Embed Size (px)
Citation preview

Institut fur Tierhaltung und Tierzuchtung der Universitat Hohenheim, Stuttgart
FettzellgroQe, lipogene und lipolytische Parameter im Riickenfettgewebe von Large White- und Piitrain-Schweinen'
Von URSULA ROTHFUSS
Eingang des Ms. 21.10.1983
1 Einleitung und Fragestellung
Das Ausmafl der Fettablagerung, das durch die Anzahl und Grofle der Fettzellen charakteri- siert werden kann, wird in erster Linie durch drei Prozesse direkt beeinfluflt: - Durch die Intentsitat der de novo Fettsaurebiosynthese, die nach O'HEA und LEVEILLE
(1969) beim Schwein zu 99 % im Fettgewebe stattfindet. Dabei ist jedoch von Bedeutung, dai3 die Fettzelle in geniigendem Ausmai3 mit Glukose versorgt wird. Der Glukoseeintritt in die Fettzelle wird durch Insulin reguliert.
- Durch das Ausmafl der Aufnahme von Fettsauren aus Bluttriglyceriden, die dann in der Fettzelle wieder zu Triglyceriden verestert und abgelagert werden konnen. Beim Schwein diirfte jedoch unter normalen Fiitterungsbedingungen (d. h. geringem Fettgehalt im Fut- ter) dieser ProzeB von nicht allzu grofler Bedeutung sein, zumal die Leber am Fettstoff- wechsel kaum beteiligt ist.
- Durch das Ausmafl der Spaltung von Triglyceriden (Lipolyse). In bezug auf die Lipolyse- stimulation wird im allgemeinen von einer hormonellen Beeinflussung der Aktivitat der Lipase ausgegangen. Wahrend iiber die lipogene Aktivitat des Fettgewebes beim Schwein zahlreiche Untersu-
chungen vorliegen, sind die Ergebnisse iiber die Bedeutung der Lipolyse fur den Fettansatz noch recht widerspriichlich.
In der vorliegenden Arbeit werden an Schweinen der Rassen Piitrain und Large White - durch mehrmalige Messung an demselben Tier wahrend der Mastperiode - altersabhangige Veranderungen in der lipogenen und lipolytischen Aktivitat des Fettgewebes erfafit und ihre Beziehung zu Kriterien der Schlachtkorperzusammensetzung und der Fleischqualitat disku- tiert. Im Vordergrund steht dabei die Frage, o b diese beiden Rassen, die sich deutlich im Fettansatz unterscheiden, auch hinsichtlich metabolischer (Lipogenese, Lipolyse) und mor- phologischer (FettzellgroBe) Parameter im Ruckenfettgewebe Unterschiede aufweisen.
Als Index der Fettsauresyntheseintensitat (Lipogenese) diente der Spiegel der NADPH- liefernden Enzyme (ROGDAKIS 1972). Die lipolytische Aktivitat des Fettgewebes wurde in vitro an isolierten Fettzellen - vor und nach Stimulation - sowie in vivo im Blut ermittelt. Beide Parameter sollen nach zahlreichen Literaturangaben von der GroBe der vorhandenen Fettzellen abhangen. Auflerdem ist die Fettzelle eine gute Bezugsbasis fur biochemische Parameter des Fettgewebes (HIRSCH, GALLIAN 1968; SALANS etal. 1968; GREENWOOD et al. 1970; SALANS, DOUGHERTY 1971; HOOD, ALLEN 1973 a).
' Mit finanzieller Unterstiitzung der Deutschen Forschungsgemeinschaft (SFB 142)
U.S. Copyright Clearance Center Code Statement: 0044-3581 /84/ 10104-O280 $2.5010 2 . Tierziichtg. Ziichtgsbiol. 101 (1984) 280-290 0 1984 Verlag Paul Parey, Hamburg und Berlin ISSN 00443581 /Intercode: ZTZBAS

FettzellgroJe irn Ruckenfettgewebe von Schweinen
2 Material und Methoden
281
2.1 Tiermaterial
Als Versuchstiere wurden 14 bzw. 11 mannliche kastrierte Large White- und Piitrain- Schweine herangezogen. Die Tiere wurden mit etwa 20 kg Gewicht angekauft und bei ad libitum Futterung bis zum Gewicht von 100 kg gemastet. Da sich Piitrains in der Regel von Large Whites durch einen geringeren Fettansatz, ein hoheres Fleischbildungsvermogen, aber auch durch eine schlechtere Fleischbeschaffenheit unterscheiden (Die Tabelle 1 besta- tigt dies fur die untersuchten Tiere), kann davon ausgegangen werden, da13 dieses Tiermate- rial fur vorliegende Fragestellung geeignet ist.
Kriterien der Mastleistung, der Schlachtkorperzusammensetzung und der Fleischqualitat (Mittelwerte und Standardfehler)
Large Whites P i k t r a i n s O i f f e r e n z S i g n i f i k a n z
( n = 14) ( n = 11) d . 0 i f f e r e n z
t a g l . Zunahrne 9
Futterverwertung 1:
Ausschlachtung
Ruckenspeckdicke cm
F e t t r e i c h e Te i l s tucke X
Fle ischre iche T e i l s t . 7:
K o t e l e t t f l a c h e cm2
F l e i s c h - F e t t v e r h a l t n i s 1:
PH45
GOFO
722,9 +14,1
3,44 k , 0 7
76,5 2 0,5
2 , 7 0 %,09
18,3 0,7
49,8 0,7
31,69+0,93
0,62+0,03
6,32+0,09
61,2+ 1,8
688,9 +15,3
3,49 %,O8
80,9 0,6
2 , 7 2 +0,10
14,6 0,7
56,l 0,8
48,3l+l ,Ol
0,33kl ,03
5,59+0,10
44,4+ 2,0
+34,0
-0,05
-4,4
-0,02
+3,7
- 6 , 3
-16,62
+ 0,729
+0,73
+16,8
n.s.
n.s.
***
n.s.
**I
***
***
***
***
***
Die Proben fur die Untersuchungen (Blut und Fettgewebe) wurden viermal wahrend der Mast bei 30,50,70 und 90 kg Gewicht entnommen. Die Biopsie von Fettgewebe erfolgte wie bei ROGDAKIS (1974 a) beschrieben. Das Blut wurde durch Punktion der Vena jugularis bzw. beim Schlachtvorgang (Gewicht der Tiere etwa 100 kg) gewonnen.
2.2 Analysen- und Rechengange
Fur die Ermittlung der Enzymaktivitat, der lipolytischen Aktivitat und der Fettzellgrofie wurden die beiden Schichten des Ruckenfettgewebes entlang des Bindegewebestranges getrennt. Das Arbeits- und Rechenschema ist in Abb. 1 dargestellt.
Auf metabolische und morphologische Unterschiede zwischen den Schichten wird an anderer Stelle eingegangen (ROTHFUSS 1983).

282 Ursula Rotbfufi
Biopsieprobe
+37"C \
-3O'C
Fet tzellenisolierung i 3 O o C I
Bestimmung Bestimrnung
der NADP- des Lipidge- n Enzymi vitat gehalt \
inkubotion /
Messung der Durchrnesser der Fettzellen von 300 Fett- A
I / \ Berechnung des mittleren Volurnens bzw. Triglyceridge- haltes einer Bestimmung BeStirnmUng Fettzelle des Lipidge- der Glycerol-
A haltes freisetzung
Berechnung der Berechnung der Anzahl der Anzahl der Fettzellen je inkubierten Gewichtseinheit Fettzellen
Berechnung der NADPH-Produktion j e Fettzelle
Berechnung der Glyceroifreisetzung j e F e t t z e l l e
Abb. 1. Analysen- und Rechengange fur die Ermittlung der Aktivitat der NADPH-liefernden Enzyme und der lipolytischen Aktivitat der
Fettzellen
Ungefahr die Halfte der Probe wurde - fur die Be- stimmung der NADP-En- zyme, des Protein- und Li- pidgehaltes - in vorgekuhl- te Gefafle auf Eis gebracht und bei -30 "C bis zur Be- stimmung gelagert.
Die andere Halfte der Probe wurde sofort nach der Entnahme in 30ml KREBS-RINGER-BI- CARBONAT-PUFFER (pH 7,4; 37°C) uberfuhrt und 30 Minuten spater fur die Isolierung der Fettzel- len prapariert.
Die Praparation des Ge- webes, die Herstellung der Gewebeextrakte und die Bestimmung der Aktivitat der NADP-Malatdehy- drogenase (NADP-MDH, E C 1.1.1.40), der Glukose- 6-Phosphat-Dehydroge- nase (G6PDH, E C 1.1.1.49) und der 6-Pho- sphoglukonat-Dehydro- genase (6PGDH, E C 1.1.1.44) erfolgte nach der von ROGDAKIS (1974a) be- schriebenen Methode.
Der losliche Proteingehalt im Gewebeextrakt wurde nach der Methode von LOWRY et al. (1951) mit Rinderserumalbumin als Standard bestimmt. Die Ermittlung des Lipidgehaltes in der Fettzellsuspension und im Fettgewebe erfolgte in Anlehnung an die von FOLCH et al. (1957) sowie von STAUFFACHER und RENOLD (1969) beschriebenen Methode nach Chloro- form-Methanol+ : 1)-Extraktion. Die Isolierung der Fettzellen, die mikroskopische Ermittlung der Fettzelldurchmesser, die Berechnung des mittleren Volumens, des Triglyce- ridgehaltes und der Anzahl der Fettzellen im Gewebe bzw. in der Fettzellsuspension, die die Bezugsbasis darstellt, wurde von ROTHFUSS (1981) beschrieben.
Der groflte Teil der isolierten Fettzellen wurde fur die Ermittlung der in vitro-lipolyse- rate (ROTHFUSS 1981) verwendet. Als Mai3 der basalen Lipolyserate diente die spontane Freisetzung von Glycerol aus den isolierten Fettzellen in das Medium, ohne Zugabe von lipolytisch oder antilipolytisch wirkenden Substanzen, innerhalb von 30 Minuten. Zur maximalen Stimulation der in vitro-Lipolyse (= Lipolysekapazitat) wurde dem Medium 10 pmol L-Adrenalin-Bitartrat pro ml zugegeben. Die Bestimmung des freien Glycerols in der Fettzellsuspension und im Blutplasma erfolgte (nach Zentrifugation) anhand der Methode von EGGSTEIN und KUHLMA" (1970).
Die Daten wurden varianzanalytisch mit Hilfe des Programms von HARVEY (1972) aus- gewertet.
Fettzellgrofie im Ruckenfettgewebe von Schweinen
3 Ergebnisse
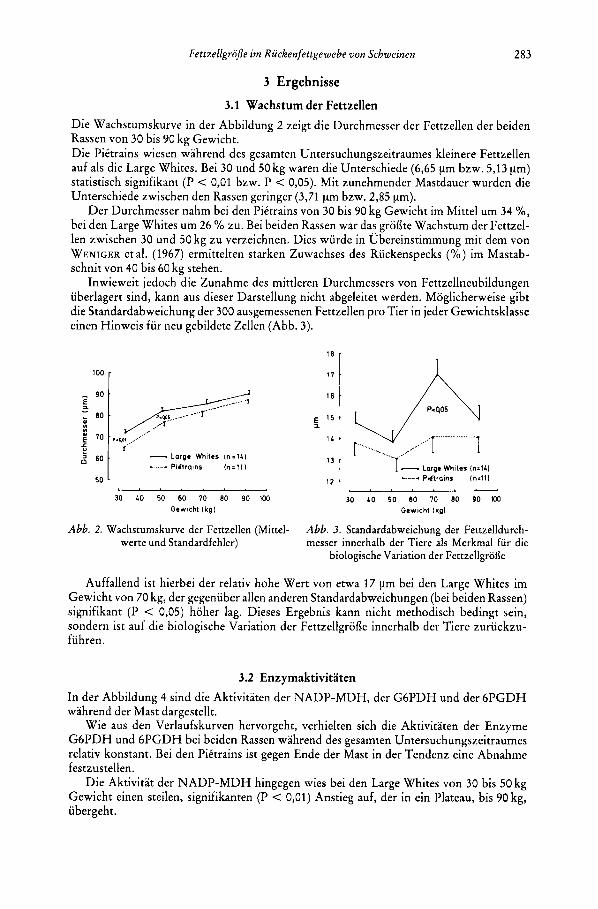
3.1 Wachstum der Fettzellen Die Wachstumskurve in der Abbildung 2 zeigt die Durchmesser der Fettzellen der beiden Rassen von 30 bis 90 kg Gewicht. Die Pittrains wiesen wahrend des gesamten Untersuchungszeitraumes kleinere Fettzellen auf als die Large Whites. Bei 30 und 50 kg waren die Unterschiede (6,65 pm bzw. 5,13 pm) statistisch signifikant (P < 0,01 bzw. P < 0,05). Mit zunehmender Mastdauer wurden die Unterschiede zwischen den Rassen geringer (3,71 pm bzw. 2,85 pm).
Der Durchrnesser nahm bei den Pittrains von 30 bis 90 kg Gewicht im Mittel um 34 %, bei den Large Whites urn 26 % zu. Bei beiden Rassen war das grodte Wachstum der Fettzel- len zwischen 30 und 50 kg zu verzeichnen. Dies wurde in Ubereinstimmung mit dem von WENIGER et al. (1967) ermittelten starken Zuwachses des Ruckenspecks (YO) im Mastab- schnit von 40 bis 60 kg stehen.
Inwieweit jedoch die Zunahme des rnittleren Durchrnessers von Fettzellneubildungen uberlagert sind, kann aus dieser Darstellung nicht abgeleitet werden. Moglicherweise gibt die Standardabweichung der 300 ausgemessenen Fettzellen pro Tier in jeder Gewichtsklasse einen Hinweis fur neu gebildete Zellen (Abb. 3).
283
I00
- 90 s : 80 UI VI
70
u 2 60
50
r
18
17
14 f"
13
12
- Large Whites In=lLl .__._. Piitrains I n = l l J
I
1 - Large Whiles ln=l4l .____.. Pi6lrains 1n:lll
30 LO 50 60 70 80 90 100 Gewicht l k g l
30 LO 50 60 70 80 90 I0 Gewicht I kgl
Abb. 2. Wachstumskurve der Fettzellen (Mittel- werte und Standardfehler)
A b b . 3. Standardabweichung der Fettzelldurch- rnesser innerhalb der Tiere als Merkmal fur die
biologische Variation der Fettzellgrofie
Auffallend ist hierbei der relativ hohe Wert von etwa 17 pm bei den Large Whites irn Gewicht von 70 kg, der gegenuber allen anderen Standardabweichungen (bei beiden Rassen) signifikant (P < 0,05) hoher lag. Dieses Ergebnis kann nicht methodisch bedingt sein, sondern ist auf die biologische Variation der Fettzellgrode innerhalb der Tiere zuruckzu- fuhren.
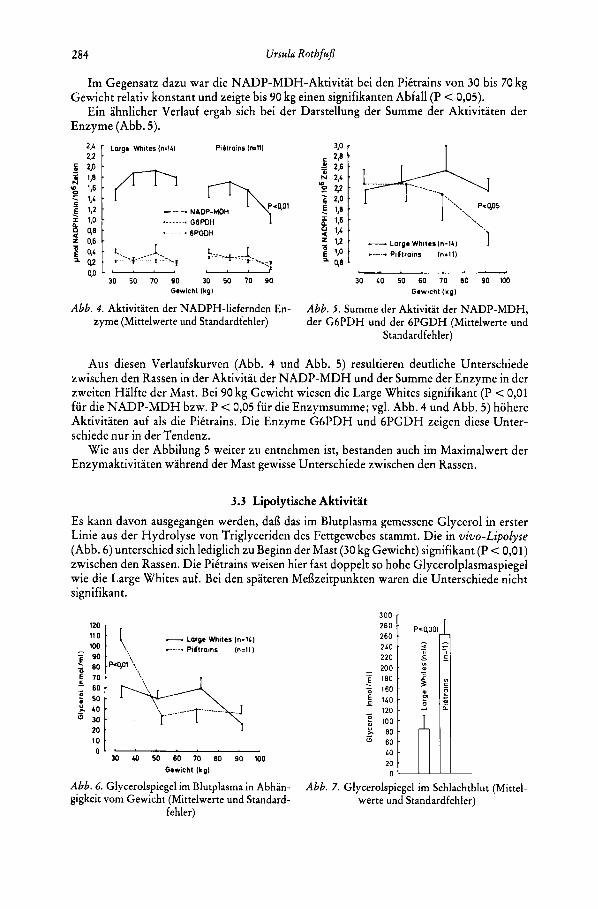
3.2 Enzymaktivitaten In der Abbildung 4 sind die Aktivitaten der NADP-MDH, der G6PDH und der 6 P G D H wahrend der Mast dargestellt.
Wie aus den Verlaufskurven hervorgeht, verhielten sich die Aktivitaten der Enzyme G6PDH und 6PGDH bei beiden Rassen wahrend des gesamten Untersuchungszeitraumes relativ konstant. Bei den Pittrains ist gegen Ende der Mast in der Tendenz eine Abnahrne festzustellen.
Die Aktivitat der NADP-MDH hingegen wies bei den Large Whites von 30 bis 50 kg Gewicht einen steilen, signifikanten (P < 0,Ol) Anstieg auf, der in ein Plateau, bis 90 kg, iibergeht.
284 Ursula Rothfuj3
Im Gegensatz d a m war die NADP-MDH-Aktivitat bei den PiCtrains von 30 bis 70 kg
Ein ahnlicher Verlauf ergab sich bei der Darstellung der Summe der Aktivitaten der Gewicht relativ konstant und zeigte bis 90 kg einen signifikanten Abfall (P < 0,05).
Enzyme (Abb. 5).
1 P14trains imtll 3.0 2.8 F
._ . . -. _. . * *6PGDH 4 ^ ^ : .^ I - Large Whitesln=lO I
*____- Pi4trains fn:ll)
0 , O L ' ' ' ' 30 50 70 90 30 50 70 90 30 40 50 60 70 eo 90 100
Gewichl lkgl Gewichl fkg1
Abb. 4 . Aktivitaten der NADPH-liefernden En- zyme (Mittelwerte und Standardfehler)
Abb. I. Summe der Aktivitat der NADP-MDH, der G6PDH und der 6PGDH (Mittelwerte und
Standardfehler)
Aus diesen Verlaufskurven (Abb. 4 und Abb. 5) resultieren deutliche Unterschiede zwischen den Rassen in der Aktivitat der NADP-MDH und der Summe der Enzyme in der zweiten Halfte der Mast. Bei 90 kg Gewicht wiesen die Large Whites signifikant (P < 0,01 fur die NADP-MDH bzw. P < 0,05 fur die Enzymsumme; vgl. Abb. 4 und Abb. 5) hohere Aktivitaten auf als die PiCtrains. Die Enzyme G6PDH und 6PGDH zeigen diese Unter- schiede nur in der Tendenz.
Wie aus der Abbilung 5 weiter zu entnehmen ist, bestanden auch im Maximalwert der Enzymaktivitaten wahrend der Mast gewisse Unterschiede zwischen den Rassen.
3.3 Lipolytische Aktivitat
Es kann davon ausgegangen werden, dafl das im Blutplasma gemessene Glycerol in erster Linie aus der Hydrolyse von Triglyceriden des Fettgewebes stammt. Die in vivo-Lipolyse (Abb. 6) unterschied sich lediglich zu Beginn der Mast (30 kg Gewicht) signifikant (P < 0,01) zwischen den Rassen. Die PiCtrains weisen hier fast doppelt so hohe Glycerolplasmaspiegel wie die Large Whites auf. Bei den spateren Meflzeitpunkten waren die Unterschiede nicht signifikant.
I20 - - Large Whites In=lhl *____. P14trains in41 1
110 ' 1
Z 6 0 . 5 0 .
E 40 . ' 30 . 20 '
t o '
\ ___-__.__- -1- ......._ ..
0 " . ' . ' ' ' 4
M 40 50 60 70 80 90 100 Gewichl Ikg)
Abb. 6. Glycerolspiegel im Blutplasma in Abhan- gigkeit vom Gewicht (Mittelwerte und Standard-
fehler)
160
: 100 80 60 LO 20
Abb. 7. Glycerolspiegel im Schlachtblut (Mittel- werte und Standardfehler)
Fettzellgrope im Ruckenfettgewebe von Schweinen 285
0,011 c 0,010 = 0,009
-
: 0,008 -> 0,001
3 2 0,001
E 0,002
; 0,006 5 g 0,005
. 0.003 =0.001 0.000
- -
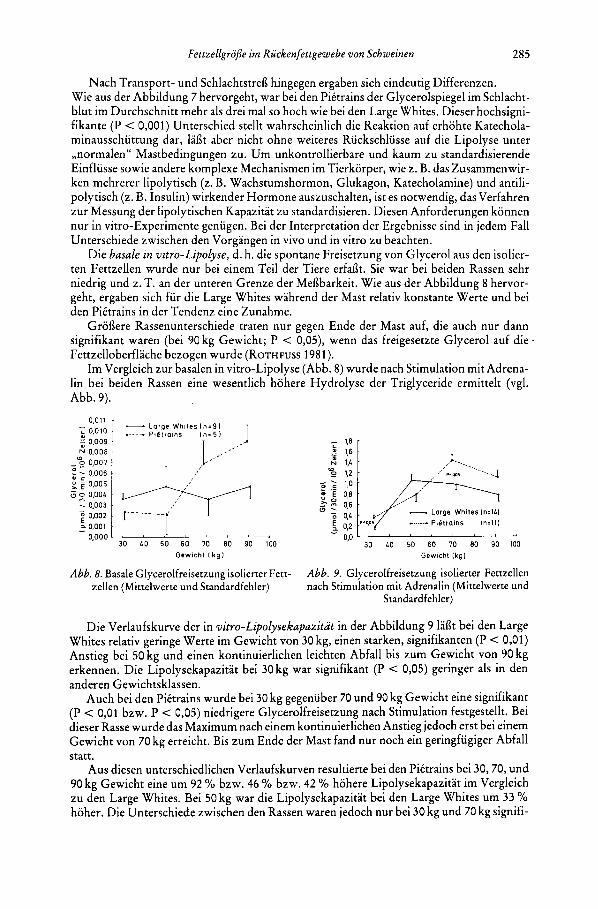
Nach Transport- und Schlachtstrefl hingegen ergaben sich eindeutig Differenzen. Wie aus der Abbildung 7 hervorgeht, war bei den Piitrains der Glycerolspiegel im Schlacht- blut im Durchschnitt mehr als drei ma1 so hoch wie bei den Large Whites. Dieser hochsigni- fikante (P < 0,001) Unterschied stellt wahrscheinlich die Reaktion auf erhohte Katechola- minausschiittung dar, Iaflt aber nicht ohne weiteres Ruckschlusse auf die Lipolyse unter ,,normalen" Mastbedingungen zu. Um unkontrollierbare und kaum zu standardisierende Einfliisse sowie andere komplexe Mechanismen im Tierkorper, wie z. B. das Zusammenwir- ken mehrerer lipolytisch (z. B. Wachstumshormon, Glukagon, Katecholamine) und antili- polytisch (z. B. Insulin) wirkender Hormone auszuschalten, ist es notwendig, das Verfahren zur Messung der lipolytischen Kapazitat zu standardisieren. Diesen Anforderungen konnen nur in vitro-Experimente genugen. Bei der Interpretation der Ergebnisse sind in jedem Fall Unterschiede zwischen den Vorgangen in vivo und in vitro zu beachten.
Die barale in wityo-lipolyse, d. h. die spontane Freisetzung von Glycerol aus den isolier- ten Fettzellen wurde nur bei einem Teil der Tiere erfaflt. Sie war bei beiden Rassen sehr niedrig und z. T. an der unteren Grenze der Meflbarkeit. Wie aus der Abbildung 8 hervor- geht, ergaben sich fur die Large Whites wahrend der Mast relativ konstante Werte und bei den Piitrains in der Tendenz eine Zunahme.
Groflere Rassenunterschiede traten nur gegen Ende der Mast auf, die auch nur dann signifikant waren (bei 90 kg Gewicht; P < 0,05), wenn das freigesetzte Glycerol auf die 1 Fettzelloberflache bezogen wurde (ROTHFUSS 1981).
Im Vergleich zur basalen in vitro-Lipolyse (Abb. 8) wurde nach Stimulation mit Adrena- lin bei beiden Rassen eine wesentlich hohere Hydrolyse der Triglyceride ermittelt (vgl. Abb. 9).
.
.
.
.
- - I 30 LO 50 60 70 80 90 100
1 p / - Large WhiteSln=lL1 W q O I c_____- Pietrains In=lO
, L ' " ' " " 30 LO 50 60 10 80 90 100
Gewichl I k g ) Gewicht ( k g )
Abb. 8. Basale Glycerolfreisetzung isolierter Fett- zellen (Mittelwerte und Standardfehler)
Abb. 9. Glycerolfreisetzung isolierter Fettzellen nach Stimulation mit Adrenalin (Mittelwerte und
Standardfehler)
Die Verlaufskurve der in vitro-lipolysekupuzitut in der Abbildung 9 laflt bei den Large Whites relativ geringe Werte im Gewicht von 30 kg, einen starken, signifikanten (P < 0,Ol) Anstieg bei 50 kg und einen kontinuierlichen leichten Abfall bis zum Gewicht von 90 kg erkennen. Die Lipolysekapazitat bei 30 kg war signifikant (P < 0,05) geringer als in den anderen Gewichtsklassen.
Auch bei den Piitrains wurde bei 30 kg gegeniiber 70 und 90 kg Gewicht eine signifikant (P < 0,01 bzw. P < 0,05) niedrigere Glycerolfreisetzung nach Stimulation festgestellt. Bei dieser Rasse wurde das Maximum nach einem kontinuierlichen Anstieg jedoch erst bei einem Gewicht von 70 kg erreicht. Bis zum Ende der Mast fand nur noch ein geringfiigiger Abfall statt.
Aus diesen unterschiedlichen Verlaufskurven resultierte bei den Piitrains bei 30,70, und 90 kg Gewicht eine um 92 % bzw. 46 % bzw. 42 % hohere Lipolysekapazitat im Vergleich zu den Large Whites. Bei 50 kg war die Lipolysekapazitat bei den Large Whites um 33 YO hoher. Die Unterschiede zwischen den Rassen waren jedoch nur bei 30 kg und 70 kg signifi-

286 Ursula RotbfuJl
kant (P < 0,05; vgl. Abb. 9). Auch der Maximalwert wahrend der Mast lag bei den Piitrains urn 37 % hoher als bei den Large Whites.
Bei Bezug der Daten auf die Fettzelloberflache (Km’) ergaben sich der gleiche Verlauf und die gleichen Rassenunterschiede.
Diese Ergebnisse sind, da sie in vitro unter kontrollierten Bedingungen durchgefiihrt wurden, nicht auf unterschiedliche Hormonkonstellation zwischen den Rassen (ROGDAKIS etal. 1979) oder auf inhibitorische Faktoren im Serum von fetten Schweinen (TRYGSTAD, Foss 1967; TRYGSTAD etal. 1972) zuriickzufuhren, sondern auf Unterschiede an oder in der Fettzelle per se (ROTHFUSS 1981).
4 Diskussion
Aus zahlreichen Untersuchungen geht hervor, daR das Wachsturn des Fettgewebes sowohl auf der Zunahme der FettzellengroRe (Hypertrophie) als auch auf der Zunahme der Gesarnt- zahl der Fettzellen (Hyperplasie) beruhen kann (z. B. fur das Schwein: ANDERSON, KAUFF- MAN 1973; STEELE etal. 1974). Nach HOOD und ALLEN (1977) liegt beim Schwein die Bedeutung eher auf der Seite der zellularen Hypertrophie. Hierdurch erklaren sich auch die von mehreren Autoren gefundenen, irn Vergleich zur Gesarntzahl der Fettzellen weitaus engeren Beziehungen zwischen der GroRe der Fettzellen und dern Verfettungsgrad (ANDER- SON, KAUFFMAN 1973; WOOD etal. 1978). In vorliegenden Untersuchungen konnten bei den Piitrains schon ab 50 kg Gewicht signifikante (P < 0,05 bzw. P < 0,Ol) positive Beziehun- gen von r = 0,67 bis r = 0,76 zwischen dem Durchmesser der Fettzellen und dem Anteil an fettreichen Teilstiicken irn Schlachtkorper errnittelt werden. Bei den Large Whites waren die Beziehungen mit r = 0,74 erst gegen Ende der Mast (90 kg Gewicht) statistisch abgesichert (P < 0,01; vgl. ROTHFUSS 1981). Auch das Wachsturn der Fettzellen im Verlauf der Mast (30 bis 90 kg Gewicht) war mit r = 0,54 (P < 0,05) fur die Large Whites bzw. r = 0,72 (P < 0,OS)fur die Piitrains positiv rnit diesem Merkmal korreliert. In Ubereinstirnmung mit den Angaben von ENSER etal. (1976) und MOODY etal. (1978), jedoch zum Teil irn Gegensatz zu den von WOOD etal. (1975) beschriebenen Ergebnissen, wurden in vorliegender Arbeit bei den Large Whites wahrend des gesamten Untersuchungszeitraumes groRere Fettzellen ermittelt als bei den mageren Piitrains (Abb. 2). Beim Vergleich von Daten iiber die FettzellengroRe miissen aber Populationsdifferenzen und Unterschiede im Verfettungsgrad der Tiere (Stichprobe) bzw. in der Methodik der Erfassung der Fettzellgroge rnit berucksichtigt werden. Die vorliegenden Ergebnisse, d. h. die groReren Fettzellen der Large Whites, das geringere Wachstum ihrer Fettzellen wahrend der Mast, die starkere Streuung der Fettzellendurch- rnesser im Gewicht von 70 kg sowie die erst gegen Ende der Mast SignifikantenBeziehungen zum Fettansatz, lassen vermuten, daR neben der Grof3e der Fettzellen vor allern die Gesamt- zahl oderhnd die Neubildung von Fettzellen bzw. die Lipideinlagerung in Pra-Adipozyten fur den starkeren Fettansatz der Large Whites gegenuber den Piitrains unter ad-fibitum- Bedingungen verantwortlich ist.
Entsprechend diesem unterschiedlichen Fettansatzvermogen waren auch Unterschiede im altersabhangigen Verlauf der Enzymaktivitat, in der maximalen Aktivitat wahrend der Mast sowie in den Spiegeln der Enzymaktivitaten in den einzelnen Gewichtsklassen zu verzeichnen. Die Untersuchungsergebnisse mehrerer Autoren iiber die lipogene Aktivitat des Fettgewebes lassen deutlich erkennen, dai3 die NADP-Enzymaktivitaten beim Schwein genetisch gesteuert werden und mit dem Fettansatz korreliert sind. Hierfur sprechen vor allem die Populationsanalysen von STRUTZ (1977), die Selektionsversuche nach dem Spiegel der NADPH-liefernden Enzyme (ROGDAKIS 1982), die Versuche zur Selektion nach der Riickenspeckdicke (ROGDAKIS 1974b; STEELE, FROBISH 1976; SCOTT et al. 1981), die Diffe- renzen im Enzyrnspiegel zwischen Rassen, die sich im Fettansatz unterscheiden (HOOD, ALLEN 1973b; LEE, KAUFFMAN 1974; MARTIN, HERBEIN 1976; ROGDAKIS etal. 1979) sowie

Fettzel1groJ.k im Ruckenfettgewebe von Schweinen 287
die positiven Beziehungen zwischen der Aktivitat der NADP-Enzyme und dem Verfet- tungsgrad der Tiere innerhalb einer Rasse oder Linie (z. B. HOOD, ALLEN 1973b; ROGDAKIS 1973; RAESIDE et al. 1980). Auch in den vorliegenden Untersuchungen wurden-vor allem in der zweiten Halfte der Mast - bei den Piitrains niedrigere Aktivitaten der NADPH-liefern- den Enzyme als bei den Large Whites gemessen. Die Beziehungen der Summe der vier Meflwerte der Enzyme wahrend der Mast zum Anteil an fettreichen Teilstucken im Schlachtkorper waren zwar nur sehr schwach und nicht signifikant (geringe Tierzahl), sie lagen jedoch mit r = 0,33 bzw. r = 0,34 fur die beiden Rassen in der erwarteten Richtung. Der Verlauf der Enzymaktivitatskurve in den Abbildungen 4 und 5, der entscheidend von der Bezugsbasis (Frischgewicht, Fettzelle, losliches Extraktprotein; ROTHFUSS 1981) beein- flufit wird, steht in Ubereinstimmung mit der starken Zunahme der Fettzellgrode, der hohen Lipidsyntheseaktivitat sowie dem groflen Fettansatzvermogen (%) von Schweinen in der ersten Halfte der Mast (WENIGER etal. 1967; ALLEE etal. 1971; ANDERSON, KAUFFMAN 1973; HOOD, ALLEN 1973a). Fur diese Parameter konnte jdoch auch die im Vergleich zu den ubrigen Gewichtsklassen geringere Lipolysekapazitat (Abb. 9) eine gewisse Rolle spielen.
Die in vitro durch Adrenalin stimulierte Lipolyse (Lipolysekapazitat, Abb. 9), die erwartungsgernafl bei beiden Rassen ein Vielfaches der basalen Lipolyse (Abb. 8) ausrnachte, deutet - mit den zu fast allen Mefizeitpunkten hoheren Werten bei den Piitrains - auf eine starkere Reaktion des Fettgewebes auf lipolytisch wirkende Substanzen im Vergleich zu den Large Whites hin. Solche Substanzen, vor allem Katecholamine, konnten unter bestimmten Bedingungen - wie Strefi, Hunger und physische Anstrengung - fur den Fettstoffwechsel beim Schwein Bedeutung erlangen.
In vorliegender in-vivo-Untersuchung stellt vermutlich das Herausfangen und Fixieren der Tiere zu Beginn der Mast (bis 30 kg Gewicht) eine solche Situation fur die Tiere dar (vgl. Abb. 6). Moglicherweise reicht die Manipulation zu den weiteren Mefizeitpunkten (50, 70 und 90 kg Gewicht) nicht mehr aus, um potentielle Unterschiede zwischen den Rassen in ihrer lipolytischen Kapazitat aufzuzeigen. Nach Transport- und Schlachtstrefi hingegen waren - entsprechend den Untersuchungen von ROGDAKIS etal. (1979) - enorme Unter- schiede zu verzeichnen (Abb. 7). Diese Ergebnisse deuten vor allem auf eine erhohte Strefi- empfindlichkeit bzw. grofiere Lipolysekapazitat der Piitrains gegenuber den Large Whites unter extremen Bedingungen hin. Nach Untersuchungen von METZ und DEKKER (1981) scheint bei diesen Rassen unter physiologischen Bedingungen die Lipolyse keine wesentli- che Bedeutung fur den Fettansatz zu haben; die Autoren schliefien jedoch einen Einflufi von antilipolytischen und lipolytischen Hormonen (Insulin, Wachstumshormon und Katecho- lamine) bei der Regulation des Fettansatzes nicht aus. Vergleiche zwischen den vorliegenden Ergebnissen und den Angaben in der Literatur uber die in-vitro-Lipolyse von genetisch differenzierten Schweinen sind aufgrund von zahlreichen Unterschieden in der Versuchsan- stellung und Methodik der Versuchsdurchfuhrung nicht ganz einfach. Eine Aufgliederung von Einfluflfaktoren findet sich bei ROTHFUSS (1981). Somit herrschen bisher hinsichtlich der Frage nach der lipolytischen Aktivitat des Fettgewebes von Rassen und Linien, die sich im Fettansatzvermogen unterscheiden, allgemein noch recht widerspruchliche Auffassun- gen (2.B. TRYCSTAD etal. 1972; STANDAL etal. 1973; WEISENBURG, ALLEN 1973; BUHLIN- GER etal. 1978; SCOTT etal. 1981). Auch in bezug auf die Abhangigkeit der Lipolyse vom Alter oder Gewicht der Schweine bestehen noch Meinungsunterschiede (z. B. MERSMANN etal. 1975; BUHLINGER et al. 1978). Vorliegende Untersuchungen ergaben eine eindeutige, signifikante (P < 0,01) Abhangigkeit der in vitro Lipolysekapazitat (vgl. Abb. 9) vom Gewicht der Tiere, sowie meist hohere Werte bei den mageren Piitrains gegenuber den fetteren Large Whites. Auflerdem deuten die bisher bei diesen Rassen erfafiten Parameter darauf hin, dafi bei den Piitrains die Sekretion kataboler Hormone erhoht und die anaboler Hormone erniedrigt ist (zusammenfassende Darstellung der Literatur siehe ROTHFUSS 1981). Somit konnte der geringere Fettansatz der Pietrains moglicherweise nicht nur auf eine verminderte Lipidsyntheserate, sondern auch auf eine erhohte Lipolyserate bei entsprechen-

288 Ursula RothfuJl
den Belastungen zuruckgefuhrt werden. Interessanterweise uben die i m Fettgewebe lipoly- tisch wirkenden Hormone im Muskel glykogenolytische Effekte (SUTHERLAND et al. 1975) aus. Diese Stoffwechsellage wiirde nicht nur den geringeren Fettansatz und die i.a. geringe- ren taglichen Zunahmen, sondern vor allem auch die schlechte Fleischqualitat der Pietrains gegenuber den Large Whites erklaren.
Zusammenfassung
Im Riickenfett ewebe von Large-White- und Piitrain-Schweinen wurden im Verlauf der Mast folgende Parameter erfak und verglichen: - Wachstum der Fettzellen - Aktivitat der NADPH-liefernden Enzyme (NADP-MDH, G6PDH, 6PGDH), als Index der lipoge-
- Lipolytische Aktivitat der isolierten Fettzellen in vitro - mit und ohne Stimulation durch Adrenalin. Die invivo-Lipolyse wurde im Blutplasma erfaI3t. 1. In Ubereinstimmung mit dem hoheren Fettansatzvermogen unter ad-libitum-Bedingungen wurden
bei Large Whites wahrend der gesamten Mast grogere Fettzellen als bei den Pietrains ermittelt. Es wird vermutet, dafl fur den starkeren Fettansatz sowohl die Gro13e als auch die Anzahl der Fettzellen verantwortlich ist.
2. Hinsichtlich der Aktivitat der NADPH-liefernden Enzyme wurden im Verlauf, im Maximalwert wahrend der Mast sowie in den einzelnen Gewichtsklassen Unterschiede zwischen den Rassen festgestellt. Die Large Whites wiesen i.a. eine hohere lipogene Aktivitat als die Piitrains auf.
3. Die Ergebnisse der invivo-Lipolyse bestatigen die hohere StreBanfalligkeit der Pietrains gegenuber den Large Whites. Die standardisierten invitro-Untersuchungen an isolierten Fettzellen ergaben zu fast allen Meheitpunkten eine hohere Stimulierbarkeit der Lipolyse durch Adrenalin bei den fettar- meren Piitrains.
nen Aktivitat
Die Ergebnisse werden diskutiert.
Summary Fat cell size, lipogene and lipolyticparameters in the back fat of Large White- and ribtrain-pigs
In the back fat of Large Whites and Piitrains during fattening the following parameters were determined andcom ared: - growtg of fat cells - activity of the NADPH-generating enzymes (NADP-MDH, G6PDH, 6PGDH), as an index of
lip0 ene activity - lipobtic activity of isolated fat cells in vitro - with or without epinephrine-stimulation. The lipolysis in vivo was determined in plasma. 1. According to hi her propensity of fat deposition under ad libitum condition, during the whole
fatteningperiod t8e Large Whites had larger fat cells compared to the Piktrains. It is suggested that in addition to the size of fat cells the number of cells is accountable therefore.
2. The breeds showed also differences in the activit of the NADPH-generating enzymes concerning the course, the maximum during the fattening anlthe levels at a definit weight. The lipogene activity was generally higher in Large Whites than in Piitrains.
3. The results of lipolysis in vivo confirm the higher stref3-susceptibility of Piitrains compared to Large Whites. Furthermore the standardized isolated fat cell studies in vitro showed in nearly all cases in the leaner Pietrains a higher ability of stimulating lipolysis with epinephrine.
The results are discussed. Resume
Taille des adipocytes, paramhtres lipoginiques et Iipolytiques dans le lard dorsal des porcs Large White et Piitrain
Pendant I’engraissement des porcs des races Large White et Piitrain les parametres suivants itaient determines dans le tissu adipeux dorsal et comparks: - I’accroissement des adipocytes - I’activiti des enzymes i NADPH (NADP-MDH, G6PDH, 6PGDH) pour I’indice de l’activiti
- I’activiti%polytique des adipocytes isolks La lipolyse en vivo itait determinee dans le plasma sanguin. 1. Nous avons constate que les Large Whites ont tout au long de I’engraissement des adipocytes plus
grandes que les Piitrains au fur et i mesure du degrk d’adipositk plus ilevi des Large Whites sous des conditions ad libitum. I1 est supposi que la taille aussi bien que le nombre des adipocytes sont responsables pour la masse adipeux augmentie.
lipogeni ue

Fettzellgrofie im Rcckenfettgewebe von Schweinen 289
2. Pour ce qui concerne I’activiti des enzymes i NADPH on a constati des diffirences entre les races au cours de la croissance, au maximum aussi bien que dans les classes de poids vif. Les Large Whites montraient en general une activiti lipoginique supirieure aux Piitrains.
3. Les resultats de lipolyse en vivo confirment une susceptibiliti particuliere aux stress des Piitrains compares aux Large Whites. Au cows des recherches en vitro avec des adipocytes isolies les Pietrains mai res montraient presque toujours une capaciti plus forte de la stimulation de I’activiti lipolytique par !adrenaline.
Les resultats sont discutis.
Resumen Tamako de las cilulas grasas, parametros lipbgenos y lipoliticos en el tejido adiposo dorsal
de cerdos Large Whi tey Piitrain En el tejido adiposo dorsal de cerdos Large White y Piitrain durante la ceba fueron registrados y comparados 10s si uientes parimetros: - crecimiento de i s cilulas grasas - actividad de las enzimas generadoras de NADPH (NADP-MDH, G6PHD, 6PGDH) como indice de
- actividad lipolitica de las cilulas grasas aisladas in vitro, con y sin estimulacion por adrenalina La lipolisis in vivo se determino en el plasma sanguineo. 1. Bajo condiciones de alimentacion a discrecion se encontraron en 10s cerdos Large White cilulas
grasas mayores que en 10s Piitrain durante toda la ceba. Se su one que tanto el tamaiio como el numero de cClulas grasas son responsables para un mis elevado Jp6s i to de grasa.
2. En lo referente a la actividad de las enzimas generadoras de NADPH se constataron diferencias entre las razas en el transcurso de la ceba, en el valor mhimo asi como entre las clases de peso. Los Large White en general mostraban una actividad lip6 ena superior a 10s Pibtrains.
3. Los resultados de la lipolisis in vivo confirman ?fa susceptibilidad mayor a1 stress de 10s Piitrains. Los examenes estandardizados de cilulas grasas aisladas en casi todos 10s momentos de medicidn com- probaron que la lipolisis en 10s Piitrains puede ser mis ficilmente estimulada por adrenalina.
la actividad lipogena
Se discuten 10s resultados.
Literatur
ALLEE, G. L.; ROMSOS, D.; LEVEILLE, G. A.; BAKER, D. H., 1971 : Influence of age on in vitro lipid biosynthesis and enzymatic activity in pig adipose tissue. P.S.E.B.M. 137,449-452.
ANDERSON, D. B.; KAUFFMAN, R. G., 1973: Cellular and enzymic changes in porcine adipose tissue during growth. J. Lipid Res. 14,160-168.
BUHLINGER, C. A.; WANGSNESS, P. J.; MARTIN, R. J.; ZIEGLER, J. H., 1978: Body composition, in vitro lipid metabolism and skeletal muscle characteristics in fast-growing, lean and slow-growing obese pigs at equal age and weight. Growth 42,225-236.
EGGSTEIN, M. ; KUHLMANN, E., 1970: Triglyceride und Glycerin. In: Methoden der enzymatischen Analyse, 2. Aufl., Bd. 11. Ed.: BERGMEYER, H. U., Weinheim: Verlag Chemie. S. 1765-1771.
ENSER, M.; MCFIE, H. J . ; MOODY, W. B.; RESTALL, D. J.; WOOD, J. D., 1976: The growth and cellularity of pig backfat. Anim. Prod. 22, 151.
FOLCH, J. ; LEES, M. ; STANLEY, G. H. S. , 1957: A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226,497-509.
GREENWOOD, M. R. C.; FOHNSON, P. R.; HIRSCH, J., 1970: Relationship of age and cellularity to metabolic activity in C57B mice. P.S.E.B.M. 133,944-947.
HARVEY, W. R., 1972: Instructions for use of LSMLMM least-squares and maximum likelihood general purpose programm 252 k mixed model version. Polycopy Ohio State Univ.
HIRSCH, J.; GALLIAN, E., 1968: Methods for the determination of adipose cell size in man and animals. J. Lipid Res. 9, 110-1 19.
HOOD, R. L.; ALLEN, C. E., 1973a: Comparative methods for the expression of enzyme data in porcine adipose tissue. Comp. Biochem. Physiol. 44,677-686.
HOOD, R. L.; ALLEN, C. E., 1973b: Lipogenic enzyme activity in adipose tissue during the growth of swine with different propensities to fatten. J. Nutr. 103, 353-362.
HOOD, R. L.; ALLEN, C. E., 1977: Cellularity of porcine adipose tissue: effects of growth and adipo- sity. J. Lipid Res. 18,275-284.
LEE, Y. B. ; KAUFFMAN, R. G . , 1974: Cellular and enzymic changes with animal growth in intramuscu- lar adipose tisue. J. Anim. Sci. 38,532-537.
LOWRY, 0. H.; ROSEBROUGH, N. J . ; FARR, A. L.; RANDALL, R. J., 1951: Protein measurement with the Folin-phenol-reagent. J. Biol. Chem. 193,265-275.
MARTIN, R. J. ; HERBEIN, J. H., 1976: A comparison of the enzyme levels and the in vitro-utilization of various substrates for lipogenesis in pair-fed lean and obese pigs. P.S.E.B.M. 151,231-235.

290 Ursula Rotbfufi
MERSMA", H. J.; PHINNEY, G.; BROWN, L. J., 1975: Ontogeny of epinephrine-induced lipolysis in adipose tissue from swine (sus domesticus). Gen. Pharmac. 6,187-191.
METZ, S. H. M., DEKKER, R. A., 1981: The contribution of fat metabolization to the regulation of fat deposition in growing Large White and Piitrain pigs. Anim. Prod. 33,149-157.
MOODY, W. G.; ENSER, M. B. ; WOOD, J. D.; RESTALL, D. J.; LISTER, D., 1978: Comparison of fat and muscle development in Pietrain and Large White piglets. J. Anim. Sci. 46,618-632.
O'HEA, E. K.; LEVEILLE, G. A,, 1969: Significance of adipose tissue and liver as sites of fatty acid synthesis in the pig and the efficiency of utilization of various substrates for lipogenesis. J. Nutr. 99,
RAESIDE, D. A.; WEEKES, T. E. C.; M; ELLIS, M.; SMITH, W. C., 1980: Influence of selection on lipogenic enzyme activity in adipose tissue of Large White pigs. Anim. Prod. 30,462.
ROGDAKIS, E., 1972: Untersuchun en iiber die Aktivitat von NADP-abhangigen Dehydrogenasen im Fettgewebe des Schweines als Iniex fur den Fettansatz. Diss. Hohenheim.
ROGDAKIS, E., 1973: Untersuchungen iiber die Beziehung zwischen der Aktivitat der NADP-abhangi- gen Dehydrogenasen im Fett ewebe und dem Fettansatz beim Schwein. Ziichtgskde. 45,412-420.
ROGDAKIS, E., 1974a: Untersucffungen iiber die Aktivitat NADPH-liefernder Enzyme im Fettgewebe des Schweines. 1. Mitt.: Biopsietechnik und Enzymteste sowie enzymatische Unterschiede zwischen verschiedenen anatomischen Stellen des Fettgewebes. Z. Tierphysiol., Tierernahrg. u. Futtermit- telkde. 33,329-338.
ROGDAKIS, E., 1974b: Einflui der Selektion nach der Riickenspeckdicke auf die Aktivitat der NADP- abhbgigen Dehydrogenasen im Fettgewebe des Schweines. Z. Tierziichtg. Zuchtungsbiol. 91,
ROGDAKIS, E.; ENSINGER, U.; FABER, H. v., 1979: Hormonspiegel im Plasma und Enzymaktivitaten im Fettgewebe von Piitrain- und Edelschwein. Z. Tierziichtg. Ziichtungsbiol. 96,108-1 19.
ROGDAKIS, E., 1982: Effekte der Selektion nach dem Spiegel der NADPH-liefernden Enzyme im Fettgewebe des Schweines. 1. Mitt.: Versuchsfrage, Versuchsanlage und erste Ergebnisse. Z. Tier- ziichtg. Ziichtungsbiol. 99,241-251.
ROTHFUSS, U., 1981: Lip0 ene und lipolytische Parameter im Fettgewebe von Large White- und Pietrain-Schweinen in Abf angigkeit vom Mastgewicht. Diss. Hohenheim.
ROTHFUSS, U., 1983: Morpholo ische und biochemische Parameter der Riickenfettgewebeschichten beim Schwein. Z. Tierpsyiol. jerernahrung u. Futtermittelkde. 50,263-269.
SALANS, L. B. ; KNI~TLE J. L.; HIRSCH, J., 1968: The role of adipose cell size and adipose tissue insulin sensivity in the carbohydrate intolerance of human obesity. J. Clin. Invest. 47,153-165.
SALANS, L. B.; DOUGHERTY, J. W., 1971: The effect of insulin and glucose metabolism by adipose cells of different size. J. Ch. Invest. 50, 1399-1410.
SCOTT, R. A.; CORNELIUS, S. G.; MERSMANN, H. J., 1981 : Effects of age on lipogenesis and lipolysis in lean and obese swine. J. Anim Sci. 52,505-51 1.
STANDAL, N. ; VOLD, E.; TRYGSTAD, 0.; Foss, I., 1973. Lipid mobilization in pigs selected for leanness or fatness. Anim. Prod. 16,37-42.
STAUFFACHER, W.; RENOLD, A. E., 1969: Effect of insulin in vivo on diaphragm and adipose tissue of obese mice. Am J. Physiol. 216,98-105.
STEELE, N. C.; FROBISH, L. T.; KENNEY, M., 1974: Lipogenesis and cellularity of adipose tissue from genetically lean and obese swine. J. Anim. Sci. 39,712-719.
STEELE, N. C.; FROBISH, L. T., 1976: Selected lipogenic enzyme activities of swine adipose tissue as influenced by genetic phenotype, age, feeding frequency and dietary energy source. Growth 40, 369-378.
STRUTZ, CH., 1977: Phenotypic and genetic parameters of NADPH-generating enzymes in porcine adipose tissue. Diss. Hohenheim.
SUTHERLAND, E. W.; DYE, I.; BUTCHER, R. W., 1965: The action of epinephrine and the role of the adenyl cyclase system in hormone action. Rec. Prog. Horm. Res. 21,623-646.
TRYGSTAD, 0.; Foss, I., 1967: Inhibition by human serum of the adipokinetic effect of a human pituitary lipid-mobilization factor (LMF) in rabbits. Acta Endocrin, 56,649-660.
TRYGSTAD, 0.; Foss, I.; VOLD, E., STANDAL, N., 1972: Suppressed lipolysis in genetically fat pigs. FEBS Letters 26,311-314.
WEISENBURG, C. L.; ALLEN, C. E., 1973: Adipose tissue metabolism in obese and lean pigs. J. Anim. Sci. 37,293.
WENIGER, J. H.; KALLWEIT, E.; STEINHAUF, D.; GLODEK, P., 1967: Einflui des Wachstums auf Teilstiickgewichte und Schlachtkorpermafle beim Schwein. Zuchtgskde. 39,188-200.
WOOD, J. D.; ENSER, M. B.; RESTALL, D. J., 1975: Fat cell size in Piitrain and Large White pigs. J. Agric. Sci. 84,221-225.
WOOD, J. D.; ENSER, M. B.; RESTALL, D. J., 1978: The cellularity of backfat in growing pigs and its relationship with carcass composition. Anim. Prod. 27,l-10.
Afithor's address: U. ROTHFUSS, Institut f. Tierhaltung und Tierziichtung der Universitat Hohenheim,
338-344.
75-87.
Garbenstr. 17, 7000 Stuttgart 70.