Embed Size (px)
Citation preview

42 | Biol. Unserer Zeit | 1/2007 (37) © 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Montanas galt als besonders ursprünglich. Diese Einschät-zung wird jetzt oft in Frage gestellt, weil die Plesiadapifor-mes im Schädelbau und Gebiss im Allgemeinen ausge-prägte Spezialisierungen aufweisen, die teilweise an dievon Nagetieren erinnern und die nur schwer mit der An-nahme vereinbar sind, dass sie die Grundgruppe der Pri-maten sind. Möglicherweise sind sie eine Schwester-gruppe der Primaten, die gemeinsame anatomische Primi-tivmerkmale aufweist und die der Konkurrenz mit denechten Nagetieren nicht standhielt und im Eozän ausstarb[4, 8].
Eozän: die Wiege der „höheren Affen“Unbestrittene echte Primaten gibt es in der Fossildoku-mentation erst ab dem Eozän [7, 8, 11]. Sie besaßen bei-spielsweise alle eine verkürzte Schnauze, die Augen wan-derten zunehmend nach vorn (binoculares, stereosko-pisches Sehen) und die � Orbita besaß hinten eineKnochenspange. In den vergangenen Jahren wurde eineganze Reihe von eozänen Skelettresten gefunden, dieschon den höheren Primaten (Simiiformes = Simiae = An-thropoidea) zuzuordnen sind und die sehr wahrscheinlichdie Grundgruppe der Neuweltaffen (Platyrrhini) und derAltweltaffen (Catarrhini) bilden.
Aus dem frühen oder mittleren Eozän Algeriens kenntman die kleinen Formen Algeripithecus und Tabelia(Zähne und Kieferreste), aus dem mittleren Eozän Chinasden kleinen Eosimias (Abbildung 1, Kiefer, Zähne undTeile des Fußskeletts). Aus Ablagerungen des mittleren Eo-zäns Burmas stammen Bahinia, Pondaungia, Amphi-pithecus (Zähne, Kiefer und Fußknochen). In Ablagerun-gen des späten Eozäns Fayums wurden Serapia, Arsinoea,Simonsius und Proteopithecus gefunden (Zähne, Kiefer-fragmente, teilweise Extemitätenknochen).
Neben einem ursprünglichen Muster der � Molarenfällt auf, dass diese Formen noch drei � Prämolaren be-saßen. Somit kommen sie – zusammen mit den Parapithe-cidae (s.u.) – auch noch als Vorfahren der südamerikani-schen Affen infrage. Es bleibt bei der Fundlage unent-schieden, ob die Wiege der Simiiformes, also der„höheren“ Affen der Neuen und der Alten Welt in Ostasienoder Nordafrika stand; die Mehrheit der etwas älterenFunde stammt aus Asien.
In den vergangenen Jahren wurden zahlreiche neue Fossilienvon Hominiden und anderen Primaten gefunden, was Anlasszu phantasievollen Spekulationen war und ist. Absicht dieseszweiteiligen Beitrages ist, sachbezogen und kritisch zu denaktuellen Erkenntnissen Stellung zu nehmen. Die vielenneuen Fossilfunde und biochemischen Befunde tragen zwareinerseits dazu bei, unsere Vorstellungen vom Ablauf der Evolution des Menschen zu präzisieren, sind aber andererseitseine Quelle neuer Fragen.
Wie aus den ersten Primaten Homo wurde (Teil 1):
Die Fossilgeschichte des MenschenULRICH WELSCH
DOI:10.1002/biuz.200610325
A B B . 1 Darstellung des eozänen primitiven Simiers Eosimias ausChina. Nach Beard et al. (1994).
Bereits über den Beginn der Entwicklung der Primatenbesteht Uneinigkeit: In der Vergangenheit wurden die
im Paleozän Nordamerikas und Europas verbreiteten undoffensichtlich erfolgreichen Plesiadapiformes oft als ältes-te und ursprünglichste Primaten angesehen. Die zu ihnengehörende Gattung Purgatorius aus dem frühen Paleozän

P R I M A T E N F O S S I L I E N , 1 . T E I L | PA L Ä O N TO LO G I E
Da im Eozän Afrika und Südamerika bereits durch den ei-nige hundert Kilometer breiten Atlantik getrennt waren,müssen die Primaten über damals in aller Wahrscheinlich-keit noch vorhandene Inselketten und mit Hilfe treiben-der floßähnlicher Ansammlungen von Bäumen nach Süda-merika gelangt sein. Sie sind erst seit dem Oligozän in derNeuen Welt nachgewiesen. Die Theorie, dass sich diesüdamerikanischen Affen parallel zu den Affen der AltenWelt aus Halbaffen, die in Europa und in Nordamerika vor-kamen, entwickelt haben, erscheint heute eher unwahr-scheinlich.
Oligozäne PrimatenWichtige neue Primatenfunde aus dem Oligozän wurdenin jüngerer Zeit in der Oase Fayum in Ägypten gemacht.Diese ergänzen die älteren Fayum-Funde und lassen sichdrei Gruppen zuordnen, den Parapithecidae, den Proplio-pithecidae und den Oligopithecidae [11].
Die Parapithecidae (Parapithecus, Apidium und Qa-trania sowie vielleicht Biretia) werden oft – wie die obengenannten eozänen Formen – in die Grundgruppe aller Si-miiformes gestellt. Diese Einschätzung beruht auf zahlrei-chen Funden (Schädel, Kiefer, Zähne und postcranialeSkelettelemente). Auch diese Tiere besaßen noch drei Prä-molaren und kommen deswegen und auch aufgrund an-derer Merkmale des Schädels und im postcranialen Skelettebenfalls als Vorfahren nicht nur der Altweltaffen, sondernauch der Neuweltaffen infrage.
Miozän: erste deutliche Ähnlichkeiten mit rezenten Menschenaffen
Aus dem Miozän Afrikas, Europas und Asiens wurden inden vergangenen Jahren viele neue fossile Menschenaffennachgewiesen, von denen einige der Familie Hominidaezugeordnet werden [3, 6, 11]. Diese Formen kommen teil-weise als Vorfahren der Homininae und Ponginae infrage.Hier sollen besonders interessante Arten kurz geschildertwerden:
Morotopithecus bishopi war ein Menschenaffe, dervor circa 20 Millionen Jahren in Ostafrika lebte [12]. Schä-del- und postcraniale Skelettreste dieser Art wurden 1997in Uganda gefunden. Morotopithecus wog vermutlich um50 kg. Eine morphologische Analyse seiner Rückenwirbeldeutet darauf hin, dass diese Form gut klettern konnte.Die Gesamtanalyse der Skelettanatomie weist eine ganzeReihe deutlicher Gemeinsamkeiten mit den großen rezen-ten afrikanischen Menschenaffen (und damit zu derGruppe, zu der auch der Mensch gehört) auf. Aufgrunddes hohen Alters und der Anatomie ist vermutet worden,dass Morotopithecus an der Basis aller Hominidae steht. Esist aber auch denkbar, dass er sogar in die Vorfahrenlinieder Homininae (Gorilla, Pan, Homo) zu stellen ist.
Dryopithecus wurde im späten Miozän Europas undanderswo gefunden und wird von manchen Autoren alsfrüher Hominide – oder sogar Hominine – angesehen [3,11]. 1999 wurde ein weitgehend vollständiger Schädel in
Ungarn gefunden, der diese Vermutung stützt. Dryopithe-cus war ein relativ großer Menschenaffe, mit Zähnen, diesehr an die des Gorillas erinnern. Er ernährte sich ver-mutlich von weichen Früchten, Knospen und Blättern.Sein Gehirn hatte dieselbe Größe wie das eines Schim-pansen. Wie moderne und andere miozäne Menschenaf-fen bewegte er sich wahrscheinlich hangelnd durch dasGeäst größerer Bäume. Wahrscheinlich gab es mehrere Ar-ten, die von Spanien bis in die Türkei und Georgien sowievermutlich bis China und Indien verbreitet waren. Fundein Spanien erhielten teilweise die Bezeichnung Hispano-pithecus. Auch in Deutschland wurden Dryopithecus-Zähne gefunden. Weitere große Menschenaffen aus demMiozän Europas sind Graecopithcus (=Ouranopithecus)und der erst im Jahr 2004 von Moya und Mitarbeitendenbeschriebene Pierolapithecus.
Gut erhaltenes Gesichtsschädelmaterial von Siva-pithecus aus dem Miozän (17 bis 14 Millionen Jahre alt)des Siwalik-Berglandes Pakistans und Nordindiens zeigteindeutige Übereinstimmungen mit dem Schädel des mo-dernen Orang Utan (Pongo), so dass an einer Verwandt-schaft der beiden Menschenaffen kaum zu zweifeln ist.
© 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2007 (37) | Biol. Unserer Zeit | 43
Die mit einem grünen Pfeil markierten Begriffewerden im Glossarauf Seite 47 erklärt.
Ü B E R B L I C K Ü B E R DA S S YS T E M D E R PR I M AT E N
Primaten gliedern sich in:
A. Plesiadapiformes (rein fossil, fragliche Primaten)
B. Strepsirrhini 1. Lorisiformes, Afrika, Süd- und Südostasien:
Galagos (Buschbabies), Loris, Potto, Bärenmaki2. Chiromyiformes, Madagaskar:
Fingertier (Daubentonia, Aye-Aye)3. Lemuriformes, Madagaskar:
Katzenmakis, Wieselmakis, Indris, Lemuren u.a.
C. Haplorrhini 4. Tarsiiformes, Südostasien:
Kobold- = Gespenstmakis (Tarsius)5. Simiiformes (Anthropoidea), Alte und Neue Welt:
a. Platyrrhini (Neuweltaffen)Kapuzineraffen, Krallenäffchen, Nachtaffen
b. Catarrhini (Altweltaffen)I Cercopithecoidea Meerkatzen, Makaken, Paviane, Languren u.a.II Hominoidea
Familie Pliopithecidae +Familie Oreopithecidae +Familie Hylobatidae (Gibbons)Famlilie Proconsulidae +Familie HominidaeUnterfamilie Sivapithecinae +Unterfamilie Ponginae (Orang Utan)Unterfamilie Dryopithecinae +Unterfamilie Homininae (Gorillas, Schimpansen, Australopithecinen, Menschen)
Nach [4].

Sivapithecus dokumentiert die frühe Eigenentwicklungdes Orang Utan. In die Ponginae (Unterfamilie der Homi-nidae, in die der Orang Utan gestellt wird) kann auch derRiese unter den fossilen Primaten, Gigantopithecus, ge-stellt werden, der im späten Miozän Indiens und im Pleis-tozän Chinas und Vietnams nachgewiesen wurde. Interes-santerweise wurden die pleistozänen Reste von Giganto-pithecus teilweise zusammen mit Resten von Homoerectus gefunden, so dass spekuliert wurde, dass Homoerectus zum Aussterben von Giganto-pithecus beigetragen haben könnte.
Miozäne und pliozäneHominini
Der Begriff Hominini bezeichnet einenso genannten Tribus der Taxonomie, d.h.eine systematische Einheit unterhalb einer Unter-familie. Sie umfassen die Formen, die zur engenVerwandtschaft des Menschen zählen. In den ver-gangenen Jahren wurden aus mio- und pliozänenAblagerungen Afrikas Formen beschrieben, die mög-licherweise in die Vorfahrenlinie des Menschengehören. Es handelt sich um Sahelanthropus, Or-rorin und Ardipithecus.
2002 wurde im Tschad (Bahr el Ghazal, 2500km westlich des afrikanischen Grabens) ein sechsbis sieben Millionen Jahre alter, recht gut erhalte-ner Schädel eines höheren Primaten gefunden,der Sahelanthropus tchadensis genannt wurde[8]. Eine vor allem in Frankreich gebräuchlicheunwissenschaftliche Bezeichnung dieses Fundesist „Toumai“. Das Gesicht war kurz und flach,über den Augen befand sich ein sehr kräftigerknöcherner � Supraorbitalwulst. Postorbital istder Schädel relativ stark eingeschnürt und be-saß hinten eine kräftige Nackenleiste (Cristanuchalis). Der Zahnbogen weist, wie der desMenschen, keine Lücken auf. Das Schädelvo-lumen betrug circa 320 – 380 ccm. Die Dicke derSchmelzkappen der Molaren lag zwischen der von Panund Australopithecus. Weiteres Skelettmaterial wurde bis-her nicht gefunden. Dieser Fund ist schwer einzuordnen.Ob er, wie teilweise behauptet wird, und wie der Gat-tungsname suggeriert, in der Vorfahrenlinie des Menschensteht, ist völlig offen. Es findet sich beispielsweise bisherkein überzeugender Befund, der für aufrechten Gangspricht.
Im Jahre 2000 wurden in den Tugen-Bergen Keniasspärliche Skelettfunde (proximale Teile zweier Ober-schenkelknochen, Abbildung 2, Unterkieferfragmenteund Zähne) gemacht, die einem sehr nahen Verwandtendes Menschen zugeschrieben wurden, der Orrorin tuge-nensis genannt wurde [8, 11]. Das Alter dieser Funde wirdauf circa sechs Millionen Jahre geschätzt. Manches (bei-spielsweise Oberschenkelhalskopf, Zähne, dicke Schmelz-kappen) spricht dafür, dass es sich um eine Form handelt,
die in den Kreis der Vorläufer von Australopithecus/Homo gehört. Die Anatomie des Oberschenkels lässt ver-muten, dass Orrorin sich biped und aufrecht fortbewegenkonnte.
Eine pliozäne Form, deren Einordnung in die Homi-nini weniger umstritten ist, ist Ardipithecus, von dem1994, 2001 und 2004 Reste bei Aramis in der nordäthiopi-schen Region Middle Awash gefunden wurden. [8]. Ardi-pithecus ist eine circa 4,4 Millionen Jahre alte, ursprüngli-che Form, die möglicherweise in direkter Beziehung zurGattung Australopithecus und somit auch zur Gattung
Homo steht. Einige Paläontologen weisen auf Ähnlich-keiten von Ardipithecus mit dem Schimpansen (Ab-
bildung 3a) hin, was nicht völlig überraschendwäre, da sich Pan und Homo nach molekul-arbiologischen Berechnungen erst vor circa
fünf bis sechs Millionen Jahren getrennt haben.
Australopithecinen – unmittelbare Vorläufer
des Menschen Die Australopithecinen umfassen verschiedene
Arten der Gattung Australopithecus und Kenyan-thropus. Sie sind eine rein fossile Gruppe, die vorgut 4 Millionen Jahren auftauchte und vor circa 1,4bis 1,5 Millionen Jahren ausstarb. Sie sind die aufden afrikanischen Kontinent beschränkte Grund-gruppe, aus der sich die Gattung Homo entwickelte.Unter den Australopithecinen gibt es in zeitlicherHinsicht jüngere Formen mit markanten, „robusten“Schädel- und Gebissmerkmalen, die teilweise in eineeigene Gattung, Paranthropus, gestellt werden. Da esjedoch Hinweise darauf gibt, dass sich diese robustenMerkmale konvergent entwickelt haben, hat eine ei-gene Gattung Paranthropus keine gesichterte Basis. Dieälteren Australopithecinen, denen die „robusten“ Merk-male fehlen, werden als „grazil“ bezeichnet. Analyse desGebisses und des Zahndurchbruchs weisen darauf hin,dass die Individualentwicklung der Australopithecinen
recht rasch verlief und der der heutigen afrikanischenMenschenaffen ähnelte.
Australopithecus anamensis wurde in Kenia (Kana-poi und Allia Bay) gefunden, ist circa 4,2 Millionen Jahre(Kanapoi) bzw. 3,9 Millionen Jahre (Allia Bay) alt. Vielesspricht dafür, dass sich diese Form schon biped fortbe-wegte.
Australopithecus afarensis (Abbildung 3b) ist relativgut bekannt. Die über 200 Einzelfunde mehrerer Indivi-duen stammen aus einer Zeit zwischen 3,9 und knapp 3Millionen Jahren vor heute. Ein Individuum, von dem einrelativ vollständiges Skelett geborgen wurde, wurde„Lucy“ getauft. Die Fundstätten liegen vor allem in Äthio-pien und Tansania. Dass A. afarensis aufrecht ging, bele-gen fossile Fußspuren. Die Schädelkapazität variiert zwi-schen 400 und 550 ccm, das Becken war schon – wiebeim Menschen – breit und kurz. Ein Australopithecinen-
44 | Biol. Unserer Zeit | 1/2007 (37) www.biuz.de © 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
A B B . 2 Ober-schenkelknochen(Femur) von Orrorin mit Hüft-gelenkskopf. Die Anatomie diesesKnochens ist gutvereinbar mit derAnnahme, dass Orrorin aufrechtging. Nach Pick-ford und Senut(2000).

P R I M A T E N F O S S I L I E N , 1 . T E I L | PA L Ä O N TO LO G I E
© 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2007 (37) | Biol. Unserer Zeit | 45
Fund (vorderer Kiefer mit Zähnen), der 1995 beschriebenwurde und aus dem Tschad stammt, wurde mit dem Na-men Australopithecus bahrelghazali belegt. Möglicher-weise repräsentiert er nur eine regionale Variante vonAustralopithecus afarensis. Dieser Fund ist vor allem des-wegen bemerkenswert, weil er 2500 Kilometer westlichdes afrikanischen Grabens erfolgte, in dem die Mehrzahlder anderen Australopithecinen gefunden wurde.
Im Jahre 2001 wurde in Kenia am Westufer des Turka-nasees circa 3,5 Millionen Jahre altes Zahn- und Schädel-material gefunden, das mit der Bezeichnung Kenyanthro-pus platyops versehen wurde. Die Entdecker verliehendem Fund einen eigenen Gattungsnamen, weil sie der An-
sicht sind, dass hier eine eigenständige Kombination vonMerkmalen vorliegt, beispielsweise ein enger knöchernerGehörgang- wie bei Pan und Australopithecus anamen-sis –, eine sehr dicke Schmelzschicht auf den Backenzäh-nen – wie bei A. anamensis und A. afarensis –, relativkleine postcanine Zähne – wie bei Homo -, kleine Schä-delkapazität – wie bei Australopithecinen generell – sowieein flacher, großer Gesichtsschädel – wie bei Homo. Obdie Errichtung einer eigenen Gattung für diese Fund-stücke Bestand haben wird, ist ungewiss, zumal der Schä-del Deformationen aufweist. Einige Forscher ordnenKenyanthropus, der gleichzeitig mit A. afarensis lebte,der Gattung Australopithecus zu.
A B B . 3 Schädel von Schimpanse, Australopithecinen und Homo. a) Schimpanse (Pan troglodytes, Männchen), b) Australopithecus afaren-sis, c) Australopithecus africanus, d) Homo habilis (KNM-ER 1813), e) Homo ergaster (KNM-ER 3733), f) klassischer Homo erectus, Peking-mensch, Fundort Zhoukoudian g) jüngerer Homo erectus, Fundort Jinniushan, Alter vermutlich ca. 280.000 Jahre, mit morphologischen An-klängen an H. heidelbergensis. h) Homo heidelbergensis (Petralona, Chalkidike, Griechenland). Maßstab unterschiedlich. Nach verschiedenenAutoren.

46 | Biol. Unserer Zeit | 1/2007 (37) www.biuz.de © 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Australopithecus garhi wurde 1999 beschrieben. Der 2,5Millionen Jahre alte Fund stammt aus dem nördlichenÄthiopien (Bouri, Middle Awash). Es wurden Schädel-bruchstücke und Zähne gefunden. Auch in der Nähe desSchädelmaterials aufgefundene Extremitätenskelettrestewerden A. garhi zugeordnet. Unter anderem wegen derzeitlichen Nähe zum ersten Auftreten der Gattung Homowird A. garhi mitunter als möglicher Vorfahr von Homoins Spiel gebracht.
Von Australopithecus africanus gibt es inzwischenviele Einzelfundstücke; ihr Alter liegt zwischen 3,2 und2,5 Millionen Jahren vor heute (Abbildung 3c). EinzelneForscher berichten über ältere Funde. Der 1924 beschrie-bene Schädel eines kindlichen A. africanus (des „Kindesvon Taung“) ist der erste beschriebene Fund eines Aus-tralopithecinen überhaupt. Bis zum heutigen Tag werdenregelmäßig, vor allem in den Höhlen von Sterkfontein inder Umgebung von Johannesburg, neue Funde gemacht.Die Schädelkapazität variierte zwischen 425 und 560 ccm,lag also im Durchschnitt über dem Wert von A. afarensis.Ein 1995 beschriebenes Fußskelett ist proximal sehr men-schenähnlich, distal besitzt es noch – wie Menschenaffen– eine abspreizbare Großzehe, was ein Indiz dafür ist, dassA. africanus sich nicht nur am Boden aufrecht gehendfortbewegte, sondern auch in Bäumen kletterte. Zufolgeneuerer Berechnungen variiert das Körpergewicht zwi-
schen 30 und gut 40 kg; Männer wurden circa 138 cm,Frauen circa 115 cm groß. Ob A. africanus schon eine ein-fache Kultur mit Holz- oder Knochengegenständen besaß,ist umstritten. Für manche Forscher kommt A. africanusals Vorfahr für die Gattung Homo in Betracht.
Robuste AustralopithecinenDie Mehrzahl der Paläoanthropologen zählt Austral-opithecus robustus (Südafrika), A. aethiopicus (Äthio-pien, Kenia) und A. boisei (verschiedene Fundstellen inOstafrika) zu den robusten Australopithecinen, die durcheinen kräftigen Gesichtsschädel und einen stark ent-wickelten Kauapparat gekennzeichnet waren. Es sind ins-besondere die Männer, die den markant-kräftigen Ge-sichtsschädel besitzen. Es ist nicht unwahrscheinlich, dassdas Merkmal „robust“ konvergent entstanden ist. A. ae-thiopicus lebte zwischen 2,6 und 2,3 Millionen Jahren vorheute, A. robustus zwischen 1,8 und 1,5 Millionen Jahrenvor heute und A. boisei zwischen 2,3 und 1,4 MillionenJahren vor heute. Mit A. robustus und A. boisei starben dieAustralopithecinen insgesamt vor circa 1,4 bis 1,5 Millio-nen Jahren aus. Der robuste Kauapparat ist primär wahr-scheinlich eine Anpassung an überwiegend harte Pflan-zennahrung (darunter Rhizome, Wurzeln, Samen undNüsse). Neuere Analysen des Strontiumgehalts der Zähnelassen vermuten, dass die Australopithecinen auch Fleisch
A B B . 4 Zeitliches Auftreten und hypothetische Verwandtschaftsbeziehungen der Gattungen Australopithecus, Kenyanthropus und Homo. Nach McKee et al., verändert (2005).

P R I M A T E N F O S S I L I E N , 1 . T E I L | PA L Ä O N TO LO G I E
zu sich nahmen, weil die Zähne, wie Fleisch, einen nied-rigen Strontiumgehalt aufweisen. Pflanzen besitzen einenhohen Strontiumgehalt, und dementsprechend sind dieZähne von fossilen pflanzenfressenden Säugern stron-tiumreich.
Erste Angehörige der Gattung HomoDie frühesten Funde der Gattung Homo (Mensch) reprä-sentieren die erste Stufe eines neuen Entwicklungsniveaus„jenseits“ von Australopithecus. Was den Menschen kenn-zeichnet und von anderen Primaten unterscheidet, wirdvermutlich immer kontrovers diskutiert werden. Wesent-lich sind der bipede Gang (mit zahlreichen anatomischenund physiologischen Anpassungen), das große und kom-plex aufgebaute Gehirn, die Sprache und eine auf Kulturbasierende Lebensweise, zu der auch die verschiedenenSozialstrukturen gehören. Diese Merkmale waren, abgese-hen vom bipeden Gang, sehr wahrscheinlich anfangs erstansatzweise vorhanden. Heute sind sie Grundbedingungunseres Daseins, wobei hinsichtlich der Ausformung derLeistungen des Gehirns, der Sprache und der Kultur einesehr starke Variabilität besteht.
Älteste bisher bekannte Steinwerkzeuge sind circa 2,3bis 2,6 Millionen Jahre alt und stammen aus Gona, Lokala-lei und Hadar in Äthiopien. Diese Werkzeuge erwiesensich bei näherer Analyse als erstaunlich komplex und di-vers und deuten auf hohe kognitive und manipulatorischeFähigkeiten ihrer Hersteller hin. Etwas jünger, circa 1,8Millionen Jahre alt, sind relativ einfache Steinwerkzeugeaus der Olduvai-Schlucht Ostafrikas („Chopper“, Oldo-wan-Kultur). Die zum Teil erhebliche Stückzahl aufgefun-dener Steinwerkzeuge deutet vielleicht darauf hin, dasshier schon fast „gewerbsmäßig“, unter Umständen auchnach „Angebot und Nachfrage“ Werkzeuge hergestelltwurden. Als Hersteller kommen eigentlich nur Homo ha-bilis und Homo rudolfensis in Betracht. Aber immer wie-der wird von einzelnen Forschern die Frage gestellt, ob
nicht auch Australopithecinen (A. garhi, A. boisei) alsWerkzeughersteller infrage kommen könnten.
Es bleibt immer noch umstritten, wie die erste Homo-Art paläoanthropologisch, d.h. im Wesentlichen morpho-logisch, ökologisch und in Hinsicht auf ihre Werkzeugkul-tur, zu definieren ist. Auch die Festlegung auf H. habilisund H. rudolfensis als erste Vertreter der Gattung Homoist umstritten. Es sei nur erwähnt, dass es Wissenschaftlergibt, die H. habilis und H. rudolfensis als Australopitheci-nen einstufen. Aus evolutionsbiologischer Sicht ist es ver-ständlich, wenn Übergangsformen oder erste Vertreter ei-ner Gattung ein Mosaik an Merkmalen aufweisen, das diesystematische Bewertung schwer macht. Die ersten Artender Gattung Homo (H. habilis und H. rudolfensis) habeneine Schädelkapazität, die zwischen 610 und 788 ccm va-riierte (Abbildung 4). Diese Zahlen liegen oberhalb derZahlen für Australopithecus. Die Variationsbreite der ge-nannten Zahlen kann auf individuelle Variabilität, die esbekanntlich in noch größerem Ausmaß beim heutigenMenschen gibt, Geschlechtsdimorphismus oder lokale Po-pulationen mit unterschiedlicher Körpergröße hindeuten.Es ist zu vermuten, dass schon die ersten Vertreter vonHomo in komplexen Sozialsystemen lebten, was mög-licherweise die Entstehung von Sprache begünstigte. Esgibt Forscher, die aufgrund der Untersuchungen von Schä-delausgüssen der Ansicht sind, dass die Hirnstruktur vonH. habilis und H. rudolfensis einen höheren Komple-xitätsgrad aufweist als diejenige von Australopithecus,speziell sollen Broca- und Wernicke-Zentrum relativ präg-nant erkennbar sein. Sprache ist aber wohl für erfolgrei-che Zusammenarbeit, beispielsweise auf der Jagd, nichtunbedingt Voraussetzung, wie gemeinschaftlich jagendeCarnivoren, beispielsweise der Afrikanische Wildhund Ly-caon pictus, zeigen.
Weitere anatomische Merkmale der Gattung Homosind: orthognathes (flaches, vertikales) Gesicht, abgerun-deter Zahnbogen, Fehlen von Zahnlücken, verkleinerte,
G LOSSA R
EEnnddooccrraanniiaallaauussgguussss:: natürlicher oder künstlicher Abguss der Innenseite desSchädels, die nicht selten die Oberfläche des Gehirn widerspiegelt.
MMoollaarr:: hinterer Backenzahn (Mahlzahn). Beim Menschen gehören der 6., 7.und 8. Zahn zu den Molaren. Es handelt sich um große, kräftige Zähne mitausgeprägten Höckern, Leisten und Furchen.
OOllddoowwaann-- uunndd AAcchheeuullééeenn--KKuullttuurr:: Als OOllddoowwaann werden die ältesten Stein-werkzeuge der Welt bezeichnet. Der Name der Steinwerkzeuge stammt vonihrem Fundort in Tansania, der Olduvai-Schlucht. Typisch ist die Verwendungvon Geröllen, denen mit wenigen Schlägen eine scharfe Kante beigebrachtwurde. Später kommen Kerngeräte und einfache Abschlagwerkzeuge hinzu.Als Produzenten der einfachen Steingeräte gelten Homo rudolfensis, Homohabilis, Homo erectus sowie Homo ergaster. Mitunter werden auch Austral-opithecinen als Hersteller in Betracht gezogen.
Das AAcchheeuullééeenn ist eine Kulturstufe des Paläolithikums, der Altsteinzeit, benannt nach einer Fundstätte der Faustkeile nahe Saint Acheul, einem Vor-
ort von Amiens (Frankreich). Das Acheuléen begann in der zweiten Eiszeit vor etwa 1.600.000 Jahren und setzte sich bis in die dritte Eiszeit fort; es endete vor etwa 100.000 Jahren. Beim Acheuléen handelte es sich um eineKultur der Jäger und Sammler. Kennzeichen sind unter anderem perfekt gearbeitete Faustkeile, die für viele verschiedene Tätigkeiten eingesetzt wer-den konnten.
OOrrbbiittaa:: Augenhöhle des Schädels, in der der Augapfel und seine Hilfsorganeliegen. Beim Menschen ist die Augenhöhle 4 bis 5 cm tief.
PPrräämmoollaarr:: einer der vorderen Backenzähne. Im Gegensatz zu den Molarenhaben Prämolaren auch Milchzahnvorgänger, üben also bereits im Milch-gebiss die Mahlfunktion aus. Im Regelfall sind die Prämolaren mindestenszweihöckrig. Neuweltaffen haben fast immer drei Prämolaren, Altweltaffen(einschließlich Menschenaffen und Mensch) stets zwei.
SSuupprraaoorrbbiittaallwwuullsstt:: knöcherner Überaugenwulst.
© 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2007 (37) | Biol. Unserer Zeit | 47

spezifisch geformte Eckzähne, relativ lange Beine, Fuß-skelett mit Längs- und Quergewölbe sowie nicht mehr ab-spreizbarem Großzeh, breiteres Becken u.a.
Homo habilis – die erste MenschenartHomo habilis ist die weithin anerkannte erste Art der Gat-tung Homo (Abbildung 3d). Er wurde im Zeitraum von 2,4bis 1,65 Millionen Jahren in Hadar (Äthiopien, 2,4 Millio-nen Jahre), in der Olduvai-Schlucht (Tansania, 1,65 Millio-nen Jahre), in Sterkfontein (Südafrika, 2,2 Millionen Jahre)und wahrscheinlich auch in Koobi Fora (Kenia, 1,9 Millio-nen Jahre) nachgewiesen. Der Zeitpunkt 2,4 MillionenJahre vor heute passt gut zum ersten Auftreten von Stein-werkzeugen in Ostafrika und Nordostafrika vor circa 2,5Millionen Jahren. Es wurde berechnet, dass H. habilis 140cm groß war. Wie bei H. sapiens war der erste Zeh nichtmehr abspreizbar und der dritte Molar etwas reduziert.Die Schädelkapazität variiert zwischen 580 und 687 ccm,eine Messung (Fund Koobi Fora) liegt bei 509 ccm.
Homo rudolfensis ist eine umstrittene frühe Homo-Form. Er wird jetzt öfter als eine etwas robustere Variantevon H. habilis angesehen, einzelne Forscher sehen in ihmsogar einen Australopithecinen. Die entsprechendenFunde sind circa 2,4 Millionen bis circa 1,8 oder 1,9 Mil-lionen Jahre alt. Die Fundstellen liegen am Malawisee, imOmogebiet (Südäthiopien) und am Ostufer (Koobi Fora)des Turkanasees. Die Schädelkapazität zweier Individuenliegt bei 752 und 824 ccm. Die Funde vom Malawisee undaus der Omoregion umfassen nur relativ spärliches Unter-kiefer- und Zahnmaterial sowie einzelne Schädelknochen,was ihre Einschätzung besonders schwer macht.
Homo ergaster – Spuren in Afrika, Asien und Europa
Homo ergaster (Abbildung 3e) wird von vielen Paläan-thropologen als eine besonders wichtige und interessanteMenschenart angesehen, die im Zeitraum von 1,9 Millio-nen Jahren bis 1 Million Jahren in Afrika lebte und von dermöglicherweise alle weiteren Homo-Arten abstammen.Eine Art, die in manchen morphologischen Details H. er-gaster ähnelt, ist H. erectus, der aber von vielen Wissen-schaftlern auf Ost- und Südostasien beschränkt wird, woer zwischen 1,8 Millionen und vielleicht 27.000 Jahren vorheute lebte. Es gibt aber auch Paläontologen und andereEvolutionsforscher, die H. ergaster und H. erectus füridentisch halten und nur die Art H. erectus anerkennen.Die afrikanischen H. ergaster-Funde werden dann mit„afrikanischer H. erectus“ bezeichnet. Es gibt weiterhinForscher, die eine Reihe von jüngeren (circa 900.000 bis400.000 Jahre alt) Funden in Europa und Asien, die oft alsH. heidelbergensis bezeichnet werden, auch H. erectuszuzählen, so dass dann H. erectus eine in der ganzen Weltverbreitete Art wäre, aus der an verschiedenen Stellen H.sapiens entstand. Es ist dennoch bei dem gegenwärtigenKenntnisstand plausibel, H. erectus – wie traditionell ge-handhabt – auf Ost- und Südostasien zu beschränken. Dies
schärft auch den Blick für Unterschiede und erleichtertdie Orientierung. Zu den morphologischen Eigenmerk-malen von H. ergaster im Vergleich mit H. erectus zählen:generell etwas grazilerer Skelettbau, längere und schma-lere Molaren, komplexe Wurzeln der Prämolaren, dünnereSchädelknochen, kein Knochenkiel auf dem Schädel,schwächerer Überaugenwulst, höhere Schädelwölbung,geringere postorbitale Schädeleinschnürung, schmalereSchädelbasis [5].
Homo ergaster ist also eine afrikanische Homo-Art,deren Entwicklungsniveau über dem von H. habilis liegt.Die Schädelkapazität variiert zwischen 750 und 850 ccm,bei den jüngsten, erst 2002 beschriebenen Funden ausDaka (nördliches Äthiopien) liegt die Schädelkapazität bei995 ccm. Ein besonders gut erhaltener und fast vollständi-ger 1,6 Millionen Jahre alter Skelettfund stammt aus Na-riokotome (Turkanasee, Kenia). Er wurde 1984 beschrie-ben. Es handelte sich um einen circa 162 cm großen Ju-gendlichen, der als Erwachsener wahrscheinlich circa 180cm groß und 67 kg schwer geworden wäre. Homo erga-ster hat Werkzeuge der � Oldowan- und Acheuléen-Kulturhergestellt und wahrscheinlich das Feuer beherrscht. Diesüdafrikanischen Funde sind schon länger bekannt undwurden zunächst als „Telanthropus capensis“ beschrie-ben.
Es ist zu vermuten, dass eine H. ergaster-Populationschon früh Afrika verlassen hat und nach Asien ausgewan-dert ist. Hier könnte aus H. ergaster die Art H. erectus ent-standen sein, deren älteste Vertreter auf Java (und vermut-lich auch in China) nach neueren Datierungen circa 1,8Millionen Jahre alt sind. Das bedeutet, dass die „Erobe-rung“ Ost- und Südasiens relativ rasch erfolgte.
Möglicherweise hat H. ergaster auch schon früh Nord-westafrika und Süd- bzw. Südwesteuropa besiedelt. Sofand man beispielsweise circa 1,8 Millionen Jahre alte pri-mitive Steinwerkzeuge in Ain Hanech (Nordostalgerien)und 1,7 bis 1,6 Millionen Jahre alte Steinwerkzeuge inOrce (Südostspanien). Diese Nachweise werden auch mitH. antecessor (s.u.) in Verbindung gebracht.
Es ist gut denkbar, dass 1991, 2000 und 2002 in Dma-nisi (Georgien) entdecktes Skelettmaterial noch H. ergas-ter zugehört, oder schon einer Übergangsform zu H. erec-tus entspricht [8]. Diese Funde sind circa 1,8 MillionenJahre alt. Die Schädel hatten eine Kapazität von 650 bis788 ccm. Die Begleitfunde sind einfache Steinwerkzeugeder Oldowan-Kultur, wie sie auch für die älteren afrikani-schen H. ergaster-Funde kennzeichnend sind. Seit 2002zählen die Entdecker der Funde das Fundmaterial einerneu geschaffenen Art „H. georgicus“ zu, wobei es sichwohl eher um eine plakative Fundbezeichnung als umbreit abgesicherte wissenschaftliche Erkenntnis handelt.
Homo erectusHomo erectus (Abbildung 3f, g) wurde schon 1891 aufJava entdeckt, ab den zwanziger und dreißiger Jahren des 20. Jahrhunderts kamen dann Funde aus Zhoukoudian
48 | Biol. Unserer Zeit | 1/2007 (37) www.biuz.de © 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim

P R I M A T E N F O S S I L I E N , 1 . T E I L | PA L Ä O N TO LO G I E
© 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2007 (37) | Biol. Unserer Zeit | 49
(bei Peking) hinzu. In den dreißiger und vierziger Jahrendes vergangenen Jahrhunderts folgten dann eine Reiheweiterer Funde sowohl auf Java, als auch in China. Bisheute werden auf Java vor allem von indonesischen Wis-senschaftlern neue Funde gemacht, und auch aus Chinaliegen viele neuere Funde vor. Die aktuellen Altersbestim-mungen lassen vermuten, dass die ältesten javanischen H.erectus-Funde (in Sangiran, nördlich des Gunung Lawu beiSolo) circa 1,8 Millionen Jahre alt sind. Ihre Schädelkapa-zität betrug circa 800 ccm, etwas jüngere (1,5 bis 1 Mil-lionen Jahre alte) Funde hatten eine Schädelkapazität voncirca 900 ccm.
Zwischen 1931 und 1933 wurden in Ostjava – bei-spielsweise bei Ngandong – Schädel gefunden, die zu-nächst einfach als spätpleistozän eingestuft wurden. ImJahr 1996 durchgeführte Altersbestimmungen ergaben einAlter von nur circa 56.000 bis 27.000 Jahren. Diese Fundewerden oft letzten überlebenden Vertretern von H. erec-tus auf Java zugeschrieben, deren Schädelkapazität beicirca 1200 ccm lag, und die in aller Wahrscheinlichkeit ne-ben H. sapiens auf Java lebten. Es gab und gibt aber auchPaläontologen, die diese Ngandong-Funde einer eigenenArt, H. soloensis, zuordnen, oder sie als Vertreter von H.sapiens ansehen, die besonders enge Beziehungen zu denaustralischen Aborigines besitzen.
Ebenfalls umstritten in ihrer Datierung sind H. erectus-Funde in Sambungmacan (Ostjava), wo bis in jüngste Zeit(2002) Funde gemacht wurden. Die Schädelkapazität ei-nes Fundstückes (SM3) beträgt circa 920 ccm, die eineszweiten (SM4) 1006 ccm. Interessant sind die Asymmetrievon linker und rechter Großhirnhemisphäre und eine Ver-größerung des Broca-Areals links. Dies deutet auf kogni-tive und sprachliche Fähigkeiten, wie sie sonst den mo-derneren Homo-Formen zugeschrieben werden, hin. DieFunde stammen vermutlich aus dem mittleren Pleistozän,die Altersangaben schwanken zwischen circa 400 000 und 800 000 Jahren.
Das Bild von H. erectus in China ist in den vergange-nen Jahrzehnten, vor allem dank reger Tätigkeit chinesi-scher Forscher, vielschichtiger geworden. Die ältesten H.erectus-Funde in dem bekannten Zhoukoudian-Höhlen-system (Abbildung 3f) sind 670.000 Jahre alt, in jüngereFunde sind 410.000 Jahre alt. Ähnlich alte Funde stamm-ten aus der Jiangshi-Höhle (Provinz Hubei, 470.000 Jahrealt), aus der Tham Khuyen Höhle (Vietnam, circa 470.000Jahre alt), aus Tangshan (bei Nanking, 600.000 Jahre alt),aus Yunxian (Provinz Hubei, um 400.000 Jahre alt) undaus der Longtang-Höhle in der Region Hexian (Provinz An-hui, circa 400.000 Jahre alt).
Deutlich älter sind H. erectus-Funde aus der Longgu-pohöhle (östliches Sechuan, wahrscheinlich circa 1,9 Mil-lionen Jahre alt, zeigen noch Anklänge an H. ergaster), ausder Nihewan Senke (Nordchina, nur Steinwerkzeuge, 1,36Millionen Jahre alt), aus Yuanmou (Yunnan, circa 1,7 Mil-lionen Jahre alt), aus der Region Lantian (Provinz Shansi,circa 1,15 Millionen Jahre alt), aus der Renzi Höhle (Pro-
vinz Anhui, Werkzeuge, vielleicht sogar etwas über 2 Mil-lionen Jahre alt).
Jüngere chinesische H. erectus-Funde stammen unteranderem aus Jinniushan (circa 280.000 oder vielleichtauch nur 20.000 Jahre alt, Schädelkapazität 1400 ccm, Abb3g), Dali (Provinz Shansi, circa 200.000 Jahre alt, vielleichtauch jünger), Maba (Provinz Guandong, circa 132.000Jahre alt, immer wieder wird auf Anklänge an den Nean-dertaler hingewiesen) und aus Changyang (Provinz Hubei,circa 195.000 Jahre alt).
Bei den jüngeren H. erectus-Funden wird einerseitseine gewisse Ähnlichkeit mit H. heidelbergensis hervor-gehoben. Andererseits werden sie von manchen Paläonto-logen als archaische H. sapiens angesehen, die Über-gangsformen seien zwischen dem „klassischen“ H. erectusund dem moderneren H. sapiens. Diese Paläontologenund andere Forscher sehen diese Funde als Beweis für dieTheorie der regionalen Entstehung des H. sapiens an (Ab-bildung 5). Das soll bedeuten, dass sich hier schrittweiseH. sapiens aus H. erectus entwickelt hat. Diese Theoriewiderspricht ausdrücklich der insbesondere von Molekul-arbiologen vertretenen Auffassung, dass alle modernenMenschen aus einer afrikanischen Vorform hervorgehen,die alle anderen Menschenarten ersetzt hat (Replacement-Theorie, Out-of-Africa-Theorie).
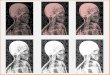
Die Flores FundeIm Jahre 2003 wurden in der Liang Bua Höhle auf Flores(Indonesien) Skelettreste von auffallend kleinen Men-schen gefunden, die verdachtsweise der Art Homo flore-siensis zugeordnet wurden. Die ältesten Funde sind circa95.000, die jüngsten circa 12.000 Jahre alt. Das am bestenerhaltene circa 12.000 Jahre alte Skelettmaterial eines In-
A B B . 5 Schematische Darstellung der Theorie der multiregionalen Konti-nuität (a) und der „Replacement Theorie“ in der Evolution des Menschen (b).Nach Tattersall, verändert (1995).
b)a)

50 | Biol. Unserer Zeit | 1/2007 (37) www.biuz.de © 2007 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
dividuums erhielt die Bezeichnung LB1. Diese Menschenwaren nur circa einen Meter groß; die Schädelkapazitätwurde auf circa 380 bis 410 ccm berechnet (Abbildung 6).Der Abdruck des Gehirns deutet trotz der geringen Hirn-größe auf ein relativ hochentwickeltes Großhirn mit aus-gedehntem präfrontalem Cortex (Area 10) hin. Hier sindbeim modernen Menschen höhere kognitive Funktionenlokalisiert, wie beispielsweise die Fähigkeit zu planen undGedächtnisinhalte zu nutzen. Diese Menschen habenStein- und Holzwerkzeuge hergestellt, konnten vermutlichmit Feuer umgehen und haben gemeinschaftlich gejagt.Diese Leistungen sind angesichts des kleinen Gehirns er-staunlich und sehr rätselhaft.
Die Interpretation dieser Funde ist schwierig und dieDiskussion darüber hält an. Ein Erklärungsansatz geht da-hin, dass es sich bei diesen Funden um eine isolierte In-selpopulation von H. erectus handelt. Möglicherweisehatte H. erectus vor circa einer Million Jahren Flores er-reicht (alte Steinwerkzeuge, die auf Flores gefunden wur-den, sind circa 800.000 Jahre alt) und in der Isolation derInsel hat sich dann diese Zwergform entwickelt. Verzwer-gung von größeren Säugern auf Inseln ist ein wiederholtzu findendes Phänomen, beispielsweise gab es Zwerg-elefanten auf Malta und Kreta [1, 2, 9].
Nach einer anderen Auffassung han-delt es sich um pathologisch verän-
derte mikrozephale moderne Men-schen, eine Ansicht, für die aber
nur wenig konkrete Befundebeigebracht werden können.Nichts spricht dafür, dass essich um eine pygmäenähnli-che Population von H. sapi-
ens handelt. Alle Pygmäen ha-ben die normale Hirngröße des
heutigen Menschen.Am besten begründet ist zurzeit, dass es sich
bei den Funden um eine eigenständige Art, H. flo-resiensis, handelt. Die sorgfältige Analyse der Schä-
delmorphologie ergibt die meisten Übereinstimmungenmit H. ergaster, die postkranialen Skelettmerkmale stim-men zum erheblichen Teil mit Australopithecus garhiüberein. Die spezifischen Übereinstimmungen (Schädel,� Endokranialausguss, Extremitäten) mit H. erectus sindnicht groß. Es ist daher zu vermuten, dass es sich um Re-präsentanten einer eigenen Art handelt, die vielleichtschon vor circa zwei Millionen Jahren aus Afrika ausge-wandert und auf noch unbekannte Weise bis nach Floresgelangt ist. Homo floresiensis zeigt altertümliche Skelett-merkmale, aber ein recht komplex gestaltetes Großhirn.
In der nächsten Ausgabe von „Biologie in unserer Zeit“folgt der zweite Teil unseres Überblicks über die Entwick-lungsgeschichte des Menschen: Von Indonesien geht eszurück nach Europa – von den frühen Funden den Gat-tung Homo zu Homo heidelbergensis, dem Neandertalerund zum modernen Homo sapiens.
ZusammenfassungDer Entwicklungsweg des Menschen begann vor circa sechsMillionen Jahren, als sich die Entwicklungslinien von Menschund Schimpansen trennten. In den vergangenen Jahrzehn-ten wurden mehr relevante Fossilfunde gemacht als in dengut 100 Jahren zuvor, so dass die Grundzüge der Evolutiondes Menschen mittlerweile recht gut bekannt sind. Die zu-nehmende Zahl an Forschern auch außerhalb Europas undder lässt erwarten, dass die Zahl der Funde in Zukunft nochwesentlich anwachsen wird, so dass Wissenslücken ge-schlossen werden, aber auch neue Interpretationen disku-tiert werden müssen. Dieser zweiteilige Artikel bietet einenÜberblick über gesicherte Erkenntnisse, spannende Funde –und offene Fragen.
Literatur[1] D. Argue, D. Dolon, G. Groves, R. Wright, Homo floresiensis: Micro-
cephalic, pygmoid, Australopithecus, or Homo?, J. Human Evolu-tion 2006.
[2] P. Brown, T. Sutikna, M. J. Morwood, R. P. Soejono, T. Jatmiko, E. Wayhu Saptomo, R. Awe Due, A new small-bodied hominin fromthe Late Pleistocene of Flores, Indonesia, Nature 2004, 431, 1055-1061.
[3] D. W. Cameron, A revised systematic scheme for the Eurasian Mio-cene fossil Hominidae, J. Hum. Evol. 1997.
[4] C. Groves, Primate Taxonomy, Smithsonian Institution, 2001.[5] W. Henke, H. Rothe, Stammesgeschichte des Menschen, Springer
Verlag, 1999.[6] J. Kelley, The hominoid radiation in Asia. In: W. Hartwig, The Pri-
mate Fossil Record, Cambridge University Press 2002, 369-384.[7] R. D. Martin, Primate Origins and Evolution: A Phylogenetic Recon-
struction, Princeton University Press, 1990.[8] J. K. McKee, F. E. Poirier, W. S. McGraw, Understanding Human Evo-
lution, Pearson Prentice Hall, New Jersey, 2005.[9] M. J. Morwood, P. Brown, T. Jatmiko Sutikna, E. Wahyu Saptomo, K.
E. Westaway, R. Awe Due, R. G. Roberts, T. Maeda, S. Wasisto, T.Djubiantono, Further evidence for small-bodied hominins from theLate Pleistocene of Flores, Indonesia, Nature 2005, 437, 1012-1017.
[10] S. Moya-Sola, M. Köhler, D. M. Alba, I. Casanovas-Vilar, J. Galindo,Pierolapithecus catalaunicus, a New Middle Miocene Great Apefrom Spain, Science 2004, 306, 1339.
[11] C. F. Ross, R. F. Kay, Anthropoid Origins. New Visions, Kluwer Aca-demic, Plenum Publishers, 2004.
[12] N. M. Young, L. Maclatchy, The phylogenetic position of Moroto-pithecus, J. Hum. Evol. 2004, 163-84.
Der AutorUlrich Welsch, geboren 1940, studierte Biologie inMünchen und Kiel. Nach Promotion und Habilita-tion im Fach Anatomie schloss er ein Medizinstu-dium an, das er 1982 mit Staatsexamen und Ap-probation abschloss. Seit 1984 ist er Lehrstuhlinha-ber an der Anatomischen Anstalt der UniversitätMünchen.
KKoorrrreessppoonnddeennzz
Prof. Dr. Dr. Ulrich WelschUniversität MünchenAnatomische AnstaltPettenkoferstraße 11D-80366 MünchenEmail: [email protected]
ABB. 6 Schädeldes kleinen Flo-res-Menschen(LB1, Schädelka-pazität ca. 390 –420 ccm). NachBrown et al.(2004).
Gimmick: © Goce Risteski – FOTOLIA