Embed Size (px)
Citation preview

Bau und Leben
extrem halophiler Archaea
Bachelorarbeit im
Studiengang Biologie der Heinrich- Heine-Universität
Düsseldorf
Chantal Jansen
Erstprüfer: Prof. Dr. Bickel Zweitprüfer: Prof. Dr. Alfermann
Bearbeitungszeitraum: 15.07.2007- 15.10.2007
Ratingen, den 15.10.2007

- 2-
Zusammenfassung Bei den halophilen Archaea handelt es sich um Bakterien, die unter extremen Lebensbedingungen gefunden werden. Sie benötigen zum Überleben eine Minimum Salzkonzentration von 9%. Bei einer maximalen Salzkonzentration von 32% ist Wachstum möglich, aber in einer verlangsamten Form. Sie werden hauptsächlich in Salinen, im Toten Meer, aber auch in unterirdischen Salzvorräten gefunden. Hier sind sie nicht nur hohen Salzkonzentrationen ausgesetzt, sondern auch hohen Temperaturen, Sauerstoffmangel und UV-Licht. Durch einen stabilen Aufbau der Membran und durch besondere Formen der Atmung wird es ihnen ermöglicht bei diesen extremen Bedingungen zu überleben. In meiner Arbeit beziehe ich mich zum größten Teil auf Halobakterium salinarum, da dies Halobakterium bis jetzt am weitesten erforscht wurde.
Schlagwörter: Halophile Archaea, extreme hohe Salzkonzentrationen, Totes Meer, 9% Salzkonzentration, Membranaufbau, H. salinarum
Abstract The extremly halophilic archaea is a microorganismus with a feature of long-term microbial survival. They need to survive a minimum of 9% of salt concentration and with a maximum of 32% it is able to grow smoothly. You can find them mainly in e.g. the Dead Sea, natural brins, alkaline salt lakes and marine solar salterns where we have high temperature, low water activity, ultraviolet-light and less oxygen. With the strong structure of their membrane and the specific way of respiration they are able to survive this extreme environment. Most of them has not been researched deeply. The results of my investigation is based mainly on Halobacteria salinarum.
Keywords: extremly halophilic archaeon, Dead Sea, hypersaline soil, 9% salts, Halobacteria salinarum

- 3-
Inhaltsverzeichnis
Zusammenfassung ....................................................................................................... 2
Abstract ......................................................................................................................... 2
Inhaltsverzeichnis......................................................................................................... 3
Abbildungsverzeichnis................................................................................................. 5
Abkürzungsverzeichnis................................................................................................ 7
1 Einleitung ........................................................................................................... 8
2 Die dritte Domäne des Lebens ....................................................................... 11
2.1 Arten .................................................................................................................. 11
2.2 Vorkommen........................................................................................................ 12
3 Extrem Halophile Archaea .............................................................................. 15
4 Beispielorganismus Halobakterium salinarum............................................. 17
5 Wie können die Archaeen in Salzgewässern leben? ................................... 19 5.1 Membranaufbau................................................................................................. 19
5.1.1 Die Purpurmembran .......................................................................................... 20 5.2 Die Energiegewinnung....................................................................................... 21
5.2.1 Fermentation (Gärung) ...................................................................................... 22 5.2.3 Lichtabhängige Ionen- Pumpen......................................................................... 22 5.2.3.1 Aufbau von Bakteriorhodopsin (BR) ................................................................. 22 5.2.3.2 Der Chromophor- dasRetinal ............................................................................ 23 5.2.3.3 Funktion von BR ............................................................................................... 23 5.2.3.4 Photozyklus von Bakterirhodopsin (BR) ........................................................... 24 5.2.3.5 Halorhodopsin (HR) .......................................................................................... 25 5.2.3.6 Photozyklus von HR ......................................................................................... 26
5.2.4 Na+/H+ Antiporter (Langzeitenergiespeicher) .................................................. 28
6 Motilität ............................................................................................................. 29
7 Signaltranduktion ............................................................................................ 31 7.1 Photoensorische Pigmente: Sensory Rhodopsine SRI und SRII ...................... 31 7.1 Rezeptoren ................................................................................................. 32
9 UV-Schutz ......................................................................................................... 34 10 Die industrielle Bedeutung von halophilen Archaeen-
Bakteriorhodopsin als photochromes Sicherheitspigment ........................ 35

- 4-
Literaturverzeichnis.................................................................................................... 38
Erklärung ..................................................................................................................... 40
Stichwortverzeichnis .................................................................................................. 41

- 5-
Abbildungsverzeichnis Abbildung 1: Phylogenetischer Stammbaum nach Woese 1990. Der anhand von rRNA Sequenzen erstellte Stammbaum zeigt die Verwandtschaftsgrade der drei Domänen des Lebens Bakterien, Archaea und Eukaryoten zueinander 9 Abbildung 2: Haloferax volcanii zeigt hier Wachstum auf einer Agarplatte und in einer Kultur. Typische Rotfärbung der Halobakterien durch Carotinoide ist hier deutlich zu erkennen (http://www.nottingham.ac.uk/genetics/people/allers/research.php). 12
Abbildung 3: Lake Magadi in Kenia eine der größten Sodaquellen der Welt. (www.lazypics.de/.../thumbnails/lake_magadi.jpg) 14
Abbildung 4: Der hohe Salzgehalt im Toten Meer verhindert das Untergehen (http://www.kultour.ch/media/images/TOTES_MEER_Zeitung.jpg). 14
Abbildung 5: Saline (Salzgewinnungsstätten), San Francisco Bay (www.uni-giessen.de/.../extrem_halophile.html). 15
Abbildung 6: Vorkommen der Archaea (www.unigiessen.de/..extrem _haloph ile.htm). 16
Abbildung 7: Extrem Halophile: Die Farben dieser Teiche, aus denen Meerwasser verdunstet, werden durch das dichte Wachstum von extrem Halopohilen hervorgerufen. Das Wasser muss einen Salzgehalt von 15-20 % erreichen. Die Teiche sind Teil einer Salzgewinnungsanlage; die halophilen Archaea sind harmlos (Neil A. Champell/ Jane B. Reece). 16
Abbildung 8: Zeigt das stäbchenförmige Halobakterium salinarum. (www.visindavefur.hi.is/myndir/halobakteria_09...) 17
Abbildung 9: Halobakterium salinarum: vom Ort des Vorkommens� Phänotyp von H.salinarum� Membranaufbau� eingelagertes Bakteriorhodopsin mit den sieben Helicen (www.bph.ruhr-uni-bochum.de/bilder/br_abb01.jpg) 18
Abbildung 10: Lipide der Purpurmembran (Besir, Hüseyin (2001):“Untersuchung der lipidvermittelten Kristallisation der Ionenpumpen Bakteriorhodopsin und Halorhodopsin aus Halobakterium salinarum“). 20
Abbildung 11: Schematischer Aufbau des Bakteriorhodopsins. All-trans- Retinal ist über SB (Schiff`sche Base) an das Protein gebunden (http://www.biophysik.uni-freiburg.de/projekte/diplom_br.html) 22
Abbildung 12: Retinalchromophor beim Übergang von all- trans zum 13-cis Retinal. (Doktorarbeit Jamila Guijarro (2006): „Untersuchung des

- 6-
lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“) 23
Abbildung 13: Die sechs wichtigsten Schritte des Photozyklus von BR mit den dazu gehörigen Intermediaten (J, K, L, M1, M2, N, O) und ihren Apsorptionsmaxima: Isomerisierung; T: Ionen-Transport; S: Zugänglichkeitsänderung/ Switch (Jamila Guijarro Doktorarbeit 2001) 24
Abbildung 14: Schematischer Photozyklus von HR (Jamila Guijarro Seite 26) (2006), „Untersuchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“). 26
Abbildung 15: Schematische Darstellung der zwei Photorezeptoren BR und HR. Das Retinal ist als Schiff` sche Base an die Proteine gebunden und liegt in der Mitte des Protonen- bzw. Chloridionenkanals (Kates, 1993) (Jamila Guijarro S.18 (2006), „Unter -suchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobak- terium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“) 27
Abbildung16: Schematische Modelldarstellung des Schaltzyklus im Flagellarmotor (Oesterhelt, Dieter; Marvan Wolfgang (01/05): „Salz in der Suppe des Lebens“ BiospektrumS.60 29
Abbildung 17: Bioenergetische Systeme und die Aktivierung des Flagellarmotor (Oesterhelt, Dieter; Marvan Wolfgang (01/05): „Salz in der Suppe des Lebens“ Biospektrum S. 60-62). 30
Abbildung 18: Schematische Darstellung der Retinalproteine aus H. salinarum und ihre Funktionen (Jamila Guijarro S.12 (2006), „Untersuchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“). 32
Abbildung 19: Schematische Beschreibung der Signaltransduktion und der bioenergetischen System und daran beteiligten Ionenbewegung (Martina Ufer, 1994 "Untersuchung über die Aktivität der Nitratreduktase in dem Archaeabakterium Haloferax volcanii") 33
Abbildung 20: Strukturen von den wichtigsten Bacterioruberinen und Carotinoiden (Susanne Bickel-Sandkötter, Martina Ufer, Kerstin Steinert, Michaela Dana: “Leben im Salz- Halophile Archaea“). 35 Abbildung 21: Die verschiedenen Farbwechsel von BR-Druckfarben: Auf der linken Seite die unbelichtet Farbe von BR und rechst der Farbwechsel nach Lichtinduzierung (Prof. Dr. Norbert Hampp: „Biologische Sicherheitspigmente und optischer Datenspeicher“ Philipps Universität Marburg) 37

- 7-
Abbildung 22: Schematische Darstellung des Bakteriorhodopsins mit den sieben Alpha-Helices und den C und N terminalen Sequenzen, die außerhalb der Membran liegen (http://www.techportal.de/uploads/publications/504/InfoPhysTech40.pdf) 38
Tabellenverzeichnis Tabelle 1: Vergleich der drei Domänen des Lebens (Neil A. Campbell
S.639 6. Auflage 2003) ..................................................................................... 13

- 8-
Abkürzungsverzeichnis AS= Aminosäure
Asp= Aspartat
BR= Bakteriorhodopsin
DMS= Dimethlysulfid
DMSO= Dimethylsulfoxid
e.g= for example
Glu= Glutamat
HCl= Salzsäure
HR= Halorhodopsin
H4= Halocin
kDa= Kilodalton
NaCl= Natriumchlorid
PMK= Protonenmotorische Kraft
PMF= proton motive force= Protonengradient
PZ= Photozyklus
SB= Schiff`sche Base
SR I /II= Sensorische Rhodopsine
TMAO= Trimethylamin N-Oxid
TMA= Trimethylamin
ZKS= Zwei-Komponenten-System
z.B.= Zum Beispiel

- 9-
1 Einleitung
Als das Leben vor mehr als vier Mrd. Jahren begann, bestand es zunächst aus einfachen organisierten Zellen. Es dauerte dann noch 2,5 Mrd. Jahre, bis Organismen entstanden, die einen Zellkern besaßen und noch weitere 600 Mio. Jahre, bis mehrzellige Lebewesen zunehmend auftraten.
Die heute auf der Erde lebenden Spezies werden in drei Domänen unterteilt: Bakterien, Archaeen und Eukaryoten. Bei den Eukaryoten kommen sowohl mit dem Auge sichtbare (makroskopische) Tiere und Pflanzen vor, so wie auch winzige Einzeller, die mit dem Auge nicht sichtbar sind. Die Archaea und auch die Bakterien lassen sich hingegen nur unter dem Mikroskop beobachten. Abgesehen von einigen filamentösen Spezies wie Streptomyceten und Cyanobakterien, welche eine primitive Form der Differenzierung zeigen, sind alle Archaeen und Bakterien Einzeller. Es wird geschätzt, dass nur eine geringe Anzahl (ca. 1%) der bakteriellen und archealen Vertreter bekannt sind und bis jetzt ist es auch nur gelungen ca. 5000 Arten zu kultivieren. Anhand von rRNA Sequenzen wurde ein phylogenetischer Stammbaum (Abb.1) erstellt. In diesem Baum wird der Verwandtschaftsgrad der archealen ribosomalen RNA zu der eukaryotischen rRNA aufgeführt.
Abb.1 : Phylogenetischer Stammbaum nach Woese 1990. Der anhand von rRNA Sequenzen erstellte Stammbaum zeigt die Verwandtschaftsgrade der drei Domänen
des Lebens Bakterien, Archaea und Eukaryoten zueinander
Heute ist bekannt, dass die Archaea Homologien zu den Eukaryoten aufweisen. Die Promotorstruktur, der Transkriptionsapparat und Teile der Proteinbiosynthese ähneln sich stark. Die Ähnlichkeit zu den Bakterien beruht auf der Morphologie: kernlos und

- 10-
einzellig. In ihrer Größe unterscheiden sie sich allerdings. Die Archaea sind kleiner als die Bakterien.
Die Halobakterien erreichen in ihren Lebensräumen eine hohe Populationsdichte, da sie kaum Konkurrenten besitzen. Aus Versuchen unter natürlichen Bedingungen sind Verdoppelungen der Population zwischen 3 und 23 Tagen gemessen worden. Sie vermehren sich durch Zweiteilung und bilden keine Dauerstadien oder Sporen aus. Unter Laborbedingungen ist es möglich, eine Populationsverdopplung in wenigen Stunden zu erhalten.
Die Halobakterien produzieren Peptid-Antibiotika, die Halocine, die das Wachstum anderer extrem halophiler Archaea inhibieren. Es gibt insgesamt 15 verschiedene Halocingruppen. Halocin H4 ist das bis jetzt am weitesten erforschte und bekannteste Peptid- Antibiotika. Es wird von Haloferax mediterranei produziert, es ist thermolabil und salzabhänging. Die Membran der Zielzelle wird durch das Halocin H4 verändert, was zur Folge eine Permeabilitätsänderung hat und schließlich dazu führt, dass sich die Zellen auflösen. Alle unsere Ergebnisse der Halocine stammen aus Versuchen aus dem Labor (Susanne Bickel-Sandkötter, Martina Ufer, Kerstin Steinert und Michaela Dane (1994) „ Leben Im Salz- Halophile Archaea“)
Eine biochemische Besonderheit der Archaea ist, dass sie keine Fettsäuresynthese haben, aber sie bilden Lipide, in denen isoprenoide Seitenketten als Ether mit dem Glycerinteil verknüpft sind. Dieses Merkmal ist nur auf die Archaea bezogen. Ihre Plasmamembran besteht bei allen bisher bekannten aus Di- und Tetraetherlipiden und nicht aus Esterlipiden. Ihre Membran wird unter anderem ausgezeichnet durch einen großen Anteil an sauren Aminosäuren (Aspartat und Glutamat). Sie enthalten einen geringen Anteil an hydophoben Aminosäuren, auf Grund des hohen Ionenanteils im Cytoplasma. Die Membran wird stabilisiert durch Natrium-Ionen. Die eingelagerten Lycopene (C-40 Carotinoide) und Bakterioruberine (C-50 Carotinoide) verleihen der Membran eine charakteristische Rotfärbung und schützen die Halobakterien vor UV-Schäden. Alle Archaea sind gramnegativ (komplexe Struktur der Membran).
Charakterisiert werden die halophilen Archaea als unbeweglich oder beweglich durch Geißelbüschel. Die Wenigen von ihnen die auch beweglich sind, besitzen einen Flagellenmotor, der durch die photosensorischen Pigmente (SRI und SRII) gesteuert wird.
Erwähnt werden sollte auch, dass die Halobakterien kleine Gasvesikel besitzen. Die Vesikel sind wasserundurchlässig, mit Gas gefüllte intrazelluläre Hohlkörper, die aus Proteinen aufgebaut sind. Durch starke lichtbrechende Eigenschaften, sind die Vesikle unter dem Phasenkontrastmikroskop gut sichtbar. Die Gasvesikel verschaffen den Zellen Auftrieb, um in Wasserschichten mit höheren Sauerstoffkonzentrationen zu kommen (Walsby 1994). Ihre lichtbrechenden Eigenschaften könnten jedoch auch als Schutz vor photooxidativen Schäden dienen. Zu den Spezialisten der Gasvesikel- bildung gehören Halobakterium salinarum, Haloferax mediterranei und Halorubrum vacuolata, aber auch andere halophile Archaea und auch Bakterien sind in der Lage Gasvesikel zu bilden.

- 11-
2 Archaea die dritte Domäne des Lebens
Die Archaeen werden in die drei Abteilungen der Crenarchaeota, Korarchaeota und der
Euryarchaeota unterteilt. Die halophilen Archaeen gehören zu der Abteilung der Euryarchaeota.
Domäne: Archaea
Stamm: Euryarchaeota
Klasse: Halobakteria
Ordnung: Halobakteriales
2.1 Arten Zu den halophilen Archaeen gehören 17 Gattungen:
Halobakterium
Haloalcalophilium
Halorubrum
Halobaculum
Haloferax
Haloarcula
Halococcus
Halogeometricum
Halorhabdus
Halosimplex
Haloterrigenea
Natronobacterium
Natronorubrum
Natrinema
Natrialba
Natronomonas
Natronococcus

- 12-
Bei den letzten sechs handelt es sich um Gattungen, die durch Alkaliphilie gekennzeichnet sind. Natronobacterium wächst bei einer niedrigen Magnesium- konzentrationen und einem pH-Wert um 9-11.
Abb. 2: Haloferax volcanii zeigt hier Wachsetum auf einer Agarplatte und in einer Kultur. Typische Rotfärbung der Halobakterien durch die Carotinoide ist hier deutlich zu
erkennen (http://www.nottingham.ac.uk/genetics/people/allers/research.php).
2.2 Vorkommen Archaea sind einzellige Mikroorganismen, die zu der einfachsten Form des Lebens zählen. Ihr häufigstes Vorkommen ist in extremen Habitaten nachgewiesen worden, wie zum Beispiel bei extrem hohen Salzkonzentrationen, Temperaturen um den Siedepunkt, Druck, aber auch bei extrem hohen und niedrigen pH-Werten. Diese Bedingungen wären für andere Organismen tödlich, aber auch unter normalen und gemäßigten Lebensbedingungen sind sie zu finden. Die extremen Habitate erinnerten Carl Woese an frühere Bedingungen auf der Erde. Er gab den Archaea Ihren Namen, den er mit dem griechischen Wort archaios für ursprünglich verband.
- 13-
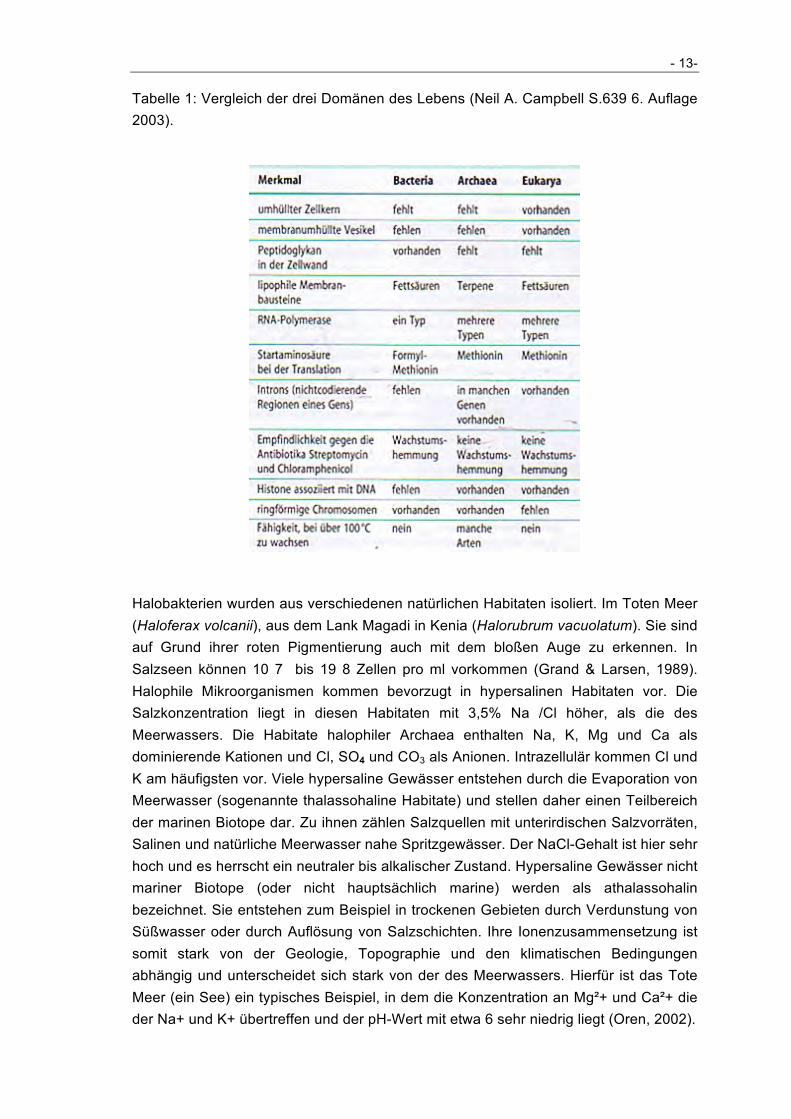
Tabelle 1: Vergleich der drei Domänen des Lebens (Neil A. Campbell S.639 6. Auflage 2003).
Halobakterien wurden aus verschiedenen natürlichen Habitaten isoliert. Im Toten Meer (Haloferax volcanii), aus dem Lank Magadi in Kenia (Halorubrum vacuolatum). Sie sind auf Grund ihrer roten Pigmentierung auch mit dem bloßen Auge zu erkennen. In Salzseen können 10 7 bis 19 8 Zellen pro ml vorkommen (Grand & Larsen, 1989). Halophile Mikroorganismen kommen bevorzugt in hypersalinen Habitaten vor. Die Salzkonzentration liegt in diesen Habitaten mit 3,5% Na /Cl höher, als die des Meerwassers. Die Habitate halophiler Archaea enthalten Na, K, Mg und Ca als dominierende Kationen und Cl, SO4 und CO3 als Anionen. Intrazellulär kommen Cl und K am häufigsten vor. Viele hypersaline Gewässer entstehen durch die Evaporation von Meerwasser (sogenannte thalassohaline Habitate) und stellen daher einen Teilbereich der marinen Biotope dar. Zu ihnen zählen Salzquellen mit unterirdischen Salzvorräten, Salinen und natürliche Meerwasser nahe Spritzgewässer. Der NaCl-Gehalt ist hier sehr hoch und es herrscht ein neutraler bis alkalischer Zustand. Hypersaline Gewässer nicht mariner Biotope (oder nicht hauptsächlich marine) werden als athalassohalin bezeichnet. Sie entstehen zum Beispiel in trockenen Gebieten durch Verdunstung von Süßwasser oder durch Auflösung von Salzschichten. Ihre Ionenzusammensetzung ist somit stark von der Geologie, Topographie und den klimatischen Bedingungen abhängig und unterscheidet sich stark von der des Meerwassers. Hierfür ist das Tote Meer (ein See) ein typisches Beispiel, in dem die Konzentration an Mg²+ und Ca²+ die der Na+ und K+ übertreffen und der pH-Wert mit etwa 6 sehr niedrig liegt (Oren, 2002).

- 14-
Abb. 3: Lake Magadi in Kenia eine der größten Sodaquellen der Welt (www.lazypics.de/.../thumbnails/lake_magadi.jpg)
Abb. 4: Der hohe Salzgehalt im Toten Meer verhindert das Untergehen (http://www.kultour.ch/media/images/TOTES_MEER_Zeitung.jpg).

- 15-
3 Extrem halophile Archaea
Ihren Namen haben die halophilen Archaea von ihrem extremen Lebensort (vom griechischen: halo für „Salz“ und philos für „Freund“), direkt übersetzt Salzliebe. Diese Organismen lieben das Salz, das heißt sie sind nicht nur an höhere Salzkonzentrationen angepasst, sondern sie benötigen das Salz zum Leben. Diese Organismen brauchen zum Leben eine minimale Salzkonzentration von 1,5 M NaCl (9%). Für ein optimales Wachstum (zum Beispiel unter Laborbedingungen) liegt eine Salzkonzentration von 3,5 und 4,5 M (21-27%) im Außenmedium vor (Grant und Larsen, 1989). Auch bei einer höheren Salzkonzentration (5,2M 32% =Sättigungs- konzentration) ist noch Wachstum möglich, dieser findet dann aber in verlangsamter Form statt. Es lässt sich über die halophilen Archaeen sagen, dass sie zur dominierenden Population werden, sobald die Salzkonzentration eine Molarität von mindestens 2,5 erreicht (Norten 1992, Grant und Larsen 1989). Die Halobakterien produzieren Halocin, die das Wachstum halophiler Konkurrenten hemmen.
Anhand dieser Organismen lässt sich untersuchen wie Leben unter extremen Bedingungen möglich ist.
Abb. 5: Saline (Salzgewinnungsstätte), San Francisco Bay (www.uni-giessen.de/.../extrem_halophile.html)

- 16-
Abb. 6: Vorkommen der Archaea (www.uni-giessen.de/.../extrem_halophile.html)
Abb. 7: Extrem Halophile: Die Farben dieser Teiche, aus denen Meerwasser verdunstet, werden durch das dichte Wachstum von extrem Halopohilen
hervorgerufen. Das Wasser muss einen Salzgehalt von 15-20 % erreichen. Die Teiche sind Teil einer Salzgewinnungsanlage (Neil A. Champell/ Jane B. Reece).

- 17-
4 Beispielorganismus Halobakterium salinarum
Abb. 8: Zeigt das stäbchenförmige Halobakterium salinarum (www.visindavefur.hi.is/myndir/halobacteria_09...)
Halaobakterium salinarum gehört zu der Familie der Halobacteria. H. salinarum ist ein stäbchenförmiger Prokaryont, der vier Flagellen besitzt, etwa 5 Mikrometer lang ist und dessen Zellen einen Durchmesser von einem halben Mikrometer aufweisen. Geringe Sauerstoffkonzentrationen und hohe Lichtintensität prägen das Habitat des Halobakterium salinarum.
Der Organismus ist stets einem osmotischen Stress ausgesetzt, da er sein optimales Wachstum bei einer außergewöhnlich hohen Natriumchlorid- Konzentration (4M) besitzt. Fällt die NaCl- Konzentration auf 3M ab, so tritt die osmotische Lyse der Zelle ein (Stoeckenius, 1976). H. salinarum ist im Toten Meer zu finden, aber auch in Salinen und in Kalifornien in dem Salzsee Orwenslake.
Die meisten Halobakterien sind aerob, chemoorganothrophe Mikroorganismen, die verschiedene Aminosäuren für ihr Wachstum benötigen. Fehlt Sauerstoff, so kann H. salinarum unter anaeroben Bedingungen Arginin zu Ornithin, Ammoniak und Kohlendioxid abbauen (Oren, 1994). In die Purpurmembran des Halobakterium salinarum ist das Bakteriorhodopsin eingelagert, das als einfaches System der Photosynthese fungiert, indem es mittels absorbierter Lichtenergie über der Membran einen Protonenkonzentrationsgradienten erzeugt (Beschreibung siehe unter 5.2.2.2).

- 18-
Abb. 9: Halobakterium salinarum zu sehen: vom Ort des Vorkommens → Phänotyp von H. salinarum → der Membranaufbau von oben → eingelagertes Bakteriorhodopsin
mit den sieben Helicen→ Bakteriorhodopsin mit dem eingelagerten Retinal (gelb makiert) (www.bph.ruhr-uni-bochum.de/bilder/br_abb01.jpg).

- 19-
5 Wie können die Archaea bei diesen extremen Bedingungen überleben?
Bei hohen Salzkonzentrationen werden normalerweise Proteine und Makromoleküle zerstört. Die Struktur der Proteine zerfällt aufgrund von hydrophoben Wechsel- wirkungen oder die Proteine aggregieren, weil ihnen die Hydrathülle entzogen wird. Außerdem werden die essentiellen elektrostatischen Wechselwirkungen innerhalb oder zwischen den Makromolekülen durch die Ladungen gestört. Die Verfügbarkeit von Wasser wird durch die Hydratation der Salzionen reduziert (Dennis& Shimmin, 1997). Die biologischen Membranen sind wasserdurchlässig, dieses würde für die halophilen Archaeen bedeuten, dass sie Wasser an ihre Umgebung abgeben müssten. Man spricht auch davon, dass hypersaline Biotope „physiologisch trocken“ genannt werden, d.h. die Umgebung entzieht den Zellen das Wasser aufgrund des hohen osmotischen Drucks. Damit sie in der Lage sind unter diesen hohen Salzkonzentrationen zu leben, mussten sie unterschiedliche Strategien entwickeln. Bei Halobakterien wurde entdeckt, dass sie durch Osmose die hohen Salzkonzentrationen der Umgebung mit den inneren Kaliumchloridkonzentrationen ausgleichen (Kuschner und Kamekura 1988; Andreas Tebbe 2005). Unter anderem spielt der Aufbau der Zellwand eine sehr große Rolle.
5.1 Membranaufbau Die Zellwand der halophilen Archaeen besteht nicht aus Peptidoglykan wie bei den Bakterien sondern aus Proteinen und Glykoproteinen, dem sogenannten S-Layer. Mit überwiegend sauren Aminosäuren, wie Glutamat, Aspartat und nur einem geringen Anteil an hydrophoben Aminosäureresten. Es wird vermutet, dass bei einem physiologischen pH- Wert die sauren Aminosäurereste aufgrund der negativ geladenen funktionellen Gruppen stärker hydratisiert werden, als die anderen Aminosäurereste. Auf diese Weise würde sich das hydrierte Salzionen-Netzwerk auf der Proteinoberfläche besser koordinieren lassen (Dennis& Shimmin, 1997). Die Hydrophobizität wird durch die negativ geladenen Seitenketten verringert und verhindert so die Aggregation (Aussalzen) der Proteine. Eine Stabilisation entsteht durch die Gegenständigkeit, der negativ geladenen Säuregruppen der Aminosäuren und der positiven Natriumionen des Außenmediums. Sind im Außenmedium keine Na+ vorhanden drängen sich die negativ geladenen Säuregruppen der Aminosäuren auseinander, das Protein entfaltet sich und somit lysieren die Zellen. Es ist also eine hohe Salzkonzentration nötig, damit die Zellwände bestehen bleiben.
- 20-
5.1.1 Die Purpurmembran (PM)
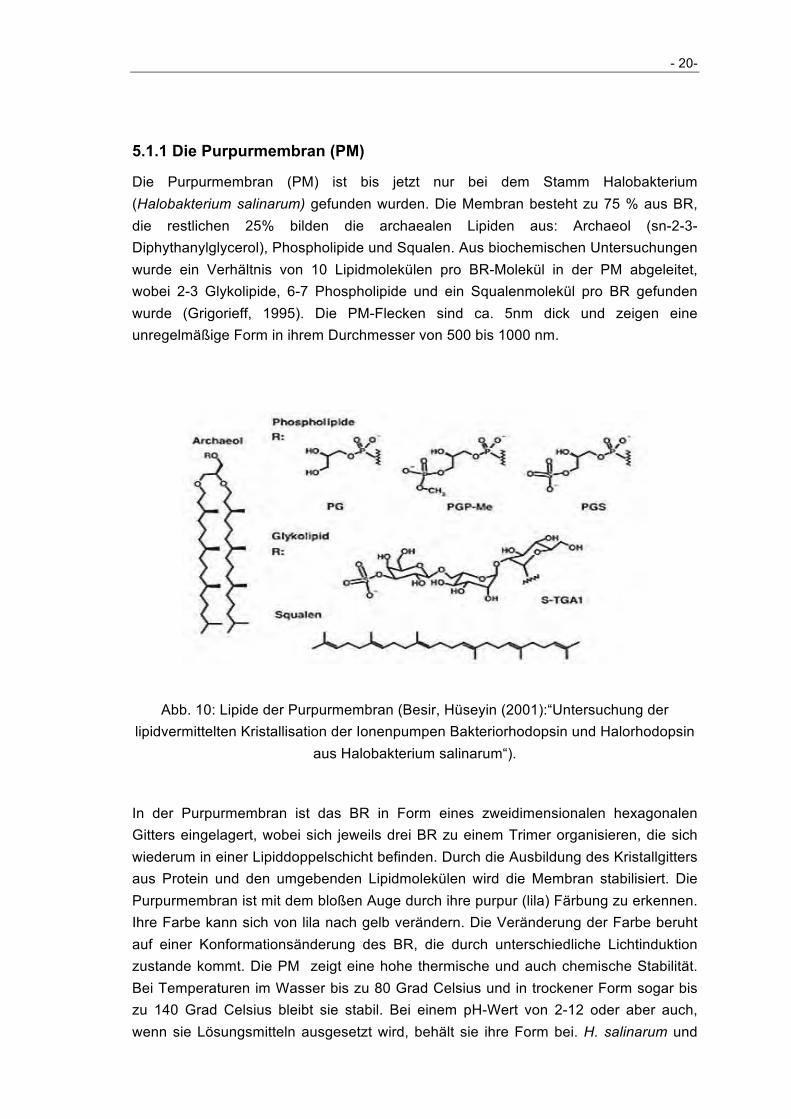
Die Purpurmembran (PM) ist bis jetzt nur bei dem Stamm Halobakterium (Halobakterium salinarum) gefunden wurden. Die Membran besteht zu 75 % aus BR, die restlichen 25% bilden die archaealen Lipiden aus: Archaeol (sn-2-3-Diphythanylglycerol), Phospholipide und Squalen. Aus biochemischen Untersuchungen wurde ein Verhältnis von 10 Lipidmolekülen pro BR-Molekül in der PM abgeleitet, wobei 2-3 Glykolipide, 6-7 Phospholipide und ein Squalenmolekül pro BR gefunden wurde (Grigorieff, 1995). Die PM-Flecken sind ca. 5nm dick und zeigen eine unregelmäßige Form in ihrem Durchmesser von 500 bis 1000 nm.
Abb. 10: Lipide der Purpurmembran (Besir, Hüseyin (2001):“Untersuchung der lipidvermittelten Kristallisation der Ionenpumpen Bakteriorhodopsin und Halorhodopsin
aus Halobakterium salinarum“).
In der Purpurmembran ist das BR in Form eines zweidimensionalen hexagonalen Gitters eingelagert, wobei sich jeweils drei BR zu einem Trimer organisieren, die sich wiederum in einer Lipiddoppelschicht befinden. Durch die Ausbildung des Kristallgitters aus Protein und den umgebenden Lipidmolekülen wird die Membran stabilisiert. Die Purpurmembran ist mit dem bloßen Auge durch ihre purpur (lila) Färbung zu erkennen. Ihre Farbe kann sich von lila nach gelb verändern. Die Veränderung der Farbe beruht auf einer Konformationsänderung des BR, die durch unterschiedliche Lichtinduktion zustande kommt. Die PM zeigt eine hohe thermische und auch chemische Stabilität. Bei Temperaturen im Wasser bis zu 80 Grad Celsius und in trockener Form sogar bis zu 140 Grad Celsius bleibt sie stabil. Bei einem pH-Wert von 2-12 oder aber auch, wenn sie Lösungsmitteln ausgesetzt wird, behält sie ihre Form bei. H. salinarum und

- 21-
auch andere extrem halophile Archaeen bilden bei niedrigem Sauerstoffgehalt purpurfarbene (lila) Bereiche in der Membran aus, wobei bei den anderen Archaeen die Färbung auf Carotinoide zurück zu führen ist, die in die Membran eingelagert sind (Doktorarbeit: Hüseyin, Besir 2001).
5.2 Energiegewinnung (Bioenergetik)
Bei den halophilen Archaeen ist der Sauerstoffgehalt bei diesen extremen Lebensbedingungen sehr knapp. Die Löslichkeit von Sauerstoff ist bei hohen Salzkonzentrationen gering. Die Archaeen sind jedoch in der Lage unter anaeroben Bedingungen weiter zu leben. Ihnen stehen weitere Elektronenakzeptoren zur Verfügung, wie zum Beispiel bei den Gattungen Haloferax und Halobakterium wurde entdeckt, dass sie in der Lage waren Fumarat als Elekronenakzeptor zu nutzen und zu Succinat zu reduzieren (Oren, 1991).
Außerdem kommt die Reduktion von Nitrat zu Nitrit über eine der Membran außen aufsitzenden Nitratreduktase vor. Die Nitrat- reduktase wurde bei Haloferax mediterranei und Haloferax volcanii entdeckt (Ufer und Bickel-Sandkötter, 1995).
Als weitere Elekronenakzeptoren stehen den Halobakterien Dimethylsulfoxid und Trimethylamin N-Oxid zu Verfügung. Bei den meisten untersuchten Stämmen wurde festgestellt, dass sie Dimethylsulfoxid (DMS) zu DMSO reduzieren oder Trimethylamin (TMA) aus Trimethylamin N-Oxid (TMAO) produzieren (Oren und Trüper, 1990). Die Elekronenakzeptoren übernehmen in diesem Fall die Rolle des Sauerstoffs. Auch die Fermentation von Arginin stellt eine Methode der Energie- gewinnung dar, die bei H. salinarum gefunden wurde.
Eine andere Möglichkeit der Energiegewinnung besitzt das Halobakterium H. salinarum. H.salinarum kann mit Hilfe von Sonnenlicht Energie gewinnen. Es lässt sich nicht mit der Photosynthese vergleichen, da hier keine CO2 Fixierung stattfindet und auch keine Kohlenhydrate erzeugt werden. Hier spielt nicht Chlorophyll eine wichtige Rolle, sondern Bakteriorhodopsin. H. salinarum erzeugt bei Abwesenheit von Sauer- stoff, mit Hilfe der lichtgetriebenen Protonenpumpe Bakteriorhodopsin eine proton- motorische Kraft, die die ATP- Synthase so effizient antreibt, dass die Zellen phototroph wachsen können. Es wird nicht Wasser gespalten, sondern es wird ein Konzentrations- unterschied zwischen innerem und äußerem Cytoplasma aufgebaut.
5.2.1 Fermentation (Gärung)
Unter Fermentation versteht man die Umsetzung von biologischem Materials mittels Bakterien zu Energiegewinnung bezeichnet, die bei Abwesenheit von Sauerstoff abläuft. Steht der Zelle kein Sauerstoff, kein Licht oder keine Kaliumbatterie (s. u Langzeitenergiespeicher) zur Verfügung, so kann H. salinarum Arginin zu Ornithin abbauen und das dabei entstehende Carbamylphosphat dazu nutzen um ATP umzu-

- 22-
wandeln (Oesterhelt, Dieter; Marvan Wolfgang (01/05): „Salz in der Suppe des Lebens“ Biospektrum S. 60-62)
Bei fermentativem Wachstum erzeugt die archeale ATPase durch ATP-Hydrolyse die notwendige ionenmotorische Kraft für Ionenaustausch, Transportprozesse und Bewegung.
5.2.2 Lichtabhängige Protonen-Pumpen
5.2.2.1 Aufbau von Bakteriorhodopsin (BR)
Eine besondere Art der Energiegewinnung ist durch das in Halobakterium salinarum entdeckte Bakteriorhodopsin möglich. Bakteriorhodopsin gehört zu den integralen Membranproteinen und ist in die Purpurmembran eingelagert.
Die Primärsequenz besteht aus 248 Aminosäuren (Stöckenius, 1971) und besitzt ein Molekulargewicht von 26784 Dalton. Die Sekundärstruktur des BR ist aus sieben annähernd parallelen Alpha-Helices aufgebaut, die sich ringförmig anordnen und senkrecht in der Membran stehen. Ein Ionenkanal wird von vier dieser Helices (B, C, F und G) gebildet. Durch diesen Ionenkanal findet der Protonentransport in der Zellmembran statt. Im Grundzustand des Proteins ist dieser cytoplasmatische Kanal geschlossen und somit kann kein Ionentransport statt finden. BR kann über hundert Protonen in der Sekunde durch die Membran transportieren. Das Bakteriorhodopsin zeigt eine große Ähnlichkeit mit dem menschlichen Sehpigment Rhodopsin. Das lichtabsorbierende Element im Bakteriorhodopsin (BR) ist das Retinal (hydrophobes Molekül).
Abb.11: Schematischer Aufbau des Bakteriorhodopsins. All- trans- Retinal ist über SB (Schiff`sche Base C═N) an das Protein gebunden (http://www.biophysik.uni-
freiburg.de/projekte/diplom_br.html).

- 23-
5.2.2.2 Der Chromophor- das Retinal
Das Retinal ist ein Isoprenoid und ein Aldehyd von Vitaminen A. Absorbiert der Chromophor Licht, wird es von der all- trans Form in die 13 cis Form überführt. Das Retinal ist zwischen cytoplasmatischer und extrazelluärer Halbpore als protonierte Schiff`sche Base (SB) an das Lysin K216 (in der G Helix) gebunden (Oesterhelt, 1998). Das Protein hat ein Molekulargewicht von 260 Dalton. Das Retinal lagert sich unter anaeroben Bedingungen dicht und regelmäßig in der Cytoplasmamembran (Purpurmembran) ein und sorgt bei den Archaeen für die sichtbare Rotfärbung. Anhand von zahlreichen biochemischen und spektroskopischen Untersuchungen mit BR-Mutanten wurde nachgewiesen, dass nur die SB unerlässlich für die Funktionsfähigkeit der Protonenpumpe ist. Alle anderen Stationen spielen keine ausschlaggebende Rolle (Lanyi 1997, Subramaniam 1999). Das Retinalmolekül kann durch Licht aktiviert werden und dadurch kommt es zu einer Konformationsänderung. Es wird reversibel von all trans zu 13-cis Retinal isomerisiert.
Abb. 11: Retinalchromophor beim Übergang von all- trans zum 13-cis Retinal. (Doktorarbeit Jarmila Guijarro (2006): „Untersuchung des lichtgetriebenen
Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“)
5.2.2.3 Funktion von Bakteriorhodopsin (BR)
Durch das Pumpen der Protonen aus dem Cytoplasma durch die Pore auf die extrazelluläre Seite entsteht ein Protonengradient (PMF= proton motive force). Die lichtgetriebene Protonenpumpe, das Bakteriorhodopsin, treibt die Bildung dieses Protonengradienten über die Zellmembran unter anaeroben Bedingungen an. Der entstandene Protonengradient wird dafür genutzt, dass ATP durch das Enzym ATPase synthetisiert werden kann. Bakteriorhodopsin schützt unter anderem die Zellen vor

- 24-
photooxidativen Schäden, die hervorgerufen werden durch die starke Sonnen- einstrahlung.
5.2.2.4 Photozyklus von Bakteriorhodopsin (BR)
Der Photozyklus kann in sechs Schritten beschrieben werden (Abb.12). Wenn Sonnenlicht auf das purpurfarbene Bakteriorhodopsin (BR 568) trifft, wird damit eine lichtgetriebene Ionenpumpe in Gang gesetzt. Es kommt zu einer Konformations- änderung des Retinalchromophors. Im unbelichteten Zustand liegt das Retinal in der trans-Form vor. Nach der Isomerierung (nach Belichtung) geht es in die 13-cis-Form über und startet den Photozyklus (Stoechenius, 1979). Die Farbe des Bakterio-rhodopsin ändert sich mit der Konformationsänderung. Durch mehrere Zwischenstufen (K, L) verändert sie sich von purpur nach gelb (M1).
Abb.12: Die sechs wichtigsten Schritte des Photozyklus von BR mit den dazu gehörigen Intermediaten (J, K, L, M, N, O) und ihren Absorptionsmaxima. Im Inneren
des Zyklus sind die zwei unterschiedlichen Konformationszustände von Retinal zu sehen (Susanne Bickel-Sandkötter, Wolfgang Gärtner, Michaela Dane: Springer
Verlag1996 „Conversion of energy in halobacteria: ATP synthesis and phototaxis“).
Das Lysin, mit dessen Hilfe die Schiff`sche Base gebildet wird, ist die zentrale Proteinbindestelle des Proteins. Im Grundzustand liegt sie protoniert vor. Beim Übergang von L(550) zum ersten langlebigen M- Intermediat wird die Schiff`sche Base deprotoniert. Gleichzeitig wechselt das Gegenion der Schiff`schen Base zum Asp 85 (Protonenakzeptor) in die protonierte Form. Außerdem wird ein Proton (H+) von der Region nahe Glutamat 194 und Glutamat 204 durch die Zellmembran transportiert und

- 25-
nach außen abgegeben. Hier liegt ein blauverschobenes Absorptionsmaximum von 410 nm vor. Bei der Reaktion von M nach N wird die deprotonierte Schiff`sche Base von Aspartat 96 reprotoniert und ein Proton wird aus der cytoplasmatischen Oberfläche wieder aufgenommen. An der letzten Station von O nach BR findet wieder eine Deprotonierung des Asp 85 statt und das Retinal kehrt durch thermale Reisomerisierung in die all-trans Form zurück. Das BR besitzt an dieser Stelle schließlich wieder seinen purpurfarbenen Ausgangszustand.
Als Photozyklus werden die reversiblen Photoreaktionen bezeichnet, die wieder in ihre Ausgangszustand zurück gehen. Das Bakteriorhodopsin und auch das Halorhodopsin durchlaufen den Photozyklus in etwas mehr als 10ms.
5.2.2.5 Halorhodopsin (HR)
Ein weiteres Retinalprotein ist das Halorhodopsin, welches nicht wie das BR bis jetzt nur bei einem Organismus gefunden wurde, sondern weiter verbreitet ist. Halorhodopsin stellt eine nach innen gerichtete, lichtgetriebene Chloridpumpe da. Die Chloridpumpe ist für Bakterien bei hohen Salzkonzentrationen lebenswichtig. Die Pumpe benutzt genau wie BR Retinal als Lichtempfänger.
Die halophilen Archaea müssen, bevor sie sich teilen können, ihr Volumen vor der Zellteilung vergrößern. Durch Absorption von zwei Photonen kann HR Protonen in die Zelle transportieren. Dazu werden negativ geladene Chlorid-Ionen in das Zellinnere gepumpt und als Gegenionen folgen Kaliumkationen durch Ionenkanäle passiv in das Innere der Zelle (KCl Konzentration innen und NaCl Konzentration außen etwa gleich = Einsalzstrategie).
Mittels Anionenimport lädt HR ebenfalls wie BR die Membran negativ auf, fungiert aber nicht als photosyntetisches Pigment, sondern erhält den osmotischen Druck des cytoplasmatischen und extrazellulären Mediums in der Balance, indem es Licht über einen gerichteten Ionentransport in elektrochemische Energie umwandelt und so die Aufnahme von Salz (K und Cl) erlaubt.
Halorhodopsin liegt unter Laborbedingungen in 3D Kristallen zu Trimeren angeordnet vor und bildet einen Kanal in der Membran. Dieser cytoplasmatische Kanal ist im Grundzustand geschlossen und somit nicht für Ionen-Transport durchlässig. Das Halorhodopsin (HR) hat eine ähnliche Struktur, wie das Bakteriorhodopsin. Jedoch gibt es einen entscheidenden Unterschied im aktiven Zentrum, dort wo das Cl-Anion gebunden wird. Bei BR sitzt an derselben Stelle die gleichfalls negativ geladene Gruppe einer AS, die für die vorübergehende Aufnahme des Protons während des Pumpprozesses zuständig ist. Da das Chlorid-Anion einen unfangreicheren Platz aus Größengründen braucht, fehlt hier die entsprechende Aminosäure. Chlorid wird nur über wenige gerichtete ionische Wechselwirkungen gehalten. Im ionischen Leitweg bindet das Chlorid wohl nur über Aminosäurereste. Eine weitere auffallende Besonderheit ist, dass das Anion von mehreren Wasserstoffatomen umgeben wird. Obwohl C-O-Bindungen nur ein schwaches Dipolmoment besitzen, reicht die Anhäufung von solchen Gruppen um das Chlorid-Ion aus, um dessen Bindung zu

- 26-
stabilisieren (http://www.mpg.de/bilderBerichteDokumente/dokumentation/PresseMitteilungen/2000/p ri49_00.htm).
5.2.2.6 Photozyklus von HR
Der Photozyklus läuft bei Halorhodopsin sehr ähnlich ab. Die Schlüsselaminosäuren Asp85 und Asp 96 in BR sind in HR durch Threonin 111 bzw. Alanin 122 ersetzt. Aus diesem Grund liegt hier die Schiff`sche Base während des gesamten Zyklus protoniert vor (Oesterhelt 1995). Es finden also keine M- Zustände statt.
Abb. 13: Schematischer Photozyklus von HR mit den dazu gehörigen Absorptionsmaxima. I: Isomerisierung; T: Ionen- Transport; S:
Zugänglichkeitsänderung/ Switch (Jarmila Guijarro Seite 26 (2006), „Untersuchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis
mit statistischer und zeitaufgelöster FTIR- Spektroskopie“)
Das erste rotverschobene Intermediat HR600 entspricht dem K590 von BR mit 13-cis-Retinal. Im darauf folgenden Schritt zu HR 520 wird das Chloridion von seiner ursprünglichen Position zur protonierten Schiff´schen Base verschoben (genaue Position noch nicht bekannt). Die Änderung der Lage des Chloridions, muss jedoch dazu führen, dass eine stärkere Lokalisation der positiven Ladungen an der protonierten Schiff´schen Base stattfindet, was durch die geänderte Absorption (von 600 zu 520nm) zu erklären wäre. Anschließend findet der Übergang von HR520 in das kurzlebige HR640 (rot) statt, wobei ein Cl-Ion in das Cytoplasma abgegeben wird und danach eine Absorption von 565 nm (O-Intermediat) gemessen wird. Nach thermischer Isomerisierung zu all- trans- Retinal wird aus der extrazellulären Seite ein Cl-Ion aufgenommen und der Ausgangszustand mit der protonierten Schiff´schen Base und

- 27-
das mit Chlorid besetzte aktive Zentrum wieder erreicht. Der Zyklus von HR ist bis jetzt noch weniger erforscht, als der von BR (Jamila Guijarro (2006), „Untersuchung des lichtgetriebenen Chloridtransports in Halo- rhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“).
Der entschiedene Unterschied der beiden Retinalproteine HR und BR lässt sich durch eine einzige Mutation einer Aminosäure aufheben. Nur der Ersatz einer negativ geladenen AS (von Asp 85) als Protonenakzeptor in BR, durch ein Cl-Anion in HR, führt zu einer kompletten Änderung der Ionenspezifität von Kationen und Anionen. BR kann somit zu einer Chloridpumpe durch eine Aminosäuremutation konvertiert werden.
Abb. 14: Schematische Darstellung der zwei Photorezeptoren 1.BR und 2.HR. Das Retinal ist als Schiff`sche Base an die Proteine gebunden und liegt genau in der Mitte des Protonen- bzw. Chloridkanals (Jarmila, Guijarro S.18 (2006), „Untersuchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“)
5.2.4 Na+/ H+ Antiporter (Langzeitenergiespeicher)
Die Halobakterien können überschüssige Lichtenergie, die nicht direkt zur ATP-Synthese benötigt wird, elektroneutral in großem Umfang speichern (der Protonengradient kann selber nicht gespeichert werden, da die elektrische Kapazität der Zellmembran limitiert ist). Der Protonengradient treibt einen H+/Na+- Antiporter an, der in der Cytoplasmamembran verankert ist. Die zuvor nach außen gepumpten

- 28-
Protonen (H+) fließen zurück in die Zelle. Na+ wird nach außen gepumpt und gleichzeitig fließt K+ durch einen selektiven Kaliumkanal nach innen und neutralisiert so die Zelle. So wird ein Kaliumspeicher molarer Konzentrationen im Inneren aufgebaut. Nach Sonnenuntergang läuft der ganze Prozess umgekehrt ab. Die Batterie wird dann entleert, indem die Zelle den Kaliumdiffusionsgradienten wieder in einen Protonengradienten umbaut, der wie das Bakteriorhodopsin (bei Licht) die ATP- Synthase antreibt (Dieter Oesterhelt, Wolfgang Marwan 2005). So ermöglicht der Kaliumgradient der Zelle eine lichtunabhängige ATP-Synthase und gewährleistet auch im Dunkeln eine Energieversorgung bis 24 h. Dieser elektrochemische Gradient kann von Sekundär-Transportsystemen genutzt werden um wiederum die Aufnahme anderer Stoffe an zutreiben (Norten, 1992).

- 29-
6 Motilität
An H. salinaum wurde das Schwimmverhalten im Laufe der letzten 20 Jahre eingehend untersucht. Die fünf bis zehn Geißeln bilden ein helikales Bündel. Im Gegensatz zu den eukaryotischen Geißeln, die peitschenartig schlagen, werden die Geißeln der Halobakterien durch einen Rotationsmotor bewegt. Der Motor kann sich gegen oder im Uhrzeigersinn drehen. Die Drehrichtung des Bündels bestimmt, in welche Richtung die Zellen schwimmen. Im Urzeigersinn bewegt sich das Halobakterium vorwärts. Der Richtungswechsel (Umkehr) wird durch äußere Reiße wie zum Beispiel ultraviolettes Licht oder aber auch ganz zufällig durch Kollision mit einem Partikel.
Die molekularen Komponenten des archealen Flagellarmotors zu identifizieren hat sich bis heute nicht erfüllt. Die zunächst angenommene Ähnlichkeit mit dem aufgeklärten Geißelmotor der Enterobakterien konnte nicht bestätigt werden. Es wird heute angenommen, dass der zweifellos nach dem rotatorischen Prinzip arbeitende archaeale Flagellarmotor aus völlig anderen Komponenten konstruiert sein muss, als der der Bakterien.
Ernst Dieter Gilles und Dieter Oesterhelt haben zusammen ein Modell der Signalkaskade entworfen. Sie stellten fest, dass ein Teil der sensorischen Informationsverarbeitung im Schaltkomplex des Geißelmotors stattfindet.
Abb.16: Schematische Modelldarstellung des Schaltzyklus im Flagellarmotor (Oesterhelt, Dieter; Marvan Wolfgang (01/05): „Salz in der Suppe des Lebens“
Biospektrum S. 60-62).

- 30-
Hier spielt das Schaltsignal (Che YP), aber auch die konformativen Zustände des Schaltkomplexes selbst eine große Rolle. Mit Hilfe des Signals und der konformativen Zustände ermittelt die Zelle die für sie besten Umweltbedingungen und die dort heraus resultierende Schwimmrichtung. In der Abb. 16 werden die acht Zustände des Schaltkomplexes und ihre Aktivitätszustände dargestellt. Die acht Zustände fassen eine komplexe Folge von Konformationsänderungen der Untereinheiten zusammen. Es können vier unter- schiedliche Aktivitätszustände vorliegen: Stopp, refraktär (ref), kompetent (comp) und aktiv (act). Außerdem kann der Kreislauf gegen den Uhrzeigersinn (ccw) oder im Uhrzeigersinn (cw) ablaufen. Die Übergänge von refraktär nach kompetent und von kompetent nach aktiv werden durch eine Stimulation reguliert, woraus eine Änderung der CheY- Phosphatkonzentration hervorgeht. Eine steigende Konzentration an CheY verlangsamt den Übergang von refraktär nach kompetent und beschleunigt den Übergang von kompetent nach aktiv. Die Schwimmphase wird dadurch aber im Ganzen verkürzt.
Die Aktivierung der Rezeptoren wird reguliert durch die Autophosphorylierungsaktivität der Histidinkinase CheA, die über ein Kopplermolekül CheW an die Rezeptoren gebunden wird. Der Phosphatrest wird auf einen Antwortregulator CheY~P übertragen, der dann am Flagellarmotor den Schaltzyklus moduliert. Parallel zur Änderung der CheY- Phosphatkonzentration wird von aktivierten Rezeptoren die Fumarat- ausschüttung bewirkt, welche ebenfalls direkt am Flagellarmotor wirkt. Bleibt der Reiz konstant, kehrt das CheY- Phosphatsignal auf den Wert vor der Stimulation zurück, was durch die konstante Aktivität der Metyltransferase (CheR) und durch die von der Histidinkinase (CheA) regulierten Esterase (CheB) bewirkt wird.
Abb. 17: Bioenergetische Systeme und die Aktivierung des Flagellarmotor (Oesterhelt, Dieter; Marvan Wolfgang (01/05):„Salz in der Suppe des Lebens“ Biospektrum S.60)

- 31-
7 Signaltransduktion
Die Signaltransduktion wird in halophilen Archaeen durch die Retinalproteine (SRI und SRII) vermittelt. Die Retinalproteine sind in die Membran eingelagert. Sie sind in der Lage, auf unterschiedliche Wellenlängen mit unterschiedlichen Reaktionen zu reagieren. Die Drehrichtung des Geißelmotors ist also abhängig von der Wellenlänge, die auf die Pigmente trifft.
7.1 Photosensorische Pigmente: Sensory Rhodopsin SRI und SRII Die Aufgabe von SRI und SRII besteht nicht darin, Ionen über die Zellmembran zu pumpen, sondern ihre Aufgabe ist es die Intensität und die Wellenlängenbereiche des einstrahlenden Lichtes zu messen.
Im Gegensatz zu den Energietransducern BR und HR agieren die beiden Sensory Rhodopsine (SRI und SRII) als Informationstransducer indem sie drei Antworten zu Lichteinwirkungen weiterleiten. Ein Hinschwimmen zu gelbem Licht wird durch SRI erzeugt, wodurch HtrI(Einleiterprotein) aktiviert wird. Eine Fluchtreaktion findet statt, wenn der Organismus mit UV Licht oder blauem Licht bestrahlt wird. Bei UV-Licht schaltet sich das SRI ein, welches HtrI aktiviert. Bei blauem Licht wird das HtrII durch das SRII aktiviert.
Durch das Einleiterprotein HtrI wird eine Signalkaskade ausgelöst, die die Dreh- richtung des Geißelapparates der Zelle ändert und so ein Wegschwimmen von UV-Licht und ein Hinschwimmen zum orangenen Licht ermöglicht. SRII ist ein Blaulichtrezeptor und löst in der Zelle durch Absorption von blauem Licht eine Fluchtreaktion aus. Die Weitergabe des Signals zu dem flagellaren Motor geschieht über einen verzweigten Aktivierungsweg (Spudich 1995; Doktorarbeit Jarmila Guijarro, 2004). Diese Sensoren ermöglichen den extrem halophilen Archaeen, sich in ihrer Umgebung zu orientieren und so die geeigneten Bedingungen für ihre Photosynthese zu finden. Die primären photochemischen Prozesse sind bei den Energietransducern und auch bei den Sensory Rhodopsinen gleich. Zuerst findet eine Isomerisierung von dem all- trans Retinal (Grundzustand) zu dem 13-cis-Retinal statt. Die Konformations- änderung führt zur Aktivierung der assoziierten Einleiterproteine HtrI und HtrII. Nach diesem Schritt folgen weitere Konformationsänderungen, bis schließlich der Grund- zustand all- trans Retinal wieder erreicht wird.
Der entscheidende Unterschied der Pumpen und auch der Sensoren besteht in der Lebensdauer des 13- cis Retinal Zustandes und in der Abgabe/Aufnahme der Ionen

- 32-
von den Pumpen. SR1 braucht 250 ms und SR2 700 ms, bis der Grundzustand im Photozyklus wieder erreicht wird. Der Zeitunterschied, ist damit zu erklären, dass sie mit Hilfe ihrer Photoprodukte katalytisch ein Signal bewirken müssen. Von den Pumpen wird jedoch nur ein effektiver Ionentransport verlangt (Marvan 1987). Die Expression von HR und auch die der von Sensory Rhodopsinen ist in den Halobakterien anteilmäßig geringer.
Abb.18: Schematische Darstellung der Retinalproteine aus H. salinarum und ihre Funktionen (Jarmila Guijarro S.12 (2006), „Untersuchung des lichtgetriebenen
Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR- Spektroskopie“).
7.2 Rezeptoren Halobakterium salinarum besitzt 18 genomisch kodierte Rezeptoren, wobei nur 7 bis jetzt identifiziert wurden. Die Überträger sind modular aufgebaut und besitzen eine cytoplasmatische Domäne, die mit der Signaltransduktionskaskade wechselwirkt. Die meisten der Übertäger sind über Transmembranhelices (besitzen 1, 2 oder 6 Helices) in der Zellmembran verankert. Die Chemorezeptoren die extrazelluläre Reize erkennen, besitzen eine extrazelluläre Ligandenbindungsdomäne. Wird an den Liganden gebunden kommt es zu einer Konformationsänderung, die über die Transmembranhelices an die cytoplasmatische Domäne weitergeleitet wird und so die Signalkaskade in den Gang setzt. Die zwei Rezeptoren BR (Bakteriorhodopsin) und HR (Halorhodopsin) analysieren die Lichtverhältnisse. Bei sinkender Lichtintensität arbeiten die Rezeptoren BR und HR nicht mehr mit voller Kapazität. In Htr1 (Überträgerprotein) und Htr2 wird der Photoaktivierungszustand, der SR1 und SR2 durch die Transmembranhelices an die cytoplasmatische Domäne weitergeleitet. Der Rezeptor SR1 kann UV-Licht und gelbes Licht wahrnehmen. SR2 nimmt blaues Licht

- 33-
wahr. Die Rezeptoren SR1 und SR2 kommunizieren über die Transduktorproteine (Htr1 und Htr2) in einem Zwei- Komponenten-System (ZKS) bestehend aus einer Histidinkinase CheA und einem Antwortregulator CheY mit dem Flagellarmotor und steuern so die lichtgetriebene Bewegung des Flagellarmotors des Archaeons.
Abb.19: Schematische Beschreibung der Signaltransduktion und der bioenergetischen Systeme und daran beteiligten Ionenbewegungen (Martina Ufer,1994 „Untersuchung über die Aktivität der Nitratreduktase in dem Archaebakterium Haloferax volcanii“).

- 34-
8 UV-Schutz
Wie schon erwähnt sind die halophilen Archaea oft in südländischen Gebieten zu finden, wo sie in den Seen und den Meeren einer starken Sonneneinstrahlung ausgesetzt sind. Die Halobakterien sind in den Gewässern durch ihre auffällige Rotfärbung mit dem bloßen Auge zu erkennen. Die typische Rotfärbung tritt nicht nur bei den Halobakterien auf, die eine Purpurmembran mit Bakteriorhodopsin besitzen, sondern auch bei anderen Halobakterein. In der Cytoplasmamembran der Halo- bakterien sind langkettige Isorenoide, Bacterioruberine (C-50 Isoprenoide) und auch Carotinoide (C-40- Einheiten) wie Lycopin (Farbstoff der Tomate) eingelagert, die die unterschiedlichen Färbungen der Salzseen hervorrufen. Welche Färbung ein Organismus ausstrahlt, kommt auf die Mengenanteile der langkettigen Isoprenoide in der Membran an.
Die Intensität der Pigmentierung ist bei den meisten Halobakterien von der Salzkonzentration des Lebensraums abhängig. Bei hoher Salzkonzentration ist eine starke Pigmentierung zu erkennen, nimmt die Konzentration ab, so wird auch die Pigmentierung weniger.
Die Organismen schützen sich durch diese Pigmente vor photooxidativen Prozessen, indem sie die UV-Strahlen abfangen (Susanne Bickel-Sandkötter und Horst Bickel, 1996: „Leben im Toten Meer: Halobakterien“). Durch das UV-Licht werden die Sauerstoffmolekühle auf ein energetisch höheres Niveau gebracht und es werden Peroxide und Oxide gebildet, die die Zellen normalerweise zerstören würden. Die in der Cytoplasmamembran eingelagerten Carotinoide sind in der Lage die Energie von den Elektronen des Sauerstoffs zu übernehmen, indem sie selber in einen energetisch höheren Zustand (Triplettzustand) übergehen und die Energie in Form von Wärme wieder abgeben (Susanne Bickel-Sandkötter, Martina Ufer, Kerstin Steinert, Michaela Dana: “Leben im Salz- Halophile Archaea“).
O2 + Carotinoide → O2 + Carotinoide→ Carotinoide + Wärme Die Färbungen beschreiben die Energiezustände: gelb= Triplett-Zustand; rot= Singulett-Zustand

- 35-
Abb. 20: Strukturen von den wichtigsten Bacterioruberinen und Carotinoiden (Susanne Bickel-Sandkötter, Martina Ufer, Kerstin Steinert, Michaela Dana: “Leben im Salz-
Halophile Archaea“).

- 36-
9 Industrieller Nutzen (Nanobiologie)
Die Nanotechnologie beschäftigt sich mit Systemen in der Größe von hundert Nanometern bis hinunter zu weniger als einem Nanometer. Dies ist genau der Größenbereich der Bestandteile von Zellen. Durch die Zusammenarbeit von Nanotechnologie und Biotechnologie ergeben sich deshalb besonders faszinierende Möglichkeiten, wie zum Beispiel der Gebrauch von Halorhodopsin als photochromes Sicherheitspigment (www.mpg.de/pdf/jahresbericht2000/jahresbericht2000.pdf)
9.1 Bakteriorhodopsin als photochromes Sicherheitspigment Aufgrund von hoch modernen Druckern, Scannern und Farbkopierern ist das Fälschen und Nachahmen von Dokumenten heute kein großes Problem mehr. Die Fälschung von Führerscheinen, Personalausweisen, Reisepässen, Banknoten und Wertpapieren wird heute stark beobachtet. Mit Hilfe des biologischen Farbstoffes Bakteriorhodopsin war es möglich ein neues leistungsfähiges, weit über den Stand der Technik hinaus gehendes Sicherungselement herzustellen. Bei dem veränderten Bakteriorhodopsin ist ein mit dem Auge sichtbarer, lichtabhängiger Farbwechsel von lila nach gelb zu sehen, wenn das Molekül belichtet wird. In diesem Spektralbereich ist die Empfindlichkeit des Auges am größten. Der Lichtinduzierte Farbstoff schützt so vor Farbkopierern, Scannern und Druckern. Das Molekül bietet des Weiteren noch nützliche Eigenschaften, die vor Fälschungen von Dokumenten schützt. Durch chemische und gentechnische Modifizierung wird aus BR ein multifunktionales Material für die Dokumentenfälschung und optische Datenspeicherung. An erster Stelle sei die Möglichkeit erwähnt, Daten zu verschlüsseln und optisch zu speichern. Magnetstreifen und Speicherchips können diese Speicherkapazitäten nicht aufbringen. Mit Hilfe von biotechnologischen Herstellungsverfahren lassen sich große Mengen dieses multifunktionellen Materials Bakteriorhodopsin preisgünstig herstellen, was eine wichtige Voraussetzung für einen technischen Einsatz ist.

- 37-
Abb. 21: Die verschiedenen Farbwechsel von BR-Druckfarben: Auf der linken Seite die unbelichtet Farbe von BR und rechst der Farbwechsel nach Lichtinduzierung (Prof. Dr.
Norbert Hampp: „Biologische Sicherheitspigmente und optischer Datenspeicher“ Philipps Universität Marburg).
Der Farbwechsel von lila nach gelb, der durch die Induktion von Licht entsteht, ist sehr auffällig. Die spektrale Verschiebung (von 160nm) von lila nach gelb kann hervorge- rufen werden durch die Belichtung mit grünem Licht. Die Umkehrung von gelb nach lila kann durch sichtbares blaues Licht bewirkt werden. Das Bakteriorhodopsin zeigt eine für ein Protein einzigartige Stabilität (siehe Aufbau der Membran). Die Membran- bereiche die eine lila Färbung zeigen, in die das Bakteriorhodopsin eingelagert ist, wird als Purpurmembran (PM) bezeichnet (siehe Aufbau PM). Das Bakteriorhodopsin zeigt eine Resistenz gegen hohe Temperaturen, Lösungsmittel und auch unterschiedliche pH-Werte, denen es ausgesetzt ist, scheint ihm nicht zu schaden. Durch eine gewisse Unempfindlichkeit ist eine großtechnische Gewinnung immer besser möglich. In diesem Fall ist jedoch die wichtigste und ausschlaggebende Eigenschaft, das BR durch Lichtinduktion seine Farbe verändert und dieser Vorgang auch reversible ist. BR ist in der Lage viele Millionen Male diesen Vorgang zu wiederholen. Schon mit einer einfachen Lampe ist es möglich BR von lila nach gelb zu verändern und in Dunkelheit wird der Vorgang reversible. Wichtig dieser Vorgang ist mit dem bloßen Auge sichtbar!
Auch im Kopierschutz ist es sehr wichtig, da es seine Farbe bereits während des Kopiervorgangs verändert. Wird ein Dokument kopiert, auf dem zuvor eine mit BR behandelte Stelle lila sichtbar war, ist diese nach dem Kopieren als gelbe sichtbar. Sollte es sich um eine Fälschung handeln würde bei dem Kopiervorgang kein Farbwechsel stattfinden.
Das Retinalprotein Bakteriorhodopsin besitzt 248 Aminosäuren (s. o. Aufbau BR). Viele von diesen AS sind für das BR nicht essentiell und können somit ausgetauscht werden gegen andere AS. Die meisten der AS sind in den sieben Alpha- Helices zu finden und nur die N- und C-Termini liegen außerhalb. Mit Hilfe einer aufwendigen Methode (Sequenzierung) ist es möglich die C-terminalen extramembranen Aminosäurekette zu

- 38-
ersetzen. Nicht das Ersetzen ist mit großem Aufwand verbunden, sondern die Identifizierung der C-terminalen extramembranen Aminosäureketten stellt das Problem dar.
Abb. 22: Schematische Darstellung des Bakteriorhodopsins mit den sieben Alpha-Helices und den C und N terminalen Sequenzen, die außerhalb der Membran liegen
(http://www.techportal.de/uploads/publications/504/InfoPhysTech40.pdf).
Werden speziell veränderten Peptidstücke für die Dokumentensicherung hergestellt, ist das veränderte BR zu seinem Hersteller zurück verfolgbar. Die veränderten Sequenzen enthalten eine charakteristische Aminosäuresequenz durch die sie identifiziert werden können. Bei einem Peptid mit 10 AS (siehe Abb. rot markierten AS) können 1024 Codes gebildet werden. Die Veränderung der AS bildet quasi eine Adresse, über die es immer wieder identifiziert werden kann. Die Funktion des BR verändert sich nicht. Bei der Datenspeicherung handelt sich um den WORM-Speicher (write- once- read- many). Der Speicher kann beschrieben werden, aber nicht mehr gelöscht werden. Auf einem Abschnitt von 6cm × 1cm ist es möglich, Daten im Megabit- Bereich zu speichern. Die Detektion kann preiswert maschinell durchgeführt werden. Es können Bilder aber auch digitale Daten auf diesem Datenspeicher festgehalten werden (http://www.uni-marburg.de/fb15/ag-hampp/forschung/BR/Info/PDF2002DInfo PhysTech_BR).

- 39-
Literaturverzeichnis Besir, Hüseyin Dissertation (2001): “Untersuchung der lipidvermittelten Kristallisation der Ionenpumpen Bakteriorhodopsin und Halorhodopsin aus Halobakterium salinarum“
Falb, Michaela Dissertation (2005): “Computational Genome and Pathway Analysis of Halophilic Archaea
Gregor, Dagmar Dissertation (2002):“Untersuchungen zur Regulation von Promotoren aus halophilen Archaea mit dem bgaH-Reportergen“
Guijarro, Jarmila Dissertation (2006): „Untersuchung des lichtgetriebenen Chloridtransports in Halorhodopsin aus Natronobakterium pharaonis mit statistischer und zeitaufgelöster FTIR-Spekroskopie“
Hausmann K., Kremer B. P.: „Extremophile Mikroorganismen in ausgefallenen Lebensräumen“
Hofacker, Annette Dissertation (2002): „Regulation der Gasvesikelbildung bei Halobacterium salinarum PHH1“
Marg, Bianca-Lucia Dissertation (2002): „Struktur und Halophilie eines archealen Ferredoxins“
Oesterhelt, Dieter; Marvan Wolfgang (01/05): „Salz in der Suppe des Lebens“ Biospektrum S. 60-62
Markus Roeßler Dissertation (2001):“ Chlorid, ein neues Signalmolekül für Bakterien: Identifizierung und molekulare Charakterisierung Cl- regulierter Prozesse in dem moderat halophilen Bakterium Halobacillus halophilus“
Sartorius-Neef, Simone Dissertation (2004): „Untersuchung zur Initiation der Translation in halophilen Archaea“
Sergiu Fendrihan, Andrea Legat, Marion Paffenhuemer, Claudia Gruber, Gerhard Weidler, Friedrich Gerbl, Helga Stan-Lotter; Review paper: “Extremely halophilic archaea and the issue of long term microbial survival” 6March 2006; Puplished online: 19 July 2006 Springer Science+ Business Media B.V 2002
Susanne Bickel-Sandkötter, Wolfgang Gärtner, Michaela Dane; Springer Verlag (1996) „Conversion of energy in halobacteria: ATP synthesis and phototaxis“
Susanne Bickel-Sandkötter und Horst Bickel (1996) „Leben im Toten Meer: Halobakterien“

- 40-
Susanne Bickel-Sandkötter, Martina Ufer, Kerstin Steinert, Michaela Dana (1995) Biologie in unserer Zeit: “Leben im Salz- Halophile Archaea“
Sinn-Meyer, Bianca Dissertation (2003): „Physiologische und molekularbiologische Untersuchungen anhydrolytischen extrazellulären Enzymen aus extremophilen marinen Mikroorganismen unter besonderer Berücksichtigung von Nucleasen“
Steinert, Kerstin Dissertation (1995) „Molekulare Charakterisierung der ATPase aus dem Archaeon Haloferax volcanii“
D. Yu. Sorokin, T. P. Tourova, E. A. Galinski, C. Belloch † International Journal of Systematic and Evolutionary Microbiology (2006) S.379-388 “Extremely halophilic denitrifying bacteria from hypersaline inland lakes, Halovibrio denitrificans sp. nov. and Halospina denitificans gen.nov., sp. nov., and evidence that the genus name Haovibrio Fendrich 1989 with the type species Halovibrio variabilis should be associated with DSM 3050”)
Tebbe, Andreas Dissertation (2005): „Das Proteom eines halophilen Archaeons und seine Antwort auf Änderung der Lebensbedingungen- Inventarisierung, Quantifizierung und posttranslationale Modifikationen“
Martina, Ufer Dissertation (1994): „ Untersuchung über die Aktivität der Nitratreduktase in dem Archaebakterum Haloferax volcanii“
Wende, Andy Dissertation (2006): “Signaltransduktion durch Zwei-Komponenten Systeme in dem halophilen Archaeon Halobacterium salinarum“
S. 12 http://www.nottingham.ac.uk/genetics/people/allers/research.php
S. 14 www.lazypics.de/.../thumbnails/lake_magadi.jpg
S. 14 http://www.kultour.ch/media/images/TOTES_MEER_Zeitung.jpg
S. 15 / 16 www.uni-giessen.de/.../extrem_halophile.html
S. 17: www.visindavefur.hi.is/myndir/halobacteria_09...
S.18 www.bph.ruhr-uni-bochum.de/bilder/br_abb01.jpg
S. 22 http://www.biophysik.uni-freiburg.de/projekte/diplom_br.html
S.25 http://www.mpg.de/bilderBerichteDokumente/dokumentation/press
emitteilungen/2000/pri49_00.htm
S. 35 www.mpg.de/pdf/jahresbericht2000/jahresbericht2000.pdf
S. 37 http://www.techportal.de/uploads/publications/504/InfoPhys Tech40.pdf
S. 37 http://www.unimarburg.de/fb15/aghampp/forschung/BR/Info/P DF_2002_D_InfoPhysTech_BR

- 41-
S. 37 http://www.uni-marburg.de/fb15/ag-hampp/forschung/BR/Praesentation BR/p08

- 42-
Erklärung Hiermit erkläre ich, dass ich die vorliegende Bachelorarbeit selbständig angefertigt habe. Es wurden nur die in der Arbeit ausdrücklich benannten Quellen und Hilfsmittel benutzt. Wörtlich oder sinngemäß übernommenes Gedankengut habe ich als solches kenntlich gemacht.
Ort, Datum Unterschrift

- 43-
Stichwortverzeichnis Abbildungsverzeichnis 6
Abkürzungsverzeichnis 8
Abstract 2
Anaerobe Atmung 21
Arten 11
Bakteriorhodopsin 24, 34, 35, 37
Carotinoide 35
Dritte Domäne 13
Einleitung 9
Erklärung 40
extrem Halophil 15
Fermentation 21
Flagellarmotor 28
Gasvesikel 9
Geißeln 28
Halobakterium salinarum 17
Halorhodopsin 26, 27
Inhaltsverzeichnis 2
Konkurrenz 9
Kopierschutz 37
Lake Magadi 14
Literaturverzeichnis 38, 39
Meerwasser 14
Membranaufbau 19
Motilität 29
Na+/H+-Antiporter 28
Photozyklus BR 24
Photozyklus HR 26
Pururmembran 20
Pylogenetischer Stammbaum 9
Photosensoren 31
Retinalproteine 21, 22
Retinal 23
Rezeptoren 32
Tabellenverzeichnis 7
UV-Schutz 33
Vorkommen 12
Zusammenfassung 2

















![Photodynamics of BLUF Domain ProteinsBesides visual rhodopsins from Eukarya and archaea, they are also found to be phototaxis receptors in chlamydomonas [Sak02]. Phytochrome photoreceptor](https://img.pdfslide.org/doc/110x75/60fb2cb54a994c3f456b22a0/photodynamics-of-bluf-domain-proteins-besides-visual-rhodopsins-from-eukarya-and.jpg)











