Embed Size (px)
Citation preview

1964 G. PFLEIDERER, C. WOENCKHAUS, K. SCHOLZ und H. FELLER 205
Die intramolekulare Wechselwirkung zwischen Heterocyclen in Coenzymen, I
NICOTINAMID-MERCAPTOPURIN-, NICOTIN AMID-METHY LMERCAPTOPURIN- UND NICOTINAMID-PURIN-DINUCLEOTID
von GERHARD PFLEIDERER, CHRISTOPH WOENCKHAUS, KLAUS SCHOLZ * und HANS FELLER
Aus dem Institut fur Biochemie im Institut fur Organische Chemie der Universitat Frankfurt/Main
Eingegangen am 20. Juli 1963
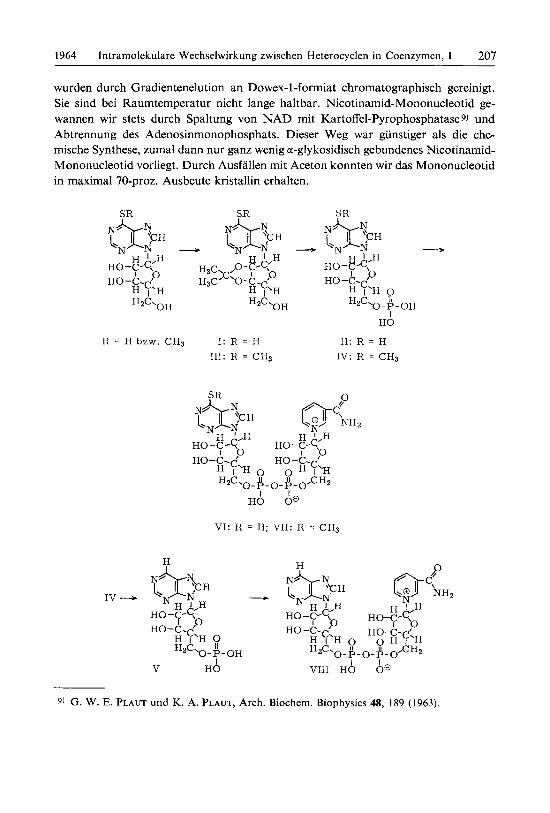
6-Mercapto-9-[2’.3’-isopropyliden-~-~-ribofuranosyl]-pur~n reagiert mit Cyan- athylphosphat und Dicyclohexylcarbodiimid zum 5‘-Phosphat. Mit Methyljodid erhalt man den Methylthioather, der sich in guter Ausbeute phosphorylieren 1aRt. Durch Abspaltung der Methylmercapto-Gruppe mit RANEY-Nickel entsteht 9-[P-~-Ribofuranosyl]-purin-5’-phosphat. Die Nucleotide werden mit Nicotin- amid-Mononucleotid zu den Coenzym-Analoga Nicotinamid-Mercaptopurin-, Nicotinamid-Methylmercaptopurin- und Nicotinamid-Purin-Dinucleotid * *)
kondensiert. Bei den S-haltigen Dinucleotiden laRt sich eine Wechselwirkung zwischen den beiden Heterocyclen im oxydierten und reduzierten Zustand UV- spektroskopisch feststellen. Die synthetisierten Dinucleotide sind biochemisch aktiv, die S-haltigen werden 2-3mal fester an das Enzym gebunden als das na- tiirliche Nicotinamid-Adenin-Dinucleotid. Das Nicotinamid-Purin-Dinucleotid zeigt gegeniiber dem natiirlichen Coenzym keinen wesentlichen Unterschied.
Adeninhaltige Coenzyme mit einem zweiten heterocyclischen Rest im Molekiil zeigen optische Anomalien, die nur durch eine Wechselwirkung zwischen den beiden heterocyclischen Systemen erklart werden konnen.
Schon lange bekannt ist die Beeintrachtigung der Fluoreszenz durch das Adenin im Flavin- Adenin-Dinucleotid gegenuber dem Flavin-Mononucleotid 1). Beim Nicotinamid-Adenin- Dinucleotid (NAD) beobachtete WEBER~) , daR man das hydrierte Coenzym mit Licht der Wellenlange 260 m p zur Fluoreszenz anregen kann, obwohl dieses nur vom Adeninteil ab- sorbiert wird. Nach Spaltung des Coenzyms mit Pyrophosphatase in die beiden Mononucleo-
*)Teil der Dissertation K. SCHOLZ, Univ. Frankfurt/Main 1963. * *) Es werden folgende Abkurzungen verwendet : Nicotinamid-Mercaptopurin-Dinucleotid =
NMD, Nicotinamid-Methylmercaptopurin-Dinucleotid = NMMD, Nicotinamid-Purin- Dinucleotid = NPD, Nicotinamid-Adenin-Dinucleotid = NAD. - Hydrierte Formen: NMDH, NMMDH, NPDH, NADH.
1) G. WEBER, Biochem. J. 47, 114 (1950). 2 ) G. WEBER, Nature [London] 180, 1409 (1957).

206 G. PFLEIDERER, C. WOENCKHAUS, K. SCHOLZ und H. FELLER Bd. 615
tide und beim enzymatisch inaktiven, stereoisomeren a-Nicotinamid-Adenin-Dinucleotid ist dieser Effekt nicht zu beobachten. - SchlieRlich haben BOCK und Mitarbeiter3) gezeigt, daR die Gesamtabsorption von Pyridin- und Purinteil im intakten Dinucleotid gegeniiber der der Spaltprodukte erniedrigt ist.
Da die Ursache dieser Wechselwirkung zwischen Adenin und Pyridinring noch nicht geklart ist und urn weitere Aufschlusse uber die Bedeutung des Adenins im NAD zu erhalten, haben wir aus Nicotinamid-Mononucleotid Dinucleotide herge- stellt, deren Purinteil systematisch abgewandelt ist4,5), und entsprechende optische und enzymatische Messungen niit diesen Coenzym-Analoga durchgefuhrt. Hier be- richten wir uber Versuche mit Dinucleotiden, die in 6-Stellung des Purins eine Mer- capto- oder Methylrnercapto-Gruppe besitzen.
Der Grund fur die Wahl dieser Gruppen ist folgender: Eine Moglichkeit der Wechsel- wirkung zwischen den Heterocyclen bestiinde in einer Wasserstoff briickenbindung zwischen dem Substituenten in 6-Stellung des Purins und der Carbonamid-Gruppe in 3-Stellung des Pyridins, ahnlich wie in Nucleinsauren. Einen Hinweis dafiir hatte die Verschiebung des Maximums nach kiirzeren Wellen bei der reduzierten Form des Nicotinamid-Hypoxanthin- Dinucleotids4) sein konnen; hier laRt sich die OH-Gruppe jedoch nur schlecht verathern, die Wasserstoff briicke fur den notwendigen Vergleichsversuch also nur schlecht ausschalten. In Dinucleotiden mit einer 6-Mercapto-purin-Komponente kann sich nun einerseits noch leicht eine Wasserstoffbriicke ausbilden, andererseits laRt sich das entsprechende S-Methyl- Derivat darstellen, bei dem diese Bindung nicht mehr moglich ist6). - In diesem Zusammen- hang haben wir auch das Nicotinamid-Purin-Dinucleotids) untersucht, das keinen Substitu- enten am Purinring besitzt.
Zur Synthese der Purinrnononucleotide gingen wir von 6-Mercapto-9-[P-~-ribo- furanosyll-purin *) aus, das nach HAMPTON 7) zuerst in die Isopropylidenverbindung iibergefuhrt und d a m nach ROY 8) rnit Cyanathylphosphat phosphoryliert wurde. Zur Darstellung von 6- Methylrnercapto - 9 - [p - ~-ribofuranosyl] -purin- 5“- phosphat wurde die Isopropylidenverbindung vor der Phosphorylierung in alkalischer Losung mit Methyljodid rnethyliert. Nebularin-5’-phosphat erhielten wir in 40-proz. Ausbeute aus 6-Methylrnercapto-9-[~-~-ribofuranosyl]-purin-5’-phosphat durch Entschwefeln mit RANw-Nickel. Die Reaktion konnte am Verschwinden der Absorptionsbande bei 320 my verfolgt werden. Als Nebenprodukt trat Inosinsaure auf. - Die Nucleotide
*) Praparat der Fa. Zellstoff Waldhof, Mannheim. 3) J. M. SIEGEL, G. A. MONTGOMERY und R. M. BOCK, Arch. Biochem. Biophys. 82, 288
4) G. PFLEIDERER, E. SANN und F. ORTANDERL, Biochim. biophysica Acta 73, 39 (1963). 5 ) G. PFLEIDERER, C. WOENCKHAUS und K. SCHOLZ, Angew. Chem. 75, 92 (1963); Angew.
Chem. internat. Edit. 2, 43 (1963). 6) Unabhangig von uns haben M. R. ATKINSON, J. F. JACKSON, R. K. MORTON und A. R.
MURRAY, Nature [London] 196, 35 (1962), Nicotinamid-Mercaptopurin-Dinucleotid dar- gestellt, ohne es jedoch naher zu charakterisieren.
(1959).
7) A. HAMPTON und M. H. MAGUIRE, J. Amer. chem. SOC. 83, 150 (1961). 8) J. K. ROY, D. C . KVAM, J. L. DAHL und R. E. PARKS, J. biol. Chemistry 236, 1158 (1961).
1964 Intramolekulare Wechselwirkung zwischen Heterocyclen in Coenzymen, I 207
wurden durch Gradientenelution an Dowex-1-formiat chromatographisch gereinigt. Sie sind bei Raumtemperatur nicht lange haltbar. Nicotinamid-Mononucleotid ge- wannen wir stets durch Spaltung von NAD mit Kartoffel-Pyrophosphatase 9) und Abtrennung des Adenosinmonophosphats. Dieser Weg war gunstiger als die che- mische Synthese, zumal dann nur ganz wenig a-glykosidisch gebundenes Nicotinamid- Mononucleotid vorliegt. Durch Ausfallen mit Aceton konnten wir das Mononucleotid in maximal 70-proz. Ausbeute kristallin erhalten.
SR SR SR
HO
R = H bzw. CH3 I : R = H 11: R = €I 111: R = CH3 IV: R = CH3
SR
HA
VI: R = H; VII: R = CH3
V H b VIII H b 8.
9) G. W. E. PLAUT und K. A. PLAUT, Arch. Biochem. Biophysics 48, 189 (1963).
208 G. PFLEIDERER, C. WOENCKHAUS, K. SCHOLZ und H. FELLER Bd. 675
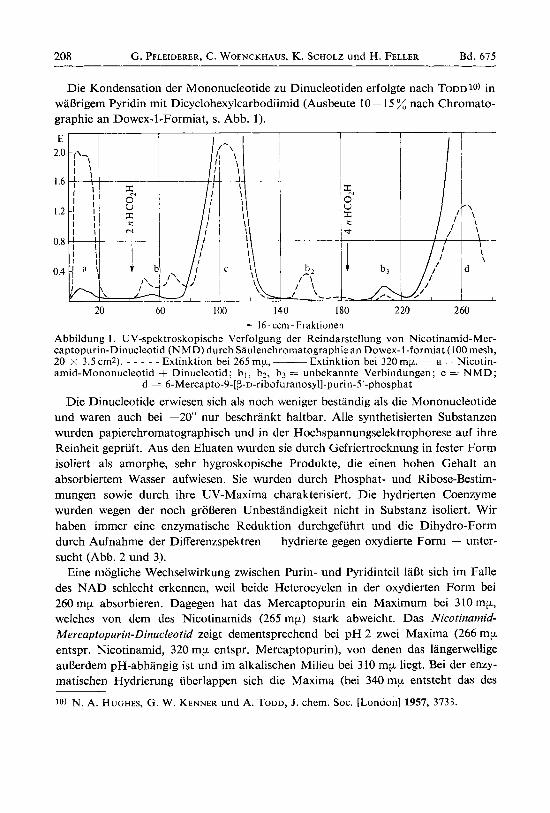
Die Kondensation der Mononucleotide zu Dinucleotiden erfolgte nach TODD 10) in wal3rigem Pyridin mit Dicyclohexylcarbodiimid (Ausbeute 10- 15 % nach Chromato- graphie an Dowex-1-Formiat, s . Abb. 1).
.\ \ \ \ -r \ \
d
I
20 60 I00 140 180 2 20 260 - 16 ~ ccm- Fraktionen Abbildung 1. UV-spektroskopische Verfolgung der Reindarstellung von Nicotinamid-Mer- captopurin-Dinucleotid (NMD) durch Saulenchromatographie an Dowex-1-formiat (100 mesh, 20 x 3.5 cm2). - - - - - Extinktion bei 265 mp, - Extinktion bei 320 mp. - a = Nicotin- amid-Mononucleotid + Dinucleotid; bl, 02, b3 = unbekannte Verbindungen; c = N M D ;
d = 6-Mercapto-9-[~-~-ribofuranosyl]-purin-5'-phosphat
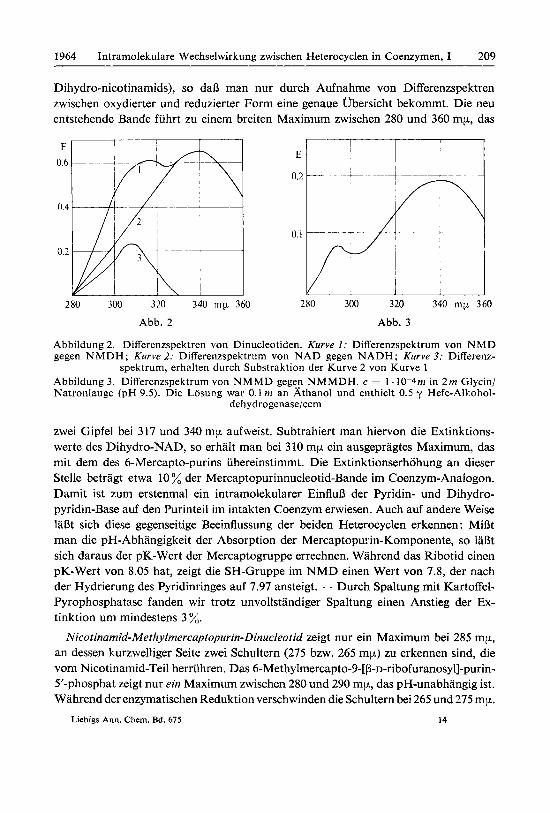
Die Dinucleotide erwiesen sich als noch weniger bestandig als die Mononucleotide und waren auch bei -20" nur beschrankt haltbar. Alle synthetisierten Substanzen wurden papierchromatographisch und in der Hochspannungselektrophorese auf ihre Reinheit gepriift. Aus den Eluaten wurden sie durch Gefriertrocknung in fester Form isoliert als amorphe, sehr hygroskopische Produkte, die einen hohen Gehalt an absorbiertem Wasser aufwiesen. Sie wurden durch Phosphat- und Ribose-Bestim- mungen sowie durch ihre UV-Maxima charakterisiert. Die hydrierten Coenzyme wurden wegen der noch groBeren Unbestandigkeit nicht in Substanz isoliert. Wir haben immer eine enzymatische Reduktion durchgefiihrt und die Dihydro-Form durch Aufnahme der Differenzspektren - hydrierte gegen oxydierte Form - unter- sucht (Abb. 2 und 3).
Eine niogliche Wechselwirkung zwischen Purin- und Pyridinteil lal3t sich im Falle des NAD schlecht erkennen, weil beide Heterocyclen in der oxydierten Form bei 260 mp absorbieren. Dagegen hat das Mercaptopurin ein Maximum bei 3 10 mp, welches von dem des Nicotinamids (265 mp) stark abweicht. Das Nicotinumid- Mercuptopurin-Dinuclotid zeigt dementsprechend bei pH 2 zwei Maxima (266 mp entspr. Nicotinamid, 320 mp entspr. Mercaptopurin), von denen das langerwellige aul3erdem pH-abhangig ist und im alkalischen Milieu bei 310 m p liegt. Bei der enzy- matischen Hydrierung uberlappen sich die Maxima (bei 340mp entsteht das des
10) N. A. HUGHES, G. W. KENNER und A. TODD, J. chem. SOC. [London] 1957, 3733.
1964 Intramolekulare Wechselwirkung zwischen Heterocyclen in Coenzymen, I 209
Dihydro-nicotinamids), so da13 man nur durch Aufnahme von Differenzspektren zwischen oxydierter und reduzierter Form eine genaue Ubersicht bekommt. Die neu entstehende Bande fiihrt zu einem breiten Maximum zwischen 280 und 360 mp, das
280 300 320 340 m p 360
Abb. 2 Abb. 3
Abbildung 2. Differenzspektren von Dinucleotiden. Kurve I : Differenzspektrum von NMD gegen NMDH; Kurve 2: Differenzspektrum von NAD gegen NADH; Kurve 3: Differenz-
spektrum, erhalten durch Substraktion der Kurve 2 von Kurve 1 Abbildung 3. Differenzspektrum von NMMD gegen NMMDH. c = 1.10-4rn in 2 m Glycin/ Natronlauge (pH 9.5). Die Losung war 0.1 rn an Athano1 und enthielt 0.5 y Hefe-Allcohol-
dehydrogenase/ccm
zwei Gipfel bei 317 und 340 mp aufweist. Subtrahiert man hiervon die Extinktions- werte des Dihydro-NAD, so erhalt man bei 310 mp ein ausgepragtes Maximum, das mit dem des 6-Mercapto-purins iibereinstimmt. Die Extinktionserhohung an dieser Stelle betragt etwa 10 % der Mercaptopurinnucleotid-Bande im Coenzym-Analogon. Damit ist zum erstenmal ein intramolekularer EinfluB der Pyridin- und Dihydro- pyridin-Base auf den Purinteil im intakten Coenzym erwiesen. Auch auf andere Weise la& sich diese gegenseitige Beeinflussung der beiden Heterocyclen erkennen : MiBt man die pH-Abhangigkeit der Absorption der Mercaptopurin-Komponente, so la& sich daraus der pK-Wert der Mercaptogruppe errechnen. Wahrend das Ribotid einen pK-Wert von 8.05 hat, zeigt die SH-Gruppe im NMD einen Wert von 7.8, der nach der Hydrierung des Pyridinringes auf 7.97 ansteigt. - Durch Spaltung mit Kartoffel- Pyrophosphatase fanden wir trotz unvollstandiger Spaltung einen Anstieg der Ex- tinktion um mindestens 3 %. Nicotinamid-Methylmercaptopurin-Dinucleotid zeigt nur ein Maximum bei 285 mp,
an dessen kurzwelliger Seite zwei Schultern (275 bzw. 265 mp) zu erkennen sind, die vom Nicotinamid-Teil herriihren. Das 6-Methylmercapto-9-[~-~-ribofuranosyl]-purin- 5'-phosphat zeigt nur ein Maximum zwischen 280 und 290 mp, das pH-unabhangig ist. Wahrend der enzymatischen Reduktion verschwinden die Schultern bei 265 und 275 mp.
Liebigs Ann. Chem. Bd. 675 14

210 G . PFLEIDERER, C. WOENCKHAUS, K. SCHOLZ und H. FELLER Bd. 675
Durch die Absorptionsbande des Purinteils wird die des Dihydropyridin-Spektrums jedoch noch teilweise uberlagert, so daR auch hier ein Differenzspektrum zwischen der oxydierten und der hydrierten Form aufgenommen werden muDte. Wie beim Dihydro- nicotinamid-Mercaptopurin-Dinucleotid zeigen sich dann auch hier zwei Gipfel (340 bzw. 293 mp). Hier betragt der Extinktionsanstieg ca. 10% der Extinktion der oxydierten Form. Durch Spaltung rnit Schlangengift-Pyrophosphatase und -Nucleo- tidase erhohte sich die Extinktion des oxydierten Coenzym-Analogons um ca. 10 %.
Das Nicotinamid-Purin-Dinucleotid weist ein Maximum bei 263 mp auf. Die redu- zierte Form hat ein sich von dem des naturlichen Dihydro-NAD kaum unterscheid- bares zweites Maximum bei 340 mp.
Biochemische Untersuchungen rnit den beschriebenen NAD-Analoga sollten Hin- weise auf die Bindungsstellen des Adeninteils mit dem Apoenzym geben. Daher haben wir von allen drei Substanzen die MICHAELIS-Konstanten im Hefe-ADH-System nach der graphischen Methode von LINEWEAVER und BURK 11) ermittelt. Innerhalb der Fehlergrenzen sind die Werte nahezu identisch (3 -6.10-4 m) und stimmen rnit der Konstanten des naturlichen P-NAD uberein. Unter vergleichbaren Testbedingungen waren in zahlreichen Dehydrogenasesystemen die Reaktionsgeschwindigkeiten des NMD trotz der sauren SH-Gruppe denen des normalen NAD sehr ahnlich. Machten schon diese Befunde eine immer wieder auftauchende Theorie, die Aminogruppe in 6-Stellung des Purins wurde von funktionellen SH-Gruppen einzeliier Dehydrogenasen gebunden werden, unwahrscheinlich, so durfte die hohe Bindungsfestigkeit des NPD vollends gegen diese Hypothese sprechen.
Bei kinetischen Messungen rnit dem Mercaptopurin-Analogon des NAD machten wir die Beobachtung, daR die Mercaptogruppe leicht eliminierbar ist. In Gegenwart von waRrigem Hydrazin bei pH 9.5 verschwindet beim Stehenlassen die Absorptionsbande bei 3 10 mp. Wir erwarteten daher eine nucleophile Substitutionsreaktion auch mit polaren Gruppen des aktiven Zentrums der Lactatdehydrogenase (LDH), nachdem wegen der geringen Dissoziation des ternaren NMD-Sulfit-LDH-Komplexes das Coenzym nahe an entsprechende Stellen herangefiihrt wird. Tatsachlich beobachteten wir sowohl bei Inkubation reiner LDH rnit Sulfit und NMD als auch mit NMMD eine langsame, nicht kompetitive irreversible Hemmung des Apoenzyms, die auch bei lang andauernder Dialyse nicht aufgehoben wurde. Nach an- schlieBender Gefriertrocknung und Elektrophorese konnte man auf dem Pherogramm rnit Methylathylketon/Ammoniak deutlich das fluoreszierende Coenzym fest an die Amido- schwarz-positive Bande gebunden erkennen. Wir hoffen, mit dieser Technik nahere Aufschlusse uber die Wirkgruppe der LDH gewinnen zu konnen.
11) H. LINEWEAVER und D. BURK, J. Amer. chem. SOC. 56, 658 (1934).

1964 Intramolekulare Wechselwirkung zwischen Heterocyclen in Coenzymen, 1 21 1
B E S C H R E I B U N G D E R V E R S U C H E
6-Mercapto-9-[2'.3'-isopropyliden-~-~-ribofuranosyl]-purin (I). - I wurde nach MAGUIRE und H A M P T O N ~ ~ ) hergestellt.
6-Mercapto-9-[~-~-ribofuranosyl]-purin-5~-phosphat (11). - 0.324 g I in 10 ccm Pyridin wurden rnit 4 ccm einer Cyanuthylphosphat-Losung (nach TENER l 3 ) hergestellt) versetzt. Es wurde bei 30" zur Trockne eingeengt, rnit 10 ccm iiber Calciumhydrid getrocknetem Pyridin aufgenommen und wiederum bei 30" i. Vak. eingeengt. Dieser TrocknungsprozeR wurde noch zweimal wiederholt. AnschlieBend wurde in 10 ccm absol. Pyridin gelost, rnit 2 g Dicyclohexyl- carbodiimid versetzt und iiber Nacht bei ca. 20" stehengelassen. Uberschiissiges Dicyclohexyl- carbodiimid wurde rnit 20 ccm Wasser zerstort. Nach 20 Min. wurde bei 30" i. Vak. zur Trockne eingeengt. Der Riickstand wurde rnit 40 ccm 0.4n LiOH 30 Min. unter RiickfluB erhitzt. Nach dem Abkiihlen wurde filtriert und das Filtrat auf eine Dowex-50-Saule (He- Form) gegeben. Das saure Eluat wurde bei 30" i. Vak. auf 3 ccm eingeengt und rnit gesattigter Ba(OH)2-Losung auf pH 7.5 eingestellt : Anorganisches Phosphat fiel aus und wurde sofort abzentrifugiert. Aus dem Uberstand kristallisierte das Ribotid 11 als Bariumsalz aus. Nach Stehenlassen iiber Nacht bei 0" wurde abfiltriert und eine zweite Fraktion durch Zugabe des zweifachen Volumens Athanol ausgefallt. Nach 2stdg. Stehenlassen bei 0" wurde abgesaugt, mit Athanol und Ather gewaschen und i. Vak. getrocknet. ~ Zur Reinigung wurde das Bariumsalz in Wasser suspendiert und rnit wenig Dowex-50 (He-Form) in Lijsung gebracht. Die Mischung wurde auf eine Dowex-50-Saute (He-Form) gegeben und das Ribotid rnit Wasser eluiert. Das Eluat wurde eingeengt, auf eine Dowex-1-Formiat-Saule gegeben und das Ribotid durch Gradienten-Elution rnit Ameisensaure steigender Konzentration gewonnen. Es kam erst rnit 2n Ameisensaure von der Saute. Die Eluate wurden im Zms-Spektralphoto- meter bei 310 m y oder im Uvicord gemessen. Durch Gefriertrocknung wurde die Verbindung in festem Zustand erhalten: gelbes, amorphes Produkt.
6-Methylmercapto-9-[2'.3'-isopropyliden-~-~-ribofuranosyll-purin (111). - 1 g I in 8 ccm 0.4n NaOH wurde rnit 0.2 ccm Methyljodid versetzt. Es wurde einige Minuten geschiittelt, und zur triiben Suspension wurden weitere 1.2 ccm 0.4n NaOH und 0.2 ccm Methyljodid hinzugegeben. Nach Stehenlassen iiber Nacht war das zunachst gebildete bl vollstandig kristallisiert. Aus Xylol 1 g (95 %) Kristalle vom Schmp. 103-107".
Cl4H18N404S (338.4) Ber. C 49.7 H 5.4 N 16.6 S 9.5 Gef. 49.8 5.6 16.6 9.8
6-Methylmercapto-9-~~-~-ribofuranosyl]-purin-5~-phosphat (IV). - Darstellung analog dem Ribotid 11. Das Bariumsalz kristallisierte nicht aus Wasser, es muBte mit k h a n 0 1 gefallt werden (Ausbeute 60% d. Th.). Das freie Ribotid wurde durch Gradienten-Elution an Dowex- I-Formiat gereinigt. Es fiel in 85-proz. Reinheit an. E = 15.0.106 [cmz/Mol]; Phosphat :Ri- bose = 0.9 : 1. Die Verbindung ist vie1 instabiler als 11. Schon beim Trocknen der Chromato- gramme zersetzte sie sich zu einer fluoreszierenden Verbindung.
9-[B-~-RibofuranosylJ-purin-5'-phosphat (V). - 0.59 g IV in 50 ccm Wasser wurden rnit 2n NaOH auf p H 7 gebracht. Es wurde 1 g frischbereitetes Raney-Nickel (Aktiv.-Stufe W2)
12) A. HAMPTON und M. H. MAGUIRE, J. Amer. chem. SOC. 83, 150 (1961). 13) G. M. TENER, J. Amer. chem. SOC. 83, 159 (1961).
14*

212 G. PFLEIDERER, C. WOENCKHAUS, K. SCHOLZ und H. FELLER Bd. 675
hinzugegeben und unter RiickfluB gekocht. Die Entschwefelung wurde UV-spektroskopisch verfolgt. Sie ist beendet, wenn das Maximum bei 285 mp verschwunden ist (u. U. nach Zusatz frischen RANEY-Nickels). Dann wurde filtriert und mit heiBem Wasser gut nachgewaschen. Das Filtrat zeigte pH 7.4. Es wurde i. Vak. bei 30" eingeengt und die konzentrierte Losung auf eine Dowex-I-Formiat-Saule gegeben. Das Ribotid wurde durch Gradienten-Elution mit Ameisensaure gereinigt. Ausbeute 200 mg (39 %). Ribose : Phosphat = 1 : I . Amax = 263 mp, E = 7.5 106 (pH = 4).
Nicotinamid-Mercaptopurin-Dinucleotid (VI). - 200 mg 6- Mercapto-9-[B-~-ribofuranosyl]- purin-5'-phosphat und 200 mg Nicotinamid-Mononucleotid in 16 ccm Wasser wurden mit 53 ccm Pyridin und anschlieBend rnit 6 g Dicyclohexylcarbodiimid versetzt. Nach 24 Stdn. wurde vom Dicyclohexylharnstoff abfiltriert und dieser mit 3 ccm heil3em Wasser gewaschen. Filtrat + Waschwasser wurden rnit 3 g Dicyclohexylcarbodiimid versetzt. Dieser Vorgang wurde so oft wiederholt, bis innerhalb von 24 Stdn. nur noch wenig Harnstoff ausfiel. Die Pyridinlosung wurde in das gleiche Volumen Wasser eingegossen und 2 Stdn. geriihrt. Vom Unloslichen wurde filtriert, der Ruckstand griindlich ausgewaschen. Filtrat + Waschwasser wurden 4mal mit je 100 ccm Chloroform ausgeschiittelt, die waBr. Phase wurde i. Vak. bei 30" auf 3 ccm eingeengt. Dann wurde auf eine Dowex-I-Formiat-Saule (2 x 30 cm) gegeben. Nach ca. 100- 150 ccm Wasserdurchlauf wurde mit Ameisensaure steigender Konzentration eluiert. Im Eluat wurde die Extinktion bestimmt ; aus entsprechenden Fraktionen wurden durch Gefriertrocknung die Coenzym-Analoga in fester Form erhalten. Ausbeute 40- 60 mg (10-15%; gelblich). Ribose:Phosphat = 1 :1 (79% Ribose, 80% Phosphat); RF = 0.55 (khano l : Wasser : Eisessig = 50 : 50 : 0.03) ; Cyanid-Konstante: 1.5.10-2.
Oxydierte Form: A,,, = 310 my (pH 9.5), 320 mp (pH 2); E = 23.2.106 (pH 9.9,
Reduzierte Form: A,,, = 310 m p (pH 9.5) bzw. 320 mp (pH 2.0), 266, 273 mp; ~ 3 1 0 =
Nicotinamid-Methylmercaptopurin-Dinucleotid (VII). - Darstellung analog VI ; VII wurde wie I1 erst mittels 2n HC02H von der Saule eluiert. Ausbeute: 10-15 % d. Th. (fast farb- 10s). Ribose :Phosphat = 1 : 1 (64% Ribose, 66% Phosphat). RF = 0.71. Cyanid-Konstante:
25.1 .106 (pH 2).
(28.1 & 0.6).106 (PH 9.5).
1.05.10-2. Oxydierte Form: Amax = 286 mp; E = 16.6.106 (pH 9.5). Reduzierte Form: A,,, = 286, 337 my; E286 = 18.8.106 (pH 9.5). Nicotinamid-Purin-Dinucleotid (VIII). - Darstellung analog VI. Die Verbindung wurde
schon mit 0.5n HC02H von der Saule eluiert. Das Dinucleotid war farblos; Ausbeute: 26% (bez. auf eingesetztes Mononucleotid). Ribose : Phosphat = 1 : 1 (75 % Ribose, 75 % Phosphat). Cyanid-Konstante: 0.98.10-2.
Oxydierte Form: A,,, = 263 my; E = 11.3 106 (pH 9.5). Reduzierte Form: A,,, = 263, 339 mp; ~ 2 6 3 = 7.8'106 (pH 9.5).




![Photoschaltbare Resorc[4]arene - uni-bielefeld.de...Photoschaltbare Resorc[4]arene Photochrome Supramolekulare Wirtsysteme durch intramolekulare [4+4] Cycloaddition von Anthracen Dissertation](https://img.pdfslide.org/doc/110x75/6080a426c1c70419872280eb/photoschaltbare-resorc4arene-uni-photoschaltbare-resorc4arene-photochrome.jpg)






