Embed Size (px)
Citation preview

Calcareous nannoplankton of the Red Sea:
A proxy for the reconstruction of the paleoenvironmental conditions during
the late Quaternary
Dissertation
zur Erlangung des akademischen Grades
eines Doktors der Naturwissenschaften
an der Fakultät für Geowissenschaften
der Ruhr-Universität Bochum
vorgelegt von
Heiko-Lars Legge
aus Bochum
Bochum,
Oktober 2007

Die vorliegende Arbeit wurde von der Fakultät für Geowissenschaften der Ruhr-Universität
Bochum als Dissertation zur Erlangung des akademischen Grades eines Doktors der
Naturwissenschaften anerkannt.
Erster Gutachter: Prof. Dr. Mutterlose
Zweiter Gutachter: Prof. Dr. Heimhofer
fachfremder Gutachter: Prof. Dr. Zepp
Tag der Disputation: 9. Januar 2008

Table of contents
I
Acknowledgements III
Abstract V
Kurzfassung VII
1 Introduction 1
1.1 Oceanography of the Red Sea 1
1.2 Paleoceanographic background 3
1.3 Calcareous Nannoplankton 4
1.4 Synopsis 7
Explanatory notes 9
References 10
2 Climatic changes in the northern Red Sea during the last 22,000 years as recorded by
calcareous nannofossils 17
Abstract 17
2.1 Introduction 17
2.2 Material and Methods 20
2.3 Results 22
2.4 Discussion 25
2.4.1 Last Glacial Maximum (LGM, circa 22-19 ka) 26
2.4.2 Heinrich Event 1 (H1, circa 18-15.5 ka) 29
2.4.3 H1-Bolling Transition (circa 15.5-14.6 ka) 30
2.4.4 Bolling-Allerod (circa 14.6-13.1 ka) 32
2.4.5 Younger Dryas (YD, 13.1-11.7 ka) 34
2.4.6 Younger Dryas Transition (circa 11.7-11.4 ka) 36
2.4.7 Earliest Holocene Period (circa 11.4-9.5 ka) 37
2.4.8 Early Holocene to Mid-Holocene Humid Period (circa 9.5-6 ka) 38
2.4.9 Mid-Holocene to Late Arid Holocene (circa 6-0.6 ka) 40
2.5 Conclusion 41
Acknowledgements 42
References 43
3 Nannoplankton successions in the northern Red Sea during the last glaciation (60 to
14.5 ka): Reactions to climate change 53
Abstract 53
3.1 Introduction 53
3.2 Material and Methods 55

Table of contents
II
3.3 Results and Discussion 56
3.4 Conclusion 64
Acknowledgements 64
References 65
4 Late Holocene environmental changes in the northern Red Sea region as indicated by
nannoplankton abundance patterns 71
Abstract 71
4.1 Introduction 71
4.2 Methods 73
4.2.1 Material and geological Background 73
4.2.2 Sampling and preparation 74
4.3 Gephyrocapsa spp. abundance patterns 75
4.4 Discussion 77
4.5 Conclusion 83
Acknowledgements 84
References 85
5 Summary 93
6 Outlook 97
Appendix 99
Curriculum Vitae 119

Acknowledgements
III
Acknowledgements
I am grateful to Dr. Jörg Mutterlose (Bochum) for initiating and supervising this thesis.
I would like to thank Dr. Jürgen Pätzold (Bremen), Dr. Helge W. Arz (Potsdam) and Dr.
Ismene A. Seeberg-Elverfeldt (Bremen) for providing sample material and geochemical data.
Varies kind of support were provided by colleagues at the Ruhr-University of Bochum. Dr.
Jens Steffahn (now Universidad Autónoma de Nuevo Léon MÉXICO) and my “fellow-
sufferers” Andreas Rexfort and Petros Hardas are thanked for a constructive and friendly
working atmosphere. I’m indebted to the working group of mineralogy/petrography for
providing access to their laboratory for sample preparation. Dr. Rolf Neuser is thanked for his
help at the SEM.
Special thanks to Dr. André Bornemann (now Leipzig) for varies kind of help and many
fruitful discussions during the last years.
Above all, I am grateful to Sylvia and my family for their continuous interest and moral
support.
This research was supported the by German Research Foundation (DFG Mu 667/23-1, -2 and
Mu 667/26-1, -2) and the Ruhr-University of Bochum (Allgemeines Promotionskolleg).

IV

Abstract
V
Abstract
The objective of this thesis is to understand the calcareous nannoplankton response to past
environmental changes in the northern Red Sea over the last 60,000 years. Small-sized pro-
and eukaryotic phytoplankton dominates the oligotrophic Red Sea environment. Calcareous
nannoplankton is well abundant in the living phytoplankton community and a key component
of the marine sediments. Due to its distinct and fast reaction to environmental changes and its
good preservation potential, it is well suited for paleoceanographic reconstructions. The study
of the calcareous nannoplankton, and the evaluation of its relative and absolute abundances, is
therefore indicative for paleoceanographic changes in the northern Red Sea. Calcareous
nannoplankton findings supplemented by geochemical and sedimentological data were
compared with other marine and terrestrial proxy records and discussed with respect to the
climate history established so far. The thesis presented here is divided into three parts, which
focus on the following aspects of the northern Red Sea history:
1.) Paleoceanographic reconstruction of the northern Red Sea over the last 22,000 years.
2.) Calcareous nannoplankton response to well-documented climate oscillations (long-term
cooling cycles, Heinrich events) of Marine Isotope Stages 3 and 2.
3.) High-resolution analysis of the late Holocene by investigating laminated sediments from a
brine-filled, submarine basin located in the northern Red Sea (Shaban Deep).
It is evident from this study that the calcareous nannoplankton community mirrors
paleoceanographic changes of the northern Red Sea, which are driven by long- and short-
termed climate variations. The paleoceanography of the northern Red Sea shows marked
similarities with the climate and vegetation history of northern Africa and the Near East.
Intervals of enhanced primary production in the marine environment go along with humid
periods on land. Another important result of the thesis is the existence of calcareous
nannoplankton in the northern Red Sea during glacial periods. Furthermore, our results
question the applicability of the geographic isolation of the Red Sea to explain calcareous
nannoplankton community changes during these periods: Due to the restricted water exchange
with the Gulf of Aden-Indian Ocean via the narrow and shallow Strait of Bab-el-Mandeb, the
modern Red Sea is characterized by high saline waters. The restricted water exchange during
glacial times (e.g., Last Glacial Maximum) is a result of the global sea level lowering and has
caused hypersaline conditions. In previous studies it has been suggested that the glacial
progression caused the absence of planktic foraminifera and the occurrence of a monospecific
pteropod population in the northern Red Sea. There is ample evidence from this study that
basic variations of the calcareous nannoplankton community structure, even during full

Abstract
VI
glacial conditions, were driven by regional hydrographic changes under the atmospheric
influence of the extratropics. Changes of surface-water stratification and thereby of the
vertical nutrient distribution within the photic zone seem to be extremely important. The
influence of these processes was probably not restricted to glacial periods; even the
calcareous nannoplankton community structure during the Holocene seems to be affected.

Kurzfassung
VII
Kurzfassung
Im Rahmen der vorliegenden Dissertation sollte die Reaktion des kalkigen Nannoplanktons
auf Veränderungen der Paläoumwelt an Sedimentkernen aus dem nördlichen Roten Meer über
den Zeitraum der letzten 60,000 Jahre untersucht werden. Unter oligotrophen Bedingungen,
wie sie im heutigen Roten Meer vorherrschen, sind die Phytoplanktonvergesellschaftungen
durch kleine Pro- und Eukaryonten gekennzeichnet, wobei das kalkige Nannoplankton die
wichtigste fossil überlieferbare Gruppe repräsentiert. Aufgrund ihrer differenzierten und
schnellen Reaktion auf Veränderungen der Umwelt und ihres guten Erhaltungspotentials
bieten Wechsel in der Zusammensetzung der kalkigen Nannoplanktongemeinschaften eine
ideale Möglichkeit ozeanographische Veränderungen zu rekonstruieren. Durch die Analyse
des kalkigen Nannoplanktons sowie die quantitative und qualitative Erfassung ihrer
Häufigkeiten, können paläoozeanographische Schwankungen im nördlichen Roten Meer
aufgezeigt werden. Die Ergebnisse aus den Untersuchungen des kalkigen Nannoplanktons
wurden, ergänzt durch umfangreiche geochemisch-sedimentologische Daten, mit bereits
publizierten Arbeiten aus dem marinen und terrestrischen Bereich verglichen und vor dem
Hintergrund der derzeit bekannten Klimageschichte diskutiert. Innerhalb der vorliegenden
Dissertation lassen sich drei Untersuchungsschwerpunkte unterscheiden:
1.) Die Rekonstruktion der Paläoozeanographie des nördlichen Roten Meeres während der
letzten 22,000 Jahre.
2.) Die Untersuchung des Einflusses der für die Marinen Isotopenstadien 3 und 2
charakteristischen Klimaschwankungen (Abkühlungszyklen, Heinrichereignisse) auf das
kalkige Nannoplankton.
3.) Eine zeitlich hochauflösende Analyse des späten Holozäns unter Zuhilfenahme laminierter
Sedimente aus einem im nördlichen Roten Meer gelegenen Solebecken (Shaban Tief).
Die durchgeführten Untersuchungen zeigen, dass sich paläoozeanographische Veränderungen
im nördlichen Roten Meer, gesteuert durch lang- und kurzfristige Klimavariationen, in den
Nannoplanktongemeinschaften widerspiegeln. Langfristige Veränderungen der Paläoozeano-
graphie zeigen deutliche Parallelen zur Klima- und Vegetationsgeschichte Nordafrikas und
des Nahen Ostens. So korrelieren beispielsweise Intervalle erhöhter Primärproduktion im
nördlichen Roten Meer meist mit humiden Klimaperioden im terrestrischen Bereich. Ein
weiteres wichtiges Ergebnis der durchgeführten Untersuchungen ist der Nachweis des
kalkigen Nannoplanktons im nördlichen Roten Meer auch während glazialer Zeitabschnitte.
Von besonderer Bedeutung ist in diesem Zusammenhang die Beobachtung, dass
Schwankungen innerhalb der glazialen Nannoplanktongemeinschaft nicht alleine über die

Kurzfassung
VIII
isolierte geographische Lage des Roten Meeres erklärt werden können: Der eingeschränkte
Austausch mit dem Indischen Ozean über die enge und flache Straße von Bab-el-Mandeb
führt zu erhöhten Salzgehalten im heutigen Roten Meer. Bedingt durch die Regression des
globalen Meeresspiegels waren kühlere Phasen in der Erdgeschichte (z.B. Letztes Glaziales
Maximum) durch hypersaline Bedingungen geprägt. Die Verschärfung des eiszeitlichen
Klimas ist im nördlichen Roten Meer durch das Verschwinden planktonischer Foraminiferen
und die Ausbildungen einer nahezu monospezifischen Pteropoden-Gemeinschaft
gekennzeichnet. Die im Rahmen der Dissertation durchgeführten Untersuchungen deuten
darauf hin, dass selbst während der Hochphase der letzten Vereisung wesentliche
Veränderungen innerhalb der kalkigen Nannoplanktongemeinschaft durch andere Faktoren
gesteuert wurden. Veränderungen der regionalen Hydrography, insbesondere Schwankungen
der Intensität und Häufigkeit der winterlichen Durchmischung bedingt durch den variierende
Einfluss außertropischer Luftmassen, scheinen Schlüsselelemente darzustellen. Dieses gilt
nicht nur für das glaziale Rote Meer. Vergleichbare Prozesse scheinen auch während des
Holozäns die Zusammensetzung des kalkigen Nannoplanktons beeinflusst zu haben.

Chapter 1: Introduction
1
1 Introduction
The chapter “Introduction” is subdivided into four sections. First, the modern oceanography
of the Red Sea is briefly summarized in section 1.1. The aim of section 1.2 is to illustrate how
basic changes of the Red Sea environment were connected to variations of the global sea
level. Section 1.3 highlights the importance of calcareous nannoplankton in modern research
and gives some remarks concerning the terminology. In addition, with special reference to the
calcareous nannoplankton, the setting of the study domain is shortly discussed. In section 1.4,
the major goals of the thesis will be summarized. An overview of the performed work will be
given briefly in conjunction with the used sediment material.
1.1 Oceanography of the Red Sea
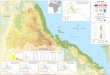
The Red Sea is a desert-surrounded ocean basin with a restricted seaway to the Indian Ocean-
Gulf of Aden via the shallow (137 m) and narrow (30 km) Strait of Bab-el-Mandeb at its
southern end (Figure 1.1). Encompassing the
Sinai Peninsula, the Gulfs of Suez and
Aqaba mark the northernmost extensions of
the Red Sea. As part of the Red Sea-Gulf of
Aden rift system (with the Afar triple
junction), the Red Sea represents an example
of a relatively young ocean basin (Bosworth
et al., 2005). It is located in the northwest-
trending rift zone where the African-Arabian
plate was broken and spread apart. The
spreading axis extends up the Red Sea main
basin. In addition, due to the movement of
the Arabian plate in northeasterly direction,
a large fracture developed at the passive
plate margin (Almond, 1986; Bosworth et al., 2005). It forms the Gulf of Aqaba and the
trough of the Jordan valley with a number of lake basins (Dead Sea, Sea of Galilee).
The modern climate is arid, with low precipitation (10–200 mm/year) and high rates
(2000 mm/year) of evaporation (Edwards, 1987; Sofianos et al., 2002). No major river system
supplies the Red Sea with freshwater and the influence of temporary run-off (e.g., from
wadis) on the hydrological budget is negligible (Morcos, 1970; Hoelzmann et al., 1998;
Siddall et al., 2003).
Figure 1.1. Map showing the Red Sea area and the Gulf of Aden-Indian Ocean region.

Chapter 1: Introduction
2
Northwesterly winds characterize the northern Red Sea during the entire year (Edwards,
1987). In summer these winds also dominate the southern parts, whereas in winter the
influence of the Indian monsoon system leads here to a reversal of the main wind direction.
The interaction between southeasterly winds over the southern Red Sea and the prevailing
northwesterly winds over the northern parts results in a convergence zone across the central
Red Sea (e.g., Edwards, 1987).
The circulation patterns of the Red Sea waters are anti-estuarine and driven by wind and
especially thermohaline forcing (e.g., Eshel and Naik, 1997). Warm and normal saline waters
enter the Red Sea from the Gulf of Aden. As this water moves northwards along the
longitudinal axis of the basin, it becomes cooler and saltier. Due to biological consumption
(Weikert, 1987) in the southernmost Red Sea, the nutrient gradients along the longitudinal
axis are less pronounced than those for salinity and temperature. Compared with the
oligotrophic central and northern Red Sea, the southern Red Sea is relatively nutrient-rich due
to the inflow from the Gulf of Aden (for details see e.g., Levanon-Spanier et al., 1979; Shaikh
et al., 1986; Weikert, 1987 and references therein). Enhanced productivity in the southern Red
Sea is displayed by an upward shift of the Oxygen Minimum Zone (OMZ) and lower values
of dissolved oxygen compared with the northern part (Weikert, 1987). Bacterial consumption,
associated with the organic decomposition, reduces the oxygen concentration rapidly below
surface waters. In the northern Red Sea lowest values of around 1–1.75 ml O2/l are located at
400–500 m, whereas the oxygen concentration in the southern Red Sea decreases (0.3–0.5 ml
O2/l) at 300–400 m water-depth. Below the OMZ, the concentration increases slowly again.
Values of around 2–3 ml O2/l are found in the deep-water (Edwards, 1987; Weikert, 1987). It
is worth mentioning that the oxygen gradient between the northern and southern Red Sea
reflect also the increasing distance from the area of intermediate and deep-water formation.
The journey of the Red Sea surface waters flowing from south to north eventually ends in
the northern most parts of the Red Sea where it sinks and forms intermediate and deep-water.
The resulting southward deep flow ventilates the Red Sea (e.g., Grasshoff, 1969; Morcos,
1970; Wyrtki, 1971; Manis, 1973; Cember, 1988; Woelk and Quadfasel, 1996; Eshel and
Naik, 1997). Different locations and modes of intermediate and deep-water formation are
identified (cf. Figure 1 in Arz et al., 2003): (1a) Deep convection as well as (1b) direct
injection beneath the pycnocline in the northern Red Sea nearby the Sinai tip (Cember, 1988).
(2) Shallow convection at the collision site of the Western and Eastern boundary current at
around 26–26.5°N (Eshel and Naik, 1997). (3) Deep-water formation in the northern Red Sea

Chapter 1: Introduction
3
initiated by the outflow of dense waters from the shallow (50–70 m) Gulf of Suez (Woelk and
Quadfasel, 1996).
1.2 Paleoceanographic background
It is well accepted that the Red Sea environment and its biological interior suffered extreme
oceanographic changes during the last glacial–interglacial cycles (e.g., Locke and Thunell,
1988; Hemleben et al., 1996; Rohling et al.,
1998; Siddall et al., 2003; Arz et al., 2007). As
stated above, the restricted water exchange with
the Gulf of Aden-Indian Ocean via the
bottleneck at Bab-el-Mandeb controls the
hydrological budget. Hence, variations of the
global sea level dictate the quantity of water
entering the Red Sea. During the Last Glacial
Maximum (LGM), for example (Figure 1.2), the
sea level decreased nearly by 120 m. This drop
reduced the water depth at the Strait of Bab-el-
Mandeb to approximately 17 m. The salinity of
the Red Sea waters increased dramatically due
to the strongly limited exchange (Winter et al., 1983; Locke and Thunell, 1988; Hemleben et
al., 1996; Siddall et al., 2003; Arz et al., 2007). Stable oxygen isotope measurements reveal
increased values in comparison to the global trend (e.g., Hemleben et al., 1996). The
hypersaline conditions (49–50‰ and more; Thunell et al., 1988; Hemleben et al., 1996;
Fenton et al., 2000) reached the tolerance limit for various planktic organisms. Final
ecological consequences of the glacial progression were the absence of planktic foraminifera
(“aplanktic zone”; e.g., Winter et al., 1983; Almogi-Labin et al., 1991; Hemleben et al., 1996)
and the occurrence of a nearly monospecific pteropod (Creseis acicula) community (Chen,
1969; Almogi-Labin, 1982; Winter et al., 1983).
In comparison to foraminifera and pteropoda, little is known about the calcareous
nannoplankton of the Red Sea (McIntyre, 1969; Müller, 1976; Winter, 1982a, 1982b). The
few calcareous nannoplankton studies, which have been carried out yet, suggest a massive
decrease or even the total absence of calcareous nannoplankton during the LGM (McIntyre,
1969; Winter et al., 1983). However, in order to understand past changes of the Red Sea
environment and its impact on the phytoplankton community, continuous and well-dated
Figure 1.2. The Red Sea at present and
during the LGM is schematically shown. For detail see text.

Chapter 1: Introduction
4
marine records are necessary. Until now a detailed reconstruction of the calcareous
nannoplankton abundance patterns is still missing.
1.3 Calcareous Nannoplankton
Like other subtropical pelagic ecosystems, the northern Red Sea is dominated by minute
primary producers (e.g., Winter et al., 1979; Kleijne et al., 1989; Lindell and Post, 1995).
Adapted to oligotrophic environmental conditions, photo-
autotrophic pico- and nan(n)oplankton are the most
common phytoplankton. The latter group comprises as one
important component of the phytocoenosis the calcareous
nannoplankton (Levanon-Spanier et al., 1979; Winter et al.,
1979; Kleijne et al., 1989).
Calcareous nannoplankton, or more precisely their
fossil remains, the calcareous nannofossils, are well suited
as biostratigraphic markers for Mesozoic and Cenozoic
sediments (e.g., Hay and Mohler, 1967; Martini, 1971;
Thierstein, 1971, 1976; Sissingh, 1977; Perch-Nielsen,
1979; Okada and Bukry, 1980; Bown et al., 1998). For a biologist, the term calcareous
nannoplankton covers rather a morphological conglomeration than a coherent phylogenetic
cluster. Based primarily on morphology and crystal
structure, it includes traditionally three major groups:
coccoliths, nannoliths and tiny calcareous dinocysts
(calcispheres). The subjects of the thesis, coccoliths
and nannoliths, are probably taxonomically closely
related, whereas the exceptional position of the
calcareous dinocysts is obvious. Coccoliths, mineral-
ized low Mg-calcite scales (Figure 1.3, Emiliania
huxleyi), are formed by coccolithophorids (or cocco-
lithophores). Coccolithophorids are algal protists,
which belong to the class Prymnesiophyceae (Green
and Jordan, 1994; Billard and Inouye, 2004). As part
of the division Haptophyta, two layers of organic
scales, two golden-brown chloroplasts, two flagella (without any hair-like appendage) and a
haptonema identify them despite their coccoliths (Figure 1.4). It is worth mentioning that a lot
Figure 1.3. E. huxleyi.
Coccolith-bearing species. Bar = 1µm.
Figure 1.4. Coccolithophorid cell;
motile live cycle stage with two
flagella and the haptonema (modified after Billard and Inouye, 2004).

Chapter 1: Introduction
5
of coccolithophorids (respectively their main live cycle stage) are non-motile (Pienaar, 1994).
Furthermore, the haptonema is often strongly reduced or even visual absent (Green and
Jordan, 1994).
Some nannoliths (Figure 1.5, Florisphaera profunda (Nannoliths incertae sedis)) are
probably also produced by coccolithophorids, whereas the origin of some other is still
unknown (cf. Young et al., 2003). Most recent plankton- and micropaleontological studies
make no differentiation between coccolith- and nannolith-bearing species and include both
under the term: coccolithophorids, coccolithophores or calcareous nannoplankton in their
description. To avoid confusion, the presented study followed this schema. The term
calcareous nannoplankton/-fossils comprise here coccoliths and nannoliths only; calcareous
dinocysts have not been considered.
Due to their distinct affinity to autecological factors (e.g., Brand, 1994; Winter et al.,
1994; Ziveri et al. 2004), calcareous nannoplankton is often applied for paleoceanographic
reconstructions (e.g., McIntyre and Bé, 1967; McIntyre,
1970; Okada and Honjo, 1973, 1975; Honjo, 1977; Okada
and McIntyre, 1977, 1979; Reid, 1980; Roth and
Coulbourn, 1982; Samtleben and Schröder, 1992; Molfino
and McIntyre, 1990; Beaufort et al., 1997; de Garidel-
Thoron et al., 2001). Information gained from intensive
biogeographic studies in the Atlantic and Pacific Ocean
allow a separation of five major calcareous nannoplankton
zones: Subartic, temperate, subtropical, tropical and
subantartic (McIntyre and Bé, 1967; Okada and Honjo,
1973, summarized in Winter et al., 1994). The
oceanographic setting of the Red Sea corresponds closely to that of the subtropical zone. In
general there is a permanent stratification of the highly transparent upper waters caused by
high solar irradiance and overall stable environmental conditions. The stratification impeded
the return of recycled inorganic nutrients from deeper waters and caused thereby oligotrophic
surface water conditions (e.g., Li et al., 1998; Post, 2005). Generated by the stratification and
characterized by specialized calcareous nannoplankton communities (Winter et al., 1994, and
references therein), a vertical subdivision of the photic zone in at least three intervals is
possibly: the upper photic zone (0–80 m; characteristic calcareous nannoplankton species e.g.,
Rhabdosphaera clavigera, Discosphaera tubifera), the middle photic zone (80–120 m; less
well defined by specific marker species) and the lower photic zone (120–220 m; characteristic
Figure 1.5. F. profunda.
Nannolith-bearing species. Bar = 1µm.

Chapter 1: Introduction
6
calcareous nannoplankton species e.g., Florisphaera profunda, Gladiolithus flabellatus (=
Thorosphaera flabellate)).
The hydrographic setting and hence the overall oligotrophic environmental conditions, as
typical for the subtropics, are modulated in our study domain during the winter season. The
annual cycle is characterized by a short winter period with lower temperatures. Cooling-
induced overturning of the water masses results in a short interval of high marine
productivity. Detailed insights into the succession patterns of various phytoplankton groups in
the northern Red Sea region are especially
available from the Gulf of Aqaba (e.g.,
Winter et al., 1979; Lindell and Post, 1995;
Post, 2005). Accompanied by distinctive
succession patterns of the phytoplankton,
the environment is characterized by a semi-
annual alternation of water column
conditions (Post, 2005). Surface waters are
characterized during most of the year by a
shallow, but stable stratification. Cold air
incursion in late fall–early winter causes a
density increase of the surface waters
(Figure 1.6). The resulting erosion of the
stratification induces a deep convective
mixing of the waters. Although limited in
strength, in relation to the Gulf of Aqaba,
the overturning of the waters during the
short winter season triggers also the
northern Red Sea environment. As
described in section 1.1, the open northern
Red Sea is the area of intermediate and
deep-water formation. Several studies have
demonstrated the sensitivity of the northern Red Sea hydrography to mid-latitude continental
climate and the influence of temperature (and evaporation) as key factors controlling surface
water stratification (e.g., Eshel et al., 2000; Arz et al., 2003; Rimbu et al., 2006).
Figure 1.6. Seasonal succession of three
major phytoplankton groups in the Gulf of
Aqaba (Lindell and Post, 1995). Shown are
cell numbers per 1 m2 for a 600 m depth
water column. Seawater temperatures at
Eilat (4 m water depth) are from Ben-David-Zaslow et al. (1999).

Chapter 1: Introduction
7
1.4 Synopsis
Late Pleistocene paleoecological conditions of the Red Sea waters were reconstructed in the
past mainly by using foraminifera (e.g., Berggren and Boersma; 1969, Geiselhart, 1998;
Fenton et al., 2000) and pteropoda (e.g., Chen et al., 1969; Reiss et al., 1980; Locke and
Thunell; 1988; Almogi-Labin et al., 1998). The current thesis presents the first continuous and
well-dated calcareous nannoplankton record from the northern Red Sea. The objective of this
study was to evaluate the calcareous nannoplankton patterns in response to paleoceanographic
changes over the last 60,000 years (Marine Isotope Stages MIS 1, MIS 2 and MIS 3). The
calcareous nannoplankton results were compared with other marine as well as terrestrial
proxy records to allow an appropriate reconstruction of the northern Red Sea paleoclimate. To
gain relative and absolute abundances, the splitting-filtration technique of Andruleit (1996)
was used for preparation. At least 300 specimens per sample were counted by using a
scanning electronic microscope (LEO 1530 Gemini FESEM). The species identification
followed the taxonomic descriptions of Jordan and Kleijne (1994) and Young et al. (2003).
More details are given in the following chapters and the Appendix.
An overview of the research performed follows. First of all, we take a brief look to the
studied sediment cores GeoB 5844-2 and GeoB 7805-1. Following this, it is shown how the
three sub-studies of the thesis are related and why the performed work focused on specific
aspects of the northern Red Sea paleoceanography.
GeoB 5844-2
Late Glacial-Holocene sediments covering the last 60,000 years
have been studied using core GeoB 5844-2 (Figure 1.7). The
gravity core was taken in 1999 near the tip of the Sinai Peninsula
(27°42´81"N, 34°40´90"E; water depth: 963 m) during RV
Meteor cruise M44/3 (Pätzold et al., 2000).
GeoB 7805-1
Late Holocene sediments spanning the last two millennia have
been analyzed using core GeoB 7805-1 (Figure 1.7). The core
was taken with a multicorer during RV Meteor Cruise M52/3
(Pätzold et al., 2003) from the eastern sub-basin of the Shaban
Deep (26°13!9" N and 35°22!6" E; water depth: 1447 m).
Figure 1.7. Sketch of the
northern Red Sea area
showing locations of the
cores GeoB 5844-2 and
GeoB 7805-1.

Chapter 1: Introduction
8
The calcareous nannoplankton findings obtained from sediment core GeoB 5844-2 are
presented in the form of two manuscripts. The first manuscript deals with the calcareous
nannoplankton variations over the last 22,000 years: Climatic changes in the northern Red
Sea during the last 22,000 years as recorded by calcareous nannofossils. An interesting result
obtained from this study is the existence of calcareous nannoplankton during the hypersaline
period in the northern Red Sea. Though faced with the overwhelming importance of the
glacial salinity increase, some evidence indicates a lasting extratropical atmospheric control
on northern Red Sea environmental changes and calcareous nannoplankton abundance
patterns especially during cold periods like the LGM, Heinrich event 1 and the Younger
Dryas.
To check these findings in more detail, we went further back in time and investigated the
development of the calcareous nannoplankton community since the onset of MIS 3 (around
60,000 years ago) in the second study. The calcareous nannoplankton in sediment core GeoB
5844-2 offered the unique possibility for the examination of millennial-scale environmental
changes during the hypersaline period of the northern Red Sea in response to the longer-term
cooling cycles and Heinrich events. One major aim was to examine whether the calcareous
nannoplankton in the northern Red Sea responses to the well-documented climate oscillations
of the North Atlantic realm or not. The results are presented in the second manuscript:
Nannoplankton successions in the northern Red Sea during the last glaciation (60 to 14.5 ka):
Reactions to climate change. The basic patterns of the North Atlantic climate oscillations
were clearly found in this study. An interesting observation was the recurrent appearance of
E. huxleyi and Gephyrocapsa spp. (G. ericsonii and G. oceanica). The longer-term cooling
cycles are characterized by increasing values of Gephyrocapsa spp. Caused by cold air
incursion from the extratropics, the Gephyrocapsa spp. are thought to indicate an intensified
winter mixing of the waters. The results of the second study lead to the conclusion that the
atmospheric influence from the mid-latitudes via its impact on the northern Red Sea
hydrography was the driving force for abundance variations in calcareous nannoplankton
during the last glacial.
One major objective of the third study undertaken on sediment core GeoB 7805-1 from
the Shaban Deep was to test if the calcareous nannoplankton is also a useful indicator for the
late Holocene climate instabilities. The results are presented in the third manuscript: Late
Holocene environmental changes in the northern Red Sea region as indicated by
nannoplankton abundance patterns. Compared to the last glacial–interglacial cycle, climate
fluctuations during the last two millennia are less pronounced. The Shaban Deep is a high

Chapter 1: Introduction
9
saline, anoxic submarine basin located in the northern Red Sea (Pätzold et al., 2003). The
extreme conditions in the Shaban Deep (e.g., Hartmann et al., 1998) prevent (multi-cellular)
benthic live. The resulting lack of bioturbation allows the sampling and examination of
undisturbed sediments with high resolution. Results presented here suggest that the
Gephyrocapsa spp. serve as a sensitive indicator for the northern Red Sea winter environment
in late Holocene times.
Following this outline, the thesis is organized as follows: First of all we take a closer look
at the paleoceanography of the northern Red Sea over the last 22,000 years (Chapter 2). The
next section deals with the calcareous nannoplankton succession during the late glacial
(Chapter 3). Following the study from the Shaban Deep (Chapter 4), the thesis concludes with
a summary (Chapter 5) and an outlook (Chapter 6) of possibly future research.
Explanatory notes
The author of the thesis has performed the micropaleontological research and has written the
presented manuscripts. The three manuscripts are either published (Climatic changes in the
northern Red Sea during the last 22,000 years as recorded by calcareous nannofossils by
H.L. Legge, J. Mutterlose and H.W. Arz; Paleoceanography, 2006, vol. 21, PA1003 - Chapter
2), submitted (Nannoplankton successions in the northern Red Sea during the last glaciation
(60 to 14.5 ka): Reactions to climate change by H.L. Legge, J. Mutterlose, H.W. Arz and J.
Pätzold; submitted for publication to EPSL - Chapter 3) or ready to submit (Late Holocene
environmental changes in the northern Red Sea region as indicated by nannoplankton
abundance patterns by H.L. Legge and J. Mutterlose - Chapter 4) for publication to an
international journal. The University of Bremen (Dr. Jürgen Pätzold) and the GFZ-Potsdam
(Dr. Helge W. Arz) provided the depth-age models and the complementary geochemical
information for the studied sediment cores. Details are given in the respective manuscripts.
The original manuscripts have been converted to achieve a uniform format of the thesis and
formal errors have been corrected. Sample material and prepared nannoplankton slides are
housed at the Ruhr-Universität Bochum.

Chapter 1: Introduction
10
References
Almond, D. C. (1986), Geological evolution of the Afro-Arabian dome, Tectonophysics, 131,
301–332.
Almogi-Labin, A. (1982), Stratigraphic and Paleoceanographic significance of Late
Quaternary pteropods from deep-sea cores in the Gulf of Aqaba (Elat) and northernmost
Red Sea, Mar. Micropaleontol., 7, 53–72.
Almogi-Labin, A., C. Hemleben, D. Meischner, and H. Erlenkeuser (1991),
Paleoenvironmental events during the last 13,000 years in the central Red Sea as
recorded by Pteropoda, Paleoceanography 6 (1), 83–98.
Almogi-Labin, A., C. Hemleben, and D. Meischner (1998), Carbonate preservation and
climatic changes in the central Red Sea during the last 380 kyr as recorded by
pteropods, Mar. Micropaleontol., 33(1-2), 87–107.
Andruleit, H. (1996), A filtration technique for quantitative studies of coccoliths,
Micropaleontol., 42, 403–406.
Arz, H. W., J. Pätzold, P. J. Müller, and M. O. Moammar (2003), Influence of Northern
Hemisphere climate and global sea level rise on the restricted Red Sea marine
environment during termination I, Paleoceanography, 18(2), 1053, doi:10.1029/
2002PA000864.
Arz, H. W., F., Lamy, A. Ganopolski, N. Nowaczyk, and P. Pätzold (2007), Dominant
Northern Hemisphere climate control over millennial-scale glacial sea-level variability,
Quat. Sci. Rev., 26, 312–321.
Beaufort, L., Y. Lancelot, P. Camberlin, O. Cayre, E. Vincent, F, Bassignot, and L. Labeyerie
(1997), Insolation cycles as a major control of equatorial Indian Ocean primary
production, Science, 278, 1451–1454.
Ben-David-Zaslow, R., G. Henning, D. K. Hofmann, and Y. Benayahu (1999), Reproduction
in the Red Sea soft coral Heteroxenia fuscescens: seasonality and long-term record
(1991 to 1997), Mar. Biol., 133, 553–559.
Berggren, W. A., and A. Boersma (1969), Late Pleistocene and Holocene planktonic
foraminifera from the Red Sea, in Hot Brines and Recent Heavy Metal Deposits in the
Red Sea, edited by E. T. Degens and D. A. Ross, pp. 282–298, Springer, New York.
Billard C., and I. Inouye (2004), What is new in coccolithophore biology?, in
Coccolithophores: From Molecular Processes to Global Impact, edited by H. Thierstein
and J. R. Young, pp. 1–30, Springer, New York.

Chapter 1: Introduction
11
Bosworth, W., P. Huchon, and K. McClay (2005), The Red Sea and Gulf of Aden Basins, J.
Afr. Earth Sci., 43, 334–378.
Bown, P. R., D. C. Rutledge, J. A. Crux, and L. T. Gallagher (1998), Lower Cretaceous, in
Calcareous nannofossil biostratigraphy, edited by P. R. Bown, pp. 86–102, Chapman
and Hall, London.
Brand, L. E. (1994), Physiological ecology of marine coccolithophores, in Coccolithophores,
edited by A. Winter and W. G. Siesser, pp. 39–49, Cambridge Univ. Press, Cambridge.
Cember, R. P. (1988), On the sources, formation, and circulation of Red Sea Deep Water, J.
Geophys. Res., 93, 8175–8191.
Chen, C. (1969), Pteropods in the Hot Brines Sediments of the Red Sea, in Hot Brines and
Recent Heavy Metal Deposits in the Red Sea, edited by E. T. Degens and D. A. Ross,
pp. 313–316, Springer, New York.
de Garidel-Thoron, T., L. Beaufort, B. K. Linsley, and S. Dannenmann (2001), Millennial-
scale dynamics of the East Asian winter monsoon during the last 200,000 years,
Paleoceanography, 16, 491–502.
Edwards, F. J. (1987), Climate and Oceanography, in Red Sea (Key Environments; 7), edited
by F. J. Edwards and S. M. Head, pp. 45–70, Pergamon, Oxford.
Eshel, G., and N. H. Naik, (1997), Climatological coastal jet collision, intermediate water
formation, and the general circulation of the Red Sea, J. Phys. Oceanogr., 27, 1233–
1257.
Eshel, G., D. P. Schrag, and B. F. Farrell (2000), Troposphere-planetary boundary layer
interactions and the evolution of the ocean surface density: Lessons from Red Sea
corals, J. Climate, 13, 339–351.
Fenton, M., S. Geiselhart, E. J. Rohling, and C. Hemleben (2000), Aplanktonic zones in the
Red Sea. Mar. Micropaleontol., 40, 277–294.
Geiselhart, S. (1998), Late Quaternary paleoceanographic and paleoclimatologic history of
the Red Sea during the last 380.000 years: Evidence from stable isotopes and faunal
assemblages, Tüb. Mikropaläontol. Mitt., 17, 87 pp.
Grasshoff, K. (1969), Zur Chemie des Roten Meeres und des inneren Golfs von Aden nach
Beobachtungen von F. S. Meteor während der Indischen Ozean Expedition 1964/65,
Meteor Forschungs-Ergebnisse, A6, 1–76.
Green, J. C., and R. W. Jordan (1994), Systematic history and taxonomy, in The Haptophyte
Algae, edited by J. C. Green and B. S. C. Leadbeater, pp. 1–22, Oxford Univ. Press,
New York.

Chapter 1: Introduction
12
Hartmann, M., J. C. Scholten, P. Stoffers, and F. Wehner (1998), Hydrographic structure of
brine-filled deeps in the Red Sea - new results from the Shaban, Kebrit, Atlantis II, and
Discovery Deep, Mar. Geol., 144, 311–330.
Hay, W. W., and H. P. Mohler (1967), Calcareous nannoplankton from early Tertiary rocks at
Pont Labau, France, and Palaeocene-Eocene correlations, J. Paleontol., 41, 1505–1541.
Hemleben, C., D. Meischner, R. Zahn, A. Almogi-Labin, H. Erlenkeuser, and B. Hiller
(1996), Three hundred eighty thousand year long stabile isotope and faunal records
from the Red Sea: influence of global sea level change on hydrography,
Paleoceanography, 11, 147–156.
Hoelzmann, P., D. Jolly, S. P. Harrison, F. Laarif, R. Bonnefille, and H. J. Pachur (1998),
Mid-Holocene land-surface conditions in northern Africa and the Arabian peninsula: A
data set for the analysis of biogeophysical feedbacks in the climate system, Glob.
Biogeochem. Cycl., 12(1), 35–51.
Honjo, S. (1977), Biogeography and provincialism of living coccolithophorids in the Pacific
Ocean, in Oceanic Micropalaeontology, Vol. 2, edited by A. T. S. Ramsay, pp. 951–
972, Academic Press, New York.
Jordan, R. W., and A. Kleijne (1994), A classification system for living coccolithophores, in
Coccolithophores, edited by A. Winter and W. G. Siesser, pp. 83–105, Cambridge
Univ. Press, Cambridge.
Kleijne, A., D. Kroon, and W. Zevenboom (1989), Phytoplankton and foraminiferal
frequencies in northern Indian Ocean and Red Sea surface waters, Neth. J. Sea Res., 24,
531–539.
Levanon-Spanier, I., E. Padan, and Z. Reiss (1979), Primary production in a desert-enclosed
sea – the Gulf of Elat (Aqaba), Red Sea, Deep-Sea Res. I, 26, 673–685.
Li, H., M. J. W. Veldhuis, and A. F. Post (1998), Alkaline phosphatase activities among
planktonic communities in the northern Red Sea, Mar. Ecol. Prog. Ser., 173, 107–115.
Lindell, D., and A. F. Post (1995), Ultraphytoplankton succession is triggered by deep winter
mixing in the Gulf of Aqaba (Eilat), Red Sea, Limnol. Oceanogr. 40 (6), 1130–1141.
Locke, S., and R. C. Thunell (1988), Paleoceanographic record of the last glacial/interglacial
cycle in the Red Sea and Gulf of Aden, Palaeogeogr. Palaeoclimatol. Palaeoecol., 64,
163–187.
Manis, P. C. (1973), A filling box model of the deep circulation of the Red Sea, Mem. Soc. R.
Sci. Liege, IV, 153–166.

Chapter 1: Introduction
13
Martini, E. (1971), Standard Tertiary and Quaternary calcareous nannoplankton zonation, in
Proccedings of the 2nd
International Conference of Planktonic Microfossils Roma,
1970, Vol. 2, edited by A. Farinacci, pp. 739–785, Tecnoscienza.
McIntyre, A., and A. W. H. Bé (1967), Modern coccolithophores of the Atlantic Ocean - I.
Placolith and cyrtoliths, Deep-Sea Res., 14, 561–597.
McIntyre, A. (1969), The Coccolithophorida in Red Sea sediments, in Hot Brines and Recent
Heavy Metal Deposits in the Red Sea, edited by E. T. Degens and D. A. Ross, pp. 299–
305, Springer, New York.
Molfino, B., and A. McIntyre (1990), Precessional forcing of nutricline dynamics in the
equatorial Atlantic, Science, 249, 766–769.
Morcos, S. A. (1970), Physical and chemical oceanography of the Red Sea, Oceanogr. Mar.
Biol. Annu. Rev., 18, 73–202.
Müller, C. (1976), Nannoplankton-Gemeinschaften aus dem Jung-Quartär des Golfes von
Aden und des Roten Meeres, Geol. Jb., D 17, 33–77.
Okada, H., and S. Honjo (1973), The distribution of oceanic coccolithophorids in the Pacific,
Deep-Sea Res., 20, 355–374.
Okada, H., and S. Honjo (1975), Distribution of coccolithophores in marginal seas along the
western Pacific Ocean and in the Red Sea, Mar. Biol., 31, 271–285.
Okada, H., and A. McIntyre (1977), Modern coccolithophores in marginal seas along the
western Pacific Ocean and in the Red Sea, Micropaleontol., 23, 1–55.
Okada, H., and A. McIntyre (1979), Seasonal distribution of modern Coccolithophores in the
western North Atlantik Ocean, Mar. Biol., 54, 319–328.
Okada, H., and D. Bukry (1980), Supplementray modification and introduction of code
numbers to the low-latitude coccolith biostratigraphic zonation, Mar. Micropaleontol.,
5, 321–325.
Pätzold, J., and cruise participants (2000), Report and preliminary results of METEOR Cruise
M 44/3 Aquaba (Jordan) - Safaga (Egypt) - Dubá (Saudi Arabia) - Suez (Egypt) - Haifa
(Israel), 12.3.–26.3.–2.4.–4.4.1999, Berichte aus dem Fachbereich Geowissenschaften
der Universität Bremen, 149, 135 pp.
Pätzold, J., G. Bohrmann, and C. Hübscher (2003), Black Sea - Mediterranean - Red Sea,
Cruise No. 52, January 2–March 27, 2002, Meteor-Berichte, Universität Hamburg, 178
pp.

Chapter 1: Introduction
14
Perch-Nielsen, K. (1985), Mesozoic calcareous nannofossils, in Plankton Stratigraphy, edited
by H. M. Bolli, J. B. Saunders and K. Perch-Nielsen, pp. 329–427, Cambridge Univ.
Press, Cambridge.
Pienaar, R. N. (1994), Ultrastructure and calcification of coccolithophores, in
Coccolithophores, edited by A. Winter and W. G. Siesser, pp. 13–34, Cambridge Univ.
Press, Cambridge.
Post, A. F. (2005), Nutrient limitation of marine cyanobacteria (Molecular ecology of
nitrogen limitation in an oligotrophic sea), in Harmful Cyanobacteria, edited by J.
Huisman, H. C. P. Matthijs and P. M. Visser, pp. 87–108, Kluwer, Dordrecht.
Reid, F. M. H. (1980), Coccolithophorids of the North Pacific Central Gyre with notes on
their vertical and seasonal distribution, Micropaleonto., 26, 151–176.
Reiss, Z., B. Luz, A. Almogi-Labin, E. Halicz, A. Winter, M. Wolf, and D. A. Ross (1980),
Late Quaternary paleoceanography of the Gulf of Aqaba (Elat), Red Sea, Quat. Res., 14,
294–308.
Rimbu, N., T. Felis, G. Lohmann, and J. Pätzold (2006), Winter and summer climate patterns
in the European-Middle East during recent centuries as documented in a northern Red
Sea coral record, The Holocene, 16, 321–330.
Rohling, E. J., M. Fenton, F. J. Jorissen, P. Bertrand, G. Ganssen, and J. P. Caulet (1998),
Magnitudes of sea-level lowstands of the past 500,000 years, Nature, 394, 162–165.
Roth, P. H., and W. T. Coulbourn (1982), Floral and solution patterns of coccoliths in surface
sediments of the north Pacific, Mar. Micropaleontol., 7, 1–52.
Samtleben, C., and A. Schröder (1992), Living coccolithophore communities in the
Norwegian-Greenland Sea and their record in sediments, Mar. Micropaleontol., 19,
333–354.
Shaikh, E. A., J. C. Roff, and N. M. Dowidar (1986), Phytoplankton ecology and production
in the Red Sea off Jiddah, Saudi Arabia, Mar. Biol., 92, 405–416.
Siddall, M., E. J. Rohling, A. Almogi-Labin, C. Hemleben, D. Meischner, I. Schmelzer, and
D. A. Smeed (2003), Sea-level fluctuations during the last glacial cycle, Nature, 423,
853–858.
Sissingh, W. (1977), Biostratigraphy of Cretaceous nannoplankton, Geol. Mijnb., 56, 37–65.
Siddall, M., E. J. Rohling, A. Almogi-Labin, C. Hemleben, D. Meischner, I. Schmelzer, and
D. A. Smeed (2003), Sea-level fluctuations during the last glacial cycle, Nature, 423,
853–858.

Chapter 1: Introduction
15
Sofianos, S. S., W. E. Johns, and S. P. Murray (2002), Heat and freshwater budgets in the Red
Sea from direct observations at Bab el Mandeb, Deep-Sea Res II, 49, 1323–1340.
Thierstein, H. R. (1971), Tentative Lower Cretaceous calcareous nannoplankton zonation,
Eclogae Geol. Helv., 64, 458–488.
Thierstein, H. R. (1976), Mesozoic calcareous nannoplankton biostratigraphy of marine
sediments. Mar. Micropaleontol., 1, 325–362.
Thunell, R. C., S. M. Locke, and D. F. Williams (1988), Glacio-eustatic sea-level control on
Red Sea salinity, Nature, 334, 601–604.
Weikert, H. (1987), Plankton and the Pelagic Environment, in Red Sea (Key Environments;
7), edited by F. J. Edwards and S. M. Head, pp. 90–111, Pergamon Press, Oxford.
Winter, A. (1982a): Paleonvironmental interpretation of Quaternary coccolith assemblages
from the Gulf of Aqaba (Elat), Red Sea, Rev. Esp. Micropaleontol., XIV, 291–314.
Winter, A. (1982b): Post-depositional shape modification in Red Sea coccoliths,
Micropaleonto., 3, 319–323.
Winter, A., Z. Reiss, B. Luz (1979), Distribution of living Coccolithophore assemblages in
the Gulf of Elat (’Aqaba), Mar. Micropaleontol., 4, 197–223.
Winter, A., A. Almogi-Labin, Y. Erez, E. Halicz, B. Luz and Z. Reiss (1983), Salinity
tolerance or marine organisms deduced from Red Sea Quaternary record. Mar. Geol.,
53, 17–22.
Winter, A., R. W. Jordan and P. H. Roth (1994), Biogeography of living coccolithophores in
oceanic waters, in Coccolithophores, edited by A. Winter and W. G. Siesser, pp. 161–
177, Cambridge Univ. Press, Cambridge.
Woelk, S., and D. Quadfasel (1996), Renewal of deep-water in the Red Sea during 1982–
1987, J. Geophys. Res., 101 (C8), 18155–18165.
Wyrtki, K. (1971), Oceanographic Atlas of the International Indian Ocean Expedition,
National Science Foundation, Washington, D.C., 531 pp.
Young J. R., M. Geisen, L. Cros, A. Kleijne, I. Probert, C. Sprengel, and J. B. Ostergaard
(2003), A guide to extant coccolithophore taxonomy, J. Nannopl. Res. Spec. Iss., 1, 124
pp.
Ziveri, P., K.-H. Baumann, B. Böckel, J. Bollmann, and J. R. Young (2004), Biogeography of
selected Holocene coccoliths in the Atlantic Ocean, in Coccolithophores: From
Molecular Processes to Global Impact, edited by H. Thierstein and J. R. Young, pp.
403–428, Springer, New York.

16

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
17
2 Climatic changes in the northern Red Sea during the last 22,000 years as recorded by
calcareous nannofossils
Abstract
We present a high-resolution record of calcareous nannofossils from the northern Red Sea for
the last 22 kyr. Extreme conditions with enhanced salinities during the Last Glacial Maximum
are characterized by high values of Gephyrocapsa ericsonii. The dominance of Emiliania
huxleyi in Heinrich event 1 indicates a climatic cooling favoring the bloom of opportunistic
species. The calcareous nannofossils record a two-step onset of the postglacial humid climate,
punctuated by the Younger Dryas. Both steps show an early oligotrophic phase dominated by
Florisphaera profunda and Gladiolithus flabellatus and a subsequent fertile phase
characterized by E. huxleyi. The Younger Dryas is described by high values of Gephyrocapsa
oceanica, indicating an increased mixing of the water column. In the late Holocene, repetitive
increases in abundance of F. profunda and G. flabellatus reflect restricted oligotrophic
conditions, caused by the high aridity following the Holocene humid period.
Key words: Red Sea; palaeoceanography; calcareous nannofossils; LGM; H1; termination;
Holocene
2.1 Introduction
The Red Sea is a restricted basin, surrounded by deserts, with a seaway to the Indian Ocean
(Gulf of Aden) via the shallow (137 m) Strait of Bab-el-Mandeb (Figure 2.1). The climate is
arid, with low precipitation (10–200 mm yr-1
) and high rates of evaporation (2000 mm yr-1
).
No major drainage network supplies the Red Sea and the influence of temporary runoff is
negligible (Morcos, 1970; Grasshoff, 1975; Hoelzmann et al., 1998).
Northwesterly winds blow to the south during the entire year over the northern Red Sea
and over the southern Red Sea during summer. In winter, the influence of the Indian monsoon
system leads to a seasonal reversal of the wind direction over the southern and central parts of
the Red Sea where southeasterly winds result in a convergence zone at 20°–25°N. The
circulation patterns are antiestuarine and parallel the longitudinal axis of the Red Sea. Warm
(26°–30°C) normal saline (36–37‰) Gulf of Aden waters enter the Red Sea. As this water
moves northward, it becomes cooler (19°–26°C) and saltier (40–41‰), eventually sinking to

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
18
form southward flowing intermediate and deep water masses, which ventilate the deep Red
Sea (e.g., Grasshoff, 1969; Morcos, 1970; Wyrtki, 1971; Cember, 1988; Woelk and
Quadfasel, 1996).
Because of biological consumption in the southernmost Red Sea, the nutrient gradients
along the longitudinal axis are less pronounced than those for salinity and temperature
(Weikert, 1987). In general, a unimodal
annual cycle in primary production,
abundance and diversity characterizes
the plankton communities of the
northern and central Red Sea (euphotic
zone ! 100 m). Low primary produc-
tion and low standing crops of the
phytoplankton were observed in the
stratified waters in summer times. In
winter cooling-induced overturning of
the waters results in a short interval of
high productivity. The southern Red
Sea is eutrophic because of the inflow
of waters from the Gulf of Aden during
the entire year (for details, see, e.g.,
Levanon-Spanier et al. (1979), Dowidar
(1983), Shaikh et al. (1986), and
Weikert (1987) and references therein). Plankton, organic detritus and inorganic particles,
especially sediment uptake from the shallow bottom and the narrow shores near the Strait of
Bab-el-Mandeb (Weikert, 1987), increase turbidity thereby reducing the depth of the euphotic
zone (" 60 m).
The higher productivity in the southern Red Sea is also displayed by an upward shift of the
oxygen minimum zone (OMZ) and lower values of dissolved oxygen compared with the north
(Weikert, 1987). In addition, the increasing distance from the area of intermediate and deep
water formation and ventilation (open northern Red Sea and Gulf of Suez (e.g., Cember,
1988; Woelk and Quadfasel, 1996; Eshel and Naik, 1997)) is noteworthy. The surface waters
are well oxygenated in all parts of the Red Sea (around 4.8–4 ml O2/l) with regard to the
oxygen saturation values in warm and saline waters (Edwards, 1987). Oxygen consumption,
due to the organic decomposition, reduces the concentration rapidly below the photic zone
Figure 2.1. Map showing the Red Sea area and the
Gulf of Aden with a detail view of the core location.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
19
with a distinctive minimum located between 300 and 500 m water depth. Beneath this depth,
the oxygen concentration increases slowly toward 2–3 ml O2/l in the deep water. In the
northern Red Sea lowest values of around 1–1.75 ml O2/l are located at 400–500 m, whereas
the oxygen concentration in the southern Red Sea decreases up to 0.3–0.5 ml O2/l already
around 300–400 m (Edwards, 1987; Weikert, 1987).
Like other oligotrophic marine ecosystems, the northern Red Sea is dominated by small
phytoplankton (Winter et al., 1979; Lindell and Post, 1995). Picoplankton and calcareous
nannoplankton, the latter consists of coccolithophorids, nannoliths and calcareous
dinoflagellates, are among the most common primary producers (Levanon-Spanier et al.,
1979; Winter et al., 1979; Kleijne et al., 1989). Coccolithophorids are biflagellate algal
protists with mineralized low magnesium-calcite plates or coccoliths, which cover the cell
surface. They belong to the class Prymnesiophyceae (division Haptophyta) and inhabit the
photic zone (0–200 m) of recent oceans (Green and Jordan, 1994). Since the fertility of
coccolithophorids depends on autecological factors (nutrients, temperature, salinity)
coccoliths are often used for paleoceanographic and paleoclimatic reconstructions (e.g.,
McIntyre and Be´, 1967; Okada and McIntyre, 1977; Molfino and McIntyre, 1990; Beaufort
et al., 1997). In contrast to other microfossils (e.g., foraminifera and pteropoda), little is
known about the distribution patterns of calcareous nannofossils (= fossilized calcareous
nannoplankton) in sediments of the Red Sea (but see McIntyre (1969), Müller (1976), and
Winter (1982a, 1982b)).
The Red Sea environment was subjected to large hydrographic changes during the last
glacial interglacial cycles. During the Last Glacial Maximum (LGM), the global sea level
dropped nearly 120 m (Locke and Thunell, 1988), reducing the water depth at the Strait of
Bab-el-Mandeb (now around 137 m) to approximately 17 m. The salinity increased
dramatically because of the reduced water exchange between the Indian Ocean and the Red
Sea. Oxygen isotopes reveal increased values in comparison to the global record (Hemleben
et al., 1996). On the basis of oxygen isotope data (compared to the open ocean record (e.g.,
Thunell et al., 1988; Hemleben et al., 1996)) and microfossil assemblage compositions (e.g.,
upper salinity limit of planktic foraminifera (cf. Hemleben et al., 1989)) the salinity estimates
for the northern Red Sea range around 50‰ and more (Fenton et al., 2000, and references
therein). The hypersaline conditions reached the tolerance limit for various planktic
organisms. Responses to this development include the absence of planktic foraminifera
(‘‘aplanktic zone’’ (e.g., Winter et al., 1983; Almogi-Labin et al., 1991; Hemleben et al.,
1996)) and the occurrence of a monospecific pteropod population (Creseis acicula) in the

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
20
northern Red Sea (Chen, 1969; Almogi-Labin, 1982; Winter et al., 1983). Previous studies
based on calcareous nannofossils suggest a massive decrease or even their total absence
during the LGM in the Red Sea. McIntyre (1969) argued that high salinities rather than cold
conditions are responsible for an impoverished flora in the central Red Sea. According to
Winter et al. (1983) high salinities caused the absence of calcareous nannofossils in the
northern Red Sea region.
In this study, we present a high-resolution record of calcareous nannofossils from core
GeoB 5844-2 in the northern Red Sea (Figure 2.1). The objective of this study is to analyze
the climatic variations in the northern Red Sea region for the last 22,000 years. The
nannofossil record, including relative and absolute abundances, is presented in combination
with geochemical results published recently for the same core (Arz et al., 2003a, 2003b). The
geochemical data consist of total organic carbon and carbonate, the vertical gradients in stable
oxygen isotopes, alkenone paleothermometry data and reconstructed paleosalinities. Our
findings are discussed in the context of the climatic history of the Red Sea and surrounding
areas for the last 22,000 years.
We want to prove that calcareous nannofossils can record paleoceanographic changes of
the past on a highresolution scale. A second objective of this study is to correlate the marine
findings with the climatic variations described from the surrounding landmasses. We further
want to test whether there is any evidence for synchronous climatic changes in the high
latitudes of the Northern Hemisphere and the northern Red Sea.
2.2 Material and Methods
The investigated gravity core GeoB 5844-2 was taken in 1999 near the tip of the Sinai
Peninsula (Figure 2.1; 27°42´81"N, 34°40´90"E, water depth: 963 m) during R/V Meteor
cruise M44/3 (Pätzold et al., 2000). The stratigraphic framework for core GeoB 5844-2 is
based on 14 calibrated 14
C AMS (Accelerated Mass Spectrometer) datings (Arz et al., 2003a,
2003b). Sedimentation rates are on average 12 cm kyr-1
during the Late Glacial and decrease
to about 6 cm kyr-1
in the Holocene. In order to achieve an average resolution of about 300
yrs, the sampling interval for the nannofossil study was 2 cm in the upper 100 cm and 4 cm
from 102 to 182 cm depth.
Samples were investigated using a scanning electron microscope (SEM). In order to
control SEM preparation and to study general changes in the floral composition light
microscope study was also undertaken using simple smear slides and pipette strew slides (e.g.,
Bown and Young, 1998). The samples were examined with an OLPYMPUS BH-2

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
21
microscope using polarizing light at a magnification of x1250 and dark field at a
magnification of x150. The samples for the SEM examination were prepared using the
filtration technique of Andruleit (1996). The freeze-dried sediment was weighted on a high-
precision balance, wet separated with a rotary splitter (FRITSCH Laborette 27) and filtered
through polycarbonate filters (pore size 0.25 µm) by means of a vacuum pump. A wedge-
shaped piece was cut out of the dry filter, mounted on an aluminum stub and sputter coated
with gold. High-resolution images were taken from the tip to the margin of the filter wedge on
a SEM (LEO 1530 Gemini FESEM) and subsequently examined on a qualitative and
quantitative basis. At least 300 specimens per sample were identified following the taxonomic
descriptions of Kleijne (1993), Jordan and Kleijne (1994) and Young et al. (2003). The
absolute abundance of the calcareous nannofossils (number of nannofossils per gram dry
sediment) was calculated with the following equation (e.g., Andruleit, 1996):
AA = (F x C x S) / (A x W)
where AA is the absolute abundance (number g-1
), F is the total sediment coated filter area
(mm2), C is the number of counted nannofossils, S is the split factor, A is the investigated
filter area (mm2), and W is the weight of the dry sample (g).
In addition the accumulation rates of calcareous nannofossils were calculated in the
following way:
AR = AA x SR x DD
where AR is the accumulation rate of the nannofossils (number cm-2
kyr-1
), AA is the absolute
abundance (number g-1
), SR is the sedimentation rate (cm kyr-1
), and DD is the dry bulk
density (g cm-3
).
The age-depth model (Figure 2.2) and the geochemical data (Figures 2.2, 2.3, and 2.4) are
taken from Arz et al. (2003a, 2003b). The paleotemperature reconstruction is based on the
alkenone unsaturation index UK 37 as defined by Prahl et al. (1988). Paleosalinities were
estimated from the local Red Sea !18
Ow-salinity relationship using the paleotemperature
equation of Bemis et al. (1998), the alkenone temperatures, and the stable oxygen isotope
measurements of planktic foraminifera and pteropoda to calculate the !18
Ow. Sediment
material and SEM slides are housed at the Department of Geology, Mineralogy and
Geophysics, Ruhr University Bochum.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
22
2.3 Results
The absolute abundances of calcareous nannofossils show a clear pattern (Figure 2.3e), with
low numbers during the Glacial (mean: 2.8E+09 number g-1
) and high numbers during the
Glacial-Holocene transition (mean: 1.3E+10 number g-1
) and the Holocene (mean: 2.4E+10
number g-1
). Our interpretations of the environmental conditions over the last 22 kyr are based
on the following species, which represent over 85% of the total nannofossil composition
(Figure 2.4).
Emiliania huxleyi shows relative abundances between 1.4 and 53.1% (mean: 24.8%) and
is the dominant species of the upper photic community since the end of the Last Glacial
Maximum (LGM). The abundance of this species increases during the Heinrich event 1 (H1),
the Bølling-Allerød and the early Holocene to mid-Holocene interval.
Figure 2.2. (a) Age-depth relation, (b) content of total organic carbon (TOC), and (c) carbonate
content of the sediment core GeoB 5844-2 (note reversed axis for TOC). Shaded vertical bars show
the Red Sea sapropels RS1a and RS1b (Arz et al., 2003a).

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
23
Figure 2.3. Comparison of (a) the stable oxygen isotope record (Stuiver and Grootes, 2000), (b) the
methane concentration of the GISP2 ice core (Blunier and Brook, 2001), (c) the record of warm
water foraminifera from the Aegean Sea (Rohling et al., 2002), (d) the UK’37 temperatures (core
GeoB 5844-2), (e) the absolute abundances (AA) of the nannofossils and the accumulation rates
(AR) of the nannofossils (core GeoB 5844-2), and (f) the sedimentation rates of core GeoB 5844-2.
Major hydrological and climatic instabilities observed in the tropics and subtropics of Africa are
marked with HC1, HC2, and HC3 (Gasse, 2000).

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
24
Figure 2.4

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
25
Figure 2.4. Comparison of the relative abundances of the calcareous nannofossils and the
geochemical data (Arz el al., 2003a, 2003b) of core GeoB 5844-2: (a) relative abundance of
Umbilicosphaera spp. (left axis) and H. carteri (right axis), (b) relative abundance of G. ericsonii,
(c) UK’37 temperatures, (d) relative abundance of G. oceanica, (e) vertical gradients in !18O
("!18Op-b, difference between planktic and benthic records; "!18Op-p, difference between shallow
planktic and deep planktic records), (f) relative abundance of E. huxleyi, (g) reconstructed sea
surface salinities, and (h) relative abundance of the lower photic community (F. profunda and G.
flabellatus). Shaded vertical bars show the Bølling-Allerød, the Younger Dryas transition, and the
Holocene humid period. The qualitative distribution of diatoms in the sediments is sketched at the top of the diagram.
Gephyrocapsa ericsonii exhibits relative abundances from 2.1 to 43.0% (mean: 10.7%),
with highest values during the LGM. The values of Gephyrocapsa oceanica fluctuate between
0.0 and 9.6% (mean: 2.6%). A prominent maximum of this species occurs in the Younger
Dryas (YD) interval. The percentages of Florisphaera profunda and Gladiolithus flabellatus
show fluctuations from 16.0 to 55.3% (mean: 37.3%) and 4.0 to 24.7% (mean: 11.4%),
respectively. Both species show maximum values after the H1, in the earliest Holocene and in
the late Holocene.
In addition we included the following less common taxa in our interpretation (Figure
2.2). Umbilicosphaera spp. (U. sibogae and U. foliosa) are rare with values between 0.0 and
4.4% (mean: 0.7%) showing a significant increase in the Holocene. Helicosphaera carteri
fluctuates between 0.0 and 7.4% (mean: 0.8%), with a prominent maximum in the lowermost
part of the investigated interval.
The calcareous nannofossils from core GeoB 5844-2 show an overall good to moderate
preservation during the Glacial-Holocene transition and the Holocene. Very good
preservation was observed in the Bølling-Allerød interval. Slightly overgrown specimens
occur in most of the investigated samples but do not hamper the investigation. In general,
overgrowth becomes more dominant in carbonate rich intervals. Samples with a different or
unusual preservation are discussed below.
2.4 Discussion
Our detailed analysis begins around 22 ka within the latest Glacial, a period of high aridity in
the northern African region (Adamson et al., 1980; Gasse et al., 1990). In the landlocked Red
Sea, the latest Glacial coincides with extreme environmental conditions. A hypersaline and
hostile environment that culminated in the LGM is documented by the amplification of the
oxygen isotope values and the absence of planktic foraminifera (e.g., Locke and Thunell,

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
26
1988; Hemleben et al., 1996; Rohling et al., 1998; Fenton et al., 2000, and references therein).
Characteristic components of northern Red Sea sediments during the last Glacial are abundant
large central diatoms (Almogi-Labin, 1982; Geiselhart, 1998). In core GeoB 5844-2 such
diatoms are concentrated in the lower part of the investigated interval. Their content decreases
after the LGM becoming absent after the H1 (Figure 2.4). According to Almogi-Labin (1982),
the monospecific Creseis acicula pteropod assemblages and their association with rich diatom
floras in the northern Red Sea indicate hypersaline conditions and high productive waters.
Geiselhart (1998) argued that high !13
C values and mass occurrences of the diatom
Cosinodiscus oculus-iridis indicate high-fertility waters and so upwelling in the northern Red
Sea. A different ecological interpretation of this species was, however, proposed by Kemp et
al. (2000) on the basis of samples from the Gulf of California and the eastern Mediterranean.
They argue that the presence of some large Cosinodiscus diatoms, including C. oculus-iridis,
characterize a stable phytoplankton community under oligotrophic conditions. The occurrence
of Cosinodiscus oculus-iridis is thought to be associated with strongly stratified waters of the
summer to early fall, whereas their sedimentation takes place after the break down of the
stratification in late fall (‘‘fall dump’’; Kemp et al., 2000).
2.4.1 Last Glacial Maximum (LGM, circa 22–19 ka)
In the northern Red Sea the LGM is characterized by high relative abundances of G. ericsonii
along with F. profunda and G. flabellatus, lower photic zone species (Figures 2.4b and 2.4h).
Furthermore higher values of H. carteri are limited to this period (Figure 2.4a).
Emiliania huxleyi, G. ericsonii and G. oceanica are the most common species of the
upper photic community observed in core GeoB 5844-2. In general, calcareous
nannoplankton tend to be dominant under stable, oligotrophic conditions (K selection). Some
taxa, especially E. huxleyi, and to a lesser extent Gephyrocapsa spp., show, however, an
opportunistic character (r selection). In recent oceans, E. huxleyi shows the widest
distribution, while G. ericsonii and G. oceanica have limited ranges. Emiliania huxleyi is a
eurytopic species which tolerates a wider range of environmental conditions than most other
calcareous nannoplankton species. It has the largest temperature range (1°–30°C), grows
under varying nutrient levels (eutrophic to oligotrophic) and reproduces more rapidly
whenever favorable conditions prevail (e.g., Birkenes and Braarud, 1952; Berge, 1962;
McIntyre et al., 1970; Okada and Honjo, 1973; Brand, 1994). This does not imply that G.
oceanica or G. ericsonii are displaced by E. huxleyi in any situation. Generally G. oceanica is
thought to prefer warm surface waters (optimum: 26°–32°C) and fertile conditions; it

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
27
dominates upwelling areas and marginal seas (Okada and Honjo, 1975; Kleijne, 1993).
Observations of G. ericsonii suggest that this species also prefers productive waters but under
cooler conditions (optimum: 17°–25°C) (Roth and Coulbourn, 1982; Okada and Wells, 1997;
Rogalla, 2002). In the hypersaline Gulf of Aqaba, G. ericsonii is observed in high numbers
within the low diversity living plankton community during the short fertile winter period
(Winter et al., 1979). High relative abundances of G. ericsonii point to fertile conditions
during the LGM, which may characterize the ‘‘winter situation’’. The isolation from the
Indian Ocean and the hypersaline conditions point to an indigenous population adapted to
high salinities that evolved during the glacial in the Red Sea. Once that normal saline
conditions were reestablished in the Red Sea G. ericsonii became less abundant. It is,
however, still common in the hypersaline Gulf of Aqaba (Winter et al., 1979).
In contrast to the Gulf of Aqaba, where no strong vertical differentiation of the
nannoplankton community is developed, the Red Sea shows a clear zonation. This is verified
by the occurrence of F. profunda and G. flabellatus. The lack of a zonation in the Gulf of
Aqaba seems to be closely related to the specific conditions with a weaker stratification and a
more uniform water column (cf. Weikert, 1987; Grossart and Simon, 2002). Florisphaera
profunda and G. flabellatus exclusively inhabit the lower photic zone (between 100–200 m),
which is characterized by low light levels and cold temperatures but a high nutrient content
compared with the upper photic zone (Okada and Honjo, 1973; Young, 1994). Abundance
variations of this ‘‘lower photic community’’ are closely related to changes of the
hydrographic setting. In general, high relative abundances of these species indicate
oligotrophic conditions in the upper photic zone and intense stratification of the water
column. High surface productivity or a deep mixing of the water column inhibits their growth
(e.g., Young, 1994). F. profunda has been successfully used as a proxy for monitoring the
depth and the stability of the nutricline (Molfino and McIntyre, 1990) and variations in
paleoproductivity (Rostek et al., 1997; Beaufort et al., 2001; de Garidel-Thoron et al., 2001).
A strong negative correlation has been proposed between primary production and the relative
abundances of F. profunda (cf. Beaufort et al., 1997). High nutrient levels in the surface
waters favor especially the upper photic community. This scenario is displayed in the
abundance pattern by high values of the upper photic species in relation to F. profunda (high
primary production = low relative abundances of F. profunda). Reduction of nutrients in the
surface waters, on the other hand, inhibits the growth of the upper photic community. Under
theses conditions the ratio of F. profunda is relatively high, favored by the higher nutrient
content in the lower photic zone (low primary production = high relative abundances of F.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
28
profunda).
In stratified subtropical waters like the northern Red Sea, which are characterized by
oligotrophic conditions, periods of high primary productivity are closely associated with
seasonally increased wind stress and surface cooling that raises the stratification (e.g., Halim,
1984; Weikert, 1987). The breakdown recycles nutrients into the surface waters. This favors
blooms of species of the upper photic zone with a preference for fertile conditions (e.g.,
Levanon-Spanier et al., 1979). In the discussion we will use the term LPC (lower photic
community) for F. profunda and G. flabellatus, and the term UPC (upper photic community)
for all other nannofossils. We take into consideration that the presence of the LPC with F.
profunda and G. flabellatus in core GeoB 5844-2 is closely related to oligotrophic conditions
in the upper photic zone. This reflects a better stratification of the water column, which
characterizes the ‘‘summer situation’’ during the LGM.
High values of H. carteri during the LGM are a significant feature. Similar observations
are documented from the central Red Sea and the Gulf of Aqaba (McIntyre, 1969; Winter,
1982a). McIntyre (1969) argued that high numbers of H. carteri are a result of its high
preservation potential, on the other hand the appearance of solution-prone species during this
period are noted by the same author. The ecological interpretation of H. carteri is not
unambiguous. Some studies point to an affinity of this species to warmer conditions, whereas
other results emphasize their preference to waters of higher fertility (e.g., McIntyre and Be´,
1967; Roth and Berger, 1975; Brand, 1994; Andruleit and Rogalla, 2002; Rogalla, 2002;
Ziveri et al., 2004). We argue that higher values of H. carteri indicate a higher nutrient
availability during the LGM. This is in accordance with the findings of Seeberg-Elverfeldt et
al. (2004), who studied diatoms in laminated sediments of the Shaban Deep (northern Red
Sea). These authors speculate that a better mixing of the water column or a high input of
terrigenous material increased the productivity during the LGM.
The geochemical record indicates that the mixing of the water column and the
productivity increased only slightly compared with the recent conditions in the northern Red
Sea (Arz et al., 2003a). We argue that the nannofossil record displays pronounced seasonal
cycles for the LGM, with prevailing fertile conditions during the winter. In summer, the water
column was oligotrophic and better stratified. This interpretation explains the co-occurrence
of calcareous nannofossils with different ecological affinities and supports the conclusions
drawn from diatom assemblages (Seeberg-Elverfeldt et al., 2004).

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
29
2.4.2 Heinrich Event 1 (H1, circa 18–15.5 ka)
The time of massive iceberg discharges of ice-rafted debris into the glacial North Atlantic
named ‘‘Heinrich events’’ (e.g., Heinrich, 1988; Bond et al., 1992; Broecker et al., 1992)
correlates with paleoclimatic anomalies at various sites outside the North Atlantic. A decrease
of sea surface temperature has been recorded from the Mediterranean Sea and the Indian
Ocean suggesting a far-reaching impact of the Heinrich events (e.g., Geraga et al., 2000;
Cacho et al., 2001 Kudrass et al., 2001; Schulte and Müller, 2001). A clear shift in the
abundance toward an E. huxleyi–dominated community can be observed during the H1 (18 to
15.5 ka (Bard et al., 2000)) in the northern Red Sea. Highest values of E. huxleyi occur during
the maximum cooling of 16.4°C at around 16.5 ka (Figures 2.4f and 2.4c). The Sea Surface
Temperature (SST) fell below the temperature optimum or even below the temperature
tolerance of certain subtropical-tropical calcareous nannoplankton species (e.g., McIntyre et
al., 1970; Winter et al., 1994). The most likely explanation for the change in the nannofossil
composition is that the climate encouraged the bloom of opportunistic species (r-selection)
with an ability to grow in moderate to cool temperate waters. We suggest that these conditions
especially favored the dominance of E. huxleyi. During H1 the calcareous nannoplankton
community gained a composition corresponding to that of the temperate climate zone in
present oceans (e.g., McIntyre and Be´, 1967; Winter et al., 1994).
An alternative explanation is that relatively wetter conditions prevailed during the H1.
The Sea Surface Salinity (SSS) record of core GeoB 5844-2 shows a minor drop of the
paleosalinity with a peak close to the maximum cooling (Figure 2.4g). The interpretations of
the climatic conditions in the northern Red Sea region are, in particular, on a short-term
timescale not unequivocal (e.g., Street and Grove, 1979; Street-Perrott and Harrison, 1985;
Fenton et al., 2000; Frumkin et al., 2000; Bartov et al., 2002, 2003). Because of low-
resolution sampling and the dating uncertainties of available paleoclimatic records it is often
impossible to distinguish between the Glacial, the LGM and the H1 period. This hinders a
detailed comparison. Most records from the northern African region display dryer and colder
conditions during the Glacial (e.g., Gasse, 2000, and references therein). The Nile was a
highly seasonal river. The sediment composition of the Nile as well as pollen spectra indicate
more open vegetation, adapted to colder and dryer conditions in the areas of its headwaters
(Adamson et al., 1980). Furthermore, lake levels in northern and eastern Africa declined or
even dried out (Adamson et al., 1980; Gasse, 2000). The climatic constellation of the Near
East is still debated controversially (e.g., COHMAP Members, 1988; Stein et al., 1999;
Bartov et al., 2003; Prasad et al., 2004). Paleoclimatic records from Israel, including lake

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
30
level records and pollen spectra, hint toward wet regional climate with glacial periods
(COHMAP Members, 1988; Frumkin et al., 2000, and references therein). In general, high
lake levels of the paleo-Dead Sea (Lake Lisan) occur during periods of colder global climate,
low lake levels are associated with warmer periods. During the Heinrich events, however,
lake levels dropped (Bartov et al., 2003; Prasad et al., 2004). Bartov et al. (2003) showed a
close connection between the Heinrich events and major lake level drops of the Lake Lisan. It
has been proposed that the cessation of the moisture supply, which caused the precipitation
over the eastern Mediterranean region and the related high lake levels, was the result of colder
conditions in the Mediterranean during the Heinrich events. Prasad et al. (2004) demonstrated
a close relationship between the deposition of Lake Lisan varved sediments, eastern
Mediterranean aridity and high-latitude cooling during marine oxygen isotope stage 2. This
suggests a close coupling between the climate of the high latitudes of the Northern
Hemisphere and the eastern Mediterranean.
On the basis of the accordance of arid conditions in northern African (Adamson et al.,
1980; Gasse et al., 1990; Gasse, 2000) and similar observations published recently from the
Near East (Bartov et al., 2003; Prasad et al., 2004), we favor temperature for explaining the
shift in the nannofossil composition to E. huxleyi–dominated assemblages. A climatic change
toward colder conditions displayed by the drop of the SST favored E. huxleyi blooms in the
northern Red Sea. At the same time the temperature drop may have caused a decrease of the
precipitation in the eastern Mediterranean. This resulted in the lake level drop of Lake Lisan
(Bartov et al., 2003; Prasad et al., 2004). We infer from the changes seen, a strong coupling
between the northern Red Sea and the climate of the higher latitude of the north. Since the
exchange with the Indian Ocean was restricted by the low sea level, a direct influence through
the Strait of Bab-el-Mandeb in the south seems to be less important for the observed
conditions in the northern Red Sea during H1 (Arz et al., 2003a). The increase of E. huxleyi,
the lowering of the SST, the timing of the changes in relation to the North Atlantic records,
and their rapidity suggest an atmospheric transfer of the cooling signal. We argue that this
mechanism was responsible for the simultaneous onset of the climatic signals observed in
both regions.
2.4.3 ‘‘H1-Bølling Transition’’ (circa 15.5–14.6 ka)
A hard layer of lithified carbonates, which contains encrusted pteropod shells and terrigenous
components, marks the end of H1 (Arz et al., 2003a). This horizon is thought to have been
deposited under rising SST and lower SSS (Milliman et al., 1969). Increased abundances of

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
31
the LPC correspond to the change from the H1 to the Bølling (Figures 2.4h and 2.5) and
suggest a well-developed water stratification and a decreased primary production. This setting
was limited to an interval here named ‘‘H1-Bølling transition’’. This interval covers the end
of the H1 and the beginning of the Bølling-Allerød, it therefore corresponds to the end of the
last glaciation and the early phase of the global sea level rise (Fairbanks et al., 1992). We
assume that during the H1-Bølling transition the Red Sea was characterized by the continued
inflow of normal marine waters from the Indian Ocean and probably a weaker seasonal
contrast. This idea is supported by the strong dependence between changes of the salinity in
the Red Sea and the global sea level rise (e.g., Rohling et al., 1998; Siddall et al., 2003, and
references therein). A stronger stratification of the Red Sea can be expected for this period.
These physical conditions with high nutrient depletion in the upper water column favored the
LPC (e.g., Beaufort et al., 1997; de Garidel-Thoron et al., 2001). The slowly improving water
Figure 2.5. Relationship between the upper photic community (UPC) and the lower photic
community (LPC) and the trends in primary production as indicated by their abundance patterns
(based on Figure 2.4h; note reversed axis). LPC-I and LPC-II mark the peak levels in abundance of
the LPC. The horizontal dashed line displays the mean value of the last 2 kyr (assumed to
correspond approximately with the modern trend; see section 2.4.9). A conclusion focused on the
two-step (I and II) onset of the postglacial humid climate is shown at the top (based on the
nannofossil record, the geochemical data, and the paleoclimatic records from the surrounding
landmasses). Shaded vertical bars show the Bølling-Allerød, the Younger Dryas transition (YD-T;
for details, see section 2.4.6) and the Holocene humid period. Arrows indicate Red Sea sapropels
RS1a and RS1b.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
32
conditions postdating the H1 are reflected by a minor but continuous decrease of the SSS
(Figure 2.4g). The SST did not yet reach Bølling-Allerød levels, but increased after the H1
(Figure 2.4c). The "!18
Op–b gradients increased up to present values, documenting a strong
stratification of the water column (Figure 2.4e). The northern Red Sea region was already
dominated by arid climatic conditions during the H1-Bølling transition (cf. Gasse et al., 1990;
Gasse, 2000). In contrast to the previous periods (see sections 4.1 and 4.2) the calcareous
nannofossils are of oligotrophic affinities as displayed by the high values of the LPC. We
propose that the increased inflow of normal saline waters into the Red Sea and a weaker
seasonality following the H1 cooling are a possible explanation.
2.4.4 Bølling-Allerød (circa 14.6–13.1 ka)
A change in the composition of the nannofossil assemblage takes place around 14.5 ka. A
sharp decrease of the LPC is followed by an interval with high values of the UPC dominated
by E. huxleyi (Figures 2.4f, 2.4h, and 2.5). This is paralleled by an increase of absolute
abundances during this interval (Figure 2.3e). Both, the onset of an E. huxleyi–dominated
community and the increase of total abundance of calcareous nannofossils correspond with
the enhanced content of atmospheric methane (circa 14.7 ka) recorded in the Greenland ice
cores and the onset of warmer conditions (circa 14.5 ka) (e.g., Dansgaard et al., 1993; Blunier
et al., 1995; Brook et al., 2000; Stuiver and Grootes, 2000; Blunier and Brook, 2001, and
references therein). According to Arz et al. (2003a), the !18
O gradient increased up to 2.5‰
and indicates, along with the SSS record, a stratified water column and a freshening of the
surface waters in the northern Red Sea (Figures 2.4e and 2.4g).
The nannofossil composition and the geochemical data of the Bølling-Allerød period
indicate stable water stratification. In contrast to the H1-Bølling transition the stable
stratification goes along with a higher productivity of the surface water as recorded by the
increase of the UPC (Figure 2.5). Because of the global sea level rise, Gulf of Aden waters
flow into the Red Sea and caused a density stratification of the water column. An increased
freshwater input from large desert wadis and a more humid climate due to the intensification
of the southwest monsoon further enhanced the development of the pycnocline in the Red Sea
(Locke and Thunell, 1988). The combination of a stable density stratification and higher
phytoplankton production went along with the deposition of ‘‘Red Sea sapropel 1a’’ (RS1a
(Arz et al., 2003a)) with maximum TOC values of 2.9 wt% (Figure 2.2b). The deposition of
TOC-rich sediments cannot directly be linked to an increase in productivity. Changes in the
deep water formation and ventilation (see section 2.1) are a second factor controlling the

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
33
depositional environment in the Red Sea (cf. Almogi-Labin et al., 1991; Arz et al., 2003a).
The increase of the UPC reflects a rise in surface water productivity, which coincides with the
deposition of RS1a. Stratification and fertilization seem to be linked to one another during the
Bølling-Allerød, probably due to humid climatic conditions. The increase in primary
production, in combination with the stratified water column, results in higher oxygen
consumption and additionally a pronounced decrease of the oxygen content. The dysoxic
conditions are maintained and further strengthened by reduced ventilation of the deeper
waters.
This relationship is supported by abundance variations between epipelagic pteropods
(restricted to the upper 100 m) and mesopelagic pteropods (diel migratory) in the Red Sea
(Almogi-Labin et al., 1998; Schmelzer, 1999). In general, the dominance of epipelagic
pteropods indicates a highly stratified water column (Almogi-Labin et al., 1991) and probably
an enhanced primary production, associated with a decrease of dissolved oxygen in the
intermediate water (reinforced and vertically extended OMZ (see section 1)). Under these
conditions the epipelagic species are favored by a good food supply. Simultaneously, the
strong stratification and the low contents of dissolved oxygen reduce the mesopelagic
pteropods (Almogi-Labin, 1982; Almogi-Labin et al., 1991, 1998). In the central Red Sea an
abundance maximum of epipelagic pteropods can be correlated with the deposition of TOC-
rich sediments (up to 1.65 wt%) during the Bølling-Allerød. Coherence with the prevailing
humid climate due to the reinforced southwest monsoon is found (Almogi-Labin et al., 1998).
A similar abundance maximum of epipelagic pteropods seems to be evident in the northern
Red Sea (cf. Schmelzer, 1999).
Humid conditions during the last 22 kyr are mainly associated with the ‘‘greening’’ of the
Sahara during the early Holocene to mid-Holocene. Various paleoclimatic records show,
however, that the onset of a more humid climate started earlier (e.g., Pachur and Altmann,
1997; deMenocal et al., 2000, and references therein). The onset of humid conditions in
subtropical northern Africa has been linked to the insolation (increased Northern Hemisphere
summer season insolation) forced intensification of the African Monsoon, associated with the
northward migration of the Intertropical Convergence Zone (ITCZ). Furthermore, the
comparison of the developing of the insolation (during the last 22 kyr, characterized by a
gradual increase toward a peak around 11 ka and a subsequent gradual decrease (cf. Berger,
1978; Berger and Loutre, 1991)) with paleoclimatic records indicate that the insolation alone
cannot explain the observed abrupt onset and termination as well as short-term variations of
the humid climate (e.g., COHMAP Members, 1988; Gasse and Van Campo, 1994; Kutzbach

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
34
et al., 1996; deMenocal et al., 2000). Various models try to explain the timing and the
intensity of the African Monsoon in response to different amplification mechanisms such as
changes in land surface conditions (e.g., vegetation cover, albedo) or the ocean surface
(deMenocal et al., 2000, and references therein). Apart from local differences the ‘‘African
humid period’’ covers the time interval from the onset of the Bølling-Allerød until the mid-
Holocene (e.g., 14.8 to 5.5 ka (deMenocal et al., 2000)). It is punctuated by an arid period
corresponding to the Younger Dryas. The two-step arid-humid transition is paralleled by
deglacial-warming events of the Greenland ice core, where the first step of the arid-humid
transition coincides with the onset of the Bølling-Allerød (Gasse et al., 1990).
Considering the geographic location of core GeoB 5844-2, which is beyond the direct
influences of the African Monsoon domain, observations from the Mediterranean borderlands
provide evidence for a regional increase in humidity. Records from the northern Sahara and
the eastern Mediterranean suggest high moisture levels expressed by the occurrence of
challenging plant species (warm and humid requirement), high water tables and an increase in
productivity probably provided by precipitation of Mediterranean origin (cf. Gasse et al.,
1990; Rossignol-Strick, 1995; Gasse, 2000, Frumkin et al., 2000). Therefore a Mediterranean
influence must be taken into account for the northern Red Sea at least additionally.
Considering the proximity of core GeoB 5844-2 to land, a nutrient supply caused by
increased precipitation and local wadi runoff may have affected the nannoplankton
community. We assume, that in addition to the former explanation, nutrient enrichment could
be already associated with an increased deepening of the surface mixed layer or enhanced
local upwelling. Apart from the overall seasonal trend in fertilization (see section 1), locally
restricted mixing processes are known to occur sporadically in the present northern Red Sea
during summer (Weikert, 1987). Such eutrophication could have reinforced during the
Bølling-Allerød period. This scenario may have resulted from a higher wind intensity
(consequence of the rapid warming, land–sea gradient) probably related to the air masses,
which caused the precipitation.
2.4.5 Younger Dryas (YD, 13.1 to 11.7 ka)
The transition to the Holocene is punctuated by the YD (Ruddiman and McIntyre, 1981;
Dansgaard et al., 1989). The YD is associated with an increase of aridity in subtropical Africa
(Gasse et al., 1990; Roberts et al., 1993; deMenocal et al., 2000) and in the Near East
(Frumkin et al., 2000). In the northern Red Sea the YD is displayed by a significant increase
of G. oceanica (Figure 2.4d). Within the E. huxleyi–Gephyrocapsa spp. group, a well-mixed

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
35
water column, that resulted in a higher nutrient content of the surface water, favored
Gephyrocapsa spp. and especially G. oceanica. This feature provides that temperature (see
section 4.4) was not a limiting factor (e.g., Winter et al., 1979; Brand, 1994; Okada and
Wells, 1997).
We argue that higher proportions of G. oceanica point to an increased mixing of the
water column during the YD. A decrease of the oxygen isotope ratios, which suggests a
deepening of the mixed layer, supports this assertion. Figure 2.4e shows the measured vertical
gradients in "!18
O. The reduction of the "!18
O values (especially "!18
Op–p) obviously
corresponds to the increase of G. oceanica. A shift in the nannoplankton community due to a
slight increase in SSS (Figure 2.4g) is rather unlikely since a preference to saltier waters of G.
oceanica was not observed during previous intervals of similar or even higher salinity (LGM,
H1). Having the warm water affinity of G. oceanica in mind, it is noteworthy that the SST
decreased, however, not reaching LGM or even H1 levels. Data from the alkenone
paleothermometry point to a maximum cooling of 23.7°C around 11.8 ka (Figure 2.4c). It is
necessary, however, to take into account that E. huxleyi already dominated the UPC and, that
the LPC provided an important component of the nannofossil assemblage (Figures 2.4f and
2.4h). The present hydrographic conditions of the northern Red Sea show higher annual
variability than other parts. Particularly during the short winter period the Red Sea is strongly
influenced by cold northerly winds (e.g., Morcos, 1970; Edwards, 1987). These conditions
cause a cooling-induced overturning of the waters (Edwards, 1987, Cember, 1988; Woelk and
Quadfasel, 1996) responsible for a fertilization of the photic zone (e.g., Levanon-Spanier et
al., 1979; Winter et al., 1979). It has been demonstrated that this break down of the
stratification had a lasting impact on the plankton communities (Levanon-Spanier et al., 1979;
Winter et al., 1979; Halim, 1984; Weikert, 1987). In the present Gulf of Aqaba the seasonal-
depth distribution of the nutrient concentration is the limiting factor in primary production
during the summer period. With the beginning of ‘‘winter’’ mixing nutrients are supplied to
the surface waters followed by phytoplankton blooms (Levanon-Spanier et al., 1979). Winter
et al. (1979) described important annual differences in the nannoplankton distribution in the
present Gulf of Aqaba and demonstrated that the appearanceof G. oceanica is mainly limited
to the winter period. According to Winter (1982a), a differentiation of the abundance pattern
between G. oceanica and E. huxleyi occurs with respect to the stratification, with G. oceanica
favoring better mixed waters.
Considering these observations, we assume that the increase of G. oceanica in core GeoB
5844-2 was associated with a reinforcement of the winter mixing during the YD. This view is

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
36
supported by diatom assemblage compositions studied by Seeberg-Elverfeldt et al. (2004).
They interpreted the shift from a Rhizosolenia-dominated assemblage to a Chaetoceros-
Bacteriastrum association as a change from the well-stratified waters of the Bølling-Allerød
period to the well-mixed surface waters of the YD. The Chaetoceros-Bacteriastrum
association of the YD displays a higher winter production as a result of an increased mixing
during this season (Seeberg-Elverfeldt et al., 2004).
The reinforcement of the winter mixing during the YD may also explain the already high
values of E. huxleyi alongside with the LPC. Subtropical conditions with a better stratification
of the waters already prevailed during the remaining year.
2.4.6 Younger Dryas Transition (circa 11.7–11.4 ka)
The end of the YD is marked by a decrease of G. oceanica (Figure 2.4d), a subsequent
slightly increase of E. huxleyi (Figure 2.4f) and the occurrence of Umbilicosphaera spp.
(mainly U. sibogae). In the geochemical results, a freshening peak centered around 11.5 ka
and an increase of the oxygen isotope ratios can be observed (Figures 2.4g and 2.4e). The
nannofossil composition and the geochemical results point to a better stratification of the
water column and a good availability of nutrients in the upper photic zone. Furthermore, the
presence of the thermophile U. sibogae, which was an inherent part of the nannofossil
associations in the Red Sea since the Younger Dryas transition (cf. McIntyre, 1969; Winter,
1982a), indicates warm surface waters (e.g., McIntyre and Be´, 1967; Okada and McIntyre,
1979; Roth, 1994). Within the dating uncertainty, we suggest that this short interval
corresponds to the abrupt warming of the ‘‘Holocene–Younger Dryas transition’’ (Dansgaard
et al., 1989; Taylor et al., 1997; Severinghaus et al., 1998). In northern Africa this interval
coincides with the early second step of the arid-humid transition (Gasse et al., 1990). High-
resolution terrestrial records, which cover this interval, are limited. Gasse et al. (1990)
described the onset of a short humid episode of less than 500 years in the Sahelian zone,
which corresponds to the Younger Dryas transition. Taking the scarce information into
account, we postulate a brief humid episode around 11.5 ka. This assumption is supported by
similar shifts of nannofossils and geochemical data, both in the Younger Dryas transition
(11.7 to 11.4 ka) and the Bølling-Allerød (14.5 to 13.1 ka). Higher values of E. huxleyi
(Figure 2.4f), accompanied by an increase of the oxygen isotope gradients (Figure 2.4e) and a
freshening of the surface waters (Figure 2.4g), can be observed during both periods.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
37
2.4.7 Earliest Holocene Period (circa 11.4–9.5 ka)
An increase of abundance of the LPC ended the short E. huxleyi–dominated interval of the
Younger Dryas transition (Figure 2.4). The following earliest Holocene period shows a
remarkable coincidence with those pattern already observed for the transition from the H1 to
the Bølling-Allerød (see section 4.3). Therefore we assume that comparable oceanographic
conditions prevailed during both periods (Figure 2.4h). In addition to Figure 2.4h, Figure 2.5
(based on Figure 2.4h; note reversed axis) shows the relationship between the UPC and the
LPC, supplemented by the conclusions drawn from their abundance patterns. Increased
relative abundances of the LPC can be observed since the mid-Holocene times toward the
present (= oligotrophic and poorly mixed environment of the present northern Red Sea (e.g.,
Weikert, 1987)). Despite this trend, highest relative abundances of the LPC occur during the
already discussed H1-Bølling transition (Figure 2.5, LPC-I) and the earliest Holocene period
(Figure 2.5, LPC-II). A clear dominance of the LPC with maximum values centered at around
10.4 ka indicates stratified and low productive waters.
The reduction of the SSS (Figure 2.4g) was driven by the inflow of normal saline water
from the Gulf of Aden. The constant increase of the SST (Figure 2.4c) and of the "!18
O
values (Figure 2.4e) as well as the lack of G. oceanica (Figure 2.4d) in significant numbers
further suggests that a reinforced overturning of the waters (e.g., winter mixing) can be
excluded (see section 4.5). Because of the inflow from the Gulf of Aden and the low
overturning, a strong stratification developed. Under these conditions the surface waters are
usually impoverished in nutrients and the primary production was low.
Near the peak level in abundance of the LPC (Figure 2.5, LPC-II), a short interval of
higher TOC values (1.1 wt%) named ‘‘Red Sea sapropel 1b’’ (RS1b (Arz et al., 2003a))
occurs. Apart from this excursion (centered around 10.6 ka), no significant fluctuations of the
TOC values were observed (Figure 2.2b). As discussed above (see section 4.4), a relationship
between sapropel formation and high primary production is not obligatory. The productivity
during the RS1b was already low, but the initial rise of the UPC shortly after the deposition
indicates a subsequent increase of fertility (Figure 2.5). We assume that the deposition of the
RS1b points to a short period of reduced ventilation, which marks the turning point in the
climatic evolution. The sapropel formation was probably induced by the climate change that
finally became manifested in the later Holocene humid period (see section 4.8). The
termination of the RS1b deposition and the absence of sapropel sediments during the humid
period is best explained by the increase of deep water formation after the flooding of the Gulf
of Suez around 11–10 ka (Arz et al., 2003a). The initiation of this important mode of deep

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
38
water formation (contribution of up to 50% to the present deep water (cf. Woelk and
Quadfasel, 1996)) probably allowed efficient ventilation independent of the surface water
conditions.
2.4.8 Early Holocene to Mid-Holocene Humid Period (circa 9.5–6 ka)
A shift in the nannofossil community, similar to that of the Bølling-Allerød (see section 4.4),
can be observed during the earliest Holocene period (see section 4.7). The decrease of the
LPC was followed again by an interval composed of E. huxleyi–dominated upper photic
assemblages that culminated between 9.5 and 6 ka (Figures 2.4h, 2.4f, and 2.5). We suggest
that the nannofossil composition of this interval reflects the ‘‘Red Sea Holocene humid
period’’ (Arz et al., 2003b). The paleosalinity record and the !18
O data indicate a reduced
salinity of the surface waters alongside with strong water stratification (Figures 2.4g and
2.4e). The vertical gradients of the oxygen isotopes increased up to 1.8‰ while the salinity
decreased to a minimum of 37‰. There are several lines of evidence suggesting that the
humidity was high during this period of the Holocene. The humid interval as indicated by the
nannofossils and geochemical data also appears in various records from Africa, Arabia and
the Mediterranean (e.g., Adamson et al., 1980; Rossignol-Strick, 1983; Ritchie et al., 1985;
COHMAP Members, 1988; Hoelzmann et al., 1998). The now extremely arid central and
eastern Sahara was dominated by savannah and steppe (Hoelzmann et al., 1998). The
extensive vegetation provided the food source for abundant large herbivorous mammals (e.g.,
antelopes, gazelles, giraffes, elephants). The areas covered by lakes and wetlands increased.
They were inhabited by crocodiles, hippos and numerous species of fishes (Pachur et al.,
1990). The high density of flora and fauna, as well as the availability of water, allowed human
settlement in what are now arid or even hyperarid regions (Gasse, 2000). Similar data were
obtained from the Arabian Peninsula, which was occupied by steppe (Hoelzmann et al.,
1998). In the eastern Mediterranean Sea, the Holocene humid period coincides with the
deposition of sapropel layer S1, which was linked to heavy freshwater input from the river
Nile (Rossignol-Strick et al., 1982; Rossignol-Strick, 1983), the Black Sea or more local
rivers (Lane-Serff et al., 1997). Rainfalls caused by the insolation forced African Monsoon
filled the headwaters of the Nile in Sudan and Ethiopia (Rossignol-Strick, 1983). The regional
humidity of the eastern Mediterranean, the northern Sahara and also the northern Red Sea,
however, is thought to have been driven by local precipitation. The moisture was provided by
a water source of Mediterranean origin (Gasse et al., 1990; Rohling and Hilgen, 1991; Arz et
al., 2003b). Paleoclimatic data (e.g., speleotherm records from Israel (Bar-Matthews et al.,

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
39
2000), Dead Sea level (Enzel et al., 2003) and snail oxygen isotopes from the Negev desert
(Goodfriend, 1988, 1990)) from the eastern Mediterranean region suggest that inflowing air
masses from the Mediterranean were the source of precipitation during the early Holocene to
mid-Holocene in the northern Red Sea (Arz et al., 2003b).
On the basis of the correlation between the well documented humid period, the !18
O
gradient, the salinity and the shift in the nannofossil assemblages, we argue that an increased
nutrient supply due to higher precipitation in the northern Red Sea region could have initiated
an increase in productivity. For the mid-Holocene slightly higher productivities, in
combination with an increase of humidity in the northern Red Sea region, are proposed
(Almogi-Labin, 1982; Winter, 1982a). Almogi-Labin (1982) assumed a strengthened
upwelling for the northernmost parts of the Gulf of Aqaba and the Red Sea during the mid-
Holocene. Winter (1982a) proposed that a high runoff in the mid-Holocene resulted in a high
nutrient supply for the Gulf of Aqaba. It is noteworthy that these data were obtained from a
core close to a major wadi fan (Wadi Dahab (cf. Winter, 1982a)). In the Holocene, however,
these runoffs must have been relatively low in the Gulf of Aqaba and high in the Red Sea.
During the LGM this trend was reversed (Winter, 1982a). This explanation is supported by
the general glacial/interglacial trends of the Lake Lisan and the paleoclimatic records from
subtropical Africa (see section 4.2). Both, the runoff from local drainage systems and the
precipitation increased (cf. Hoelzmann et al., 1998), supplying nutrients to the Red Sea. In
addition the inflow of the air masses from the Mediterranean may have enforced the mixing
of the surface waters or local upwelling, supporting a high productivity of the UPC (see
section 4.4).
An alternative explanation for the observed increase in primary production is the
increased inflow of Gulf of Aden waters. These added nutrients to the Red Sea (Weikert,
1987). The primary production in the present northern Red Sea is, however, not directly
related to the nutrient input from the Gulf of Aden. Because of the rapid biological
consumption the influence of the fertilization through the inflow is limited to the southern
Red Sea. It does not reach farther north than 19°–21°N. Differences in primary production,
plankton biomass and abundances are relatively small between the northern and central Red
Sea whereas both parts differ distinctively from the southern Red Sea (cf. Khmeleva, 1970;
Weikert, 1987).
The interpretations based on the calcareous nannofossils from GeoB 5844-2 are in
agreement with various terrestrial paleoclimatic records. The Red Sea record of a two-step
onset of the postglacial humid climate, separated by the YD, corresponds with numerous

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
40
paleohydrological studies (Gasse, 2000, and references therein). It is apparent, however, that
there are some anomalies in the nannofossils records. Bearing in mind the brief humid
interval of the Younger Dryas transition (11.7 to 11.4 ka), the Holocene humid period (9.5 to
6 ka) cannot be interpreted as a direct continuation of this interval because of its separation by
a more oligotrophic/arid period during the earliest Holocene period (11.7 to 9.5 ka).
Direct comparison between marine and terrestrial records on a short timescale is
problematic. Paleoclimatic signals achieved from land can be affected by the interplay of
climate, vegetation and water cycle (e.g., Gasse and Van Campo, 1994). Abrupt climatic
changes in the terrestrial records are often buffered and delayed by the response time of the
water cycle (Gasse and Van Campo, 1994). Evaporation of lakes, swamps and the vegetation
can result in self-stabilizing regional precipitation regimes that compensated periods of
droughts (Pachur and Altmann, 1997). Furthermore, signals of humid periods can be lost by
erosion (Hoelzmann et al., 1998).
The regional effects of the climatic reorientation during the early Holocene are more
complex, but already detectable by the calcareous nannofossils. We assume that the marine
record and the variations of the nannofossil community are a direct response to abrupt climate
changes.
2.4.9 Mid-Holocene to Late Arid Holocene (circa 6–0.6 ka)
The LPC shows a distinct increase in abundance after the humid period (Figure 2.5), with F.
profunda being the most common species in the late Holocene (Figure 2.4h). We argue that
the F. profunda–dominated assemblages in the northern Red Sea reflect geographically
restricted, oligotrophic conditions, caused by an increased aridity following the Holocene
humid period. Enhanced aridity is recorded in the eastern Sahara after 6 ka and in the whole
Sahara after 4.5 ka. Nowadays conditions were established around 2 ka (e.g., Maley, 1997;
Gasse, 2000).
Apart from the long-term trend, minor fluctuations in the nannofossil composition
occurred during the later part of the Holocene, which are still under investigation. Our current
data suggest four intervals of reduced nannofossil abundances, centered around 8.2, 6.4 and
4.0 and after 2.0 ka (Figure 2.3e). Though the number of samples is limited compared to the
frequency of the fluctuations, it seems that these intervals coincide with major cooling events
in the Aegean Sea (Figure 2.3c). These are thought to be related to variations of the
winter/spring intensity of the Siberian High with a periodicity around 2500 years (Figure
2.3c) (Rohling et al., 2002). Hydrological fluctuations, as compiled from various subtropical

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
41
African lake level records, are centered around 8.5–7.8, 7.0–6.6 and 4.5–3.5 ka (Figure 2.3)
(Gasse, 2000, and references therein). These seem to correspond to an increased aridity and a
stepwise transition to recent conditions in the northern Red Sea region. The 8.2 and the 4.0
events are the most prominent climate events. The 8.2 event can be correlated with a
distinctive !18
O shift and a decrease of the atmospherical methane as observed in the
Greenland ice core. The 4.0 ka event is thought to be related with the collapse of the Old
World societies (e.g., Gasse, 2000; deMenocal, 2001).
2.5 Conclusions
The calcareous nannofossil assemblages of the northern Red Sea mirror the climatic trends of
the last 22,000 years for this region and are closely coupled to the climatic changes of the
northern mid-latitudes and high latitudes. The changes in the nannofossil composition derived
from core GeoB 5844-2 indicate, that the calcareous nannofossil communities are not only
affected by variations of the salinity. Other factors, especially temperature and nutrients,
control the observed distribution patterns of nannoplankton in the northern Red Sea.
The nannofossil record allows differentiation of the Last Glacial Maximum (LGM) and
the Heinrich event 1 (H1) during the Glacial. Gephyrocapsa ericsonii and Helicosphaera
carteri characterize the LGM, whereas an increase of Emiliania huxleyi marks the H1.
Nannofossil assemblages with common G. ericsonii suggest pronounced seasonality during
the LGM. During the H1 the cooler climate encouraged the dominance of E. huxleyi. In the
Younger Dryas higher values of Gephyrocapsa oceanica are closely related to the intensified
‘‘winter’’ mixing of the water column.
The nannofossil assemblages of both the Bølling-Allerød transition and of the Holocene
show two subsequent, distinctive short-term phases. The early phase is dominated by the
lower photic community (LPC). This indicates a strong stratification of the water column and
oligotrophic conditions (H1-Bølling transition and earliest Holocene period). The later phase
shows a significant shift toward an E. huxleyi–dominated upper photic community that points
to more fertile conditions in the surface waters (Bølling-Allerød and Holocene humid period).
The late Holocene is characterized by a continuous increase of the LPC reflecting the
increase of aridity following the Holocene humid period. Apart from the long-term trend
fluctuations, the nannofossil record suggests a close link to minor climatic variations as
detectable during the Holocene.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
42
Acknowledgments
Sample material has been supplied by Jürgen Pätzold (Bremen). The work benefited from
discussions with Sylvia Rückheim and André Bornemann. We thank Rolf Neuser for
technical assistance at the SEM. This research was supported by the Deutsche
Forschungsgemeinschaft (Mu 667/23-1, -2). We thank David Watkins and an anonymous
reviewer for useful comments and improvements of the paper. Jeremy Young spent quite
some time on smoothing the English.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
43
References
Adamson, D. A., F. Gasse, F. A. Street, and M. A. J. Williams (1980), Late Quaternary
history of the Nile, Nature, 288, 50–55.
Almogi-Labin, A. (1982), Stratigraphic and Paleoceanographic significance of Late
Quaternary pteropods from deep-sea cores in the Gulf of Aqaba (Elat) and northernmost
Red Sea, Mar. Micropaleontol., 7, 53–72.
Almogi-Labin, A., C. Hemleben, and D. Meischner (1998), Carbonate preservation and
climatic changes in the central Red Sea during the last 380 kyr as recorded by
pteropods, Mar. Micropaleontol., 33(1-2), 87–107.
Almogi-Labin, A., C. Hemleben, D. Meischner, and H. Erlenkeuser (1991),
Paleoenvironmental events during the last 13,000 years in the central Red Sea as
recorded by Pteropoda, Paleoceanography 6 (1), 83–98.
Andruleit, H. (1996), A filtration technique for quantitative studies of coccoliths,
Micropaleontol., 42, 403–406.
Andruleit, H., and U. Rogalla (2002), Coccolithophores in surface sediments of the Arabian
Sea in relation to environmental gradients in surface waters, Mar. Geol., 186, 505–526.
Arz, H. W., J. Pätzold, P. J. Müller, and M. O. Moammar (2003a), Influence of Northern
Hemisphere climate and global sea level rise on the restricted Red Sea marine
environment during termination I, Paleoceanography, 18(2), 1053, doi:10.1029/
2002PA000864.
Arz, H. W., F. Lamy, J. Pätzold, P. J. Müller, and M. Prins (2003b), Mediterranean Moisture
Source for an Early-Holocene Humid Period in the Northern Red Sea, Science, 300,
118–121.
Bar-Matthews, M., A. Ayalon, and A. Kaufman (2000), Timing and hydrological conditions
of Sapropel events in the Eastern Mediterranean, as evident from speleothems, Soreq
Cave, Israel, Chem. Geol., 169, 145–156.
Bard, E., F. Rostek, J.-L. Turon, and S. Gendreau (2000), Hydrological impact of Heinrich
events in the subtropical Northeast Atlantic, Science, 289, 1321–1324.
Bartov Y., M. Stein, Y. Enzel, A. Agnon, and Z. Reches (2002), Lake levels and sequence
stratigraphy of Lake Lisan, the Late Pleistocene precursor of the Dead Sea, Quat. Res.,
57, 9–21.
Bartov, Y., A. Agnon, Y. Enzel, and M. Stein (2003), Catastrophic arid episodes in the
Eastern Mediterranean climate linked with the North Atlantic Heinrich events, Geology,
31, 439–442.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
44
Beaufort, L., Y. Lancelot, P. Camberlin, O. Cayre, E. Vincent, F. Bassignot, and L. Labeyerie
(1997), Insolation cycles as a major control of equatorial Indian Ocean primary
production, Science, 278, 1451–1454.
Beaufort, L., T. de Garidel-Thoron, A. C. Mix, and N. G. Pisias (2001), ENSO-like forcing on
the oceanic primary production during the late Pleistocene, Science, 293, 2440-2444.
Bemis, B. E., H. J. Spero, J. Bijma, and D. W. Lea (1998), Reevaluation of the oxygen
isotopic composition of planktonic foraminifera: Experimental results and revised
paleotemperature equations, Paleoceanography, 13, 150–160.
Berge, G. (1962), Discoloration of the sea due to Coccolithus huxleyi blomm, Sarsia, 6, 27–
40.
Berger, A. L. (1978), Long-term variations of caloric solar radiation resulting from the earth’s
orbital elements, Quat. Res., 9, 139–167.
Berger, A. L., and M. F. Loutre (1991), Insolation values for the climate of the last 10 million
years, Quat. Sci. Rev., 10, 297–317.
Birkenes, E., and T. Braarud (1952), Phytoplankton in the Oslo Fjord during “Coccolithus
huxleyi-summer”, Avh. Norske Vidensk Akad. Oslo I. Mat.-Nat. Kl., 2, 1–23.
Blunier, T., and E. J. Brook (2001), Timing of Millennial-Scale Climate Change in Antarctica
and Greenland during the Last Glacial Period, Science, 291, 109-112, doi:10.1126/
science.291.5501.109.
Blunier, T., J. A. Chappellaz, J. Schwander, B. Stauffer, and D. Raynaud (1995), Variations in
atmospheric methane concentration during the Holocene epoch, Nature, 374, 46–49.
Bond, G., et al. (1992), Evidence for massive discharges of icebergs into the North Atlantic
ocean during the last glacial period, Nature, 360, 245–249.
Bown, P. R., and J. R. Young (1998), Techniques, in Calcareous Nannofossil Biostrati-
graphy, edited by P. R. Bown, pp 16–28, Chapman and Hall, London.
Brand, L. E. (1994), Physiological ecology of marine coccolithophores, in Coccolithophores,
edited by A. Winter and W. G. Siesser, pp. 39–49, Cambridge Univ. Press, Cambridge.
Broecker, W., G. Bond, M. Klas, E. Clark, and J. McManus (1992), Origin of the northern
Atlantic's Heinrich events, Clim. Dyn., 6, 265–273.
Brook, E. J., S. Harder, J. Severinghaus, E. J. Steig, and C. M. Sucher (2000), On the origin
and timing of rapid changes in atmospheric methane during the last glacial period, Glob.
Biogeochem. Cycl., 14, 559–572.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
45
Cacho, I., J. O. Grimalt, M. Canals, L. Sbaffi, N. J. Shackelton, J. Schönfeld, and R. Zahn
(2001), Variability of the western Mediterranean Sea surface temperature during the last
25,000 years and its connection with the Northern Hemisphere climatic changes,
Paleoceanography, 16(1), 40–52.
Cember, R. P. (1988), On the sources, formation, and circulation of Red Sea Deep Water, J.
Geophys. Res., 93, 8175–8191.
Chen, C. (1969), Pteropods in the Hot Brines Sediments of the Red Sea, in Hot Brines and
Recent Heavy Metal Deposits in the Red Sea, edited by E. T. Degens and D. A. Ross,
pp. 313–316, Springer, New York.
COHMAP members (1988), Climatic changes of the last 18,000 years: observations and
model simulations, Science, 241, 1043–1052.
Dansgaard, W., J. W. C. White, and S. J. Johnson (1989), The abrupt termination of the
Younger Dryas climate event, Nature, 339, 532–534.
Dansgaard, W., S. J. Johnson, H. B. Clausen, D. Dahl-Jensen, N. S. Gundestrup, C. U.
Hammer, C. S. Hvidberg, J.P. Steffensen, A. E. Sveinbjörnsdottir, J. Jouzel, and G.
Bond (1993), Evidence for general instability of past climate from a 250-kyr ice-core
record, Nature, 364, 218–220.
de Garidel-Thoron, T., L. Beaufort, B. K. Linsley, and S. Dannenmann (2001), Millennial-
scale dynamics of the East Asian winter monsoon during the last 200,000 years,
Paleoceanography, 16, 491–502.
deMenocal, P. J. (2001), Cultural responses to climate change during the Late Holocene,
Science, 292, 667-673, doi:10.1126/science.1059827.
deMenocal, P., J. Ortiz, Z. Guilderson, J. Adkins, M. Sarnthein, L. Baker, and M. Yarusinsky
(2000), Abrupt onset and termination of the African Humid Period: rapid climate
reponses to a gradual insolation forcing, Quat. Sci. Rev., 19, 347–361.
Dowidar, N. M. (1983), Primary production in the Red Sea off Jeddah, Proc. int. Conf. mar.
Sci. Red Sea, Bull. Inst. Oceanogr. Fish., 9, 160–170.
Edwards, F. J. (1987), Climate and Oceanography, in Red Sea (Key Environments; 7), edited
by F. J. Edwards and S. M. Head, pp. 45–70, Pergamon, Oxford.
Enzel, Y., R. Bookman (Ken-Tor), D. Sharon, H. Gvirtzman, U. Dayan, B. Ziv, and M. Stein
(2003), Late Holocene climates of the Near East deduced from Dead Sea level
variations and modern regional winter rainfall, Quat. Res. 60, 263–273.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
46
Eshel, G. and N. H. Naik (1997), Climatological coastal jet collision, intermediate water
formation, and the general circulation of the Red Sea, J. Phys. Ocenogr., 24(7), 1233–
1257.
Fairbanks, R. G., C. D. Charles, and J. D. Wright (1992), Origin Of Global Meltwater Pulses,
in Radiocarbon after four decades, edited by R. E. Taylor, A. Long, and R. S. Kra, pp.
473–500, Springer, New York.
Fenton, M., S. Geiselhart, E. J. Rohling, and C. Hemleben (2000), Aplanktonic zones in the
Red Sea. Mar. Micropaleontol., 40, 277–294.
Frumkin, A., D. C. Ford, and H. P. Schwarcz (2000), Paleoclimate and vegetation of the last
glacial cycles in Jerusalem from a speleotherm record, Glob. Biogeochem. Cycl. 14,
863–870.
Gasse, F. (2000), Hydrological changes in the African tropics since the Last Glacial
Maximum, Quat. Sci. Rev., 19, 189–211.
Gasse F., and E. Van Campo, (1994), Abrupt post-glacial climate events in West Asia and
North Africa monsoon domains, Earth and Planet. Sci. Lett., 126, 435–456.
Gasse, F., R. Téhet, A. Durand, E. Gilbert, and J. C. Fontes (1990), The arid-humid transition
in the Sahara and Sahel during the last deglaciation, Nature, 346, 141–146.
Geiselhart, S. (1998), Late Quaternary paleoceanographic and paleoclimatologic history of
the Red Sea during the last 380.000 years: Evidence from stable isotopes and faunal
assemblages, Tüb. Mikropaläontol. Mitt., 17, 87 pp.
Geraga, M., S. Tsaila-Monopolis, C. Ioakim, G. Papatheodorou, and G. Ferentinos (2000),
Evaluation of palaeoenvironmental changes during the last 18,000 years in the Myrtoon
basin, SW Aegean Sea, Palaeogeogr. Palaeoclimatol. Palaeoecol., 156, 1–17.
Goodfriend G. A. (1988), Mid-Holocene rainfall in Negev Desert from 13
C of snail shell
organic matter, Nature, 333, 757–760.
Goodfriend, G. A. (1990), Rainfall in the Negev Desert during the Middle Holocene, based on
13C of organic matter in land snail shells, Quat. Res., 34, 186–197.
Grasshoff, K. (1969), Zur Chemie des Roten Meeres und des inneren Golfs von Aden nach
Beobachtungen von F. S. Meteor während der Indischen Ozean Expedition 1964/65,
Meteor Forschungs-Ergebnisse, A6, 1–76.
Grasshoff, K. (1975), The Hydrochemistry of Landlocked Basins and Fjords, in Chemical
Oceanography, vol. 2, edited by J. P. Riley and G. Skirrow, pp. 455–597, Academic
Press, New York.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
47
Green, J. C., and R. W. Jordan (1994), Systematic history and taxonomy, in The Haptophyte
Algae, edited by J. C. Green and B. S. C. Leadbeater, pp. 1–22, Oxford Univ. Press,
New York.
Grossart, H.-P., and M. Simon (2002), Bacterioplankton dynamics in the Gulf of Aqaba and
the northern Red Sea in early spring, Mar. Ecol. Prog. Ser., 239, 263–276.
Halim, Y. (1984), Plankton of the Red Sea and the Arabian Gulf, Deep-Sea Res., 31, 969–
982.
Heinrich, H. (1988), Origin & consequences of cyclic ice-rafting in the NE Atlantic Ocean,
Quat. Res., 29, 142–152.
Hemleben, C., M. Spindler, and O. R. Anderson (1989), Modern planktonic foraminifera,
Springer, New York, 363 pp.
Hemleben, C., D. Meischner, R. Zahn, A. Almogi-Labin, H. Erlenkeuser, and B. Hiller
(1996), Three hundred eighty thousand year long stabile isotope and faunal records
from the Red Sea: influence of global sea level change on hydrography,
Paleoceanography, 11, 147–156.
Hoelzmann, P., D. Jolly, S. P. Harrison, F. Laarif, R. Bonnefille, and H. J. Pachur (1998),
Mid-Holocene land-surface conditions in northern Africa and the Arabian peninsula: A
data set for the analysis of biogeophysical feedbacks in the climate system, Glob.
Biogeochem. Cycl., 12(1), 35–51.
Jordan, R. W., and A. Kleijne (1994), A classification system for living coccolithophores, in
Coccolithophores, edited by A. Winter and W. G. Siesser, pp. 83–105, Cambridge
Univ. Press, Cambridge.
Kemp, A. E. S., J. Pike, R. B. Pearce, and C. B. Lange (2000), The “Fall dump”- a new
perspective on the role of a “shade flora” in the annual cycle of diatom production and
export flux, Deep-Sea Res. II, 47, 2129–2154.
Khmeleva, N. N. (1970), On the primary production in the Red Sea and the Gulf of Aden,
Biol. morja, Kiew 21, 107–133.
Kleijne, A. (1993), Morphology, taxonomy and distribution of extant coccolithophorids
(calcareous nannoplankton), Ph.D. thesis, 320 pp., Vrije Universiteit, Amsterdam.
Kleijne, A., D. Kroon, and W. Zevenboom (1989), Phytoplankton and foraminiferal
frequencies in northern Indian Ocean and Red Sea surface waters, Neth. J. Sea Res., 24,
531–539.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
48
Kudrass, H. R., A. Hofmann, H. Doose, K. Emeis, and H. Erlenkeuser (2001), Modulation
and amplification of climatic changes in the Northern Hemisphere by the Indian
summer monsoon during the past 80 k.y., Geology, 29, 63–66.
Kutzbach, J. E., G. Bonan, J. Foley, and S. P. Harrison (1996), Vegetation and soil feedbacks
on the response of the African monsoon to orbital forcing in the early to middle
Holocene, Nature 384, 623–626.
Lane-Serff, G. F., E. J. Rohling, H. L. Bryden, and H. Charnock (1997), Postglacial
connection of the Black Sea to the Mediterranean and its relation to the timing of the
sapropel formation, Paleoceanography, 12, 169–174.
Levanon-Spanier, I., E. Padan, and Z. Reiss (1979), Primary production in a desert-enclosed
sea – the Gulf of Elat (Aqaba), Red Sea, Deep-Sea Res. I, 26, 673–685.
Lindell, D., and A. F. Post (1995), Ultraphytoplankton succession is triggered by deep winter
mixing in the Gulf of Aqaba (Eilat), Red Sea, Limnol. Oceanogr. 40 (6), 1130–1141.
Locke, S., and R. C. Thunell (1988), Paleoceanographic record of the last glacial/interglacial
cycle in the Red Sea and Gulf of Aden, Palaeogeogr. Palaeoclimatol. Palaeoecol., 64,
163–187.
Maley, J. (1997), Middle to Late Holocene changes in Tropical Africa and other continents:
palaeomonsoon and sea surface temperature variations, in Third Millennium BC Climate
Change and Old World Collapse, edited by H. Dalfes, G. Kukla and H. Weiss, pp 611–
640, NATO ASI Series, 149, Springer, New York.
McIntyre, A. (1969), The Coccolithophorida in Red Sea sediments, in Hot Brines and Recent
Heavy Metal Deposits in the Red Sea, edited by E. T. Degens and D. A. Ross, pp. 299–
305, Springer, New York.
McIntyre, A., and A. W. H. Bé (1967), Modern coccolithophores of the Atlantic Ocean-I.
Placolith and cyrtoliths, Deep-Sea Res. I, 14, 561–597.
McIntyre, A., A. W. H. Bé, and M. B. Roche (1970), Modern Pacific Coccolithophorida; a
paleontological thermometer, Trans. NY Acad. Sci., 32, 720–731.
Milliman J. D., D. A. Ross, and I. L. Ku (1969), Precipitation and lithification of deep-sea
carbonates in the Red Sea, J. Sediment. Petrol., 39, 724–736.
Molfino, B., and A. McIntyre (1990), Precessional forcing of nutricline dynamics in the
equatorial Atlantic, Science, 249, 766–769.
Morcos, S. A. (1970), Physical and chemical oceanography of the Red Sea, Oceanogr. Mar.
Biol. Annu. Rev., 18, 73–202.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
49
Müller, C. (1976), Nannoplankton-Gemeinschaften aus dem Jung-Quartär des Golfes von
Aden und des Roten Meeres, Geol. Jb., D 17, 33–77.
Okada, H., and S. Honjo (1973), The distribution of oceanic coccolithophorids in the Pacific,
Deep-Sea Res. I, 20, 355–374.
Okada, H., and S. Honjo (1975), Distribution of coccolithophores in marginal seas along the
western Pacific Ocean and in the Red Sea, Mar. Biol., 31, 271–285.
Okada, H., and A. McIntyre (1977), Modern coccolithophores of the Pacific and North
Atlantic oceans, Micropaleontol., 23, 1–54.
Okada, H., and A. McIntyre (1979), Seasonal distribution of modern Coccolithophores in the
western North Atlantik Ocean, Mar. Biol., 54, 319–328.
Okada, H., and P. Wells (1997), Late Quaternary nannofossil indicators of climate change in
two deep-sea cores associated with the Leeuwin Current off western Australia,
Palaeogeogr. Palaeoclimatol. Palaeoecol., 131, 413–432.
Pachur, H.-J., and N. Altmann (1997), The Quaternary (Holocene, ca. 8000 a B.P.), in
Palaeogeographic-Palaeotectonic Atlas of North-Eastern Africa, Arabia, and Adjacent
Areas, edited by H. Schandelmeier and P. O. Reynolds, pp. 111–125, Balkeema,
Rotterdam.
Pachur, H.-J., S. Kroepelin, P. Hoelzmann, M. Gaschin, and N. Altman (1990), Late
Quaternary fluvio-lacustrine environments of Western Nubia, Berl. Geowiss. Abh. A.,
120, 203–260.
Pätzold, J., and cruise participants (2000), Report and preliminary results of METEOR Cruise
M 44/3 Aquaba (Jordan) - Safaga (Egypt) - Dubá (Saudi Arabia) - Suez (Egypt) - Haifa
(Israel), 12.3.–26.3.–2.4.–4.4.1999, Berichte aus dem Fachbereich Geowissenschaften
der Universität Bremen, 149, 135 pp.
Prahl, F. G., L. A. Muehlhausen, and D. L. Zahnle (1988), Further evaluation of long-chain
alkenones as indicators of paleoceanographic conditions, Geochem. Cosmochim. Acta,
52, 2303–2310.
Prasad, S., H. Vos, J. F. W. Negendank, N. Waldmann, S. L. Goldstein, and M. Stein (2004),
Evidence from Lake Lisan of solar influence on decadal to centennial scale climate
variability during marine oxygen isotope stage 2, Geology, 32, 581–584.
Ritchie, J. C., C. H. Eyles, and C. V. Haynes (1985): Sediment and pollen evidence for an
early to mid-Holocene humid period in the eastern Sahara, Nature, 314, 352–355.
Roberts, N., M. Taieb, P. Barker, B. Damnati, M. Icole, and D. Williamson (1993), Timing of
the Younger Dryas event in East Africa from lake-level changes, Nature, 366, 146–148.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
50
Rogalla, U. (2002), Rekonstruktion von Paläoumweltbedingungen mit Coccolithophoriden
aus spätquartären Sedimenten des nördlichen Arabischen Meeres, Boch. Geow. Arb. 2,
130 pp.
Rohling, E. J., and F. J. Hilgen (1991), The eastern Mediterranean climate at times of sapropel
formation: a review, Geol. Mijnb., 70, 253–264.
Rohling, E. J., M. Fenton, F. J. Jorissen, P. Bertrand, G. Ganssen, and J. P. Caulet (1998),
Magnitudes of sea-level lowstands of the past 500,000 years, Nature, 394, 162–165.
Rohling, E. J., P. A. Mayewski, R. H. Abu-Zied, J. S. L. Casford, and A. Hayes (2002),
Holocene atmosphere-ocean interactions: records from Greenland and the Aegean Sea,
Climate Dyn., 18, doi:10.1007/s00382-001-0194-8.
Rossignol-Strick, M. (1995), Sea-land correlation of pollen records in the eastern
Mediterranean for the glacial-interglacial transition: Biostratigraphy versus radiometric
time-scale, Quat. Sci. Rev., 14, 893–915.
Rossignol-Strick, M. (1983), African monsoons, an immediate climate response to orbital
insolation, Nature, 304, 46–49.
Rossignol-Strick, M., W. D. Nesteroff, P. Olive, and C. Vergnaud-Grazzini (1982), After the
deluge: Mediterranean stagnation and sapropel formation, Nature, 295, 105–110.
Rostek, F., E. Bard, L. Beaufort, C. Sonzogni, and G. Ganssen (1997), Sea surface
temperature and productivity records for the last 240 kyr in the Arabian Sea, Deep-Sea
Res. II, 44, 1461–1480.
Roth, P. H. (1994), Distribution of coccoliths in oceanic sediments, in Coccolithophores,
edited by A. Winter and W. G. Siesser, pp. 199–217, Cambridge Univ. Press,
Cambridge.
Roth, P. H., and W. H. Berger (1975), Distribution and dissolution of coccoliths in the south
and central Pacific, Spec. Publ. Cushman Found. Foraminif. Res., 13, 87–113.
Roth P. H., and W. T. Coulbourn (1982), Floral and solution patterns of coccoliths in surface
sediments of the north Pacific, Mar. Micropaleontol., 7, 1–52.
Ruddimen, W. F., and A. McIntyre (1981), The North Atlantic Ocean during the last
deglaciation, Palaeogeogr. Palaeoclimatol. Palaeoecol., 35, 145–214.
Schmelzer, I. (1999), High-frequency event-stratigraphy and paleoceanography of the Red
Sea, 124, Ph.D. thesis, 124 pp., Eberhard Karls Universität, Tübingen.
Schulte, S., and P. J. Müller (2001), Variations of sea-surface temperature and primary
productivity during Heinrich and Dansgaard-Oeschger events in the northeastern
Arabian Sea, Geo-Mar. Lett., 168–175.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
51
Seeberg-Elverfeldt, I. A., C. B. Lange, H. W. Arz, J. Pätzold, and I. Pike (2004), The
significance of diatoms in the formation of laminated sediments of the Shaban Deep,
Northern Red Sea, Mar. Geol., 209, 279–301.
Severinghaus, J. P., T. Sowers, E. J. Brook, R. B. Alley, and M. L. Bender (1998), Timing of
the abrupt climate change at the end of the Younger Dryas interval from thermally
fractionated gases in polar ice, Nature, 391, 141–146.
Shaikh, E. A., J. C. Roff, and N. M. Dowidar (1986), Phytoplankton ecology and production
in the Red Sea off Jiddah, Saudi Arabia, Mar. Biol., 92, 405–416.
Siddall, M., E. J. Rohling, A. Almogi-Labin, C. Hemleben, D. Meischner, I. Schmelzer, and
D. A. Smeed (2003), Sea-level fluctuations during the last glacial cycle, Nature, 423,
853–858.
Stein, M., Y. Bartov, Y. Enzel, S. L. Goldstein, M. Machlus, R. Ken-Tor, and S. Marco
(1999), High-resolution chronology of Lake Lisan levels, EOS Transactions, AGU Fall
Meeting 1999 Supplement, F501, 80(46).
Street, F. A., and A. T. Grove (1979), Global maps of lake-level fluctuations since 30,000
years BO, Quat. Res., 12, 83–118.
Street-Perrott, F. A., and S. P. Harrison (1985), Lake levels and climatic reconstruction, in
Palaeoclimate Analysis and Modelling, edited by A. D. Hecht, pp. 291–331, John
Wiley, New York.
Stuiver, M., and P. M. Grootes (2000), GISP2 oxygen isotope ratios, Quat. Res., 53, 277–284.
Taylor, K. C., et al. (1997), The Holocene-Younger Dryas Transition Recorded at Summit,
Greenland, Science, 278, 825–827.
Thunell, R. C., S. M. Locke, and D. F. Williams (1988), Glacio-eustatic sea-level control on
Red Sea salinity, Nature, 334, 601–604.
Weikert, H. (1987), Plankton and the Pelagic Environment, in Red Sea (Key Environments;
7), edited by F. J. Edwards and S. M. Head, pp. 90–111, Pergamon Press, Oxford.
Winter, A. (1982a), Paleoenvironmental interpretation of Quaternary coccolith assemblages
from the Gulf of Aqaba (Elat), Red Sea, Rev. Esp. Micropaleontol., XIV, 291–314.
Winter, A. (1982b), Post-depositional shape modification in Red Sea coccoliths,
Micropaleontol., 3, 319–323.
Winter, A., Z. Reiss, and B. Luz (1979), Distribution of living Coccolithophore assemblages
in the Gulf of Elat (’Aqaba), Mar. Micropaleontol., 4, 197–223.

Chapter 2: Climatic changes in the Red Sea during the last 22,000 years…
52
Winter, A., A. Almogi-Labin, Y. Erez, E. Halicz, B. Luz, and Z. Reiss (1983), Salinity
tolerance or marine organisms deduced from Red Sea Quaternary record. Mar. Geol.,
53, 17–22.
Winter, A., R. W. Jordan and P. H. Roth (1994), Biogeography of living coccolithophores in
oceanic waters, Coccolithophores, in Coccolithophores, edited by A. Winter and W. G.
Siesser, pp. 161–177, Cambridge Univ. Press, Cambridge.
Woelk, S., and D. Quadfasel (1996), Renewal of deep-water in the Red Sea during 1982-
1987, J. Geophys. Res., 101 (C8), 18155–18165.
Wyrtki, K. (1971), Oceanographic Atlas of the International Indian Ocean Expedition,
National Science Foundation, Washington, D.C., 531 pp.
Young, J. R. (1994). Functions of coccoliths, in Coccolithophores, edited by A. Winter and
W. G. Siesser, pp. 63–82, Cambridge Univ. Press, Cambridge.
Young, J. R., M. Geisen, L. Cros, A. Kleijne, I. Probert, C. Sprengel, and J. B. Ostergaard,
(2003), A guide to extant coccolithophore taxonomy, J. Nannopl. Res. Spec. Iss., 1, 124
pp.
Ziveri, P., K.-H. Baumann, B. Böckel, J. Bollmann, and J. R. Young (2004), Biogeography of
selected Holocene coccoliths in the Atlantic Ocean in Coccolithophores: From
Molecular Processes to Global Impact, edited by H. Thierstein and J. R. Young, pp.
403–428, Springer, New York.

Chapter 3: Nannoplankton successions during the last glaciation…
53
3 Nannoplankton successions in the northern Red Sea during the last glaciation (60 to
14.5 ka): Reactions to climate change
Abstract
Due to its restricted connection with the Indian Ocean, the desert-enclosed Red Sea is
extremely sensitive to global sea level changes and thus ideally suited for paleoceanographic
studies of what occurred during the last glaciation. The understanding of its glacial history is,
however, still limited. A serious obstacle to obtain satisfactory paleoecological information
has been the rarity of microfossil proxy species caused by high salinities. Here, we present a
continuous and well-dated calcareous nannoplankton record from the northern Red Sea,
covering the interval from 60–14.5 ka. Our investigation shows that the composition of the
calcareous nannoplankton community varied between ca. 32 ka and 14.5 ka in response to
rapid environmental changes which are closely correlated to climatic fluctuations described
from the North Atlantic region. Heinrich events H3, H2 and H1 are dominated by Emiliania
huxleyi. Gephyrocapsa oceanica and especially Gephyrocapsa ericsonii are abundant
between H3–H2 and H2–H1. A less pronounced response of the calcareous nannoplankton to
the high latitudinal climatic oscillations is documented prior to 32 ka, suggesting that a strong
atmospheric coupling between the northern Red Sea and the North Atlantic realm was
established in the late Marine Isotope Stage 3. In contrast to the previously held view of a sea
level related salinity increase as the major cause for changes of the plankton communities
within the glacial Red Sea, we interpret the documented variations as being caused by local
hydrographic changes under the atmospheric control from the extratropics. Temperature
changes and especially variations of the water stratification appear to be critical selective
factors for the calcareous nannoplankton composition.
Keywords: Red Sea; calcareous nannoplankton; paleoclimate; glacial environment; Heinrich
events
3.1 Introduction
Continuous microfossil records from the northern Red Sea, which cover Marine Isotope
Stages (MIS) 3 and 2 (e.g., Imbrie et al., 1984), are sparse. As a result of the globally low sea
level during the last glacial (up to around 120–125 m), water exchange between the Red Sea

Chapter 3: Nannoplankton successions during the last glaciation…
54
and the Gulf of Aden (Indian Ocean) via the shallow (now around 137 m) and narrow (now
around 18 km) Strait of Bab-el-Mandeb was drastically reduced (Siddall et al., 2003; Arz et
al., 2007). Hypersaline conditions, amounting to 50‰ or even above, prevailed during MIS 3
and especially MIS 2 (Thunell et al., 1988; Hemleben et al., 1996; Arz et al., 2003). The
disappearance of various microfossil species prelude the “aplanktic zone”, a period
characterized by the absence of planktic foraminifera and by a pteropod community mainly
consisting of the euryhaline Creseis acicula (Almogi-Labin, 1982; Winter et al., 1983; Fenton
et al., 2000). Recent results indicate the presence of calcareous nannoplankton (simply named
nannoplankton in the ongoing text) during this hypersaline “aplanktic zone” in the northern
Red Sea (Legge et al., 2006).
Figure 3.1. Location of core GeoB 5844-2 in the northern Red Sea and a schematic view of the precipitation regimes at both margins of the subtropical desert belt (north: Mediterranean winter rain,
south: southwest summer monsoon). Dashed line shows position of the Intertropical Convergence Zone (ITCZ); thick black arrow indicates wintertime mean position of the Subtropical Jet (STJ) which roughly mark the southern limit of the extratropical Mediterranean winter climate features (Air Ministry, 1962; Blackmon et al., 1984; Martyn, 1992; Hoskins and Hodges, 2002). The desert belt is shaded in light gray (vegetation-based; Schmithüsen, 1976). A detail view of the northern Red Sea with the Gulf of Aqaba (right) and the Gulf of Suez (left) is shown in the upper right panel.
We present a nannoplankton record from the desert-enclosed northern Red Sea (Figure 3. 1)
covering the period between around 60 ka and the onset of the Bølling-Allerød warm period
at around 14.5 ka (Figure 3.2 and Figure 3.3). Short-term climatic variations, including
Dansgaard-Oeschger (D-O) cycles and Heinrich events, are well documented in the North
Atlantic region during the time period under consideration (Bond et al., 1993; Dansgaard et
al., 1993). D-O cycles, rapid and large amplitude changes observed in the !18O ice core
records from Greenland, reflect variations in air temperature over the northern ice sheets.
Several D-O cycles are grouped into multi-millennial intervals characterized by an overall
progressive cooling (longer-term cooling cycle). Massive iceberg discharges into the North
Atlantic are indicated by extensive layers of coarse-grained debris (Broecker et al., 1992).
These Heinrich events, or more precisely their climatic consequences are documented as

Chapter 3: Nannoplankton successions during the last glaciation…
55
pronounced cold periods that occurred at the end of the longer-term cooling cycles (Broecker
et al., 1992; Bond et al., 1993). D-O cycles, longer-term cooling cycles, and the climatic
consequences of Heinrich events (Figure 3.3) are not restricted to the North Atlantic region.
Various records in the mid-latitudes document similar variations and indicate a strong relation
to the climate of the North Atlantic realm (e.g., Voelker, 2002). Given the sensitivity of
nannoplankton to autoecological factors (e.g., Brand, 1994; Ziveri et al., 2004), our record
offers a unique possibility for the examination of millennial-scale environmental changes
during the hypersaline period of the Red Sea in response to the longer-term cooling cycles
and Heinrich events of the North Atlantic realm.
3.2 Material and Methods
The studied sediment core GeoB 5844-2 (Figure 3.1) was recovered in 1999 during Meteor
cruise 44/3 (Pätzold et al., 2000) in the vicinity of the Sinai Peninsula (27°42´81"N,
34°40´90"E, water depth: 963 m). For this study, we investigated fossil nannoplankton from
the time-interval 60–22 ka. To allow a more comprehensive view of the results, we present a
complete record of the 60–14.5 ka time period by including recently published data (Legge et
al., 2006). Sample preparation follows the filtration technique as described by Andruleit
(1996). At least 300 specimens of each sample were examined under scanning electron
microscope. We describe the abundance patterns of the three Noelaerhabdaceae Emiliania
huxleyi (Figure 3.3f), Gephyrocapsa ericsonii (Figure 3.3g) and Gephyrocapsa oceanica
(Figure 3.3h), which dominate the upper-photic
community of the Red Sea. In addition, we figure the
relative abundance of Florisphaera profunda (Figure
3.5d). We integrate F. profunda in our discussion
because this lower-photic species contributes
significantly to the fossil record. Based on AMS
(Accelerated Mass Spectrometer) 14C dating and
additional paleomagnetic dating points, the
stratigraphy framework of core GeoB 5844-2 (Figure
3.2) has been established by Arz et al. (2007). Ages
(on the SFCP timescale; Shackleton et al., 2004) are
announced in calibrated calendar years before
present.
Figure 3.2. Age depth relation of the sediment core GeoB 5844-2 (Arz et al. 2007). The core section/ time interval studied here is gray shaded.

Chapter 3: Nannoplankton successions during the last glaciation…
56
3.3 Results and Discussion
The varying composition of nannoplankton assemblages defines two intervals (60–32 ka and
32–14.5 ka). The most striking feature in the younger interval (32–14.5 ka) is the recurrent
appearance of E. huxleyi, G. ericsonii and G. oceanica. Those intervals, which correspond
stratigraphically to Heinrich events H3, H2 and H1 (Figure 3.3), are dominated by E. huxleyi
(Figure 3.3f). The longer-term cooling cycles, or more precisely the intervals between H3–H2
and H2–H1, are characterized by increasing values of G. ericsonii (Figure 3.3g). In addition,
brief increases of G. oceanica are displayed shortly after H3 and especially after H2 (Figure
3.3h). Fluctuations in the abundance of these three species are also documented in the older
part of the record (60–32 ka), there are, however, no analogue abundance patterns as in the
younger interval. A comparable distinct species succession of G. oceanica, G. ericsonii and E.
huxleyi (order in view of the North Atlantic climate/temperature trend) is virtually absent. In
the following discussion we will focus on two aspects: first, the recurrent sequence of G.
oceanica, G. ericsonii, and E. huxleyi (= Noelaerhabdaceae-succession) in the younger
interval, and second, the timing of the onset of the succession at around 32 ka.
Biogeographical distribution patterns emphasize the affinity of G. oceanica to the warm
waters of the tropical and subtropical latitudes (e.g., Brand, 1994; Ziveri et al., 2004).
Gephyrocapsa ericsonii flourishes under cooler conditions, while cold waters also inhibit its
growth (e.g., Gartner, 1988; Okada and Wells, 1997). Emiliania huxleyi tolerates a broad
temperature range as documented by its abundant occurrence from the tropics to the subarctic
(e.g., Gartner, 1988; Brand, 1994).
The alkenone-sea surface temperatures (SSTs) available from core GeoB 5844-2 (Arz et
al., 2007) show that the Heinrich events H3, H2 and H1 are correlated with pronounced
cooling events in the northern Red Sea (Figure 3.3e). A weaker response to the high-
latitudinal cooling is evident in the older part of the alkenone-record, which also corresponds
well to the nannoplankton record. Prior to around 32 ka, fluctuations were of lower intensity
and the SST always remained above 22.1°C. Due to the good correlation of our
nannoplankton findings with the SSTs, it seems reasonable to envisage a temperature-related
explanation for the Noelaerhabdaceae-succession.
The specific temperature preferences may explain the dominance of E. huxleyi during the
maximum cooling intervals (H3, H2, H1) but leaves us with one problem: It remains open
why this eurythermal species was outcompeted by Gephyrocapsa spp. in the transitional
intervals (H3–H2, H2–H1). Corroborated by plankton studies, we speculate on temperature-
driven variations of the water stratification as the key mechanism.

Chapter 3: Nannoplankton successions during the last glaciation…
57
Figure 3.3

Chapter 3: Nannoplankton successions during the last glaciation…
58
Figure 3.3. Comparison of (a) the SPECMAP !18O record (stacked; Imbrie et al., 1984) and (b) the !
18O GRIP ice-core record (on the SFCP timescale; Shackleton et al., 2004) with results from core GeoB 5844-2 (northern Red Sea): (c) the planktic !18O record (Arz et al., 2007), (d) the recon-structed sea level curve (two temperature end-member scenarios: with and without taking the full range of alkenone SSTs changes into account; for details please see Arz et al., 2007), (e) the alkenone-sea surface temperatures (Arz et al., 2007), (f) the relative abundance of E. huxleyi, (g) the relative abundance of G. ericsonii, (h) the relative abundance G. oceanica and (i) the absolute abundance (AA) and accumulation rates (AR). The qualitative distribution of diatoms (large Cosinodiscus) is sketched in Figure 3.3i above the absolute nannoplankton values. Age control points of core GeoB 5844-2 are indicated by triangles at the bottom (for details see Figure 3.2). The trend of the longer-termed cooling cycles and the position of the Heinrich events (H) are sketched at the top. Interstadials in !18O GRIP ice-core record are numbered (1–17). The extent of the “aplanktic zone” (salinity !49–50‰) is sketched at the bottom of the diagram. Gray shaded vertical bars show intervals of major SST-drops and E. huxleyi-peaks, which correspond to H3, H2 and H1.
The present-day nannoplankton community in the northern Red Sea region, overall
dominated by E. huxleyi (within the Noelaerhabdaceae; Winter, 1982), is controlled by the
stable stratification of the waters resulting in oligotrophic conditions (regenerated
production). Only during the short winter period does the breakdown of stratification allow an
upwards-mixing of nutrient-rich subsurface waters. The result of this winter mixing is an
elevated nutrient level in the photic zone, which favors new production (Levanon-Spanier, et
al., 1979; Weikert, 1987). The hydrographic changes are initiated (i.e. the formation of dense
surface waters) by the invasion of cold air from northerly direction. Data from the Gulf of
Aqaba, the northernmost extension of the Red Sea (Figure 3.1) shows high abundances of G.
ericsonii and G. oceanica associated with the deeply mixed waters of the winter period
(Winter et al., 1979). Independent evidence from plankton studies of other algal groups
indicates that the structure of the phytoplankton community in the Gulf of Aqaba is strongly
affected by the deep (600 m water depth or more) convective mixing in winter (e.g., Levanon-
Spanier, et al., 1979; Lindell and Post, 1995; Post, 2005), whereas generally a year-round
stable stratification (mixing limited to the surface waters) characterizes the open northern Red
Sea. Another useful modern analogue for the G. ericsonii-dominated intervals in core GeoB
5844-2 can be observed in the northern parts of the western Mediterranean Sea, the Gulf of
Lion, and adjoining areas up to the Balearic Islands. The comparison of plankton data
(Knappertsbusch, 1993) and hydrological observations (e.g., Send et al., 1999) reveals a
possible relationship between abundance maxima of G. ericsonii and the overturning of the
waters caused by the strong katabatic mistral-wind in winter.
We argue that a deep convective mixing due to the surface buoyancy loss favors
especially Gephyrocapsa spp. We infer that the increase of G. ericsonii in core GeoB 5844-2

Chapter 3: Nannoplankton successions during the last glaciation…
59
reflects significant hydrographic changes driven by the increased strength and influence of
cold northern winds in the northern Red Sea region. Still relative high SSTs during the early
parts of the cooling cycles may account for the brief increases of G. oceanica.
Evidence for the above-described scenario may also comes from planktic diatoms. The
deposition of opal-rich sediments, characterized by abundant large Centrales of the genus
Cosinodiscus (nearly solely Cosinodiscus oculus-iridis) and low absolute values of
nannoplankton (Figure 3.3i), coincide roughly with the Gephyrocapsa spp. dominated
intervals. The mass occurrence of C. oculus-iridis in marine sediments is thought to be the
result of the “fall-dump” (Kemp et al., 2000), a sedimentation-event initiated by the onset of
water mixing in late fall–early winter. It was probably this breakdown of stratification that led
later in the cold season to the increase of the Gephyrocapsa spp.
Though confronted with the unique glacial setting of the Red Sea and the prevailing
“aplanktic zone” (for planktic foraminifera), the nannoplankton abundance patterns are
probably not adequately explained by a salinity increase. The mechanisms described above
provide a plausible explanation for the Noelaerhabdaceae-succession where the glacial
salinity increase is only of secondary importance. Because there is rough accordance between
the absolute abundance patterns (Figure 3.3i) and the long-term trends, documented for
different parameters (e.g., sea level, salinity, temperature), it is difficult to relate the absolute
values to one specific factor. Similarities with the sea level curve (Figure 3.3d; Arz et al.,
2007) could lead to the assumption that the salinity played a crucial role. On the other hand, it
seems to be unlikely that a salinity-related reduction of the absolute values have occurred
without a marked imprint in the species composition. If nannoplankton was affected by the
salinity increase, one could anticipate a gradual disappearance of species (i.e. a non-repeating
species succession somehow similar as documented for planktic foraminifera species). Such
patterns are, however, not recognizable in our record. Instead we observed the reiterated
Noelaerhabdaceae-successions, which are closely correlated with the longer-term cooling
cycles and Heinrich events of the North Atlantic realm. Corroborating evidence against a
salinity-related approach to explain the Noelaerhabdaceae-succession in the northern Red Sea
may come from the Mediterranean. Nannoplankton data from the western Mediterranean
(Colmenero-Hidalgo et al., 2004), although not identical in detail, show comparable patterns
to those from the northern Red Sea. Abundance-peaks of E. huxleyi during Heinrich events
and high values of small Gephyrocapsa between the Heinrich events have been observed
(Colmenero-Hidalgo et al., 2004). It is worth mentioning that even other micro-
paleontological studies undertaken in the northern Red Sea region question a simple salinity-

Chapter 3: Nannoplankton successions during the last glaciation…
60
related approach to explain all shifts within the planktic foraminifera and pteropod
communities. Temperature variations, changes of the water stratification and the availability
of food (i.e. productivity of the surface waters) seem to be highly important even in glacial
times (Almogi-Labin, 1982; Fenton et al., 2000).
Moreover, it should be recalled that the trend towards hypersaline conditions mirrors a
diminishing communication with the open ocean (e.g., Siddall et al., 2003; Arz et al., 2007).
Hence, the imprints of climatic signals transmitted from the Gulf of Aden to the Red Sea
should be weaker during full glacial conditions. Due to the great distance from the Strait of
Bab-el-Mandeb (nearly 2000 km), this should be especially true for our study area. The
Noelaerhabdaceae-succession, by contrast, points to a sensitive and fast acting mechanism
linking high latitudinal climate oscillations and environmental changes in the northern Red
Sea region especially after 32 ka.
A plausible mechanism fulfilling the above requirements is the atmospheric
teleconnection with the extratropics and the associated hydrographic changes driven by an, in
relation to early–mid MIS 3 conditions, reinforced influence of cold air from the mid-
latitudes. Data from the North Atlantic (e.g., Shackleton et al., 2000; Figure 3.4a, planktic
!18O record from the Iberian Margin) and the Mediterranean Sea (e.g., Cacho et al., 1999; Fig.
Figure 3.4. Comparison of (a) the North Atlantic planktic !18O record (Shackleton et al., 2000) and (b) the Western Mediterranean alkenone-sea surface temperatures (Cacho et al., 1999) with (c) the alkenone-sea surface temperatures from the northern Red Sea (Arz et al., 2007). The onset/range of the Noelaerhabdaceae-succession is sketched at the bottom of the diagram.

Chapter 3: Nannoplankton successions during the last glaciation…
61
3.4b, alkenone record from the Alboran Sea) reveal large and rapid environmental
fluctuations in response to the high latitudinal cooling already before 32 ka. Considering the
onset of the succession at 32 ka and the alkenone-SST variations (Figure 3.4c), a stronger
atmospheric connection between the northern Red Sea and the extratropics was established in
the late MIS 3. The present-day Mediterranean winter climate (Figure 3.1) is affected, in
addition to the westerlies responsible for the characteristic winter cyclonic rain, by cold
continental and altered maritime polar and arctic air reaching the region via southern and
eastern Europe (e.g., Air Ministry, 1962; Martyn, 1992; Hoskins and Hodges, 2002). The
onset of the succession at around 32 ka in the northern Red Sea may reflect an expansion of
the extratropical winter climate features into the subtropical desert belt (a wintertime mean
position of the “Mediterranean front” further south; Air Ministry, 1962). This shift and
accordingly the more direct and more prolonged influence of the extratropical circulation
systems on the northern Red Sea may account for the now undamped atmospheric transfer of
the high latitudinal cooling signals to the northern Red Sea as indicated by the nannoplankton
abundance patterns and the alkenone data.
Terrestrial records from the northern parts of the Saharan desert belt provide a good link
between a climatic change towards cooler and wetter conditions (respectively a positive
precipitation-evaporation balance) and the changes of the nannoplankton community
structure. The onset of a distinctive “pluvial period” (in absolute terms probably semi-arid or
at most semi-humid in large part of the present desert belt) is documented at around 32–30 ka
(e.g., Nicholson and Flohn, 1980; Servant and Servant-Vildary, 1980; Rognon, 1996;
Hoelzmann et al., 2004). The increased atmospheric pressure gradient, associated with the
expansion of the northern ice sheets and the enhanced latitudinal temperature contrast,
modulated the atmospheric circulation. A southward shift of the westerlies that allowed
Atlantic depression to supply winter-rain towards the desert belt has been proposed
(Nicholson and Flohn, 1980; Hoelzmann et al., 2004).
This late Pleistocene pluvial period ended at around 22–20 ka with the onset of arid or
even hyper-arid climatic conditions, which characterize the Last Glacial Maximum (LGM;
22–19 ka) and the subsequent interval up to the onset of the Bølling-Allerød (at around 14.5
ka) in wide areas of Africa and Arabia (e.g., Nicholson and Flohn, 1980; Hamilton and
Taylor, 1991; Gasse, 2000; Hoelzmann et al., 2004). The climatic re-orientation at around 22
ka did not disturb the continuation of the Noelaerhabdaceae-succession (Figure 3.3). On
closer examination, however, it seems to be manifested in the nannoplankton record by the
secular trend of F. profunda (Figure 3.5d). Apart from a moderate decrease at around 21 ka,

Chapter 3: Nannoplankton successions during the last glaciation…
62
overall high values of F. profunda since around 22 ka indicate low paleoproductivities (cf.
Beaufort et al., 1997). The highest primary production/lowest F. profunda-abundance is
documented between 32 ka and 22 ka.
The coincidence (in time) between the interval of higher productivity in the northern Red
Sea and the subtropical pluvial period on land (Figure 3.5) suggests a linkage between the
moisture climate and the trophic state of the northern Red Sea waters. Fenton et al. (2000)
propose that the glacial conditions were milder in the Gulf of Aqaba than in the Red Sea.
Unlike modern conditions, a freshwater inflow to the Gulf of Aqaba supplied by
Mediterranean depressions is assumed (Fenton et al. 2000). Higher water levels of the Lake
Lisan (Figure 3.5c), the progenitor of the Dead Sea, may give evidence for higher regional
Figure 3.5. Comparison of the paleoproductivity in the northern Red Sea and the changes in regional precipitation/water balance: (a) variations of the southern desert margin (Reichelt et al., 1992), (b) Chad basin precipitation/evaporation variability (Servant and Servant-Vildary, 1980), (c) Lake level record (in meter below mean sea level) of Lake Lisan (Bartov et al., 2003) and (d) the relative abundance of F. profunda (note reversed axis). Shaded vertical bar indicate the subtropical pluvial (ca. 32 to 22-20 ka) as documented by terrestrial proxy records in the subtropics of Africa (e.g., Nicholson and Flohn, 1980; Rognon, 1996; Hoelzmann et al., 2004). The arrows in Figure 3.5c mark lake level drops of Lake Lisan associated with Heinrich events H5–H1 (Bartov et al., 2003). The paleoflood-record (interval of extreme floods) from the Negev desert (Greenbaum et al., 2006) is sketched in Figure 3.5c together with the Lake Lisan curve.

Chapter 3: Nannoplankton successions during the last glaciation…
63
rainfall (Bartov et al., 2002; 2003). There are similarities between fluctuations of the
reconstructed lake level and the F. profunda abundance patterns/productivity changes on
longer time scale, which may indicate an influence of extratropical precipitation on northern
Red Sea fertility. Another possibility is that the African monsoon (Figure 3.1), which affected
the subtropical desert belt (Figure 3.4a) during the late Pleistocene from the south (Rossignol-
Strick, 1983; Reichelt et al., 1992; Rognon, 1996), influenced marine fertility. According to
Almogi-Labin et al. (1998), the late Pleistocene humid period is documented in the central
Red Sea by abundance maxima of epipelagic pteropod species, which should indicate a
reduced ventilation of the surface waters and an increase in productivity caused by a strong
southwest monsoon. After all, any explanation linking Red Sea fertility and extratropical or
tropical rainfall remains debatable. The high productivity period in the northern Red Sea
cannot be linked to the African monsoon without problems; a strong direct influence of the
southwest monsoon should be limited to the southern and central Red Sea. Furthermore,
differences in the spatial rainfall in the Near East (e.g., Horowitz 1979; Amit et al., 2006;
Dayan and Morin, 2006) make it difficult to compare wetter periods in the northern
subhumid/semiarid parts (e.g., as deduced from the Lake Lisan record) with environmental
changes further south (southern Negev-Gulf of Aqaba-northern Red Sea region). Oxygen
isotopic data from the northern Red Sea (Figure 3.3c), overall characterized by a substantial
salinity overprint, reveal no distinctive period of surface freshening between 32 ka and 22 ka.
This does not necessarily exclude a local restricted fresh water supply (e.g., from wadis), but
wind-driven variations of the water column properties were probably more important. Due to
a wetter atmosphere and the related decrease of thermal stability, stronger winds/a higher
frequency of storms, may have caused stronger upwelling/mixing of ocean waters. This
speculative approach offers an alternative mechanism that explains the high productivity
period in accordance with the lack of a distinct freshening signal in the isotopic data.
Evidence of increased storminess under overall still dry regional conditions is documented by
a recently published paleoflood-record from the hyperarid southern Negev desert (Greenbaum
et al., 2006). A conspicuously higher frequency of storms/paleofloods is indicated during the
late MIS 3–early MIS 2 (Figure 3.4). Interestingly, the authors (Greenbaum et al., 2006)
argued for variations of the Red Sea Trough systems (Kahana et al., 2002; for details) as a
possible cause. This implies a profound role of the tropical convection intensity and hence of
the African monsoon system (cf. Kahana et al., 2002; Dayan and Morin, 2006).
The information available so far only allow speculative statements. Although we note a
high productivity period in the northern Red Sea contemporaneous with the pluvial period on

Chapter 3: Nannoplankton successions during the last glaciation…
64
land, further research is needed to understand the causal relationship between F. profunda
abundance patterns (and deduced primary production) and wetness documented by terrestrial
proxy records.
3.4 Conclusion
Our record demonstrates that the nannoplankton responded differently to high latitudinal
cooling before and after 32 ka. A strong atmospheric influence from the extratropics is
indicated since the late MIS 3. The climatic evolution of the northern subtropics lends support
to this finding. A reiterated succession, closely correlated with the longer-term cooling cycles
and Heinrich events of the North Atlantic realm, is detected after 32 ka. The three Heinrich
events H3, H2, and H1 are dominated by Emiliania huxleyi. Gephyrocapsa oceanica and
especially Gephyrocapsa ericsonii are abundant between H3 and H2 and between H2 and H1.
The present data led to the assumption that the glacial salinity increase did not cause the
nannoplankton successions. Although the glacial Red Sea represents a unique marine
environment with regard to the impressive sea level driven salinity increase, the documented
nannoplankton changes are probably better explained by a more unspectacular approach. A
plausible mechanism seems to be the atmospheric teleconnection with the extratropics and the
associated hydrographic changes driven by an, in relation to present-day and early–mid MIS 3
conditions, reinforced influence of cold air from northerly direction. Furthermore, we
documented a low F. profunda/high productivity interval contemporaneous with a pluvial
period on land between 32 and 22 ka. This finding raises further questions with respect to the
specific mechanism linking F. profunda abundance patterns, marine productivity and
terrestrial environmental changes. Although we cannot present a final explanation for this
relationship here, it may provide an interesting approach for further studies to investigate past
climatic changes in the northern Red Sea region.
Acknowledgements
We thank Rolf Neuser (Bochum) for technical support at the SEM and Alistair Ruffell
(Belfast) for comments on a previous version of the manuscript. This research was supported
by the Deutsche Forschungsgemeinschaft (Mu 667/23-1, -2).

Chapter 3: Nannoplankton successions during the last glaciation…
65
References
Air Ministry Meteorological Office (1962), Weather in the Mediterranean, Vol. 1, 362 pp.,
Her Majesty's Stationery Office, London.
Almogi-Labin, A. (1982), Stratigraphic and Paleoceanographic significance of Late
Quaternary pteropods from deep-sea cores in the Gulf of Aqaba (Elat) and northernmost
Red Sea, Mar. Micropaleontol., 7, 53–72.
Almogi-Labin, A., C. Hemleben, and D. Meischner (1998), Carbonate preservation and
climatic changes in the central Red Sea during the last 380 kyr as recorded by
pteropods, Mar. Micropaleontol., 33(1-2), 87–107.
Amit, R., Y. Enzel, and D. Sharon (2006), Permanent Quaternary hyperaridity in the Negev,
Israel, resulting from regional tectonics blocking Mediterranean frontal systems,
Geology, 34, 509–512.
Andruleit, H. (1996), A filtration technique for quantitative studies of coccoliths,
Micropaleontol., 42, 403–406.
Arz, H. W., J. Pätzold, P. J. Müller, and M. O. Moammar (2003), Influence of Northern
Hemisphere climate and global sea level rise on the restricted Red Sea marine
environment during termination I, Paleoceanography, 18(2), 1053, doi:10.1029/
2002PA000864.
Arz, H. W., F. Lamy, A. Ganopolski, N. Nowaczyk, and J. Pätzold, (2007), Dominant
Northern Hemisphere climate control over millennial-scale glacial sea-level variability,
Quat. Sci. Rev., 26, 312–321.
Bartov Y., M. Stein, Y. Enzel, A. Agnon, and Z. Reches (2002), Lake levels and sequence
stratigraphy of Lake Lisan, the Late Pleistocene precursor of the Dead Sea, Quat. Res.,
57, 9–21.
Bartov, Y., A. Agnon, Y. Enzel, and M. Stein (2003), Catastrophic arid episodes in the
Eastern Mediterranean climate linked with the North Atlantic Heinrich events, Geology,
31, 439–442
Beaufort, L., Y. Lancelot, P. Camberlin, O. Cayre, E. Vincent, F. Bassignot, and L. Labeyerie
(1997), Insolation cycles as a major control of equatorial Indian Ocean primary
production, Science, 278, 1451–1454.
Bond, G., W. Broecker, S. Johnsen, J. McManus, L. Labeyrie, J. Jouzel, and G. Bonani
(1993), Correlation between climate records from North Atlantic sediments and
Greenland ice, Nature, 365, 143–147.

Chapter 3: Nannoplankton successions during the last glaciation…
66
Brand, L. E. (1994), Physiological ecology of marine coccolithophores, in Coccolithophores,
edited by A. Winter and W. G. Siesser, pp. 39–49, Cambridge Univ. Press, Cambridge.
Broecker, W., G. Bond, M. Klas, E. Clark, and J. McManus (1992), Origin of the northern
Atlantic's Heinrich events, Clim. Dyn., 6, 265–273.
Colmenero-Hidalgo, E., J.-A. Flores, F. J. Sierro, M. Ángeles Bárcena, L. Löwemark, J.
Schönfeld, and J. O. Grimalt (2004), Ocean surface water response to short-term
climate changes revealed by coccolithophores from the Gulf of Cadiz (NE Atlantic) and
Alboran Sea (W Mediterranean), Palaeogeogr., Palaeoclimatol., Palaeoecol., 205,
317–336.
Cacho, I., J. O. Grimalt, C. Pelejero, M. Canals, F. J. Sierro, J.-A. Flores, and N. J. Shackleton
(1999), Dansgaard-Oeschger and Heinrich event imprints in the Alboran Sea
paleotemperatures, Paleoceanography, 14, 698–705.
Dansgaard, W., S. J. Johnson, H. B. Clausen, D. Dahl-Jensen, N. S. Gundestrup, C. U.
Hammer, C. S. Hvidberg, J.P. Steffensen, A. E. Sveinbjörnsdottir, J. Jouzel, and G.
Bond (1993), Evidence for general instability of past climate from a 250-kyr ice-core
record, Nature, 364, 218–220.
Dayan, U., and E. Morin (2006), Flash flood-producing rainstorms over the Dead Sea: A
review, in New frontiers in Dead Sea paleoenvironmental research, edited by Y. Enzel,
A. Agnon and M. Stein, Geol. Soc. Am., Spec. Pap., 401, doi: 10.1130/2006/2401(04).
Fenton, M., S. Geiselhart, E. J. Rohling, and C. Hemleben (2000), Aplanktonic zones in the
Red Sea. Mar. Micropaleontol., 40, 277–294.
Gartner, S. (1988), Paleoceanography of the Mid-Pleistocene, Mar. Micropaleontol., 13, 23–
46.
Gasse, F. (2000), Hydrological changes in the African tropics since the Last Glacial
Maximum, Quat. Sci. Rev., 19, 189–211.
Greenbaum, N., N. Porat, E. Rhodes, and Y. Enzel (2006), Large floods during late Oxygen
Isotope Stage 3, southern Negev desert, Israel, Quat. Sci. Rev., 25, 704–719.
Hamilton, A., and D. Taylor (1991), History of climate and forests in tropical Africa during
the last 8 million years, Clim. Change, 19, 65–78.
Hemleben, C., D. Meischner, R. Zahn, A. Almogi-Labin, H. Erlenkeuser, and B. Hiller
(1996), Three hundred eighty thousand year long stabile isotope and faunal records
from the Red Sea: influence of global sea level change on hydrography,
Paleoceanography, 11, 147–156.

Chapter 3: Nannoplankton successions during the last glaciation…
67
Hoelzmann, P., F. Gasse, L. M. Dupont, and U. Salzmann (2004), Palaeoenvironmental
changes in the arid and subarid belt (Sahara-Sahel-Arabian Peninsula) from 150 kyr to
present, in Past climate variability through Europe and Africa, edited by R. W.
Battarbee, C. E. Stickley and F. Gasse, pp. 219–256, Springer, Dordrecht.
Horowitz, A. (1979), The Quaternary of Israel, 394 pp., Academic Press, New York.
Hoskins, B. J., and K. I. Hodges (2002), New perspectives on the Northern Hemisphere
winter storm tracks, J. Atmos. Sci., 59, 1041–1061.
Imbrie, J., J. D. Hays, D. G. Martinson, A. McIntyre, A. C. Mix, J. J. Morley, N. G. Pisias, W.
L. Prell, and N. J. Shackleton (1984), The orbital theory of Pleistocene climate: support
from a revised chronology of the marine !18O record, in Milankovitch and Climate,
edited by A. Berger, J. Imbrie, J. D. Hays, G. Kukla and B. Saltzman, pp. 269–305,
Reidel, Dordrecht.
Kahana, R., B. Ziv, Y. Enzel, and U. Dayan (2002), Synoptic Climatology of Major Floods in
the Negev Desert, Israel, Int. J. Clim., 22, 867–882.
Kemp, A. E. S., J. Pike, R. B. Pearce, and C. B. Lange (2000), The “Fall dump”- a new
perspective on the role of a “shade flora” in the annual cycle of diatom production and
export flux, Deep-Sea Res. II, 47, 2129–2154.
Knappertsbusch, M. (1993), Geographic distribution of living and Holocene coccolithophores
in the Mediterranean Sea, Mar. Micropaleontol., 21, 219–247.
Legge, H. L., J. Mutterlose, and H. W. Arz (2006), Climatic changes in the northern Red Sea
during the last 22,000 years as recorded by calcareous nannofossils, Paleoceanography,
21, doi:10.1029/2005PA001142.
Levanon-Spanier, I., E. Padan, and Z. Reiss (1979), Primary production in a desert-enclosed
sea - the Gulf of Elat (Aqaba), Red Sea, Deep-Sea Res. I, 26, 673–685.
Lindell, D., and A. F. Post (1995), Ultraphytoplankton succession is triggered by deep winter
mixing in the Gulf of Aqaba (Eilat), Red Sea, Limnol. Oceanogr. 40 (6), 1130–1141.
Martyn, D. (1992), Climates of the world, Developments in atmospheric science, 18, 435 pp.,
Elsevier, Amsterdam, Elsevier.
Nicholson, S. E., and H. Flohn (1980), African environmental and climatic changes and the
general atmospheric circulation in late Pleistocene and Holocene, Clim. Change, 2,
313–348.
Okada, H., and P. Wells (1997), Late Quaternary nannofossil indicators of climate change in
two deep-sea cores associated with the Leeuwin Current off western Australia,
Palaeogeogr. Palaeoclimatol. Palaeoecol., 131, 413–432.

Chapter 3: Nannoplankton successions during the last glaciation…
68
Pätzold, J., and cruise participants (2000), Report and preliminary results of METEOR Cruise
M 44/3 Aquaba (Jordan) - Safaga (Egypt) - Dubá (Saudi Arabia) - Suez (Egypt) - Haifa
(Israel), 12.3.–26.3.–2.4.–4.4.1999, Berichte aus dem Fachbereich Geowissenschaften
der Universität Bremen, 149, 135 pp.
Post, A. F. (2005), Nutrient limitation of marine cyanobacteria (Molecular ecology of
nitrogen limitation in an oligotrophic sea), in Harmful Cyanobacteria, edited by J.
Huisman, H. C. P. Matthijs and P. M. Visser, pp. 87–108, Kluwer, Dordrecht.
Reichelt, R., H. Faure, and J. Maley (1992), Die Entwicklung des Klimas im randtropischen
Sahara-Sahelbereich während des Jungquartärs - ein Beitrag zur Klimakunde,
Petermanns geogr. Mitt., 136, 69–79.
Rognon, P. (1996), Climatic change in the African deserts between 130,000 and 10,000 y BP,
C. R. Acad. Sci. Paris, Série II a, 323, 549–561.
Rossignol-Strick, M. (1983), African monsoons, an immediate climate response to orbital
insolation, Nature, 304, 46–49.
Schmithüsen, J. (1976), Atlas zur Biogeographie, 80 pp., Meyers Kartographischer Verlag,
Mannheim.
Send, U., J. Font, G. Krahmann, C. Millot, M. Rhein, and J. Tintoré (1999), Recent advances
in observing the physical oceanography of the western Mediterranean Sea, Prog.
Oceanogr., 44, 37–64.
Servant, M., and S. Servant-Vildary (1980), L’environnement Quaternaire du bassin du
Tchad, in The Sahara and the Nile - Quaternary environments and prehistoric
occupation in northern Africa, edited by M. A. J. Williams and H. Faure, pp. 133–162,
Balkema, Rotterdam.
Shackleton, N. J., M. A. Hall, and E. Vincent (2000), Phase relationships between millennial-
scale events 64,000–24,000 years ago, Paleoceanography, 15, 565–569, doi:10.1029/
1999PA000513
Shackleton, N. J., R. G. Fairbanks, T.-C. Chiu, and F. Parrenin (2004), Absolute calibration of
the Greenland time scale: implications for Antarctic time scales and for "14C, Quat. Sci.
Rev., 23, 1513–1522.
Siddall, M., E. J. Rohling, A. Almogi-Labin, C. Hemleben, D. Meischner, I. Schmelzer, and
D. A. Smeed (2003), Sea-level fluctuations during the last glacial cycle, Nature, 423,
853–858.
Thunell, R. C., S. M. Locke, and D. F. Williams (1988), Glacio-eustatic sea-level control on
Red Sea salinity, Nature, 334, 601–604.

Chapter 3: Nannoplankton successions during the last glaciation…
69
Voelker, A. H. L., and workshop participants (2002), Global distribution of centennial-scale
records for Marine Isotope Stage (MIS) 3, a database, Quat. Sci. Rev., 21, 1185–1212.
Weikert, H. (1987), Plankton and the Pelagic Environment, in Red Sea (Key Environments;
7), edited by F. J. Edwards and S. M. Head, pp. 90–111, Pergamon Press, Oxford.
Winter, A. (1982), Paleoenvironmental interpretation of Quaternary coccolith assemblages
from the Gulf of Aqaba (Elat), Red Sea, Rev. Esp. Micropaleontol., XIV, 291–314.
Winter, A., Z. Reiss, and B. Luz (1979), Distribution of living Coccolithophore assemblages
in the Gulf of Elat (’Aqaba), Mar. Micropaleontol., 4, 197–223.
Winter, A., A. Almogi-Labin, Y. Erez, E. Halicz, B. Luz, and Z. Reiss (1983), Salinity
tolerance or marine organisms deduced from Red Sea Quaternary record. Mar. Geol.,
53, 17–22.
Ziveri, P., K.-H. Baumann, B. Böckel, J. Bollmann, and J. R. Young (2004), Biogeography of
selected Holocene coccoliths in the Atlantic Ocean in Coccolithophores: From
Molecular Processes to Global Impact, edited by H. Thierstein and J. R. Young, pp.
403–428, Springer, New York.

70

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
71
4 Late Holocene environmental changes in the northern Red Sea region as indicated by
nannoplankton abundance patterns
Abstract
A sediment core retrieved from the Shaban Deep, a brine-filled, submarine basin located in
the northern Red Sea, was studied with respect to its calcareous nannoplankton content. We
focus on the abundance patterns of Gephyrocapsa spp. (Gephyrocapsa oceanica and
Gephyrocapsa ericsonii) and the reaction of this group to environmental changes in late
Holocene times. The changes of the abundance patterns observed for the last two millennia
correlate strongly with climatic fluctuations documented by proxy records from the
extratropics of the Northern Hemisphere. The calcareous nannoplankton record documents
two longer-lasting intervals with high Gephyrocapsa spp. values (A.D. 300–600 and A.D.
1350–1820), which coincide with the so-called “Migration Period Pessimum” and “Little Ice
Age” in Europe. Another short termed Gephyrocapsa spp. maximum has been observed
around A.D. 1050, punctuating the “Mediaeval Warm Period”. The Gephyrocapsa spp.
abundance patterns suggest that during cooler periods, documented particularly for Europe
but also for other areas of the Northern Hemisphere, the northern Red Sea was characterized
by a reinforced winter convective mixing. This in turn was probably caused by a
strengthened/more frequent cold air incursion from northerly direction. These findings are
compared with other proxy records of the late Holocene and discussed with respect to the
atmospheric background conditions potentially being responsible for the changes of the
northern Red Sea winter climate.
Key words: Northern Red Sea; Shaban Deep; nannoplankton; winter climate; late Holocene
4.1 Introduction
The Red Sea (Figure 4.1), located within the Afro-Arabian desert belt, is a long (around 1932
km) and narrow (averaged around 280 km) ocean basin, connected to the Gulf of Aden-Indian
Ocean by the shallow (137 m water depth) Strait of Bab-el-Mandeb at its southern end (e.g.,
Morcos, 1970; Edwards, 1987). Due to the negligible fresh water supply and the arid climate,
evaporation (2000 mm yr-1
) clearly outbalances precipitation (10– max. 200 mm yr-1
), surface
waters are characterized by a distinctive increase in salinity from 36–37‰ close to the Strait

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
72
of Bab-el-Mandeb to around 40–41‰ in
the northern Red Sea (Edwards, 1987;
Sofianos et al., 2002). The sea surface
temperatures (SST) reveal the opposite
trend; waters are getting continuously
cooler to the north. Following the
longitudinal axis of the basin, the
prevailing winds are basically from
northwest-to-southeast. Only south of
around 19°N the main direction reverses
in winter under the influence of the
Asian monsoon system (Edwards, 1987).
The anti-estuarine circulation patterns of
the Red Sea waters, the northwards
surface flow and the southwards sub-
surface return flow, are driven by wind
and especially thermohaline forcing with
an intermediate and deep-water formation in the northern Red Sea (Cember, 1988; Woelk and
Quadfasel, 1996; Eshel and Naik, 1997).
The northern Red Sea is situated in the northern margin of the subtropical desert belt, in
vicinity of the Mediterranean climate zone. Due to its sensitive hydrography, which is
influenced by extratropical meteorological elements (e.g., Eshel et al., 2000; Arz et al., 2003;
Felis et al., 2004; Lamy et al., 2006), it represents an interesting area to study paleoclimatic
changes. Fluctuations of the water stratification, driven by air temperature and humidity
changes, are influenced by variations of the North Atlantic winter climate and associated
pressure fields. In particular the North Atlantic Oscillation (NAO; Hurrell, 1995) and its
supra-regional counterpart, the Artic Oscillation (AO; Thompson and Wallace, 1998) play a
key role. Owning to a weak stratification of the northern Red Sea waters, relatively cold and
arid regional conditions are linked to positive NAO/AO index winters (e.g., Felis et al., 2000;
Eshel et al. 2000). A high-pressure anomaly, which extends over large parts of the
Mediterranean, favors an anticyclonic flow of surface winds in the eastern Mediterranean and
causes thereby the advection of northerly air from southeastern Europe towards the northern
Red Sea region (Rimbu et al., 2001; 2006; see also Wallace and Gutzler, 1981).
In an attempt to reveal changes of the northern Red Sea environment on a multidecal to
Figure 4.1. Map showing the Red Sea and the
Gulf of Aden. A detailed view of the northern
Red Sea with the Gulf of Aqaba, the Gulf of
Suez and the Shaban Deep is shown in the upper
right panel.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
73
multicentennial time-scale for the last two millennia, we investigated variations of the
calcareous nannoplankton (further simply named nannoplankton). Sediment surface samples
as well as plankton studies have shown that the most common nannoplankton species in the
overall well-stratified waters of the northern Red Sea is Emiliania huxleyi (Winter, 1982).
Recent results have indicated that during colder periods in earth climate history different
hydrographic conditions prevailed; high abundances of Gephyrocapsa spp. are thought to
indicate an intensified convective mixing of the waters (Legge et al., 2006). The
Gephyrocapsa spp. abundance patterns can therefore probably be understood as an indicator
for the water column conditions, i.e. the strength and frequency of winter mixing in the
northern Red Sea.
This study aims to test whether Gephyrocapsa spp. (Gephyrocapsa ericsonii and G.
oceanica) is a useful indicator for variations of the environmental conditions in the northern
Red Sea region during the late Holocene. Further on we want to view and discuss potential
variations of Gephyrocapsa spp. abundances in the light of paleoceanographic changes caused
by tropical-extratropical teleconnection.
4.2 Methods
4.2.1 Material and geological background
The studied sediment core GeoB 7805-1 was retrieved from the Shaban Deep, an axial
depression of the northern Red Sea (Figure 4.1 and 4.2). It is one of 25 brine-filled, submarine
basins located in the central and northern
Red Sea (Karbe, 1987; Hartmann et al.,
1998). The Red Sea occupies a relatively
young tectonic basin. It is situated in the
long and narrow northwest-trending rift
zone where the African-Arabian continent
broke and spreads apart (Almond, 1986).
The main contour of the Red Sea
depression was probably achieved during
the Oligocene (Bohannon et al., 1989;
Daradich et al., 2003). In Miocene times,
the precipitation of salts resulted in the
buildup of evaporites on the bottom of the young basin. Today, leaching and diffusion of
these evaporites result in the formation of the brines (Manheim, 1974; Hartmann et al., 1998).
Figure 4.2. Simplified topographic map of the
Shaban Deep with its four sub-basins and side
of core GeoB 7805-1.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
74
Two intersected ridges subdivide the Shaban Deep into four smaller sub-basins (Figure
4.2). High-resolution seismic profiles (e.g., Pätzold et al., 2003), in which the top of the
Miocene evaporites is marked by the so-called “S-Reflector”, indicate that the salt crops out
at the basins flanks. Due to its high density, the brine remains “pond-like” in the sub-basins
(brine body thickness around 200 m). The same brine-interface depth (1324.5 ± 0.5 m) and a
similar chemical composition indicate a sub-surface connection of all four sub-basins
(Hartmann et al., 1998). In the current study, we investigated core GeoB 7805-1, taken by a
multicorer during RV Meteor Cruise M52/3 (Pätzold et al., 2003) from the eastern sub-basin
(26°13´9"N and 35°22´6"E; water depth: 1447 m) of the Shaban Deep.
The extreme high salinity (chlorinities 150.7 ± 0.2‰ Cl) and the low oxygen content (<
0.3 mg O2/l) below the brine-seawater interface (Hartmann et al., 1998) have prevented
eukaryotic live and thereby bioturbation, preserving the fine lamination of the brine sediment.
The lack of benthic activity allows a high-resolution paleoceanographic reconstruction. A
detailed description of core GeoB 7805-1 is given by Seeberg-Elverfeldt et al. (2005). Most of
the 48 cm long core shows laminated sediments. From 7.6 cm to 9.5 cm (= HI1) and from
10.9 cm to 18.4 cm (= HI2) core depth two homogenous, non-laminated intervals are
intercalated. Due to the particle size distribution with coarser material at the bottom, both
intervals have been interpreted by Seeberg-Elverfeldt et al. (2005) to be of turbiditic origin.
4.2.2 Sampling and preparation
We tried to balance the sample density between resolution and totally covered time span.
Therefore, core GeoB 7805-1 was consecutively sampled every 0.5 cm. In order to gain
relative and absolute data of the nannoplankton, the filtration technique of Andruleit (1996)
was used for preparation. The freeze-dried sediments were weighted on a high-precision
balance, wet separated with a rotary splitter (FRITSCH Laborette 27) and filtered by means of
a vacuum pump on polycarbonate filters with 0.4 µm pore size. After drying at 40°C, a
wedge-shaped piece of the filter was mounted on an aluminum stub with self-adhesive tape,
outlined with a silver colloidal suspension and coated with gold in argon atmosphere using a
sputter coater (BioRad SC-500). At least 300 specimens per sample were identified and
counted by use of a scanning electronic microscope (LEO 1530 Gemini FESEM).
We investigated the complete core (also the homogeneous intervals/turbidties HI1 and
HI2). The nannoplankton results are prepared in two forms. (1) Primary data set: The absolute
and relative abundance patterns of Gephyrocapsa spp. and the Gephyrocapsa spp./E. huxleyi
ratios are shown in Figure 4.3 on the depth scale (all data are figured, intervals HI1 and HI2

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
75
are included). (2) Corrected for turbities: The relative abundance patterns of Gephyrocapsa
spp. and the Gephyrocapsa spp./Emiliania huxleyi ratio are shown in Figure 4.4 and 4.5 on
the age scale (intervals HI1 and HI2 are excluded; details will be given below).
4.3 Gephyrocapsa spp. abundance patterns
The abundance patterns of Gephyrocapsa spp. are shown in Figure 4.3 on the depth scale.
Two extensive intervals (around 40–45 cm and around 3–20 cm) of higher Gephyrocapsa spp.
values are documented. Throughout the reminder of the core, the abundances of
Gephyrocapsa spp. are, despite a minor peak at around 26 cm, characterized by overall low
values. Figure 4.4 and 4.5 shows the nannoplankton data on the age scale. The depth-age
model (corrected for turbidites) is taken from Seeberg-Elverfeldt et al. (submitted). Core
GeoB 7805-1 covers the time span from A.D. 160 to 1820 (1790 to 130 B.P.). The turbidite
Figure 4.3. Counted (a) absolute and (b, c) relative abundances of Gephyrocapsa spp. as a function
of the core depth. The relative abundances of Gephyrocapsa spp. are shown as their % proportion
on the total nannoplankton community (b; Gephyrocapsa spp. – TOTAL) and as their % propotion
in relation to Emiliania huxleyi (c; Gephyrocapsa. spp – Emiliania huxleyi). The homogeneous
intervals/turbidities HI1 (7,6–9,5 cm) and HI2 (10,9–18,4 cm) are outlines by gray shaded bars.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
76
HI1 removed no previously deposited sediments, whereas turbidite HI2 eroded sediments
presenting a time interval of around one century. Based on this depth-age model, the two
long-termed Gephyrocapsa spp. maxima occur between A.D. 300 and 600 (1650 to 1350
B.P.) and between A.D. 1350 and 1820 (600 to 130 B.P.), respectively. The short-termed
maximum at 26 cm is dated A.D. 1050 (900 B.P.).
Figure 4.4. Comparison of (a) changes in alpine glacier extent (Holzhauser et al., 2005), (b)
variations of petrologic tracers of ice-rafted debris in the eastern North Atlantic (Bond et al., 2001),
(c) reconstructed NH temperature anomalies (Mann and Jones, 2003), (d) stalagmite growth rates
from the Uamh-an-Tartair cave in northwest Scotland (Proctor et al., 2000; 2002) and (e)
reconstructed solar irradiance changes (Bard et al., 2000; with different scaling factors: 0.25%,
0.4%, 0.55% and 0.65%) with the relative abundances of Gephyrocapsa spp. (f and g). Age control
points for core GeoB 7805-1 are indicated by triangles at the bottom.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
77
4.4 Discussion
A general observation is that the core intervals, which are characterized by high values of
Gephyrocapsa spp. coincide in time with periods of overall colder conditions, documented in
most parts of Europe but also elsewhere in the Northern Hemisphere (NH). Changes in alpine
glacier extent (Holzhauser et al., 2005) and variations of petrologic tracers of ice-rafted debris
in the eastern North Atlantic (Bond et al., 2001) as well as reconstructed NH temperature
anomalies (Mann and Jones, 2003), documented overall similar trends (Figure 4.4). Periods of
glacier advance (Figure 4.4a), increased drift ice (Figure 4.4b) and lower NH temperatures
(Figure 4.4c) co-occur with higher values of Gephyrocapsa spp. in the northern Red Sea
(Figure 4.4f and g). The most prominent features in core GeoB 7805-1 are the two long-
lasting Gephyrocapsa spp. maxima (A.D. 300–600 and A.D. 1350–1820), which seem to
correlate with the “Migration Period Pessimum” (roughly between A.D. 300 and 700;
sometimes also “Dark Ages Cold Period”, e.g., Keigwin and Pickart, 1999) and the “Little Ice
Age” (LIA, roughly between A.D. 1350 and 1850). In addition, the Gephyrocapsa spp.
maximum at A.D. 1050, although less well reflected in the above records, probably matches a
short-lasting cool interval within the “Mediaeval Warm Period” (MWP) and the Oort solar
minimum (cf. Pfister et al., 1998; Damon and Peristykh, 2000; Pla and Catalan, 2005;
Ammann et al., 2007).
In order to integrate the observed nannoplankton changes into a more regional context,
we compare our data with records from the eastern Mediterranean (Figure 4.5). Based on the
interpretation of proxy records and archaeological evidence, Issar (1998) created an integrated
climate scheme for the eastern Mediterranean, which shows alternating cold-humid (Roman
Period = 2200–1700 B.P., Byzantine Period = 1800–1500 B.P., Crusader Period = 1000–800
BP, LIA = 500–100 B.P.) and warm-dry (Moslem-Arab Period = 1300–1000 BP, Moslem-
Turkish Period = 800–500 B.P.) phases for the late Holocene (Figure 4.5a). The long-lasting
Gephyrocapsa spp. maxima (Figure 4.5f and g) probably coincide with the Byzantine Period
and the LIA of Issar (1998). The short-lasting Gephyrocapsa spp. maximum at A.D. 1050
may fall within the Crusader Period. A detailed correlation of single proxy records from the
eastern Mediterranean with the nannoplankton abundance patterns of core GeoB 7805-2 is
more difficult. The Dead Sea lake level record for the last two millennia is shown in Figure
4.5b (Enzel et al., 2003; Bookman et al., 2004). Lake level high stands occurred at the end of
the Byzantine and Crusader period as well as from the end of the 19th century to the onset of
the 20th century. With regard to the proposed precipitation patterns, a deviation from the
scheme of Issar (1998) is provided by terrestrial and marine !18
O and !13
C records (!13
C not

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
78
further discussed here) from the southeastern Mediterranean (Figure 4.5c and d). Oxygen
isotope variations of the foraminifera Globigerinoides ruber) from the southeastern
Mediterranean Sea (Figure 4.5c; Schilman et al., 2001) and of the Israeli Soreq cave
speleothem (Figure 4.5d; Bar-Matthews et al., 1997; Bar-Matthews and Ayalon, 2004) are
linked to changes of temperature and especially to variations of the regional hydrological
balance. The information provided by both, the foraminiferal and the speleothem record, are
summarized by Schilman et al. (2002), which propose two dry (and/or cold) “events” (Event I
Figure 4.5. Comparison of paleoclimatic records from the eastern Mediterranean region with the
nannoplankton data: (a) simplified illustration of the cold-humid/warm-dry series of Issar (1998),
(b) Dead Sea lake level (Enzel et al., 2003), (c) foraminiferal (Schilman et al., 2002) and (d)
speleothem !18O (Bar-Matthews et al., 1997; Bar-Matthews and Ayalon, 2004) data from the south
eastern Mediterranean and (e) a schematic view of the precipitation/evaporation changes in the Nar
Gölü crater-lake in central Turkey (Jones et al., 2006). The displayed nannoplankton record (f and g) is identical with Figure 4.4f and 4.4g. Age control points are indicated as in Figure 4.4.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
79
= 300 B.P., Event III = 900 B.P.) and two humid (and/or warm) “events” (Event II = 700
B.P., Event IV = 1300 B.P.) during the last two millennia.
Evidence, at least partly, for the basic NH climate trends can be also seen in the eastern
Mediterranean. However, even if we take dating uncertainties as well as different approaches
to interpret the impact of temperature and rainfall on the paleoclimatic archives for granted,
the overall impression from the above records is that they reflect also the complexity of
Mediterranean climate caused by orography, land-sea contrasts and the diversity of climate
regimes affecting the region (e.g., Kutiel et al., 1996; Kutiel and Benaroch, 2002; Alpert et
al., 2005; Ziv et al., 2006).
Our approach to explain the nannoplankton variations in core GeoB 7805-1 is based on
the intimate relationship between the physical state of the water column and the structure of
the phytocoenosis. Vertical mixing of the water column controls the nutrient distribution
within the photic zone and triggers the composition and annual succession of the
phytoplankton (e.g., Lindell and Post, 1995). A preference for higher levels of inorganic
macronutrients and an adaptation to relatively unstable environmental conditions is typical of
Gephyrocapsa spp. Gephyrocapsa oceanica and G. ericsonii are abundant in upwelling areas
and nutrient-enriched coastal regions, as well as in overall oligotrophic waters in response to
seasonal mixing events (e.g., Winter et al., 1979; Okada and Wells; 1997, Ziveri et al., 2004).
Since no nutrient-rich fresh waters enter the Red Sea (Morcos, 1970; Hoelzmann et al., 1998;
Siddall et al., 2003) and the relatively eutrophic waters from the Gulf of Aden-Indian Ocean
effect only the south (Weikert, 1987), higher trophic levels in the open northern Red Sea
photic zone mainly result from the upwards convective mixing of the nutrient-richer
subsurface waters in winter. Convection can penetrate to sizeable depths only if an adequate
increase in density results from cooling and evaporation of the surface waters. Reinforced
winter mixing, most likely induced by a strengthened/more frequent penetration of cold
northerly air into the northern Red Sea region during periods of higher Gephyrocapsa spp.
values, is therefore assumed.
Detailed insights into the succession patterns of various phytoplankton groups in the
northern Red Sea region are especially available from the Gulf of Aqaba (e.g., Levanon-
Spanier et al., 1979; Winter et al., 1979; Lindell and Post, 1995). The environmental
conditions in the Gulf are characterized by an annual cycle of water stratification and mixing,
accompanied by distinctive succession patterns of the phytoplankton community. Surface
waters are characterized during most of the year by a shallow, but stable thermocline. Air
temperature lowering in late fall causes a rapid erosion of the stratification and induces a deep

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
80
convective mixing up to 600 m water depth in winter (December, January, February).
Levanon-Spanier et al. (1979) investigated the larger sized phytoplankton (! 65 "m). The
community is dominated by dinoflagellates followed by diatoms and cyanobacteria
(Trichodesmium) in summer. In contrast, diatoms nearly solely dominate (within the spectrum
of investigated species) the winter period. Winter et al. (1979) studied the calcareous
nannoplankton. Emiliania huxleyi is the major component of the assemblage composition
during most time of the year. In winter, G. ericsonii, along with G. oceanica, increases in
abundance and characterizes the nannoplankton community in addition to E. huxleyi.
Furthermore, G. ericsonii is able to displace E. huxleyi as the dominating species during this
season. Lindell and Post (1995) compared the abundance patterns of three small sized
phytoplankton-groups: small eukaryotic algae (2–8 "m), the cyanobacteria Synechococcus (1–
2 "m) and the prochlorophyte Prochlorococcus (0.6–1 "m). The small eukaryotic algae
dominate in late winter, stimulated by the deep convective mixing and the associated injection
of nutrients into the photic zone. With the onset of stratification, eukaryotic algae decline and
Synechococcus increases in abundance. Prochlorococcus, absent in winter and rare during the
first two months after the onset of stratification, dominates the stratified and nutrient poor
waters in summer. In sum, the change of the surface water conditions, caused by the winter
convection, seems to be the most important factor affecting the composition of the
phytoplankton in the Gulf of Aqaba (Lindell and Post, 1995; Post, 2005). Previous studies
portray the influence of low temperature and evaporation as a key factor controlling the water
column stability in the northern Red Sea (see 4.1). Though limited in strength and depth, in
relation to the Gulf of Aqaba, the mixing of the waters during the short winter season triggers
the northern Red Sea environment and its biological interior (Wyrtki, 1974; Weikert, 1987;
Eshel et al., 2000).
As stated in the Introduction section, severe conditions in the northern Red Sea region are
thought to be favored today during positive NAO/AO index winters. Hence, the
Gephyrocapsa spp. maxima may be explained in term of prolonged positive NAO/AO index
winters. Taking the modern phase opposition between the northern Red Sea region and the
European sector as a response to NAO/AO for granted, such approach would probably be in
conflict with paleoclimatic information provided from other study sides (e.g., Proctor et al.,
2000; Shindell et al., 2001; Noren et al., 2002). The NAO index is based on the normalized
winter sea level atmospheric pressure differences between the Azores high and the Icelandic
low. Atmospheric circulation patterns, which cause stormy but relatively warm conditions in
northwestern Europe are favored today during positive NAO index winters (strong westerlies

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
81
over the eastern North Atlantic), whereas a negative index (weak westerlies) coincide with
cold and dry conditions (Hurrell, 1995). Enhanced European winter cooling, driven by strong
advection of northeasterly air under the influence of continental anticyclones during the LIA,
or at least during their severest stages (Maunder Minimum, Late Maunder Minimum), is
probably rather consistent with a negative than with a positive NAO (e.g., Luterbacher et al.,
2001; Shindell et al., 2001). Growth rate variations of stalagmites from the Uamh-an-Tartair
cave in northwest Scotland are interpreted to reflect variations of the local precipitation,
strongly controlled by the winter NAO (Figure 4.4d). High precipitation throughout the MWP
is linked to persistently positive index winters, whereas predominantly negative indexes mark
the LIA (Proctor et al., 2000; 2002). It has to be noted, that not all records agree on a
persistently negative state of the NAO during the LIA (Luterbacher et al., 2001; Cook et al.,
2002; Brückner and Mackensen; 2006). In fact, multiproxy reconstructions illustrate large
variability. For example, comparable periods of high positive NAO index winters, as
documented during the twentieth century, seem to have occurred from the mid 15th century
towards the mid 16th century (Cook et al., 2002). Irrespective by whether the NAO/AO
phenomenon exhibited a long lasting preferred mode or was restricted to an interannual and
interdecal timescale, the Gephyrocapsa spp. abundance patterns documented in core GeoB
7805-1 are difficult to ascribe to NAO/AO on the basis of its modern atmospheric flow
characteristics (cf. Keigwin and Pickart, 1999; Bond et al., 2001).
Other teleconnections than NAO/AO may have played a role in our study domain.
Prominent but less well-studied extratropical teleconnections detected in the eastern
Mediterranean region are the zonal oriented East Atlantic-West Russia Pattern (Barnston and
Livezey, 1987; Krichak and Alpert, 2005) and the comparable North Sea-Caspian Pattern
(NCP; Kutiel and Benaroch, 2002). A possible relationship between the NCP and the eastern
Mediterranean winter climate was recently suggested on the basis of a proxy record (for
precipitation) from the Nar Gölü crater-lake (Cappadocia region of Turkey; Jones et al.,
2006). Associated with an increased northeasterly atmospheric circulation across the
Balkan/Black Sea-region (Kutiel et al., 2002), below-normal temperature and precipitation is
documented in the Cappadocia region today during positive NCP episodes. In the Nar Gölü
crater-lake record (schematically illustrated in Figure 4.5e) overall dryer conditions are
documented from A.D. 300 to 500, around A.D. 800–900 and from A.D. 1400 to 1950.
Wetter conditions, on the other hand, prevailed from A.D. 560 to 750 and from A.D. 1000 to
1350. Though the NCP may play a significant role in the northeastern Mediterranean, the
conclusion that it also influenced the northern Red Sea winter environment is highly

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
82
speculative at this moment. More detailed records are essential to decipher the regional
importance of the NCP/East Atlantic-West Russia Pattern during the late Holocene.
Another approach to explain the Gephyrocapsa spp. maxima is that the linkage between
the northern Red Sea and the mid-latitudes changed more generally during the NH cold
periods. The reaction of the nannoplankton in the subtropical northern Red Sea to
extratropical forcing should be finally critical to the poleward development of the Hadley cell
and hence to strength and position of the associated upper-tropospheric subtropical jet
(located today at around 30°N in winter; Held and Hou, 1980; Chang, 1995). Due to the
exposed position of our study domain, even a subtle equatorward shift/weakening of the
Hadley cell/subtropical jet may increase the likelihood of extratropical air to affect the
northern Red Sea setting. Similar dynamics of the subtropical jet are observed during the last
decades in the opposite direction caused by the global warming (Fu et al., 2006) and may
account, at least partly, for the strong correlation between winter rainfall in Israel and ENSO
(El Niño-Southern Oscillation) events since the mid-1970s (Price et al., 1998; Alpert et al.,
2006). The signature of ENSO is also documented in a 245-year !18
O coral record from the
northern Red Sea (Felis et al., 2000). The authors consider variations in the strength of
NAO/ENSO atmospheric teleconnection (i.e., changing regional importance of extratropical
and tropical teleconnection modes) to explain the relationship between northern Red Sea
SSTs and coral !18
O variations on a multi-decadal time scale.
There is reason to assume that the influence from the extratropics could have been
intensified during the NH cold periods beyond those changes associated with phenomena like
the NAO/AO or other extratropical teleconnections. Following this line, the Gephyrocapsa
spp. abundance variations documented in core GeoB 7805-1 seem to reflect first and foremost
changes of the tropical-extratropical connection which may have been caused by a more or
less latitudinal-parallel shift/modification of the polewards branch of the Hadley cell. In
addition, a modulation of the regional characteristic of the Hadley cell/subtropical jet stream
patterns coupled with variations of the tropical circulation should be considered. The intensity
of tropical diabatic heating and associated tropical atmospheric circulation strength is seen to
modulate the location and shape of the ridges within the subtropical jet (Krishnamurti, 1979;
Yang and Webster, 1990). A weaker diabatic heating in central equatorial Africa may have a
lasting influence on the Mediterranean ridge of the jet. A recently published proxy record (Mg
% in authigenic calcite) from Lake Edwards provides evidence for droughts/reduced tropical
convection at A.D. 540–890, 1000–1200 and 1400–1750 in equatorial Africa (Russel and
Johnson, 2007).

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
83
Finally there is also the possibility that the late Holocene climatic changes are related to
variations of solar output. With regard to the mechanism treated above (i.e., Hadley
cell/Subtropical jet stream pattern), the effect of irradiance changes on the thermally structure
of the stratosphere and on the tropospheric winter climate is of particular interest. Model
studies (e.g., Haigh, 1996; Shindell et al., 1999) of the eleven-year sunspot cycle indicate a
broadening (retreating) of the Hadley circulation and poleward (equatorward) shifts of the
subtropical jets during periods of solar maximum (solar minimum). To the extent that it is
appropriate to apply the above model results to the late Holocene epoch, it seems possible that
solar variability played a crucial role on the observed nannoplankton changes by modifying
the poleward branch of the Hadley cell and thereby the tropical-extratropical connection.
Figure 4.4e shows the solar irradiance changes for the last 1200 years as indicated by
fluctuations of the cosmogenic nuclides 10-Beryllium (10
Be) and 14-Carbon (14
C) (Bard et al.,
2000). In comparison with the nannoplankton record (Figure 4.4f and g), similarities between
the Gephyrocapsa spp. maxima and intervals of reduced solar output are indicated.
It is worth mentioning that the relevance of solar variations in forcing late Holocene
climate is still the subject of much discussion (Lean and Rind, 1998; Bond et al., 2001;
Ammann et al., 2007) and that we cannot present here a final key for the understanding of the
sun-climate interaction. Whether solar activity was the dominating external forcing factor or
not, variations of the subtropical jet/of the Hadley cell are probably an important approach
that could help to explain the distinctive nannoplankton changes documented during the last
two millennia in the northern Red Sea and the coincidence of Gephyrocapsa spp. maxima and
lower NH temperatures.
4.5 Conclusions
The calcareous nannoplankton from the Shaban Deep supplies crucial information for a better
understanding of the Red Sea paleoenvironment. Throughout the last two millennia, the
northern Red Sea region was affected by climatic fluctuations, which left a clear imprint in
the nannoplankton record. The record revealed three intervals of higher Gephyrocapsa spp.
values, which seem to coincide in time with periods of overall lower temperatures.
Convective mixing, prefaced by the buoyancy loss of the surface waters, probably triggered
the increase of Gephyrocapsa spp. The changes of the regional hydrography can be explained
by a reinforced wintertime penetration of cold northerly air. Our observations underscore that
the linkage between the northern Red Sea and the mid-latitudes is complicated and varies
through time. The indicated changes of the northern Red Sea hydrographic are probably not

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
84
adequately explained by NAO/AO (supposing modern patterns). Suggestions are made,
linking the documented calcareous nannoplankton changes with other phenomena. Overall, it
remains important to pay attention to the exposed position of the northern Red Sea near the
tropical-extratropical atmospheric “boundary”. Thermally driven changes of the Hadley cell
in winter and of the associated subtropical jet may have contributed to the observed
nannoplankton changes by modulating the tropical-extratropical connection. Variations of
solar output are perhaps the primary trigger; further research is, however, needed to
understand its influence on the climate, especially with regard to the conditions in the North
Atlantic realm and associated atmospheric circulation patterns.
Acknowledgements
This study was supported by the Deutsche Forschungsgemeinschaft (Mu 667/26-1, -2).
Sample material has been supplied by Ismene A. Seeberg-Elverfeldt and Jürgen Pätzold
(Bremen). Special thanks go to Rolf Neuser for technical support at the SEM and to Sylvia
Rückheim for helpful comments and discussions.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
85
References
Almond, D. C. (1986), Geological evolution of the Afro-Arabian dome, Tectonophys., 131,
301–332.
Alpert, P., M. Baldi, R. Ilani, S. Krichak, C. Price, X. Rodo, H. Saaroni, B. Ziv, P. Kischa, J.
Barkan, A. Mariotti, and E. Xoplaki (2006), Relations between climate variability in the
Mediterranean region and the Tropics: ENSO, South Asian and African Monsoons,
Hurricanes and Saharan dust, in The Mediterranean Climate: an overview of the main
characteristics and issues, edited by P. Lionello, P. Malanotte-Rizzoli and R. Boscolo,
pp. 149–177, Elsevier, Amsterdam.
Ammann, C. M., F. Joos, D. S. Schimel, B. L. Otto-Bliesner, and R. A. Tomas (2007), Solar
influence on climate during the past millennium: Results from transient simulations
with the NCAR Climate System Model, PNAS, 104, 3713–3718.
Andruleit, H. (1996), A filtration technique for quantitative studies of coccoliths,
Micropaleontol., 42, 403–406.
Arz, H. W., F. Lamy, J. Pätzold, P. J. Müller, and M. Prins (2003), Mediterranean Moisture
Source for an Early-Holocene Humid Period in the Northern Red Sea, Science, 300,
118–121.
Bar-Matthews, M., A. Ayalon, and A. Kaufman (1997), Late Quaternary paleoclimate in the
eastern Mediterranean region from stable isotope analysis of Speleothems at Soreq
Cave, Israel, Quat. Res., 47, 155–168.
Bar-Matthews, M., and A. Ayalon (2004), Speleothems as palaeoclimate indicators, a case
study from Soreq Cave located in the eastern Mediterranean region, Israel, in Past
climate variability through Europe and Africa, edited by R. W. Battarbee, F. Gasse and
C. E. Stickley, pp. 363–391, Springer, New York.
Barnston, A. G., and R. E. Livezey (1987), Classification, seasonality and persistence of low-
frequency atmospheric circulation patterns, Mon. Weather Rev., 115, 1083–1126.
Bard, E., G. Raisbeck, F. Yiou and J. Jouzel (2000), Solar irradiance during the last 1200
years based on cosmogenic nuclides, Tellus, 52, 985–992.
Bohannon, R. G., C. W. Naeser, D. L. Schmidt, and R. A. Zimmermann (1989), The timing of
uplift, volcanism, and rifting peripheral to the Red Sea: A case of passive rifting?, J.
Geophys. Res., 94, 1683–1701.
Bond, G., B. Kromer, J. Beer, R. Muscheler, M. N. Evans, W. Showers, S. Hoffmann, R.
Lotti-Bond, I. Hajdas, and G. Bonani (2001), Persistent solar influence on North
Atlantic climate during the Holocene, Science, 294, 2130–2136.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
86
Bookman (Ken-Tor), R., Y. Enzel, A. Agnon, and M. Stein (2004), Late Holocene lake levels
of the Dead Sea, Geol. Soc. Am. Bull., 116, 555–571.
Brückner, S., and A. Mackensen (2006), Deep-water renewal in the Skagerrak during the last
1200 years triggered by the North Atlantic Oscillation: evidence from benthic
foraminiferal !18
O, The Holocene, 16, 331–340.
Cember, R. P. (1988), On the sources, formation, and circulation of Red Sea Deep Water, J.
Geophys. Res., 93, 8175–8191.
Chang, E. K. M. (1995), The influence of Hadley circulation intensity changes on
extratropical climate in an idealized model, J. Atmos. Sci., 52, 2006–2024.
Cook, E. R., R. D. D’Arrigo, and M. E. Mann (2002), A well-verified, multiproxy
reconstruction of the winter North Atlantic Oscillation Index since A.D. 1400, J.
Climate, 15, 1754–1764.
Daradich, A., J. X. Mitrovica, R. N. Pysklywec, S. D. Willett, and A. M. Forte (2003), Mantle
flow, dynamic topography, and the rift-flank uplift of Arabia, Geology, 31, 901–904.
Damon, P. E., and A. N. Peristykh (2000), Radiocarbon calibration and application to
geophysics, solar physics, and astrophysics, Radiocarbon, 42, 137–150.
Edwards, F. J. (1987), Climate and Oceanography, in Red Sea (Key Environments; 7), edited
by F. J. Edwards and S. M. Head, pp. 45–70, Pergamon, Oxford.
Enzel, Y., R. Bookman (Ken-Tor), D. Sharon, H. Gvirtzman, U. Dayan, B. Ziv, and M. Stein
(2003), Late Holocene climates of the Near East deduced from Dead Sea level
variations and modern regional winter rainfall, Quat. Res., 60, 263–273.
Eshel, G., and N. H. Naik (1997), Climatological coastal jet collision, intermediate water
formation, and the general circulation of the Red Sea, J. Phys. Ocenogr., 24(7), 1233–
1257.
Eshel, G., D. P. Schrag, and B. F. Farrell (2000), Troposphere-planetary boundary layer
interactions and the evolution of the ocean surface density: Lessons from Red Sea
corals, J. Climate, 13, 339–351.
Felis, T., J. Pätzold, Y. Loya, M. Fine, A. H. Nawar, and G. Wefer (2000), A coral oxygen
isotope record from the northern Red Sea documenting NAO, ENSO, and North Pacific
teleconnections on Middle East climate variability since the year 1750,
Paleoceanography, 15, 679–694.
Felis, T., G. Lohmann, H. Kuhnert, S. J. Lorenz, D. Scholz, J. Pätzold, S. A. Al-Rousan, and
S. M. Al-Moghrabi (2004), Increased seasonality in Middle East temperatures during
the last interglacial period, Nature, 429, 164–168.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
87
Fu, Q., C. M. Johanson, J. M. Wallace, and T. Reichler (2006), Enhanced mid-latitude
tropospheric warming in satellite measurements, Science, 312, 1179.
Haigh, J.H. (1996), The impact of solar variability on climate, Science, 272, 981–984.
Hartmann, M., J. C. Scholten, P. Stoffers, and F. Wehner (1998), Hydrographic structure of
brine-filled deeps in the Red Sea - new results from the Shaban, Kebrit, Atlantis II, and
Discovery Deep, Mar. Geol., 144, 311–330.
Held, I. M., and A. Y. Hou (1980), Nonlinear axially symmetric circulations in a nearly
inviscid atmosphere, J. Atmos. Sci., 37, 515–533.
Holzhauser, H., M. Magny, and H. J. Zumbühl (2005), Glacier and lake-level variations in
west-central Europe over the last 3500 years, The Holocene, 15, 789–801.
Hoelzmann, P., D. Jolly, S.P. Harrison, F. Laarif, R. Bonnefille, and H. J. Pachur (1998),
Mid-Holocene land-surface conditions in northern Africa and the Arabian peninsula: A
data set for the analysis of biogeophysical feedbacks in the climate system, Glob.
Biogeochem. Cycl., 12(1), 35–51.
Hurrell, J. W. (1995), Decadal trends in the North Atlantic Oscillation: Regional temperatures
and precipitation, Science, 269, 676–679.
Issar, A. S. (1998), Climate change and history during the Holocene in the Eastern
Mediterranean region, in Water, environment and society in times of climatic change,
edited by A. S. Issar and N. Brown, pp. 113–128, Kluwer, Dordrecht.
Jones, M. D., C. N. Roberts, M. J. Leng, and M. Türkes (2006), A high-resolution late
Holocene lake isotope record from Turkey and links to North Atlantic and monsoon
climate, Geology, 34, 361–364.
Karbe, L. (1987), Hot Brines and the Deep Sea environment, in Key Environments: Red Sea,
edited by A. J. Edwards and S. M. Head, pp. 70–89, Pergamon, Oxford.
Keigwin, L. D., and R. S. Pickart (1999), Slope water current over the Laurentian fan in
interannual to millennial time scale, Science, 286, 520–523.
Kutiel, H., P. Maheras, and S. Guika (1996), Circulation indices over the Mediterranean and
Europe and their relationship with rainfall conditions across the Mediterranean, Theor.
Appl. Climatol., 54, 125–138.
Kutiel, H., and Y. Benaroch (2002), North Sea-Caspian Pattern (NCP) – an upper level
atmospheric teleconnection affecting the Eastern Mediterranean: Identification and
definition, Theor. Appl. Climatol., 71, 17–28.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
88
Kutiel, H., P. Maheras, M. Türkes, and S. Paz (2002), North Sea-Caspian Pattern (NCP) – an
upper level atmospheric teleconnection affecting the Eastern Mediterranean –
implication on the regional climate, Theor. Appl. Climatol., 72, 173–192.
Krichak, S., and Alpert, P. (2005), Decadal trends in the East Atlantic-West Russia Pattern
and Mediterranean precipitation, Int. J. Clim., 25, 183–192.
Krishnamurti, T.N. (1979), Compendium of Meteorology, v. 2, part 4 Tropical Meteorology,
Rep. 364, World Meteorological Organization, Geneva.
Lamy, F., H. W. Arz, G. C. Bond, A. Bahr, and J. Pätzold (2006), Multicentennial-scale
hydrological changes in the Black Sea and the northern Red Sea during the Holocene
and the Arctic/North Atlantic Oscillation, Paleoceanography, 21,
doi:10.1029/2005PA001184.
Lean, J., and D., Rind (1998), Climate forcing by changing solar radiation, J. Climate, 11,
3069–3094.
Legge, H. L., J. Mutterlose, and H. W. Arz (2006), Climatic changes in the northern Red Sea
during the last 22,000 years as recorded by calcareous nannofossils, Paleoceanography,
21, doi:10.1029/2005PA001142.
Levanon-Spanier, I., E. Padan, and Z. Reiss (1979), Primary production in a desert-enclosed
sea – the Gulf of Elat (Aqaba), Red Sea, Deep-Sea Res. I, 26, 673–685.
Lindell, D., and A. F. Post (1995), Ultraphytoplankton succession is triggered by deep winter
mixing in the Gulf of Aqaba (Eilat), Red Sea, Limnol. Oceanogr. 40 (6), 1130–1141.
Luterbacher, J., R. Rickli, E. Xoplaki, C. Tinguely, C. Beck, C. Pfister, and H. Wanner
(2001), The Late Maunder Minimum (1675–1715) – A key period for studying decadal
scale climatic changes in Europe, Clim. Change, 49, 441–462.
Manheim, F. T. (1974), Red Sea geochemistry, Initial Report DSDP, 23, 975–998.
Mann, M. E., and P. D. Jones (2003), Global surface temperatures over the past two
Millennia, Geophys. Res. Lett., 30, doi: 10.1029/2003GL017814.
Morcos, S. A. (1970), Physical and chemical oceanography of the Red Sea, Oceanogr. Mar.
Biol. Ann. Rev., 18, 73–202.
Noren, A. J., P. R. Biermann, E. J. Steig, A. Lini, and J. Southon (2002), Millennial-scale
storminess variability in the northeastern United States during the Holocene epoch,
Nature, 419, 821–824.
Okada, H., and P. Wells (1997), Late Quaternary nannofossil indicators of climate change in
two deep-sea cores associated with the Leeuwin Current off western Australia,
Palaeogeogr. Palaeoclimatol. Palaeoecol., 131, 413–432.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
89
Pätzold, J., G. Bohrmann, and C. Hübscher (2003), Black Sea-Mediterranean-Red Sea, Cruise
No. 52, January 2–March 27, 2002, Meteor-Berichte, Universität Hamburg, 178 pp.
Pfister, C., J. Luterbacher, G. Schwarz-Zanetti, and M. Wegmann (1998), Winter air
temperature variations in western Europe during the early and high middle ages (AD
750-1300), The Holocene, 8, 535–552.
Pla, S., and J. Catalan (2005), Chrysophyte cysts from lake sediments reveal the submillennial
winter/spring climate variability in the northwestern Mediterranean region throughout
the Holocene, Climate Dyn., 24, 263–278.
Post, A. F. (2005), Nutrient limitation of marine cyanobacteria (Molecular ecology of
nitrogen limitation in an oligotrophic sea), in Harmful Cyanobacteria, edited by J.
Huisman, H. C. P. Matthijs and P. M. Visser, pp. 87–108, Kluwer, Dordrecht.
Price, C., L. Stone, A. Huppert, B. Rajagopalan, and P. Alpert (1998), A possible link
between El Niño and precipitation in Israel, Geophys. Res. Lett., 25, 3963–3966.
Proctor, C. J., A. Baker, W. L. Barnes, and M. A. Gilmour (2000), A thousand year
speleothem proxy record of North Atlantic climate from Scotland, Climate Dyn., 16,
815–820.
Proctor, C. J., A. Baker, and W. L. Barnes (2002), A three thousand year record of North
Atlantic climate, Climate Dyn., 19, 449–454.
Rimbu, N., G. Lohmann, T. Felis, and J. Pätzold (2001), Arctic Oscillation signature in a Red
Sea coral, Geophys. Res. Lett., 28, 2959–2962.
Rimbu, N., T. Felis, G. Lohmann, and J. Pätzold, (2006), Winter and summer climate patterns
in the European-Middle East during recent centuries as documented in a northern Red
Sea coral record, The Holocene, 16, 321–330.
Russell, J. M., and T. C. Johnson, (2007), Little Ice Age drought in equatorial Africa:
Intertropical Convergence Zone migrations and El Niño-Southern Oscillation
variability, Geology, 35, 21–24.
Schilman, B., M. Bar-Matthews, A. Almogi-Labin, and B. Luz (2001), Global climate
instability reflected by Eastern Mediterranean marine records during the late Holocene,
Palaeogeogr. Palaeoclimatol. Palaeoecol., 176, 157–176.
Schilman, B., A. Ayalon, M. Bar-Matthews, E. J. Kagan, and A. Almogi-Labin (2002), Sea-
land paleoclimate correlation in the Eastern Mediterranean region during the late
Holocene, Isr. J. Earth Sci.51, 181–190.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
90
Seeberg-Elverfeldt, I. A., C. B. Lange, J. Pätzold, and G. Kuhn (2005), Laminae type and
possible mechanisms for the formation of laminated sediments in the Shaban Deep,
northern Red Sea, Ocean Science, 1, 113–126.
Seeberg-Elverfeldt, I.A., J. Pätzold, H. W. Arz, and J.-B. W. Stuut (2007), Late Holocene
climate variability in the northern Red Sea (submitted to Paleoceanography).
Shindell, D. T., D. Rind, N. Balachandran, J. Lean, and P. Lonergan (1999), Solar cycle
variability, ozone, and climate, Science, 284, 305–308.
Shindell, D. T., G. A. Schmidt, M. E. Mann, D. Rind, and A. Waple (2001), Solar forcing of
regional climate change during the Maunder minimum, Science, 294, 2149–2152.
Siddall, M., E. J. Rohling, A. Almogi-Labin, C. Hemleben, D. Meischner, I. Schmelzer, and
D. A. Smeed (2003), Sea-level fluctuations during the last glacial cycle, Nature, 423,
853–858.
Sofianos, S. S., W. E. Johns, and S. P. Murray (2002), Heat and freshwater budgets in the Red
Sea from direct observations at Bab el Mandeb, Deep-Sea Res. II, 49, 1323–1340.
Thompson, D. W. J., and J. M. Wallace (1998), The Arctic Oscillation signature in the
wintertime geopotential height and temperature fields, Geophys. Res. Lett., 25, 1297–
1300.
Wallace, J. M., and D. S. Gutzler (1981), Teleconnections in the geopotential height field
during the Northern Hemisphere winter, Mon. Wea. Rev., 109, 784–812.
Weikert, H. (1987), Plankton and the Pelagic Environment, in Red Sea (Key Environments;
7), edited by F. J. Edwards and S. M. Head, pp. 90–111, Pergamon, Oxford.
Winter, A., Z. Reiss, and B. Luz (1979), Distribution of living Coccolithophore assemblages
in the Gulf of Elat (’Aqaba), Mar. Micropaleontol., 4, 197–223.
Winter, A. (1982a), Paleoenvironmental interpretation of Quaternary coccolith assemblages
from the Gulf of Aqaba (Elat), Red Sea, Rev. Esp. Micropaleontol., XIV, 291–314.
Woelk, S., and D. Quadfasel (1996), Renewal of deep-water in the Red Sea during 1982–
1987, J. Geophys. Res., 101 (C8), 18155–18165.
Wyrtki, K. (1974), On the deep circulation of the Red Sea, La Formation des Eaux
Oceaniques Profondes: Centre National de la Recherche Scientifique, p. 91–106.
Yang, S., and P. J. Webster (1990), The effect of summer tropical heating on the location and
intensity of the extratropical westerly jet streams, J. Geophys. Res., 95, 18705–18721.
Ziv, B., U., Dayan, Y. Kushnir, C. Roth, and Y. Enzel (2006), Regional and global
atmospheric patterns governing rainfall in the southern Levant, Int. J. Clim., 26, 55–73.

Chapter 4: Late Holocene environmental changes in the northern Red Sea…
91
Ziveri, P., K.-H. Baumann, B. Böckel, J. Bollmann, and J. R. Young (2004), Biogeography of
selected Holocene coccoliths in the Atlantic Ocean in Coccolithophores: From
Molecular Processes to Global Impact, edited by H. Thierstein and J. R. Young, pp.
403–428, Springer, New York.

92

Chapter 5: Summary
93
5 Summary
Following the outline of the thesis, the basic findings are summarized in the order of the three
manuscripts. Finally, some general remarks concerning the calcareous nannoplankton
abundance patterns and the responsible background mechanisms are given.
1.) Covering the last 22,000 years, the calcareous nannoplankton record of core GeoB 5844-2
allows the identification of the Last Glacial Maximum, the Heinrich Event 1, the
Bølling/Allerød warm period, the Younger Dryas and the Holocene in the northern Red Sea.
Furthermore, the subdivision of the Holocene and the identification of two distinctive short-
term phases subsequent to the Bølling/Allerød warm period and the “Holocene Humid
period” are possible.
Extreme conditions during the hypersaline “aplanktic zone” are characterized by
Gephyrocapsa ericsonii and Emiliania huxleyi. High abundances of Gephyrocapsa ericsonii
during the Last Glacial Maximum point to moderately cool and fertile conditions, whereas the
dominance of Emiliania huxleyi during Heinrich Event 1 goes ahead with a further climatic
cooling favoring the bloom of opportunistic species. Punctuated by the Younger Dryas with
high values of Gephyrocapsa oceanica, a two-step onset of the postglacial humid climate
characterized the calcareous nannoplankton community. Both steps show an early
oligotrophic phase dominated by Florisphaera profunda and Gladiolithus flabellatus, and a
subsequent fertile phase characterized by Emiliania huxleyi. Following the Emiliania huxleyi-
dominated “Holocene Humid period”, the repetitive increases in abundance of F. profunda
and G. flabellatus reflect modern oligotrophic conditions accompanied by high aridity on
land.
2.) The glacial northern Red Sea represents a unique marine environment with regard to the
hypersaline “aplanktic zone”. The findings summarized above, however, indicate the presence
of calcareous nannoplankton during this period. Northern Red Sea environmental changes
were analyzed in the second study with special reference to the longer-term cooling cycles
and Heinrich events of the North Atlantic realm. The calcaroues nannoplankton record
obtained from core GeoB 5844-2 allows a precise and detailed correlation with the North
Atlantic climate trends. It is shown that the calcareous nannoplankton responded differently to
high latitudinal cooling before and after ca. 32 ka. A reiterated succession, closely correlated
with the longer-term cooling cycles and Heinrich events, is detected after 32 ka. The three
Heinrich events (H3, H2, and H1) are dominated by Emiliania huxleyi. Gephyrocapsa
oceanica and especially Gephyrocapsa ericsonii are abundant between H3 and H2 and

Chapter 5: Summary
94
between H2 and H1. Fluctuations in the abundance of Emiliania huxleyi, Gephyrocapsa
oceanica and Gephyrocapsa ericsonii are also documented before 32 ka. However, no
analogue succession as in the younger interval was found. The record presented here, and the
comparison with other microfossil data, led to the assumption that the glacial salinity increase
did not cause the documented calcareous nannoplankton succession. The calcareous
nannoplankton record points to a sensitive and fast acting mechanism linking high latitudinal
climate oscillations and environmental changes in the northern Red Sea region, especially
after 32 ka. The key mechanism seems to be the atmospheric teleconnection with the
extratropics and the associated hydrographic changes driven by a reinforced influence of cold
air. A strong influence from the mid-latitudes is evident since the late Marine Isotope Stage 3.
This approach to explain the calcareous nannoplankton abundance patterns is in good
agreement with climatic evolution of the northern subtropical desert belt.
3.) The third study focuses on the abundance patterns of Gephyrocapsa spp. (Gephyrocapsa
oceanica and Gephyrocapsa ericsonii). The response of Gephyrocapsa spp. to environmental
changes over the course of the last two millennia was analyzed. The documentation of
calcareous nannoplankton changes in late Holocene times corroborates the findings from core
GeoB 5844-2. Overall, the record from the Shaban Deep provides further evidence for the
linkage between the northern Red Sea and the mid-latitudes. The calcareous nannoplankton
record documents three intervals with high Gephyrocapsa spp. values in the northern Red
Sea. The Gephyrocapsa spp. abundance peaks seem to coincide with cold periods in Europe.
Reinforced convective mixing has probably triggered their increase. The changes of the
regional hydrography can be explained by a strengthened penetration of cold air from
northerly direction. The Gephyrocapsa spp. seem to be a sensitive indicator for the
hydrography of the northern Red Sea winter environment. Similar conditions, as documented
during the Gephyrocapsa spp.-maxima, are thought to be associated today with positive index
winters of the North Atlantic Oscillation/Artic Oscillation (NAO/AO). However, the
Gephyrocapsa spp. maxima in late Holocene times are probably inadequately explained on
the basis of the modern NAO/AO patterns. It is tentatively assumed here that the
Gephyrocapsa spp. record may reflect fluctuations in the strength and/or position of the
subtropical jet/development of the poleward branch of the Hadley cell.
In summary, this thesis represents the first well-dated, comprehensive calcareous
nannoplankton study from the northern Red Sea region. As remarkable as the glacial salinity
increase seems, the changes documented in core GeoB 5844-2 indicate that the calcareous
nannoplankton were less affected by variations of the salinity than other fossil groups. Other

Chapter 5: Summary
95
factors explain the observed distribution patterns probably better. Environmental factors
affecting phytoplankton distribution can interact in complex ways. The data from core GeoB
5844-2 point to variations of the water stratification and its influence on the nutrient
distribution within the photic zone as a crucial factor for the structure of calcareous
nannoplankton community. Closely coupled to the climate of the northern mid and high
latitudes, basic changes documented in the fossil assemblage compositions seem to be driven
by local hydrographic changes under the atmospheric influence from the extratropics. These
findings are consistent with modern observations, which illustrate the sensitivity of the
northern Red Sea hydrography to mid-latitude continental climate. The investigation of core
GeoB 7805-1 may reveal more about the atmospheric elements involved. One of the most
distinctive features of the northern Red Sea is its close position to the tropical-extratropical
atmospheric “boundary”. The results from core GeoB 7805-1 seem to indicate a crucial role
of the position and/or strength of the subtropical jet/the development of the poleward branch
of the Hadley cell on environmental changes in late Holocene times.

96

Chapter 6: Outlook
97
6 Outlook
The Near East-North African region represents a key area of human civilization where
important changes took place in prehistoric and historic time. Controlling the vegetation cover
and the water storage, temperature and especially precipitation changes in this dry part of the
world were key factors for human settlement and cultural development. Furthermore, the
availability of fresh water (Figure 6.1) is
seen as a crucial factor for the regional
development and the prevention of social
and military crises in the near future. Since
observational data cover only a restricted
time span, the study of paleoclimatic
archives is crucial for the understanding of
the climate system. Continuous and well-
dated records are needed for this attempt.
Unfortunately the arid environment with its
unprotected land surface limited the
availability of such records from the
terrestrial setting. One answer to this
problem is the use of records from nearby
marine sites. The study of microfossils and
certain geochemical-sedimentological data
can provide insights into the past oceanographic setting and hence into the climate history.
The calcareous nannoplankton community, and the study of its composition, represents one
fruitful attempt. It has been unequivocally shown here that this fossil group mirrors the
climatic trends of the last 60,000 years for the northern Red Sea region. The calcareous
nannoplankton data extended and improved past studies undertaken in this region: Fortunately
the calcareous nannoplankton seems to be less affected by the glacial salinity increase than
other marine organisms; even during hypersaline periods a detailed reconstruction of the
paleoceanographic conditions is possible. In order to update the analysis of past
environmental changes, more detailed plankton studies are needed. Better information on the
seasonal succession patterns of calcareous nannoplankton species, together with detailed
physical and chemical measurements, are probably an important requirement to refine the
explanatory power of the fossil records. Paleoceanographic studies of Red Sea sediments
older than Marine Isotope Stage 3 may help to gain deeper insights into the climatic evolution
Figure 6.1. Map showing the Red Sea area
and the water availability in 2000 (based on
information provided by the United Nations
Environment Programme).

Chapter 6: Outlook
98
of the African-Arabia desert belt. In addition, the accompanying investigation of marine
records from the Mediterranean Sea could be an important approach to get more detailed
information about the extratropical influence on northern Red Sea environmental changes and
the role of the North Atlantic Oscillation/Artic Oscillation (NAO/AO). The hypothesized role
of the subtropical jet in modulating marine environmental conditions in the northern Red Sea
region raises further questions with respect to its development and specific regional impact.
There is no final answer to the question of how variations of the sun are included. Direct
observations of how solar variations modulate the atmospheric structure are limited to short-
term effects. Furthermore, our knowledge about the specific role of the El Niño-Southern
Oscillation (ENSO) and the NAO/AO in influencing jet stream patterns on a longer time scale
is far from being complete.

99
Appendix

100

Methods
101
All nannoplankton samples were investigated using a
scanning electron microscope (SEM; LEO 1530 Gemini
FESEM). The filtration technique of Andruleit (Andruleit,
H., 1996, A filtration technique for quantitative studies of
coccoliths, Micropaleontol., 42, 403–406) was applied for
preparation. The freeze-dried sediment was weighted on a
high-precision balance, wet-separated with a rotary splitter
(FRITSCH Laborette 27; picture on the right hand) and
filtered by means of a vacuum pump on polycarbonate
filters. After drying at 40°C (2-3 hours), a wedge-shaped
piece of the filter was mounted on an aluminum stub with
self-adhesive tape, outlined with a silver colloidal suspension and coated with gold in argon
atmosphere using a sputter coater (BioRad SC-500). At least 300 specimens per sample were
identified and counted.
The absolute abundance was calculated with the following equation:
AA = (F x C x S)/(A x W)
AA = absolute abundance (number g-1
)
F = total sediment coated filter area (mm2)
C = number of counted coccoliths/nannoliths
S = split factor
A = investigated filter area (mm2)
W = weight of the dry sample (g)
The accumulation rates were calculated with the following equation:
AR = AA x SR x DD
AR = accumulation rate (number cm-2
kyr-1
)
SR = sedimentation rate (cm kyr-1
)
DD = dry bulk density (g cm-3
)

Methods
102
In addition, in order to control SEM preparation and to study general changes in the sediment
composition, all sample were studied with an OLPYMPUS BH-2 light microscope using
polarizing light at a magnification of x1250 and dark field at a magnification of x150. For
slide preparation please see Bown et al. (Bown, P. R., and J. R. Young, 1998, Techniques, in
Calcareous Nannofossil Biostratigraphy, edited by P. R. Bown, pp 16–28, Chapman and Hall,
London).

Taxonomy
103
Species identification followed the description of Kleijne (Kleijne, A., 1993, Morphology,
taxonomy and distribution of extant coccolithophorids (calcareous nannoplankton), Ph.D.
thesis, 320 pp., Vrije Universiteit, Amsterdam), Jordan and Kleijne (Jordan, R. W., and A.
Kleijne, 1994, A classification system for living coccolithophores, in Coccolithophores,
edited by A. Winter and W. G. Siesser, pp. 83–105, Cambridge Univ. Press, Cambridge) and
Young et al. (Young, J. R., M. Geisen, L. Cros, A. Kleijne, I. Probert, C. Sprengel, and J. B.
Ostergaard, 2003, A guide to extant coccolithophore taxonomy, J. Nannopl. Res. Spec. Iss., 1,
124 pp.).
Below an overview (up to the genus level) of the classification schema of Young et al.
(2003) is given:
Classification system
Order ISOCHRYSIDALES
Family Isochrysidaceae (non-calcifying)
Isochrysis
Chrysotila
Family Noelaerhabdaceae
Emiliania
Gephyrocapsa
Reticulofenestra
Order COCCOSPHAERALES
Family Coccolithaceae
Coccolithus
Cruciplacolithus
Family Calcidiscaceae
Calcidiscus
Hayaster
Oolithotus
Umbilicosphaera
Family Pleurochrysidaceae
Pleurochrysis
Jomonlithus
Family Hymenomonadaceae
Hymenomonas
Ochrosphaera

Taxonomy
104
Order ZYGODISCALES
Family Helicosphaeraceae
Helicosphaera
Family Pontosphaeraceae
Pontosphaera
Scyphosphaera
Order SYRACOSPHAERALES
Family Syracosphaeraceae
Calciopappus
Michaelsarsia
Ophiaster
Coronosphaera
Syracosphaera
Family Calciosoleniaceae
Calciosolenia
Alveosphaera
Family Rhabdosphaeraceae
Acanthoica
Algirosphaera
Anacanthoica
Cyrtosphaera
Discosphaera
Palusphaera
Rhabdosphaera
Saturnulus
HETEROCOCCOLITHS INCERTAE SEDIS, Heterococcolith families and genera incertae sedis
Family Alisphaeraceae
Alisphaera
Canistrolithus
(Polycrater)
Family Umbellosphaeraceae
Umbellosphaera
Narrow-rimmed placoliths
“Calyptrosphaera”
Tetralithoides
Turrilithus
Placorhombus
Family Papposphaeraceae
Papposphaera
Pappomonas

Taxonomy
105
Narrow-rimmed muroliths, Genera incertae sedis aff. Papposphaeraceae
Picarola
Vexillarius
Wigwamma
NANNOLITHS
Family Braarudosphaeraceae
Braarudosphaera
Family Ceratolithaceae
Ceratolithus
(Neosphaera)
Nannoliths incertae sedis
Florisphaera
Gladiolithus
Ericiolus
HOLOCOCCOLITHS [Calyptrosphaeraceae]

Stratigraphic framework
106
Age model of core GeoB 5844-2 used in the first manuscript/Chapter 2 (Arz, H. W., J.
Pätzold, P. J. Müller, and M. O. Moammar, 2003, Influence of Northern Hemisphere climate
and global sea level rise on the restricted Red Sea marine environment during termination I,
Paleoceanography, 18(2), 1053, doi:10.1029/2002PA000864):
Core Depth 14
C AMS Age Calibrated Age
(cm) (years BP) (calendar years BP)
0 1015 ±35 516
10 2490 ±80 2002
20 3920 ±45 3764
30 5325 ±50 5584
40 6320 ±100 6658
55 8525 ±55 8922
70 9990 ±50 10630
85 12030 ±55 13430
100 13120 ±60 14497
125 14960 ±120 17205
150 17030 ±90 19587
175 18580 ±100 21371
200 20540 ±120 23626
225 22090 ±160 25433

Stratigraphic framework
107
Age model of GeoB 5844-2 used in the second manuscript/Chapter 3 (Arz, H. W., F., Lamy,
A. Ganopolski, N. Nowaczyk, and J. Pätzold, 2007, Dominant Northern Hemisphere climate
control over millennial-scale glacial sea-level variability, Quat. Sci. Rev., 26, 312–321):
Core Depth 14
C AMS Age Calibrated Age (calendar years BP)
(cm) (years BP) CalPal Fairbanks
0 1015 ±35 510 ±30 505 ±19
10 2490 ±80 1870 ±100 1866 ±90
20 3920 ±45 3590 ±70 3589 ±60
30 5325 ±50 5470 ±90 5506 ±76
40 6320 ±100 6560 ±110 6546 ±118
55 8525 ±55 8820 ±120 8814 ±131
70 9990 ±50 10650 ±60 10653 ±67
85 12030 ±55 13360 ±80 13369 ±110
100 13120 ±60 14840 ±150 14745 ±130
125 14960 ±120 17680 ±160 17438 ±215
150 17030 ±90 19670 ±140 19581 ±116
175 18580 ±100 21700 ±130 21478 ±144
200 20540 ±120 23870 ±170 23927 ±151
225 22090 ±160 25720 ±310 25921 ±239
250 25110 ±210 29580 ±370 29179 ±325
275 26360 ±240 30660 ±220 30952 ±202
300 28890 ±290 32890 ±710 33005 ±313
325 32060 ±400 36500 ±400 36468 ±407
Paleomagnetic excursions
361 Laschamp event 41800 calendar years BP
417 – 49400 calendar years BP
517 Norwegian-Greenland-Sea event 66100 calendar years BP

Stratigraphic framework
108
Age model for the core GeoB 7805-1 (Shaban Deep) used in the third manuscript/Chapter 4
(Seeberg-Elverfeldt, I. A., J. Pätzold, H. W. Arz, and J.-B. W. Stuut, 2007, Late Holocene
climate variability in the northern Red Sea; submitted to Paleoceanography):
Core Core Depth 14
C AMS Age Cal. Age (cal. years)
Orig./trans. (cm) (years BP) BP AD
GeoB 7805-1 0,5/0,5 *725 ±25 175 1775
10/8,2 935 ±25 385 1565
19/9,7 1115 ±25 550 1400
25/15,7 1535 ±30 880 1070
35/25,7 1825 ±30 1200 750
45/35,7 2245 ±30 1630 320
GeoB 7803-2_2 10,5/10,5 *705 ±30 150 1800
38/16,7 1295 ±30 785 1165
52,2/29,7 1865 ±30 1230 720
GeoB 7804-4_1 7,5/7,5 870 ±25 345 1605
38/18,2 1570 ±25 945 1005
*Combined since both data points lie within their respective error range
Note: Since cores from the Shaban Deep show distinct lamination that can be used for cross-
correlation, 14
C-ages were transferred from other cores (GeoB 7803-2, GeoB 7804-4) to core
GeoB 7805-1 to improve the age model.

Chapter 2
Figure 2.3
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Abso
lute
abundan
ces
(no.g
-1 s
ed.)
Acc
um
ult
ion r
ates
(no. cm
-2 k
a-1)
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Abso
lute
abundan
ces
(no.g
-1 s
ed.)
Acc
um
ult
ion r
ates
(no. cm
-2 k
a-1)
1 665 1,99E+10 1,54E+11 90 13786 1,67E+10 1,57E+11
2 813 2,47E+10 1,91E+11 92 13928 1,46E+10 1,22E+11
4 1110 2,38E+10 1,84E+11 94 14070 1,35E+10 1,20E+11
6 1408 2,30E+10 1,88E+11 96 14212 1,26E+10 1,12E+11
8 1705 2,85E+10 2,44E+11 98 14355 1,13E+10 1,74E+11
10 2002 2,62E+10 2,05E+11 100 14497 6,63E+09 1,09E+11
12 2354 3,34E+10 2,05E+11 102 14714 6,89E+09 7,61E+10
14 2707 2,84E+10 1,70E+11 106 15147 6,57E+09 6,39E+10
16 3059 1,90E+10 1,17E+11 110 15580 6,30E+09 4,34E+10
18 3412 1,58E+10 1,03E+11 114 16013 6,69E+09 4,30E+10
20 3764 2,26E+10 1,44E+11 118 16447 5,94E+09 3,36E+10
22 4128 1,82E+10 1,14E+11 122 16880 3,76E+09 1,62E+10
24 4492 2,22E+10 1,40E+11 126 17300 3,26E+09 1,85E+10
26 4856 2,89E+10 1,88E+11 130 17681 2,15E+09 1,24E+10
28 5220 2,07E+10 1,35E+11 134 18063 2,45E+09 1,47E+10
30 5584 2,13E+10 1,37E+11 138 18444 2,24E+09 1,43E+10
32 5799 2,19E+10 2,47E+11 142 18825 2,59E+09 1,61E+10
34 6014 2,02E+10 2,09E+11 145 19111 2,50E+09 1,51E+10
36 6228 1,49E+10 1,58E+11 150 19587 3,06E+09 1,87E+10
38 6443 1,64E+10 1,73E+11 154 19872 2,62E+09 2,00E+10
40 6658 2,57E+10 2,65E+11 158 20158 2,16E+09 1,70E+10
42 6960 2,63E+10 1,95E+11 162 20443 2,46E+09 1,97E+10
44 7262 3,01E+10 2,18E+11 166 20729 2,40E+09 1,92E+10
46 7413 2,09E+10 1,56E+11 170 21014 2,12E+09 1,92E+10
48 7865 2,71E+10 1,99E+11 174 21300 2,60E+09 2,11E+10
50 8167 2,46E+10 1,68E+11 178 21642 3,06E+09 1,86E+10
52 8469 3,35E+10 2,36E+11 182 22002 2,27E+09 1,47E+10
54 8771 3,48E+10 2,35E+11
56 9036 2,84E+10 2,58E+11
58 9264 3,51E+10 3,14E+11
60 9492 2,44E+10 2,22E+11
62 9720 2,36E+10 2,06E+11
64 9948 2,65E+10 2,45E+11
66 10176 2,25E+10 2,02E+11
68 10404 2,63E+10 2,25E+11
70 10632 2,23E+10 1,69E+11
72 11005 1,53E+10 7,79E+10
74 11378 1,40E+10 7,03E+10
76 11751 1,58E+10 7,86E+10
78 12124 1,46E+10 6,66E+10
80 12497 1,43E+10 6,67E+10
82 12870 1,85E+10 8,08E+10
84 13243 1,71E+10 7,21E+10
86 13501 1,80E+10 1,74E+11
88 13643 1,94E+10 1,79E+11
109

Chapter 2
Figure 2.4 and 2.5C
ore
dep
th (
cm)
Cal
endar
age
(B.P
.)
Um
bil
icosp
haer
e sp
p. (%
)
Hel
icosp
haer
a c
art
eri
(%)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
Em
ilia
nia
huxl
eyi
(%)
Flo
risp
haer
a p
rofu
nda (
%)
Gla
dio
lith
us
flabel
latu
s (%
)
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Um
bil
icosp
haer
e sp
p. (%
)
Hel
icosp
haer
a c
art
eri
(%)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
Em
ilia
nia
huxl
eyi
(%)
Flo
risp
haer
a p
rofu
nda (
%)
Gla
dio
lith
us
flabel
latu
s (%
)
1 665 1,5 0,3 8,9 1,2 24,3 49,2 5,5 90 13786 0,0 0,7 7,5 2,4 47,3 17,0 12,9
2 813 1,1 0,3 5,7 4,2 25,2 40,8 11,6 92 13928 0,0 0,9 9,4 2,5 43,9 25,4 8,2
4 1110 2,9 0,3 5,1 1,6 19,9 52,1 8,7 94 14070 0,0 0,9 4,0 3,4 52,2 16,0 9,2
6 1408 2,5 0,6 8,4 1,2 23,0 50,6 5,3 96 14212 0,0 0,0 3,0 2,0 53,1 24,5 4,0
8 1705 1,0 0,0 9,9 3,0 19,1 46,7 9,2 98 14355 0,0 0,4 7,3 1,1 15,8 29,0 23,5
10 2002 1,5 0,0 8,2 2,9 23,1 40,9 15,5 100 14497 0,0 0,7 10,5 1,4 17,3 48,0 15,5
12 2354 0,6 0,0 8,9 4,6 19,6 41,3 18,3 102 14714 0,3 0,0 5,8 1,0 19,9 54,0 16,2
14 2707 2,2 0,0 8,8 4,4 21,3 40,0 16,9 106 15147 0,0 0,0 6,0 0,4 27,3 55,3 4,6
16 3059 1,3 0,0 6,9 3,3 24,3 39,0 18,4 110 15580 0,0 0,0 4,8 0,7 43,5 35,3 5,8
18 3412 1,0 0,3 6,6 2,8 27,3 39,9 14,3 114 16013 0,0 0,3 7,7 1,0 40,8 37,5 5,4
20 3764 1,9 0,0 7,0 5,7 23,5 48,3 4,8 118 16447 0,3 0,7 7,4 0,3 42,6 35,5 7,1
22 4128 1,2 0,3 7,8 3,4 27,0 38,8 11,2 122 16880 0,0 0,3 10,3 0,7 29,0 36,0 11,0
24 4492 2,8 0,0 6,5 1,8 25,5 40,0 10,5 126 17300 0,0 0,7 14,7 0,7 20,3 36,0 15,0
26 4856 1,2 0,3 7,2 3,2 21,4 44,6 10,4 130 17681 0,0 0,7 43,0 2,7 7,6 29,9 9,3
28 5220 1,7 0,0 6,6 3,6 29,1 40,7 8,6 134 18063 0,0 0,7 31,5 2,7 4,8 37,7 7,2
30 5584 1,9 0,3 5,2 1,6 28,5 48,3 6,2 138 18444 0,0 0,7 33,2 1,0 4,7 40,0 11,9
32 5799 1,9 0,0 6,9 1,2 30,6 41,5 8,7 142 18825 0,0 0,6 33,3 0,0 2,9 37,9 14,6
34 6014 1,9 0,0 5,1 1,9 29,4 44,3 11,1 145 19111 0,0 1,7 32,4 0,3 3,7 43,5 9,7
36 6228 1,9 0,6 4,5 3,2 35,2 42,6 6,5 150 19587 0,0 2,4 34,9 1,0 4,1 32,1 9,1
38 6443 1,6 0,3 4,5 1,9 33,9 40,3 5,8 154 19872 0,0 2,0 31,0 1,0 3,3 29,7 7,9
40 6658 1,2 0,0 9,2 4,0 21,0 37,9 13,6 158 20158 0,0 2,8 23,4 0,7 5,9 25,2 7,3
42 6960 1,5 0,0 10,2 1,5 26,5 29,3 14,5 162 20443 0,0 1,0 23,7 1,0 1,4 32,5 8,8
44 7262 0,4 0,4 7,0 3,2 25,7 28,5 17,2 166 20729 0,0 6,1 24,5 3,2 3,5 27,4 5,4
46 7413 1,6 0,3 7,5 3,3 32,1 34,0 12,8 170 21014 0,0 5,7 20,0 1,3 5,0 40,0 6,7
48 7865 2,1 0,7 8,3 3,1 28,1 37,2 9,0 174 21300 0,0 6,4 23,8 1,0 5,7 40,3 5,4
50 8167 0,7 0,3 6,2 1,3 35,8 37,8 10,4 178 21642 0,0 3,6 22,0 2,0 3,6 44,4 7,2
52 8469 1,3 0,3 6,9 4,6 33,0 33,3 10,4 182 22002 0,0 7,4 22,8 2,3 4,4 30,5 7,7
54 8771 0,3 0,0 7,6 2,9 32,6 33,9 8,9
56 9036 2,4 0,3 5,4 1,4 30,1 38,2 13,2
58 9264 0,0 0,0 8,3 3,3 24,3 37,3 16,3
60 9492 1,3 0,0 2,7 2,0 28,9 38,0 18,5
62 9720 1,7 0,0 4,8 3,1 25,9 39,7 19,0
64 9948 1,4 0,0 4,5 2,4 27,7 38,8 20,1
66 10176 0,0 0,3 3,7 1,7 26,9 41,9 16,6
68 10404 0,6 0,0 3,5 1,9 18,7 41,8 24,7
70 10632 0,7 0,0 3,7 1,7 26,5 38,9 22,1
72 11005 0,3 0,6 4,6 0,9 38,7 35,3 12,7
74 11378 0,4 0,0 3,9 5,0 40,2 30,2 12,9
76 11751 0,0 2,4 2,8 4,5 30,0 34,9 13,3
78 12124 0,0 0,0 4,3 8,9 34,2 35,2 10,9
80 12497 0,3 0,7 2,1 9,6 27,8 31,9 18,5
82 12870 0,0 0,4 4,2 7,1 37,0 20,5 9,9
84 13243 0,0 1,3 2,4 6,4 28,3 35,4 14,5
86 13501 0,0 0,7 6,9 2,3 43,2 30,4 5,2
88 13643 0,0 1,0 4,0 3,7 40,8 32,0 9,8
110

Chapter 3
Figure 3.3
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Em
ilia
nia
huxl
eyi
(%)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
Abso
lute
abundan
ces
(no.g
-1 s
ed.)
Acc
um
ula
tion r
ates
(no.c
m-2 k
a-1)
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Em
ilia
nia
huxl
eyi
(%)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
Abso
lute
abundan
ces
(no.g
-1 s
ed.)
Acc
um
ula
tion r
ates
(no.c
m-2 k
a-1)
96 14335 53,1 3,0 2,0 1,26E+10 8,11E+10 270 30384 44,3 13,8 0,9 9,45E+09 1,68E+11
98 14533 15,8 7,3 1,1 1,13E+10 1,26E+11 274 30557 53,9 12,7 1,0 7,94E+09 1,25E+11
100 14730 17,3 10,5 1,4 6,63E+09 7,86E+10 278 30889 61,9 8,8 1,0 9,99E+09 8,18E+10
102 14967 19,9 5,8 1,0 6,89E+09 7,26E+10 282 31275 75,2 4,1 2,5 5,69E+09 4,73E+10
106 15440 27,3 6,0 0,4 6,57E+09 6,09E+10 286 31660 52,1 7,8 7,8 5,44E+09 4,84E+10
110 15914 43,5 4,8 0,7 6,30E+09 4,14E+10 290 32046 49,3 6,6 15,6 6,01E+09 5,24E+10
114 16388 40,8 7,7 1,0 6,69E+09 4,10E+10 294 32432 20,5 19,1 21,1 5,49E+09 4,31E+10
118 16861 42,6 7,4 0,3 5,94E+09 3,20E+10 298 32817 40,4 9,9 6,0 6,21E+09 5,00E+10
122 17335 29,0 10,3 0,7 3,76E+09 1,55E+10 302 33299 37,5 12,2 2,2 7,55E+09 4,26E+10
126 17776 20,3 14,7 0,7 3,26E+09 2,21E+10 306 33876 16,9 4,8 16,9 3,96E+08 2,27E+09
130 18118 7,6 43,0 2,7 2,15E+09 1,48E+10 310 34454 28,4 8,4 6,9 7,60E+09 4,49E+10
134 18460 4,8 31,5 2,7 2,45E+09 1,76E+10 314 35032 35,7 15,4 7,2 7,68E+09 4,50E+10
138 18803 4,7 33,2 1,0 2,24E+09 1,71E+10 318 35609 43,4 9,9 4,3 7,84E+09 4,59E+10
142 19145 2,9 33,3 0,0 2,59E+09 1,92E+10 322 36187 40,5 7,2 8,9 7,37E+09 4,44E+10
145 19402 3,7 32,4 0,3 2,50E+09 1,81E+10 326 36764 48,9 5,9 12,1 6,12E+09 3,95E+10
150 19830 4,1 34,9 1,0 3,06E+09 2,20E+10 330 37339 36,1 6,5 20,6 6,97E+09 4,50E+10
154 20147 3,3 31,0 1,0 2,62E+09 1,76E+10 334 37915 30,7 14,1 15,7 8,64E+09 5,34E+10
158 20464 5,9 23,4 0,7 2,16E+09 1,49E+10 338 38491 37,7 8,9 10,1 1,03E+10 6,23E+10
162 20780 1,4 23,7 1,0 2,46E+09 1,73E+10 342 39066 32,8 10,3 3,2 8,09E+09 4,99E+10
166 21097 3,5 24,5 3,2 2,40E+09 1,69E+10 346 39642 28,3 10,4 8,1 1,00E+10 6,29E+10
170 21414 5,0 20,0 1,3 2,12E+09 1,69E+10 350 40217 40,3 7,9 12,1 6,20E+09 3,44E+10
174 21731 5,7 23,8 1,0 2,60E+09 1,85E+10 354 40793 29,2 14,9 13,2 7,19E+09 3,88E+10
178 22063 3,6 22,0 2,0 3,06E+09 1,93E+10 358 41368 31,3 12,4 21,5 6,81E+09 4,35E+10
182 22401 4,4 22,8 2,3 2,27E+09 1,53E+10 362 41936 39,0 6,5 9,4 6,07E+09 3,97E+10
186 22738 4,9 27,8 4,9 2,48E+09 1,50E+10 366 42479 32,4 10,2 15,2 6,71E+09 5,10E+10
190 23076 5,5 30,7 9,3 3,07E+09 2,09E+10 370 43021 26,9 12,8 20,2 5,52E+09 4,15E+10
194 23414 3,2 25,0 18,8 3,11E+09 2,12E+10 374 43564 22,4 19,8 9,4 8,57E+09 6,39E+10
198 23751 7,2 18,8 21,6 3,11E+09 2,02E+10 378 44107 21,7 14,0 13,7 1,12E+10 8,41E+10
202 24071 6,1 28,2 4,4 3,10E+09 2,69E+10 382 44650 12,5 9,9 18,6 7,02E+09 5,68E+10
206 24374 10,7 21,3 2,7 3,44E+09 3,31E+10 386 45193 11,9 12,2 19,2 1,14E+10 8,55E+10
210 24676 12,6 30,1 2,0 3,01E+09 2,36E+10 390 45736 19,3 8,0 17,6 8,27E+09 6,22E+10
214 24978 22,1 24,8 0,7 2,67E+09 1,98E+10 394 46279 26,1 12,0 4,9 1,37E+10 7,94E+10
218 25281 31,3 16,3 0,7 4,91E+09 3,83E+10 398 46821 19,6 11,9 23,1 1,10E+10 8,39E+10
222 25583 43,8 19,0 0,7 4,85E+09 3,76E+10 402 47364 19,9 10,4 21,5 1,07E+10 8,59E+10
226 25958 43,4 8,4 1,0 4,35E+09 1,85E+10 406 47907 23,6 6,5 17,5 5,81E+09 4,76E+10
230 26552 37,3 9,3 1,7 3,80E+09 1,32E+10 410 48450 14,9 10,5 15,9 1,03E+10 8,30E+10
234 27146 8,9 43,4 1,4 3,17E+09 1,12E+10 414 48993 13,5 19,3 10,8 1,14E+10 8,95E+10
238 27739 14,4 33,4 1,6 3,60E+09 1,41E+10 418 49568 22,9 7,6 12,7 1,17E+10 7,73E+10
242 28333 15,9 28,0 2,4 4,20E+09 1,78E+10 422 50238 25,7 13,1 14,7 1,20E+10 7,60E+10
245 28778 29,3 29,0 3,8 3,74E+09 1,62E+10 426 50908 34,1 10,1 10,1 1,29E+10 8,20E+10
250 29520 31,2 18,8 10,5 5,97E+09 2,69E+10 430 51578 25,9 11,6 11,0 1,00E+10 6,46E+10
254 29693 34,8 18,5 2,3 5,70E+09 9,54E+10 434 52248 17,8 27,9 8,7 1,32E+10 7,87E+10
258 29866 40,7 14,3 5,5 5,95E+09 9,02E+10 438 52918 19,3 17,8 13,2 1,12E+10 7,25E+10
262 30038 40,9 9,7 3,5 7,27E+09 8,34E+10 442 53588 21,5 19,6 11,7 9,24E+09 5,93E+10
266 30211 33,7 14,4 2,2 6,96E+09 7,93E+10 446 54258 18,4 16,8 10,8 9,48E+09 6,40E+10
111

Chapter 3
Figure 3.3
Core
dep
th (
cm)
Cal
endar
age
(ka
B.P
.)
Em
ilia
nia
huxl
eyi
(%)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
Abso
lute
abundan
ces
(no.g
-1 s
ed.)
Acc
um
ula
tion r
ates
(no.c
m-2 k
a-1)
450 54928 29,0 14,4 10,9 9,68E+09 6,00E+10
454 55598 20,3 30,5 8,5 1,36E+10 9,00E+10
458 56268 20,9 23,2 12,2 1,02E+10 6,45E+10
462 56938 30,6 12,3 11,7 1,06E+10 6,49E+10
466 57608 31,9 12,6 8,8 1,11E+10 7,14E+10
470 58278 29,1 12,9 12,3 1,01E+10 6,45E+10
474 58948 31,6 26,5 5,8 1,10E+10 6,88E+10
478 59618 33,1 24,8 5,1 7,42E+09 4,66E+10
112

Chapter 3
Figure 3.5
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Flo
risp
haer
a p
rofu
nda
(%
)
Core
dep
th (
cm)
Cal
endar
age
(B.P
.)
Em
ilia
nia
huxl
eyi
(%)
96 14335 24,5 270 30384 27,4
98 14533 29,0 274 30557 13,6
100 14730 48,0 278 30889 18,9
102 14967 54,0 282 31275 10,2
106 15440 55,3 286 31660 24,6
110 15914 35,3 290 32046 21,2
114 16388 37,5 294 32432 24,2
118 16861 35,5 298 32817 30,5
122 17335 36,0 302 33299 34,1
126 17776 36,0 306 33876 22,9
130 18118 29,9 310 34454 42,2
134 18460 37,7 314 35032 25,9
138 18803 40,0 318 35609 32,9
142 19145 37,9 322 36187 34,2
145 19402 43,5 326 36764 22,3
150 19830 32,1 330 37339 21,3
154 20147 29,7 334 37915 26,6
158 20464 25,2 338 38491 27,8
162 20780 32,5 342 39066 38,3
166 21097 27,4 346 39642 40,7
170 21414 40,0 350 40217 31,1
174 21731 40,3 354 40793 22,4
178 22063 44,4 358 41368 23,8
182 22401 30,5 362 41936 36,1
186 22738 28,8 366 42479 29,2
190 23076 24,1 370 43021 28,3
194 23414 18,5 374 43564 36,7
198 23751 17,1 378 44107 30,4
202 24071 22,4 382 44650 44,6
206 24374 27,1 386 45193 41,3
210 24676 35,4 390 45736 41,2
214 24978 20,7 394 46279 40,2
218 25281 19,2 398 46821 27,2
222 25583 14,7 402 47364 31,3
226 25958 25,6 406 47907 40,5
230 26552 32,3 410 48450 43,8
234 27146 16,7 414 48993 30,4
238 27739 28,9 418 49568 25,4
242 28333 23,8 422 50238 30,9
245 28778 15,6 426 50908 24,3
250 29520 18,5 430 51578 30,8
254 29693 14,9 434 52248 26,5
258 29866 26,7 438 52918 32,8
262 30038 31,8 442 53588 30,9
266 30211 34,6 446 54258 29,4
113

Chapter 3
Figure 3.5
Core
dep
th (
cm)
Cal
endar
age
(ka
B.P
.)
Flo
risp
haer
a p
rofu
nda
(%
)
450 54928 26,8
454 55598 21,6
458 56268 21,9
462 56938 25,3
466 57608 30,0
470 58278 27,0
474 58948 24,5
478 59618 20,9
114

Chapter 4
Figure 4.3, 4.4 and 4.5
Core
dep
th (
cm)
Cal
endar
age
(A.D
.)
Cal
endar
age
(B.P
.)
Gep
hyr
oca
psa
spp. ab
solu
te
abundan
ces
(no.g
-1 s
ed.)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
G. sp
p.
— T
OT
AL
(%
)
Em
ilia
nia
huxl
eyi
(%)
G. sp
p.
— E
. huxl
eyi
(%)
0,25 1820 130 9,66E+08 1,9 4,7 6,6 48,1 12,0
0,75 1804 146 1,65E+09 4,4 6,2 10,6 39,6 21,1
1,25 1788 162 5,76E+08 0,3 2,8 3,1 31,5 8,9
1,75 1773 178 8,18E+08 0,5 4,3 4,8 31,2 13,4
2,25 1757 193 8,64E+08 1,4 3,4 4,9 33,5 12,7
2,75 1741 209 5,74E+08 1,0 3,8 4,8 35,6 11,8
3,25 1725 225 8,25E+08 2,3 3,6 6,0 30,5 16,4
3,75 1709 241 1,15E+09 3,6 4,5 8,1 36,7 18,1
4,25 1694 257 1,44E+09 2,9 5,1 7,9 35,9 18,1
4,75 1678 272 7,79E+08 1,5 5,1 6,6 26,0 20,2
5,25 1662 288 1,30E+09 2,6 3,9 6,5 35,6 15,4
5,75 1646 304 9,00E+08 3,2 5,7 8,8 30,6 22,4
6,25 1630 320 9,95E+08 1,6 6,9 8,5 24,3 26,0
6,75 1614 336 7,04E+08 2,8 3,4 6,2 34,2 15,3
7,25 1598 353 2,74E+09 6,0 3,5 9,4 38,4 19,7
7,75 1593 358 9,09E+08 2,0 5,9 7,9 36,0 18,0
8,25 1588 363 1,23E+09 4,6 5,2 9,9 25,3 28,1
8,75 1583 368 1,19E+09 6,8 4,8 11,6 32,3 26,5
9,25 1578 373 5,35E+08 2,0 2,7 4,7 29,2 13,7
9,75 1573 378 1,89E+09 4,8 5,1 9,9 30,7 24,4
10,25 1559 391 7,27E+09 10,8 5,4 16,2 37,6 30,1
10,75 1550 400 3,68E+09 7,8 3,7 11,5 38,5 23,0
11,25 1542 408 6,63E+08 3,4 5,6 9,1 46,9 16,2
11,75 1534 416 2,63E+09 7,2 7,2 14,4 28,1 33,8
12,25 1525 425 2,35E+09 8,6 2,4 11,0 36,1 23,4
12,75 1517 433 2,46E+09 7,4 4,8 12,2 30,9 28,3
13,25 1509 441 1,86E+09 5,3 3,9 9,3 32,0 22,4
13,75 1500 450 2,61E+09 10,0 4,4 14,3 25,9 35,7
14,25 1492 458 1,53E+09 7,1 3,8 10,9 31,2 25,9
14,75 1484 466 2,55E+09 7,7 3,8 11,5 35,5 24,5
15,25 1475 475 8,02E+08 1,6 3,6 5,2 36,4 12,5
15,75 1467 483 1,49E+09 3,8 4,4 8,1 37,2 17,9
16,25 1459 491 1,42E+09 3,3 5,1 8,4 25,4 24,8
16,75 1451 499 1,71E+09 4,0 4,6 8,6 33,0 20,7
17,25 1442 508 2,55E+09 9,6 3,5 13,2 28,9 31,3
17,75 1434 516 1,49E+09 3,2 3,4 6,6 34,2 16,2
18,25 1426 524 1,28E+09 3,9 4,5 8,3 24,4 25,5
18,75 1417 533 2,48E+09 8,2 3,4 11,6 29,0 28,6
19,25 1388 562 1,37E+09 3,0 3,0 5,9 29,3 16,8
19,75 1358 592 8,16E+08 2,3 2,3 4,6 39,3 10,5
20,25 1329 621 2,08E+09 3,6 5,2 8,8 34,9 20,1
20,75 1299 651 9,10E+08 2,0 2,6 4,6 37,4 10,9
21,25 1270 680 1,37E+09 3,1 2,5 5,6 34,4 14,1
21,75 1240 710 1,30E+09 2,8 2,8 5,6 34,5 14,1
22,25 1211 739 1,10E+09 3,3 3,3 6,5 38,7 14,5
115

Chapter 4
Figure 4.3, 4.4 and 4.5
Core
dep
th (
cm)
Cal
endar
age
(A.D
.)
Cal
endar
age
(B.P
.)
Gep
hyr
oca
psa
spp. ab
solu
te
abundan
ces
(no.g
-1 s
ed.)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
G. sp
p.
— T
OT
AL
(%
)
Em
ilia
nia
huxl
eyi
(%)
G. sp
p.
— E
. huxl
eyi
(%)
22,75 1181 769 1,42E+09 2,6 5,5 8,1 37,5 17,8
23,25 1154 796 1,15E+09 3,0 4,5 7,5 36,1 17,2
23,75 1130 820 1,97E+09 3,2 4,4 7,6 31,6 19,4
24,25 1105 845 1,44E+09 3,2 3,8 6,9 31,2 18,2
24,75 1081 869 5,08E+08 0,7 2,6 3,3 26,3 11,1
25,25 1065 885 9,95E+08 3,1 3,7 6,9 32,4 17,5
25,75 1058 892 1,67E+09 4,0 5,0 9,0 31,6 22,1
26,25 1051 899 3,21E+09 10,9 3,4 14,3 27,2 34,5
26,75 1044 906 1,31E+09 5,6 3,3 8,9 24,7 26,4
27,25 1037 913 7,99E+08 2,4 4,0 6,4 36,2 15,0
27,75 1030 920 8,51E+08 1,7 2,6 4,3 27,4 13,5
28,25 1023 927 1,45E+09 2,5 4,3 6,7 24,2 21,8
28,75 1016 934 7,65E+08 2,1 3,0 5,2 27,3 15,9
29,25 1009 941 9,68E+08 4,3 2,0 6,2 29,2 17,6
29,75 994 956 2,53E+09 6,0 2,6 8,7 34,6 20,0
30,25 970 980 1,48E+09 4,5 4,8 9,3 36,1 20,4
30,75 947 1003 2,30E+09 5,6 1,8 7,4 34,0 17,9
31,25 923 1027 8,12E+08 3,1 1,8 4,9 35,9 12,0
31,75 900 1050 1,41E+09 2,9 5,1 8,0 36,0 18,2
32,25 876 1074 1,67E+09 6,3 2,8 9,1 34,9 20,7
32,75 853 1097 1,17E+09 4,2 4,9 9,2 48,4 15,9
33,25 829 1121 1,66E+09 5,1 4,1 9,2 37,5 19,7
33,75 806 1144 1,39E+09 2,2 3,8 6,1 36,9 14,2
34,25 782 1168 1,24E+09 1,9 4,5 6,4 30,5 17,4
34,75 759 1191 1,06E+09 1,6 3,8 5,4 35,2 13,3
35,25 745 1205 1,08E+09 1,8 4,2 6,0 29,5 16,9
35,75 740 1210 1,38E+09 3,8 1,6 5,4 29,8 15,5
36,25 736 1214 1,99E+09 3,3 3,3 6,5 32,4 16,8
36,75 731 1219 1,44E+09 5,0 2,2 7,2 32,1 18,4
37,25 727 1224 7,11E+08 3,4 1,2 4,6 25,6 15,3
37,75 722 1228 2,35E+09 8,0 2,4 10,3 33,6 23,5
38,25 718 1233 1,08E+09 3,4 4,0 7,3 36,6 16,7
38,75 691 1260 9,57E+08 1,0 6,7 7,7 33,0 18,9
39,25 661 1289 1,50E+09 5,6 2,7 8,3 31,8 20,7
39,75 632 1318 1,61E+09 5,9 2,2 8,1 33,1 19,7
40,25 602 1348 1,94E+09 5,2 2,6 7,8 38,0 17,0
40,75 573 1377 3,19E+09 12,1 2,1 14,2 26,3 35,0
41,25 543 1407 3,27E+09 7,2 5,7 12,9 19,8 39,4
41,75 514 1436 1,32E+09 5,1 2,9 8,0 31,4 20,3
42,25 484 1466 1,27E+09 7,4 4,2 11,6 27,3 29,8
42,75 455 1495 2,74E+09 11,7 5,8 17,5 27,8 38,6
43,25 426 1525 3,32E+09 11,7 4,2 16,0 25,1 38,9
43,75 396 1554 2,55E+09 10,3 3,0 13,3 23,9 35,8
44,25 367 1583 1,90E+09 5,8 3,7 9,5 25,5 27,2
44,75 337 1613 1,22E+09 5,9 2,6 8,5 37,7 18,4
116

Chapter 4
Figure 4.3, 4.4 and 4.5
Core
dep
th (
cm)
Cal
endar
age
(A.D
.)
Cal
endar
age
(B.P
.)
Gep
hyr
oca
psa
spp. ab
solu
te
abundan
ces
(no.g
-1 s
ed.)
Gep
hyr
oca
psa
eri
csonii
(%
)
Gep
hyr
oca
psa
oce
anic
a (
%)
G. sp
p.
— T
OT
AL
(%
)
Em
ilia
nia
huxl
eyi
(%)
G. sp
p.
— E
. huxl
eyi
(%)
45,25 308 1642 1,94E+09 5,9 6,3 12,2 28,1 30,2
45,75 278 1672 1,72E+09 7,9 2,6 10,5 31,5 25,0
46,25 249 1701 1,93E+09 5,2 4,3 9,6 37,3 20,4
46,75 220 1731 1,69E+09 5,5 5,5 10,9 34,1 24,3
47,25 190 1760 2,60E+09 7,4 6,5 13,9 27,2 33,8
47,75 161 1789 2,66E+09 8,0 6,8 14,8 23,8 38,3
117

118

119
Curriculum Vitae
Personal data
Name: Heiko-Lars Legge
Date of birth: 26. January 1973
Place of birth: Bochum
Marital status: single
Nationality: German
School education
1979 – 1983 Gemeinschaftsgrundschule, Alte Wittener Str. 19, Bochum
1983 – 1992 Erich Kästner-Schule, Markstr. 189, Bochum
“Compulsory community service” (Ziviler Ersatzdienst)
1992 – 1993 Behinderten-Schule, Bochum-Langendreer
University education and scientific activity
1994 – 2002 Study of biology at the Department of Biology, Ruhr-University
Bochum
Diploma-Thesis (“Die Coccolithophoriden des Roten Meeres:
Ökologie und Biogeographie der letzten 24.000 Jahre”) at the
department of Geology, Mineralogy and Geophysics, Ruhr-
University Bochum.
2002 Participant Meteor cruise M52/3 (Red Sea)
2003 – 2007 Ph.D. student and research assistant at the department of
Geology, Mineralogy and Geophysics, Ruhr-University Bochum