Embed Size (px)
Citation preview

Histochemie 25, 268--285 (1971) �9 by Springer-Verlag 1971
Enzym-Topochemie von Friihentwicklung und Implantation des Kaninchens
II. Glykosidasen
H. W. DENKER Anatomisches Insti tut der Universit~t Marburg a.d. Lahn
(Direktor: Prof. Dr. G. Perry)
Eingegangen am 30. November 1970
E n z y m e T o p o c h e m i s t r y of t h e E a r l y D e v e l o p m e n t
a n d I m p l a n t a t i o n in t h e R a b b i t
II . Glycosidases
~ummary. Early stages of development and the surrounding tissues of the Fallopian tube and the uterus axe studied in the rabbit from ovulation to implantation. The topochemistry of the following glycosidases is demonstrated: fl-galactosidase, neurami~ddase, glucosami~tidase, 8-glucuro~tidase, a-glur~osidase. The distribution given for neuraminidase is the result of preliminary attempts to develop histochemical procedures for this enzyme.
Results. In deavi~j eggs, the activities of 8-glucuronidase and ghcosaminidase are domi- nant. The epithelium of the Fallop~n tube shows an activity of glucosaminidase and 8- galactosid~se possibly correlated to the formation of the mucoproteid layer. These enzymes are also demonstrated in the epithelium of the uterus in all stages, whereas 8-glucuronidase has its maximum activity before eggs enter the uterus and at implantation. In the tropho- blastic knobs, these three enzymes could have a physiological role in the processes of dissolution of blastocyst coverings and implantation. Neuraminidase activity is found in the epithelium of the uterus.
Zusammen~assung. Untersueht werden Friihstadien der Entwicklung des Kaninehens yon der Ovulation bis zur Implantation, einsehliel3lich Tube und Uterus. Registriert wird die Ver- teilung yon Glykosidasen (8-Galaktosidase, ~Veuraminida~e, Olukosaminidase, fl-Glulcuronidase, a-Glukosidase), vor allem im Hinblick auf den Stoffwechsel von Mukosubstanzen. Die ver- wendete Methode zum Neuraminidasenachweis ist ein erster Versuch zur histochemischen Lokalisation dieses Enzyms.
Ergebnisse. In Furchungsstadien sind fl-Glukuronidase und Glukosaminidase auff~llig aktiv. Eine Funktion bei den wichtigen morphogenetischen Prozessen dieser Phase wird ver- mutet. Das Epithel der Tube zeigt vor allem eine Aktivita't yon Glukosaminidase und fl- Galaktosidase, die m6glicherweise eine Beziehung zur Bildung der Mukoproteidschicht haben. Im Uteru~epithel sind in allen Stadien fl-Galaktosidase und Glukosaminidase aktiv; 8- Glukuronidase t r i t t vor allem vor dem Eintri t t des Keims in den Uterus und bei der Implanta- tion hervor. Fiir die Aufl6sung der BlastozystenhiiUen und fiir die Implantation ist wahr- scheinlich die 8-Galaktosidase-, fl-Glukuronidase- und Glukosaminidaseaktivit~t der Tro~pho- bla~tsprosse yon Bedeutung. Neuraminidase ist dagegen vor allem im Uterusepithel lokalisiert.
Abk~rzungen. EDTA ~-~thylendiamintetraazetat, F B B = Fast Blue B Salt (Eehthlau- salz B), MS = Mukosubstanz, -en, NA ~ Neuramins~ure, Salins~uren, NAase ~ Neuramini- dase, N i t r o - B T ~ N i t r o Blue Tetrazolium Chloride, nMS~neu t r a l e Mukosubstanz,-en, p. c. ~ post coitum, h p.c., d p. e . = Stunden p.c., Tage p.c., PMS ~ Phenazinmethosul~at, PVP~Polyvinylpyrro l idon , s M S ~ s a u r e Mukosubstanz, -en, v/v~Mischungsverhii l tnis zweier Volumina. wJv ~ Gewicht pro Endvolumen.

Frfihentwieklung und Implantation. II 269
Einleitung
Mit dieser Arbeit werden Untersuchungen fiber den Stoffwechsel hochmole- kularer Kohlenhydratsubstanzen in Friihentwicklung und Implantat ion des Kaninchens fortgesetzt. Sie bauen auf der Kenntnis der Substratverteilung auf (Denker, 1969). In einer ersten Mitteflung (Denker, 1971a) wurde die Topochemie yon Enzymen des Glykogenstoffwechsels abgehandelt. Nun soil die Verteilung yon Glykosidasen, die hn Stoffwechsel yon Mukosubstanzen (Spicer u. Mitarb., 1965) eine Rolle spielen kSnnen, aufgezeigt werden. Einige neue Methoden, die dabei angewendet werden mu~ten, werden beschrieben und diskutiert. - - Zur Definition der verwendeten Begriffe vgl. Denker (1969).
Material und histologisehe Teehnik
Verwcndet warden Kryostatnativschnitte yon Tuben bzw. Uteri des Kaninchens mit Keimen verschiedener Entwick]ungsst~lien. NKhere Angaben fiber dieses Material und die histologisehe Verarbeitung linden sich in der crsten Mitteilung (Denker, 1971a). Ffir den Naehweis yon fl-Galaktosidase nach Rutenburg u. Mitarb. und fl-Glukuronidase wurden die Schnitte in Fomol-Ca, fiir Glukosaminidase in ~thanol naelffixiert (jewefls bei 4 ~ C).
Chemikalle~ (vgl. Denker, 1971a). Abweichcude Quellen: N-Acetyl-Neurarnirdc Acid Lactose, crystalline 99% pure: General Biochemicals; 6-Bromo-2-naphthyl-fl-D-galakto- pyranosid: Serva; Fast blue B salt (tetrazotiertes di-o-Anisidin. ZnC18) : Serva; Fomaldehyd- 16sung s~urefrei rain. 37 Gew.- % fiir die Histologie: Merck; fl-Galaktosidase alas Here, B-grade, Act. 250 NPG units/rag: Calbiochem; Gelatine: Merck; Glukose-Oxydase 10000 E/g fiir biochemische Zwecke: Merck; Neuraminidase 500E/mh Behring-Werke; Pararcsanilin: Bayer; Phcuazinmethosulfat rein: Serva.
Resaltate
1. fl-Ga~ktosid~e (EC 3.2.1.23) Fiir die Spaltung der fl-galaktosidischen Bindungen kommen mehrere ver-
schiedene Enzyme in Frage. Sie unterseheiden sich hinsichtlich des pH-Optimums, der Strukturgebundenheit, der Resistenz gegeniiber Fixierung und der Inhibitoren (Lojda, 1970), vieHeicht such in gewissem Umfang hinsiehtlich der Substrat- spezifit~t (Dahlqvist u. Brun, 1962). In dieser Arbeit werden die Ergebnisse yon zwei Methoden verglichen: a) kiinstliches Substrat, b) ,,natiirliches" Substrat; verschiedene Bedingungen. Die Bestimmung mit ,,natiirliehem" Substrat dient vor ahem als Kontroile fiir den Neuraminidase-Nachweis (S. 273). Die neuen indogogenen Veffahren (Lojda, 1970) konnten noeh nieht ber~eksiehtigt werden.
a) Methode nach Rutenburg u. Mitarb.
Methodik. Substr~t: 6-Brom-2-naphthyl-fl-D-galaktopyranosid. ,,Post-incubation coup- ling", Azofarbstoffbildung (s. Rutenburg u. Mitarb., 1958; Pearse, 1960).
Ergebniase (vgl. Tabelle 1; Abb. l a, b). Tubeneier ailer Stadien zeigen im Zytoplasma eine ]eichte Aktivit~t. Die Zona war in den 2 F/illen yon besamten Eiern schwach, aber doch deutlich angef~rbt, w~hrend sie bei unbesamten Eiern und Furchungsstadien so gut wie nicht reagiert, ebenso die Mukoproteidschicht. Das Sekret der Tube verh~lt sich praktisch negativ, das Epithel wird mittelstark gef~rbt, und zwar erscheint die Aktivit~t in der Pars ampullaris vorwiegend

to
Tab
elle
1. f
l-Oal
abto
sid~
e
Unb
esam
t B
esam
t 35
h
45 h
3
d 4
d 5
d 6
d 7
d 7
d I0
h
8 d
Kei
m
insg
esam
t
Kei
mse
heib
e
Tro
phob
last
Hii
llen
+
+
+
+
+
%%
+
%
%%
his
+
++
0 (%
) 0
0 0
0 0
0 O
bis
%
Ob
is%
+b
is
+b
is
%b
is
++
+
++
+
+
++
+
+
++
Tub
e S
ekre
t 0
,0
0 .0
0
Epi
thel
%
+
+ +
+
+
+ +
%
+
Str
oma
+
%
+
+
+
,~
Ute
rus
Sek
ret
%
%
+
0 hi
s %
0
bis
%
0 hi
s %
,0
bis
%
0 bi
s +
0
bis
Epi
thel
+
bi
s +
bi
s +
hi
s %
% %
+
+ %
%
+ +
+
+
bis
+ +
bi
s +
+
bis
+%
+
+
%%
%
++
+
++
+
++
Str
oma
(+)
(+)
(+)
%
+
+
his
+
bis
+
bis
+
bis
-}-+
+
+
+%
-+
%
+,
+ %
, +
+ +
, +
+ +
% :
Pos
itiv
er b
is s
ehr
star
k po
siti
ver
Aus
fall
der
Rea
ktio
n.
% %
+
his
+ +
% %
: Z
wis
chen
wer
t zw
iseh
en %
% %
un
d %
% +
+,
oder
Stii
rke
der
Rea
ktio
n an
ver
schi
eden
en O
rten
wec
hsel
nd.
0: N
egat
iver
Aus
faU
der
Rea
ktio
n.
(%):
Ein
gekl
amm
erte
Wer
te s
tehe
n ff
ir u
nsic
here
Rea
ktio
nsau
sfR
lle.

Friihentwicklung und Implantation. II 271
Abb. la u. b. ~-Galaktosidase nach Rutenburg u. Mitarb. Keim im Uterus, 5 d p.c.a 30• Das Uterusepithel ist stark, der Keim nur sehr schwach angefgrbt, b 180 • Das Uterusepithel
reagiert stark, der Trophoblast mgBig, die Blastozystenhiillen negativ
apikal, in der Pars isthmica in den SohleimzeUen vorwiegend basal, in den Flim- merzeUen in ihrer ganzen Ausdehnung. Das Stroma reagiert besonders sub- epithelial leicht positiv, die glatte Muskulatur der Tube wie die des Uterus ist in allen Stadien mg6ig positiv. Bei Blastozysten wird der Trophoblast stets besonders deutlich gefgrbt (Abb. l b), 71/s d p.c. vor aUem die Trophoblastsprosse. Die Keimseheibe zeigt ab 7 d p.c. eine starke Aktivitgt der guBeren Oberflgche des Ektoblastems; im iibrigen ist sie nur angedeutet positiv, ebenso verhalten sich zu allen Zeiten die Blastozystenhiillen (Abb. 1). Das Uterussekret reagiert 35 h his 3 d p.c. schwach, aber deutlich positiv, danach nut angedeutet und wechselnd. Das Uterusepithel wird bis 3 d p.c. leicht, ab 4 d p.e. stark gefgrbt; bJs 5 d p.c. ist allenthalben eine Betonung der apikalen Partien des Zytoplasmas deutlich. Ab 6 d p.c. ist die Anfgrbung mesometral genereli stgrker als antimesometral; 6 und 7 d p.c. reagieren in den oberflgehlichen und mittleren Abschnitten der Falten bzw. Krypten besonders stark die basalen Zellteile, in den tiefen Ab- schnitten und in den unmittelbar dem Keim anliegenden ZeUen die apikalen. Ab 71/s d p.c. liegt das Maximum iiberall apikal. Im Stroma des Endometriums ist die Anfgrbung besonders subepithelial in der Nghe des Lumens und in der glatten GefgBmuskulatur zu finden.
Zur Krit ik der Methode ist anzufiihren, dab es sich hierbei um einen Nachweis handelt, der in zwei zeitlich getrennten Schritten durchgefiihrt wird (,,Post- incubation coupling"), wobei es stets zu Diffusionsartefakten kommen kann. Auch besteht eine Affinitgt des bier verwendeten Naphtholderivates zu bestimm- ten Gewebsstrukturen (bestimmte Proteine; Lipid]6slichkeit; vgl. Pearse, 1968; Lojda, 1970; S. 276). Eine zytochemische Lokalisation (lysosomal) erhglt man daher nicht.

272 H.W. Denker:
b) Methode nach Dah lqv i s t u. Brun
Methodik. Substrat ist Laktose (,,natiirliches Substrat"). Die durch die Enzymwirkung freigesetzte Glukose wird mit Glukoseoxydase durch gekoppelte Formazanbildung nach- gewiesen (Dahlqvist u. Brun, 1962; Lojda, 1965). Damit sind die Probleme der Dehydroge- nasen-Histochemie zu beachten (vgl. ReiB, 1967).
Mit dieser Prozedur werden demnach die Orte demonstriert, an denen eine Freisetzung von Glukose erfo|gt. Das System ist sehr kompliziert (vgl. auch Abb. 2, S. 273). Auf allen Stufen ist die Entstehung yon Diffusionsartefakten m6glich, vor allem wenn eine der be- teiligten Komponenten eine besondere Affinit~t zu bestimmten Gewebsstrukturen hat. Die Diffusion kann vermindert werden dutch Zusatz yon Polyvinylpyrrolidon (PVP) oder durch Ausfiihrung in einem Agarge|medium (s. Lojda, 1965). Im iibrigen konnte Lojda (1965) fl-Galaktosidaseaktivit~t mit dieser Methode nur dann deut|ich naehweisen, wenn die In- kubation in besonderen Inkubationskammern (s. Bartonicek u. Lojda, 1963) unter ann~hernd anaeroben Bedingungen erfolgte. Sie wurden auch hier verwende~.
In der vor|iegenden Arbeit diente dieser fl-Galaktosidasenachweis nach Dahlqvist und Brun vor allem zur Kontrolle der Ergebnisse des Neuraminidasenaehweises (S. 273). Einzel- hei~n der Methode warden daher der NAase-Prozedur angepaBt.
Material. 1..4u~ga~Jsl~gunge~ (bis auf A. frisch bereitet): A. Veronal~cetatpuffer nach Michaelis 0,1 m pH 6,0; B. LaktoselSsung in Aqua dest. 10 % (w/v); C. Phenazinmethosulfat- |Ssung (PMS) in Puffer A. 0,033 mg/ml; D. Nitro-BT-LSsung in Aqua dest. 2 mg/ml. I1. In- kubaJionsmedlum: Das Inkubationsmedium wird aus entspreehenden Volumina der Ausgangs- 16sungen (I.) in der folgenden Weise hergestellt: Glukoseoxydase in der PMS-LSsung 15sen, dann Nitro-BT-LSsung (D.), Puffer (A.), zur Vo|umenkorrektur ggf. Aqua dest. und zuletzt Laktosel6sung (B.) zugeben.
Konzentrationen im Inkubationsmedinm: Laktose 1,5 (oder 10) mg/ml; G|ukoseoxydase 10000 E/g 1 mg/ml; PMS 0,01 mg/ml; Nitro-BT 0,2 (oder 0,4) mg/ml; Puffer 0,05 m.
Kontrollen. Ebenso ohne Laktose, Glukoseoxydase oder Nitro-BT. Aus]~hrung. Auf Objekttr~gern montierte Nativschnitte werden mit einer Inkubations-
kammer (s. o.) bedeckt, die mit dem Inkubationsmedium gef~illt ist; Luftblasen sind zu ver- meiden. Inkubation bei 37 ~ C 14 h, spiilen in Aqua dest., fixieren in Formol (Formaldehyd- 16sung etwa 40 Gew.-% 10 ml, Aqua dest. 90 ml) 10 rain, flieBend Leitungswasser 10 min, Aqua dest., Einschlul3 in Glyceringelatine.
Resultat. Die blauschwarze F~rbung des reduzierten Nitro-BT zeigt die Orte der Enzym- aktivit~t an.
Ergebnisse. I n den Versuch kamen nur Ke ime und Uter i der S~adien 62/8 und 7 d p .c . Gewer te t wurden ausschlieBlich blauschwarze Granula , die s te ts rein- g ranu la r sind. Braune F~rbungen zeigen sich im gesamten Uterusepi the l , mi t e iner Be tonung der F u r c h e n und der basalen Zel lpar t ien, wo sich grob granul~re S t r u k t u r e n t ingieren, die auch im ungef~rbten P r ~ p a r a t durch anderes Brechungs- vermSgen e rkennba r sind. Diese S t ruk tu r en speiehern viele Farbs tof fe und werden bei verschiedenen E n z y m r e a k t i o n e n dargesteUt; es dfirfte wohl die Region der Lysosomen sein. I n Kon t ro l l en zu dieser f l -Galak tos idasereak t ion (ohne Subs t ra t , ohne Glukoseoxydase) i s t bier die braune Anf~rbung ebenfalls, wenn auch e twas schw~cher, e rkennbar , w~hrend die b lauschwarzen Granula fehlen. I n Abwesen- heir yon Ni t ro -BT bleiben die P r~pa ra t e p rak t i sch vSllig farblos. Bei der b raunen F~rbung mag es sich daher um eine unspezifisehe Anlagerung yon Ni t ro -BT hande ln ; sie wurden n ich t als Zeichen einer E n z y m a k t i v i ~ t gewertet .
Die Ergebnisse en tsprechen hinsicht l ich der b lauschwarzen Pr~z ip i ta te im Pr inz ip denen, die m a n mi t der Methode yon R u t e n b u r g u. Mitarb . (S. 269) er- h~lt : Der Fa rbs to f f f inder sich im Bereich des Uterusepi thels , weniger im Bereich des Myomet r i ums und nur angedeu te t im Ute rus -S t roma und im Ke im. Die Kehnhf i l len reagieren negat iv , fiber dem Uterussekre t s ind in den Schle imhaut -

Friihentwicklung und Implantation. II 273
furchcn Granuls vorhanden, doch ist wegen der aUgcmein nnscharfen Lokslisation der Methode eine Bewertung nicht gut mSglich. Auf f~ ig ist eine allgemeine Be- tonung der Region der Psraplszentsrfurchen. Die blsusehwarzen Formazan- niederschl~ge sind hier fiber das Epithel und das subepitheliale Stroms ziemlich regellos vcrteilt. Eine schiirfere Lokalisstion ergibt sich such nicht bci kiirzeren Inkubationszeiten. Die Lokalisation wird offenbar durch Diffusionsartefakte stark becintr~chtigt. Da die Methode sls Kontrollc bewuBt eng an das Vorgehen beim Neuraminidsse-Nschweis (S. 274) sngeglichen wurdc, wurde suf eine Diffusions- minderung in Gelmedien (Lojds, 1965) verzichtct. Von den verschiedenen Kon- zentrstionen an Substrst und Nitro-BT bringt jeweils die geringere einen st~rkeren Resktionssusfall.
2. Neuraminidaze (NAase ; N-Acetylneuraminat-Glylmhydrolaze ; EC 3.2.1.18)
Eine histochemische Prozedur zum Nschweis yon NAase-Aktivit/it ist in der Literstur bislang nicht sngegeben. Ich hsbe mich im Rahmen dieser Arbeit be- miiht, eine solche Methode zu finden, d~ meine friiheren Kohlenhydrst-Substrst- Untersuchungen eine Beteiligung yon NAase bci der AuflSsnng der Blastozystcn- hiillen und der Implantation denkbar crscheinen lieBen (Denker, 1969). Die Er- gebnissc dieser Versuche sollcn bier vorgelegt werdcn.
MeShodilr Zv_r Spezifit~t des Enzyms vgl. Gottschalk (1957), Rafelson u. Mit~rb. (1966), Drzeniek (1967). Ein geeignetes synthetisches Substrat steht zur Zeit nicht zur Verfiigung, Synthesen mit der Neuramins~ure sind schwierig und erst in jiingster Zeit m6glich. Daher
NA-Laktose
NAose
NA § Lak tose
Gotoktose
- Gatal~osidase
Gtukoso
I ~ .- Gtukons~ure / \ red u z ierte
Gtukose- oxydose G|ukose -
oxydose
reduz ie r tes P H S P H S
X reduziertes
Nitro - B T Nitro - BT
( b touschworzes p r6z ip i ta t )
Abb. 2. NAase-Nachweis

274 H.W. Denker:
wurden Versuche mit dem einfachsten verfiigbaren natiirlichen Oligosaccharid der Neuramin- s~ure unternommen: der Sialyllaktose (NANA-Laktose), einem Trisaceharid vom Aufbau N-Acetyl-Neuramins~ure (NANA)-Galaktose-Glukose (vgl. Trucco u. Caputto, 1954; Schneir u. Rafelson, 1966). Verwendet wird das Prinzip yon Dahlqvist u. Brun (S. 272). Durch die Aktion yon NAase entstehen NANA und Laktose. Soll nach dem Prinzip yon Dahlqvist u. Brun freigesetzte Glukoso (oder Galaktose) nachgewiesen werden, so muB zun~ehst die Glykosidbindung der Laktose gespalten werden. Deswegen wird dem Inkubationsmedium ~-Galaktosidase zugesetzt. Damit nimmt das verwendete System die in Abb. 2 dargestellte Form an. Es ist gegeniiber dem yon Dahlqvist u. Brun verwendeten um eine Stufe erweitert und damit noch komplexer.
Hier liegt der Grund fiir eine Reihe yon Problemen und Schwierigkeiten: 1. Die Lokali- sation wird schlecht sein wegen der M6glichkeit der Entstehung yon Diffusionsartefakten. 2. Fflr dieses System miissen Bedingungen gew~hlt werden, unter denen drei Enzyme gleich- zeitig aktiv sind: NAase, ~-Galaktosidase und Glukoseoxydase. Die bisher bekannten NAase- Pr~parationen haben ihr pH-Optimum fast alle im Sauren (vgl. Ada, 1963; Cook u. Ada, 1963; Di Donate u. Tettamanti, 1967; Kuratowska u. Kubicka, 1967; Mahadewn u. Mitarb., 1967; Sandhoff u. Jatzkewitz, 1967; Taha u. CarubeUi, 1967; Wilson u. Rafelson, 1967). Allerdings ist eine Varianz gegeben durch eine deutliche Abh~ngigkeit yon verschiedenen Komponenten des Inkubationsmediums, u.a. yore Substrat (Ra~elson u. Mitarb., 1966; Schneir u. Rafelson, 1966). Bei starker sauren pH~Werten (~ 6) war die mir zur Veffiigung stehende ~-Galaktosi- dasepr~paration instabiL Andererseits sollte auch gepriiit werden, ob eine NAase-Aktivit~t bei dem leicht alkalischen pH der Uterusfliissigkeit demonstrabel ist. Daher babe ich pH- Werte zwischen 6,0 und 7,3 verwendet. Leider wurde mein ~-Galaktosidasepr~parat auch durch andere Komponenten des Gemischs inaktiviert, n~mlich dutch Nitro-BT und das Sub- strat, Sialyllaktose, ferner dutch PVP und Agarose. Auch die Art des Puffers spielte eine Rolle. Ca++-Ionen inaktivierten die ~-Galaktosidase ebenfalls, aus diesem Grunde mul3te ein solcher Zusatz fortfaUen, den ich geplant hatte, da eine Aktivierung der NAase durch Ca++ in den meisten untersuehten F~tlien beobachtet worden ist (Boschman u. Jacobs, 1965; Wilson u. Rafelson, 1967; vgl. dagegen zu einem fSrdernden Einflul3 yon EDTA: Wooley u. Gommi, 1.~64). Um gewebseigenes Ca++ nicht noch zu f~iUen, babe ich keinen Phosphatpufer, sondern Veronal-Acetat-Puffer benutzt.
In Reagenzglas-Vorversuchen mit NAase, ~-Galaktosidase, Glukoseoxydase und den anderen Bestandteilen des Inkubationsgemischs wurden folgende Kompromil3ansiitze ent- wickelt:
I. Au~]angsl~sungen (bis auf Puffer frisch hergestellt): A. Veronalacetatpuffer nach Michaelis 0,1 m pH 6,0; 6,5; 7,0 oder 7,3; oder Phosphatpuffer nach SSrensen 1/15 m pH 6,0; B. Sialyl-Laktose-LSsung: Sialyl-Laktose (N-Acetyl-Neuramins~ure-Laktose, NANA- Laktose) wird im Puffer (A.) des gewiinschten pH gelSst und der pH durch Zugabe yon 0,1 n KOH unter Kiihlung in Eiswasser korrigiert. Endkonzentration der Sialyl-Laktose etwa 9 mg/ml; C. PMS-LSsung in Pufer (A.) 0,033 mg/ml; D. Nitro-BT-LSsung in Aqua dest. 2 mg/ml; E. fl-GalaktosidaselSsung (250 NPG units/rag) 13,3 mg/ml.
II . Inbubation~medium. Ansatz unter Mischung der oben angegebenen Ausgangsl6sungen (I.) in der Reihenfolge: B, C, Zugabe und L~sen yon Glukoseoxydase (Subst~nz), D, E. End- konzentrationen im Inkubationsmedium: Sialyl-Laktose 3,8 mg/ml; 8-Galaktosidase 2,6mg/ml; Glukoseoxydase 1,5mg/ml; PMS 0,01 mg/ml; Nitro-BT 0,2mg/ml; Veronal- acetatpuffer 0,05 m (oder Phosphatpuffer 0,037 m).
Kontrollen. 1. Ohne fl- Galaktosidase; 2. ohne Substrat Sialyl-Laktose; 3. ohne fl- Galak- tosidase und ohne Substrat; 4. ohne Glukoseoxydase bzw. andere Komponenten des farbstoff- bildenden Systems.
Ausf~hrung. K.ryostat-Nativschnitte montieren. Inkubation in Inkubationskammern (S. 272) bei 37 ~ C 15--22 Std. Wasehen in 2real Aqua dest., fixieren in Formol (Formaldehyd- lSsung 40 Gew.-% 10 ml, Aqua dest. 90 ml) 10 rain, waschen in flieflend Leitungswasser 10 rain, Aqua dest., Einschlul3 in Glyceringelatine.
R~dtat. Die Orte einer NAase-Aktivit~t werden im Pr~parat (ira Gegensatz zu den Kontrollen) dutch blauschwarze Granula dargestellt.

Friihentwieklung und Implantation. II 275
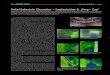
Abb. 3a u. b. Neuraminidase. Keim im Uterus 7 d p.c. a Mesometrales Endometrium und Keim, 75 • Die Formazangranula liegen vorwiegend fiber dem Uterusepithel in der Tiefe der Krypten, dagegen kaum im Bereich des Keims. b Mesometrales Endometrium, 180•
Feine Formazangranula (oben) und grobgranul~re, unspezifische (ira Original braune) Anf~rbungen (unten)
Ergebn~se (Abb. 3). Nach Inkubat ion in einem Medium mit Veronal-Aeetat- Puffer yon pH 6,0 t reten bei den geprfiften Stadien yon 62/8 und 7 d p.c. die typisehen blauschwarzen Nitro-BT-Formazan-Granula auf, und zwar vor allem fiber dem Endometrium der Paraplacentar]urchen, besonders in den tieferen Ab- schnitten der Furchen. Die Granula reichen in das Gebiet des subepithelialen Stromas und andererseits des Sekrets hinein und sind fiber all diese Orte ziemlich regellos verstreut. Die fibrigen Abschnitte des Endometr iums und der Keim zeigen nur sp~rliche Granula. Die oben unter fl-Galaktosidase (S. 273) beschriebe- hen braunen F~rbungen sind in diesem Fall ebenso zu sehen (Abb. 3b), abet wiederum auch bei Kontrollen ohne fl-Galaktosidase und ohne Substrat.
Bei den Kontrollen ohne Substrat fehlen die blauschwarzen Granula praktisch vollst~ndig; dagegen sind sie bei Inkubat ion mit Substrat, abet ohne ~-Galakto- sidase zwar vermindert, aber doch deutlich vorhanden und wie im Hauptversuch verteflt. Die ~berpri ifung des Inkubationsmediums nach der Inkubat ion auf die Aktivit~t der einzelnen Komponenten zeigt, dab die ~-Galaktosidase so gut wie nicht mehr akt iv ist. Die Aktivit~t ist in Vergleichstests schon nach wenigen Stunden sehr stark abgefallen. Andererseits entspricht die erhaltene Lol~alisation der Formazan-Granula recht gut dem Maximum der Aktivit~t der uteruaeigenen
19 Histochemie, Bd. 25

276 H.W. Denker:
fl-Gab~ktasidaze, wie sie sieh in den l~achweisen nach Dahlqvist/Lojda (gleiehes Prinzip) und naeh Rutenburg u. Mitarb. (anderes Prinzip) darstellt (S. 273). Es ist damit die MSglichkeit gegeben, dab die Granula bei diesem NAase-Nachweis am Oft der ~-Galaktosidaseaktivit/it des Endometriums entstehen, da sie die instabfle ~-Galaktosidase des Inkubationsmediums ersetzt hat. Ich habe ~-Galaktosidase w~hrend der Inkubat ion naehgegegen; dies fiihrte zu einer Inten- sivierung der F~rbung, abet zu keiner wesentliehen ~nderung des Befundes. Es ist bei der Komplexi t~t des verwendeten Systems also mSglich, dal3 NAase an einem anderen Ort als dem dargestellten akt iv ist, d~B dort die Sialyl-Laktose gespalten wird und dal~ Laktose wegen nut geringer ~-Galaktosidaseaktivit/it des Inkubat ionsmediums unver~ndert diffundieren kann zum Oft der gewebseigenen ~-Galaktosidase; dort k/~me es dann zur Glukosefreisetzung und Formazan- bfldung. Andere MSgliehkeiten fiir die Entstehung yon Artefakten seheinen mir unwahrscheinlich: Glukose oder Laktose des Schnitts kSnnen keine Rolle spielen, da die Kontrollen ohne Substrat negativ ausfielen; Glukose oder Laktose des Inkubat ionsmediums waren in den Testans~tzen nicht nachweisbar; eine bevor- zugte Bindung yon Nitro-BT an bestimmte Gewebsstrukturen gibt es zwar often- bar bei den bier untersuehten Objekten (eigene Beobaehtung), das resultierende Verteilungsmuster ist aber ein anderes. Diese Tatsache spricht ebenfalls gegen die Annahme einer Verffilsehung dutch Glukose oder Laktose des Inkubations- mediums.
Da eine Reaktion nur bei Anwesenheit des Substrats zustandekommt, scheint es mir gerechtfertigt anzunehmen, dab dieser Test tats/ich]ieh eine NAase- Alctiv~t~A demonstriert; ob die erhaltene Lokalisation mit der Lokalisation der NAase iibereinstimmt oder durch den Ort der gewebseigenen ~-Galaktosidase best immt ist, ist nicht sicher zu entscheiden. Es ist jedoeh unwahrsche/nlich, daft beide Orte sehr weir auseinander liegen. Das wfirde bedeuten, dab ein Laktose- t ransport diese Kluft iiberbriicken miiBte; sp~tere Zugabe yon ~-Galaktosidase demonstriert keine solche Laktoseansammlung an bestimmten Orten.
Bei pH-Werten yon 7--7,3 (Veronal-Acetat-Puffer) wurden nur schw~chere Ausf~lle der oben angegebenen F~rbungen bei gleichem Verteibmgsmuster beobachtet. Bei Anwendung yon Phosphatpuffer babe ich auch auf dem Trophoblasten Formazangranula erhalten, doch waren die Ergebnisse der Kontrollen widerspriichlich. Diese Versuche sollen deswegen nut am Rande erwiihnt sein.
3. N-Acetyl-fl-D-Glukosaminidaze (fl-A cet ylaminodeoxyglu]~osidase ; ,,Glukosaminidase" ; EC 3.2.1.30)
Methodik. ,,Simultaneous capture" nach Hayashi (1965a, b). Ergebnisse (vgl. Tabelle 2 ; Abb. 4, 5). Zum Nachweis dieses Enzyms erwies es
sieh als vorteilhaft, die Sehnitte auf einem diinn mit Gelatine fiberzogenen Objekt- tr~ger zu montieren (s. Denker, 1971a). Offenbar wurde dadurch eine Diffusion des Enzyms behindert.
Die Lysosomen sind in den verwendeten naehfixierten Kryostatschnit ten beseh~digt. Dadurch treten mehr diffuse Anf~rbungen hervor.
Der Keim zeigt in allen Stadien eine Aktivit~t; sie ist besonders stark in den mitt leren und sp/~teren Furchungsstadien und in den Trophoblastsprossen. Die Keimh/illen werden bei Tubeneiern (bis auf eine gelegentlich schwache Reaktion

Tab
elle
2.
N-A
cety
l.p-D
-glu
ko~m
inid
~e
Unb
esam
t B
esam
t 35
h
45 h
3
d 4
d 5
d 6
d 7
d 7
d 10
h
8 d
Kei
m
insg
esam
t
Kei
msc
heib
e
Tro
phob
last
+ +
++
+
+
++
Hii
llen
0
0 O
bis
+b
0
bis
+b
O
+ +
+
+ +
+ +
+ bi
s +
bis
++
~
++
~
~,
O
0 +
+c
++
c +
bis
++
~
Tub
e S
ekre
t
Epi
thel
Stro
ma
Ute
rus
O b
is +
O
bis
+
+ fl
bis
+
(+)
bis
(+)
bis
++
+
+
++
d
++
~t
(+)
bis
+ (+
) bi
s +
(+)
bis
+ (+
) bi
s +
O (b
is +
) +
+
(+)
bis
+
Sek
ret
Epi
thel
Str
oma
bis
+ 0
bis
+ 0
bis
+ 0
bis
+ ,0
bis
+
++
+
bis
+ bi
s +
+
++
+
+
++
(+)
(+)
(+)
(+)
(+)
bis
+ bi
s +
bis
-+-
bis
+
s T
roph
obla
stsp
ross
e.
b M
ukop
rote
idsc
hich
t.
c A
ussa
gekr
aft f
mgl
ich,
vgl
. T
ext.
d
Par
s am
pull
aris
. E
rkli
irun
g de
r Sy
mbo
M u
nd A
bki~
unge
n s.
Tab
elle
1.
bis
+ 0
bis
+ 0
bis
+ 0
bis
++
+
+
++
+
+
(bis
++
+)(
bis
+
++
)
bis
+ 0
bis
-}-
~ bi
s +
0 bi
s
tO
...j

278 H.W. Denker:
Abb. 4. N-Acetyl-fl-D-glukosaminidase. Tube, Pars isthmica, 121/4 h p.o., besamt. 160• Posi- tive Reaktion dcr basalen Abschnitte der Epi-
thelzellen
Abb. 5. N-Acetyl-fl-D-glukosaminidase. Keim im Uterus, 6 d p.c., mesometral. 26• Starke
Reaktion des Uterusepithels BlastozystenhShle
der Mukoproteidschicht) nieht, bei Blastozysten aber ab 6 d p.c. leicht gef~rbt (Abb. 5). AUerdings kommt es bei der Entws (die hier im Gegensatz zu den anderen Enzymtests durehgeffihrt wurde) zu einer Schrumpfung der H/illen, wodurch eine st~rkere Anf~rbung vorget~uscht werden kann. Die Coronazel[en sind sehr schwach positiv, ebenso, und zwar weehselnd, das Sekret yon Tube und Uterus. Das Tubenepithel dagegen reagiert deutlich positiv, und zwar vor- wiegend in den basalen Zellpartien (Abb. 4). Das Stroma yon Tube und Uterus zeigt nur sehr schwaehe Aktivit~t, im Uterus reagieren besonders die Stromazellen der epithelnahen Partien; es ist eine deutliche Betonung der Umgebung der Furehen zu sehen. ])as Uterusepithel wird in den Frtihstadien gleiehm~l~ig gef~rbt (Abb. 5), 71/s d p.e. fs auf, da~ es in der Tiefe der Krypten allgemein st~ixker reagiert als oberfl~chlich; innerhalb der Zellen ist ein F~rbungsmaximum apikal zu erkennen beim Uterusepithel, das mesometral das Uteruslumen begrenzt; 8 d p.c. ist dies auch antimesometral deutlich.
4. fl-Glukuronid~ze ( EC 3.2.1.31)
Methodik. Siehe Hayashi (1964), Hayashi u. Mitarb. (1964). Ergebnis~e (vgl. Tabelle3; Abb. 6--11). Die mit dieser Methode erhaltene
Lokalisation ist relativ seharf, obwohl bei den verwendeten nachfixierten Kryo- statsehnitten mit einer gewissen Seh~digung der Lysosomen gereehnet werden mu~.
Alle Tubenkeime reagieren m~Big bis stark positiv (vgl. Abb. 6), ebenso Granulosaepithelzellen, wenn sie nach der Besamung offenbar in Nekrobiose begriffen sind (zuvor sind sie nur sehwaeh positiv). Das Tubenepithel wird in allen Stadien sehwaeh gef~rbt, in der Pars isthmiea stellenweise, besonders tier

Tab
elle
3. [
J-O
luku
roni
dase
Unb
esam
t B
esam
t 35
h
45 h
3
d 4
d 5
d 6
d 7
d 7
d 10
h
8 d
Kei
m
insg
esam
t
Kei
msc
heib
e
Tro
phob
last
+b
is
+b
is
++
bis
+
+b
is
++
bis
+
++
+
++
+
++
+
++
+
++
HfiH
en
0 0
0 0
0
Tub
e
(§
++
+
(+)
(+)
(+)
-t-
bis
,0
,0
0 bi
s (+
) +
(+)
(+)
(+)
bis
Sek
ret
0 0
0 0
0
Epi
thel
+
+ +
+
Str
oma
(~)
(+)
(-~)
(~
) (+
)
Ute
rus
Sek
ret
0 bi
s 0
bis
0 bi
s +
++
+
++
+
+
Epi
thel
+
bis
+ bi
s +
bis
++
+
++
+
++
Str
oma
+ +
+
0 bi
s +
,0 b
is
0 hi
s +
0 bi
s ~
bis
0 bi
s +
(+)
++
§
++
+
~ bi
s +
~ bi
s -t
- (+
) hi
s (+
) bi
s (+
) +
+b
+
++
b
+ +
+
++
+
++
+
++
+
++
a T
roph
obla
stsp
ross
e.
b A
ntim
esom
etra
L
Erk
l~ru
ng d
er A
bkii
rzun
gen
und
Sym
bole
s.
Tab
elle
1.
tO
~J

280 H.W. Denker:
Abb. 6. fl-Glukuronidase. Keim in der Tube, 35 h p.e., 100x. Starke Reaktion des Keims Abb. 7. fl-Glukuronidase. Uterus. 7 d p.c., mesometral. 100 x. Stark angef~btes globul6ses
Sekret. Myometrium schwaeh positiv Abb. 8. fl-Glukuronidase. Keim im Uterus, 6 d p.c., 160 x. Stark reagieren einige Stromazellen
und das an die Blastozystenhfillen angelagerte Uterussekret Abb. 9. fl-Glukuronidase. Keim im Uterus, 7 d p.c., Keimscheibe. 160x. Starke Reaktion
der Oberfl~che der Keimscheibe und der Stromazellen des Uterus
in den Krypten, m~Big stark, die glatte Muskulatur der Tube ist allentha|bcn sehr sehwach positiv, das Sekret negativ. Das Uter~epithel ist 35 h p.c. besonders in der Ticfe der Krypten reich an m ~ i g bis stark positiven tropfig-granul~rcn Ein- schlfissen, das Sekret dort stark gef~rbt; diese Reaktionen nehmen in den folgen- den Stadicn an Intensit~t ab, so dab 5 d p.c. nur noch wcnig Aktivit~t in den tiefsten Tcilcn der Krypten und praktisch kein positives Sekret zu erkennen ist. Ab 6 d p.c. ist wieder positiv reagiercndes Sekret zu sehen, das in einzelnen globul6s geformten KSrpern hier und da im Lumen der Krypten licgt (Abb. 7) oder an die AuBenseite der B]astozystenhfillen angelagert ist (Abb. 8). Das Epithel wird von 7 d p.c. bis 7a/3 d p.c. zunchmend positiv, vor allem antimesometra], rnit eincr Bctonung wieder der tiefcn Abschnitte der Krypten, wo das Maximum

Friihentwicklung und Implantation. II
Throphoblastsprosse beim Durchdringen der Blastozystenhiillen Trophoblastsprol3
281
Abb. 10. fl-Glukuronidase. Keim im Uterus, 7 d p.c., antimesometml. 160• Stark reagieren Stromazellen, weniger Uterusepithel und Trophoblastsprosse. Keimhiillen negativ
Abb. 11. fl-Glukuronidase. Keim im Uterus, 71/8 d p.c., antimesometral. 160• Starke Reaktion des Trophoblastsprosses
intrazellul~r basal liegt. 71]8 d p.c. sind auch die mehr oberfl~chlichen Zellen des antimesometralen Endometriums gut positiv, und zwar mit Betonung der apikalen ZeUpartie. 8 d p. c. reagieren Uterusepithel und Sekret nur noeh schwach. Junge Blas~ozysten zeigen so gut wie keine Aktivit~t (ira Gegensatz zu den Furchungsstadien), allenfalls noch 4 d p.c. im Trophoblasten. 7 d p.e. dagegen ist eine deutliche Reaktion der oberfl/iehliehen Region des Ektoblastems zu erkennen (s. Abb. 9), sie ist 71/a d p.c. abgeschw~cht, aber noch deutlich, 8 d p.c. dagegen nicht mehr wahrnehmbar. Meso- und Entoblastem ergeben maximal (7---T/a d p.c.) nur eine schwache Reaktion. Der Trophoblast wird nur sehr leicht gef~rbt mit Ausnahme der Trophoblastsprosse, die 71/a d p.c. oft stark positiv sind (Abb. 11); 8 d p.c. sind nur noch geringe Reste dieser Reaktion erhalten. Die Kelmhiillen reagieren stets negativ bis auf 7---~/a d p.c., wo sie geringffigige Fiixbungen zeigen. Im Stroma des Uterus linden sich m/~13ig bis stark positive S~romazelle 35 h p.c. in geringer Zahl, ab 5 d p.c. mesometral reichlicher, ab 6 d p.e. au antimesometral, und zwar dort besonders dicht gelagert (Abb. 8 bis 10). Das lvlyometrium reagiert stets sehwach, 7 d p.e. ist eine Betonung der Ringmuskulatur deutlich. 8 d p.c. sind die Reaktionen aUer Strukturen bis auf die Stromazellen stark abgesehw~cht.
5. ~-Olukosidase (EC 3.2.1.20) Methodik. ,,Simultaneous capture", s. Lojda (1965). Ergebnis. Weder im Keim noeh in der Tube oder im Uterus lieB sich in
irgendeinem Stadium ~-Glukosidase nachweisen.

282 H.W. Denker:
Diskuss ion Die in dieser Arbeit untersuchten Glykosidasen wurden unter dem Gesichts-
punkt ausgewghlt, dal3 sie im Sto/fweohsel yon Mukosubstanzen (MS) eine Rolle spielen k6nnen (zum Begriff ,,Mukosubstanzen" s. Denker, 1969). Proteasen, die ffir den Abbau des in den MS meist vorhandenen Proteinanteils in Betraeht kommen, werden in einer weiteren Mitteflung besproehen (Denker, 1971b). Fiir die meisten dieser Enzyme kommen als physiologisehe Substrate auch nieder- molekulare Stoffe in Frage. Da fiber die topoehemisehe Verteflung dieser Stoffe in den bier interessierenden Strukturen abet gegenwgrtig niehts bekannt ist, soil dieser Gesiehtspunkt vorlgufig unberfieksiehtigt bleiben.
Der Keim ist in Furchungsstadien vor allem auff~llig reich an fl-Glukuronidase. Ffir dieses Enzym wird sowohl eine Rolle im Stoffwechsel der sauren Muko- substanzen (sMS) als auch der Steroide diskutiert (vgl. Fishman u. Fishman, 1944; Pearse, 1960; Barka u. Anderson, 1965). Dem sMS-Stoffwechsel kann in diesen Stadien insofern eine besondere Bedeutung zukommen, als bier die Schei- dung von zwei Kategorien von Blastomeren erfolgt: prgsumptive Embryonal- knotenzellen und prgsumptive Trophoblastzellen (Denker, 1969). Diese Blasto- meren unterseheiden sieh m6glicherweise durch die Ladung der sMS ihrer Zelloberflgchen. Neben fl-Glukuronidase ist aueh Glukosaminidase in Furehungs- stadien sehr aktiv. Hinsiehtlich der physiologischen Bedeutung ist zu beachten, dab das letztere Enzym auch N-Acetyl-fl-D-galaktosaminide hydrolysiert {,,N- Aeetyl-fl-hexosaminidase") (Walker, 1966).
Im Sekret der Tube kommt den MS beim Kaninchen eine besondere Be- deutung ffir die Physiologie des Keims zu. Wghrend der Tubenwanderung wird auf die Zona eine weitere, tertigre Keimhfille aufgelagert, die Muko19roteidschicht. Sie entsteht aus diesem Sekret, das sich aueh um versehiedene FremdkSrper zu /~hnliehen geschiehteten Massen verdiehten kann (vgl. Greenwald, 1958). Sieher hat sie spezielle physiologisehe Aufgaben ffir den Keim: Ganz ohne diese Hfillen ist er nieht lebensfghig. Weleher Art diese Aufgaben sind, ist abet noeh unklar (vgl. Denker, 1969). Ebensowenig geklgrt sind die physikaliseh-ehemisehen Grundprozesse ihrer Bfldung. Gesiehert ist nur eine Abh/ingigkeit vom Hormon- haushalt, die abet bereits die Sekretion selbst betrifft (Greenwald, 1958). Als rein passiver Vorgang ist die Ablagerung aber nieht zu verstehen: Eine Betefligung des Keims wird nahegelegt dutch die Beobaehtung yon Yoshinaga und Adams (1967), dab auf Mguseblastozysten in der Tube des Kaninehens keine Muko- proteidsehieht aufgelagert wird. Nennenswerte Enzymaktivit/tten konnte ieh allerdings in den Hfillen nieht naehweisen. Das Tubenepithel besitzt eine relativ hohe Glukosaminidaseaktivitgt; das Enzym geht offenbar in das Sekret fiber. fl-Galaktosidase ist mgBig und fl-Glukuronidase nur sehwaeh aktiv. Die letztere finder sieh dagegen reiehlieh in einzelnen degenerierenden FoUikelepithelzellen, die frei im Lumen liegen. Diese Zellen seheinen aueh bei anderen bioehemischen Prozessen eine Bedeutung ffir die Physiologie des Keims zu haben. So k6nnen sie zumindest in vitro Pyruvat liefern (Kennedy u. Donahue, 1969). Das erst kfirzlieh entdeekte Enzym, das an der Bindung der Kohlerdaydratbausteine der MS an die Proteinkomponente angreift, die 0-Seryl-N-aeetylgalaetosaminid-glykosidase, be- sitzt nach Sehauer u. Gottsehalk (1967) im Ovidukt des Frosehes kurz vor dem Laiehen eine relativ hohe Aktivitgt.

Frfihentwicklung und Implantation. I I 283
I m Uteru~ des Kaninehens ist ~-Glukuronidase vor allem in StromazeUen und Sekret lokallsiert. W/i~rend des Beobaehtungszeitraums sind Ver~nderungen der Aktivit/~t deutlieh: I m Epithel und Sekret zun/i.chst ein Abfall, dann erneuter Anstieg (gleiche Beobachtung bei biochemiseher Bestimmung, die allerdings das Stroma mit erfal3te, an der Ra t te : s. Wood u. Psychoyos, 1967; zum 0strogen- einfluB s. ebenda und Fishman u. Fishman, 1944; zum Vorkommen in der Uterus- fliissigkeit allgemein s. Ringlet, 1961). I m Stroma steigen Zahl und F~rbungs- intensit~t der reagierenden ZeUen zum Zeitpunkt und Ort der Implanta t ion bin an.
fl-Galaktosidase und Glukosaminidase zeigen im Uterus keine Auff/~lligkeiten. ~.Galaktosidase scheint in das Uterussekret iiberzutreten. Zum Vorkommen dieses Enzyms im Uterus vgl. aueh Bulmer (1964), zur Glukosaminidase Coleman u. Mitarb. (1967). Fiir beide Enzyme ist an eine Funktion im Stoffwechsel der sMS zu denken. Die histochemische Charakteristik dieses sMS ist im Uterussekret in allen Stadien relativ konstant. Sie ~ndert sich dagegen in den Blastozysten- hiillen. Ferner lassen sieh mit biochemischer Methodik Versehiebungen hn Spektrum der Uterussekretproteine (Glykoproteine!) in den einzelnen Stadien zeigen. Eine Bedeutung der Glykosidasen bei diesen Prozessen daft zwar vermute t werden, l ~ t sieh abet dureh die histoehemischen Tests mi t den erhaltenen gleich- f6rmigen Reaktionsausf~llen nicht belegen.
Eine Bedeutung haben die Glykosidasen vermutlich fiir die Aufl6sung der Blastozystenhiillen und fiir das Eindringen der Trophoblasten zwischen die Uterusepithelze]len bei der Implantation, denn diese Strukturen sind reich an sMS (vgl. Denker, 1969). Die Beobaehtung einer Aktivit~t yon fl-Glukuronidase, fl-Galaktosidase und Glukosaminidase in den Trophoblastsprossen stiitzt diese Vorstellung. Neuraminidase andererseits l~13t sich nicht in diesen Strukturen, sondern im Uterusepithel demonstrieren. Sollte diese Lokalisation richtig sein, was aus methodischen Griinden nicht sicher ist (S. 275 f.), so miil3te man sich vor- stellen, dab die Neuraminidase der Uterusfliissigkeit (Sehwick, 1963) vom Endo- metr ium sezerniert wird und dann in diesem Sekret ubiquit~r, d.h. nicht lokal wirkt. Fiir die streng lokaUsierte Invasion des Trophoblasten in das Uterus- epithel ist wahrscheinlieh die hohe proteolytisehe Aktivit~t der Trophoblast- sprosse wesentlieh (Denker, 1971b).
Literatur
Ada, G.I.: Purification and properties of avian neuraminidase. Biochim. biophys. Acta (Amst.) 78, 276--284 (1963).
Barka, T., Anderson, P. J.: Histochemistry. - - Theory, practice, and bibliography. New York-Evanston-London: Hoeber 1963, 1965.
Bartonicek, V., Lojda, Z.: Incubation chambers made of plastic. Acta histochem. (Jena) 51, 389---391 (1963).
Beier, H. M. : Biochemisch-entwicklungsphysiologische Untersuchungen am Proteinmilieu fi~r die Blastozystenentwicklung des Ka~inchens (Orycytolagus cuniculus). Diss. Nat. Fak. Marburg, 1967. Zoo]. Jb. Abt. Anat. u. Ontog. 85, 72--190 (1968).
Boschman, T. A. C., Jacobs, J.: The influence of ethylenediamine tetra~etate on various neuraminidases. Biochem. Z. 342, 532--541 (1965).
Bulmer, D. : The histochemical distribution of ~-galactosidase activity in the ovary and female genital tract. Anat. Rec. 149, 699--705 (1964).
Cassidy, J. T., Jourdian, G. W., Roseman, S. : The sialic acids. VI. Purification and properties of salidase from Clostridium perfringens. J. biol. Chem. ~.40, 3501--3506 (1965).

284 H.W. Denker:
Coleman, R. L., Seroggs, R. A., Whittington, A. : Purification and properties of ~-N-acetyl- glueosaminidase from bovine uterus. Biochim. biophys. Acta (Amst.) 146, 290--292 (1967).
Cook, B., Ada, G. L. : Neuraminidase in tissues of the chick embryo and chick. Biochim. biophys. Acta (Amst.) 78, 451 !61 (1963).
Dahlqvist, A., Brun, A.: A method for the histochemical demonstration of disaccharidaso activities: application to invertase and treahalase in some animal tissues. J. Histochem. Cytochem. 1O, 294--302 (1962).
Denker, H. W.: Topochemie hochmolekularer Kohlenhydratsubstanzen in Friihentwicklung und Implantation des Kaninchens. Diss. Nat. Fak. Marburg 1969. Zool. Jb. Abt. allg. Zool. u. Physiol. 75, 141--245 und 246---308 (1970). Enzym-Topochemie yon Frflhentwicklung und Implantation des Kaninchens. I. Glykogen- stoffwechsel. Histochemie 2~, 256--267 (1971a).
- - Enzym-Topochemie yon Friihentwicklung und Implantation des Kaninchens. III. Prote- asen. Histochemie im Druck (1971b).
DiDonato, S., Tettamanti, G.: Ricerche sulla localizzazione endocellulare della sialidasi cerebrale. Boll. Soc. ital. Biol. sper. 48, 1032--1035 (1967).
Drzeniek, R . : Differences in splitting capacity of virus and V. cholerae neuraminidases on sialic acid type substrates. Biochem. biophys. Res. Commun. 26, 631--638 (1967).
Fishman, W.H., Fishman, L.W.: The elevation of uterine fl-glucuronidase activity by estrogenic hormones. J. biol. Chem. 152, 487--488 (1944).
Gottsehalk, A. : Neuraminidase: the specific enzyme of influenza virus and Vibrio cholerae. Biochim. biophys. Aeta (Amst.) 28, 645--646 (1957).
Greenwald, G. S. : Endocrine regulation of the secretion of muein in the tubal epithelium of the rabbit. Anat. Rec. 180, 477--496 (1958).
Hayashi, M. : Distribution of fl-glueuronidase activity in rat tissues employing the naphthol AS-BI ghicuronide hexazonium pararosanilin method. J. Histochem. Cytochem. 12, 659--669 (1964).
- - Histochemical demonstration of N-acetyl-fl-glueosaminidase employing naphthol AS-BI N-acetyl-fl-glueosaminide as substrate. J. Histochem. Cytochem. 18, 355--360 (1965a).
- - The cytologic demonstration of N-acetyl-fl-glucosaminidase employing naphthol AS-BI N-aeetyl-fl-glucosaminide and hexazonium pararosanilin. J. Histochem. Cytochem. 18, 26 (1965b).
- - Nahajima, Y., Fishmau, W. H. : The cytologic demonstration of fl-glucuronidase employing naphthol AS-BI glueuronide and hexazonium pararosanilin; a preliminary report. J. Histochem. Cytochem. 12, 293--297 (1964).
Kennedy, J. F., Donahue, R. P. : Human oocytes: maturation in chemically defined media. Science 164, 1292--1293 (1969).
Kuratowska, Zofia, Kubicka, T.: Purification and some properties of the neuraminidase from rabbit kidney. Acta biochim, pol. 14, 255--259 (1967).
Lojda, Z.: Some remarks concerning the histochemical detection of disaccharidases and glucosidases. Histochemie 5, 339--360 (1965).
- - Indigogenic methods for glyeosidases. IL An improved method for ~-D-galactosidase and its application to localization studies of the enzymes in the intestine and in other tissues. Histochemie 28, 266---288 (1970).
Mahadevan, S., Nduaguba, J. C., Tappel, A. L. : Sialidase of rat liver and kidney. J. biol. Chem. 242, 4409 ~13 (1967).
Pearse, A. G. E. : Histochemistry. Theoretical and applied, 2nd ed., 1960, 3rd ed., part I. London: Churchill 1968.
Pearson, B., Standen, A. C., Esterly, J. R.: Histoehcmical ~-ghicuronidase distribution in mammalian tissue as detected by 5-bromo-4-chloroindol-3-yl-~-D-glucopyruroniside. Lab. Invest. 17, 217--224 (1967).
Rafelson, E. M., Jr., Sclmeir, M., Wilson, V.W., Jr.: Neuraminidase. The amino sugars, vol. II, part B (Balazs, E.A., Jeanloz, R. W., eds.), p. 171--179. New York-London: Academic Press 1966.
ReiB, J. : Grundiagen und neuere Ergebnisse des histochemischen Nachweises von Dehydro- genasen mit Tetrazoliumsalzen. Z. wiss. Mikr. 68, 169--189 (1967).
Ringler, I. : The composition of rat uterine luminal fluid. Endocrinology 68, 281--291 (1961).

Friihentwicklung und Implantation. II 285
Rutenburg, A. M., Rutenburg, S. H., Monis, B., Teague, R., Seligman, A. M.: Histochemical demonstration of ~-galactosidase in the rat. J. Histochem. Cytoehem. 6, 122--129 (1958).
Sandhoff, K., Jatzkewitz, H.: A particle-bound sialy! lactosidoceramide splitting mawmaUan sialidase. Biochim. biophys. Acta (Amst.) 141, 442--444 (1967).
Schauer, Huberta, Gottschalk, A. : On the distribution of 0-seryl-N-acety]galactosaminide glycosidase and N-acetyl-~-D-hexosaminidase in vertebrates and invertebrates. Z. Natur- forsch. 22b, 1030--1032 (1967).
Schneir, M. L., Rafelson, M. E., Jr. : Isolation and characterization of two structural isomers of N-acetylneuraminyllactese from bovine colostrum. Biochim. biophys. Acta (Amst.) 180, 1--11 (1966).
Taha, B. H., Carubelli, R.: Mammalian neuraminidase: Intracellular distribution and changes of enzyme activity during lactation. Arch. Biochem. 119, 55--61 (1967).
Trucco, R. E., Caputto, R. : Neuramin-lactose, a new compound isolated from the mammary gland of rats. J. biol. Chem. 205, 901--909 (1954).
Walker, P. G. : Hexosaminidases. The amino sugars, vol. II, part B (Balazs, S.A., Jeanloz, R. W., eds.), p. 155--169. New York-London: Academic Press 1966.
Wilson, V.W., Jr., Rafelson, M.E., Jr.: Studies on the neuraminidase of influenza virus. III. Stimulation of activity by bivalent cations. Biochim. bipohys. Acta (Amst.) 146, 160--166 (1967).
Wood, C., Psyehoyos, A.: Activit6 de certaines enzymes hydrolytiques dans l'endom~tre et le myom&tre au cours de la pseudogestation et de divers 6tats de r6ceptivit6 ut6rine chez la Ratte. C. R. Acad. Sci., S6r. D 26~, 141--144 (1967).
Wooley, D. W., Gommi, B. W. : Serotonin receptors: V. Selective destruction by neuraminid- �9 sc plus EDTA and reactivation with tissue lipids. Nature (Lond.) 202, 1074--1075 (1964).
Yoshinaga, K., Adams, C. E. : Reciprocal transfer of blastocysts between the rat and rabbit. J. Reprod. Fertil. 14, 325--328 (1967).
Dr. H. W. Denker Arbeitsgruppe Prof. Dr. G. H. M. Gottschewski Max-Planck-Institut fi~r Immunbiologie BRD-7800 l~reiburg i.Br. Stefan-Meier-Str. 8 Deutschland