Embed Size (px)
Citation preview

(Aus dem Ins t i tuut Pasteur, Bandoeng, Java).
H A E M A G G L U T I N A T I O N D U R C H P O C K E N V I R U S
6. Ueber Inhibine aus normaler Lunge fiir die Vaceinevirus- Haemagglutination 1)
v o n
W. A. COLLIER (Eingegangen am 5 Apri l 1949).
Es ist bekannt, dass die Haemagglutination einzelner Virusarten dutch die verschiedensten Stoffe gehemmt werden kann. STONE and BURNET (15) fanden ein derartiges Inhibin in roher Kalbslymphe und sprachen ibm. die Natur eines Antistoffes zu, 5ahnlich den Serumantistoffen. Eine derartige Inhibinwirkung durch Extrakte aus normalen Organen wurde auch in diesem Institut beizahl- reichen Versuchen fiber die Haemagglutination durch Karbouwen- Vaccine gefunden. Am regelm~issigsten waren d.ie Hemmungswir- kungen bei Lungenextrakten. Da ffir die Pockeninfektion die porte d'entrde in den Atmungsorganen zu suchen ist, schien eine genauere Untersuchung der in der Lunge nachweisbaren Hemmungsstoffe ge- rechtfertigt. Von einem Inhibin muss man eine deutliche Affinit~it zu dem betreffenden Virus annehmen, die mindestens ebenso gross ist wie die Affinit~it tier benutzten Erythrocyten.
M e t h o d i k .
Benutzt wurde Vaccinevirus vom Karbouw, das 3 Tage nach cutaner Impfung geerntet war. Die Aufbewahrung erfolgte bei - -15 ~ C. Die sehr fein verriebene 10%ige Suspension wurde ca. 10 Minuten centrifugiert, und die fiberstehende Flfissigkeit ver- wendet.
Das Blut s tammt von javanischen Landhfihner, wurde 3 mal gewaschen und meist frisch benutzt, nur gelegentlich nach 24 Stunden. Es wurden nur Hfihner gebraucht, die eine sehr hohe Empfindlichkeit aufwiesen.
1) Mittei lung 5: Antonie van Leeuwenhoek 15, 53, 1949. 7

98 \V. A. Collier,
Die Lungen stammten von den verschiedensten Tieren, auch kamen einige Menschenlungen zur Untersuchung, welche von durchaus gesunden Personen stammten, die durch Unfall ums Leben gekommen waren. Die Versuche wurden sowohl w~ihrend der trocknen Zeit (J: 22 ~ C.) als auch w~ihrend der Regenzeit (_+_ 18 ~ C.) durchgefiihrt, wobei sich keine erkennbaren Unter- schiede zeigten.
Die Bindung zwischen Inhibin aus den Lungen und Vaccinevirus erfolgte 30 Minuten lang bei 37 ~ C. Nach Blutzufiigung blieben die R6hrchen bei Zimmertemperatur stehen, und die Ablesung konnte in der Regel nach ca. 1 �89 Stunden erfolgen.
Antisera gegen Inhibine wurden vom Kaninchen gewonnen. Benutzt wurden die schwachen javanischen Landkaninchen von ca. 1 Kilo Gewicht. Die Immunisierung erfolgte intravenbs mit 10 ~ Lungen-Suspension, die ca. 10 Minuten bei 2000 Umdrehungen zentrifugiert worden war. Von der tiberstehenden Fltissigkeit wurde je 0,5 oder 1,0 ccm an 6 aufeinander folgenden Tagen intraven6s injiziert. Die Blutentnahme erfolgte 8--10 Tage nach der letzten Injektion. Kaninchenlunge erwies sich als sehr toxisch, M~iuselunge wurde dagegen gut vertragen. Die Sera wurden im inaktiven Zustande untersucht. Es wurden nur Kaninchen benutzt, die vor der ImmunisieruI N keine hemmenden Serumstoffe fiir die Vaccine- virus-Haemagglutination aufwiesen und die ebensowenig die In- hibinwirkung yon Lungenextrakten hemmten.
EXPERIMENTELLER TEIL.
V i r u s a d s o r p t i o n d u r c h L u n g e n b r e i u n d H e m - m u n g d u r c h L u n g e n e x t r a k t . Von 5 bunten Ratten wurde je 1 g Lunge, Milz, Leber und Gehirn
im Mbrser sehr fein verrieben, in 2 ccm physiologischer Kochzalz- 16sung suspendiert und bei 2000 Umdrehungen centrifugiert. Die iiberstehende Fliissigkeit wurde zum Hemmungsversuch benutzt: Fallende Verdiinnungen hiervon wurden in der Menge yon 0,2 ccm mit 0,2 ccm der doppelten agglutinierenden Dosis Vaccine gemengt und 30 Minuten bei 37 ~ C. gehalten. Die Ergebnisse finden sich in Tabelle 1 zusammengestellt.
Der Niederschlag wurde noch einmal mit je 10 ccm physiologi- scher Kochsalzl6sung gewaschen und hiernach met 3 ccm Vaccine- virus 1/10 gemischt und 1 Stunde lang bei 37 ~ C. gehalten. Hierauf
Haemagglutinat ion durch Pockenvirus. 99
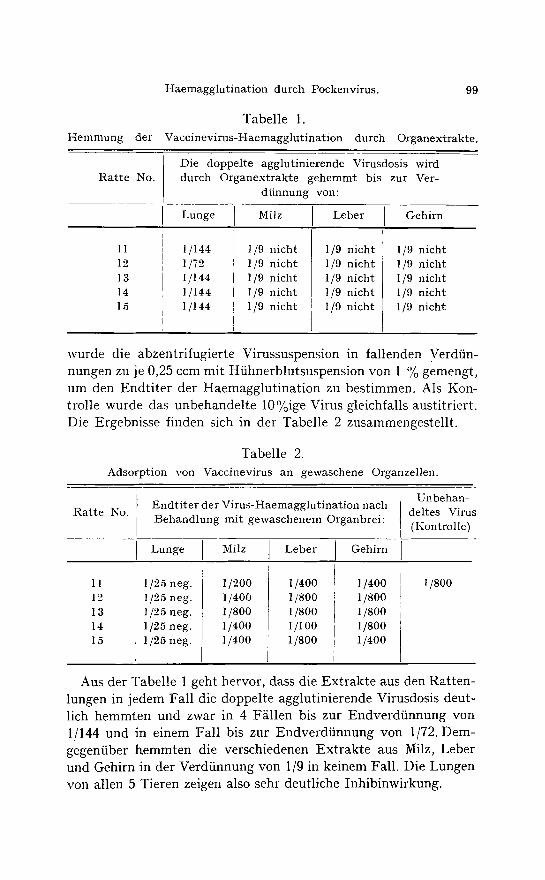
Tabelle 1. Hemmung der Vaccinevirus-Haemaggtut inat ion durch Organextrakte.
Die doppelte agglutinierende Virusdosis wird Ra t t e No. durch Organextrakte gehemmt bis zur Ver-
dfinnung yon:
Lunge Milz Leber Gehirn
1I 12 13 14 15
1/144 1/72 1/144 1/144 1/144
1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht
1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht
1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht 1/9 nicht
wurde die abzentrifugierte Virussuspension in fallenden Verdfin- nungen zu je 0,25 ccmmi t Htihnerblutsuspension von 1 % gemengt, um den Endti ter der Haemagglutination zu bestimmen. Als Kon- trolle wurde das unbehandelte 10%ige Virus gleichfalls austitriert. Die Ergebnisse finden sich in der Tabelle 2 zusammengestellt.
Tabelle 2. Adsorption yon Vaccinevirus an gewaschene Organzellen.
Unbehan- Ra t te No. E n d t i t e r d e r Virus-Haemaggtut inat ion nach deltes Virus
Behandlung mit gewaschenem Organbrei: (Kontrolle)
Lunge Milz Leber Gehirn
11 12 13 14 15
1/25 neg. 1/25 neg. 1/25 neg. 1/25 neg. 1/25 neg.
1/2oo 1/400 1/8oo 1/4o0 1/400
1/4oo 1/800 ]/800 1/loo ~/8oo
1/40o 1/8oo 1/800 U8oo 1/400
~/8oo
Aus der Tabelle 1 geht hervor, class die Extrakte aus den Ratten- lungen in jedem Fall die doppelte agglutinierende Virusdosis deut- lich hemmten und zwar in 4 F~llen bis zur Endverdfinnung von 1/144 und in einem Fall bis zur Endverdtinnung yon 1/72. Dem- gegenfiber hemmten die verschiedenen Extrakte aus Milz, Leber und Gehirn in der Verdfinnung von 1/9 in keinem Fall. Die Lungen von allen 5 Tieren zeigen also sehr deutliche Inhibinwirkung.

100 VC A. Collier,
Die F~ihigkeit der gewaschenen Organzellen, Vaccinevirus zu adsorbieren, geht aus der Tabelle 2 hervor. In der Kontrollereihe agglutinierte das unbehandelte Virus noch bis zur Verdiinnung yon 1/800. Demgegeniiber zeigte das Virus nach Adsorption mit Ratten- lungenbrei in keinem Fall Agglutination, selbst nicht in den st~irk- sten untersuchten Konzentrationen yon 1/25. Es hatte also eine starke Adsorption des Virus an die Lungenzellen stattgefunden. Bei den Adsorptionsversuchen mit anderen Organen hatte in diesem Versuch allerdings auch einmal die Milz den ursprtinglichen Virus Titer auf 1/4 gesenkt, und einmal die Leber auf 1/8.
Die Rattenlunge diirfte also im Vergleich mit den anderen Organen wohl die meisten virusaffihen Rezeptoren aufweisen. Diese sind nicht nur zellst~indig, sondern auch imstande, in den Gewebs- extrakt iiberzugehen.
Bei 3 Kaninchen erwiesen sich zwar die Lungenextrakte als stark hemmend, aber Extrakte aus Leber, Milz, Niere, Herz und Gehirn waren wirkungslos.
Etwas anders verhielten sich Meersehweinchen, da bier auch in anderen Organen nicht selten Inhibine nachgewiesen werden konnten. So zeigten beispielsweise von 12 untersuchten Tieren Inhibinwirkung gegen die doppelte agglutinierende Virusdosis:
10 Tiere im Extrakt der Lungen 6 Tiere im Extrakt der Nieren 5 Tiere im Extrakt der Milz
kein Tier im Extrakt der Leber.
Bei allen diesen 12 Meerschweinchen erwies sich das Blut sowohl im aktiven als aueh im inaktiven Zustande als vollkommen frei yon hemmenden Stoffen.
U.a. wurden auch zwei Affen (Macaca irus mordax) untersucht, die nach cerebraler Infektion mit Newcastle disease-Virus im mori- bunden Zustand get6tet waren. Eines der Tiere wies in Lunge und Niere reichlich Inhibine auf, in Leber und Herz wenig und in der Milz gar nicht. Der andere Affe hatte nur in Niere und Leber Inhibine, nicht dagegen in Lunge, Milz, Herzmuskel, Pancreas und Skelettnmskulatur. Das Serum des ersten Affen wirkte im aktiven Zustande bis zur Verdtinnung yon 1/20, das des anderen bis zu 1,/40 hemmend. Inaktiv waren beide Seren wirkungslos.
In den Organextrakten yon zahlreichen Meerschweinchen und Ratten, die gegen Vaccinevirus immunisiert waren, fanden sich
Haemagglutination durch Pockenvirus. 101
sehr h~ufig noch in relativ hohen Verdfinnungen hemmende Stoffe, die aber fast stets einigermassen mit den entsp~echenden Anti- Haemagglutininen des Serums a n St~rke fibereinstimmten und wohl mit diesen zu identifizieren waren.
W i r k s a m k e i t v e r s c h i e d e n e r L u n g e n e x t r a k t e .
Nicht bei allen untersuchten Tierarten fand sich die Wirkung der Lungenextrakte gleichm~issig ausgebildet.
Wirksam erwiesen sich stets in hohen Verdfinnungen die Extrakte aus Rinderlungen, die vom Schlachthaus bezogen wurden. Ferner waren alle 7 untersuchten Menschenlungen deutlich aktiv. Von l0 M~iusen zeigten 2 Inhibinwirkung auf die doppelte agglutinierende Virusdosis in der Verdiinnung von 1/400, 6 in der Verdtinnung von 1,,/200 und 2 in der Verdfinnung yon 1/100.
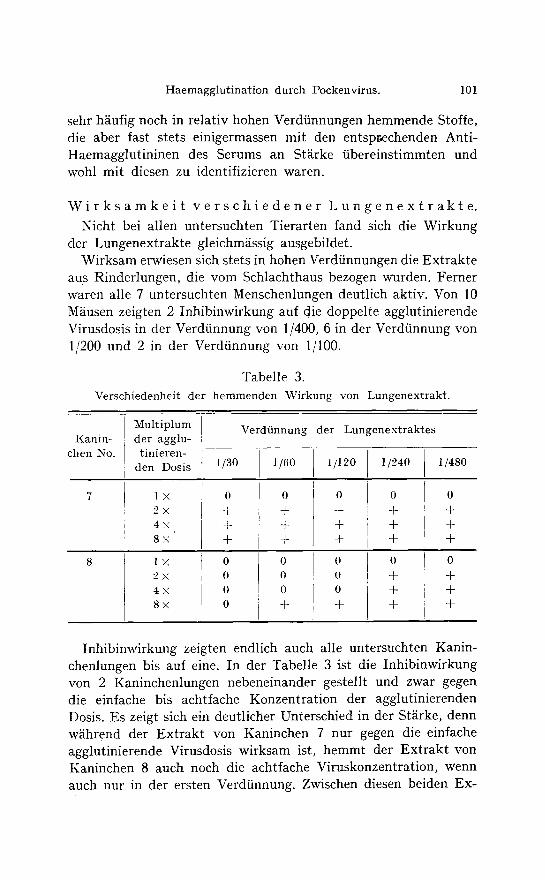
Tabelle 3. Verschiedenheit der hemmenden \u yon Lungenextrakt.
Kanin- chen No.
Multiplum der agglu- tinieren-
den Dosis
l x 2 x 4x 8X'
Verdfinnung der Lungenextraktes
1/30
o + + +
J/Go
0 + + +
1/120
0 + + +
1/240
0 + + +
lX 2 x 4 x 8 x
0 0 0 +
o o o +
0 + + +
1/480
o + + +
o + + +
Inhibinwirkung zeigten endlich auch alle untersuchten Kanin- chenlungen bis auf eine. In der Tabelle 3 ist die Inhibinwirkung von 2 Kaninchenlungen nebeneinander gestellt und zwar gegen die einfache bis achtfache Konzentration der agglutinierenden Dosis. Es zeigt sich ein deutlicher Unterschied in der St~irke, denn wiihrend der Extrakt von Kaninchen 7 nur gegen die einfache agglutinierende Virusdosis wirksam ist, hemmt der Extrakt von Kaninchen 8 auch noch die achtfache Viruskonzentration, wenn auch nur in der ersten Verdiinnung. Zwischen diesen beiden E x -

102 W . A . Collier,
tremen fanden sich bei anderen Tieren alle mSglichen Zwischen- stufen.
Die Lunge eines jungen Hundes enthielt viel Inhibin, die Lunge eines Reisvogels (Padda oryzivorus) sehr wenig.
Nicht regelm~issig fanden sick Inhibine in den Lungen yon schwarz-weissen Ratten und Meerschweinchen. Bei einer Meer- schweinchenserie waren beispielweise nur 10 yon 12 Tieren positiv, in einer anderen 16 yon 20. Es handelte sich hierbei stets um Tiere, die mit Diphtherie- oder Tetanustoxin vorbehandelt waren. Bei den Ratten waren mitunter alle Tiere positiv, gelegentlich aber fanden sich verschiedene Tiere ohne Inhibine in den Lungen. So hemmten beispielweise die Lungenextrakte yon 9 Tieren einer Serie in 4 F~illen bis 1/400 (deren Serum einmal bei 1/20 und dreimal nicht bei 1/5), in 2 F~illen bis 1/200 (deren Serum einmal bei 1/20 und einmal nicht bei 1/5), und in 3 F/illen nicht in der V'erdtinnung 1/25 (deren Serum einmal bei 1/10 und zweimal nicht bei 1/5) die doppelte agglutinierende Virusdosis.
T h e r m o l a b i l i t ~ i t d e r L u n g e n e x t r a k t e .
Wichtig erschien die Feststellung der Thermolabilit~it der In- hibine in den Lungenextrakten. Es zeigte sich, dass es keineswegs gleichgiiltig war, in welcher Konzentration die Extrakte erhitzt wurden. In Konzentrationen yon beispielsweise 1 : 4 wurde bei hSheren Temperaturen sehr viel Eiweiss ausgeflockt und riss anscheinend dabei wirksame Stoffe mit sodass infolgedessen kon- zentrierte Extrakte schneller inaktiviert wurden als verdtinntere. Als brauchbar erwies sich eine Lungensuspension von 1/8--1/10.
Tabelle 4. Thermolabili t~tt yon Rinder lunge: H e m m u n g der Haemagglu t ina t ion .
Elaviir- H e m m u n g der Agglu t ina t ion nach Erw~irmung yon mung auf: 0 10' 20' 30' 1 h 2 h
nat iv: 45 ~ C. 50 ~ C. 55 ~ C. 65 ~ C. 70 ~ C. 80 ~ C.
11200; - -
1/800 1/4oo 1/lO0
11/2oo /1200
i1/25 nicht
b
1/8oo 1/4oo 1/5o 1/5o 1/2oo 1/25 nicht
1/400 1/800 1/25 nicht 1/25 n ich t 1/25 n ich t 1/25 nicht
1/200 1/100 1/25 n ich t 1/25 n ich t 1/25 n ich t 1/25 n ich t
m
1/4oo 1/2oo 1/25 n ich t

Haemagglut inat ion dutch Pockenvirus. 103
In Tabelle 4 findet sich eine Versuchsserie wiedergegeben, in welcher Rinderlungenextrakt in der Verdiinnung l/8,33 verschieden lange auf Temperaturen yon 45 ~ bis 80 ~ C. erw~rmt wurde, 45 ~ und 50 ~ C. erh6hten bei kurzer Einwirkung zun~chst die hemmende Wirkung des Extraktes, dann aber trat nach l Stunde wieder ein Rtickgang ein. Bei Erw~rmen auf 55 ~ 65 ~ und 70 ~ C. waren die Ext rakte nach 30 minuten unwirksam geworden, wtihrend Er- w~rmen auf 80 ~ C. schon in 10 Minuten inaktivierte.
W~hrend in diesem Versuch der Ex t rak t bereits nach 30 Minuten bei 55 ~ C. inaktiv wurde, zeigte sich bei anderen Extrakten von Rinderlunge eine Inaktivierung erst bei 63 ~ C. Dies war aber die Temperatur, bei der Rinderlungenextrakte nach 30 Minuten stets wirkunKslos wurden.
Analog reagierten die Ext rakte aus den Lungen yon weissen M~usen, yon schwarz-weissen Rat ten von Hund und Affe: Bei 60--63 ~ C. wurde jegliche Inhibinwirkung nach 30 Minuten zer- st6rt.
Auch die Inhibine in Kaninchenlungenextrakt wurden bei 60--63 ~ C. zerst6rt, doch zeigte sich einmal, dass der Lungen- extrakt eines Tiere bei 60 ~ C. noch nicht v611ig zerstSrt war.
Lungenextrakte von Meerschweinchen schienen noch empfind- licher zu sein, denn in 7 Versuchen wurde die Inhibinwirkung schon nach 30 Minuten Erwttrmen auf 56 ~ C. aufgehoben und in 12 Versuchen nach 15 Minuten Erw~rmen auf 63 ~ C. Auch die Wirkung yon Reisvogel-Lunge wurde nach 30 Minuten Erw~rmen auf 56 ~ C. aufgehoben.
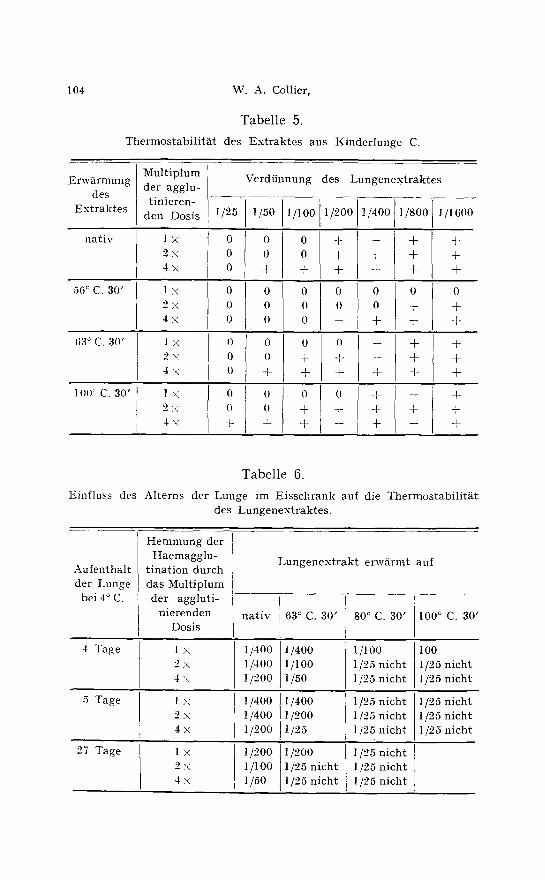
Ganz anders verhielten sich in jedem Fall Ext rakte aus frischen Menschenlungen, gleichgtiltig ob sie yon ebengeborenen Kindern oder yon Erwachsenen stammten. In der Tabelle 5 ist ein Versuch mit dem Ext rakt von Kinderlunge C gegen die einfache, doppelte und vierfache agglutinierende Virusdosis nach Erwtirmen auf 56 ~ 63 ~ und 100 ~ C. zusammengestellt. Erw~rmen auf 56 ~ C. steigerte die Wirkung, 63 ~ und ebenso 80 ~ C. (in der Tabelle nicht aufge- nommen) war ohne jeden Einfluss, und erst 30 Minuten Erwttrmen auf 100 ~ C. schw~chte etwas ab. Doch war hier noch deutliche Inhibinwirkung zu erkennen.
W~hrend also alle anderen untersuchten Inhibine deutlich ther- molabil waren, zeigte sich bei Menschenlunge eine ausgesprochene Thermostabilit~t. Es wurde bald deutlich, dass diese Thermo- stabilit~t nur bei frischen Menschenlungen nachweisbar war. Je
104 W. A. Collier,
T a b e l l e 5.
Thermostabilit~t des Extraktes aus Kinderlunge C.
t~rw~trmung des
Extraktes
nat iv
56 ~ C. 30'
Multiplum der agglu- t inieren-
den Dosis
I x 2 x 4 x
1x 2 x 4 x
i /25
Verdtinnung des Lungenextraktes
1 /5o 11100
0 0 0 0 + +
1/200 1/40o
+ + + + + +
0 0 0 0 + +
l /8oo
+ + +
0 + +
0 0 0 0 0 0
1/1600
+ + +
0 + +
tJ3 c C. 30'
10W C. 3(1'
l x 2 x 4 x
0 0 +
0 + +
0 + +
+ + +
1x 2 ~., 4X
0 o
0 o +
0 + +
0 + +
+ + +
+ + +
+ + +
+ + +
+ + +
T a b e l l e 6.
Einfluss des Alterns der Lunge im Eisschrank auf die Thermostabilitfit des Lungenextraktes.
Aufenthalt der Lunge
bei 4 ~ C.
Hemmung der Haemagglu-
t inat ion durch das Multiplum der aggluti-
nierenden Dosis
nat iv
Lungenextrakt erw~irmt auf
63 ~ 30 ' 80 ~ 30 ' 100 ~ C. 30'
4 Tage l x 2• 4 x
1/4oo 1/4oo 1/200
1/400 1/100 1/5o
l / l o o l/25 nicht l /25 nicht
I00 1/25 nicht 1/25 nicht
5 Tage
27 Tage
1x 2 x 4x
1 x 2• 4•
1/400 1/400 1/2oo
i 1200 1/100 1/5o
1/400 l /200 1/25
1/20o 1/25 nicht 1/25 nicht
]/25 nicht 1/25 nicht l/25 nicht
!/25 nicht l/25 nicht l/25 nicht
1/25 nicht 1/25 nicht 1/25 nicht

Haemagglutination durch Pockenvirus. 105
l~inger die Lunge im Eisschrank bei 4 ~ C. oder im Frigolo bei - -15 ~ C. bewahrt wurde, je mehr lief die Thermostabilit~it zurfick, um schliesslich vollkommen zu verschwinden. Ein Beispiel hierffir ist in Tabelle 6 gegeben. Nach einer Aufbewahrungszeit von 27 Tagen bei - -15 ~ C. waren die Inhibine in der Mensctlenlunge genau so thermolabil, wie die untersuchten Inhibine aus den verschiede- nen Tierlungen. Da kaum vorzustellen ist, dass ein und dieselbe Substanz yon Thermostabilit~it nach Thermolabilit~it umschlagen sollte, geht man wohl nicht fehl, wenn mann in der Menschenlunge z w e i v e r s c h i e d e n e Inhibine annimt: Ein thermostabiles, aber nut kurz haltbares, und ein thermolabiles, aber lange halt- bares Inhibin.
In verschiedenen F~illen konnte man die Beobachtung machen, dass die in toto bei 4 ~ C. aufbewahrte Lunge nach einigen Tagen in den daraus bereiteten Extrakten st~irkere Inhibinwirkung zeigte als im frischen Zustand. Ein Beispiel hierffir findet sich in Tabelle 7. Spiiter nahm die Wirkung Wieder etwas ab.
Tabelle 7. Einfluss des Altern's auf die hemmende Wirksamkeit yon Lungenextrakt.
Cavialunge
frisch
Multiplum der agglu- tinieren-
den Dosis
1 x 2 • 4 • 8 •
Agglutination bei Verdfinnflng des Lungen- extraktes
1/12o 1/24o
0 0 0 0 + + + +
1/3o 1/6o
0 0 0 0 + + + +
1/48o
0 0 + +
48 h 4 ~ C. ] x 2 x 4 • 8 •
0 0 0 0
0 0 0 +
0 0 0 +
H a l t b a r k e i t d e r L u n g e n e x t r a k t e .
Die Haltbarkeit der Lungenextrakte war nicht besonders gut. Aus diesem Grunde wurden sie stets frisch hergestellt. Bei 4 ~ C.
war Rinderlungenextrakt nach 3 Tagen zwar manchmal noch gut wirksam, nach 8 Tagen aber schon sehr deutlich schw~icher und nach 20 Tagen vollkommen wirkungslos. Bei - - 1 5 ~ C. war der

11)6 W.A. Collier,
gleiche Extrakt bis zu 8 Tagen unver/indert, nach 20 Tagen abei erwies auch er sich als fast unwirksam.
B i n d u n g s f ~ i h i g k e i t d e s L u n g e n e x t r a k t e s .
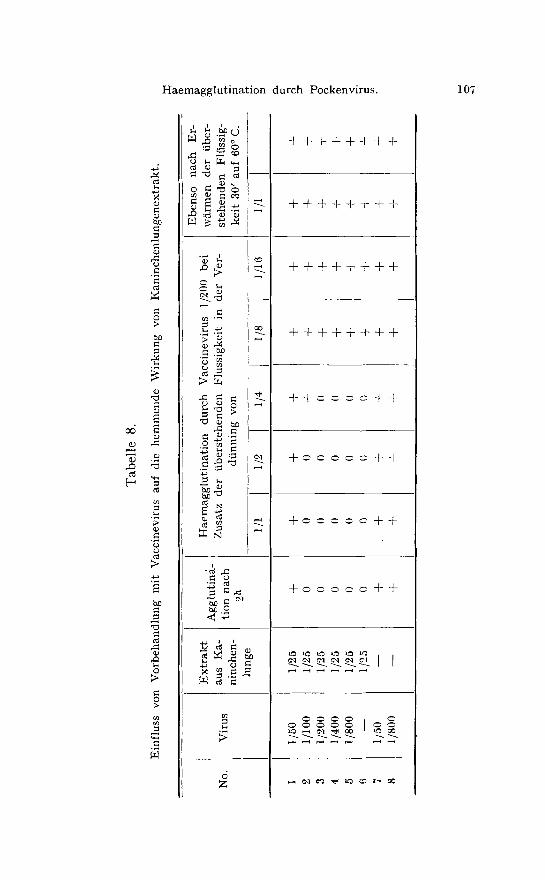
Wird st/irker Lungenextrakt mit Vaccinevirus gemengt, so wird das gebundene Virus ftir Haemagglutination ungeeignet, nicht ver- brauchtes Inhibin aber bleibt in seiner Wirkungsf~ihigkeit erhalten. Als Beispiel sei folgender Versuch wiedergegeben: Fallende Mengen yon Virus wurden mit der konstanten Menge Kaninchenlungen- extrakt 1/25 gemengt und 2 Stunden lang stehen gelassen. Wie aus Tabelle 8 hervorgeht, war die hohe Virusdosis yon 1/50 im ersten R6hrchen noch imstande zu agglutinieren, w~ihrend die Virusdosen 1:100 bis 1/800 (Grenzdosis) nach 2 Stunden infolge Inhibinwirkung keine Agglutination zeigten. Von den 3 Kontrollr6hrchen agglu- tinierte die Kaninchenlunge allein nicht, wohl aber die beiden Viruskontrollen ohne Lungenextrakt. Die tiberstehende Fliissigkeit wurde nunmehr abgegossen und auf noch vorhandene hemmende Wirkung in fallenden Dosen gegen eine Virusverdtinnung von 1200 untersucht. Die Fltissigkeit des ersten RShrchens hemmte nicht, da alles Inhibin durch die hohe Virusdosis gebunden war. Die i:ltissigkeit des 2. RShrchen hemmte bis zur Verdtinnung 1/2, aber die Fltissigkeiten aus dem 3.-5.R6hrchen hemmten die Hae- magglutination genau so stark, wie die reineLungenextraktkon- trolle. Die tiberstehende Fltissigkeit aus dem Viruskontroll-R6hr- chen hemmte nicht. Dass es sich in der Tat um die gew6hnliche Inhibinwirkung handelte, wurde durch die vollkommen fehlende Hemmung der auf 60 ~ C. erhitzten iiberstehenden Fliissigkeiten bewiesen.
I n h i b i n w i r k u n g v o n E x t r a k t e n n i c h t n o r - m a l e r L u n g e n .
Weiterhin wurde untersucht, wie sich die Inhibinwirkung ver- hielt, wenn die Extrakte nicht aus normalen, sondern aus Lungen stammten, die mit Keuchhustenbacillen, Pestbacillen oder Vaccine- virus infiziert waren.
14 Meerschweinchen wurden mit 10 Milliarden Pestbaci l len (apathogener Stamm Tjiwidej) in 0,5 ccm phys. Kochsalzl6sung in leichter Aethernarkose pulmonal infiziert. 4 Tiere wurden nach 24 Stunden, 3 nach 48 Stunden, 3 nach 72 Stunden und 3 nach 120
Ta
be
lle
8.
Ein
flu
ss v
on
Vo
rbeh
and
lun
g m
it
Vac
cin
evir
us
auf
die
hem
men
de
\Vir
ku
ng
vo
n
Kan
inch
enlu
ng
enex
~tr
akt.
r
No.
V
iru
s
l/50
l/lo
o
1120
0 1/
400
1/8o
o
1/5o
1/
8oo
Ex
trak
t au
s K
a-
nin
chen
- lu
ng
e
U'2
5 1/
25
1/25
1/
25
1/25
Ag
glu
tin
a-
tio
n n
ach
+ o o o o 0 +
+
Hae
mag
glu
tin
atio
n
du
rch
Z
usa
tz
der
fib
erst
ehen
den
d
fin
nin
g y
on
ill
1/2 + o o o o +
+
1/4 +
+ 0 0 0 +
+
Vac
cin
evir
us
1/20
0 be
i F
luss
igk
eit
in
der
V
er-
Eb
enso
n
ach
E
r-
w~
irm
en
der
fi
ber-
st
ehen
den
F
lfis
sig-
k
eit
30
' au
f 60
~ C
.
1/1 +
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
2 k
§ o o o (}
o + +
1/8
1/1(
~
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
0q
5 < 7~

108 \V. A. Collier,
Stunden untersucht. Alle wiesen deutlich Inhibinwirkung auf, analog entsprechenden Kontroll-Meerschweinchen.
Ebensowenig Einfluss hatte auch die pulmonale Infektion yon Meerschweinchen mit je 10 Milliarden Keuchhustenbacillen.
Interessant sind auch die Ergebnisse nach Infektion mit Vaccine- virus, wobei 0,5 ccm einer Virussuspension von 1/10 unter Aether- narkose pulmonal instilliert wurde. Die Ergebnisse sind in Tabelle 9 zusammengestellt. In diesem Versuch wurde vorher untersucht, ob mittels der Haemagglutinationsmethode in den Lungenextrakten Vaccinevirus nachweisbar war. Dies war bei einem Tier der Fall, das nach 2 Tagen, und ausserdem bei 2 Tieren, die beide am 5. Tage get6tet wurden. Alle anderen Lungenextrakte waren nicht imstande Hiihnerblut zu agglutinieren. Dagegen wiesen alle diese Lungen eindeutig hemmende Stoffe auf und zwar die am 1. und 2. Tage untersuchten etwas weniger stark als die sp~iter untersuchten.
Tabelle 9. Hemmung der Haemagglutination durch Lungenextrakt und Virusnachweis in der Lunge mittels Haemagglutination bei pulmonal mit Vaccinevirus
infizierten Meerschweinchen (0,5 ccm 1/10).
Tier No.
1 2 3 4 5 i; 7 S 9
lO 11 12 13 14
Untersucht nach Tagen
Hemmung der I
doppelten vierfachen
1 1 1 2 2 2 3 3 5 5 9
10 10 10
Virusdosis Virusdosis
1/100 1/;~o 1/5(.) )/loo ! / 1 0 0
1/200 ]/200
1/400 I /200 ]/soo 112oo
1/25 1/25 I/25 1/25 ]/25
1/100 1120(I
1/200 l/! oo ]/25 ]/5()
Haemagglut inat ion durch Lungen-
extrakt
neg. neg. neg. neg. neg. 1/5o neg. neg. 1/400 1/400 neg. neg. neg. neg.
Aus den hier mitgeteilten Versuchen geht hervor dass die In- hibinwirkung aueh vorhanden ist, wenn die Lungen dureh nicht

Haemagglutination durch Pockenvirus. 109
t6dliche Infektion mit Bakterien oder durch Vaccinevirus selbst beeinflusst sind.
E i n f l u s s d e r I n h i b i n e a u f d i e I n f e k t i o s i t / i t y o n V a c c i n e v i r u s .
Eine Suspension yon Vaccinevirus 1/10 wurde 10 Minuten lang bei 2000 Umdrehungen zentrifugiert und die tiberstehende Flfissig- keit wurde beginnend mit 1/500 in fallenden Verd/innungen mit der konstanten Menge 1/25 einer Suspension aus Rinderlunge ver- dtinnt. Eine gleiche Virus-Verdtinnungsreihe wurde mit physiolo- gischer Kochsalzl6sung als Kontrolle verdtinnt. Beide Versuchs- reihen wurden 2 Stunden lang bei 37 ~ C. bewahrt und hierauf nach der GINsschen Methodik am skarificierten Meerschweinchenauge geprtift. Am gleichen Meerschweinchen wurde rechts Virus _a Inhibin und links die entsprechende Kontrolle untersucht, aus- serdem wurden ffir jede Verdiinnung je 2 Meerschweinchen ge- braucht. Der gleiche Versuch wurde wiederholt, wobei das Gemisch Virus + Lungenextrakt und die Kontrollreihe erst 2 Stunden bei 37 ~ C., dann 18 Stunden bei 4 ~ C. und noch einmal 1 Stunde bei 37 ~ C. bewahrt wurden. In beiden Versuchserien ergab die Virus- verdtinnung 1/100.000 mit und ohne Lungenextrakt ein positives Impfergebnis. Bei einem Tier war ausserdem die Kontrollimpfung mit 1/200.000 schwach positiv.
In genau der gleichen Weise verliefen Versuche, wobei Kaninchen mit 0,1 ccm Virus + 1/5 Lungenextrakt intracutan injiziert wumen. Virus + Lungenextrakt und Virus allein ergaben genau die gleichen Werte: Die Konzentrationen von l/l.000.000 und 1/'500.000 waren in jedem Falle gleichm~issig positiv.
Aus diesen Versuchen geht eindeutig hervor, dass die Inhibine keinerlei Einfluss auf die Infektiosit~it des Vaccinevirus auszutiben imstande sind. Diese Schlussfolgerung ist zweifellos richtig, wenn uns auch im Verlauf yon anderen Versuchen deutlich geworden ist, dass Bindungsversuche zwischen Vaccinevirus und Antistoffen sowohl im Meerschweinchenversuch (Skarifikation) als auch im Kaninchenversuch (i. cut.) nur mit der allergr6ssten Vorsicht zu bewerten sind. Im Meerschweinchenversuch kann die Streuung sehr gross sein, und man hat mit Minimalwerten zu tun. Dies ist zwar bei der Kontrolle yon Vaccine keine Hinderniss, wohl aber bei Neutralisierungsversuchen. Dagegen sind die kleinen und schwachen javanischen Kaninchen infolge unverhiiltnism~issig
110 \V. A. Collier,
grosser Unterscliiede in ihrer Reaktionsf~ihigkeit v611ig unbrauch- bar.
A u f h e b u n g d e r I n h i b i n w i r k u n g d u r c h L u n - g e n a n t i s e r e n .
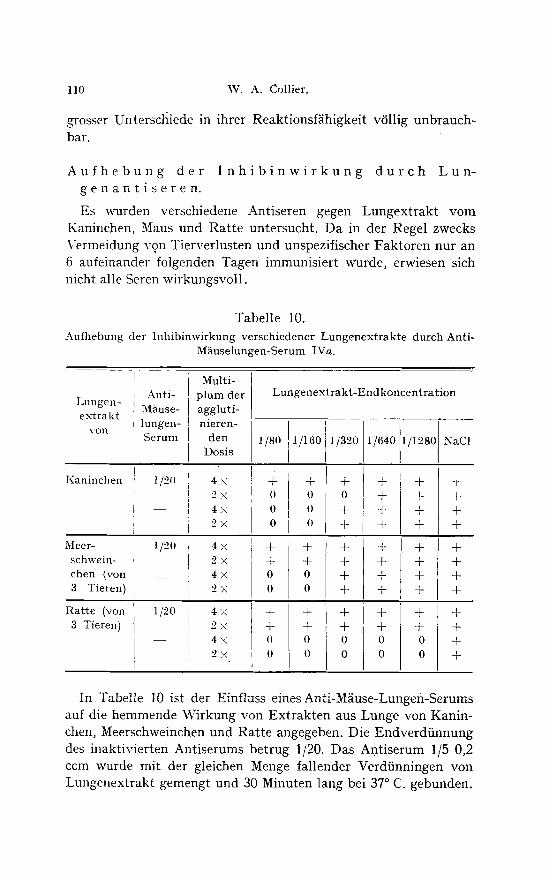
Es ~a~rden verschiedene Antiseren gegen Lungextrakt vom Kaninchen, Maus und Ratte untersucht. Da in der Regel zwecks Vermeidung yon Tierverlusten und unspezifischer Faktoren nur an 6 aufeinander folgenden Tagen immunisiert wurde, erwiesen sich nicht alle Seren wirkungsvoll.
Tabelle 10. Aufhebung der Inhibinwirkung verschiedener Lungenextrakte durch Anti-
M~iuselungen-Serum IVa.
Anti- Lungen- M~iuse- extrakt
lungen- yon
Serum
Kaninchen
Meer- schwein- chen (von 3 Tieren)
Ratte (yon 3 Tieren)
t /2o
1/2 o
/2o
Multi- plum der aggluti- nleren-
den Dosis
4 x 2• 4• 2",<
Lungenextrakt-End koncentration
1/32o
4 x 2 x 4• 2•
1/64o 1/128o J/so 1/16o
t
-~ I + + + 0 0 0 + I
0 I o + + o o + +
+ + + + + + + + o o + + o o + +
+ + + +
+ + + +
+
+o o
4X 2 x 4• 2 x
+ + + + + + + +
O 1~ 0 0 o o o
NaC1
+ + + +
+ + + +
+ + + +
In Tabelle 10 ist der Einfluss eines Anti-M~iuse-Lungen-Serums auf die hemmende Wirkung von Extrakten aus Lunge von Kanin- chen, Meerschweinehen und Ratte angegeben. Die Endverdiinnung des inaktivierten Antiserums betrug 1/20. Das Antiserum 1/5 0,2 ccm wurde mit der gleichen Menge fallender Verdiinningen von Lungenextrakt gemengt und 30 Minuten lang bei 37 ~ C. gebunden.
Haemagglut/nation durch Pockenvirus. 111
Eine Verdtinnungsreihe von Lungenextrakt mit phys. Kochsalz- 16sung diente als Kontrolle. Hierzu wurden je 0,2 ccm Vaccinevirus gefiigt und nach 30 Minuten bei 22 ~ C. je 0,2 ecm 2 % Hfihner- blutsuspension. Aus den Kochsalzkontrollen ist zu ersehen, dass dieses Antiserum keine eigene Hemmung auf die Virus-Haemag- glutination ausiibte. Dagegen wurde die Hemmungswirkung des Kaninchenlungenextraktes teilweise und die der Lungenextrakte yon Meerschweinchen und Ratte vollkommen aufgehoben. Da dieses Anti-Miiuse-Lungen-Serum auch gegen die Inhibine aus den Lungen anderer Tierarten wirksam war, handelt es sich bei dessen neutralisierender Wirkung sicher nicht um speeies-specifisehe Antistoffe, vielmehr um echte Antistoffe gegen Inhibine. Diese scheinen demnach bei den verschiedenen Tierspecies wenn auch nicht identisch, so doch mindestens miteinander verwandt zu sein.
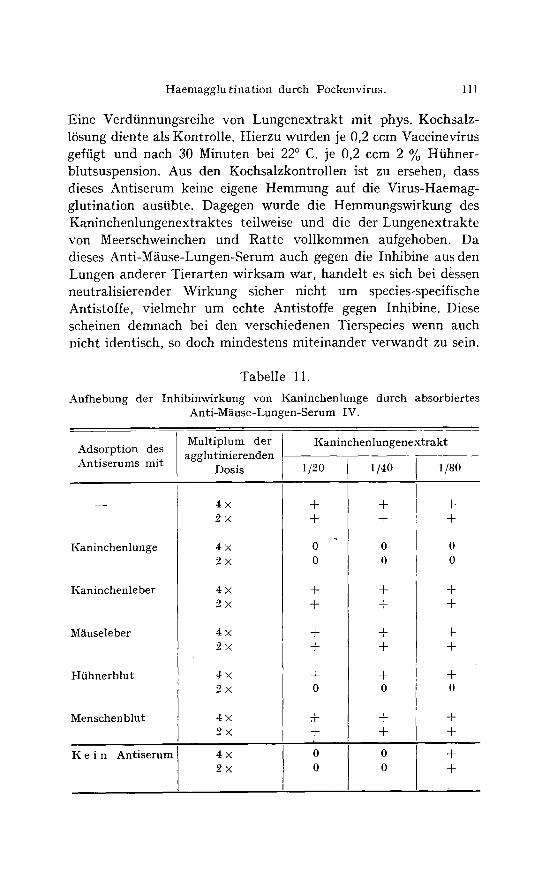
Tabelle 11.
Aufhebung der Inhibinwirkung yon Kaninchenlunge durch absorbiertes Anti-M~iuse-Lungen-Serum IV.
Adsorption des Antiserums mit
I ( a n i n c h e n l u n g e
Kaninchenleber
M~use leber
Hfihnerblut
M e n s c h e n b l u t
I ( e i n A n t i s e r u m
Multiplum der K a n i n c h e n l u n g e n e x t r a k t
1/40 agglutinierenden
Dosis
+
+ +
+ +
+ 0
1/20
x + x +
x o
x o
x + x +
x +
x +
x + x o
x .+
x + T
x o x o
i/8o
+ +
+ +
+ +
+ 0
4 + + 2 + +
4 0 + �9 2 0 +

t12 W . A . Collier,
I)a manche Antiseren von den verschiedenen Tierar ten spontane
Agglutinine gegen Hfihnerblut enthal ten, die durch Adsorpt ion
an Hfihnerblut und oftmals auch an Hammelb lu t entfernt werden kSnnen, wurden die gewonnenen Antiseren meistens erst nach
erfolgter Adsorpt ion untersucht . Daneben wurden auch Adsorp-
tionen an andere Organsuspensionen untersucht , wovon folgendes Beispiel in Tabelle 11 gegeben sei.
Inakt ivier tes Anti-M~iuse-Lungen-Serum IV wurde in der Ver-
dfinnung yon 1/5 mit je 1 g Kaninchenlunge, Kaninchenleber , M~iuseleber oder je 1 ccm gewaschenen conc. Ht ihnenblutk6rperchen
oder Menschenblutk6rperchen gemengt , 1 Stunde bei 37 ~ C. ge-
halten und danach abzentrifugiert . Je 0,25 ccm wurde mit 0,25 ccm fallender Konzentra t ionen yon Kaninchenlungenext rak t gemengt
Tabelle 12.
Atlfhebullg der Inhibinwirkung yon Rinderhmge durch verschiedene Konzentrationen absorbierten Anti-M~iuse-Lungen-Serums 119.
Antiserum absorbiert
lint
Endver- dfinnung des An- tiserums
Hfilmerblut 1/20
Hammelblut
1 /40
!/8()
1/2 o
l / 4 0
Multi- plum der aggluti- nleren-
den Dosm
4 )<
o x
4 >: '2 X
4 x 2 x
4x 2 >"
Rinderlungenextrakt-Endver- dfinnung
1/200 1/400 NaC1
+ +
+ +
+ +
4 ).~ 2 X
7 /25 1 /5o l/lO0
+ + + + 0 + + +
0 ~- + , - f -
o 0 + +
(I 0 o + o 0 0 o
+ + + + + + -- T +
0 0 + + o o o +
0 0 0 + 0 0 0 +
+ +
§ +
+ +
+ +
+ +
+ +
+ +
1 /80 4 • 2 •
K e i 11 4 • 0 0 0 + + + Antiserum 2 • 0" 0 0 0 + +

Haemagglutination .durch Pockenvirus. 113
und 30 Minuten lang bei 37 ~ C. gehalten. Hierauf wurden 0,25 ccm der zweifachen und vierfachen wirksamen Virusverdiinnungen zugeftigt und das Ganze wieder 30 Minuten lang bei 22 ~ C. gehalten. Zum Schluss wurde 0,25 2%ige Hiihnerblutsuspension beigegeben. Das nicht adsorbierte Antiserum hob die Inhibinwirkung auf. Unver~indert war das Serum auch durch Vorbehandlung mit Kaninchenleber, M~iuseleber und Mensehenblut. Htihnerblut hob die neutralisierende Wirkung teilweise auf und Kaninchenlunge vollkommen. In beiden F~illen handelt es sich nicht um eine wirk- liche Aufhebung der Antiserumwirkung. Htihnerblut enth~ilt viel- fach ein schwach wirkendes Inhibin, und in der Kaninchenlunge war sehr viel Inhibin enthalten, sodass die Antiserumwirkung durch die bei der Vorbehandlung dazugekommene Inhibinmenge abgeschw~icht bzw. g~inzlich aufgehoben wurde.
Schliesslich ist in Tabelle 12 noch ein Beispiel gegeben, wie ein Anti-Mtiuse-Lungen-Serum nach Adsorption an Htihner- und Ham- melblut austitriert wurde. In den Verdiinnungen von 1/20 war der Serumeinfluss sehr deutlich zu sehen, auch wirkte das Hiihnerblut etwas abschw~ichend, im Vergleich mit dem Hammelblut. In beiden F~illen war die Verdiinnung von 1/40 schon viel schwiicher, und die Verdtinnung yon 1/80 wirkte ungef~ihr wie die nicht Serum ent- haltenden Kontrollen des Lungenextraktes. Die Antiserumwirkung ist also zwar deutlich feststellbar, die benutzten Seren waren aber doch relativ schw~ich.
B e s p r e c h u n g d e r E r g e b n i s s e .
Bei der Untersuchung der Inhibinwirkung auf die Haemagglu- tination durch verschiedene Virusarten sind durch BEVERIDGE und LIND (1), BOVARNICK und DE BURGH (2), DE BURGH, YU, HOWE und BOVARNICK (3), BURNET und BEVERIDGE (4), BURNET, MCCREA und ANDERSON (5), FRIEDEWALD, MILLER und WHATLEY (8), GREEN and WOOLEY (10), HENLE, HENLE und HARRIS (12), STONE und BURNET (15), SVEDMYR (16) u.a. die verschiedensten Extrakte und Stoffe als wirksam gefunden. Wichtig sind in diesem Zusam- menhang vor allem die Hemmungen der Influenza- und Mumps- Haemaggh~tination durch Extrakte aus menschlicher Lunge, Leber, Niere und Milz und durch Organextrakte v0n normalen Kaninchen und Meerschweinchen. Die Wirksamkeit derartiger Organextrakte wurde durch Erw~irmen auf 65 ~ C. reduziert.
Inhibinwirkung gegen Vaccinevirus wurde bisher nur in roher
8

I 14 ~,u A. -CoIIier,
K~Iberlymphe durch STONE und BURNET (15) nachgewiesen. Wenn bisher auch noch nicht viel fiber die Natfir der Inhibine
bekannt ist, so bekommt man doch den Eindruck, dass hier zwei verschiedenen TypeD vorliegen. Die Inhibine der einen Gruppe, die m6glicherweise ffir den Infektionsmechanismus wichtig sind und als Virus-Acceptoren angesehen werden k6nnen, stellen an- scheinend einen normalen Bestandteil mit bestimmten Funktionen im Lebensablauf bestimmter Zellen dar. In dieser Eigenschaff muss ein solcher Acceptor mindestens zum grosseD Tell aus Protein- substanzen bestehen und ist jedenfalls wegen dieser seiner Eiweiss- natur auch mehr oder weniger ausgesprochen thermolabil. Eine zweite Gruppe von Inhibinen hat jedenfalls einen anderen Aufbau, vorwiegend yon Nicht-Eiweisstruktur, wodurch seine Thermosta- bilit/it zu erkl~iren w/ire. Hierher wfirde beispielsweise der von SVEDMYR (16) in normaler Allantoisflfissigkeit nachgewiesene In- hibitor gegen Grippevirus geh6ren, der erst nach 1 Stunde Kochen eine gewisse Abschw~iehung erf~ihrt. Auch der in den vorliegenden Versuchen in Menschenlunge gefundene thermostabile Inhibin- faktor darf hierher gerechnet werden, wie auch die verschiedenen Inhibitoren der Influenze-Haemagglutination (Flachssaat-Gummi, Pectine, Akazien-Gummi usw.) von GREEN und WOOLEu (10) und die ebenso wirkende Blutgruppen A- und O-Substanz nach BURNET, MCCREA und ANDERSON (5).
Man darf annehmen, dass ein in Organextrakten nachweisbarer Inhibitor, der den Viruspartikelchen die Bindung an empf/ingliche Erythrozyten m6glich macht, auch im lebenden Organismus vor- handeD ist und in vivo besondere virusaffine Eigenschaften besitzt. In Wirklichkeit liegt also ein Virus-Acceptor vor, der das in den K6rper eingedrungene Virus bindet.
Diese Verankerung an die Acceptoren der Lunge kann keines- wegs den Zweck haben, das Virus unsch/idlich zu roacheD. Die Acceptoren haben nichts zu tun mit Serum-Antistoffen, sondern wirken vollkommen unabh~ingig von diesen. In vaccineimmunisierten TiereD, die einen m/issig hohen Serumtiter aufweisen, l~isst sich die hemmende Wirkung des Lungenextraktes dutch Erhitzen auf 56 ~ und 63 ~ C. zwar verringern, abet nicht v611ig zum Verschwinden briDged. Es bleiben nach Inaktivierung der Acceptoren imlner noch die spezifischen Antistoffe fibrig, die ebenfalls die Haemagglutina- tion hemmeD. Schliesslich wurde auch in den Virus-Neutralisie- rungsversuchen gezeigt, dass die Acceptoren nicht imstande sind

Haemagglutination durch Pockenvirus. 115
die Infektiosit~it von Vaccinevirus nur im geringsten Masse zu be- einflussen.
Die Bedeutung der Acceptoren ist jedenfalls in etwas anderem zu suchen: Das Angehen der Infektion wird gew~ihrleistet. Bei der Variola-infektion sind die Atemwege die wichtigste porte d'entrde. Die Acceptoren fiir das eingedrungene Virus finden sich in den Epithelien der Atemwege. Wenn die bier geschilderten Versuchen vorwiegend mit Lungenextrakten durchgeffihrt sind, so liegt das daran, dass mit diesen besonders gut quantitativ zu arbeiten ist. Abgeschabtes Trachealepithel wirkte gleichfalls agglutinations- hemmend. Fiir eine besondere Affinit~it des Epithels der Atemor- gane spricht auch der durch GINS, HACKENTHAL und KAMENT- ZEWA (9) gefiihrte Nachweis von Virus auf den Schleimhaut der oberen Luftwege von Kindern 3--5 Tage nach der Erstvaccination und ebenso bei cutan, intracutan und intraven6s infizierten Kanin- chen und Meerschweinchen.
Gegentiber der Bedeutung der Atmungsorgane.fiir das Zustande- kommen der Infektion tritt die Haut an Wichtigkeit zuriick. Es w~ire sonst in frtiheren Zeiten infolge der damals getibten Vario- lation viel h~iufiger zu echten Variolaerkrankungen gekommen. Es ist daran zu denken, class in der Haut neben Acceptoren noch andere die Virushaemagglutination hemmende Stoffe vorhanden sind, denn STONE und BURNET (15) fanden Anweisungen daftir, dass in Kalbslymphe eine Art Antistoff anwesend war, der den Serum- Antistoffen ~hnelte.
Wenn auch in den Atmungsorganen zweifellos besonders viele Acceptoren ffir Pockenvirus vorhanden sind, so finden sich solche ebenfalls in anderen Organen, allerdings nicht so regelm~issig und nicht in gleicher Menge. Dies stimmt fiberein mit den frfiheren Befunden fiber den Nachweis yon Vaccinevirus in den inneren Organen. Unabh~ingig v o n d e r Infektionsweise wiesen HAAGEN und KODAMA (11) bei Kaninchen und M~iusen in fast allen Organen Elementark6rperchen nach, nachdem schon vorher BIjL und I~'RENKEL (6), PASCHEN (14) und DOUGLAS und PRICE (7) Meta- stasen in fast allen Organen gefunden hatten. Schwieriger war der Nachweis in den Organen geimpfter Rinder, worauf ZELLER, GILDEMEISTER und HILGERS (17) und LEHMANN (13) hinwiesen.
Antisera gegen Lungenextrakt verm6gen die Inhibinwirk~lng auf- zuheben. Diese Wirkung erstreckt sich nicht allein auf den Lungen- extrakt, mit dem die Antiseren gewonnen sind, sondern auch auf

116 W . A . Collier,
Lungenext rakte yon ganz anderen Tierarten. Daher ist bei diesem Neutralisierungseffekt die species-specifische Komponen te aus-
zuschliessen. Auch Adsorpt ion an anderes Gewebe, beispielsweise
Lebergewebe, v o n d e r gleichen Tierart hebt die Wi rkung nicht auf. Es ist auf eine gewisse Verwandtschaf t der verschiedenen Vaccine-
virus-Acceptoren i n den Lungen der verschiedenen Tierspecies zu schliessen.
Wenn man sich auch besonders auf dem Gebiete der Virusfor- schung vor Veral lgemeinerungen htiten muss, so liegt doch der
Gedanke nahe, dass die Empf~inglichkeit verschiedener Tierspecies ftir eine natiirliche Infekt ion mit einem bes t immten Erreger ab-
h~ngig sein dfirfte yon gleichen oder ~ihnlichen empfiinglichen
Acceptoren an den Zellen jener Organsysteme, die ftir diesen Erreger die por te ,d 'entr~e darstellen.
Z . u s a m m e n f a s s u n g .
In dem Lungenex t rak t verschiedener Tierspecies findet sich ein Stoff, der die Vaccine-Haemagglut ina t ion hemmt . Dieses Inhibin
ist thermolabil und wird bei 56 ~ oder 63 ~ C. nach 30 Minuten zer- stSrt. In der Lunge des Menschen findet sich daneben noch ein
thermostabi ler Stoff, der im Eisschrank schnell verschwindet. Durch Immunis ie rung mit Lungenex t rak t l~isst sich vom Kanin-
chen Ant iserum gegen die Inhibine nicht nut aus M~uselunge,
sondern auch gegen solche aus Lungen anderer Tierar ten gewinnen.
Es wird die Bedeutung dieser die Haemagglu t ina t ion hemmenden Stoffe als , ,Virus-Acceptoren" besprochen, die jedenfalls ftir das Zus t andekommen der Infekt ion yon Bedeutung sind.
L i t e r a t u r .
1. \V. I. B. BEVERIDGE und P. E. LI.~D, Austral. J. exp. Biol. med. Sci. 24, 127, 1946. - 2. M. BOVARNICK und P. M. DE BURGH, Science 105, 550, 1947. - 3. P. M. DE BURGH, P. C. YU, C. HOWE und M. BOVARNICK, J. exp. Med. 87, ], 1948. - 4 . F. M. BURNET und V~r. I. B. BEVERIDGE, Austral. J. exp. Biol. med. Sci. 23, 186, 1945. - 5. F. M. BURNET, J. F. McCREA und S. C. ANDERSON, Nature 160, 404, 1947. - 6. J. P. BIJL und H. S. FRENKEL, Zentralbl. f. BakL I, 112, 412, 1929. - 7. S. R. DOUGLAS und L. R. W. PRICE, J. Path. a. Bact. 32, 99, 1929. - 8. W. F. FRIEDEVc'ALD, E. S. MILLER und L. R. W'HATLEu J. exp. Med. 86, 65, 1947. - 9. H. A. GINS, H. HACKENTHAL und iN. ~AMENTZEVCA, Z. f. Hyg. 110, 429, 1929. i 10. R. H. GREEN und D. W. WOOLEY, j. exp. Med. 86, 55, 1947. - 11. E. HAAGEN und M. Ko-

H a e m a g g l u t i n a t i o n d u r c h Pockenv i rus . 117
DAMA, Z e n t r a l b l . f. Bak t . I, 133, 23, 1934. - 12. G. HENLE, W. HENLE und S. HARRIS, Proc. Soc. exp. Biol . a. Med. 64, 290, 1947. - 13. W. LEHMANN, Z e n t r a l b l . f. B a k t . I, 131, 336, 1934. - 14. E. PASCHEN, in KOLLE--KRAuS-- UHLENHUTH, H a n d b . d. p a t h . Mikroorg . Bd. V I I I / 2 , 82], 1930. - 15. J. D. STONE und F. M. BURNET, Aus t r a l . J . exp. Biol . med. Sci. 24, 9, 1946. - 16. A. SVEDMYRj Bri t . J . exp. P a t h o l . 29, 295, 1948. - 17. H. ZELLER, E. GILDEMEISTER u n d P. HILGERS, Z e n t r a l b l . f. B a k t . I, 128, 21, 1933.





























