Embed Size (px)
Citation preview

Zeitschrift fur Allg. Mikrobiologie 1 4 1961 274-285
Institut fur Mikrobiologie der Universitiit Frankfurt/Main
Auslosung von Mutationen verschiedenen Typs durch Behandlung des extrazellularen Phagen ,,Kappa"
von Serratia mit Nitrit
R. W. KAPLAN und S. K. BOSE*)
(Eingegangen am 4. 4. 61) I. Probleme
Der temperierte Phage Kappa aus dem lysogenen Stamm K des rote Kolonien bildenden Bakteriums Serratia marcesceiis (Bact. prodigiosum) hat die nngewohn- liche Eigenschaft, bei Bestrahlung mit UV oder Rontgenstrahlen der extrazellularen Partikel bis einige Prozent Mutationen vom Klarloch-Typ zu ergeben (ELLMAUER u. KAPLAN 1959). Eine Bestrahlung auch des Wirtes wie bei anderen Phagen (WEIGLE 1953, JAKOB 1954) ist dazu nicht notwendig, wenn eine solche auch die Mutationsaus- beute erhoht ( WINKLER). Da es sich hier bei anscheinendum Initierung von Erbande- rungen mit hoher Rate in extrazellularer DNS, also ,,in vitro", handelt, 1aBt das Stu- dium dieses Vorgangs Beitrage zur Erkenntnis der Natur von strahleninduzierten Mutationen in der Erbsubstanz erhoffen. Bisher zeigte sich bereits, daB solche zu Klarlochmutationen fiihrende W-Lasionen zu einem erheblichen Teil durch Nach- behandlung des freien Phagzn mit Licht, Warme oder anorganischen Ionen revertiert werden konnen (KAPLAN, WINKLER u. WOLF-ELLMAUER 1960; PARK 1961 ; KAPLAN 1960).
Auffallig ist, daB die UV-Bestrahlung fast nur Mutationen erzeugt, welche die Triibheit des Wildtyp-Plaques verschwinden laBt (klares Loch, c-Typ) oder sie stark vermindert (f leicht getriibtes Loch, 1-Typ). Zunachst wurde vermutet, da13 diese Mutationen vielleicht zu Verlnderungen des Lysogenisierungsvermogens fiihren, wenn auch noch andere Ursachen in Frage kommen. Inzwischen haben aber Ver- suche gezeigt ( KAPLAN, unveroffentlicht), daB die Lysier- sowie Lysogenierfahigkeit der c- und 1-Mutanten gegeniiber dem Wildtyp iiberraschenderweise nicht merklich geandert ist. AuBer solchen c- und 1-Mutationen, die etwa im Verhaltnis 2 : l auftreten, finden sich andere Mutationstypen nach UV nur auBerst selten. Diese betreffen Veranderungen des violettrot gefarbten Hofes, welcher den Plaque in dem gelbrot pigmentierten Rasen der Indikatorbakterirn (Stamm HY von S-rratia marc.) umgibt. Solche Farbhofmutationen sind nach UV-Bestrahlungen bisher unter vielen Tausenden von c- u. 1-Mutationen nur in wenigen Exemplaren gefunden worden. Es scheint also zunachst, daB das UV seh r e l e k t i v nur die c- u. 1-Gene zur Mutation bringt, wahrend das iibrige Genom weitgehend UV-resistent hinsichtlich Mutation erscheint. Eine andere Hypothese ware, daB das Phagengenom zwar in allen seinen Teilen etwa gleichmaBig UV-ssnsibel ist, aber die iibsrall induzierten Mutationen sich
*) Dr. BOSE: z. Zt. c/o Dr. H. E. SWIM, Dept. of Microbiol. Western Reserve Univ. Cleve- land/Ohio, USA.

Mutations- Auslosung bei extrazelluliiren Phagen 275
unter den angewandten Bedingungen (Plattungsmethode, Indikator u. a.) phanoty- pisch fast ausschliel3lich als Bnderung der Lochtrubung auszuwirken vermogen. Um zwischen diesen beiden Hypothesen zu entscheiden, besteht ein Weg darin, ein niu- tagenes Agens anzuwenden, welches sicher nicht nur eine bestimmte Region des Ge- noms mutieren l&&, sondern Mutationen weit verstreut iiber das ganzr Genom erzeugt. Wenn die 2. obige Moglichkeit nicht zutrifft, also die Mutationen des Phagen Kappa sich nicht nur vorwiegend als geanderte Lochtrubung, sondern haufig auch als Bnde- rung des Farbhofes und anderer Merkmale auszuwirken vermogen, so sollten mit einem solchen nicht-elektiven Mutagen aul3er c- u. 1- auch andere Mutationstypen in grol3erer Zahl ausgelost werden.
Als ein Mutagen dieser Art kommt N i t r i t in Frage. Dieser Stoff lost Mutationen varschiedenen Typs beim TIM-Virus (MUNDRY u. GIERER 1958) und beim Phagen T2 (VIELMETTER u. WIEDER 1959) durch Dasaminierungan an den Basen der Nuklein- sauren aus.
In der DNS wird dabei Adenin (A) in Hypoxanthin (HX), Guanin (G) in Xanthin (X) u. Cytosin ( C ) in Uracil (U) uberfuhrt. Thymin bleibt unverlndert, da es keine NH,-Gruppe besitzt. Versuche von VIELMETTER u. SCHUSTER (1960) zeigen, daB die Mutationen bei T 2 wohl vorwiegend durch die Umlagerung A -+ HX sowie auch C (hier HMC) -+ U verursacht werden, wahrend der flbergang G+ X Abtotung bewirkt. Da A u. C nicht grob ungleich- maI3ig uber das Phagengenom (DNS) verteilt sein diirfte, sollten alle moglichen Gene vom Nitrit mutiert werden. Das wird durch die Verschiedenartigkeit der gefundenen Plaquemutationen bei T 2 bestatigt.
Es wurde daher in den unten geschilderten Versuchen der extrazellulke Phage Kappa der Wirkung von Nitrit ausgesetzt uiid die erscheinenden Plaquemutanten qualitativ und quantitativ erfal3t. Nebenher wurde die Inaktivierung des Wildtyps sowie einiger der isolierten Mutanten untersucht, wobei sich z. T. geringe Unter- schiede in der Inaktivierungsrate zeigten. Das Auftreten sehr vieler Farbhofmu- tanten neben c- u. 1-Typen nach Nitritbehandlung zeigt eine starke Elektivitat des U V fur die Auslosung von c- u. 1-Mutationen an und bestatigt damit die erste der obigen Hypothesen.
I I . Methodik Die mit Nitrit behandelten Suspensionen des Phagen K a p p a waren durch
Vermebrung in flussiger NBhrbouillon (NB) (Difco Nutrient Broth) mit dem alyso- genen lndikatorstamm HY (unter Ablauf von 3-4 Infektionsrunden wahrend der Kultur) gawonnen. Die Phagen der steril-filtrierten NB-Lysate wurden durch mehr- faches hochtouriges Zentrifugieren und Resuspendieren in Puffersaline nach HERS- HEY (1952) gewaschen. Der Titer betrug dann um lOl1/ml.
Das Reakt ionsgemisch bestand aus 7,4 ml 0,25 mol. Azetatpuffer + 0,l ml Phagensuspsnsion + 2,5 ml25% Essigsaure + 0,l ml1,25 mol. NaN0,-Losung. Die NaN0,-Konzentration war also 0,0125 mol. Der pH-Wert des Gemisches lag bei 4,O und blieb wahrend der Reaktionszeit bis 20min praktisch konstant. Die Re- aktionsmischung wurde im Wasserbad mit Ruhrwerk auf 30 "C gehalten. Nach den angegebenen Zeiten wurden aus ihr Proben entnommen und sogleich 1 : 100 in Puffer- saline verdunnt und, gegebenenfalls nach weiterer Verdunnung, ausgeplattet. Die Plattung geschah nach der ublichen Weichagarschicht-Methode 1 (9. ADAMS). Die 3 ml Weichagar (0,5% Agar) wurden mit 0,l ml (verdunnter) Phagensuspension + 0,2 ml einer beluftet zur Sattigung gewachsenen ubernachtkultur des Indikator- bakteriums Serratia marc. HY beimpft, die durch Verdiinnung ca. 1 : 7 auf den Titer 1 - lo8 Zellen ml eingestellt worden war. Als Grundschicht dienten 20ml Difco- Nutrientbrothagar. Das mit der Phagensuspension eingebrachte Nitrit war auch bei
19u I
2
3
4
5
15 %o
Mid.
I0
5 70 15 20 h I I I I
min
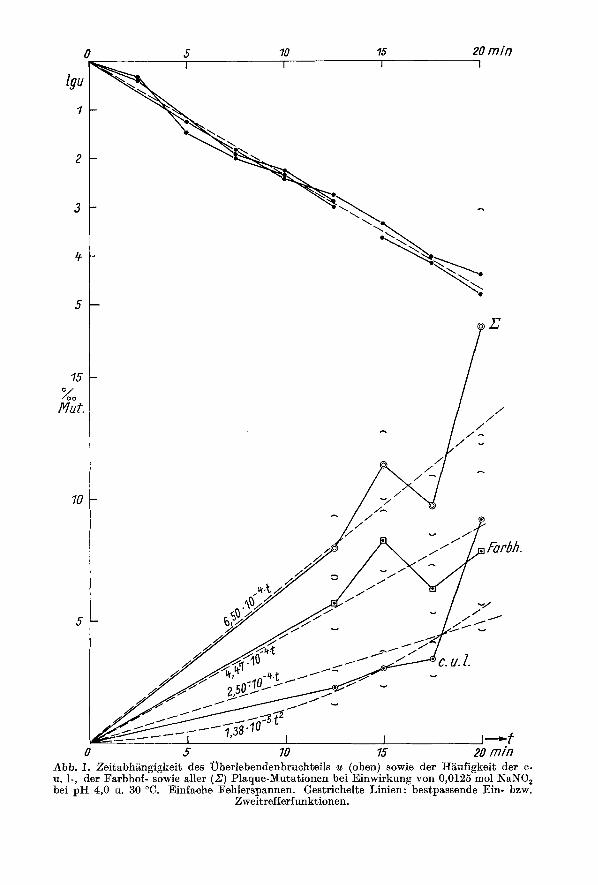
0 5 70 15 20 min Abb. I. ZeitabhBngigkeit des uberlebendenbruchteils u (oben) sowie der Haufigkeit der c- u. 1-, der Farbhof- sowie aller (Z) Plaque-Mutationen bei Einwirkung yon 0,0125 mol NaNO, bei pH 4,0 u. 30 "C. Einfache Fehlerspannen. Gestrichelte Linien: bestpassende Ein- bzw.
Zweitrefferfunktionen.

Mutations-Ausliisung bei extrazellularen Phagen 277
den schwachst verdunnten Suspensionen so verdunnt, daB es nicht mehr merklich auf Bakterien oder Phagen wirken konnte.
Zur Auszahlung kamen nur solche Platten, die nicht mehr als einige Hundert Plaques enthielten, so daD dieplaques gut getrennt waren und moglichst alle Mu- tantentypen identifiziert werden konnten. Serien von Parallelplatten dienten dazu, eine genugend groDe Zahl priifbarer Plaques zu erhalten. Nur bei 20 min Behand- lungsdauer war die uberlebendenrate so gering, da13 die erwunschte Zahl von > 1000 gepriiften Plaques nicht erreicht wurde. Bei den langeren Behandlungsdauern ent- standen zunehmend viele & kleine Plaques, z. T. mit nicht deutlich ausgebildetem Loch und daher nur als kleine violette Farbflecken erscheinend. Nachkulturen zeig- ten, daD sie keine Mutanten darstellen, also wohl auf Latenzzeitverlangerung beruhen. Da deren Zuordnung zu den Mutantentypen unsicher war, wurden fur die Peststellung des Mutantenbruchteils nur die groDen, mit einem deutlichen Loch ausgestatteten Plaques ausgewertet.
Fiir die Nachku l tu ren von Mutantenplaques wurden nur solche verwendet, die mindestens 1 cm vom nachsten Plaque entfernt waren, denn friihere Versuche hatten ergeben, da13 vereinzelte Phaganpartikel bis etwa 3 mm iiber den Lochrand hinaus- diffundieren, somit zur Vernnreinigung benachbarter Plaques und dadurch zur Vor- tauschung von Mischplaques (Heterozygote oder Labilmutierte) fuhren konnen. Die Nachkulturen wurden so hergestellt, daD rnit dem Impfdraht das Zentrum des Loches angestochen und dann die Drahtspitze in 10 ml Puffersaline abgespult wurde. Die so hergestellte Suspension wurde 10-1 verdunnt, 'der Verdiinnung sowie der Stamm- suspension 0,l ml entnommen und mit Weichagar + Indikator wie iiblich Schicht- platten gegossen. Auf diesen erschienen nach Bebriitung wenige Hundert bis etwa lo3 Nachkommenplaques. Die 2. Nachkulturen wurden in gleicher Weise aus Ein- zelplaques dieser 1. Nachkulturen hergestellt. Sofern diese Nachkulturen genetisch rein waren, wurden von denen mit sehr dichtem, z. T. konfluierendem Plaquebewuchs (2 103) Phagensuspens ionen du rch Abschwemmen des Weichagars mit 10 ml NB, Schutteln mit Glasperlen, 4-5 h Bebriitung, falls die Plaquezahl < lo3 war, und Sterilfiltration gewonnen. Der Titer dieser Mutantensuspensionen lag meist zwischen 1O1O und 101l/ml. Solche ungewaschene Abschwemmsuspensionen von Mn- tanten wurden in den Mischungsversuchen vermengt und zusammen mit ge- waschenen Wildtypphagen rnit Nitrit behandelt. Es zeigte sich, daB die Nitritwirkurig von der geringen Menge an NB-Lysatstoffen im Reaktionsgemisch 0, l ml Phagen- mischung + 10 ml Nitrit-Azetatpuffer nicht merklich beeinflufit wurde.
III. Ergebnisse
1. Inak t iv i e rung des Wild typphagen x+
Die Ergebnisse uber die Inaktivierung des Lochbildungsvermogens unabhangig von der Einwirkungsdauer des 0,0125 molaren Nitrit bei pH 4 und 30" sind aus Abb. 1 zu ersehen. Der Logarithmus des Uberlebensbruchteils (lg u = Ig (N/NO)) fallt iiber fast 5 Zehnerpotenzen etwa linear mit der Zeit t ; die Inaktivierung eines Phagen- partikels geschieht also schon durch einen ,,Treffer" eines Nitritmolekels. Der Phage Kappa verhalt sich also insofern wie die ubrigen von anderer Seite studierten Phagen (T 2 u. P 22; VIELMETTER und WIEDER, VIELMETTER und SCHUSTER). Die Totungs- rate errechnet sich zu z = 0,54 Inaktivierungen pro Partikel und Minute. Nach VIEL- METTER und SCHUSTER ist der Wert fur Phagen T 2 etwa 10 Letaltrefferlmin, fur Phage P 22 etwa 20 Trefferlmin bei 20°, 1 mol NaNO, u. pH NN 4,O. Da das Nitrit konzentrationsproportional wirkt, ergibt sich 0,l min fur T 2 und 0,2 min fur P 22 bei 0,Ol mol. Da die Temperatur in unseren Versuchen um 10" hoher lag als in jenen

278 R. W. KAPLAN und S. K. BOSE
Versuchen, ist zu schlieoen, dal3 die Inaktivierungssensibilitat dcs Phagen Kappa nicht erheblich von der der anderen beiden Phagen abweicht, vielleicht ein wenig hoher ist.
2. Auslosung verschiedener Muta t ions typen Tab. 1 enthalt die Befunde iiber die bei variabler Einwirkungszeit (12,5-20 min)
beobachteten Mutanten. Der Wildtypphage X + erzeugt auf dem gelblichroten Rasen des Indikatorbakteriums HY stark triibe Locher von 1-2 mm Durchmesser, die von einen intensiv violettroten Farbhof von etwa 1-2 mm Breite mit diffuser AuBengrenze umgeben sind. Die Triibung des Loches ist durch viele darin wachsende winzige weil3e Kolonien verursacht, die wohl aus den lysogen infizierten Bakterien entstehen. Da lysogene Bakterien HY(x) normalrote Kolonien bilden, ist die weil3e Kolonienfarbe im Loch wold durch Hemmstoffe verursacht. Es wurden nach Nitrit- behaiidlung von 1c' folgende phanotypischen Gruppen von Mutan t e n gefunden und differenziert :
e: klares bis fast klares Loch, normaler violettroter Hof. 1: mehr oder weniger schwacher alsnormal ge t r i ib tes Loch , normaler Hof . z: etwa normaltriibes Loch, Hoffarbe f gelbrot (zinnober), Hof oft scharfer begrenzt und bisweilen schmaler. dz: iihnlich z, jedoch Hoffarbe dunk le r und intensiver rot. b: etwa normal triibes Loch, Hof aber 2 blasser als normal. 0 : etwa normal triibes, jedoch meist & kleineres Loch, ohne oder rnit nur sehr schwachem Hof. Da die zu einer Typengruppe gezahlten Mutanten untereiiiander, auch in Nachkul-
tur, nicht gleich waren, gehoren sie wohl zu verschiedenen Genotypen (Genen oder Allelen). Besonders heterogen erscheint die Gruppe der b-Mutanten. AuSer den aufge- fiihrten Mutantentypen erschienen noch wenige andere, z. B. rnit unregelmaoig ge- zacktem Hofrand oder blaSzinnober Hof (bz), die in Tab. l unter ,,sonstige" gezahlt sind. Die Manifestation des Phans der Mutanten ist von der Plattungsmethode erheblich abhangig. Insbesondere zeigten sich die Farbhofmutanten in ihrem Aus- sehen beeinflnl3bar von der Dichte der Indikatorbakterien im Weichagar, von der Dicke der Weichagarschicht und von der 0,-Versorgung wahrend des Wuchses des Indikatorrasens. Die Beschreibungen gelten daher genau nur fiir die obige Plattungs- methode.
In der Tab. 1 sind die G und 1 zur Gruppe der Lochtriibungsmutanten, die dz und z als Farbton-, und die b und o (+ die wenigen sonstigen) als Farbgradmutanten zu- sammengruppiert. Wie die unterste Zeile (Summen) ausweist, machen die c + 1-Mu- tanten nur etwa ein Drittel aller durch Nitrit induzierten Typen aus, wahrend die Hofmutan ten ( F a r b t o n + Farbgrad ) doppe l t so hauf ig siiid wie diese Locht r i ibungsmutanten . Da nach UV-Bestrahlung Hofmutanten nur als sel- t-ne Ausnahme entstehen - bisher nur einige neben mehreren tausend von c u. l - ist es sicher, daB das UV die c- und 1- Gene m i t sehr hoher E l e k t i v i t a t zu r Muta t ion br ing t.
Das Verhaltnis der c- zu den 1-Mutanten ist nach Nitritwirkung etwa 2:1, und mit diesem Haufigkeitsverhaltnis erscheinen beide auch durch UV. Die ,,Treff- wahrscheinlichkeiten" fur Nitrit und UV sind hinsichtlich dieser c- und 1-Gene also nicht unterschiedlich, wahrend die Treffwahrscheinlichkeiten bezuglich der Farb- hof-Mutationen fiir UV im Vergleich rnit denjenigen fur Nitrit extrem gering sind. Innerhalb der Gruppe der Triibungs-Gene wirkt also das U V nicht elektiv, gegeniiber den ubrigen Genen zeigt es jedoch nur auBerst geringe Wirksamkeit.
Mutations-Auslosung bei extrazellularen Phagen 279
a 0 0 rxg G
.. -i L-v 'Ij m a
3. Pri i fung d e s Einf lusses se lek t iver To tung der
Mutan ten
Wic Tab. 1 und Abb. 1 er- kennen lassen, nimmt der Bruchteil an Mutanten unter den Uberlebenden mit der Nitritdosis (Behandlungszeit) zu. Diese Zunahme konnte auBer Mutationsauslosung auch durch selektiv besseres uberleben, also hohere Nitrit- resistenz, von spontan in der Phagensuspension vorhan- denen Mutanten verursacht sein. Da bisher spontane c-Mutanten (jedoch kein an- derer Typ) in unbehandelten Suspensionen nur mit der Haufigkeit von etwa ge- funden wurden, ist solche Se- lektion a priori nicht sehr wahrscheinlich.
Dieszeigt eine kleineRechnung: Es sei m, der spontane Mutanten- bruchteil und m der nach Behand- lung gefundene, u die Uberleben- denrate des Wildtyps und ~11.1 die der Mutauten. Daun ist, falls m
und m, < 1, m w ?no % undalso m
UM w u - . Fur gegen 1 merk- ma
liche m und m,, gibt
U
Fur die experimentell gefunde- nen Werte u = 10-4, m = 4 . mo = lov5 ware dann UM = 4 lop2 ; wenn also der Wildtyp nach 17,5 min auf u = abgetotet ist, durfen die c-Mutanten nur auf 4 . lop2 getotet worden sein, falls ihre gesamte Zunahme (auf 4"/,,) auf Selektion beruhen wiirde. Sie muaten also sehr vie1 nitrit- resistenter als der Wildtyp sein.
Um zu priifen, ob solche Selektion beteiligt ist, wurden 2 Versuche durchgefiihrt, in denen Mischungen verschie- dener Mutanten mit Normalen behandelt und die Veriin-

280 R. W. KAPLAN und S. K. BOSE
__ 37,l 38,9 38,l 44.0
derung der Mutaiitenanteile durch die Behandlung festgestellt wurden. Das Er- gebnis eines dieser Versuche ist in Tab. 2 wiedergegeben. Beide Versuche ergaben keine signifikante Zunahme des Anteils der Mutanten, sodaB der Anstieg des Mutanten- gehaltes durch Nitrit sicher auf Muta t ionsaus losung durch den Stoff im freien Phagen, nicht auf Selektion beruht.
I n dem einen Versuch mit 1, dz und o-Mutanten geschah iiberhaupt keine merkliche Veranderung des Bruchteiles der Mutanten (ca 40%) bis zum uberleben u = 10-29s7; demnach werden diese Mutanten gleich stark wie der Normaltyp abgetotet. Der in Tab. 2 dargestellte Versuch dagegen zeigt eine leichte Zunahme der Normalen und
32,7 18,9 11,3 33,4 17,4 10,2 32,2 18,O 11,s 37.1 13.0 5.4
Tabelle 2 EinfluB selektiver Totung in einer Mischung von Wildtyp und Mutanten
Behand1.-Zeit in rnin
0 295
795 10 12,5
5,o
log Bruchteil uberlebender
k u -. .
0 -0,34 -1,47 -2,oo -2,37 -2,99
Gesamtzahl gepriifter Plaques
159 688 644 855 652 201
entsprechend eine leichte Abnahme der zugemischten z-Mutante, vielleicht auch der Mutante dz,. Dies deutet auf negative Selektion, also Ausmerzen dieser Mutanten infolge starkerer Totung als die Normalen. Fur die z-Mutante la& sich die Abnahme statistisch gut sichern, wenn man die Werte fiir 0 + 2,5 + 5,O min zu- sammenfaot (266/1491 = 17,9 f 0,99%) und mit dem entsprechenden Summen- wert fur 7,5 + 10 + 12,5 min (239/1708 = 14,O 5 0,85%) vergleicht. Es ergibt sich dann ein x = A/o = 3,9/1,30 = 3,O mit Zufallswahrscheinlichkeit P = 0,27%. Die Abnahme der dz bei gleicher Rechnungsart (11,O & 0,81 auf 8,55 f 0,6S%) sowie die entsprechende Zunahme des Wildtyps (38,4 1,22 auf 42,2 f 1,20%) zeigen sich mit P = 2,0% bzw. P = 2,5% als weniger gut gesichert, wenn auch immerhin als recht wahrscheinlich auBerzufallig. Die in der Mischung untersuchte z-Mutante, vielleicht auch die dz, wird also vom Nitrit etwas starker abgetotet als der Normltyp x+. Einerseits ist damit gesichert, daR all diese Mutanten infolge der Nitritbehand- lung neu durch Mutation entstanden sein miissen ; andererseits zeigt sich hier das interessante Problem, daB e in M u t a t i o n s s c h r i t t (x '+ x/z) niclit nur zur Bnde- rung des Plaquephans fiihrte sondern auch zu etwas hohe re r Sens ib i l i t a t d e r z -Pa r t ike l f u r I n a k t i v i e r u n g d u r c h N i t r i t . Die VerstLrkung der Inaktivierung von x/z ist allerdings gering, sie 15Bt sich rnit der Mischungsmethode gerade noch nachweisen, ware jedoch nur muhsam durch die Aufnahme der Inaktivierungskurve zu finden gewesen.
Die GroBe der Anderung der Inaktivierungskonstante z bei der Mutation + -+ z ergibt sich aus folgender Rechnung : Der Mutantenanteil nach Nitriteinwirkung ware
M , e-il t 1 m=----------
1 + -3 e A t (l ) Ho e-zlt + A o e-rt -
wobei M , die Zahl der Mutanten-, A , die der Nichtmutantenpartikel in der Misehung vor der Behandlung, ma = M,/(M, + A,) der Ausgangs-Mutantenanteil, t die Behandlungszeit, z die Totungswahrscheinlichkeit pro min fiir Nichtmutan-

Mutations- Auslosung bei extrazellularen Phagen 281
dz,
dz,
1* 11 0 2
bl b2
ten, z1 fur Mutanten und A = zl--t die Zunahme der Totungswahrscheinlich- keit infolge der Mutation x' -+ XIZ bedeuten. Durch Umformung obiger Gleichung
ergibt sich A = r l g 7 * ~--m, mo ) . Setzen wir die Experimentalwerte fur die
z-Mutante m, = lS,9yo und m = 12,0y0 bei t = 12,5 min (lg u = - 3,O) ein, so ergibt sich eine Zunahme d e r To tungskons tan te f u r die z -Mutante von A = 0,043 min gegenuber der Totungskonstante des Wildtyps von z = 0,54 min (s. o.), also um e t w a 8%.
2230 r -*
90% dz, 10% + 90% dz, 10% + 25% c, 72% + meist 1, wenige + meist 0, 0,3% + meist b, wenige yo + meist b, wenige yo +
4. Die Nachkommenschaf t f r i sch mut i e r t e r Phagenpar t ike l i n P l a qu e-Na c h k u l t ur e n
Nachkulturen von frisch durch UV induzierten c- oder 1-Mutantenplaques zeigjti fast immer nur den Mutantentyp. Unter 83 c- und 12 1-Mutanten-Nachkulturen ent- hielten nur 4 c aul3er dem Mutantentyp noch einige yo Wildtypplaques; diese erschie- nen in der 2. und weiteren Nachkulturen nicht wieder (KAPLAN 1960 und unveroff.). Es handelt sich bei letzteren Ausnahmen also wohl um ,,heterozygot" mutierte Pha- gen, also mit Mutation in nur einem der beiden Nukleotidstriinge der DNS. W erzeugt demnach ganz vorwiegend ,,homogene" Mutationen, d. h. solche, in denen entweder beide DNS-Strange mutiert sind oder in denen einer eine Mutation, der andere einen ,,rezessiven'' Letalschaden enthalt.
Von insgesamt 30 frisch durch N i t r i t induzierten Mutantenplaques verschiedenen Typs (7 c, 6 1,8 dz, 1 z, 1 bz, 4 b, 3 0) ergaben 22 in Nachkulturen iiur den Mutanten- typ, 8 enthielten Beimischungen, meist vom Wild-(+)Typ. Die Befunde uber die heterogenen Nachkulturen sind in Tab. 3 zusammengestellt. In jeder Nachkultur wurden viele hundert, oft uber 1000 Plaques, z. T. in Parallelplatten, gepriift.
Die in der Tab. 4 zuoberst aufgefiihrten 4 Mutanten zl, dz,, d, und ,,li ' sind als ,,heterozygot" anzusehen, da der Mutantentyp in der 2. Nachkultur nicht mehr ,,spaltet". Der frisch induzierte Plaque J,," machte wegen leichter Trubung zunachst den Eindruck einer 1-Mutante, erwies sich jedoch in der Nachkultur als Mischung von c- und f-Typ. Da hier der +-Typ uberwiegt, hat er beim Wuchs im Plaque wohl selektiven Vorteil gegenuber den c-Phagen. Bei den ubrigen heterozygoten Mutanten erscheint der Wildtyp im Nachteil.
Tabelle 3 Beschaffenheit der heterogenen Nachkulturen
(%-Angaben meist geschatzt)
2. Nachkulturen
von z: nur z von +: nur + von dz: nur dz von +: nur + von dz: nur dz von +: nur + von c: nur c von 1: 94% 1, 5% c, 1%+ von 0: meist 0, 0,l% + von +: nur + von b: meist b, wenige yo + von b: meist b, wenige yo +
3. Nachkultur von b: 98% b, 2% +

282 R. W. KAPLAN und S. K. BOSE
Bei den 4 folgenden Mutanten l,, b,, b, und o2 ,,spaltete" der Mutantentyp in der 2. Nachkultur wiederum in gleicher Weise wie in der ersten. Der die Heterogenitat erzeugende Zustand wird hier also vermehrt, weshalb keine echte Spaltung aus einem partiell mutierten (heterozygoten) Genom vorliegen kann. Es liegt also ein m u t a bles Allel vor, welches wahrend der Vermehrung spontan mit hoher Rate zum +-ahn- lichen Typ weiter- (,,zuriick-") mutiert. Der Anteil dieser Weitermutanten betragt in der 2. (u. 3.) Nachkultur nur wenige Prozent, was auf ihre kleine Entstehungs- chance (Mutation) deutet. Bei 0, erwies sich der +-Typ als stabil, bei den anderen wurde er nicht gepriift. 1, mutierte zu 2 verschiedenen Typen (Allelen '1) weiter: ,,zuriick" zu + und ,,vorwarts" zu c. (In der 1.Nachkultur wurden die wenigen c- wohl unter den 1-Plaques iibersehen.) Dies Verhalten kann durch ,,Heteroiygotie" uberhaupt nicht erklart werden, es kommt hier nur Labilmutation in Frage.
5 . Ab h a n g i g k e. i t de r Muta t ion s h a u f ig k e i t v on d e r Be h a n d l u n g sz e it Die Versuche waren nicht ausdriicklich zur Feststellung der Zeitabhangigkeit der
Mutationsausbeute angelegt, sondern vor allem zur Klarung der Elektivitatsfrage. Trotz des dadurch bedingten Fehlens niederer ,,Dosen" (kurzer Behandlungszeiten) der Nitritwirkung erlauben sie die Zeitabhangigkeitskurve aus den gewonnenen Daten festzustellen. Dadurch ergaben sich Schliisse auf die Anzahl derNitrittreffer und also Desaminierungen, die fur die Initiierung einer Mutation notig sind. Abb. 1 und Tab. 1 enthalt die Befunde. Die Zeitabhangigkeit der Summe, aller Mutationstypen (,Pin der Abb. 1) zeigt einen starken Ausschlag des Punktes bei 20 min, der vermuten la&, daB vielleicht keine Ein-, sondern eine Zweitrefferkurve zugrunde liegt. Aller- dings ist wegen der geringen Stichprobengrofle die Fehlerspanne (a = y)p(l- p) /n ) dieses Wertes sehr groB. Die Punktfolge der Hiiufigkeiten nur der c + 1-Mutationen zeigt, daB der Ausschlag bei 20 min nur durch diese Mutationsgruppe verursacht ist ; die Farbhofmutationen deuten eher auf eine Eintrefferfunktion. Zur Entscheidung zwischen 2- und 1-Trefferabhangigkeit wurden X2-Rechnungen herangezogen.
Da die Mutantenhaufigkeiten (m = p t im Eintrefferfall und m = p2 t2 im Zweit- trefferfall, p = Trefferwahrscheinlichkeit/min) 1 yo nicht erheblich uberschreiten, kann mit Poissonverteilung gerechnet werden. Zur Berechnucg der bestangepaBten (effizienten) Mutationskonstanten p bzw.pa dient die met.hodof maximum likelihood (z. B. MATHER). Aus der nach ihr anzusetzenden Maximumbedingung bei Eintreffer-
funktion ____ = - 2 (ai In p ti - ni ,u t ) = 0 ergibt sich p = 2 a i / 2 n, ti ,
wobei uSi die gefundene Zahl Mutanten, ni die gepriifte Gesamtplaquezahl in der zu Zeit ti gehorigen Stichprobe bedeuten, die aus Tab. 1 zu entnehmen sind. Analog ergibt sich das bestpassende p2 = 2 a i / 2 ni t! fiir die Zweitrefferkurve. Zunachst werden die mit diesem p bzw. p2 zu erwartenden Werte fur die theoretischcn Mu- tationshaufigkeiten bei Auslosung durch 1 Treffer mth = p t bzw. durch 2 Treffer mth= = $2 berechnet; dann werden deren Abweichungen A i = mi - m i t h gegenuber den Experimentalwerten mi sowie die mittleren Fehler dcr letzteren (q = lmi (1 -nii)/ni) bestimmt. Daraus ergibt sich schliefllich das x2 = 2 (Ai/ai)2 fur die ganze Versuchs- reihe von i Proben zu den Zeiten ti (12,5, 15, 17,5 und 20 min).
Da Abb. 1 ein vielleicht unterschiedliches Verhalten der Lochmutationen c + 1 und der Farbhofmutationen andeutet, wurden beide Mutationsgruppen getrennt auf die Trefferzahl ihrer Induktion hin gepriift. Die F a r b h o f m u t a t i o n e n liefern fiir die Eintrefferfunktion p = 134/30,06 - = 4,47 . lO-*/rnin. Das zugehorige xi = 4,42 entspricht einer Zufallswahrscheinlichkeit von P = 22%. Die Befunde sind also innerhalb des Zufallsbereiches mit der E in t r e f f e r theo r i e vertraglich.
d Z h Wi d dP
____ -

Mutations- Auslosung bei extrazellularen Phagen 283
Priifen wir analog die 2-Trefferhypothese, so ergibt sich ,u2 = 2,82. 10-5/min und damit xi = 11,21. Die zugehorige Zufallswahrscheinlichkeit P = l , O % entscheidet gegen die Entstehung dieser Mutationen durch 2 Nitrittreffer. Die entsprechenden Rechnungen fur die c + 1-Mutationen liefern einerseits ,u = 2,5 . low4 ur,d xi = 5,'i bzw. P ='13%, also Moglichkeit der Eintrefferfunktion, andrerseits ,u2 = 1,38 . 10-5 und xi = 2,21 mit P = 54%, also auch Zulassigkeit der Zweitrefferfunktion. Unter- suchen wir die Gesamtheit aller Mutationstypen, so erhalten wir pz = 6,9 . l0-4/min sowie xi = 3,79 u. P = 28% fur die 1-Trefferhypothese, xi = 12,l u. P = 0,7% fur die 2-Trefferhypothese. Die Analyse zeigt also, da13 alle oder wenigstens die meisten Muta t ionen d u r c h einen Ni t r i t t r e f f e r entstehen. Bei den c + 1-Mu- tationen kann allerdings die Beteiligung von 2-Trefferereignissen nicht ausgeschlossen werden, wenn auch hier 1 Treffer durchaus moglich und wahrscheinlich ist. Dies Ergebnis am x-Phagen entspricht somit dem von VIELMETTER und WIEDER uber die Erzeugung von r-Mutationen beim Phagen T 2.
Der Vergleich der jetzt gefundenen Chancen p fur einen Mutationstreffer pro Zeit- einheit und die oben berechnete fur einen Totungstreffer z = 0,54/min ergibt fol- gendes: Betrachten wir die Gesamtheit aller festgestellten Plaque-Mutationstypen, so ist das Verhaltnis von Totungs- zu Mutationstreffwahrscheinlichkeit z :,uz = 0,54 : : 6,9 - = 8,83. Auf die Auslosung einer c-oder 1-Mutation kommen analog 0,54:2,5 - = 2160 Totungstreffer. Die Versuche obiger Autoren beim Phagen T 2 ergaben ca 260 Totungstreffer auf 1 r-Mutationstreffer bei pH 4,O. Da die Zahl dcr an den r- wie auch an den c- und 1-Mutationen beteiligten Gene klein ist und ahnlicher sein diirfte als die an den gesamten Plaquemutationen von x, erscheint der Vergleich des Wert>es von T 2 mit dem der c- und 1-Mutationen von x sinnvoller als mit dem fur alle x-Plaquemutationen. Demnach ist der Phage x f u r die Aus- losung von Muta t ionen r e l a t iv zu Inak t iv i e rungen d u r c h N i t r i t erheb- lich unempf indl icher a l s der P h a g e T 2.
I V . Folgerungen
Die eingangs dargelegte Hauptfrage nach der extrem e lek t iven Auslosung von c- u n d 1-Mutat ionen durch UV ist durch die Ergebnisse positiv beantwortet: Nitrit erzeugt etwa doppelt soviele Farbhofmutationen verschiedenen Typs wie c- und 1-Mutationen. Wenn W andere als Lochtrubheits-Mutationen induzieren wiirde, miifiten sie unter den verwendeten Bedingungen erscheinen. Da sie nicht auftreten, wirkt W extrem elektiv auf diese Gengruppe. Wie WINKLER nachwies, liegen alle gepruften W-induzierten c-Mutationen im selben Funktionsgen (Cistron), die elektive Wirkung des Wr ich te t sich also vorwiegend auf dieses Gen. Andererseits ist nach W wie Nitrit das Verhaltnis c- : 1-Mutationen gleich (2 : 1) ; UV wahlt also nicht zwischen diesen Genen. Am leichtesten ware dies zu erklaren, wenn die c- und I-Gene eng be- nachbart in einer Region des Genoms besonders UV-sensibler Struktur lagen. Da auf Grund der Ergebnisse WINKLERS uber die Erhohung der Mutationsausbeute durch W-Bestrahlung des Wirtes die Moglichkeit besteht, da13 die W-induzierten c- und 1 keine :,echten" Mutationen sind, sondern durch Rekombination mit dem Wirts- genom entstehen, konnte diese c- und 1-Region durch besonders intensive Paarung mit dem Wirt ausgezeichnet sein. Allerdings verursaohen, wie einleitend dargelegt, diese Mutationen keine verminderte Lysogenisation wie die clearplaque-Mutationen des PhageniZ (KAISER), welche vielleicht an der Anheftungsstelle des Prophagen an das Wirtsgenom mutiert sind. Falls aber die W-induzierten c und 1 echte Mutationen ohne einrekombiniertes Wirtsmaterial sind, wofiir z. B. die Ruckmutabilitat spricht, miil3te die Besonderheit der c-1-Region andere Ursachen haben (z. B. konnte in ihr

284 R. W. KAPLAN und S. K. BOSE
eine ,,Storstelle" in der DNS-Periodizitat liegen, die als Energiefalle wirkt). Moglicher- weise liegt eine Analogie zu der hohen UV-Mutabilitat der Pigmentbildungsgene von Serratia vor.
Die Nachkulturen frisch induzierter Mutationsplaques ergaben einen Anteil , ,heterozygoter" Muta t ionen . Diese diirften wie die ,,heterozygoten" r von T 2 (VIELMETTER und WIEDER) auf Desaminierung nur einer Base eines Paares in der DNS beruhen. Ihr Anteil ist mit 4 unter 30 (= 13 & 6,2%) wohl niedriger als der Anteil heterozygoter r-Mutationen von T 2, wo sie 3 0 4 0 % ausmachen. Die Mu- tationen mit homogener Nachkultur gehen wahrscheinlich auf Phagengenome zuriick, die in dem einen DNS-Strang einen Mutations-, im anderen einen ,,rezessiven" Letaltreffer erhalten haben (der die Vermehrung nur dieses Stranges verhindert, wahrend die Inaktivierungstreffer die Vermehrung des ganzen Genoms hemmen, also ,,dominant" sind). Der hohere Anteil Homogenmutierter bei x deutet auf etwas starkere Auslosung von ,,rezessiven" Lasionen relativ zu Mutationen als bei T 2.
Dies ist eine Parallele zu dem Befund, daR auch das Verhaltnis der I n a k t i v i e r u n - gen z u den c- und 1-Muta t ionen von x mit ca. 2100 hoher liegt als das von In- aktivierungen zu r-Mutationen bei T 2 (260). Nach den Ergebnissen von VIELMET- TER und SCHUSTER bei verschiedenem pH ist anzunehmen, daB Inaktivierungen vor- wiegend durch Desaminierung des Guanin, Mutationen durch solche des Adenin und Cytosin entstehen. Man konnte daher vermuten, daB der %-Phage relativ mehr Guanin enthalt als T 2 . Das (A + T):(G + C)-Verhiltnis in der DNS von T 2 ist etwa 1,s - 2,0 (ADAMS, p. 91 ; VIELMETTER und SCHUSTER), dieser Phage ist also relativ arm an Guanin. Serratia hat ein Verhaltnis von CIL. 0,76 (cit. in RAVIN), und da temperierte Phagm Lhnliche Verhaltnisse haben wie ihre Wirte (LANNI) diirfte auch Kappa ein Verhaltnis < 1 haben, also relativ reich an Guanin sein. Die obigen Befunde iiber das Verhaltnis von ,,dominanten" bzw. ,,rezessiven" Letal- zu Muta- tionsdesaminierungen waren also auf Grund dieses DNS-Baus gut verstiindlich. Da die (,,dominanten") Inaktivierungen nach obigen Autoren vorwiegend durch Desami- nierungen des Guanins entstehen, konnte man fiir die ihrer Natur nach andersartigen, aber in x auch haufigeren ,,rezessiven" Letallasionen vermuten, daB sie durch Um- wandlungen des in gleicher Menge wie Guanin vorhandenen Cytosins entstehen. Die im Vergleich zu beiden Liisionen seltenen Mutationen waren d a m Desaminierungen nur des in relativ geringer Menge vorhaiidenen Adenins.
Das Anffinden von heterogenen Nachkulturen mit Mutanten, die auch weiterhin heterogene Nachkommen liefern, zeigt an, daB ein Teil der Nitritmutationen zu s tark-mutablen Allelen gefuhrt hat. (Vereinzelte Riickmutationen kommen auch in manchen ,,stabilen" Nitrit- oder UV-Mutanten vor.) Die Befunde stiitzen die Annahme, daR frisch mutierte Allele alle moglichen Mutabilitatsgrade zeigen konnen und daR das Vorwiegen der stabilen Allele in langer geziichteten Stammen oder in der Natnr durch ,,Sslektion" infolge Weitermutation der instabilen zu sta- bilen Allelen zustande kommt. Die hoch-mutablen unter den Mutanten von x ent- sprechen also den sektorierte Kolonien ergebendeii Mutationen von Serratia (KAP- LAN 1959). Es mag vielleicht an der DNS-Struktur dieses Bakteriums und seines Phageii liegen, daR solche Labilmutationen hier besonders haufig sind, wahrend sie in T 2 anscheinend nicht beobachtet wurden.
Die Selektionsversuche zeigteii an, daR die Mutante z (vielleicht auch dz) eine etwa 8% hohere Inaktivierungswahrscheinlichkeit besitzt als der Wildtyp. Da die Mutation durch Nitrit zu der chemischen Anderung nur eines Basenpaares unter tausenden in der DNS fiihrt, ist es nicht moglich, daI3 dieses Basenpaar (z. B. durch Zunahme des Guaningehaltes in der DNS) allein die hohere Inaktivierungsrate ergibt. Vielmehr muR es sich um einen sekundaren phanotypischen Effekt dieser

Mutations-Anslosung bei extrazellularen Phagen 2 85
BasenSnderung handeln, wobei am ehesten an eine veranderte Struktur des Hullen- oder Schwanzprotcins, daneben an eine veranderte Faltung der DNS im Phagenkopf zu denken ist. Diese konnte z. B. die Permeabilitat fur Nitrit verbessert haben. Aber auch eine Erhohung der Sensibilitat des Proteins, z. B. der Schwanzspitze, gegen Nitritwirkung komint in Frage. Nach HARM konnten etwa 1 / G m 17% der Inakti- vierungen a d Desaminierungen am Protein beruhen. Wiirde die Mutation z (neben- her) die Proteiniiiaktivierung andern, so wurde die Zunahme um 8% eine Erhohung der Proteinsensibilitat urn etwa die Halfte bedeuten.
Der Deutschen Forschungsgemeinschaft sei fur die Ermoglichung der Arbeit durch finanzielle Unterstutzung gedankt. Der Experimentator (S. K. BOSE) dankt den Mit- gliedern des Instituts fur Mikrobiologie der Universitat FrankfurtlM. fur viele Rat- schlage und fur die Aufnahme als Gast.
Summary
The inactivation as well as the induction of different mutations in the free temperate phage kappa of Serratia by nitrite are oneliit processes. Many different types of plaque-mutations are induced of which only are clearplaque-types (c + 1, which have no changed lysogenization). Since the latter ones are nearly the only types induced by UV-irradiation this radiation acts highly electivly on the c- and 1-genes. This may indicate some special structure of this genomic region of phage kappa. After 20 min of treatment by nitrite (0,0125 m NaNO,, p H 4,0, 30 "C) about 1,3% of different mutations appear. Reconstruction experiments show that they are induced, not merely selected. One of the induced mutants, z (cinnabar color halo), has an inactivation rate about 8% higher than the wild type, perhaps because of a changed protein coat. The inactivation rate (0,54/rnin) of the wild type is not much different from the rate of other phages (P22, T2). About 2100 inactivation hits are produced by nitrite for every c- or 1-mutation hit. This higher inactivation: mutation ratio compared with phage T2 (260 for r-mutations) can be understood if inactivation is mainly due t o deaminations of guanine and the Serratia-phage has a higher guanine content than T2, as the host bacterium does. Among 30 different mutants 4 were highly mutable and 4 were ,,heterozygous". Compared with T2 the ratio of ,,heterozygous" mutations appears smaller in kappa. This may be due to a higher rate of ,,recessive" lethal hits (leading to ,,homogenous" mutations if in the non-mutated DNA-strand) than in T2 and can be understood if ,,recessive" lethal lesions are produced mainly by cytosine or guanine deaminations.
L i t e r a t u r ADAMS, U. H., 1959. Bacteriophages. Interscience Publ. Inc. New York. ELLMAUER, H. u. KAPLAN,R. W. 1959. Naturwiss. 46, 150. HARM, W., 1960. Z. Vererbl. 91, 52-62. JACOB, F., 1954. C. R. Acad. Sci. (Paris) 238, 732. KAPLAN, R. W., 1959. Arch. f. Mikrobiol. 32, 138-60. KAPLAN, R. W., 1960. Internat. Congr. Photobiol. Copenhagen 1960. KAPLAN, R. W., WINKLER U. u. H. WOLF-ELLMAUER, 1960. Nature 186, 330. KAISER, A. D., 1957. Virology 3, 42. LANNI, F., 1960. Perspect. Biol. Med. 3, 418-32. MATHER, K., 1946. Statistische Analpen in der Biologie. London 1946: Methuen & Co.,
MUNDRY, K. W. u. A. GIERER, 1958. Z. Vererbl. 89, 614. PARK, S. W., 1961. Diss. Nat. Fak. Univ. Frankfurt/M. 1961. RAVIN, A. W., 1960. Bact. Rev. 24, 201-20. VIELMETTER, W. u. C. U. WIEDER, 1959. Ztschr. f. Naturforschg. 14b, 312-317. VIELMETTER, W. u. H. SCHUSTER, 1960. Zschr. f . Naturforschg. 15b, 304-311. WEIGLE, J. J., 1953. Proc. Nat. Acad. Sci. USA 39, 628. WINKLER, U., 1960. Unveroffentlichte Ergebnisse.
in Kommission bei Springer-Verlag, Wien.
20 Zeit,sclirift f . Allg. Mikrobiologie
























![Entwicklung einer API-Produktionsplattform mittels CRISPR ... · legte George P. Smith 1985 [15], indem er Fremd-DNA in das Genom filamentöser Phagen einfügte, die das Protein des](https://img.pdfslide.org/doc/110x75/6043c1e458e21d79be005561/entwicklung-einer-api-produktionsplattform-mittels-crispr-legte-george-p-smith.jpg)




