Embed Size (px)
DESCRIPTION
Ingo Rechenberg. PowerPoint-Folien zur 7. Vorlesung „Evolutionsstrategie I“. Von der (1 + 1) - ES mit 1/5 - Erfolgsregel zur (1, l ) - ES mit mutativer Schrittweitenregelung. Rückblick auf die letzte Vorlesung. Algorithmus der ( 1 + 1) – ES mit 1/5 -Erfolgsregel - PowerPoint PPT Presentation
Citation preview

Ingo Rechenberg
PowerPoint-Folien zur 7. Vorlesung „Evolutionsstrategie I“
Von der (1 + 1) - ES mit 1/5 - Erfolgsregel zur
(1, ) - ES mit mutativer Schrittweitenregelung

Rückblick auf die letzte Vorlesung

ggg zxx EN
Algorithmus der (1 + 1) – ES mit 1/5 -Erfolgsregel
in der originalen Form
1Egx
)() ENN (für ggg QQ xxx sonst E
gx
1,5 für We > 1 / 5
eiltnormalvert -1,0 / )( ngz
1,5 für We < 1 / 5
Nach jeweilsn Generationen
Interpretiert als isotrope Stichprobe der Weite 1
Breite der Gaußschen Glockenkurve
Verstärkungs- oder Abschwächungsfaktor
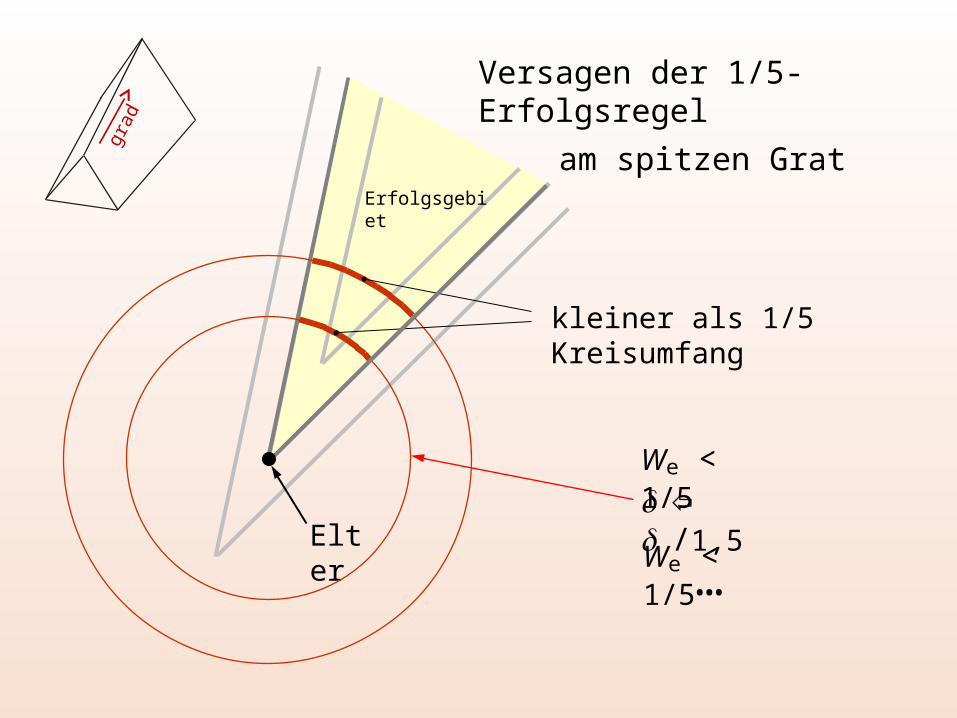
Versagen der 1/5-Erfolgsregel
am spitzen Grat
Elter
kleiner als 1/5 Kreisumfang
We < 1/5
We < 1/5
/1,5
…
Erfolgsgebiet
grad
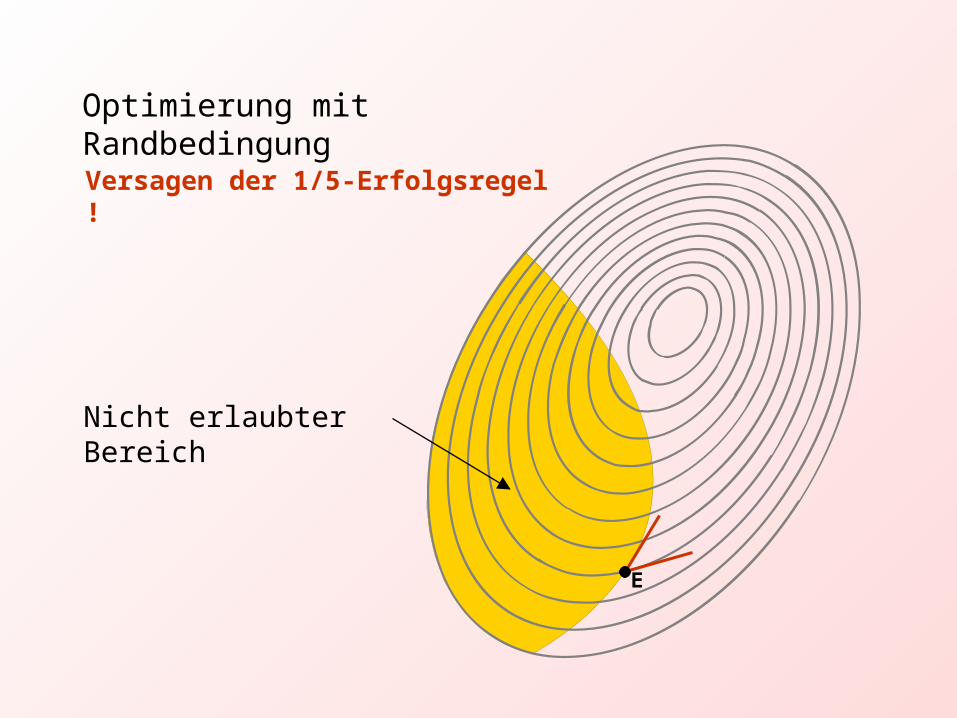
Nicht erlaubter Bereich
Optimierung mit Randbedingung
Versagen der 1/5-Erfolgsregel !
E

Ideale Funktion in der mathematischen Welt
Rauher Berg in der experimentellen Welt
Versagen der 1/5-Erfolgsregel !

2 3 4 57 8
69
Eine fehlerhafte Messung:
Q = 6 statt Q = 4
Fehlmessung bei Störungen
We < 1/ 5
/1,5
We < 1/ 5…
Erfolgsgebiet
Q =
E
~

Die (1 + 1)-ES kann an Unstetigkeiten versagen
… und sie ist unbiologisch !
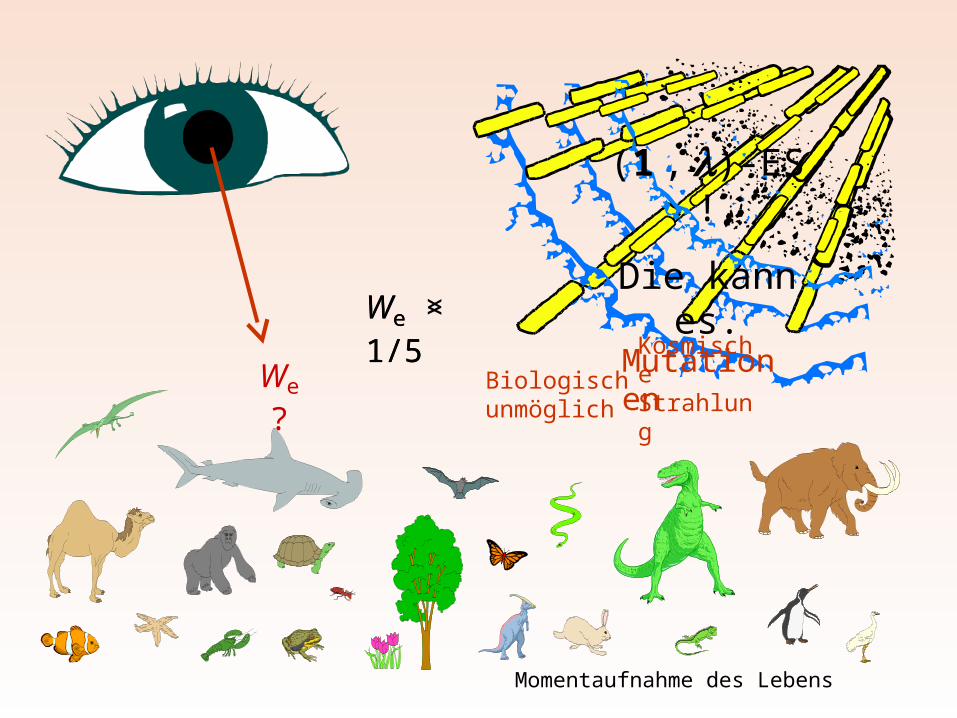
We > 1/5We < 1/5
Mutationen
Biologisch unmöglich
Kosmische StrahlungWe ?
Momentaufnahme des Lebens
(1 , )-ES !
Die kann es.
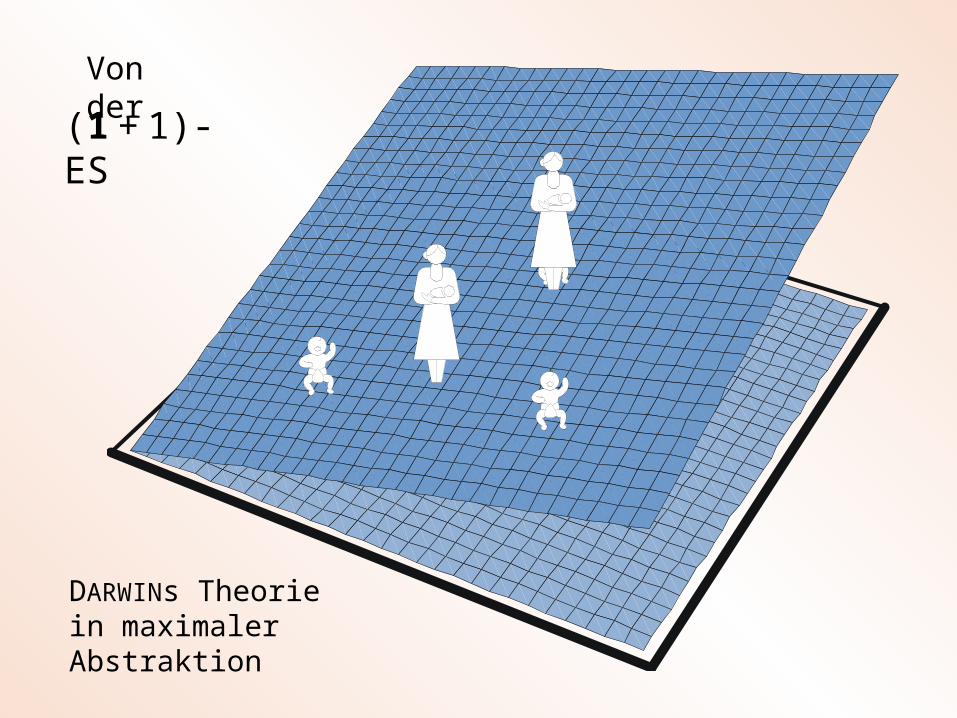
(1 + 1)-ES
DARWINs Theorie in maximaler Abstraktion
Von der
(1 , )-ES
ES mit mehr als einem Nachkommen
= 6
zur

Basis-Algorithmus der (1, ) - Evolutionsstrategie
1E1N zxx gg
2E2N zxx gg
zxx ggEN
eiltnormalvert)1,0(,, /21 nzzz n
ggNBE
1 xx )(),(),()( NNNNB 21minmax/ gggg QQQQ xxxx
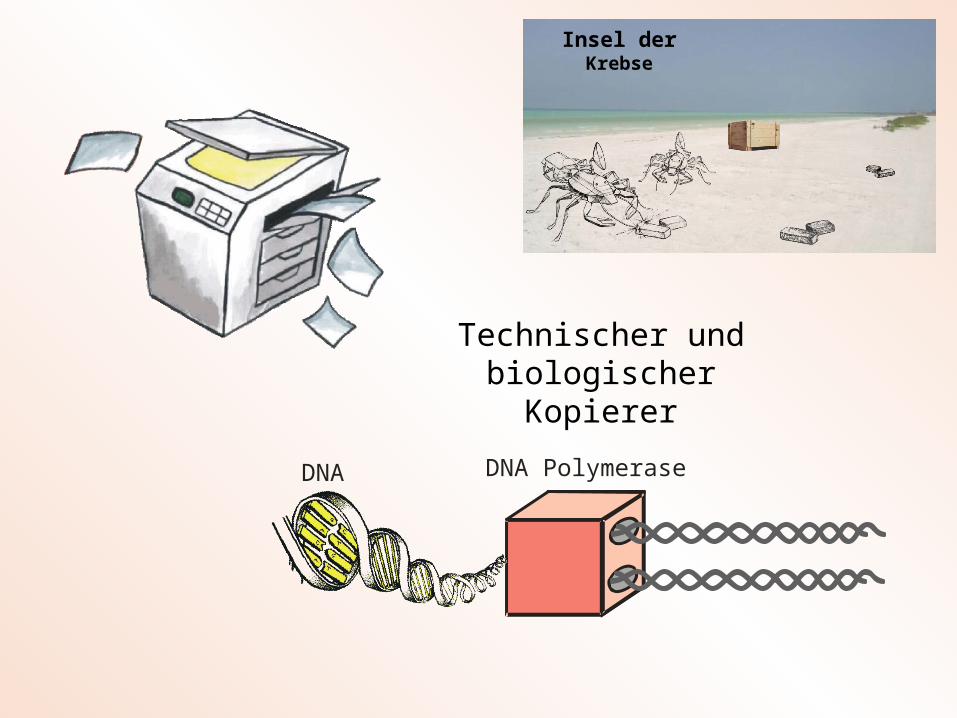
Kopiervorgang mit Fehlern
Bester
DNA PolymeraseDNA
Technischer und biologischer Kopierer
Insel der Krebse

Wenn ich mich nicht an meinem eigenen Schopfe
herausgezogen hätte
Baron von Münchhausen
Ein Lebewesen kopiert sich selbst ?

DNA-Kopierer
DNA
Der DNA-Kopierer ist zugleich ein Reparaturgerät. Falsche Buchstaben, z. B. durch kosmische Strahlung erzeugt, werden berichtigt!
Text
Eine nicht 100-prozentige Reparatur-leistung des DNA-Kopierers lässt Fehler (Mutationen in der DNA) übrig.
Verdopplung der Erbinformation durch Anlagerung komplementärer Buchstabenmoleküle (G,A,T,C) an die aufgetrennte DNA-Doppelhelix

DNA-Kopierer
DNA
Hat Kopierer hergestellt
Genetische Individualität der übrig gelassenen Mutationen
Text
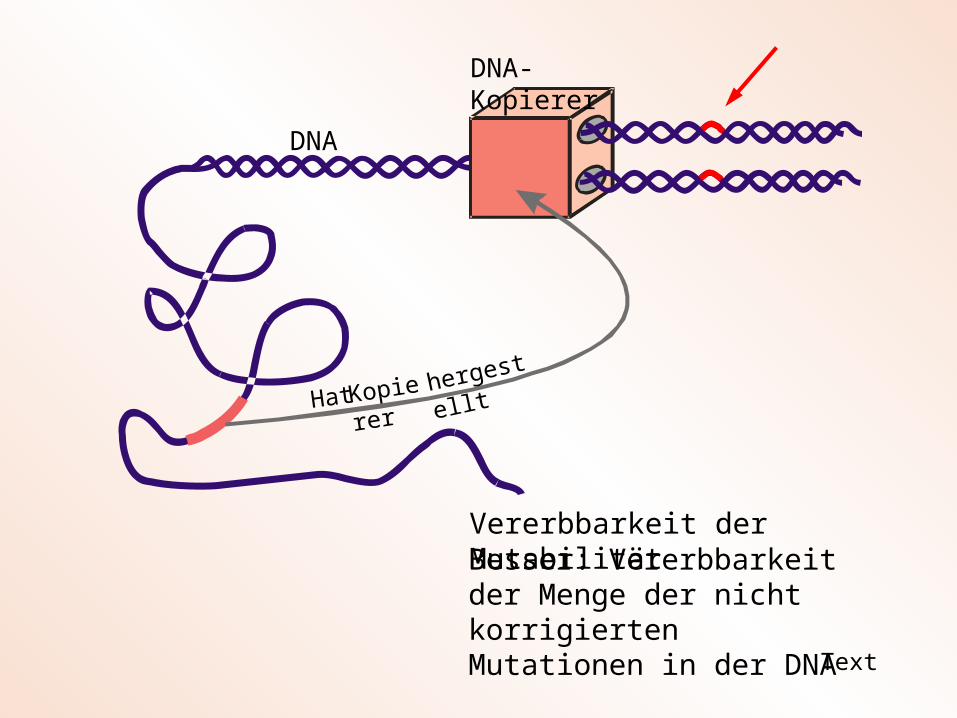
DNA-Kopierer
DNA
Hat Kopierer hergestellt
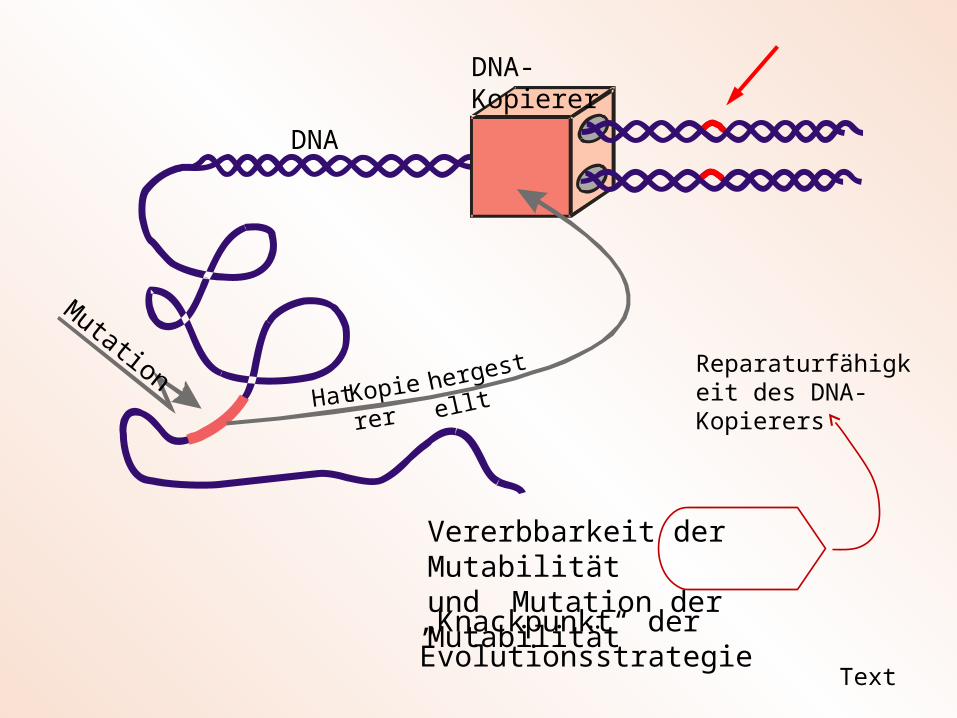
Vererbbarkeit der Mutabilität
Text
Besser: Vererbbarkeit der Menge der nicht korrigierten Mutationen in der DNA
Mutation
DNA-Kopierer
DNA
Hat Kopierer hergestellt
Vererbbarkeit der Mutabilitätund Mutation der Mutabilität
„Knackpunkt“ der Evolutionsstrategie
Text
Reparaturfähigkeit des DNA-Kopierers

Basis-Algorithmus der (1, ) - Evolutionsstrategie
1E1N zxx gg
2E2N zxx gg
zxx ggEN
eiltnormalvert)1,0(,, /21 nzzz n
ggNBE
1 xx )(),(),()( NNNNB 21minmax/ gggg QQQQ xxxx
Schrittweiten derNachkommen gleich
Aber kann variiert
und vererbt werden

Basis-Algorithmus der (1, ) - Evolutionsstrategie
11NE1N zxx ggg
22NE2N zxx ggg
zxx gggNEN
eiltnormalvert)1,0(,, /21 nzzz n
ggNBE
1 xx )(),(),()( NNNNB 21minmax/ gggg QQQQ xxxx
Schrittweiten derNachkommen
verschieden(Keimzellen eines Elters !)

Basis-Algorithmus der (1, ) - Evolutionsstrategie
1g11 NEN zxx gg
22NE2N zxx ggg
zxx gggNEN
eiltnormalvert)1,0(,, /21 nzzz n
ggNBE
1 xx )(),(),()( NNNNB 21minmax/ gggg QQQQ xxxx
ggNBE
1
Vererbung der Mutabilität

Algorithmus der (1, ) - Evolutionsstrategie mit MSR
11NE1N zxx ggg
22NE2N zxx ggg
zxx gggNEN
eiltnormalvert)1,0(,, /21 nzzz n
ggNBE
1 xx )(),(),()( NNNNB 21minmax/ gggg QQQQ xxxx
ggNBE
1
1E1N gg
2E2N gg
ggEN
eiltnormalvert schlogarithmi
Mutation der Mutabilität
Vererbung der Mutabilität
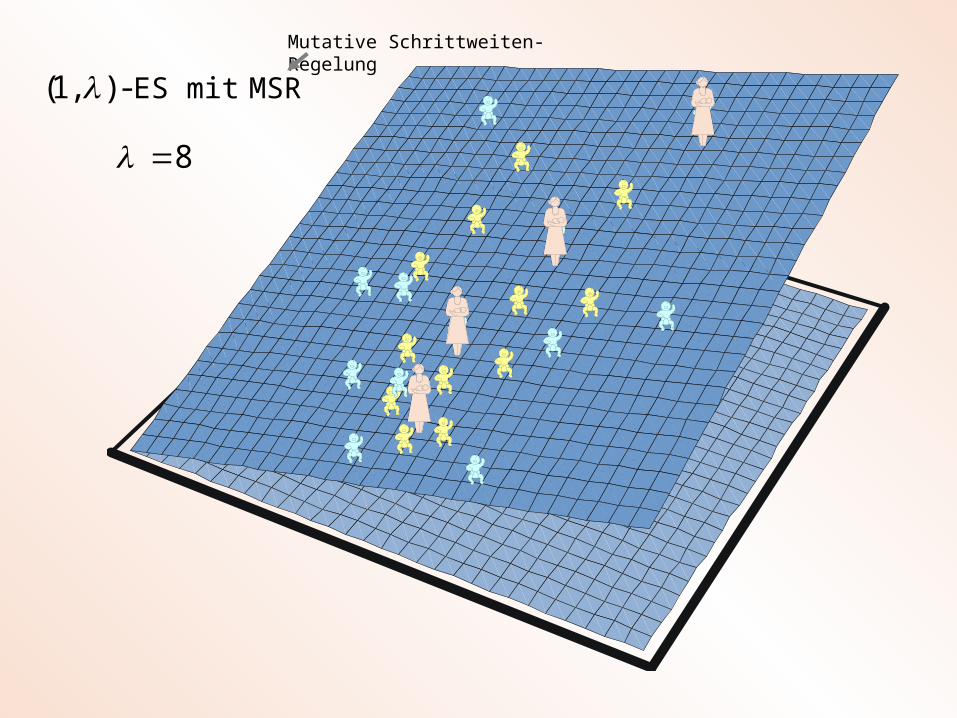
MSR mit ES-),1(
8
Mutative Schrittweiten-Regelung

? ? ?
ich bin Spitze
Einschätzung des Kletterstils
im Solo- und im Gruppenklettern
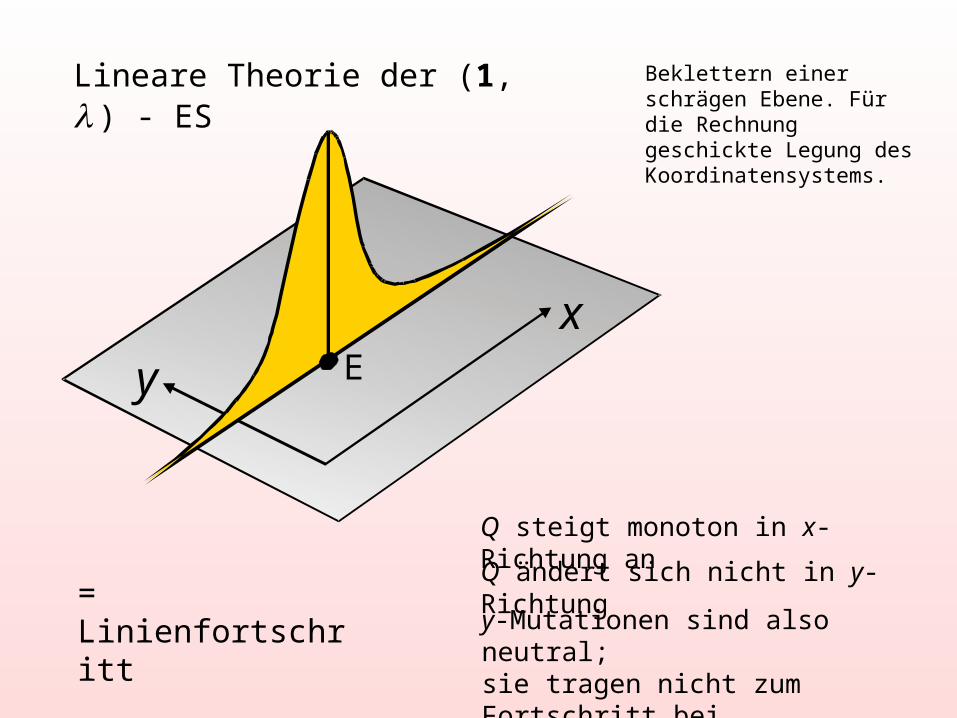
Lineare Theorie der (1, ) - ES
= Linienfortschritt
Q steigt monoton in x-Richtung an
Q ändert sich nicht in y-Richtung
y-Mutationen sind also neutral;sie tragen nicht zum Fortschritt bei
xy E
Beklettern einer schrägen Ebene. Für die Rechnung geschickte Legung des Koordinatensystems.
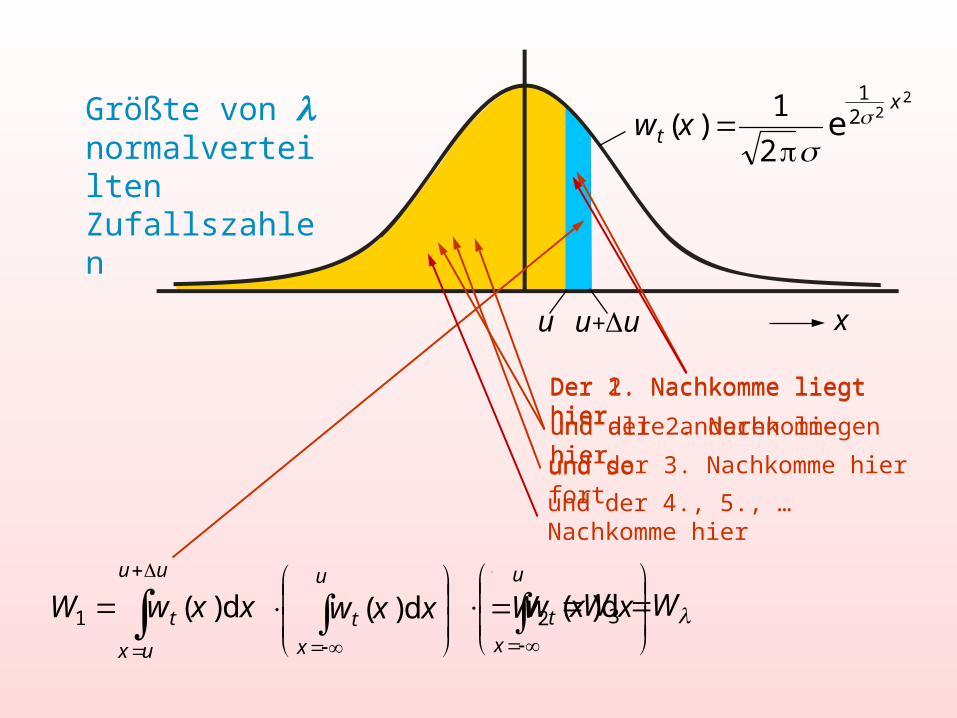
u u+u x
uu
ux
t xxwW d)(1
222
1
e2
1)(
x
t xw
2W1
d)(
u
xt xxw
Größte von normalverteilten Zufallszahlen
Der 1. Nachkomme liegt hierund der 2. Nachkomme hierund der 3. Nachkomme hier
Der 2. Nachkomme liegt hierund alle anderen liegen hierund so fort
u
xt xxw d)(
und der 4., 5., … Nachkomme hier
WW 3

u u+u x
222
1
e2
1)(
x
t xw
Größte von
normalverteilten Zufallszahlen
Die Wahrscheinlichkeit, dass der 1. oder der 2. oder der 3. oder… der . Nachkomme im blauen Bereich liegt
1
2121
u
xt
uu
uxt xxwxxwWWWW d)(d)(,,
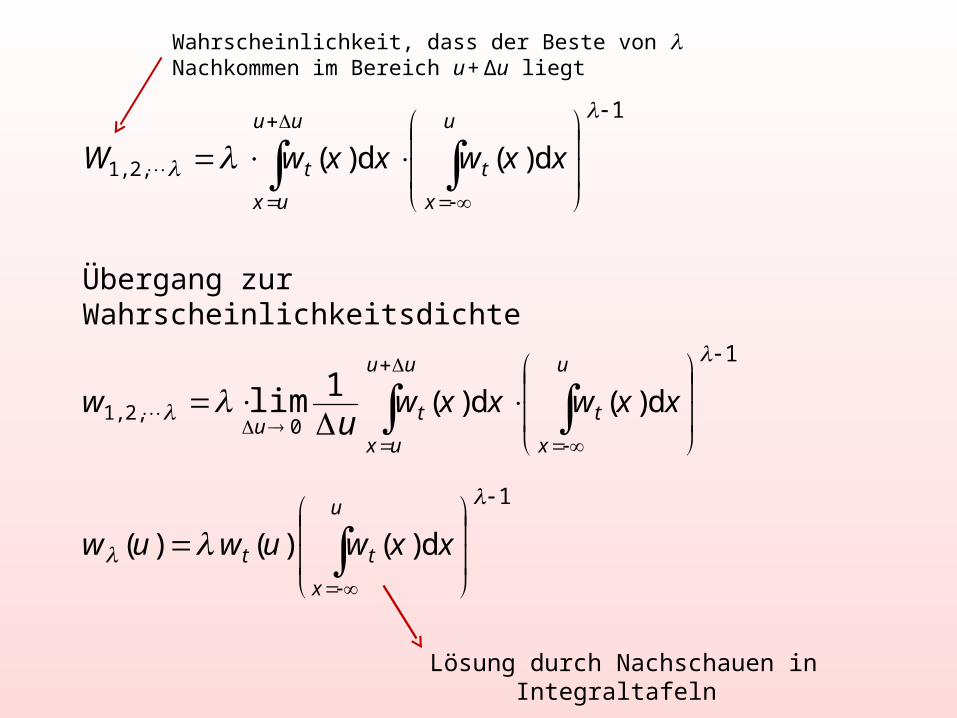
Übergang zur Wahrscheinlichkeitsdichte
1
d)(d)(,2,1
u
x
t
uu
ux
t xxwxxwW
1
d)(d)(1lim0
,2,1
u
x
t
uu
ux
tu
xxwxxwwu
1
d)()()(
u
x
tt xxwuwuw
Lösung durch Nachschauen in Integraltafeln
Wahrscheinlichkeit, dass der Beste von Nachkommen im Bereich u + Δu liegt
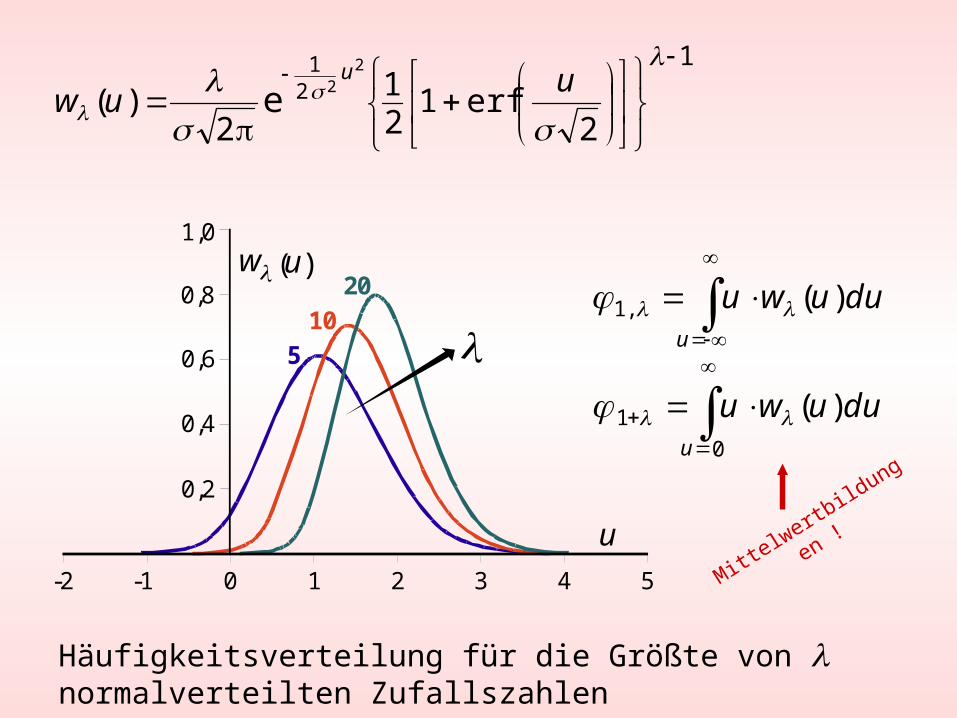
Häufigkeitsverteilung für die Größte von normalverteilten Zufallszahlen
1
2erf1
21
2)(
222
1
e
u
uwu
-1-2 0 1 2 3 4 5
u
w
0,2
0,4
0,6
1,0
0,8
5
10
20u(
)duuwu
u
)(,1
duuwuu
01 )(
Mittelw
ertbild
ungen
!

duuwuu
)(,1
0
d2
erf121
2
1222
1
,1 eu
u
uu
u
,1,1 c zzzcz
z d)erf(1e2
2 1
1,10
2
mit
duuwuu
01 )(
Lineare Fortschrittsgeschwindigkeit
Nur noch eine Funktion von

Die Fortschrittsbeiwerte der Komma-Strategie (exakte Werte)
01,1 c
5642,01
2,1
c
8463,02
33,1
c
0294,12arctan6
4,1
c
1630,112arctan62
55,1
c
,1lin,1 c
40,0lin11
21/

Die Fortschrittsbeiwerte der Plus-Strategie (exakte Werte)
3989,02
111
c
6810,02
11
2
121
c
8881,02arctan2
2
121
23
2
131
c
0458,12
321
2
141
c

Fortschrittsvergleiche der linearen Theorie
(Plus versus Komma-Strategie)
111,1 0
212,1 828,0
313,1 953,0
414,1 984,0
111141 621,22
321
4 für1,1
?

Parallele und serielle Fortschrittsgeschwindigkeit
g Weg
Nachkommenaller Zahl Weggter zurückgele
seriell
/parallelseriell
262,04046,1 /seriell 41
3989,012
1 /seriell 11
Text
gWeg
enGenerationder Zahl Weggter zurückgele
parallel

Ende
www.bionik.tu-berlin.de

Fehlerhaftigkeit des DNA-Kopieres heißt nicht, dass dieser Fehler selbst erzeugt! Kopierfehler werden ursächlich durch Umwelteinflüsse (kosmische Strahlung, thermisches Rauschen, chemische Agenzien) erzeugt. Der biologische Kopierer kann aber diese Fehler erkennen und korrigieren. Je größer die Korrigierleistung des DNA-Kopierers ist, um so kleiner ist die übrig bleibende Zahl der Mutationen (Mutationsrate). Daraus folgt: Die Mutationsfreudigkeit eines Organismus kann dieser selbst kontrollieren. Die Mutationsfreudigkeit (Mutabilität) ist eine inhärente Eigenschaft des jeweiligen Lebewesens.

Der Kopierer wird nicht von außen zu Verfügung gestellt (wir kaufen uns einen Kopierer in einem Geschäft), sondern der biologische Kopierer (die DNA-Polyme-rase) wird von dem Lebewesen selbst hergestellt. Die Bauanweisung steht irgend-wo in dem lange DNA-Erbmolekül. Daraus folgt, dass jedes Lebewesen seinen individuellen Kopierer (z. B. mit unterschiedlicher Kopiergenauigkeit) besitzt, so wie jeder Mensch sich durch seine individuelle Haarfarbe auszeichnet. Deshalb ordnen wir in dem Algorithmus der (1, )-ES jedem Nachkommen eine andere Kopierge-nauigkeit, sprich eine andere Mutationsschrittweite zu.

Aus der Tatsache, dass die Bauanleitung für den DNA-Kopierer eines Lebewesens im Erbmolekül niedergeschrieben ist, folgt die Vererbbarkeit dieses Merkmals. Die individuelle DNA-Sequenz für die DNA-Polymerase wird auf die Nachkommen über-tragen. Dies bedeutet für den Evolutionsalgorithmus, dass im abschließenden Selektionsschritt einer Generation nicht nur der Objektvariablenvektor x des besten Nachkommen, sondern auch die Mutationsschrittweite des besten Nachkommen an den Elter der neuen Generation übergeben wird.

Alles, was im DNA-Molekül niedergeschrieben ist, kann durch eine Mutation verän-dert werden. Eine Mutation des Abschnitts, der die Bauanleitung für den Kopierer enthält, ändert die Arbeitsweise des Kopierers. Der Kopierer kann genauer oder ungenauer arbeiten. Er kann Fehler mehr oder weniger korrigieren. Damit wird die Mutationsfreudigkeit (sprich Mutationsschrittweite) eines Organismus mutativ ver-ändert. Im Algorithmus der Evolutionsstrategie wird der Mutationsmechanismus ergänzt, indem nicht nur der Objektvariablenvektor x, sondern auch die Strategie-variable einer Mutation unterworfen wird. Mutationsschrittweiten müssen multipli-kativ verändert werden (eine Verdopplung der Schrittweite muss genauso häufig auftreten wie eine Halbierung. Deshalb werden die Schrittweitenmutationen mit einer logarithmischen Normalverteilung erzeugt.

Die biologische Evolution arbeitet parallel. Es bedeutet keinen zusätzlichen Auf-wand, wenn in einem gegebenen Lebensraum statt mit einem Individuum mit Tau-senden von ihnen experimentiert wird. In der Evolution zählt der Fortschritt pro Generation (parallele Fortschrittsgeschwindigkeit). Ganz anders ist es bei der An-wendung der Evolutionsstrategie auf einem gewöhnliche Computer. Hier müssen die Nachkommen einzeln nacheinander durchgerechnet werden. Für die Compu-teranwendung muss die Fortschrittsgeschwindigkeit auf die Zahl der Nachkommen bezogen werden. Diese serielle Fortschrittsgeschwindigkeit ergibt sich, indem die parallele Fortschrittsgeschwindigkeit zusätzlich durch dividiert wird. Nur wenn in der Zukunft sich die Parallelrechner durchsetzen, ist die biologische Bewertung des Fortschritts auch bei der Computeranwendung der Evolutionsstrategie richtig.





















