Embed Size (px)
Citation preview

Roux ' Arehiv ftir Entwieklungsmechanik 158, 13--51 (1967)
H I S T O L O G I S C H E U N D E X P E R I M E N T E L L E
U N T E R S U C H U N G E N U B E R D I E E N T W I C K L U N G
D E S O P I S T H O N E P H I ~ O S B E I T R I T U R U S A L P E S T R I S *
ROLF LEHMANN
Zoologisches Institut der Universit~t Freiburg i. Br. (Direktor: Prof. B. I-IASSENSTEIN)
Eingegangen am 17. November 1965
Inha l t seite
A. Einlei tung . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14 I. Fragestel lung . . . . . . . . . . . . . . . . . . . . . . . . . . 14
I I . Der Opis thonephros . . . . . . . . . . . . . . . . . . . . . . . 15 B. Unte rsuehungsobjek te und Arbe i t smethoden . . . . . . . . . . . . . 16 C. Histogenese der 0p is thonephroskan~lchen . . . . . . . . . . . . . . 17
I. Differenzierung des Blastems . . . . . . . . . . . . . . . . . . 17 I I . Ausbi ldung der Nephroblas tb lasen . . . . . . . . . . . . . . . . 19
I I I . Sonderung der Kani / lehenabschni t te . . . . . . . . . . . . . . . 20 1. NierenkSrperchen . . . . . . . . . . . . . . . . . . . . . . 20 2. Nephros tom . . . . . . . . . . . . . . . . . . . . . . . . 22
a) Ventrale Nephros tome . . . . . . . . . . . . . . . . . . . 22 b) Dorsale Nephros tome . . . . . . . . . . . . . . . . . . . 23
3. Tubulusepi thel . . . . . . . . . . . . . . . . . . . . . . . 25 4. Miindung in den pr im~ren Harnle i ter . . . . . . . . . . . . . . 29 5. Dorsolateraler Bogen . . . . . . . . . . . . . . . . . . . . . 30 6. Differenzierungsfolge innerhalb eines Kan~ilchens . . . . . . . . 30
IV. He rkun f t dorsaler Anlagen . . . . . . . . . . . . . . . . . . . 31 D. Defekte am intermedi~iren Mesoderm . . . . . . . . . . . . . . . . . 32
I. Deiektse tzung . . . . . . . . . . . . . . . . . . . . . . . . . 32 I I . Folgen der Defektsetzung . . . . . . . . . . . . . . . . . . . . 33
1. Fehlende Organteile . . . . . . . . . . . . . . . . . . . . . 33 2. Beeinflussung des DifferenzierungsgefSJles . . . . . . . . . . . . 35 3. Regenerat ive Prozesse . . . . . . . . . . . . . . . . . . . . . 36
E. Transp lan ta t ionen des intermedi~ren Mesoderms . . . . . . . . . . . . 38 I. Ausfi ihrung der Exper imente . . . . . . . . . . . . . . . . . . . 38
I I . Differenzierung der Transp lan ta te . . . . . . . . . . . . . . . . . 39 1. Muskula tur . . . . . . . . . . . . . . . . . . . . . . . . . 39 2. Urgesehleehtszellen . . . . . . . . . . . . . . . . . . . . . . 39 3. Gef~Be und Blur . . . . . . . . . . . . . . . . . . . . . . . 4I 4. Nierenanlagen . . . . . . . . . . . . . . . . . . . . . . . . 41
a) Nierenkani~lehen . . . . . . . . . . . . . . . . . . . . . . 41 b) NierenkSrperehen . . . . . . . . . . . . . . . . . . . . . 43 e) Nephros tom . . . . . . . . . . . . . . . . . . . . . . . . 44 d) Vergleieh der Nierenkan~/lehen . . . . . . . . . . . . . . . . 44
F. I ) iskussion der Ergebnisse . . . . . . . . . . . . . . . . . . . . . . 46 Zusammenfas sung . . . . . . . . . . . . . . . . . . . . . . . . . . 49 Li te ra tur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
* Angeregt yon H e r i n Dr. 0. J . ST3i~K.

14 g. L E H ~ N :
A. Einleitung I. Fragestellung
Die Entwicklung der Urodelenniere ist schon frfiher an Salamandra salamandra (Fff~BnI~GER, 1877) und ffir Triturus vulgaris (GRAY, 1932) beschrieben women. In der zuerst erw~hnten Arbeit mugte haupts~ch- rich aus Grfinden datums noch unzureiehender techniseher Methoden auf die Beschreibung histologiseher Einzelheiten verzichtet werden. Da- bei erscheinen aueh keine Angaben fiber Flimmerepithelien. In der Arbeit von GnAY (1932) sind nur wenige histologische l~bersichtsbilder enthalten, die fiber den cytologischen Differenziernngsvorgang eines Opisthonephroskan/~lchens nut tmgenfigend Aufschlug geben. In zu_ sammenfassenden Werken (H~sRTWm, 1906; F~A~CIS, 1934; NOBL~, 1954) sind nur Teilangaben zu finden, die sich auf die vorgenannten Autoren beziehen. Die genaue Kenntnis der histologischen Struktur des untersuchten Organs aber ist Voraussetzung ffir die Auswertung der im Transplantat differenzierten Strukturen, ffir die Interpretation elektro- nenmikroskopischer Befunde und zur sp/iteren autoradiographischen Untersuchung. Deshalb ist es notwendig, zuerst die ttistogenese der Opisthonephroskan~lehen zu studieren.
Keine einheitliche Anffassung besteht fiber die Entwicklung der Nierenkan~lchen. Nach F~RBRISG~R (1877) entstehen bei S. salaman- dra sekund~re Kan~lehen zungchst getrennt yon den prim~ren Nephro- hen, mit deren obersten Windnngen sie spi~ter verschmelzen (S. 85), auch die terti~ren Anlagen entwickeln sich in gleieher Weise und treten mit den lateralen Tubuli der sekundaren Kan~lchen in Verbindung, so dab die lateralen Abschnitte der primi~ren Kan~tlchen zu Sammelr6hren ffir die prim~ren, sekund/~ren und terti~ren Tubuli werden (S. 117). Nach G~AY (1932) entstehen bei Tr. vulgaris seknnd~re und tertii~re Tubuli dutch Sprossung aus dem primgren Kanglchen (S. 459), Auch bei NosLE (1954) steht, dab bei Urodelen aus den prim~ren Tubuli, nachdem sie funktionell t~tig sind, sekundgre aussprossen (S. 268). - - Jedoch sind Verschmelzung und Sprossung yon Kani~lchenabschnitten zwei gegens~tzliche Prinzipien, die sehr unterschiedliche Potenzen des betreffenden Zellmaterials voraussetzen. - - GRAY (1932) ffihrt an, dal~ keiner der frfiheren Autoren in ihrer Entwicklung ausreichend fortge- schrittene Anlagen rekonstruiert babe (S. 452). Da diese Aussage kei- nesfalls zur L6sung des Problems beitr~gt, soll an einem weiteren Ver- treter der Salamandriae das Prinzip der Kan~lchenbildung untersucht werden.
Die Entwieklung yon Keim- und 0rganteilen geht yon Blastemen aus (Ls~MA~N, F. E., 1945). Ffir die Niere wird ein solider Blastem- strang angegeben (F~gBm~G~R, 1877; HAJ~n, 1904; F~n~x, 1906; GRAY, 1932). Dieser Strang leitet sieh vom Blastem der Randzone ab, die

Entwicklung des 0pisthonephros bei Triturus alpestris 15
auger Niere Chorda, Somiten, Sei tenpla t ten u n d Blut liefert. Aus der
Li te ra tur konnte keine befriedigende Vorstellung fiber das Nierenbla- s tem bei Tr i turus gewonnen werden. I t ier eine Kla rung zu finden, war eine weitere Aufgabe, die ieh mir stellte.
Welehe Folgen haben Defekte am intermedi/ tren Mesoderm, insbe- sondere ffir die En twiek lung des Opisthonephros ? Naeh NmEvxooP (1948), tier die Pronephrosanlage und den p rgsumpt iven primi~ren Harn- leiter im frfihen Neuru las tad ium ent fern t hat, beeinf lugt der prim/~re Harnle i te r die Differenzierung des , ,Mesonephrosblastems". Das Bla- s tem braueht n u t einen kleinen induk t iven Anstog. Naeh MACHENE~ (i929) iSt die pr / isumptive , ,Urnierenanlage" im frfihen Neuru las tad ium noch nieht determinier t . I s t der primfi, re t Ia rn le i te r aueh nach der De- t e rmina t ion des Opisthonephrosblastems yon Einf lug auf die Differen- zierung der Kan~lehenanlagen ? Weitere Fragen fi ihrten zu Transplan- ta t ionsversuehen: Dis zu welehem Differenzierungsgrad k6nnen sieh Nierenkan/i lchen am fremden Ort entwiekeln ? Gliedern sie sieh in histologisch folgeriehtige Abschni t te ? Von welehen Einflfissen k a n n die En twick lung abh/~ngig sein ?
I I . Der Opisthonephros Obgleich die Niere einer 3 0 ~ 1 mm langen Larve yon Triturus alpestris noch
nicht ihre endgiiltige Gestalt erreicht hat, sind in der gesamten Region, in der ws der Normogenese die Entwicklung yon Nierengewebe re~lisiert wird, be- reits Kanalchen angelegt. Auf die KSrperoberflache projeziert, erstreckt sich die Niere yon etwa caudal der Vorderextremitat bis dorsal hinter die Beckenextremi- tat. In bezug auf benachbarte Organe beginnt der Opisthonephros mit seinen ersten Kanalchenschleifen dorsal fiber der l~Iitte des Magens. Nach etwa einem Vier- tel seiner Gesamtliinge erreicht er den Gonadenbereich und zieht parallel zum Dickdarm bis an das caudale Ende der LeibeshShle. Im vorderen Bereich liegt die Niere sehr weit medial, im Gonadenbereich etwas mehr lateral, und caudal wird sie durch die Begrenzung des Beckens wieder zur KOrpermitte gedrangt. Die ge- samte Nierenregion einschlieNich des Pronephros, der fiber dem vorderen Leber- lappen liegt, reicht veto dritten bis zum sechzehnten metotischen Segment. Nur das fanfte Segment, das zwischen Vorniere und Opisthonephros liegt, bleibt bei Triturus alpestris ohne Nierenbildung, so dab sich der Opisthonephros fiber elf Segmente erstreckt.
Der vordere Tell der Niere tritt bei den mannlichen Tieren in Beziehung zu den Gonaden, wahrend aus dem hinteren Abschnitt durch weitere Anlage zahl- reicher Kanalchen die Niere des adulten Tieres entsteht.
Ein Frontalschnitt dureh die Niere stellt uns ihre Architektur vor. Der Schnitt fahrt durch drei verschiedene Regionen: Cranial liegen hauptsachlich Anschnitte yon NierenkSrperchen (Abb. 1), im mittleren Bereich sind vorwiegend Tubuli ge- troffen, und caudal treten uns Anschnitte noch wenig differenzierter Kanalchen und yon Nephroblastblasen entgegen.
Des erste Abschnitt des Nierenkanalchens, der an der Bowmanschen Kapsel seinen Anfang nimmt, wird als Halsstfick bezeiehnet und ist bewimpert. Die Tu- bull verlaufen scheinbar ungeordnet in vielfachen Schleifen und Windungen. Sie gliedern sieh auf Grund ihrer histologischen Differenzierung in mehrere Abschnitte.
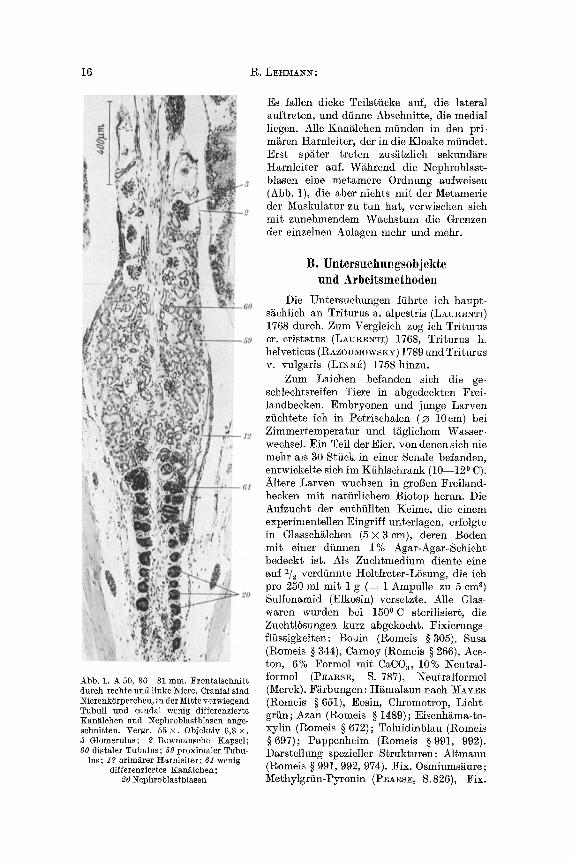
16 R. LEIEVIA~I~:
Abb. 1. A 50, 30 31 ram. Frontalschnitt dutch rechte und linke LNiere. Cranial sind ~ierenk6rperchen, in der Mitre vorwiegend Tubuli und caudal wenig differenzierte Kani~Ichen und Nephroblastblasen ange- schnitten. Vergr. 55 • Objektiv 6,3 • 3 Glomerulus; 2 Bowmansche Kapsel; 60 distaler Tubulus; 59 proximaIer Tubu-
lus; 12 prim~rer ttarnleiter; 61 wenig differenziertes Kan~lchen;
20 l%phroblastblasen
Es fallen dicke Teils~ficke auf, die lateral auftreten, und diinne Abschnitte, die medial liegen. Alle Kan~lchen miinden in den prL m~ren Harnleiter, der in die K]oake miindet. Erst sp~ter treten zus~tzlich sekund~re Harnleiter auf. W~hrend die 1Nephroblast- blasen eine metamere Ordnung aufweisen (Abb. 1), die aber nichts mit der Metamerie der !Yiuskulatur zu tun hat, verwischen sich mit zunehmendem Wachstum die Grenzen der einzelnen Anlagen mehr und mehr.
B. U n t e r s u c h u n g s o b j e k t e
und A r b e i t s m e t h o d e n
Die Untersuchungen ffihrte ich haupt- s~chlich an Triturus a. Mpestris (LAuRENTI) 1768 durch. Zum Vergleich zog ich Triturus cr. cristatus (LAuIaEI~TI) 1768, Triturus h. helveticus (RAzouMows~Y) 1789 und Triturus v. vulgaris ( L I ~ ) 1758 hinzu.
Zum LMchen befanden sieh die ge- schlechtsreifen Tiere in abgedeckten Frei- landbecken. Embryonen und junge Larven zfichtete ich in PetrischMen (;~ 10cm) bei Zimmertemperatur und t~glichem Wasser~ weehsel. Ein Tell der Eier, yon denen sich hie mehr als 30 Stiick in einer SchMe beianden, entwiekelte sich im Kiihlschrank (10--12 o C). Altere Larven wuchsen in gro•en Freiland- becken mit natfirlichem Biotop heran. Die Aufzucht der enthfillten Keime, die einem experimentellen EingriH unterlagen, erfolgte in Gl~ssch~lehen (5 • 3 era), deren Boden mit einer diinnen 1% Agar-Agar-Schicht bedeckt ist. Als Zuchtmedium diente eine auf 1/2 verdfinnte Holtfreter-LSsung, die ich pro 250 ml mit 1 g ( = 1 Ampulle zu 5 cm a) Sulfonamid (Elkosin) versetzte. Alle Glas- waren wurden bei 150 ~ C sterilisiert, die ZuchtlSsungen kurz abgekocht. Fixierungs- fliissigkeiten: Bouin (t~omeis w Susa (Romeis w 344), C~rnoy (Romeis w 266), Ace- ton, 6% Formol mit CaCOs, 10% l~eutral- formo] (PE~a~SE, S. 787), NeutrMformol (lYierek). F~rbungen: tt~malaun nach ~r (l~omeis w 651), Eosin, Chromotrop, Licht- grfin; Azan (Romeis w 1489); Eisenh~ma-to- xylin (~omeis w 672); Toluidinblau (Romeis w 697); Pappenheim (l~omeis w 992). Darstellung spezieller Strukturen: Altmann (l~omeis w 991, 992, 974), :Fix. Osmiums~ure; Methylgrfin-Pyronin (PEA~SE, S.826), Fix.

Engwieklung des Opist~honephros bei Triturus Mpestris 17
Carnoy; Gallocyanin-Chromalaun (Spannhof, Vers. 44), Fix. Athanol; Feulgen (Spannhof, Vers. 42), Fix. Carnoy ohne Chloroform.
Die verschiedenen Fixierungsfliissigkeiten nnd Fgrbungen dienten zum Ver- gleich der Strukturen des Untersuchungsobjektes. Alle Mikrophotos nach BovI~ fixierten Pr~paraten, F/irbung mit It~malaun (MAYER) und Eosin oder Chromotrop.
Larven sowie Keime, die sich bereits bewegten, narkotisierte ich vor der Fixierung durch ~ropfenweise Zugabe yon 1% Trichlorbutylalkohol. Um ein schnelles Durchfixieren des Nierengewebes zu sichern, 5ffnete ich bei gr6geren Tieren die Leibesh6hle. Die Einbettung der Objekte geschah nach der iiblichen Paraffinmethode. Die Sehnittdicke betr/~gt einheitlich bei allen Serien 10 ~m. EinschluBmitteh Eukitt und Entellan.
Die Transplantationen und Defektversuche fiihrte i ch unter dem Leitz-Binoku- lar (12,5fache VergrdBerung) mit den gebr~uchlichen Instrumenten der Entwick- lungsphysiologie aus: Glasnadeln, Haarsehlingen, feine Lanze~tmesser, Uhrmacher- Pinzetten.
Die Beobaehtung der Eingriffe erfolgte bei i2,5-, 25- und 50faeher Vergrdl3e- rung. Fiir Operationen benu~zte ieh flaehe Seh~lehen, die mit niedrig sehmelzen- dem Paraffin odor I0% Agar-Agar ausgegossen waren; beide Stoffe enthielten zur Kontraststeigerung pulverisierte Lindenholzkohle.
Zur Anfertigung der Zeiehnungen naeh mikroskopisehen Pr~paraten kam der Zeiss-Zeiehenaufsatz zur Anwendung. Die YIikrophotos wurden mit dem Zeiss- Photomikroskop hergestellt. Soweit keine anderen Daten angegeben sind, ver- wendete ich das Zeiss-Objektiv Apo 40/1,00el. Als Filmma~erial diente aussehlieg- lieh Adox KB 14. Um Kontroll- und Betegaufnahmen der operierten Tiere anzu- fertigen, setzte ieh ein Praetiea-Geh/iuse unter Gebraueh eines Zwischenstiickes auf einen Binokulartubus.
C. Histogenese der 0pisthonephroskan~ilchen
I. Di//erenzierung des Blastems
Bei einer sehr jungen La rve (8 - -9 m m lung) t r e ten u n m i t t e l b a r nach dem Schlfipfen im Bereich der sp&teren Nierenregion Zellen mi t re la t iv grogen Kernen auf, die sich mi t H/~malaun s t a rk anf/~rben. Die ovoiden Kerne , die mehrere Nukleol i en tha l ten , bef inden sich zwischen pr im~- rem I t a rn l e i t e r und ven t r a l em Myotomrand , einige l iegen auch a m me- d ia len oder la te ra len R a n d der Myotome. Von den Nachbarze l len lassen sic sich e indeut ig unterscheiden. Die Kerne der Myotomzel len sind lich- te r angef/ irbt , rundl icher und fal len meis tens durch einen grol?en Nu- kleolus auf. Die in der NiChe l iegenden Bindegewebskerne t r e t en durch ihre langges t reck te Ges ta l t hervor . Die Ep idermis und das sehr do t te r - reiche en todermale Mater ia l g l iedern sich k lar ab. I m Verlauf yon Ge- s t a l tungsbewegungen werden zusammen mi t dem pr ims t t a rn ]e i t e r die yon e inem dfinnen C y t o p l a s m a m a n t e l umgebenen , s t a rk anfi~rb- baren Kerne in media le R ich tung ver lager t . Diese Zellen s ind die Bi]- dungszel len der Nierenkan/ i lchen.
Die ers ten l i eh tmikroskopiseh beobaeh tba r e n Differenzierungen des Opis thonephrosb las tems s ind bei einer 9 - - 1 0 m m langen Larve zu sehen. Uber dem p r imgren t I a rn l e i t e r haben sieh Zel lgruppen ausgebi ldet .
2 Wilhelm Roux ' Arch. Entwickl.-~clech. Org., Bd. 158

18 R.L~MAN~:
Die Kerne sind denen der Blastemzellen in Gestalt und Struktur /~hn- lich. Auffallend ist bei beiden ein starkes Randchromatin, das den Eindruek einer besonders scharf gezeichneten Kernmembran hervorruft. Die Zellen enthalten nur sehr wenig Cytoplasma, das sich als sehwaeh eosinophil erweist. Die Zel]gruppen bilden keinen einheitlichen Strang, sondern eine unregelm/~gige Folge kleinerer und gr6Berer Zellanh/~ufun-
gen. Bereits zu diesem Zeitpunkt tr i t t ein cranio-caudales Differenzie- rungsgef~lle hervor. W~hrend im cranialen Bereieh mehrere Zellen pro Quersehnitt auftreten, ist bei demselben Tier welter caudal nur ein grol3er Kern naehweisbar.
Die Differenzierung zu Zell- gruppen ist eine Vorstufe der Ne- phroblastblase. Sic geht yon cranial naeh caudal fortschreitend yon Ein- zelzellen aus und f/ihrt zu Zell- assoziationen verschiedenen Grades. Die Gesamtheit der ZellanMulungen ist als primitive Opisthonephros- anlage aufzufassen. Da sieh aber im Verlaufe des Entwicklungsganges noch an anderen Orten, n~mlich
Abb. 2. A 603, 37--38 m m . Querschni t t du tch dorsal tier bisherigen Folge, Zell- die caudale Nierenregion. I n der Mitre eine Nephroblastblase der zweiten dorsalen gruppen bilden, kann die ventrale Schicht, darfiber eine Zellgruppe der dr i t t en Folge nur eine Teilanlage sein. dorsalen Anlagenschicht . Vergr . 625 x .
35 Kerne tier dorsalen Zel lgruppe; Wenn die letzten Anlagen der 14 TubuIus~nsehni t te
ventralen Schieht als Nephroblast- blasen vorliegen, sind in diesem Bereich keine Blastemzellen oder Ab- k6mmlinge yon ihnen mehr nachweisbar. Aber dorsal der ventralen Anlagen treten locker gef/igte Zellgruppen auf, deren Zellen eine groBe Ahnlichkeit mit denen ventraler Zellgruppen in jfingeren Larven haben. Es fgllt auf, dab die Zellen der jfingeren Nierenanlage nieht so dieht gedr~ngt liegen wie bei der/~lteren. Die Streuung der Zellen im loekeren Bindegewebe ist sieher eine Folge des zur Verfiigung stehenden vergrS- l]erten Raumes. Wghrend bei einer jungen Larve der Zwiekel zwischen Aorta, Venen und C61omepithel einerseits nnd rechtem und linkem Myotomrand andererseits den Zellen nur wenig BewegungsmSgliehkeit bietet, l~Bt bei einem ~lteren Tier das loekere Netzwerk des Binde- gewebes eine gr6Bere Ortsvergnderung zu.
t Iaben sieh aus den Zellgruppen der ersten dorsalen Sehieht gleieh der ventralen Anlagenfolge Nephroblastblasen differenziert, sind dorsal

Entwicklung des Opisthonephros bei Triturus alpestris 19
yon ihnen abermals Gruppen des bereits bckannten Zellentypus zu er- kennen. Iqachdem sich diese Zellgruppen zu INephroblastblascn ent- wiekelt haben, treten fiber ihrer Dorsalseite erneut Kanglchenbildungs- zellen auf, die die Gruppen der dritten dorsalen An]agenfolge bilden (Abb. 2).
Von einer 8--9 mm-Larve bis zu einem 54--55 mm langen Tier treten in kontinuierlicher Folge AbkSmmlinge yon Nierenblastemzellen auf, die sich yon cranial naeh caudal fortsehreitend differenzieren. Unter einem Nierenblastem haben wit uns eine Folge yon Zellen vorzu- stellen, die in unregelmgl3igen Abstgnden voneinander am ventralen Myo~omrand liegen. Sic kommen aus dem intermedigren Mesoderm, das neben Nierenbildungszellen auch noch andere Zellen enthglt. Die Differenzierungen des Blastems treten in mindestens vier Schichten nacheh~ander auf, yon denen jede dorsal dcr bereits in ihrer Differen- zierung fortgeschrittenen Kanglchenanlage liegt.
Diese schichtmgBige Differenzierung findet nur im kiinftigen Bek- kennierenbereieh start. Sowohl bei der jiingeren als auch bei der glteren Larve treten im cranialen Nierenbereich, der keine dorsalen Zellgruppen enthglt, nur ventrale Anlagen auf.
II. Ausbildung der Nephroblastblasen Die Zellml, die in dichten Gruppen zusammenliegen, weiohen mi~
zunehmender Zahl unter Bildung eines Lumens auseinander (Abb. 7 a, 4). Aus der locker gefiigten Gruppe ist eine Blase entstanden, die yon einem einschichtigen Epithel gebfldet wird. Drei Prozesse kennzeichnen den Weg yon der relativ einfachen Zellgruppe zur komplizierteren Nephroblastblase. Mit der Blasenbildung kehrt in das Zellgefiige eine vermehrte Ordnung ein, die bereits mit dem Zusammentreten der locker nebeneinander liegenden Zellen zu einer gesehlossenen Gruppe eingeleitet wird. Gleichzeitig finden Wachstumsvorggnge statt, die Ge- stalt und Gr6Be der primitiven Anlagen vergndern. Daneben lguft die histologische Differenzierung ab, die sich in einer Vergnderung der Zell- strukturen zdgt. Besonders auffgllig ist der Formwechsel der Kerne yon einer anfangs ovalen GestMt zu einer tanggestreekten Form (Abb. 7a, 5). Mit dem Einsetzen der Blasenbildung werden bereits Uberggnge zu langgezogenen Kernen sichtbar. Die glteren Nephroblastblasen enthal- ten fast nur Zellen mit z. T. extrem gestreekten Kernen.
Die junge Nephroblastblase der ventralen Anlagenserie bfldet sich dorsolateral vom primgren Harnleiter. Bei einer etwas gr5Beren Larv~ hat eine relative Jmderung der Lage stattgefunden. Die gltere Blase liegt jetzt dorsomedial yore primgren tIarnleiter. Wghrend der Binde- gewebszwickel zwisehen rechten und linken Myotomen mit steigender
2*

20 R.L~HMA~:
K6rpergrSSe breiter wird, bleibt die Lagebeziehnng zwischen Blasen und prim/~rem Harnleiter bei allen Anlagen erhalten.
Das cranio-caudale Differenzierungsgef~lle innerhalb der ventralen Anlagen- folge tritt bei Gegenfiberstellung je eines Quersehnittes dureh den grSBten Durch- messer der Nephroblastblasen z.B. der 24, 29. und 36. Anlage einer 2 6 4 7 mm- Larve hervor. Wghrend die vordere Blase bereits Kontakt mit dem prim~ren Harnleiter aufgenommen hat, ist die etwas weiter zum K6rperende liegende Blase kleiner and enth~lt rundere Kerne als die vordere. Die letzte Blase, die fiber dem nach ventral zur Kloake verlaufenden primgren Harnleiter liegt, bildet gerade ein Lumen aus, ihr Durchmesser ist geringer und die Gestalt der Kerne noch rand- licher als bei der vorhergehenden.
Die Nephroblastblasen in der ersten dorsalen Anlagenschicht bilden sich h~ der gleiehen Weise aus wit die in der ventralen Lage. Bei Be- traehtung der gr6Bten Durehmesser cranialer und eaudaler Nephro- blastblasen ist zu sehen, dab auch hier die Differenzierung yon vorn nach hinten abnimmt. Die dorsalen Nephroblastblasen entwiekeln sieh immer in der N/ihe eines dorsolateralen Bogens. Wird die Nephroblast- blase s t r i t t sie mit ihm in Verbindung. Die Kontaktnahme der Nephroblastblase mit dem dorsolateralen Bogen ist vergleiehbar mit der Versehmelzung der Epithelien einer ventralen Nephroblastblase mit dem prim/tren Harnleiter. Die liehtmikroskopiseh beobaehtbaren Pha- sen dieses Vorganges sind die gleichen. Das Epithel des dorsolateralen Bogens ist dem Epithel des prim/tren I-Iarnleiters zur Zeit der Ver- sehmelznng ~hnlieh.
Auch ffir die zweite dorsale Schieht der Nephroblastblasen trifft zu, dag die cranialen welter differenziert sind als die caudalen Anlagen. Die Nephroblastblasen der dritten Anlagensehicht geben ebenfalls ein Beispiel fiir das cranial nach caudal anftretende Differenzierungsgefalle. Ffir alle dorsalen Anlagenfolgen gilt aber, daft keine strenge Differen- zierungsfolge eingehalten wird, sondern immer wieder einzelne Anlagen in ihrer Entwieklung im Vergleich zu ihren Nachbarblasen zu weir vor- angeschritten sind.
III . Sonderung der Kani~Ichenabschnitte
1. Nie~'enkSrperchen
Nachdem sich die Nephroblastblase zu einem S-f6rmigen Strang (Abb. 7a, 6) entwiekelt hat, dessen tines Ende mit dem Epithel des prim/tren Harnleitcrs verschmolzen ist, deutet sich am entgegengesetz- ten Abschnitt, der blind endet, eine Ver/inderung der Zellen an. Die Kerne der dicht gedr/~ngt liegenden Zellen beginnen sich am proxima- len Absehnitt gerade abzurunden und damit dig Differenzierung zum Anfangsteil des Nierenkan/~lchens einzuleiten. Mit weiterer Entwiek- lung werden die Kerne der ventral liegenden Zellen kleiner und das

En~wicklung des Opist.honephros bei Triturus Mpestris 21
Epithel wird flaeher. Diese Differenziernng setzt medial ein und sehreitet naeh lateral fort. Gleiehzeitig riieken mit der Abflachung der Kerne die Zellen welter auseinander. Das bislang kubisehe Epithel formt sieh zu einem Plattenepithel urn, das mit seinen sehr flaehen Zellen ein Epithelbl/~sehen (Abb. 3 , Bowmansehe Kapsel) bildet, dessen Hohl- raum tiber das bewimperte Hals- sttiek (Abb. 3 und 4) mit dem Tu- bulus in Verbindung steht.
Die dorsalen Zellen des blind- endenden proximalen Absehnittes nehmen einen ~onderen Entwiek- lungsgang als die unteren. Ausgehend yon gleiehartigen Zellen runden sieh die Kerne ab, ohne sieh wie im ven- tralen Tell abzuflaehen. Bei diesem Vorgang senken sieh die Zellen in d~s en~stehende Epithelblg~sehen ein (Abb. 3). Gleiehzeitig entsteht ein Hilus, dutch den eine Kapillare einsproBg, kenntlieh an den zwei Erythrozyten (Abb. 3).
Aus der Glomerulusanlage ent- stehg der Glomerulus. Deckzelien, die sieh von kubisehen Epithelzellen ableiten, tiberziehen das arterielle Kapillarnetz. Mit zunehmendem Alter der Larven werden die Nieren- k6rperehen volumin6ser. Ein Quer- sehnitt dureh die Nierenregion zeigt die membranartige Oberfl~ehe des Epithelbls den Glomerulus
~ylhrozften 10p/1/are //~ Bowmonsehe/(apse/
?ii !
..... . . .[(. ' ~ ' ~ ~ i i ;:2O ":7..~'~'~'::""'<"/":'":::"3"~-'. 3. ~i:<.~, ~'
I / &hseo~n~ i //alsstJcX ~/n/~ C/Z/
~ubulusaz;chn/~ Abb. ~:, A 54, 20--21 ram. Querschni~t dutch ~t~e c~udale N i e r e ~ r e g ~ . Glomerulusb~ld~ng: Dorsale Zellen des l )roximalen Absehni t tes
senken sich in das ents tehende EpithelbI~schen ein
und das zu- und abftihrende GefgB (Abb. 4). Ein- und Austritt der A. afferens nnd A. efferens liegen nicht so dieht beieinander, dab man yon einem Gefggpo] spreehen k~nn. Die Arterie ~ritt yon dorsaI ein und naeh lateral wieder aus (Abb. 4).
Gleichzeitig mit dem Glomerulus und der Bowmansehen Kapsel differenziert sieh das tIalsst/iek, das die Verbindung des Nierenk6rper- ehens mit dem Kan/~lehen herstellt. Dieser Absehnitt entwiekelt sieh im AnsehluB an die Glomerulusanlage. Er tr i t t deutlieh hervor, wenn sioh die Nephrostomanlage naeh ventral ausbuelltet. Bereigs zu diesem Zeitpunkt treten die ersten Cilien auf.
W/ihrend die Nephrostomanlage ausw/tehst, bleibt das Halsstiiek noeh sehr kurz. Alter geworden, verl/~ngert sieh der Halsabsehnitt, und

22 R. LEHhIANlV :
das NierenkSrperchen sitzt wie auf einem kurzen Rohransatzstfick. Deutlich ist zu sehen, wie sich die beiden Wimperflammen des Hals- sttiekes und des Nephrostoms vereinigen (Abb. 4) und in die Cilien des Abschnittes TI fibergehen.
Die lkTierenkSrperehen der ersten dorsalen Sehieht entwiekeln sich entspreehend ihrer sp/iteren Anlage nach Ausbildung der ventralen NierenkSrperchen. Der Entwicklungsgang ist der gleiche. Ein Vergleich von cranial naeh caudal zeigt aueh hier ein Entwicklungsgef/ille. Wiih-
Abb. 4. Querschnitt durch die ~Nierenregion. Yergr. 240 • 25 Aorta; 42 Abschnitt TVI; 48 ~us- kulatur; 9 Nephrostom; 23 A. afferens; 24 A. effe~ens; 26 V. cava post; 27 u portae renis
rend das Nierenk6rperehen des weiter vorn liegenden Kan&lchens Merkmale fortgeschrittener Differenzierung zeigt (flaches Epithel der Kapsel, das in kubisehes mit rundlichen Kernen des I-Ialsstfickes fibergeht, Cilien des ttalsstfickes, aufgelockerter Glomerulus), ist bei den Nierenk6rperchen nachfo]gender Kan~lchen zunehmender embryonaler Charak~er zu bemerken. Das Epithel des Halsstfickes ist noch h6her, die Kerne sind gr6Ber und rund-oval bis l&uglich.
2. Nephrostom
a) Ventrale Nephrostome Die Nephrostomanlage tr i t t gleichzeitig mit der ers~en sichtbaren
Differenzierung der Bowmanschen Kapsel auf. Am proximalen Ende des teilweise mit einem Lumen versehenen Zellstranges entsteht ven- tral eine Zellproliferation, die sich gegen das dorsale C61omepithel aus- dehnt. Aus dieser Zellknospe w&chst ein solider Zellstrang aus, in dem sich bald eiu kleines Lumen bildet.

Entwicklung des Opisthonephros bei Triturus ~lpestris 23
Zu diesem Zeitpunkt der Entwieklung liegen in der Regel der Be- ginn eines Nierenkanalchens (NierenkSrperehen) und sein Ende (Mfin- dung in den primaren Harnleiter) in derselben Querschnittsebene. Das Lumen tier Nephrostomanlage kommuniziert noch nicht mit dem CSlom. Das CSlomepithe], das mit den Zel]en des kfinftigen Nephrostoms in engem Kontakt steht, fiberzieht als diinne Membran die spatere Off- nung des ~qephrostoms. Spater besteht dieser Verschlul3 nicht mehr und das Nierenkanalchen steht mit der LeibeshShle fiber das Nephrostom in oftener Verbindung (Abb. 4).
Die Zellen, aus denen die INephrostomanlage hervorgeht, unter- seheiden sieh anfangs nur wenig yon den Zellen der gesamten Kanal- chenanlage. Sie sind ausgezeiehnet dureh langlieh-ovale Kerne, die yon sehr wenig Plasma umgeben sind. Mit der Entwicklung der Nephro- stomanlage erfolgt ein Wechsel zu grSSeren Zellen mit rundlicheren Kernen. Das differenzierte Nephrostom wird yon einem kubischen Epi- thel gebildet, dessen Kerne kuglig bis oval sind. Der grSl]te Teil der Zelle wird vom Kern ausgeftillt. Zum Lumen hin bleibt ein Plasmasaum frei, in dem die Basis der Cilien liegt. Die Nephrostome haben im cra- nialen Bereieh der Niere eine andere Lage als im caudalen. Vorn liegen die ()ffnungen ventral der Niere, im mittleren Bereich medial nnd im caudalen Abschnitt kommt die TrichterSffnung mediodorsal zu liegen. Sehr versehieden sind die 0ffnungen der Nephrostome ausgebfldet. Wahrend bei einer jungen Niere das Nephrostom wie ein offenes l~ohr beginnt, 5ffnet es sich bei der alteren Niere mit einem ausgepragten Triehter. Das Nephrostomepithel bfldet eine Kreisflache um die eigent- fiche TriehterSffnung. Auch dieses Epithel ist mit Cilien versehen. Nach der Verbindung des Lumens der RTephrostomanlage mit der LeibeshShle geht das Nephrostomepithel am Trichterrand stufenlos in das CSlom- epithel fiber.
b) Dorsale Nephrostome Die Nephrostome der ersten dorsalen Schicht werden als solide Zell-
strange angelegt (Abb. 5). Sie sind bedeutend langer als bei den ven- tralen Flimmertriehtern, da ihr Ausgangspunkt, das Halssttiek der Bow- mansehen Kapsel, vie] welter yore CSlom entfernt liegt, als das bei den ventralen Kanalehen der Fall ist. Der auswaehsende Zellstrang bahnt sich seinen Weg dnrch die Kana]chenwindungen in l~iehtung auf das CSlom (Abb. 5). Wenn der Strang das CSlomepithel erreieht hat, nimmt er mit ihm Kontakt auf. Dann bfldet sich das Lumen, das noch nicht bis zur LeibeshShle durchgeht. Wenn die Nephrostome differenziert sind, werden sie vom gleichen Epithel gebildet wie die ventralen.
Die Flimmertriehter liegen immer im dorsomedialen CSlom, das durch das dorsale Mesenterium in zwei Teilraume unterteflt wird. Da
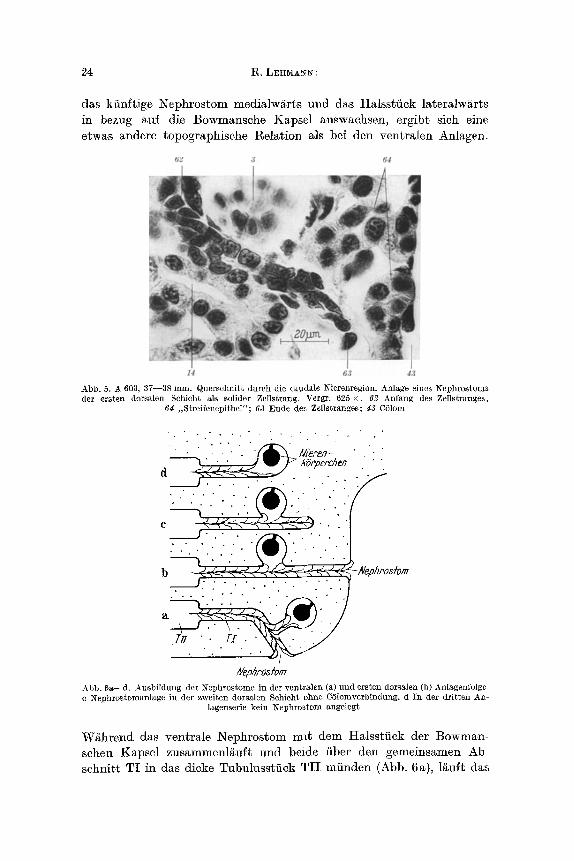
24 ~,LERMA~g:
das kfinftige Nephrostom media,lwarts und das Halsstfick lateralw/irts in be~ug aui die Bowmansche Kapse] ~uswaehsen, ergibt sich eine etwas andcre topographisehe Relation al~ bei den ventralen Anlagen
Abb. 5, A 603, 37--38 ram. Querschnitt durch die caudgle Nierenregion. Anl~go eines Nephrostoms der er~ten dorsalen Schicht als solider Zellstrang. Vergr. 625 • 62 Anfang des Zellstrange~;
64 ,,StreifenepitheI"; 63 Ende des Zellstranges; 43 Cblom
~ kOrper-cken +
d . . 5 - /
e J - . �9 . . . . �9 . .
b'" " " ~ ~
': " " . ' . ' . ' ' " " ' ' " " ' i ' " �9 . - , ] , . . , - . " , - , .
A/epk,,'os/am
Nephro#om
Abb. 6a--d, Ausbildung der Nephrostome in der ventlaler~ (a) und ersten dorsalen (b) Anlagenfolge. c I~ephro~tomanlage in der zweiten dorsalen Schicht ohne C61omverbindung. d In cler dritten Aa-
lagenserie kein Nephrostom angelegt
W~hrend das ventrale Nephrostom mi~ dem Halsstfiek der Bowman schen Kapsel zusammenl/~uft und beide fiber den gemcinsamen Ab schnit~0 TI in das dicke Tubulusstfick TII mfinden (Abb. 6a), 1/~uft das

EntwicMung des Opisthonephros bei Triturus alpestris 25
dorsale Nephrostom an der Ventralseite der Bowmansehen Kapsel vor- bei und geht direkt in den Absehnitt TI fiber (Abb. 6b). Ausnahms- weise kommt es vor, dag in der ventralen Anlagenfolge einzelne Ka- nglehen ohne Nephrostom anftreten. Besonders im eaudalen Bereieh k6nnen Nierentriehter Iehlen. IIier liegen die Anlagen so welt im Kloa- kenbereieh, dab sie nur sehr sehwer eine Verbindung zum C61om auf- nehmen k6nnten. Bei den Kanglehen der zweiten dorsalen Sehieht sind meist eine Nephrostomanlage (Abb. 6e) und nur gelegentlieh ein Nephrostom ausgebildet. In der dritten dorsalen Sehieht sind keine Nephrostome mehr vorhanden (Abb. 6 d). Alle Nephrostomanlagen tragen bereits Wimperfahnen, bevor der Durehbrueh zum C61om stattfindet.
3. Tubulusepithel Die Wand eines Nierenkan~lchens besteht aus einem einsehichtigen
Epithel, das auf einer Basalmembran sitzt und das in den histologisch unterseheidbaren Abschnitten yon wechselnder Besehaffenhei~ ist. Der Form naeh tr i t t es als kubisehes Epithel auf, das einerseits bis zu einem Plattenepithel abgellaeht sein kann, andererseits bis an ein niedrigpris- matisehes Epithel heranreieht. Es erhebt sich die Frage, wie die Diffe- renzierung des Kan~tlehenepithds abl~uit. In der Reihenfolge des Kan~lehenverlaufs lassen sieh sechs Absehnitte (TI--TVI) unterseheiden (Abb. 7b, 39--45). Das Teilstfiek TV gliedert sieh h~ufig in zwei Ab- sehnitte (Abb. 7 b, 4 3 - - ~ ) .
Im folgenden wird die Ausbildung des Epithels fiir die einzelnen Absehnitte in der Reihenfolge des Kan~lehenverlaufes besehrieben. Um eine m5gliehst gute Ubersicht zu bekommen, fiihre ieh aus dem ganzen Nierenbereieh Beispiele an, vorwiegend yore ersten, neunten, siebzehn- ten, ffinfundzwanzigsten und zweiunddreigigsten Nierenkan~lehen. Zu Beginn der Kan~lehenbildung kann man das Epi~hel des ersten Ab- sehnittes nur sieher dureh seine Lagebeziehung zum sich gleiehzeitig differenzierenden Nierenk6rperchen yon den Zellen anderer Teilstiieke unterscheiden. An einem Quersehnitt ist ein Lumen zu erkennen, doeh sind noeh keine Cilien ausgebfldet (Abb. 7a, 7). Die gro6en, l~tnglich- ovalen Kerne liegen dieht beieinander, mit ihrer L~ngsaehse senkreeht zum Kan~lehenlumen. Bei einer etwas/~lteren Anlage beginnt das Epi- thel sieh abzuflaehen (Abb. 7a, 10). In dieser Difierenzierungsphase treten zum ersten Male Cilien auf. Mit ihrem Erseheinen wird wie bei den Flimmerzellen des Nephrostoms und des Halsstiiekes an der api- kalen Seite der Zellen ein Plasmasaum siehtbar, und die Kerne runden sieh ab, doeh liegen sie immer sehr dieht beieinander. Im Laufe der weiteren Entwieklung versehiebt sieh die Kernplasmarelation zugun- sten des Cytoplasmas. Die Kerne sind lgnglieh-ova], passen sieh in ihrer Form der Rundung des Lumens an und stehen jetzt mit ihrer kurzen Aehse senkreeht zu ihm (Abb. 7b, 20).
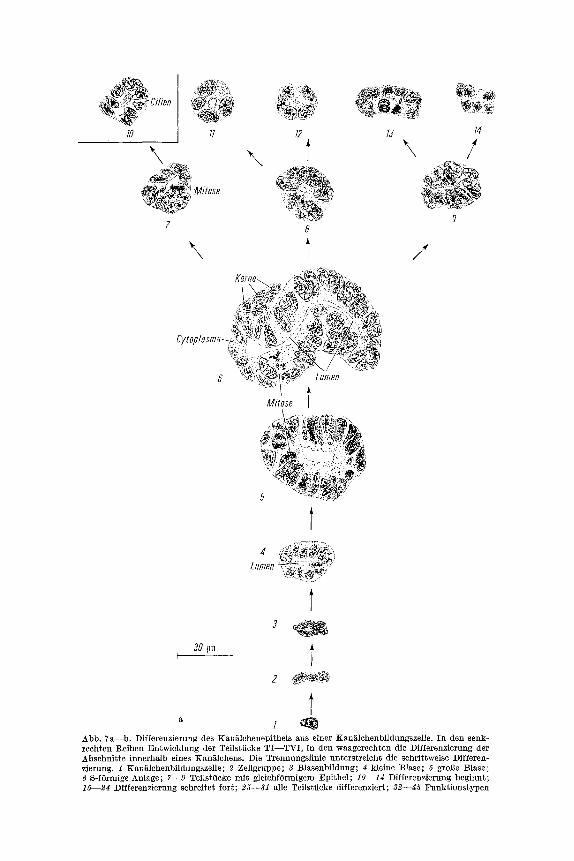
1o
?
/1
Mitose
\
/2
\ l
8
/3
\ /4
/
9
/
30 ~m
Mltose ]
t
t
, l
t Abb. 7a--b. Differenzierung des Kan~lchenepithels aus einer Kan~lehenbildungszelle. In den senk- rechten l~eihen Entwieklung der Teilstticke TI- -TVI , in den waagereehten die Differenzierung der Absehnitte innerhalb eines KanE~lchens. Die Trennungslinie unterstreieht die schrittweise Differen- zierung. 1 Kan~ilehenbildangszelle; 2 Zellgruppe; 3 Blasenbildung; 4 kleine Blase; 5 gro•e Blase; 6 S-f6rmige Anl~ge; 7--9 Teitstticke mit gleiehf0rmigem Epithel; 10--1d Differenzierung beginnt; 15--Z4 Differenzierung schreitef~ fort; 25--31 a11e Teilstiicke dffferenzier~; 32--g5 ~"unktionstypea
R. L~n~A~: Entwicklung des Opisthonephros bei Triturus alpestris 27
T$ T~7 T~ TZI 7 T_V z T~, r s
M~krowlk "~Ng'~> 40 41 42 43 44 45
32
l l l l I
7_5
2#
15
28 27 2~
+ . ~ Mikr~wlh - - CHien
21 22 23
.... ~ , , Mikrovllll . . . . . .
16 17
s
29 30 31
18
\ l / %
24
19
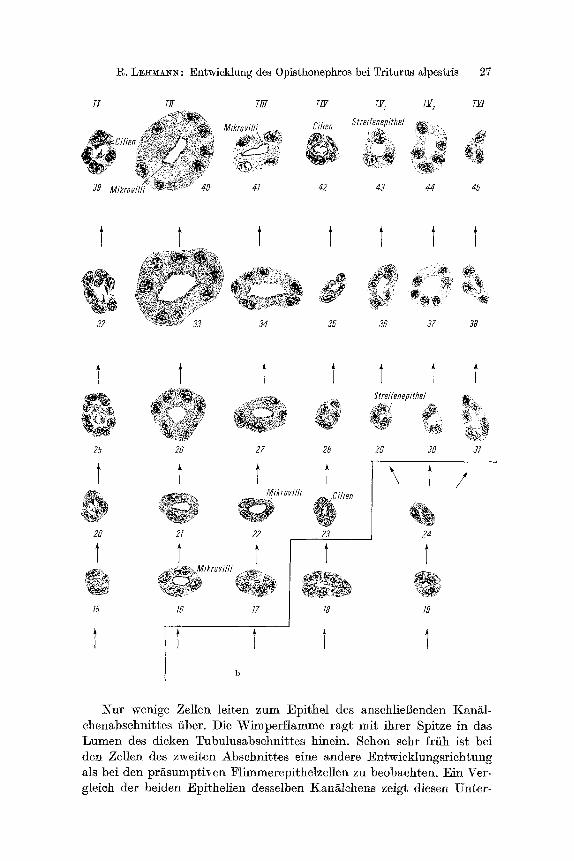
Nur wenige Zellen leiten zum Epithel des anschliel3enden Kangl- ehenabsehnittes fiber. Die Wimperflamme ragt mit ihrer Spitze in das Lumen des dieken Tubulusabsehnittes hinein. Sehon sehr frfih ist bei den Zellen des zweiten Abschnittes eine andere Entwieklungsriehtung als bei den prgsumptiven Flimmerepithelzellen zu beobaehten. Ein Ver- gleich der beiden Epithelien desselben Kanglehens zeigt diesen Unter-

28 R.L~MA~:
schied (Abb. 7a, 10--11). Die Zellkerne sind groB und abgerundet, ihre Lage ist aufgeloekerter als bei TI, und es ist mehr Cytoplasma vorhan- den. Im weiteren Verlauf wachsen die Zellen durch Vermehrung des Zytoplasmas, und die erste Andeutung der Mikrovilli wird sichtbar (Abb. 7). Ansehliel3end setzt eine enorme VergrSBerung der Zellen dureh Vervielfaehung des Zytoplasmas ein. Gleichzeitig wird durch Zunahme der Mikrovilli die lumenseitige Zelloberfl/~che stark vergrSBert. Das Epithel dieses Tubulusabschnittes ist nieht nur dureh auff/~lhges Waehs- turn und gut histologisch siehtbare Differenzierung gekennzeiehnet, sondern es tr i t t im Epithelverband neben der ErhShung seiner Zellzahl in der L~ngsriehtung des Kan~lehens aueh eine Vervielfaehung der Zellen pro Quersehnitt auf. Die Fo]ge ist sowohl eine Diekenzunahme als aueh eine VergrSBerung der lichten Weite des Tubulus.
Im distalen Verlauf des Nierenkan/s wird das Epithel wieder etwas flacher. Die Entwieklung dieses Abschnittes (TIII) ist mit der des vorigen (TII) am Anfang seiner Differenzierung vergleiehbar (Abb. 7 b, 21--22). W/~hrend sich die Zellen des Teilstfickes TI I sehr stark ver- grSl~ern, nimmt diese Entwieklung mit dem allm/~hlichen Ubergang zum Abschnitt T I I I ab. Zwar zeichnen sieh diese Zellen auch dureh Mikrovilli aus, aber sie werden nicht so hoeh wie bei TII. Ebenfalls ist dieses Tubulusstfiek aus einem flaeheren Epithel aufgebaut als TII. Beiden Epithelien (TII, TIII) gemeinsam ist eine polare Differenzie- rung ihrer Zellen. Die Kerne liegen immer im basalen Tell der Zelle, w/~hrend der apikale Tell durch die Ausbildung yon Mikrovilli ausge- zeiehnet ist.
Der n/~ehste Abschnitt ist wieder ein mit Cilien versehenes Teil- stfiek. Erst relativ sp/it, wenn die voranliegenden Tubulusabschnitte histologiseh schon gut zu unterscheiden sind, beginnt die Cihendiffe- renzierung (Abb. 7b, 23). Die Kerne gehen yon einer 1/s in eine rundliche Form fiber (Abb. 7b, 28). Aueh wenn die Wimper- flammen roll ausgebfldet sind, behalten die Kerne die rundliehe Ge- stalt zun~chst bei. Mit fortsehreitender Entwieklung tr i t t eine Form- ver~nderung der Kerne zu einer l/s Gestalt auf (Abb. 7b, 35 und 42). Wie die Kerne des Abschnittes TI stehen sie ebenfalls mit ihrer kurzen Aehse senkrecht zum Kan/~lchenlumen.
Der folgende Kan/s entwiekelt sieh aus einem /~hn- lichen Epithel wie alle anderen Teilst/icke des ~qephrons. Auf Grund ihrer gegenw~rtigen Lage und im Hinbliek auf ihre kfiTfftige Differen- zierung lassen sieh sehon zu einem relativ frfihen Entwieklungszeit- punkt zwei Teilstficke unterscheiden: der Abschnitt, in dem sieh das sog. Streifenepithel ausbildet~ und der dorsolaterale Bogen. Die weitere Differenzierung dieser beiden Teilstficke soll nacheinander besproehen werden.

Entwicklung des Opisthonephros bei Triturus alpest.ris 29
Die Zellen werden zytoplasmareieher, und die Kerne runden sieh ab. Spgter kommen sie in dem apikalen Teil der Zellen zu liegen und ragen oft bis in das Lumen vor. Mit dieser Ver~nderung tr i t t aueh im Zytoplasma eine senkreeht zur Oberflgehe gestreifte Struktur auf, die bei glteren Nephronen sehr deutlieh hervortritt . Der Durehmesser und die lichte Weite des vom ,,Streifenepithel" gebildeten Teilstiiekes weeh- seln im Verlauf des Kanglehenabsehnittes.
Zu einem etwas anders gearteten Epithel differenziert sieh der bei alien Kanglehen ausgebildete dorsolaterale Bogen, in den im eandalen Teil des Opisthonephros die dorsal liegenden Kanglehen einmfinden (L~MA~S, 1964, Abb. 7). Die Differenzierung ffihrt zu einem kubi- schen Epifhel, dessert runde Kerne etwa die ]3reite der Zellen einneh- men. Das fiir den vorangehenden Absehnitt typisehe Streifenmuster fehlt. Das ngehste kurze Teilstiiek stellt die Verbindung zum primgren H~rnleiter her.
4. Miindung in den primiiren Harnleiter
Die Einmfindung des kfinftigen Nierenkanglchens in den primgren ILarnMter wird mit der Anlage der Nephroblastblase schon vorbereitet. Sie liegt so fiber dem Ableitungsrohr, dab die Zellen ihrer Ventralseite in engen Kontakt mit den Zellen des primgren Ha.rnleiters kommen (L~IMA~I% 1963, Abb. 2). Anschliegend verschmelzen beide Epithelien miteinander. Die Zellen der auswaehsenden Nephroblastblase unter- scheiden sich deutlieh yon denen des primgren tIarnleiters. Dazwischen bildet sieh eine TJbergangszone aus mit Zellen, deren Kerne in bezug auf die der Kanglchenanlage weiter und im Verhgltnis zu denen des primgren Harnleiters weniger differenziert sind. Das sehr feine, spalt- f6rmige Lumen setzt sich noch nicht bis in das Ableitungsrohr fort. Erst bei der etwas glteren Anlage kommunizieren beide Tefle mitein- ander. Der anfangs sehr weitlumig angelegte Einmtindungsteil wird bei den meisten Kanglchen zu einem engen Rohr (Abb. 4). Nur die crania- len Nephrone haben hgufig ein weitlumiges Miindungsstiick. Mit zu- nehmender Entwicklung gleichen sich die Epithelien des primgren Harn- leiters und des Miindungsabschnittes so weir an, dab beide nicht yon- einander unterschieden werden kannen.
Wie sind die Differenzierung des Nierenk6rperchens, des Einmiin- dungsteils und des Nephrostoms zeitlieh aufeinander abgestimmt ? Der kontinuierlieh ablaufende Differenzierungsprozeg lggt sieh in drei Pha- sen unterteilen: 1. Mtindung in den primgren tIarnleiter noeh nieht vorhanden, Nierenk6rperehen angelegt, kein Nephrostom, 2. Miindung ausgebildet, Glomerulus durehbluteL Nephrostomanlage mit Flimmer- epithel ohne Verbindung zum C61om, 3. Miindung, Nierenk6rperehen und Nephrostom vorhanden.

30 R. LSH~A~S:
5. Dorsolateraler Bogen
Die Analyse s/~mtlicher Kan/~lehenanlagen eines 41--42 m m langen Tieres zeigt, dab alle Nierenkani~lehen ihren Endabschni t t als dorso- lateralen Bogen anlegen (LEHMAN~, 1964, Tabelle 1, Abb. 3). Aber nu t in der caudalen I-Ii~lfte der ventralen Kan/~lehenfolge und bei den Kan/ilchen der ersten und zweiten dorsalen Schicht kann der dorsolaterale Bogen als gemeinsames Sammelrohr dienen.
6~ Di]/erenzierungs/olge innerhalb ~ines KanSlchens Die in Abb. 7a und b zusammengesteliten Differenzierungsfolgen ~i~4 nach
dem Gesiehtspunk~ ~usgewii.Mt, einen m6giiclas~ fiir ~tle Ax~t~gen zutreffende~ Aussch~itt aus dem Entwicklungsgeschehen widerzuspiegeln. Selbstverstiindlich gibt es yon den clargestellten Kaniilchenabschnitten Abweichungen innerhalb der m6glichen Reaktionsbreite. Zum Tell mag d~e Ursache dafiir darin liegen, dab die Nierenkani~lchen sich nicht gleichzeitig an einer eng umschriebenen Stelle, son- dern naeheinander fast fiber die ganze L~inge der Bauchh6hle entwickeln und je nach Alter und Ort in vergnderter Umgebung. Die histologische Entwicklungsreihe ist das Ergebnis yon Untersuchungen an l~ckenlosen Schnittserien durch etwa 1000 A~lagen verschiedenen Entwicklungsgrades, die sich aus fiber 5300 Einzel- sehnitten zusammensetzen. In je einer waagerechten Reihe (Abb. 7a und b) sind Quer- und Liingsschnitte dureh verschiedene Regionen eines Kanglchens, dann in den senkrechten Reihen durch die gleichen Abschnitte verschieden welt diffe- renzierter Kanglchen angeordnet. Die Pfei[e geben die Differenzierungsfolge der Kan~ichenahsehnitte mit zunehmendem Alter der Nephrone an.
Auf Grund d~r L~gebeziehung zum primgre~ H~rnleiter, der Ahn- lichkeit in der Kerns t ruktur , der Ani~rbbarkeit und des geringen Zyto- plasmagehaltes der gellen, die bereits zu einer Gruppe vereinigt sind (2, 3), lal~t sich die Entwieklung eines Nierenkan/ilehens auf einzelne Zellen zurtiekf/ihren (1, 2). Ob nut eine Zelle am Anfang steht oder ob mehrere verstreutliegende Zellen an der Bildung einer Kanglehenan- lage beteiligt sind, kann zur Zeit noch nieht entsehieden werden. Aus einer kleinen Gruppe (2) entsteht ein Zellhaufen (3), der dutch Lumen- bildung und Zellvermehrung zu einem B1/~sehen heranw/~ehst (4). Durch eine enorme Zunahme der Zellzahl entwiekelt sieh eine groge Nephro- blastb[ase (5), aus der eine S-fSrmige Anlage ents teht (6). Indem sie zum Kang[chen ~usw/~ehst, iassen sich im Hinblick auf die kiinftigen KanMehenabsehnit te mindestens drei Teilstfieke unterseheiden: das Halsstiiek (7), der dieke Tubulusabsehnit t (8) und das Verbindungs- stiick aura prim/~ren Harnlei ter (9). Von hieraus t r i t t mit fortsehreiten- dem Kan~ilehenalter eine zunehmende Spezialisierung der Nephronab- sehnitte ein.
Als erste bleibende Differenzierung t r i t t das Flimmerepithel im Ilalssgfick auf (10), obgleieh es bis zum voll entwiekelten Kan/~lehen einer stetigen Formvergnderung unterworfen ist (I5, 20, 25, 32, 39). Die Ab8chnitte vom Typ I I und I I I sind sehr /~hnlieh s trukturier t (11, 12),

Entwieklung des Opisthonephros bei Triturus ~]pestris 31
doch ist das Teilstiick I I in seiner Entwicklung etwas voraus, wie z.B. die Ausbildung der Mikrovilh zeigt (16), die im Tubulusabschnitt I I I erst spgter auftreten (22). Die weitere Entwicklung dieser Absehnitte ist sowohl durch eine VergrSgerung der Zellzahl pro Querschnitt als aueh dutch eine GrSBenzunahme der einzelnen Zellen gekennzeiehnet (21, 26, 33, 40; 22, 27, 3r 41). Das Teilsttick IV, ebenfalls mit Cilien versehen (23, 28, 35, 42), ist in der Entwicklungsphase, in der der Typ I seine Cilien ausbildet, noch ein mit einem schmalen Spalt versehenes wimperloses Rohr (18). Der Abschnitt V (14, 19, 24) gliedert sich in drei Abschnitte: 1. ein englumiges Teilstfiek, das von einem ,,Streifen- epithel" gebildet wh-cl (29, 36, 43), 2. einen weith~migeren Kan~]ehen- abschnitt, der den dorsolateralen Bogen bildet (30, 37, 44), und 3. ein besonders bei einem /klteren Kan~lehen sehr enges Verbindungsstiiek, das im eaudalen Nierenbereieh als Sammelrohr dient (31, 38, 45).
Aus der Gegenfiberstellung verschieden alter Kan&lehen kann man erkennen, dab die Entwicklung eines Nephrons nieht in alien Teilen gleiehzeitig erfolgt. Zuerst entsteht das Flimmerepithel im Halssttick (10), dann treten dig Mikrovilli im Aniangstefl des dieken Tubulus anf (16), sp/~ter erscheinen sie aueh im ansehlieBenden Abschnitt (22), und der Typ IV ist ]etzt mit Cilien versehen (23). Zuletzt bfldet sigh das ,,Streifenepithel" aus (29), und gleichzeitig sondern sieh die /ibrigen beiden AbsGhnitte (30, 31). Die Differenzierung der fflr Gin Nieren- kan~lehen typisehen Zellen findet von proximal naeh distal statt (10, 16, 22, 23, 29, 30, 31).
IV. Herkun/t dorsaler Anlagen Nach der bisherigen Betrachtung verschmelzen die neu angelegten
Tubuli der dorsalen Schichten mit dem unter ihnen liegenden Ab- schnitt eines dorsolateralen Bogens. Naeh Gl~.Xu (1932) sollen neue Tubuli durch Sprossung aus den zuerst angelegten Kan~lchen entste- hen. Die Untersuchungen ergaben, da[~ zu dem Zeitpunkt, da noch nicht alle ventralen Kans differenziert bind, dorsal von ihnen wei- tere Nephroblastblasen auftreten. Sie liegen night in einem gr6geren Abstand fiber den ventralen Tubuli, sondern etwas in die Kan/~lehen- windungen der Sehicht, die unter ihnen liegt, eingesenkt (Abb. 2). Den- noeh stehen die dorsalen Nephroblastblasen an keiner Stelle mit ir- gendeinem Absehnitt der bereits ausgebfldeten Kanglchen in Beziehung. Sp/~ter nehmen sie mit dem ihnen zuni~ehst liegenden dorsolateralen Schenkel Kontakt auf; er ist der erste siehtbare Sehritt zum sp~teren Zusammentreffen der Tubuli. Naeh der Verschmelznng beider Epithe- lien zu einem einheitlichen Zellverband vereinigen sich die Lumen bei- der Kanglehen. Der Entwicklungsgang aller dorsalen und ventralen Kans ist prinzipiell der gleiehe. Es besteht kein Zweifel, dab alle
32 R. LEHMANN:
n e u a n g e l e g t e n d o r s a l e n A n l a g e n i n b e r e i t s v o r h a n d e n e K a n a l c h e n a b -
s c h n i t t e e i n w a e h s e n . N i e m a l s f i n d e t d e r u m g e k e h r t e P r o z e g s t a r t , d a b
a u s S p r o s s u n g s z o n e n K a n / ~ l e h e n a u s w a e h s e n .
D. Defekte am intermedi~iren Mesoderm
I. De/ektsetzung Im Bereich des 8.--14. metotischen Segments werden rechteckige Gewebe-
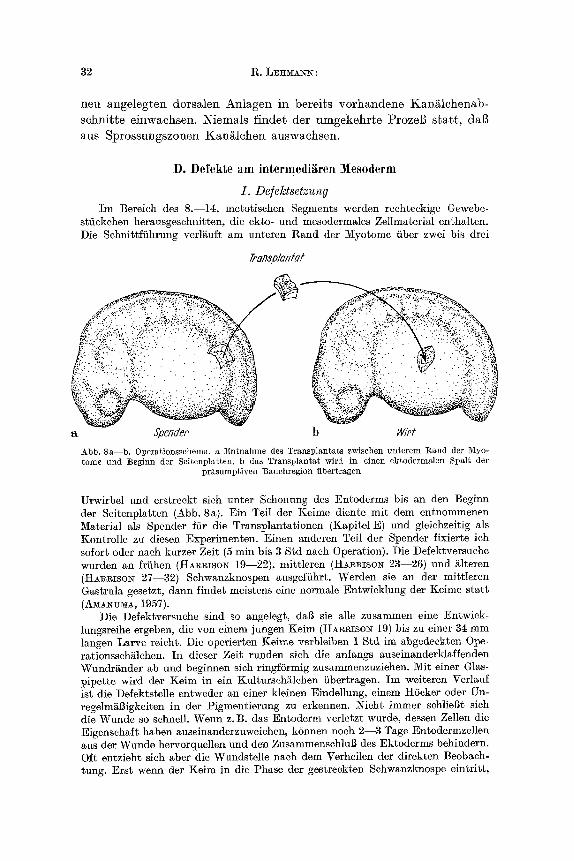
stfickchen herausgesehnitten, die ekto- und mesodermales Zellmaterial enthaiten. Die Sehnittfi ihrung verl~uft am unteren Rand der Myotome fiber zwei bis drei
~ansplazlar
a Spender b g/t~t
Abb. 8a--b. Operationsschelna. a Entnahme des TranspIantats zwisehen unterem Rand der 3'[yo- tome und Beginn der Seitenplatten, b das Transplantat wird in einen ektodermalen Spalt der
Dr~sumptiven Bauchregion abertragen
Urwirbel und erstreckt sich unter Schonung des Entoderms bis an den Beginn der Seitenplat ten (Abb. 8a). E in Teil der Keime diente mt t dem entnommenen Material als Spender fiir die Transplantat ionen (Kapitel E) und gleiehzeitig als Kontrolle zu diesen Experimenten. Einen anderen Teil der Spender fixierte ich sofort oder nach kurzer Zeit (5 rain bis 3 Std naeh Operation). Die Defektversuehe wurden an friihen (HAgmso~ 19--22), mit t leren (H~A~RIso~ 23--26) und ~lteren (HAgmsoN 27--32) Sehwanzknospen ausgeffihrt. Werden sie an der mit t leren Gastrula gesetzt, dann findet meistens eine normMe Entwicklung der Keime s ta t t (A~IANIr~IA, 1957).
Die Defektversuche sind so angelegt, dab sie alle zusammen eine Entwiek- lungsreihe ergeben, die yon einem jungen Keim ( H A ~ I s o ~ 19) bis zu einer 34 mm ]angen Larve reieht. Die operierten Keime verbleiben 1 Std im abgedeekten Ope- rationsschiilchen. In dieser Zeit runden sich die anfangs auseinanderklaffenden Wundr~inder ab und beginnen slch ringfSrmig zusammenzuziehen. Mit einer Glas- pipet te wird tier Keim in ein Kulturschalehen fibertragen. I m weiteren Verlauf ist die Defektstelle entweder an einer kleinen Eindellung, einem HScker oder Un- regelmi~Bigkeiten in der Pigmentierung zu erkennen. Nicht immer schlieBt sich die Wunde so schnell. Wenn z.B. das Entoderm verletzt wurde, dessen Zellen die Eigenschaft haben auseinanderzuweichen, k f n n e n noch 2--3 Tage Entodermzellen aus der Wunde hervorquellen und den ZusammensehluB des Ektoderms behindern. Oft entzieht sich aber die Wundstelle nach dem Verheilen der direkten Beobach- tung. Ers t wenn der Keim in die Phase der gestreekten Sehwanzknospe eintri t t ,

Entwicklung des Opisthonephros bei Triturus Mpestris 33
ist der hinter der Operationsstelle liegende K6rperabschnitt h~ilffig zur Defekt- seite hin ~bgebogen. Besonders eindeutig abet gibt sieh die Defektregion durch die ehar~kteristische Anordnung der Chrom~tophoren zu erkennen, die zu diesem Zeitpunkt der Entwicklung dutch ihr Pigment sichtbar werden. Die sehwarzen Farbstofftri~ger liegen bei ihrem Erseheinen ~uf jeder Seite der L~rve in zwei Rei- hen in der L~ngsrichtung des KSrpers angeordnet (Abb. 9~). Normalerweise liegt zwischen beiden Streifen eine in dieser En~wieklungsphase chromatophorenfreie Zone. An der Defektstelle ~ber gehen die Farbstofftriiger beider Reihen inein~nder fiber (Abb. 9a). Mit zunehmendem Alter der L~rve ver~ndern die iKelanophoren
Abb. 9 a--b . a AD 880, 9--10 ram. Im Defektbereich gehen der dorsMe und der laterale Pigmentstreifen ineinander fiber, b AD 880, 21--22 ram. Dis scharfe A.bgrenzung der Pigmentb~nder verwiseht.
u 7 • 65 ventraler Pigmentstreifen; 66 dorsMer Figmentstreifen; 67 Beriihrungszone der Pigmentstreifen
~hren Ort ~ sicher we~rden aueh neue r si~htbar -- und die soharfe Abgrenzung der Pigmentbinder yon ihrer Umgebung verwischt (Abb. 9b). Bis dahin aber bietet die Berfihrungsstelle der Pigmentstreifen, unter der der meso- dermale Defekt liegt, eine ausgezeichnete Kontrollm6glichkeit, den Oloerations- bereich zu lokalisieren.
II . Folffen der De]e/ctsetzung
1. Fehlende Organteile Einen ~berblick fiber Teile yon Organen, Organanlagen und Ge-
weben sowie Zellgruppen und Einzelzellen, die infolge der Defektset- zung nicht vorhanden oder nieht typisch differenziert ausgebildet sind, gibt Tabe]le 1.
Mit groBer Regelms feh]en in der Nierenregion e~nzelne Kan~lehenanl~gen. Der K6rpergr6Be der Tiere entspreehend sind es Kan~lehenbildungszellen, Zellgrulopen , ~ephroblastblasen oder Kan~Iehen, die sieh im Defektbereieh nieht entwiekelt haben (Abb. 10).
3 Wilhelm ~oux ' Arch. Entwield.-Meeh. Org., Bd. 158

34 ~. LEHMANN :
T~belle 1. Folgen der De/el~texperimente
7--14 rim1 ] 5 - - 3 4 m m
:Niere pr im~rer Musku- Ke im- :Niere pr im~rer i~usku- Ke im- Harn- la tur zenen I t a rn - la tur zellen leiter und leiter und
Gonaden Gonaden
Defekt 17 15 2 15 21 14 - - 13
Atypisch differen- ziert - - 12 (5) 6 2 - - 6 (3) 6 1
Kein Defekt - - 3 17 2 - - 5 16 4
Aul3erhMb der Gonadenregion - - - - - - 1 - - - - - - 3
Nicht zu beurteilen 8 - - - - 5 1 - - - - 1
( ) = zusatzlich Defekt.
A n der Musku l~ tu r , die sich y o n den M y o t o m e n ab le i t e t , s ind n u r
ger inge A b w e i c h u n g e n v o n d e r N o r m M e n t w i c k l u n g zu b e o b a c h t e n .
Meis tons s t i m m t die oper ie r t e Sei te m i t de r n i c h t o p e r i e r t e n hinsichr
Abb. 10. A D 821, 21--22 m m . Querschni t t du tch die Nierenregion im Defektbereich. Auf der operierten Seite s ind keine Nierentubul i entwickelt . Vergr. 100 • Objek t iv 6,3 x . 70 5Tiere
l ich der M u s k u l a t u r i iberein . I n e in igen F~ l l en is t die M u s k u l a t u r ver -
l ager t . F e h l e n d e s M a t e r i a l is t n u r be i j u n g e n L a r v e n u n d E m b r y o n e n
fes tzus te l len . Sofe rn de r D e f e k t im p r ~ s u m p t i v e n G o n a d e n b e r e i e h gese t z t wurde ,
f eh len in dieser l~egion Urgesch lech t sze l l en , ode r es t r i t t ke ine t yp i s ehe D i f f e r enz i e rung de r G o n a d e auf. F e r n e r s ind Ms Fo lge der D e f e k t e
die V. p o r t a e renis u n d k le inere Blutgefi~ge sowie das Se i t en l i n i eno rgan
a t y p i s c h en twieke l t . Alle D e f e k t e r s c h e i n u n g e n t r e t e n in bezug zur K6rper l /~nge in ver -
g l e i chba re r F o r m sowohl be i j u n g e n Ms auch bei s L a r v e n auf
( A u s n a h m e : Musku la tu r ) .

Entwieklung des Opisthonepkros bei Triturus Mpestris 35
2. Beein/lussung des Di//erenzierungsgeJiilles Wh'd durch den Eingriff in das intermedi/~re Mesoderm das eranio-
caudale Differenzierungsgef/~lle beeinfluBt ? Um diese Frage zu beant- worten, m6ehte ieh die Kan/~lehenanlagen der intakten Seite mit denen auf der Defektseite vergleichen. Vor der Operationsstelle sind reehte und linke Seite bis aut geringffigige Abweiehungen, die aueh in der Normalentwieklung auftreten, gleieh weir differenziert. Hinter dem Be- reich, in dem der Defekt auftritt, bleiben die Kan~lehenanlagen gegen- fiber der intakten K6rperseite in ihrer Entwieklung zurfiek (Abb. 11).
Abb. 11. AD 43, 22--23 mm. Querschnitt dutch den Defektbereich. AIs Folge des Defektes fehlt links der prim~re ]-Iarnleiter. Die Nierenanlage is~ im Vergleich zur in tak ten Seite zurfickgeblieben.
Vergr. 240 • Objekt iv 16 x . 58 Gonade
Doch nicht immer tr i t t dieser Befnnd Mar hervor. In allen Beispie- len, in denen der Untersehied in der Differenzierung nur sehr geI~ng ist, ist der lorim/~re Harnleiter in seiner ganzen L/~nge ausgebildet. Da- neben treten aueh F/tlle auf, bei denen der I-Iarnleiter hinter dem De- fekt fehlt. Wird hier der Differenzierungsgrad der Nierenanlagen beider K6rperseiten zum Vergleich herangezogen, so ist der Unterschied in ihrer Entwieklung sehr deutlich (Abb. 12). Auf der rechten, intakten Seite entwiekeln sieh die Anlagen in typiseher Weise dem Differenzie- rungsgefs entspreehend. W/~hrend cranial ausgebildete Nierenkan/~l- then liegen, nimmt ihr Differenzierungsgrad nach hinten ab; es folgen groBe Nephroblastblasen, die sehlieBlich caudal am kleinsten sind. Auf der oloerierten Seite (links) haben die Kan/tlchen vor dem Defektbe- reich den gleichen Differenzierungsgrad wie auf der Gegenseite, der zn Beginn der Defektregion sprunghaft abf/~llt. Den Zellgruppen stehen anf der intakten Seite Kan~lchen gegenfiber, die den Unterschied in
3*

36 R. LE~MA~:
der Entwieklung der Anlagen beider Seiten hervorheben. Anschlie~end steigt der Differenzierungsgrad auf der Defektseite wieder an, darauf f~llt er erneut ab und nimmt gegen Ende der Defektregion abermals zu. Danaeh stimmen die Anlagen der rechten und linken Seite in ihrer EntwieklungshShe wieder fiberein.
Der prim~re Harnleiter zieht am Anfang der Defektregion naeh lateral und versehmilzt mit der Epidermis, die sieh an dieser Stelle
eingesenkt hat. Dahinter ist zu- nrz~n#re ttarnle/ler
Kan#lchen-
\
9~
~'nsen kun g k /~ Epidermis ~
,'~'Zellgruppen
o ~ o
//arn/e/;,-t:,r o AlephroblaM -0
_ (~bla~en
ii o o //)7/~e mc/~ /e
ven/ra/e An/agezfo/#e Abb. 12. A D 77~-, 15--16 m m . Aussehni t t aus der vent ra len Anlagenfolge. Differenzierung der Kani i lehenanlagen m i t und ohne pr im~ren
I Iarn le i te r
n~tchst kein Harnleiter ausgebildet, dann folgt ein kurzer AbschnitL und darauf fehlt er wieder. Vom Ende der Defektregion ab ist der prim/~re Harnleiter regelms angelegt. Es f~tllt auf, dab die Kans gerade dann einen h6heren Diffe- renzierungsgrad erreichen, wenn sie in der Naehbarschaft des prim/~ren ttarnleiters ]iegen.
Diese Befunde zeigen, dab eine St6rung des Zellkomplexes, der anch die Nierenbildnngszellen enth~Llt, mit einer verminderten Differenzierungs- leistung der Kan~lehenanlagen be- antwortet wird, besonders dann, wenn der prim~re Harnleiter sich nur unvollst~ndig ausbildet. Ein Ein- fluB des prim~tren Harnleiters auf die Differenzierung der Niere wird aueh yon NIEUKOOP (1948) ffir Ambly- stoma und yon KoTA~I (1962) ffir Triturus pyrrhogaster angegeben.
3. Regenerative Prozesse
Durch die Defektsetzung entsteht im Ekto- und Mesoderm eine Materia]lficke. Das vorma]s gegen die AuBenwelt abgegrenzte Zellge- ffige steht an der Operationsste]le unmittelbar mit der Kulturf]iissig- keit in Verbindung. Die entodermalen Zellen quellen naeh der Entfer- nung des Ektoderms und des darunterliegenden intermedigren Meso- derms aus der Wunde hervor. Bereits 1 Std nach der Operation wird der WundverschluB siehtbar. Innerhalb yon 3 Tagen versehlieBt das Ektoderm die Wunde vollstgndig. ])as AusmaB der Defektsetzung l~gt sieh erst erkennen, wenn sich die embryonalen Zellen in der Umgebung des Defektes differenziert haben.

Entwicklung des Opisthonephros bei Triturus alpestris 37
In einigen F~llen verklebt der primgre Harnleiter mit dem Ekto- derm. Die Verschmelzung kann so weir gehen, da~ eine neue Ansmiin- dung des Vornierenganges entsteht. Anstatt fiber die Kloake tr i t t der prim~re Harnleiter lateral dutch die I-Iaut nach au~en (Abb. 13). Auf Grund grol~er Xhnlichkeit bietet das Hautepithel far dan prim~ren Harnleiter ebenso ein Reaktionsgewebe wie das Epithel der Kloake. Ftir die Regeneration des primgren Harnleiters treten drei MSglichkei- ten auf: 1. er regeneriert vollst~ndig; 2. hinter der Defektstelle bilden sich einzelne unabhs Teilstiieke; 3. die Regeneration unterbleibt; er miindet an einer atypischen Stelle aus oder endet blind.
Abb. 13. AD 780, 9--10 mm. Querschnitt dutch die Defektregion (KSrpermitte). Auf der Defekt- seite miindet der prim~ire ttarnleiter lateral (rechts). Vergr. 230 • Objektiv 16 • 68 Darln;
71 Mtindung des prim~ren Iiarnleiters
Nur nach ungewdhnlich grol3en Zellverlusten des Myotommaterials ist bei einer glteren Larve noch ein Mangel an Muskelzellen zu bemer- ken. Im allgemeinen ist die Muskulatur so welt regeneriert, da~ gegen- fiber der intakten KSrperseite kein Untersehied festzustellen ist. Da- gegen ist ein Verlnst des Nierengewebes nach wie vor sichtbar, obgleich sich ein Ausgleieh der ursprtinglichen Materialliicke bemerkbar macht, indem sich die Iqierenkanglchen vor und hinter der Operationsstelle mit ihren Tuhulusschleifen einander nghern. Bei alteren Larven wird die in bezug auf das ganze Nierenorgan nur geringffigige Lticke vollst~ndig ausgeglichen, falls nicht dureh eine zus~tzliche atypische Differenzie- rung eine St6rung der Regeneration eintritt (z.B. bindegewebige Wucherungen in der defekten Nierenregion).
Die Regeneration setzt erst ein, nachdem sich die embryonalen Zellen g e m ~ ihrer prospektiven Bedeutung ganz oder teilweise diffe- renziert haben. Zuerst regeneriert das Ektoderm, dann die Muskulatur und erst sehr spgt das Nierengewebe. W/~hrend die Muskelfasern durch Zuwachs die Gewebslfieke erg/~nzen, wird in der Niere der Defekt dutch

38 ~.LEHMANN:
eine allm~hliche Verlagerung der Kan/~lchen w~hrend ihres Wachs tums ausgeglichen.
I n enger Verbindung mit den regenerat iven Prozessen steht die Wiederherstellung der durch die Operation gestSrten KSrpersymmetrie. Die Zellen eines Organs bedingen durch ihr Vorhandensein ihre gegen- seitige Lage. Fehlt insbesondere mesodermales Material, kommen Chorda und Neuralrohr dicht unter das Ek tode rm zu liegen. Der KSrper bietet eine asymmetrisehe Gestalt, die mit zunehmender l~egeneration der Muskulatur ausgeglichen wird. Es kann sogar in Bereiehen, in denen Nierenbfldung nnterbleibt, das renale Areal durch Muskulatur ausge- ffillt werden. Solange die Muskulatnr noeh nicht vollst~ndig regeneriert ist, kann man die Stelle des Nierendefektes schon makroskopisch an der lebenden Larve erkennen, abgesehen yon tier typisehen Pigment- verteilnng an der Defektstelle. Unabh/~ngig davon, wieweit sieh die AusmaSe der Operation auf die Nierenkan~lehen und den prim/~ren Harnlei ter erstreeken, wird durch Regenerat ion die ffir die Individual- entwieklung festgelegte harmonische KSrpergestal t wieder hergestellt.
E. Transplantationen des intermedi~iren Mesoderms
I. Aus/i~hrung der Experimente Die Transplantationsversuche sind so angelegt, dal~ sie einerseits als Kon-
trollen zu den Defektversuchen (Kapitel D) dienen, andererseits sollen sie zeigen, in welcher Weise sieh aul~erhalb der normalen Umgebung der ZeIlkomplex des
Tabelle 2. ~/berblick iiber die Transplantationen. H.-St. =Harrison.Stadium
Entwicklungs- Entwicklungsstadium des Wirtes bei der stadium des Fixierung Spenders
~.-St. 8--15 15--2] 21--40 31--38 mm ram mm
tt.-St 19--22 6 1 4 - - H.-St. 23--26 1 16 11 7 tt.-St. 27--32 1 3 8 8
intermedi~ren Mesoderms differenziert. Die Schnittffihrung ist die gteiche wie bei den Defektversuchen (Abb. 8). Da sich die presumptive Nierenregion vom 6.--16. metotischen Segment erstreekt, mul]te alas entnommene Material auch Zellen ent- h~lten, die gem~l~ ihrer prospektiven Bedeutung Nierengewebe bilden. In den meisten F~llen wurde das isolierte Gewebestfick in die presumptive Bauchregion verpflanzt (Abb. 8 b). Zu diesen Versuchen verwandte ich Keime im Schwanzknos- penstadium, bei denen das Nierenm~terial schon determiniert ist (~ACHEMEt~,
1929). Wirt und Spender sind in den meisten F~llen gleieh ~lt, bei verschiedenem Alter betrKgt der Unterschied bis zu fiinf Stadien nach HArrisoN. Die Tabelle 2 gibt einen t:Tberblick fiber das Entwicklungsst~dium der Spender und fiber das Alter des Wirtes zum Zeitpunkt der Fixierung.

Entwicklung des Opisthonephros bei Triturus alpestris 39
W~Mt man Spender und Wirt so aus, dal~ sie yon mSglichst gegens~tzlicher FarbtSnung sind, kann man die Lage des Transplantats such nach dem Einwach- sen leicht erkennen. Meist hebt es sieh auBerdem als kleiner Buckel yon der iibrigen KSrperoberfl~ehe ab. Zur Zeit, da das Pigment in den Chromatophoren syntheti- siert und ausgebreitet ist und sie als schwarze sternfSrmige Zellen sichtbar wer- den, treten auch an der Stelle Farbstofftr~ger auf, an der das transplantierte Ge- webestfick eingewachsen ist. Diese Zellen kamen - - noeh unpigmentier~ - - mit dem Spendermaterial dort hin. Oft ordnen sich die Chromatophoren ringfSrmig an and umgrenzen so das Transplantat, oder sie sind fiber die ganze Fl~tche des implantierten Gewebes verteilt. Auf Grund der aufgeffihrten Kriterien und zahl- reicher Einzelbeobaehtungen, die sich aus der Erfahrung ergeben, kann das Trans- plantat vom Beginn bis zum Ende des Versuches eindeutig lokalisiert und beob- achtet werden.
II . DiHerenzierung der Transplantate
1. Muskulatur
Da die T ransp l an t a t e meis tens auch Zellen des ven t ra l en Myotom- randes enthie l ten , h a t sich bis auf drei Fa l le (Tabe]le 3) immer Muskula- t u r entwickel t , abgesehen yon den Larven , in denen die Differenzierung zu Muskelgewebe zum Z e i t p u n k t der F ix ie rung noeh n ieh t s ta t tgefun- den hat . Die Musku la tu r zeigt teilweise eine typ isehe Myo tomanord - nung, teilweise ver laufen einige Fase rn wel t in die Se i t enp la t t enmuskn- l a tn r hinein. Neben den Muskelfasern entwiekeln sich aueh Binde- gewebsziige, die oft ex t r em lange Zel lkerne entha] ten.
T a b e l l e 3. Di]]erenzierung yon Muskulatur, Urgeschlechtszellen und Kan~ilchenanlagen im Transplantat
Differen- Keine Noch nicht ziert differenziert
Muskulatur 58 3 5 Urgeschlechtszellen 50 10 5 Kan~lchenanlagen 60 1 5
2. Urgeschlechtszellen Zur Zeit der T r a n s p l a n t a t i o n bef inden sich im in te rmedi~ren Meso-
d e r m aueh die Zellen, deren p rospek t ive Bedeu tung es ist , sieh zu Ur- geschlechtszel len zu dJfferenzieren. U n t e r exper imente] len Bedingungen ist auch eine Differenzierung yon Urgeseh]eehtsze]len aus dem Seiten- p l a t t e n m e s o d e r m mSglieh (AsAYAMA, 1950, 1961). Sie t r e t en mi t einer Ausnahme (Tabelle 3) immer zusammen mi t Nierenan]agen auf. Mei- s tens k o m m e n sie einzeln vor und ]iegen innerha lb des t r ansp l an t i e r t en Gewebes oder d ieh t a m Mesodermrand . I n dem Transp l an t a t , das nur Urgesehlechtszel len enth~l t , l iegen sie abgekapse l t in e inem Nes t zu e twa 20 St i ick (Abb. 14).
40 R. LE~MANN:
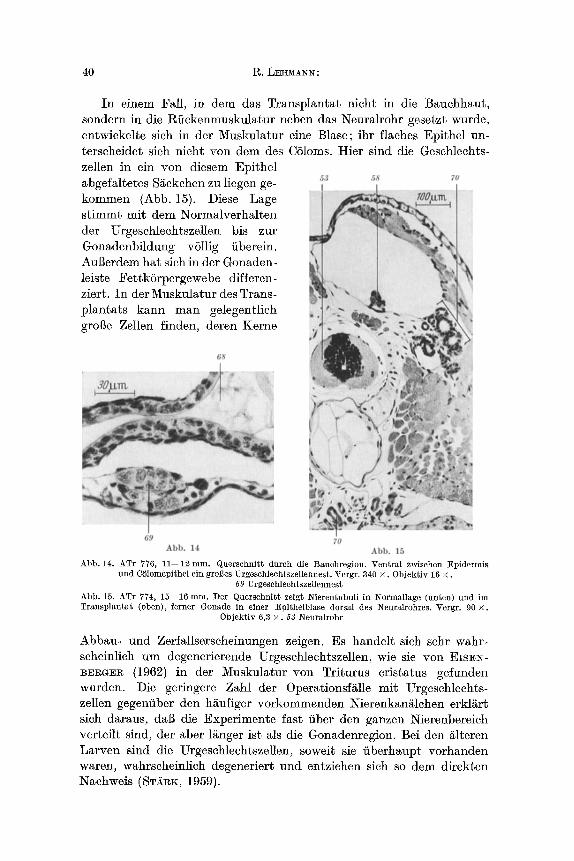
In einem Fall, in dem das Transplantat nicht in die Bauchhaut, sondern in die Rfiekenmuskulatur neben das Neuralrohr gesetzt wurde, entwicke]te sieh in der Muskulatur eine Blase; ihr flaehes Epithel un- terseheidet sich nicht yon dem des CSloms. Hier sind die Geschlechts- zellen in ein yon diesem Epithel abgefaltetes S~ckchen zu liegen ge- kommen (Abb. 15). Diese Lage s t immt mit dem Normalverhalten der Urgeschleehtszellen his zur Gonadenbildung vSllig fiberein. Augerdem hat sich in der Gonaden- leiste FettkSrpergewebe differen- ziert. In der Muskulatur des Trans- plantats kann man gelegentlieh groBe Zellen finden, deren Kerne
Abb. 14. ATr 776, 11--12 mm. Querschnitt durch die Bauchregion. Ventral z~schell Epidermis und CSlomepithel ein groi]es Urgeschlechtszellennest. Vergr. 340 • 0bjekt iv 16 x .
69 Urgeschlechtszellennest
Abb. 15. ATr 774, 15--16 ram. Der Querschllitt zeigt Nierentubuli in Normallage (unten) and im Transplantat (oben), ferner Gollade in einer Epithelblase dorsal des Neuralrohres. Vergr. 90 •
0bjekt iv 6,3 • 53 Neuralrohr
Abbau- und Zerfallserseheinungen zeigen. Es handelt sich sehr wahr- scheinlich uln degenerierende Urgeschlechtszellen, wie sie von EISE~- BEI~G]~I~ (1962) in der Muskulatur yon Triturus cristatus gefunden wurden. Die geringere Zahl der Operationsfs mit Urgeschlechts- zellen gegenfiber den hs vorkommenden Nierenkan~lchen erkli~rt sieh daraus, daft die Experimente fast fiber den ganzen Nierenbereich verteilt sind, der aber ls ist als die Gonadenregion. Bei den ~lteren Larven sind die Urgesehlechtszellen, soweit sie fiberhaupt vorhanden waren, wahrscheinlich degeneriert und entziehen sich so dem direkten Naehweis (STX~K, 1959).

Entwieklung des 0pisthonephros bei Triturus alpestris hl
3. Ge/ii[3e und Blur
Bei besonders intensiver Entwieklung, schon makroskopisch sieht- bar, erseheinen im Stadium zwisehen 7--10 m m K6rperlgnge ira Trans- plantar grol~e rote Blasen. I m Schnittbild erweisen sie sieh als kleine Epithelsgckehen, die prall mit roten BlutkSrperchen gefiillt sind. Die starke Vermehrung der Blutzellen wird dutch Mitosen angezeigt.
Die Verbindung des Transplantatgewebes mit dem Kreislaufsystem des Wirtes kann fiber eine Kapillare oder auch ein grSgeres Gefgg zu- stande kommen. Es kann z.B. fiber eine Gewebebrficke, die einventrales Mesenterium vortguscht, vom Transplantat zum Darm eine Verbindung mit einem Blutgef~l~ des Darmes hergestellt werden. Die Beziehung zum Kreislaufsystem des Wh'tes kann in zeitlieh versehiedenen Phasen stattfinden und ist z.B. ffir die Differenzierung tier Glomeruli yon Be- deutung. Warm und in welehem Ma6e mit den Blutgefgl~en des Wirtes eine Verbindung eintritt, ist sicher yon der rgumliehen Beziehung zwi- schen Spender-Wirtsgewebe sowie yon der F~thigkeit abhgngig, Gefg6e in das Transplantat hinein- oder aus ihm heraussprossen zu lassen. J, ltere Transplantate sind mehr oder weniger stark yon Kapillaren durehsetzt. Welehe Anteile vom Wirt oder vom Spender sind, kann nieht mehr entsehieden werden.
st. Nierenanlagen
a) Nierenkani~lchen
Bei den Versuchen entwickelten sich in der fiberwiegenden Mehrzahl der Fglle (Tabelle 3) Kanglchenanlagen versehiedenen Differenzierungs- grades. Bei ffinf Tieren wurde die Entwieklung vor der siehtbaren Dif- ferenzierung von Nierenzellen unterbrochen (Tabelle 3). Von allen ande- ren zeigt nur eine Larve keine eindeutig erkennbaren Nierenbildnngen.
Die Larven, in denen sich das Transplantat differenziert hat, fixierte ich so, dab nach den gewonnenen Kenntnissen fiber die Normalentwicklung der Niere eine Entwicklungsreihe yon der Nephroblastblase bis zum ~usdifferenzierten Kan~lchen zu erwarten war.
Die Ubersichtsaufnahme (Abb. 16) zeigt eine Nephroblastblase in der Bauehmuskulatur. Auch die anderen Transplantate liegen his anf wenige Ausnahmen in diesem Bereich, tells etwas ngher zur t Iaut , tells welter median. Ein Aussehnitt aus Abb. 16 ]&6t die genaue Lage der Iqephroblastblase erkennen (Abb. 17). Sie ist durch ein loekeres Binde- gewebe v o n d e r Bauehmuskulatur getrennt und ragt, yore CSlomepithel /iberzogen, in das CSlom hinein. Sie gleicht mit ihren ]&nglichovalen Kernen den IkTephroblastblasen in der l~ormallage (Abb. 2). Die Wei- terentwieklung fiihrt zu einem Kanglchen, das in seinen Teflstfieken zun~ehst noch einheitlich strukturiert ist. In der Folge entstehen eine Vielzahl von Tubu]usschleifen, die dieht beieinander liegen (Abb. 18)
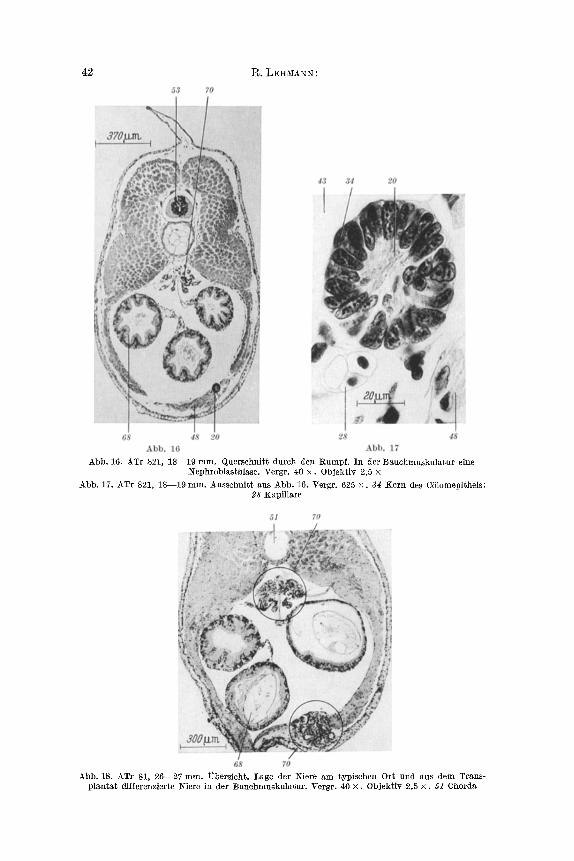
42 R . L E n ~ A ~ :
Abb. 16. ATr 821, 18--19 ram, Querschnitt durch den Rumpf. In der Bauchmuskula~ur eine Nephroblastblase. Vergr. 40 • Objektiv 2,5 •
Abb. 17. ATr 821, 18--19 ram. Ausschnitt aus Abb. 16. Vergr. 625 • 3g Kern des CSlomepi~hels; 28 Kapillare
Abb. 18. ATr 81, 26--27 ram. ~bersicht. Lage der ~iere am typischen Oft und aus dem Trans- plantar differenzierte :Niere in der Bauchmuskulatur. Vergr. 40 • Objektiv 2,5 • 51 Chorda
Entwicklung des Opisthonephros bei Triturus ~lpestris 43
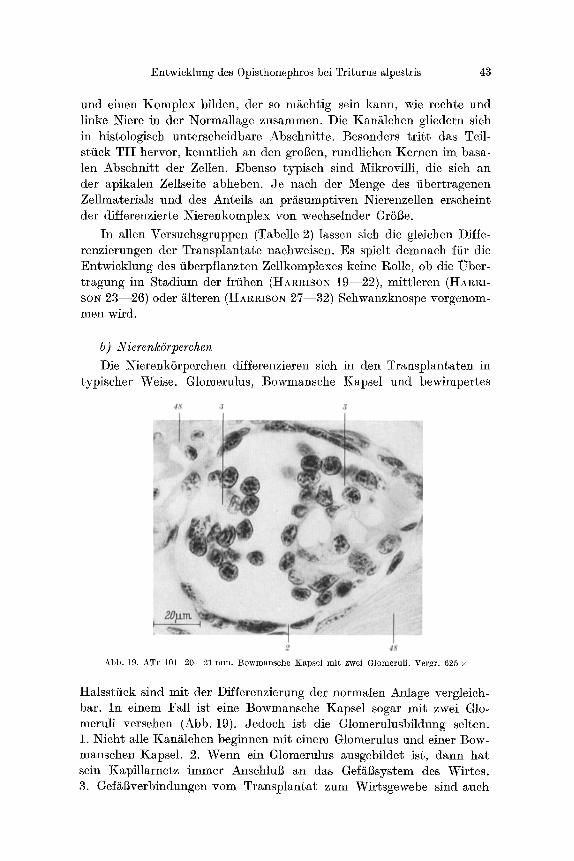
und einen Komplex bilden, der so m/~chtig sein kann, wie rechte und linke Niere in dcr Normallage zusammen. Die Kan/~lchen gliedern sich in histologisch unterscheidbare Abschnitte. Besonders t r i t t das Teil- stfick TI I hervor, kenntlich an den grol3en, rundlichen Kernen im basa- len Abschnitt der Zellen. Ebenso typisch sind Mikrovflli, die sich an der apikalen Zellseite abheben. Je nach der Menge des fibertragenen Zellraaterials und des Anteils an pr/~sumptiven ~ierenzellen erscheint der differenzierte Nierenkomplex yon wechselnder Gr613e.
In allen Versuchsgruppen (Tabelle 2) lassen sich die gleichen DJffe- renzierungen der Transplantate nachweisen. Es spielt demnach ffir die Entwicklung des fiberpflanzten Zellkomplexes keine l~olle, ob die IJber- tragung im Stadium der frfihen (HARRISO~ 19--22), mittleren (HAlc~I- SO~ 23--26) oder is (HAn~ISO~ 27--32) Schwanzknospe vorgenom- men wird.
b) Nieren/cdrperchen Die Nierenk6rperchen differenzieren sich in den Transplantaten in
typischer Weise. Glomerulus, Bowmansche Kapsel und bewimpertes
Abb. 19. ATr 101 20--21 ram. Bowmansche Kapsel mi t zwei Glomeruli. Vergr. 625 •
Halsstfick sind mit der Differenzierung der normalen Anlage vergleich- bar. In einem Fall ist eine Bowmansche Kapsel sogar mit zwei Glo- meruli versehen (Abb. 19). Jedoch ist die Glomerulnsbildung selten. 1. Nicht alle Kan~tlchen beginnen mit einem Glomerulus und einer Bow- mansehen Kapsel. 2. ~u ein Glomerulus ausgebildet ist, dann hat sein Kapfllarnetz immer Anschlul3 an das Gef~Bsystem des Wirtes. 3. Gef~l~verbindungen yore Transplantat zum Wirtsgewebe sind auch

44 t~. L ~ A ~ :
vorhanden, wenn kein Glomerulus ausgebildet ist. Diese Befunde m6ehte ich wie folgt interpretieren: Zur Ausbildung eines Glomerulus ist es notwendig, dab Kapillaren, die mit dem Gef/~Bsystem des Wirtes in Verbindung stehen, in der N/~he der Glomerulusanlage vorhanden sind. Das Zusammentreffen yon Kapillare und Glomerulusanlage muB reeht- zeitig stattfinden. Nur wghrend einer kurzen Entwieklungsphase kann der proximale Absehnitt der Kanglchenanlage mit einer Glomerulns- bildung reagieren. Der Normalentwieklung folgend, ist diese Phase sehr wahrseheinlich auf die auswachsende Nephroblastblase besehr/~nkt. Fin- det keine zeitgereehte Verbindung mit einer Kapillare start, dann ent- steht kein Glomerulus oder in besonderen Fi~llen eine atypisehe Anlage.
Die geringe Anzahl der gebildeten Glomeruli 1/~l~t sieh mit dem zah- lenm/~Bigen und zeitliehen Auftreten yon Kapillaren im Operationsbe- reich begrfinden. Die Kapillaren, die das Transplantatgewebe durchzie- hen, sind nieht yon Anfang an vorhanden, denn der K6rperbereieh, der das Spendermaterial aufnimmt, ist normalerweise nur sehwaeh mit Ge- fagen versorgt. Erst im Laufe der nachfolgenden Entwieklung wird das Transplantatgewebe s~grker kapillarisiert. Aus der Normalentwiek]ung ist aber bekannt, dab sehr frfihzeitig eine Kapillare in die Glomerulus- anlage einsprogt (Abb. 3). So hat die auswaehsende Nephroblastblase in der ortsfremden Region nur eine geringe Chance, mit einer K~pillare in Berfihrung zu kommen. Nimmt die Zahl der Kapillaren zu, ist die Dffferenzierung der Kanglehenanlage so weir vorangesehritten, dag es dem Epithel offenbar nicht mehr m6glieh ist, auf einen Gefiigkontakt mit einer Glomerulusbfldung zu antworten.
c) Nephrostom In keinem der Transplantate konnte ein Nephrostom beobachtet
werden. Auch dann nieht, wenn sich ein Nephron mit typischen Ab- schnitten sowie die Enmfindung in den mittransplantierten prim~ren I-Iarnleiter und das Nierenk6rperchen entwickelt haben. Nur sehr selten ist eine Nephrostomanlage zu sehen; ihr Epithel tritgt zwar Cilien, aber eine Kommunikation mit der Leibesh6hle, selbst wenn das C61omepi- thel unmittelbar anschlieBt, ist nicht ausgebildet.
d) Vergleich der Nierenkandlchen Ein Vergleich der Nierenkan~lchen, die sich im Transplantat dif-
ferenziert haben, mit denen, die sieh in der Normallage entwickelt h~ben, ergibt ~uf den ersten Blick eine erstaunliche ~bereinstimmung (Abb. 20a--b). Beide Nierenkani~lchen beginnen mit einem Nieren- kSrperchen, an das sich das bewimperte I-Ialsstfick und der Teflab- schnitt TI anschlieBen. Darauf folgt das dicke Tubnlusstfick TII, das allmghlich dfinner wird und in TI I I fibergeht. Nach einem sehr engen

Entwicklung des Opisthonephros bei Triturus alpestris 45
mit Cilien versehenen Kan//lchenabsehnitt miindet das Nephron fiber das Teilstfick TV bei dem normal entwickelten Kan~lehen in den pri- ms Harnleiter (Abb. 20a). Der im Transplantat gebildete Tubulus ist kfirzer und endet blind (Abb. 20b). Aueh ein Nephrostom ist nieht vorhanden. Soweit sieh abet das Kan~lchen (Abb. 20b) entwiekelt hat, ist es in morphologisch folgeriehtige Abschnitte gegliedert.
Zu welchem Ergebnis ffihrt nun der histologisehe Vergleich der Kan/ilchenabschnitte ? Das Epithel der Bowmanschen Kapsel und die
] / z
pr/mo?~er~l ~ r
II \ Mg'ndung
Harnle//e/" I I b 7~J;/stubk
Ne~hros/om', "/IV ~ N / ~ , , " e n -
-117 '-:j ] / I \Ha/~sCdc/~
I /-~r I
I ender r
Abb. 20a- -b . Yergleich eines Opisthonephroskan~lchens (a) mit einem Transplantatkan~lchen (b), Vergleichbare Kaniilchenabschnittr eingerahmt, mit Cilien versehene gestrichelt
Ausdifferenzierung des Glomerulus stimmen sehr gut mit der Normal- entwieklung fiberein. In gfinstigen Fgllen sind aueh die bewimperten Absehnitte TI und TIV ausgebildet. Die Zellen des dicken Tubulus- absehnittes TI I tragen zwar eharakteristische Merkmale (basal liegende Kerne, Mikrovilli), aber ein Vergleieh mit einem normal differenzierten Opisthonephroskan~ichen ergibt auffgllige Untersehiede. Zellgr6Se und I-I6he der Mikrovilli werden nieht erreieht ; die Kerne sind unregelm/~Big in ihrer Ausbildung und haben teihveise nierenf6rmige Gestalt ange- nommen. Aueh die Differenzierung des distalen Kan~lehenteils in einen weitlumigen und einen engen Absehnitt konnte nieht naehgewiesen wet- den. Zusammenfassend 1/~l]t sieh sagen, dal3 sieh die Kanglehen im Transplantat in Absehnitte gliedern, die histologiseh unt, erseheidbar sind. Obwohl die Epithelien typisehe Merkmale tragen, besteht ein entseheidender Untersehied zum normMen Kanglehenepithel, indem keine Funktionstypen ausgebildet werden. Besonders trifft das ffir die Absehnitte TII, T I I I und TV (mit ,,Streifenepithel") zu, die der Sekretion bzw. Resorption dienen und in besonderem MaBe mig ihrer

46 ~.L~HMA~:
Umgebung in Beziehung stehen. In der Normallage werden die Tubulus- sehleifen yon einem starken Blutstrom in lakunenartigen Kapillaren umspiilt, der yon der V. portae renis zur V. cava post. flie[R. Im Trans- plantar verlaufen nut wenige Kapillaren, die entsprechend ihrer gerin- gen Gr613e und peripheren Lage nur eine kleine Blutmenge an die Kanglehensehlingen heranffihren. Das Fehlen des ven6sen Blutstromes hat mutmaBlieh Einflul3 auf die Differenzierung des ~XTierenepithels.
In einigen Fgllen entwiekeln sich Nierenzellen, die sich in einem Zu- stand befinden, der normalerweise nie auftritt. Das sehr lieht vorh~ndene Zytoplasma lg~t die Kerne in einem breiten, hellen Hof erscheinen. Die Zellgrenzen, die zwischen zwei benaehbarten Nierenzellen eines normal entwickelten Kanglchens nut sehr schwer zu erkennen sind, treten hier ~Is dicke Wgnde hervor. Die fibrige Zelloberflgehe fgrbt sich ebenfalls krgRig an und ruft den Eindruek einer verstgrkten Grenzflgche hervor. Aueh ffir den mittransplantierten primgren Harnleiter treffen diese Merk- male zu. Mit dieser besonderen Ausbildung bekommt das abgewandeRe Nierenepithel Ahnlichkeit mit einem anderen Zellverband, ngmlich der Epidermis. Ihre Zellen sind gleiehermal3en dutch ein sehr liehtes Zyto- plasma und eine verstgrkte Zellmembran ausgezeichnet.
Ein Nephron im Transplantat erreieht nicht die gleiehe Differenzie- rungsh6he wie in der Opisthonephrosregion. Man kann sieh ffir das Differenzierungsgeschehen im Transplantat vorstellen, dal3 an irgend- einer Stelle im zeitliehen Ablauf eine St6rung auftritt. Es k6nnen Ein- fliisse aus den benaehbarten Geweben sein, die hemmend auf die I)iffe- renzierung der ortsfremden Nierenkanglehen einwirken, oder es kann ein Faktor, der fiir die Weiterentwicklung unbedingt notwendig ist, fehlen, z.B. ausreiehende Blutversorgung. Der letzten Annahme ist der Vorzug zu geben, da der Blutstrom in direkter funktioneller Beziehung zum Nierenepithel steht.
Die Entwieklung einer Kanglchenanlage im Transplantat kann so eharakterisiert werden, dal3 sich Nephroblastblase und junges Kangl- then zungchst herkunftsgemgl3 entwickeln. Die Weiterentwicklung zu einem yell ausgebildeten Nephron geht aber fiber einen kritisehen Be- reich nicht hinaus, da ein normalerweise vorhandener Einflul3 aus der Umgebung, der jetzt eingreifen mfilRe, unterbleibt.
F. Diskussion der Ergebnisse iiber:
1. Herl~un/t der NierenbildungszeUen ~ber die Herkunft des Nierenblastems besteht bei allen Autoren
eine iibereinstimmende Auffassung. Abgesehen yon Ft)RBRI~G~R (1877, dort weitere Literaturangaben), der die Niere yon Wucherungen des ,,parietalen Peritoneums" (S. 54) ableitet, geben alle anderen (I-IALL, 1904; FELIX, 1906; GaAY, 1932; NELSOn, 1953; NOBL~, 1954) das

Entwicklung des Opisthonephros bei Triturus alpestris 47
intermedi~tre Mesoderm (Ursegmentstiel, Nephrotom, Mesomer) als Ur- sprungsort an, der durch Experimente yon MACI~MER (1929), HuM- PUu~Y (1928) und eigene eindeutig best~tigt ist. Eine unterschiedliehe Meinung herrscht aber fiber die spezielle Ausbfldung des Nierenblastems. Ft~RsuIsG~u (1877) spricht yon ,,Urnierenstri~ngen" (S. 54) und nach GRAY (1932), dessen Angaben sieh auf HALL (1904) sttitzen, gliedert sieh vom intermedi/tren Mesoderm ein solider Blastemstrang ab, der dann in eine Anzahl runder oder ovaler Blasen zerfiillt (S. 431). FsLIX (1906, mit ausfiihrlichen Literaturangaben), der auch eine Abstammung der ,,Urnieren"-Anlagen yore C61omepithel nicht ganz aussehlieBt (S. 256), vertr i t t die Ansieht, dab sich bei dan ,,urodelen Batrachiern" die Ursegmentstiele im Mesenchymgewebe aufl6sen. Spgter kommt es dutch Konzentrierung zu einer Neubildung des nephrogenen Gewebes, und dann warden ,,in ihnen Zellkugeln und Zellblgsehen hergestellt" (S. 205, 207, 255). NOBL~ (1954) ffihrt in diesem Zusammenhang eine Abbildung nach FELIX (1906) an, in der die Entstehung des Vornieren- kan~lehens als Abfaltung vom Ursegmentstiel erl/~utert wird. Weiter heilR as, dab die Opisthonephroskanglehen in derselben Weise ent- stehen wie die Tubuli des Pronephros (S. 268).
Naeh meinen Untersuehungen leiten sieh die Kanglehenbildungs- zellen als Einzelzellen aus dem intermedi~ren Mesoderm ab. Sie liegen anfangs weir lateral am ventralen Myotomrand und gelangen zusam- man mit dam ebenfalls noah lateral liegenden prim~ren Harnleiter als Folge yon Gestaltungsbewegungen nach medial. Dureh Verbraueh des entodermalen Dotters wird es m6glieh, dab der primate Harnleiter und die Nierenbildungszellen der lateral waehsenden Muskulatur naeh me- dial ausweiehen k6nnen. Dag meehanisehe Kr/tfte ffir die Verlagerung verantwortlieh sind, zeigen Defekttiere, die viel Myotommaterial ver- loren haben. Bei diesen Larven hat der prim/~re Itarnleiter seine ]aterale Lage im Operationsbereieh beibehalten, w/~hrend er in dan Regionen davor und gegebenenfalls dahinter die mediale Normallage erreieht hat. Aueh die Nierenbildungszellen, sofern sic nicht im entnommenen Trans- plantatmaterial enthalten waren, sind dann nicht naeh medial gelangt.
Die Gesamtheit der Nierenbildungszellen kann man nieht als einen soliden Strang betraehten. Es handelt sich immer um eine unregel- m~gige Folge yon Einzelzellen, die dorsal des prim~ren Itarnleiters liegen. Spgter entsteht yon cranial naeh caudal fortsehreitend eine Folge yon Zellgruppen und daraus eine Folge yon Nephroblastblasen.
2. Entstehung dorsaler Kaniilchen
Von GRAY (1932) werden keine dorsalen oder ihnen entsprechende Kan/~lehen beschrieben. Auch seinen Abbildungen sind keine Hinweise fiir diese Anlagen zu entnehmen. Den Rekonstruktionen, die er yon

den Nierenkan~lehen yon Tr. vulgaris angefertigt hat, sowie aus den dazugehSrigen Erl/~uterungen ist zu entnehmen, dab yon den zuerst angelegten Nierenkan/tlchen dureh Sprossung weitere Kan~lehen ent- stehen. Bei Tr. alpestris entwiekeln sieh dagegen dorsale Kan/~lehen als selbstgndige Anlagen, die in bereits ausgebildete KanNehen einmilnden. Da ffir S. salamandra (FOxBRI~GE~, 1877) eine/~hnliche Entwieklung der Nierenkan/~lehen besehrieben ist, wie f/it Tr. alpestris gefunden, kSnnte man daraus ableiten, dag sieh in bezug auf die Nierenentwicklung Alpenmoleh und Feuersalamander n~her stehen als A1pen- und Teich- molch. Eine wei~ere Tatsaehe 1/~13t die Befunde yon G~Au sehr unwahr~ scheinlieh erseheinen. Die sekund/~ren und tertiN'en Kan~lchen yon Tr. vulgaris sprossen an einem ganz anderen Tubulusabsehnitt aus, als die dorsalen in der Alpestrisniere einwaehsen. Aber aueh in der Niere yon Neeturus maeulosus (CHASE, 1923) vereinigen sieh die Nierenkan~L ehen im gleiehen Teilsttick (dorsolateraler Bogen) wie bei Tr. alpestris. Wenn der yon GRAY besehriebene Entwieklungsgang s t immt - - es ver- einigen sieh immer drei Kanglehen --~ mfil~ten z.B. auf 13 Mfindungen in den prim/iren Harnleiter 3 • 13-~ 39 Nierenk6rperehen kommen. Augerdem diirfte kein Kan~lchen in den dorsolateralen Bogen miinden. Eine Sehnittserie dutch ein 27--28 mm langes Tier (Tr. vulgaris) nach der Metamorphose ergab folgende Zahlen und Zusammenh/mge (Tabelle 4):
T~belle 4. Anzahl tier Nephrcme der linken Niere bei Tr. vulgaris (27--28 ram)
Nierenk6rper- Miindungen in den Anzahl der Kan~lchen chert
primgren dorsolateralen ventrMe dorsMe Harnleiter Bogen
Gesamte Niere 45 33 12 33 12 Cranialer Teil 20 20 - - 20 - - Caudaler Tell 25 13 12 13 12
Die linke Niere enth/~lt 45 Nierenk6rperehen, yon denen 20 auf die cranialen Anlagen entfallen und ebenso 20 Mtindungen in den prim//ren Harnleiter. Naeh diesem Abzug verbleiben ffir den eaudalen Nierem bereieh 25 NierenkSrperehen, 13 Miindungen in den prim/iren }[arnleiter und 12 Mtindungen in den dorsolateralen Bogen. Anders ausgedr/iekt verteilen sieh die 25 Nierenk6rperchen auI 13 ventrale und 12 dorsMe Kan/ilehen. Drei Nephroblastblasen, die auf dem gleichen Niveau liegen wie die dorsalen Nephrone, weisen auf die gMehe selbstgndige Ent- stehnngsweise der dorsalen Kan~lehen bin wie bei Tr. Mpestris, zumal innerhalb der dorsalen Anlagenfolge aaeh ein eranio-eaudales Differen- zierungsgef/ille vorliegt. Die yon GRAY (1932) besehriebene Kan/~lehen- entwicklung dutch Sprossung kann in keiner Weise best/~tigt werden.

Entwicklung des Opisthonephros bei Triturus alpestris 49
Zusammenfassung 1. Der Opisthonephros wh'd in mehreren fibereinanderliegenden An-
lagenfolgen ausgebfldet, die sich nacheinander in einem cranio-caudalen Differenzierungsgefi~lle entwiekeln, Abweiehungen treten in der cau- dalen H/ilfte und bei den dorsalen Anlagenfolgen auf.
2. Das I~Tierenblastem, das bei Triturus aus einer Folge yon Einzel- zellen besteht, die in unregelm/il~igen Abst/inden fiber dem prim/iren tIarnleiter liegen, kommt aus dem Zellkomplex des intermedi/iren Meso- derms.
3. Die Entwicklung eines Opisthonephroskan/ilehens geht yon Ein- zelzellen fiber Zellgruppen und -blasen aus und nimmt nicht ihren Anfang durch Faltung aus dem Nephrotom. Die Differenzierung inner- halb eines Kan/ilchens sehreitet yon proximal nach distal fort.
4. Die Nephrostomanlage tritt nach Differenzierungsbeginn der Bow- mansehen Kapsel auf. Bevor die Verbindung mit dem CSlom eintritt, sind Cilien ausgebildet; auch in den Nephrostomanlagen dorsaler Kan/ilchen, die z.T. niemals mit dem CSlom kommunizieren.
5. Neue Kan/ilehen entstehen nieht dureh Sprossung aus bereits vorhandenen Tubuli, sondern aus selbst/indigen Anlagen, die fiber ihre dorsolateralen Bogen miteinander in Verbindung treten.
6. Im Operationsbereich fehlen h/iufig Kan/ilchenanlagen; liegt er in der pr/isumptiven Gonadenregion, fehlen aueh Urgesehlechtszellen. l~egenerationscrscheinungen laufen im Defektbereich in folgender zeit- lieher Reihenfolge ab: Ektoderm, Muskulatur, Nierengewebe. Die durch die Operation gestSrte Symmetrie des Embryos wird in kurzer Zeit wieder hergestellt.
7. Als Operationsfolge ist der prim/ire Harnleiter ganz, teilweise oder nicht vorhanden. Er kann mit dem Ektoderm verkleben und eine neue Ausmfindung erhalten. Die V. portae renis und das Seitenlinienorgan sind atypisch entwiekelt. Die Muskulatur weicht nur geringfiigig yon der Normalentwieklung ab.
8. Durch den Defekt ist die zeitgereehte Differenzierung der Kan/il- ehenanlagen, die hinter der Defektstelle liegen, verzSgert. Besonders, wenn der prim/ire Harnleiter unvollst/indig oder gar nieht ausgebildet ist.
9. Die Transplantate enthalten herkunftsgem/iBe Differenzierungen: Muskulatur, Keimzellen, Gef/ige, Blur und Kan/ilehenanlagen verschie- denen Differenzierungsgrades.
10. Das Transplantatkanglchen zeigt eine Gliederung in morpho- logiseh folgerichtige Absehnitte, aber es werden keine Funktionstypen ausgebildet. Das Fehlen des ven6sen Blutstromes hat mSglicherweise EinfluB auf die Differenzierung des Nierenepithels.
4a Wilhelm !~oux' Arch. Entwickl.-Mech. Org., Bd. 158

50 1~. LEm~n~:
Literatur
AMA~U]VL~, A. : Effect of exstirpation of the presumptive intermediate mesoderm upon the differentiation of the primordial germ cells. Zool. Mug. (Tokyo) 66, 310~313 (1957).
ASAY~IA, S. : The developmental potencies of the intermediate mesoderm of Tri- turus pyrrhogaster when transplanted into orthotopic or heterotopic site in the body wall of another embryo. J. Inst. Polytech., Osaka City Univ. 1, 13--32 (1950).
- - Potency of lateral plate mesoderm relating to the formation of reproductive tissue. Zool. Mag. (Dobutsugaku Zasshi) 70, 7--11 (1961).
C~sE, S.W.: The mesonephros and urogenital ducts of Necturus maculosus t~afinesque. J. Morph. 37, 457~531 (1923).
EIs~]3E~o~, W. : Die streuenden Geschlechtszel]en bei Triturus eristatus. Un- ver6ffentl. (1962).
FELIX, W., u. A. Bih~L~R: Die Entwicklung der Ham- und Geschlechtsorgane. In O. HEB~Wm, Handbuch der vergleichenden und experimente]]en Entwick- lungslehre der Wirbeltiere, Bd. 3. Jena: Gustav Fischer 1906.
FRANCIS, E. T. B. : The anatomy of the salamander. Oxford: Clarendon Press 1934.
Fff~B~I~GE~, ~ . : Zur Entwicklung der Amphibienniere. Heidelberg: J. H5rning 1877.
GRAY, P.: Development of the amphibian kidney. Part II. The development of the kidney of Triton vulgaris and a comparison of this form with I~ana tem- poraria. Quart. J. micr. Sci. 75, 4 2 5 ~ 6 5 (1932).
I-DmL, I~.W.: The development of the mesonephros and Mullerian ducts in am- phibia. Bull. Mus. compar. Zool. Harvard 44, 31--125 (1904).
tt~RTWlG, O. : Lehrbuch der Entwicklungsgeschichte des Mensehen und der Wir- beltiere, 10. Aufl. Jena: Gustav Fischer 1915.
HHv~Ptrp~EY, t~. 1~.: The developmentM potencies of the intermediate mesoderm of Amblystoma, when transplanted into ventrolateral sites in other embryos: The primordial germ cells of such grafts and their role in the development of a gonad. Anat. t~ec. 40, 67--90 (1928).
KoT~m, M.: The differentiation of Wolffian duct, mesonephros, and primordial germ cells in the newt Triturus pyrrhogaster after exstirpation of the prim- ordium of Wolffian duct. J. Biol., Osaka City Univ. 13, I00--118 (1962).
LEm~A~, F. E. : Einftihrung in die physiologische Embryologie. Basel: Birkhiiu- set 1945.
L~ENAN~r, 1~.: I-Iistologisch-cytologische Untersuchungen fiber den Ditferenzie- rungsvorgang der Urnierenkanglchen bei Triturus alpestris. Zool. Anz. 27 Suppl., 92--99, Verh. d. Dtsch. Zool. Ges. Miinchen 1963.
- - Bauprinzipien des 3/iesonephros yon Triturus Mpestris. Zool. Anz. 28 Suppl., 300--311, Verb. d. Dtsch. Zool. Ges. Kiel 1964.
M~e~NEg, H. : Bifferenzierungsfghigkeit der Urnierenanlage yon Triton alpestris. Wilhelm Roux' Arch. Entwickl.-Meeh. Org. 118, 200--251 (1929).
N ~ s ~ , O. E.: Comparative embryology of the vertebrates. Toronto: The Bla- kiston Co. Inc. 1953.
NInvw~ooe, P. D. : Some further data concerning the determination of the meso- nephros. Experientia (Basel) 4, 391--394 (1948).

Entwieklung des Opisthonephros bei Triturus ~lpestris 51
NOBL~, K. G.: The biology of the ~ml0hibia. NewYork: Dover Publ., Inc. 1954. P~Ags:s, A. C. E. : ttistochemistry theoretical and applied, 2nd ed. London: Chur-
chill, Ltd. 1960. I~o~EIs, B. : Mikroskopische Technik, 15. Aufl. M~nchen: R. Oldenbourg 1948. SPA~z~zao~', L. : Einfiihrung in die Praxis der ttistochemie. Jen~: VEB G. Fischer
1964. STXz~, O. J.: Die streuenden Geschlechtszellen yon Triturus ulpestris. Z. Zell-
forsch. 50, 693--748 (1959).
Dr. ROLF LEI]:I~J[ANN Department of Biology University of Koehester Rochester, New York 14627/USA
4'





























