Embed Size (px)
Citation preview

Ein Versuch zur statistisch-graphischen Untersuchung und Analyse der zeitlichen Eigenschaften der Regene-
rationsverg ,nge. Von
Jal'oslav K~i~eneck~ ~.
Aus dem Institut ftir allgemeine Biologie und experimentelle Morpho- logie an der medizinischen Fakult~tt der biihmischen Universitiit in Prag.
Mit 1 Figur im Text, 3 Tabellen und 1 Kurventafel XXXVII.
Eingegangen am 28. Januar 1916.
W~hrend die morphologische Seite der Regenerationsvorgange ich meine hier den ersten Typus der Restitution, nKmlich die
Regeneration in engerem Sinne oder die sog. ,Regeneration durch Sprossung, naeh Rovx (1905, S. 83) - - sehon seit 1Kngerer Zeit einer genauen Untersuchung und Analyse immer wieder unterzogen wird, sind die ze i t l i ehen Eigensehaften, welchen besonders in der letzten Zeit beim normalen Wachstum eine groBe Aufmerksamkeit gewidmet wurde, bei den Regenerationsprozessen fast vollstandig ohne Beachtung geblieben. Man hat zwar untersucht, wie verschiedene auBere Ein- flUsse, wie Licht, Temperatur, verschiedenfarbige Strahlen, Radio- aktivit~tt, Elektrizit~t, Chemismus des Mil ieus . . . usw. auf die Re- generation Uberhaupt und auch speziell auf ihre Geschwindigkeit einwirken (vgl. darUber BARFURTH, 1910, S. 14--16), abet der Ver- such, den Regenerationsvorgang als solehen in seinen zeitlichen Eigen- sehaften zu analysieren, wurde noch nieht ausgefUhrt.
In seinem BUchlein Uber die Anwendung elementarer Mathematik auf biologisehe Probleme ist zwar PRZIBRAM (1908) auf diese Frage ein wenig eingegangen, aber seine AusfUhrungen beziehen sich teil- weise auf andere Erseheinungen, z. B. auf den EinfluB der Regene- ration auf die H~tutungsgeschwindigkeit, teilweise, soweit sie den

Ein Versueh zur statistisch-graphisehen Untersuehung und Analyse usw. 623
eigentliehen Regenerationsvorgang betreffen, sind sic bloB hypothetiseh, rein abstrakt-mathematiseh begrUndet. Was abet die Experimente betrifft, auf welehe sieh PRZIBRAM bezieht, go waren diese damals noch nieht abgesehlossen, wie mir n~tmlieh PRZmRAM brieflieh mit- geteilt hat. Ubrigens werde ieh auf PRZmRAMS AusfUhrungen noeh sp~tter eingehen.
Die Versuehe, Uber welche im folgenden beriehtet werden soil, warden auf Grand folgender Betraehtungen ausgefUhrt: wird yon paarigen Organen (z. B. Extremit~tten) das eine zum Teil entfernt, dann besteht seine Regeneration darin, dab es sieh zur GrfiBe des anderen, welches unberUhrt geblieben ist, wiederherstellt. Es ist dabei gleieh, oh dieses normale Organ w~thrend der Regeneration des auderen weiterw~tehst oder nieht; immer erreieht das regenerierende Organ naeh einer gewissen Zeit die Grt~lle des normalen, nieht ent- fernten, ungeaehtet ob dies mittels einfachen Waehstums des Regene- rates ;in dem Falle, wenn das normale Organ dabei nieht weiter- w~tehst), oder mittels b e s e h l e u n i g t e n Waehstums im Gegensatz zu dem normalen des niehtentfernten Organes vor sieh geht. lqur selten finden Abweiehungen yon diesem Schema statt: KAMMERER beobaehtete z. B. bei Ciona (zitiert naeh SEMO~, 1912~ S. 58--59), weleher er einen Siphon entfernt hatte, dab dieses bei seiner Regeneration gr~ller wird als das normale, MEGU~AR (1907) hat umgekehrt naeh Entfernung einer Mandibel bei dem Wasserk~fer Hydrocharis caraboides eine Verkleinerung der anderen festgestellt.
Die zitierten Beobachtungen bilden jedoch nur eine Ausnahme yon der Regel, dab die paarigen Organe unabh~tngig voneinander regenerieren, und lassen sieh his zu einem gewissen Grade als funk- tionelle Sekund~trerseheinungen erkl~tren: die yon i~IEGU~An beobaehtete Mandibelreduktion k~nnte z. B. ganz gut als umgekehrte Parallele zur kompensatorisehen Hypertrophie der lqiere nach Entfernung der anderen, also sozusagen als kompensatorisehe Atrophic, betraehtet werden.
l~iemals warden solehe kompensatorische Korrelationen an Fallen beobaehtet. Aus diesem Grunde warden die Ftil~e, ebenso wie z. B. die Ftihler bei Arthropoden, ein sehr gUnstiges Objekt zum Stadium des Regenerationsverlaufes abgeben, denn, gibt es bier wirklieh keine Korrelationen, so ist uns die Vergleiehung des Regenerates mit dem anderen nieht operierten Fulle oder FUhler gut mtiglieh.
Aus diesem Grunde habe ieh meine Versuche an Fallen aus- gefiihrt.

624 Jaroslav Kiitieneck~-
Das gestattet uns den RegenerationsprozeB unter Anwendung des sog. �9 R e g e n e r a t i o n s q u o t i e n t e n , zu studieren. Der Regenerations- quotient ist durch das ~Verhaltnis der GrSBe des normalen FuBes zu der des operierten in einem bestimmten Zeitintervall naeh der Ope- ration gegeben. Bezeichnen wir die GrSBe des normalen Organes zur Zeit t mit ~Vl~ die GrSBe des Regenerats zu derselben Zeit mit Rt, dana ist der Regenerationsquotient
Kt-:N. Rt
Die Feststellung dieses Regenerationsquotienten kann stattfinden, ohne dab man darauf sieht, ob der normale FuB noeh weiter waehst oder nieht, denn derselbe bestimmt~ wieweit das Regenerat auf seinem Wege~ das Verlorengegangene wiederherzustellen, gelangt ist, wobei das Verlorengegangene dureh den Zustand des normalen FuBes: bzw. seine GrSBe gegeben wird. Der Regenerationsquotient bedeutet also die y e r h a l t n i s m a B i g e Geschwindigkeit der Regeneration. Ist nam- lieh ein Regenerat naeh einer bestimmten Zeit mehr oder minder ausgewaehsen, regenerierte es schneller oder langsamer, dann ist auch der Regenerationsquotient entweder kleiner oder grSBer - - die GriiBe des Regenerates steht namlich, da sie sich im ~enner der den Quotienten bezeichnenden Formel befindet, zu der GriJlle des Regene- rationsquotienten, also zur Regenerationsgeschwindigkeit~ in negati- vem Verhaltnis, bzw. sie verh~tlt sich umgekehrt proportional zu der letzteren.
Auf der anderen Seite ist es aber miiglich, mittels dieses Re- generationsquotienten zu untersuchen~ ob in Fallen~ in welchen wahrend der Regeneration noeh ein Wachstum des normalen Organes, in unserem Falle des anderen~ nicht operierten FuBes stattfindet, der RegenerationsprozeB langsamer oder schneller vor sich geht, als in den Fallen, in welehen das normale Organ nieht mehr weiter waehst und umgekehrt. ~atUrlieh ermSglieht die Feststellung des Regene- rationsquotienten zu untersuchen, mit welcher Gesehwindigkeit die Regeneration versehiedenster p a a r i g e r Organe vor sich geht.
Meine Versuehe babe ieh an Larven yon Tenebrio molitor aus- gefUhrt und, wie sehon bemerkt~ als Objekt die FUBe derselben ge- wahlt.
Die praziseste Methode fUr meine Zweeke ware wohl, die Grtil~e des Regenerates sowie aueh des entspreehenden normalen Ful]es in gewissen Perioden, bzw. nach jeder Hautung gleichzeitig zu messen

Ein Versueh zur statistiseh-graphischen Untersuehung und Analyse usw. 625
und dureh das Verh~tltnis der beiden den Regenerationsquotienten far diese oder jene bestimmte Zeit nach der Operation festzustellen. Leider zeigte sich diese Methode bei meinem Objekte als nieht an- wendbar. Die FUBehen der Tenebrio-Larven sind sehon normaler- weise sehr klein; yon den Regeneraten derselben gilt das noeh in erh0htem MaBe, so dab eine genauere Messung dieser Objekte ohne ihre Entfernung yon der Larve nicht ansgefUhrt werden kann.
Um dieser Sehwierigkeit zu entgehen, bediente ich reich der Messung der Dimensionen yon normalen und regenerierten FUBen bei den Puppen , wo diese betriichtlich griiBer sind, bzw. die Dimen- sionen der imaginalen sehon besitzen. Das war aus dem Grunde miiglieh, weil heute als Tatsache feststeht, dab bei den Inse.kten- puppen keine Regeneration oder weiteres Waehstum des Regenerates stattfindet - - obzwar ihre detailierte Differenzierung nicht geleugnet worden soll - - ; es repr~tsentieren ni~mlich die Insektenpuppen naeh PRZrBIr Terminologie (vgl. PRZIBR.&~-WEI~BER~ 1907, S. 623) einen �9 vo l lkommenenr und , e r w a e h s e n e n , Zustand. Man kann also die Verpuppung als ein Stadium auffassen, in welehem die Regene- rationsprozesse unterbroehen sind. Bis zu dem Momente der Ver- puppung besitzt die Insektenlarve die Fahigkeit, ihre verlorenge- gangenen GliedmaBen zn regenerieren; nach der Verpuppung geht diese Fiihigkeit verloren. Den Puppen beigebraehte Defekte werden niebt mehr dutch Wiederherstellung des Verlorengegangenen restituiert, sondern nut dureh eine Umordnung des Gewebes in der n~ehsten Umgebung der Wunde repariert, wie ich dies genauer fur die FlUgel- anlagen (K~ti~C.~ECK~:, 1914) auseinandergesetzt habe.
Deswegen ist es aueh begreiflieh, warum Tiere, welehe als jUngere Larven operiert wnrden, im Imaginalzustande eine voll- kommenere Regeneration zeigen, als Tiere, welche als iiltere Larven der Operation unterworfen worden sind: Die i~lteren Larven ver- puppen sieh viel frUher naeh der Operation als die jtingeren und haben deswegen eine viel kUrzere Zeit zur Wiederherstellung des Verlorengegangenen als die jUngeren zur Disposition 1). Es entspreehen
t) De facto seheinen aber diese Verh~ltnisse nieht ausschliel31ieh so einfach zu sein, denn die ~ilteren Larven regenerieren nieht nur deswegen unvollst~tndiger, wr ihnen kiirzere Zait dazu vergt~nnt ist als den jiingeren. Richtig hat darauf KOPE~ hingewiesen (1913), dab die ,Erscheinung, dal3 die Regenerationsi~,ihigkeit mit dem Alter der Raupen abnimmt, wahrscheinlieh nicht aussehliel31ich durch die abnehmende Zahl der zur Regeneration verfiigbaren Tage, sondern aUem Ansehein naeh gleiehzeitig dureh die mit dem Alter der Tiere zustande kommenden

626 Jaroslav. K~'~eneek~
also die Verhaltnisse bei den Pnppen in gewissem Sinne den Ver- h~Itnissen an der Larve, welche sich zu dieser Puppe verwandelt hat, wenigstens ist das Verh~ltnis des normalen Fu~es zu dem regene- rierten, also der Regenerationsquotient, yon demselben Wert.
Es ist also mSglich, den Regenerationsquotienten mittels Messung an Puppen festzustellen, an welehen die letztere wegen der betr~eht- licheren GreBe der PuppengliedmaBen leiehter als bei den Larven auszu- fiihren ist, wobei die Zeit, welehe zwischen der Operation and der Ver- puppung der operierten Larven verflossen ist, als sog. ~Regenerations- zeit, angesehen werden kann. In meinen Versuchen schnitt ich also den Larven den FuB dicht am KSrper ab und habe das Regenerat erst, als sich die Lar~en zur Puppe verwandelt haben, gemessen and mit dem normalen FuB derselben verglichen and damit den Regene- rationsquotient bestimmt.
Es ist bekannt, dab bei Tieren, welehe als Larven l~ngere Zeit zum Regenerationsprozesse zur VerfUgnng haben, endlich der verloren- gegangene FuB vollst~ndig wiederhergestellt wird; in einem solchen Falle ist der Regenerationsquotient gleieh 1, denn im :Nenner wie im Z~hler des Bruehes
Bt finden sich gleiche Werte. Je kUrzere Zeit aber die Tiere regene- rieren kSnnen, desto grSBer wird der Regenerationsquotient sein, denn sein Wert ist. wie sehon oben gesagt wnrde, der GrSBe des Regene- rates umgekehrt proportional. Bestimmen wir den Regenerations- quotienten gleieh naeh der Operation, so sehen wir, dab er unendlieh groB ist, da wir n~mlieh bei seiner Bestimmung eine reale Zahl mit :Null zu dividieren haben, well ja die GrS~e des Regenerates gleich naeh der Operation dem Werte :Null entsprieht. Es gilt also: K o = c ~ ,
denn N o ist einer realen Zahl, /~o aber :Null gleich, also a O
Ist aber t ~ O dann besitzt K t einen realen Weft and zwar: ist t2 ~ tl, dann gilt, dal] Ks ~ KI.
Warden wir also die Verandernngen des Regenerationsquotienten mit verschiedenen Regenerationszeiten graphisch darstellen, auf die
Veriinderungen ihrer physiologischen Eigenschaften ableitbar,, (S, 465) ist; es ist n~mlich die kleinere Regenerationsfiihigkeit ~lterer Larv~en in gewissem Grade auch dutch ihr absolut gr~13eres Alter als solches vernrsacht, l~iiheres dariiber vgl. in meiner Abhandlung aus dem Jahre 1914, {S. 190--192). Doch erscheint abet dieser absolute Einflu~ des Alters verh~ltnism~iBig klein, so da~ man ihn im folgenden vernachliissigen kann.

Ein Versueh zur stat ist iseh-graphisehen Untersuehung und Analyse usw. 627
Abszisse die Regenerationszeiten, auf die Ordinate die entspreehenden Werte der Regenerationsquotienten auftragen, so bek~men wir eine regelm~Bige Kurve, welehe vom unendliehen Durehsehnittspunktd mit der Aehse Y (Ordinate) - - der Wert des Regenerationsquotienten naeh tier Operation ist n~mlieh gleieh c ~ - langsam herabsinkt, bis sie endlieh in tier H~he derOrdinate ~ I mit tier Aehse X parallel verl~uft, was bedeutet, dab das Regenerat sehon die GreBe des nor- malen FuBes erreieht hat. Je fr|iher nun diese Kurve - - man kann sie ~ R e g e n e r a t i o n s k u r v e ~ nennen - - die Ordinate yon Weft ~--- 1 erreieht, desto raseher muflte sieh fie Regeneration abspielen, denn desto frUher erreiehte alas Regenerat die GreBe des normalen FuBes. Es kann also der Abfall der Regenerationskurve als Kriterium der Regenerationsgesehwindigkeit betraehtet werden: je sehroffer dieser Abfall tier Regenerationskurve ist, ttesto sehneller mnBte der Verlauf der Regeneration gewesen sein. Man kann also sehon aus tier bloBen Form der Regenerationskurve auf die Regenerations- gesehwindigkeit urteilen.
Bei meinen Versuehen handelte es sieh erstens datum, festzustellen, ob diese obzitierten logiseh konstruierten Vorstellungen riehtig sind oder niehti die zweite Aufgabe war, mittels dieser Regenerations- kurven zu untersnehen, ob sieh nieht etwa bei den Insekten ein Analogon zu der yon Z~LENY (1903 und 1905) an Seesternen und Crustaeeen besehriebenen Erseheinung feststellen lieBe, dab naeh Ent- fernung yon mehreren K~rperanh~ngen die Regeneration raseher vor sieh geht, als naeh Entfernnng eines einzigen. WUrde n~mlieh aueh bei den Insekten eine ~hnliehe Erseheinung stattfinden, so mtiBte die Regenerationskurve far einen FuB, wenn gleiehzeitig noeh das eine oder die beiden anderen FuSpaare mit entfernt worden w~ren, einen schrofferen Abfall aufweisen, als wenn nnr der eine Fu~ allein ent- fernt worden ware.
Es hat zwar schon MEGU~AR (1907) bei seinen Regenerations- versnehen an Coleopteren einigemal gefunden, daB, wenn mehrere Extremit~ten entfernt wnrden, die Regeneration sehneller vor sieh ging. Da es sieh dabei aber nur um einige vereinzelte Beobaeh- tungen an wenigen Individuen handelt, so ist es nieht gut m~glieh, daraus einen allgemein gtiltigen SehluB za ziehen, besonders wenn man die groBe individuelle Variabilit~t tier Regenerationsf'~higkeit bei den Insekten, welehe schon mehrere Exper imenta to ren- beson- ders z. B. JANDA, KOPE(~ - - betont haben, erw~gt. Dazu sind en masse ausgeftlhrte nnd statistiseh bearbeitete Experimente n~tig, in

628 Jaroslav KH~.eneek~
welehen man alle individuellen Abweichungen und Sehwankungen berUeksiehtigen wUrde.
Bei meinen Experimenten habe ieh drei Serien yon Larven ver- sehiedensten Alters operiert, und zwar in jeder Serie in drei Gruppen: bei der einen Gruppe sehnitt ich bloB den linken FuB des hinteren Paares ab, bei der zweiten auiSerdem ~oeh den linken FuB des mitt - leren Paares, and in der dritten Gruppe die linken FUBe aller drei Paare.
Diese dritte Gruppe ergab in keiner Serie benutzbare Resultate: es kam ni~mlich naeh Absehneiden aller drei FiiBe zu einem groBen Blutverlust, und aus der ausflieiSenden und gerinnenden Blutlymphe hat sieh fast am ganzen AbdomGn Gin miichtiger WundverschluB aus-
t t
o o..--"
Schema des PuppenfuBes yon Tenebrio molitor. Die Kreuzchen )< zeigen die Punkte, you wel- ehen die Messung d-_r einzelaen Fuf~teile ausge-
ffihrt wurde. a L~.nge des Femurs, b L~.nge der Tibia,
c L~nge des Tarsus.
gebildet, weleher die Larven ver- hinderte, sieh zu hauten, so dab sie bald zugrunde gingen. We dies nicht geschah, waren die Regenerate miB- bildet, so daiS es vollstandig unmiig- lieh war, sie zu meinem Zwecke zu benUtzen. •ur die Versuehe in der ersten und zweiten Gruppe aller Serien ergaben positive Resultate. In jeder Gruppe wurden je 50 Lar- ven versehiedensten Alters operiert, um in den versehiedensten Perioden naeh der Operation Puppen zu be- kommen. Leider herrsehte in alien Serien eine groBeMortalitat--wahr-
seheinlieh infolge yon Wundinfektion, so dab ieh aus 150 operierten Larven in einem Falle nur 17, im anderen 19 Puppen bekommen konnte. An diesen Exemplar~n wurden die Messungen in der oben angedeuteten Weise ausgefUhrt.
Gemessen wurden Femur, Tibia und Tarsus, jedes fur sich. Es war niimlieh meine Absieht, die Regeneration aller dieser Teile ein- zeln zu studieren, leider zeigte sich aber mein Material als ungenti- gend zu diesem Zweek. Aueh ist Tenebrio molitor night das pas- sendste Objekt zu solehen Untersuehungen~ da aueh bei den Puppen die Dimensionen der FUBe, besonders der regenerierten, zu klein sind, was natUrlieh yon ihren einzelnen Bestandteilen in noeh hiiherem MaBe gilt. Geeigneter dUrften z.B. Odonateu, Mantiden usw. sein. Gemessen wurde in den auf der Textfigur dargestellten Linien; dureh Addition

Ein Versuch zur statistisch-graphisehen Untersuchung and Analyse usw. 629
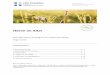
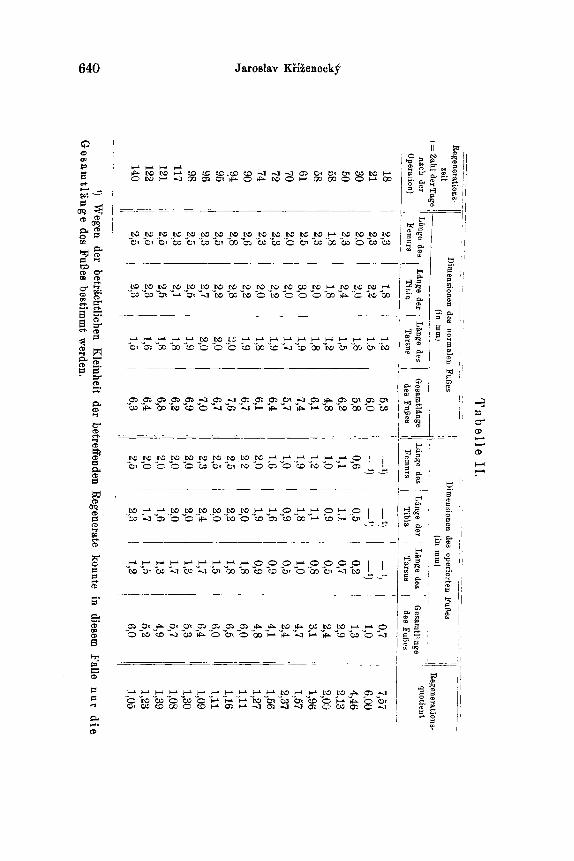
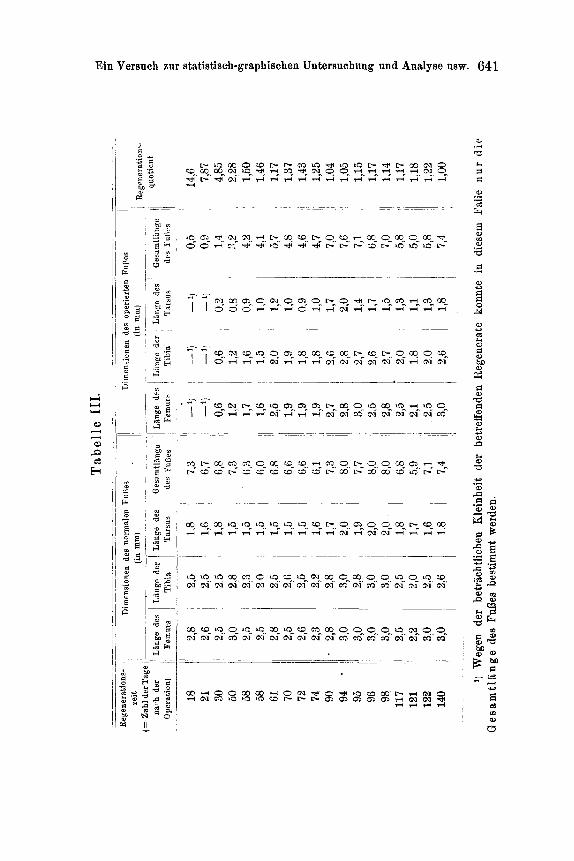
der Femur-, Tarsus- und Tibialiingen wurde die Gesamtl~tnge des ganzen FuBes festgestellt. Aber bei den kleinsten Regeneraten war es nicht mSglich, sich eines gewShnlichen, auf Millimeterzehntel ge- teilten MaBstabes zu bedienen; da benutzte ieh zur direkten Messung der Gesamtliinge des FuBes REICI-IERTS Objektivmikrometer (2 mm lung geteilt in 200 Teile) mit Hilfe eines Pr~iparationsmikroskopes. Die Resultate dieser Messungen sind in den Tabellen I - - I I I nieder- gelegt; auf Grund derselben wurden auch die Graphen (Tar. XXXVII) konstruiert 1).
Diese Resultate haben erstens die obzitierten logisehen Vorstel- lungen~ welche ich mir tiber den Zusammenhang des Regenerations- quotienten mit der Regenerationszeit gebildet habe, als richtig be- sti~tigt. Die graphische Darstellung dieses Zusammenhanges ergab in der Tat eine regelm~tBige Kurve, welche veto unendlichen Durch- schnittspunkte mit der Achse Y (Ordinate) langs~tmer herabsinkt, his sie endlieh in der HShe der Ordinate ~ 1 mit der Achse X parallel verl~iuft; dies gilt sowohl fur den Fall, dab bloB ein FuB yon dem hinteren l%are entfernt wurde, als aueh fUr den Fall, dab aui~er diesem noch ein FuB des mittleren Paares mit entfernt wurde, and zwar in gleieher Weiss fUr den FuB des hinteren oder mittleren Paares~ wie aus den betreffenden Regenerationskurven auf der Ta- fel XXXVII zu ersehen ist.
Was die Frage anbetrifft, ob in dem Falle, dab mehrere Glied- mai%n entfernt wurden, die Regeneration schneller vor sich geht 7 wie dies ZELE~Y 1903 und 1905 bei den Seesternen und Crustaeeen beobaehtet hat~ so geht aus meinen Versuchen nichts derartiges her- vor. Hiitte in dem Falle, in welchem auBer dem FnBe des hinteren Paares noeh ein FuB aus dem mittleren Paare entfernt wurde, eine schnellere Regeneration des ersteren stattgefunde.n, dann wUrde sich dies dutch den schrofferen Abfall der Regenerationskurve kundgeben. Vergleichen wir aber die Graphen I u. III (Tar. XXXVII), so erkennen wir, dab beide Kurven im allgemeinen den gleichen Abfall zeigen, dab also in beiden Fallen die gleiehe Regenerationsgeschwindigkeff geherrscht haben muBte. Ja, man kiinnte sogar noch die Meinung aus- spreehen, dab in dem Falle, we noch ein FuB aus dem mitfleren raare mit entfernt worden war (Graph III, Taf. XXXV]I), die Regeneration
l) Die Eintragung der Regenerationsquotienten in die Graphen geschah unter Korrektion yon der zweiten Dezimalstelle, so da6 z.B. der berechnete Regenerationsquotient 2,37 im Graphe als 2,4 eingetragen ist, wobei 1 bier gleich 10 mm an der Ordinate ist; an der Abszisse bedeutet 1 mm einen Tag.
Archiv f. Entwicklungsmechanik . XLII. 4~0

630 Jaroslav Ki'i~eneck~
des FuBes aus dem hinteren Paare langsamer vor sich gegangen ist. ttier erreieht namlieh die Kur~e den parallelen Verlauf mit der Abszisse ein wenig spater, als in dem Falle, in welehem bloB der eine FuB entfernt worden ist (Graph I, Tar. XXXVII). Es ist jedoeh nicht ausgesehlossen, dab dieseErscheinung elne s ekund i i r e ist, dadurch verursaeht, dab in dem Falle, wo zwei FUBe entfernt wurden, ein grSBerer Blutverlust stattgefunden hat, weleher die Ausbildung eines maehtigeren Wundverschlusses zur Folge hatte, so dab die Hautung der Larven sehr erschwert war; dadurch konnte endlieh die Vitalitat der Larven schwer gesehitdigt und natUrlich auch der Verlauf des Regenerationsprozesses verz(igert werden.
Vergleiehen wir den Graph I[ (Taf. XXXVII), weleher die Regene- rationskurve eines FuBes des mittleren Paares darstellt, mit dem Graph I, weleher den FuB des hinteren Paares betrifft, so sehen wit, dab die Kurve I[ in deutlieher Weise weniger schroff ist, als die Kurve I, ja dab sie eigentlieh den parallelen Verlauf mit der Abszisse in der HShe der Ordinate ~ 1 tiberhaupt nicht erreicht, und ich habe auch keinen einzigen Fall erhalten, bei welehem der Regenerationsquotient den Weft 1 erreicht hatte. Dies zengt augen- seheinlieh davon, dab der Regenerationsverlauf des FuBes des mitt- leren Paares entgegen den Fi|Ben des hinteren Paares langsamer war, denn der geringere Abfall der Regenerationskurve kann nut dureh die langsamere Abnahme des Regenerationsquotienten verursacht sein. Der Grund dieser Erseheinung ist - - da der Regenerationsquotient das Verhaltnis der Gri~Be des normalen FuBes zu der des operierten darstellt, wobei sich die erstere im Zahler, die zweite im l'~enner be- finder - - in der geringeren Gr(iBenzunahme des Regenerates zu er- blicken. Ginge die Regeneration schneller vor sich, dann niihme aueh die GriiBe des Regenerationsquotienten raseher ab und der Ab- fall tier Regenerationskurve ware ein schrofferer.
Es ist also klar, - - soviel glaube ieh aus meinen Befnnden sehlieBen zu dUrfen - - dab bei Tenebrio die FtiBe des mittleren Paares langsamer regenerieren als die des hinteren Paares. Dieser Untersehied ist ein primarer und nicht sekundiir, z. B. dureh un- gUnstige Waehstumsverhaltnisse naeh sehwerer Verwundung, infolge m~iehtigen Blut~erlustes and daran sieh knUpfender schwieriger Hitu- tung verursacht. Das geht daraus hervor, daB, wenn hier solche ungUnstige Verhiiltnisse ins Spiel gekommen waren, dieselben zwei- fellos aueh das hinters FuBpaar getroffen und in gleicher Weise in s~incr Regeneration und seinem Waehstum beeinfluBt h~itten, wie das

Ein Versuch zur statistisch-graphischen Untersuchung und Analyse usw. 631
mittlere. Die primaren Unterschiede zwisehen den beiden FuBpaaren sind also in meinen Versuehen unberUhrt geblieben. Daher sind die Unterschiede zwischen der Regenerationsgeschwindigkeit des hinteren und mittleren FuBpaares bei Tenebrio als primar zu betraehten.
Die festgestellte Erscheinung, dab dis Regeneration des FuBes des mittleren Paares langsamer vor sich geht, als die des Ful]es vom hinteren Paare, ist besonders deswegen yon Bedeutung, well sie im Gegensatze zu der yon CmLD (1909) gefundenen Regel steht, dal~ ~the rapidity of restitution decrease with increasing distance from the oral end of the body% welche Regel neuerdings yon Umscrl (1915) auch fiir die Insekten, namlich Larven yon C/o~, bestatigen konnte. Er land, dab das RegenerationsmaB um so gr(il3er ist , je weiter das Regenerat nach vorn liegt, dab namlich die Femora des vorderen Paares schneller reg~nerierten als die Femora des mittleren Paares und diese wieder schneller als die des hinteren Paares, dab also, entspreehend der CmLoschen Regel, die Regenerationspotenzen mit der Entfernung vom oralen K(irperende abnehmen.
Auf Grund meiner eigenen Befunde kann ich diese Regel fur Tenebrio nicht bestatigen: hier zeigten sieh die Verhaltnisse eben umgekehrt; die mehr naeh vorn liegenden GliedmaBen regenerierten langsamer, als die mehr naeh hinten liegenden. Es ware aber - - eben wegen dieser D i f f e r e n z - sehr erwUnscht, wenn man diese Umsti~nde einer erneuten genaueren Untersuchung unterzieben wUrde, als es mir auf Grund meines Materiales m~iglieh war. Besonders ware die Regeneration des FuBes des mittleren Paares aueh in dem Falle zu untersuehen, wenn nur dieser selbst entfernt wurde; dies ware - - meiner Ansicht naeh - - unter Benutzung meiner hier beschrie- benen statistisch-graphisehen Methode keine sehwere Aufgabe. - -
Wie schon oben erwahnt wurde, hat schon einmal PRZiBRA)L (1908) versucht, die zeitlichen Eigenschaften des Regenerationsver- laufes analytisch zu studieren. Er ging aber yon einem anderen Standpunkte aus. PRZIBRAM vergleicht, indem er yon seiner Ansieht ausgeht, dab die Regeneration nur besehleunigtes Wachstum dar- stellt, nicht die GriiBe des normalen und regeneriel'enden Organes in einzelnen Perioden, wie ieh, sondern ihr~ Waehstumsgeschwindig- keit. Sonst abet geht er in derselben Weise vor wie ich, bestimmt auch das Verhaltnis des normalen und regenerierenden Organs, wel- ches er dann zur graphischen Darstellung benutzt ; nut darin differiert PrtZmRAM Yon mir, dab er im Ziihler die Wachstumsgesehwindigkeit des Regenerates (l~.), im Nenner die Wachstumsgeschwindigkeit des
42*

632 Jaroslav Ki'i~eneck~
normalen Org.anes (V~c) anbringt, w~ihrend ieh mit den Gr~Ben des Regenerates und des normalen Organes in umg.ekehrter WeisG ver- fahre. Ebenso wie bei meiner Methode tier Reg.enerationsquotient,
so nimmt aueh bei PRZIBRAMS Methode der Quotient Vr mit fort-
sehreitender Regeneration ab und, wenn das Regenerat die Gri}l~e des normalen Organes erreieht, wird dieser Quotient g.leieh 1 - yon diesem Punkte ab waehsen n~mlieh sowohl alas normale Organ, als aueh das reg.enerierte mit gleicher Gesehwindigkeit fort.
Bei der graphisehen Darstellung erh~tlt 19RZIBRA~t eine Kurve, die yon der Aehse Y lang.sam herabsinkt, um naeh Erreiehung. des Wertes 1 horizontal zu verlaufGn. Darin hat PRZm~A~I vollst~tndig reeht, aber so wie er diese Kur~e auf S. 38 seines BUehleins auf Grund yon VersuGhen an einer Krabbe darstellt, dUrfte dieselbe wohl kaum verlaufen; es ist nieht anzunehmen, dab sie die Aehse Y dureh- sehneiden wtirde, wie es PRZIBRA3[ zeiehnet.
Dies ergibt sieh aus der folgenden Erw~tgung: PRZIBr~A3I g'ibt zwar nieht an, wig er die Regenerations- bzw. Waehstumsgesehwin- digkeit bestim'mt hat, aber wenn wir yon der physikalisehen Defini- tion ausg.ehen, dab n~tmlieh die Gesehwindigkeit die in Zeiteinheit durehlaufene Bahn bedeutet, dann ergibt sieh die Reg.enerationsge- sehwindig.keit als der Zuwaehs eines Org.anes bzw. das Verhaltnis dieses Zuwaehses - - da wir hier nut mit verh~tltnism~tBig'en Werten reehnen k~nnen -- zu der vorherigen Grt~Be des betrefl'enden Organes in einer Zeiteinheit. Ist also ein Organ, das gestern A maB, bis heute um a ausg.ewaehsen, dann i s t seine Wachstums- oder Regene- rationsg.esehwindig.keit, wenn es sich n~tmlieh um ein Regenerat handelte, in dieser Zeit, d. h. in 24 Stunden, dutch das Verhaltnis a ]- gegeben. Betraehten wir nun den regenerierenden FuB: naeh der
Exstirpation maB dieser Null, A = O; naeh 24 Stunden reg.enerierte er zwar sehon ein wenig., wenn aueh kaum feststellbar, und ist um nnd zu a (yon einem r ea l en und positiven Wert) ausgewaehsen.
g Setzen wir denselben nun in die Formel ~- ein, so ergibt sieh,
dab in diesen ersten 24 Stunden die Regenerationsgesehwindigkeit a
eine unendliehe war, ~ - = ~ - - es ersehien hier etwas, was hier
vorher night war; V,. betr~tg.t also in den ersten 24 Stunden naeh der Operation den Wert ~ . In denselben 24 Stunden ist der nor-

Ein Versueh zur statistiseh-graphischen Untersuchung und Analyse usw. 633
male, frUher A' messende FuB u m a ' ausgewachsen, welehe beide Zahlen reale und positive Werte vorstellen; es war also die Wachs- tumsgesehwindigkeit des normalen Organes in den ersten 24 Stunden
a t
~%~ = ~ , also einem ebenso positiven und realen Werte gleieh. Setzen
wir denselben nun in die Formel far das Verh~tltnis der Reg.enerations- und Wachstumsg.esehwindigkeit, dcssen Weft die GrSl~e der Ordinate
vorstellt, so bekommen wir das Verh~iltnis V~, was wieder ~ er-
g.ibt. Es betr~g.t also Y fur X ~ 1 den Weft cr fur den Fall, dab O
.X ~ 0, ist Y, mathematisch genommen ~ gleich, stellt also einen
unbestimmten Wert vor. Mit anderen Worten: auch die PRZlBaAM- sche Kurve schnitt, ebenso wie die meinig'e, weder die Achse X, noch die Achse Y, denn schon fUr x = 1 ist die Ordinate y---~ c~. Beide Kurven trag.en also, obzwar sic auf Grnnd verschiedener Methoden konstruiert worden sind, denselben Charakter und dieselbe Form.
Ans beiden Kurven, scwohl aus der, welehe auf Grund meiner eigenen, als aueh aus der, welche auf Grund yon 1)RZIBRAMS Methode konstruiert worden war, l~tl3t sich eine Parallele zwischen der Re- generation und dem normalen Wachstum feststellen.
Das Waehstum ist schematisch als ein negativ autokatalytiseher Vorg.ang anzunehmen, was bedeutet, dab die verhi~ltnismM~igen, z. B. prozentualen Zuwiichse in gleichen Zeiteinheiten - - a l s o die Wachs- tumsgesehwindig.keit - - sich mit dem fortschreitenden absoluten Waehs- turn regelmaBig verkleinern, so dab sic endlich, nach Einstellnng. des
Waehstums, l~ull gleiehen. Besonders MlXOT (1906, vg.1. aueh 1913 hat5 das Wachstum yon digser Seite her einer eingehenden Analyse unterzogen.
Dasselbe erg.ibt sieh auch ftir die Regenerationsvorg.iing.e und zwar aus 1)ICZIBt~AMS Methode unmittelbar, da er die Reg.enerationsg.eschwin- digkeit direkt bestimmt hat. Aus meiner Methode aber ergibt sieh dies auf Grund folg.ender Erw~g.ung.: Sollte die Gesehwindig.keit der
Regeneration dauernd die g.leiehe ble, iben, dann mtiBte sieh das Re- generat dem normalen Organe in jeder Zciteinheit und verhaltnis- mM,~ig, um dasselbe ann~hern; in diesem Falle wUrde sieh also die Reg'enerationskurve nieht als Kurve, sondern als eine g.erade sinkende Linie darstellen lassen. De facto ist aber die Regenerationskurve eine wirkliehe regelmiil~ige Kurve .-- vielleieht wird einmal jemand

634 Jaroslav KH~eneek)'-
aueh ihre analytiseh-geometrische Formel feststellen ~) - - , deren Form und Lage den Achsen gegenUber klar und zweifellos ergibt, daft die erwahnte Annaherung mit fortschreitender GrSl~enzunahme des Re- generates regelm~tfiig abnimmt, woraus auf eine entsprechende Ab- nahme der Regenerationsgeschwindigkeit zu schlieBen ist. Es besteht also zwischen dem normalen Wachstum und der Regeneration, was ihren dynamischen Charakter anbetrifft, eine vollstiindige Parallele, beide sind n~tmlich als negativ autokatalytisehe Vorgange anzunehmen.
Es kann das auch nicht anders sein. Sind doch diese beiden Vorgi~nge, sowohl das normale Wachstum als auch die Regeneration, in ihren Grundlagen einer und derselben Natur, in beiden handelt es sieh um Reproduktion und Differenzierung, welche Fiihigkeiten zu den Grundeigenschaften des lebenden Plasmas gehSren. Bei dem Fortschreiten jeder organischen Differenzierung kommt es gleich- zeitig immer zu einer parallel fortschreitenden Bildung hSherer und komplexerer, dutch den Stoffwechsel schwerer angreifbarer EiweiB- stoffverbindungen, wie RI~IZICKA (1908) fur niedere einzeltige 0rga- nismen, besonders aueh Bakterien, und far Tritone und Frtisehe ge- zeigt hat; dies hat aber eine Herabsetzung aller Assimilationsvorgitnge zur Folge, da die genannten Verbindungen mit fortsehreitender Diffe- renziernng des Organismus eine immer stabilere Natur gewinnen2). Diesen Vorgang, der eine Teilerseheinung des Tatsaehenkomplexes des morphologisehen Metabolismus bildet, hat Rfi~I~KA a l s , Hysteresis<< des Protoplasmas bezeiehnet und als Hauptursaehe des Alterns be- wertet. Im Liehte 'd ieser Tatsaehe erseheint uns also die negative Autokatalyse der Lebensvorg~inge als ein automatiseher Vorgang. Zugleieh ist aueh selbstversti~ndlieh, dab in jedem Falle, in welehem es zu einer Differenzierung und Entwieklnng lebender Substanz
1) Auf den ersten Blick scheinen racine Regenerationskurven ebenso wie die Kurve von I)RZIBRAM Ityperbolen bzw. H~lften yon Hyperbolen vorzustellen, bei welchen die Achsen X und Y als Assymptoten zu betrachten w~iren. Trotz- dem handelt es sich nicht um solche: ich versuchte schon einige Male, diese Kurven nach der assymptotischen Gleichung der Hyperbole (flit unseren Fall: x �9 y ~ Konst.) zu berechnen, aber immer vergeblich, der Wert x �9 y ergab nie- mals eine Konstante, sondern nahm mit zunehmendem x immer ab. Es wird sich hier also mn Kurven hiiherer "Ordnung handeln, obzwar dabei noch die M~iglichkeit often bleibt, da• ihre Formel vielleicht eine unendliche sein wird, wie dies z. B. bei der Kurve der Liingenausdehnung bei Temperaturver~inde- rungen der Fall zu sein scheint.
2) Siehe diesbeziiglich auch RS~I(~KA8 Arbeiten ,Kausal-analytische Unter- suchungen fiber die Herkunft des Chromatins. I.~ und >>Besehleunigung der Hiiutung durch Hunger<,. (Dieses Arch. Bd. 42.)

Ein Versuch zur statistisch-graphisehen Untersuchung und Analyse usw. 635
kommt, aueh solehe negativ autokatalytisehe Vorg~nge - - welehe sieh besonders in z e i t l i e h e r H i n s i e h t ~ u B e r n - stattfinden mnssen. Ihre Ursaehe ist wohl eben in der Anh~ufung der erw~hnten kom- plexen, stabilen EiweiBstoffverbindungen zu suehen, worin naeh Rfi- ~t~iX (1908, 1910) aueh die Ursaehe des >~Alterns~ zu erblieken ist, ob es sieh nun um das Altern eines ganzen Organismus, oder bloB eines seiner Bestandteile (eines Organes) handelt. Deswegen ist die Abnahme der Waehstums- und Regenerationsgeschwindigkeit eben aueh ein Anzeiehen des ,Alterns,~.
Was die regenerierenden Organe anbetriff~, so scheint, dab bei ihnen das ~Altern~ schneller vor sieh geht. Es nimmt n~imlieh bei der Regeneration die Waehstumsgesehwindigkeit in raseherem Tempo ab~ als bei normalerweise waehsenden Organen. Dies geht aus dem Folgenden hervor: W~re die Abnahme der Regenerationsgesehwindig- keit der Abnahme der Waehstumsgesehwindigkeit gleieh, dann mUBte das Regenerat schlieBlieh grSBer werden, als das normale Organ der anderen Seite, wie dies z. B. bei tier Ascidie naeh KAMMERE~S Be- fund tats~ehlich aueh der Fall ist. Denn das Reffenerat weist, im ~ergleieh mit dem entspreehenden normalen Organe, stets eine be- tr~ehtlieh gr~l~ere Waehstumsgesehwindigkeit auf. 1%rmalerweise finden wit aber, wie z. B. auch bei den Arthropoden-FtiBen,. dab das Regenerat nur die GrtiBe des normalen Organes erreicht, dann aber entweder mit derselben Geschwindigkeit wie dieses w~iter zunimmt, oder mit ihm gemeinsam wachst. Daraus ist wohl za schlieBen, daB dis Waehstumsgesehwindigkeit beim ~Regenerat schneller abnimmt, als bei dem normalen Organe.
Dies ist aber nur selbstverstSndlich: denn dadurch, dab das Regenerat schneller als das normale Organ w '~ ichs t - yon 1)ltzInaA~l wurde die Regeneration als b e s c h l e u n i g t e s Wachstum aufgefaBt - - leistet es auch eine grSBere >~metabolistische Arbeit~ als das normale Organ in derselben Zeit. Es mUssen in ihm auch intensivere Stoff- wechselvorgiinge vor sieh gehen, mit diesen geht Hand in Hand auch dis m~ehtigere Bildung und Ablagerung der komplexen und stabilen EiweiBstoffverbindungen. Diese aber muB eine schnellere Abnahme der Wachstumsgesehwindigkeit zur Folge haben.
Als Parallele mSchte ich hier Ct~ILDS Versuehe an Planarien (1911, 1914) anfUhren: CHILD beobaehtete, dab bei jungen Tieren eine grSBere Resistenz gegen Alkohol herrscht als bei alteren; er bringt diese Erscheinung mit den Unterschieden in der Intensiti~t des Stoffweehsels zusammen, der bei den jUngeren Tieren miichtiger ist

636 Jaroslav Ki'i~.eneek:~
ale bei den iilteren. LiiBt man die Planarien (such Jilters) hungers, dann kann man beobachten, dab bei ibsen in den ersten Phases tier Inanition die Resistenz gegen den Alkohol sich bis zu dem Grade vergr(iBert, wie s~e bei den jUngeren Tieren sich findet. Das ist zwar, wie Rll~I~KA aufmerksam.gemacht hat1), ass den Ergebnissen der Methode CHILDS nicht direkt zu entnehmen, erseheint jedoeh ganz verst~ndlich, denn wie wit aus anderen Versuehen wissen (vgl. BAn- ~)IER), nimmt in der ersten Zeit des Hungerns dis Intensit~it des Stoff- weehsels zu. Die hungernden Planarien zeigen also eincn intensi- veren Stoffwechsel, womit such ihrs griiBere Resistenz gegen Alkohol zusammenhiingt, was such bei den jUngeren Tieren der Fall ist. Bald nimmt abet diese Resistenz bei den hungernden Tieren ab, so dab die ausgehungerten Planarien hinsichtlich ihrer Resistenz dem Alkohol gegsnUber den alten Tieren nahestehen; sie besitzen such ein seniles Aussehen. l~ach der erst'en ,Veliitingung~ des Organismus durch Inanition finder also bald sine ~Alterung,, start, die rascher eintritt ale Enter normalen Verh~tltnissen. (Siehe die AnsfUhrungen yon Rfi~ISKA, Besehleunigung der Hi,stung dureh Hunger. Dieses Arehiv. Bd. 42.)
Dies liiBt sich nur dadurch versti~ndlich maehen, dab mit dem intensiveren Stoffwechsel in den ersten Phases des Hungerns zu- gleieh sine machtigere Ablagerung jener stabileren EiweiBstoffver- bindungen, w~lche doch als Produkte des Stoffwechsels anzusehen sind~ stattfindet,.was zu siner schnelleren ,Alterung,, des Organismus flihren muB. Jedes intensivere Leben mnB also notwendigerweise ein schnelleres AltErs zur Folge babes. Umgekehrt wieder muB Enter EinflUssen, die eine Herabsetzung der Intensitiir der Stoffwechssl- vorgange verursaehen (wie z. B. niedere Temperatur), eine Verliinge- rung des Lebens stattfinden. Von diesem Standpunkte aus erscheint uns LoEss Lehre von dem das Alters verzSgernden Einflusse der niederen Temperatur in vollst~ndigem Einklange mit Rfi~ISKAS kau- saler Auffassung des Vorganges des Alterns, n~imlieh: die Intensit~it und L~nge des Lebens sind indirekt proportional untereinander. Alles, was sin intensiveres Leben des Plasmas herbeifiihrt, beschleunigt das Altwerden und Herbeifuhren aller dasselbe begleitenden Phiino- msne, in erster Reihe also eine Retardation des Wachstums End zu der daran geknUpften Entwicklung. Alles also, was den Lebensvorgang
t) Vgl. dazu such die Arbeit yon R~I~I~KA, Besehleunigung der H'2utung dutch Hunger. Diesos Arehiv. Bd. 42.

Ein Versuch z u ~ s t a t i s t i s - c h - g r a p h i s c h e n Untersuchung und A n a l y s e u s w . 637
beschleunigt, wird ihn zugleieh verkUrzen; mit vollem Recht sagt
also KAMMERER (1910): , D a s Wachs tum an sich ist rascher in der
Witrme; aber die Fi ihigkei t zu waehsen bleibt liinger erhal ten in der
K~tlte. In dieser Be leuchtung erscheint uns - - um zu unserem eigenen
Thema zurUckzukehren - - das schnel lere Al tern eines regener ie renden 0 r g a n e s als eine vol lkommen k lare und selbstvers tandl iche Erschei-
hung: die Regenera t ion repr~tsentiert sich uns als eine Beschleunigung
des Waehstums uud der E n t w i c k l ung ; sic bedeute t also eine Besehleu-
nigunff aller Lebensvorgange , des g'anzen Metabolisinus, und hicrmit
notwendigcrweise auch eine Beschleunig'unff des Alterns.
P r a g , im Mai 1915.
Literaturverzeichnis,
BARDIER, E., Inanition. Darts ,Dictionaire de physiotogie,( par ChARLeS RICHET. Premier fascicule du tome 9. Paris, F. Alcan.
BARFURTH, D., Die Regeneration und Transplantation in der Medizin. (Samml. Anatom. u. Physiol. Vortr. u. Anfs. Heft 10.) Jena, Fischer. 1910.
CrIIiD, CH. ,~I., Factors of Form Regulation in Harenactis attenuata. Journ. of Exper. Zool. Vol. 6. 4. 1909.
- - A Study of Senescence and Reiuvenescence based on Experiments with Planaria dorotocepbala. Arch. f. Entw.-Mech. Bd. 31. 1911. Starvation, Rejuvenescence and Acclimation in Planaria dorotocephala. Arch. f. Entw.-Mech. Bd. 38. 1914.
KA~IMERER, Das Beibehalten jugendlich uureifer Formzust'Xnde (Neotenie und Progenesel. Ergebnisse der wi~senschaftlichen Medizin. 1910.
KOPEr STm'rL, Untersuchungen fiber die Regeneration yon Larvalorganen und Imaginalscheiben bei Schmetterliegen. Arch. f. Entw.-Mech. Bd. 37. 1913.
K~,I~E~ECK~, JAR., Experimentelle und theoretische Untersuchungen fiber die Restitution der Insektenflfigel. Arch. f. Entw.-Meeh. Bd. 39. 1914.
LOEB, J., Uber den Temperaturkoeffizienten fiir die Lebensdauer kaltblfitiger Tiere und die Ur.-~che des natfirlichen Todes. Pr'L/3GERS Arch. f. d. ges. Physiol. Bd. 124. 1908.
MEGU~AE~, FR., Die Regeneration bei Coleopteren. Arch. f. Entw.-Mech. Bd. 25. 1907.
MINOT, CH. S., The problem of Age, Growth and Death. New York 1908. - - Moderne Probleme der Biologie. Jena. Fischer. 1913. 0STWALD, WOLFG., Uber die zeitlichen Eigensehaften der Entwicklungsvorg~inge.
(Vortr. u. Aufsiitze fiber Entw.-Mech. d. 0rg. Heft 5.) Leipzig, Engelmann. 1908.
PI~ZIBRA~, H., Quantitative Wachstumstheorie der Regeneration. Zentralbl. f. Physiol. Bd. 19. 1905.
- - Anwendung elementarer Mathematik auf biologische Probleme. (Vortr. u. Aufs. fiber Entw.-Mecb. d. 0rg. Heft 3.) Leipzig, Engelmann. 1908. Experimental-Zoologic. 2. Regeneration. Leipzig u. Wien, Deuticke. 1909.

638 Jaroslav Ki'i~eneck~
PRZIBRAM, H., und WERBER, E. J., Regenerationsversuche allgemeinerer Bedeu- tung bei Borstenschwiinzen (Lepismatidae). Arch. f. Entw.-Mech. Bd. 23. 1907.
Roux, W., Die Entwicklungsmechanik, ein neuer Zweig der biologischen Wissen- schaft. (Vortr: u Aufs. tiber Entw.-Mech. d. Org. Heft 1.) Leipzig, Engel- mann. 1905.
R~I~I~KA, VLAD., Zur Kenntn[s der Natur und Bedeutung des Plastins. Arch. f. Zellforsch. Bd. 1. 1908. S. 600ft. Das Chromatin und Plastin in ihren Beziehungen zur Regsamkeit des Stoff- weehsels.' Festschr. f. R. HERTWIG. Jena, Fischer. 1910. S. 63. THEOD. SCltWAN~ Cas. 15k. 5esk. Prag 1910.
SEMen, R., Das Problem der Vererbung ,erworbener Eigensehaften,. Leipzig, Engehnann. 191"2.
vo~ UBIS(~, LEOP., Uber den Einflu~ von Gleichgewichtsstiirungen auf die Regenerationsgeschwindigkeit (Versuche an Cloe diptera). Arch. f. Entw.- Mech. Bd. 41. 1915.
ZELE~u CH., Tim Relation of the Degree of Injury to the Rate of Regene- ration. Journ. of Exper. Zoology. Vol. 2. 1905.
- - - - A Study of the Rate of Regeneration of the Arms in the Brittle star, 0phioglypha lacertosa. Biolog. Bullet. Vol. 6. 1903.-
Erkl~irung der Abbildungen, Tafel XXXVII .
i. G rap h: Regenerationskurve des Fui3es des hinteren Paares in dem Falle, da~ dieser allein entfernt wurde. Zahlenangaben hierzu siehe in Tabelle I (S. 639).
II. G r a p h : Regenerationskurve des Fui3es des mittleren Paares in dem Falle, da~ auch ein FuI~ des hinteren Paares mit entfernt wurde. Zahlenangaben hierzu siehe in Tabelle ]i (S. 640).
I I I G r a p h : Regenerationskurve des Ful3es des hinteren Paares in dem Falle, da~ auch ein Ful3 des mittleren Paares mit entfernt wurde. Zahlenangaben hierzu siehe in Tabelle III (S. 6411.
Reg
ener
atio
ns-
zeit
{=
Zah
l d~
r T
age
I na
ch
der
Ope
rati
on}
16
21
25
34
40
44
46
51
58
62
70
71
86
100
114
117
121
Tab
elle
I.
Dim
ensl
onen
(lo
s no
rmal
en
FnB
es
(in
mm
} !!
Lgu
ge d
er
Tib
ia
2,6
2,8
2,4
2,9
2,3
2,6
2,5
2,6
2,5
2,3
3,0
2,6
2,5
2,8
:3,0
2,
3 2,
2
Ls
des
Tar
sus
1,9
1,0
1,6
2,0
1,8
1,8
1,8
2,0
1,7
1,6
2,0
1,8
1,8
1,8
1,8
1,8
1,0
jl (~
esam
tl[i
nge
Lin
ge
des
des
Fur
ies
I l F
emur
s
,i 'l
7,1
I __
t) (;.
3 _
I)
6,5
, 0,
6 7,
8 !
0,9
6,5
il 0,
8 7,
1 I
1,1
7,0
:, 1,
3 7,
1 1,
8 7,
1 2,
3 6,
4 2,
0 8,
0 ,'
2,5
7,1
2,5
7,0
' 2,
1 7,
6 3,
0 i
7,8
~! 3,
0 6,
4 ,:
2,0
~,~;
!l e,4
l)im
ensi
onen
de
s op
erie
rten
F
uBes
(i
n ra
m)
Lin
ge
der
Lhn
go d
es
Tib
ia
" T
arsu
s
i __
1)
__
l ~
_ t)
__
i)
0,5
0,2
0,4
0,3
0,7
0,8
0,4
1,2
1,0
1,5
1,2
1,2
1,3
1,8 1',3
1,
0
Ges
amtl
inge
de
s F
uBes
0,5
0,7
1,3
2,2
1,9
3,0
3,0
2,8
5,2
3,8
6,5
6,0
5,1
7,1
7,8
5,3
5,6
Lhn
ge
des
Fem
urs
2,6
2,5
2,5
2,9
2,4
2,7
2,7
2,5
2,9
2,5
3,0
2,7
2,7
3,0
3,0
2,3
2,4
Reg
ener
atio
ns-
quot
ient
14,2
0 9,
00
5,00
3,
54
3,42
2,
37
2,33
2,
53
1,36
1,
68
1,23
1,
18
1,37
1,
06
1,00
1,
20
1,00
1) W
egen
de
r be
tri~
chtli
chen
Kbi
nhei
t de
r be
tref
fend
en R
egen
erat
e ko
nute
in
di
esem
Fa
lle
nur
die
Ges
amtl
~in
ge
des
FuB
es b
estim
mt
wer
den.
g = E C~q g m-
r cD
r162
Tab
elle II.
II D
imensionen de
ormalen FuB
es '
D1m
ensienen des epenerten FuB
es I
zei~ "
mm
:' "
i
nach
der
I Lan
.....
, uot*en%
(= Zahl derTage
~ ~
ln
ram
} .
..
..
..
..
!!n ram
) -
-'!1 Regenerations-
, "
go des L~nge der
Ls des
I G
esamtl~,nge
IJ Ls des
LS~n~e tier L~nge des
Gesam
tFinge ~,'
q "
Operauon)
i1,, Fem
urs TitA
a T
arsus I
des Ful~es I}
Femurs.
Tibia T
arsus des FuB
es ,,'
IJ 18
i 2,3
1,8 1,2
5,3 __ t)
-- ~) -- ~j
0,7 7,57
21 2,3
2,2 1,5
6,0 _
_
1) _
_ 1)
-- t) 1,0
6,00 30
2,0 2,0
1,8 5,8
0,6 0,5
0,2 1,3
4,46 50
2,3 2,4
1,5 6,2
1,1 1,1
0,7 2,9
2,13 58
1,8 1,8
1,2 4,8
1,0 0,9
0,5 2,4
2,0~) 58
2,3 2,0
1,8 6,1
1,2 1,1
0,8 3,1
1,96 61
2,5 3,0
1,9 7,4
1,9 1,8
1,0 4~7
1,57 70
2,0 2,0
1,7 5,7
1,0 0,9
0,5 2,4
2,37 72
2,3 2,2
1,9 6,4
1,6 1,6
0,9 4,1
1,56 74
2~3 2,0
1,8 6,1
2,0 1,9
0,9 4,8
1,27 90
2,6 2,2
1,9 6,7
22 2,0
1,8 6,0
1,11 94
2,8 2,8
2,0 7,6
2,5 2,2
1,8 6,5
1,16 95
2,5 2,2
2,0 6,7
2,5 2,0
1.5 6,0
1,11 96
2,3 2,7
2,0 7,0
2,3 2,4
117 6,4
1,09 98
2,5 2,5
] ,9 6,9
2,0 2,0
1,3 5,3
1,30 117
2,3 2,1
1,8 6,2
2,0 2,0
1,7 5,7
1,08 121
2,5 2,5
1,8 6,8
2,0 1,6
1,3 4,9
1,39 122
2,5 2,3
1,6 6,4
2,0 1,7
1,5 5,2
1,23 140
2,5 2,3
] ,5 6,3
2,5 2,3
1,2 6,0
1,05
1) Wegen
der betr~ichtlichen
Kleinheit
der betreffenden Regenerate
konute in
diesem
FaUe
aur die
Gesam
tliing
e des Ful3es bestim
mt w
erden.
Tab
elle
II
L
Reg
ener
atio
ns-
Dim
ensi
onen
des
nor
mal
en F
uBes
D
hnem
, ioa
en
des
oper
ierS
en F
uBes
ze
it
(in
ram
) (i
n ra
m)
(= Z
ahl
derT
age
I~--
-
--~
..
..
..
..
..
II
--
---
na~'
h de
r L
~nge
des
]
L~n
ge d
er
L~n
ge
des
Ges
amtl
~nge
L
~nge
des
L
iing
e de
r L
fnge
de
s G
esam
tlS
ege
! O
pera
tion
F
emur
s I
Tib
ia
Tar
sus
des
FuB
es
Ii F
emur
s T
ibia
T
arsu
s de
s Fu
B~,
s /
I, !
18
2,8
2,5
1,8
7,3
, __
1)
-- t)
--
t)
0,5
21
2,6
2,5
1,6
6,7
i __
1)
__1~
_1
) 0,
9 30
2,
5 2.
5 1,
8 6,
8 i:
0,6
0,6
0,2
1,4
i 50
3,
0 2,
8 1,
5 7,
3 1,
2 1,
2 0,
8 .'4
,2 58
2,
5 2,
3 1,
5 6,
3 1,
7 1,
6 0,
9 4,
2 58
2,
5 2
0 1,
5 6,
0 '
1,6
1,5
1,0
4,1
61
2,8
2,5
1,5
6.8
J[ 2,
5 2,
0 1,
2 5,
7 70
2,
5 2,
6 1,
5 6,
6 1,
9 1,
9 1,
0 4,
8 '
72
2,6
2,5
1,5
6,6
1,9
1,8
0.9
4,6
74
2,3
2,2
1,6
6,1
1,9
1,8
1,0
4,7
90
2,8
2,8
1,7
7,3
2,7
2,6
1,7
7,0
94
3,0
3,0
2,0
8,0
2,8
2,8
2,0
7,6
95
3,0
2,8
1,9
7,7
3,0
2,7
1,4
7,1
3,0
96
3,0
2,0
8,0
2,5
2,6
1,7
6,8
98
3,0
3,0
2,0
. 8,
0 2,
8 2,
7 1,
5 7,
0 I!
117
2,5
2,5
1,8
6,8
2,5
2,0
1,3
5,8
',I 12
1 2,
2 2,
0 1,
7 5,
9 2,
1 1,
8 1,
1 5,
0 12
2 3,
0 2,
5 1,
6 7,
1 .2
,5
2,0
1,3
5,8
140
3,0
2,6
1,8
7,4
3,0
2,6
1,8
7,4
1) W
egeu
de
r be
triic
htlic
hen
Kle
inhe
it
der
betr
effe
nden
Reg
ener
ate
konn
te
in
dies
em
Falle
G
esam
tl~n
ge
des
Ful]
es b
esti
mm
t w
erde
n.
[I i i R
egen
erat
ions
- qu
otie
nt
14,6
7,
87
4,85
2,
28
1,50
1,
46
1,17
1,
37
1,43
1,
25
1,04
1,
05
1,15
1,
17
1,14
1,
17
1,18
1,
22
1,00
nur
die
i 0~
t~
t~