Embed Size (px)
Citation preview

Institut far technische Chemie, Leipzig, (Direktor: Prof. Dr. sc. M. RINGPFEIL) der Akademie der Wissenschaften der DDR
Enzymatische Untersuchungen zur Citrat-Isocitrat-Akkumulation bei Hefen
U. BEHREWS, K. HIRZEL und E. SCHULZE
Die Aktivitaten der Citratsynthase. Aconitathydratase, Isocitratdehydrogenase und Isocitrat- lyase wurden in Zellhomogenaten eines Vie1 Citrat plus Isocitrat akkumulierenden Stammes von Candida lipolytica und eines wenig akkumulierenden Stammes von C. guilliermondii be- stimmt. Beim ubergang von der Tropho- zur Idiophase fie1 die Aktiviat der Enzyme deutlich ab. Besonders stark war der Abfall der Isocitratlyase bei C. guilliermondii (CQuelle n-Paraffin). WLhrend der Idiophase blieb die AktivitAt der Enzyme bis auf die der Isocitratlyase, die weiter langsam abnahm, erhalten. Es wird diskutiert, dafi die Abnahme der Enzymaktivittiten beim ubergang zur Idiophase m6glicherweise auf methodische Unzuknglichkeiten des Aufschlusses. durch Veranderung der Membranstrukturen hervorgerufen, beruht, daL3 aber die Abnahme der Lyaseaktivitat einer der FaMoren sein kann, der die Produktionsrate beeinflufit. Die Verringerung der Isocitrat- lyaseaktivitat wird als Beispiel ekes verschwindenden (disappearing) Enzyms, das fiir die normale Zellfunktion nicht mehr benotigt wird, gedeutet.
I. Eilzfiiltrung
Seit iiber 50 Jahren wird Citronensaure aus Melasse mit Stammen von As@ergillus iriger im in- dustriellen MaBstab hergestellt. Die Ursache der Uberproduktion konnte noch nicht aufgeklart werden. Einige Autoren sehen in einer Abnahme der ACH1 und ICDH die Ursache [14, 251, warend andere [13] eine Veranderung der Mengen bzw. Ekeinflussungen der AktivitSten der ACH und ICDH nicht mit einer uberproduktion in Zusammenhang briigen k6nnen. Nach neueren Anschauuugen soll die energetische Regulation (negative Energieladungskontrolle) gestort sein [IS], was m6glicherweise durch einen abnormen Stoffwechsel des c-AMP hervorgerufen wird. Andererseits konnte aber auch c-;IMP die Ausscheidung des Citrats aus der Zelle aktivieren [IS. 271.
Vor einigen Jahren entdeckten japanische Wissenschaftler, daO einige Hefearten in Medien mit Paraffin, Glucose aber auch anderen Verbindungen als Kohlenstoffquelle, groI3e Mengen an Citrat plus Isocitrat akkumulieren [I, 211. Neben Untersuchungen iiber Ursachen der uberproduktion [ZO]
bei Hefen wurden in der Folgezeit insbesondere solche zur Beeinflussung des Citrat-Isocitratverhalt- nisses durchgefiihrt [6]. Der Aktivitat der ACH wird bei der Beeinflussung des Citrat-Isocitratver- haltnisses groI3e Bedeutung beigemessen, wahrend die Aktivitit der ICDH in Medien mit Paraffinen alz Kohlenstoffquelle die Ausscheidung von Citrat und Isocitrat kontrollieren soll [ZI - 231. Gleich- zcitig mu8 eine volle Funktion des Glyoxplatcyclus gewahrleistct sein, was durch eine hohe Aktivitat tlrr ICL auch wahrend der Idiophase angezeigt wird [11, 121.
In vorliegender Arbeit soll durch Messung der Aktivitaten der CS, ACH, ICDH und ICL wahrend der Tropho- und Idiophase die Veranderung der Enzymmengen und die Produktbildung untersucht werden.
Abkarzungen: ACH = Aconitrathydratase; CS = Citratsynthase; ICDH = Isocitratdehydro- genase; ICL = Isocitratlyase.

526
2. Material zlnd Methoden
2. I . Mikrobiologisclrc Methoden
Strimme ; Candida lifiolytica und Candida guilliermondii aus dcr Stammsanimlung des Instituts
Medim (fur 10 g/l Hefetrockensubstanz, HTS) : fiir technische Chemie.
Hefeextraktmedium: NH,C1 2,o g; Hefeextrakt ,,Bramsch" 2 4 g; KH,PO, 0.5 g; MgSO, - 7 H,O 0,25 g; Spurensalzlosung 7,5 ml. Thiaminmedium: NH,Cl 3.0 g; KH,PO, 0,7 g; MgSO, . 7 H,O 0,35 g ; SpurensalzlBsiing 7,5 nil; Thiamin-HC1 O,I mg; FeSO, 3,5 mg. Die Medien werden in dest. Wasser angesetzt und auf I 1 aufgeftillt.
Spurensalzlosung: In I 1 dest. Wasser CuSO, 4 g; ZnC1, P,I g; CoSO, 0.5 g ; MnSO, - 5 H,O 4,o g;
Kohlenstoffquelle: Die Medien enthielten entweder 12% Glucose odcr es wurden IOO g Paraffin auf I 1 zugesetzt.
Die Sterilisation erfolgte in bekannter Weke, lediglich Thiamin wurde als Lijsuiig getrennt durch Filtration sterilisiert. Die Kultivierung erfolgte unter aseptischen Bedingungen in einem 16 1-Fer- ~nenter mjt 7 1 Fiillvolumen bei pH 4,5: T 30 'C; Luftdurchsatz 500 1 h-l.
Die Animpfung erfolgte im Verhatnis I : 10 mit Kulturen, die 24 h beliiftet waren und wrlche die gleiche Medium-Zusammensetzung hatten wie die Hauptkultur, jedoch nur 30 g 1-1 Parafiin odcr Glucose und zusatzlich 5 g - 1-1 CaCO, enthielten. 2.2. Bestimmung der Ewyrnaktivitdten
Die Hefezellen wurden durch Zentrifugation vom Medium getrennt und dreimal mit SORENSEN- Phosphatpuffer (pH 7,s) gewaschen. Die so erhaltene feuchte Biomasse wurde rnit Seesand bei 4 "C 15 min im Morser zerrieben, rnit Phosphatpuffer (pH 7.8) aufgenommen und bei o 'C 60 min zentri- fugiert (15000 U/min). Der uberstand - Rohextrakt - wurde for die Enzymbestimrnuiigcn ver- wendet.
Die Messung der Extinktionsanderung/Zeit erfolgte im Zweistrahlspektralphotomcter CARY 17 bei 23-25 "C. Dic spozifische Aktivitat ist durch pMol Substratumsatz min-l mg-l Protein defi- niert. CS (EC 4.1.3.7.) wurde ~ ~ ~ ~ B E R G M E ~ R [3]; ACH (EC4.2.1.3.) nach REEVES u. a. [16]; ICL (EC 4.1.3.1.) nach DIXON u. a. [4] und ICDH (EC 1.1.1.4.2.) nach KORNBERG [IO] bestimmt.
Die Bestimmung des Proteingehaltes des Rohextraktes erfolgte nach der Biuret-Methode mit dem Reagenz nach BEISENHERZ [z]. Als Standard diente Humanserum-Albumin. Der Farbkomplex wurde bei 555 nm geinessen. Bei Glucose als C-Quelle wurde Wasser als Blindwert beriicksichtigt; bei Paraffin trat eine leichte Opaleszenz auf. hier wurde eine Rohextraktprobe, entsprechend ver- diinnt, fiir die Blindwertbcstimmung herangczogen.
H,BO, 5 7 g.
2.3. Bestimmung der !iauren
Citronensawe wurcle in der von Biomasse befreiten Losung nach der Kupferkomplexmethodc von LEOPOLD u. a. [q] bestimmt. Citronen- plus Isocitronensaure wurde nach der Methode von SAFRAN u. a. [17] modifiziert nach TANAKA [24] bestimmt. Beide Sauren geben nach Reaktion rnit Pyridin und Essigsaure gelblich gefiirbte Losungen. Als Eichlosung wurde eine Citronensaurelosung verwendet. Zur Errechnung des Isocitronensaure-Anteils wurde die Differenz der Werte zwischen der Kupferkomplexmethode und der Pyridinmethode rnit dem Faktor 1.5 multipliziert.
Die Gesamtsaure (Citronensawe plus Isocitronensaure) ist die Summe aus dem Wert ffir die Ci- troncnsaure nach der Pyridinmethode und dem ftir die Isocitronensaure errechneten Wert.
3. Ergebnisse
Es wurden die spezifischen Enzymaktivitaten (CS, ACH, ICL und ICDH) von zwei Stammen bestimmt, wovon der eine, C. lipolytica, ein guter und der andere, C. guillier- mondii, ein schlechter Produzent ist. Die Kultivierung erfolgte in Medien mit Glucose oder n-Paraffin als Kohlenstoffquelle. Die Proben zur Charakterisierung der Tropho- phase wurden gegen Ende des Wachstums bei noch vorhandener Stickstoffquelle und

Citrat-Isocitrat-.4kkumulation bei Hefen 527
I I Paraffin
; Glucose 1
Tabelle I
Spezifische Aktivitaten ($101. min-’ * mg-’ Protein) der CS, ACH, ICDH und ICL yon C. lipolytica (guter Siiurebildncr) und C. guilliermondii (maBiger Saurebildner). Hefecxtraktmedium mit n- Paraffin bzw. Glucose als C-Quelle. Robenahme wkhrend dcr ausklingenden Trophophase und nach aostt~ndiger Produktion. Die Ergebnisse sind Mittelwerte aus mehreren Fermentationen (2 - 7) _ _ _ _ _ _ _ _ _ ~ -
Hefe I C-Quelle j Phase I cs I ACH I ICDH I JCL
0,2 1 0,2
Produktion 0 , 3 i 0,I 0, I 0 , O l
I Wachstum 182 0.4 F’roduktion Or9 02 0, I 0, I
Wachstum o,G ’ 082 0,2 1 0 , 0 4
C. lipolytica
C. girillierniondii i Paraffin I , Glucose
I Wachstum 029 I 0822 0,2 1 0.2
PI oduktion 0 , 5 ~ O917 0,13 I o,oz Wachstum O,G I 092 0.23 ~ z;;:3 F’roduktion 0 ,3 I 0.1 0.15
4. Diskussion
TABUCHI u. a. [23] bestimmten wahrend der Citronensauresynthese die Aktivitaten der ACH und ICDH bei A . niger und C. li$olytica. Im Gegensatz zu A . niger, wo die Enzymmengen stark abfielen, blieben diese bei der Hefe weitgehend unverandert.
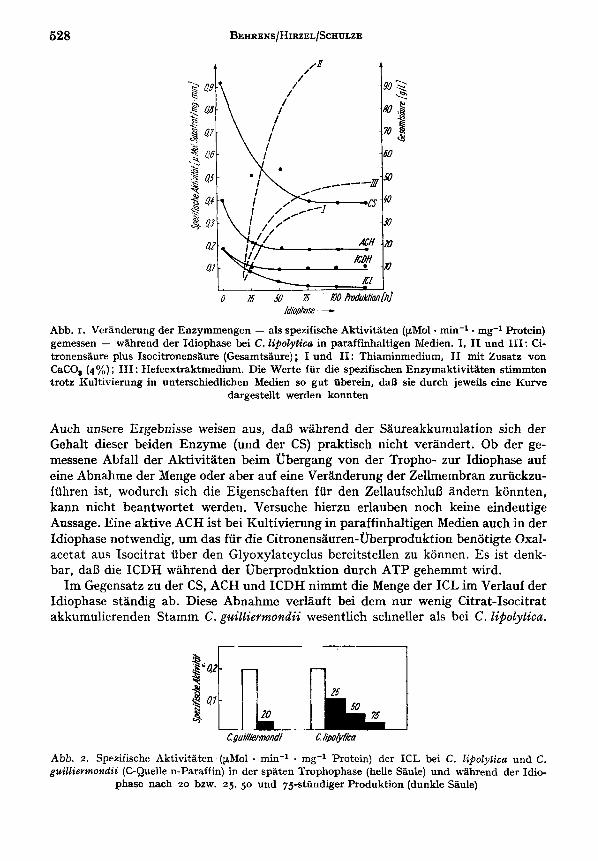
528
fb7ophse - Abb. I. Vcrandcrurig der Enzymmengen - als spezifische Aktivitaten (pMol rnin-' mg-1 Protein) gemessen - wahrend der Idiophasc bei C. lipolytica in paraffinhaltigen Medien. I , I1 und 111: Ci- tronensaure plus Isocitronensanre (Gesamtsaure) ; I und I1 : Thiaminmedium, I1 mit Zusatz von CaCO, (4%) ; I11 : Hefcextraktmedium. Die Wcrte for die spezifischen Enzymaktivitaten stimmten trotz Kultivicrung in unterschiedlichen Medien so gut iiberein, daB sie durch jeweils eine Kurve
dargestellt werden konnten
Auch unsere Ergebnisse weisen aus, daD wahrend der Saureakkuniulation sich der Gehalt dieser beiden Enzyme (und der CS) praktisch nicht verandert. Ob der ge- messene Abfall der Aktivitaten beim ubergang von der Tropho- zur Idiophase auf eine Abnahme der Menge oder aber auf eine Veranderung der Zellmembran zuriickzu- fuhren ist, wodurch sich die Eigenschaften fur den ZellaufschluB andern konnten, kann nicht beantwortet werden. Versuche hierzu erlauben noch keine eindeutige Aussage. Eine ak:tive ACH ist bei Kultivierung in paraffinhaltigen Nedien auch in der Idiophase notwendig, urn das fur die Citronensauren-fiberproduktion benotigte Oxal- acetat aus Isocit rat iiber den Glyoxylatcyclus bereitstellen zu konnen. Es ist denk- bar, daI3 die ICDH wahrend der uberproduktion durch ATP gehemmt wird.
Im Gegensatz zu der CS, ACH und ICDH nimmt die Menge der ICL im Verlauf der Idiophase standig ab. Diese Abnahme verlauft bei dem nur wenig Citrat-Isocitrat akkumulierenden Stamm C. guilliermolzdii wesentlich schneller als bei C . Zipolytica.
Abb. 2. Spezifischc Aktivitaten'(pMo1 - min-' . mg-' Protein) der ICL bei C. liPoZytica und C. guilliermondii (C-Quelle n-Paraffin) in der spaten Trophophase (helle Saule) und wahrend der Idio-
phase nach 20 bzw. 25, 50 und 75stondiger Produktion (dunkle Saule)

Citrat-Isocitrat-Akkumulation bei Hefen 529
Der ICL-Gehalt konnte einer der Faktoren sein, der die Rate der UberschuBpro- duktion steuert.
Es stellt sich die Frage, worauf die Abnahme der Enzyme im allgemeinen und die der ICL im besonderen zuriickzufiihren ist. Nach GOLDBERG u. a. pi3 sol1 der Protein- abbau als Ergebnis einer Adaptation an eine nahrstoffiirmere Umwelt (,,partieller Hunger") eintreten, urn Aminosauren fur die Energiegewinnung und Synthese be- stinmter Enzyme bereitstellen zu konnen. THURSTON [z6] spricht von verschwin- denden (disappearing) Enzymen und findet diesen Vorgang hauptsachlich bei indu- zierbaren Enzymen. Bei Chlorella tyrenoides wird beim Ubergang von Acetat- auf Glucosemedien eine schnelle Abnahme der ICL-Menge registrier t, die in einem stick- stofffreien Medium beschleunigt wird. Es sollen jene Enzyme am schnellsten abge- baut werden, die fur die Zelle sinnlos geworden sind. Mit Aufhoren des Wachstums ist die anaplerotische Sequenz sinnlos geworden, da ja die in der Idiophase ablaufende f'berproduktion von Citronen- und Isocitronensaure einem pathologischen Zustand gleichkommt.
Es ist somit zu erwarten, daB die ICL, als Schlusselenzym fur ein Wachstum auf n-Paraffinen, bevorzugt abgebaut wird. Ein spezifischer Abbau der ICL kann durch eine erhohte Aktivitat intrazellularer proteolytischer Enzyme oder durch eine ver- anderte Konformation, die gegenuber proteolytischen Enzymen empfindlicher ist. verursacht werden [8].
S u m m a r y
U. BEHRESS. I<. HIRZEL and E. SCHULZE : Enzymatic examination on the citratc-isocitrate-accu- mulation by yeasts
The activities of the enzymes citrate synthase, aconitate hydratase, isocitrate dehydrogenase and isocitrate lyase in cell homogenates of n-paraffin grown citrate plus isocitrate accumulating yeasts (C. ZipoZytica high rate accumulating strain and C. guilliermondii low rate accumulating strain) were determined. I t is shown that the activities of the enzymes decline after transition from tropho- to idiophase and remain constant with exception of isocitrate lyase which diminishes slowly. The decline in activity of the isocitrate lyase was greatest in the cells of C. guilliernwndii. It is discussed that the differences of the enzymatic activities in the tropho- and idiophase, resp. may be artefacts due to changes in the structure of cellular envelope, but that the decline of lyase activity in the idiophase could be one factor determining the rate of citric- and isocitric acid overproduction. The diminishing of isocitrate lyase in the course of idiophase is interpreted as example of a disappearing enzyme no further needed for normal function of the cells.

L i t e r a t u r
[I] ABE, hI., T. TABUCIII u. Y. TAHARA, Nippon Nogei Kagaku Kaishi 44, 493-498 (1970). [z] BEISENHERZ, G., H.-J. BOLTZE, TH. BUCHER, R. CZOK, K. H. GARBADE, E. MEYER-ARENDT
[3] BERGMEYER, H. U., Methoden der enzymatischen Analyse, Bd. I, S. 404, Akademic-Verlag.
[4] DIXON, G. H., 11. H. L. KORNBERG, Biochem. J. 72, 3P (1959). [5] GOLDBERG, A. I-.. 11. J. F. DICE, Annual Rev. Biochem. 43, 835-864 (1974). [GI HATTORI, K., S. Y o ~ o o u. 0. IMADA, J. Ferment. Technol. 52, 542-550 (1974). [7] HILDEBRANDT, W., u. H. WEIDE, 2. ,Illgem. Mikrobiol. 13, 569-576 (1973). [8] ILLARIONOWA, ’J. J., T. V. FINOGENOVA u. L. M. GLAZUNOVA, F’rilrladn, Biokhim. Mikrobiol. 11,
[g] JOHN, P. C. L., C. F. THURSTON u. P. J. SYRETT, Eiochem. J. 119, 913-919 (1970).
u. G. PFLEIDERER, 2. Naturforsch. Sb, 555-577 (1953).
Berlin 1970.
172-178 (1975).
[IO] KORFBERG, A,, in Methods in Enzymology (Eds. P. P. COLOWICK u. N. 0. KAPLAN) Vol. I,
[11] LATYSHEVA, N. N., Prikladn. Biokhim. Mikrobiol. 7. 515-520 (1971). [IZ] LATYSHEVA, N. N., A. D. GOLOBOV u. S. A. KONOVALOV, Prikladn. Biokhim. Mikrobiol. 7. 632
[13] LA N a v z ~ , J. &I., J. Gen. Microbiol. 44, 33-81 (1966). [14] LEOPOLD, H.. u. 2. VALTR, Nahrung 6, 532-576 (1958). [IS] RAMAKRISHNAB’, C. V., R. STEEL, u. C. P. LENTZ, Arch. Biochem. Biophys. 55.270-~77 (1955). [16] REEVES, H. L., R. RABIN, W. S. WEGENERU. S. AJC, in Methods in Microbiology (Ed. J. R. NOR-
[17] SAFRAN, M., u. 0. F. DENSTEDT, J. biol. Chemistry 175, 849-855 (1948). [IS] SEIDEL, H., in DECHEMA Nr. 71 (8. Tutzing Symp. 1972, Techn. Biochem.), S. 79-96, Verlag
Chernie, Weinhcim 1973. [19] SMITH, J. E., A. NOWAKOWSKA-WASZCUK u. J. G. ANDERSON, in FEBS, Vol. 30, Pt. I (Industrial
Aspects of Biochemistry, Ed. B. SPENCER), S. 297-3x7, North Holland Publ. Comp., Amster- dam 1974.
S. 705 - 709, Academic Press, New York 1955.
bis 636 (1971).
RIS) 6A, 433-434, Academic Press, London 1970.
[PO] TABUCHI, T., u. S. HARA, Nippon Nogei Kagaku Kaishi 48, 417-424 (1974). [PI] TABUCHI, T., M. TANAKA u. M. ABE, Nippon Nogei Kagaku Kaishi 43, 154-1158 (1969). [PZ] TABUCHI, T., 34. TANAKA, Y. TAHARA u. M. ABE, Nippon Nogei Kagaku Kaishi 44, 562-566
[23] TABUCHI, T.. Y. TAHARA, M. TANAKA u. S. YANAGUICIII, Nippon Nogei Kagaku Kaishi 47,
[q] TANAKA, M., Nippon Nogei Kagaku Kaishi 44. 499-5504 (1970). [25] THUFEL, K., u. U. BEHNKE, Nahrung 10, 443-451 (1966). [26] THURSTON, C. F., Process Biochem. 7 (7). 18-20 (1972). [27] WOLD, W. S. X I , u. I. SUZUKI, Biochem. biophysic. Res. Commun. 50, 237-214 (1973).
(1970).
617/622 (197.7).
Dr. sc. U. BEHRENS, K. HIRZEL und E. SCHULZE, Institut fiir technische Chemie der Akademie der Wissenschaften der DDR, 705 Leipzig. PermoserstraSe 15
Emgegangen 22. 10. 1976