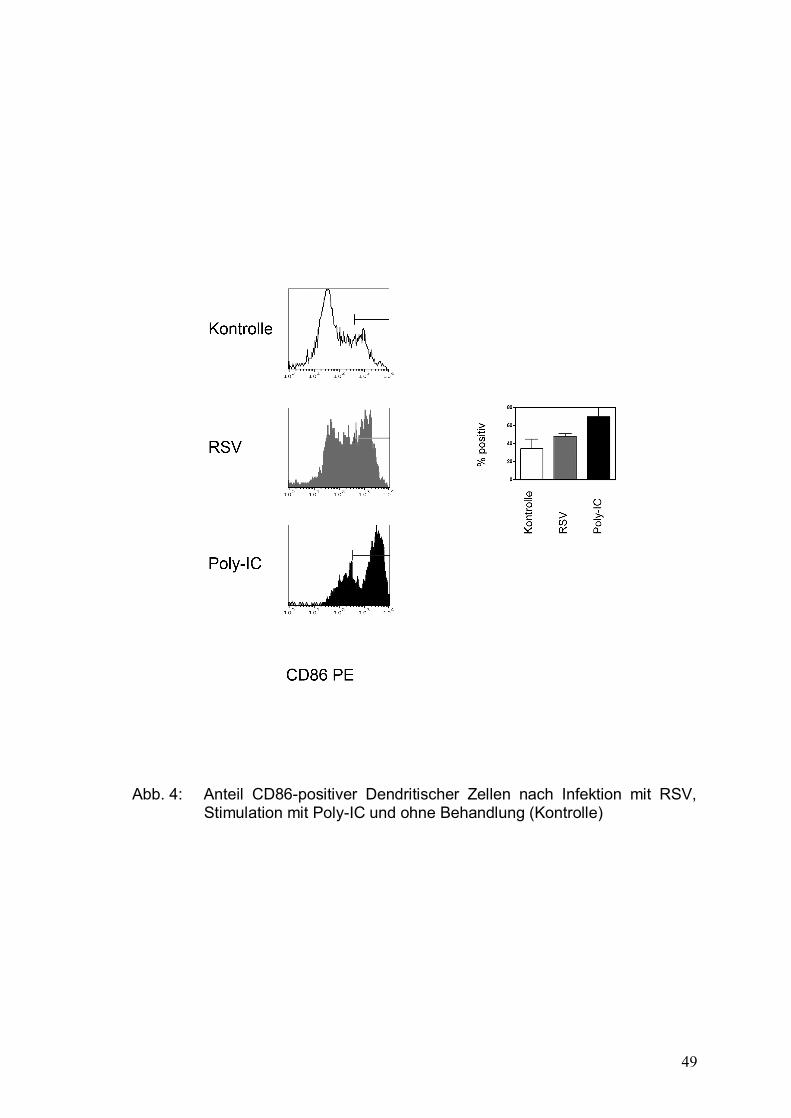
49
Abb. 4: Anteil CD86-positiver Dendritischer Zellen nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle)
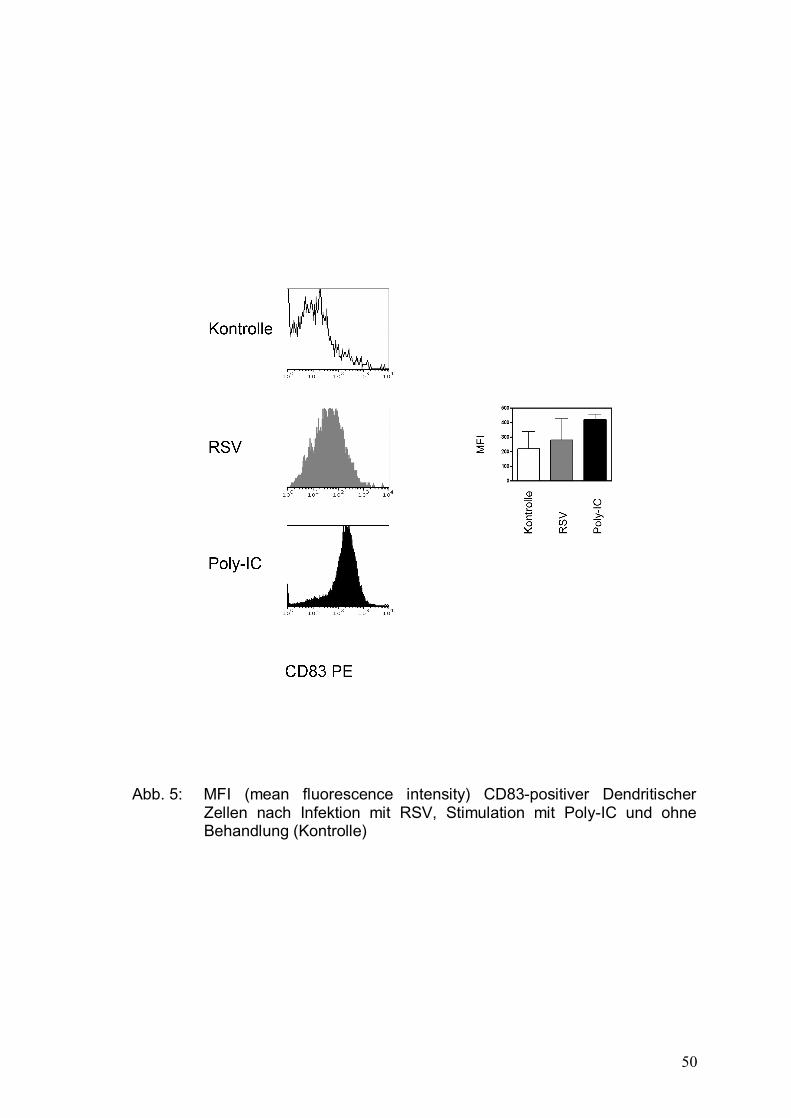
50
Abb. 5: MFI (mean fluorescence intensity) CD83-positiver Dendritischer Zellen nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle)

51
Somit zeigt sich zwar eine Reifung Dendritischer Zellen durch Kontakt zu RSV,
im Vergleich zur Poly-IC-Positiv-Kontrollgruppe sind der Effekt der Ausbildung
co-stimulierender Moleküle und die Ausreifung Dendritischer Zellen jedoch
deutlich abgeschwächt.
4.2 DIE RSV-INFEKTION HAT KEINEN NEGATIVEN EINFLUSS AUF DIE AUSBILDUNG VON MEMORY-T-ZELLEN BEI DER IMMUNO-LOGISCHEN SEKUNDÄRANTWORT
Um die Primärimmunantwort zu untersuchen, wurden naive T-Zellen,
Dendritische Zellen und 0,1 ng/ml TSST-1 co-inkubiert. TSST-1 aktiviert
bevorzugt T-Zellen, deren T-Zell-Rezeptoren eine Vβ2-Kette tragen. Wir nutzten
dies zur Simulation einer Antigen-Stimulation. Analysen wurden jeweils an den
Tagen 0, 1, 3 und 7 durchgeführt.
An den Tagen 0 und 1 waren fast 100% der Zellen CD45RA+, CD45R0- und
CD49a-. Demzufolge handelte es sich um naive, nicht-aktivierte T-Zellen. Etwa
5% der Zellen waren Vβ2+.
Zwischen Tag 1 und 3 sank der Anteil der CD45RA+ Zellen auf etwa 20% ab,
etwa 75% waren CD45R0+ und um 20% CD49a+. Der Anteil Vβ2-tragender
Zellen stieg auf etwa 13%.
Am 7. Tag waren annähernd 100% der Zellen CD45R0+ und CD45RA-. Sie
hatten somit einen Memory-/Effektor-Memory-T-Zell-Phänotyp angenommen.
Etwa 20% trugen zudem den Oberflächenmarker CD49a. 50% trugen eine Vβ2-
Kette. Es gab zu keinem Zeitpunkt signifikante Unterschiede im Vergleich der
RSV-Gruppe zur Kontroll- und zur Poly-IC-Gruppe.
An Tag 7, 14 und 21 wurden naive T-Zellen erneut mit nichtinfizierten, RSV-
infizierten oder Poly-IC-behandelten Dendritischen Zellen restimuliert, um auch
die Memory-/Effektor-Memory-Phase untersuchen zu können, deren Analyse an
Tag 22 erfolgte.
Hier zeigte sich, dass weiterhin 0% der Zellen CD45RA+ waren. Etwa 85%
trugen den Oberflächenmarker CD45R0 und etwa ein Viertel waren CD49a+.

52
Alle Zellen trugen eine Vβ2-Kette. Auch hier gab es keine signifikanten Unter-
schiede zwischen den einzelnen untersuchten Gruppen (s. Abb. 6 bis 9).
Dies zeigt, dass die T-Zellen ihren naiven CD45RA+ Charakter ablegten und
einen CD45R0+/CD49a+ Memory-/Effektor-Memory-T-Zell-Phänotyp akquirier-
ten, unabhängig davon, ob sie mit RSV-infizierten, Poly-IC-behandelten oder
nichtinfizierten Dendritischen Zellen kultiviert worden waren.
Demzufolge beeinflusst die RSV-Infektion die Bildung eines immunologischen
Gedächtnisses nicht, da trotz der Infektion durch das RS-Virus T-Zellen ihren
naiven Phänotyp ablegten und einen Memory-/Effektor-Memory-T-Zell-Phäno-
typ annahmen, was sich anhand der geänderten Oberflächenantigene nach-
vollziehen ließ.
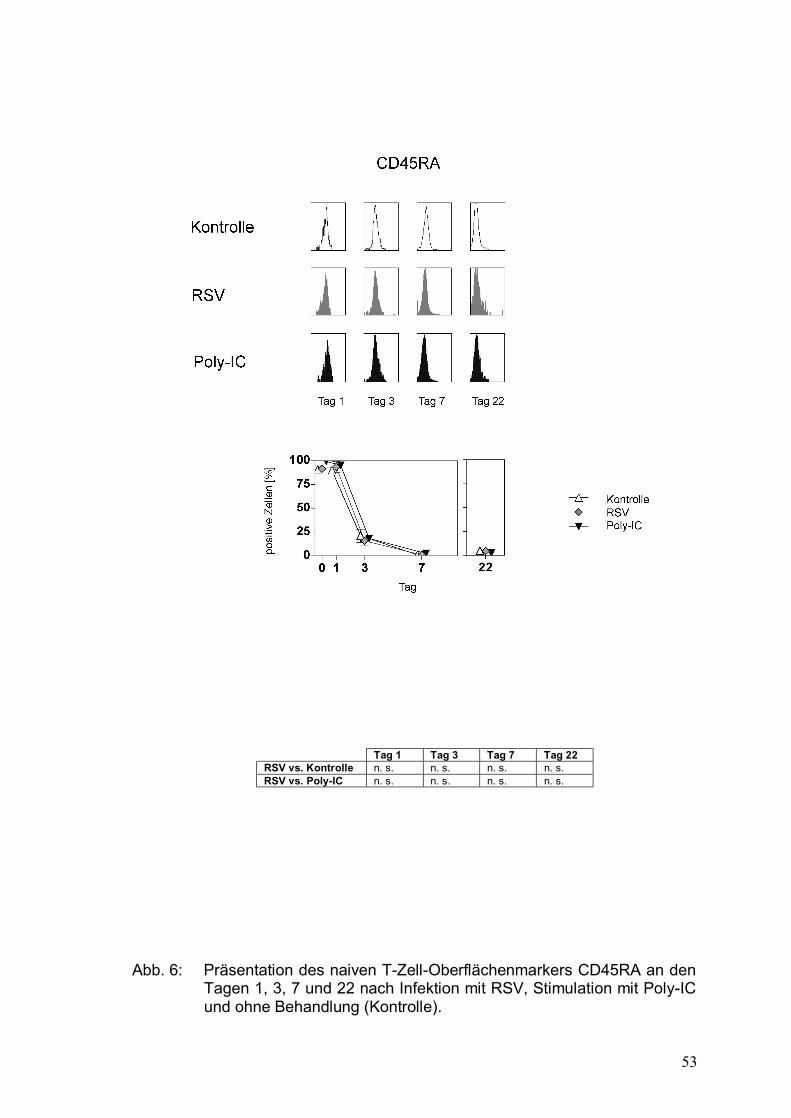
53
Tag 1 Tag 3 Tag 7 Tag 22
RSV vs. Kontrolle n. s. n. s. n. s. n. s. RSV vs. Poly-IC n. s. n. s. n. s. n. s.
Abb. 6: Präsentation des naiven T-Zell-Oberflächenmarkers CD45RA an den Tagen 1, 3, 7 und 22 nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle).
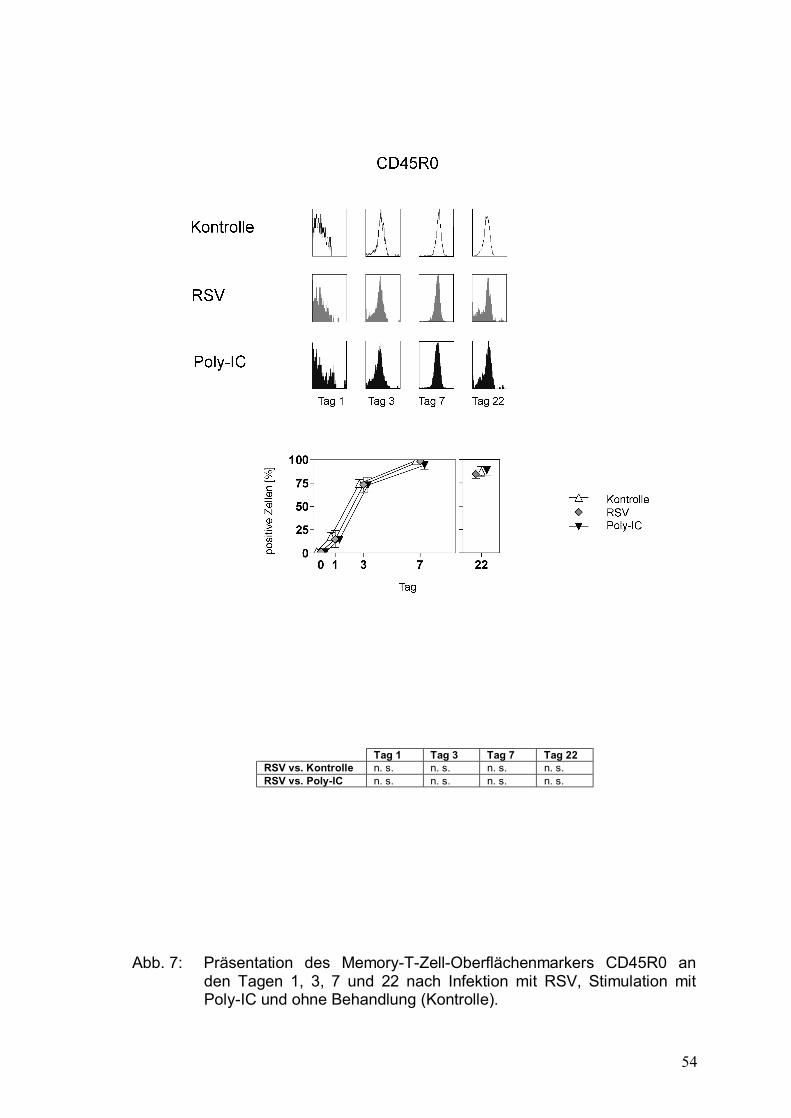
54
Tag 1 Tag 3 Tag 7 Tag 22
RSV vs. Kontrolle n. s. n. s. n. s. n. s. RSV vs. Poly-IC n. s. n. s. n. s. n. s.
Abb. 7: Präsentation des Memory-T-Zell-Oberflächenmarkers CD45R0 an
den Tagen 1, 3, 7 und 22 nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle).
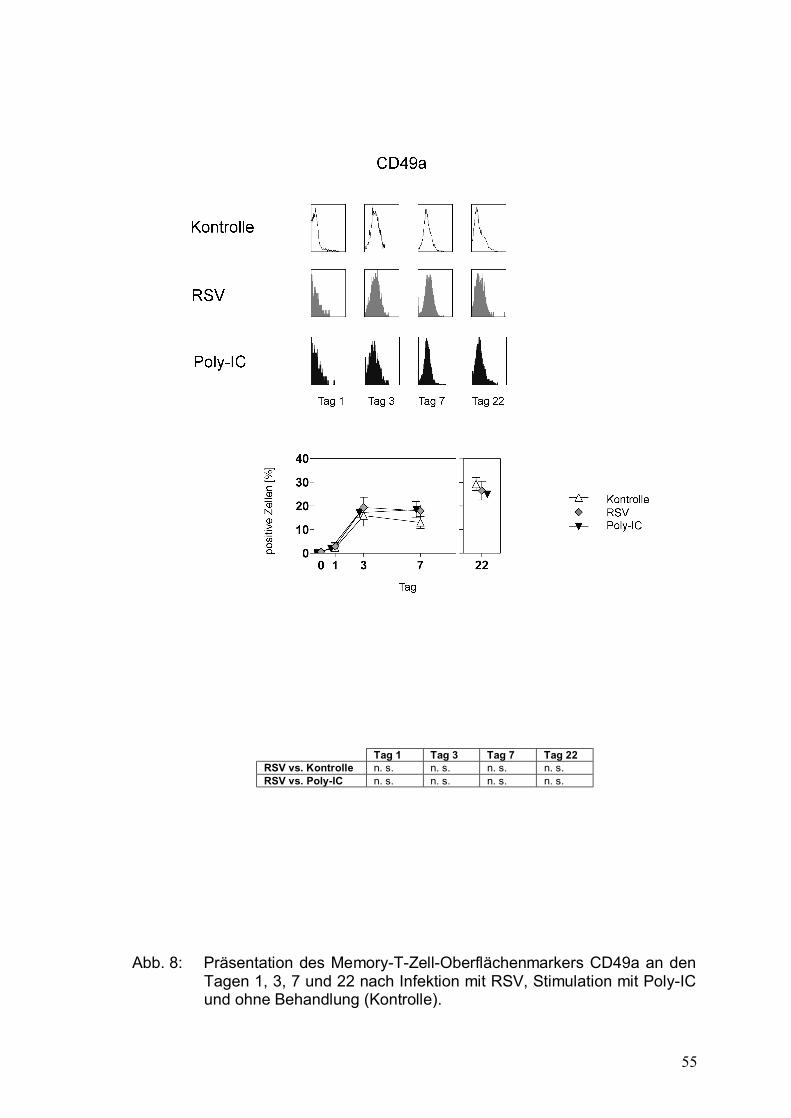
55
Tag 1 Tag 3 Tag 7 Tag 22
RSV vs. Kontrolle n. s. n. s. n. s. n. s. RSV vs. Poly-IC n. s. n. s. n. s. n. s.
Abb. 8: Präsentation des Memory-T-Zell-Oberflächenmarkers CD49a an den
Tagen 1, 3, 7 und 22 nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle).
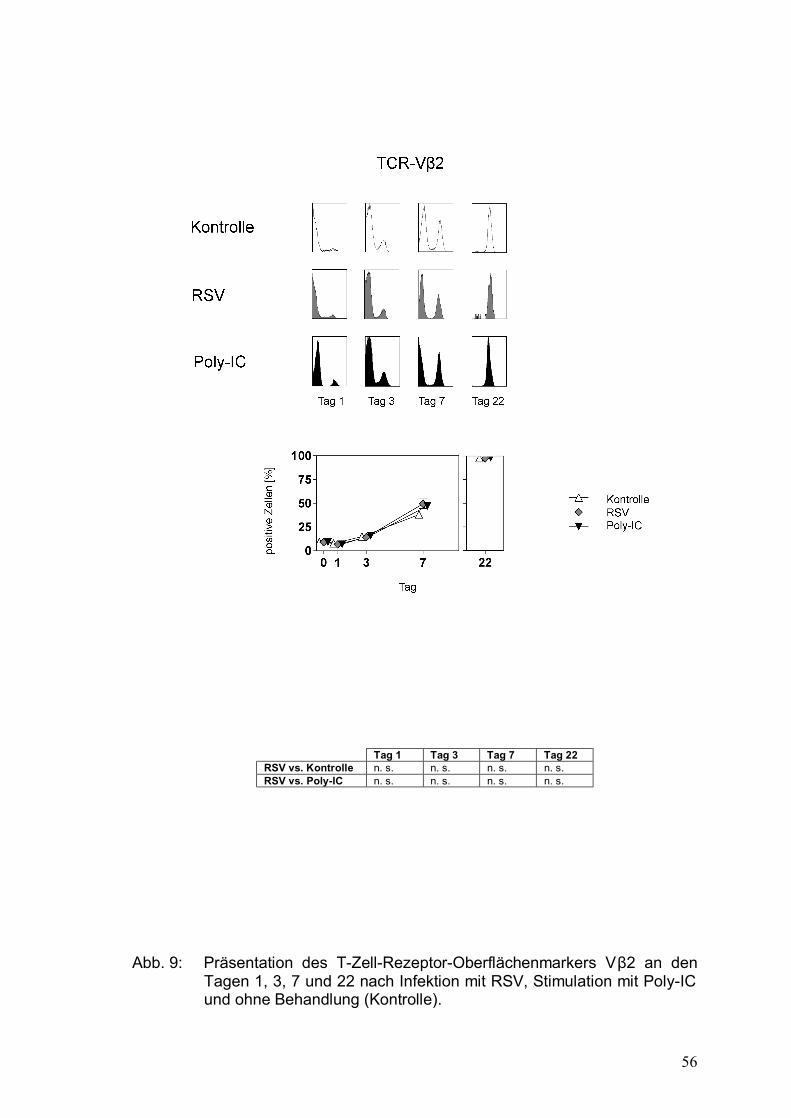
56
Tag 1 Tag 3 Tag 7 Tag 22
RSV vs. Kontrolle n. s. n. s. n. s. n. s. RSV vs. Poly-IC n. s. n. s. n. s. n. s.
Abb. 9: Präsentation des T-Zell-Rezeptor-Oberflächenmarkers Vβ2 an den
Tagen 1, 3, 7 und 22 nach Infektion mit RSV, Stimulation mit Poly-IC und ohne Behandlung (Kontrolle).

57
5. DISKUSSION
5.1 DIE RSV-INFEKTION FÜHRT ZU EINER IMMUNOLOGISCHEN SEKUNDÄRANTWORT MIT AUSBILDUNG VON MEMORY-T- ZELLEN
Es ist bekannt, dass die RSV-Infektion keinen lang anhaltenden Schutz vor
einer Reinfektion hinterlässt. Im Gegenteil, schon kurz nach überstandener
Erstinfektion wurden Reinfektionen mit dem genetisch identischen Virus
beschrieben, und auch Erwachsene erkranken immer wieder im Laufe ihres
Lebens symptomatisch an RSV-Infektionen, wenngleich auch mit abge-
schwächtem Krankheitsbild (Scott et al., 2006, Hall et al., 1991, Henderson et
al., 1979). Bisher gibt es keinen aktiven, lang anhaltenden Impfschutz; lediglich
für Hochrisikogruppen wie ehemals sehr junge Frühgeborene oder Säuglinge
mit Lungen- oder Herzerkrankungen steht ein Passivimpfstoff zur Verfügung.
Uns stellte sich daher die Frage, ob die Ursache der häufigen symptomatischen
Reinfektionen in der fehlenden Ausbildung von Memory-T-Zellen liegt, es also
zu keiner ausreichenden immunologischen Sekundärantwort kommt.
Hierfür untersuchten wir den Effekt RSV-infizierter Dendritischer Zellen
zunächst auf die T-Zell-Primär- und dann auch auf die -Sekundärantwort in
vitro. Die Zellen separierten wir aus Nabelschnurblut, welches eine hohe Anzahl
naiver T-Zellen und adulter hämatopoetischer Stammzellen enthält, aus denen
große Mengen Dendritischer Zellen gewonnen werden können. Um die Antigen-
Stimulation in vitro zu simulieren, nutzen wir das Superantigen TSST-1,
welches sich in vielen Versuchsreihen als Simulator der Virusantigen-Reaktion
bewährt hat. Als Kontrollgruppen dienten uninfizierte und mit Poly-IC, einem
Simulator viraler Infektionen in vitro, behandelte Dendritische Zellen.
Naive T-Zellen (CD45RA+) reifen durch wiederholten Antigenkontakt zu
CD45R0+ aktivierten oder Memory-/Effektor-Memory-T-Zellen heran und
erwerben so ein immunologisches Gedächtnis (Koning et al., 1996).

58
In unseren Untersuchungen konnten wir zeigen, dass es nach wiederholter
Stimulation zu einer Sekundärantwort mit der Ausbildung von Memory-/Effektor-
Memory-T-Zellen kommt: Nach Re-Stimulation der Co-Kulturen naiver T-Zellen,
Dendritischer Zellen und TSST-1 an den Tagen 7, 14 und 21 zeigte die Analyse
an Tag 22, dass die T-Zellen ihren naiven CD45RA+-Charakter verloren und
einen CD45R0+/CD49a+-Phänotyp akquirierten. Dieser Phänotyp entspricht
Memory-/Effektor-Memory-T-Zellen.
Ähnliche Ergebnisse konnten bei neugeborenen Kälbern, die mit bovinem RSV
infiziert wurden, erzielt werden. In ihrer Studie zeigten McInnes et al., dass es
nach RSV-Infektion zu einem Anstieg der Memory-Zellen im Untersuchungs-
material der Kälber kam. Während an Tag 1 die Mehrheit der Zellen im Blut, in
der Lunge und in der Bronchoalveolären Lavage (BAL) uninfizierter Kälber
naive T-Zellen waren, entsprach am fünften Tag nach Infektion die Anzahl
naiver T-Zellen bereits der Menge CD45R0-positiver T-Zellen. An Tag 10 nach
Infektion war der Anteil von Memory-/Effektor-Memory-T-Zellen signifikant
größer als der von naiven T-Zellen (McInnes et al., 1999).
Zudem wurde das Sputum infizierter Kälber und an RSV-Bronchiolitis erkrankter
Kinder untersucht: Im Vergleich der humanen mit den bovinen RSV-Infektionen
zeigte sich, dass bei infizierten Kälbern in der Bronchoalveolären Lavage eher
CD8+ T-Zellen dominierten, wohingegen es bei Kindern mit RSV-Bronchiolitis
zu einem Überwiegen der CD4+ T-Zellen kommt.
Die Vergleichbarkeit dieser Studie mit unseren Ergebnissen unterstreicht die
Theorie, dass es zur Ausbildung eines immunologischen Gedächtnisses kommt
und der eingeschränkten Immunität nach RSV-Infektion nicht eine fehlende
Ausbildung von Memory-/Effektor-Memory-T-Zellen zugrunde liegt.
Offen bleibt in unseren Untersuchungen jedoch die Differenzierung der
Untergruppen der zentralen Memory- und Effektor-Memory-T-Zellen.
Mithilfe des Chemokinrezeptors CCR7 und der Oberflächenmarker CD45RA
und CD45R0 lassen sich drei Typen von T-Zellen unterscheiden: Naive T-
Zellen (CCR7+, CD45RA+), zentrale Memory-T-Zellen (CCR7+, CD45R0+) und
Effektor-Memory-T-Zellen (CCR7-, CD45R0+).

59
De Bree et al. zeigten, dass RSV-spezifische CD8+ T-Zellen von Erwachsenen
zumeist (92%) einen CCR7- Phänotyp aufweisen, also die Effektor-Memory-T-
Zellen deutlich überwiegen (de Bree et al., 2005).
In unseren Untersuchungen unterschieden wir nicht, ob es sich um zentrale
Memory-T-Zellen handelt, die hauptsächlich das „Langzeitgedächtnis“ reprä-
sentieren, oder um Effektor-Memory-T-Zellen, die eher kürzer wirksam sind und
die Ausbreitung des Erregers verhindern. Möglicherweise besteht auch hier
eine Imbalance in der Ausbildung dieser beiden Subgruppen, die dazu führen
könnte, dass die Immunität nicht optimal generiert werden kann. Dies ist eine
Frage, die durch weiterführende Untersuchungen noch zu klären sein wird.
Eine andere mögliche Erklärung für die unzureichende Immunität liefern die
Ergebnisse weiterer, in dieser Arbeit nicht beschriebener Untersuchungen der
Zytokingeneration und der Proliferation.
Wir untersuchten die Zytokingeneration während der immunologischen Primär-
und der Sekundärantwort (Rothoeft et al., 2007). Bekannt ist nicht nur, dass
RSV unter anderem die Interferon-γ-Generation deutlich negativ beeinträchtigt,
sondern auch, dass die Schwere der klinischen Symptome von der Interferon-γ-
Generation abhängig ist (Bartz et al., 2003, Legg et al., 2003, Schauer et al.,
2004). Wir konnten zeigen, dass, während es bei der Primärantwort zu einer
deutlichen Beeinflussung der Zytokinmuster kam, die Memory-Antwort hin-
gegen nicht in diesem Maße beeinträchtigt wurde (Rothoeft et al., 2007).
Während der Primärantwort kam es zu einer zeitlichen Abfolge der maximalen
Ausschüttung der verschiedenen Zytokine (IL-2, TNF-α, IL-10, IL-4, IL-13 und
IFN-γ); zudem beeinträchtigte die RSV-Infektion die Anzahl zytokin-
produzierender Zellen einerseits und die Konzentration der ausgeschütteten
Zytokine andererseits. Insbesondere ergaben sich signifikante Unterschiede in
der IFN-γ-Reihe sowie beim TNF-α.
Im Vergleich hierzu kam es während der Sekundärantwort an Tag 22 zu einem
zeitlich simultanen Peak aller Zytokine, deren Konzentration jedoch zum Teil
signifikante Unterschiede zeigte: An Tag 3 erreichte IFN-γ seine maximale
Ausschüttung, wobei die Interferon-γ-Produktion in der RSV-Gruppe deutlich
niedriger war. Auch die Sekundärantwort an Tag 22 war gekennzeichnet durch
eine signifikant herabgesetzte IFN-γ-Ausschüttung im Vergleich zu Poly-IC.

60
Sowohl die Anzahl IFN-γ-produzierender Zellen als auch die Konzentration des
ausgeschütteten Zytokins zeigten signifikante Unterschiede, die klinischen
Beispielen entsprechen, in denen reduzierte Interferon-γ-Spiegel bei an RSV-
Bronchiolitis Erkrankten gemessen wurden. Dass es während der
Sekundärantwort zu einer zwar schwachen, jedoch ausreichenden IFN-γ-
Generation kommt, könnte ein Grund für die abnehmende Schwere der
klinischen Symptomatik bei einer Reinfektion im Erwachsenenalter sein, da
IFN-γ möglicherweise eine protektive Wirkung hat (Henderson et al., 1979,
Kumar et al., 1999).
Ebenfalls ist bekannt, dass RSV neben der Zytokingeneration auch die
Proliferation von T-Zellen beeinflusst. Schlender et al. beschreiben eine
herabgesetzte T-Zell-Proliferation durch Kontakt zum RSV-F-Protein, das auf
der Zelloberfläche des RS-Virus exprimiert wird (Schlender et al., 2002).
Wir untersuchten in weiteren, in dieser Arbeit nicht beschriebenen
Experimenten die Induktion der T-Zell-Proliferation durch Dendritische Zellen
mittels Durchflusszytometrie (Rothoeft et al., 2007).
In Co-Kulturen unbehandelter, Poly-IC-behandelter oder RSV-infizierter
Dendritischer Zellen mit TSST-1 und entweder naiven oder Memory-/Effektor-
Memory-T-Zellen zeigte sich eine signifikant eingeschränkte Proliferation bei
RSV-infizierten im Vergleich zu unbehandelten und Poly-IC-behandelten
Kulturen. Naive T-Zellen, die den höchsten Proliferationsgrad zeigten, sprachen
dabei deutlich stärker auf die Kontaktinhibition an als Memory-/Effektor-
Memory-T-Zellen.
Zusammenfassend lässt sich sagen, dass Memory-/Effektor-Memory-T-Zellen
zwar ausreifen und Zytokine gebildet werden, die Anzahl der gebildeten
Memory-/Effektor-Memory-T-Zellen jedoch eventuell zu gering ist, so dass bei
einer Reinfektion möglicherweise zu wenige kompetente Zellen zur Immun-
abwehr vorliegen und es daher zu einer Infektion mit abgeschwächter klinischer
Symptomatik kommt.

61
5.2 DIE RSV-INFEKTION INDUZIERT DIE REIFUNG DENDRITISCHER
ZELLEN WENIGER STARK ALS POLY-IC
Dendritische Zellen sind als professionelle Antigenpräsentierende Zellen die
wichtigsten Induktoren der T-Zell-vermittelten Immunantwort. Während der
RSV-Infektion kommt es zur Ausreifung Dendritischer Zellen, jedoch ist ein
viraler Mechanismus, die Immunantwort des Wirtes zu umgehen, die
Beeinflussung der Funktion Dendritischer Zellen.
Wir untersuchten daher die Ausbildung der Oberflächenmoleküle CD83 und
HLA-DR als Reifemarker Dendritischer Zellen und die Ausbildung des co-
stimulierenden Moleküls CD86 in Kulturen Dendritischer Zellen, die wir ent-
weder unbehandelt ließen, mit Poly-IC behandelten oder mit RSV infizierten.
Unsere Untersuchungen zeigten eine mangelnde Reifung Dendritischer Zellen
trotz erfolgreicher Infektion mit dem RS-Virus (etwa 25% der Zellen trugen das
RSV-F-Protein). In der Positiv-Kontrollgruppe der mit Poly-IC behandelten
Dendritischen Zellen zeigte sich eine deutliche Expression der Ober-
flächenmoleküle CD83 (MFI 423 ± 36) und HLA-DR (MFI 1061 ± 223),
wohingegen es bei der Infektion mit RSV zu einer abgeschwächten Expression
von CD83 (MFI 283 ± 144) kam und zu keinem Effekt beim HLA-DR (MFI 705 ±
106). Hier lag die Negativ-Kontrollgruppe der nicht-infizierten Dendritischen
Zellen mit einer MFI von 727 ± 211 sogar noch höher in der HLA-DR-
Expression als die RSV-Gruppe.
Einen ähnlichen Effekt sahen wir bei der Expression des co-stimulierenden
Faktors CD86: 69,2% ± 10,2% der mit Poly-IC behandelten Dendritischen
Zellen trugen nach 24-stündiger Stimulation diesen Oberflächenmarker, bei den
mit dem RS-Virus infizierten Zellen lag die Rate lediglich bei 47,7% ± 2,8%, bei
den unbehandelten Zellen bei 33,8% ± 9,6%.
Vergleichbares beschrieben bereits de Graaff et al.: Sie beobachteten eine
Reifung Dendritischer Zellen nach Infektion mit RSV, die im Vergleich zur
Behandlung mit Poly-IC jedoch deutlich herabgesetzt war (de Graaff et al.,
2005).

62
Poly-IC ist ein starker Stimulus für Dendritische Zellen, deren Reifung zu
induzieren und einen Phänotyp zu generieren, der hohe Level von HLA-DR,
CD83 und CD86 exprimiert. Dies zeigten bereits Verdijk et al., als sie 1999 die
Reifung von Dendritischen Zellen mit Poly-IC auslösten (Verdijk et al., 1999).
Als synthetische dsRNA ist Poly-IC ein Molekül, welches mit dem Toll-like-
Rezeptor 3 (TLR3) interagiert und eine virale Infektion simuliert. Es gilt als
starker Ligand des TLR3. In unseren Experimenten setzten wir Poly-IC daher
als Positivkontrolle ein.
Über rezeptorvermittelte Signalwege nach Bindung von Poly-IC an den TLR3
werden sowohl Zytokine ausgeschüttet als auch die Reifung Dendritischer
Zellen induziert. Somit stellt es einen annähernd optimalen Reifungsstimulus für
Dendritische Zellen in vitro dar.
Neben Poly-IC bindet virale Doppelstrang-RNA (dsRNA) an TLR3 (Matsumo et
al., 2008). RSV hingegen ist ein Einzelstrang-RNA-Virus (ssRNA), das nur
während eines kurzen Momentes der Virusreplikation - wenn die messenger-
RNA (mRNA) an den ssRNA-Strang bindet - doppelsträngig vorliegt. Die
Bindung an einen Rezeptor wird somit durch seine Verfügbarkeit zu diesem
Zeitpunkt des Replikationszyklus limitiert, womit die abgeschwächte Ausbildung
der Oberflächenmarker bei der RSV-Infektion im Vergleich zur Poly-IC-
Behandlung möglicherweise erklärt werden könnte.
Bekannt ist auch, dass ssRNA mit dem TLR7 interagiert, wie die Arbeitsgruppe
von Diebold et al. zeigte (Diebold et al., 2004). Zudem interagiert das RS-Virus
mit TLR4. Der TLR4 erkennt virale Oberflächenproteine wie das RSV-F-Protein
(Kurt-Jones et al., 2000). Poly-IC hingegen interagiert ausschließlich mit TLR3,
womit sich die Bindungswahrscheinlichkeit an TLR3 im Vergleich zu RSV
deutlich erhöht.
Daher ist es möglich, dass es bei Behandlung mit Poly-IC zu einer stärkeren
Ausbildung der Oberflächenmarker kommt als nach Infektion mit dem RS-Virus.
Einen weiteren Effekt auf die DC-Reifung konnten Kruse et al. in Studien mit
Herpes simplex Virus-1 (HSV-1) beschreiben: 10 Stunden nach Infektion kam
es zu einer starken Herabregulation von CD83 an der Zelloberfläche reifer DCs.
Da die fehlende CD83-Oberflächenexpression nicht durch die Inhibition der
CD83-mRNA-Synthese hervorgerufen wurde, vermutete man, dass ein

63
posttranskriptionelles Ereignis für die Herabmodulation von CD83 und damit
auch für die Reifung Dendritischer Zellen verantwortlich sein müsse, da die
CD80/86 Expression nicht beeinflusst wurde: Es konnte gezeigt werden, dass
CD83 in Lysosomen abgebaut wird (Kruse et al., 2000).
Arrode et al. zeigten an mit humanem Cytomegalievirus (hCMV) spätinfizierten
Fibroblasten, dass diese vermindert CD83 exprimierten, was wiederum zur
Inhibition der DC-Reifung führte (Arrode et al., 2002).
Diese Ergebnisse weisen darauf hin, dass die CD83-Herabmodulation und die
Inhibition der DC-Reifung virale Mechanismen sind, eine Induktion antiviraler
Immunantworten zu verhindern. Möglicherweise kann ein ähnlicher Effekt durch
Infektion mit RSV ausgelöst werden, so dass auch hier die Immunantwort
zugunsten des Virus abgeschwächt wird.
Eine weitere mögliche Ursache der weniger stark ausgeprägten Reifung
Dendritischer Zellen könnte sein, dass es bei der Virusinfektion noch zu
anderen Begleitreaktionen von Virusproteinen in der Zelle kommt, die den
Effekt auf die Oberflächenmarkerexpression herabsetzen. Beispielsweise
beschreiben Jeffree et al. eine Interaktion von Aktin und RSV, wodurch es zu
einer „Lähmung“ der RSV-infizierten Zelle kommt (Jeffree et al., 2007). Das
Aktin-Zytoskelett spielt eine wichtige Rolle bei der Zellmotilität, -bewegung und
-morphologie. Aktin selbst ist ein wichtiger viraler Transkriptionsfaktor (Bitko et
al., 2003).
Zusammenfassend lässt sich sagen, dass die RSV-Infektion im Vergleich zur
Behandlung mit Poly-IC zu einer verminderten Ausreifung Dendritischer Zellen
in vitro führt. Aufgrund der rezeptorspezifischen Bindung von Poly-IC erhöht
sich die Bindungswahrscheinlichkeit und damit die Ausreifung Dendritischer
Zellen deutlich. Das RS-Virus ist hingegen weiteren Rezeptorinteraktionen
ausgesetzt, die zu einer herabgesetzten Bindungswahrscheinlichkeit führen.
Ebenfalls unterliegt RSV anderen zellulären Mechanismen, die die Oberflächen-
markerexpression herabregulieren können.

64
5.3 DIE REAKTION DER T-ZELLEN AUF TSST-1 IST EHER SUPER-
ANTIGEN- ALS ANTIGEN-SPEZIFISCH
TSST-1, ein Superantigen, welches bevorzugt an die Vβ2-Kette des T-Zell-
Rezeptors bindet, wurde genutzt, um die Antigen-Stimulation in vitro nach-
zuahmen. Dass an Tag 22 annähernd 100% der T-Zellen Vβ2+ waren, ist ein
Effekt der klonalen Expansion nach Stimulation mit TSST-1, somit ist die
Reaktion eher superantigen- als antigenspezifisch.
Zudem ist bekannt, dass TSST-1 eher MHC-Klasse II-Reaktionen fördert,
wohingegen RSV MHC-Klasse I-spezifisch reagiert. Aus diesem Grunde ist das
genutzte in vitro-Modell nicht vollständig auf die in vivo-Situation übertragbar,
liefert jedoch einige Hinweise und Erklärungen, um die Pathogenese der RSV-
Infektion verstehen zu können.

65
6. ZUSAMMENFASSUNG
In dieser Arbeit sollte an einem humanen in vitro-Modell untersucht werden, ob
es nach einer Infektion mit dem RS-Virus zu einer potenten Primärantwort des
Immunsystems mit Ausreifung professioneller Antigenpräsentierender Zellen
kommt und ob während der Sekundärantwort die Ausbildung von Memory-T-
Zellen als Zellen des immunologischen Gedächtnisses induziert wird.
Folgende Oberflächenmoleküle als Marker der Reifung Dendritischer Zellen
wurden durchflusszytometrisch bestimmt: CD86, CD83, CD49a und HLA-DR.
Als Negativkontrolle diente die Co-Kultur naiver T-Zellen mit uninfizierten DCs,
als Positivkontrolle die Co-Kultur naiver T-Zellen mit Poly-IC-stimulierten DCs.
Poly-IC gilt als Simulator viraler Infektionen in vitro.
In der syngenen Langzeit-Co-Kultur untersuchten wir die immunologische
Sekundärantwort. Hier diente das Superantigen TSST-1 als exogener Stimulus,
der hauptsächlich an die Vβ2-Untereinheit des T-Zell-Rezeptors bindet, welcher
schnell durchflusszytometrisch erkannt werden kann. Die Langzeit-Co-Kultur
wurde an den Tagen 7, 14 und 21 restimuliert. Eine Messung der
Oberflächenmarker CD45RA, CD45R0, CD49a und Vβ2 erfolgte jeweils an den
Tagen 1, 3, 7 und 22. Den Oberflächenmarker CD45RA tragen naive T-Zellen,
die Marker CD45R0 und CD49a Memory-T-Zellen.
In den Untersuchungen konnte gezeigt werden, dass das RS-Virus eine
Reifung Dendritischer Zellen induziert, wenngleich auch schwächer als Poly-IC.
Dies kann erklärt werden mit der hochspezifischen Bindung des Poly-IC an den
Toll-like-Rezeptor 3, womit Poly-IC bei der Bindungsspezifität dem RS-Virus,
das unter anderem noch mit dem Toll-like-Rezeptor 4 interagiert, deutlich
überlegen ist. Auch kann das einzelsträngige RS-Virus nur während eines
kurzen Moments der Virusreplikation, wenn es doppelsträngig vorliegt, an den
TLR3 binden, da dieser lediglich Doppelstrang-Nukleide bindet.

66
In der Langzeit-Co-Kultur konnten wir zeigen, dass sich ein immunologisches
Gedächtnis in Form von Memory-T-Zellen bildet:
An Tag 22 trugen etwa 85% der T-Zellen den Oberflächenmarker CD45R0 und
entsprachen somit dem Memory-T-Zell-Typ. Das initial vorliegende Ober-
flächenmolekül CD45RA als Charakteristikum der naiven T-Zelle hatten sie
abgelegt.
Somit ist die Tatsache, dass es bereits kurze Zeit nach durchlaufener, schwerer
RSV-Infektion zu Reinfektionen mit dem genetisch identischen Virus kommt,
nicht ursächlich auf die fehlende Ausbildung eines immunologischen Gedächt-
nisses zurückzuführen. Hier müssen andere Faktoren eine Rolle spielen.

67
7. LITERATURVERZEICHNIS
Alexopoulou, L., Holt, A.C., Medzhitov, R., Flavell, R.A. (2001). Recognition of
double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature
413, 732-738
Arrode, G., Boccaccio, C., Abastado, J.P., Davrinche, C. (2002). Cross-
presentation of human cytomegalovirus pp65 (UL83) to CD83+ T cells is
regulated by virus-induced, soluble-mediator-dependent maturation of dendritic
cells. J. Virol. 76, 142-150
Bartz, H., Türkel, Ö., Hoffjan, S., Rothoeft, T., Gonschorek, A., Schauer, U.
(2003). Respiratory syncytial virus decreases the capacity of myeloid dendritic
cells to induce interferon-γ in naive T cells. Immunology 109, 49-57
Bekanntmachung eines Beschlusses des Gemeinsamen Bundesausschusses
über eine Änderung der Arzneimittel-Richtlinie in Anlage 4: Therapiehinweis zu
Palivizumab (2008). (Zugriff vom 22.02.2010).
http://www.g-ba.de/downloads/39-261-694/2008-06-19-AMR4-Palivizumab_BAnz.pdf
Bitko, V., Oldenburg, A., Garmon, N.E., Barik, S. (2003). Profilin is required for
viral morphogenesis, syncytium formation and cell-specific stress fiber induction
by respiratory syncytial virus. BMC Microbiology 3, 9
Caux, C., Dezutter-Dambuyant, C., Schmitt, D., Banchereau, J. (1992). GM-
CSF and TNF-alpha cooperate in the generation of dendritic Langerhans cells.
Nature 360, 258-261
Chanock, R.M., Roizman, B., Myers, R. (1957). Recovery from infants with
respiratory illness of a virus related to chimpanzee coryza agent (CCA). Am. J.
Hyg., 66, 281-290

68
Charlton, B., Auchincloss jr., H., Fathman, C.G. (1994). Mechanisms of
transplantation tolerance. Annu. Rev. Immunol. 12, 707-734
Choi, Y., Kotzin, B., Heeron, L., Callahan, J., Marrack, P., Kappler, J. (1989).
Interaction of Staphylococcus aureus toxin „superantigens“ with human T cells.
Immunology 86, 8941-8945
Choi, Y., Lafferty, J.A., Clements, J.R., Todd, J.K., Gelfand, E.W., Kappler, J.,
Marrack, P., Kotzin, B.L. (1990). Selective expansion of T cells expressing Vβ2
in toxic shock syndrome. J. Exp. Med. 172, 981-984
Cramer, S.O., Trumpfheller, C., Mehlhoop, U., Moré, S., Fleischer, B., von
Bonin, A. (2000). Activation-induced expression of murine CD83 on T cells and
identification of a specific CD83 ligand on murine B cells. Int. Immunol. 12,
1347-1351
Cyster, J.G., Fowell, D., Barclay, A.N. (1994). Antigenic determinants encoded
by alternatively spliced exons of CD45 are determined by the polypeptide but
influenced by glycosylation. Int. Immunol. 6, 1875-1881
de Bree, G.J., Heidema, J., van Leeuwen, E.M., van Bleek, G.M., Jonkers, R.E.,
Jansen, H.M., van Lier, R.A., Out, T.A. (2005). Respiratory syncytial virus-
specific CD8+ memory T cell responses in elderly persons. J Infect Dis 191,
1710-1718
de Graaff, P.M.A., de Jong, E.C., van Capel, T.M., van Dijk, M.E.A., Roholl,
P.J.M., Boes, J., Luytjes, W., Kimpen, J.L.L., van Bleek, G.M. (2005).
Respiratory Syncytial Virus Infection of Monocyte-Derived Dendritic Cells
Decreases Their Capacity to Activate CD4 T Cells. The Journal of Immunology
175, 5904-5911
Dellabona, P., Peccoud, J., Kappler, J., Marrack, P., Benoist, C., Mathis, D.
(1990). Superantigens interact with MHC class II molecules outside of the
antigen groove. Cell 62, 1115-1121

69
Diebold, S.S., Kaisho, T., Hemmi, H., Akira, S., Reis e Sousa, C. (2004). Innate
Antiviral Responses by Means of TLR7-Mediated Recognition of Single-
Stranded RNA. Science 303, 1529-1531
Fischer, E.H., Charbonneau, H., Tonks, N.K. (1991). Protein tyrosine phospha-
tases: a diverse family of intracellular and transmembrane enzymes. Science
253, 401-406
Fleischer, B., Schrezenmeier, H. (1988). T cell stimulation by staphylococcal
enterotoxins. Clonally variable response and requirement for major
histocompatibility complex class II molecules on accessory or target cells. J.
Exp. Med. 167, 1697-1707
Fortier, M., Kent, S., Ashdown, H., Poole, S., Boksa, P., Luheshi, G. (2004).
The viral mimic, polyinosinic:polycytidylic acid, induces fever in rats via an
interleukin-1 dependant mechanism. Am. J. Physiol. Regul. Integr. Comp.
Physiol. 287, 759-766
Fujimoto, Y., Tu, L., Miller, A.S., Bock, C., Fujimoto, M., Doyle, C., Steeber,
D.A., Tedder, T.F. (2002). CD83 Expression Influences CD4+ T Cell Develop-
ment in the Thymus. Cell 108, 1-20
Gascoigne, N.R., Ames, K.T. (1991). Direct binding of secreted T-cell receptor
beta chain to superantigen associated with class II major histocompatibility
complex protein. Proc. Natl. Acad. Sci. USA 88, 613-616
Hall, C.B., Walsh, E.E., Long, C.E., Schnabel, K.C. (1991). Immunity to and
frequency of reinfection with respiratory syncytial virus. J. Infect. Dis. 163, 693-
698
Hathcock, K.S., Laszlo, G., Pucillo, C., Linsley, P., Hodes, R.J. (1994).
Comparative analysis of B7-1 and B7-2 costimulatory ligands: expression and
function. J. Exp Med. 180, 631-640

70
Henderson, F.W., Collier, A.M., Cycle jr., W.A., Denny, F.W. (1979). Respiratory
syncytial virus infections, reinfections and immunity. A prospective, longitudinal
study in young children. N Engl J Med 300, 530-534
Hock, B.D., Kato, M., McKenzie, J.L., Hart, D.N.J. (2001). A soluble form of
CD83 is released from activated dendritic cells and B lymphocytes, and is
detectable in normal human sera. Int. Immunol. 13, 959-967
Inaba, K., Inaba, M., Witmer-Pack, M., Hatchcock, K., Hodes, R., Steinman,
R.M. (1995). Expression of B7 costimulator molecules on mouse dendritic cells.
Adv. Exp. Med. Biol. 378, 65-70
Jeffree, C.E., Brown, G., Aitken, J., Su-Yin Dawn, Y., Tan, B.H., Sugrue, R.J.
(2007). Ultrastructural analysis of the interaction between F-actin and
respiratory syncytial virus during virus assembly. Virology 369, 309-323
June, C.H., Bluestone, J.A., Nadler, L.M., Thompson, C.B. (1994). The B7 and
CD28 receptor families. Immunol. Today 15, 321-331
Karron, R.A., Wright, P.F., Belshe, R.B., Thumar, B., Casey, R., Newman, F.,
Polack, F.P., Randolph, V.B., Deatly, A., Hackell, J., Gruber, W., Murphy, B.R.,
Collins, P.L. (2005). Identification of a recombinant live attenuated respiratory
syncytial virus vaccine candidate that is highly attenuated in infants. J Infect Dis
191, 1093-1104
Khoury, S.J., Akalin, E., Chandraker, A., Turka, L.A., Linsley, P.S., Sayegh,
M.H., Hancock, W.W. (1995). CD28-B7 costimulatory blockade by CTLA4lg
prevents actively induced experimental autoimmune encephalomyelitis and
inhibits Th1 but spares Th2 cytokines in the central nervous system. J. Immunol
155, 4521-4524
Koning, H., Baert, M.R.M., Oranje, A.P., Savelkoul, H.F.J., Neijens, H.J. (1996).
Development of Immune Functions Related to Allergic Mechanisms in Young
Children. Pediatric Research 40, 363-375

71
Krakauer, T. (1999). Immune response to staphylococcal superantigens.
Immunol. Res. 20, 163-173
Kruse, M., Rosorius, O., Krätzer, F., Stelz, G., Kuhnt, C., Schuler, G., Hauber,
J., Steinkasserer, A. (2000). Mature Dendritic Cells Infected with Herpes
Simplex Virus Type 1 Exhibit Inhibited T-Cell Stimulatory Capacity. J. Virol. 74,
7127-7136
Kumar, M., Behera, A.K., Matsuse, H., Lockey, R.F., Mohapatra, S.S. (1999).
Intranasal IFN-gamma gene transfer protects BALB/c mice against respiratory
syncytial virus infection. Vaccine 18, 558-567
Kurt-Jones, E.A., Popova, L., Kwinn, L., Haynes, L.M., Jones, L.P., Tripp, R.A.,
Walsh, E.E., Freeman, M.W., Golenbock, D.T., Anderson, L.J., Finberg, R.W.
(2000). Pattern recognition receptors TLR4 and CD14 mediate response to
respiratory syncytial virus. Nat. Immunol. 1, 398–401
Lechmann, M., Krooshoop, D.J.E.B., Dudziak, D., Kremmer, E., Kuhnt, C.,
Figdor, C.G., Schuler, G., Steinkasserer, A. (2001). The Extracellular Domain of
CD83 Inhibits Dendritic Cell–mediated T Cell Stimulation and Binds to a Ligand
on Dendritic Cells. J. Exp. Med. 194, 1813-1821
Legg, J.P., Hussain, I.R., Warner, J.A., Johnston, S.L., Warner, J.O. (2003).
Type 1 and type 2 cytokine imbalance in acute respiratory syncytial virus
bronchiolitis. Am J Resp Crit Care Med 168, 633-639
Lu , P., Wang, Y.L., Linsley, P.S. (1997). Regulation of self-tolerance by
CD80/CD86 interactions. Current Opinion in Immunology 9, 858-862
Mackay, C.R. (1993). Immunological Memory. Adv Immunol 53, 217-265
Mackay, C.R. (1999). Dual personality of memory T cells. Nature 401, 659-660

72
Matsumoto, M., Seya, T. (2008). TLR3: interferon induction by double-stranded
RNA including poly(I:C). Adv. drug Deliv. Rev. 60, 805-812
Matzinger, P. (1994). Tolerance, danger, and the extended family (1994). Annu.
Rev. Immunol. 12, 991-1045
McInnes, E., Sopp, P., Howard, C.J., Taylor, G. (1999). Phenotypic analysis of
local cellular responses in calves infected with bovine respiratory syncytial virus.
Immunology 96, 396-403
Medzhitov, R. (2001). Toll-like receptors and innate immunity. Nature 1,135-145
Morris, J.A., Blount jr., R.E., Savage, R.E. (1956). Recovery of cytopathogenic
agent from chimpanzees with coryza. Proc. Soc. Exper. Biol. & Med. 92, 544-
549
Moscona, A. (2005). Entry of parainfluenza virus into cells as a target for
interrupting childhood respiratory disease. J. Clin. Invest. 115, 1688-1698
O´Connell, K., Boo, T.W., Keady, D., Niriain, U., O´Donovan, D., Commane, M.,
Faherty, C., Cormican, N. (2011). Use of palivizumab and infection control
measures to control an outbreak of respiratory syncytial virus in a neonatal
intensive care unit confirmed by real-time polymerase chain reaction. J. Hosp.
Infect. 77, 338-342
Pichler, W.J., Peter, H.H. (2001). Immunsystem.
In Siegenthaler, W. (Hrsg.). Klinische Pathophysiologie.
Georg Thieme Verlag, Stuttgart, New York, 491-562
Punnoose, A. (2012). Respiratory Syncytial Virus Bronchiolitis. JAMA 307, 213

73
Resch, B., Sommer, C., Nuijten, M.J.C., Seidinger, S., Walter, E., Schoellbauer,
V., Mueller, W.D. (2012). Cost-effectiveness of palivizumab for respiratory
syncytial virus infection in high-risk children, based on long-term epidemiologic
data from Austria. Pediatr. Infect. Dis. J. 31, E1-E8
Respiratorische Synzytial-Viren (RSV). RKI-Ratgeber für Ärzte. Robert-Koch-
Institut, Ratgeber Infektionskrankheiten A-Z, RSV-Infektionen (2011). (Zugriff
vom 16.12.2011).
http://www.rki.de/cln_116/nn_504478/DE/Content/Infekt/EpidBull/Merkblaetter/R
atgeber__RSV.html
Rothoeft, T., Fischer, K., Zawatzki, S., Schulz, V., Schauer, U., Koerner-
Rettberg, C. (2007). Differential response of human naive and memory/effector
T cells to dendritic cells infected by respiratory syncytial virus. Clinical and
Experimental Immunology 150, 263-273
Sallusto, F., Lenig, D., Förster, R., Lipp, M., Lanzavecchia, A. (1999). Two
subsets of memory T lymphocytes with distinct homing potentials and effector
functions. Nature 401, 708-712
Schauer, U., Hoffjan, S., Bittscheidt, J., Köchling, A., Hemmis, S., Bongarzt, S.,
Stephan, V. (2002). RSV bronchiolitis and risk of wheeze and allergic
sensitisation in the first year of life. Eur. Respir. J. 20, 1277-1283
Schauer, U., Hoffjan, S., Rothoeft, T., Bartz, H., König, S., Fuchs, E.,
Bittscheidt, J., Köchling, A., Stephan, V. (2004). Severe respiratory syncytial
virus infections and reduced interferon-γ generation in vitro. Clin. Exp. Immunol.
138, 102-109
Schlender, J., Walliser, G., Fricke, J., Conzelmann, K.K. (2002). Respiratory
syncytial virus fusion protein mediates inhibition of mitogen-induced T-cell
proliferation by contact. J Virol 76, 1163-1170

74
Scholler, N., Hayden-Ledbetter, M., Dahlin, A., Hellström, K.E., Hellström, I.,
Ledbetter, J.A. (2001). CD83 is a sialic acid-binding Ig-like lectin (Siglec)
adhesion receptor that binds monocytes and a subset of activated CD8 + T cells.
J. Immunol. 166, 3865-3872
Scholler, N., Hayden-Ledbetter, M., Dahlin, A., Hellström, I., Hellström, K.E.,
Ledbetter, J.A. (2002). CD83 Regulates the Development of Cellular Immunity.
J. Immunol. 168, 2599-2602
Scott, P.D., Ochola, R., Ngama, M., Okiro, E.A., Nokes, D.J., Medley, G.F.,
Cane, P.A. (2006). Respiratory syncytial virus re-infections in infants from
coastal Kenya. J. Infect. Dis. 193, 59-67
Singh, B.R., Kokan-Moore, N.P., Bergdoll, M.S. (1988). Molecular topography of
toxic shock syndrome toxin 1 as revealed by spectroscopic studies.
Biochemistry 27, 8730-8735
Sullender, W.M. (2000). Respiratory Syncytial Virus Genetic and Antigenic
Diversity. Clinical Microbiology Reviews 1, 1-15
Thurau, A.M., Streckert, H.J., Rieger, C.H.L., Schauer, U. (1998). Increased
number of T cells committed to IL-5 production after respiratory syncytial virus
(RSV) infection of human mononuclear cells in vitro. Clin Exp Immunol 114,
450-455
Tripp, R.A., Moore, D., Jones, L., Sullender, W., Winter, J., Anderson, L.A.
(1999). Respiratory Syncytial Virus G and/or SH Protein Alters Th1 Cytokines,
Natural KillerCells, and Neutrophils Responding to Pulmonary Infection in
BALB/c Mice. Journal of Virology, 73, 7099-7107
Trowbridge, I.S. (1991). CD45. A prototype for transmembrane protein tyrosine
phosphatases. J. Biol. Chem. 266, 23517-23520

75
Trowbridge, I.S., Ostergaard, H.L., Johnson, P. (1991). CD45: a leukocyte-
specific member of the protein tyrosine phosphatase family. Biochim. Biophys.
Acta 1095, 46-56
Trowbridge, I.S., Johnson, P., Ostergaard, H.L., Hole, N. (1992). Structure and
function of CD45: A leucocyte-specific protein tyrosine phosphatase. Advances
in Experimenatl Medicine and Biology 323, 29-37
Turley, S.J., Inaba, K., Garrett, W.S., Ebersold, M., Unternaehrer, J., Steinman,
R.M., Mellman, I. (2000). Transport of Peptide-MHC Class II Complexes in
Developing Dendritic Cells. Science 288, 522-527
Ullmann, A.J., Maschmeyer, G. (2012). Respiratory Syncytial Virus (RSV) –
Infektionen bei Patienten nach hämatopoetischer Stammzelltransplantation.
(Zugriff vom 05.05.2012).
http://www.dgho-onkopedia.de/onkopedia/leitlinien/respiratory-syncytial-virus-rsv-
2013-infektionen
Ventre, K., Randolph, A.G. (2004). Ribavirin for respiratory syncytial virus
infection of the lower respiratory tract in infants and young children. Cochrane
Database Syst. Rev. 4, CD000181
Verdijk, R., Mutis, T., Esendam, B., Kamp, J., Melief, C.J., Brand, A., Goulmy,
E. (1999). Polyriboinosinic polyribocytidylic acid (poly(I:C)) induces stable
maturation of functionally active human dendritic cells. Immunol. 163, 57-61
White, J., Herman, A., Pullen, A.M., Kubo, R., Kappler, J.W., Marrack, P.
(1989). The Vß-specific superantigen staphylococcal enterotoxin B: stimulation
of mature T cells and clonal deletion in neonatal mice. Cell 56, 27-35

DANKSAGUNG
Mein besonderer Dank gilt meinem Doktorvater, Herrn Professor Doktor Uwe
Schauer, für die Überlassung des interessanten Themas und die engagierte
und geduldige Betreuung meiner Arbeit, für die begleitende Unterstützung und
fachliche Beratung sowie für die kritische Korrektur.
Frau Veronika Baumeister und Frau Angelika Michel führten mich in freund-
licher Arbeitsatmosphäre in labortechnische Arbeitsweisen ein und standen
jederzeit und uneigennützig bei Fragen und Problemen zur Verfügung.
Für seine Hilfe bei allen computertechnischen Schwierigkeiten und die persön-
liche Unterstützung meiner Arbeit danke ich Herrn Martin Schulte.
Unter der Leitung von Herrn Doktor Bernhard Ibach durfte ich meine
pädiatrische Ausbildung absolvieren und lernen, dass ärztliches Handeln und
Denken stets dem Wohl des Kindes zu unterstellen sind: Primum nil nocere!
Ich danke Linda Sender für den seit Kindertagen gemeinsam und in aufrichtiger
Freundschaft beschrittenen Lebensweg, der mich zu einem erfüllenden Beruf
führte.
Mein ganz persönlicher Dank gilt meinen Eltern, die meinen Werdegang ermög-
licht, mich zu jeder Zeit gefördert und gefordert haben und mir Rückhalt und
Ansporn gaben, meine Ziele zu erreichen.

LEBENSLAUF
PERSÖNLICHE DATEN
Name Sandra Zawatzki
Geburtsdatum 12. November 1978
Geburtsort Gelsenkirchen
SCHULISCHE AUSBILDUNG
08 / 1985 - 06 / 1989 Städtische Grundschule an der Devensstraße in
Gelsenkirchen-Horst
08 / 1989 - 06 / 1998 Annette-von-Droste-Hülshoff-Gymnasium in
Gelsenkirchen-Buer
12. Juni 1998 Erlangen der Allgemeinen Hochschulreife
BERUFLICHE AUSBILDUNG
10 / 1998 - 09 / 2001 Katholische Krankenpflegeschule am St. Josef-
Hospital in Gelsenkirchen-Horst
01. Oktober 2001 Erlangen der Berufsbezeichnung Krankenschwester
10 / 2001 - 09 / 2005 Teilzeitbeschäftigung als Krankenschwester im
St. Josef-Hospital in Gelsenkirchen-Horst, Klinik für
Medizinische & Radiologische Onkologie,
Hämatologie & Palliativmedizin

10 / 2005 - 10 / 2007 Teilzeitbeschäftigung als Krankenschwester in den
Evangelischen Kliniken Gelsenkirchen, Klinik für
Radioonkologie & Strahlentherapie
STUDIUM & MEDIZINISCHE AUSBILDUNG
10 / 2001 - 12 / 2007 Studium der Humanmedizin an der Ruhr-Universität
Bochum
10. September 2003 Bestehen der Ärztlichen Vorprüfung
08 / 2006 - 07 / 2007 Praktisches Jahr in der Klinik für Innere Medizin, der
Klinik für Chirurgie sowie der Klinik für Kinder &
Jugendliche am Sana-Klinikum in Remscheid
27. November 2007 Bestehen des Zweiten Staatsexamens
18. Dezember 2007 Approbation als Ärztin
01 / 2008 - 03 / 2013 Assistenzärztin in der Klinik für Kinder & Jugendliche
am Sana-Klinikum in Remscheid
20. März 2013 Anerkennung als Fachärztin für Kinder- &
Jugendmedizin
seit 01. April 2013 Fachärztin in der Uniklinik Köln, Klinik & Poliklinik für
Kinder- & Jugendmedizin, Abteilung für Neonatologie
& pädiatrische Intensivmedizin