Embed Size (px)
Citation preview

Aus dem Institut für Pflanzenernährung der Universität Hohenheim
Prof. Dr. V. Römheld
Einfluß der arbuskulären Mykorrhiza auf
Nährstoffaufnahme und Wachstum bei Reben (Vitis sp.)
Dissertation zur Erlangung des Grades eines Doktors
der Agrarwissenschaften vorgelegt
der Fakultät III - Agrarwissenschaften I (Pflanzenproduktion und Landschaftsökologie)
der Universität Hohenheim
von
Matthias Petgen
Diplom-Agraringenieur aus Saarburg
Stuttgart - Hohenheim, Juni 1998


Die vorliegende Arbeit wurde im Fachbereich Phytomedizin der Staatlichen Lehr- und
Forschungsanstalt für Landwirtschaft, Weinbau und Gartenbau (SLFA) in Neustadt/
Weinstraße unter der Leitung von Herrn Dr. F. Louis und Betreuung durch Herrn
Dr. A. Schropp in Zusammenarbeit mit dem Institut für Pflanzenernährung der Uni-
versität Hohenheim unter der Leitung von Herrn Prof. Dr. V. Römheld angefertigt. In den
ersten beiden Versuchsjahren wurde die Arbeit wissenschaftlich von Herrn Prof. Dr. Drs. h. c.
H. Marschner (†) betreut.

DANKSAGUNG
An dieser Stelle möchte ich Herrn Prof. Dr. Drs. h. c. H. Marschner posthum für die
Zusammenarbeit, sein stetiges Interesse und seine anregende und kritische Diskussion an der
vorliegenden Arbeit danken. Für die weitere wissenschaftliche Betreuung und die gute
Zusammenarbeit danke ich Herrn Prof. Dr. V. Römheld vielmals.
Für die Überlassung des Themas und die bereitwillige Betreuung der Arbeit an der SLFA
Neustadt danke ich Herrn Dr. A. Schropp. Herrn Dr. F. Louis danke ich für seine großzügige
Unterstützung im Fachbereich Phytomedizin und seine Hilfsbereitschaft.
Herrn Dr. N. Merkt vom Institut für Obst-, Gemüse- und Weinbau, Fachbereich Weinbau an
der Universität Hohenheim, danke ich für die ständige Diskussionsbereitschaft und
Unterstützung im Bemühen um wissenschaftliche Kontakte und Gedankenaustausch.
Für die wertvollen fachlichen Anregungen und Diskussionen vom Beginn bis zum Ende
meiner Arbeit bedanke ich mich herzlich bei Herrn Dr. E. George vom Institut für
Pflanzenernährung der Universität Hohenheim.
Folgende Mitarbeiterinnen und Mitarbeiter der SLFA Neustadt haben wesentlich durch
Anregungen, Ratschläge und tatkräftige Hilfe zum Gelingen der Arbeit beigetragen, bei denen
ich mich an dieser Stelle bedanken möchte: Frau I. Johann bei der Verarbeitung des
Probenmaterials und der Mineralstoffanalysen; Herrn M. Jutzi für die Hilfe bei den
statistischen Auswertungen der Versuchsergebnisse; Herrn W. Klein und Herrn M. Schreieck
für die Betreuung der Ozonversuche; Herrn Dr. F. Schumann sowie Herrn B. Ziegler für die
Überlassung von Versuchsflächen; Herrn R. Rueff und Herrn C. Lauria für die Unterstützung
bei der Erstellung von Grafiken; Herrn Dr. K.-J. Schirra, W. Köglmeier, P. Lambert und A.
Schmidt-Tiedemann für die kritische Durchsicht des Manuskripts; bei meinem Freund und
Büronachbarn A. Kopf für die oftmals aufmunternden Worte während der Tiefphasen der
Arbeit sowie seine hilfreiche Diskussionsbereitschaft; stellvertretend für die Rebveredlung
Herrn H. Wadle für Unterstützung bei der Betreuung der laufenden Versuche; allen
Hilfskräften bei der schweißtreibenden Wurzelprobenahme im Weinberg.
Nicht zuletzt gilt mein besonderer Dank meiner Freundin Vera, die mich in allen Phasen der
Arbeit motiviert hat und viel Verständnis gezeigt hat. Ganz besonders danken möchte ich
meinem Vater, der durch seine Förderung meine Ausbildung ermöglicht hat und der durch
seinen Glauben an mich und meine Arbeit sehr zum Gelingen der Arbeit beigetragen hat.
Ich danke dem Forschungsring des Deutschen Weinbaues (FDW) bei der Deutschen
Landwirtschafts-Gesellschaft, der die vorliegende Arbeit finanziell unterstützt hat.

LEBENSLAUF
Name: Matthias Petgen Geburtsdatum: 22.03.1970 Geburtsort: Saarburg Familienstand: ledig Staatsangehörigkeit: deutsch
Schulbildung:
1976 - 1980 Grundschule in Perl
1980 - 1989 Privates St.-Josef-Gymnasium in Biesdorf (Eifel)
Studium:
Oktober 1989 - Studium der Allgemeinen Agrarwissenschaften an der Oktober 1994 Universität Hohenheim, Fachrichtung: Pflanzenproduktion
Prüfungen:
Mai 1989 Erwerb der Allgemeinen Hochschulreife
Juni 1993 Landwirtschaftliche Praktikantenprüfung
Oktober 1994 Diplomprüfung: Diplom-Agraringenieur
Beruflicher Werdegang:
Juni - landwirtschaftliches Praktikum auf dem Martinshof in September 1989 St.-Wendel-Osterbrücken
Februar - weinbauliches Praktikum auf dem Weingut Alfons Petgen- Juli 1990 Dahm in Perl-Sehndorf
Mai - landwirtschaftliches Praktikum auf dem Betrieb von Franz September 1991 Sassen und Sohn GbR in Hoyerhagen/Weser
seit August 1994 Doktorand an der Staatlichen Lehr- und Forschungsanstalt für Landwirtschaft, Weinbau und Gartenbau in Neustadt/Weinstraße, im Fachbereich Phytomedizin
seit Mai 1998 Landwirtschaftsreferendar beim Ministerium für Wirtschaft, Verkehr, Landwirtschaft und Weinbau in Mainz

MEINER MUTTER

Teile der Arbeit sind veröffentlicht in:
PETGEN, M., SCHROPP, A. und RÖMHELD, V. (1997): Mykorrhiza - Ein Wurzelpilz und seine
Bedeutung für die Rebe.
Der Deutsche Weinbau 16-17, 30-32.
PETGEN, M., SCHROPP, A., MARSCHNER, H. und RÖMHELD, V. (1997): Die Bedeutung der
Mykorrhiza in der Rebschule.
Deutsches Weinbau-Jahrbuch 49, 79-92.
PETGEN, M., SCHROPP, A., MARSCHNER, H. und RÖMHELD, V. (1997): Untersuchungen über
das Vorkommen der arbuskulären Mykorrhiza in verschiedenen Rebschulböden der
Pfalz sowie deren praktische Anwendung in der Rebschule.
Mitt. Biol. Bundesanst. Land- und Forstwirtsch. Berlin Dahlem, Heft 332, 32-46.
PETGEN, M., SCHROPP, A. und RÖMHELD, V. (1998): Einfluß verschiedener Bodenpfle-
gemaßnahmen und Begrünungsvarianten auf die autochthone Mykorrhiza in einem
Weinberg.
Vitic. Enol. Sci. 53, 11-17.
PETGEN, M., SCHROPP, A., GEORGE, E. und RÖMHELD, V. (1998): Einfluss unterschiedlicher
Inokulationstiefen mit dem arbuskulären Mykorrhizapilz Glomus mosseae auf die
Mykorrhizierung bei Reben (Vitis sp.) in Wurzelbeobachtungskästen.
Vitis 37, 99-105.

I
INHALTSVERZEICHNIS
A ALLGEMEINER TEIL - EINLEITUNG UND METHODEN.......................... 1
1 Allgemeine Einleitung und Zielsetzung ............................................ 1
2 Material und Methoden (allgemeiner Teil) ...................................... 7
2.1 Impfmaterial ............................................................................................................ 7
2.2 Gewinnung der Wurzelproben ................................................................................ 8
2.3 Anfärben der arbuskulären Mykorrhiza (AM)-Infektion......................................... 9
2.4 Bestimmung des AM-Infektionsgrades und der Wurzellänge................................. 9
2.5 Ernte und Aufbereitung der Pflanzen .................................................................... 10
2.6 Mineralstoffanalysen ............................................................................................. 10
2.7 Statistische Auswertung ........................................................................................ 11
B SPEZIELLER TEIL................................................................................ 12
B 1 FREILANDVERSUCHE.................................................................................... 12
3 Vorkommen der autochthonen Mykorrhiza in Rebschulen und Ertragsanlagen .................................................... 12
3.1 Einleitung ............................................................................................................. 12
3.2 Material und Methoden ...................................................................................... 14
3.2.1 Standorte der Rebschulen...................................................................................... 14
3.2.2 Standorte der Ertragsanlagen ............................................................................... 15
3.2.3 Einfluß von Begrünungseinsaaten und Bodenbearbeitung auf die
autochthone Mykorrhiza........................................................................................ 18
3.2.3.1 Versuchsstandort und Versuchsvarianten.............................................................. 18

II
3.2.3.2 Witterungsbedingungen......................................................................................... 20
3.2.3.3 Wurzelprobenahme ............................................................................................... 21
3.2.3.4 Biotest mit Weißklee (Gefäßversuch im Gewächshaus) ....................................... 22
3.2.3.5 Wurzelprobenaufbereitung.................................................................................... 23
3.3 Ergebnisse ............................................................................................................ 23
3.3.1 Untersuchungen zum AM-Infektionsgrad in Rebschulen ...................................... 23
3.3.2 Untersuchungen zum AM-Infektionsgrad in Ertragsanlagen und
Adaptionsanlagen.................................................................................................. 25
3.3.3 Einfluß von Begrünungseinsaaten und Bodenbearbeitung auf die
autochthone Mykorrhiza........................................................................................ 28
3.3.3.1 AM-Infektionsgrad der Rebwurzeln...................................................................... 28
3.3.3.2 AM-Infektionsgrad der Weißkleewurzeln (Bioindikatorpflanzen) im Gefäßversuch nach 3- und 9wöchiger Versuchsphase .......................................... 30
3.4 Diskussion ............................................................................................................ 32
3.4.1 AM-Infektionsgrad von Rebwurzeln aus Rebschulen und Ertragsanlagen........... 32
3.4.2 Einfluß der Begrünung auf die AM....................................................................... 36
4 Praktische Anwendung von arbuskulären Mykorrhizapil-zen in Rebschulen und in der Topfrebenproduktion .................... 42
4.1 Einleitung ............................................................................................................. 42
4.2 Material und Methoden ...................................................................................... 44
4.2.1 Rebschulen............................................................................................................. 44
4.2.1.1 Versuchsstandorte, Pflanzenmaterial und Versuchsaufbau................................... 44
4.2.1.2 Inokulationsmethode und Wurzelprobenahme...................................................... 45
4.2.2 Topfrebenproduktion ............................................................................................. 47
4.2.2.1 Pflanzenmaterial, Versuchsgefäße und Kultursubstrat.......................................... 47

III
4.2.2.2 Inokulationsmethode ............................................................................................. 48
4.2.2.3 Pflanzenanzucht, Pflanzenpflege und Ernte der Versuchspflanzen ...................... 49
4.2.2.4 Versuchsstandort (Boden, Witterungsdaten) und Versuchsanlage........................ 49
4.3 Ergebnisse ............................................................................................................ 52
4.3.1 Rebschulen............................................................................................................. 52
4.3.1.1 Verlauf der AM-Infektion an Reb- und Weißkleewurzeln (Standort „Harthäuser“)......................................................................................... 52
4.3.1.2 Verlauf der AM-Infektion an Reb- und Maiswurzeln (Standort „Steingebiß“) ......................................................................................... 54
4.3.1.3 Einfluß der AM-Inokulation auf das Wachstum der Reben (Standort „Harthäuser“ und „Steingebiß“) ............................................................ 56
4.3.2 Topfrebenproduktion ............................................................................................. 57
4.3.2.1 Verlauf der AM-Infektion ..................................................................................... 57
4.3.2.2 Trieblängenwachstum und Trockensubstanzproduktion....................................... 58
4.3.2.3 Mineralstoffgehalte in den Blättern (Anzucht)...................................................... 62
4.3.2.4 Trieblängenwachstum der vorkultivierten Topfreben im Freiland........................ 63
4.4 Diskussion ............................................................................................................ 65
4.4.1 Einfluß der AM in Rebschulen.............................................................................. 65
4.4.2 Einfluß der AM in der Topfrebenproduktion ........................................................ 69
B 2 GEFÄßVERSUCHE ........................................................................................... 73
5 Inokulationsversuche mit AM-Pilzen an Rebenstecklingen......... 73
5.1 Einleitung ............................................................................................................. 73
5.2 Material und Methoden ...................................................................................... 75
5.2.1 Einfluß der nativen Mykorrhiza eines Rebschulbodens auf AM-
Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünstecklingen
unterschiedlichen Genotyps .................................................................................. 75
5.2.1.1 Pflanzenmaterial, Versuchsgefäße und Boden ...................................................... 75

IV
5.2.1.2 Versuchsanordnung ............................................................................................... 76
5.2.2 Einfluß unterschiedlicher Inokulationstiefen mit Glomus mosseae auf
AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünsteck-
lingen in Wurzelbeobachtungskästen .................................................................... 77
5.2.2.1 Pflanzenmaterial, Versuchsgefäße und Boden ...................................................... 77
5.2.2.2 Versuchsanordnung ............................................................................................... 78
5.2.2.3 Ernte der Versuchspflanzen................................................................................... 79
5.2.3 Einfluß einer Inokulation mit Glomus intraradices auf AM-Infektions-
grad, Nährstoffaufnahme und Wachstum von grünveredelten Stecklingen........... 80
5.2.3.1 Pflanzenmaterial, Versuchsgefäße und Boden ...................................................... 80
5.2.4 Einfluß einer AM-Inokulation bei einer erhöhten Ozonbelastung der Luft
auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Reben ............... 82
5.2.4.1 Pflanzenmaterial, Versuchsgefäße und Boden ...................................................... 82
5.2.4.2 Versuchsaufbau und Versuchsdurchführung......................................................... 82
5.2.4.3 Klimadaten ............................................................................................................ 84
5.3 Ergebnisse ............................................................................................................ 85
5.3.1 Einfluß der nativen Mykorrhiza eines Rebschulbodens auf AM-
Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünstecklingen
unterschiedlichen Genotyps .................................................................................. 85
5.3.1.1 AM-Infektionsgrad ................................................................................................ 85
5.3.1.2 Trockensubstanzproduktion und Sproß-/Wurzelverhältnis................................... 86
5.3.1.3 Mineralstoffgehalte................................................................................................ 88
5.3.2 Einfluß unterschiedlicher Inokulationstiefen mit Glomus mosseae auf
AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünsteck-
lingen in Wurzelbeobachtungskästen .................................................................... 91
5.3.2.1 Entwicklung und Verlauf des AM-Infektionsgrades im Versuchsgefäß ............... 91
5.3.2.2 Verteilung der Wurzeln im Versuchsgefäß ........................................................... 92
5.3.2.3 Trieblängenwachstum und Trockensubstanzproduktion....................................... 94
5.3.2.4 Mineralstoffgehalte................................................................................................ 96
5.3.3 Einfluß einer Inokulation mit Glomus intraradices auf AM-Infektions-
grad, Nährstoffaufnahme und Wachstum von grünveredelten Stecklingen........... 98
5.3.3.1 AM-Infektionsgrad ................................................................................................ 98
5.3.3.2 Wachstumsparameter und Sproß-/Wurzelverhältnis............................................. 99

V
5.3.3.3 Mineralstoffgehalte.............................................................................................. 101

VI
5.3.4 Einfluß einer AM-Inokulation bei einer erhöhten Ozonbelastung der Luft
auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Reben ............. 103
5.3.4.1 AM-Infektionsgrad .............................................................................................. 103
5.3.4.2 Ozon-Konzentrationen ........................................................................................ 104
5.3.4.3 Trockensubstanzproduktion ................................................................................ 106
5.3.4.4 Mineralstoffgehalte.............................................................................................. 107
5.4 Diskussion .......................................................................................................... 109
5.4.1 Nährstoffaufnahme und Wachstum der Rebenstecklinge ................................... 109
5.4.2 Wurzelwachstum und Ausbreitung der AM-Infektion........................................ 118
5.4.2 Auswirkungen erhöhter Ozonbelastung .............................................................. 122
C ZUSAMMENFASSENDER TEIL............................................................ 125
6 Abschließende Diskussion .............................................................. 125
7 Zusammenfassung........................................................................... 132
D ANHANG ............................................................................................ 136
E LITERATURVERZEICHNIS ................................................................. 146

Kapitel 1 Einleitung und Zielsetzung 1
A ALLGEMEINER TEIL - EINLEITUNG UND METHODEN
1 Allgemeine Einleitung und Zielsetzung
Der moderne Weinbau hat zum Ziel, Lesegut mit hohem Qualitätsstandard bei gleichzeitiger
Erhaltung der Leistungsfähigkeit der Rebe mit Hilfe von umweltschonenden Maßnahmen zu
erzeugen. Die Bewahrung bzw. Steigerung der natürlichen Bodenfruchtbarkeit im Ökosystem
Weinberg wird durch eine, dem Nährstoffbedarf der Rebe angepaßte Düngung erreicht. Dabei
können im Boden vorkommende Mykorrhiza-Pilze, die aufgrund ihrer symbiotischen Le-
bensweise in einer engen Wechselwirkung mit der Kulturpflanze Rebe stehen, erheblich zur
Nährstoffaufnahme beitragen. Die Besiedlung der Wurzeln höherer Pflanzen mit Pilzen aus
der zur Familie der Endogonaceen gehörenden Gattungen Glomus, Gigaspora, Sclerocystis,
Acau-
lospora, Entrophospora und Scutellospora (HALL, 1984) führt zu einer arbuskulären Mykor-
rhiza (AM). Die arbuskulären Mykorrhiza-Pilze gehen mit fast allen Pflanzenfamilien eine
symbiotische Beziehung ein, wobei die Pflanzenfamilien der Cyperaceae, Chenopodiaceae,
Brassicaceae, Caryophyllaceae, Juncaceae und Protaceae keine bzw. nur schwach ausgebil-
dete Mykorrhizen bilden. Die arbuskuläre Mykorrhiza ist gekennzeichnet durch in der Wur-
zelrinde inter- und intrazellulär wachsende Hyphen sowie dem externen Hyphengeflecht au-
ßerhalb der Wurzel (BONFANTE-FASOLO, 1984). Die Arbuskeln entstehen intrazellulär durch
Verzweigungen von Hyphen und ermöglichen den bidirektionalen Stoffaustausch zwischen
beiden Partnern. Bei einigen AM-bildenden Pilzen werden Dauerorgane ausgebildet, die als
Vesikel bezeichnet werden. Hierbei handelt es sich um bläschenförmige Hyphenanschwellun-
gen, die stark verdichtete Zellwände aufweisen und besonders lipidreich sind. Eine detaillierte
Anatomie der AM wird von BONFANTE-FASOLO (1984) beschrieben.
Bereits 1885 hat FRANK in seiner Veröffentlichung „Ueber die auf Wurzelsymbiose beruhen-
de Ernährung gewisser Bäume durch unterirdische Pilze“ bei der Untersuchung von Saugwur-
zeln an verschiedenen Baumarten von einem Pilzmantel berichtet, der die Wurzel vollständig
einhüllt. FRANK bezeichnete dieses Zusammenleben zweier verschiedener Partner als Mykor-
rhiza (griech. mycos = Pilz; rhiza = Wurzel). 1887 hat FRANK die Begriffe ectotrophische und
endotrophische Mykorrhiza geprägt. Die ectotrophische Mykorrhiza-Form besteht aus einem

Kapitel 1 Einleitung und Zielsetzung 2
dichten Mantel aus pilzigem Gewebe, das die Wurzel als Mantel umhüllt. Bei der endotrophi-
schen Form fehlt dieser charakteristische Wurzelmantel.
Die Bedeutung der Mykorrhiza als Wachstumsförderer aufgrund erhöhter Nährstoffaufnahme
an zahlreichen Pflanzenspezies ist bekannt und mehrfach beschrieben worden (ABBOTT und
ROBSON, 1984; MARSCHNER und DELL, 1994). Eine Hauptwirkung der Mykorrhiza auf die
Nährstoffaufnahme insbesondere von immobilen Nährstoffen liegt in der Oberflächenvergrö-
ßerung durch die weit über die Rhizosphäre hinausreichenden Pilzhyphen. Dadurch können
mykorrhizierte Pflanzen ein größeres Bodenvolumen erschließen, was zu einer gesteigerten
Aufnahme, unter anderem von P, Zn und Cu führt. Die Diffusionswege werden verringert und
die Aufnahmeeffizienz erhöht sich. Dies gilt insbesondere für Phosphat, welches in der Bo-
denlösung nur in geringen Konzentrationen von oftmals unter 1 mg/l vorliegt. LI et al. (1991
a) fanden in Gefäßversuchen mit Weißklee (Trifolium repens L.), daß bei den mit dem AM-
Pilz Glomus mosseae inokulierten Pflanzen die Gesamt-P-Aufnahme in zwei unterschiedli-
chen Böden zu 70 % bzw. 80 % über die Pilzhyphen erfolgte. In einer weiteren Untersuchung
stellten LI et al. (1991 b) fest, daß AM-Hyphen bis aus einer Entfernung von 11 cm von der
Wirtswurzel aus Phosphat aufnehmen können und über die Hyphen zur Wirtswurzel transpor-
tieren. In den aufgeführten Untersuchungen wurde mit sterilisierten Böden mit anschließender
Inokulation mit AM-Pilzarten gearbeitet. Über die Wirkung von nativen AM-Pilzen in Wein-
bergsböden auf die Nährstoffaufnahme und das Wachstum an Reben gibt es bisher kaum
Hinweise. Neben der Oberflächenvergrößerung besitzen AM-Hyphen noch eine weitere wich-
tige Eigenschaft, die die Nährstoffaufnahme verbessern. Die Hyphen können aufgrund ihres
geringen Durchmessers je nach Pilzart von nur 4 µm auch in kleinere Bodenporen eindringen,
die von den Wurzelhaaren nicht erreicht werden (LI et al., 1991 a). Dadurch kann die räumli-
che Verfügbarkeit von Boden-P zusätzlich erhöht werden.
In landwirtschaftlichen Kulturpflanzen wurden in Gefäßversuchen mit sterilisierten Böden
nach Beimpfung mit AM-Pilzen erhöhte Zn- und Cu-Gehalte in Sojabohne (PACOVSKY,
1986), Gartenbohne (KUCEY und JANZEN, 1987) und Hirse (RAJU et al., 1990 a) festgestellt.
Bei mehrjährigen Kulturpflanzen fanden GNEKOW und MARSCHNER (1989 b) in der Wurzel
und in den Blattspreiten bei Apfel erhöhte Zn- und Cu-Gehalte. Dabei werden ähnlich wie
beim Phosphat verbesserte Zn- und Cu-Aufnahmemechanismen über die AM-Hyphen disku-

Kapitel 1 Einleitung und Zielsetzung 3
tiert (KOTHARI et al., 1991 a; LI et al., 1991 c). Auch an mykorrhizierten Reben wurden erhöh-
te Zn- und Cu-Gehalte festgestellt (BAVARESCO und FOGHER, 1992; KARAGIANNIDIS et al.,
1995; BIRICOLTI et al., 1997).
DEHNE (1994) berichtet von einer erhöhten Resistenz der Pflanzen gegenüber bodenbürtigen
Schadorganismen. TAUBE-BAAB und BALTRUSCHAT (1993) prüften in Gefäßversuchen mit
apfelmüdem Boden fünf Mykorrhizapilze, von denen Glomus intraradices und in geringerem
Maße auch G. fasciculatum in der Lage waren, die durch Bodenmüdigkeit hervorgerufene
Wachstumsdepression an der mit Cox Orange veredelten Unterlage M9 aufzuheben. WASCH-
KIES et al. (1994) konnten in ihren Versuchen mit rebmüdem Boden durch Inokulation mit
dem AM-Pilz Glomus mosseae die typischen Merkmale der Rebenmüdigkeit wie Verkürzung
der Internodien und der Triebspitze an den Grünstecklingen der Unterlagssorte 5C aufheben.
Die Inokulation bewirkte einen signifikanten Rückgang der Anzahl fluoreszierender Pseudo-
monaden auf der Rhizoplane der Reben, was gleichzeitig zu einem verbesserten Sproß- und
Wurzelwachstum führte. In weiteren Untersuchungen wurde bei guter Nährstoffversorgung
der Wirtspflanzen bei der Besiedlung der Wurzelsysteme mit arbuskulärer Mykorrhiza gene-
rell eine verbesserte Streßresistenz der Wirtspflanzen festgestellt. So wird die Empfindlichkeit
der Pflanzen z. B. gegenüber Trockenheit (RUNJIN, 1989; RUIZ-LOZANO und AZCÓN, 1996),
Salzstreß (AZCÓN und EL-ATRASH, 1997) oder phytotoxischen Substanzen herabgesetzt. Eine
höhere Widerstandsfähigkeit mykorrhizierter Pflanzenwurzeln gegenüber Schadpilzen (COR-
DIER et al., 1996) oder Nematoden (CARLING et al., 1996; CALVET et al., 1995) kann indirekt
über ein verbessertes Wurzelwachstum zu einer erhöhten Nährstoffaufnahme beitragen und
die Schadwirkungen am Wurzelsystem kompensieren.
Seit einigen Jahren werden die symbiotischen Mykorrhizapilze insbesondere in den Tropen in
nährstoffarmen oder P-fixierenden Böden bei Maniok (HOWELER und SIEVERDING, 1983; SIE-
VERDING, 1991) oder Kautschukbäumen (IKRAM et al., 1992; FELDMANN, 1990) unter praxis-
nahen Anwendungsbedingungen eingesetzt. BALTRUSCHAT (1987 b) konnte in landwirtschaft-
lich genutzten Böden mit hohem Düngungsniveau weitere Ertragssteigerungen bei Mais durch
Freilandinokulationen herbeiführen. Es gibt bislang keine Arbeiten, die sich mit dem Einsatz
von AM-Isolaten in Rebschulen beschäftigt haben. Denkbar sind Bekämpfungsmöglichkeiten

Kapitel 1 Einleitung und Zielsetzung 4
auf rebmüden Standorten, die eine längere Nutzungsdauer von Rebschulstandorten
zulassen.
Erstmals wird 1923 von PEYRONEL über das Vorkommen der Mykorrhiza an Reben berichtet.
Anfang der 70er Jahre beschreiben POSSINGHAM und GROOT OBBINK (1971), daß die Mykor-
rhiza in allen Weinbaugebieten auf dem australischen Kontinent zu finden ist. DEAL et al.
(1972) konnten in 20 Weinbergen im Staat New York an den Rebsorten Concord, Delaware
und Catawba (Vitis lubrasca) und an den Unterlagen Baco, C3309 und Geneva 1613 sowie an
den französischen Hybriden Seibel 5279 und Seibel 100 eine Mykorrhizierung der jungen
Saugwurzeln beobachten. HAYMAN et al. (1976) fanden anhand von Wurzelproben aus ver-
schiedenen Weinbaugebieten in Südspanien eine mehr oder weniger intensive Besiedlung mit
AM-Pilzen.
In Deutschland machten GEBBING et al. (1977) erste Beobachtungen über das Vorkommen der
arbuskulären Mykorrhiza in Wurzelproben aus allen deutschen Weinbaugebieten. Die Autoren
wiesen auf die wachstumssteigernde Wirkung der Mykorrhiza an Reben hin. BONFANTE-
FASOLO (1978) führte morphologische Untersuchungen an Rebwurzeln der Pfropfkombinati-
on Barbera/5BB durch und beschrieb die verschiedenen Stadien der Arbuskelbildung in den
Wurzelzellen. EIBACH (1982) führte zahlreiche Inokulationsversuche mit Sämlingen, Steck-
lingen und Pfropfreben verschiedener Rebsorten durch. Die Autorin hielt den großflächigen
Einsatz von AM-Inokulum in Weinbergen für unwahrscheinlich und empfahl, die AM in
Weinbergsböden durch indirekte Maßnahmen, beispielsweise durch Reduktion der P-
Düngung zu fördern. In Wurzelproben aus 18 Weinbergen in Kalifornien identifizierten
MENGE et al. (1983) unterschiedliche Sporen von verschiedenen Mykorrhiza-Gattungen. Da-
bei fanden die Autoren hohe Anzahlen von Sporen der Gattungen Glomus fasciculatus, Scle-
rocystis sinuosa und Glomus macrocarpus. In Italien wurde eine Mykorrhizierung der Reb-
wurzeln aus den Weinbaugebieten von Südtirol (NAPPI et al., 1980/81), dem Piemonte (NAPPI
et al., 1985) und aus dem Südwesten beobachtet (SCHUBERT und CRAVERO, 1985). Neuere
Untersuchungen beschäftigen sich mit dem Vorkommen der Mykorrhiza in deutschen Anbau-
gebieten, z. B. des Rheingaus (BRENDEL et al., 1990) oder der Mosel (MOHR, 1993 und 1997).
MOHR (1993) sieht in der Topfrebenproduktion eine Möglichkeit, die AM-Pilze im prakti-
schen Weinbau einzusetzen, allerdings fehlen hierzu bisher gezielte Untersuchungen. In Grie-

Kapitel 1 Einleitung und Zielsetzung 5
chenland konnten KARAGIANNIDIS et al. (1997) in einem Weinberg bei Thessaloniki an den
Unterlagen 110R, 41B, 140Ru und 1103P unterschiedliche AM-Infektionsgrade der Rebwur-
zeln feststellen. Während es sich bei den aufgeführten Untersuchungen um Ertragsanlagen
handelte, liegen über das Vorkommen von AM-Pilzen in Rebschulböden, auf denen Reben-
pflanzgut erzeugt wird, keine Erkenntnisse vor.
Im Rahmen des gestiegenen Umweltbewußtseins wurde in den letzten Jahren eine Reduzie-
rung des Einsatzes von Pflanzenschutz- und Düngemitteln in Weinbergen angestrebt. Die
Weinbergsflächen wurden zunehmend mit einjährigen oder mehrjährigen Begrünungspflanzen
eingesät, von denen einige AM-Wirtspflanzen sind. Es fehlen bisher Untersuchungen, die die
Wechselwirkungen verschiedener Begrünungspflanzen mit der Rebe im Freiland in Zusam-
menhang mit der AM-Infektion aufzeigen. Bei der Erstellung einer Neuanlage erfolgt die
Auswahl der Unterlagssorte nach Gesichtspunkten wie Bodenverträglichkeit, Affinität zum
Edelreis, Reblausresistenz, Lebensdauer und Gesundheit (SCHMID et al., 1998). Dabei besitzen
Unterlagen ein unterschiedliches genotypisches Nährstoffaneignungsvermögen (RÜHL, 1992;
FARDOSSI et al., 1994). Manche Unterlagssorten haben eine hohe Nährstoffeffizienz, d. h. die-
se Sorten erbringen bei niedrigem Nährstoffstatus eine optimale Leistung und werden deshalb
als sogenannte „low-input“-Sorten bezeichnet (FARDOSSI et al., 1988). Über die im deutschen
Weinbau verwendeten Unterlagssorten gibt es hinsichtlich der Mykorrhizierung bisher kaum
Hinweise.
Die vorliegende Arbeit soll einen Beitrag zur Erweiterung des Kenntnisstandes leisten, in-
wieweit die veränderten Rahmenbedingungen im praktischen Weinbau das Vorkommen der
arbuskulären Mykorrhiza beeinflußt haben. Hierzu wurden in Rebschulen und Ertragsanlagen
Untersuchungen durchgeführt, die das quantitative Vorkommen der Mykorrhiza anhand des
Infektionsgrades (infizierte Wurzellänge in %) aufzeigen sollen (Kapitel 3). Dabei wurden
neben dem Einfluß der unterschiedlichen Unterlagssorten auf die Mykorrhizierung die unter-
schiedlichen Bewirtschaftungsweisen (konventionell, kontrolliert-umweltschonend, ökolo-
gisch) und verschiedene Begrünungspflanzen mit berücksichtigt. Ergänzend dazu wurde das
Mykorrhizierungspotential im Boden eines Versuchsweinberges mit Hilfe eines Biotests mit
Weißklee bestimmt.

Kapitel 1 Einleitung und Zielsetzung 6
Während von landwirtschaftlichen Kulturpflanzen bereits Erkenntnisse über Freilandinokula-
tionen mit AM-Pilzen vorliegen, gibt es in der Literatur keine Hinweise über die Anwendung
der AM im Freiland an Reben. Daher war ein weiteres Ziel der vorliegenden Arbeit, verschie-
dene AM-Isolate in der Rebenpflanzguterzeugung unter praxisnahen Bedingungen einzuset-
zen. Hierzu wurden neben Freilandinokulationen in Rebschulen Inokulationsversuche an
Topfreben durchgeführt, die als präinokulierte Reben nach der Anzuchtphase ins Freiland
gepflanzt wurden. Mit Hilfe geeigneter Inokulationsmethoden sollte überprüft werden, ob eine
praktische Nutzung der AM-Pilze bei der Rebenpflanzguterzeugung zu Wachstumsförderun-
gen an den Reben führt.
Begleitend zu den Freilanduntersuchungen wurden Inokulationsversuche im Gewächshaus mit
unterschiedlichen Fragestellungen durchgeführt (Kapitel 5). In einem Versuch wurde der Ein-
fluß der nativen Mykorrhiza eines Rebschulbodens auf Nährstoffaufnahme und Wachstum an
verschiedenen Grünstecklingen überprüft. Weiterhin wurde die Ausbreitung der Infektion des
arbuskulären Mykorrhiza-Pilzes Glomus mosseae in Wurzelbeobachtungskästen untersucht.
Darüber hinaus wurde der Frage nachgegangen, wie durch die Pfropfung von Grünstecklingen
die Infektion durch die Inokulation mit dem AM-Pilz Glomus intraradices an Rebwurzeln
beeinflußt wird und wie sich Wachstum und Nährstoffaufnahme bei den wurzelechten und
gepfropften Stecklingen veränderten. Im Hinblick auf die zunehmende Schadstoffbelastung
der Luft und die in der Literatur beschriebenen Wechselwirkungen von ozongeschädigten
Pflanzen und AM-Pilzen (MCCOOL et al., 1979; BREWER und HEAGLE, 1983) wurde unter-
sucht, inwieweit eine erhöhte Ozonbelastung der Luft den AM-Infektionsgrad von Rebwur-
zeln sowie das Wachstum beeinflußt.
Die einzelnen Fragestellungen werden im folgenden kapitelweise bearbeitet, wobei zwischen
Freilandversuchen (Kapitel 3 und 4) sowie Gefäßversuchen (Kapitel 5) unterschieden wird.

Kapitel 2 Material und Methoden (allgemeiner Teil) 7
2 Material und Methoden (allgemeiner Teil)
2.1 Impfmaterial
Die eingesetzten Inokula setzten sich aus Sporen, Hyphen, infizierten Wurzelstücken und
Blähton zusammen und wurden zuvor an Tagetes (Tagetes erecta L. cv. Citronenprinz) oder
Weißklee (Trifolium repens L. cv. Huja) weitervermehrt (s. Tab. 2.1). Es wurde auf verschie-
dene Bezugsquellen für AM-Inokulum zurückgegriffen. Dadurch ergaben sich die unter-
schiedlich verwendeten Inokula1 (s. Tab. 2.1), die bereits zum Teil an Reben erfolgreich ein-
gesetzt wurden (WASCHKIES et al., 1994). Bei den in Kapitel 5.2.3 und 5.2.4 beschriebenen
Gefäßversuchen wurde pro Pflanzgefäß 5 % Inokulum dem Versuchsboden beigemischt. Die
nicht mykorrhizierten Varianten (Kontrolle) in den Versuchen in Kapitel 4.2.1, 4.2.2 [1995],
5.2.3 und 5.2.4 erhielten die gleiche Menge Inokulum von nicht mykorrhizierten Tagetes-
bzw. Kleepflanzen (4.2.2 [1997] und 5.3.2). Das Inokulum für den Versuch mit den Wurzel-
beobachtungskästen enthielt AM-infizierte Wurzeln aus einer Vermehrungskultur von Glomus
mosseae an Mais und wurde drei Monate vor Versuchsbeginn nochmals an Weißklee in Bläh-
ton weitervermehrt (s. Tab. 2.1).
Die angewandten Inokulationsmethoden in der Rebschule sowie bei der Topfrebenproduktion
werden in den entsprechenden Kapiteln 4.2.1 und 4.2.2 aufgeführt.
1 Im Versuchsjahr 1997 gab es bereits mehrere Firmen, die Mykorrhizainokulum kommerziell in größeren Men-
gen (> 10.000 l) vertrieben. BACKHAUS und FELDMANN (1996) geben in ihrer Arbeit eine aktuelle Übersicht über
Anbieter von AM-Inokula.
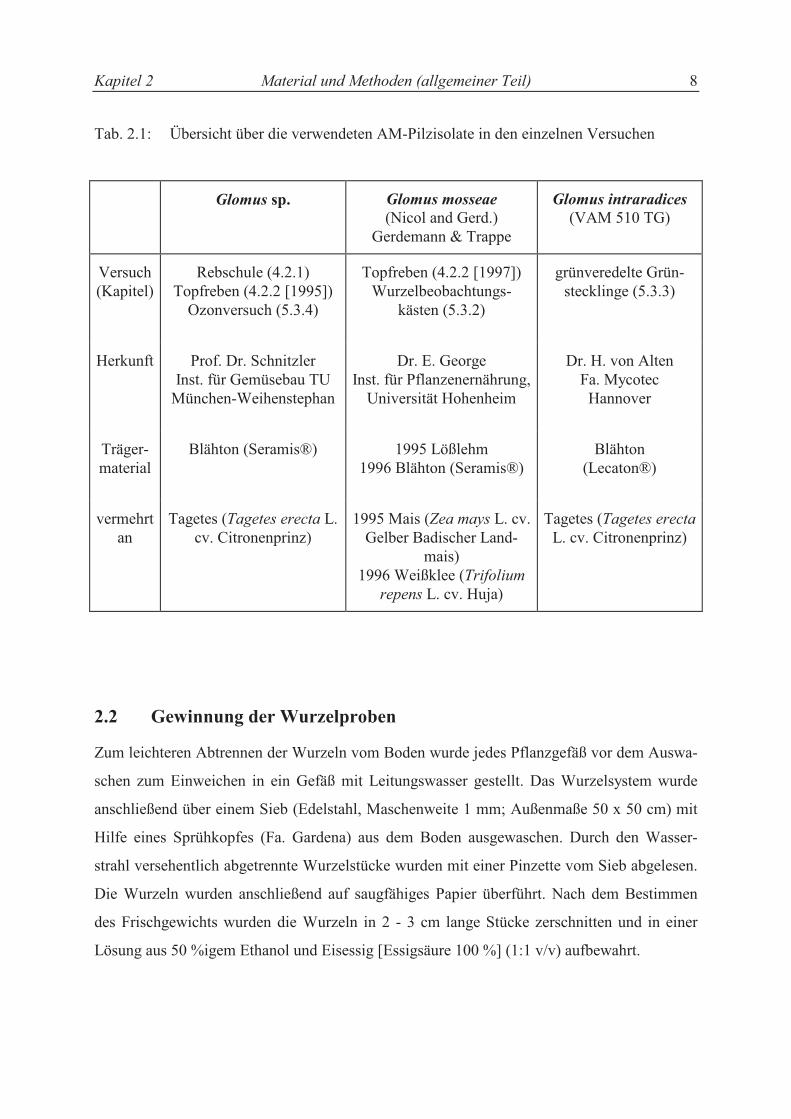
Kapitel 2 Material und Methoden (allgemeiner Teil) 8
Tab. 2.1: Übersicht über die verwendeten AM-Pilzisolate in den einzelnen Versuchen
Glomus sp. Glomus mosseae
(Nicol and Gerd.) Gerdemann & Trappe
Glomus intraradices (VAM 510 TG)
Versuch (Kapitel)
Rebschule (4.2.1) Topfreben (4.2.2 [1995])
Ozonversuch (5.3.4)
Topfreben (4.2.2 [1997]) Wurzelbeobachtungs-
kästen (5.3.2)
grünveredelte Grün-stecklinge (5.3.3)
Herkunft Prof. Dr. Schnitzler Inst. für Gemüsebau TU
München-Weihenstephan
Dr. E. George Inst. für Pflanzenernährung,
Universität Hohenheim
Dr. H. von Alten Fa. Mycotec
Hannover
Träger- material
Blähton (Seramis®) 1995 Lößlehm 1996 Blähton (Seramis®)
Blähton (Lecaton®)
vermehrtan
Tagetes (Tagetes erecta L. cv. Citronenprinz)
1995 Mais (Zea mays L. cv. Gelber Badischer Land-
mais) 1996 Weißklee (Trifolium
repens L. cv. Huja)
Tagetes (Tagetes erecta
L. cv. Citronenprinz)
2.2 Gewinnung der Wurzelproben
Zum leichteren Abtrennen der Wurzeln vom Boden wurde jedes Pflanzgefäß vor dem Auswa-
schen zum Einweichen in ein Gefäß mit Leitungswasser gestellt. Das Wurzelsystem wurde
anschließend über einem Sieb (Edelstahl, Maschenweite 1 mm; Außenmaße 50 x 50 cm) mit
Hilfe eines Sprühkopfes (Fa. Gardena) aus dem Boden ausgewaschen. Durch den Wasser-
strahl versehentlich abgetrennte Wurzelstücke wurden mit einer Pinzette vom Sieb abgelesen.
Die Wurzeln wurden anschließend auf saugfähiges Papier überführt. Nach dem Bestimmen
des Frischgewichts wurden die Wurzeln in 2 - 3 cm lange Stücke zerschnitten und in einer
Lösung aus 50 %igem Ethanol und Eisessig [Essigsäure 100 %] (1:1 v/v) aufbewahrt.

Kapitel 2 Material und Methoden (allgemeiner Teil) 9
Für die Bestimmung des AM-Infektionsgrades wurde eine Mischprobe aus der Gesamtwur-
zelprobe entnommen, wobei keine Klassifizierung nach Alter oder Dicke der Wurzeln erfolg-
te. Die Methodik zur Gewinnung und Aufbereitung der Wurzelproben im Freiland wird in den
Kapiteln 3.2.1, 4.2.1 und 4.2.2 sowie im Gefäßversuch mit Wurzelbeobachtungskästen in Ka-
pitel 5.2.2 im einzelnen beschrieben.
2.3 Anfärben der arbuskulären Mykorrhiza (AM)-Infektion
Die Wurzelproben von Reben, Mais und Weißklee wurden in Anlehnung an die Methode von
KOSKE und GEMMA (1989) in 5 %iger KOH für eine Stunde bei 90 °C im Trockenschrank
entfärbt. Bei Rebwurzeln mußte je nach Alter und Pigmentierung mit 10 %iger KOH für zwei
Stunden entfärbt werden, da die Wurzeln aus dem Freiland zum Teil erheblich lignifiziert
waren. Dabei wurde die KOH-Lösung 2 - 3 mal gewechselt. Im Anschluß daran erfolgte bei
den Rebwurzeln eine zusätzliche Bleichung in frisch angesetzter alkalischer 3 %iger H2O2-
Lösung für 10 Minuten. Nach gründlichem Spülen mit Leitungswasser wurden die Wurzel-
proben über Nacht in 1 %iger HCl angesäuert und anschließend in saurer Glycerinlösung
(0,05 %ig an Trypanblau) für 10 Minuten bei 90 °C im Trockenschrank gefärbt. Überschüssi-
ger Farbstoff wurde mit saurer Glycerinlösung aus der Wurzelrinde gespült.
2.4 Bestimmung des AM-Infektionsgrades und der Wurzellänge
Die Bestimmung des Infektionsgrades und der gesamten Wurzellänge erfolgte mit der von
GIOVANNETTI und MOSSE (1980) für die Untersuchung AM-infizierter Wurzeln angepaßten
Intersektionsmethode nach TENNANT (1975). Dazu wurden die angefärbten Wurzelstücke auf
einer PVC-Platte (Kantenlänge 10 cm), in die auf der Unterseite ein Gitter eingeritzt war, zu-
fällig verteilt und bei 50-facher Vergrößerung unter dem Binokular untersucht. Aus der Zahl
der Schnittpunkte der Linien mit den Wurzeln kann nach TENNANT (1975) die Wurzellänge
der Probe berechnet werden. Zur Bestimmung des AM-Infektionsgrades der Wurzeln wurden
horizontal und vertikal in der Regel jeweils 150 - 200 Schnittpunkte ausgezählt. Der Infekti-
onsgrad der Probe wurde aus dem Anteil (%) der infizierten Schnittpunkte an der gesamten

Kapitel 2 Material und Methoden (allgemeiner Teil) 10
Anzahl der Schnittpunkte berechnet. Pro Variante wurden die Wurzelproben in zwei Portio-
nen ausgezählt.
2.5 Ernte und Aufbereitung der Pflanzen
Am Ende der Versuchsphasen wurden die Reben am basalen Ende des Sprosses dekapitiert
und nach Blattspreiten („Blätter“) und Sproßachse mit Blattstiel getrennt geerntet. Beim Ino-
kulationsversuch mit Grünstecklingen (s. Kap. 5.2.1) wurde der Sproß in verschiedene Frakti-
onen aufgeteilt:
• junge Blätter = Blätter (Blattspreiten, ohne Blattstiele) der Triebspitze und noch nicht ausgewachsene Blätter im oberen Sproßteil (bis einschließlich zum sechsten Blatt)
• alte Blätter = Blätter (Blattspreiten, ohne Blattstiele) des darunterliegenden Sproßabschnittes
• Blattstiele
• Sproßachse
Die Bestimmung der Frisch- bzw. Trockenmasse der Reben erfolgte am gesamten oberirdi-
schen Aufwuchs (Blätter + Stiele + Sproßachse) und wird im folgenden als Sproßfrisch- bzw.
Sproßtrockenmasse bezeichnet. Nach der Bestimmung des Frischgewichts wurde das Tro-
ckengewicht des Pflanzenmaterials nach Trocknung bei 80 °C für 48 Stunden ermittelt. Je
nach Versuchsansatz wurde die Trieblänge der Reben an verschiedenen Terminen bestimmt.
Hierbei wurde der Zuwachs des Sprosses von der Veredlungsstelle aus berücksichtigt.
2.6 Mineralstoffanalysen
Nach der Bestimmung des Trockengewichts wurden die Proben in einer Achatmühle
(Fa. Fritsch) staubfein gemahlen. Jeweils 0,5 g der Pflanzenprobe wurde in Quarzglastiegeln
bei 500 °C für 12 Stunden verascht. Die Asche wurde auf einem Sandbad mit 10 ml 10 %iger
HCl eingedampft und erneut in 10 ml 10 %iger HCl aufgenommen. Nach 2minütigem Sieden

Kapitel 2 Material und Methoden (allgemeiner Teil) 11
wurde die Lösung über einen Blauband-Filter in 100 ml Meßkolben mit heißem Wasser über-
führt und mit aqua bidest. aufgefüllt. Die Lösung wurde direkt oder nach Verdünnung gemes-
sen. Die Bestimmung der Mikronährstoffe Eisen (Fe), Mangan (Mn), Kupfer (Cu) und Zink
(Zn) erfolgte am Atomabsorptionsspektrometer (Perkin-Elmer, 420). Kalium (K) und Calcium
(Ca) wurden flammenphotometrisch bestimmt, Phosphat (P) kolorimetrisch mit der
Ammonium-Molybdat-Vanadat-Methode nach GERICKE und KURMIES (1952) am Spektral-
photometer (Perkin-Elmer, 554).
2.7 Statistische Auswertung
Die statistische Verrechnung der Daten erfolgte mit Hilfe der Varianzanalyse. Die Prozentzah-
len der Infektionsgrade (y) wurden nach y => arc sin y / 100 transformiert, da die Daten
nicht normalverteilt waren. Bei Signifikanzen des F-Wertes wurden die Mittelwerte generell
mit dem Scheffé-Test verglichen, bei ausschließlich paarweisen Vergleichen (-AM/+AM)
wurde der Dunnett-Test eingesetzt. Zur Kennzeichnung von gesicherten Unterschieden zwi-
schen den Mittelwerten wurden je nach besserer Übersichtlichkeit entweder die Grenzdiffe-
renz angegeben oder Buchstaben benutzt. Signifikante Unterschiede sind jeweils (falls nicht
anders angegeben) mit 5 % Irrtumswahrscheinlichkeit (p ≤ 5 %) angegeben.

Kapitel 2 Material und Methoden (allgemeiner Teil) 12
2 Material und Methoden (allgemeiner Teil)
2.1 Impfmaterial
Die eingesetzten Inokula setzten sich aus Sporen, Hyphen, infizierten Wurzelstücken und
Blähton zusammen und wurden zuvor an Tagetes (Tagetes erecta L. cv. Citronenprinz) oder
Weißklee (Trifolium repens L. cv. Huja) weitervermehrt (s. Tab. 2.1). Es wurde auf verschie-
dene Bezugsquellen für AM-Inokulum zurückgegriffen. Dadurch ergaben sich die unter-
schiedlich verwendeten Inokula2 (s. Tab. 2.1), die bereits zum Teil an Reben erfolgreich ein-
gesetzt wurden (WASCHKIES et al., 1994). Bei den in Kapitel 5.2.3 und 5.2.4 beschriebenen
Gefäßversuchen wurde pro Pflanzgefäß 5 % Inokulum dem Versuchsboden beigemischt. Die
nicht mykorrhizierten Varianten (Kontrolle) in den Versuchen in Kapitel 4.2.1, 4.2.2 [1995],
5.2.3 und 5.2.4 erhielten die gleiche Menge Inokulum von nicht mykorrhizierten Tagetes-
bzw. Kleepflanzen (4.2.2 [1997] und 5.3.2). Das Inokulum für den Versuch mit den Wurzel-
beobachtungskästen enthielt AM-infizierte Wurzeln aus einer Vermehrungskultur von Glomus
mosseae an Mais und wurde drei Monate vor Versuchsbeginn nochmals an Weißklee in Bläh-
ton weitervermehrt (s. Tab. 2.1).
Die angewandten Inokulationsmethoden in der Rebschule sowie bei der Topfrebenproduktion
werden in den entsprechenden Kapiteln 4.2.1 und 4.2.2 aufgeführt.
2 Im Versuchsjahr 1997 gab es bereits mehrere Firmen, die Mykorrhizainokulum kommerziell in größeren Men-
gen (> 10.000 l) vertrieben. BACKHAUS und FELDMANN (1996) geben in ihrer Arbeit eine aktuelle Übersicht über
Anbieter von AM-Inokula.
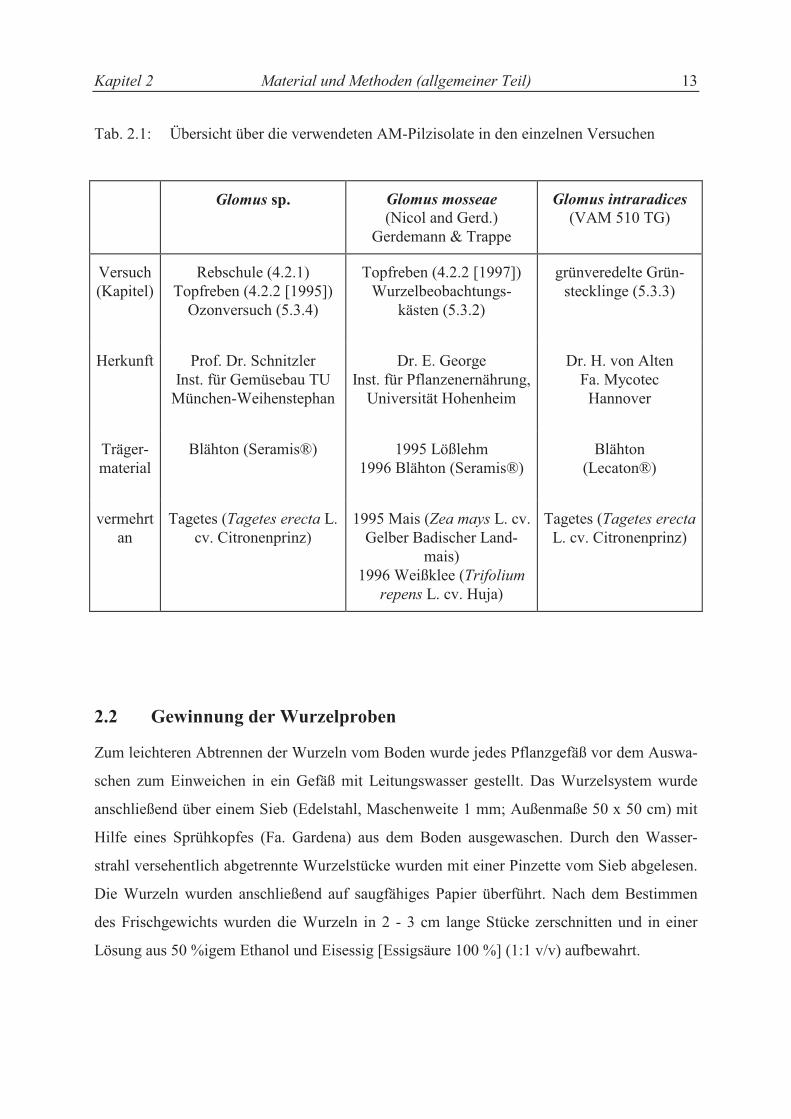
Kapitel 2 Material und Methoden (allgemeiner Teil) 13
Tab. 2.1: Übersicht über die verwendeten AM-Pilzisolate in den einzelnen Versuchen
Glomus sp. Glomus mosseae
(Nicol and Gerd.) Gerdemann & Trappe
Glomus intraradices (VAM 510 TG)
Versuch (Kapitel)
Rebschule (4.2.1) Topfreben (4.2.2 [1995])
Ozonversuch (5.3.4)
Topfreben (4.2.2 [1997]) Wurzelbeobachtungs-
kästen (5.3.2)
grünveredelte Grün-stecklinge (5.3.3)
Herkunft Prof. Dr. Schnitzler Inst. für Gemüsebau TU
München-Weihenstephan
Dr. E. George Inst. für Pflanzenernährung,
Universität Hohenheim
Dr. H. von Alten Fa. Mycotec
Hannover
Träger- material
Blähton (Seramis®) 1995 Lößlehm 1996 Blähton (Seramis®)
Blähton (Lecaton®)
vermehrtan
Tagetes (Tagetes erecta L. cv. Citronenprinz)
1995 Mais (Zea mays L. cv. Gelber Badischer Land-
mais) 1996 Weißklee (Trifolium
repens L. cv. Huja)
Tagetes (Tagetes erecta
L. cv. Citronenprinz)
2.2 Gewinnung der Wurzelproben
Zum leichteren Abtrennen der Wurzeln vom Boden wurde jedes Pflanzgefäß vor dem Auswa-
schen zum Einweichen in ein Gefäß mit Leitungswasser gestellt. Das Wurzelsystem wurde
anschließend über einem Sieb (Edelstahl, Maschenweite 1 mm; Außenmaße 50 x 50 cm) mit
Hilfe eines Sprühkopfes (Fa. Gardena) aus dem Boden ausgewaschen. Durch den Wasser-
strahl versehentlich abgetrennte Wurzelstücke wurden mit einer Pinzette vom Sieb abgelesen.
Die Wurzeln wurden anschließend auf saugfähiges Papier überführt. Nach dem Bestimmen
des Frischgewichts wurden die Wurzeln in 2 - 3 cm lange Stücke zerschnitten und in einer
Lösung aus 50 %igem Ethanol und Eisessig [Essigsäure 100 %] (1:1 v/v) aufbewahrt.

Kapitel 2 Material und Methoden (allgemeiner Teil) 14
Für die Bestimmung des AM-Infektionsgrades wurde eine Mischprobe aus der Gesamtwur-
zelprobe entnommen, wobei keine Klassifizierung nach Alter oder Dicke der Wurzeln erfolg-
te. Die Methodik zur Gewinnung und Aufbereitung der Wurzelproben im Freiland wird in den
Kapiteln 3.2.1, 4.2.1 und 4.2.2 sowie im Gefäßversuch mit Wurzelbeobachtungskästen in Ka-
pitel 5.2.2 im einzelnen beschrieben.
2.3 Anfärben der arbuskulären Mykorrhiza (AM)-Infektion
Die Wurzelproben von Reben, Mais und Weißklee wurden in Anlehnung an die Methode von
KOSKE und GEMMA (1989) in 5 %iger KOH für eine Stunde bei 90 °C im Trockenschrank
entfärbt. Bei Rebwurzeln mußte je nach Alter und Pigmentierung mit 10 %iger KOH für zwei
Stunden entfärbt werden, da die Wurzeln aus dem Freiland zum Teil erheblich lignifiziert
waren. Dabei wurde die KOH-Lösung 2 - 3 mal gewechselt. Im Anschluß daran erfolgte bei
den Rebwurzeln eine zusätzliche Bleichung in frisch angesetzter alkalischer 3 %iger H2O2-
Lösung für 10 Minuten. Nach gründlichem Spülen mit Leitungswasser wurden die Wurzel-
proben über Nacht in 1 %iger HCl angesäuert und anschließend in saurer Glycerinlösung
(0,05 %ig an Trypanblau) für 10 Minuten bei 90 °C im Trockenschrank gefärbt. Überschüssi-
ger Farbstoff wurde mit saurer Glycerinlösung aus der Wurzelrinde gespült.
2.4 Bestimmung des AM-Infektionsgrades und der Wurzellänge
Die Bestimmung des Infektionsgrades und der gesamten Wurzellänge erfolgte mit der von
GIOVANNETTI und MOSSE (1980) für die Untersuchung AM-infizierter Wurzeln angepaßten
Intersektionsmethode nach TENNANT (1975). Dazu wurden die angefärbten Wurzelstücke auf
einer PVC-Platte (Kantenlänge 10 cm), in die auf der Unterseite ein Gitter eingeritzt war, zu-
fällig verteilt und bei 50-facher Vergrößerung unter dem Binokular untersucht. Aus der Zahl
der Schnittpunkte der Linien mit den Wurzeln kann nach TENNANT (1975) die Wurzellänge
der Probe berechnet werden. Zur Bestimmung des AM-Infektionsgrades der Wurzeln wurden
horizontal und vertikal in der Regel jeweils 150 - 200 Schnittpunkte ausgezählt. Der Infekti-
onsgrad der Probe wurde aus dem Anteil (%) der infizierten Schnittpunkte an der gesamten

Kapitel 2 Material und Methoden (allgemeiner Teil) 15
Anzahl der Schnittpunkte berechnet. Pro Variante wurden die Wurzelproben in zwei Portio-
nen ausgezählt.
2.5 Ernte und Aufbereitung der Pflanzen
Am Ende der Versuchsphasen wurden die Reben am basalen Ende des Sprosses dekapitiert
und nach Blattspreiten („Blätter“) und Sproßachse mit Blattstiel getrennt geerntet. Beim Ino-
kulationsversuch mit Grünstecklingen (s. Kap. 5.2.1) wurde der Sproß in verschiedene Frakti-
onen aufgeteilt:
• junge Blätter = Blätter (Blattspreiten, ohne Blattstiele) der Triebspitze und noch nicht ausgewachsene Blätter im oberen Sproßteil (bis einschließlich zum sechsten Blatt)
• alte Blätter = Blätter (Blattspreiten, ohne Blattstiele) des darunterliegenden Sproßabschnittes
• Blattstiele
• Sproßachse
Die Bestimmung der Frisch- bzw. Trockenmasse der Reben erfolgte am gesamten oberirdi-
schen Aufwuchs (Blätter + Stiele + Sproßachse) und wird im folgenden als Sproßfrisch- bzw.
Sproßtrockenmasse bezeichnet. Nach der Bestimmung des Frischgewichts wurde das Tro-
ckengewicht des Pflanzenmaterials nach Trocknung bei 80 °C für 48 Stunden ermittelt. Je
nach Versuchsansatz wurde die Trieblänge der Reben an verschiedenen Terminen bestimmt.
Hierbei wurde der Zuwachs des Sprosses von der Veredlungsstelle aus berücksichtigt.
2.6 Mineralstoffanalysen
Nach der Bestimmung des Trockengewichts wurden die Proben in einer Achatmühle
(Fa. Fritsch) staubfein gemahlen. Jeweils 0,5 g der Pflanzenprobe wurde in Quarzglastiegeln
bei 500 °C für 12 Stunden verascht. Die Asche wurde auf einem Sandbad mit 10 ml 10 %iger
HCl eingedampft und erneut in 10 ml 10 %iger HCl aufgenommen. Nach 2minütigem Sieden

Kapitel 2 Material und Methoden (allgemeiner Teil) 16
wurde die Lösung über einen Blauband-Filter in 100 ml Meßkolben mit heißem Wasser über-
führt und mit aqua bidest. aufgefüllt. Die Lösung wurde direkt oder nach Verdünnung gemes-
sen. Die Bestimmung der Mikronährstoffe Eisen (Fe), Mangan (Mn), Kupfer (Cu) und Zink
(Zn) erfolgte am Atomabsorptionsspektrometer (Perkin-Elmer, 420). Kalium (K) und Calcium
(Ca) wurden flammenphotometrisch bestimmt, Phosphat (P) kolorimetrisch mit der
Ammonium-Molybdat-Vanadat-Methode nach GERICKE und KURMIES (1952) am Spektral-
photometer (Perkin-Elmer, 554).
2.7 Statistische Auswertung
Die statistische Verrechnung der Daten erfolgte mit Hilfe der Varianzanalyse. Die Prozentzah-
len der Infektionsgrade (y) wurden nach y => arc sin y / 100 transformiert, da die Daten
nicht normalverteilt waren. Bei Signifikanzen des F-Wertes wurden die Mittelwerte generell
mit dem Scheffé-Test verglichen, bei ausschließlich paarweisen Vergleichen (-AM/+AM)
wurde der Dunnett-Test eingesetzt. Zur Kennzeichnung von gesicherten Unterschieden zwi-
schen den Mittelwerten wurden je nach besserer Übersichtlichkeit entweder die Grenzdiffe-
renz angegeben oder Buchstaben benutzt. Signifikante Unterschiede sind jeweils (falls nicht
anders angegeben) mit 5 % Irrtumswahrscheinlichkeit (p ≤ 5 %) angegeben.

Kapitel 3 Vorkommen der AM in Weinbergsböden 17
B SPEZIELLER TEIL
B 1 FREILANDVERSUCHE
3 Vorkommen der autochthonen Mykorrhiza in Rebschulen und Ertragsanlagen
3.1 Einleitung
An Reben wurde die arbuskuläre Mykorrhiza (AM) bereits in zahlreichen Weinbergsböden
weltweit nachgewiesen (DEAL et al., 1972; EIBACH, 1982; NAPPI et al., 1985; SCHUBERT und
CRAVERO, 1985; BRENDEL et al., 1990). Diese Untersuchungen wurden in Ertragsanlagen
durchgeführt, die bereits mehrere Jahre bewirtschaftet wurden. Ein Ziel der vorliegenden Un-
tersuchung war es festzustellen, inwieweit die arbuskuläre Mykorrhiza im Vergleich zu Er-
tragsanlagen auch in Rebschulen vorkommt, in denen die veredelten Pfropfreben für die Dau-
er einer Vegetationsperiode kultiviert und erst danach an den endgültigen Standort im Wein-
berg verpflanzt werden. Aus der Literatur sind bisher zu diesem Themenkomplex keine Un-
tersuchungen bekannt.
Durch die Entscheidung über die richtige Unterlagenwahl wird die Grundlage für den langfris-
tigen Erfolg einer Rebanlage gelegt. Die Wahl der jeweiligen Unterlage erfolgt nach verschie-
denen Gesichtspunkten, wie z. B. Wüchsigkeit, Ertragshöhe und Kalkverträglichkeit (CURRLE
et al., 1983). Im zweiten Teil der vorliegenden Untersuchung sollten Wurzelproben aus ver-
schiedenen Weinbergsstandorten aus dem Anbaugebiet „Pfalz“ Aufschluß über das Vorkom-
men der Mykorrhiza an Reben in dieser Region geben. Um verschiedene Unterlagssorten auf
ihre Mykorrhizierung hin vergleichen zu können, wurde ein Teil der Proben aus Adaptionsan-
lagen entnommen, in denen verschiedene Unterlagssorten mit einer Edelreissorte auf dem
gleichen Standort stehen. Neben dem Einfluß der Unterlage auf die Mykorrhizierung an Re-
ben (KARAGIANNIDIS et al., 1997) sind mehrere Untersuchungen von landwirtschaftlichen
Ackerbaukulturen bekannt, die von einem unterschiedlichen Einfluß der Bewirtschaftungs-
weisen auf die Mykorrhizierung berichten. FELDMANN (1997) beschrieb eine potentielle Re-

Kapitel 3 Vorkommen der AM in Weinbergsböden 18
duktion der AM-Pilzartenzahl in konventionell betriebenen Monokulturen, die der Autor auf
die eingesetzten Pflanzenbaumaßnahmen wie Düngung und Pestizideinsatz zurückführt. Da-
her wurden Wurzelproben aus konventionell, kontrolliert umweltschonend und ökologisch
bewirtschafteten Weinbergen auf ihre Mykorrhizierung untersucht. Ergänzend zur Bestim-
mung des AM-Infektionsgrades an den Rebwurzeln wurden die Nährstoffgehalte im Boden
sowie in der Trockensubstanz der Blätter quantitativ bestimmt, um mögliche Zusammenhänge
zwischen Mykorrhizierung und Nährstoffversorgung der Reben erfassen zu können.
Im Weinanbaugebiet „Pfalz“ werden circa 6 % der Gesamtrebfläche (Stand 1997) nach den
Richtlinien für den kontrolliert umweltschonenden Weinbau (KUW) bewirtschaftet, wobei in
den Richtlinien eine Begrünung der Weinbergsböden grundsätzlich vorgeschrieben ist (ANO-
NYM, 1997 a). Auch im ökologischen Weinbau ist eine Begrünungseinsaat im Weinberg eine
wesentliche Voraussetzung für das Funktionieren der Stoffkreisläufe im Ökosystem Weinberg
und wird in den Bioland-Richtlinien (ANONYM, 1997 b) vorgeschrieben, so daß eine Begrü-
nungseinsaat im Weinberg einen immer größeren Stellenwert im praktischen Weinbau erlangt
hat.
Durch die Begrünung kann die Bodenerosion vermindert und den damit verbundenen Nähr-
stoffverlusten und der Eutrophierung von Oberflächengewässern entgegengewirkt werden. Bei
der Dauerkultur Rebe kann die Begrünung als Ersatz einer Fruchtfolge angesehen werden, die
das Wachstum der Reben positiv beeinflußt (HOFMANN, 1994). Die ganzjährige Bodenbede-
ckung verbessert die Bodenstruktur und die Wasserhaltekraft durch eine intensive Durchwur-
zelung des Bodens und durch die Lebendverbauung der Begrünungswurzeln. Ebenso kann die
Nitratauswaschung ins Grundwasser vermieden werden, indem in Zeiten niedrigen N-Bedarfs
der Rebe überschüssiger Stickstoff im Boden festgelegt wird (PERRET, 1993). Je nach Wuchs-
stärke der Begrünungseinsaat und dem Wasserhaushalt im Weinberg wird die Begrünung ab-
gemäht oder umgebrochen. Durch die Einsaaten von unterschiedlichen Begrünungspflanzen
wird die Fauna im Ökosystem Weinberg gefördert (REMUND et al., 1989). Diese Begrünungs-
pflanzen werden einzeln oder als Gemenge einjährig, zweijährig oder bei Dauerbegrünung
mehrjährig verwendet. Eine Vielzahl dieser Pflanzen wird mit arbuskulären Mykorrhizapilzen
infiziert, die in fast allen Böden vorzufinden sind.

Kapitel 3 Vorkommen der AM in Weinbergsböden 19
Verschiedene Begrünungspflanzen sind in der Lage, die Mykorrhiza an der Rebe zu fördern.
MOHR (1994) stellte in einem Gefäßversuch mit wurzelechten Rieslingstecklingen
(Vitis vinifera L.) fest, daß Rotes Straußgras (Agrostis tenuis L.) und Weißklee (Trifolium
repens L.) in einem sterilisierten Boden nach der Inokulation mit einem Mykorrhizapilz die
Mykorrhizierung an den Rieslingreben förderte. NAPPI et al. (1980/81) berichteten von einem
positiven Einfluß der Mykorrhiza bei verschiedenen Rebsorten mit der Unterlage 5BB in Süd-
Tirol und fanden unter einer Grasmulchdecke ein signifikant höheres Mykorrhizasporenvor-
kommen im Boden als in den offen gehaltenen Varianten ohne Bewuchs. Daher war ein weite-
res Ziel der vorliegenden Untersuchung, den Einfluß verschiedener Begrünungspflanzen im
Weinberg auf die autochthone Mykorrhiza an Rebwurzeln zu untersuchen. In einer seit vier
Jahren bestehenden Versuchsanlage wurden aus sieben Begrünungsvarianten und einer offen
gehaltenen Variante Wurzelproben für die Mykorrhiza-Untersuchungen entnommen. Ergän-
zend wurde im Gewächshaus das Infektionspotential der Mykorrhiza im Boden aus den jewei-
ligen Varianten anhand eines Biotests mit Weißklee bestimmt.
3.2 Material und Methoden
3.2.1 Standorte der Rebschulen
Auf zehn verschiedenen Rebschulflächen aus dem Anbaugebiet „Pfalz“ wurden in der Zeit
vom 15.07. - 19.07.1996 Wurzelproben von den Pfropfkombinationen Riesling (Vitis vinifera
L.) auf SO4 (V. berlandieri x V. riparia) und Müller-Thurgau (V. vinifera L.) auf 5BB (V.
berlandieri x V. riparia) entnommen. Die Wurzeln wurden mit einem Spaten im Bereich von
5 - 7 Reben an einer Seite der Pflanzfolie aus einer Bodentiefe von 30 cm entnommen und in
angefeuchtete Polyethylenbeutel überführt. Das Anfärben der Rebwurzeln sowie die Bestim-
mung des AM-Infektionsgrades (s. Kap. 2.3 und 2.4) erfolgten im Labor. Pro Pfropfkombina-
tion und Standort wurden jeweils vier Proben entnommen, die zufällig verteilt auf dem Stand-
ort ausgewählt wurden. Bei der Wurzelprobenahme wurden gleichzeitig Bodenproben aus
dem Oberboden (0 - 30 cm) gezogen, die später im Labor auf Phosphat untersucht wurden.

Kapitel 3 Vorkommen der AM in Weinbergsböden 20
3.2.2 Standorte der Ertragsanlagen
Für die vorliegende Untersuchung wurden Wurzelproben aus verschiedenen Adaptionsanla-
gen2 bzw. Ertragsanlagen aus dem Anbaugebiet „Pfalz“ entnommen und der AM-
Infektionsgrad bestimmt. Ein Teil der Rebflächen wurde nach den Richtlinien der EU-
Verordnung 2092/91 (ANONYM, 1997 b) ökologisch (BIOLAND-Mitgliedsbetriebe) bzw.
nach den Richtlinien für den kontrolliert umweltschonenden Weinbau in Rheinland-Pfalz
(KUW) bewirtschaftet (ANONYM, 1997 a). Bei den ökologisch bewirtschafteten Weinbergs-
flächen dürfen keine chemisch synthetischen Fungizide, Insektizide, Herbizide, Stickstoffdün-
gemittel sowie leicht lösliche Phosphate verwendet werden (ANONYM, 1997 b). Die Richtlinie
für den KUW läßt ebenfalls keine Herbizidanwendung zu, Fungizide dürfen je nach Zulas-
sung angewendet werden (ANONYM, 1997 a). Bei zwei Rebflächen handelte es sich um kon-
ventionell geführte Rebanlagen ohne Bindung an Richtlinien. In Tabelle 3.1 sind die Standorte
der einzelnen Rebanlagen mit Angaben über Sorte (Pfropfkombination), Anbaurichtung (öko-
logisch, kontrolliert umweltschonend, konventionell), Begrünungsmanagement sowie boden-
chemische Kenndaten auf-
gelistet. Falls keine näheren Angaben zur Dauerbegrünung angegeben sind, handelt es sich um
eine naturbelassene Dauerbegrünung oder eine Einsaat mit einer Gramineenmischung.
2Adaptionsanlagen sind Ertragsanlagen, bei denen die Möglichkeit besteht, dieselbe Edelreissorte am gleichen
Standort mit verschiedenen Unterlagssorten auf ihre Eignung zu prüfen. Jede Unterlage ist in Adaptionsanlagen
in zwei Feldwiederholungen angelegt. Bei der Wurzelprobenahme wurden die Werte aus den zwei Feldwiederho-
lungen gemittelt.

Kapitel 3 Vorkommen der AM in Weinbergsböden 21

Kapitel 3 Vorkommen der AM in Weinbergsböden 22
Die Wurzelprobenahmen erfolgten in der Zeit vom 23.07. - 05.08.1996. Gleichzeitig wurden
Bodenproben in vierfacher Wiederholung pro Standort gezogen. In den Adaptionsanlagen
wurden pro Unterlage vier Rebstöcke beprobt, in den herkömmlichen Ertragsanlagen pro
Standort ebenfalls vier Rebstöcke zufällig über die Parzelle verteilt. Zur Wurzelprobenahme
wurde zwischen zwei Rebstöcke innerhalb der Rebzeile ein Holzrahmen (Innenmaße
55 x 22 cm) gelegt. Aus diesem Bereich wurde mit einem Spaten der Boden 22 cm tief ent-
nommen. Damit war gewährleistet, daß die zu untersuchenden Wurzeln immer aus einem
gleichen Bodenvolumen (circa 26.620 cm3) stammten. Aus dem definierten Bodenbereich
wurden für die Bestimmung des AM-Infektionsgrades nur Wurzeln mit weniger als 2 mm
Durchmesser berücksichtigt. Die Wurzeln wurden in Polyethylendosen, gefüllt mit Leitungs-
wasser, bis zur weiteren Verarbeitung aufbewahrt. Nach dem Trocknen auf saugfähigem Pa-
pier wurde im Labor das Frischgewicht der Rebwurzeln bestimmt. Aus der Gesamtmenge der
Wurzeln wurde eine repräsentative Wurzelprobe entnommen, an der gleichzeitig der AM-
Infektionsgrad sowie die Wurzellänge (s. Kap. 2.4) bestimmt wurden. Die Gesamtwurzellänge
konnte somit anteilsmäßig errechnet werden. Von jedem Standort wurden in der Zeit vom
23.07. - 05.08.1996 Blattproben entnommen und die Nährstoffgehalte in der Trockensubstanz
der Blätter nach der in Kapitel 2.6 beschriebenen Methode quantitativ bestimmt. Von jeder
Pfropfkombination wurde an zehn Rebstöcken jeweils ein ausgewachsenes Blatt aus der Trau-
benzone gesammelt.
Ein direkter Vergleich zwischen unterschiedlichen Bewirtschaftungsweisen auf verschiedenen
Standorten war aufgrund zahlreicher Einflußfaktoren wie unterschiedlicher Bodenverhältnisse
und mikroklimatischer Differenzen zwischen den Standorten nicht möglich. Daher erfolgte
eine statistische Verrechnung der Ergebnisse nur innerhalb der gleichen Adaptionsanlage.

Kapitel 3 Vorkommen der AM in Weinbergsböden 23
3.2.3 Einfluß von Begrünungseinsaaten und Bodenbearbeitung auf die autochthone Mykorrhiza
3.2.3.1 Versuchsstandort und Versuchsvarianten
Die Untersuchungen wurden in einem Weinberg des Staatsweingutes der Staatlichen Lehr-
und Forschungsanstalt in Neustadt/Weinstraße in der Lage „Mußbacher Hundertmorgen“
durchgeführt. Der Weinberg hatte eine Größe von 66,6 Ar, war eben und als Drahtrahmenan-
lage erzogen. Die 22 Rebzeilen verliefen in Nord-Süd-Richtung. Die Zeilenbreite betrug 2,00
m und der Stockabstand 1,20 m. Die Anlage wurde 1987 mit Silvaner (Vitis vinifera L.) [Klon
N60 und N62] auf 5BB (V. berlandieri x V. riparia) [Klon N101] bepflanzt.
In der Versuchsfläche wurde vom Fachbereich Weinbau der SLFA Neustadt seit 1991 ein
Versuch über den Einfluß verschiedener Bodenpflegeverfahren auf den Nitrataustrag durchge-
führt. Hierzu wurden sieben verschiedene Begrünungsvarianten und eine offen gehaltene Va-
riante in vier Wiederholungen angelegt und über vier Jahre entsprechend betreut (vgl. Tab.
3.2). Die Wiederholungsparzellen erstreckten sich jeweils über eine Länge von 50 Metern.
Um unnötige Leerfahrten für die Bodenpflegearbeiten zu vermeiden, wurden die einzelnen
Gassen mit gleichartigen Varianten belegt (z. B. in einer Gasse hintereinander zwei verschie-
dene Gründüngungsvarianten (Cruciferen/Gramineen)). Die Einsaaten erfolgten nach einer
Saatbettbereitung mit Fräse oder Scheibenegge als Breitsaat. Der Umbruch der einzelnen Be-
grünungsvarianten wurde mit einer Fräse durchgeführt. Die mechanische Offenhaltung der
Varianten, die im Sommer nicht begrünt waren, erfolgte mit Grubber oder Scheibenegge. Die
Begrünung ist während des Jahres mehrfach mit Kreiselmulchgeräten abgemäht worden, wo-
bei das Mulchgut im Weinberg verblieb. Der Unterzeilenbereich wurde mit Flachscharräu-
mern bearbeitet und offen gehalten. Die Pflanzenschutz- und Düngungsmaßnahmen erfolgten
in praxisüblicher Weise bei allen Varianten einheitlich. Die Versuchsanlage wurde in den Jah-
ren 1991 bis 1994 gleichmäßig mit 30 kg N/ha und Jahr (Kalkammonsalpeter) gedüngt, wobei
1995 nach dem Umbruch der gesamten Begrünung keine N-Düngung erfolgte.
Die ersten Einsaaten der Varianten 2 - 4 erfolgten im August 1991. Der Aufwuchs aus diesen
Varianten sowie die natürliche Winterbegrünung, die sich in der Kontrolle im offen gehalte-
nen Boden entwickelte, wurde im Mai 1992 wieder umgebrochen. Diese Vorgehensweise

Kapitel 3 Vorkommen der AM in Weinbergsböden 24
wurde weiter fortgeführt. Im Mai 1991 wurden die Varianten 5 - 8 eingesät. Die Zweijahres-
begrünung der Varianten 5 und 6 wurde im Mai 1993 umgebrochen und im darauffolgenden
August wieder eingesät. Im Versuchsjahr 1995 wurden aus betriebstechnischen Gründen alle
Varianten einheitlich umgebrochen und über Sommer offen gehalten.
Tab. 3.2: Übersicht über die Versuchsvarianten (Versuchsanlage „Mußbacher Hundertmorgen“)
Variante Bodenpflegesystem Pflanzeneinsaaten Saatjahr
1 Offenhaltung(Kontrolle)
keine Einsaaten im Winter: Wildflora
-
2 Wintergründüngung(Cruciferen) im Sommer offen
Ölrettich1 / Winterraps2
Gelbsenf 3 / Winterrübsen4
Ölrettich1 / Winterraps2
Gelbsenf 3 / Winterrübsen4
1991 1992 1993 1994
3 Wintergründüngung(Gramineen) im Sommer offen
Einjähriges Weidelgras5
Winterroggen6
Winterroggen6
Einjähriges Weidelgras5
1991 1992 1993 1994
4 Wintergründüngung(Leguminosen) im Sommer offen
Winterwicken7
Wintererbsen8
Winterwicken7
Futter9-, Platt10-, Wintererbsen8
Winterwicken7
1991 1992 1993 1994
5 Zweijahresbegrünung (Gramineen)
Deutsches Weidelgras11
Welsches Weidelgras121991/93 1993/95
6 Zweijahresbegrünung(Legumino-sen/Gramineen)
Deutsches Weidelgras11 / Erdklee13 Rotklee14
Winterroggen6 / Winterwicken7
1991/93
1993/95
7 Dauerbegrünung(Gramineen)
Deutsches Weidelgras11 Horstrotschwingel15 / Wiesenris-pe16
1991
8 Dauerbegrünung(Leguminosen)
Weiß17-, Rot14-, Hornschotenklee18 1991
1Raphanus sativus L., 2
Brassica napus L., 3Sinapsis arvensis L., 4
Brassica rapa L., 5Lolium
multiflorum Lam., 6Secale cereale L., 7Vicia villosa L., 8Pisum sativum L., 9Pisum arvense L.,

Kapitel 3 Vorkommen der AM in Weinbergsböden 25
10Pisum sativum L., 11
Lolium perenne L., 12Lolium multiflorum L., 13
Trifolium subterraneum
L., 14Trifolium pratense L., 15
Festuca rubra commutata L., 16Poa pratensis L., 17
Trifolium
repens L., 18Lotus corniculatus L.
Für die Bestimmung von Humusgehalt, pH-Wert sowie der Bodenart wurden stichprobenartig,
über die Versuchsfläche verteilt, Bodenproben gezogen. In der Versuchsanlage Hundertmor-
gen handelte es sich um einen sandig-schluffigen Lehm. Nähere Eigenschaften sind in der
Tabelle 3.3 aufgeführt. Aus jeder Versuchsvariante wurden aus den Tiefenstufen 0 - 25, 25 -
50 und 50 - 75 cm Bodenproben entnommen und die Nährstoffe Phosphat und Kalium analy-
siert (s. Tab. A1 im Anhang).
Tab. 3.3: Chemische und physikalische Eigenschaften des Bodens in der Versuchsanlage „Mußbacher Hundertmorgen“
Bodentiefe Humus pH Korngrößenzusammensetzung Bodenart
(KCl) Ton Schluff Sand
(cm) (%) (%)
0 - 30 2,2 7,3 17,5 51,4 31,1 sandig-schluffiger Lehm
30 - 60 1,4 7,5 16,8 51,4 31,8 "
60 - 90 0,8 7,7 13,5 68,7 17,8 "
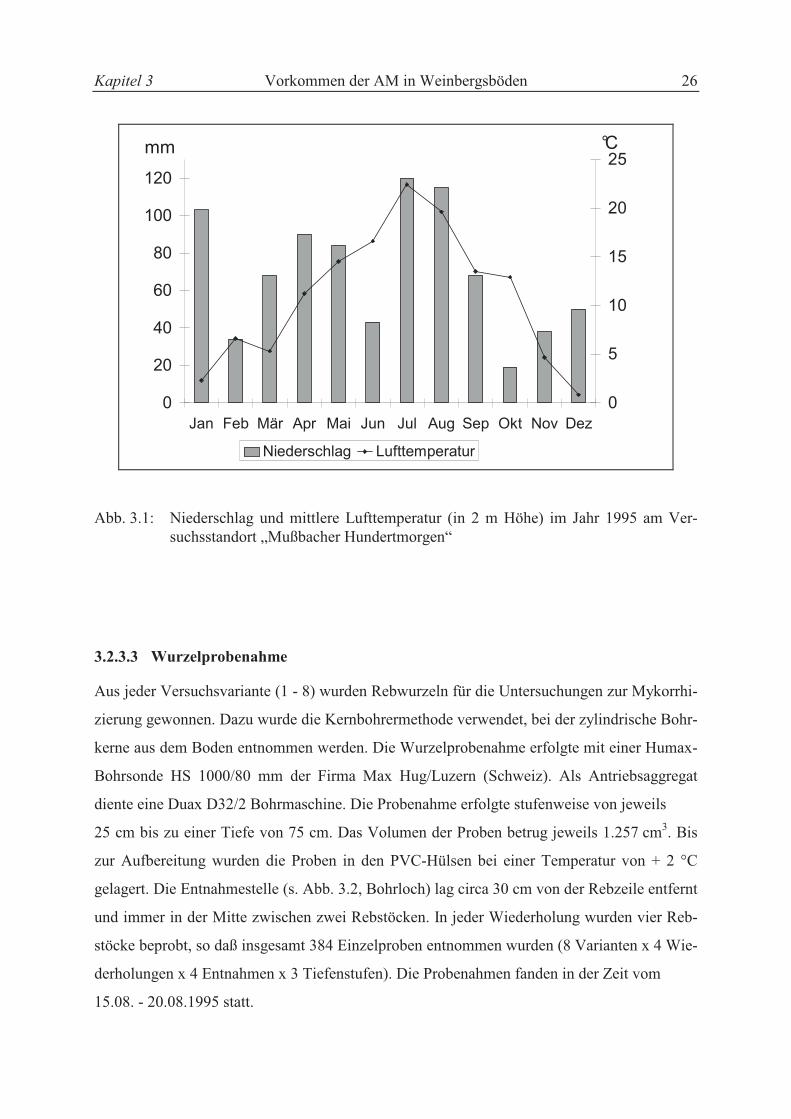
3.2.3.2 Witterungsbedingungen
Die Klimadaten des Versuchsstandortes wie Niederschlag und mittlere Tagesdurchschnitts-
temperatur wurden von einer agrarmeterologischen Station, die etwa 300 m von dem Ver-
suchsstandort entfernt lag, aufgezeichnet. Die Jahressumme des Niederschlages lag 1995 mit
831 mm erheblich über dem 30jährigen Mittel (621 mm). Zum Zeitpunkt der Wurzelprobe-
nahme fielen ergiebige Niederschläge (s. Abb. 3.1). Das Jahresmittel der Lufttemperatur in 2
m Höhe lag mit 10,9 °C 1995 über dem 30jährigen Durchschnitt (10,4 °C).
Kapitel 3 Vorkommen der AM in Weinbergsböden 26
0
20
40
60
80
100
120
Jan Feb Mär Apr Mai Jun Jul Aug Sep Okt Nov Dez
mm
0
5
10
15
20
25°C
Niederschlag Lufttemperatur
Abb. 3.1: Niederschlag und mittlere Lufttemperatur (in 2 m Höhe) im Jahr 1995 am Ver-suchsstandort „Mußbacher Hundertmorgen“
3.2.3.3 Wurzelprobenahme
Aus jeder Versuchsvariante (1 - 8) wurden Rebwurzeln für die Untersuchungen zur Mykorrhi-
zierung gewonnen. Dazu wurde die Kernbohrermethode verwendet, bei der zylindrische Bohr-
kerne aus dem Boden entnommen werden. Die Wurzelprobenahme erfolgte mit einer Humax-
Bohrsonde HS 1000/80 mm der Firma Max Hug/Luzern (Schweiz). Als Antriebsaggregat
diente eine Duax D32/2 Bohrmaschine. Die Probenahme erfolgte stufenweise von jeweils
25 cm bis zu einer Tiefe von 75 cm. Das Volumen der Proben betrug jeweils 1.257 cm3. Bis
zur Aufbereitung wurden die Proben in den PVC-Hülsen bei einer Temperatur von + 2 °C
gelagert. Die Entnahmestelle (s. Abb. 3.2, Bohrloch) lag circa 30 cm von der Rebzeile entfernt
und immer in der Mitte zwischen zwei Rebstöcken. In jeder Wiederholung wurden vier Reb-
stöcke beprobt, so daß insgesamt 384 Einzelproben entnommen wurden (8 Varianten x 4 Wie-
derholungen x 4 Entnahmen x 3 Tiefenstufen). Die Probenahmen fanden in der Zeit vom
15.08. - 20.08.1995 statt.
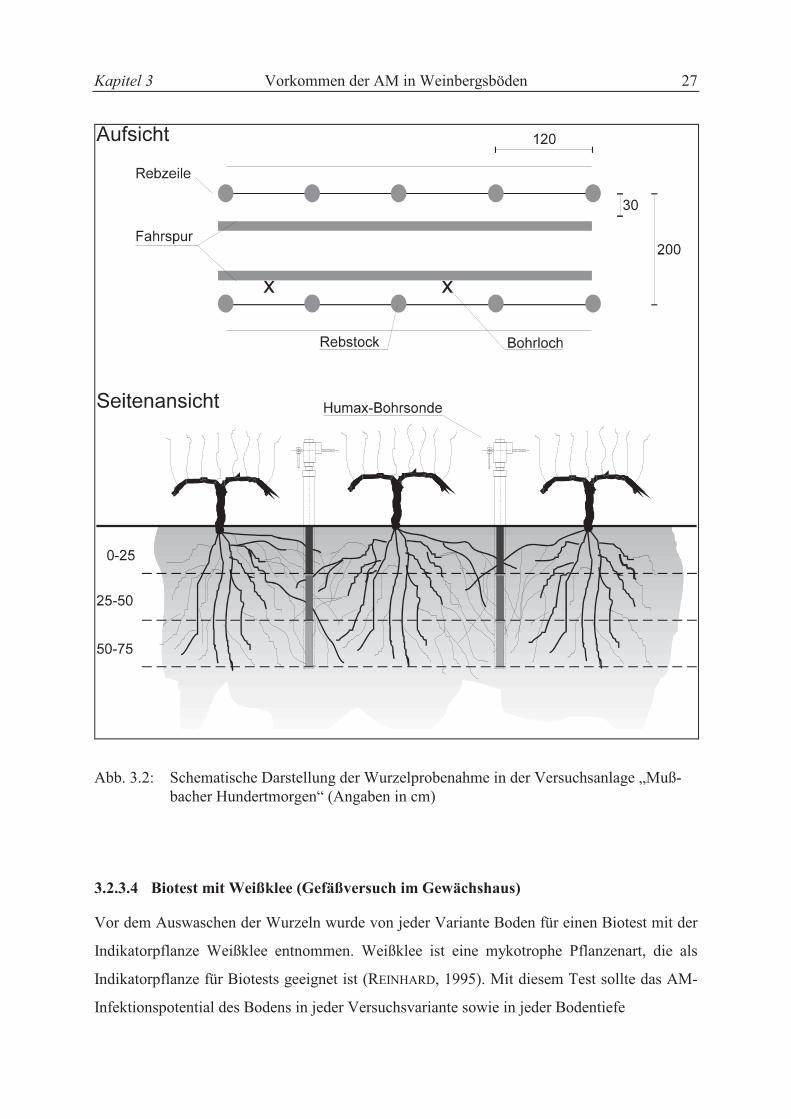
Kapitel 3 Vorkommen der AM in Weinbergsböden 27
Abb. 3.2: Schematische Darstellung der Wurzelprobenahme in der Versuchsanlage „Muß-bacher Hundertmorgen“ (Angaben in cm)
3.2.3.4 Biotest mit Weißklee (Gefäßversuch im Gewächshaus)
Vor dem Auswaschen der Wurzeln wurde von jeder Variante Boden für einen Biotest mit der
Indikatorpflanze Weißklee entnommen. Weißklee ist eine mykotrophe Pflanzenart, die als
Indikatorpflanze für Biotests geeignet ist (REINHARD, 1995). Mit diesem Test sollte das AM-
Infektionspotential des Bodens in jeder Versuchsvariante sowie in jeder Bodentiefe

Kapitel 3 Vorkommen der AM in Weinbergsböden 28
(0 - 25, 25 - 50, 50 - 75 cm) der Versuchsanlage festgestellt werden. Von jeder Variante wur-
den 6 Töpfe (275 cm3) mit Boden gefüllt, anschließend Weißklee (Trifolium repens L.,
cv. Huja) eingesät und im Gewächshaus bei einer durchschnittlichen Temperatur von 25 ± 2
°C am Tag (16 h) bzw. 22 ± 3 °C in der Nacht (8 h) kultiviert. Der Weißklee wurde 3 bzw. 9
Wochen nach der Aussaat abgeerntet und nach dem Auswaschen der Wurzeln der AM-
Infektionsgrad nach der in Kapitel 2.4 beschriebenen Methode bestimmt. Von jeder Variante
(8 Varianten x 4 Wiederholungen x 3 Tiefenstufen) standen 3 Töpfe pro Erntetermin zur Ver-
fügung, die beim Auswaschen zu einer Probe zusammengefaßt wurden. Dadurch stand genü-
gend Wurzelmaterial für Untersuchungszwecke zur Verfügung.
3.2.3.5 Wurzelprobenaufbereitung
Die Proben wurden einzeln für 20 Minuten in Wasser eingeweicht, damit die Rebwurzeln
leichter aus den Bohrkernen zu entnehmen waren. Die teilweise zerfallenden Proben wurden
auf ein Sieb (Edelstahl, Maschenweite 1 mm; Außenmaße 50 x 50 cm) überspült und die Fei-
nerde unter einem mäßig starken Wasserstrahl abgespült. Auf dem Sieb wurden die Rebwur-
zeln mit einer Pinzette ausgelesen, wobei eine Klassifizierung anhand von Farbe und Durch-
messer der Wurzeln durchgeführt wurde. Für die Mykorrhiza-Untersuchung wurden nur Wur-
zeln mit einem Durchmesser von weniger als 2 mm berücksichtigt. Beim Auswaschen der
Bohrkerne stellte sich heraus, daß die Wurzelausbeute sehr gering war. Deshalb wurden die
Wurzeln der vier Entnahmestellen getrennt für jede Wiederholung und Tiefenstufe zu einer
Probe zusammengefaßt.
3.3 Ergebnisse
3.3.1 Untersuchungen zum AM-Infektionsgrad in Rebschulen
Auf allen zehn untersuchten Versuchsflächen im Anbaugebiet „Pfalz“ konnte an den Rebwur-
zeln arbuskuläre Mykorrhiza festgestellt werden. Der niedrigste Infektionsgrad bei der Unter-
suchung über das Vorkommen der arbuskulären Mykorrhiza in Rebschulen lag mit 2,1 % bei
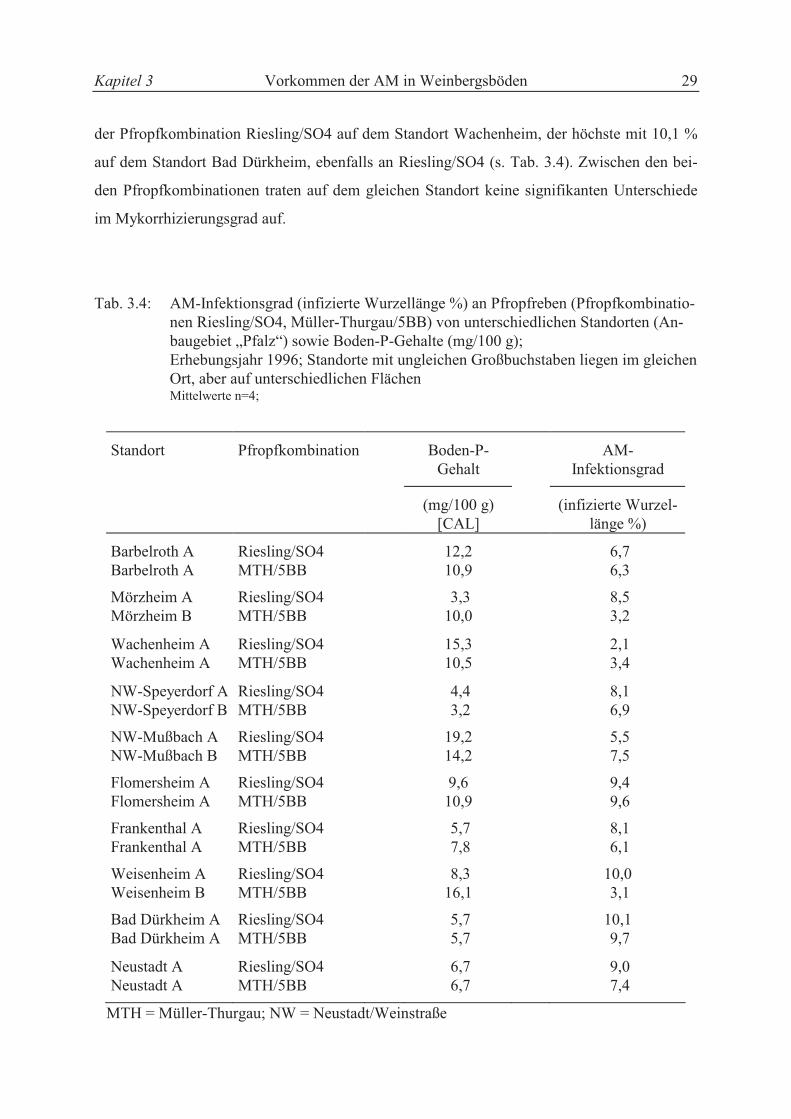
Kapitel 3 Vorkommen der AM in Weinbergsböden 29
der Pfropfkombination Riesling/SO4 auf dem Standort Wachenheim, der höchste mit 10,1 %
auf dem Standort Bad Dürkheim, ebenfalls an Riesling/SO4 (s. Tab. 3.4). Zwischen den bei-
den Pfropfkombinationen traten auf dem gleichen Standort keine signifikanten Unterschiede
im Mykorrhizierungsgrad auf.
Tab. 3.4: AM-Infektionsgrad (infizierte Wurzellänge %) an Pfropfreben (Pfropfkombinatio-nen Riesling/SO4, Müller-Thurgau/5BB) von unterschiedlichen Standorten (An-baugebiet „Pfalz“) sowie Boden-P-Gehalte (mg/100 g);
Erhebungsjahr 1996; Standorte mit ungleichen Großbuchstaben liegen im gleichen Ort, aber auf unterschiedlichen Flächen
Mittelwerte n=4;
Standort Pfropfkombination Boden-P-Gehalt
AM- Infektionsgrad
(mg/100 g) [CAL]
(infizierte Wurzel-länge %)
Barbelroth A Riesling/SO4 12,2 6,7 Barbelroth A MTH/5BB 10,9 6,3
Mörzheim A Riesling/SO4 3,3 8,5 Mörzheim B MTH/5BB 10,0 3,2
Wachenheim A Riesling/SO4 15,3 2,1 Wachenheim A MTH/5BB 10,5 3,4
NW-Speyerdorf A Riesling/SO4 4,4 8,1 NW-Speyerdorf B MTH/5BB 3,2 6,9
NW-Mußbach A Riesling/SO4 19,2 5,5 NW-Mußbach B MTH/5BB 14,2 7,5
Flomersheim A Riesling/SO4 9,6 9,4 Flomersheim A MTH/5BB 10,9 9,6
Frankenthal A Riesling/SO4 5,7 8,1 Frankenthal A MTH/5BB 7,8 6,1
Weisenheim A Riesling/SO4 8,3 10,0 Weisenheim B MTH/5BB 16,1 3,1
Bad Dürkheim A Riesling/SO4 5,7 10,1 Bad Dürkheim A MTH/5BB 5,7 9,7
Neustadt A Riesling/SO4 6,7 9,0 Neustadt A MTH/5BB 6,7 7,4
MTH = Müller-Thurgau; NW = Neustadt/Weinstraße
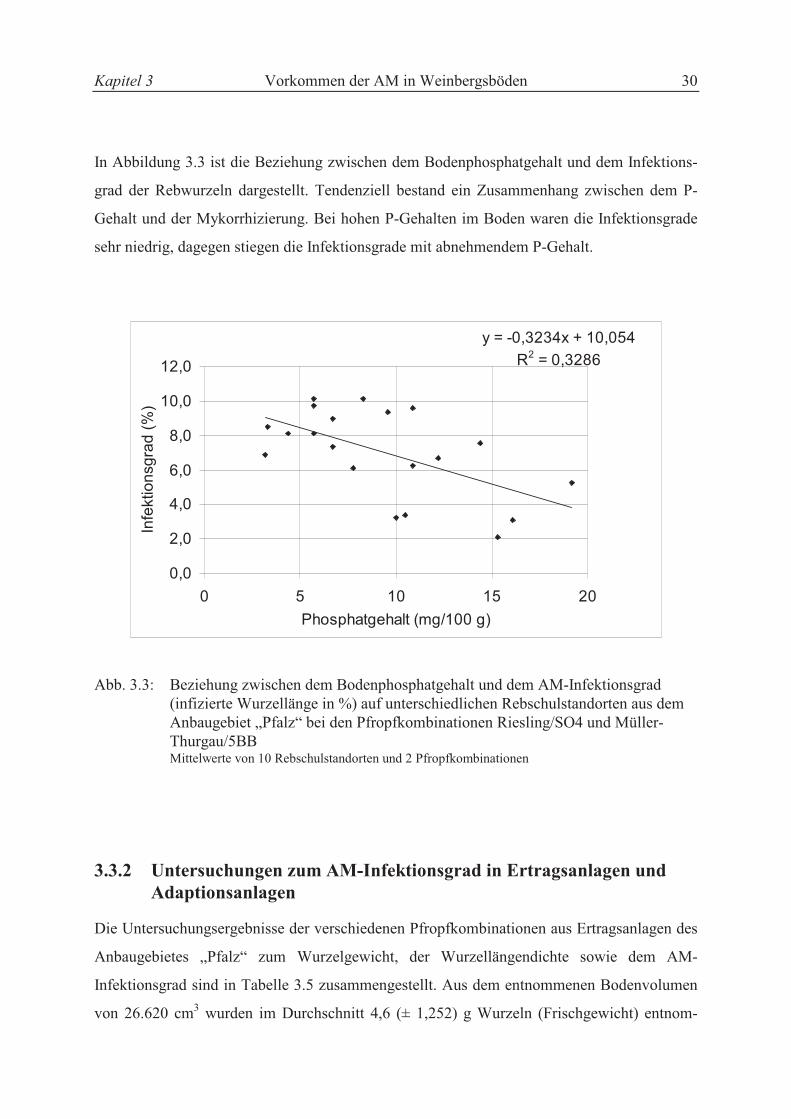
Kapitel 3 Vorkommen der AM in Weinbergsböden 30
In Abbildung 3.3 ist die Beziehung zwischen dem Bodenphosphatgehalt und dem Infektions-
grad der Rebwurzeln dargestellt. Tendenziell bestand ein Zusammenhang zwischen dem P-
Gehalt und der Mykorrhizierung. Bei hohen P-Gehalten im Boden waren die Infektionsgrade
sehr niedrig, dagegen stiegen die Infektionsgrade mit abnehmendem P-Gehalt.
y = -0,3234x + 10,054
R2 = 0,3286
0,0
2,0
4,0
6,0
8,0
10,0
12,0
0 5 10 15 20
Phosphatgehalt (mg/100 g)
Infe
ktio
nsg
rad
(%
)
Abb. 3.3: Beziehung zwischen dem Bodenphosphatgehalt und dem AM-Infektionsgrad (infizierte Wurzellänge in %) auf unterschiedlichen Rebschulstandorten aus dem Anbaugebiet „Pfalz“ bei den Pfropfkombinationen Riesling/SO4 und Müller-
Thurgau/5BB Mittelwerte von 10 Rebschulstandorten und 2 Pfropfkombinationen
3.3.2 Untersuchungen zum AM-Infektionsgrad in Ertragsanlagen und Adaptionsanlagen
Die Untersuchungsergebnisse der verschiedenen Pfropfkombinationen aus Ertragsanlagen des
Anbaugebietes „Pfalz“ zum Wurzelgewicht, der Wurzellängendichte sowie dem AM-
Infektionsgrad sind in Tabelle 3.5 zusammengestellt. Aus dem entnommenen Bodenvolumen
von 26.620 cm3 wurden im Durchschnitt 4,6 (± 1,252) g Wurzeln (Frischgewicht) entnom-

Kapitel 3 Vorkommen der AM in Weinbergsböden 31
men. Die aus den Daten errechnete Wurzellängendichte betrug im Durchschnitt 0,041
(± 0,0136) cm/cm3. Zwischen den Unterlagen auf den Standorten Böchingen I, Wollmesheim
I, Maikammer, Gleisweiler I und II traten im AM-Infektionsgrad keine signifikanten Unter-
schiede auf. In den ökologisch bewirtschafteten Weinbergen lag der AM-Infektionsgrad zwi-
schen 8,5 % (Böchingen II) und 18 % (Wollmesheim II). In der Adaptionsanlage von Assel-
heim traten im AM-Infektionsgrad signifikante Unterschiede zwischen den Unterlagensorten
auf
(s. Tab. 3.5). An der Unterlage Fercal und C3 war die AM-Infektion mit 6,7 % signifikant
geringer als an der Unterlage A9 mit 20,1 %. Zwischen den übrigen Unterlagen gab es keine
Unterschiede. Die durchschnittliche AM-Infektion lag beim Standort Asselheim bei 10,6 %.
In einem Vorversuch aus dem Jahr 1995 konnten ähnliche Ergebnisse festgestellt werden. Der
durchschnittliche AM-Infektionsgrad des Vorversuches lag mit 12,2 % etwas höher als der im
vorliegenden Versuch. Der höchste Infektionsgrad in einer Einzelprobe mit 24,7 % wurde
ebenfalls an der Unterlagssorte A9 bestimmt.
Die Mineralstoffgehalte in den Blättern waren bei den einzelnen Unterlagssorten aus der glei-
chen Adaptionsanlage nicht signifikant unterschiedlich (vgl. Tab. A2 und Tab. A3). Der P-
Gehalt in den Blättern der Unterlagen von Asselheim lag zwischen 1,5 und 1,7 mg P/g Tro-
ckensubstanz (TS) (s. Tab. A2) bei einem Boden-P-Gehalt von 7,0 mg/100 g Boden (Tab.
3.1). Die höchsten P-Gehalte in der Trockensubstanz der Blätter traten auf dem Standort Mai-
kammer (gemittelt über alle Varianten) mit 2,5 mg/100 g TS auf. Die Mikronährstoffe in den
Blättern waren ähnlich wie die Makronährstoffe der einzelnen Unterlagen aus der gleichen
Adaptionsanlage nicht signifikant unterschiedlich (s. Tab. A3). Insbesondere in den ökolo-
gisch bewirtschafteten Flächen traten zum Teil sehr hohe Cu-Gehalte (z. B. Mörzheim mit 97
mg/kg TS) in den Blättern auf, die auf den Einsatz von Cu-haltigen Pflanzenschutzmitteln
zurückgeführt werden (KOPF, pers. Mitteilung). Die erhöhten Fe-Gehalte in den Blättern vom
Standort Asselheim im Vergleich zu den anderen Standorten (Tab. A3) gehen auf die Applika-
tion von eisenhaltigem Blattdünger (Fetrilon Combi) zurück (GAUL, pers. Mitteilung).
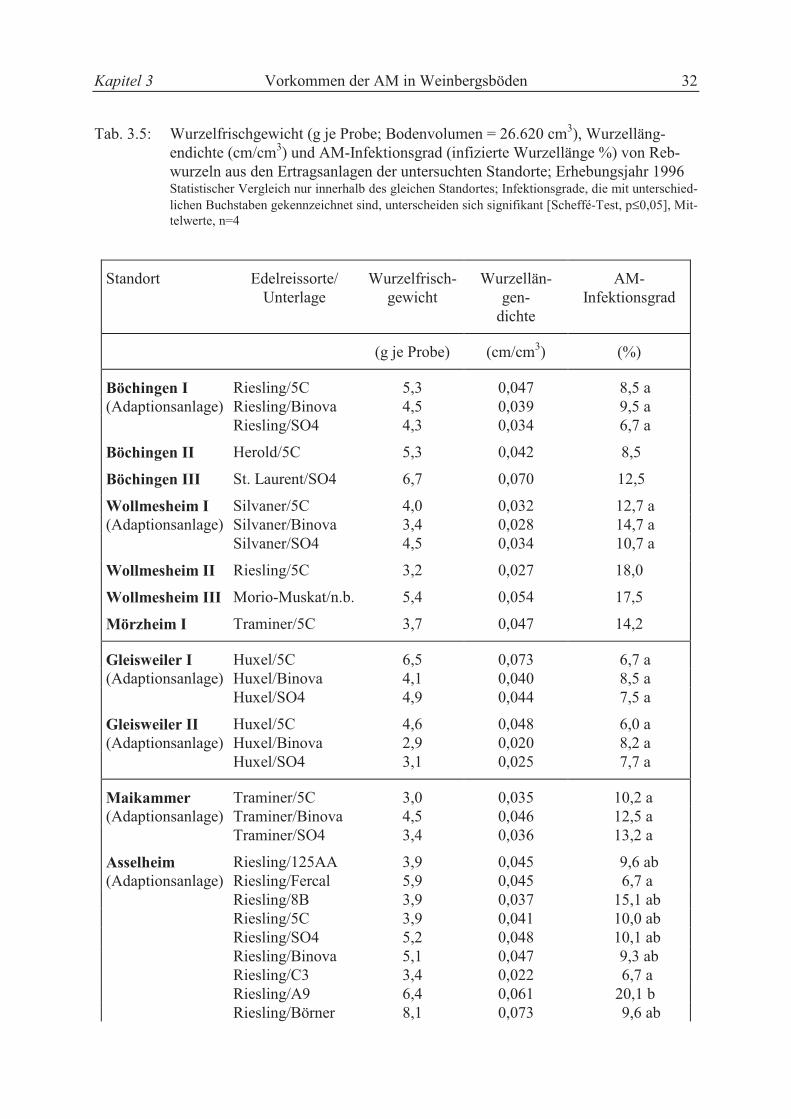
Kapitel 3 Vorkommen der AM in Weinbergsböden 32
Tab. 3.5: Wurzelfrischgewicht (g je Probe; Bodenvolumen = 26.620 cm3), Wurzelläng-endichte (cm/cm3) und AM-Infektionsgrad (infizierte Wurzellänge %) von Reb-wurzeln aus den Ertragsanlagen der untersuchten Standorte; Erhebungsjahr 1996
Statistischer Vergleich nur innerhalb des gleichen Standortes; Infektionsgrade, die mit unterschied-lichen Buchstaben gekennzeichnet sind, unterscheiden sich signifikant [Scheffé-Test, p≤0,05], Mit-telwerte, n=4
Standort Edelreissorte/ Unterlage
Wurzelfrisch- gewicht
Wurzellän-gen-
dichte
AM-Infektionsgrad
(g je Probe) (cm/cm3) (%)
Böchingen I Riesling/5C 5,3 0,047 8,5 a (Adaptionsanlage) Riesling/Binova 4,5 0,039 9,5 a Riesling/SO4 4,3 0,034 6,7 a
Böchingen II Herold/5C 5,3 0,042 8,5
Böchingen III St. Laurent/SO4 6,7 0,070 12,5
Wollmesheim I Silvaner/5C 4,0 0,032 12,7 a (Adaptionsanlage) Silvaner/Binova 3,4 0,028 14,7 a Silvaner/SO4 4,5 0,034 10,7 a
Wollmesheim II Riesling/5C 3,2 0,027 18,0
Wollmesheim III Morio-Muskat/n.b. 5,4 0,054 17,5
Mörzheim I Traminer/5C 3,7 0,047 14,2
Gleisweiler I Huxel/5C 6,5 0,073 6,7 a (Adaptionsanlage) Huxel/Binova 4,1 0,040 8,5 a Huxel/SO4 4,9 0,044 7,5 a
Gleisweiler II Huxel/5C 4,6 0,048 6,0 a (Adaptionsanlage) Huxel/Binova 2,9 0,020 8,2 a Huxel/SO4 3,1 0,025 7,7 a
Maikammer Traminer/5C 3,0 0,035 10,2 a (Adaptionsanlage) Traminer/Binova 4,5 0,046 12,5 a Traminer/SO4 3,4 0,036 13,2 a
Asselheim Riesling/125AA 3,9 0,045 9,6 ab (Adaptionsanlage) Riesling/Fercal 5,9 0,045 6,7 a Riesling/8B 3,9 0,037 15,1 ab Riesling/5C 3,9 0,041 10,0 ab Riesling/SO4 5,2 0,048 10,1 ab Riesling/Binova 5,1 0,047 9,3 ab Riesling/C3 3,4 0,022 6,7 a Riesling/A9 6,4 0,061 20,1 b Riesling/Börner 8,1 0,073 9,6 ab

Kapitel 3 Vorkommen der AM in Weinbergsböden 33
Riesling/26G 5,6 0,040 9,0 ab
3.3.3 Einfluß von Begrünungseinsaaten und Bodenbearbeitung auf die autochthone Mykorrhiza
3.3.3.1 AM-Infektionsgrad der Rebwurzeln
In allen Varianten konnte an den Rebwurzeln arbuskuläre Mykorrhiza festgestellt werden. Der
durchschnittliche Infektionsgrad in den Versuchsvarianten 1 - 8 lag bei 11,3 % (s. Abb. 3.4).
Der niedrigste Infektionsgrad betrug 6 % in der Kontrollfläche, in der keine Begrünungsein-
saat erfolgte und der Boden über das ganze Jahr offen gehalten wurde. Der höchste Infekti-
onsgrad wurde mit 15 % in der Variante 4 (Wintergründüngung mit Leguminoseneinsaat)
ermittelt. In der Tendenz waren die Rebwurzeln aus der Kontrollfläche im Vergleich zu den
anderen Versuchsvarianten (2 - 8) weniger mykorrhiziert. Dieser Unterschied war bei den
Varianten Wintergründüngung mit Gramineen und Leguminosen sowie bei der Zweijahresbe-
grünung mit Gramineen statistisch abgesichert (p≤0,05 %). Die Unterschiede zwischen den
Varianten 1 und 4 (Wintergründüngung mit Leguminosen) konnten bei der Irrtumswahr-
scheinlichkeit von p≤0,01 % abgesichert werden. Die Rebwurzeln aus der Variante 2 (Winter-
gründüngung mit Cruciferen) zeigten tendenziell niedrigere Infektionsgrade als alle anderen
Begrünungsvarianten. Signifikante Unterschiede zwischen den Begrünungsvarianten traten
bei der Versuchsvariante Wintergründüngung mit Leguminosen und Wintergründüngung mit
Cruciferen auf. Zwischen den zweijährig- bzw. dauerbegrünten Varianten wurden keine Un-
terschiede im Infektionsgrad festgestellt (s. Abb. 3.4).
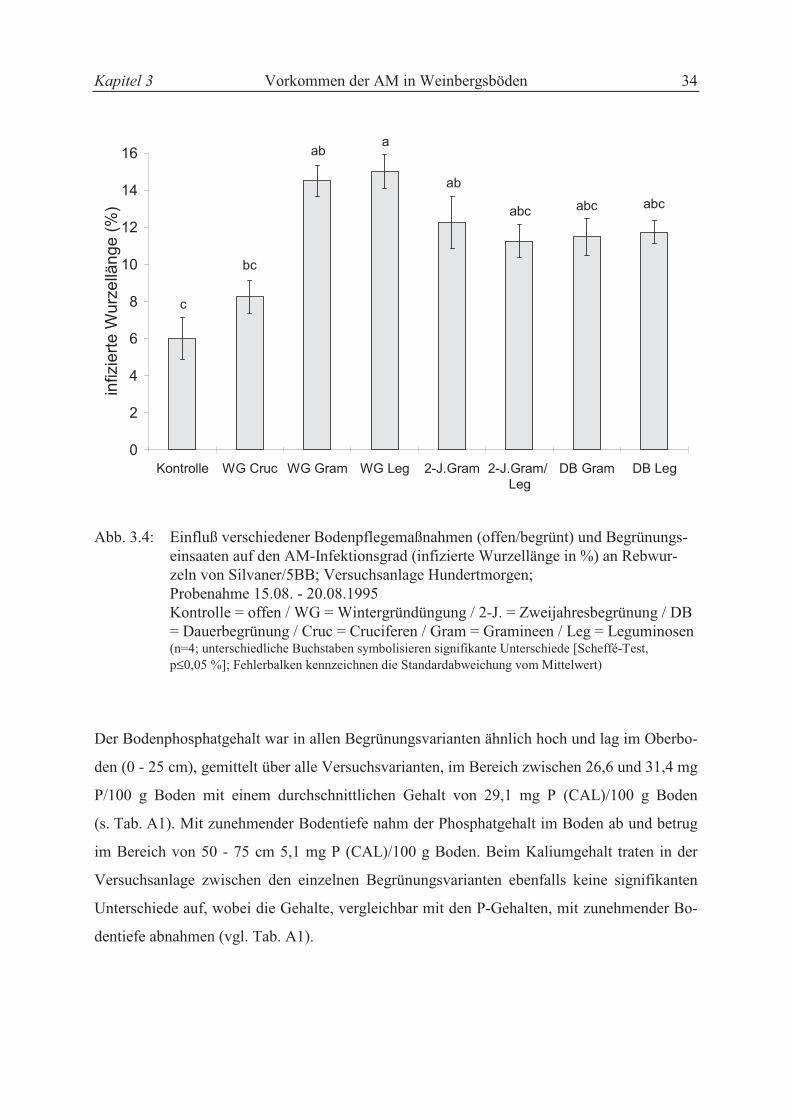
Kapitel 3 Vorkommen der AM in Weinbergsböden 34
c
bc
aba
ab
abc abc abc
0
2
4
6
8
10
12
14
16
Kontrolle WG Cruc WG Gram WG Leg 2-J.Gram 2-J.Gram/
Leg
DB Gram DB Leg
infizie
rte
Wu
rze
llän
ge
(%
)
Abb. 3.4: Einfluß verschiedener Bodenpflegemaßnahmen (offen/begrünt) und Begrünungs-einsaaten auf den AM-Infektionsgrad (infizierte Wurzellänge in %) an Rebwur-zeln von Silvaner/5BB; Versuchsanlage Hundertmorgen;
Probenahme 15.08. - 20.08.1995 Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung / DB
= Dauerbegrünung / Cruc = Cruciferen / Gram = Gramineen / Leg = Leguminosen (n=4; unterschiedliche Buchstaben symbolisieren signifikante Unterschiede [Scheffé-Test,
p≤0,05 %]; Fehlerbalken kennzeichnen die Standardabweichung vom Mittelwert)
Der Bodenphosphatgehalt war in allen Begrünungsvarianten ähnlich hoch und lag im Oberbo-
den (0 - 25 cm), gemittelt über alle Versuchsvarianten, im Bereich zwischen 26,6 und 31,4 mg
P/100 g Boden mit einem durchschnittlichen Gehalt von 29,1 mg P (CAL)/100 g Boden
(s. Tab. A1). Mit zunehmender Bodentiefe nahm der Phosphatgehalt im Boden ab und betrug
im Bereich von 50 - 75 cm 5,1 mg P (CAL)/100 g Boden. Beim Kaliumgehalt traten in der
Versuchsanlage zwischen den einzelnen Begrünungsvarianten ebenfalls keine signifikanten
Unterschiede auf, wobei die Gehalte, vergleichbar mit den P-Gehalten, mit zunehmender Bo-
dentiefe abnahmen (vgl. Tab. A1).

Kapitel 3 Vorkommen der AM in Weinbergsböden 35
3.3.3.2 AM-Infektionsgrad der Weißkleewurzeln (Bioindikatorpflanzen) im Gefäßversuch nach 3- und 9wöchiger Versuchsphase
Die Ergebnisse aus dem Biotest mit Weißklee sind in Abbildung 3.5 dargestellt. Nach
3wöchiger Wachstumsphase waren die Weißkleewurzeln im Boden aus den verschiedenen
Varianten der Versuchsanlage Hundertmorgen nur gering infiziert. Der durchschnittliche In-
fektionsgrad betrug 3 %. Sowohl zwischen den Begrünungsvarianten als auch innerhalb der
gleichen Varianten in unterschiedlicher Bodentiefe traten keine signifikanten Unterschiede im
Infektionsgrad auf. Nach weiteren 6 Wochen stieg der durchschnittliche Infektionsgrad auf
10,4 % an. Tendenziell waren die Weißkleewurzeln, die im Boden aus dem Bereich von 0 - 25
und 25 - 50 cm wuchsen, höher infiziert als die Varianten, die in dem Boden aus der untersten
Tiefenstufe (50 - 75 cm) wuchsen (vgl. Abb. 3.5). Bei den Weißkleewurzeln aus dem Oberbo-
den konnte ein durchschnittlicher Infektionsgrad von 15,1 % ermittelt werden, im mittleren
Bodenbereich (25 - 50 cm) von 10,4 % und im unteren Bereich von 5,7 %. Diese Unterschie-
de konnten bei fast allen Varianten, außer den Varianten 2 und 3, statistisch abgesichert wer-
den.
Bei der Tiefenstufe 0 - 25 cm traten zwischen den Varianten signifikante Unterschiede auf
(s. Abb. 3.5). Der Infektionsgrad des Weißklees aus der Variante mit der Zweijahresbegrü-
nung mit Gramineen und Leguminosen lag bei 19,2 % und war signifikant höher als bei der
Wintergründüngung mit Cruciferen mit 9,7 %. Signifikante Unterschiede in den Varianten aus
dem mittleren Tiefenbereich (25 - 50 cm) traten bei der Dauerbegrünung mit Gramineen und
der Kontrolle sowie bei der Wintergründüngung mit Leguminosen und der Kontrolle auf, wo-
bei die letztgenannte Variante mit 6,2 % den niedrigsten Infektionsgrad aufzeigte (s. Abb.
3.5). Im unteren Bodenbereich traten keine signifikanten Unterschiede im Infektionsgrad an
den Weißkleewurzeln auf.
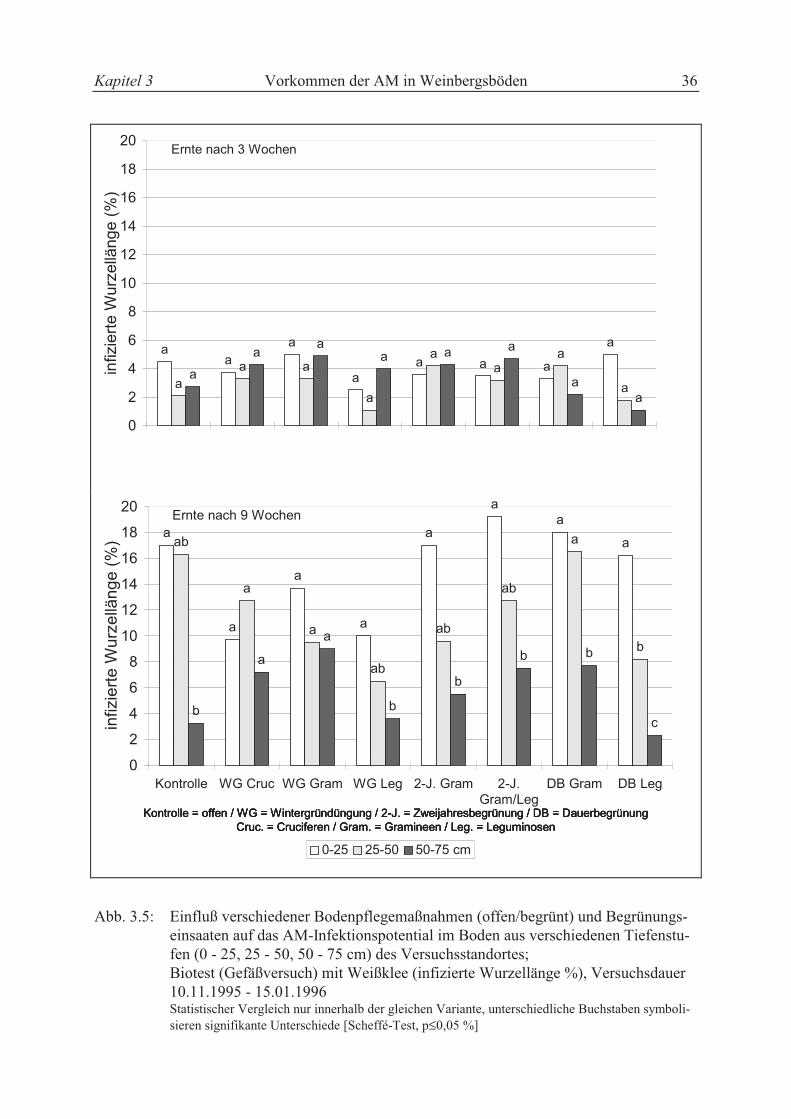
Kapitel 3 Vorkommen der AM in Weinbergsböden 36
a
aaa
a
a
aa
aa
a a
a
aa
a
a
a
aa
a a a
a
0
2
4
6
8
10
12
14
16
18
20
infizie
rte W
urz
ellä
nge (
%)
Ernte nach 3 Wochen
a
a
a
a
a
a
a
aa
ab
ab
ab
a
a
ab
bbb
b
b
a
a
bc
0
2
4
6
8
10
12
14
16
18
20
Kontrolle WG Cruc WG Gram WG Leg 2-J. Gram 2-J.
Gram/Leg
DB Gram DB Leg
infizie
rte
Wu
rze
llän
ge
(%
)
0-25 25-50 50-75 cm
Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung / DB = Dauerbegrünung
Cruc. = Cruciferen / Gram. = Gramineen / Leg. = Leguminosen
Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung / DB = Dauerbegrünung
Cruc. = Cruciferen / Gram. = Gramineen / Leg. = Leguminosen
Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung / DB = Dauerbegrünung
Cruc. = Cruciferen / Gram. = Gramineen / Leg. = Leguminosen
Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung / DB = Dauerbegrünung
Cruc. = Cruciferen / Gram. = Gramineen / Leg. = Leguminosen
Ernte nach 9 Wochen
Abb. 3.5: Einfluß verschiedener Bodenpflegemaßnahmen (offen/begrünt) und Begrünungs-einsaaten auf das AM-Infektionspotential im Boden aus verschiedenen Tiefenstu-fen (0 - 25, 25 - 50, 50 - 75 cm) des Versuchsstandortes;
Biotest (Gefäßversuch) mit Weißklee (infizierte Wurzellänge %), Versuchsdauer 10.11.1995 - 15.01.1996
Statistischer Vergleich nur innerhalb der gleichen Variante, unterschiedliche Buchstaben symboli-sieren signifikante Unterschiede [Scheffé-Test, p≤0,05 %]

Kapitel 3 Vorkommen der AM in Weinbergsböden 37
3.4 Diskussion
3.4.1 AM-Infektionsgrad von Rebwurzeln aus Rebschulen und Ertragsanlagen
Die aus den verschiedenen Rebschulflächen gewonnenen Daten zeigen, daß die arbuskuläre
Mykorrhiza bei der Rebe auch an jungen Pfropfreben vorkommt, allerdings waren die Infekti-
onsgrade nur sehr niedrig. Im Mittel aller untersuchter Rebschulflächen betrug der Infektions-
grad bei den Pfropfkombinationen Riesling/SO4 und Müller-Thurgau/5BB 6,7 % (s. Tab.
3.4). Diese Pfropfkombinationen wurden für die vorliegenden Untersuchungen gewählt, weil
es sich bei den Unterlagssorten SO4 und 5BB um die am häufigsten verwendeten Unterlagen
im deutschen Weinbau handelt. SCHUBERT und CRAVERO (1985) fanden in Rebwurzeln aus
verschiedenen Ertragsweinbergen in Nord-West-Italien deutlich höhere Infektionsgrade (> 25
%). In Rebschulen ist möglicherweise die Mykorrhizierung im allgemeinen geringer als in
Ertragsanlagen. In Ertragsanlagen breitet sich das Wurzelsystem über mehrere Jahre aus. Die
von der arbuskulären Mykorrhiza gebildeten Dauersporen können über einen längeren Zeit-
raum auskeimen und somit das Wurzelsystem der Rebe intensiver infizieren. Dagegen ist bei
den Pfropfreben in der Rebschule nur ein Kontakt der infektiösen Strukturen und den Wurzeln
von etwa vier Monaten gegeben. Es ist davon auszugehen, daß aufgrund der relativ kurzen
Kulturzeit in Rebschulen die Pfropfreben weniger intensiv mit AM-Pilzen besiedelt werden
als in Ertragsanlagen. Durch die hohe Pflanzdichte in Rebschulen mit etwa 12 - 15 Reben je
Meter Pflanzfolie kommt es zu einer wesentlich intensiveren Durchwurzelung des Oberbo-
dens und somit liegt eine größere Infektionsmöglichkeit mit AM-Pilzen vor als in Ertragsan-
lagen, in denen der Stockabstand zwischen 0,90 m und 1,20 m liegt. Trotzdem waren die AM-
Infektionsgrade in den untersuchten Rebschulen niedriger als in den Ertragsanlagen. Es wurde
nachgewiesen, daß es in Rebschulen unter der Mulchfolie zu Temperaturerhöhungen und
Nährstoffakkumulationen (CURRLE et al., 1983) kommt. Dadurch wurden die Infektionsbe-
dingungen offensichtlich gestört.
In den Rebschulflächen wurde tendenziell eine negative Korrelation zwischen dem Boden-
P-Gehalt und dem AM-Infektionsgrad festgestellt (vgl. Abb. 3.3). Dies steht im Einklang mit
MILLER et al. (1985), die ebenfalls unter Feldbedingungen eine negative, allerdings nicht sig-
nifikante Korrelation des Infektionsgrades von Apfelstecklingen mit der P-Verfügbarkeit des
Bodens feststellten. EIBACH (1982) konnte nach dem Topfen von Müller-Thurgau-Stecklingen

Kapitel 3 Vorkommen der AM in Weinbergsböden 38
in Böden aus sechs verschiedenen deutschen Weinanbaugebieten bei der Ermittlung des Infek-
tionsgrades eine signifikante negative Korrelation zum P-Gehalt des Bodens feststellen. In
ihren Versuchen lagen die Infektionsgrade im phosphatreichen Boden auf einem ähnlich nied-
rigen Niveau wie bei den hier vorgestellten Ergebnissen. Die Ursachen für die geringere My-
korrhizierung bei hoher P-Verfügbarkeit liegen nach SANDERS (1975) und SCHWAB et al.
(1983) in einer verminderten Ausbildung des externen Pilzmyzels, wodurch die Anzahl der
Sekundärinfektionen reduziert wird. Folglich kommt es zu einer geringen Ausbildung der
Mykorrhizastrukturen, wie in den eigenen Ergebnissen anhand der Wurzeluntersuchungen in
Rebschulen gezeigt werden konnte.
Das Sporenvorkommen in Weinbergsböden wird durch hohe P-Gehalte im Boden negativ
beeinflußt (NAPPI et al., 1985; SCHUBERT und CRAVERO, 1985; BRENDEL et al., 1990). NAGA-
HASHI et al. (1996) fanden in ihren Untersuchungen bei hohen P-Gehalten eine Hemmung des
Hyphenwachstums bei Gigaspora margarita. TAWARAYA et al. (1996) stellten qualitative und
quantitative Veränderungen im Wurzelexsudat bei Zwiebel (Allium cepa L.) fest, die für das
geringere Hyphenwachstum bei Gigaspora margarita bei hohen P-Gehalten verantwortlich
gemacht werden konnten. Die Ergebnisse aus den Untersuchungen von TAWARAYA et al.
(1996) und NAGAHASHI et al. (1996) können die geringen Infektionsgrade in der vorliegenden
Untersuchung jedoch nur bedingt erklären, da keine detaillierten Beobachtungen zu Hyphen-
wachstum und Wurzelexsudationen erfolgten. Ein bedeutender Grund für die geringe My-
korrhizierung in Rebschulen liegt möglicherweise im Fehlen von Ackerbeikräutern, die zu-
mindest in landwirtschaftlichen Kultursystemen einen wichtigen Beitrag zum Erhalt des My-
korrhizierungspotentials im Boden leisten (WULFF, 1996). In einer sogenannten „low-input-
Variante“ ohne Unkrautbekämpfung und Düngung fanden KURLE und PFLEGER (1994) bei
einer Fruchtfolge mit Mais und Sojabohne ein signifikant höheres Sporenvorkommen und
einen höheren Infektionsgrad als in der konventionell bewirtschafteten Variante. Die Autoren
führten das niedrigere Sporenvorkommen auf das Fehlen der Unkräuter, die AM-Symbionten
sind, in der konventionell bewirtschafteten Variante zurück. Dies steht im Einklang mit den
eigenen Untersuchungen aus dem Begrünungsversuch (s. Kap. 3.3.3, Abb. 3.4) und könnte
auch für Rebschulen gelten. In Rebschulen werden mehrmals im Jahr mechanische Unkraut-
bekämpfungen durchgeführt, um die aufgelaufene Unkrautflora, die als direkte Konkurrenten
für die Pfropfreben gelten, auszuschalten. Durch das Fehlen der Unkräuter fällt die natürliche

Kapitel 3 Vorkommen der AM in Weinbergsböden 39
Verbreitung oder der direkte Eintrag von AM-Infektionspotential durch AM-Wirtspflanzen in
Rebschulen weg. Dadurch könnte es auf Standorten, die mehrere Jahre als Rebschule genutzt
werden, auch langfristig zu einer Abnahme des Mykorrhizierungspotentials im Boden kom-
men, was WULFF (1996) in landwirtschaftlichen Kultursystemen ebenfalls vermutet.
In den Ertragsanlagen hatten die zur Bestimmung des AM-Infektionsgrades ausgegrabenen
Rebwurzeln aus den Bodenblöcken (26.620 cm3) ein durchschnittliches Frischgewicht von
4,6 (± 1,25) g (s. Kap. 3.3.2, Tab. 3.5). Ähnliche Ergebnisse erzielten REIMERS et al. (1994).
Zwischen den verschiedenen Pfropfkombinationen vom gleichen Standort gab es keine signi-
fikanten Unterschiede in der Wurzelmasse. In Texas fanden dagegen PERRY et al. (1983) an
vier Vitis-Arten ein unterschiedliches Wurzelgewicht, bezogen auf ein bestimmtes Bodenvo-
lumen. Dabei hatte Vitis champini cv. Dogridge die größte Wurzelmasse ausgebildet, während
V. rotundifolia cv. Nobel und V. vinifera cv. Barbera die geringste Wurzelmasse ausgebildet
hatten. Die Autoren führten diese Unterschiede auf die Standortbedingungen, unter denen
diese Arten vorkommen, zurück. Ein weitaus wichtigerer Parameter bei Wurzeluntersuchun-
gen als das Gewicht ist die Wurzellängendichte im Boden. MOHR (1996) fand in seinen Un-
tersuchungen bei Müller-Thurgau/5BB an der Mosel Wurzellängendichten von durchschnitt-
lich 0,035 cm Wurzellänge je ml Boden (in 0 - 100 cm Tiefe). Dabei teilte MOHR (1996) die
Rebwurzeln in fünf Klassen ein (A - E), wobei der Wert von 0,035 cm je ml Bodenvolumen
die Klasse A und B (Saugwurzeln mit primärer Rinde mit heller bzw. bräunlicher Farbe) be-
rücksichtigt. Diese Werte stimmen mit eigenen Ergebnissen aus den Ertragsanlagen überein.
Im Durchschnitt aller Standorte wurde in den Weinbergen eine Wurzellängendichte von 0,041
(± 0,014) cm/cm3 ermittelt (s. Tab. 3.5). Im Gegensatz zu den Untersuchungen von MOHR
(1996) wurden die Rebwurzeln in den eigenen Versuchen nicht nach Klassen eingeteilt, son-
dern für die Bestimmung des AM-Infektionsgrades wurden nur Wurzeln mit < 2 mm Durch-
messer berücksichtigt (s. Kap. 3.2.2). Dadurch war die Probenahme vereinfacht und es war
gewährleistet, daß nur Saugwurzeln in die Untersuchung einbezogen wurden. Diese können
nach MOHR (1996) mykorrhiziert werden, während die Leitungswurzeln mit einem Durch-
messer von > 2 mm keine Mykorrhizierung aufweisen.
Ähnlich wie in den Rebschulen wurde in den Ertragsanlagen an jeder Pfropfkombination eine
Infektion mit AM an Rebwurzeln gefunden (s. Tab. 3.5). Signifikante Sortenunterschiede in-

Kapitel 3 Vorkommen der AM in Weinbergsböden 40
nerhalb einer Adaptionsanlage traten lediglich am Standort Asselheim auf, wobei an der
Pfropfkombination Riesling/A9 eine höhere Mykorrhizierung vorhanden war als bei Ries-
ling/Fercal und Riesling/C3. Die P-Gehalte in den Blättern zwischen den einzelnen Pfropf-
kombinationen waren nicht signifikant unterschiedlich. KARAGIANNIDIS et al. (1997) ermittel-
ten in einer Adaptionsanlage mit den Unterlagen 110R, 41B, 140Ru und 1130P mit verschie-
denen Edelreissorten unterschiedliche Mykorrhizierungsgrade zwischen den Pfropfkombinati-
onen. Allerdings konnten die Autoren bei gleichen Boden-P-Gehalten signifikant unterschied-
liche P-Gehalte in den Blättern der Reben feststellen. Bei den Untersuchungen von KARAGI-
ANNIDIS et al. (1997) war möglicherweise die P-Aufnahme durch die Mykorrhizahyphen um
ein Mehrfaches höher als in der vorliegenden Arbeit, denn die Reben waren fast dreimal so
stark infiziert wie in den eigenen Ergebnissen. In den hier beprobten Weinbergen lagen die P-
Gehalte in den Blättern zwischen 1,7 und 2,7 mg P/g Trockenmasse (s. Tab. A2). BERGMANN
(1986) gibt Gehalte zwischen 2,5 und 4,5 mg P/g Trockenmasse für ausreichendes Pflanzen-
wachstum bei Reben an. Obwohl in der Adaptionsanlage Asselheim die niedrigsten P-Gehalte
in den Blättern mit 1,5 - 1,7 mg P/g Trockenmasse auftraten, wurden keine P-Mangel-
symptome, wie z. B. dunkelgrüne Verfärbung der Mangelblätter (MENGEL, 1991), an den Re-
ben beobachtet.
Verschiedene Autoren berichteten darüber, daß die Bewirtschaftungsweise einen bedeutenden
Einfluß auf das Mykorrhiza-Potential hat (DOUDS et al., 1993; RYAN et al., 1994; DOUDS et
al., 1995; FELDMANN, 1997). So wurde festgestellt, daß in ökologisch genutzten Standorten im
Vergleich zu konventionell genutzten ein wesentlich höheres Mykorrhizierungspotential beo-
bachtet wurde. SATTELMACHER et al. (1991) fanden an Roggen, der nach biologisch-
dynamischen Richtlinien angebaut wurde, deutlich höhere Infektionsgrade und mehr AM-
Sporentypen als an konventionell angebautem Roggen. Die Autoren führten die Unterschiede
neben dem unterschiedlichen Boden-P-Gehalt auf das Ergebnis mehrerer sich gegenseitig be-
einflussender Faktoren zurück. Dabei hatten die sehr enge Fruchtfolge und die regelmäßige
Anwendung von Herbiziden, die zu einem stark eingeschränkten Auftreten der Ackerbeikräu-
ter führten, eine große Bedeutung. Nach SAIF (1981) wirkt sich ein höherer Humusgehalt über
eine verbesserte Bodendurchlüftung positiv auf die Sporenbildung aus. In den nach den Richt-
linien von BIOLAND ökologisch bewirtschafteten Weinbergen lag der Humusgehalt um 1 %
höher als in den Weinbergen, die nach kontrolliert umweltschonenden Richtlinien bewirt-

Kapitel 3 Vorkommen der AM in Weinbergsböden 41
schaftet wurden (s. Tab. 3.1). Trotzdem traten in der durchgeführten Untersuchung in den
nach BIOLAND-Richtlinien bewirtschafteten Weinbergen im Vergleich zu den nach KUW-
Richtlinien bzw. den konventionell geführten Weinbergen keine deutlichen Unterschiede in
der AM-Infektion der Rebwurzeln auf. Während SATTELMACHER et al. (1991) das geringe
Infektionspotential im konventionell angebauten Roggen unter anderem auf das Fehlen von
Unkräutern zurückführten, trifft dies für die vorliegende Arbeit nicht zu. Die untersuchten
Weinberge waren bis auf die Adaptionsanlage in Asselheim zwischen den Rebgassen ständig
begrünt.
Bei einjährigen Ackerbaukulturen konnten hinsichtlich des Mykorrhizierungspotentials zwi-
schen verschiedenen Bewirtschaftungssystemen (ökologisch und konventionell) häufig Unter-
schiede nachgewiesen werden. Für die Dauerkultur Rebe müßten hierzu weitere Untersuchun-
gen durchgeführt werden. Im ökologischen Landbau werden oftmals vielgliedrige Fruchtfol-
gen mit einem hohen Leguminosenanteil angebaut, die insbesondere durch die hohe My-
kotrophie der Leguminosen zu einer Erhöhung des AM-Infektionspotentials im Boden beitra-
gen. Um gezielte Aussagen über einen Vergleich zwischen ökologisch und konventionell be-
wirtschafteten Weinbergen hinsichtlich der Mykorrhizierung an den Rebwurzeln treffen zu
können, müßten beide Anbausysteme bei ansonsten gleichen Standortbedingungen miteinan-
der verglichen werden.
3.4.2 Einfluß der Begrünung auf die AM
In den Bodenproben aller Begrünungsvarianten (Wintergründüngungen/2-Jahresbegrünun-
gen/Dauerbegrünungen mit unterschiedlichen Pflanzeneinsaaten) und der Kontrolle ohne Bo-
denbedeckung konnten Rebwurzeln gefunden werden, die mit arbuskulären Mykorrhizen infi-
ziert waren. Allerdings waren die Infektionsgrade relativ niedrig (vgl. Abb. 3.4). Im Mittel
aller untersuchten Varianten betrug der Infektionsgrad bei den Rebwurzeln der Sorte Silva-
ner/5BB 11,3 % und war somit auf einem ähnlichen Niveau wie die Infektionsgrade der Reb-
wurzeln aus den verschiedenen Adaptions- bzw. Ertragsanlagen mit durchschnittlich 10,7 %
(s. Tab. 3.5). Die Versuchsanlage Hundertmorgen zeigte Bodenphosphatgehalte im Oberbo-
den von
29,1 mg P (CAL)/100 g Boden auf und war mit P überversorgt (s. Tab. A1). Über alle Tiefen-

Kapitel 3 Vorkommen der AM in Weinbergsböden 42
stufen gemittelt betrug der P-Gehalt 18,3 mg P (CAL)/100 g. In diesem phosphatreichen Bo-
den kommt es nur bedingt zu einer Infektion mit Mykorrhiza, was bereits in Kapitel 3.4.1 in
der Untersuchung in Rebschulen diskutiert wurde.
KIEFER und WEBER (1986) fanden in ihren Untersuchungen eine leichte Zunahme der Anzahl
von Wurzelspitzen bis zur Zeilenmitte, wobei in der Fahrspur des Bearbeitungsschleppers
weniger Wurzelspitzen gefunden wurden. Daher erfolgte die Wurzelprobenahme mit der Hu-
max Bohrsonde unmittelbar neben der Fahrspur (vgl. Abb. 3.2). Trotzdem war die Wurzel-
ausbeute so gering, daß die Wurzelproben aus den jeweiligen Tiefenstufen und den vier Ent-
nahmestellen innerhalb einer Wiederholung als Mischprobe zusammengefaßt wurden. Auf-
grund der geringen Wurzelausbeute konnte deshalb über die unterschiedliche Mykorrhizie-
rung der Rebwurzeln in verschiedenen Bodentiefen keine Aussage getroffen werden, aber der
Vergleich der AM-Infektion der Rebwurzeln aus den verschiedenen Varianten war gewähr-
leistet und konnte statistisch verrechnet werden (s. Abb. 3.4).
Der positive Einfluß der Begrünung auf das Vorkommen der arbuskulären Mykorrhiza war in
den Begrünungsvarianten Wintergründüngung mit Gramineen und Leguminosen und der
2-Jahresbegrünung mit Gramineen zu erkennen und statistisch absicherbar (s. Abb. 3.4).
Durch die Begrünungseinsaaten wurde der Infektionsgrad der Rebwurzeln erhöht. Die Gräser
und Leguminosen aus den Varianten 3 - 8 sind Wirtspflanzen der AM (HARLEY und HARLEY,
1987). Durch die höhere Wurzellängendichte der Gräser und Leguminosen im Vergleich zu
den Rebwurzeln könnte es zu einem intensiveren Kontakt der Begrünungswurzeln mit den
Rebwurzeln gekommen sein, wodurch die Rebwurzeln stärker mykorrhiziert wurden. Von
ähnlichen positiven Effekten der Begleitflora für die Mykorrhizierung an Kautschukbäumen
(Hevea spec.) berichteten FELDMANN und LIEBEREI (1992). Die Autoren fanden auf Planta-
genstandorten mit Hevea spec. an den Wurzeln der Kautschukbäume einen Mykorrhiza-
Besiedlungsgrad von 38,2 % gegenüber 72,2 % am Naturstandort, der durch eine hohe Anzahl
an primären Pflanzengesellschaften am gleichen Standort gekennzeichnet war. FELDMANN
und LIEBEREI (1992) forderten deshalb die Einführung eines intensiven Mischkultursystems
für Kautschukbaumplantagen mit zahlreichen tropischen Nutzpflanzen, die als AM-Wirte die
Mykorrhizierung an Hevea spec. fördern könnten. Diese Forderung ergäbe sich auch aus den
hier vorgestellten Ergebnissen und würde im Weinbau eine Begrünung der Rebgassen, insbe-

Kapitel 3 Vorkommen der AM in Weinbergsböden 43
sondere mit Leguminosen und Gramineen, die die native Mykorrhiza im Weinberg fördern,
befürworten.
In der Kontrolle wurde mehrmals im Jahr der Boden in einem Bereich bis circa 20 cm tief
bearbeitet und in den oberen Bodenschichten Wurzeln von auflaufenden Unkräutern abge-
schnitten, die das Mykorrhizierungspotential beeinflussen. Die Gefäßversuche von JASPER et
al. (1989) zeigten, daß durch eine simulierte Bodenbewegung im Gefäß das externe Hyphen-
geflecht der Mykorrhiza zerstört wurde und das Mykorrhizierungspotential zurückging. Eine
weitere Erklärung für die geringe Mykorrhizierung an den Rebwurzeln der offen gehaltenen
Variante (Kontrolle) kann sich durch das mehrfache Befahren der Rebgasse mit dem Wein-
bergsschlepper ergeben. Der Boden wird wesentlich stärker verdichtet als in den begrünten
Varianten (WUNDERER, 1992). Diese Bodenverdichtungen, die nach SOMMER (1985) auch
unmittelbar neben der Fahrspur vorkommen, könnten in der vorliegenden Untersuchung eben-
falls aufgetreten sein. NADIAN et al. (1996) führten in Gefäßversuchen eine simulierte Boden-
verdichtung in Pflanzgefäßen mit Erdklee (Trifolium subterraneum L.) durch und stellten mit
zunehmender Bodenverdichtung einen geringeren Beitrag der Mykorrhiza für die P-Aufnahme
und das Wachstum fest. Dabei wurde der Sauerstoffgehalt von 0,18 m3 O2/m3 Boden im nicht
verdichteten Boden auf 0,10 m3 O2/m3 im verdichteten Boden abgesenkt. Die Abnahme des
Sauerstoffgehaltes in verdichteten Böden reduziert nach SAIF (1981) die Intensität der Besied-
lung mit dem AM-Pilz. In einem weiteren Versuchsansatz fanden NADIAN et al. (1997) mit
zunehmender Bodendichte (von 1,1 - 1,6 g/cm3) eine Verringerung der Gesamtwurzellänge an
T. subterraneum und einen geringeren AM-Infektionsgrad; ebenso wurde das Außenhyphen-
geflecht des AM-Pilzes G. intraradices reduziert. Rebwurzeln können bei 1,1 - 1,2 g/cm3 Bo-
dendichte ungehindert wachsen, während es ab einer Bodendichte von 1,5 g/cm3 zu Wuchs-
hemmungen an den Rebwurzeln kommen kann (RICHARDS, 1983). Folglich muß angenom-
men werden, daß die nativen Mykorrhizapilze im Weinberg durch mehrfaches Befahren, ins-
besondere in den nicht begrünten Rebgassen, stark in ihrer Effektivität eingeschränkt worden
sind.
In den vorliegenden Untersuchungen war der Infektionsgrad der Rebwurzeln aus der Varian-
te 2 (Wintergründüngung mit Cruciferen) tendenziell niedriger als bei den anderen Begrü-
nungsvarianten und signifikant niedriger als bei der Variante 4 (Wintergründüngung mit Le-

Kapitel 3 Vorkommen der AM in Weinbergsböden 44
guminosen; vgl. Abb. 3.4). Es gibt zahlreiche Untersuchungen, die in den Wurzeln von Nicht-
wirtspflanzen für AM fungizide Substanzen, wie z. B. Isothiocyanate, vermuten (TESTER et
al., 1987; VIERHEILIG und Ocampo, 1990). Ausgangspunkt für die Bildung der Isothiocyanate
stellen die Glukosinolate dar. Glukosinolate sind eine Gruppe schwefelhaltiger Glykoside, die
in unterschiedlicher Konzentration in Cruciferen vorkommen (VIERHEILIG und OCAMPO,
1990). Durch das Enzym Myrosinase werden die Glukosinolate in verschiedene Produkte ab-
gebaut. Ein Abbauprodukt ist das Isothiocyanat, welches SCHREINER und KOIDE (1993) mit
Hilfe der Dünnschichtchromatographie im Wurzelextrakt verschiedener Brassica-Arten ge-
funden haben. Als Hauptkomponente fanden die Autoren das 4-Hydroxybenzyl-Isothiocyanat,
welches bei Brassica kaber L. die Sporenkeimung von Glomus etunicatum hemmte.
GLENN et al. (1988) fanden in den Wurzeln von Kohl (Brassica campestris L.) und Raps
(Brassica napus L.) Glukosinolat-Konzentrationen von 7 - 524 µmol/g Frischgewicht. EL-
ATRACH et al. (1989) stellten in ihren Untersuchungen mit Luzerne (Medicago sativa L.) und
Kohl (B. oleracea cv. Brunswick) nach einer Inokulation mit Glomus mosseae an der Luzerne
geringere Infektionen fest, wenn beide Pflanzenarten im gleichen Topf wuchsen. VIERHEILIG
und OCAMPO (1990) ermittelten in den Wurzeln von Kohl (B. oleracea) ebenfalls Isothiocya-
nat, welches die Sporenkeimung bei Glomus mosseae hemmte. In einem Agarsystem fanden
GLENN et al. (1988), daß die AM-Pilze Glomus mosseae und Gigaspora gigantea mit ihren
Hyphen zwar in den Apoplasten von Brassica-Wurzeln eindrangen, eine Penetration in das
Wurzelsystem durch die AM-Pilze jedoch nicht erfolgte. Die Isothiocyanate können allerdings
nicht alleine für das Nichtzustandekommen einer AM-Infektion verantwortlich gemacht wer-
den. Neben den fungitoxischen Eigenschaften der Glukosinolate vermuten VIERHEILIG et al.
(1996) noch andere Wirkungsmechanismen, die die Infektion der Nichtwirtswurzeln mit AM
verhinderten. Hierzu zählen die Autoren das Fehlen von Signalen, die von dem AM-Pilz be-
nötigt werden, um eine Pflanzenwurzel infizieren zu können. Die erwähnten Wirkungsmecha-
nismen (Ausscheidung von Hemmstoffen bei Nicht-AM-Symbionten und fehlende Signale
der Nichtwirtspflanzen, die Wurzeln mit AM zu penetrieren) könnten für den negativen
Einfluß der Cruciferen auf die Mykorrhizierung der Rebwurzeln verantwortlich sein.
Um den Einfluß der verschiedenen Begrünungseinsaaten auf die Mykorrhizierung näher zu
beleuchten, wurde neben der Bestimmung des AM-Infektionsgrades an den Rebwurzeln von
jeder Variante mit einem Teil des Bodens aus jeder Tiefenstufe ein Biotest mit Weißklee im

Kapitel 3 Vorkommen der AM in Weinbergsböden 45
Gewächshaus durchgeführt. Die Mykorrhizierungsgrade der Weißkleewurzeln waren relativ
gering (s. Abb. 3.5). JASPER et al. (1991) schlossen aus geringen Infektionsgraden an den
Testpflanzen in Biotests auf ein niedriges Infektionspotential im untersuchten Boden. Dies
wurde an den niedrigen Infektionsgraden der Rebwurzeln im Versuchsweinberg deutlich
(11,3 %, gemittelt über alle Versuchsvarianten). Beim Biotest wurde in der oberen Boden-
schicht (0 - 25 cm) das größte Mykorrhizierungspotential (gemessen am Infektionsgrad der
Weißkleewurzeln) gefunden, obwohl in dieser Bodenschicht der P-Gehalt mit 29,1 mg P
(CAL)/100 g Boden am höchsten war (vgl. Tab. A1). Die Hauptwurzelmasse der Begrü-
nungspflanzen befindet sich in den oberen 20 cm (HUSSE und STEINBERG, 1983), was sich
positiv auf das Mykorrhizierungspotential ausgewirkt haben könnte, weil ein intensiverer
Kontakt der Begrünungswurzeln mit den Rebwurzeln bestand. DOUDS et al. (1995) fanden
anhand eines Bioassays im Gewächshaus im oberen Bodenbereich ebenfalls ein signifikant
höheres Mykorrhizapotential. Von ähnlichen Ergebnissen berichteten NAPPI et al. (1985), die
in den oberen Bodenschichten (0 - 20 cm) von Weinbergsböden aus Piemonte/Italien die
höchste Sporenanzahl mit 144 Sporen/g Boden ermittelten, während sie in einer Tiefe ab 60 -
70 cm keine Mykorrhiza-Sporen fanden. Für das hohe Mykorrhizapotential im Oberboden
könnten auch P-tolerante Pilzspezies verantwortlich sein, die an die hohen P-Gehalte im Bo-
den angepaßt sind. Solche Befunde wurden ebenfalls von DOUDS und SCHENK (1990) und
MORIN et al. (1994) in Gefäßversuchen beschrieben. Auch in Freilandversuchen wurden P-
tolerante AM-Pilze beobachtet, die bei hohen P-Gehalten im Boden präsent sind (SYLVIA und
SCHENK, 1983) bzw. nach einer Freilandinokulation an Rotesche (Fraxinus pennsylvanica
Mush) (LAMAR und DAVEY, 1988) oder im Reisanbau (Oryza sativa L.) (SOLAIMAN und HI-
RATA, 1996 und 1997) sogar zu Wachstumssteigerungen führten.
Im Biotest mit Weißklee trat der Unterschied zwischen Cruciferenanbau und den anderen Be-
grünungsvarianten nur im Oberboden (0 - 25 cm) auf (s. Abb. 3.5). Dabei war der Weißklee
aus der Variante mit der Cruciferen-Wintergründüngung mit 9,7 % tendenziell am geringsten
infiziert. Dies deutet darauf hin, daß Weißklee ähnlich wie bei den Rebwurzeln mit einer ge-
ringeren Infektion reagiert. Der Weißklee im Boden aus der offenen Variante im oberen Be-
reich (0 - 25 und 25 - 50 cm) wurde relativ stark infiziert (17 %) und lag in etwa auf dem glei-
chen Niveau wie die Begrünungsvarianten mit Leguminosen und Gramineen (s. Abb. 3.5).
Der Infektionsgrad der Rebwurzeln gibt die momentane Situation der Mykorrhizierung an.

Kapitel 3 Vorkommen der AM in Weinbergsböden 46
Untersuchungen bezüglich des Sporenvorkommens im Boden könnten über das Infektionspo-
tential im Boden Auskunft geben. Es kann davon ausgegangen werden, daß die aufgelaufene
Unkrautflora in der Kontrolle über die Herbst- und Winterperiode das Mykorrhizierungspo-
tential positiv beeinflußt hat. Diese Wildflora könnte durch die Ausbildung von infektiösen
Strukturen im Boden (Sporen, Hyphen, infizierte Wurzeln) das Mykorrhizierungspotential
während der Winterperiode aufrechterhalten haben. Diese Annahme wird durch die Untersu-
chungen von SCHUBERT und CRAVERO (1985) bestätigt. Nach 9wöchiger Wachstumsphase
war der Weißklee im Gefäßversuch stärker infiziert als die Rebwurzeln im Freiland. Der
Weißklee könnte anders auf die autochthone Mykorrhiza im Boden reagieren als die Rebe.
GIOVANNETTI et al. (1988) fanden bei der Inokulation mit Glomus monosporum an den ver-
schiedenen Pflanzenarten Rebe (Sorte Nebbiolo), Zwiebel (Allium cepa), Rotklee (T. praten-
se) und Wiesenrispe (Poa pratensis) unterschiedliche Infektionsgrade und erklärten dies mit
einer unterschiedlichen Wirtsspezifität des Pilzes. Andererseits wird die Erde in Pflanzgefä-
ßen meistens intensiver durchwurzelt als im Freiland, so daß die Infektionswahrscheinlichkeit
für AM-Pilze und somit der Infektionsgrad in den Gefäßen erhöht wird.
Zusammenfassend läßt sich feststellen, daß die AM die Reben sowohl in Ertragsanlagen als
auch in Rebschulen besiedelt. Eine eindeutige Aussage über den Einfluß der Unterlagssorte
auf die Mykorrhizierung konnte aufgrund der vorliegenden Ergebnisse nicht getroffen werden.
In Ertragsanlagen können bestimmte Begrünungspflanzen wie Gramineen und Leguminosen
die autochthone Mykorrhiza der Rebe positiv beeinflussen. Von Nicht-Wirtspflanzen für die
AM, wie z. B. Ölrettich, Winterraps oder Gelbsenf geht in Ertragsanlagen ein hemmender
Einfluß auf die Mykorrhizierung der Reben aus. Die Praxis kann sich diese Effekte beispiels-
weise durch Einsaat eines Gräser-Leguminosen-Gemisches zunutze machen.

Kapitel 4 Praktische Anwendung der AM 47
4 Praktische Anwendung von arbuskulären Mykorrhizapilzen in Rebschulen und in der Topfrebenproduktion
4.1 Einleitung
Die Erzeugung von Rebenpflanzgut erfolgt in mehreren Schritten. Nach einer bestimmten
Vortreibphase im Gewächshaus werden die veredelten Holzstecklinge für eine Vegetationspe-
riode in den Rebschulen kultiviert und im darauffolgenden Jahr als fertiges Pflanzgut im
Weinberg eingesetzt. In Deutschland wurden 1997 auf einer Fläche von 200 Hektar etwa 24
Mio. Pfropfreben angebaut (amtliche Anerkennungsstellen für Rebenpflanzgut, pers. Mittei-
lung). Verkaufsfähiges Rebenpflanzgut muß mindestens drei gleichmäßig verteilte, gut entwi-
ckelte Wurzeln aufweisen (ANONYM, 1986). Voraussetzung für die optimale Rebenentwick-
lung während der Wurzelbildungsphase ist ein gut vorbereitetes Pflanzbeet in der Rebschule
und günstige Temperaturen und Niederschläge zur Pflanzzeit.
Durch Inokulation mit unterschiedlichen Mykorrhizastämmen konnten in Gefäßversuchen
enorme Wachstumssteigerungen bei Reben erzielt werden (SCHUBERT et al., 1988; SCHUBERT
et al., 1990; KARAGIANNIDIS et al., 1995). Im Freiland wurden bei anderen Pflanzenarten
durch Inokulationen mit arbuskulären Mykorrhizen, z. B. bei Mais (Zea mays L.), die Biomas-
seproduktion deutlich erhöht (BALTRUSCHAT, 1987 b). Durch die Herstellung von Mykorrhi-
za-Inokulum, welches an anorganische Trägerstoffe gebunden ist, besteht die Möglichkeit, die
AM-Pilze mit Hilfe von praxisgängigen Inokulationsmethoden in landwirtschaftlichen Kul-
turpflanzen einzusetzen (DEHNE und BACKHAUS, 1986; BALTRUSCHAT, 1987 a, b, c; GRUNE-
WALDT-STÖCKER und DEHNE, 1989).
In Kapitel 3 konnte gezeigt werden, daß die Wurzeln von Pfropfreben in allen untersuchten
Rebschulen mit AM infiziert waren. In einem weiteren Versuchsansatz sollte geprüft werden,
inwieweit positive Wachstumseffekte durch die Inokulation mit Glomus sp. in Rebschulen,
vergleichend zu nativ vorkommenden AM-Pilzen, bei Pfropfreben erreicht werden können.
Der Inokulationserfolg sollte neben den Reben zusätzlich an Indikatorpflanzen, die zwischen
den Pfropfreben in den Versuchsvarianten ausgesät wurden, bestimmt werden.

Kapitel 4 Praktische Anwendung der AM 48
Neben der Verwendung von Pfropfreben aus Rebschulen besteht für den Winzer die Möglich-
keit, bereits vorgetriebene Reben in den Weinberg zu pflanzen. Bei diesem sogenannten Topf-
reben- bzw. Kartonagerebenverfahren werden die Pfropfreben veredelt und nach dem Vortrei-
ben in Jiffy-Pots, Kartonagen oder Gittertöpfe kartoniert. Als Pflanzerde werden tonhaltige
Anzuchterden verwendet. In Gewächshäusern oder in Frühbeetkästen bildet sich während der
Anzuchtphase bereits ein Wurzelsystem mit einem mehr oder weniger stark ausgeprägten
Trieb. Die Topfreben können noch im gleichen Jahr an ihren endgültigen Standort im Wein-
berg gepflanzt werden. Somit ist der Weg über die Rebschule eingespart (VOGT und GÖTZ,
1987). Der Anteil der Topfreben an der Gesamtproduktion von Pfropfreben (etwa 2,5 %)
schwankt sehr stark von Jahr zu Jahr. 1997 wurden etwa 600.000 Topfreben hergestellt (amt-
liche Anerkennungsstellen für Rebenpflanzgut, pers. Mitteilung).
MENGE (1983) gibt an, daß der kommerzielle Einsatz von AM-Inokulum sich auf die Standor-
te und Einsatzbereiche beschränkt, auf denen die native AM-Population nur sehr gering oder
überhaupt nicht vorkommt, z. B. beim Einsatz von sterilisierten Substraten bei Gewächshaus-
kulturen oder in entseuchten Böden. Über präinokulierte Pflanzen mit AM-Pilzen gibt es in
der Literatur zahlreiche Hinweise (PLENCHETTE et al., 1981 an Apfel (Malus pumila Mill. cv.
Beautiful arcade), FERGUSON und MENGE, 1986 an Citrus (Citrus aurantium L.) sowie KRI-
KUN et al., 1987 an Paprika (Capsicum annuum cv. Maor)). An Reben wurden Inokulations-
versuche mit in-vitro-vermehrten Stecklingen durchgeführt (RAVOLANIRINA et al., 1989;
SCHUBERT et al., 1990 und SCHELLENBAUM et al., 1991). Während bei Zier- und Gemüse-
pflanzen bereits Berichte über zahlreiche Nutzeffekte wie Verkürzung der Kulturzeit oder
bessere Widerstandsfähigkeit gegenüber bodenbürtigen Krankheitserregern durch effektive
AM-Pilzstämme in gärtnerischen Substraten vorliegen (DEHNE, 1994; BACKHAUS und FELD-
MANN, 1996), sind über die Anwendung von AM-Pilzen in der Topfrebenproduktion bisher
keine Untersuchungen bekannt. Das Ziel des vorliegenden Versuches war es daher, durch eine
frühzeitige Inokulation der Kultursubstrate mit den AM-Pilzen Glomus sp. und Glomus mos-
seae die jungen Rebwurzeln vor der Anzuchtphase zu infizieren und das Wachstum der Topf-
reben sowohl während der Anzuchtphase im Gewächshaus als auch später im Jungfeld zu
beschleunigen. Dabei wurden zwei unterschiedliche, in der Praxis häufig verwendete Pflanz-
erden eingesetzt, um die Übertragbarkeit auf die Praxis zu erleichtern.

Kapitel 4 Praktische Anwendung der AM 49
4.2 Material und Methoden
4.2.1 Rebschulen
4.2.1.1 Versuchsstandorte, Pflanzenmaterial und Versuchsaufbau
Die vorliegenden Untersuchungen wurden auf zwei Versuchsflächen der Staatlichen Lehr-
und Forschungsanstalt (SLFA) in Neustadt durchgeführt. Die Versuchsfläche Standort „Hart-
häuser“ wurde seit 1974 als Rebschule genutzt, wobei diese Fläche nur alle drei Jahre mit
Reben bepflanzt wurde. In den dazwischenliegenden Jahren erfolgte mehrmals die Einsaat
von Ölrettich (Raphanus sativus oleiformis L.) als Zwischenfrucht. Auf dem Standort „Stein-
gebiß“ wurden 1995 erstmals Reben gepflanzt. Von 1990 bis 1994 wurden auf dieser Fläche
Getreide und Zuckerrüben angebaut. Der Boden der Versuchsfläche „Harthäuser“ war ein
sandiger Lehm, beim Boden des Standortes „Steingebiß“ handelte es sich um einen schluffig-
lehmigen Sandboden (s. Tab. 4.1). Als Pfropfreben wurden Riesling/5C (Standort „Harthäu-
ser“) bzw. Müller-Thurgau/5BB (Standort „Steingebiß“) gepflanzt. Um den Inokulationser-
folg an weiteren Pflanzenarten zu überprüfen, wurden auf dem Standort „Harthäuser“ für die
Bestimmung des Infektionsgrades die Indikatorpflanze Weißklee (Trifolium repens L., cv.
Huja) und auf der Versuchsfläche „Steingebiß“ Mais (Zea mays L., cv. Gelber Badischer
Landmais) ausgesät. Die Aussaat der Indikatorpflanzen erfolgte für jede Variante und Wie-
derholung auf einer Länge von einem Meter im Bereich der Mulchfolie.
Tab. 4.1: Chemische und physikalische Eigenschaften der Versuchsböden in den Rebschu-len „Harthäuser“ und „Steingebiß“
Standort Humus P K Mg pH Korngrößenbestimmung
(CAL) (CaCl2) (KCl) Ton Schluff Sand
(%) mg/100 g Boden (%)
Harthäuser 3,6 28,4 28,1 14,2 7,1 8,4 25,4 66,2
Steingebiß 1,8 12,1 14,5 7,7 7,5 15,9 41,4 42,7
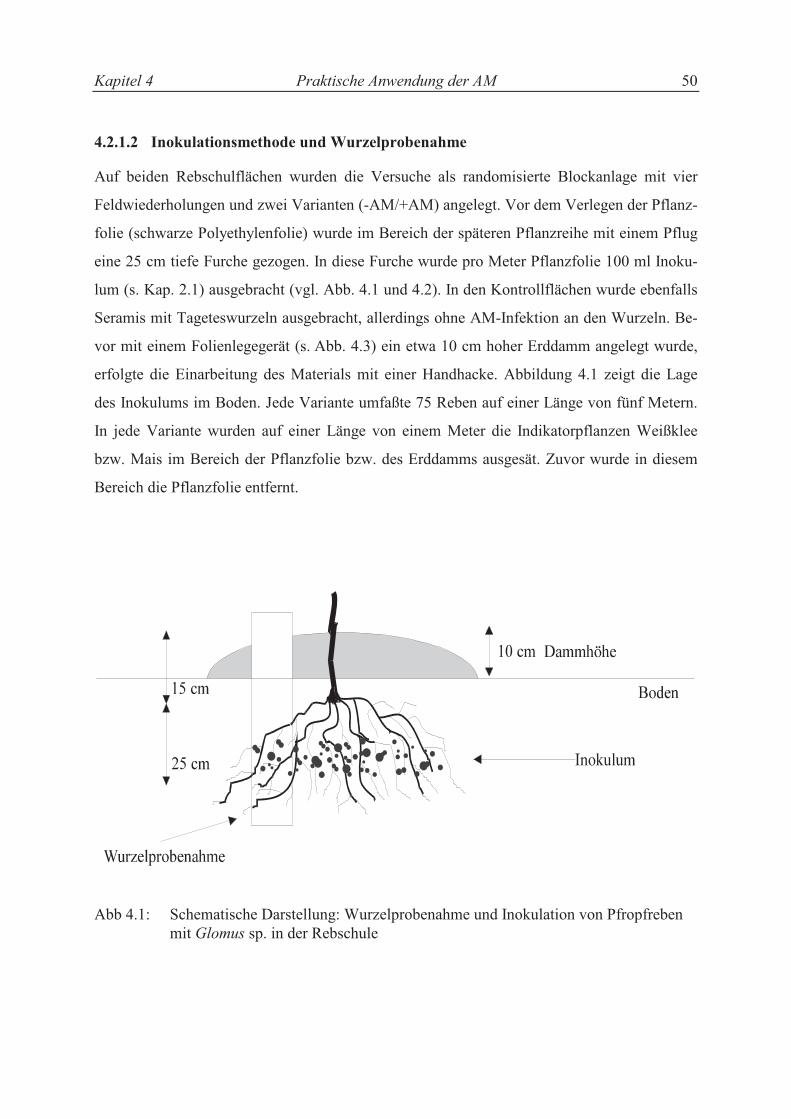
Kapitel 4 Praktische Anwendung der AM 50
4.2.1.2 Inokulationsmethode und Wurzelprobenahme
Auf beiden Rebschulflächen wurden die Versuche als randomisierte Blockanlage mit vier
Feldwiederholungen und zwei Varianten (-AM/+AM) angelegt. Vor dem Verlegen der Pflanz-
folie (schwarze Polyethylenfolie) wurde im Bereich der späteren Pflanzreihe mit einem Pflug
eine 25 cm tiefe Furche gezogen. In diese Furche wurde pro Meter Pflanzfolie 100 ml Inoku-
lum (s. Kap. 2.1) ausgebracht (vgl. Abb. 4.1 und 4.2). In den Kontrollflächen wurde ebenfalls
Seramis mit Tageteswurzeln ausgebracht, allerdings ohne AM-Infektion an den Wurzeln. Be-
vor mit einem Folienlegegerät (s. Abb. 4.3) ein etwa 10 cm hoher Erddamm angelegt wurde,
erfolgte die Einarbeitung des Materials mit einer Handhacke. Abbildung 4.1 zeigt die Lage
des Inokulums im Boden. Jede Variante umfaßte 75 Reben auf einer Länge von fünf Metern.
In jede Variante wurden auf einer Länge von einem Meter die Indikatorpflanzen Weißklee
bzw. Mais im Bereich der Pflanzfolie bzw. des Erddamms ausgesät. Zuvor wurde in diesem
Bereich die Pflanzfolie entfernt.
Abb 4.1: Schematische Darstellung: Wurzelprobenahme und Inokulation von Pfropfreben mit Glomus sp. in der Rebschule

Kapitel 4 Praktische Anwendung der AM 51
Abb. 4.2: Ausbringung von Mykorrhiza-Inokulum (Pfeil) in der Rebschule (Standort „Hart-häuser“); links: vor dem Verlegen der Mulchfolie; rechts: eingearbeitetes Inoku-lum mit Mulchfolie
Abb. 4.3: Folienlegegerät in der Rebschule (Standort „Harthäuser“)
Die Ausbringung des Mykorrhiza-Materials erfolgte am 07.05.1996. Die Pfropfreben wurden
am 14.05.1996 gesteckt. Weißklee bzw. Mais wurden am 05.06.1996 ausgesät. Nach dem
Austrieb der Reben wurden die Pflanzen auf einen Trieb gestellt. In 14tägigem Abstand wurde

Kapitel 4 Praktische Anwendung der AM 52
die Trieblänge von jeweils 20 Reben pro Variante bestimmt. Die Pflanzenschutz- und Dün-
gungsmaßnahmen erfolgten in allen Varianten einheitlich und gemäß den Empfeh-
lungen für die weinbauliche Praxis.
Die ersten Wurzelproben wurden am 19.06.1996 genommen, fünf Wochen nach dem Stecken
der Reben. Hierbei wurde mit einem Spaten im Bereich von 5 - 7 Reben an einer Seite der
Pflanzfolie 30 cm tief gegraben (vgl. Abb. 4.1), die Wurzeln entnommen und in angefeuchtete
Polyethylenbeutel überführt. Das Anfärben der Rebwurzeln sowie die Bestimmung des AM-
Infektionsgrades (s. Kap. 2.3 und 2.4) erfolgten im Labor. Um Angaben über den zeitlichen
Verlauf der Infektion zu erhalten, wurden an zwei weiteren Terminen (7 und 17 Wochen nach
dem Stecken der Reben) Wurzelproben entnommen. In gleicher Weise wurden die Wurzel-
proben von Mais ausgewertet. Die Probenahme von Weißklee erfolgte 11 Wochen nach Aus-
saat.
4.2.2 Topfrebenproduktion
4.2.2.1 Pflanzenmaterial, Versuchsgefäße und Kultursubstrat
Der Inokulationsversuch mit Topfreben wurde 1995 und 1997 in einem Foliengewächshaus
über einen Zeitraum von zwei Monaten bzw. einem Monat durchgeführt. Als Versuchspflan-
zen dienten in beiden Jahren veredelte und vorgetriebene Pfropfreben der Sorte Riesling
[Klon N90] auf 125 AA [Klon FR25]. Während sich der Versuch 1995 auf die Anzuchtphase
unter Gewächshausbedingungen beschränkte, wurde 1997 ein Teil der Topfreben ins Freiland
gepflanzt.
Als Pflanzgefäße wurden im Versuchsjahr 1995 Kartonagehüllen (Fa. Ehm, Trier) eingesetzt.
Hierbei handelte es sich um gelochte Kartonhüllen (Höhe 10 cm, Breite 4 cm) mit einem Vo-
lumen von 160 cm3. Für die Kultivierung im Gewächshaus wurde die tonhaltige Anzuchterde
Floraton 1 (Fa. Floragard, Oldenburg) verwendet (s. Tab. 4.2). Der Versuch wurde am
02.05.1995 angesetzt und nach einem bzw. zwei Monaten ausgewertet.
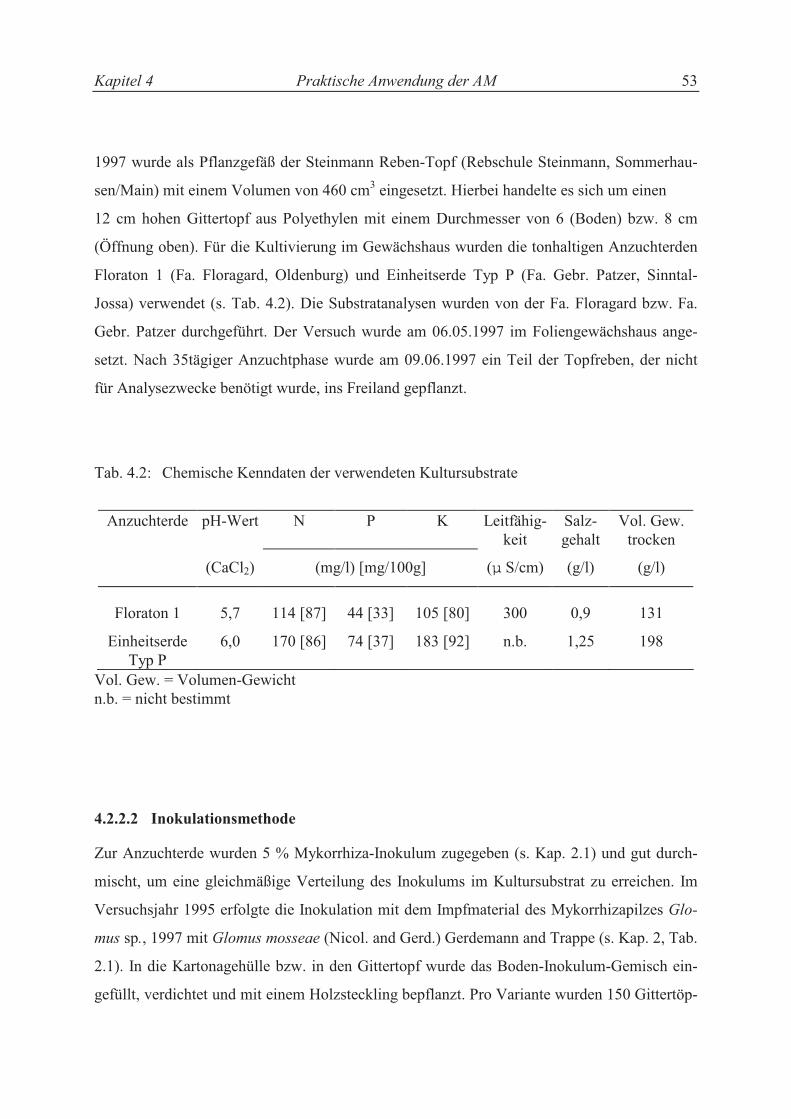
Kapitel 4 Praktische Anwendung der AM 53
1997 wurde als Pflanzgefäß der Steinmann Reben-Topf (Rebschule Steinmann, Sommerhau-
sen/Main) mit einem Volumen von 460 cm3 eingesetzt. Hierbei handelte es sich um einen
12 cm hohen Gittertopf aus Polyethylen mit einem Durchmesser von 6 (Boden) bzw. 8 cm
(Öffnung oben). Für die Kultivierung im Gewächshaus wurden die tonhaltigen Anzuchterden
Floraton 1 (Fa. Floragard, Oldenburg) und Einheitserde Typ P (Fa. Gebr. Patzer, Sinntal-
Jossa) verwendet (s. Tab. 4.2). Die Substratanalysen wurden von der Fa. Floragard bzw. Fa.
Gebr. Patzer durchgeführt. Der Versuch wurde am 06.05.1997 im Foliengewächshaus ange-
setzt. Nach 35tägiger Anzuchtphase wurde am 09.06.1997 ein Teil der Topfreben, der nicht
für Analysezwecke benötigt wurde, ins Freiland gepflanzt.
Tab. 4.2: Chemische Kenndaten der verwendeten Kultursubstrate
Anzuchterde pH-Wert N P K Leitfähig-keit
Salz-gehalt
Vol. Gew. trocken
(CaCl2) (mg/l) [mg/100g] (µ S/cm) (g/l) (g/l)
Floraton 1 5,7 114 [87] 44 [33] 105 [80] 300 0,9 131
Einheitserde Typ P
6,0 170 [86] 74 [37] 183 [92] n.b. 1,25 198
Vol. Gew. = Volumen-Gewicht n.b. = nicht bestimmt
4.2.2.2 Inokulationsmethode
Zur Anzuchterde wurden 5 % Mykorrhiza-Inokulum zugegeben (s. Kap. 2.1) und gut durch-
mischt, um eine gleichmäßige Verteilung des Inokulums im Kultursubstrat zu erreichen. Im
Versuchsjahr 1995 erfolgte die Inokulation mit dem Impfmaterial des Mykorrhizapilzes Glo-
mus sp., 1997 mit Glomus mosseae (Nicol. and Gerd.) Gerdemann and Trappe (s. Kap. 2, Tab.
2.1). In die Kartonagehülle bzw. in den Gittertopf wurde das Boden-Inokulum-Gemisch ein-
gefüllt, verdichtet und mit einem Holzsteckling bepflanzt. Pro Variante wurden 150 Gittertöp-

Kapitel 4 Praktische Anwendung der AM 54
fe bzw. Kartonagehüllen verwendet. Für die Kontrollen wurde Seramis mit Tagetes- bzw.
Weißkleewurzeln ohne Mykorrhizainfektion zugegeben.
4.2.2.3 Pflanzenanzucht, Pflanzenpflege und Ernte der Versuchspflanzen
Jede Versuchsvariante wurde separat in beheizte Beete in eine Torfschicht eingebettet. Wäh-
rend der Anzuchtphase betrug die Wurzelraumtemperatur etwa 24 °C. In der darauffolgenden
Abhärtungsphase wurden die Reben durch Senkung der Wurzelraumtemperatur auf 20 °C
langsam an die Außenbedingungen adaptiert.
Die Topfreben wurden je nach Bedarf mit Leitungswasser gegossen. Eine Düngung während
der Anzuchtphase erfolgte in beiden Versuchsjahren nicht. Gegen Echten Mehltau (Uncinula
necator (Schwein.) Burr.) wurde einmal Folicur E (0,25 %ig [Dichlofluanid + Tebuconazol])
eingesetzt. Nach dem Austrieb wurden die Reben auf einen Trieb gestellt. Aus jeder Variante
wurden stichprobenartig 20 Reben ausgewählt, an denen wöchentlich die Trieblänge bestimmt
wurde. Im Versuchsjahr 1995 wurden vier bzw. acht Wochen nach Versuchsbeginn an 10 Re-
ben pro Variante Wurzelproben entnommen und der AM-Infektionsgrad (s. Kap. 2.4) ermit-
telt. Für eine Wurzelprobe wurden aufgrund des sehr geringen Wurzelmaterials Wurzeln von
zwei Reben zu einer Mischprobe zusammengefaßt. Die Topfreben wurden vorsichtig aus dem
Beet entnommen und die Wurzeln unter fließendem Wasser vom Kultursubstrat befreit. Da
bereits zum ersten Erntezeitpunkt die Wurzeln die Kartonagehülle durchwachsen hatten, wur-
den für die Bestimmung des Infektionsgrades auch solche Wurzeln mit berücksichtigt, die
sich außerhalb des Pflanzgefäßes befanden.
Im Versuchsjahr 1997 wurden die Wurzeln 35 Tage nach Versuchsbeginn entnommen.
Gleichzeitig wurden von 20 Reben, die stichprobenartig aus jeder Variante ausgewählt wur-
den, das Frisch- und Trockengewicht der Blätter und Sprosse (mit Blattstielen) bestimmt. Au-
ßerdem wurden die Makronährstoffe P, K, Mg und Ca sowie die Mikronährstoffe Fe, Mn, Zn
und Cu in den Blättern von sechs Reben pro Variante quantitativ bestimmt (s. Kap. 2.6).

Kapitel 4 Praktische Anwendung der AM 55
4.2.2.4 Versuchsstandort (Boden, Witterungsdaten) und Versuchsanlage
Im Versuchsjahr 1997 wurden die vorkultivierten Topfreben nach 5wöchiger Anzuchtphase
im Foliengewächshaus ins Freiland in einen neu angelegten Weinberg gepflanzt. Der Ver-
suchsweinberg lag in der Gemarkung Ruppertsberg, Lage „Ruppertsberger Linsenbusch“ und
hatte eine Größe von 80 Ar. Auf der Parzelle standen zwei Rebzeilen von 200 m Länge zur
Verfügung, in denen der Versuch als randomisierte Blockanlage mit je vier Feldwiederho-
lungen angelegt wurde. Die Zeilenbreite betrug 2,0 m und der Stockabstand 1,1 m. In jeder
Versuchsvariante standen 20 Reben. Bei vier Versuchsvarianten (-AM/+AM mit jeweils zwei
unterschiedlichen Kultursubstraten) ergab dies insgesamt 320 Reben. An jeder Pflanzstelle
wurde mit einem Spaten ein 30 cm tiefes Loch ausgehoben, in die die Topfrebe gesetzt wurde.
Das Pflanzloch wurde soweit mit Boden aufgefüllt, bis sich die Veredelungsstelle oberhalb
der Bodenoberfläche befand. Der emporwachsende Trieb wurde am Pflanzpfahl (Bambusstab)
befestigt.
Die Bodenprobenahme für die Bodenanalysen erfolgte aus zwei Tiefen (0 - 30 und 30 - 60
cm) und wurde getrennt in Polyethylenbeutel gesammelt. Aus jeder Feldwiederholung wurde
eine Mischprobe von fünf Bohrstockeinstichen gewonnen. Beim Boden handelte es sich um
einen schwach (Feldwiederholung A), mittel (Feldwiederholung B/D) bzw. stark lehmigen
Sandboden (Feldwiederholung C) (s. Tab. 4.3). Die Bodenanalysen wurden vom Fachbereich
Bodenkunde und Bodenschutz der Staatlichen Lehr- und Versuchsanstalt in Trier durchge-
führt. In der Feldwiederholung C traten in jeder Versuchsvariante einheitlich Wachstumsde-
pressionen auf, so daß bei der Verrechnung der Versuchsdaten die Ergebnisse aus diesem Ab-
schnitt eliminiert wurden und mit drei Feldwiederholungen gerechnet wurde.
Tab. 4.3: Chemische und physikalische Eigenschaften des Versuchsstandortes („Rupperts-berger Linsenbusch“), Boden aus 0 - 30 cm Tiefe
Feldwieder- holung
Humus P K Mg pH Korngrößenbestim-
mung
(CAL) CaCl
2
(KCl) Ton Schluff Sand
Kapitel 4 Praktische Anwendung der AM 56
(%) mg/100 g Boden (%)
A 1,4 12,2 20,8 12,0 7,0 7,9 18,4 73,7
B 1,7 16,6 26,6 14,0 6,9 11,3 23,8 64,8
C 2,1 29,2 29,2 17,0 7,0 14,4 28,3 57,2
D 1,4 20,1 20,0 14,0 6,8 9,0 31,5 59,4
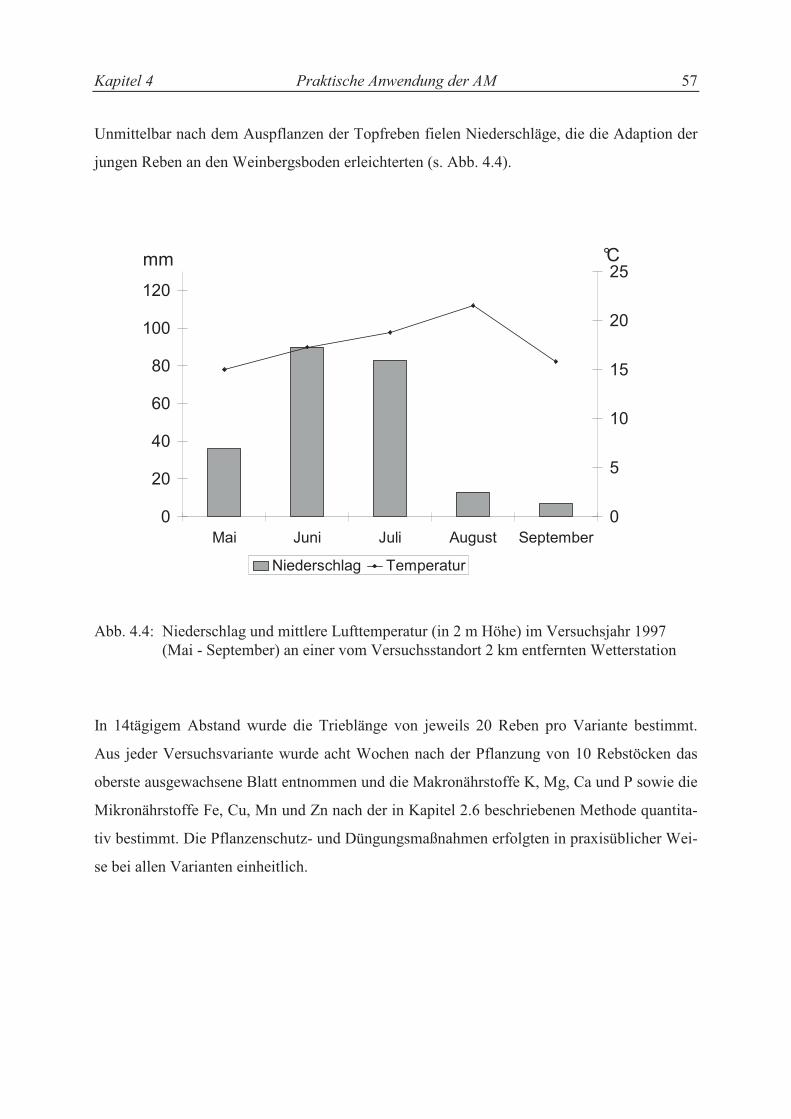
Kapitel 4 Praktische Anwendung der AM 57
Unmittelbar nach dem Auspflanzen der Topfreben fielen Niederschläge, die die Adaption der
jungen Reben an den Weinbergsboden erleichterten (s. Abb. 4.4).
0
20
40
60
80
100
120
Mai Juni Juli August September
mm
0
5
10
15
20
25°C
Niederschlag Temperatur
Abb. 4.4: Niederschlag und mittlere Lufttemperatur (in 2 m Höhe) im Versuchsjahr 1997 (Mai - September) an einer vom Versuchsstandort 2 km entfernten Wetterstation
In 14tägigem Abstand wurde die Trieblänge von jeweils 20 Reben pro Variante bestimmt.
Aus jeder Versuchsvariante wurde acht Wochen nach der Pflanzung von 10 Rebstöcken das
oberste ausgewachsene Blatt entnommen und die Makronährstoffe K, Mg, Ca und P sowie die
Mikronährstoffe Fe, Cu, Mn und Zn nach der in Kapitel 2.6 beschriebenen Methode quantita-
tiv bestimmt. Die Pflanzenschutz- und Düngungsmaßnahmen erfolgten in praxisüblicher Wei-
se bei allen Varianten einheitlich.

Kapitel 4 Praktische Anwendung der AM 58
4.3 Ergebnisse
4.3.1 Rebschulen
4.3.1.1 Verlauf der AM-Infektion an Reb- und Weißkleewurzeln (Standort „Harthäuser“)
Bei der ersten Wurzelprobenahme war die Verteilung des Inokulums im Wurzelbereich der
Pfropfreben sehr deutlich (vgl. Abb. 4.5). Die Reben zeigten fünf Wochen nach dem Aus-
pflanzen sowohl in den Kontrollen (< 1 %) als auch in den inokulierten Varianten (3 %) nur
sehr geringe Infektionsgrade (s. Abb. 4.6). Ein Einfluß der Inokulation mit Glomus sp. war
nach fünf Wochen tendenziell erkennbar. Nach weiteren zwei Wochen hatte die Mykorrhizie-
rung in den Rebwurzeln der nicht inokulierten Varianten, die durch die natürliche Mykorrhi-
zapopulation vom Standort hervorgerufen wurde, geringfügig zugenommen (2 % Infektions-
grad). Dagegen war durch die Inokulation mit Glomus sp. der Infektionsgrad auf etwa 11 %
angestiegen. Dieser Unterschied konnte statistisch abgesichert werden (vgl. Abb. 4.6). Bis
zum Ende des Versuches nach 17 Wochen blieb der Infektionsgrad an den Kontrollreben un-
verändert. Der Infektionsgrad in den mit AM inokulierten Varianten stieg bis Versuchsende
geringfügig auf
12 % an.
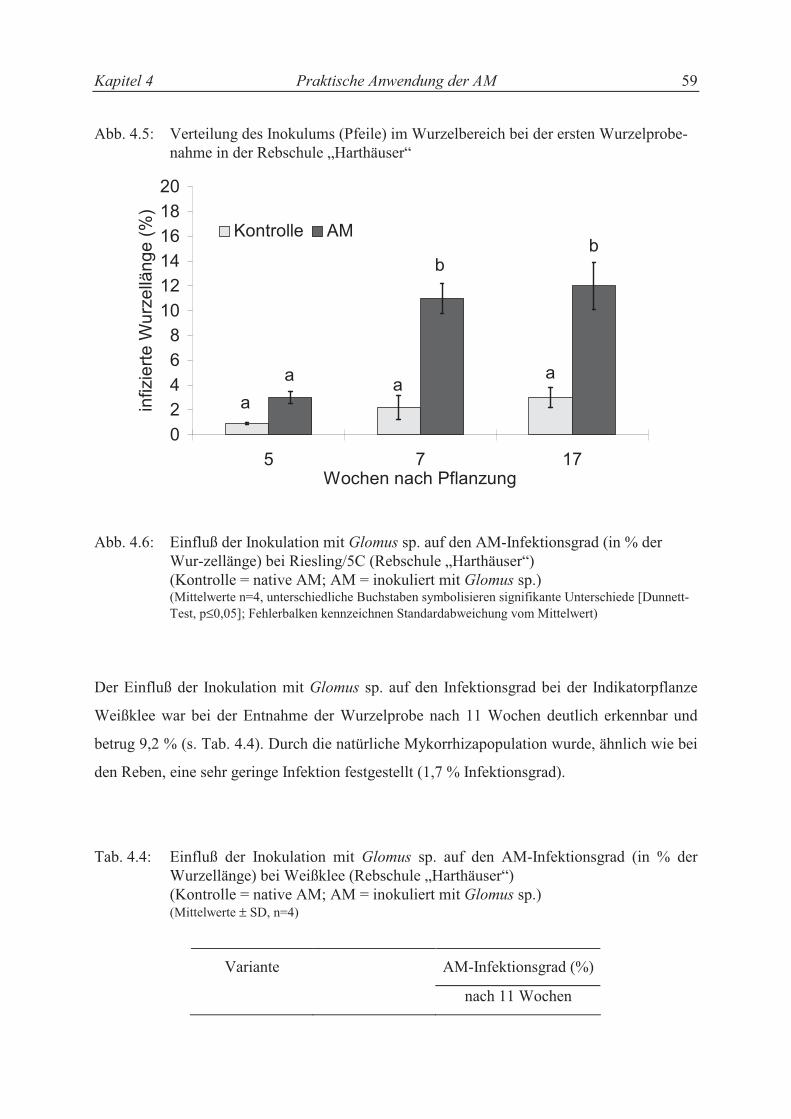
Kapitel 4 Praktische Anwendung der AM 59
Abb. 4.5: Verteilung des Inokulums (Pfeile) im Wurzelbereich bei der ersten Wurzelprobe-nahme in der Rebschule „Harthäuser“
aa
aa
bb
0
2
4
6
8
10
12
14
16
18
20
5 7 17
infizie
rte
Wu
rze
llän
ge
(%
)
Kontrolle AM
Wochen nach Pflanzung
Abb. 4.6: Einfluß der Inokulation mit Glomus sp. auf den AM-Infektionsgrad (in % der Wur-zellänge) bei Riesling/5C (Rebschule „Harthäuser“)
(Kontrolle = native AM; AM = inokuliert mit Glomus sp.) (Mittelwerte n=4, unterschiedliche Buchstaben symbolisieren signifikante Unterschiede [Dunnett-
Test, p≤0,05]; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
Der Einfluß der Inokulation mit Glomus sp. auf den Infektionsgrad bei der Indikatorpflanze
Weißklee war bei der Entnahme der Wurzelprobe nach 11 Wochen deutlich erkennbar und
betrug 9,2 % (s. Tab. 4.4). Durch die natürliche Mykorrhizapopulation wurde, ähnlich wie bei
den Reben, eine sehr geringe Infektion festgestellt (1,7 % Infektionsgrad).
Tab. 4.4: Einfluß der Inokulation mit Glomus sp. auf den AM-Infektionsgrad (in % der Wurzellänge) bei Weißklee (Rebschule „Harthäuser“)
(Kontrolle = native AM; AM = inokuliert mit Glomus sp.) (Mittelwerte ± SD, n=4)
Variante AM-Infektionsgrad (%)
nach 11 Wochen

Kapitel 4 Praktische Anwendung der AM 60
Kontrolle 1,7 ± 0,4 AM 9,2 ± 2,1
4.3.1.2 Verlauf der AM-Infektion an Reb- und Maiswurzeln (Standort „Steingebiß“)
Nach 5wöchiger Wachstumsphase betrug der AM-Infektionsgrad an den Reben in den Kon-
trollen 1,5 % und in den inokulierten Varianten 8 % (s. Abb. 4.7, oben). Im Vergleich zur
Rebschule „Harthäuser“ machte sich in der Rebschule „Steingebiß“ der Einfluß der Inokulati-
on auf die Infektion der Rebwurzeln früher bemerkbar. Die Unterschiede zwischen Kontrolle
und Inokulation waren zu diesem Zeitpunkt aber nicht statistisch abgesichert (Abb. 4.7). Ge-
gen Ende der Versuchsphase (Wurzelprobenahme nach 17 Wochen) war die Mykorrhizierung
in den Rebwurzeln durch die natürliche Mykorrhizapopulation auf 5 % angestiegen. Durch die
Inokulation mit Glomus sp. konnte der Infektionsgrad auf 17 % erhöht werden.
In Abbildung 4.7 (unten) ist der Einfluß der Inokulation mit Glomus sp. auf den Infektions-
grad bei Mais dargestellt. Die Kontrollen zeigten zu allen Zeitpunkten höhere Infektionsgrade
als die Kontrollen der Reben. Nach 17wöchigem Wachstum lag der Infektionsgrad bei 8 %.
Auch hier machte sich die Inokulation mit Glomus sp. bemerkbar. Nach sieben Wochen waren
die Unterschiede statistisch abgesichert und nach 17 Wochen betrug der Infektionsgrad an
Mais
14 % und war damit geringer als bei den inokulierten Reben (17 %, vgl. Abb. 4.7).
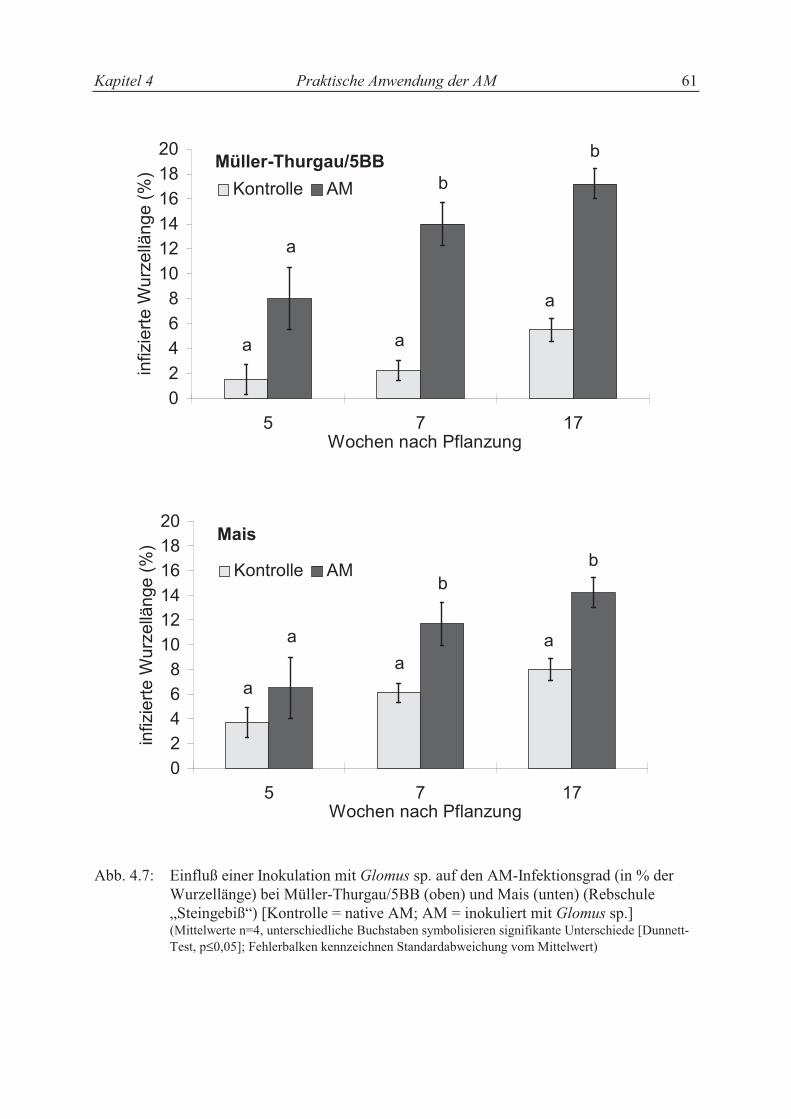
Kapitel 4 Praktische Anwendung der AM 61
a a
a
a
b
b
0
2
4
6
8
10
12
14
16
18
20
5 7 17
infizie
rte
Wu
rze
llän
ge
(%
)
Kontrolle AM
Wochen nach Pflanzung
Müller-Thurgau/5BB
a
a
aa
b
b
0
2
4
6
8
10
12
14
16
18
20
5 7 17
infizie
rte W
urz
ellä
nge (
%)
Kontrolle AM
Wochen nach Pflanzung
Mais
Abb. 4.7: Einfluß einer Inokulation mit Glomus sp. auf den AM-Infektionsgrad (in % der Wurzellänge) bei Müller-Thurgau/5BB (oben) und Mais (unten) (Rebschule „Steingebiß“) [Kontrolle = native AM; AM = inokuliert mit Glomus sp.]
(Mittelwerte n=4, unterschiedliche Buchstaben symbolisieren signifikante Unterschiede [Dunnett-Test, p≤0,05]; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
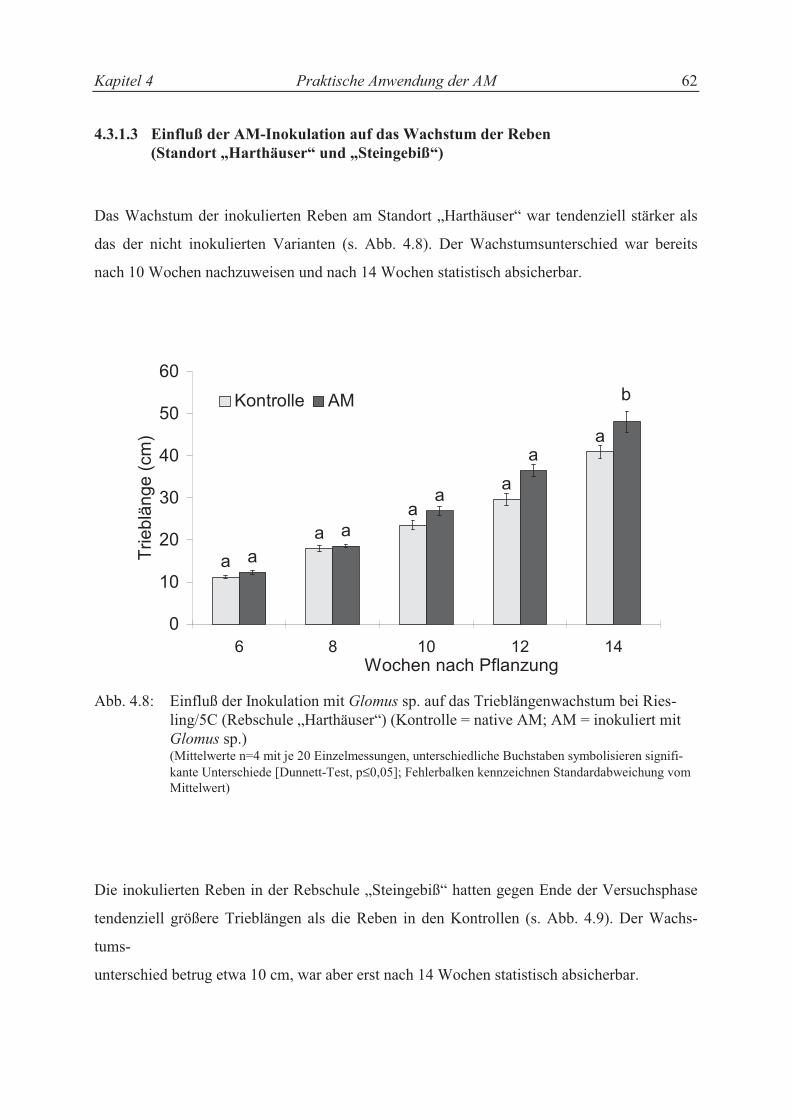
Kapitel 4 Praktische Anwendung der AM 62
4.3.1.3 Einfluß der AM-Inokulation auf das Wachstum der Reben (Standort „Harthäuser“ und „Steingebiß“)
Das Wachstum der inokulierten Reben am Standort „Harthäuser“ war tendenziell stärker als
das der nicht inokulierten Varianten (s. Abb. 4.8). Der Wachstumsunterschied war bereits
nach 10 Wochen nachzuweisen und nach 14 Wochen statistisch absicherbar.
a
a
a
a
a
a
a
a
a
b
0
10
20
30
40
50
60
6 8 10 12 14
Tri
eb
län
ge
(cm
)
Kontrolle AM
Wochen nach Pflanzung
Abb. 4.8: Einfluß der Inokulation mit Glomus sp. auf das Trieblängenwachstum bei Ries-ling/5C (Rebschule „Harthäuser“) (Kontrolle = native AM; AM = inokuliert mit Glomus sp.)
(Mittelwerte n=4 mit je 20 Einzelmessungen, unterschiedliche Buchstaben symbolisieren signifi-kante Unterschiede [Dunnett-Test, p≤0,05]; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
Die inokulierten Reben in der Rebschule „Steingebiß“ hatten gegen Ende der Versuchsphase
tendenziell größere Trieblängen als die Reben in den Kontrollen (s. Abb. 4.9). Der Wachs-
tums-
unterschied betrug etwa 10 cm, war aber erst nach 14 Wochen statistisch absicherbar.
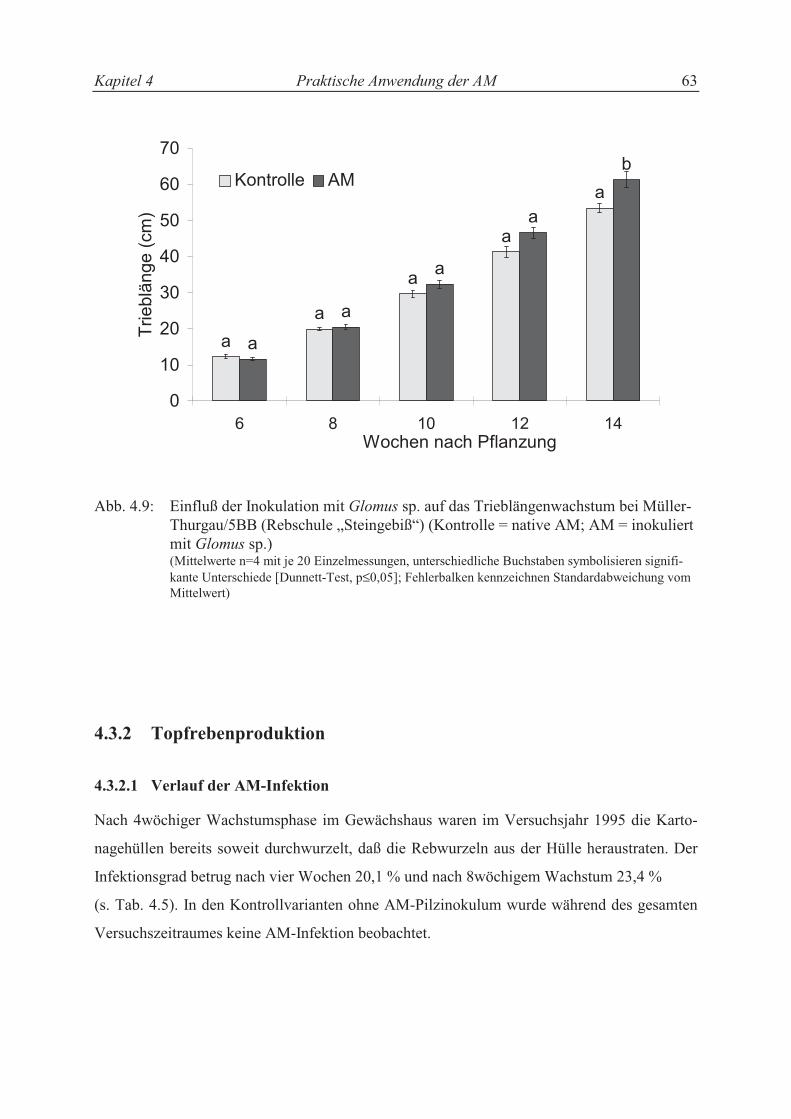
Kapitel 4 Praktische Anwendung der AM 63
a
a
a
a
a
a
a
a
a
b
0
10
20
30
40
50
60
70
6 8 10 12 14
Tri
eb
län
ge
(cm
)
Kontrolle AM
Wochen nach Pflanzung
Abb. 4.9: Einfluß der Inokulation mit Glomus sp. auf das Trieblängenwachstum bei Müller-Thurgau/5BB (Rebschule „Steingebiß“) (Kontrolle = native AM; AM = inokuliert mit Glomus sp.)
(Mittelwerte n=4 mit je 20 Einzelmessungen, unterschiedliche Buchstaben symbolisieren signifi-kante Unterschiede [Dunnett-Test, p≤0,05]; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
4.3.2 Topfrebenproduktion
4.3.2.1 Verlauf der AM-Infektion
Nach 4wöchiger Wachstumsphase im Gewächshaus waren im Versuchsjahr 1995 die Karto-
nagehüllen bereits soweit durchwurzelt, daß die Rebwurzeln aus der Hülle heraustraten. Der
Infektionsgrad betrug nach vier Wochen 20,1 % und nach 8wöchigem Wachstum 23,4 %
(s. Tab. 4.5). In den Kontrollvarianten ohne AM-Pilzinokulum wurde während des gesamten
Versuchszeitraumes keine AM-Infektion beobachtet.

Kapitel 4 Praktische Anwendung der AM 64
Tab 4.5: Einfluß einer Beimpfung mit dem AM-Pilz Glomus sp. auf den AM-Infektionsgrad (infizierte Wurzellänge %) von vier und acht Wochen alten Topf-reben (Riesling/125 AA) im Foliengewächshaus; Bodensubstrat Floraton 1;
Versuchsdauer 02.05. - 05.07.1995 Mittelwerte ± SD, n=10
Variante AM-Infektionsgrad (%)
nach 4 Wochen nach 8 Wochen
-AM 0 0 +AM 20,1 ± 2,1 23,4 ± 3,7
Im Versuchsjahr 1997 betrug der AM-Infektionsgrad nach 35tägiger Anzuchtphase im Folien-
gewächshaus im Bodensubstrat Floraton 1 18,3 % und im Substrat Einheitserde Typ P
16,3 % (s. Tab. 4.6). Die Mykorrhizierung der Topfreben war in beiden Bodensubstraten ver-
gleichbar. Ähnlich wie im Versuchsjahr 1995 waren die Pflanzgefäße zum Zeitpunkt der Ern-
te bereits intensiv durchwurzelt. In den Kontrollvarianten ohne AM-Inokulum wurde keine
AM-Infektion festgestellt.
Tab 4.6: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf den AM-Infektionsgrad (infizierte Wurzellänge %) von vier Wochen alten Topfreben (Riesling/125 AA) im Foliengewächshaus mit zwei unterschiedlichen Bodensub-straten (Floraton 1 [Floraton] und Patzer Einheitserde Typ P [Patzer])
Versuchsdauer 06.05. - 09.06.1997 Mittelwerte ± SD, n=10
Bodensubstrat Behandlung AM-Infektionsgrad (%)
nach 4 Wochen
„Floraton“ -AM 0 +AM 18,3 ± 2,1
„Patzer“ -AM 0 +AM 16,3 ± 2,3
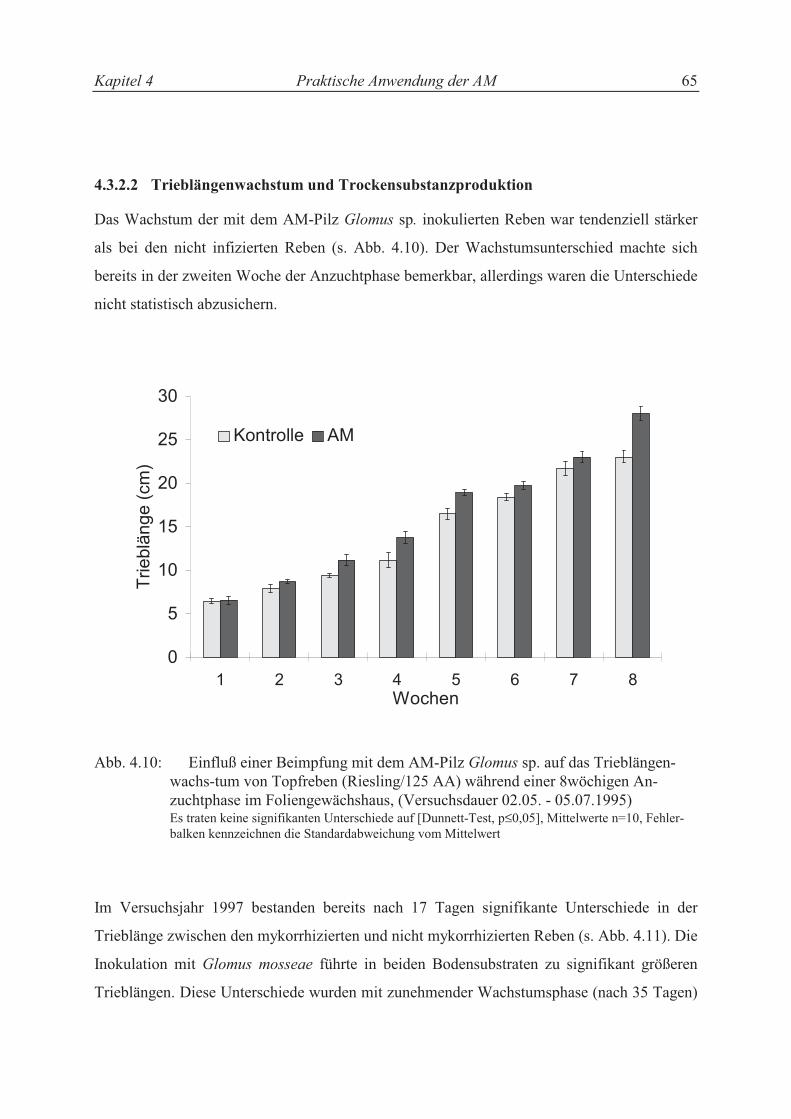
Kapitel 4 Praktische Anwendung der AM 65
4.3.2.2 Trieblängenwachstum und Trockensubstanzproduktion
Das Wachstum der mit dem AM-Pilz Glomus sp. inokulierten Reben war tendenziell stärker
als bei den nicht infizierten Reben (s. Abb. 4.10). Der Wachstumsunterschied machte sich
bereits in der zweiten Woche der Anzuchtphase bemerkbar, allerdings waren die Unterschiede
nicht statistisch abzusichern.
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8
Trieblä
nge (
cm
)
Kontrolle AM
Wochen
Abb. 4.10: Einfluß einer Beimpfung mit dem AM-Pilz Glomus sp. auf das Trieblängen-wachs-tum von Topfreben (Riesling/125 AA) während einer 8wöchigen An-zuchtphase im Foliengewächshaus, (Versuchsdauer 02.05. - 05.07.1995)
Es traten keine signifikanten Unterschiede auf [Dunnett-Test, p≤0,05], Mittelwerte n=10, Fehler-balken kennzeichnen die Standardabweichung vom Mittelwert
Im Versuchsjahr 1997 bestanden bereits nach 17 Tagen signifikante Unterschiede in der
Trieblänge zwischen den mykorrhizierten und nicht mykorrhizierten Reben (s. Abb. 4.11). Die
Inokulation mit Glomus mosseae führte in beiden Bodensubstraten zu signifikant größeren
Trieblängen. Diese Unterschiede wurden mit zunehmender Wachstumsphase (nach 35 Tagen)
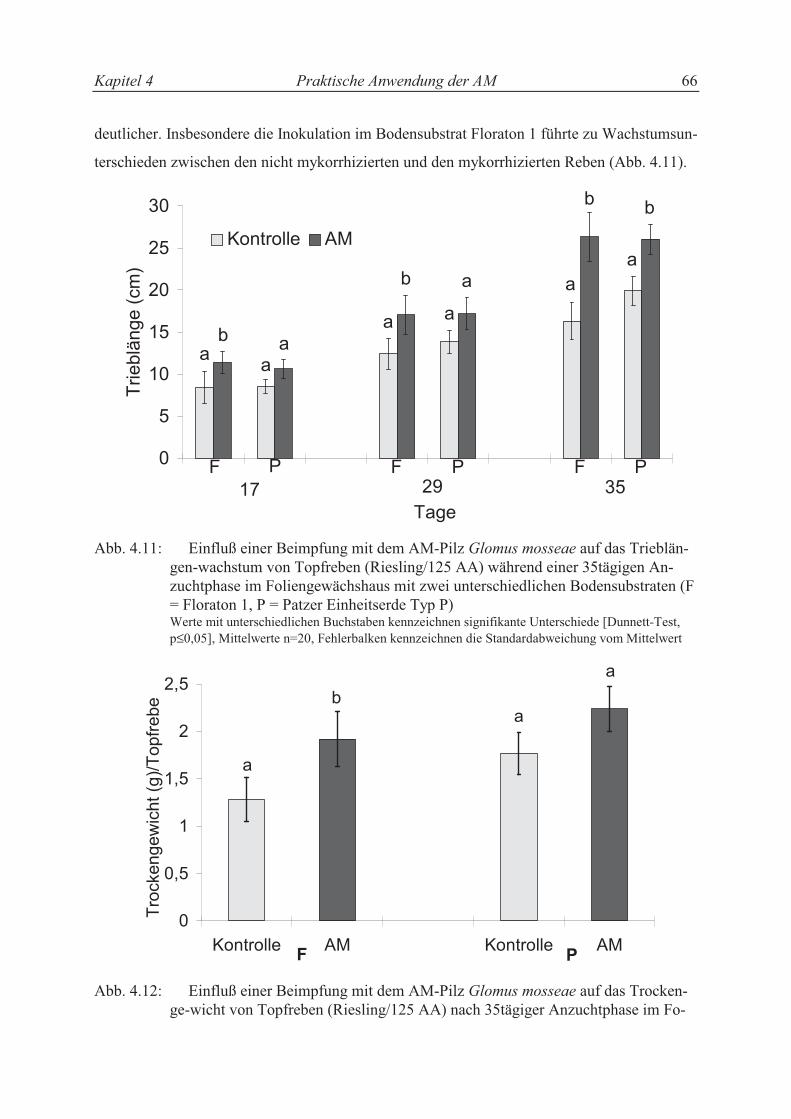
Kapitel 4 Praktische Anwendung der AM 66
deutlicher. Insbesondere die Inokulation im Bodensubstrat Floraton 1 führte zu Wachstumsun-
terschieden zwischen den nicht mykorrhizierten und den mykorrhizierten Reben (Abb. 4.11).
aa
aa
a
ab
ab
a
bb
0
5
10
15
20
25
30
Tri
eblä
nge (
cm
)
Kontrolle AM
Tage
F F FP P P
17 29 35
Abb. 4.11: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf das Trieblän-gen-wachstum von Topfreben (Riesling/125 AA) während einer 35tägigen An-zuchtphase im Foliengewächshaus mit zwei unterschiedlichen Bodensubstraten (F = Floraton 1, P = Patzer Einheitserde Typ P)
Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test, p≤0,05], Mittelwerte n=20, Fehlerbalken kennzeichnen die Standardabweichung vom Mittelwert
a
a
ba
0
0,5
1
1,5
2
2,5
Kontrolle AM Kontrolle AM
Tro
ckengew
icht (g
)/T
opfr
ebe
F P
Abb. 4.12: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf das Trocken-ge-wicht von Topfreben (Riesling/125 AA) nach 35tägiger Anzuchtphase im Fo-

Kapitel 4 Praktische Anwendung der AM 67
liengewächshaus mit zwei unterschiedlichen Bodensubstraten (F=Floraton 1, P=Patzer Einheitserde Typ P)
Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test, p≤0,05], Mittelwerte n=20, Fehlerbalken kennzeichnen die Standardabweichung vom Mittelwert
In Abbildung 4.12 ist der Einfluß der Beimpfung mit dem Mykorrhizapilz Glomus mosseae
auf das Trockengewicht und in Abbildung 4.13 auf den Sproßzustand der Topfreben nach
35tägiger Anzuchtphase dargestellt. Wie erwartet war das Trockengewicht in den beimpften
Varianten bei beiden Bodensubstraten höher als in den unbeimpften Varianten. Die Unter-
schiede waren in den mit Floraton 1 verwendeten Varianten statistisch abzusichern.
Abb. 4.13: Sproßzustand von Topfreben (Riesling/125AA) nach 35tägiger Anzuchtphase im Foliengewächshaus nach Inokulation mit dem AM-Pilz Glomus mosseae (Bo-densubstrat Floraton 1)
Kapitel 4 Praktische Anwendung der AM 68
(links: ohne Mykorrhiza [-AM]; rechts: mit Mykorrhiza [+AM])
4.3.2.3 Mineralstoffgehalte in den Blättern (Anzucht)
In den Tabellen 4.7 bzw. 4.8 sind die Mikro- und Makronährstoffgehalte in den Blattspreiten
der mit Glomus mosseae beimpften und unbeimpften Topfreben wiedergegeben. Ein deutli-
cher Effekt der Mykorrhizainokulation auf die Cu-Gehalte trat in beiden Bodensubstraten in
gleichem Maße auf und führte bei den mykorrhizierten Pflanzen zu signifikant höheren Cu-
Gehalten in den Blattspreiten. Die Zn-Gehalte in den Blättern waren bei beiden Bodensubstra-
ten in den beimpften Varianten höher als in den unbeimpften Varianten. Die Unterschiede
waren nur im Bodensubstrat Floraton statistisch abzusichern. Die Mn- und Fe-Gehalte der
Blätter waren durch die Mykorrhizierung verringert und im Bodensubstrat Patzer (Mn-Gehalt)
bzw. Floraton (Fe-Gehalt) im Vergleich zur unbeimpften Variante signifikant unterschiedlich.
Tab. 4.7: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf die Mikronähr-stoffgehalte in Blättern von Topfreben (Riesling/125 AA) nach 4wöchiger An-zuchtphase im Foliengewächshaus (Bodensubstrate Floraton 1 [Floraton] und Pat-zer Einheitserde Typ P [Patzer])
(Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test, p≤0,05], Mittelwerte ± SD, n=6)
Bodensubstrat Behand-lung
Cu Zn Fe Mn
(mg/kg TS)
„Floraton“ - AM 8,8 ± 1,1 a 37,0 ± 5,0 a 206 ± 19,4 a 157 ± 11,5 a + AM 10,7 ± 0,7 b 46,5 ± 5,5 b 143 ± 22,9 b 136 ± 26,8 a
„Patzer“ - AM 7,5 ± 2,1 a 31,8 ± 3,0 a 156 ± 23,4 a 178 ± 23,5 a + AM 9,0 ± 1,2 b 35,5 ± 4,2 a 135 ± 16,2 a 125 ± 10,4 b
Der P-Gehalt in den Blattspreiten lag gemittelt über beide Bodensubstrate mit 2,9 mg P/g Tro-
ckensubstanz (TS) in den mykorrhizierten Reben deutlich über den Werten der nicht my-

Kapitel 4 Praktische Anwendung der AM 69
korrhizierten Reben (2,5 mg P/g TS) (s. Tab. 4.8). Die Ca-Gehalte wurden durch die Inokula-
tion mit dem AM-Pilz nicht beeinflußt. Bei den Nährstoffgehalten Mg und K traten zwischen
den nicht infizierten und infizierten Reben in beiden Bodensubstraten keine signifikanten Un-
terschiede auf.
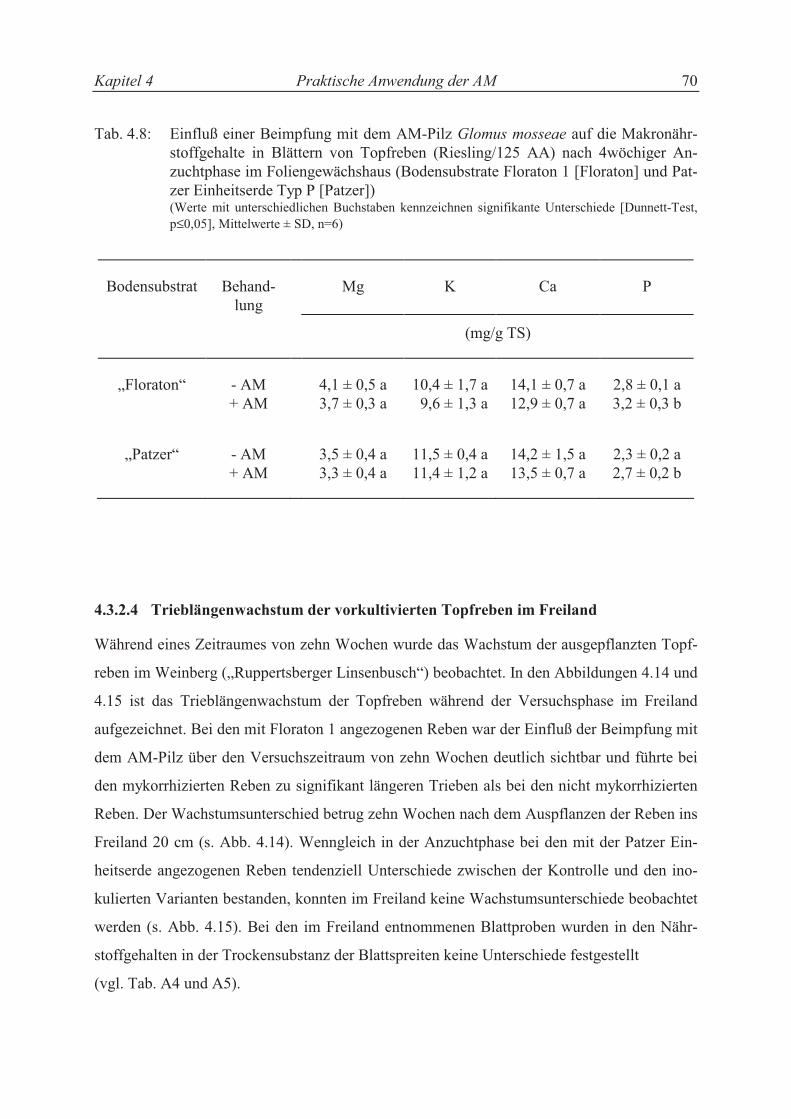
Kapitel 4 Praktische Anwendung der AM 70
Tab. 4.8: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf die Makronähr-stoffgehalte in Blättern von Topfreben (Riesling/125 AA) nach 4wöchiger An-zuchtphase im Foliengewächshaus (Bodensubstrate Floraton 1 [Floraton] und Pat-zer Einheitserde Typ P [Patzer])
(Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test, p≤0,05], Mittelwerte ± SD, n=6)
Bodensubstrat Behand-lung
Mg K Ca P
(mg/g TS)
„Floraton“ - AM 4,1 ± 0,5 a 10,4 ± 1,7 a 14,1 ± 0,7 a 2,8 ± 0,1 a + AM 3,7 ± 0,3 a 9,6 ± 1,3 a 12,9 ± 0,7 a 3,2 ± 0,3 b
„Patzer“ - AM 3,5 ± 0,4 a 11,5 ± 0,4 a 14,2 ± 1,5 a 2,3 ± 0,2 a + AM 3,3 ± 0,4 a 11,4 ± 1,2 a 13,5 ± 0,7 a 2,7 ± 0,2 b
4.3.2.4 Trieblängenwachstum der vorkultivierten Topfreben im Freiland
Während eines Zeitraumes von zehn Wochen wurde das Wachstum der ausgepflanzten Topf-
reben im Weinberg („Ruppertsberger Linsenbusch“) beobachtet. In den Abbildungen 4.14 und
4.15 ist das Trieblängenwachstum der Topfreben während der Versuchsphase im Freiland
aufgezeichnet. Bei den mit Floraton 1 angezogenen Reben war der Einfluß der Beimpfung mit
dem AM-Pilz über den Versuchszeitraum von zehn Wochen deutlich sichtbar und führte bei
den mykorrhizierten Reben zu signifikant längeren Trieben als bei den nicht mykorrhizierten
Reben. Der Wachstumsunterschied betrug zehn Wochen nach dem Auspflanzen der Reben ins
Freiland 20 cm (s. Abb. 4.14). Wenngleich in der Anzuchtphase bei den mit der Patzer Ein-
heitserde angezogenen Reben tendenziell Unterschiede zwischen der Kontrolle und den ino-
kulierten Varianten bestanden, konnten im Freiland keine Wachstumsunterschiede beobachtet
werden (s. Abb. 4.15). Bei den im Freiland entnommenen Blattproben wurden in den Nähr-
stoffgehalten in der Trockensubstanz der Blattspreiten keine Unterschiede festgestellt
(vgl. Tab. A4 und A5).
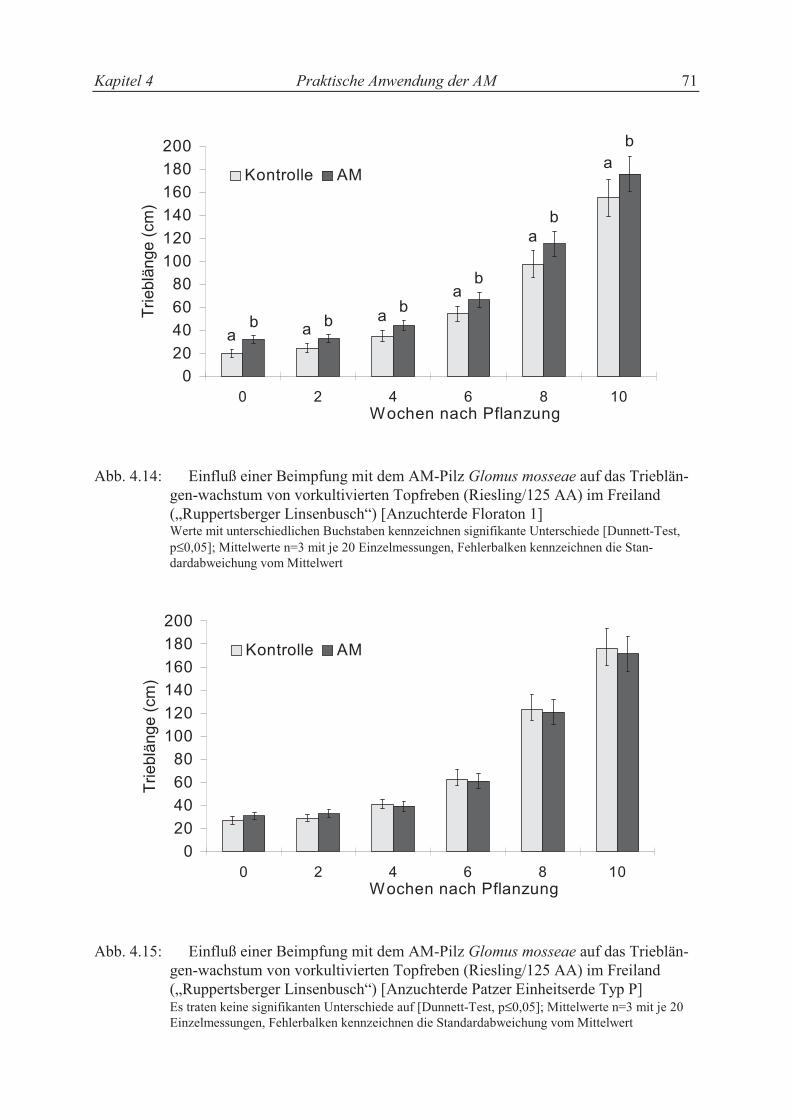
Kapitel 4 Praktische Anwendung der AM 71
a
a
aaa
a
b
bbb
b
b
0
20
40
60
80
100
120
140
160
180
200
0 2 4 6 8 10
Tri
eblä
nge (
cm
)
Kontrolle AM
Wochen nach Pflanzung
Abb. 4.14: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf das Trieblän-gen-wachstum von vorkultivierten Topfreben (Riesling/125 AA) im Freiland („Ruppertsberger Linsenbusch“) [Anzuchterde Floraton 1]
Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test, p≤0,05]; Mittelwerte n=3 mit je 20 Einzelmessungen, Fehlerbalken kennzeichnen die Stan- dardabweichung vom Mittelwert
0
20
40
60
80
100
120
140
160
180
200
0 2 4 6 8 10
Tri
eblä
nge (
cm
)
Kontrolle AM
Wochen nach Pflanzung
Abb. 4.15: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf das Trieblän-gen-wachstum von vorkultivierten Topfreben (Riesling/125 AA) im Freiland („Ruppertsberger Linsenbusch“) [Anzuchterde Patzer Einheitserde Typ P]
Es traten keine signifikanten Unterschiede auf [Dunnett-Test, p≤0,05]; Mittelwerte n=3 mit je 20 Einzelmessungen, Fehlerbalken kennzeichnen die Standardabweichung vom Mittelwert

Kapitel 4 Praktische Anwendung der AM 72
4.4 Diskussion
4.4.1 Einfluß der AM in Rebschulen
In der vorliegenden Untersuchung wurde überprüft, inwieweit Freilandinokulationen mit ar-
buskulären Mykorrhizapilzen das Wachstum junger Pfropfreben beeinflussen. Die Versuche
wurden an den Standorten „Harthäuser“ und „Steingebiß“ durchgeführt. Durch die angewen-
dete Inokulationsmethode konnte der AM-Infektionsgrad auf beiden Rebschulflächen an den
Pfropfreben Riesling/5C bzw. Müller-Thurgau/5BB erhöht werden. Auf dem Standort „Hart-
häuser“ betrug der AM-Infektionsgrad 17 Wochen nach der Inokulation mit Glomus sp. 12 %
und auf dem Standort „Steingebiß“ 17 % (s. Abb. 4.6 und 4.7). Dabei setzte sich die Mykor-
rhiza-Population aus dem ausgebrachten AM-Pilz Glomus sp. und der nativen Mykorrhiza des
Standortes zusammen. An den Indikatorpflanzen Weißklee (Standort „Harthäuser“) und Mais
(Standort „Steingebiß“) war der Inokulationserfolg mit 9,2 % bzw.
14 % infizierter Wurzellänge geringer als an den Reben (s. Tab. 4.4 und Abb. 4.7). Über er-
folgreiche Inokulationsversuche berichtet HAMEL (1996) insbesondere in entseuchten und
nährstoffarmen Böden. SIEVERDING (1991) stellt bei Freilandinokulationen mit dem AM-Pilz
Glomus manihotis an Maniok (Cassava cv. MCol 1522) in den Tropen fest, daß auf Standor-
ten mit hohen qualitativen und quantitativen bodeneigenen AM-Pilzen keine Ertragserhöhung
durch Inokulation möglich ist. Allerdings sind die Böden der Tropen und Subtropen mit dem
hohen Anteil unfruchtbarer P-fixierender Böden anders zu beurteilen als die Böden der gemä-
ßigten Zone mit ihren in der Regel hohen P-Versorgungsgraden und hohem Düngungsniveau.
Die eigenen Versuchsflächen waren gut mit Nährstoffen versorgt (s. Tab. 4.1). Trotzdem führ-
te die Inokulation mit Glomus sp. zu deutlich verbesserten Wachstumsbedingungen der
Pfropfreben. Dies steht im Einklang mit den Untersuchungen von BALTRUSCHAT (1987 a, b),
der in landwirtschaftlich genutzten Böden mit standortspezifischer Mykorrhizapopulation und
hohem Düngungsniveau durch Inokulation mit AM-Pilzen weitere Ertragssteigerungen her-
beiführen konnte.
Der ausgebrachte AM-Pilz konnte die Rebwurzeln infizieren und sich gegen die autochthone
Mykorrhizapopulation, die in den Kontrollen ohne AM-Inokulation an den Rebwurzeln und
an Weißklee und Mais nachgewiesen werden konnte, durchsetzen. SIEVERDING (1991) stellte
an Maniok fest, daß neu gebildete Wurzeln mindestens zwei Tage mit dem Mykorrhizainoku-

Kapitel 4 Praktische Anwendung der AM 73
lum Kontakt haben müssen, bis eine erfolgreiche Infizierung erfolgen kann. Beim Ausgraben
der Rebwurzeln im vorliegenden Versuch konnte sehr deutlich die vertikale Verteilung des an
Blähton gebundenen Inokulums im Bereich der Pflanzfolie auch in tiefere Bodenzonen
beobachtet werden (vgl. Abb. 4.5). Es war gewährleistet, daß die Rebwurzeln mit dem Inoku-
lum Kontakt hatten und infiziert wurden.
Auf der Versuchsfläche „Harthäuser“ wurde an den Rebwurzeln in den Kontrollen ein sehr
niedriger Infektionsgrad festgestellt, der mit 3 % infizierter Wurzellänge niedriger war als in
den Kontrollflächen des Standortes „Steingebiß“ (s. Abb. 4.6). Dies läßt darauf schließen, daß
das Potential der autochthonen Mykorrhiza dieses Standortes sehr gering war. Die niedrige
Infektionsrate des Weißklees bestätigt diese Annahme. Das geringe Auftreten der nativen My-
korrhiza an diesem Standort könnte neben der hohen P-Versorgung des Bodens (s. Tab. 4.1)
auf die Fruchtfolge der letzten Jahre zurückgeführt werden. Auf der Versuchsfläche Harthäu-
ser wurde in den letzten Jahren mehrmals Ölrettich (R. sativus oleiformis L.) als Zwischen-
frucht angebaut. Ölrettich gehört zu den Cruciferen und gilt nach HARLEY und HARLEY (1987)
zu den Nichtwirtspflanzen für die arbuskuläre Mykorrhiza. Von landwirtschaftlich genutzten
Kulturarten gibt es in der Literatur zahlreiche Hinweise, daß nicht mykotrophe Pflanzen in der
Fruchtfolge das Mykorrhizierungspotential im Boden reduzieren (KRUCKELMANN, 1975;
BALTRUSCHAT und DEHNE, 1988; LAND, 1990). Der mehrmalige Zwischenfruchtanbau mit
Ölrettich könnte deshalb das AM-Potential im Rebschulboden gesenkt haben. Unterstützt
wird diese Annahme durch den Versuch „Einfluß der Begrünung auf die AM“ (s. Kap. 3). Bei
diesem war der Infektionsgrad an den Rebwurzeln aus der Variante mit den Cruciferen Ölret-
tich, Winterraps, Gelbsenf und Winterrübsen tendenziell geringer als in den anderen Begrü-
nungsvarianten mit AM-Wirtspflanzen. Der Einfluß der Ölrettichwurzeln auf das Infektions-
potential in der Rebschule „Harthäuser“ könnte auch nach dem Abmulchen und Umbrechen
des Bodens zur Pflanzvorbereitung für die Pfropfreben bestehen bleiben, denn der Großteil
der Wurzeln bleibt eine gewisse Zeit im Boden erhalten und wird erst nach und nach abge-
baut. DÜTSCH et al. (1997) fanden in einem Weinberg mit unterschiedlichen Winterbegrü-
nungseinsaaten bei Perko (Chinakohl x Winterrübsen) im Herbst eine neunfach höhere Wur-
zeltrockensubstanz als z. B. bei Phacelia. Möglicherweise können bereits Wurzelstücke von
AM-Nichtwirtspflanzen, die nicht mehr in Kontakt mit der restlichen Pflanze stehen, einen
hemmenden Einfluß auf die Mykorrhizierung der Rebwurzeln haben, was aus den Untersu-

Kapitel 4 Praktische Anwendung der AM 74
chungen von BÉCARD und PICHÉ (1990) hervorgeht. Durch die Inokulation mit dem AM-Pilz
konnte der Infektionsgrad im eigenen durchgeführten Versuch auf 12 % angehoben werden
(vgl. Abb. 4.6). Dieser lag damit aber trotzdem deutlich niedriger als auf dem Standort
„Steingebiß“. Vor der Nutzung des Standortes „Steingebiß“ als Rebschule wurde drei Jahre
lang Getreide angebaut. Möglicherweise führte dies zu einer Erhöhung des AM-
Infektionspotentials, was von WULFF (1996) an Winterweizen beobachtet werden konnte.
Über Freilandinokulationen an Reben sind in der Literatur bisher keine Untersuchungen be-
kannt. Von anderen Pflanzenarten liegen erfolgsversprechende Berichte von Inokulationen
vor, die vor dem Verpflanzen an ihren endgültigen Standort während des Anzuchtstadiums in
Pflanzschulen oder Baumschulen mit AM-Pilzen inokuliert wurden (PLENCHETTE et al., 1981;
SIEVERDING, 1985; FERGUSON und MENGE, 1986; AZCÓN-AGUILAR und BAREA, 1997). FELD-
MANN (1990) inokulierte in Baumschulen des Kautschukbaumes (Hevea spec.) veredelte
Pfropfunterlagen und begründete den Inokulationserfolg damit, daß zum Zeitpunkt der Inoku-
lation das Wurzelsystem sehr stark reduziert ist und die jungen, nachwachsenden Wurzeln
sofort mit den inokulierten AM-Pilzen besiedelt werden. In Rebschulen kann von vergleichba-
ren Bedingungen ausgegangen werden. Unmittelbar nach Ausbringung der veredelten Pfropf-
reben hatte sich noch kein Wurzelsystem ausgebildet. Die ersten Rebwurzeln konnten in den
Wurzelproben nachgewiesen werden, die fünf Wochen nach der Pflanzung entnommen wur-
den. Die Infektion mit AM-Pilzen war zu diesem Zeitpunkt mit 3 % („Harthäuser“, s. Abb.
4.6) bzw.
8 % („Steingebiß“, s. Abb. 4.7) sehr gering.
Im Rebschulversuch wurden pro Meter Mulchfolie 100 ml Mykorrhizainokulum (Blähton als
Trägermaterial mit infizierten Wurzeln, Hyphen und Sporen) ausgebracht. Auf einem Hektar
Rebschulfläche werden circa 10.000 Meter Mulchfolie ausgelegt. Im Bereich von einem Me-
ter Mulchfolie befinden sich etwa 12 Reben. Hieraus ergibt sich eine Aufwandmenge von
1.000 l Inokulum für die Inokulation von einem Hektar Rebschule, auf dem circa 120.000
Reben erzeugt werden. HOWELER und SIEVERDING (1983) kamen bei einem Boden-Inokulum-
Gemisch auf eine Menge von 1 - 1,5 t/Hektar, wenn jede Maniok-Pflanze mit 100 g inokuliert
wurde, wobei pro Hektar circa 10 - 20.000 Maniokpflanzen angebaut wurden. IKRAM et al.
(1992) inokulierten nach einer 20tägigen Anzuchtphase 10 g eines AM-Boden-Gemisches an

Kapitel 4 Praktische Anwendung der AM 75
Parakautschukbäumen (Hevea brasiliensis) unmittelbar ins Pflanzloch. BALTRUSCHAT (1987
b) brachte sporenbeladene poröse Blähtonpartikel aus Mykorrhiza-Dauerkulturen mit Glomus
etunicatum mit herkömmlichen Unterfußdüngungsgeräten seitlich zur Saatreihe in Mais aus.
Dabei führte die Inokulation bei einer Aufwandmenge von 150 kg/Hektar zu einer Förderung
der AM-Infektion und zu höheren Kolbentrockengewichten. Um die Freilandinokulationen in
Rebschulen praxisrelevanter zu gestalten und eine Reduzierung der Inokulummengen zu er-
reichen, müßten weitere Untersuchungen durchgeführt werden. Ansatzpunkte wären bei-
spielsweise Modifizierungen am Folienlegegerät, auf das aus einem Vorratsbehälter je nach
Fahrgeschwindigkeit in den Wurzelbereich der Pfropfreben definierte Inokulummengen abge-
legt werden könnten.
Etwa 17 Wochen nach der Inokulation betrug der AM-Infektionsgrad an den Rebwurzeln
12 % (Standort „Harthäuser“) bzw. 17 % (Standort „Steingebiß“). EIBACH (1982) fand in Ge-
wächshausversuchen mit einer phosphatarmen Braunerde an Zwei-Augen-Stecklingen der
Rebsorte Bacchus nach Inokulation mit einem Sand-Sporen-Gemisch des AM-Pilzes Glomus
macrocarpus 26 Tage nach dem Austrieb bereits einen Infektionsgrad von 26 %. Nach weite-
ren 14 Tagen stieg der Infektionsgrad auf 39 % an. An Pfropfreben der Sorte Kerner/5BB fand
EIBACH (1982) im nicht gedüngten Boden (2,2 mg P/100 g Boden) einen Infektionsgrad von
32 %, der nach einer P-Düngung (Phosphatgehalt von 12 mg P/100 g Boden) auf 24 % zu-
rückging. Es kann davon ausgegangen werden, daß die höhere Infektionswahrscheinlichkeit in
Pflanzgefäßen die höheren Infektionsgrade von EIBACH (1982) im Vergleich zu den Freiland-
versuchen erklärt. Allerdings könnten die hohen P-Gehalte im Boden in der vorliegenden Un-
tersuchung mit 28,4 („Harthäuser“) bzw. 12,1 („Steingebiß“) mg P/100 g Boden ebenfalls
mitverantwortlich für die geringen Infektionen der Rebwurzeln sein. Dieser Sachverhalt wur-
de bereits in den Untersuchungen in Rebschulen deutlich (s. Kapitel 3) und würde mit den
Ergebnissen von MOHR (1997) übereinstimmen. Der Autor stellte in einer Untersuchung in 28
Rebflächen der Mittleren Mosel eine sehr enge Beziehung zwischen dem Boden-P-Gehalt und
der Mykorrhizierung der Rebwurzeln fest. Dabei nahm mit steigendem Phosphatgehalt des
Bodens
(3 - 35 mg P/100 g) die Mykorrhizierungsrate ab.

Kapitel 4 Praktische Anwendung der AM 76
Durch die Inokulation wurde das Wachstum der Pfropfreben verbessert. Dieser Effekt war 12
bzw. 14 Wochen nach dem Stecken der Reben signifikant (s. Abb. 4.8 und 4.9). Am Ende der
Vegetationsperiode werden die Reben ausgeschult und beim Rebveredler pflanzfertig herge-
richtet. Dabei wird ein Großteil des Wurzelsystems abgeschnitten und die gesamte Pfropfrebe
mit einer Chinosol-Lösung [8-Hydroxychinolin] gegen Grauschimmel (Botrytis cinerea Pers.)
behandelt. Durch diese Pflanzenschutzbehandlung ist eine Eliminierung der AM-Pilze an dem
verbliebenen Wurzelsystem möglich. Diese Annahme wird durch die Untersuchungen von
EIBACH (1982) bestätigt, die nach einer Applikation mit den Fungiziden DuPont Benomyl
[Wirkstoff Benomyl] und Bayleton [Wirkstoff Triademifon] eine geringere Mykorrhizierung
an Bacchus-Stecklingen fand. Dies bedeutet, daß eine Übertragung der AM-Pilze über das
Pflanzgut an den endgültigen Standort bei den in der Rebschule erzeugten Pfropfreben nur
bedingt möglich ist. Allerdings können die im Boden verbliebenen Wurzelreste eine sehr gute
Inokulumquelle für die nachfolgenden Kulturen darstellen und das AM-Infektionspotential im
Boden erhöhen (DEAL et al., 1972).
Für das verbesserte Wachstum der Pfropfreben durch die AM können zahlreiche Faktoren,
insbesondere im Zusammenhang mit der Wurzelgesundheit, eine Rolle spielen, die die
Wachstumssteigerungen an den Reben erklären könnten. Da auf der Versuchsfläche Harthäu-
ser keine Rebenmüdigkeit aufgetreten ist, bedarf es weiterer Untersuchungen, ob sich die in
Gefäßversuchen nachgewiesene Bekämpfung der Rebenmüdigkeit mit AM-Pilzen (WASCH-
KIES et al., 1994) auch im Freiland in Rebschulen bestätigt.
4.4.2 Einfluß der AM in der Topfrebenproduktion
Im Versuchsjahr 1995 führte die Inokulation mit dem AM-Pilz Glomus sp. an den Holzsteck-
lingen Riesling/125AA zu tendenziell größeren Trieblängen als die nicht infizierten Reben der
Kontrolle (s. Abb. 4.10). Bereits vier Wochen nach dem Topfen der Stecklinge betrug der
AM-Infektionsgrad 20,1 % und war mit 23,4 % nach weiteren vier Wochen kaum angestiegen
(s. Tab. 4.5). Die Beimpfung mit Glomus mosseae führte im Versuchsjahr 1997 zu Infektions-
graden von 18,3 % (Floraton 1) bzw. 16,3 % (Patzer-Erde) (s. Tab. 4.6).Verschiedene Mykor-
rhiza-Arten können sich in ihrer Wirksamkeit auf Pflanzenwachstum und Nährstoffaufnahme
unterscheiden. In den eigenen Versuchen zeigte sich der AM-Pilz Glomus mosseae als geeig-

Kapitel 4 Praktische Anwendung der AM 77
neter Symbiosepartner für Topfreben und führte bereits 17 Tage nach Inokulation zu signifi-
kant größeren Trieblängen.
FERGUSON und MENGE (1986) untersuchten verschiedene Inokulationsmethoden an Citrus
(Citrus aurantium L.) und stellten bei der Präinokulation der Citrus-Sämlinge im Gewächs-
haus mit Glomus deserticola deutliche Wachstumsverbesserungen nach der Anzuchtphase
fest. MENGE (1983) gibt an, daß die Inokulation von verschiedenen Containerpflanzen in der
Anzuchtphase im Gewächshaus unter praxisüblichen Bedingungen in großer Anzahl durchge-
führt werden kann. KRIKUN et al. (1987) inokulierten Paprika im Gewächshaus und pflanzten
die infizierten Sämlinge ins Freiland. Dabei liefen alle Arbeitsschritte praxisnah ab, wobei die
Durchführung der Inokulation des Anzuchtmediums einfach durchzuführen war. Dies kann
durch die eigenen Versuche bestätigt werden, bei denen die Beimischung des AM-Inokulums
zu den Kultursubstraten Floraton 1 und Patzer Einheitserde der einzige Schritt war, der von
der herkömmlichen Topfrebenproduktion abweicht.
PLENCHETTE et al. (1981) inokulierten Apfelsämlinge (Malus pumilla) in Pflanztöpfen mit
einem Durchmesser von 15 cm und pflanzten die Sämlinge nach sechs Wochen ins Freiland.
Dabei konnten die Autoren keine Wachstumsunterschiede zwischen den nicht mykorrhizierten
und mykorrhizierten Pflanzen feststellen. BIERMANN und LINDERMAN (1983 b) fanden bei
präinokulierten Geranien-Pflanzen (Pelargonium x hortorum L.H. Bailey) ebenfalls keine
Wachstumsunterschiede. KRIKUN et al. (1987) kamen zu ähnlichen Ergebnissen und ermittel-
ten bei präinokulierten Paprikasämlingen vor der Auspflanzung ins Feld keine signifikanten
Wachstumsunterschiede zwischen den beimpften Varianten und der nicht beimpften Kontrol-
le. Im Gegensatz zu den genannten Autoren wurde bei den Reben in den eigenen Untersu-
chungen das Wachstum der Topfreben bereits nach der 4- bzw. 8wöchigen Anzuchtphase im
Gewächshaus in beiden Versuchsjahren verbessert (s. Abb. 4.10 und Abb. 4.11). Es muß da-
von ausgegangen werden, daß sich der angewendete AM-Pilz frühzeitig in den Rebwurzeln
etablierte, so daß die Reben mit einem stärkeren Wachstum reagierten. SOLAIMAN und HIRA-
TA (1996 und 1997) kamen zu vergleichbaren Ergebnissen bei Reis und fanden nach einer
Anzuchtphase höhere Biomasseerträge bei den inokulierten Sämlingen im Vergleich zu den
nicht inokulierten. Die Mineralstoffanalysen bei den Topfreben zeigten insbesondere nach der
Anzuchtphase erhöhte P-Gehalte in der Trockensubstanz der Blattspreiten (s. Tab. 4.8). Wei-

Kapitel 4 Praktische Anwendung der AM 78
terhin wurden durch die Beimpfung mit dem AM-Pilz Glomus mosseae die Cu- und Zn-
Gehalte in den Blättern der Topfreben bei beiden Kultursubstraten erhöht (s. Tab. 4.7), so daß
möglicherweise die höheren Nährstoffgehalte in der Trockensubstanz der Blätter für das
Wachstum der inokulierten Reben verantwortlich gemacht werden können. Von ähnlichen
Mykorrhizaeffekten berichten JOHNSON et al. (1980) sowie MORIN et al. (1994), die in Sub-
straten mit hohen Nährstoffgehalten ebenfalls Wachstumssteigerungen an den inokulierten
Pflanzen beobachteten. Obwohl im Versuch von 1995 bei den Topfreben (s. Kapitel 4.3.2)
keine Nährstoffanalyse durchgeführt wurde, waren vermutlich auch in diesem Versuchsansatz
höhere Nährstoffgehalte in der Trockensubstanz der mykorrhizierten Reben Ursache für das
verbesserte Wachstum. Inwieweit die AM-Pilze die Nährstoffgehalte beeinflussen, wird in
Kapitel 5 näher beschrieben.
Die Notwendigkeit der Inokulation von Kulturpflanzen mit AM-Pilzen wird besonders in
nährstoffarmen Böden oder P-fixierenden Böden deutlich, wie sie beispielsweise im heiß-
feuchten Klima der Tropen vorzufinden sind (NURLAENY, 1995). Daneben kann der Einsatz
von sterilisierten Bodensubstraten zu Kümmerwuchs an den Pflanzen führen (KOCH et al.,
1997). SCHUBERT et al. (1990) inokulierten in-vitro-vermehrte Reben der Unterlagssorte
1103P
(V. berlandieri x V. rupestris) mit dem AM-Pilz Glomus caledonicum in unterschiedlichen
Kultursubstraten. In einem Torf-Sand-Gemisch stellten die Autoren zwar Infektionen fest, die
Reben zeigten jedoch keine Wachstumsunterschiede im Vergleich zu den nicht infizierten
Kontrollen. Erst durch Zugabe von 10 % sterilisierter Weinbergserde waren signifikante
Wachstumsunterschiede in Form von vergrößerten Gesamtblattflächen bei den mykorrhizier-
ten Reben nachweisbar. Von ähnlichen Versuchsergebnissen berichteten BIERMANN und LIN-
DERMAN (1983 a) an mykorrhizierten Kleepflanzen. In den eigenen Untersuchungen konnten
sich dagegen die AM-Pilze Glomus sp. (Versuch 1995) und Glomus mosseae (Versuchsjahr
1997) in beiden Kultursubstraten an den Rebwurzeln etablieren und das Wachstum der Topf-
reben Riesling/125AA nach der Anzuchtphase fördern. Dies steht im Einklang mit den Ver-
suchsergebnissen von VARMA und SCHUEPP (1994), die in-vitro-vermehrte Hortensia-
Pflanzen (Hydrangea var. Leuchtfeuer) mit Glomus intraradices in einem Torfsubstrat inoku-
lierten. Dabei zeigten die mykorrhizierten Pflanzen besseres Wachstum und eine größere
Blattfläche als die Kontrollpflanzen, obwohl das eingesetzte Torfsubstrat mit 105 mg/l P we-

Kapitel 4 Praktische Anwendung der AM 79
sentlich höher mit Phosphat versorgt war als die eingesetzten Kultursubstrate in der vorlie-
genden Untersuchung (vgl. Tab. 4.2).
Die Verwendung der unterschiedlichen Bodensubstrate in der Anzuchtphase hatte keine signi-
fikanten Auswirkungen auf den AM-Infektionsgrad (s. Tab. 4.6). Während die Wachstumsun-
terschiede bei den mit Patzer-Erde angezogenen Reben nach der Anzuchtphase zwischen
Kontrolle (-AM) und mykorrhizierten Reben (+AM) statistisch abzusichern waren, konnten
nach dem Auspflanzen in den Weinberg keine Unterschiede mehr beobachtet werden (s. Abb.
4.15). Die Wachstumsunterschiede deuten auf eine verbesserte Nährstoffaufnahme hin, die in
den höheren P-Gehalten der Blattspreiten in dem mit dem AM-Pilz Glomus mosseae inoku-
lierten Reben bei gleichzeitig höherer Trockensubstanzproduktion im Vergleich zu den nicht
inokulierten Reben bestätigt wurde (s. Tab. 4.8). Neben dem Nährstoffeffekt können andere
Faktoren bei den mykorrhizierten Reben zu einer Wachstumsverbesserung geführt haben.
Denkbar sind Veränderungen im Phytohormonhaushalt (DRÜGE, 1992) oder eine Förderung
der Wurzelgesundheit durch die Beeinflussung von phytopathogenen Pilzen im Boden (DEH-
NE, 1994). Diese Annahme wird dadurch verstärkt, daß beim Auspflanzen der Topfreben in
den Weinberg zwischen den Behandlungen (-AM/+AM) keine signifikanten Unterschiede in
den Nährstoffgehalten bestanden. Trotzdem traten bei den mit Floraton 1 angezogenen Reben
auch im Freiland Wachstumsunterschiede auf. Von ähnlichen Ergebnissen berichten PLEN-
CHETTE et al. (1981) an präinokulierten Apfel-Sämlingen. Es ist denkbar, daß die Rebwurzeln
nach dem Durchwachsen des Boden-Inokulum-Gemisches aus dem Gittertopf nicht mehr aus-
reichend infiziert waren und die Ausbildung des externen Hyphengeflechts eingeschränkt
wurde. Dies wurde durch einen späteren Versuch bestätigt, bei dem für die Bestimmung des
AM-Infektionsgrades nur die Wurzeln berücksichtigt wurden, die bereits aus dem Boden-
Inokulum-Gemisch des Pflanzgefäßes heraustraten. Anhand dieser Wurzeln konnte keine
bzw. nur eine geringe AM-Infektion festgestellt werden. BIERMANN und LINDERMAN (1983 b)
kamen dagegen zu anderen Ergebnissen. Die Autoren inokulierten Geranien- (Pelargonium x
hortorum L.H. Bailey) Stecklinge in Pflanzgefäße mit einem Durchmesser von 6 cm und be-
stimmten nach sechs Wochen die Mykorrhizierung. Anschließend wurden die Pflanzen in
größere Töpfe mit sterilisiertem Boden umgetopft. Nach weiteren sechs Wochen konnten
BIERMANN und LINDERMAN (1983 b) auch an den neu gebildeten Wurzeln außerhalb der Ino-
kulationsschicht im sterilisierten Boden mit 74,7 % ähnlich hohe AM-Infektionen feststellen

Kapitel 4 Praktische Anwendung der AM 80
wie an den Wurzeln aus dem Boden-Inokulum-Gemisch mit 70,2 %. Möglicherweise hat der
AM-Pilz in der vorliegenden Untersuchung nur unzureichende externe Hyphen ausgebildet
und es kam nicht zu Sekundärinfektionen, so daß die Rebwurzeln außerhalb des Pflanzgefäßes
nicht mehr infiziert wurden.
Zusammenfassend haben die Ergebnisse der vorliegenden Arbeit bezüglich der Anwendung
der AM in der Rebenpflanzguterzeugung gezeigt, daß die Umsetzung in die Praxis bei der
Topfrebenproduktion relativ einfach durchzuführen ist. Beide AM-Pilzisolate besiedelten die
Rebwurzeln und führten zu einem stärkeren Trieblängenwachstum als die nicht beimpften
Topfreben. Dagegen sind Freilandinokulationen in Rebschulen erst einsatzreif, wenn die Ino-
kulummenge reduziert werden kann und die Anwendung neben den Wachstumsverbesserun-
gen an den Pfropfreben eine Bekämpfungsmöglichkeit für rebenmüde Standorte darstellt.

Kapitel 5 Inokulationsversuche 81
GEFÄßVERSUCHE
5 Inokulationsversuche mit AM-Pilzen an Rebenstecklingen
5.1 Einleitung
Die meisten Inokulationsversuche in Gefäßversuchen wurden mit sterilen Böden oder Kultur-
substraten und anschließender Beimpfung mit Mykorrhiza-Inokulum durchgeführt. Dabei
wurden oftmals AM-Pilzisolate eingesetzt, die zuvor aus Böden isoliert und unter Gewächs-
hausbedingungen kultiviert wurden. Wie in Kapitel 4 beschrieben konnte auf dem Rebschul-
standort „Steingebiß“ eine geringe Mykorrhizainfektion an den Pfropfreben festgestellt wer-
den, die durch die bodeneigene AM-Spezies verursacht wurde. In der vorliegenden Untersu-
chung sollte der native Mykorrhizabeitrag aus dem gleichen Rebschulboden ohne Beimi-
schung von Substraten im Vergleich zum sterilisierten Boden ohne native Mykorrhiza auf die
Nährstoffaufnahme und das Wachstum an verschiedenen Grünstecklingen untersucht werden.
In den Untersuchungen in Kapitel 3 wurden in einer Adaptionsanlage signifikante Sortenun-
terschiede im AM-Infektionsgrad zwischen verschiedenen Pfropfkombinationen festgestellt.
Möglicherweise reagieren die einzelnen Unterlagsrebsorten unterschiedlich auf die in Wein-
bergsböden vorkommende native AM-Pilzspezies. Deshalb wurde ein Sortenscreening mit
neun Unterlagssrebsorten und einer Edelreissorte durchgeführt. Dabei waren u. a. die in der
Praxis gängigen Unterlagssorten 125AA, SO4, 5BB und 5C, die Sämlingsnachkommen aus
der Kreuzung Vitis riparia x V. berlandieri sind, Gegenstand der Untersuchungen.
Durch die Inokulation mit AM-Pilzen konnte der Infektionsgrad der jungen Pfropfreben in
Rebschulen erhöht werden (s. Kapitel 4.3.1), allerdings waren der AM-Infektionsgrad der
Rebwurzeln im Vergleich zu Inokulationsversuchen in Gefäßen (EIBACH, 1982; SCHUBERT et
al., 1988; MOHR, 1993; KARAGIANNIDIS et al., 1995) relativ niedrig. Es ist denkbar, daß die
Rebwurzeln verhältnismäßig schnell durch die Inokulumschicht hindurchgewachsen sind,
ohne daß es zu einer ausgeprägten AM-Infektion kam. Die Inokulationsversuche an Topfreben
haben diese Annahme bestätigt (s. Kapitel 4.3.2). Über die Ausbreitung einer Mykorrhizain-
fektion an Rebwurzeln in einem definierten Bodenvolumen gibt es in der Literatur bisher kei-

Kapitel 5 Inokulationsversuche 82
ne Hinweise. Daher war ein weiteres Ziel der vorliegenden Untersuchung, mit Hilfe von Wur-
zelbeobachtungskästen und einer unterschiedlichen Bandinokulation mit dem AM-Pilz Glo-
mus mosseae die Entwicklung des AM-Pilzes innerhalb des Wurzelsystems sowie den Einfluß
auf das Wachstum und die Nährstoffaufnahme in einem Gefäßversuch zu überprüfen.
Aus der Literatur sind zahlreiche Untersuchungen bekannt, in denen an in-vitro-vermehrten
Reben durch Inokulation mit AM-Pilzen signifikante Wachstumssteigerungen erzielt werden
konnten (RAVOLANIRINA et al., 1989; MAZZITELLI und SCHUBERT, 1989; SCHUBERT et al.,
1990; SCHELLENBAUM et al., 1991). Diese Untersuchungen wurden mit wurzelechten Reben
durchgeführt. In der Weinbaupraxis werden allerdings Pfropfreben verwendet, die aus einer
Unterlage und dem Edelreis bestehen. Durch die Pfropfung zweier unterschiedlicher Genoty-
pen besteht die Möglichkeit, daß die Stoffkreisläufe innerhalb der Pflanze gestört werden.
Denkbar sind modifizierte Kohlenstoffakkumulationen im Wurzelbereich der Rebwurzeln, die
die AM beeinflussen könnten. Ob es tatsächlich eine Wirkung des Pfropfeingriffs auf die My-
korrhizierung an Rebwurzeln gibt, sollte Gegenstand der vorliegenden Untersuchung sein. Mit
Hilfe des „Hohenheimer Verfahrens“ wurden Grünveredlungen angefertigt, wobei im Gegen-
satz zu konventionellen Pfropfungen Grünstecklinge anstatt Holzstecklinge verwendet wur-
den. Um unterschiedliche Pfropfeffekte erfassen zu können, wurden sowohl homo- (Pfropfun-
gen von gleichen Genotypen aufeinander) als auch heteroplastische (Pfropfungen von unter-
schiedlichen Genotypen aufeinander) Pfropfungen mit wurzelechten Stecklingen verglichen.
Neben biotischen Streßfaktoren ist die Rebe auch abiotischen Streßfaktoren ausgesetzt. Hierzu
zählt die Schadstoffbelastung der Luft, wobei die wichtigsten pflanzentoxischen Schadstoffe
Schwefeldioxid (SO2), Ozon (O3), Fluorwasserstoff (HF), Stickstoffoxide (NO2, NO) und
Peroxyacetylnitrat (PAN) sind (HOCK und ELSTNER, 1988). Ozon zählt zu den phytotoxisch
wirksamsten Luftschadstoffen und kann zu beträchtlichen Schäden an landwirtschaftlichen
Kulturpflanzen führen. In Kalifornien wurden erste Ozonschäden an Reben bereits in den 50er
Jahren beobachtet (RICHARDS et al., 1958). Aber auch aus deutschen Anbaugebieten berichten
HERRMANN (1991) sowie VON TIEDEMANN und HERRMANN (1992) von Blattschäden an Re-
ben, die auf Ozoneinfluß zurückgeführt werden können. Infolge der Einwirkung von photo-
chemischen Oxidantien kommt es zu veränderten Stofftransporten zwischen Assimilationsor-
ganen und Wurzeln (ANONYM, 1983), die z. B. bei Leguminosen zu einer Verringerung von

Kapitel 5 Inokulationsversuche 83
Zahl, Größe und Gewicht der Rhizobium-Knöllchen geführt haben (BLUM und TINGEY, 1977).
Aber auch die Mykorrhizierung verschiedener Pflanzenarten wurde durch Ozoneinfluß
nachteilig beeinflußt (MCCOOL et al., 1979). Bei Citrus-Sämlingen war nach langfristiger O-
zoneinwirkung die AM-Infektion an den Wurzeln verringert, was zu einer Minderung der
Wuchsleistung führte (MCCOOL et al., 1979). Aus Untersuchungen von BREWER und HEAGLE
(1983) geht hervor, daß bei mykorrhizierten Sojabohnen im Vergleich zu nicht mykorrhizier-
ten Sojabohnen weniger durch O3 bedingte Schadsymptome auftraten. Ziel der vorliegenden
Untersuchung war herauszufinden, ob eine erhöhte Ozonbelastung der Luft den AM-
Infektionsgrad von Rebwurzeln sowie das Wachstum und die Nährstoffaufnahme beeinflußt.
Pflanzen werden im Freiland in der Regel durch unterschiedliche Kombinationen verschiede-
ner Schadstoffe belastet, die sich gegenseitig beeinflußen können (HOCK und ELSTNER, 1988).
Deshalb wurde in der Versuchsanstellung neben der „Ozonvariante“ und der „Kontrolle“ mit
gefilterter Luft eine Variante mit natürlichen Schadstoffkonzentrationen berücksichtigt, die
Aufschluß über den Einfluß der natürlich vorkommenden Schadstoffkonzentrationen auf die
Mykorrhizierung an Reben geben sollte.
5.2 Material und Methoden
5.2.1 Einfluß der nativen Mykorrhiza eines Rebschulbodens auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünsteck-lingen unterschiedlichen Genotyps
5.2.1.1 Pflanzenmaterial, Versuchsgefäße und Boden
Als Versuchspflanzen wurden Grünstecklinge (Zweiaugenstecklinge) der Unterlagssorten
Börner (Vitis riparia x V. cinerea), C3309 [FR 465/5] (V. riparia x V. rupestris), 5BB [N101]
(V. berlandieri x V. riparia), 125AA [FR 26] (V. berlandieri x V. riparia), SO4 [N201]
(V. berlandieri x V. riparia), 5C [6Gm] (V. berlandieri x V. riparia), 8B [Gm349/7]
(V. berlandieri x V. riparia), A9 (V. riparia x V. cinerea), C3 (125AA x V. cinerea) sowie der
Edelreissorte Riesling [N90] (V. vinifera) verwendet. Die Grünstecklinge wurden aus grünen
Rebtrieben aus dem Freiland von Vermehrungsanlagen der SLFA Neustadt zugeschnitten und
in Perlit (Agriperl) unter einer Sprühnebelanlage im Gewächshaus angezogen. Nach etwa drei

Kapitel 5 Inokulationsversuche 84
Wochen konnten die ausgetriebenen und bewurzelten Stecklinge für den Versuch verwendet
werden.
Als Versuchsboden wurde ein Boden eingesetzt, der drei Monate vor Versuchsbeginn aus
einer Rebschule in Neustadt/Weinstraße entnommen wurde. Der Boden wurde gesiebt (< 25
mm) und anschließend ein Teil mit einer Mikrowelle (Micro-Chef FM 3515, Fa. Moulinex)
sterilisiert. Es handelte sich um einen schwach sandigen Lehm. Nähere Eigenschaften des
Versuchsbodens sind in der Tabelle 5.1 dargestellt. Die Bodenanalysen wurden vom Fachbe-
reich Bodenkunde und Bodenschutz der Staatlichen Lehr- und Versuchsanstalt in Trier durch-
geführt. Bis zum Versuchsbeginn wurde der Boden bei + 5 °C gelagert.
Tab. 5.1: Chemische und physikalische Eigenschaften des Versuchsbodens
Humus pH P K Mg Korngrößenbestimmung
(KCl) (CAL) CaCl2 Ton Schluff Sand
(%) (mg/100 g Boden) (%)
1,7 7,5 12,2 14,2 8,0 19,0 45,9 35,1
5.2.1.2 Versuchsanordnung
Nach der Anzuchtphase wurden die bewurzelten Grünstecklinge am 28.07.1994 in 2 l fassen-
de Plastiktöpfe („Teku“, Fa. Pöppelmann, Hannover) mit einem Durchmesser von 14 cm ge-
topft. Jedes Gefäß wurde mit einer Rebe bepflanzt und die Bodenoberfläche zur Verringerung
der Evaporation mit Perlit (Agriperl) abgedeckt. Beim Einfüllen in die Pflanzgefäße hatte der
Versuchsboden einen Wassergehalt von 13 %. Die Töpfe wurden je nach Bedarf mit Lei-
tungswasser gegossen.
Um die Wirkung der nativen Mykorrhiza eines Rebschulbodens an Reben zu überprüfen, wur-
de ein Teil des Bodens mit Hilfe einer Mikrowelle sterilisiert. Bei zehn verschiedenen Sorten

Kapitel 5 Inokulationsversuche 85
mit jeweils zwei unterschiedlichen Bodenbehandlungen (sterilisiert [-AM]; nicht sterilisiert
[+ native AM]) wurden 20 Varianten angelegt. Der Versuch wurde in fünf Wiederholungen
durchgeführt. Einmal wöchentlich wurde die Trieblänge der Reben bestimmt. Der vorliegende
Versuch wurde in der Zeit vom 28./29.07. - 27.09.1994 im Gewächshaus bei einer durch-
schnittlichen Temperatur von 26 ± 5 °C am Tag (16 h) bzw. 20 ± 6 °C in der Nacht (8 h)
durchgeführt. Die Ernte der Reben erstreckte sich über einen Zeitraum von vier Tagen
(s. Kap. 2.5). In der Trockensubstanz der Blätter wurden die Makro- und Mikronährstoffe
nach der in Kapitel 2.6 beschriebenen Methode quantitativ bestimmt. Ab dem 12.08.1994
wurde Zusatzlicht (Osram HQI Lampen, 400 W, DH) gegeben. Gegen Echten Mehltau (Unci-
nula necator (Schwein.) Burr.) wurde fünfmal Bayfidan spezial WG [Triadimenol,
0,05 %ig] eingesetzt. Zuvor wurden die Töpfe mit Plastikfolie verschlossen. Drei Wochen
nach Versuchsbeginn wurde wöchentlich eine N-Düngung in Form von Calciumnitrat
(Ca(NO3)2) mit jeweils 0,05 g N/Topf durchgeführt (MERKT, pers. Mitteilung, 1994).
5.2.2 Einfluß unterschiedlicher Inokulationstiefen mit Glomus mosseae
auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünstecklingen in Wurzelbeobachtungskästen
5.2.2.1 Pflanzenmaterial, Versuchsgefäße und Boden
Als Pflanzenmaterial wurden Grünstecklinge (Zweiaugenstecklinge) der Unterlagssorte SO4
[N201] (Vitis berlandieri x V. riparia) eingesetzt (s. Kapitel 5.2.1.1). Als Versuchsgefäße
wurden Schrägwandgefäße aus PVC mit 70 cm Höhe, 37 cm Breite und 9 cm Tiefe verwen-
det, deren Vorderseite aus einer abnehmbaren Plexiglasplatte bestand. Um die Wurzeln vor
Licht zu schützen, wurden die Gefäße mit schwarzer Folie abgedeckt.
Für den Inokulationsversuch wurde ein Rebschulboden verwendet, der sieben Wochen vor
Versuchsbeginn aus dem Freiland (Rebschule in Neustadt/W.) entnommen wurde. Der Boden
wurde gesiebt (< 25 mm) und anschließend mit einem Bodensterilisator (Sterilo W,
Fa. Schwarz, Alpirsbach) bei einer Temperatur von 110 °C acht Stunden lang sterilisiert. Es
handelte sich um einen mittel-lehmigen Sand. Nähere Eigenschaften des Versuchsbodens sind

Kapitel 5 Inokulationsversuche 86
in Tabelle 5.2 dargestellt. Die Bodenanalysen wurden vom Fachbereich Bodenkunde und Bo-
denschutz der Staatlichen Lehr- und Versuchsanstalt in Trier durchgeführt. Bis zum Ver-
suchsbeginn wurde der Boden bei 2,5 °C gelagert.
Tab. 5.2: Chemische und physikalische Eigenschaften des Versuchsbodens
Humus pH P K Mg Korngrößenbestimmung
(KCl) (CAL) CaCl2 Ton Schluff Sand
(%) (mg/100 g Boden) (%)
2,9 5,9 2,2 15,0 14,0 11,4 19,5 69,1
5.2.2.2 Versuchsanordnung
Beim Befüllen der Schrägwandgefäße wurden je nach Variante Inokulumbänder mit dem ar-
buskulären Mykorrhizapilz Glomus mosseae (Nicol. and Gerd.) Gerdemann & Trappe in einer
Tiefe von 9 - 18 („oben“) bzw. 36 - 45 cm („unten“) eingebracht (vgl. Abb. 5.1). Pro Inoku-
lumband wurde eine Menge von 2.500 ml Inokulum mit einer Höhe von 9 cm in die Gefäße
eingefüllt. Die nicht mykorrhizierten Varianten (Kontrolle) erhielten die gleiche Menge Ino-
kulum von nicht mykorrhizierten Kleepflanzen. Der Versuchsboden hatte beim Einfüllen in
die Gefäße einen Wassergehalt von 15 %. Der Boden wurde schichtweise auf eine Bodendich-
te von d = 1,0 g/cm3 eingefüllt, d. h. die einer Schichtdicke von 2 cm entsprechende Boden-
menge bzw. 9 cm dicke Inokulummenge wurde jeweils eingefüllt, verteilt und angedrückt. Die
Gefäße wurden je nach Bedarf mit Leitungswasser gegossen. Der Versuch wurde im Ge-
wächshaus bei einer durchschnittlichen Temperatur von 25 ± 2 °C am Tag (16 h) bzw. 22 ± 3
°C in der Nacht (8 h) in der Zeit vom 07.04. - 20.05.1997 durchgeführt. Gegen Echten Mehl-
tau (Uncinula necator (Schwein.) Burr.) wurde zweimal Folicur E [Dichlofluanid + Tebuco-
nazol, 0,25 %ig]) eingesetzt.

Kapitel 5 Inokulationsversuche 87
Abb. 5.1: Schematische Darstellung der Versuchsgefäße mit Inokulationsbereich „oben“ (Inokulumband in einer Gefäßtief von 9 - 18 cm) und „unten“ (Inokulumband in einer Gefäßtiefe von 36 - 45 cm)
Zur Verringerung der Evaporation wurden die Versuchsgefäße mit einer 5 cm dicken Quarz-
kieselschicht (Ø 5 - 8 cm) abgedeckt. Jedes Gefäß wurde mit zwei Reben bepflanzt
(s. Abb. 5.1). Pro Variante und Erntetermin standen vier Wiederholungen zur Verfügung, wo-
bei die beiden Pflanzen aus einem Gefäß zu einem Mittelwert zusammengefaßt wurden.
5.2.2.3 Ernte der Versuchspflanzen
Die erste Ernte der Versuchspflanzen erfolgte nach 4wöchiger Wachstumsphase nach der in
Kapitel 2.5 beschriebenen Methode. Zu diesem Zeitpunkt wurde der Infektionsgrad der Wur-

Kapitel 5 Inokulationsversuche 88
zeln (infizierte Wurzellänge in %) ermittelt. Eine Bestimmung der Trockenmasse des oberir-
dischen Aufwuchses erfolgte nicht. Die zweite Ernte erfolgte sechs Wochen nach Versuchs-
beginn. In der Trockensubstanz der Blätter wurden die Makro- und Mikronährstoffe nach der
in Kapitel 2.6 beschriebenen Methode quantitativ bestimmt.
Die Wurzeln wurden schichtweise in verschiedenen Blöcken (9 cm hoch, 9 cm tief, 37 cm
breit) aus den Versuchsgefäßen entnommen. Hierbei wurden die Wurzeln sorgfältig aus dem
Bodenblock ausgelesen und das Frischgewicht bestimmt. Pro Pflanzgefäß wurden aus den
Tiefenstufen 0-9 / 9-18 / 18-27 / 27-36 / 36-45 / 45-54 / 54-66 cm sieben Wurzelproben unter-
sucht.
5.2.3 Einfluß einer Inokulation mit Glomus intraradices auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von grün- veredelten Stecklingen
5.2.3.1 Pflanzenmaterial, Versuchsgefäße und Boden
Für den Inokulationsversuch wurden Grünstecklinge (Zwei- bzw. Dreiaugenstecklinge) der
Unterlagssorte SO4 und der Edelreissorte Riesling aus grünen Rebtrieben von im Gewächs-
haus kultivierten Reben zugeschnitten. Um den Einfluß der Veredlung auf die Mykorrhizie-
rung an den Rebwurzeln zu überprüfen, wurden verschiedene Pfropfkombinationen angefer-
tigt. Hierzu wurden sowohl homo- (Pfropfung von gleichen Genotypen aufeinander) als auch
heteroplastische (Pfropfung von unterschiedlichen Genotypen aufeinander) Pfropfungen
durchgeführt (s. Tab. 5.3).
Tab. 5.3: Übersicht über die Versuchsvarianten
Varianten wurzelecht / Pfropfung
SO4 wurzelecht Riesling wurzelecht Riesling/SO4 heteroplastische Pfropfung Riesling/Riesling homoplastische Pfropfung

Kapitel 5 Inokulationsversuche 89
SO4/SO4 homoplastische Pfropfung
Die Stecklinge von SO4 und Riesling, die im Versuch als wurzelechte Reben verwendet wur-
den, bestanden aus zwei Internodien mit einem basalen Nodium zur besseren Bewurzelung.
Die Pfropfpartner, die als Edelreis verwendet wurden, wurden auf ein Internodium mit apikal
belassenem Nodium und Knospe zurechtgeschnitten. Darunter verblieb ein Triebstück von
mindestens 2 cm, um den maschinellen V-Schnitt durchführen zu können. Die Grünveredlun-
gen wurden mit der Pfropfmaschine Vitimat (Fa. Wahler, Weinstadt-Schnait) nach dem „Ho-
henheimer Verfahren“ durchgeführt. Nach dem Pfropfvorgang wurden die Reben zur Bewur-
zelung in Steinwollwürfel (Fa. Grodania) gesteckt, die sich auf Kunststoffschalen befanden.
Die Anzucht der Reben erfolgte in einer Klimakammer bei einer Temperatur von 28 °C (± 1)
am Tag (16 h) bzw. 26 °C (± 1) in der Nacht (8 h). Die relative Luftfeuchte betrug 94 %. Die
3wöchige Anzuchtphase diente vorrangig der Kallusbildung. Danach wurde die Temperatur
auf 24 °C am Tag bzw. 22 °C in der Nacht abgesenkt. Während dieser Phase sollte die Wur-
zelbildung angeregt werden. Die Reben wurden nach Bedarf mit Leitungswasser gegossen.
Während der gesamten Anzuchtphase wurde mehrmals ein Spezialbotrytizid (Rovral [Vinclo-
zolin, 0,1 %ig]) eingesetzt.
Nach der 4wöchigen Anzuchtphase wurden die bewurzelten Stecklinge in 2 l fassende Plastik-
töpfe getopft (s. Kap. 5.2.1). Als Versuchsboden wurde der in Kapitel 5.2.2 beschriebene Reb-
schulboden verwendet. Ein Teil des sterilisierten Bodens wurde mit dem AM-Pilz Glomus
intraradices (s. Kap. 2.1) inokuliert. Die nicht mykorrhizierten Varianten (Kontrolle) erhielten
die gleiche Menge Inokulum von nicht mykorrhizierten Tagetespflanzen. Der Versuch wurde
in sechs Wiederholungen durchgeführt. Die Reben wurden in der Zeit vom 11.06. -
25.08.1997 im Gewächshaus bei einer durchschnittlichen Temperatur von 24 °C (± 4) am Tag
(16 h) bzw. 20 °C (± 4) in der Nacht (8 h) kultiviert. Gegen Echten Mehltau (Uncinula neca-
tor (Schwein.) Burr.) wurde in 14tägigem Abstand Folicur E [Dichlofluanid + Tebuconazol,
0,25 %ig] eingesetzt. Zuvor wurden die Pflanztöpfe mit Plastikfolie verschlossen. Vier Wo-
chen nach Versuchsbeginn wurde wöchentlich N gedüngt (s. unter 5.2.1.2). Die Ernte der
Versuchspflanzen, das Bestimmen des AM-Infektionsgrades sowie die Mineralstoffanalysen
erfolgten nach den in Kapitel 2.4, 2.5 und 2.6 beschriebenen Methoden.

Kapitel 5 Inokulationsversuche 90
5.2.4 Einfluß einer AM-Inokulation bei einer erhöhten Ozonbelastung der Luft auf AM-Infektionsgrad, Nährstoffaufnahme und Wachs-tum von Reben
5.2.4.1 Pflanzenmaterial, Versuchsgefäße und Boden
Für den Ozonversuch wurden vorkultivierte Topfreben der Rebsorte Riesling [N 90] mit der
Unterlagssorte 125 AA [FR 25] verwendet. Ein Teil der Reben war mit Glomus sp. inokuliert
worden (Herstellung der Topfreben und Inokulation s. Kap. 4.2.2.). Nach der Anzuchtphase
unter Gewächshausbedingungen wurden die Topfreben mit den Kartonagehüllen am
30.05.1995 in 5 l fassende Plastiktöpfe („Teku“, Fa. Pöppelmann, Hannover) getopft. Jedes
Gefäß wurde mit einer Rebe bepflanzt. Als Versuchsboden wurde der in Kapitel 5.2.1 be-
schriebene Rebschulboden verwendet. Jeder Topf enthielt 4,1 kg lufttrockenen Boden. Vor
dem Topfen wurde dem Boden 5 % Inokulum (Glomus sp., s. Kap. 2.1) zugegeben und
durchmischt, um eine gleichmäßige Verteilung des Inokulums im Boden zu erreichen. Die
nicht mykorrhizierten Varianten (Kontrolle) erhielten die gleiche Menge Inokulum von nicht
infizierten Tagetespflanzen. Pro Variante wurden 36 Töpfe getopft. Beim Einfüllen in die Ge-
fäße hatte der Versuchsboden einen Wassergehalt von 11 %. Die Gefäße wurden je nach Be-
darf mit Leitungswasser gegossen. Am 08.06.1995 und 26.06.1995 erfolgte eine N-Düngung
in Form von Calciumnitrat (Ca(NO3)2) mit jeweils 0,1 g N/Gefäß. Gegen Echten Mehltau
(Uncinula necator (Schwein.) Burr.) wurde zweimal Bayfidan spezial WG [Triadimenol, 0,05
%ig]
eingesetzt.
5.2.4.2 Versuchsaufbau und Versuchsdurchführung
Bei Versuchsbeginn verfügten die Reben über vier voll entwickelte Blätter bei einer durch-
schnittlichen Trieblänge von etwa 30 cm. In der Zeit vom 03.07. - 20.07.1995 fand die eigent-
liche Versuchsphase statt. Hierzu wurden die Reben in UV-durchlässige Plexiglaskammern
mit einem Volumen von 2,2 m3 gestellt (s. bei BERAN, 1994 a). Der den Begasungskammern
zugeführte Luftstrom wurde vor Eintritt in die Kammer durch Kohlefilter (Norit) schadstoff-
frei gemacht. Anschließend wurde das von Generatoren erzeugte Ozon zudosiert und das
Luftgemisch in den Innenraum geleitet (vgl. Abb. 5.2). Die Einheitlichkeit der Schadstoffbe-

Kapitel 5 Inokulationsversuche 91
lastung in den Versuchskammern wurde durch einen 30fachen Luftwechsel/h gewährleistet.
Die Windgeschwindigkeit in den Kammern betrug 0,75 m/s. Das Klima der im Freien befind-
lichen Begasungskammern folgte mehr oder weniger dem natürlichen Verlauf und wurde kon-
tinuierlich aufgezeichnet (s. Abb. 5.3). Kontrollmessungen des Ozongehaltes (Geräte von Fa.
Dasibi) wurden stündlich in allen Kammern sowie in drei Meter Höhe oberhalb der Bega-
sungsanlage während des gesamten Versuchszeitraumes durchgeführt.
Abb. 5.2: Schematischer Aufbau einer Begasungskammer
Für den Ozonversuch standen drei Kammern zur Verfügung, in denen jeweils sechs Reben mit
und ohne Mykorrhiza (-AM/+AM) standen (vgl. Abb. 5.2). In der ersten Kammer wurden die
Reben an vier Tagen (Montag - Donnerstag)/Woche vier Stunden lang jeweils von 9 - 13oo
Uhr mit einer Ozonkonzentration von circa 250 µg/m3 begast („Ozon“). Nach dieser Bega-
sungsphase wurde die Kammer mit schadstofffreier gefilterter Außenluft versorgt. In die
zweite Kammer wurde immer schadstofffreie Außenluft geleitet („Außenluft gereinigt“). Die
Reben in der dritten Kammer wurden mit ungereinigter Außenluft mit natürlich vorkommen-
der Schadstoffbelastung der Luft versorgt („Außenluft“). Wöchentlich wurde einmal die
Trieblänge bestimmt. Vier Tage nach Beenden der Begasungen wurden die Pflanzen nach der
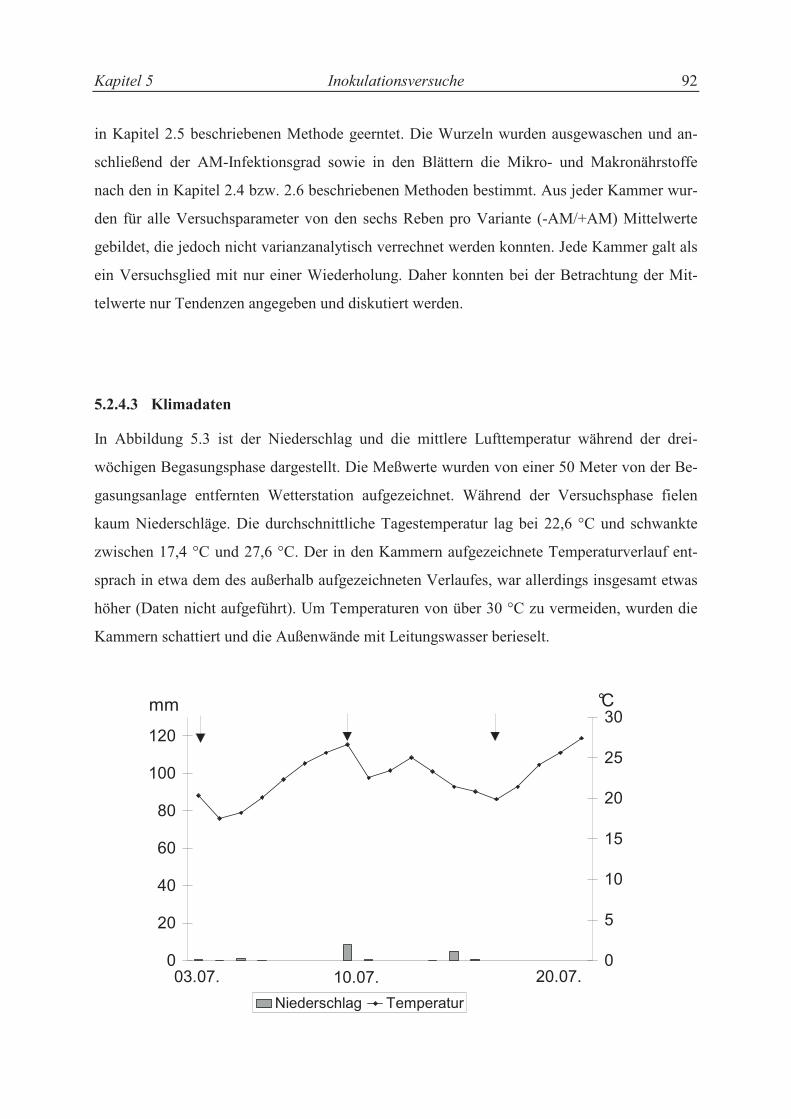
Kapitel 5 Inokulationsversuche 92
in Kapitel 2.5 beschriebenen Methode geerntet. Die Wurzeln wurden ausgewaschen und an-
schließend der AM-Infektionsgrad sowie in den Blättern die Mikro- und Makronährstoffe
nach den in Kapitel 2.4 bzw. 2.6 beschriebenen Methoden bestimmt. Aus jeder Kammer wur-
den für alle Versuchsparameter von den sechs Reben pro Variante (-AM/+AM) Mittelwerte
gebildet, die jedoch nicht varianzanalytisch verrechnet werden konnten. Jede Kammer galt als
ein Versuchsglied mit nur einer Wiederholung. Daher konnten bei der Betrachtung der Mit-
telwerte nur Tendenzen angegeben und diskutiert werden.
5.2.4.3 Klimadaten
In Abbildung 5.3 ist der Niederschlag und die mittlere Lufttemperatur während der drei-
wöchigen Begasungsphase dargestellt. Die Meßwerte wurden von einer 50 Meter von der Be-
gasungsanlage entfernten Wetterstation aufgezeichnet. Während der Versuchsphase fielen
kaum Niederschläge. Die durchschnittliche Tagestemperatur lag bei 22,6 °C und schwankte
zwischen 17,4 °C und 27,6 °C. Der in den Kammern aufgezeichnete Temperaturverlauf ent-
sprach in etwa dem des außerhalb aufgezeichneten Verlaufes, war allerdings insgesamt etwas
höher (Daten nicht aufgeführt). Um Temperaturen von über 30 °C zu vermeiden, wurden die
Kammern schattiert und die Außenwände mit Leitungswasser berieselt.
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
mm
0
5
10
15
20
25
30°C
Niederschlag Temperatur
03.07. 10.07. 20.07.

Kapitel 5 Inokulationsversuche 93
Abb. 5.3: Niederschlag und mittlere Lufttemperatur (in 2 m Höhe) während der 3wöchigen Versuchsphase (03.07. - 20.07.1995) an der Begasungsanlage
Markierungen zeigen jeweiligen Wochenbeginn
5.3 Ergebnisse
5.3.1 Einfluß der nativen Mykorrhiza eines Rebschulbodens auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünsteck-lingen unterschiedlichen Genotyps
5.3.1.1 AM-Infektionsgrad
Die Mykorrhizainfektion im nicht sterilisierten Boden wurde durch die bodeneigene Pilzflora
im verwendeten Rebschulboden hervorgerufen und betrug, gemittelt über die Sorten, 12 %
(s. Tab. 5.4). Die Sortenunterschiede variierten zwischen 6,8 % bei der Unterlagssorte A9 und
17,4 % bei der Unterlagssorte Börner. Nach der Sterilisation des Bodens konnte bei keiner
Sorte eine AM-Infektion festgestellt werden (s. Tab. 5.4). In einem Vorversuch wurde die
Beimpfung mit einem AM-Pilz (Glomus sp.) in einem nicht sterilisierten und sterilisierten
Boden überprüft. Dabei traten, ähnlich wie im vorliegenden Versuch, durch die native Mykor-
rhiza signifikante Sortenunterschiede im AM-Infektionsgrad zwischen den einzelnen Sorten
auf.
Tab. 5.4: AM-Infektionsgrad von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte) in sterilisiertem (-AM) und nicht sterilisiertem Boden (+AM = native Mykorrhiza aus einem Rebschulboden); Versuchsdauer 28.07. - 27.09.1994. (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Scheffé-Test, p≤0,05], Mittelwerte, n=5)
Sorten Behandlungen
sterilisierter Boden nicht sterilisierter Bo-den
-AM +AM
Riesling 0 12,2 ab
Börner 0 17,4 a
C3309 0 10,3 ab
5BB 0 11,1 ab
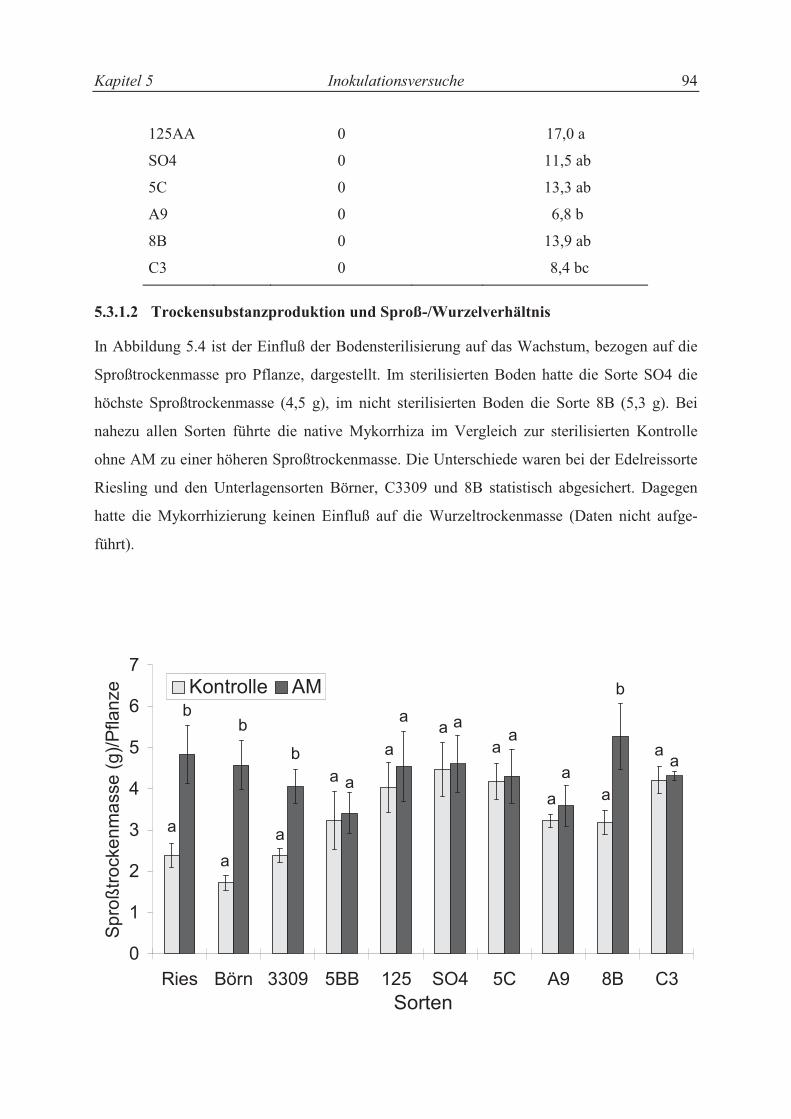
Kapitel 5 Inokulationsversuche 94
125AA 0 17,0 a
SO4 0 11,5 ab
5C 0 13,3 ab
A9 0 6,8 b
8B 0 13,9 ab
C3 0 8,4 bc
5.3.1.2 Trockensubstanzproduktion und Sproß-/Wurzelverhältnis
In Abbildung 5.4 ist der Einfluß der Bodensterilisierung auf das Wachstum, bezogen auf die
Sproßtrockenmasse pro Pflanze, dargestellt. Im sterilisierten Boden hatte die Sorte SO4 die
höchste Sproßtrockenmasse (4,5 g), im nicht sterilisierten Boden die Sorte 8B (5,3 g). Bei
nahezu allen Sorten führte die native Mykorrhiza im Vergleich zur sterilisierten Kontrolle
ohne AM zu einer höheren Sproßtrockenmasse. Die Unterschiede waren bei der Edelreissorte
Riesling und den Unterlagensorten Börner, C3309 und 8B statistisch abgesichert. Dagegen
hatte die Mykorrhizierung keinen Einfluß auf die Wurzeltrockenmasse (Daten nicht aufge-
führt).
a
a
a
a
a
a
a
a a
a
a
b
bb a a
a
a
b
a
0
1
2
3
4
5
6
7
Ries Börn 3309 5BB 125 SO4 5C A9 8B C3
Sorten
Sp
roß
tro
cke
nm
asse
(g
)/P
fla
nze Kontrolle AM
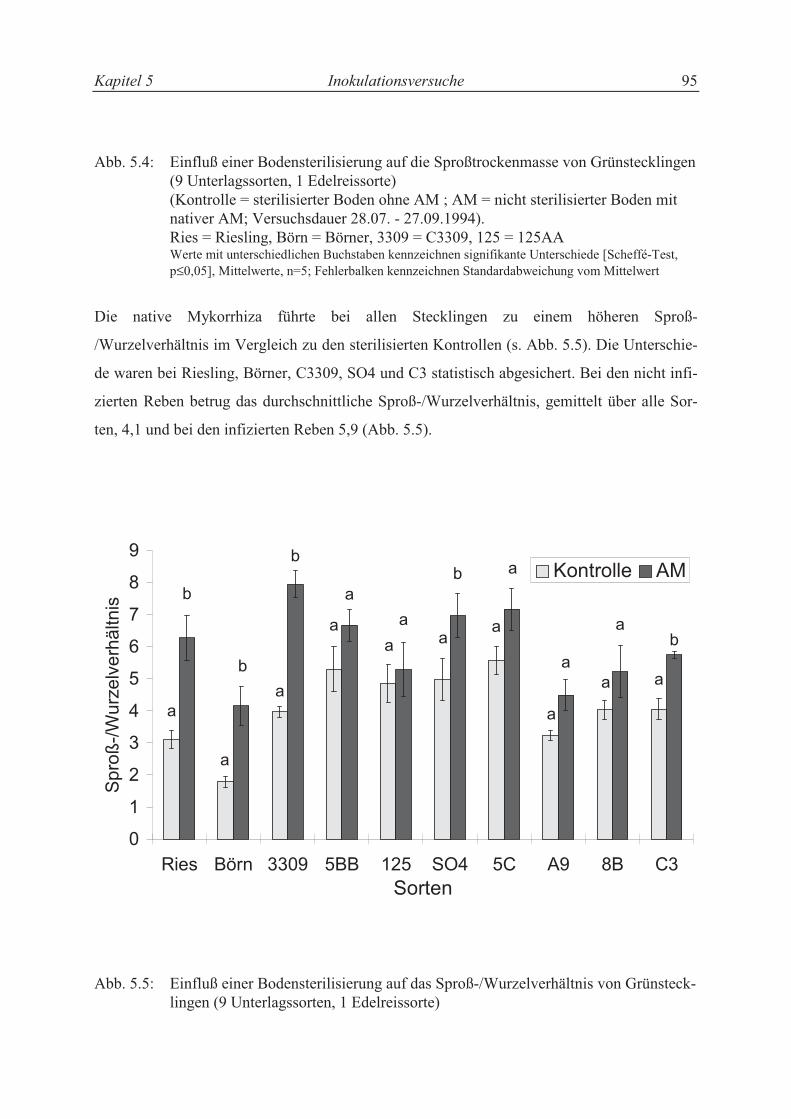
Kapitel 5 Inokulationsversuche 95
Abb. 5.4: Einfluß einer Bodensterilisierung auf die Sproßtrockenmasse von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte) (Kontrolle = sterilisierter Boden ohne AM ; AM = nicht sterilisierter Boden mit
nativer AM; Versuchsdauer 28.07. - 27.09.1994). Ries = Riesling, Börn = Börner, 3309 = C3309, 125 = 125AA
Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Scheffé-Test, p≤0,05], Mittelwerte, n=5; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert
Die native Mykorrhiza führte bei allen Stecklingen zu einem höheren Sproß-
/Wurzelverhältnis im Vergleich zu den sterilisierten Kontrollen (s. Abb. 5.5). Die Unterschie-
de waren bei Riesling, Börner, C3309, SO4 und C3 statistisch abgesichert. Bei den nicht infi-
zierten Reben betrug das durchschnittliche Sproß-/Wurzelverhältnis, gemittelt über alle Sor-
ten, 4,1 und bei den infizierten Reben 5,9 (Abb. 5.5).
aa
a
aaa
a
a
a
ab
a
a
ab
a
b
b
b
a
0
1
2
3
4
5
6
7
8
9
Ries Börn 3309 5BB 125 SO4 5C A9 8B C3
Sorten
Sp
roß
-/W
urz
elv
erh
ältn
is
Kontrolle AM
Abb. 5.5: Einfluß einer Bodensterilisierung auf das Sproß-/Wurzelverhältnis von Grünsteck-lingen (9 Unterlagssorten, 1 Edelreissorte)

Kapitel 5 Inokulationsversuche 96
(Kontrolle = sterilisierter Boden ohne AM; AM = nicht sterilisierter Boden mit nativer AM); Versuchsdauer 28.07. - 27.09.1994).
Ries = Riesling, Börn = Börner, 3309 = C3309, 125 = 125AA Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Scheffé-Test, p≤0,05], Mittelwerte, n=5; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert

Kapitel 5 Inokulationsversuche 97
5.3.1.3 Mineralstoffgehalte
Im folgenden Teil wird der Einfluß der nativen Mykorrhiza auf Makro- und Mikronährstoff-
gehalte in der Sproßfraktion „alte Blätter“ vorgestellt. Die Ergebnisse über die Wirkung der
nativen AM auf Makro- und Mikronährstoffe in den „jungen Blättern“, Blattstielen und der
Sproßachse sind im Anhang aufgelistet (s. Tab. A6, A7, A8, A9, A10 und A11).
Die native Mykorrhiza führte bei allen Sorten zu höheren P-Gehalten in den „alten Blättern“
(s. Tab. 5.5). Die Unterschiede waren bei den Unterlagssorten Börner, C3309, SO4, 8B und
C3 statistisch abgesichert. Dieser Effekt war in den „jungen Blättern“, Blattstielen und der
Sproßachse ebenfalls deutlich sichtbar (s. Tab. A6, A8 und A10). Die höchsten P-Gehalte
traten mit 3,5 mg/g Trockensubstanz (TS), gemittelt über alle Sorten, in den Blattstielen auf,
die geringsten P-Gehalte waren mit 0,8 mg/g TS in der Sproßachse vorzufinden (s. Tab. A8
und A10). Die Mg-Gehalte in den „alten Blättern“ lagen zwischen 2,3 mg/g TS bei der Unter-
lagssorte SO4 im sterilisierten Boden (-AM) und 3,7 mg/g TS bei C3309, ebenfalls im sterili-
sierten Boden (s. Tab. 5.5). Zwischen den nicht mykorrhizierten und den mykorrhizierten Re-
ben traten keine signifikanten Unterschiede im Mg-Gehalt auf. Die Ca-Gehalte waren sowohl
im sterilisierten Boden ohne AM (-AM) als auch im nicht sterilisierten Boden mit AM (+AM)
bei der Unterlagssorte C3309 mit 24,6 bzw. 26,0 mg/g TS am höchsten (s. Tab. 5.5). Die My-
korrhizierung hatte keinen Einfluß auf die Ca-Gehalte in den „alten Blättern“. Dagegen führte
die Mykorrhizierung in den Sproßfraktionen „junge Blätter“, Blattstiele und Sproßachse bei
einigen Sorten zu signifikant niedrigeren Ca-Gehalten in der Trockensubstanz (vgl. Tab. A6,
A8 und A10). Bei den Sorten Börner, 8B und C3 führte die native Mykorrhiza zu höheren K-
Gehalten in den „alten Blättern“ im Vergleich zu den Varianten ohne Mykorrhiza (s. Tab.
5.5). Dieser Unterschied zeigte sich ebenfalls in den „jungen Blättern“, der Sproßachse und
den Blattstielen (vgl. Tab. A6, A8 und A10). In den Blattstielen waren die K-Gehalte, unab-
hängig von der Mykorrhizierung, deutlich höher als in den „älteren Blättern“ (s. Tab. A8).
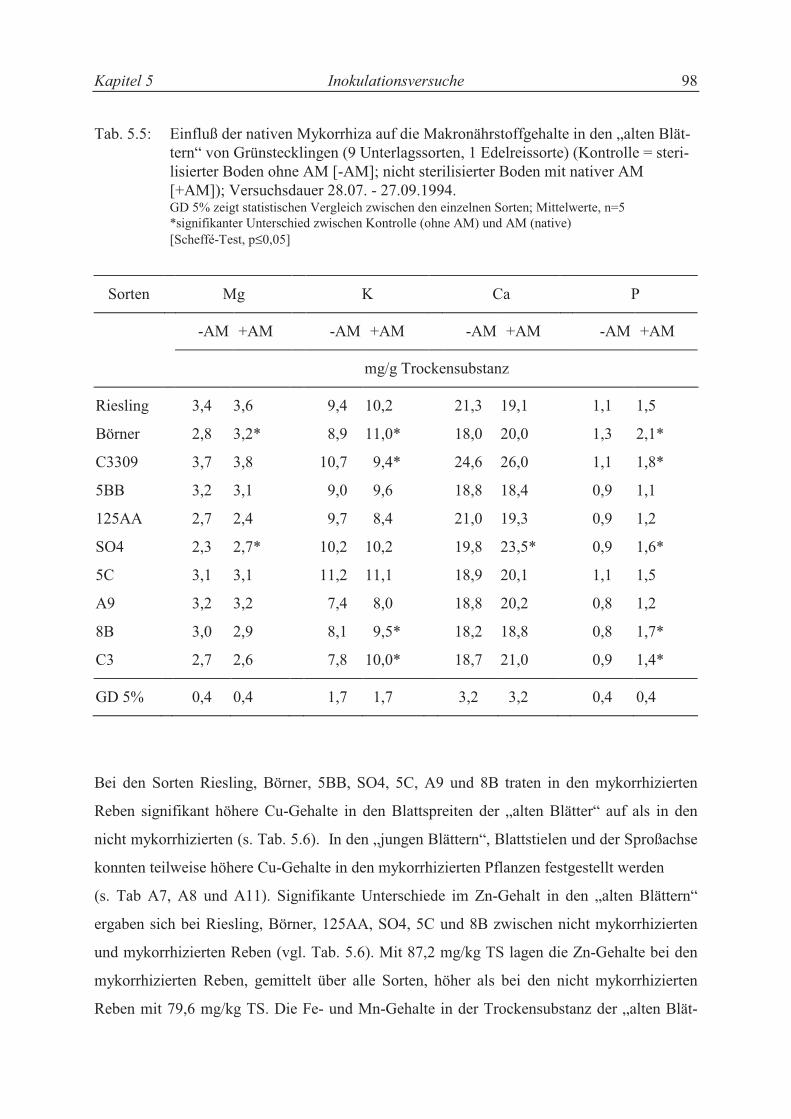
Kapitel 5 Inokulationsversuche 98
Tab. 5.5: Einfluß der nativen Mykorrhiza auf die Makronährstoffgehalte in den „alten Blät-tern“ von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte) (Kontrolle = steri-lisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994.
GD 5% zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Mg K Ca P
-AM +AM -AM +AM -AM +AM -AM +AM
mg/g Trockensubstanz
Riesling 3,4 3,6 9,4 10,2 21,3 19,1 1,1 1,5
Börner 2,8 3,2* 8,9 11,0* 18,0 20,0 1,3 2,1*
C3309 3,7 3,8 10,7 9,4* 24,6 26,0 1,1 1,8*
5BB 3,2 3,1 9,0 9,6 18,8 18,4 0,9 1,1
125AA 2,7 2,4 9,7 8,4 21,0 19,3 0,9 1,2
SO4 2,3 2,7* 10,2 10,2 19,8 23,5* 0,9 1,6*
5C 3,1 3,1 11,2 11,1 18,9 20,1 1,1 1,5
A9 3,2 3,2 7,4 8,0 18,8 20,2 0,8 1,2
8B 3,0 2,9 8,1 9,5* 18,2 18,8 0,8 1,7*
C3 2,7 2,6 7,8 10,0* 18,7 21,0 0,9 1,4*
GD 5% 0,4 0,4 1,7 1,7 3,2 3,2 0,4 0,4
Bei den Sorten Riesling, Börner, 5BB, SO4, 5C, A9 und 8B traten in den mykorrhizierten
Reben signifikant höhere Cu-Gehalte in den Blattspreiten der „alten Blätter“ auf als in den
nicht mykorrhizierten (s. Tab. 5.6). In den „jungen Blättern“, Blattstielen und der Sproßachse
konnten teilweise höhere Cu-Gehalte in den mykorrhizierten Pflanzen festgestellt werden
(s. Tab A7, A8 und A11). Signifikante Unterschiede im Zn-Gehalt in den „alten Blättern“
ergaben sich bei Riesling, Börner, 125AA, SO4, 5C und 8B zwischen nicht mykorrhizierten
und mykorrhizierten Reben (vgl. Tab. 5.6). Mit 87,2 mg/kg TS lagen die Zn-Gehalte bei den
mykorrhizierten Reben, gemittelt über alle Sorten, höher als bei den nicht mykorrhizierten
Reben mit 79,6 mg/kg TS. Die Fe- und Mn-Gehalte in der Trockensubstanz der „alten Blät-
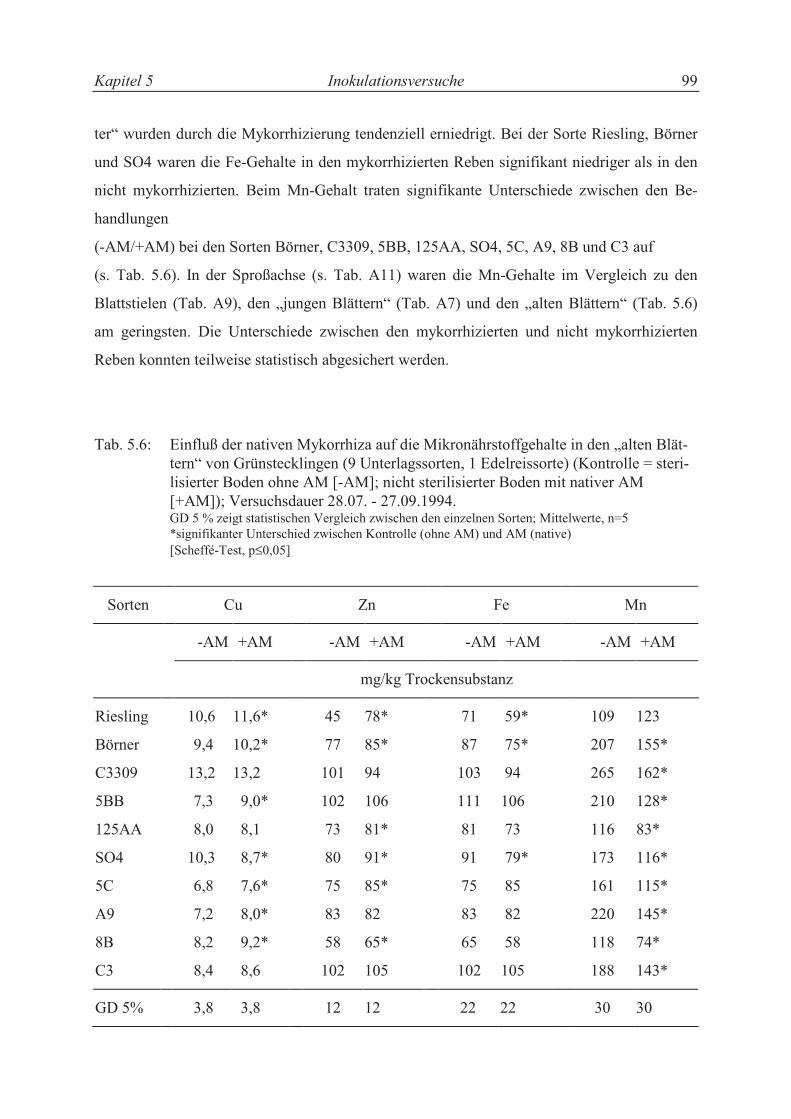
Kapitel 5 Inokulationsversuche 99
ter“ wurden durch die Mykorrhizierung tendenziell erniedrigt. Bei der Sorte Riesling, Börner
und SO4 waren die Fe-Gehalte in den mykorrhizierten Reben signifikant niedriger als in den
nicht mykorrhizierten. Beim Mn-Gehalt traten signifikante Unterschiede zwischen den Be-
handlungen
(-AM/+AM) bei den Sorten Börner, C3309, 5BB, 125AA, SO4, 5C, A9, 8B und C3 auf
(s. Tab. 5.6). In der Sproßachse (s. Tab. A11) waren die Mn-Gehalte im Vergleich zu den
Blattstielen (Tab. A9), den „jungen Blättern“ (Tab. A7) und den „alten Blättern“ (Tab. 5.6)
am geringsten. Die Unterschiede zwischen den mykorrhizierten und nicht mykorrhizierten
Reben konnten teilweise statistisch abgesichert werden.
Tab. 5.6: Einfluß der nativen Mykorrhiza auf die Mikronährstoffgehalte in den „alten Blät-tern“ von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte) (Kontrolle = steri-lisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994.
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Cu Zn Fe Mn
-AM +AM -AM +AM -AM +AM -AM +AM
mg/kg Trockensubstanz
Riesling 10,6 11,6* 45 78* 71 59* 109 123
Börner 9,4 10,2* 77 85* 87 75* 207 155*
C3309 13,2 13,2 101 94 103 94 265 162*
5BB 7,3 9,0* 102 106 111 106 210 128*
125AA 8,0 8,1 73 81* 81 73 116 83*
SO4 10,3 8,7* 80 91* 91 79* 173 116*
5C 6,8 7,6* 75 85* 75 85 161 115*
A9 7,2 8,0* 83 82 83 82 220 145*
8B 8,2 9,2* 58 65* 65 58 118 74*
C3 8,4 8,6 102 105 102 105 188 143*
GD 5% 3,8 3,8 12 12 22 22 30 30

Kapitel 5 Inokulationsversuche 100
5.3.2 Einfluß unterschiedlicher Inokulationstiefen mit Glomus mosseae
auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von Grünstecklingen in Wurzelbeobachtungskästen
5.3.2.1 Entwicklung und Verlauf des AM-Infektionsgrades im Versuchsgefäß
Nach 4wöchiger Wachstumsphase betrug der AM-Infektionsgrad der Rebwurzeln im unmit-
telbaren Inokulationsbereich (9 - 18 cm, „oben“ bzw. 36 - 45 cm, „unten“) 30 % bzw. 17 %
(s. Abb. 5.6). Die Rebwurzeln im angrenzenden Bodenbereich waren sowohl oberhalb als
auch unterhalb des Inokulumbandes bei beiden Inokulationsbereichen („oben“, „unten“) deut-
lich geringer infiziert. In der Versuchsvariante mit dem Inokulationsband „oben“ wurde ab
einer Tiefe von 36 cm keine Mykorrhizainfektion an den Rebwurzeln festgestellt. Beim Ino-
kulationsbereich „unten“ waren die Rebwurzeln bis zu einer Tiefe von 27 cm nicht mykorrhi-
ziert (vgl. Abb. 5.6).
Nach 6wöchiger Wachstumsphase waren die Rebwurzeln in beiden Versuchsvarianten deut-
lich stärker mykorrhiziert. Im unmittelbaren Inokulationsbereich (9 - 18 cm, „oben“ bzw.
36 - 45 cm „unten“) betrug der Infektionsgrad 45 % bzw. 35 %. Die Wurzeln in den angren-
zenden Bodenbereichen waren, ähnlich wie nach vier Wochen Wachstum, geringer mykorrhi-
ziert. Ab einer Tiefe von 36 cm konnte bei der Versuchsvariante „oben“ keine Mykorrhizain-
fektion an den Rebwurzeln festgestellt werden (s. Abb. 5.6). In der Versuchsvariante „unten“
wurde, ähnlich wie nach 4wöchiger Wachstumsphase, in den oberen 18 cm keine Mykorrhi-
zainfektion beobachtet. In den Kontrollvarianten ohne AM-Pilzinokulum wurde bei beiden
Inokulationsvarianten zu keinem Erntetermin eine AM-Infektion ermittelt.
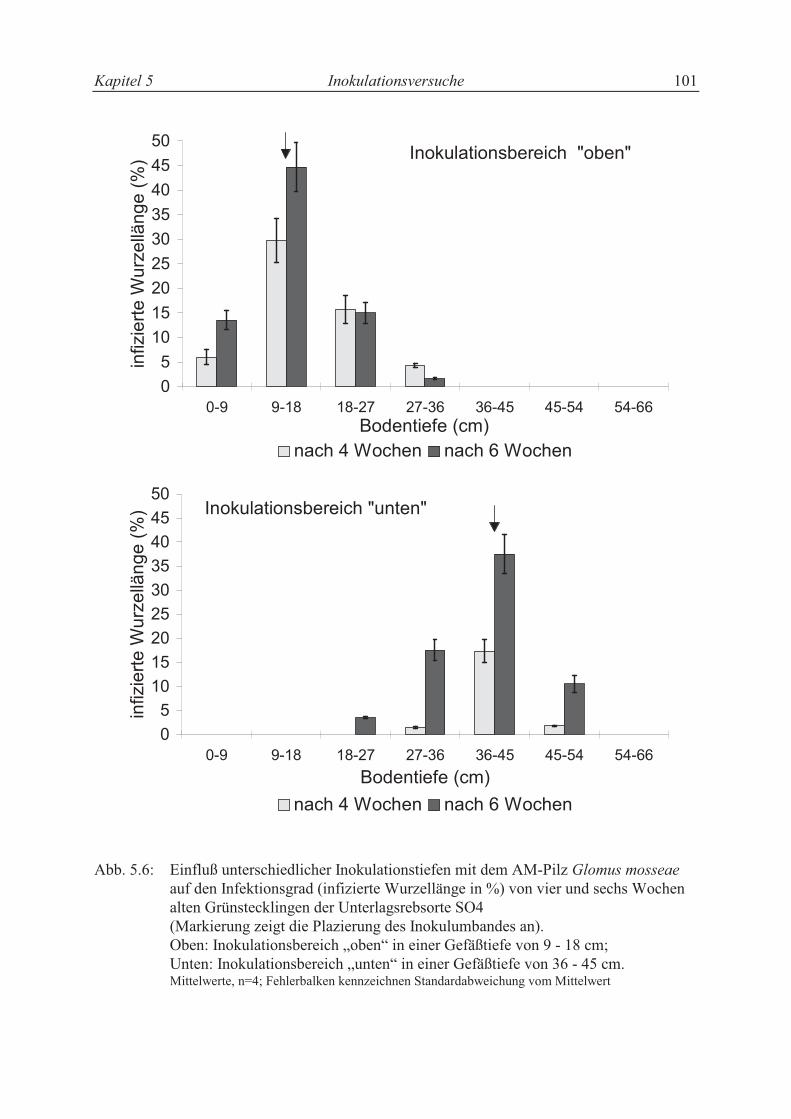
Kapitel 5 Inokulationsversuche 101
0
5
10
15
20
25
30
35
40
45
50
0-9 9-18 18-27 27-36 36-45 45-54 54-66
infizie
rte
Wu
rze
llän
ge
(%
)
nach 4 Wochen nach 6 Wochen
Inokulationsbereich "oben"
Bodentiefe (cm)
0
5
10
15
20
25
30
35
40
45
50
0-9 9-18 18-27 27-36 36-45 45-54 54-66
Bodentiefe (cm)
infizie
rte
Wu
rze
llän
ge
(%
)
nach 4 Wochen nach 6 Wochen
Inokulationsbereich "unten"
Abb. 5.6: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf den Infektionsgrad (infizierte Wurzellänge in %) von vier und sechs Wochen alten Grünstecklingen der Unterlagsrebsorte SO4
(Markierung zeigt die Plazierung des Inokulumbandes an). Oben: Inokulationsbereich „oben“ in einer Gefäßtiefe von 9 - 18 cm; Unten: Inokulationsbereich „unten“ in einer Gefäßtiefe von 36 - 45 cm. Mittelwerte, n=4; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert
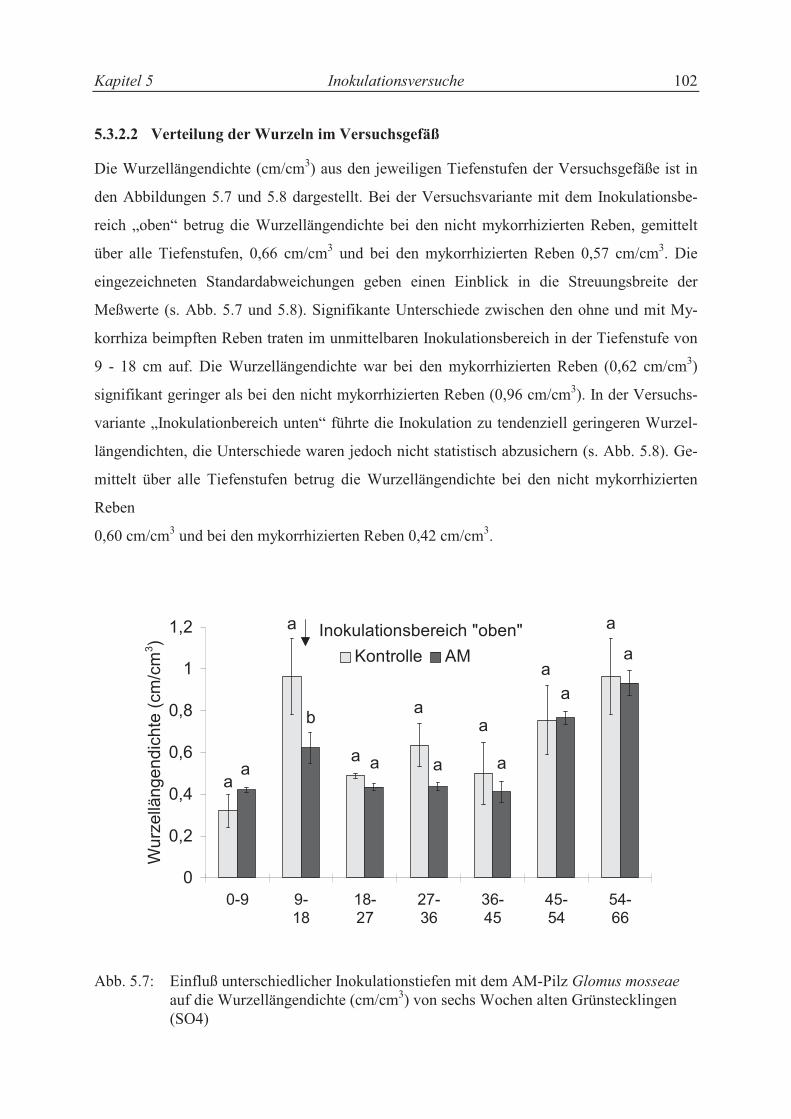
Kapitel 5 Inokulationsversuche 102
5.3.2.2 Verteilung der Wurzeln im Versuchsgefäß
Die Wurzellängendichte (cm/cm3) aus den jeweiligen Tiefenstufen der Versuchsgefäße ist in
den Abbildungen 5.7 und 5.8 dargestellt. Bei der Versuchsvariante mit dem Inokulationsbe-
reich „oben“ betrug die Wurzellängendichte bei den nicht mykorrhizierten Reben, gemittelt
über alle Tiefenstufen, 0,66 cm/cm3 und bei den mykorrhizierten Reben 0,57 cm/cm3. Die
eingezeichneten Standardabweichungen geben einen Einblick in die Streuungsbreite der
Meßwerte (s. Abb. 5.7 und 5.8). Signifikante Unterschiede zwischen den ohne und mit My-
korrhiza beimpften Reben traten im unmittelbaren Inokulationsbereich in der Tiefenstufe von
9 - 18 cm auf. Die Wurzellängendichte war bei den mykorrhizierten Reben (0,62 cm/cm3)
signifikant geringer als bei den nicht mykorrhizierten Reben (0,96 cm/cm3). In der Versuchs-
variante „Inokulationbereich unten“ führte die Inokulation zu tendenziell geringeren Wurzel-
längendichten, die Unterschiede waren jedoch nicht statistisch abzusichern (s. Abb. 5.8). Ge-
mittelt über alle Tiefenstufen betrug die Wurzellängendichte bei den nicht mykorrhizierten
Reben
0,60 cm/cm3 und bei den mykorrhizierten Reben 0,42 cm/cm3.
a
a
a
a
a
aa
a
a
aa
b
a a
0
0,2
0,4
0,6
0,8
1
1,2
0-9 9-18
18-27
27-36
36-45
45-54
54-66
Wu
rze
llän
ge
nd
ich
te (
cm
/cm
3)
Kontrolle AM
Inokulationsbereich "oben"
Abb. 5.7: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf die Wurzellängendichte (cm/cm3) von sechs Wochen alten Grünstecklingen (SO4)
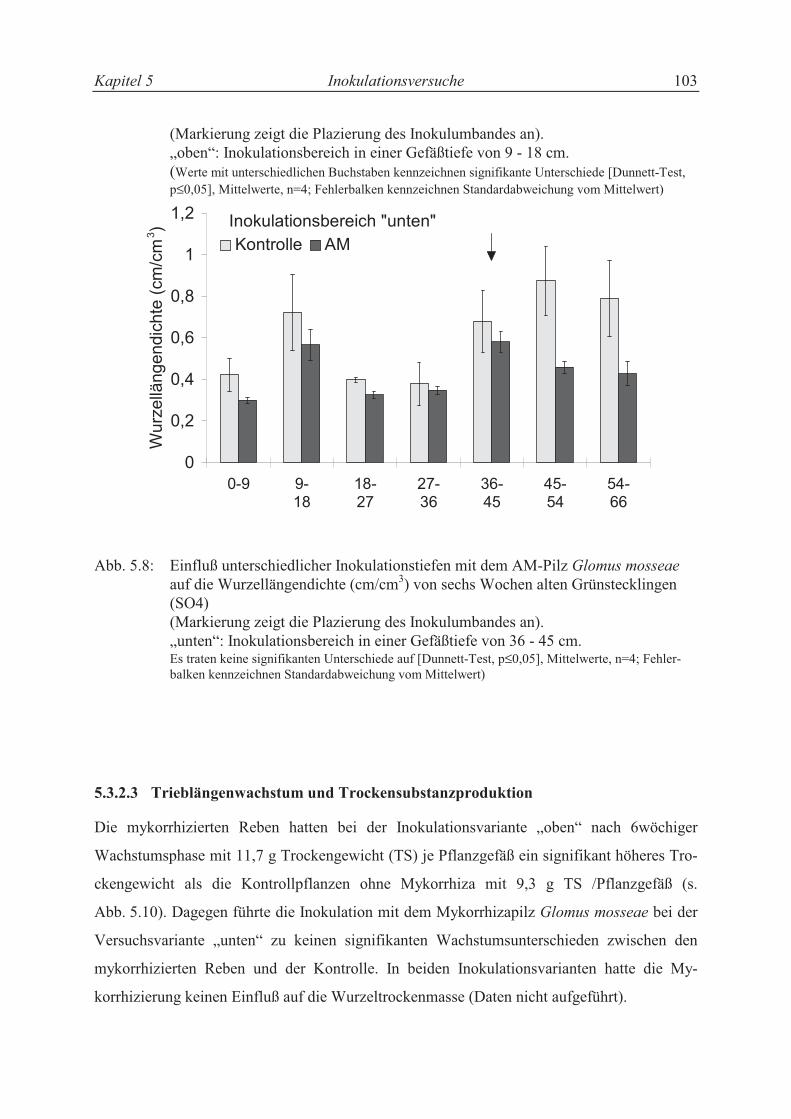
Kapitel 5 Inokulationsversuche 103
(Markierung zeigt die Plazierung des Inokulumbandes an). „oben“: Inokulationsbereich in einer Gefäßtiefe von 9 - 18 cm. (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test,
p≤0,05], Mittelwerte, n=4; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
0
0,2
0,4
0,6
0,8
1
1,2
0-9 9-18
18-27
27-36
36-45
45-54
54-66
Wurz
ellä
ngendic
hte
(cm
/cm
3)
Kontrolle AM
Inokulationsbereich "unten"
Abb. 5.8: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf die Wurzellängendichte (cm/cm3) von sechs Wochen alten Grünstecklingen (SO4)
(Markierung zeigt die Plazierung des Inokulumbandes an). „unten“: Inokulationsbereich in einer Gefäßtiefe von 36 - 45 cm.
Es traten keine signifikanten Unterschiede auf [Dunnett-Test, p≤0,05], Mittelwerte, n=4; Fehler-balken kennzeichnen Standardabweichung vom Mittelwert)
5.3.2.3 Trieblängenwachstum und Trockensubstanzproduktion
Die mykorrhizierten Reben hatten bei der Inokulationsvariante „oben“ nach 6wöchiger
Wachstumsphase mit 11,7 g Trockengewicht (TS) je Pflanzgefäß ein signifikant höheres Tro-
ckengewicht als die Kontrollpflanzen ohne Mykorrhiza mit 9,3 g TS /Pflanzgefäß (s.
Abb. 5.10). Dagegen führte die Inokulation mit dem Mykorrhizapilz Glomus mosseae bei der
Versuchsvariante „unten“ zu keinen signifikanten Wachstumsunterschieden zwischen den
mykorrhizierten Reben und der Kontrolle. In beiden Inokulationsvarianten hatte die My-
korrhizierung keinen Einfluß auf die Wurzeltrockenmasse (Daten nicht aufgeführt).

Kapitel 5 Inokulationsversuche 104
In Abbildung 5.9 ist der Sproßzustand der Grünstecklinge vier Wochen nach der Inokulation
mit dem AM-Pilz Glomus mosseae bei der Inokulationsvariante „oben“ dargestellt. Die Be-
impfung führte sechs Wochen nach Versuchsbeginn zu signifikant längeren Trieben an den
Stecklingen im Vergleich zu den Kontrollen.
Abb. 5.9: Sproßzustand von Grünstecklingen (SO4) in Wurzelbeobachtungskästen unter Gewächshausbedingungen drei Wochen nach Inokulation mit dem AM-Pilz Glo-
mus mosseae (Inokulationsbereich „oben“) rechts: ohne Mykorrhiza [-AM]; links: mit Mykorrhiza [+AM]
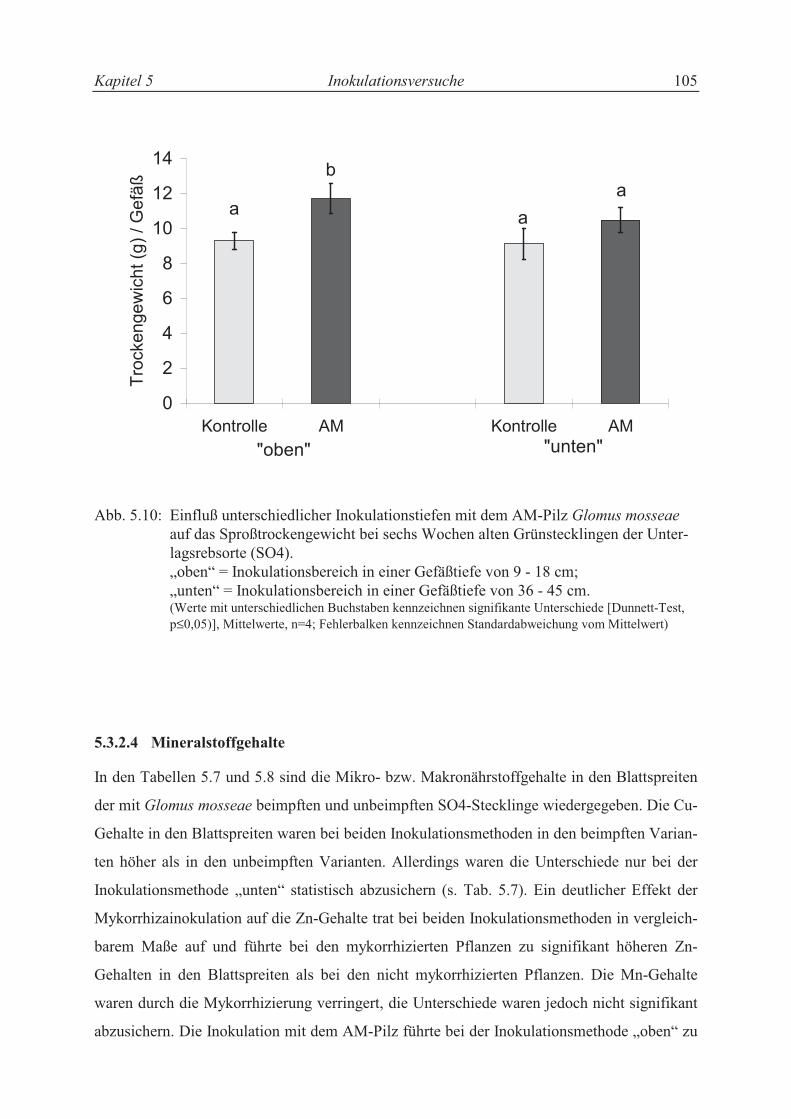
Kapitel 5 Inokulationsversuche 105
"unten"
a
b
a
a
0
2
4
6
8
10
12
14
Kontrolle AM Kontrolle AM
"oben"
Tro
cke
ng
ew
ich
t (g
) / G
efä
ß
Abb. 5.10: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf das Sproßtrockengewicht bei sechs Wochen alten Grünstecklingen der Unter-lagsrebsorte (SO4).
„oben“ = Inokulationsbereich in einer Gefäßtiefe von 9 - 18 cm; „unten“ = Inokulationsbereich in einer Gefäßtiefe von 36 - 45 cm. (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test,
p≤0,05)], Mittelwerte, n=4; Fehlerbalken kennzeichnen Standardabweichung vom Mittelwert)
5.3.2.4 Mineralstoffgehalte
In den Tabellen 5.7 und 5.8 sind die Mikro- bzw. Makronährstoffgehalte in den Blattspreiten
der mit Glomus mosseae beimpften und unbeimpften SO4-Stecklinge wiedergegeben. Die Cu-
Gehalte in den Blattspreiten waren bei beiden Inokulationsmethoden in den beimpften Varian-
ten höher als in den unbeimpften Varianten. Allerdings waren die Unterschiede nur bei der
Inokulationsmethode „unten“ statistisch abzusichern (s. Tab. 5.7). Ein deutlicher Effekt der
Mykorrhizainokulation auf die Zn-Gehalte trat bei beiden Inokulationsmethoden in vergleich-
barem Maße auf und führte bei den mykorrhizierten Pflanzen zu signifikant höheren Zn-
Gehalten in den Blattspreiten als bei den nicht mykorrhizierten Pflanzen. Die Mn-Gehalte
waren durch die Mykorrhizierung verringert, die Unterschiede waren jedoch nicht signifikant
abzusichern. Die Inokulation mit dem AM-Pilz führte bei der Inokulationsmethode „oben“ zu
Kapitel 5 Inokulationsversuche 106
signifikant höheren P-Gehalten in den Blattspreiten (s. Tab. 5.8). Bei den Gehalten von K, Ca
und Fe traten zwischen den nicht infizierten und infizierten Reben bei beiden Inokulationsme-
thoden keine signifikanten Unterschiede auf.
Tab. 5.7: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf die Mikronährstoffgehalte in der Trockensubstanz (TS) der Blattspreiten bei sechs Wochen alten Grünstecklingen der Unterlagsrebsorte SO4
„oben“ = Inokulationsbereich in einer Gefäßtiefe von 9 - 18 cm; „unten“ = Inokulationsbereich in einer Gefäßtiefe von 36 - 45 cm. (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test,
p≤0,05], Mittelwerte ± SD, n=4)
Inokulati-ons-tiefe
Behandlung Cu Zn Fe Mn
(mg/kg TS)
„oben“ - AM 14,3 ± 1,0 a 33,1 ± 3,4 a 144 ± 11,1 a 150 ± 11,4 a + AM 15,0 ± 1,9 a 40,4 ± 8,7 b 151 ± 9,3 a 136 ± 9,1 a
„unten“ - AM 13,6 ± 0,8 a 30,8 ± 0,9 a 146 ± 10,5 a 211 ± 37,6 a + AM 17,1 ± 1,4 b 34,5 ± 5,2 b 139 ± 10,2 a 185 ± 28,5 a
Tab. 5.8: Einfluß unterschiedlicher Inokulationstiefen mit dem AM-Pilz Glomus mosseae
auf die Makronährstoffgehalte in der Trockensubstanz (TS) der Blattspreiten bei sechs Wochen alten Grünstecklingen der Unterlagsrebsorte SO4
„oben“ = Inokulationsbereich in einer Gefäßtiefe von 9 - 18 cm; „unten“ = Inokulationsbereich in einer Gefäßtiefe von 36 - 45 cm. (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Dunnett-Test,
p≤0,05], Mittelwerte ± SD, n=4)
Inokulations-tiefe
Behandlung Mg K Ca P
(mg/g TS)
„oben“ - AM 2,8 ± 0,1 a 12,5 ± 1,0 a 16,8 ± 0,8 a 2,1 ± 0,1 a + AM 2,6 ± 0,2 a 13,2 ± 0,4 a 15,9 ± 1,7 a 2,5 ± 0,2 b
„unten“ - AM 2,9 ± 0,1 a 13,2 ± 1,0 a 17,5 ± 2,1 a 2,4 ± 0,4 a

Kapitel 5 Inokulationsversuche 107
+ AM 2,7 ± 0,1 a 13,2 ± 0,7 a 17,6 ± 0,7 a 2,2 ± 0,2 a
5.3.3 Einfluß einer Inokulation mit Glomus intraradices auf AM-Infektionsgrad, Nährstoffaufnahme und Wachstum von grün- veredelten Stecklingen
5.3.3.1 AM-Infektionsgrad
In Abbildung 5.11 sind Teile einer Wurzel der Unterlagsrebsorte SO4 mit einer intensiven
Besiedlung mit dem AM-Pilz Glomus intraradices dargestellt, wobei die Pilzstrukturen wie
Vesikel und Hyphen nach Färbung mit Trypanblau deutlich sichtbar wurden. Zwischen den
einzelnen Pflanzenvarianten traten zum Teil signifikante Unterschiede im AM-Infektionsgrad
auf
(s. Tab. 5.9). Der höchste Infektionsgrad wurde in der wurzelechten Variante mit Riesling, der
niedrigste in der heteroplastischen Pfropfkombination mit Riesling/SO4 festgestellt. In den
Kontrollvarianten ohne AM-Pilzinokulum wurde keine Mykorrhizainfektion beobachtet.
Abb. 5.11: AM-Infektion einer Rebwurzel der Unterlagsrebsorte SO4 mit Vesikeln und Hy-phen nach Färbung mit Trypanblau (Vergrößerung x 50)

Kapitel 5 Inokulationsversuche 108
Tab. 5.9: AM-Infektionsgrad von Grünstecklingen der Sorten SO4 und Riesling (hetero- bzw. homoplastische Pfropfungen und wurzelecht) beimpft mit dem AM-Pilz Glomus intraradices;
Versuchsdauer 11.06. - 25.08.1997 (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede [Scheffé-Test,
p≤0,05], Mittelwerte ± SD, n=6)
Variante AM-Infektionsgrad
(infizierte Wurzellänge %)
Riesling 38,5 ± 4,04 a SO4 29,6 ± 2,80 ab Riesling/SO4 26,2 ± 2,99 b Riesling/Riesling 32,1 ± 4,79 ab SO4/SO4 30,0 ± 3,89 ab
5.3.3.2 Wachstumsparameter und Sproß-/Wurzelverhältnis
Die Wurzellängendichte wurde durch die Pfropfung sowohl in den nicht mit Glomus intrara-
dices inokulierten Pflanzenvarianten als auch den inokulierten Varianten nicht beeinflußt
(s. Tab. 5.10). Dagegen führte die Mykorrhizierung in allen Pflanzenvarianten zu einer niedri-
geren Wurzellängendichte. Die Unterschiede zwischen Kontrolle (-AM) und beimpften Vari-
anten (+AM) waren bei den wurzelechten Varianten Riesling und SO4 signifikant (Tab. 5.10).
Tab. 5.10: Wurzellängendichte (cm/cm3) von Grünstecklingen der Sorten SO4 und Riesling (hetero- bzw. homoplastische Pfropfungen und wurzelecht) ohne (-AM) und mit (+AM) Beimpfung mit dem AM-Pilz Glomus intraradices;
Versuchsdauer 11.06. - 25.08.1997 *signifikanter Unterschied zwischen unbeimpfter und mit G. intraradices beimpfter Variante
[Scheffé-Test, p≤0,05])
Variante Wurzellängendichte Abnahme bei +AM
(cm/cm3) (%) -AM +AM
Riesling 3,30 2,75 -16,7* SO4 3,58 3,00 -16,2* Riesling/SO4 2,97 2,83 - 4,7 Riesling/Riesling 2,98 2,52 -15,4
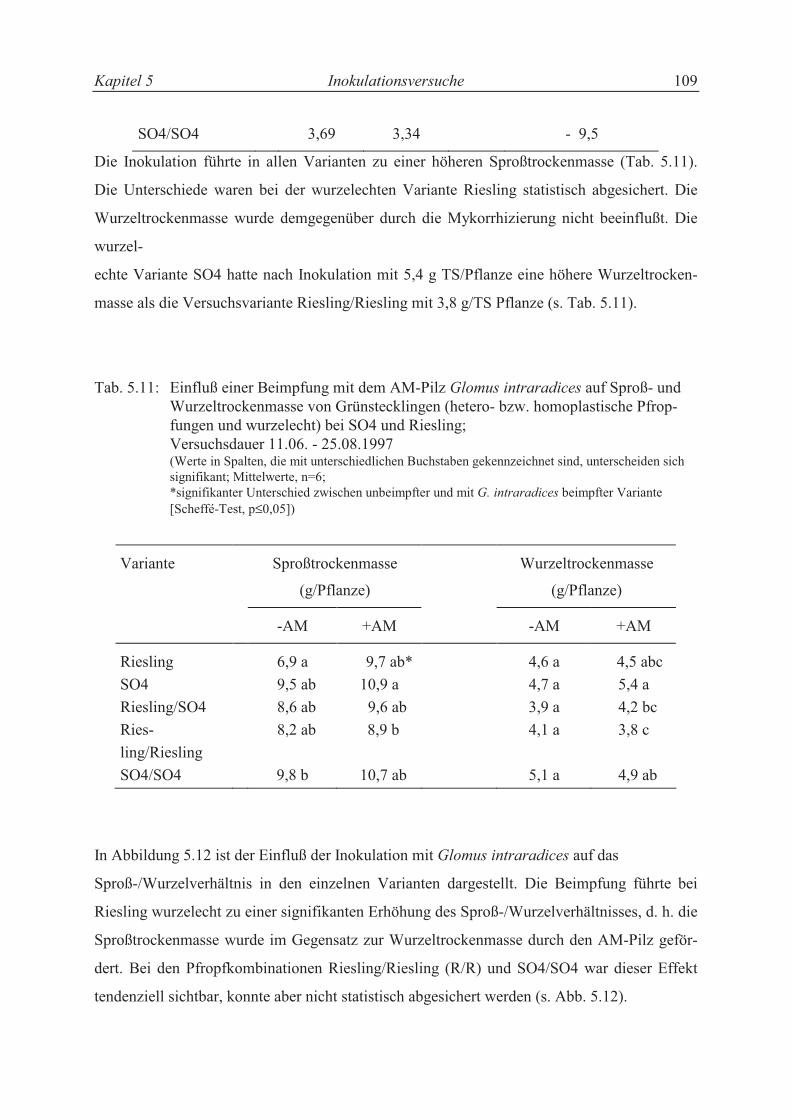
Kapitel 5 Inokulationsversuche 109
SO4/SO4 3,69 3,34 - 9,5
Die Inokulation führte in allen Varianten zu einer höheren Sproßtrockenmasse (Tab. 5.11).
Die Unterschiede waren bei der wurzelechten Variante Riesling statistisch abgesichert. Die
Wurzeltrockenmasse wurde demgegenüber durch die Mykorrhizierung nicht beeinflußt. Die
wurzel-
echte Variante SO4 hatte nach Inokulation mit 5,4 g TS/Pflanze eine höhere Wurzeltrocken-
masse als die Versuchsvariante Riesling/Riesling mit 3,8 g/TS Pflanze (s. Tab. 5.11).
Tab. 5.11: Einfluß einer Beimpfung mit dem AM-Pilz Glomus intraradices auf Sproß- und Wurzeltrockenmasse von Grünstecklingen (hetero- bzw. homoplastische Pfrop-fungen und wurzelecht) bei SO4 und Riesling;
Versuchsdauer 11.06. - 25.08.1997 (Werte in Spalten, die mit unterschiedlichen Buchstaben gekennzeichnet sind, unterscheiden sich signifikant; Mittelwerte, n=6;
*signifikanter Unterschied zwischen unbeimpfter und mit G. intraradices beimpfter Variante [Scheffé-Test, p≤0,05])
Variante Sproßtrockenmasse
(g/Pflanze)
Wurzeltrockenmasse
(g/Pflanze)
-AM +AM -AM +AM
Riesling 6,9 a 9,7 ab* 4,6 a 4,5 abc
SO4 9,5 ab 10,9 a 4,7 a 5,4 a
Riesling/SO4 8,6 ab 9,6 ab 3,9 a 4,2 bc
Ries-
ling/Riesling
8,2 ab 8,9 b 4,1 a 3,8 c
SO4/SO4 9,8 b 10,7 ab 5,1 a 4,9 ab
In Abbildung 5.12 ist der Einfluß der Inokulation mit Glomus intraradices auf das
Sproß-/Wurzelverhältnis in den einzelnen Varianten dargestellt. Die Beimpfung führte bei
Riesling wurzelecht zu einer signifikanten Erhöhung des Sproß-/Wurzelverhältnisses, d. h. die
Sproßtrockenmasse wurde im Gegensatz zur Wurzeltrockenmasse durch den AM-Pilz geför-
dert. Bei den Pfropfkombinationen Riesling/Riesling (R/R) und SO4/SO4 war dieser Effekt
tendenziell sichtbar, konnte aber nicht statistisch abgesichert werden (s. Abb. 5.12).
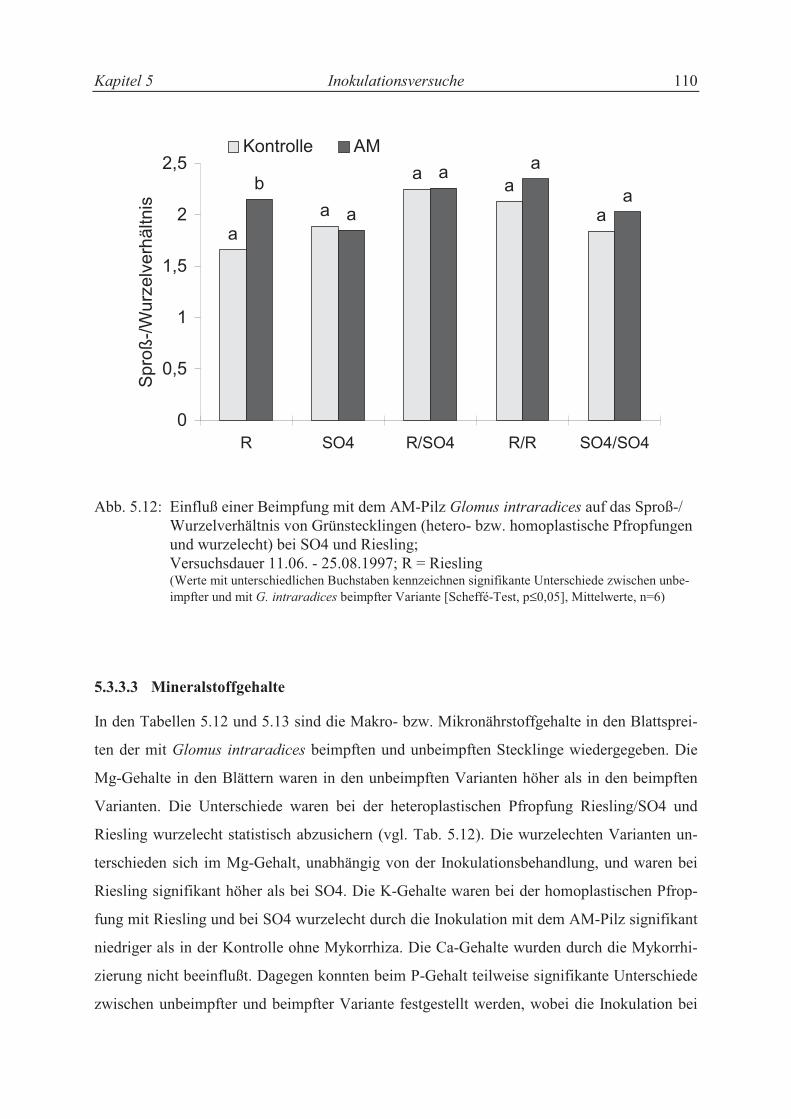
Kapitel 5 Inokulationsversuche 110
a
aa
a
a
a
aa
a
b
0
0,5
1
1,5
2
2,5
R SO4 R/SO4 R/R SO4/SO4
Sp
roß
-/W
urz
elv
erh
ältn
isKontrolle AM
Abb. 5.12: Einfluß einer Beimpfung mit dem AM-Pilz Glomus intraradices auf das Sproß-/ Wurzelverhältnis von Grünstecklingen (hetero- bzw. homoplastische Pfropfungen und wurzelecht) bei SO4 und Riesling;
Versuchsdauer 11.06. - 25.08.1997; R = Riesling (Werte mit unterschiedlichen Buchstaben kennzeichnen signifikante Unterschiede zwischen unbe-
impfter und mit G. intraradices beimpfter Variante [Scheffé-Test, p≤0,05], Mittelwerte, n=6)
5.3.3.3 Mineralstoffgehalte
In den Tabellen 5.12 und 5.13 sind die Makro- bzw. Mikronährstoffgehalte in den Blattsprei-
ten der mit Glomus intraradices beimpften und unbeimpften Stecklinge wiedergegeben. Die
Mg-Gehalte in den Blättern waren in den unbeimpften Varianten höher als in den beimpften
Varianten. Die Unterschiede waren bei der heteroplastischen Pfropfung Riesling/SO4 und
Riesling wurzelecht statistisch abzusichern (vgl. Tab. 5.12). Die wurzelechten Varianten un-
terschieden sich im Mg-Gehalt, unabhängig von der Inokulationsbehandlung, und waren bei
Riesling signifikant höher als bei SO4. Die K-Gehalte waren bei der homoplastischen Pfrop-
fung mit Riesling und bei SO4 wurzelecht durch die Inokulation mit dem AM-Pilz signifikant
niedriger als in der Kontrolle ohne Mykorrhiza. Die Ca-Gehalte wurden durch die Mykorrhi-
zierung nicht beeinflußt. Dagegen konnten beim P-Gehalt teilweise signifikante Unterschiede
zwischen unbeimpfter und beimpfter Variante festgestellt werden, wobei die Inokulation bei
Kapitel 5 Inokulationsversuche 111
Riesling mit 2,1 mg P/g TS zu höheren Werten führte als bei der Kontrolle mit 1,3 mg P/g TS.
Die Pfropfung hatte keinen Einfluß auf die P-Gehalte (s. Abb. 5.12).
Signifikante Unterschiede im Cu-Gehalt der Blätter ergaben sich bei Riesling wurzelecht und
bei SO4/SO4 zwischen unbeimpfter und beimpfter Variante (s. Tab. 5.13). Die Zn-Gehalte
waren in den mykorrhizierten Varianten Riesling, SO4/SO4 und Riesling/Riesling signifikant
höher als in den nicht mykorrhizierten. Beim Vergleich der wurzelechten Varianten Riesling
und SO4 waren die Zn-Gehalte, unabhängig von der Inokulation, bei Riesling höher als bei
SO4. Der Fe-Gehalt war bei Riesling wurzelecht durch die Beimpfung mit G. intraradices mit
174 mg Fe/kg Trockensubstanz (TS) niedriger als in der unbeimpften Variante mit
304 mg Fe/kg TS (s. Tab. 5.13). Beim Mn-Gehalt traten zwischen den nicht infizierten und
infizierten Reben keine signifikanten Unterschiede auf.
Tab. 5.12: Makronährstoffgehalte in den Blättern von Grünstecklingen der Sorten SO4 und Riesling (hetero- bzw. homoplastische Pfropfungen und wurzelecht) ohne (-AM) und mit (+AM) Beimpfung mit dem AM-Pilz Glomus intraradices;
Versuchsdauer 11.06. - 25.08.1997; R = Riesling (Werte in Spalten, die mit unterschiedlichen Buchstaben gekennzeichnet sind, unterscheiden sich
signifikant; Mittelwerte, n=6) *signifikanter Unterschied zwischen unbeimpfter und mit G. intraradices beimpfter Variante
[Scheffé-Test, p≤0,05]
Variante Mg K Ca P
-AM +AM -AM +AM -AM +AM -AM +AM
mg/g Trockensubstanz
R 5,4a 4,4a* 12,7ab 13,1ab 14,8a 14,3a 1,3a 2,1a*
SO4 2,9b 2,7b 12,5b 11,2c* 12,3bc 13,1ab 1,9a 2,0a
R/SO4 3,2b 2,8b* 14,9a 14,2a 13,1b 12,8ab 1,9a 1,9a
R/R 5,0a 4,6a 13,7ab 12,1bc* 13,1b 14,0a 1,9a 2,0a
SO4/SO4 2,8b 2,6b 11,7b 11,1c 11,6c 12,2b 1,8a 1,9a
Kapitel 5 Inokulationsversuche 112
Tab. 5.13: Mikronährstoffgehalte in den Blättern von Grünstecklingen der Sorten SO4 und Riesling (hetero- bzw. homoplastische Pfropfungen und wurzelecht) ohne (-AM) und mit (+AM) Beimpfung mit dem AM-Pilz Glomus intraradices;
Versuchsdauer 11.06. - 25.08.1997: R = Riesling (Werte in Spalten, die mit unterschiedlichen Buchstaben gekennzeichnet sind, unterscheiden sich
signifikant; Mittelwerte, n=6) *signifikanter Unterschied zwischen unbeimpfter und mit G. intraradices beimpfter Variante
[Scheffé-Test, p≤0,05]
Variante Cu Zn Fe Mn
-AM +AM -AM +AM -AM +AM -AM +AM
mg/kg Trockensubstanz
R 7,3b 12,1b* 31,5a 39,0a* 304a 174a* 243a 217ab
SO4 13,6a 14,9a 28,3b 32,0b 148c 158ab 193a 191ab
R/SO4 12,7a 13,1b 32,4a 32,2b 143c 134b 212a 222a
R/R 11,4a 13,1ab 31,5a 36,5ab* 244b 157ab 185a 209ab
SO4/SO4 14,2a 15,2a* 29,2ab 34,2ab* 154bc 156ab 170a 184b
5.3.4 Einfluß einer AM-Inokulation bei einer erhöhten Ozonbelastung der Luft auf AM-Infektionsgrad, Nährstoffaufnahme und Wachs-tum von Reben
5.3.4.1 AM-Infektionsgrad
Nach der Begasungsphase von drei Wochen lagen die AM-Infektionsgrade zwischen 20,5 %
(„Ozon“) und 23,3 % („Außenluft“) (vgl. Tab. 5.14). Vor dem Überführen der Kartonagere-
ben in die Pflanzgefäße lag der Infektionsgrad bei 23,4 % (s. Kap. 4.3.2) und war nach der
3wöchigen Versuchsphase nahezu unverändert. Die unterschiedlichen Ozonkonzentrationen
hatten keinen Einfluß auf den AM-Infektionsgrad der Rebwurzeln (s. Tab. 5.14). In den Kon-
trollvarianten ohne AM-Pilzinokulum wurde in keiner Kammer an den Reben eine AM-
Infektion beobachtet.

Kapitel 5 Inokulationsversuche 113
Tab. 5.14: Einfluß unterschiedlicher Ozonkonzentrationen (nach 3wöchiger Versuchsphase, 03.07. - 20.07.1995) auf den AM-Infektionsgrad (infizierte Wurzellänge %) in den Versuchsvarianten „Außenluft (gereinigt)“ [ = gereinigte Außenluft ohne Ozon], „Außenluft“ [= mit natürlichem Ozongehalt der Luft] und „Ozon“ [= 250 µg O3 /m
3 für 4 h an vier Tagen/Woche] Mittelwerte, n=6
Variante AM-Infektionsgrad
(infizierte Wurzellänge (%)
„Außenluft 20,6 (gereinigt)“
„Außenluft“ 23,3
„Ozon“ 20,5
5.3.4.2 Ozon-Konzentrationen
Durch die Kohlefilter wurde die Außenluft ohne Ozonbelastung schadstofffrei in die Kammer
(„Außenluft gereinigt“) geleitet. Innerhalb der Begasungszeit (9°°-13°° Uhr) wurde in der
Kammer mit Ozonbegasung („Ozon“) die angestrebte Ozonkonzentration von 250 µg O3/m3
in etwa eingehalten (vgl. Abb. 5.13). Die durchschnittliche Schwankung lag bei 9,4 %. In der
Kammer mit Außenluft wurde eine durchschnittliche Ozonkonzentration während der Bega-
sungszeit von 9°°-13°° Uhr von 36,7 µg O3/m3 ermittelt. Die niedrigste Ozonkonzentration
trat am 19.07.1995 auf und lag bei 19,9 µg O3/m3, die höchste O3-Konzentration wurde am
10.07.1995 gemessen und betrug 50,4 µg O3/m3 (s. Abb. 5.13). Die ermittelten Ozonkonzent-
rationen außerhalb der Versuchskammer in drei Meter Höhe lagen während des gesamten
Versuchszeitraumes deutlich über den Werten in der Kammer mit Außenluft (s. Abb. 5.13).
Nach dem Eintritt des Ozons in die Kammer nehmen die Reben über ihre Stomata O3 auf, so
daß die Konzentrationen innerhalb der Kammer niedriger ausfallen als außerhalb der Kam-
mer.
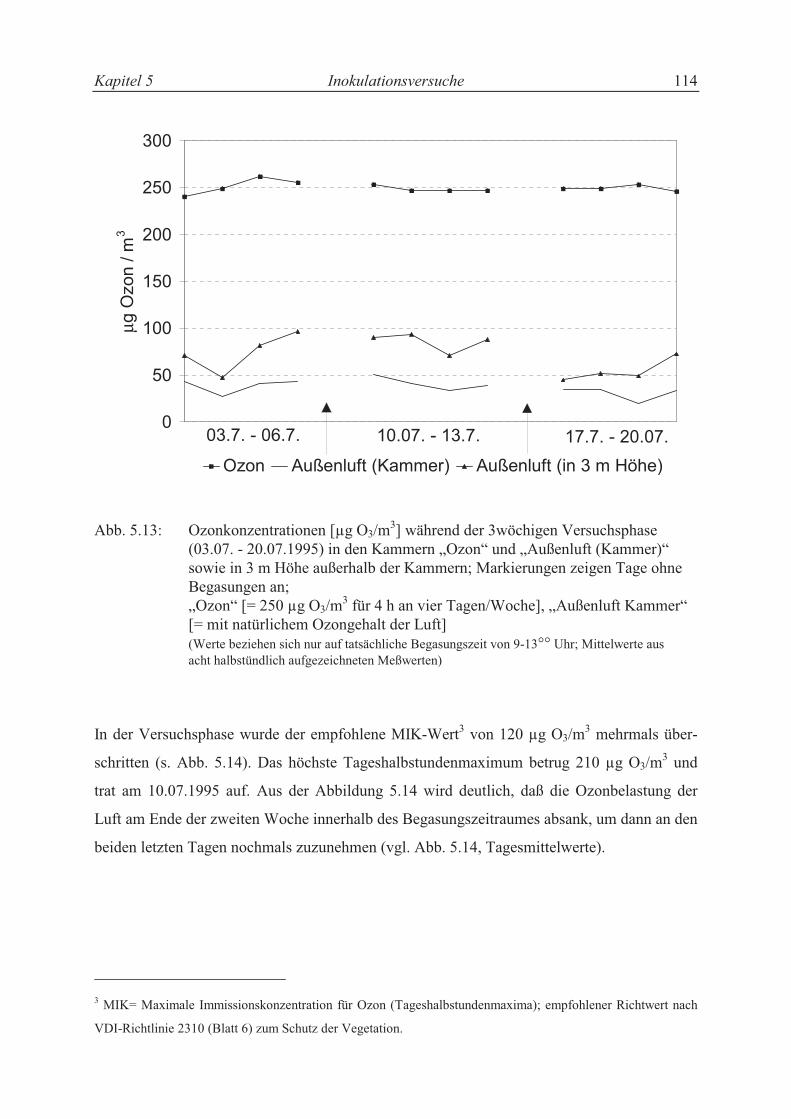
Kapitel 5 Inokulationsversuche 114
0
50
100
150
200
250
300µ
g O
zo
n / m
3
Ozon Außenluft (Kammer) Außenluft (in 3 m Höhe)
03.7. - 06.7. 10.07. - 13.7. 17.7. - 20.07.
Abb. 5.13: Ozonkonzentrationen [µg O3/m3] während der 3wöchigen Versuchsphase
(03.07. - 20.07.1995) in den Kammern „Ozon“ und „Außenluft (Kammer)“ sowie in 3 m Höhe außerhalb der Kammern; Markierungen zeigen Tage ohne Begasungen an;
„Ozon“ [= 250 µg O3/m3 für 4 h an vier Tagen/Woche], „Außenluft Kammer“
[= mit natürlichem Ozongehalt der Luft] (Werte beziehen sich nur auf tatsächliche Begasungszeit von 9-13°° Uhr; Mittelwerte aus
acht halbstündlich aufgezeichneten Meßwerten)
In der Versuchsphase wurde der empfohlene MIK-Wert3 von 120 µg O3/m3 mehrmals über-
schritten (s. Abb. 5.14). Das höchste Tageshalbstundenmaximum betrug 210 µg O3/m3 und
trat am 10.07.1995 auf. Aus der Abbildung 5.14 wird deutlich, daß die Ozonbelastung der
Luft am Ende der zweiten Woche innerhalb des Begasungszeitraumes absank, um dann an den
beiden letzten Tagen nochmals zuzunehmen (vgl. Abb. 5.14, Tagesmittelwerte).
3 MIK= Maximale Immissionskonzentration für Ozon (Tageshalbstundenmaxima); empfohlener Richtwert nach
VDI-Richtlinie 2310 (Blatt 6) zum Schutz der Vegetation.
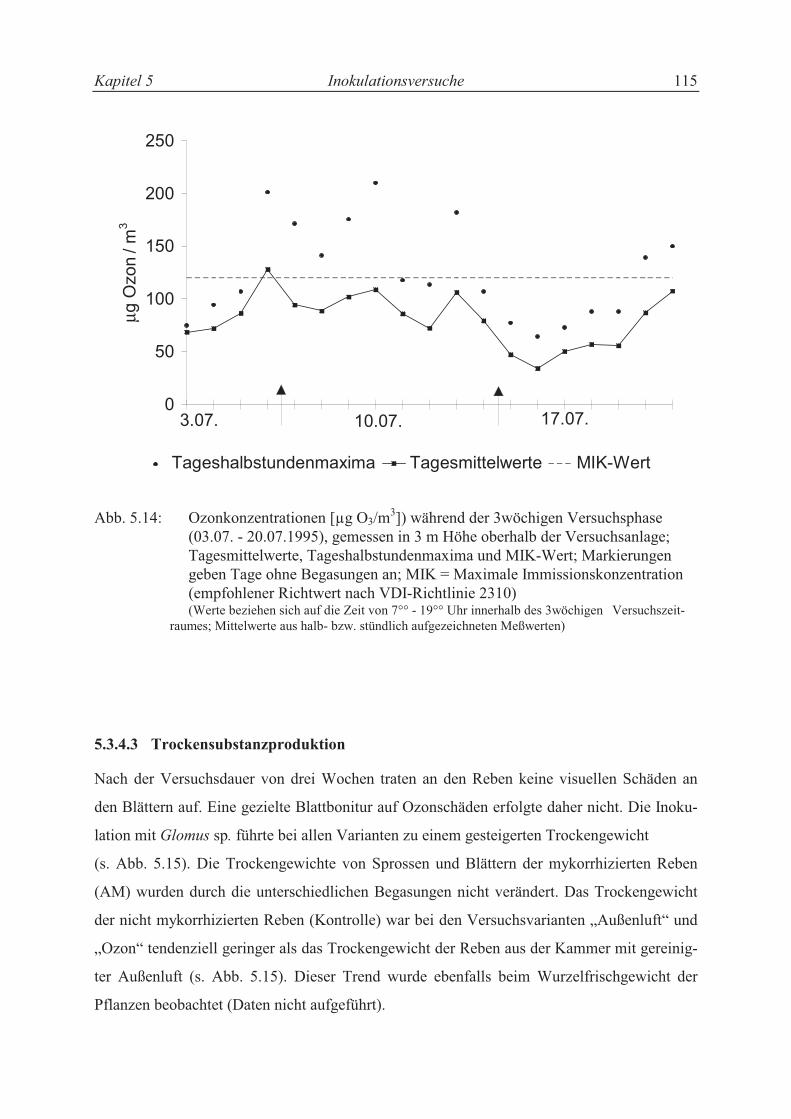
Kapitel 5 Inokulationsversuche 115
0
50
100
150
200
250µ
g O
zon
/ m
3
Tageshalbstundenmaxima Tagesmittelwerte MIK-Wert
3.07. 10.07. 17.07.
Abb. 5.14: Ozonkonzentrationen [µg O3/m3]) während der 3wöchigen Versuchsphase
(03.07. - 20.07.1995), gemessen in 3 m Höhe oberhalb der Versuchsanlage; Tagesmittelwerte, Tageshalbstundenmaxima und MIK-Wert; Markierungen geben Tage ohne Begasungen an; MIK = Maximale Immissionskonzentration (empfohlener Richtwert nach VDI-Richtlinie 2310)
(Werte beziehen sich auf die Zeit von 7°° - 19°° Uhr innerhalb des 3wöchigen Versuchszeit-raumes; Mittelwerte aus halb- bzw. stündlich aufgezeichneten Meßwerten)
5.3.4.3 Trockensubstanzproduktion
Nach der Versuchsdauer von drei Wochen traten an den Reben keine visuellen Schäden an
den Blättern auf. Eine gezielte Blattbonitur auf Ozonschäden erfolgte daher nicht. Die Inoku-
lation mit Glomus sp. führte bei allen Varianten zu einem gesteigerten Trockengewicht
(s. Abb. 5.15). Die Trockengewichte von Sprossen und Blättern der mykorrhizierten Reben
(AM) wurden durch die unterschiedlichen Begasungen nicht verändert. Das Trockengewicht
der nicht mykorrhizierten Reben (Kontrolle) war bei den Versuchsvarianten „Außenluft“ und
„Ozon“ tendenziell geringer als das Trockengewicht der Reben aus der Kammer mit gereinig-
ter Außenluft (s. Abb. 5.15). Dieser Trend wurde ebenfalls beim Wurzelfrischgewicht der
Pflanzen beobachtet (Daten nicht aufgeführt).
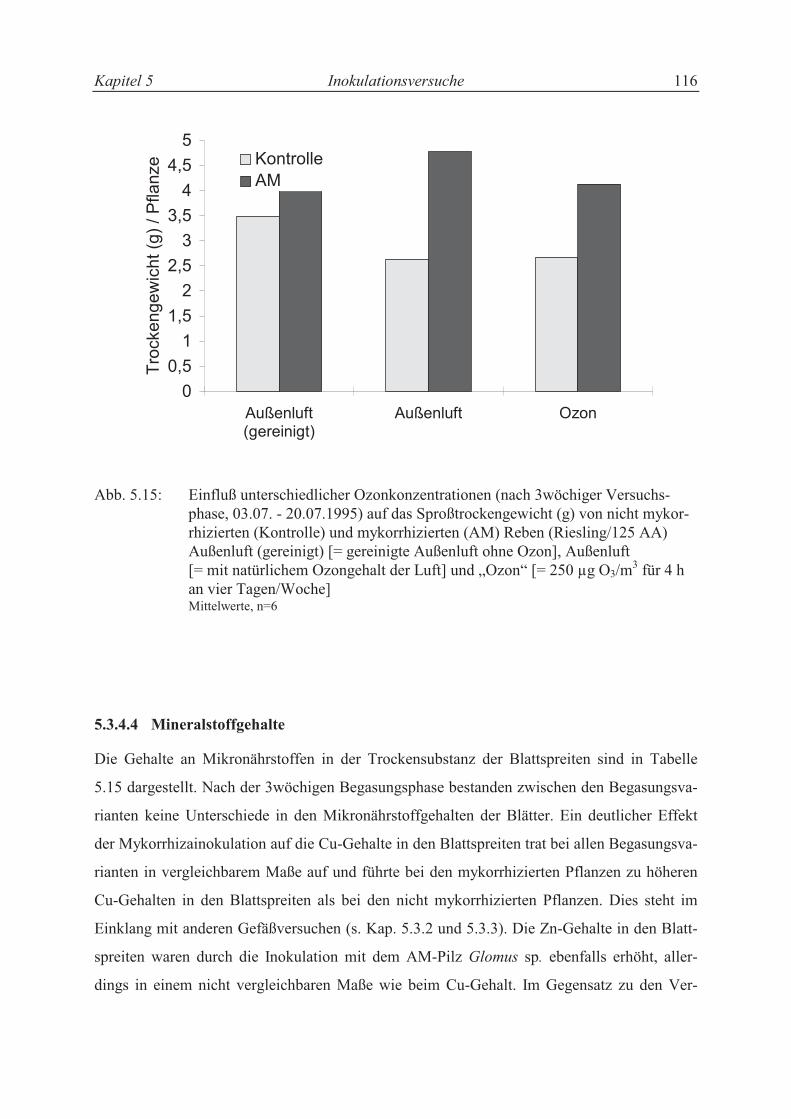
Kapitel 5 Inokulationsversuche 116
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
Außenluft(gereinigt)
Außenluft Ozon
Tro
cke
ng
ew
ich
t (g
) /
Pfla
nze Kontrolle
AM
Abb. 5.15: Einfluß unterschiedlicher Ozonkonzentrationen (nach 3wöchiger Versuchs- phase, 03.07. - 20.07.1995) auf das Sproßtrockengewicht (g) von nicht mykor- rhizierten (Kontrolle) und mykorrhizierten (AM) Reben (Riesling/125 AA)
Außenluft (gereinigt) [= gereinigte Außenluft ohne Ozon], Außenluft [= mit natürlichem Ozongehalt der Luft] und „Ozon“ [= 250 µg O3/m
3 für 4 h an vier Tagen/Woche]
Mittelwerte, n=6
5.3.4.4 Mineralstoffgehalte
Die Gehalte an Mikronährstoffen in der Trockensubstanz der Blattspreiten sind in Tabelle
5.15 dargestellt. Nach der 3wöchigen Begasungsphase bestanden zwischen den Begasungsva-
rianten keine Unterschiede in den Mikronährstoffgehalten der Blätter. Ein deutlicher Effekt
der Mykorrhizainokulation auf die Cu-Gehalte in den Blattspreiten trat bei allen Begasungsva-
rianten in vergleichbarem Maße auf und führte bei den mykorrhizierten Pflanzen zu höheren
Cu-Gehalten in den Blattspreiten als bei den nicht mykorrhizierten Pflanzen. Dies steht im
Einklang mit anderen Gefäßversuchen (s. Kap. 5.3.2 und 5.3.3). Die Zn-Gehalte in den Blatt-
spreiten waren durch die Inokulation mit dem AM-Pilz Glomus sp. ebenfalls erhöht, aller-
dings in einem nicht vergleichbaren Maße wie beim Cu-Gehalt. Im Gegensatz zu den Ver-

Kapitel 5 Inokulationsversuche 117
suchsergebnissen aus Kapitel 5.3.2 wurden die Mn-Gehalte durch die Mykorrhizierung nicht
signifikant vermindert.
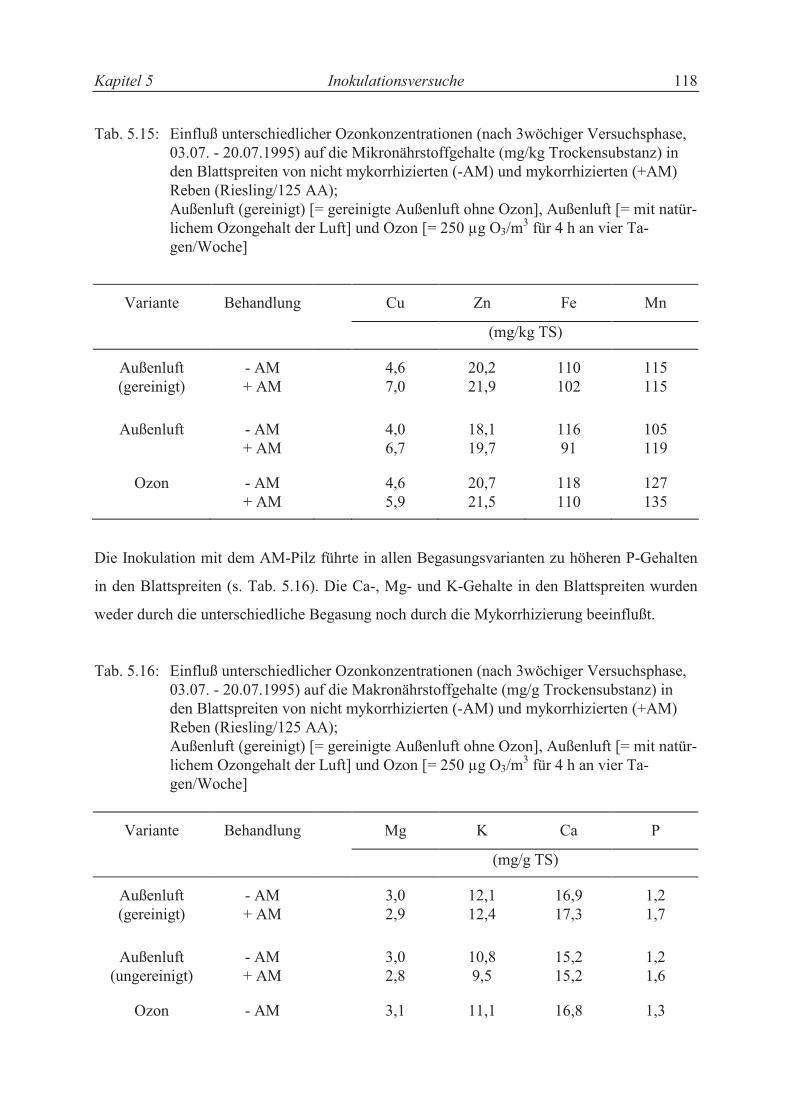
Kapitel 5 Inokulationsversuche 118
Tab. 5.15: Einfluß unterschiedlicher Ozonkonzentrationen (nach 3wöchiger Versuchsphase, 03.07. - 20.07.1995) auf die Mikronährstoffgehalte (mg/kg Trockensubstanz) in den Blattspreiten von nicht mykorrhizierten (-AM) und mykorrhizierten (+AM) Reben (Riesling/125 AA);
Außenluft (gereinigt) [= gereinigte Außenluft ohne Ozon], Außenluft [= mit natür-lichem Ozongehalt der Luft] und Ozon [= 250 µg O3/m
3 für 4 h an vier Ta-gen/Woche]
Variante Behandlung Cu Zn Fe Mn
(mg/kg TS)
Außenluft - AM 4,6 20,2 110 115 (gereinigt) + AM 7,0 21,9 102 115
Außenluft - AM 4,0 18,1 116 105 + AM 6,7 19,7 91 119
Ozon - AM 4,6 20,7 118 127 + AM 5,9 21,5 110 135
Die Inokulation mit dem AM-Pilz führte in allen Begasungsvarianten zu höheren P-Gehalten
in den Blattspreiten (s. Tab. 5.16). Die Ca-, Mg- und K-Gehalte in den Blattspreiten wurden
weder durch die unterschiedliche Begasung noch durch die Mykorrhizierung beeinflußt.
Tab. 5.16: Einfluß unterschiedlicher Ozonkonzentrationen (nach 3wöchiger Versuchsphase, 03.07. - 20.07.1995) auf die Makronährstoffgehalte (mg/g Trockensubstanz) in den Blattspreiten von nicht mykorrhizierten (-AM) und mykorrhizierten (+AM) Reben (Riesling/125 AA);
Außenluft (gereinigt) [= gereinigte Außenluft ohne Ozon], Außenluft [= mit natür-lichem Ozongehalt der Luft] und Ozon [= 250 µg O3/m
3 für 4 h an vier Ta-gen/Woche]
Variante Behandlung Mg K Ca P
(mg/g TS)
Außenluft - AM 3,0 12,1 16,9 1,2 (gereinigt) + AM 2,9 12,4 17,3 1,7
Außenluft - AM 3,0 10,8 15,2 1,2 (ungereinigt) + AM 2,8 9,5 15,2 1,6
Ozon - AM 3,1 11,1 16,8 1,3

Kapitel 5 Inokulationsversuche 119
+ AM 3,0 10,7 17,1 1,6
5.4 Diskussion
5.4.1 Nährstoffaufnahme und Wachstum der Rebenstecklinge
Ausgangspunkt der vorliegenden Arbeit war die Frage, inwieweit die arbuskuläre Mykorrhiza
die Nährstoffversorgung und das Wachstum der Rebe beeinflußt. In zahlreichen Untersuchun-
gen wurde nachgewiesen, daß AM-Pilze die Nährstoffgehalte verschiedener Pflanzenarten
insbesondere auf nährstoffarmen oder P-fixierenden Böden erhöhen oder erniedrigen können
(FABER et al., 1990; KOTHARI et al., 1990 b; LI et al., 1991 c; ARINES et al., 1992; MARSCH-
NER und DELL, 1994; DIAZ et al., 1996; MONZON und AZCÓN, 1996). In der vorliegenden Ar-
beit wurden Versuche mit Böden von Rebschulstandorten mit einer mittleren
(s. Tab. 5.2) bzw. hohen P-Versorgung (s. Tab. 5.1) durchgeführt, die anschließend mit Glo-
mus mosseae (Versuch mit Wurzelkästen, s. Kap. 5.2.2), Glomus intraradices (grünveredelte
Stecklinge, s. Kap. 5.2.3) und Glomus sp. (Ozonversuch, s. Kap. 5.2.4) inokuliert wurden.
Beim Versuch mit Grünstecklingen (s. 5.2.1) wurde die Hälfte des Versuchsbodens sterilisiert,
um die Wirkung der nativen Mykorrhiza eines Rebschulbodens zu überprüfen. Wie erwartet,
führte die Inokulation bei den meisten Reben zu einer erhöhten Trockenmassebildung, was
mit den Ergebnissen von Inokulationsversuchen an Reben aus der Literatur übereinstimmt
(EIBACH, 1982; KARAGIANNIDIS et al., 1995; BIRICOLTI et al., 1997). Bei optimaler bis hoher
Nährstoffversorgung des Bodens wird das Sproßwachstum von Pflanzen stärker gefördert als
das Wurzelwachstum, was in einem höheren Sproß-/Wurzelverhältnis zum Ausdruck kommt.
Bei einem ausgewogenen Nährstoffstatus der Wirtspflanze kann diese Eigenschaft bei my-
korrhizierten Pflanzen noch gesteigert werden (MARSCHNER, 1995). Dies konnte in der vor-
liegenden Untersuchung bestätigt werden. Beim Versuch mit grünveredelten Stecklingen (s.
Kap. 5.2.3, Tab. 5.11) sowie dem Inokulationsversuch mit Wurzelkästen
(s. Kap. 5.2.2) hatte die Mykorrhizierung keinen Einfluß auf die Wurzeltrockenmasse. Aller-
dings führte die Mykorrhizierung bei nahezu allen Grünstecklingen zu einem erhöhten Sproß-/
Wurzelverhältnis (s. Abb. 5.5), was ebenfalls bei der Beimpfung der grünveredelten Stecklin-
ge mit dem AM-Pilz Glomus intraradices tendenziell festgestellt werden konnte (s. Abb.
5.12). Dies stimmt mit den Ergebnissen von SKINNER et al. (1988) überein, die an verschiede-
nen Pfropfkombinationen mit der Edelreissorte Cabernet sauvignon und den Unterlagssorten
St. George (V. rupestris), AXR#1 (V. rupestris x V. vinifera) und 110Richter (V. rupestris x

Kapitel 5 Inokulationsversuche 120
V. berlandieri) nach Inokulation mit Glomus deserticola oder mit nativer Mykorrhiza eben-
falls ein höheres Sproß-/Wurzelverhältnis bei den mykorrhizierten Reben feststellten. Dabei
war der Effekt der bodeneigenen Mykorrhizapopulation sogar noch größer als bei den mit G.
deserticola inokulierten Reben. BAVARESCO und FOGHER (1996) fanden nach der Inokulation
mit dem AM-Pilz Glomus mosseae an Holzstecklingen der Pfropfkombinationen Pinot blanc
mit C3309 und 41B ebenfalls signifikant niedrigere Wurzeltrockengewichte. Dagegen stellten
die Autoren bei der Pfropfkombination mit der Unterlagssorte SO4 keine Veränderungen im
Wurzeltrockengewicht fest. In weiteren Untersuchungen wurde neben einer erhöhten Tro-
ckenmasseproduktion des Sprosses auch erhöhte Wurzeltrockengewichte nach Inokulation mit
AM-Pilzen an Apfelsämlingen (MATSUBARA et al., 1996) und Rebensämlingen mit der Unter-
lagssorte 420A
(V. berlandieri x V. riparia) festgestellt (SCHUBERT et al., 1988). In der vorliegenden Untersu-
chung wurden im Ozonversuch an den mykorrhizierten Rebenstecklingen ebenfalls höhere
Wurzeltrockenmassen festgestellt (nicht dargestellt). Möglicherweise wurde das Wurzel-
wachstum in beiden Versuchsvarianten (-AM/+AM) durch das Umtopfen der Kartonagereben
in größere Pflanzgefäße kurz vor Versuchsbeginn stimuliert, so daß die inokulierten Reben
tendenziell höhere Wurzeltrockenmassen aufwiesen und somit keine Unterschiede mehr er-
mittelt werden konnten.
Bei dem Versuch über die Ausbreitung der AM in Wurzelkästen war die Wurzellängendichte
im Inokulationsbereich signifikant niedriger als bei den nicht mykorrhizierten Reben
(s. Abb. 5.7). Dieser Befund konnte anhand des Versuchs mit den grünveredelten Stecklingen
bestätigt werden. Bei den wurzelechten Varianten Riesling und SO4 führte der AM-Pilz Glo-
mus intraradices zu signifikant geringeren Wurzellängendichten (s. Tab. 5.10). Dieses Ergeb-
nis wurde tendenziell auch bei den gepfropften Varianten erzielt. Dies steht im Einklang mit
den Versuchen von GNEKOW und MARSCHNER (1989 a), die bei ausreichender P-
Verfügbarkeit im Boden an mykorrhizierten Haferpflanzen eine Reduktion der Gesamtwurzel-
länge ermittelten. Die Ausbildung kleinerer Wurzelsysteme in Form von reduzierten Gesamt-
wurzellängen bei ausreichendem P-Versorgungsgrad ist nach MARSCHNER (1995) allgemein
ein Ausdruck von Konkurrenz um Kohlenstoffverbindungen im Wurzelbereich zwischen dem
Pilz und der Wurzel. Die Ausbildung des externen Hyphengeflechts mit seiner enormen Ober-
fläche kann dagegen das durch Mykorrhizierung reduzierte Wurzelsystem teilweise wieder

Kapitel 5 Inokulationsversuche 121
ausgleichen. Daneben fanden BERTA et al. (1990) an mykorrhizierten Wurzelsystemen von
Allium porrum morphogenetische Veränderungen in Form von kürzeren, aber zahlreicheren
Seitenwurzeln, die nach den Autoren auf eine geringere Aktivität der apikalen Meristemzellen
in der Wurzelspitze zurückgeführt werden. SCHELLENBAUM et al. (1991) bestätigten dies an
in-vitro vermehrten Grünstecklingen der Unterlagssorte SO4.
Im folgenden soll zunächst auf den Einfluß der nativen Mykorrhiza auf die Infizierung, die
Nährstoffgehalte und das Wachstum der Grünstecklinge eingegangen werden. Die bodeneige-
ne Mykorrhiza in dem verwendeten Rebschulboden führte nach 8wöchigem Gewächshausver-
such zu einem Infektionsgrad an den Rebwurzeln von durchschnittlich 12 % (s. Tab. 5.4).
Dies entspricht etwa dem Infektionsgrad der Reben aus den Ertragsanlagen, wie in Kapitel
3.3.2 dargestellt wurde. Im Gegensatz dazu wurde im Gewächshausversuch (s. Tab. 5.4) bei
der Unterlagssorte A9 der niedrigste AM-Infektionsgrad festgestellt, wohingegen bei der Frei-
landerhebung in Asselheim die Sorte A9 mit 20,1 % den höchsten Infektionsgrad aufwies
(s. Kap. 3.3.2, Tab. 3.5). Ursachen für diese unterschiedlichen Ergebnisse können in den ver-
schiedenen AM-Pilzspezies in beiden Versuchsböden begründet sein, die zu unterschiedlichen
Reaktionen zwischen Pflanze und AM-Pilzen geführt haben. Signifikante Unterschiede traten
zwischen den Unterlagen Börner, 125AA und A9 auf, wobei die Unterlagssorten Börner und
125AA stärker infiziert waren als die Sorte A9. Da Börner und A9 beides Vitis riparia x
V. cinerea-Kreuzungen sind, scheint die genetische Herkunft der Unterlagssorten keinen Ein-
fluß auf die Mykorrhizierung zu haben, da V. riparia x V. cinerea-Kreuzungen sowohl hohe
Infektionsgrade im Fall von Börner aufwiesen als auch niedrige im Fall von A9 (s. Tab. 3.5,
Kap. 3.3.2). GEISLER (1957) unterscheidet zwischen einem extensiven und einem intensiven
Wurzelsystem. Die amerikanischen Unterlagssorten werden dem extensiven Wurzelsystem
zugeordnet, die europäischen Edelreissorten dem intensiven. Der extensive Wurzeltyp durch-
wurzelt den Boden weiträumiger mit Hauptwurzeln von großer Länge, aber nur mit wenigen
Verzweigungen. Die Wurzelmasse je Bodenvolumen ist gering. Dagegen durchwurzelt der
intensive Wurzeltyp nur ein geringes Bodenvolumen, zeigt jedoch aufgrund der starken Wur-
zelverzweigungen eine relativ große Masse je Bodenvolumen auf (GEISLER, 1957). Riesling
wurde als einzige Europäersorte im Inokulationsversuch mit Grünstecklingen ähnlich stark
mykorrhiziert wie die amerikanischen Unterlagssorten (s. Tab. 5.4). Dagegen wurden im Ino-
kulationsversuch mit grünveredelten Stecklingen an Riesling tendenziell höhere Infektions-

Kapitel 5 Inokulationsversuche 122
grade ermittelt als an der Unterlagssorte SO4 (s. Tab. 5.9). KARAGIANNIDIS et al. (1995) stell-
ten bei der Inokulation mit verschiedenen AM-Pilzen an Einaugenstecklingen der Unterlags-
sorten 41B, 110R und 5BB sowie der Edelreissorte Razaki unterschiedliche Infektionsintensi-
täten fest. Bei einem AM-Pilzisolat fanden die Autoren bei der Edelreissorte Razaki höhere
Infektionsgrade als bei den Unterlagen, was im Einklang mit dem Inokulationsversuch mit den
grünveredelten Stecklingen steht. Da Riesling als Europäerrebe den Boden intensiver durch-
wurzelt (GEISLER, 1957), könnte es zu höheren Infektionsgraden gekommen sein. Warum
Riesling im Versuch mit der nativen Mykorrhiza ähnliche AM-Infektionsgrade aufwies wie
einige Unterlagssorten hängt möglicherweise mit den unterschiedlichen Wurzelexsudationen
zusammen, die nach CURRLE et al. (1983) zwischen Rebsorten variieren und die aufgrund
ihrer verschiedenartigen Zusammensetzung die Mykorrhizierung unterschiedlich beeinflussen
können.
Der verwendete Boden stammte aus der Rebschule Steingebiß (s. Kap. 4). An den Müller-
Thurgau/5BB-Pfropfreben konnte ein AM-Infektionsgrad von 5 % festgestellt werden,
(s. Abb. 4.7) was auf ein geringes AM-Infektionspotential in diesem Boden hindeutete. Die
höheren Infektionsgrade im Gefäßversuch können auf die intensivere Durchwurzelung der
Pflanzgefäße im Vergleich zum Freiland zurückgeführt werden. Eine Übertragung der Ergeb-
nisse aus dem Gefäßversuch auf die Verhältnisse im Freiland in Rebschulen ist problematisch,
da die im Vergleich zum Freiland verschiedenen physikalisch-chemischen Bedingungen in
Gefäßversuchen zu unterschiedlichen Reaktionen zwischen Pilz und Reben führen können.
Interessant war der Befund, daß es trotz des hohen P-Gehalts des Bodens (12,2 mg P/100 g
Boden [CAL]) überhaupt zu einer Mykorrhizainfektion gekommen ist. Die bodeneigene AM
wies offensichtlich eine hohe P-Toleranz auf, was ebenfalls nach den eigenen Ergebnissen in
der Versuchsanlage „Hundertmorgen“ (s. Kap. 3.2.3) vermutet wurde. Über solche Populatio-
nen wurden bisher beispielsweise von MORIN et al. (1994) an Apfel berichtet.
Bei der wurzelechten Variante Riesling wurde das Sproßtrockengewicht signifikant, bei SO4
tendenziell durch den AM-Pilz Glomus intraradices erhöht (s. Tab. 5.11). Bei den gepfropften
Varianten war kein Einfluß der AM sichtbar. Dies deutet auf einen Pfropfeffekt hin, der die
Effizienz der Mykorrhiza beeinflußt haben könnte. MERKT (1989) fand in seinen Untersu-
chungen zur Affinität von Unterlage und Edelreis, daß durch die Pfropfung das Wurzeltro-

Kapitel 5 Inokulationsversuche 123
ckengewicht deutlich reduziert wurde. Gründe hierfür vermutete der Autor in einer Hemmung
des basipetalen Assimilationstransports durch die Pfropfung, wodurch es über der Pfropfstelle
zu einem Assimilatstau kommt. In der vorliegenden Untersuchung wurde tendenziell sowohl
bei den mykorrhizierten als auch bei den nicht mykorrhizierten Riesling-Stecklingen das
Wurzeltrockengewicht durch die Pfropfung ebenfalls verringert. Es ist denkbar, daß durch den
Pfropfeingriff die Rebwurzeln mit weniger Assimilaten versorgt werden und folglich weniger
Wurzelexsudate in die Rhizosphäre ausgeschieden werden. Da AM-Pilze C-heterotroph sind
und von der Wirtspflanze mit Energie- und Kohlenstoffverbindungen versorgt werden müssen
(PEARSON und JAKOBSEN, 1993), kann ein durch Pfropfung hervorgerufenes geringeres Ange-
bot an Assimilationsprodukten in der Rhizosphäre zu einer geringeren Infektion an den Reb-
wurzeln führen. Dies konnte in der vorliegenden Untersuchung bestätigt werden. Während die
Sorte Riesling wurzelecht mit 38,5 % den höchsten Infektionsgrad aufwies (s. Tab. 5.9), führ-
te die heteroplastische Pfropfung mit der Unterlagssorte SO4 mit 26,2 % zu einem signifikant
niedrigeren Infektionsgrad, wobei bei der homoplastischen Pfropfung der gleiche Effekt als
Tendenz sichtbar war. Dadurch könnten die fehlenden Wachstumsbeeinträchtigungen bei den
veredelten Pfropfkombinationen erklärt werden. Eine andere Ursache liegt möglicherweise in
dem durch die Pfropfung veränderten Phytohormonhaushalt der Reben. Nach Untersuchungen
von EDRISS et al. (1984) und DRÜGE (1992) sind die erhöhten Cytokinin-Gehalte in mykorrhi-
zierten Pflanzen für das verbesserte Sproßwachstum verantwortlich. Im Wurzelexsudat von
mykorrhizierten Citrus-Sämlingen stellten DIXON et al. (1988) höhere Cytokininkonzentratio-
nen fest, wobei in den mykorrhizierten Sämlingen größere Mengen in den Sproß transportiert
wurden als in den nicht mykorrhizierten. Möglicherweise lagen in den vorliegenden Untersu-
chungen in den mykorrhizierten wurzelechten Varianten ebenfalls höhere Cytokiningehalte
vor, die das verbesserte Wachstum im Vergleich zu den veredelten Stecklingen erklären könn-
ten. Allerdings liegen bisher keine ausreichenden Erkenntnisse über den Einfluß der Pfrop-
fung auf den Phytohormonhaushalt bei Reben vor.
Das verbesserte Wachstum der mykorrhizierten Reben kann auf eine verbesserte Nährstoff-
versorgung der mykorrhizierten Pflanzen zurückgeführt werden. Der P-Gehalt in den Blatt-
spreiten wurde durch die Beimpfung mit dem AM-Pilz in allen Inokulationsversuchen erhöht
(s. Tab. 5.5, 5.8, 5.12 und 5.16). Die P-Gehalte in den Blättern der nicht mykorrhizierten Re-
ben lagen teilweise im Mangelbereich, insbesondere bei dem Versuch mit der nativen Mykor-

Kapitel 5 Inokulationsversuche 124
rhiza (s. Kap. 5.3.1, Tab. 5.5). Gemittelt über alle Sorten betrug der P-Gehalt in den Blättern
der mykorrhizierten Grünstecklinge 1,51 mg P/g TS. In den nicht mykorrhizierten Reben wur-
de ein Gehalt von 0,98 mg P/g TS in den Blättern ermittelt. BERGMANN (1986) gibt Gehalte
zwischen 2,5 und 4,5 mg P/g Trockenmasse für ausreichendes Pflanzenwachstum bei Reben
an. Es wurden jedoch keine P-Mangelsymptome wie dunkel- bis schmutzig-graugrüne Verfär-
bung der Mangelblätter (MENGEL, 1991) beobachtet. Ähnliche Ergebnisse fanden BAVARESCO
und FOGHER (1996), die nach Inokulation mit dem AM-Pilz Glomus mosseae bei einjährigen
Stecklingen der Pfropfkombinationen Weißburgunder (Vitis vinifera L. cv. Pinot blanc) mit
den Unterlagen SO4 und 41B in den Blättern höhere P-Gehalte feststellten. Zwischen den
einzelnen Unterlagen traten im Inokulationsversuch mit der nativen Mykorrhiza zum Teil un-
terschiedliche Nährstoffgehalte, unabhängig von der Mykorrhizabehandlung, auf. Diese gehen
auf das genotypisch unterschiedliche Nährstoffaneignungsvermögen der Unterlagen zurück,
was bereits mehrfach nachgewiesen wurde (SCIENZA et al., 1986; RÜHL, 1992; FARDOSSI
et al., 1994).
Eine Nutzung des organisch gebundenen Phosphats in den Versuchsböden, was TARAFDAR
und MARSCHNER (1994) in mykorrhizierten Weizenwurzeln beobachteten, könnte auch eine
Erklärung für die höheren P-Gehalte in den mykorrhizierten Reben sein. Die Hauptwirkung
der Mykorrhiza auf die Nährstoffaufnahme, vor allem von immobilen Nährstoffen, liegt je-
doch in der Oberflächenvergrößerung durch die weit über die Rhizosphäre hinausreichenden
Pilzhyphen. Die Ausbildung des externen Hyphengeflechts kann nach KOTHARI et al. (1990 a)
sowie ZHAO et al. (1997) eine Länge von 3 bzw. 8 - 10 m/g Bodentrockengewicht erreichen.
Bei der Bestimmung des AM-Infektionsgrades in der vorliegenden Arbeit konnte beobachtet
werden, daß das Hyphenwachstum, insbesondere bei dem AM-Pilz Glomus intraradices, in-
tensiv ausgeprägt war (s. Abb. 5.11). Ein positiver Einfluß der AM-Hyphen auf die Nährstoff-
aufnahme, wie von LI et al. (1991 a) an Weißklee und KOTHARI et al. (1991 a) an Mais ver-
mutet wurde, ist daher auch für die eigenen beschriebenen Versuche mit Reben möglich. Im
Versuch mit den grünveredelten Stecklingen wurde bei der Variante SO4 (wurzelecht) die
Sproßtrockensubstanz durch den AM-Pilz Glomus intraradices nicht signifikant verändert (s.
Tab. 5.11). Die Wurzellängendichte der mykorrhizierten SO4-Stecklinge wurde signifikant
um 16,2 % reduziert (s. Tab. 5.10). Trotz des kleineren Wurzelsystems wurde der P-Gehalt in
den Blattspreiten signifikant erhöht. Diese Tatsache zeigt sehr deutlich, daß das kleinere Wur-

Kapitel 5 Inokulationsversuche 125
zelsystem und folglich die verringerte Wurzeloberfläche durch die Ausbildung der Hyphen im
Boden wieder ausgeglichen wird. Von ähnlichen Ergebnissen berichten KOTHARI et al. (1990
a), die nach der Beimpfung mit dem AM-Pilz Glomus mosseae an Mais eine um 31 % redu-
zierte Gesamtwurzellänge fanden, wobei das Sproßtrockengewicht unverändert blieb, die P-
Gehalte im Sproß aber signifikant anstiegen.
Ein Mykorrhizabeitrag zur P-Aufnahme über die Hyphen erscheint im Inokulationsversuch
mit der nativen Mykorrhiza fraglich, da die Reben mit durchschnittlich 12 % nur relativ ge-
ring infiziert waren (s. Tab. 5.4). Trotzdem traten in allen Sproßfraktionen neben der gestie-
genen Sproßtrockenmasse (s. Abb. 5.4) erhöhte P-Gehalte in den mykorrhizierten Reben auf.
Neben dem direkten Mykorrhizabeitrag über die Hyphen gibt es indirekte Mykorrhiza-Effekte,
die zu einer erhöhten P-Aufnahme in mykorrhizierten Pflanzen führen. So kann die Ausschei-
dung von organischen Verbindungen, wie z. B. Citrat, Siderophoren oder Chelat-
Verbindungen eine pH-Absenkung des Rhizosphärenbodens herbeiführen, die die P-
Verfügbarkeit im Boden durch Mobilisierung von schwer löslichen Ca-Phosphaten in neutra-
len oder alkalischen Böden erhöht. Allerdings wurden diese Beobachtungen bisher nur bei
Ektomykorrhizen festgestellt (MARSCHNER und DELL, 1994).
Neben der Beeinflussung der Nährstoffaufnahme durch Bodeneigenschaften hängt die Nähr-
stoffaufnahme insbesondere von schwer beweglichen Nährstoffen vom jeweiligen Aneig-
nungsvermögen der Pflanzenarten ab. Dieses wird in besonderem Maße von den morphologi-
schen Eigenschaften der Wurzeln beeinflußt (SCHWEIGER et al., 1995). Bei Pflanzenarten mit
grobem Wurzelsystem und nur wenigen Wurzelhaaren, wie z.B bei der Rebe (RICHARDS,
1983; MOHR, 1988), hat die Mykorrhiza möglicherweise einen wesentlichen Einfluß auf die
Nährstoffaufnahme. Anhand des Versuches mit Wurzelbeobachtungskästen (s. Kap. 5.3.2)
konnte nachgewiesen werden, daß die Wurzellängendichte bei den Versuchsvarianten mit
dem Inokulationsbereich „oben“ bzw. „unten“ bei den nicht mykorrhizierten Reben, gemittelt
über die Tiefenstufen, 0,66 cm/cm3 bzw. 0,60 cm/cm3 und bei den mykorrhizierten Reben
0,57 cm/cm3 bzw.
0,42 cm/cm3 betrug (s Abb. 5.7 und 5.8). Im Vergleich dazu durchwurzeln Gramineen den
Boden viel intensiver, wie aus den Untersuchungen von GNEKOW (1988) hervorgeht, der in
Gefäßversuchen mit Hafer eine Wurzellängendichte von 38 - 68 cm/cm3 Boden ermittelte. Im

Kapitel 5 Inokulationsversuche 126
Freiland muß bei Reben von noch kleineren Werten ausgegangen werden, wie die Ergebnisse
von MOHR (1996) sowie die eigenen Ergebnisse aus den Ertragsanlagen (s. Tab. 3.5) bestäti-
gen. An Gramineen sind die Werte im Freiland ebenfalls wesentlich niedriger als in Gefäßen
(3 cm/cm3 an Winterweizen; BÖHM, 1979), aber immer noch um ein Mehrfaches höher als bei
Reben. Ein hoher Mykorrhizabeitrag zur P-Aufnahme wird ebenfalls für Zwiebel und Maniok
diskutiert, da diese Pflanzenarten, ähnlich wie die Rebe, kaum bzw. keine Wurzelhaare besit-
zen (KOIDE, 1991). Im Gegensatz dazu ist bei Pflanzenarten mit feinem Wurzelsystem, hoher
Wurzellängendichte und folglich großer Oberfläche, wie z. B. bei Gramineen, der Einfluß der
Mykorrhiza oftmals geringer.
In der vorliegenden Untersuchung wurden in den inokulierten Varianten neben den erhöhten
P-Gehalten erhöhte Gehalte der Mikronährstoffe Zn und Cu in den Blattspreiten ermittelt
(s. Tab. 5.6, 5.7, und 5.13). Bei mehrjährigen Kulturpflanzen fanden GNEKOW und MARSCH-
NER (1989 b) bei Apfel in der Wurzel und in den Blattspreiten erhöhte Zn- und Cu-Gehalte. In
den Inokulationsversuchen waren die Reben ausreichend mit Zn (31,8 - 106 mg/kg Trocken-
substanz [TS] und Cu (7,5 - 17,1 mg/kg TS) versorgt. Lediglich im Ozonversuch lagen die
Cu-Gehalte in den Blattspreiten bei den nicht mykorrhizierten Reben, gemittelt über die Ver-
suchsvarianten mit 4,4 mg/kg TS, im Mangelbereich (s. Tab. 5.15). Ausreichende Cu-Gehalte
in Rebblättern liegen nach BERGMANN (1986) im Bereich zwischen 6 - 12 mg/kg TS. Durch
die Inokulation mit dem AM-Pilz Glomus sp. wurde der Cu-Gehalt im Durchschnitt auf 6,5
mg/kg TS erhöht. In Versuchen mit kompartimentierten Gefäßen mit Weißklee konnten LI et
al. (1991 c) durch räumliche Trennung von Wurzel- und Hyphenraum einen Hyphenbeitrag
für die Cu-Gehalte im Sproß von 53 - 62 % ermitteln. In einer ähnlichen Versuchsanordnung
fanden KOTHARI et al. (1991 a) im Sproß einen Hyphenbeitrag von 16 - 25 % für die Zn-
Gehalte. Die höheren Zn- und Cu-Gehalte in den mykorrhizierten Reben könnten in der vor-
liegenden Untersuchung ebenfalls auf einen Mykorrhizabeitrag über die AM-Hyphen zurück-
geführt werden, was bereits für P erwähnt wurde.
Im Ozonversuch traten nach 3wöchiger Versuchsphase zwischen den mykorrhizierten und
nicht mykorrhizierten Rieslingstecklingen in den Blättern keine Unterschiede im Zn-Gehalt
auf (s. Tab. 5.15). Der fehlende Mykorrhizabeitrag für die Zn-Aufnahme könnte auf die ver-
wendete Pilzart zurückzuführen sein. Andererseits hängt der Mykorrhizaeffekt auf die Mikro-

Kapitel 5 Inokulationsversuche 127
nährstoffaufnahme von zahlreichen Faktoren ab, wie z. B. dem pH-Wert im Boden (MARSCH-
NER, 1993), der Nährstoffversorgung des Bodens (RAJU et al., 1990 a) und der Bodentempera-
tur (RAJU et al., 1990 b). Es ist denkbar, daß die Umweltbedingungen, insbesondere die hohen
Temperaturen in den Begasungskammern (s. Kap. 5.2.4, Abb. 5.3) einen Einfluß auf die Aus-
prägung des AM-Pilzes gehabt haben könnten.
Während die Cu- und Zn-Gehalte in der vorliegenden Untersuchung teilweise in den my-
korrhizierten Reben erhöht waren, wurden im Versuch mit der nativen Mykorrhiza (Kap.
5.3.1, Tab. 5.6) sowie beim Topfrebenversuch (Kap. 4.3.2, Tab. 4.7) in den inokulierten Re-
ben zum Teil signifikant niedrigere Mn-Gehalte in den Blattspreiten festgestellt. Gemittelt
über die Sorten waren die Mn-Gehalte in den Blattspreiten („alten Blätter“) der mykorrhizier-
ten Reben um 30 % geringer als in den nicht mykorrhizierten Reben. Als Ursache für die ge-
ringeren Mn-Gehalte in mykorrhizierten Pflanzen werden in der Literatur verschiedene Mög-
lichkeiten diskutiert. Die Mn-Aufnahme wird durch Oxidations- und Reduktionsprozesse ge-
steuert, die im Boden gleichzeitig ablaufen. Die mikrobielle Aktivität in der Rhizosphäre er-
höht oder erniedrigt die Verfügbarkeit von Mn für die Pflanzenwurzeln (MARSCHNER, 1988).
Die AM-Pilze induzieren in der Rhizosphäre qualitative Veränderungen der Mikroorganis-
menpopulationen und quantitative Modifikationen bei den Wurzelausscheidungen. KOTHARI
et al. (1991 b) und POSTA et al. (1994) machen die Verminderung der Anzahl bakterieller Mn-
Reduzierer in der Mykorrhizosphäre und somit die entsprechend geringere Mn-Verfügbarkeit
für die geringere Mn-Aufnahme mykorrhizierter Pflanzen verantwortlich. Demgegenüber
könnte auch eine hohe Populationsdichte Mn-oxidierender Bakterien in der Mykorrhizosphäre
zu niedrigerer Mn-Verfügbarkeit führen (ARINES et al., 1992). Daß es sich in den eigenen
Versuchen bei den niedrigeren Mn-Gehalten in den Blättern durch das verbesserte Wachstum
der mykorrhizierten Reben um einen Verdünnungseffekt handeln könnte, wird von den Er-
gebnissen von KOTHARI et al. (1991 b) widerlegt, die in mykorrhizierten Maispflanzen im
Sproß erhöhte Mn-Gehalte fanden. Im Versuch mit den grünveredelten Stecklingen mit dem
AM-Pilz Glomus intraradices wurden zwischen den AM-Behandlungen keine Unterschiede
im Mn-Gehalt festgestellt
(vgl. Tab. 5.13). Die unterschiedliche Wirkung verschiedener AM-Pilze auf die Mn-
Aufnahme könnte, wie GEORGE et al. (1994) ebenfalls vermuten, auf die unterschiedliche
Wurzelexsudation mykorrhizierter Pflanzen zurückgeführt werden.

Kapitel 5 Inokulationsversuche 128
Die durch die AM-Pilze bedingten niedrigeren Mn-Gehalte können zu einer höheren Mn-
Toleranz der Pflanzen führen. Dies gilt insbesondere in sterilisierten Bodensubstraten, in de-
nen z. B. durch Bodendämpfung das leicht reduzierbare Mn im Boden in das leicht pflanzen-
verfügbare, austauschbare Mn überführt wird. Dadurch kann es zu hohen bis toxischen Mn-
Gehalten kommen. WASCHKIES et al. (1992) stellten in Blättern von Grünstecklingen der Un-
terlags-
sorte 5C, die in einem sterilisierten Rebschulboden wuchsen, Mn-Gehalte von
484 mg Mn/kg TS fest, die möglicherweise zu Wachstumsbeeinträchtigungen geführt haben.
In den vorliegenden Untersuchungen lagen die Mn-Gehalte teilweise weit über dem optimalen
Versorgungsbereich von 30 - 100 mg/kg TS (BERGMANN, 1986). Die höchsten Mn-Gehalte
mit 211 mg/kg TS traten im Versuch mit den Wurzelbeobachtungskästen (Kap. 5.3.2,
Tab. 5.7) bei der Inokulationsvariante „unten“ in den nicht mykorrhizierten Reben auf. Mn-
Toxizitätserscheinungen wie chlorotische Aufhellungen und braune Flecken in den Blättern
(MENGEL, 1991) traten nicht auf.
In dem Versuch mit der nativen Mykorrhiza (s. Kap. 5.3.1) und dem Inokulationsversuch mit
Glomus intraradices (s. Kap. 5.3.3) wurden bei verschiedenen Stecklingen in der Trockensub-
stanz der Blätter niedrigere Fe-Gehalte in den mykorrhizierten Pflanzen festgestellt (s. Tab.
5.6 bzw. 5.13). Die geringeren Fe-Gehalte in der Sproßmasse mykorrhizierter Pflanzen im
Vergleich zu nicht mykorrhizierten sind ebenso wie bei Mn bekannt (PACOVSKY, 1986;
KOTHARI et al, 1991 b). Ursache hierfür könnte eine qualitative Veränderung in der Rhi-
zosphärenmikrobenpopulation sein, die durch die AM-Pilze hervorgerufen wird. POSTA et al.
(1994) stellten neben der reduzierten Anzahl von Mn-reduzierenden Bakterien auch eine ge-
ringere Anzahl an Fe-reduzierenden Bakterien fest, die in der Rhizosphäre mykorrhizierter
Maispflanzen um das 20 - 30fache niedriger war. BIRICOLTI et al. (1997) ermittelten an 5BB-
Stecklingen tendenziell niedrigere Fe-Gehalte in den Blättern von mykorrhizierten Reben, was
mit den eigenen Ergebnissen übereinstimmt. BAVARESCO und FOGHER (1996) fanden dagegen
an mit dem AM-Pilz Glomus mosseae inokulierten Reben verschiedener Pfropfkombinationen
(Pinot blanc mit 3309, SO4 und 41B) signifikant höhere Fe-Gehalte in den Blättern. Es kann
davon ausgegangen werden, daß diese unterschiedlichen Ergebnisse auf die Verwendung ver-
schiedener Rebsorten oder Pilzarten zurückzuführen sind.

Kapitel 5 Inokulationsversuche 129
Zusammenfassend weisen die Inokulationsversuche darauf hin, daß sowohl der AM-Pilz
Glomus intraradices als auch die native Mykorrhiza in einem Rebschulboden das Wachstum
von wurzelechten bzw. veredelten Grünstecklingen verschiedener Rebsorten verstärkt. Die
Mykorrhizierung führte bei den meisten Versuchen zu erhöhten Nährstoffgehalten in den Blät-
tern, insbesondere bei den immobilen Nährstoffen wie P, Cu und Zn. Demgegenüber konnten
in den mykorrhizierten Reben oftmals geringere Mn- und Fe-Gehalte festgestellt werden.
5.4.2 Wurzelwachstum und Ausbreitung der AM-Infektion
Die Verwendung von Wurzelkästen mit einer Höhe von 70 cm ermöglichten ein ungehinder-
tes Wachstum der Rebwurzeln über einen bestimmten Zeitraum. Die Grünstecklinge der Un-
terlagssorte SO4 hatten nach 4wöchiger Wachstumsphase die Versuchsgefäße nicht vollstän-
dig durchwurzelt. In der Bodentiefe von 54 - 66 cm wurden keine Rebwurzeln gefunden. Erst
nach sechs Wochen war der gesamte Wurzelkasten durchwurzelt. Dies entsprach bei einer
Höhe des Versuchsgefäßes von 70 cm (abzüglich 4 cm Kiesauflage) und einer Wachstums-
dauer von
42 Tagen einem durchschnittlichen Tiefenwachstum der Rebwurzeln von 1,6 cm/d. Ähnliche
Ergebnisse erzielte MOHR (1988) an mit Weinbergserde gefüllten Wurzelkästen bei wurzel-
echten Rieslingstecklingen. Bei den Wurzeln 1. Ordnung stellte der Autor Tiefenwachstums-
raten von durchschnittlich 1,2 cm/d fest, bei den Wurzeln 2. Ordnung 0,5 cm/d. HILTON und
KHATAMIAN (1973) fanden in Freilandversuchen an Reben im späten Frühjahr Wurzelausbrei-
tungsraten von 1 cm/d. Das Wachstum und die Ausbreitungsrichtung von Rebwurzeln wird
durch zahlreiche pedogene Faktoren wie Nährstoffgehalt des Bodens, pH-Wert und Belüftung
des Bodens beeinflußt (MORLAT und JACQUET, 1993). Der in den eigenen Versuchen einge-
setzte Rebschulboden ermöglichte den Stecklingen aufgrund des hohen Sandgehalts
(s. Tab. 5.2) ideale Wachstumsmöglichkeiten, wie sie auch im Freiland in rebschulfähigen
Böden vorzufinden sind.
Durch die Inokulation mit dem AM-Pilz Glomus mosseae betrug der AM-Infektionsgrad der
Rebwurzeln im unmittelbaren Inokulationsbereich nach sechs Wochen 45 % („oben“) bzw.
35 % („unten“) (s. Abb. 5.6). BIRICOLTI et al. (1997) inokulierten Einaugenstecklinge der Un-

Kapitel 5 Inokulationsversuche 130
terlagssorte 5BB mit den AM-Pilzen Glomus constrictus, G. deserticola und G. mosseae und
fanden ähnlich hohe Infektionsgrade an den Rebwurzeln wie in der vorliegenden Untersu-
chung. Bei einer durchschnittlichen Wachstumsgeschwindigkeit der Rebwurzeln von 1,6 cm/d
und einer Versuchsdauer von 42 Tagen befanden sich die jungen Wurzeln etwa acht Tage lang
im unmittelbaren Inokulationsbereich. Während dieser Zeit konnten die Rebwurzeln intensiv
mit dem AM-Pilz infiziert werden. Verschiedene Autoren haben mehrere Modelle entwickelt,
mit denen das Wachstum von Mykorrhizainfektionen in pflanzlichen Wurzelsystemen be-
schrieben werden kann (SANDERS und SHEIKH, 1983; BUWALDA et al., 1984; AMIJEE et al.,
1986). So gibt z. B. BUWALDA et al. (1984) eine Verzögerungszeit mit Werten von fünf Tagen
an, die zwischen Beimpfung und Sichtbarwerden der Infektion verstreicht. Die Zeitdauer für
die Ausbreitung des Mykorrhizapilzes wird durch den Entwicklungszyklus des Pilzes be-
stimmt. Nach der Keimung einer Spore breiten sich die Pilzhyphen soweit aus, bis eine Wirts-
pflanzenwurzel erreicht wird. TOMMERUP (1984) stellte in ihren Untersuchungen fest, daß die
Bildung einer Keimhyphe aus einer Spore von Glomus caledonium sechs Tage benötigte. In
ihrem Modell zur Beschreibung der Infektionsausbreitung im Wurzelsystem von Lauchpflan-
zen bestimmten BUWALDA et al. (1984) die konstante Längenwachstumsrate des infizierten
Segmentes der Einzelwurzel. Ebenso stellten AMIJEE et al. (1986) und GNEKOW (1988) Werte
von 2 -
5,3 mm/d fest. Dieser Wert konnte annähernd in den eigenen Untersuchungen für Rebwurzeln
bestätigt werden. Ausgehend vom Inokulumband, welches in der Bodentiefe von 9 - 18 cm
lag, hat sich der AM-Pilz Glomus mosseae bei einer durchschnittlichen Ausbreitungsge-
schwindigkeit von 5 mm/d nach 42tägiger Wachstumsphase bis in eine Bodentiefe des
Pflanzgefäßes von etwa 29 cm ausgebreitet. Der Infektionsgrad der Rebwurzeln nahm von der
Inokulationsschicht ausgehend mit zunehmender Bodentiefe ab. Bei der Inokulation „oben“
(Inokulumband in 9 - 18 cm Bodentiefe) war ab einer Tiefe von 36 cm keine AM-Infektion
mehr nachweisbar (vgl. Abb. 5.6). Ein möglicher Grund für die geringe Ausbreitung des AM-
Pilzes G. mosseae im vorliegenden Versuchssystem könnte folglich in der hohen Wachstums-
geschwindigkeit der Rebwurzeln liegen.
Das Wurzelsystem der SO4-Grünstecklinge war durch die Inokulation mit G. mosseae nur
teilweise mit AM infiziert. Trotzdem war die Trieblänge und das Trockengewicht in der Ino-
kulationsvariante „oben“ deutlich erhöht (s. Abb. 5.9). Dies deutet darauf hin, daß die Rebe

Kapitel 5 Inokulationsversuche 131
bereits bei einer Teilbesiedlung des Wurzelsystems mit einem AM-Pilz mit verstärktem
Wachstum reagiert. Das Zusammentreffen der Rebwurzeln mit dem AM-Inokulum bei der
Inokulationsmethode „unten“ erfolgte sehr spät. Eine mykorrhizabedingte Wachstumsförde-
rung und erhöhte P-Gehalte in den Blättern wie bei der Inokulationsmethode „oben“ konnte
bei der Inokulationsmethode „unten“ nicht beobachtet werden. BIRICOLTI et al. (1997) stellten
in ihren Untersuchungen fest, daß nach Inokulation mit verschiedenen Mykorrhizapilzen Ein-
augenstecklinge der Unterlagssorte 5BB mit unterschiedlichem Wachstum auf die Infektion
reagierten. Während die mit G. mosseae und G. constrictus beimpften Stecklinge mit stärke-
rem Wachstum reagierten, war die Wachstumssteigerung bei den mit G. deserticola beimpften
Stecklingen deutlich geringer, obwohl G. deserticola zu den höchsten Infektionsgraden an den
Wurzeln führte. Folglich kann der durch einen AM-Pilz hervorgerufene Infektionsgrad nicht
allein für das verbesserte Wachstum verantwortlich gemacht werden.
Nach der Freilandinokulation mit einem AM-Pilz in beiden Rebschulen wurden erhöhte Infek-
tionsgrade an den Pfropfreben festgestellt (s. Kap. 4.3.1). Daraus resultierten nach 14wöchiger
Wachstumsphase signifikant größere Trieblängen an den inokulierten Reben bei einem durch-
schnittlichen Infektionsgrad der Wurzeln von 12 % bzw. 17 %. Die eigene Versuchsanstellung
mit den Wurzelkästen entsprach in etwa einer im Gefäßversuch simulierten Feldinokulation,
bei der die Reben, ähnlich wie in der Rebschule, durch ein Inokulumband hindurchwachsen.
Dabei nahm der AM-Infektionsgrad an den Rebwurzeln mit zunehmender Entfernung vom
Inokulumband ab. Im Freilandversuch in Kapitel 4 wurden die Wurzelproben nicht schicht-
weise im Abstand vom Inokulum geerntet, sondern der AM-Infektionsgrad wurde an einer
Wurzelmischprobe bestimmt. Der dort festgestellte niedrige Infektionsgrad weist darauf hin,
daß die Rebwurzeln im Freiland nach dem Durchwachsen des Inokulumbandes nur geringfü-
gig infiziert werden. SIEVERDING (1985) berichtet dagegen von Inokulationserfolgen mit Ma-
niok (Manihot esculenta Crantz) und über unterschiedliche Plazierungsmethoden von VA-
Mykorrhizapilzen im Freiland. Insbesondere bei der Plazierung des Beimpfungsmaterials
Glomus manihotis unterhalb des Steckholzes konnte der Autor eine Infektion der Wurzel mit
G. manihotis auch außerhalb des Inokulationsbereiches beobachten. Die größte Infektionsrate
fand SIEVERDING (1985) drei Monate nach der Inokulation. BIERMANN und LINDERMAN (1983
b) stellten bei Inokulationsversuchen mit Geranien-Stecklingen (Pelargonium x hortorum
L.H. Bailey) auch an den neu gebildeten Wurzeln außerhalb der Inokulationsschicht ähnlich

Kapitel 5 Inokulationsversuche 132
hohe AM-Infektionen fest wie an den Wurzeln aus dem Boden-Inokulum-Gemisch. Über die
Ausbildung der externen Mykorrhiza-Hyphen werden über Sekundärinfektionen neue Wur-
zeln infiziert. Möglicherweise hat der AM-Pilz in der vorliegenden Untersuchung nur unzu-
reichende externe Hyphen ausgebildet. Es kam nicht zu Sekundärinfektionen, so daß die
Rebwurzeln unterhalb des Inokulumbandes nicht mehr infiziert wurden. Dies stimmt mit den
Ergebnissen aus Kapitel 4.3.2 überein, bei dem die aus dem Boden-Inokulum-Gemisch des
Gittertopfes herausgewachsenen Wurzeln kaum bzw. gar nicht infiziert waren.
Die Ergebnisse aus der vorliegenden Untersuchung zeigen, daß in Rebwurzeln nach Bandino-
kulation mit AM und einer Wachstumsdauer von sechs Wochen das Wurzelsystem im unmit-
telbaren Bodenbereich sowie im angrenzenden Bodenbereich an das Bandinokulum hinrei-
chend besiedelt wird. Die erhöhte Trockensubstanzbildung der mykorrhizierten Reben zeigt
jedoch eindeutig, daß bereits eine Teilbesiedlung des Wurzelsystems mit AM ausreicht, die
Rebe mit mehr Nährstoffen zu versorgen. Inwieweit dieser Effekt im Freiland in nicht sterili-
siertem Boden auch für autochthone Mykorrhizapilze gilt, muß in weiteren Untersuchungen
geprüft
werden.
5.4.2 Auswirkungen erhöhter Ozonbelastung
Die Menge der Schadstoffkonzentration von 250 µg O3/m3 entsprach einer Konzentration, die
auch im Freiland in Weinbergen auftreten kann und die nach BERAN (1994 a) je nach Dauer
der Expositionszeit zu irreparablen Blattschäden an Reben führen kann. In der vorliegenden
Untersuchung wurde der empfohlene Grenzwert (MIK-Wert) von 120 µg O3/m3 während des
Versuchszeitraumes in der Außenluft mehrmals überschritten, wobei das höchste Tages-
Halbstundenmaximum 210 µg O3/m3 betrug (s. Abb. 5.14). Entsprechend dem natürlichen
Verlauf der Ozonkonzentration, die bis zum Nachmittag ihren Höhepunkt erreicht und dann
langsam wieder absinkt, wurde eine vierstündige Begasungszeit an vier Tagen in der Woche
mit dazwischenliegenden Erholungsphasen ohne Schadstoffbelastung durchgeführt. Somit
konnten die Freilandverhältnisse im Gegensatz zu kontinuierlichen Schadstoffapplikationen
annähernd simuliert werden. Durch die vierstündige Expositionszeit an den Begasungstagen
ergab sich mit 250 µg O3/m3 eine Ozondosis von 1.000 µg/m3.

Kapitel 5 Inokulationsversuche 133
Ozon gelangt über die Spaltöffnungen ins Blattinnere und schädigt in erster Linie das Palisa-
denparenchym, während Schwammparenchym und Epidermis nicht geschädigt werden. Die
Folge sind chlorotische Aufhellungen, die durch den Abbau des Chlorophylls verursacht wer-
den. Bedingt durch Permeabilitätsänderungen der Membranen kommt es zu dunkel gefärbten,
teils wässrig aussehenden Flecken, die als „water soaked spots“ bezeichnet werden (BERAN,
1994 a). Bei kurzzeitig einwirkenden Konzentrationen werden scharf abgegrenzte, punktför-
mige Symptome auf der Blattoberfläche von Reben sichtbar, die als „oxidant stipple“ be-
zeichnet werden (RICHARDS et al., 1958; MUSSELMAN et al., 1978; ANONYM, 1983). Bei
makroskopischen Untersuchungen stellte BERAN (1994 a) punktförmige, braune bis schwarze
nekrotische Flecke an den Blättern verschiedener V. vinifera-Sorten fest, die sich je nach Hö-
he und Dauer der Schadstoffbelastung auch zu größeren, unregelmäßig begrenzten Einheiten
vereinigten. Nach der 3wöchigen Versuchsdauer wurden sowohl an den mykorrhizierten als
auch an den nicht mykorrhizierten Reben in keiner Versuchsvariante sichtbare Blattschäden
beobachtet. Aus der Literatur liegen Erkenntnisse vor, die bei der Klassifikation von Immissi-
onswirkungen auf Pflanzen neben den sichtbaren Schäden wie Nekrosen, Chlorosen oder Ver-
färbungen von äußerlich nicht feststellbaren Schädigungen berichten (ANONYM, 1983; REN-
NENBERG et al., 1996). Dies trifft für den vorliegenden Versuch zu und steht im Einklang mit
BERAN (1994 a), der bei geringen, aber langandauernden Schadstoffbelastungen von nicht
sichtbaren Eingriffen in die Photosynthese berichtet. MUSSELMAN und MELIOUS (1984) ver-
glichen verschiedene amerikanische und französische Hybriden mit sechs Vitis vinifera-Sorten
hinsichtlich ihrer Anfälligkeit gegenüber Ozonschäden und fanden bei den V. vinifera-Sorten
eine geringere Sensibilität im Vergleich zu den Hybridsorten. Bei der Schadensbonitur stellten
die Autoren innerhalb der V. vinifera-Sorten keine Unterschiede in der Befallshäufigkeit fest.
In Freilandversuchen fanden dagegen HASLER et al. (1990) und HERRMANN (1991) teilweise
deutliche Sortenunterschiede in der Befallshäufigkeit zwischen verschiedenen Europäersorten,
was BERAN (1994 b) in Gefäßversuchen mit Begasungskammern bestätigen konnte. Der Autor
fand bei Müller-Thurgau-Stecklingen die größte Sensibilität gegenüber Ozon, während Portu-
gieser in den meisten Fällen eine höhere Verträglichkeit erkennen ließ. Die unterschiedliche
Sensibilität zwischen den verschiedenen Europäersorten hinsichtlich Ozon hängt unter ande-
rem von der durch das Blatt tatsächlich aufgenommenen Ozon-Menge ab, die durch die unter-
schiedliche Stomatareaktion der Pflanze bestimmt wird (WEINSTEIN, 1984). BERAN (1994 b)

Kapitel 5 Inokulationsversuche 134
stellte bei seinen Untersuchungen über unterschiedlich lange Expositionszeiten und O3-
Konzentrationen an verschiedenen Rebsorten fest, daß der O3-Gehalt der Umgebungsluft ei-
nen höheren Einfluß auf die Reben ausübte als die Dauer der Begasungszeit. Es wurde bewußt
eine O3-Konzentration von 250 µg O3/m3 nicht überschritten, um freilandähnliche Verhältnis-
se herzustellen. In einem weiteren Versuch wurde die Expositionsdauer bei gleicher Schad-
stoffkonzentration um vier Wochen verlängert. Dabei konnten ebenfalls keine Blattschäden
festgestellt werden.
In der vorliegenden Untersuchung wurde die Mykorrhizierung der Rebwurzeln durch die un-
terschiedliche Schadstoffapplikation nicht beeinflußt (s. Tab. 5.14). Im Gegensatz dazu sind
Untersuchungen bekannt, die nach O3-Einwirkung von reduzierten AM-Infektionsgraden be-
richten (MCCOOL et al., 1979; MCCOOL et al., 1983; MCCOOL und MENGE, 1984). Durch O-
zoneinfluß hervorgerufene Blattschäden führen zunächst zu einer Akkumulation von Stärke
und Kohlenhydraten in den Blättern, was zu einer meßbaren Verringerung in der Photosynthe-
seleistung führt (RENNENBERG et al., 1996). Die Folge ist eine Abnahme in der Verlagerung
von Assimilaten in die Wurzel, wodurch die Kohlenhydratversorgung der Wurzeln zurück-
geht. Im Wurzelexsudat von Ozon exponierten Tomatensämlingen fanden MCCOOL et al.
(1983) geringere Gehalte an reduzierenden Zuckern und Aminosäuren, die den AM-
Infektionsgrad im Vergleich zu den nicht begasten Pflanzen je nach Schadstoffkonzentration
um 46 % bzw. 63 % reduzierten. Da die Reben im vorliegenden Versuchssystem zumindest
keine sichtbaren Blattschäden aufwiesen, war eine Beeinflussung der Mykorrhizierung nicht
zu erwarten. Die Schadstoffbelastung in den Varianten „Ozon“ sowie „Außenluft“ führte im
Vergleich zu den nicht begasten Reben bei den nicht mykorrhizierten Reben tendenziell zu
einer geringeren Sproßmasse (vgl. Abb. 5.15). Dieser Effekt konnte bei den mykorrhizierten
Reben nicht beobachtet werden. Von ähnlichen Ergebnissen berichten MAHONEY et al. (1985)
an Kiefer-Sämlingen, allerdings mit Ektomykorrhiza-Pilzen. Die Autoren führten die protek-
tive Eigenschaft der mykorrhizierten Kiefern gegenüber Ozoneinfluß auf das größere Wurzel-
system der mykorrhizierten Sämlinge zurück. Möglicherweise kann in der vorliegenden Ar-
beit ebenfalls von positiven Eigenschaften der mykorrhizierten Reben gegenüber Ozoneinfluß
ausgegangen werden, da diese, ähnlich wie in den Untersuchungen von MAHONEY et al.
(1985), größere Wurzelsysteme aufwiesen. Es kann davon ausgegangen werden, daß sich die
Immissionsempfindlichkeit gegenüber Ozon in den mykorrhizierten Reben aufgrund der er-

Kapitel 5 Inokulationsversuche 135
höhten Nährstoffgehalte in den Blättern soweit verändert hat, daß die mykorrhizierten Reben
gegenüber den nicht mykorrhizierten einen Wachstumsvorteil hatten. Die Wachstumsbeein-
flussung der nicht mykorrhizierten Reben in der Variante mit „Außenluft“ war vergleichbar
der mit Ozonbegasung. Im Freiland werden Reben nicht nur durch einzelne Schadstoffe, son-
dern durch unterschiedliche Kombinationen verschiedener photochemisch wirkender Oxidan-
tien in wechselnden Konzentrationen belastet (WEINSTEIN, 1984; MUSSELMAN et al., 1985;
BERAN, 1994 b). Dabei kann die Wirkung der Gemische in der Luft aus Ozon, Schwefeldioxid
und Stickstoffdioxid synergistischer oder antagonistischer Natur sein. Die eigenen Ergebnisse
an Reben haben gezeigt, daß mykorrhizierte Reben sowohl bei hohen Ozonkonzentrationen
(250 µg O3/m3) als auch bei natürlicher Schadstoffbelastung der Luft über einen bestimmten
Zeitraum einen Wachstumsvorteil gegenüber nicht mykorrhizierten Reben haben können.

Kapitel 6 Abschließende Diskussion 136
C ZUSAMMENFASSENDER TEIL
6 Abschließende Diskussion
In der vorliegenden Arbeit wurde das Vorkommen der AM in Ertragsanlagen und Rebschulen
ermittelt sowie Anwendungsmöglichkeiten der AM-Pilze in der Rebenpflanzguterzeugung
aufgezeigt. Die Inokulation mit dem AM-Pilz Glomus sp. erhöhte an Pfropfreben den Infekti-
onsgrad in zwei Rebschulstandorten signifikant (s. Kap. 4.3.1). Nach 12- bzw. 14wöchiger
Wachstumsphase wurden an den mykorrhizierten Reben signifikant größere Trieblängen er-
mittelt. Die Anwendung der AM-Pilze Glomus sp. und Glomus mosseae in der Topfrebenpro-
duktion führte, ähnlich wie in den Gefäßversuchen an Rebenstecklingen gezeigt, zu signifi-
kant größeren Trieblängen. Daneben wurden Inokulationsversuche mit den AM-Pilzen Glo-
mus mosseae (s. Kap. 5.3.2), Glomus intraradices (s. Kap. 5.3.3) und Glomus sp. (s. Kap.
5.3.4) an Rebenstecklingen durchgeführt, die oftmals zu Wachstumssteigerungen an den my-
korrhizierten Reben geführt haben. Außerdem konnte gezeigt werden, daß die nativen AM-
Pilze eines Rebschulbodens im Vergleich zu einer Variante, bei der durch Bodensterilisierung
die nativen AM-Pilze eliminiert wurden, das Wachstum von Rebenstecklingen förderten
(s. Kap. 5.3.1).
Durch die bodeneigene AM in einem Rebschulboden wurde an Grünstecklingen ein durch-
schnittlicher AM-Infektionsgrad von 12 % ermittelt (s. Kap. 5.3.1, Tab. 5.4). Bei der Wirkung
der nativen Mykorrhiza auf die einzelnen Rebenstecklinge deutete sich eine positive Korrela-
tion zwischen dem Grad der Mykorrhizierung und der oberirdischen Trockenmassebildung an,
die jedoch nicht statistisch abgesichert werden konnte (r2 = 0,526). Von vergleichbaren Er-
gebnissen berichtet FELDMANN (1990) an Kautschukbäumen. Folglich ist eine intensive Be-
siedlung von Wurzeln mit AM-Pilzen nicht unbedingt ein Kriterium für die Effizienz eines
Mykorrhizapilzes, was die eigenen Ergebnisse mit den Wurzelkästen (s. Kap. 5.3.2) belegen.
Bereits eine Teilbesiedlung des Wurzelsystems mit dem AM-Pilz Glomus mosseae führte zu
Wachstumsverbesserungen, die unter anderem durch die erhöhte Trockensubstanzbildung und
die höheren Nährstoffgehalte in den Blättern auf eine erhöhte Nährstoffaufnahme der my-
korrhizierten Reben zurückgeführt werden konnte. In einigen Versuchen blieb das Wurzel-

Kapitel 6 Abschließende Diskussion 137
wachstum durch die Inokulation mit AM-Pilzen unbeeinflußt (s. Kap. 5.3.2 und 5.3.3.2, Tab.
5.11). Hieraus resultierten bei den mykorrhizierten Reben erhöhte Sproß-/Wurzelverhältnisse,
was mit den Ergebnissen aus der Literatur übereinstimmt (SKINNER et al., 1988; GNEKOW und
MARSCHNER, 1989 a). Das verstärkte Sproßwachstum verbunden mit einer verringerten Tro-
ckensubstanzbildung der mykorrhizierten Rebwurzeln deutet auf eine verbesserte P-
Ernährung der Reben hin. Dies wurde anhand der höheren P-Gehalte der Blätter bestätigt (s.
Tab. 5.8
und 5.12).
Der positive Einfluß der Mykorrhiza auf die Sproßtrockenmasse war bei den wurzelechten
Stecklingen größer als bei den gepfropften Stecklingen (s. Tab. 5.11). Die Pfropfung stellt
einen erheblichen Eingriff in die Stoffwechselprozesse der Rebe dar. Durch die Pfropfung
kann es zu einer Hemmung des basipetalen Assimilationstransportes kommen, wodurch es
über der Pfropfstelle zu einem Assimilatstau kommt (MERKT, 1989). Es ist davon auszugehen,
daß die reduzierte Ausbildung der AM-Strukturen in den gepfropften Varianten (s. Tab. 5.9)
auf die begrenzte Verfügbarkeit der Assimilate in den Wurzeln zurückgeführt werden kann.
Trotz der durch die Mykorrhizierung reduzierten Wurzellängendichte wurden insbesondere
von den nur schwer beweglichen Nährstoffen P, Cu und Zn erhöhte Gehalte in den Blättern
ermittelt
(s. Tab. 5.7, 5.8, 5.12 und 5.13). Die eigenen Ergebnissen zeigen, daß die Besiedlung der
Wurzeln mit AM zu verstärktem Triebwachstum führt. Es kann davon ausgegangen werden,
daß die Ausbildung des externen Hyphengeflechts eine wichtige Rolle bei der Nährstoffauf-
nahme spielt. Die von LI et al. (1991 c) beschriebene Wechselwirkung bei der Nährstoffauf-
nahme von Cu und Zn mit der P-Aufnahme über die Hyphen wurde in der vorliegenden Arbeit
bestätigt. Die höheren Cu- und Zn-Gehalte gingen oftmals mit erhöhten P-Gehalten einher.
Die Erfassung des Wurzelsystems bei der Dauerkultur Rebe stellt eine sehr arbeitsaufwendige
Aufgabe dar. Bereits 1903 hat MÜLLER-THURGAU auf die Schwierigkeiten bei der Erfassung
des Wurzelsystems von Reben hingewiesen. In der vorliegenden Arbeit konnte eine durch-
schnittliche Wurzellängendichte an Reben, ausgegraben aus einem definierten Bodenvolumen
unmittelbar in Stocknähe, von 0,041 cm/cm3 Boden in Ertragsanlagen (s. Kap. 3.3.2, Tab. 3.5)
ermittelt werden. Die eigenen Versuchsergebnisse stehen in Einklang mit den Ergebnissen
von MOHR (1996), der bei Untersuchungen mit Müller-Thurgau/5BB durchschnittliche Wur-

Kapitel 6 Abschließende Diskussion 138
zellängendichten von 0,035 cm Wurzellänge je ml Boden feststellte. Die niedrigen Werte ver-
anschaulichen die geringe Durchwurzelungsintensität von Reben und weisen auf die Proble-
matik für die Wurzelgewinnung mit der Kernbohrermethode hin. Um nach BÖHM (1979) auf-
grund der ungleichmäßigen Verteilung der Rebwurzeln reproduzierbare Ergebnisse zu erhal-
ten, wurde in der vorliegenden Untersuchung mit einer großen Anzahl von Wiederholungen
gearbeitet. Die Wurzelausbeute war allerdings sehr gering. Zu gleichen Ergebnissen kamen
PERRY et al. (1983) und empfahlen beim Einsatz von Kernbohrern zur Untersuchung der
Wurzelverteilung im Boden Bohrkerne mit größerem Durchmesser zu verwenden. MOHR
(1994) untersuchte verschiedene Methoden zur Erfassung der Wurzelverteilung im Weinberg
und berichtet von unzureichenden Ergebnissen der Kernbohrermethode, insbesondere bei stei-
nigen und trockenen Böden. In der vorliegenden Arbeit konnte in der Versuchsanlage „Hun-
dertmorgen“ im tiefgründigen Lößlehm ohne großen Arbeitsaufwand die Kernbohrermethode
angewendet werden. Jedoch sollten für weitere Wurzeluntersuchungen in Weinbergen die in
den Ertragsanlagen in Kapitel 3.3.2 beschriebene Untersuchungsmethode angewendet werden,
bei der aus einem definierten Bodenvolumen ausreichende Mengen an Rebwurzeln für Unter-
suchungszwecke gewonnen werden konnte.
Die in dieser Arbeit durchgeführten Untersuchungen in Rebschulen in Kapitel 3.3.1 haben
gezeigt, daß neben dem Vorkommen der AM in Ertragsanlagen auch die jungen Pfropfreben
in Rebschulen mykorrhiziert sind. Die Untersuchung ergab einen Hinweis darauf, daß das
Vorkommen der AM-Pilze an den Pfropfreben negativ mit dem Boden-P-Gehalt korrelierte.
Die insgesamt geringeren AM-Infektionsgrade in Rebschulen im Vergleich zu den eigenen
Ergebnissen aus Ertragsanlagen und zu Literaturangaben (SCHUBERT und CRAVERO, 1985;
NAPPI et al., 1985; BRENDEL et al., 1990) resultieren möglicherweise aus der kurzen Kulturzeit
von nur wenigen Monaten in Rebschulen. Weitaus wichtiger könnte jedoch das Fehlen von
Ackerbeikräutern, insbesondere der AM-Wirtspflanzen in Rebschulen sein, da durch die
mehrmalige Bodenbearbeitung während der Vegetationsperiode der direkte Eintrag von AM-
Infektionspotential durch AM-Wirtspflanzen nicht gegeben ist. Ähnliche Ergebnisse wurden
bisher von WULFF (1996) in landwirtschaftlichen Kultursystemen beschrieben. Trotz der ins-
gesamt geringen Mykorrhizierung der Rebwurzeln im Freiland kann von einer Wachstums-
förderung der nativen Mykorrhiza im Weinberg ausgegangen werden, wie der Versuch in
Kapitel 5.3.1 zeigte. Bei einem durchschnittlichen AM-Infektionsgrad von nur 12 %, hervor-

Kapitel 6 Abschließende Diskussion 139
gerufen durch die native AM eines Rebschulbodens, wurden an den mykorrhizierten Grün-
stecklingen teilweise signifikant erhöhte Sproßtrockenmassen ermittelt.
In den Gefäßversuchen traten beim gezielten Einsatz von AM-Pilzen und dem Einpflanzen
von Rebenstecklingen in Böden mit nativer AM Wachstumsförderungen an den Reben auf.
Die durchgeführten Mineralstoffanalysen zeigten bei fast allen Stecklingen erhöhte Nährstoff-
gehalte in den Blättern auf. Bei gleichzeitig gesteigerter Trockensubstanzproduktion läßt sich
somit auf eine erhöhte Nährstoffaufnahme der mykorrhizierten Reben schließen. Daneben
werden in der Literatur zahlreiche nicht nährstoffspezifische Effekte der AM-Pilze diskutiert.
EIBACH (1982) registrierte in ihren Untersuchungen mit Zweiaugenstecklingen der Rebsorte
Bacchus eine frühere Seitenwurzelbildung an den mit Glomus macrocarpus inokulierten Re-
ben, wobei die Autorin diese Effekte auf eine innere Beeinflussung des Phytohormonhaushal-
tes zurückführte. DRÜGE (1992) konnte an Leinpflanzen zeigen, daß eine höhere Cytoki-
ninproduktion des mykorrhizierten Wurzelsystems maßgeblich für Wachstumssteigerungen
verantwortlich waren. Einen Nährstoffeffekt konnte der Autor ausschließen, denn die Nähr-
stoffgehalte bei mykorrhiziertem und nicht mykorrhiziertem Lein waren nahezu unverändert.
Da in den Versuchen mit Wurzelkästen (s. Kap. 5.3.2) und grünveredelten Stecklingen (s.
Kap. 5.3.3) ein P-armer Rebschulboden eingesetzt wurde, konnten die aufgetretenen Mykor-
rhizaeffekte auf einen P-Effekt zurückgeführt werden. Dagegen lassen die deutlichen Wachs-
tumssteigerungen bei den Topfreben (s. Kap. 4.3.2) während der Anzuchtphase im Gewächs-
haus weitere Wirkungsmechanismen vermuten, da beide Kultursubstrate sehr hohe P-Gehalte
aufwiesen
(s. Tab. 4.2). Zwar konnten nach 35tägiger Anzuchtphase signifikant höhere P-Gehalte in den
mykorrhizierten Pflanzen beobachtet werden, die allerdings im Freiland nach dem Auspflan-
zen an den präinokulierten Topfreben nicht mehr festgestellt wurden. Trotzdem bestanden in
den mit Floraton 1 angezogenen Reben im Freiland auch nach 10 Wochen Versuchsdauer
signifikante Wachstumsunterschiede zwischen den Reben ohne und mit AM-Beimpfung. Ne-
ben der Wachstumsförderung führt eine Besiedlung mit AM-Pilzen gegenüber biotischen und
abiotischen Faktoren zu einer verbesserten Streßresistenz der Wirtspflanze. FELDMANN (1990)
konnte bei seinen Untersuchungen mit Holzstecklingen von Kautschukbäumen keine mykor-
rhizabedingten Wachstumssteigerungen beobachten. Dagegen fand der Autor an den Blättern
der Kautschukbaumpflanzen mykorrhizainduzierte Veränderungen im Resistenzverhalten ge-
gen Blattpathogene. Es ist denkbar, daß die mykorrhizierten Topfreben im Weinberg eine hö-

Kapitel 6 Abschließende Diskussion 140
here Toleranz gegenüber biotischen oder abiotischen Stressoren erhalten haben, was bereits in
zahlreichen Untersuchungen an Reben und anderen Pflanzenarten bestätigt werden konnte
(MCCOOL et al., 1979; DEHNE, 1994; WASCHKIES et al., 1994). In der Versuchsfläche „Rup-
pertsberger Linsenbusch“ wurden vor dem Pflanzen der Topfreben im Rahmen einer Flurbe-
reinigung Erdbewegungsmaßnahmen durchgeführt. Möglicherweise wurden durch die um-
fangreichen Bodenbewegungen mit zum Teil schweren Gerätschaften das natürliche My-
korrhizierungspotential durch die Zerstörung des externen Hyphengeflechts deutlich reduziert
(JASPER et al., 1989, 1991). Dies hatte zur Folge, daß einige Unkräuter, die wichtige AM-
Wirtspflanzen sind und nach den eigenen Ergebnissen aus Kapitel 3 die Mykorrhizierung der
Rebwurzeln verbessern, entfernt wurden. Durch die gezielte Ausbringung präinokulierter Re-
ben dürfte sich das Infektionspotential auf diesem Standort erhöhen bzw. wieder neu aufbau-
en.
Die Anwendung der AM in der Topfrebenproduktion (s. Kap. 4.3.2) führte während der An-
zuchtphase unter Gewächshausbedingungen an den mykorrhizierten Topfreben zu signifikant
größeren Trieblängen. Anhand des Versuchs mit Wurzelkästen (s. Kap. 5.3.2) konnte darge-
stellt werden, daß Rebwurzeln relativ schnell durch das Inokulumband hindurchwachsen und
die weitere Ausbreitung der Mykorrhiza-Infektion innerhalb des Wurzelsystems nur unzurei-
chend ist. Das gleiche gilt für Topfreben, bei denen die Wurzeln frühzeitig das Boden-
Inokulum-Gemisch verlassen und in mykorrhizafreies Substrat wachsen. Um höhere Infekti-
onsgrade an den Topfreben zu erreichen und den Mykorrhizabeitrag zu optimieren, wäre eine
weitere Inokulation des Torfbeets denkbar, auf dem sich die Topfreben während der An-
zuchtphase befinden. Somit hätten die wachsenden Rebwurzeln nach dem Durchwachsen der
Pflanzgefäße weiterhin Kontakt mit dem Mykorrhiza-Inokulum und die Wahrscheinlichkeit
einer höheren Besiedlung mit dem AM-Pilz würde ansteigen. Nachteilig an diesem modifi-
zierten System wäre eine höhere Inokulummenge, die für die Inokulation des Pflanzbeets auf-
gebracht werden müßte. Ein weiterer Lösungsansatz könnte durch den gezielten Einsatz einer
Pilzspezies erbracht werden, die eine große Menge an externen Hyphen ausbildet, über die
durch Sekundärinfektionen wachsende Rebwurzeln infiziert werden. Die Anwendung der
AM-Pilze in Rebschulen erscheint aufgrund der aus dem Gefäßversuch mit den Wurzelkästen
gewonnenen Ergebnisse nur schwer unter praxisrelevanten Bedingungen durchführbar. Die
Infektionswahrscheinlichkeit in Rebschulen ist bedeutend kleiner als in Pflanzgefäßen. Trotz
der hohen Inokulummenge von 100 ml/laufendem Meter Mulchfolie lag der Infektionsgrad

Kapitel 6 Abschließende Diskussion 141
mit
12 % bzw. 17 % (vgl. Kap. 4.3.1, Tab. 4.6 und 4.7) wesentlich niedriger als der in Gefäßver-
suchen erzielte. Um ein Hektar Rebschule inokulieren zu können, müßten etwa 1.000 l My-
korrhiza-Inokulum als sporenbeladene Blähtonpartikel ausgebracht werden. Freilandinokula-
tionen wären dann für den Praktiker von Vorteil, wenn die Aufwandmenge reduziert wird und
die Applikationsmethode mit praxisüblichen Geräten mit verhältnismäßig geringen Modifizie-
rungen durchgeführt werden könnte, was BALTRUSCHAT (1987 a, b) an Mais feststellen konn-
te. Eine Bekämpfungsmöglichkeit von rebmüdem Boden durch die Interaktion zwischen den
AM-Pilzen und den Mikroorganismen der Rhizosphäre, wie WASCHKIES et al. (1994) sie in
Gefäßversuchen aufzeigen konnte, wäre eine interessante Anwendungsmöglichkeit für AM-
Pilze in Rebschulen. Die Folge wäre eine längere Nutzung von rebschulfähigen Böden als
Rebschulflächen.
Neben der gezielten Anwendung der AM-Pilze in der Rebenpflanzguterzeugung (s. Kap. 4)
zeigten die Ergebnisse der vorliegenden Arbeit, daß indirekte Maßnahmen die native Mykor-
rhiza im Weinbergsboden erhalten oder sogar fördern können. Wie aus dem Begrünungsver-
such in Kapitel 3.3.3 hervorging, führte eine Begrünungseinsaat mit Gramineen oder Legumi-
nosen im Vergleich zur offenen Bodenhaltung zu einer Erhöhung des AM-Infektionsgrades
der Rebwurzeln. Demgegenüber konnte bei den Varianten mit Cruciferen als Begrünungs-
pflanzen an den Rebwurzeln eine niedrigere AM-Infektion festgestellt werden als in den Vari-
anten, in denen mykotrophe Begrünungspflanzen wie Leguminosen oder Gräser eingesät wa-
ren. Dies konnte ebenfalls in einem Rebschulboden („Standort Harthäuser“,
s. Kap. 4.3.1) bestätigt werden. Die eigenen Ergebnisse stimmen mit denen aus landwirt-
schaftlichen Fruchtfolgesystemen überein, in denen durch die Verwendung von Nicht-
Wirtspflanzen für die AM das Mykorrhiza-Potential im Boden verringert wurde (BALTRU-
SCHAT und DEHNE, 1988; HARINIKUMAR und BAGYARAJ, 1988; LAND, 1990; WULFF, 1996).
Hieraus ergeben sich Empfehlungen für die Praxis, beim Einsatz von Begrünungspflanzen in
Ertragsanlagen gezielt AM-Wirtspflanzen wie z. B. Gramineen oder Leguminosen zu verwen-
den und auf Cruciferen weitgehend zu verzichten. Aufgrund von auftretenden Bodenmüdig-
keitserscheinungen werden Rebschulböden nur in einem Turnus von 3 - 8 Jahren als Rebschu-
len genutzt. Deshalb sollten in den dazwischenliegenden Jahren nur landwirtschaftliche
Wirtspflanzen für die AM kultiviert werden. Ölrettich hat in Weinbergen als Begrünungs-
pflanze eine Bedeutung als Feindpflanze gegen einige virusübertragende Nematoden. Hierzu

Kapitel 6 Abschließende Diskussion 142
zählen die ektoparasitisch lebende Nematodenart Xiphinema index, die als Überträger des
Grapevine-Fanleaf-Virus gilt (RÜDEL et al., 1992; BLEYER und FOX, 1995). Es ist deshalb zu
empfehlen, Ölrettich in Weinbergen als Begrünungspflanze nur in einem geringen Anteil in
der Begrünungsmischung einzusetzen, um einerseits die nematoden-reduzierenden Effekte
ausnutzen zu können, aber andererseits die native AM nicht nachhaltig zu verringern. Auch
bei der Auswahl von Rebschulflächen sollte verstärkt darauf geachtet werden, daß die Pflan-
zenarten der Vorkulturen AM-Wirtspflanzen sind.
Insgesamt tragen die AM-Pilze zu einer effizienteren Nährstoffaufnahme der Rebe bei. Um
das Mykorrhizierungspotential im Boden zu verbessern, haben die Inokulationsversuche in
Rebschulen und in der Topfrebenproduktion gezeigt, daß an mykorrhizierten Reben das
Wachstum gefördert wurde. Dazu haben die nicht nährstoffspezifischen Eigenschaften der
Mykorrhiza möglicherweise genauso beigetragen wie die direkte Beteiligung der AM-Pilze an
der Nährstoffaufnahme. EIBACH (1982) testete verschiedene Mykorrhiza-Arten auf ihre Eig-
nung als Symbiosepartner für die Rebe und stellte in Bezug auf Wachstum und Substanzertrag
deutliche Unterschiede zwischen den Arten fest. Für weitere Inokulationsversuche an Reben
würden sich die in der vorliegenden Arbeit getesteten AM-Arten anbieten. Durch indirekte
Maßnahmen wie Begrünungseinsaaten mit mykotrophen Pflanzenarten läßt sich in Ertragsan-
lagen und Rebschulen das AM-Potential erhöhen.

Zusammenfassung 143
7 Zusammenfassung
Im umweltschonenden Weinbau wird die Erhaltung bzw. Förderung der natürlichen Boden-
fruchtbarkeit im Ökosystem Weinberg angestrebt. Dazu können arbuskuläre Mykorrhiza
(AM)- Pilze, die in Symbiose mit der Rebe leben, einen erheblichen Beitrag leisten. In der
vorliegenden Arbeit wurden im Freiland Untersuchungen über das Vorkommen von AM-
Pilzen sowie Möglichkeiten über deren praktische Anwendung in der Rebenpflanzguterzeu-
gung durchgeführt. Weiterhin wurde die Bedeutung der AM für die Nährstoffaufnahme und
das Wachstum an Rebenstecklingen untersucht. Hierzu wurden in den Jahren 1994 - 1997
Inokulationsversuche mit Rebenstecklingen im Gewächshaus durchgeführt.
Der AM-Infektionsgrad in Ertragsanlagen und Rebschulen aus dem Anbaugebiet „Pfalz“ wur-
de bestimmt. Ziel war es festzustellen, ob das quantitative Vorkommen der AM an Reben mit
der Bewirtschaftungsweise (organisch-biologisch, kontrolliert umweltschonend, konventio-
nell), dem Boden-P-Gehalt und/oder der Unterlagssorte korrelierte. In einer weiteren Untersu-
chung wurde der Einfluß verschiedener Begrünungspflanzen und Bodenpflegemaßnahmen
(offene Gassen/begrünte Gassen) auf die native Mykorrhizierung der Rebwurzeln im Wein-
berg überprüft. Die Probenahme zur Überprüfung des AM-Infektionsgrades von Rebwurzeln
aus unterschiedlichen Bodentiefen eines Versuchsweinberges mit sieben verschiedenen Be-
grünungsvarianten und einer offen gehaltenen Variante erfolgte mit Hilfe eines Kernbohrers.
Ergänzend dazu wurde das Mykorrhizierungspotential im Boden aus den jeweiligen Varianten
mit Hilfe eines Biotests mit Weißklee im Gewächshaus ermittelt. In einem weiteren Versuch
wurden verschiedene AM-Isolate in der Rebenpflanzguterzeugung unter praxisnahen Bedin-
gungen eingesetzt. Ziel war es festzustellen, ob durch die Inokulation an Pfropfreben in der
Rebschule sowie an Topfreben im Gewächshaus das Wachstum der Reben verbessert werden
kann. Besonderes Augenmerk galt dem Verlauf des AM-Infektionsgrades nach Inokulation in
der Rebschule, der neben den Reben auch an den Indikatorpflanzen Weißklee und Mais be-
stimmt
wurde.
In Gefäßversuchen, die die Freilanduntersuchungen begleiteten, wurde der Einfluß der nativen
Mykorrhiza eines Rebschulbodens auf Nährstoffaufnahme und Wachstum an verschiedenen

Zusammenfassung 144
Grünstecklingen überprüft. Weiterhin wurde die Ausbreitung der AM-Infektion im Wurzel-
system untersucht. Ziel war es festzustellen, inwieweit Rebwurzeln nach Durchwachsen eines
Inokulumbandes (Glomus mosseae), welches je nach Versuchsvariante in zwei unterschiedli-
chen Bodentiefen in Wurzelkästen angelegt wurde, mit dem AM-Pilz infiziert werden. In ei-
nem weiteren Versuchsansatz wurde geprüft, ob die Pfropfung von Grünstecklingen die My-
korrhizierung beeinflußt und welche Auswirkungen dies auf Nährstoffgehalte und Wachstum
der Reben hat. Weiterhin wurde der Frage nachgegangen, welchen Einfluß eine erhöhte O-
zonbelastung der Luft auf den Infektionsgrad und das Wachstum von Reben hat.
Im einzelnen können die gewonnenen Ergebnisse wie folgt zusammengefaßt werden:
1. In Rebschulflächen wurde an den Pfropfkombinationen Riesling/SO4 und Müller-
Thurgau/5BB ein durchschnittlicher AM-Infektionsgrad von 6,7 % ermittelt. Es konnte eine
negative, statistisch nicht abgesicherte Korrelation zwischen dem Boden-P-Gehalt und dem
Auftreten der AM-Infektion in Rebschulen festgestellt werden. In Ertragsanlagen wurde im
Durchschnitt aller beprobten Standorte ein AM-Infektionsgrad von 12,2 % ermittelt. Es konn-
te eine durchschnittliche Wurzellängendichte an Reben, ausgegraben aus einem definierten
Bodenvolumen, von 0,041 cm/cm3 Boden ermittelt werden. Eine eindeutige Aussage über den
Einfluß der Unterlagssorte sowie der Bewirtschaftungsweise auf die Mykorrhizierung konnte
aufgrund der vorliegenden Ergebnisse nicht getroffen werden.
2. In den Begrünungsvarianten „Wintergründüngung mit Gramineen“, „Wintergründüngung
mit Leguminosen“ und „2-Jahresbegrünung mit Gramineen“ wurden mit 14,5 % und 15,0 %
bzw. 12,2 % signifikant höhere AM-Infektionsgrade an den Rebwurzeln ermittelt als in der
unbewachsenen Kontrolle mit 6 %. Dagegen wurden in den Begrünungsvarianten mit Crucife-
reneinsaaten tendenziell niedrigere AM-Infektionsgrade an den Rebwurzeln festgestellt als in
den anderen Begrünungseinsaaten. Es konnte nachgewiesen werden, daß von den AM-
Nichtwirtspflanzen Winterraps, Ölrettich, Gelbsenf und Winterrübsen ein hemmender Einfluß
auf die Mykorrhizierung der Rebwurzeln ausging. Obwohl in der oberen Bodenschicht
(0 - 25 cm) eine hohe P-Verfügbarkeit auftrat, wurde im Oberboden das größte Mykorrhizie-
rungspotential anhand eines Biotests mit Weißklee bestimmt. Dies wird im Zusammenhang
mit P-toleranten AM-Pilzspezies in Weinbergsböden diskutiert.

Zusammenfassung 145
3. Durch die Inokulation mit dem AM-Pilz Glomus sp. wurde der Infektionsgrad an Pfropfre-
ben (Riesling/5C bzw. Müller-Thurgau/5BB) auf zwei Rebschulstandorten signifikant erhöht.
Der Inokulationserfolg war an den Indikatorpflanzen Weißklee und Mais mit 9,2 % bzw. 14 %
deutlich erkennbar. Nach 12- bzw. 14wöchiger Wachstumsphase wurden an den inokulierten
Pfropfreben signifikant längere Triebe gemessen als an den nicht inokulierten. In den Kon-
trollvarianten ohne zusätzliche Inokulation wurden an Rebwurzeln und an den Indikatorpflan-
zen Weißklee und Mais nur in geringem Umfang eine native AM festgestellt, die im Zusam-
menhang mit der hohen P-Verfügbarkeit und der eventuell auftretenden Fruchtfolgeeffekte
diskutiert wird.
4. Die Inokulation mit Glomus sp. und Glomus mosseae führte an Topfreben nach 4- bzw.
8wöchiger Anzuchtphase zu einer Mykorrhizierung der Rebwurzeln und bewirkte signifikant
größere Trieblängen. Die Wachstumsunterschiede lassen sich durch die höheren Nährstoffge-
halte in den Blättern der mykorrhizierten Reben erklären. Nach dem Auspflanzen der präino-
kulierten Topfreben ins Freiland trat in der mit Floraton eingesetzten Versuchsvariante ein
stärkeres Trieblängenwachstum auf. Da keine unterschiedlichen Nährstoffgehalte im Freiland
zwischen den nicht beimpften und beimpften Versuchsvarianten bestanden, sind offensicht-
lich nicht nährstoffspezifische Mykorrhizaeffekte für das verbesserte Wachstum verantwort-
lich.
5. Die native AM eines Rebschulbodens führte in einem Gefäßversuch an Grünstecklingen
verschiedener Rebsorten zu einem durchschnittlichen AM-Infektionsgrad von 12 %. Hinsicht-
lich der Mykorrhizierung wurden signifikante Unterschiede an den Sorten Börner (17,4 %)
und A9 (6,8 %) festgestellt. An einigen Sorten bewirkte die native AM signifikant höhere
Sproßtrockenmassen, das Wurzeltrockengewicht war dagegen unbeeinflußt. Bei den infizier-
ten Stecklingen wurde ein signifikant vergrößertes Sproß-/Wurzelverhältnis mit durchschnitt-
lich 5,9 ermittelt als in den nicht infizierten (4,1). Es konnte gezeigt werden, daß durch My-
korrhizierung bei den meisten Sorten die P-, Cu- und Zn-Gehalte in den Blättern erhöht wa-
ren, während die Gehalte an Mn und Fe in den Blättern mykorrhizierter Reben oftmals signi-
fikant verringert waren.
6. Die Inokulation mit Glomus mosseae führte in Wurzelkästen an Wurzeln der Unterlagssor-
te SO4 unmittelbar im Inokulationsbereich in beiden Versuchsvarianten zu AM-
Infektionsgraden von 45 % bzw. 35 %. Mit zunehmendem Abstand vom Inokulumband waren

Zusammenfassung 146
die Rebwurzeln geringer mykorrhiziert bzw. es konnte keine AM-Infektion festgestellt wer-
den. Durch die Inokulation im oberen Bodenbereich des Wurzelkastens (9 - 18 cm) wurde das
Trockengewicht der SO4-Stecklinge signifikant erhöht. Bereits eine Teilbesiedlung des Wur-
zelsystems mit Glomus mosseae führte zu Inokulationserfolgen in Form von erhöhter Tro-
ckensubstanzbildung und verbesserter Nährstoffaufnahme.
7. Die Inokulation mit Glomus intraradices führte in der wurzelechten Variante Riesling
(38,5 %) zu signifikant höheren Infektionsgraden als in der heteroplastischen Pfropfvariante
mit der Unterlagssorte SO4 (26,2 %). In der homoplastischen Pfropfvariante war der Infekti-
onsgrad tendenziell verringert. Es konnte gezeigt werden, daß in den wurzelechten Varianten
Riesling und SO4 die Sproßtrockenmasse in den inokulierten Varianten teilweise signifikant
erhöht war, die Wurzellängendichte wurde dagegen signifikant verringert. Das Sproß-
/Wurzelverhältnis war bei Riesling in der inokulierten Variante signifikant erhöht.
8. Eine 3wöchige Ozonbelastung der Luft führte bei mykorrhizierten und nicht mykorrhizier-
ten Rebenstecklingen gleichermaßen zu keinen sichtbaren Blattschäden. Zwischen den Ver-
suchsvarianten (ohne bzw. mit Ozon und Außenluft) traten an den mykorrhizierten Reben
keine unterschiedlichen AM-Infektionsgrade auf. Die mykorrhizierten Reben der Ozon- und
Außenluftvariante (mit natürlicher Schadstoffbelastung) reagierten mit einem verbesserten
Wachstum im Vergleich zu den nicht mykorrhizierten Reben. Dies hängt möglicherweise mit
den höheren Nährstoffgehalten in den Blättern der mykorrhizierten Reben zusammen.
Die Untersuchungsergebnisse zeigen, daß die AM-Pilze Glomus sp., Glomus mosseae und
Glomus intraradices die Sproßtrockenmasse und die Mineralstoffgehalte von P, Cu und Zn in
den Blättern von Rebenstecklingen erhöhten. Es konnte gezeigt werden, daß durch den geziel-
ten Einsatz von geeigneten AM-Pilzen in der Rebenpflanzguterzeugung eine Wachstumsför-
derung eintritt. Weiterhin wurde nachgewiesen, daß durch indirekte Maßnahmen wie bei-
spielsweise eine Einsaat von mykotrophen Begrünungspflanzen wie Gramineen oder Legumi-
nosen in Ertragsanlagen und in rebschulfähigen Böden das AM-Potential im Boden erhöht
wird. Die Praxis kann sich diese Effekte beispielsweise durch Einsaat eines Gräser-
Leguminosen-Gemisches zunutze machen.
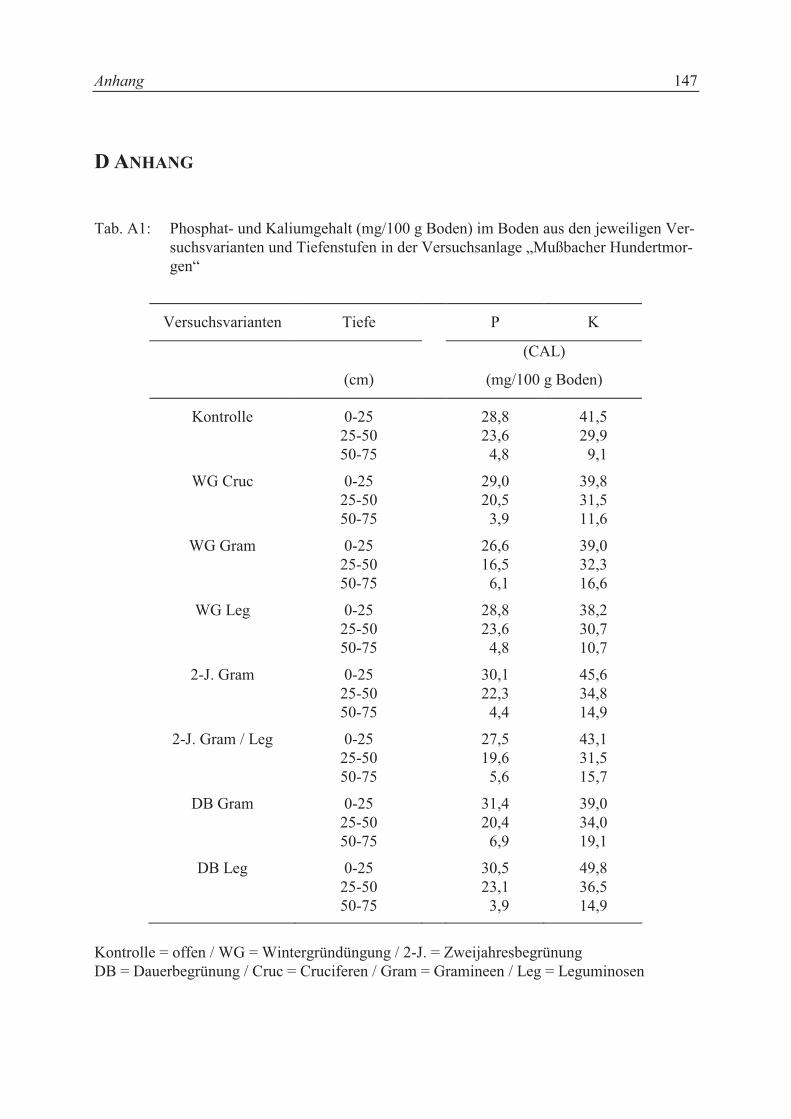
Anhang 147
D ANHANG
Tab. A1: Phosphat- und Kaliumgehalt (mg/100 g Boden) im Boden aus den jeweiligen Ver-suchsvarianten und Tiefenstufen in der Versuchsanlage „Mußbacher Hundertmor-gen“
Versuchsvarianten Tiefe P K
(CAL)
(cm) (mg/100 g Boden)
Kontrolle 0-25 28,8 41,5 25-50 23,6 29,9 50-75 4,8 9,1
WG Cruc 0-25 29,0 39,8 25-50 20,5 31,5 50-75 3,9 11,6
WG Gram 0-25 26,6 39,0 25-50 16,5 32,3 50-75 6,1 16,6
WG Leg 0-25 28,8 38,2 25-50 23,6 30,7 50-75 4,8 10,7
2-J. Gram 0-25 30,1 45,6 25-50 22,3 34,8 50-75 4,4 14,9
2-J. Gram / Leg 0-25 27,5 43,1 25-50 19,6 31,5 50-75 5,6 15,7
DB Gram 0-25 31,4 39,0 25-50 20,4 34,0 50-75 6,9 19,1
DB Leg 0-25 30,5 49,8 25-50 23,1 36,5 50-75 3,9 14,9
Kontrolle = offen / WG = Wintergründüngung / 2-J. = Zweijahresbegrünung DB = Dauerbegrünung / Cruc = Cruciferen / Gram = Gramineen / Leg = Leguminosen
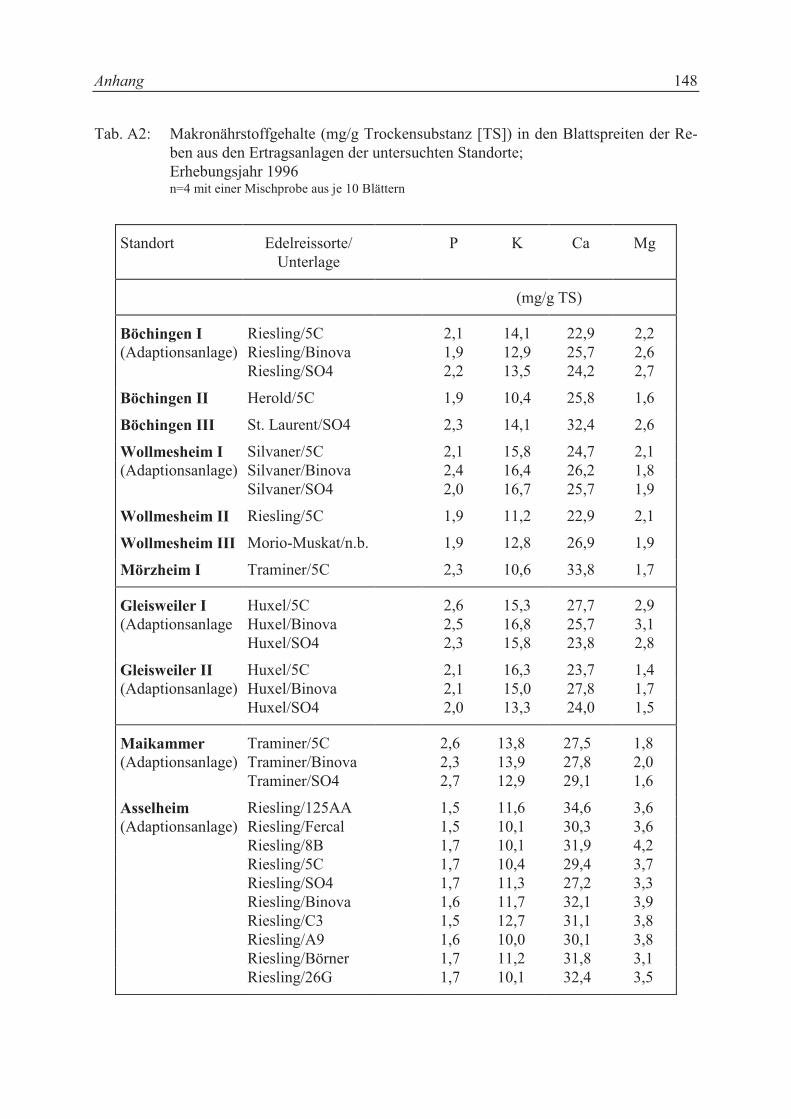
Anhang 148
Tab. A2: Makronährstoffgehalte (mg/g Trockensubstanz [TS]) in den Blattspreiten der Re-ben aus den Ertragsanlagen der untersuchten Standorte;
Erhebungsjahr 1996 n=4 mit einer Mischprobe aus je 10 Blättern
Standort Edelreissorte/ Unterlage
P K Ca Mg
(mg/g TS)
Böchingen I Riesling/5C 2,1 14,1 22,9 2,2 (Adaptionsanlage) Riesling/Binova 1,9 12,9 25,7 2,6 Riesling/SO4 2,2 13,5 24,2 2,7
Böchingen II Herold/5C 1,9 10,4 25,8 1,6
Böchingen III St. Laurent/SO4 2,3 14,1 32,4 2,6
Wollmesheim I Silvaner/5C 2,1 15,8 24,7 2,1 (Adaptionsanlage) Silvaner/Binova 2,4 16,4 26,2 1,8 Silvaner/SO4 2,0 16,7 25,7 1,9
Wollmesheim II Riesling/5C 1,9 11,2 22,9 2,1
Wollmesheim III Morio-Muskat/n.b. 1,9 12,8 26,9 1,9
Mörzheim I Traminer/5C 2,3 10,6 33,8 1,7
Gleisweiler I Huxel/5C 2,6 15,3 27,7 2,9 (Adaptionsanlage Huxel/Binova 2,5 16,8 25,7 3,1 Huxel/SO4 2,3 15,8 23,8 2,8
Gleisweiler II Huxel/5C 2,1 16,3 23,7 1,4 (Adaptionsanlage) Huxel/Binova 2,1 15,0 27,8 1,7 Huxel/SO4 2,0 13,3 24,0 1,5
Maikammer Traminer/5C 2,6 13,8 27,5 1,8 (Adaptionsanlage) Traminer/Binova 2,3 13,9 27,8 2,0 Traminer/SO4 2,7 12,9 29,1 1,6
Asselheim Riesling/125AA 1,5 11,6 34,6 3,6 (Adaptionsanlage) Riesling/Fercal 1,5 10,1 30,3 3,6 Riesling/8B 1,7 10,1 31,9 4,2 Riesling/5C 1,7 10,4 29,4 3,7 Riesling/SO4 1,7 11,3 27,2 3,3 Riesling/Binova 1,6 11,7 32,1 3,9 Riesling/C3 1,5 12,7 31,1 3,8 Riesling/A9 1,6 10,0 30,1 3,8 Riesling/Börner 1,7 11,2 31,8 3,1 Riesling/26G 1,7 10,1 32,4 3,5
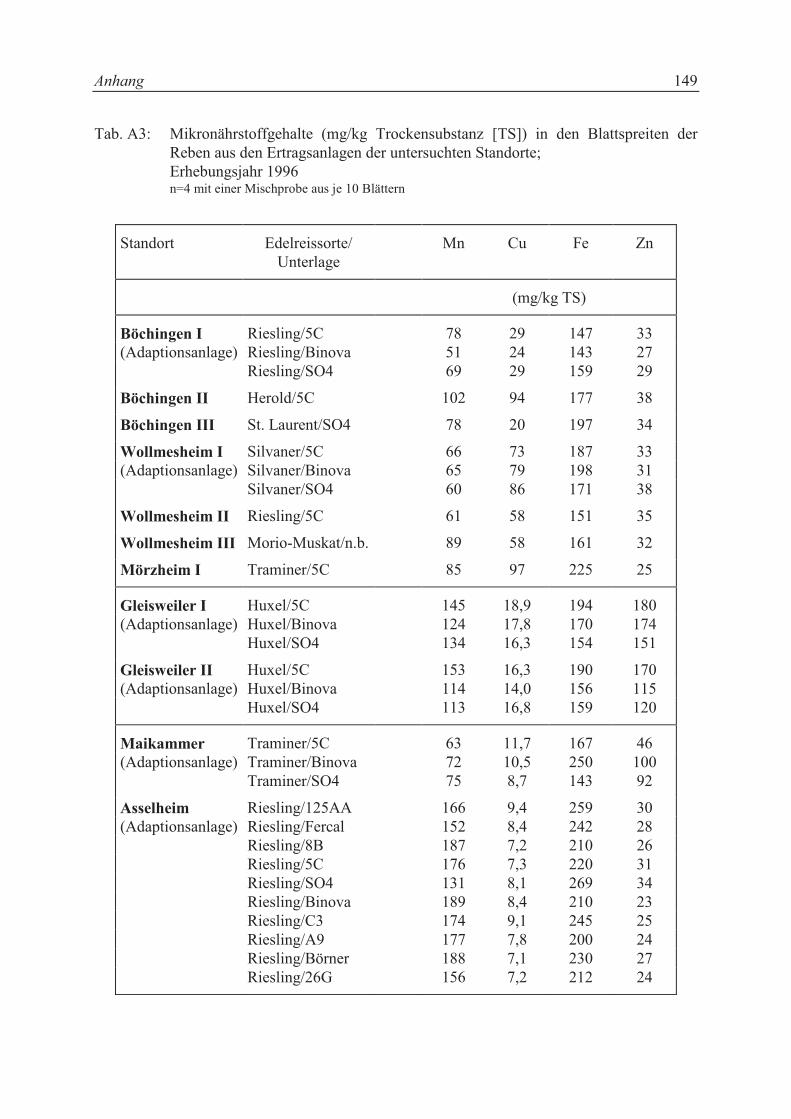
Anhang 149
Tab. A3: Mikronährstoffgehalte (mg/kg Trockensubstanz [TS]) in den Blattspreiten der Reben aus den Ertragsanlagen der untersuchten Standorte;
Erhebungsjahr 1996 n=4 mit einer Mischprobe aus je 10 Blättern
Standort Edelreissorte/ Unterlage
Mn Cu Fe Zn
(mg/kg TS)
Böchingen I Riesling/5C 78 29 147 33 (Adaptionsanlage) Riesling/Binova 51 24 143 27 Riesling/SO4 69 29 159 29
Böchingen II Herold/5C 102 94 177 38
Böchingen III St. Laurent/SO4 78 20 197 34
Wollmesheim I Silvaner/5C 66 73 187 33 (Adaptionsanlage) Silvaner/Binova 65 79 198 31 Silvaner/SO4 60 86 171 38
Wollmesheim II Riesling/5C 61 58 151 35
Wollmesheim III Morio-Muskat/n.b. 89 58 161 32
Mörzheim I Traminer/5C 85 97 225 25
Gleisweiler I Huxel/5C 145 18,9 194 180 (Adaptionsanlage) Huxel/Binova 124 17,8 170 174 Huxel/SO4 134 16,3 154 151
Gleisweiler II Huxel/5C 153 16,3 190 170 (Adaptionsanlage) Huxel/Binova 114 14,0 156 115 Huxel/SO4 113 16,8 159 120
Maikammer Traminer/5C 63 11,7 167 46 (Adaptionsanlage) Traminer/Binova 72 10,5 250 100 Traminer/SO4 75 8,7 143 92
Asselheim Riesling/125AA 166 9,4 259 30 (Adaptionsanlage) Riesling/Fercal 152 8,4 242 28 Riesling/8B 187 7,2 210 26 Riesling/5C 176 7,3 220 31 Riesling/SO4 131 8,1 269 34 Riesling/Binova 189 8,4 210 23 Riesling/C3 174 9,1 245 25 Riesling/A9 177 7,8 200 24 Riesling/Börner 188 7,1 230 27 Riesling/26G 156 7,2 212 24
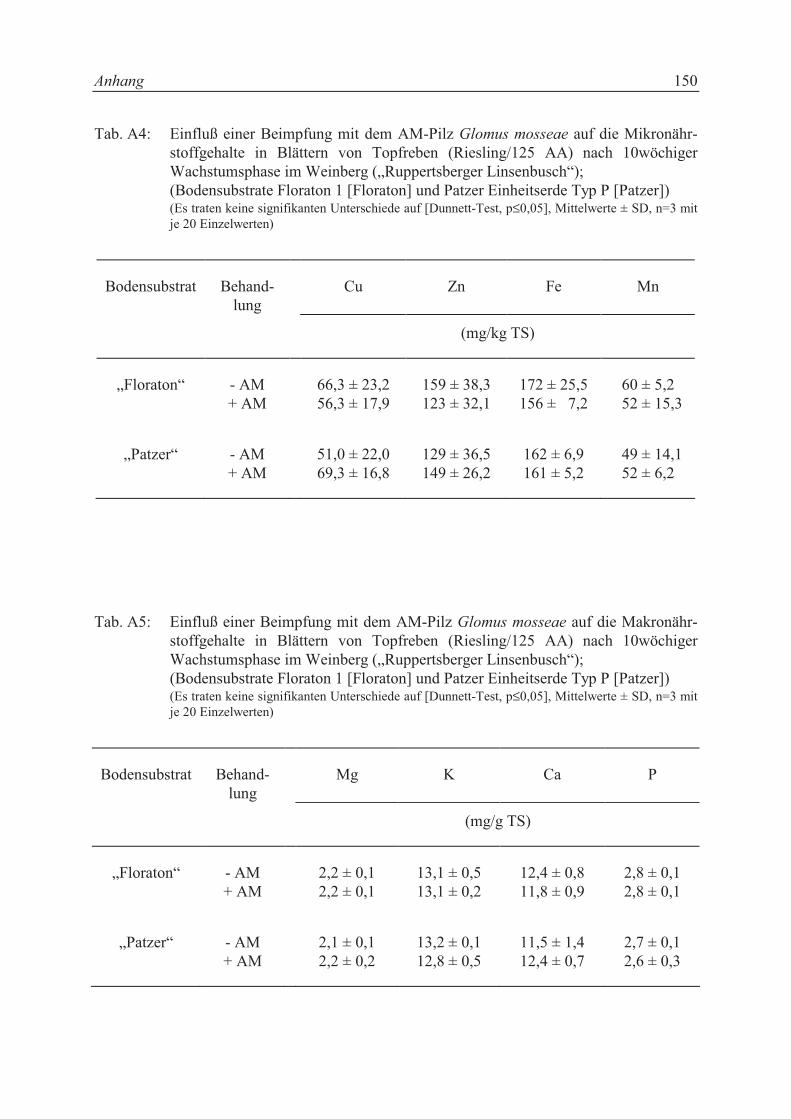
Anhang 150
Tab. A4: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf die Mikronähr-stoffgehalte in Blättern von Topfreben (Riesling/125 AA) nach 10wöchiger Wachstumsphase im Weinberg („Ruppertsberger Linsenbusch“);
(Bodensubstrate Floraton 1 [Floraton] und Patzer Einheitserde Typ P [Patzer]) (Es traten keine signifikanten Unterschiede auf [Dunnett-Test, p≤0,05], Mittelwerte ± SD, n=3 mit
je 20 Einzelwerten)
Bodensubstrat Behand-lung
Cu Zn Fe Mn
(mg/kg TS)
„Floraton“ - AM 66,3 ± 23,2 159 ± 38,3 172 ± 25,5 60 ± 5,2 + AM 56,3 ± 17,9 123 ± 32,1 156 ± 7,2 52 ± 15,3
„Patzer“ - AM 51,0 ± 22,0 129 ± 36,5 162 ± 6,9 49 ± 14,1 + AM 69,3 ± 16,8 149 ± 26,2 161 ± 5,2 52 ± 6,2
Tab. A5: Einfluß einer Beimpfung mit dem AM-Pilz Glomus mosseae auf die Makronähr-stoffgehalte in Blättern von Topfreben (Riesling/125 AA) nach 10wöchiger Wachstumsphase im Weinberg („Ruppertsberger Linsenbusch“);
(Bodensubstrate Floraton 1 [Floraton] und Patzer Einheitserde Typ P [Patzer]) (Es traten keine signifikanten Unterschiede auf [Dunnett-Test, p≤0,05], Mittelwerte ± SD, n=3 mit
je 20 Einzelwerten)
Bodensubstrat Behand-lung
Mg K Ca P
(mg/g TS)
„Floraton“ - AM 2,2 ± 0,1 13,1 ± 0,5 12,4 ± 0,8 2,8 ± 0,1 + AM 2,2 ± 0,1 13,1 ± 0,2 11,8 ± 0,9 2,8 ± 0,1
„Patzer“ - AM 2,1 ± 0,1 13,2 ± 0,1 11,5 ± 1,4 2,7 ± 0,1 + AM 2,2 ± 0,2 12,8 ± 0,5 12,4 ± 0,7 2,6 ± 0,3
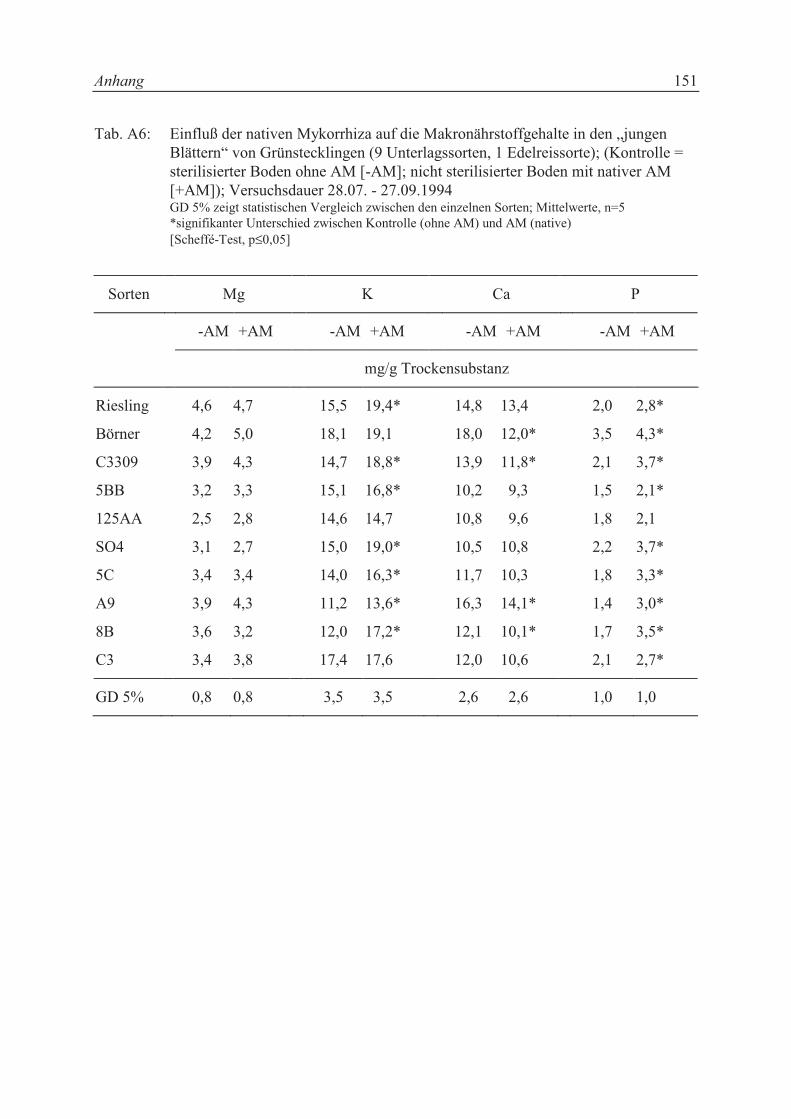
Anhang 151
Tab. A6: Einfluß der nativen Mykorrhiza auf die Makronährstoffgehalte in den „jungen Blättern“ von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte); (Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994
GD 5% zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Mg K Ca P
-AM +AM -AM +AM -AM +AM -AM +AM
mg/g Trockensubstanz
Riesling 4,6 4,7 15,5 19,4* 14,8 13,4 2,0 2,8*
Börner 4,2 5,0 18,1 19,1 18,0 12,0* 3,5 4,3*
C3309 3,9 4,3 14,7 18,8* 13,9 11,8* 2,1 3,7*
5BB 3,2 3,3 15,1 16,8* 10,2 9,3 1,5 2,1*
125AA 2,5 2,8 14,6 14,7 10,8 9,6 1,8 2,1
SO4 3,1 2,7 15,0 19,0* 10,5 10,8 2,2 3,7*
5C 3,4 3,4 14,0 16,3* 11,7 10,3 1,8 3,3*
A9 3,9 4,3 11,2 13,6* 16,3 14,1* 1,4 3,0*
8B 3,6 3,2 12,0 17,2* 12,1 10,1* 1,7 3,5*
C3 3,4 3,8 17,4 17,6 12,0 10,6 2,1 2,7*
GD 5% 0,8 0,8 3,5 3,5 2,6 2,6 1,0 1,0
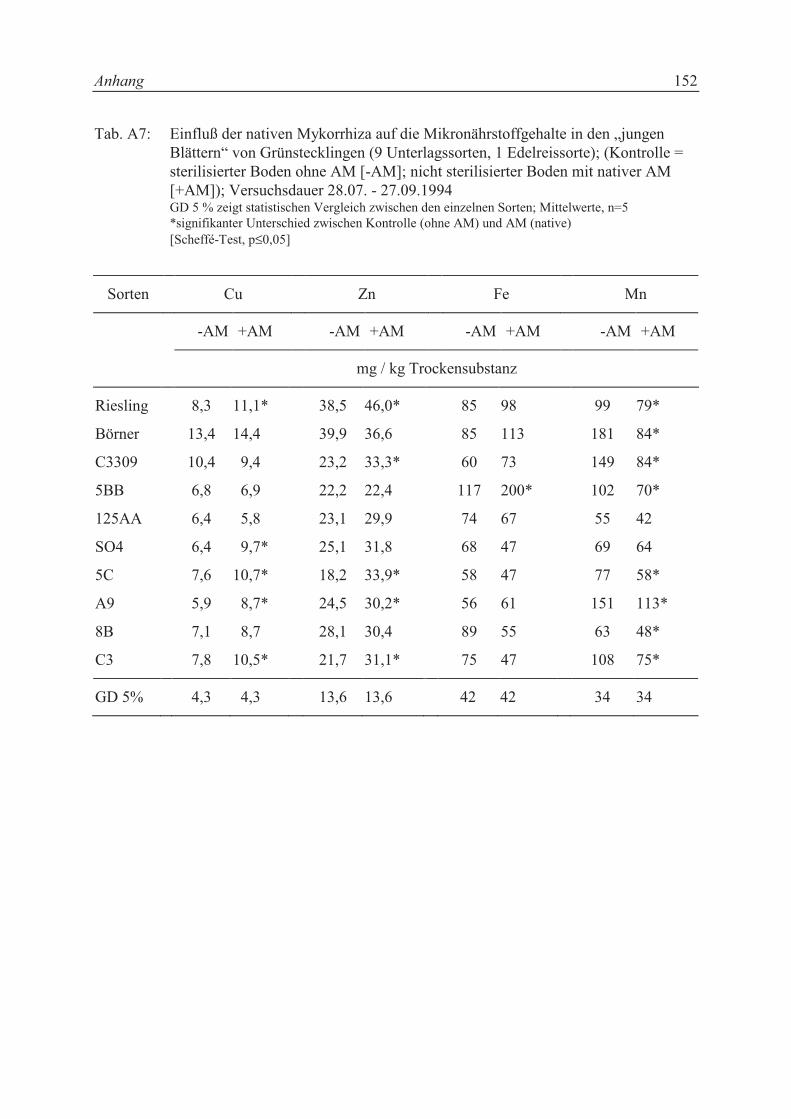
Anhang 152
Tab. A7: Einfluß der nativen Mykorrhiza auf die Mikronährstoffgehalte in den „jungen Blättern“ von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte); (Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Cu Zn Fe Mn
-AM +AM -AM +AM -AM +AM -AM +AM
mg / kg Trockensubstanz
Riesling 8,3 11,1* 38,5 46,0* 85 98 99 79*
Börner 13,4 14,4 39,9 36,6 85 113 181 84*
C3309 10,4 9,4 23,2 33,3* 60 73 149 84*
5BB 6,8 6,9 22,2 22,4 117 200* 102 70*
125AA 6,4 5,8 23,1 29,9 74 67 55 42
SO4 6,4 9,7* 25,1 31,8 68 47 69 64
5C 7,6 10,7* 18,2 33,9* 58 47 77 58*
A9 5,9 8,7* 24,5 30,2* 56 61 151 113*
8B 7,1 8,7 28,1 30,4 89 55 63 48*
C3 7,8 10,5* 21,7 31,1* 75 47 108 75*
GD 5% 4,3 4,3 13,6 13,6 42 42 34 34
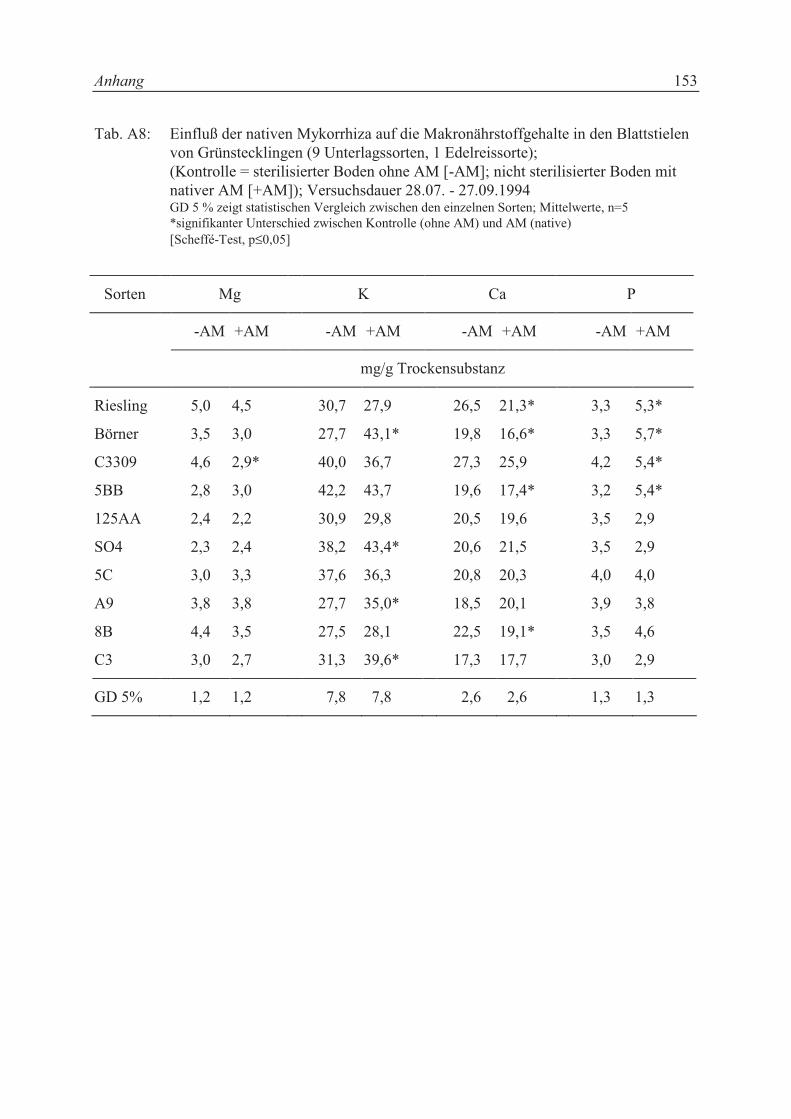
Anhang 153
Tab. A8: Einfluß der nativen Mykorrhiza auf die Makronährstoffgehalte in den Blattstielen von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte);
(Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Mg K Ca P
-AM +AM -AM +AM -AM +AM -AM +AM
mg/g Trockensubstanz
Riesling 5,0 4,5 30,7 27,9 26,5 21,3* 3,3 5,3*
Börner 3,5 3,0 27,7 43,1* 19,8 16,6* 3,3 5,7*
C3309 4,6 2,9* 40,0 36,7 27,3 25,9 4,2 5,4*
5BB 2,8 3,0 42,2 43,7 19,6 17,4* 3,2 5,4*
125AA 2,4 2,2 30,9 29,8 20,5 19,6 3,5 2,9
SO4 2,3 2,4 38,2 43,4* 20,6 21,5 3,5 2,9
5C 3,0 3,3 37,6 36,3 20,8 20,3 4,0 4,0
A9 3,8 3,8 27,7 35,0* 18,5 20,1 3,9 3,8
8B 4,4 3,5 27,5 28,1 22,5 19,1* 3,5 4,6
C3 3,0 2,7 31,3 39,6* 17,3 17,7 3,0 2,9
GD 5% 1,2 1,2 7,8 7,8 2,6 2,6 1,3 1,3
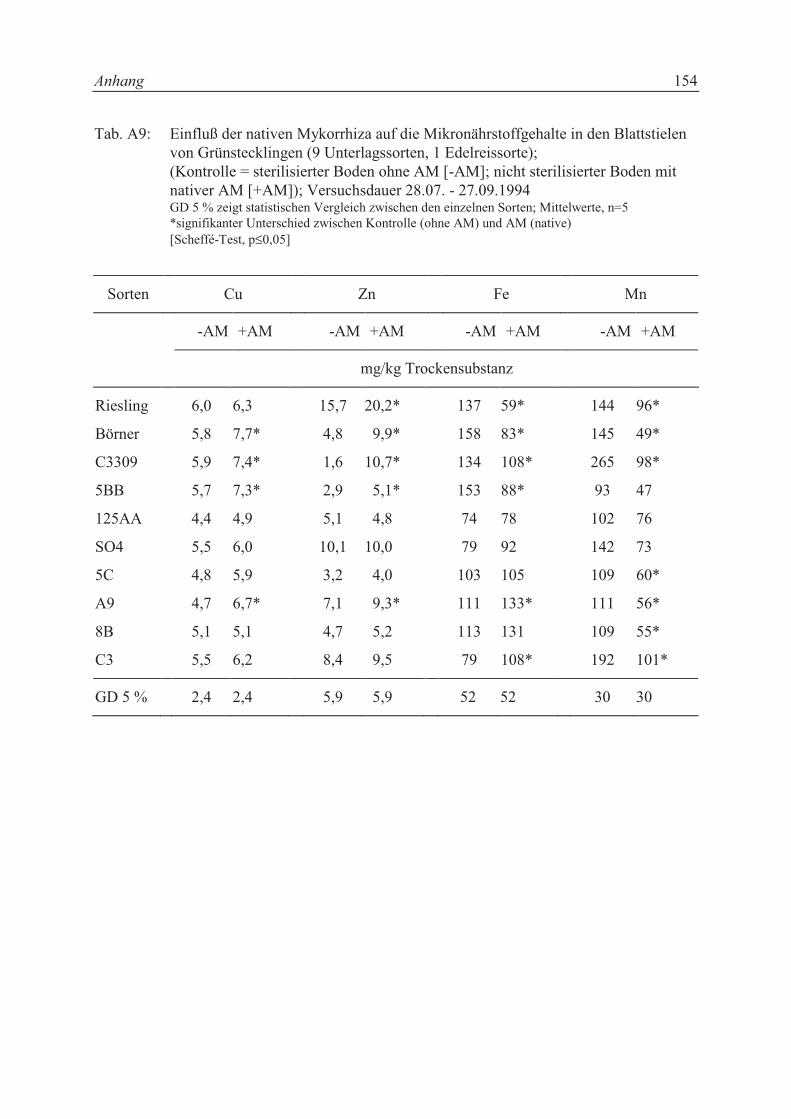
Anhang 154
Tab. A9: Einfluß der nativen Mykorrhiza auf die Mikronährstoffgehalte in den Blattstielen von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte);
(Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Cu Zn Fe Mn
-AM +AM -AM +AM -AM +AM -AM +AM
mg/kg Trockensubstanz
Riesling 6,0 6,3 15,7 20,2* 137 59* 144 96*
Börner 5,8 7,7* 4,8 9,9* 158 83* 145 49*
C3309 5,9 7,4* 1,6 10,7* 134 108* 265 98*
5BB 5,7 7,3* 2,9 5,1* 153 88* 93 47
125AA 4,4 4,9 5,1 4,8 74 78 102 76
SO4 5,5 6,0 10,1 10,0 79 92 142 73
5C 4,8 5,9 3,2 4,0 103 105 109 60*
A9 4,7 6,7* 7,1 9,3* 111 133* 111 56*
8B 5,1 5,1 4,7 5,2 113 131 109 55*
C3 5,5 6,2 8,4 9,5 79 108* 192 101*
GD 5 % 2,4 2,4 5,9 5,9 52 52 30 30
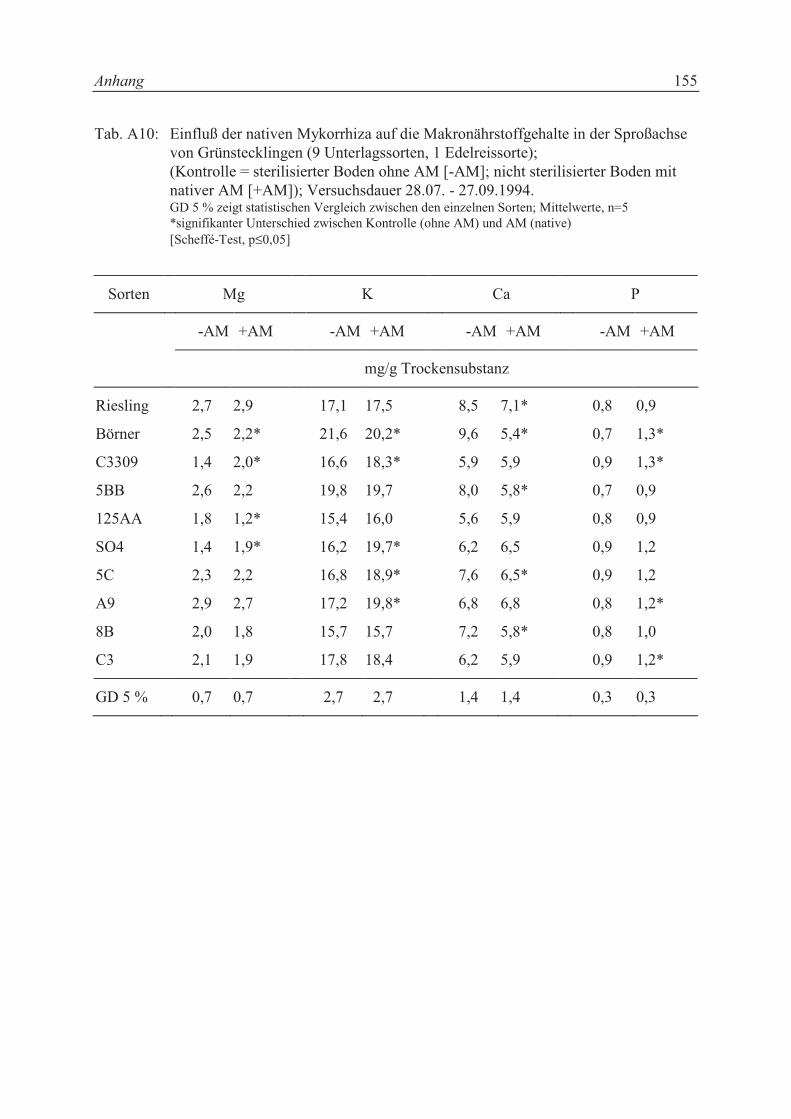
Anhang 155
Tab. A10: Einfluß der nativen Mykorrhiza auf die Makronährstoffgehalte in der Sproßachse von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte);
(Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994.
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Mg K Ca P
-AM +AM -AM +AM -AM +AM -AM +AM
mg/g Trockensubstanz
Riesling 2,7 2,9 17,1 17,5 8,5 7,1* 0,8 0,9
Börner 2,5 2,2* 21,6 20,2* 9,6 5,4* 0,7 1,3*
C3309 1,4 2,0* 16,6 18,3* 5,9 5,9 0,9 1,3*
5BB 2,6 2,2 19,8 19,7 8,0 5,8* 0,7 0,9
125AA 1,8 1,2* 15,4 16,0 5,6 5,9 0,8 0,9
SO4 1,4 1,9* 16,2 19,7* 6,2 6,5 0,9 1,2
5C 2,3 2,2 16,8 18,9* 7,6 6,5* 0,9 1,2
A9 2,9 2,7 17,2 19,8* 6,8 6,8 0,8 1,2*
8B 2,0 1,8 15,7 15,7 7,2 5,8* 0,8 1,0
C3 2,1 1,9 17,8 18,4 6,2 5,9 0,9 1,2*
GD 5 % 0,7 0,7 2,7 2,7 1,4 1,4 0,3 0,3
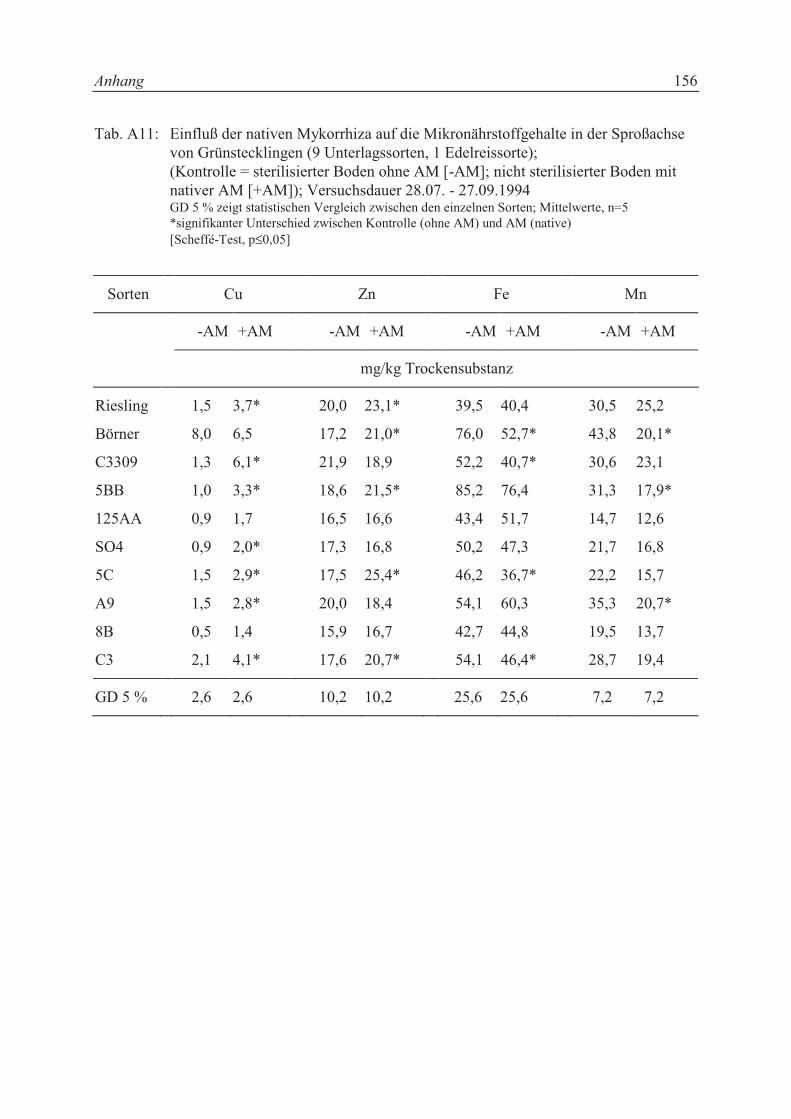
Anhang 156
Tab. A11: Einfluß der nativen Mykorrhiza auf die Mikronährstoffgehalte in der Sproßachse von Grünstecklingen (9 Unterlagssorten, 1 Edelreissorte);
(Kontrolle = sterilisierter Boden ohne AM [-AM]; nicht sterilisierter Boden mit nativer AM [+AM]); Versuchsdauer 28.07. - 27.09.1994
GD 5 % zeigt statistischen Vergleich zwischen den einzelnen Sorten; Mittelwerte, n=5 *signifikanter Unterschied zwischen Kontrolle (ohne AM) und AM (native) [Scheffé-Test, p≤0,05]
Sorten Cu Zn Fe Mn
-AM +AM -AM +AM -AM +AM -AM +AM
mg/kg Trockensubstanz
Riesling 1,5 3,7* 20,0 23,1* 39,5 40,4 30,5 25,2
Börner 8,0 6,5 17,2 21,0* 76,0 52,7* 43,8 20,1*
C3309 1,3 6,1* 21,9 18,9 52,2 40,7* 30,6 23,1
5BB 1,0 3,3* 18,6 21,5* 85,2 76,4 31,3 17,9*
125AA 0,9 1,7 16,5 16,6 43,4 51,7 14,7 12,6
SO4 0,9 2,0* 17,3 16,8 50,2 47,3 21,7 16,8
5C 1,5 2,9* 17,5 25,4* 46,2 36,7* 22,2 15,7
A9 1,5 2,8* 20,0 18,4 54,1 60,3 35,3 20,7*
8B 0,5 1,4 15,9 16,7 42,7 44,8 19,5 13,7
C3 2,1 4,1* 17,6 20,7* 54,1 46,4* 28,7 19,4
GD 5 % 2,6 2,6 10,2 10,2 25,6 25,6 7,2 7,2

Literaturverzeichnis 157
E LITERATURVERZEICHNIS
Abbott, L.K. and Robson, A.D. (1984): The effect of VA mycorrhizae on plant growth. In: VA mycorrhiza, Powell, C.L. and Bagyaraj, D.J., Eds., CRC Press, Boca Raton, Florida, 113-130.
Amijee, F., Stribley, D.P. and Tinker, P.B. (1986): The development of endomycorrhizal root systems. 6. The relationship between development of infection and intensity of infecti-on in young leek roots. New Phytol. 102, 293-301.
Anonym (1983): Luftqualitätskriterien für photochemische Oxidantien. (Umweltbundesamt, Hrsg.). Berichte 5/83, Erich Schmidt Verlag Berlin.
Anonym (1986): Rebenpflanzgutverordnung vom 21.01.1986: Bundesgesetzblatt, Jahrgang 1986, Teil 1.
Anonym (1997 a): Richtlinien für den kontrolliert umweltschonenden Weinbau in Rheinland- Pfalz (Stand 01.01.1997). Ministerium für Wirtschaft, Verkehr, Landwirtschaft und Weinbau, Rheinland-Pfalz.
Anonym (1997 b): Bioland-Richtlinien für Pflanzenbau, Tierhaltung und Verarbeitung (Fas-sung vom 19. April 1997). BIOLAND Verband für organisch-biologischen Land- bau e.V., Göppingen.
Arines, J., Porto, M.E. and Vilarino, A. (1992): Effect of manganese on vesicular-arbuscular mycorrhizal development in red clover plants and on soil Mn-oxidizing bacteria. Mycorrhiza 1, 127-131.
Azcón, R. and El-Atrash, F. (1997): Influence of arbuscular mycorrhizae and phosphorus fer-ti- lization on growth, nodulation and N2 fixation (15N) in Medicago sativa at four salinity levels. Biol. Fertil. Soils 24, 81-86.
Azcón-Aguilar, C. and Barea, J.M. (1997): Applying mycorrhiza biotechnology to horticultu- re: Significance and potentials. Scientia Horticulturae 68, 1-24.
Backhaus, G.F. und Feldmann, F. (1996): Mykorrhiza in gärtnerischen Substraten - endlich einsatzreif? Taspo Gartenbaumagazin, 12-14.
Baltruschat, H. (1987 a): Evaluation of the suitability of expanded clay as carrier material for VAM spores in field inoculation of maize. Angew. Bot. 61, 163-169.

Literaturverzeichnis 158
Baltruschat, H. (1987 b): Field inoculation of maize with vesicular-arbuscular mycorrhizal fungi by using expanded clay as carrier material for mycorrhiza. Journal of Plant Diseases and Protection 94, 419-430.
Baltruschat, H. (1987 c): Zur möglichen Nutzung der VAM in der Pflanzenproduktion. Gesunde Pflanzen 39, 510-518.
Baltruschat, H. and Dehne, H.W. (1988): The occurence of vesicular-arbuscular mycorrhiza in agro-ecosystems. I. Influence of nitrogen fertilization and green manure in continuous monoculture and in crop rotation on the inoculum potential of winter wheat. Plant and Soil 107, 279-284.
Bavaresco, L. and Fogher, C. (1992): Effect of root infection with Pseudomonas fluorescens
and Glomus mosseae in improving Fe-efficiency of grapevine ungrafted rootstocks. Vitis 31, 163-168.
Bavaresco, L. and Fogher, C. (1996): Lime-induced chlorosis of grapevine as affected by rootstock and root infection with arbuscular mycorrhiza and Pseudomonas fluorescens. Vitis 35, 119-123.
Bécard, G. and Piché, Y. (1990): Physiological factors determining vesicular-arbuscular my-corrhizal formation in host and non-host Ri T-DNA transformed roots. Can. J. Bot. 68, 1260-1264.
Beran, N. (1994 a): Untersuchungen über die Auswirkungen des Luftschadstoffes Ozon auf die Rebe (Vitis vinifera L.). I. Charakteristische Schädigungssymptome an Blättern. Vitic. Enol. Sci. 49, 55-61.
Beran, N. (1994 b): Untersuchungen über die Auswirkungen des Luftschadstoffes Ozon auf die Rebe (Vitis vinifera L.). II. Der Einfluß der Konzentration und ihrer Einwirkungs- dauer auf das Ausmaß der sichtbaren, irreversiblen Schädigungen in Abhängigkeit vom Alter der Blätter sowie von der Sorte. Vitic. Enol. Sci. 49, 96-109.
Bergmann, W. (1986): Ernährungsstörungen bei Kulturpflanzen. VEB Gustav Fischer Verlag Jena, 2. erweiterte Auflage, S. 31.
Berta, G., Fusconi, A., Trotta, A. and Scannerini, S. (1990): Morphogenetic modifications induced by the mycorrhizal fungus Glomus strain E3 in the root system of Allium por-
rum L. New Phytol. 114, 207-215.
Biermann, B.J. and Linderman, R.G. (1983 a): Effect of container plant growth medium and fertilizer phosphorus on establishment and host growth response to vesicular-arbuscular mycorrhizae. J. Amer. Soc. Hort. Sci. 108, 962-971.

Literaturverzeichnis 159
Biermann, B.J. and Linderman, R.G. (1983 b): Increased geranium growth using pretransplant inoculation with a mycorrhizal fungus. J. Amer. Soc. Hort. Sci. 108, 972-976. Biricolti, S., Ferrini, F., Rinaldelli, E., Tamantini, I. and Vignozzi, N. (1997): VAM fungi and soil lime content influence rootstock growth and nutrient content. Am. J. Enol. Vitic. 48, 93-99.
Bleyer, G. und Fox, R. (1995): Auswirkungen von unterschiedlichen Rigoltiefen und Bra- chenutzungen auf die Populationsdichte des Nematoden Xiphinema index. Deutsches Weinbau-Jahrbuch 46, 101-107.
Blum, U. and Tingey, D.T. (1977): A study of the potential ways in which ozone could reduce root growth and nodulation of soybean. Atmospheric Environment 11, 737-739.
Böhm, W. (1979): Methods of studying root systems. Springer Verlag Berlin, Heidelberg New York.
Bonfante-Fasolo, P. (1978): Some ultrastructural features of the vesicular-arbuscular my- corrhiza in the grapevine. Vitis 17, 386-395.
Bonfante-Fasolo, P. (1984): Anatomy and morphology of VA mycorrhizae. In: VA mycorrhi- za, Powell, C.L. and Bagyaraj, D.J., Eds., CRC Press, Boca Raton, Florida., 5-33.
Brendel, G., Büscher, E. und Steinberg, B. (1990): Untersuchungen über das Vorkommen der vesikulär-arbuskulären Mykorrhiza in Weinbergsböden des Rheingaus. Vitic. Enol. Sci. 45, 97-100.
Brewer, P.F. and Heagle, A.S. (1983): Interactions between Glomus geosporum and exposure of soybeans to ozone or simulated acid rain in the field. Phytopathology 73, 1035-1040.
Buwalda, J.G., Stribley, D.P. and Tinker, P.B. (1984): The development of endomycorrhizal root systems. 5. The detailed pattern of development of infection and the control of infection level by host in young leek plants. New Phytol. 96, 411-427.
Calvet, C., Pinochet, J., Camprubí, A. and Fernández, C. (1995): Increased tolerance to the root-lesion nematode Pratylenchus vulnus in mycorrhizal micropropagated BA-29 quince rootstock. Mycorrhiza 5, 253-258.
Carling, D.E., Roncadori, R.W. and Hussey, R.S. (1996): Interactions of arbuscular mycorrhi- zae, Meloidogyne arenaria and phosphorus fertilization on peanut. Mycorrhiza 6, 9-13.

Literaturverzeichnis 160
Cordier, C., Gianinazzi, S. and Gianinazzi-Pearson, V. (1996): Colonisation patterns of root tissues by Phytophtora nicotianae var. parasitica related to reduced disease in my- corrhizal tomato. Plant and Soil 185, 223-232.
Currle, O., Bauer, O., Hofäcker, W., Schumann, F. und Frisch, W. (1983): Biologie der Rebe. Verlag Meininger, Neustadt, 1. Auflage.
Deal, D.R., Boothroyd, C.W. and Mai, W.F. (1972): Replanting of vineyards and its rela-tionship to vesicular-arbuscular mycorrhiza. Phytopathology 62, 172-175.
Dehne, H.-W. and Backhaus, G.F. (1986): The use of VA-mycorrhizal fungi in plant producti- on. 1. Inoculum production. Journal of Plant Diseases and Protection 93, 415-424.
Dehne, H.-W. (1994): Interaktionen bei der Regulation von Schaderregerpopulationen - Mykorrhiza und Pflanzengesundheit. Berichte über Landwirtschaft. Sonderheft 209, 93-101.
Diaz, G., Azcón-Aguilar, C. and Honrubia, M. (1996): Influence of arbuscular mycorrhiza on heavy metal (Zn and Pb) uptake and growth of Lygeum spartum and Anthyllis cyti-
soides. Plant and Soil 180, 241-249.
Dixon, R.K., Garrett, H.E. and Cox, G.S. (1988): Cytokinins in the root pressure exudate of Citrus jambhiri Lush. colonized by vesicular-arbuscular mycorrhizae.
Tree Physiol. 4, 9-18.
Douds, D.D. and Schenk, N.C. (1990): Relationship of colonization and sporulation by VA mycorrhizal fungi to plant nutrition and carbohydrate contents. New Phytol. 116, 621-627.
Douds, D.D., Janke, R.R. and Peters, S.E. (1993): VAM fungus spore populations and colo- nization of roots of maize and soybean under conventional and low-input sustainable agriculture. Agric. Ecosyst. Environ. 43, 325-335.
Douds, D.D., Galvez, L., Janke, R.R. and Wagoner, P. (1995): Effect of tillage and farming system upon populations and distribution of vesicular-arbuscular mycorrhizal fungi. Agric. Ecosyst. Environ. 52, 111-118.
Drüge, U. (1992): Zur Wachstumsförderung von Lein (Linum usitatissimum L.) durch VA Mykorrhiza unter besonderer Berücksichtigung der Cytokinine. Dissertation Universität Hannover.
Dütsch, H., Rückert, F.-E. und Steinberg, B. (1997): Die Eignung verschiedener Winterteil- zeitbegrünungen im umweltschonenden Weinbau.

Literaturverzeichnis 161
Mitt. Klosterneuburg 47, 61-73.
Edriss, M.H., Davis, R.M. and Burger, D.W. (1984): Effect of vesicular-arbuscular mycorrhi-za infection on transpiration, photosynthesis and growth of flax (Linum usitatissimum L.) in relation to cytokinin levels. J. Plant Physiol. 141, 40-48.
Eibach, H. (1982): Die Vesikulare-Arbuskulare Mykorrhiza der Rebe. Dissertation Universität Hohenheim.
El-Atrach, F., Vierheilig, H. and Ocampo, J.A. (1989): Influence of non-host plants on vesicu- lar-arbuscular mycorrhizal infection of host plants and on spore germination. Soil Biol. Biochem. 21, 161-163.
Faber, B.A., Zasoski, R.J., Burau, R.G. and Uriu, K. (1990): Zinc uptake by corn as affec-ted by vesicular-arbuscular mycorrhizae. Plant and Soil 129, 121-130.
Fardossi, A., Mayer, G. und Barna, J. (1988): Untersuchungen über die Zusammenhänge zwi- schen Mn-Aufnahme und Chlorose bei Vitis vinifera L. Mitt. Klosterneuburg 38, 39-47.
Fardossi, A., Hepp, E., Mayer, C. und Kalchgruber, R. (1994): Über die Nährstoffeffizienz von Unterlagsrebsorten. Mitt. Klosterneuburg 44, 117-126.
Feldmann, F. (1990): Die Mykorrhiza des Kautschukbaumes Hevea spec.: Vorkommen am Naturstandort und in Plantagen, Wirkung auf das Resistenzverhalten und Nutzung im Plantagenbau. Dissertation Technische Universität Braunschweig.
Feldmann, F. und Lieberei, R. (1992): Kulturmaßnahmen im Pflanzenbau des Kautschukbau- mes (Hevea spec.) und ihr Einfluß auf das Vorkommen von Mykorrhiza in Plantagen des Amazonasgebietes. Angew. Bot. 4, 67-87.
Feldmann, F. (1997): Charakterisierung des Vorkommens von arbuskulären Mykorrhizasym- biosen in Pflanzennutzungssystemen des Braunschweiger Raumes. Braunschw. naturkdl. Schr. 5, 491-503.
Ferguson, J.J. and Menge, J.A. (1986): Response of citrus seedlings to various field inoculati- on methods with Glomus deserticola in fumigated nursery soils. J. Amer. Soc. Hort. Sci. 111, 288-292.
Frank, B. (1885): Ueber die auf Wurzelsymbiose beruhende Ernährung gewisser Bäume durch unterirdische Pilze. Ber. d. dtsch. bot. Ges. 3, 128-145.

Literaturverzeichnis 162
Frank, B. (1887): Ueber neue Mycorhiza-Formen. Ber. d. dtsch. bot. Ges. 5, 395-409.
Gebbing, H., Schwab, A. und Alleweldt, G. (1977): Mykorrhiza bei Reben. Vitis 16, 279-285.
Geisler, G. (1957): Die Bedeutung des Wurzelsystems für die Züchtung dürreresistenter Re- benunterlagssorten. Vitis 1, 14-31. George, E., Römheld, V. and Marschner, H. (1994): Contribution of mycorrhizal fungi to micronutrient uptake by plants. In: Biochemistry of Metal Micronutrients in the Rhi- zosphere, Manthey, J.A., Crowley, D.E. and Luster, D.G., Eds., CRC Press Inc., Boca Raton, FL, 93-109.
Gericke, S. und Kurmies, B. (1952): Die kolorimetrische Phosphorbestimmung mit Ammoni- um-vanadat-molybdat und ihre Anwendung in der Pflanzenanalyse. Z. Pflanzenern. Düng. Bodenk. 59, 235-247.
Giovanetti, M. and Mosse, B. (1980): An evaluation for measuring vesicular arbuscular my-corrhizal infection. New Phytol. 84, 489-500.
Giovannetti, M., Schubert, A., Cravero, M.C. and Salutini, L. (1988): Spore production by the vesicular-arbuscular mycorrhizal fungus Glomus monosporum as related to host spe- cies, root colonization and plant growth enhancement. Biol. Fertil. Soils 6, 120-124.
Glenn, M.G., Chew, F.S. and Williams, P.H. (1988): Influence of glucosinolate content of Brassica (Cruciferae) roots on growth of vesicular-arbuscular mycorrhizal fungi.
New Phytol. 110, 217-225.
Gnekow, M.A. (1988): Die Rolle von VA-Mykorrhiza bei der Phosphaternährung von Kul- turpflanzen in Abhängigkeit von Phosphatdüngung und Bewirtschaftungsweise. Dissertation Universität Hohenheim.
Gnekow, M.A. and Marschner, H. (1989 a): Influence of the fungicide pentachloronitrobenze- ne on VA-mycorrhizal and total root length and phosphorus uptake of oats (Avena sativa). Plant and Soil 114, 91-98.
Gnekow, M.A. and Marschner, H. (1989 b): Role of VA-mycorrhiza in growth and mineral nutrition of apple (Malus pumila var. domestica) rootstock cuttings. Plant and Soil 119, 285-293.
Grunewaldt-Stöcker, G. and Dehne, H.W. (1989): The use of vesicular-arbuscular mycorrhizal fungi in plant production. II. Characterization of inocula on organic carrier material. Journal of Plant Diseases and Protection 96, 615-626.

Literaturverzeichnis 163
Hall, I.R. (1984): Taxonomy of VA mycorrhizal fungi. In: VA mycorrhiza. Powell, C.L. and Bagyaraj, D.J., Eds., Boca Raton, Florida. CRC Press, 57-94.
Hamel, C. (1996): Prospects and problems pertaining to the management of arbuscular my- corrhizae in agriculture. Agric. Ecosyst. Environ. 60, 197-210.
Harinikumar, K.M. and Bagyaraj, D.J. (1988): Effect of crop rotation on native vesicular ar- buscular mycorrhizal propagules in soil. Plant and Soil 110, 77-80.
Harley, J.L. and Harley, E.L. (1987): A check-list of mycorrhiza in the British flora. New Phytol. 105, 1-102.
Hasler, T., Frey, B. und Schüepp, H. (1990): Ozonschäden an Reben. Landwirtschaft Schweiz 3, 383-387.
Hayman, D.S., Barea, J.-M. and Azcón, R. (1976): Vesicular-arbuscular mycorrhiza in Sou-thern Spain: its distribution in crops growing in soil of different fertility. Phytopath. medit. 15, 1-6.
Herrmann, J.V. (1991): Schädigungen an Reben in Franken durch Ozon. Rebe und Wein 12, 389-391.
Hilton, R.J. and Khatamian, H. (1973): Diurnal variation in elongation rates of roots of woody plants. Can. J. Plant Sci. 53, 699-700.
Hock, B. und Elstner, F. (1988): Schadwirkungen auf Pflanzen. Lehrbuch der Pflanzentoxiko- logie. BI-Wissenschaftsverlag Mannheim, Wien, Zürich.
Hofmann, U. (1994): Begrünung im ökologischen Weinbau. Das deutsche Weinmagazin 14, 13-18.
Howeler, R.H. and Sieverding, E. (1983): Potentials and limitation of mycorrhizal inoculation illustrated by experiments with field-grown cassava. Plant and Soil 75, 245-261.
Husse, B. und Steinberg, B. (1983): Wurzel- und Sproßwachstum von Begrünungspflanzen. Der deutsche Weinbau 36, 1849-1854.
Ikram, A., Mahmud, A.W., Ghani, M.N., Ibrahim, M.T. and Zainal, A.B. (1992): Field nurse-ry inoculation of Hevea brasiliensis Muell. Arg. seedling rootstock with vesicular- ar-buscular mycorhizal (VAM) fungi. Plant and Soil 145, 231-236.
Jasper, D.A., Abbott, L.K. and Robson, A.D. (1989): Soil disturbance reduces the infec-tivity of external hyphae of vesicular-arbuscular mycorrhizal fungi.

Literaturverzeichnis 164
New Phytol. 112, 93-99.
Jasper, D.A., Abbott, L.K. and Robson, A.D. (1991): The effects of disturbance on vesicular- arbuscular mycorrhizal fungi in soils from different vegetation types. New Phytol. 118, 471-476.
Johnson, C.R., Joiner, J.N. and Crews, C.E. (1980): Effects of N, K and Mg on growth and leaf nutrient composition of three container grown woody ornamentals inoculated with mycorrhiza. J. Amer. Soc. Hort. Sci. 105, 286-288.
Karagiannidis, N., Nikolaou, N. und Mattheou, A. (1995): Wirkung dreier VA- Mykorrhiza-pilze auf Ertrag und Nährstoffaufnahme von drei Unterlagen und einer Tafeltraubensorte. Vitis 34, 85-89.
Karagiannidis, N., Velemis, D. and Stavropoulos, N. (1997): Root colonization and spore po- pulation by VA-mycorrhizal fungi in four grapevine rootstocks. Vitis 36, 57-60.
Kiefer, W. und Weber, M. (1986): Einfluß unterschiedlicher Standweiten auf die Ausbreitung der Rebwurzeln. Die Weinwissenschaft 41, 404-412.
Koch, M., Tanami, Z., Bodani, H., Wininger, S. and Kapulnik, Y. (1997): Field application of vesicular-arbuscular mycorrhizal fungi improved garlic yield in disinfected soil. Mycorrhiza 7, 47-50.
Koide, R.T. (1991): Nutrient supply, nutrient demand and plant response to mycorrhizal in- fection. New Phytol 117, 365-386.
Koske, R.E. and Gemma, J.N. (1989): A modified procedure for staining roots to detect VA- mycorrhizas. Mycol. Res. 92, 486-488.
Kothari, S.K., Marschner, H. and George, E. (1990 a): Effect of VA mycorrhiza fungi and rhizosphere microorganisms on root and shoot morphology, growth and water relations in maize. New Phytol. 116, 303-311.
Kothari, S.K., Marschner, H. and Römheld, V. (1990 b): Direct and indirect effects of VA mycorrhizal fungi and rhizosphere microorganisms on aquisition of mineral nutrients by maize (Zea mays L.) in a calcareous soil. New Phytol. 116, 637-645.

Literaturverzeichnis 165
Kothari, S.K., Marschner, H. and Römheld, V. (1991 a): Contribution of the VA mycorrhizal hyphae in acquisition of phosphorus and zinc by maize grown in a calcareous soil. Plant and Soil 131, 177-185.
Kothari, S.K., Marschner, H. and Römheld, V. (1991 b): Effects of a vesicular-arbuscular my- corrhizal fungus and rhizosphere micro-organisms on manganese reduction in the rhi- zosphere and manganese concentrations in maize (Zea mays L.). New Phytol. 117, 649-655.
Krikun, J., Haas, J.H. and Bar-Yosef, B. (1987): Use of VA mycorrhizal-fungus inoculum in soils in arid and semi-arid climates: A field study with bell pepper and transplants. Angew. Bot. 61, 97-105.
Kruckelmann, H.W. (1975): Effects of fertilizers, soils, soil tillage and plant species on the frequency of Endogene chlamydospores and mycorrhizal infection in arable soils. In: Endomycorrhizas, Sanders, F.E., Mosse, B. and Tinker, P.B., Eds., Academic Press, London, pp. 511-525.
Kucey, R.M.N. and Janzen, H.H. (1987): Effects of VAM and reduced nutrient availability on growth and phosphorus and micronutrient uptake of wheat and field beans under greenhouse conditions. Plant and Soil 104, 71-78.
Kurle, J.E. and Pfleger, F.L. (1994): Arbuscular mycorrhizal fungus spore populations respond to conversions between low-input and conventional management practices in a corn- soybean rotation. Agron. J. 86, 467-475.
Lamar, R.T. and Davey, C.B. (1988): Comparative effectivity of three Fraxinus pennsylvanica
Marsh. vesicular-arbuscular mycorrhizal fungi in a high-phosphorus nursery soil. New Phytol. 109, 171-181.
Land, S. (1990): Auftreten und Charakterisierung der vesikulär-arbuskulären (VA) Mykorrhi-za in intensiv genutzten Ackerböden. Dissertation Universität Hannover.
Li, X.-L., George, E. and Marschner, H. (1991 a): Phosphorus depletion and pH decrease at the root-soil and hyphae-soil interfaces of VA mycorrhizal white clover fertilized with ammonium. New Phytol. 119, 397-404.
Li, X.-L., George, E. and Marschner, H. (1991 b): Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil. Plant and Soil 136, 41-48.
Li, X.-L., Marschner, H. and George, E. (1991 c): Aquisition of phosphorus and copper by VA-mycorrhizal hyphae and root-to-shoot transport in white clover. Plant and Soil 136, 49-57.

Literaturverzeichnis 166
Mahoney, M.J., Chevone, B.I., Skelly, J.M. and Moore, L.D. (1985): Influence of mycorrhizae on growth of loblolly pine seedlings exposed to ozone and sulfur dioxide. Phytopathology 75, 679-682.
Marschner, H. (1988): Mechanisms of manganese acquisition by roots from soils. In: Manga- nese in soils and plants, Graham, R.D., Hannam, R.J. and Uren, N.C., Eds., Kluwer academic publishers, London, 191-204.
Marschner, H. (1993): Zinc uptake from soils. In: Zinc in soils and plants. Developments in plant and soil science 55, 59-77.
Marschner, H. and Dell, B. (1994): Nutrient uptake in mycorrhizal symbiosis. Plant and Soil 159, 89-102.
Marschner, H. (1995): Mineral Nutrition of Higher Plants. 2nd ed., Academic Press, London.
Matsubara, Y-I., Karikomi, T., Ikuta, M., Hori, H., Ishikawa, S. and Harada, T. (1996): Effect of arbuscular mycorrhizal fungus inoculation on growth of apple (Malus ssp.) seedlings. J. Japan. Soc. Hort. Sci. 65, 297-302.
Mazzitelli, M and Schubert, A. (1989): Effect of several VAM endophytes and artificial sub-strates on in vitro propagated Vitis berlandieri x rupestris „1103P“. Agric. Ecosyst. Environ. 29, 289-293.
McCool, P.M., Menge, J.A. and Taylor, O.C. (1979): Effects of ozone and HCl gas on the development of the mycorrhizal fungus Glomus fasciculatus and growth of „Troyer“ citrange. J. Amer. Soc. Hort. Sci. 104, 151-154.
McCool, P.M. and Menge, J.A. (1983): Influence of ozone on carbon partitioning in tomato: Potential role of carbon flow in regulation of the mycorrhizal symbiosis under conditi- ons of stress. New Phytol. 94, 241-247.
McCool, P.M. and Menge, J.A. (1984): Interaction of ozone and mycorrhizal fungi on tomato as influenced by fungal species and host variety. Soil Biol. Biochem. 16, 425-427.
Menge, J.A. (1983): Utilization of vesicular-arbuscular mycorrhizal fungi in agriculture. Can. J. Bot. 61, 1015-1024.
Menge, J.A., Raski, D.J., Lider, L.A., Johnson, E.L.V., Jones, N.O., Kissler, J.J. and Hem- street, C.L. (1983): Interactions between mycorrhizal fungi, soil fumigation and growth of grapes in california. Am. J. Enol. Vitic. 34, 117-122.

Literaturverzeichnis 167
Mengel, K. (1991): Ernährung und Stoffwechsel der Pflanze. Gustav Fischer Verlag, Jena, 7., überarbeitete Auflage.
Merkt, N. (1989): Untersuchungen zur Affinität von Unterlage und Reis bei Reben. Dissertation Universität Hohenheim.
Miller, D.D., Domoto, P.A. and Walker, C. (1985): Mycorrhizal fungi at eighteen apple root- stock plantings in the United states. New Phytol. 100, 379-391.
Mohr, H.D (1988): Untersuchungen zum Wachstum von Rebwurzeln in Wurzelbeobachtungs- kästen. Deutsches Weinbau-Jahrbuch 40, 87-96.
Mohr, H.D. (1993): Mykorrhiza bei Reben: Eine Lebensgemeinschaft mit Perspektiven. Deutsches Weinbau-Jahrbuch 44, 133-148.
Mohr, H.D. (1994): Verteilung und Mykorrhizierung der Wurzeln von Reben und Be-grünungspflanzen im Boden. Bundesministerium für Land- und Forstwirtschaft Wien. Internationaler Arbeitskreis Begrünung im Weinbau: Kolloquium. Sonderausgabe der Zeitschrift Förderungsdienst, 60-68.
Mohr, H.D. (1996): Periodicity of root tip growth of vines in the Moselle valley. Vitic. Enol. Sci. 51, 83-90.
Mohr, H.D. (1997): Wachstum und Mykorrhizierung von Rebwurzeln in Weinbergen des Mo- seltals. In: Boden und Landschaft, Schriftenreihe zur Bodenkunde, Landeskultur und Landschaftsökologie Band 17, Justus-Liebig-Universität Gießen, 63-76.
Monzon, A. and Azcón, R. (1996): Relevance of mycorrhizal fungal origin and host plant ge- notype to inducing growth and nutrient uptake in Medicago species. Agric. Ecosyst. Environ. 60, 9-15.
Morin, F., Fortin, J.A., Hamel, C., Granger, R.L. and Smith, D.L. (1994): Apple rootstock response to vesicular-arbuscular mycorrhizal fungi in a high phosphorus soil. J. Amer. Soc. Hort. Sci. 119, 578-583.
Morlat, R. and Jacquet, A. (1993): The soil effects on the grapevine root system in several vineyards of the Loire valley (France). Vitis 32, 35-42.
Müller-Thurgau (1903): Verhalten der Wurzeln bei zunehmendem Alter der Rebstöcke. Mitteilungen über Weinbau und Kellerwirtschaft 15, 33-38.
Musselman, R.C., Kender, W.J. and Crowe, D.E. (1978): Determining air pollutant effects on the growth and productivity of Concord grapevines using open-top chambers. J. Amer. Soc. Hort. Sci. 103, 645-648.

Literaturverzeichnis 168
Musselman, R.C. and Melious, R.E. (1984): Sensitivity of grape cultivars to ambient ozone. HortScience 19, 657-659.
Musselman, R.C., Forsline, P.L. and Kender, W.J. (1985): Effects of sulfur dioxide and am- bient ozone on concord grapevine growth and productivity. J. Amer. Soc. Hort. Sci. 110, 882-888.
Nadian, H., Smith, S.E., Alston, A.M. and Murray, R.S. (1996): The effect of soil compaction on growth and P uptake by Trifolium subterraneum: interactions with mycorrhizal co- lonisation. Plant and Soil 182, 39-46.
Nadian, H., Smith, S.E., Alston, A.M. and Murray, R.S. (1997): Effects of soil compaction on plant growth, phosphorus uptake and morphological characteristics of vesicular- ar-buscular mycorrhizal colonization of Trifolium subterraneum. New Phytol. 135, 303-311.
Nagahashi, G., Douds Jr, D.D. and Abney, G.D. (1996): Phosphorus amendment inhibits hy- phal branching of the VAM fungus Gigaspora margarita directly and indirectly through its effect on root exudation. Mycorrhiza 6, 403-408.
Nappi, P., Jodice, R. and Kofler, A. (1980/81): Vesicular-arbuscular mycorrhizas in vi-neyards, given different soil treatments in southern Tyrol. Allionia 24, 27-42.
Nappi, P., Jodice, R., Luzzati, A. and Corino, L. (1985): Grapevine root system and VA my- corrhizae in some soils of Piedmont (Italy). Plant and Soil 85, 205-210.
Nurlaeny, N. (1995): Bedeutung von Mykorrhiza und Kalkung für die Aufnahme von Phos-phat und Mikronährstoffen durch Mais und Sojabohne aus zwei sauren tropischen Böden Indonesiens. Dissertation Universität Hohenheim, Grauer Verlag, Stuttgart.
Pacovsky, R.S. (1986): Micronutrients uptake and distribution in mycorrhizal or phosphorus- fertilized soybeans. Plant and Soil 95, 379-388.
Pearson, J.N. and Jakobsen, I. (1993): Symbiotic exchange of carbon and phosphorus between cucumber and three vesicular-arbuscular mycorrhizal fungi. New Phytol. 124, 481-488.
Perret, P. (1993): Begrünung im Weinbau: Optimierung des zeitlichen Stickstoffangebotes durch Bodenpflegemaßnahmen. Deutsches Weinbau-Jahrbuch 44, 81-88.

Literaturverzeichnis 169
Perry, R.L., Lyda, S.D. and Bowen, H.H. (1983): Root distribution of four Vitis cultivars. Plant and Soil 71, 63-74.
Peyronel, M.B. (1923): Fructification de l `endophyte à arbuscules et à vésicules des mycorhi- zes endotrophes. Bull. Soc. Mycol. France 39, 119-126.
Plenchette, C., Furlan, V. and Fortin, A. (1981): Growth stimulation of apple trees in unsterili- zed soil under field conditions with VA mycorrhiza inoculation. Can. J. Bot. 59, 2003-2008.
Possingham, J.V. and Groot Obbink, J. (1971): Endotrophic mycorrhiza and the nutrition of grapevines. Vitis 10, 120-130.
Posta, K., Marschner, H. and Römheld, V. (1994): Manganese reduction in the rhizosphere of mycorrhizal and nonmycorrhizal maize. Mycorrhiza 5, 119-124.
Raju, P.S., Clark, R.B., Ellis, J.R. and Maranville, J.W. (1990 a): Mineral uptake and growth of sorghum colonized with VA mycorrhiza at varied soil phosphorus levels. J. Pl. Nutr. 13, 843-859.
Raju, P.S., Clark, R.B. , Ellis, J.R. and Maranville, J.W. (1990 b): Effects of species of VA- mycorrhizal fungi on growth and mineral uptake of sorghum at different temperatures. Plant and Soil 121, 165-170.
Ravolanirina, F., Gianinazzi, S., Trouvelot, A. and Carre, M. (1989): Production of endomy- corrhizal explants of micropropagated grapevine rootstocks. Agric. Ecosyst. Environ. 29, 323-327.
Reimers, H., Steinberg, B. und Kiefer, W. (1994): Ergebnisse von Wurzeluntersuchungen an Reben bei offenem und begrüntem Boden. Vitic. Enol. Sci. 49, 136-145.
Reinhard, S. (1995): Beeinflussung von N2-Fixierung und Pflanzenwachstum durch die arbus- kuläre Mykorrhiza bei Erbse (Pisum sativum L.). Dissertation Universität Hohenheim, Grauer-Verlag, Stuttgart.
Remund, U., Niggli, U. und Boller E.F. (1989): Faunistische und botanische Erhebungen in einem Rebberg der Ostschweiz. Einfluß der Unterwuchsbewirtschaftung auf das Öko- system Rebberg. Landw. Schweiz 2, 393-408.
Rennenberg, H., Herschbach, C. and Polle, A. (1996): Consequences of air pollution on shoot- root interactions. J. Plant Physiol. 148, 296-301.

Literaturverzeichnis 170
Richards, B.L., Middleton, J.T., Hewitt, W.B. (1958): Air pollution with relation to agrono-mic crops: V. Oxidant stipple of grape. Agron. J. 50, 559-561.
Richards, D. (1983): The grape root system. Horticultural Reviews 5, 127-168.
Rüdel, M., Dechet, F. und Eichhorn, K.-W. (1992): Feindpflanzen virusübertragender Nema- toden - Möglichkeiten für den Weinbau? Rebe und Wein 5, 190-192.
Rühl, E.H. (1992): Effect of K+-supply and relative humidity on ion uptake and distribution on two grapevines rootstock varieties. Vitis 31, 23-33.
Ruiz-Lozano, J.M and Azcón, R. (1996): Mycorrhiza colonization and drought stress as fac- tors affecting nitrate reductase activity in lettuce plants. Agric. Ecosyst. Environ. 60, 175-181.
Runjin, L. (1989): Effects of vesicular-arbuscular mycorrhizas and phosphorus on water status and growth of apple. J. Pl. Nutr. 12, 997-1017.
Ryan, M.H., Chilvers, G.A. and Dumaresq, D.C. (1994): Colonisation of wheat by VA- my-corrhizal fungi was found to be higher on a farm managed in an organic manner than on a conventional neighbour. Plant and Soil 160, 33-40.
Saif, S.R. (1981): The influence of soil aeration on the efficiency of the vesicular-arbuscular mycorrhizas. I. Effects of soil oxygen on growth and mineral uptake of Eupatorium
odoratum L. inoculated with Glomus macrocarpus. New Phytol. 88, 649-659.
Sanders, F.E. (1975): The effect of foliar-applied phosphate on the mycorrhizal infections of onion roots. In: Endomycorrhizas, Sanders, F.E., Mosse, B. and Tinker, P.B., Eds., Academic Press, London, pp. 261-276.
Sanders, F.E. and Sheikh, N.A. (1983): The development of vesicular-arbuscular mycorrhizal infection in plant root systems. Plant and Soil 71, 223-246.
Sattelmacher, B., Reinhard, S. and Pomikalko, A. (1991): Differences in mycorrhizal coloni-za- tion of rye (Secale cereale L.) grown in conventional or organic (biological-dynamic) farming systems. J. Agr. Crop Sci. 167, 350-355.

Literaturverzeichnis 171
Schellenbaum, L., Berta, G., Ravolanirina, F., Tisserant, B., Gianinazzi, S. and Fitter, A.H. (1991): Influence of endomycorrhizal infection on root morphology in a micropropaga- ted woody plant species (Vitis vinifera L.) Ann. Bot. 68, 135-141.
Schmid, J., Manty, F. und Rühl, E.H. (1998): Welche Unterlage für welchen Standort ? Das Deutsche Weinmagazin 2, 26-30.
Schreiner, R.P. and Koide, R.T. (1993): Antifungal compounds from the roots of my- cotrophic and non-mycotrophic plant species. New Phytol. 123, 99-105.
Schubert, A. and Cravero, M.C. (1985): Occurence and infectivity of vesicular-arbuscular my- corrhizal fungi in north-western Italy vineyards. Vitis 24, 129-138.
Schubert, A., Cammarata, S. and Eynard, J. (1988): Growth and root colonization of grapevi- nes inoculated with different mycorrhizal endophytes. HortScience 23, 302-303.
Schubert, A., Mazzitelli, M., Ariusso, O. and Eynard, I. (1990): Effects of vesicular- ar-buscular mycorrhizal fungi on micropropagated grapevines: Influence of endophyte strain, P fertilization and growth medium. Vitis 29, 5-13.
Schwab, S.M., Menge, J.A. and Leonard, R.T. (1983): Quantitative and qualitative effects of phosphorus on extracts and exudates of sudangrass roots in relation to vesicular- ar-buscular mycorrhiza formation. Plant Physiol. 73, 761-765.
Schweiger, P.F., Robson, A.D. and Barrow, N.J. (1995): Root hair length determines benefi- cial effect of a Glomus species on shoot growth of some pasture species. New Phytol. 131, 247-254.
Scienza, A., Failla, O. und Romano, F. (1986): Untersuchungen zur sortenspezifischen Mine- ralstoffaufnahme bei Reben. Vitis 25, 160-168.
Sieverding, E. and Leihner, D.E. (1984): Influence of crop rotation and intercropping of cassa- va with legumes on VA mycorrhizal symbiosis of cassava. Plant and Soil 80, 143-146.
Sieverding, E. (1985): Influence of method of VA mycorrhizal inoculum placement on the spread of root infection in field-grown cassava. Z. Acker- und Pflanzenbau 154, 161-170.

Literaturverzeichnis 172
Sieverding, E. (1991): Vesicular-arbuscular mycorrhiza management in tropical agrosystems. Schriftenreihe der GTZ (No. 224), Eschborn.
Skinner, P.W., Grant, R.S. and Matthews, M.A. (1988): Interaction of rootstock and my- corrhizae on dry matter distribution and nutrient levels of Cabernet Sauvignon (Vitis
vinifera L.) lamina. In: Smart, R., Thomton, R., Rodriguez, S. and Joung, J., Eds.: Proceedings second international cool climate viticulture and oenology symposium, Auckland, New Zealand, 165-168.
Solaiman, M.Z. and Hirata, H. (1996): Effectiveness of arbuscular mycorrhizal colonization at nursery-stage on growth and nutrient in wetland rice (Oryza sativa L.) after transplan- ting under different soil fertility and water regimes. Soil Sci. Plant Nutr. 42, 561-571.
Solaiman, M.Z. and Hirata, H. (1997): Effect of arbuscular mycorrhizal fungi inoculation of rice seedlings at the nursery stage upon performance in the paddy field and greenhou-se. Plant and Soil 191, 1-12.
Sommer, C. (1985): Ursachen und Folgen von Bodenverdichtungen sowie Möglichkeiten zu ihrer Verminderung. Landtechnik 40, 378-384.
Sylvia, D.M. and Schenk, N.C. (1983): Application of superphosphate to mycorrhizal plants stimulates sporulation of phosphorus-tolerant vesicular-arbuscular mycorrhizal fungi. New Phytol. 95, 655-661.
Tarafdar, J.C. and Marschner, H. (1994): Phosphatase activity in the rhizosphere and hy- phosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biol. Biochem. 26, 387-395.
Taube-Baab, H. and Baltruschat, H. (1993): Effect of vesicular-arbuscular mycorrhizal fungi on the growth of young apple trees in apple replant desease soil. Z. PflKkrankh. PflSchutz 100, 474-481.
Tawaraya, K., Watanabe, S., Yoshida, E. and Wagatsuma, T. (1996): Effect of onion (Allium
cepa) root exudates on the hyphal growth of Gigaspora margarita. Mycorrhiza 6, 57-59.
Tennant, D. (1975): A test of a modified line intersect method of estimating root length. J. Ecol. 63, 995-1001.
Tester, M., Smith, S.E. and Smith, F.A. (1987): The phenomenon of „nonmycorrhizal“ plants. Can. J. Bot. 65, 419-431.

Literaturverzeichnis 173
Tommerup, I.C. (1984): Persistance of infectivity by germinated spores of vesicular-arbuscular mycorrhizal fungi in soil. Trans. Br. mycol. Soc. 82, 275-282.
Varma, A. and Schuepp, H. (1994): Positive influence of arbuscular mycorrhizal fungus on in
vitro raised hortensia plantlets. Angew. Bot. 68, 108-115.
Vierheilig, H. and Ocampo, J.A. (1990): Effect of isothiocyanates on germination of spores of G. mosseae.
Soil Biol. Biochem. 22, 1161-1162.
Vierheilig, H., Iseli, B., Alt, M., Raikhel, N., Wiemken, A. and Boller, T. (1996): Resistance of Urtica dioica to mycorrhizal colonization: a possible involvement of Urtica dioica ag- glutinin. Plant and Soil 183, 131-136.
Vogt, E. und Götz, B. (1987): Weinbau. Verlag Eugen Ulmer, Stuttgart.
Von Tiedemann, A. und Herrman, J.V. (1992): First record of grapevine oxidant stipple in Germany and effects of field treatments with Ethylenediurea (EDU) and Benomyl on the disease. Journal of Plant Diseases and Protection 99, 533-541.
Waschkies, C., Schropp, A. and Marschner, H. (1992): Relations between replant disease, growth parameters and mineral nutrition status of grapevines (Vitis sp.) Vitis 32, 69-76.
Waschkies, C., Schropp, A. and Marschner, H. (1994): Relations between grapevine replant desease and root colonization of grapevine (Vitis sp.) by fluorescent pseudomonas and endomycorrhizal fungi. Plant and Soil 162, 219-227.
Weinstein, L.H. (1984): Effects of air pollution on grapevines. Vitis 23, 274-303.
Wulff, K. (1996): Einfluß von Kulturmaßnahmen auf die arbuskuläre Mykorrhiza in landwirt- schaftlichen Nutzflächen. Dissertation Christian-Albrecht-Universität Kiel, Grauer Verlag, Stuttgart.
Wunderer, W. (1992). Vergleich verschiedener Mulch- und Bodenabdeckverfahren und deren Auswirkung auf Boden und Erntegut. In: Umweltgerechte Weinbautechnik. KTBL- Schrift 353. Landwirtschaftsverlag GmbH, Münster-Hiltrup.
Zhao, B., Trouvelot, A., Gianinazzi, S. and Gianinazzi-Pearson, V. (1997): Influence of two legume species on hyphal production and activity of two arbuscular mycorrhizal fungi. Mycorrhiza 7, 179-185.

Literaturverzeichnis 174













