Embed Size (px)
Citation preview

MATERIAL UND METHODEN
4 Material und Methoden 4.1 Untersuchungsgebiet Das Untersuchungsgebiet ist Teil des Ravensberger Hügellandes und liegt zwischen den
Bielefelder Stadtteilen Schildesche und Jöllenbeck. Es handelt sich dabei um eine Reihe von
vier kleineren stehenden Gewässern (s.u.), die im Rahmen von Renaturierungsmaßnahmen
angelegt wurden und ca. 500 m nördlich des Schildescher Obersees am Zulauf der Jölle
lokalisiert sind. MEIER (1999) und REDECKER (1999) führten in diesem Gebiet bereits in den
Jahren 1998 – 1999 stichpunktartig Untersuchungen zum Vorkommen von Trematodenlar-
ven durch.
Das Renaturierungsgebiet liegt im Tal der Jölle und wird östlich von der Jölle und westlich
vom im Sommer trocken fallenden Wiesenbach begrenzt. Die genaue Lokalisation kann den
in den Arbeiten von MEIER (1999) und REDECKER (1999) veröffentlichten Karten entnommen
werden. Jenseits des Wiesenbachs erstreckt sich aus dem Tal ansteigend ein kleiner Rotbu-
chenbestand (Fagus sylvatica L.), östlich der Jölle wird das Tal von einigen Schwarzpappeln
(Populus nigra L.) und der erhöht liegenden Engerschen Straße begrenzt. Dem Lauf der Jöl-
le folgend finden sich Schwarzerlen (Alnus glutinosa (L.) Gaertner) und Weiden (Salix sp.).
Da sich die Jölle tief in ihr Bett eingegraben hat, das Tal ein deutliches Gefälle zum Obersee
hin aufweist und der Wiesenbach den Charakter eines Entwässerungsgrabens mit nur gerin-
gem Wasserstand besitzt, ist davon auszugehen, dass die Wasserkörper der stehenden
Gewässer unabhängig von den beiden Fließgewässern sind und lediglich von Regenwasser
gespeist werden.
Die Vegetation der unmittelbaren Umgebung der stehenden Gewässer ist gekennzeichnet
durch folgende Arten:
Alnus glutinosa (L.) Gaertner (Schwarzerle), Betula sp. (Birke), Rumex aquaticus L. (Wasserampfer), Epilobium hirsutum L. (Zottiges Weidenröschen), Iris pseudacorus L. (Sumpfschwertlilie), Phalaris arundinacea L. (Rohrglanzgras), Glyceria maxima (Hart-man) Holmberg (Wasserschwaden), Juncus effusus L. (Flatterbinse), Urtica dioica L. (Brennnessel), Galium aparine L. (Klettenlabkraut), Caltha palustris L. (Sumpfdotterblu-me), Lythrum salicaria L. (Blutweiderich), u.a.. Als Wasserpflanzen finden sich aspektbil-dend Elodea canadensis Michaux (Wasserpest), Ceratophyllum demersum L. (Rauhes Hornblatt) und Lemna minor L. (Kleine Wasserlinse).
Die Fauna des Renaturierungsgebiet weist keine Besonderheiten auf. Bezüglich potentieller
End- und Zwischenwirte von Trematoden ist ein weites Spektrum vorhanden. Als mögliche
1. Zwischenwirte wurden in den stehenden Gewässern neben L. stagnalis Linnaeus 1758
folgende Mollusken gefunden:
27

MATERIAL UND METHODEN
Planorbarius corneus L. (Posthornschnecke), Planorbis planorbis L., Planorbis carinatus Müller 1774, Anisus vortex L., Gyraulus albus Müller 1774, Bathyomphalus contortus L., Acroloxus lacustris L. (Teichnapfschnecke), Physa fontinalis L. (Quellenblasenschnecke), Radix baltica 4 L., Bithynia tentaculata L. und Sphaerium corneum L. (Kugelmuschel).
Am häufigsten gefunden wurden dabei P. corneus, A. lacustris und L. stagnalis.
Als potentielle 2. Zwischenwirte finden sich in den Gewässern neben den genannten Mollus-
ken eine Vielzahl von Arthropoden und ihren Larvalstadien. An dieser Stelle seien nur jene
aufgeführt, für die eine Funktion als Zwischenwirt im Labor nachgewiesen werden konnte:
Erpobdella octoculata L., Asellus aquaticus L., Simocephalus sp., Cloeon sp. Aeshna cy-anea Müller 1764, Chaoborus sp., Chironomidae.
Als potentielle Endwirte sind eine Vielzahl von Wirbeltieren in der Umgebung der Gewässer
und in ihnen anzutreffen. So konnte für Rana esculenta L. der Befall mit O. ranae Froelich
1791 nachgewiesen werden. Der in den untersuchten Gewässern häufige Trematode Asti-
otrema trituri Grabda 1959, als dessen Zwischenwirt P. corneus festgestellt wurde, nutzt
vermutlich den Teichmolch (Triturus vulgaris L.) als Endwirt (GRABDA, 1959). Auch für die
Erdkröte (Bufo bufo L.) sind verschiedene Trematoden beschrieben worden (SPIELER, 1990).
Sie nutzt die Tümpel und Weiher als Laichgewässer. Fische sind in den stehenden Gewäs-
sern nicht angetroffen worden. An Wasservögeln, die als Hauptwirte von Trematoden be-
kannt sind, wurden folgende Arten nachgewiesen: Anas platyrhynchos L. (Stockente), Ardea
cinerea L. (Fischreiher), Gallinula chloropus L. (Teichralle). Allerdings ist anzunehmen, dass
vereinzelt auch Wasservögel des nahe gelegenen Obersees das Renaturierungsgebiet und
seine Gewässer aufsuchen. Hier konnten zusätzlich folgende Wasservogelarten beobachtet
werden:
Aythya fuligula L. (Reiherente), Aythya ferina L. (Tafelente), Branta canadensis L. (Kana-dagans), Anser anser L. (Graugans), Cygnus olor Gmelin 1789 (Höckerschwan), Anas crecca L. (Krickente), Mergus serrator (Mittelsäger), Fulica atra L. (Bläßralle), Larus ridi-bundus Linnaeus 1766 (Lachmöwe), Podiceps cristatus L. (Haubentaucher).
Aber auch andere Vögel sind als potentielle Endwirte nicht auszuschließen. So wurden für
einige Plagiorchis-Arten verschiedene Hirundinidae als Endwirte nachgewiesen (BOCK &
JANSSEN, 1987, JANSSEN & BOCK, 1990). Auch Leucochloridium sp., der Succinea sp. als
Zwischenwirt nutzt, und dem verschiedene Singvögel als Endwirt dienen, ist in dem Gebiet
regelmäßig anzutreffen. Folgende Vögel wurden weiterhin nachgewiesen:
Accipiter nisus L. (Sperber), Milvus milvus L. (Rotmilan), Phasianus colchicus L. (Fasan), Alcedo atthis L. (Eisvogel), Corvus frugilegus L. (Saatkrähe), Prunella modularis L. (He-ckenbraunelle), Sylvia borin Boddaert 1783 (Gartengrasmücke), Sylvia atricapilla L. (Mönchsgrasmücke), Phylloscopus collybita Vieillot 1817 (Zilpzalp), Phoenicurus phoeni-
4 GLÖER (2002) beendet die Diskussion um die Validität der Arten Radix ovata Müller 1774 und Radix peregra Draparnaud 1805 und sieht sie als Varietäten von R. baltica Linnaeus 1758 an.
28

MATERIAL UND METHODEN
curus L. (Gartenrotschwanz), Erithacus rubecula L. (Rotkehlchen), Luscinia megarhyn-chos Brehm 1831 (Nachtigall), Turdus merula L. (Amsel), Turdus philomelos Brehm 1831 (Singdrossel), Fringilla coelebs L. (Buchfink), Carduelis chloris L. (Grünfink).
An Säugetieren wurden folgende Arten nachgewiesen:
Erinaceus europaeus L. (Igel), Crocidura sp., Nyctalus noctula Schreber 1774 (Abend-segler), Oryctolagus cuniculus L. (Wildkaninchen), Ondatra zibethicus Linnaeus 1766 (Bi-sam), Clethrionomys glareolus Schreber 1780 (Rötelmaus), Apodemus agrarius Pallas 1771 (Brandmaus), Sus scrofa L. (Wildschwein), Capreolus capreolus L. (Reh).
Obwohl zwischen den stehenden Gewässern im Untersuchungsgebiet nur eine geringe Dis-
tanz liegt, weisen sie zum Teil erhebliche ökologische Unterschiede auf.
Das südlichste dieser Gewässer hat
den Charakter eines Weihers, da er
ganzjährig Wasser führt und somit
nicht trocken fällt. Der Weiher weist
eine Fläche von ca. 80 m2 auf. Die
maximale Tiefe von 1,20 m gewähr-
leistet eine im Winter dauerhaft eis-
freie Zone am Grund des Teiches.
Der Uferbereich des Weihers ist stark
verkrautet, die Gewässermitte aber
ganzjährig vegetationsfrei. Aufgrund
des fehlenden Fischbesatzes weist
der Weiher eine hohe Dichte an Süsswassermollusken auf. Aspektbildend diesbezüglich sind
P. corneus, A. vortex und L. stagnalis. Neben den genannten Arten wurden auch S. corneum
und A. lacustris häufig angetroffen. Weiterhin wurden vereinzelt B. tentaculata, P. fontinalis
gefunden.
Abb. 4.1: Weiher J1 im Renaturierungsgebiet der Jölle (Mai 2001)
Die Untersuchungen der vorliegenden Arbeit wurden an diesem Weiher (J1) (s. Abb. 4.1)
durchgeführt, da er als einziger der stehenden Gewässer eine konstante Probennahme über
das ganze Jahr hinweg zuließ (s.u.).
Das Gewässer J2 liegt ca. 20 m nördlich von Weiher J1 und besitzt trotz einer ähnlich gro-
ßen Fläche wie J1 den Charakter eines Tümpels. Obwohl Weiher J1 und Tümpel J2 nahe
beieinander lokalisiert sind, ist ein Austausch der Schneckenpopulationen auszuschließen,
da keine wasserführende Verbindung besteht. So konnte in keinem Fall eine der entspre-
chend des Gewässers markierten L. stagnalis im anderen Gewässer nachgewiesen werden.
Die maximale Tiefe des Gewässers beträgt lediglich 80 cm, sodass ein Durchfrieren des
29

MATERIAL UND METHODEN
Gewässers in strengen Wintern möglich erscheint. Aufgrund der geringen Wassertiefe ist die
gesamte Fläche des Tümpels mit E. canadensis dicht bewachsen, was Probenahmen von
Plankton bei den regelmäßig auftretenden niedrigen Wasserständen im Sommer nicht zu-
lässt. Tümpel J2 wird östlich und westlich von A. glutinosa und Betula sp. gesäumt und weist
daher eine ganztägige Beschattung auf.
Die Molluskenfauna des Tümpels unterscheidet sich von der in Weiher J1 vorgefundenen.
So ist die Besatzdichte von P. corneus und L. stagnalis deutlich geringer, die Anzahl von
A. vortex deutlich höher als in J1. B. tentaculata konnte nicht nachgewiesen werden, wohin-
gegen P. planorbis, P. carinatus, G. albus, B. contortus, R. baltica und P. fontinalis regelmä-
ßig anzutreffen sind. Aufgrund der geringen Distanz zwischen den Gewässern J1 und J2 ist
davon auszugehen, dass Endwirte zu beiden gleichermaßen Zugang haben. Es ist daher
anzunehmen, dass die in den Schnecken persistierenden Trematoden von den gleichen
Endwirten in die Gewässer eingebracht wurden. Tatsächlich konnten innerhalb der Mollus-
kenarten die gleichen Trematodenarten nachgewiesen werden, wenn auch mit unterschiedli-
cher Prävalenz (s. Kap. 5.4.2). Tümpel J2 wurde daher für Vergleichsuntersuchungen he-
rangezogen, die, wären sie in Weiher J1 durchgeführt worden, aufgrund der notwendigen
Tötung der Schnecken zu einer Verfälschung der dort parallel laufenden Probenahmen ge-
führt hätten.
Die beiden weiteren Gewässer des Renaturierungsbereiches sollen an dieser Stelle nur kurz
Erwähnung finden, da sie zu den Untersuchungen in keiner direkten Beziehung standen.
Der nächstliegende Tümpel befindet sich ca. 25 m nordwestlich von J2. Er ist sehr flach und
führt nur bis ca. Juli/August Wasser. Nur in regenreichen Sommern trocknet er vermutlich
nicht aus. Süsswasserschnecken sind in diesem Tümpel nicht anzutreffen.
Weitere 20 m nördlich befindet sich ein Weiher, der durch einen angrenzenden Bestand von
Schwarzerlen (A. glutinosa) stark beschattet wird. Trotz seiner nur geringen Tiefe von maxi-
mal 60 cm führt er daher ganzjährig Wasser. Ein Durchfrieren bis zum Grund in starken Win-
tern ist anzunehmen. Der Weiher wird im Laufe des Jahres von einer dichten Schicht aus
L. minor vollständig bedeckt, was die Entnahme von Planktonproben verhinderte. Die
Schneckenfauna des Weihers setzt sich ähnlich wie in Tümpel J2 zusammen, allerdings
bleiben die Individuen deutlich kleiner. Vereinzelt wurde auch dieser Tümpel beprobt, um
Aufschluss über die dort vorkommende Trematodenfauna zu erhalten.
30

MATERIAL UND METHODEN
4.2 Material 4.2.1 Untersuchungsmaterial 4.2.1.1 Planktonproben Die Planktonproben wurden mit einem Planktonnetz (Maschenweite: 150 µm) gewonnen,
das am Ende einer Teleskopstange befestigt war. Die Proben wurden dabei oberflächennah
gewonnen, d.h. das Planktonnetz wurde nur zur Hälfte eingetaucht und mit seihender Bewe-
gung durch das Wasser geführt. Es wurde jeweils eine Strecke von ca. 15 m beprobt, wobei
5 Schwünge von jeweils ca. 3 m durchgeführt wurden. Die dabei abgefischte Menge Teich-
wasser errechnete sich mit ca. 25 l. Die daraus gewonnenen Planktonproben hatten ein Vo-
lumen von 15 ml und wurden bis zur weiteren Untersuchung in 100 ml PE-Flaschen bei
-20°C aufbewahrt. An jedem Probentag wurden jeweils zwei Planktonproben aus unter-
schiedlichen Bereichen des Weihers entnommen. Während anfänglich bevorzugt in unmittel-
barer Nähe des Ufers beprobt wurde, musste im Laufe des Jahres aufgrund der zunehmen-
den Verkrautung des Weihers mehr auf die freiere Wasserfläche der Gewässermitte ausge-
wichen werden. Die dort vereinzelt anzutreffenden Schwimmpflanzen wie C. dermersum und
E. canadensis stellten bei der Probennahme kein Hindernis dar.
Nachfolgend fand die Untersuchung der Planktonproben auf T. ocellata mit Hilfe der Tricho-
bilharzia – nPCR statt (s. Kap. 4.3.4.2 und 4.3.6.2) und Verifikation positiver Proben mit einer
Restriktionsanalyse (s. Kap. 4.3.6.6) statt.
Da die gewonnen Planktonproben mit einer spezifischen nPCR auf T. ocellata hin untersucht
wurden, wurden besondere Maßnahmen getroffen um Kontaminationen der Proben durch
anhaftende Organismen vorheriger Probenahmen zu verhindern. Dazu wurden Planktonnetz
und Probenbehälter zunächst gründlich bei 70°C in der Geschirrspülmaschine gewaschen
und nachfolgend unter einem Sterisol®-Intensivstrahler Typ NN 15/44 der Fa. Heraeus No-
belight mit UV-C-Licht bei einer Leistung von 1900 µW/cm2 für 15 min dekontaminiert. Vorun-
tersuchungen mit anderen Gerätschaften (s. Kap 4.3.6.5) zeigten, dass nach dieser Proze-
dur mit Hilfe der angewandten nPCR keine DNA von T. ocellata mehr nachgewiesen werden
kann.
31

MATERIAL UND METHODEN
4.2.1.2 Schnecken Die für die verschiedenen Untersuchungen benötigten L. stagnalis wurden durch Aufsam-
meln dem untersuchten Tümpel entnommen. Dabei wurde unter Verwendung einer Wathose
der Teich im Uferbereich abgeschritten und angetroffene Schnecken eingesammelt. Hierbei
ist zu erwähnen, dass jede angetroffene L. stagnalis auch eingesammelt und im Labor unter-
sucht wurde, um die Bevorzugung einer bestimmten Größengruppe der Schnecken auszu-
schließen (s. Kap. 5.7.2).
Die Sammlungen und der nachfolgende Cercarienschlupftest wurden in der Regel von April
bis Oktober im Abstand von 7 -14 Tagen in Abhängigkeit von der Witterung (s.u.) genom-
men, sodass zumeist 2 Probenahmen pro Monat stattfanden. Da L. stagnalis in Abhängigkeit
von den Witterungsbedingungen erst im April im oberflächennahen Bereich des Gewässers
anzutreffen ist und bereits im September/Oktober wieder in tiefere Schichten abwandert,
konnte in diesen Monaten meist nur eine Probenahme durchgeführt werden. An sonnigen
Tagen konnten aufgrund der besseren Durchleuchtung des Gewässers die höchsten Stück-
zahlen entnommen werden, weshalb wenn möglich entsprechende Tage für Probennahmen
bevorzugt wurden. Lang andauernde Schlechtwetterperioden wie z.B. im August 2002 führ-
ten dabei ebenfalls zu einem Abweichen von der 2-maligen Probennahme pro Monat.
Ab dem Jahr 2002 wurde zum Auffinden größerer Stückzahlen von Schnecken eine Polarisa-
tionsbrille benutzt, die durch Filtern eines Teiles der Lichtreflexe der Wasseroberfläche einen
besseren Einblick in das Gewässer gewährleistete. Durch Verwendung dieser Brille konnte
die pro Jahr gesammelte Stückzahl in etwa verdoppelt werden (s. Kap. 5.6).
4.2.2 Nukleinsäuren Synthetisch hergestellte Oligonukleotide: Die als Primer verwendeten Oligonukleotide wurden von den Firmen Whatman Biometra,
Carl Roth GmbH und IIT GmbH bezogen.
32

MATERIAL UND METHODEN
4.2.2.1 Primer zur Sequenzierung der 18s rDNA von T. ocellata Zur Auswahl der Primer s. Kap. 4.3.5.2 Primer Nukleotidsequenz
18s-x1 5’ GGTTGATCCTGCCAGTAGTC 3’
to18s-x2 5’ GGAGAGGGAGCCTGAGAAAT 3’
to18s-x3 5’ CAGGCCTTTGTGCCTAGAAA 3’
18s-y1 5’ CTTCCGCAGGTTCACCTACG 3’
to18s-y2 5’ GTGCCGTCTGTCCCTCTAAG 3’
to18s-y3 5’ CCGTCTGTCCCTCTTAACCA 3’
4.2.2.2 Primer der Trichobilharzia – nPCR Zur Auswahl der Primer s. Kap. 4.3.6.1 Primer Nukleotidsequenz
toc1 5’ GGCAGTTTCGGCTGTCTCTGTTGA 3’
toc2 5’ CGACACCAATCCAACGGCTTCA 3’
toc3 5’ ACGGAGATGGGTGAGCTTGT 3’
toc4 5’ TCGATGTGCCGTCTGTCC 3’
4.2.2.3 DNA-Längenmarker
Abb. 4.2: Für alle in dieser Arbeit durchgeführten
Gelelektrophoresen wurde der DNA-
Längenmarker GeneRulerTM 100 bp DNA
Ladder Plus der Fa. MBI Fermentas GmbH
verwendet (s. Abb. 4.2).
Darstellung des DNA-Längenmarkers GeneRulerTM 100 bp DNA Ladder Plus
33

MATERIAL UND METHODEN
4.2.3 Enzyme 4.2.3.1 Restriktionsendonuklease
Enzym Herkunftsorgamismus Erkennungssequenz Lieferant
BamH I Bacillus amylolyquefa-ciens H
5' G^G A T C C 3'
3' C C T A G^G 5'
MBI Fermentas, St. Leon-Rot
4.2.3.2 Weitere Enzyme
Taq Polymerase Eppendorf AG, Hamburg
HotMaster® Taq Polymerase Eppendorf AG, Hamburg
Proteinase K Carl Roth GmbH, Karlsruhe
Ribonuklease A MBI Fermentas, St. Leon-Rot
4.2.4 Reagenzien und Verbrauchsmaterial 10x HotMaster® Taq Reaktionspuffer Eppendorf AG, Hamburg
10x Taq Reaktionspuffer Eppendorf AG, Hamburg
Agarose High Resolution Carl Roth GmbH, Karlsruhe
Bromphenolblau Carl Roth GmbH, Karlsruhe
Chloroform Carl Roth GmbH, Karlsruhe
CTAB (N-Cetyl-N,N,N-trimethyl-ammoniumbromid) Merck AG, Darmstadt
Desoxynukleotid – Set (dATP, dCTP, dGTP, dTTP) Eppendorf AG, Hamburg
EDTA Carl Roth GmbH, Karlsruhe
EDTA-Na2 Carl Roth GmbH, Karlsruhe
Einwegspritzen 2 ml mit stumpfer Kanüle Carl Roth GmbH, Karlsruhe
Eisessig Carl Roth GmbH, Karlsruhe
EtOH 99,8% Carl Roth GmbH, Karlsruhe
Ethidiumbromidlösung Carl Roth GmbH, Karlsruhe
Falcongefäß Rotilabo®-Zentrifugenröhrchen 15 ml Carl Roth GmbH, Karlsruhe
Isoamylalkohol Carl Roth GmbH, Karlsruhe
34
MATERIAL UND METHODEN
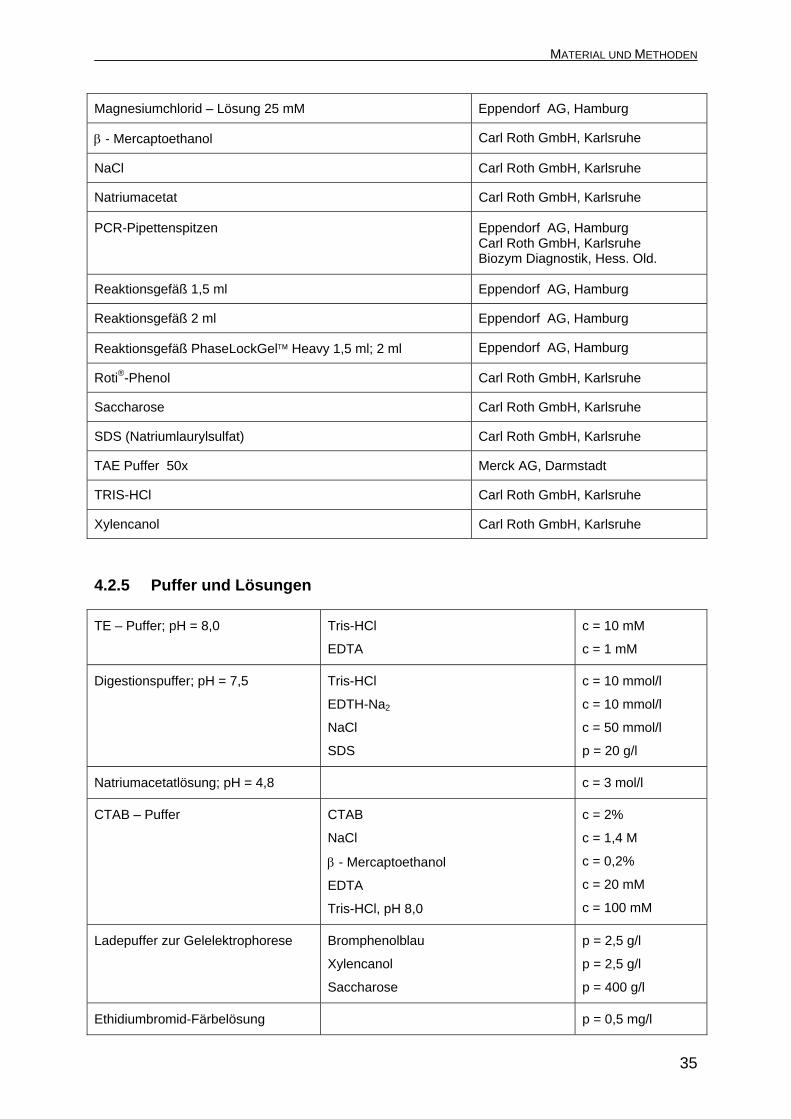
Magnesiumchlorid – Lösung 25 mM Eppendorf AG, Hamburg
β - Mercaptoethanol Carl Roth GmbH, Karlsruhe
NaCl Carl Roth GmbH, Karlsruhe
Natriumacetat Carl Roth GmbH, Karlsruhe
PCR-Pipettenspitzen Eppendorf AG, Hamburg Carl Roth GmbH, Karlsruhe Biozym Diagnostik, Hess. Old.
Reaktionsgefäß 1,5 ml Eppendorf AG, Hamburg
Reaktionsgefäß 2 ml Eppendorf AG, Hamburg
Reaktionsgefäß PhaseLockGel™ Heavy 1,5 ml; 2 ml Eppendorf AG, Hamburg
Roti®-Phenol Carl Roth GmbH, Karlsruhe
Saccharose Carl Roth GmbH, Karlsruhe
SDS (Natriumlaurylsulfat) Carl Roth GmbH, Karlsruhe
TAE Puffer 50x Merck AG, Darmstadt
TRIS-HCl Carl Roth GmbH, Karlsruhe
Xylencanol Carl Roth GmbH, Karlsruhe
4.2.5 Puffer und Lösungen
TE – Puffer; pH = 8,0 Tris-HCl
EDTA
c = 10 mM
c = 1 mM
Digestionspuffer; pH = 7,5 Tris-HCl
EDTH-Na2
NaCl
SDS
c = 10 mmol/l
c = 10 mmol/l
c = 50 mmol/l
p = 20 g/l
Natriumacetatlösung; pH = 4,8 c = 3 mol/l
CTAB – Puffer CTAB
NaCl
β - Mercaptoethanol
EDTA
Tris-HCl, pH 8,0
c = 2%
c = 1,4 M
c = 0,2%
c = 20 mM
c = 100 mM
Ladepuffer zur Gelelektrophorese Bromphenolblau
Xylencanol
Saccharose
p = 2,5 g/l
p = 2,5 g/l
p = 400 g/l
Ethidiumbromid-Färbelösung p = 0,5 mg/l
35
MATERIAL UND METHODEN
4.2.6 Geräte
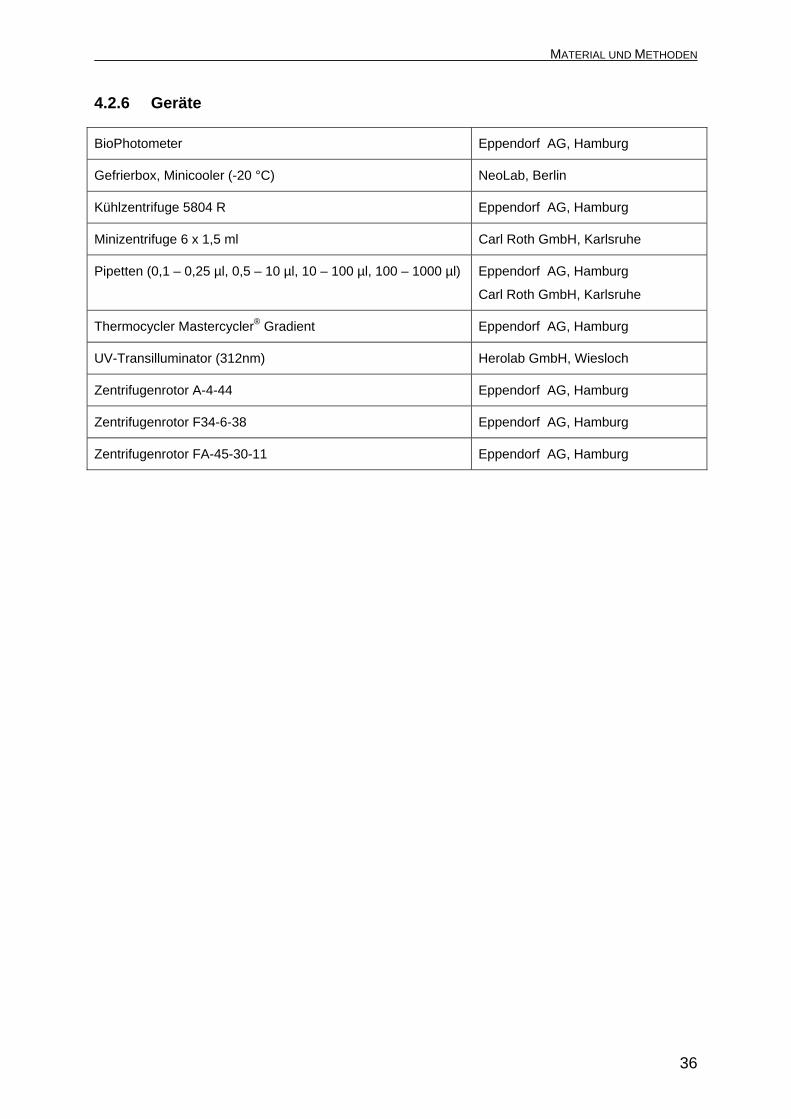
BioPhotometer Eppendorf AG, Hamburg
Gefrierbox, Minicooler (-20 °C) NeoLab, Berlin
Kühlzentrifuge 5804 R Eppendorf AG, Hamburg
Minizentrifuge 6 x 1,5 ml Carl Roth GmbH, Karlsruhe
Pipetten (0,1 – 0,25 µl, 0,5 – 10 µl, 10 – 100 µl, 100 – 1000 µl) Eppendorf AG, Hamburg
Carl Roth GmbH, Karlsruhe
Thermocycler Mastercycler® Gradient Eppendorf AG, Hamburg
UV-Transilluminator (312nm) Herolab GmbH, Wiesloch
Zentrifugenrotor A-4-44 Eppendorf AG, Hamburg
Zentrifugenrotor F34-6-38 Eppendorf AG, Hamburg
Zentrifugenrotor FA-45-30-11 Eppendorf AG, Hamburg
36

MATERIAL UND METHODEN
4.3 Methoden 4.3.1 Sammlung und Markierung von L. stagnalis Die regelmäßige Sammlung von L. stagnalis wurde durchgeführt (s. Kap. 4.2.1.2), um über
den Jahresverlauf Infektionsraten der Schnecken mit Trematoden zu erheben (s. Kap. 4.2.1).
Gleichzeitig dienten das regelmäßige Sammeln und das Anbringen einer individuellen Mar-
kierung der Schnecken (s.u.) dazu, Erkenntnisse zum Lebenszyklus dieser Schnecken im
Untersuchungsgebiet zu gewinnen. Insbesondere sollten dabei Erkenntnisse zur
Lebensdauer sowie der aktiven Phase5 von L. stagnalis gewonnen werden. Die Erforschung des Lebenszyklus von L. stagnalis scheint mit Ende der 70er Jahre ab-geschlossen zu sein, da seit diesem Zeitpunkt keine weiteren Veröffentlichungen zur Bio-logie dieser Schnecke erschienen sind. Dies ist verwunderlich, da die existierende Litera-tur durchaus unterschiedliche Lebenszyklen beschreibt. BAKER (1911) beschrieb für die Art Lebensdauern von 4 – 5 Jahren, während MCDONALD (1969) in ihrer Untersuchung i.d.R. eine ca. einjährige Lebensdauer von L. stagnalis beobachtete, wobei die Adulti im Spätsommer zur Eiablage gelangten und nachfolgend starben. Die überwinternden Jung-schnecken pflanzten sich dann im nachfolgenden Jahr fort. BERRIE (1965) untersuchte Populationen in Schottland und fand unter den dort vorherrschenden klimatischen Bedin-gungen obligatorisch einen mehrjährigen Lebenszyklus von L. stagnalis. Er sah dies als Anpassung an die dortigen klimatischen Verhältnisse, die bei den vorherrschenden kur-zen Sommern die Schnecken i.d.R. erst nach mehreren Jahren zur Geschlechtsreife ge-langen lassen. CALOW (1978) ordnet in seiner Arbeit L. stagnalis einen ein- bis mehrjähri-gen Lebenszyklus zu mit Fortpflanzung jeweils im Frühjahr. BOAG & PEARLSTONE (1979) untersuchten eine Population von L. stagnalis in Alberta (Kanada) und beschrieben obli-gatorisch mehrjährige Zyklen für die Schnecken mit einer erstmaligen Oviposition im drit-ten Sommer.
Die Erfassung von Infektionsraten bei gleichzeitiger Markierung von Schneckenindividuen
sollte die Verfolgung der Verläufe von Trematodeninfektionen in einzelnen Schnecken er-
möglichen. Dies diente dem Erhalt weiterer epidemiologischer Erkenntnisse, wie z.B. der
Duration erworbener Infektionen, ob der Parasit in der Schnecke überwintert, und ob inner-
halb der Schnecke Verdrängungsprozesse zwischen den Trematoden im Sinne einer inter-
spezifischen Konkurrenz beobachtet werden können.
Die Sammlung und Markierung der Schnecken wurden in den Jahren 2001 und 2002 durch-
geführt. Für das Jahr 2003 wurden die Markierung und Vermessung nicht weitergeführt, da
im strengen Winter 2002 / 2003 ein Großteil der adulten Schnecken starb. Infektionsraten
wurden für dieses Jahr aber weiterhin ermittelt.
5 Zeitraum, während dessen L. stagnalis die Überwinterungszone in den eisfreien Regionen des Ge-wässers verlässt und im oberflächennahen Bereich des Gewässers anzutreffen ist. Infektionen von L. stagnalis finden vermutlich bevorzugt in den oberflächennahen Bereichen des Gewässers statt.
37

MATERIAL UND METHODEN
Die dem Gewässer entnommenen Schnecken (s. Kap. 4.2.1.2) wurden zunächst in einem
Kunststoffaquarium gehältert, mit Kopfsalat gefüttert und über Nacht bei 4°C im Kühlraum
aufbewahrt. Am nachfolgenden Tag wurde dann die Ge-
häusehöhe der Schnecken ermittelt und das Gehäuse von
Algenaufwuchs mit Hilfe einer Pinzette schabend gerei-
nigt. Die Vermessung der Gehäusehöhen erfolgte mit ei-
nem Messschieber auf 5-Zehntel cm genau. Abb. 4.3
zeigt schematisch die Ermittlung der Gehäusehöhe. Die
Ermittlung der Gehäusehöhe war dabei zunächst nicht zur
Protokollierung des Schneckenwachstums, sondern viel-
mehr als zusätzlicher Anhaltspunkt bei der Identifizierung
der Schnecken gedacht. Allerdings erwiesen sich die Mar-
kierungen als ausgesprochen dauerhaft. So konnte z.B.
eine Schnecke (Nr. 0042, s. Anh. S. 159) nach 13 Mona-
ten erstmalig wieder eingesammelt werden und war an-
hand ihrer Beschriftung problemlos zu identifizieren.
Abb. 4.3: Ermittlung der Gehäusehöhe bei L. stagnalis.
Das gesäuberte und trockene Gehäuse wurde im Anschluss mit Hilfe eines weißen Lackstif-
tes Nr. 751 der Fa. Edding mit einer individuellen Nummer versehen. Die im Laufe der Un-
tersuchung wieder gefundenen Schnecken wurden protokolliert und bei Bedarf die Numme-
rierung erneuert. Die Schnecken wurden am Untersuchungstag oder am darauf folgenden
Tag wieder in ihr Ursprungsgewässer ausgesetzt. Bei Aussetzen am nächsten Tag wurden
die Schnecken eine weitere Nacht wie oben beschrieben gehalten.
4.3.2 Cercarienschlupftest
4.3.2.1 Durchführung Der Cercarienschlupftest wurde im Anschluss an das Sammeln und Markieren (s. Kap. 4.3.1)
durchgeführt. Dieser Test weist eine Trematodeninfektion anhand der aus der Schnecke
austretenden Larvalstadien (Cercarien) nach. Die so ermittelte Infektionsrate trifft eine Aus-
sage über die Patenz der Infektionen im Zwischenwirt. Präpatente oder ruhende Infektionen
werden durch den Cercarienschlupftest nicht nachgewiesen. Die Ermittlung patenter Infekti-
onen kann daher zur Abschätzung eines tatsächlichen Infektionsrisikos herangezogen wer-
den.
38

MATERIAL UND METHODEN
Für die Untersuchung wurden die Schnecken zunächst einzeln in Makrolongefäße gegeben
und mit Wasser bedeckt. Anschließend wurden die Schnecken mit einer 1500 Lux Tages-
lichtanlage bei 26°C in einer Klimakammer für ca. 2 Stunden bestrahlt. Die Veränderung der
Tageslichtstärke in Kombination mit der erhöhten Temperatur induziert den Cercarienschlupf
aller in L. stagnalis im untersuchten Weiher vorgefundenen Trematoden. Anschließend er-
folgt die Untersuchung auf ausgetretene Cercarien unter dem Binokular. Zur weiteren Be-
stimmung der Cercarien (s. Kap. 4.3.2.2) wurde ein Stereomikroskop benutzt.
Eine andere Möglichkeit zur Ermittlung einer Infektionsrate ist die Sektion der Schnecken,
die neben den patenten Infektionen auch nicht patente erfasst (s. Kap. 4.3.3 und 5.2). Da
aber in dieser Untersuchung über einen längeren Zeitraum hinweg Infektionsraten ermittelt
werden sollten, konnte lediglich der Cercarienschlupftest durchgeführt werden, da die stän-
dige Entnahme großer Stückzahlen von L. stagnalis für eine Sektion einen massiven Eingriff
in die Population dargestellt hätte.
4.3.2.2 Zur Bestimmung der in L. stagnalis gefundenen Cercarien
Die Artbestimmung der Trematoden anhand des Larvalstadiums der Cercarie blickt auf eine
lange Tradition zurück. Erste Beschreibungen datieren bereits in das 18. Jahrhundert. An-
fang des 20. Jahrhunderts und vermutlich stimuliert durch die Entdeckung des Bilharziose-
Erregers Schistosoma haematobium durch Theodor Bilharz im Jahre 1851 wurden erste um-
fassende systematische Einordnungen sowohl der Adulti als auch der Cercarien publiziert
(LÜHE, 1909). Die Einordnung erfolgte dabei anhand morphologischer Merkmale, die als art-
spezifisch und konstant angesehen wurden. Für adulte Trematoden sind dies besonders die
Anordnung, Struktur und relative Position der Geschlechtsorgane sowie die Struktur des Ex-
kretionssystems. Weiterhin wurden die Körperabmessungen, besondere charakteristische
Merkmale wie z.B. zusätzliche Haftorgane sowie Position und Struktur der Saugnäpfe be-
rücksichtigt. Nachfolgend soll an dieser Stelle auf die Artbestimmung anhand von Cercarien
ausführlicher und kritisch eingegangen werden, da sie für die vorliegende Arbeit von beson-
derer Bedeutung war.
Cercarien wurden erstmalig von LÜHE (1909) aufgrund bestimmter morphologischer Merkma-
le in Hauptgruppen unterteilt. Basierend auf Lühe´s System ordnete DUBOIS (1929) Cercarien
zehn Typen (s. Tab. 4.1) zu, die noch heute weitgehend Verwendung finden. Dabei ist an-
zumerken, dass diese Klassifizierung lediglich zu Bestimmungszwecken herangezogen wird,
da eine Übereinstimmung dieser Cercarientypen mit der gültigen Taxonomie nicht immer
gegeben ist, d.h. die erwähnten 10 Gruppen spiegeln keine entsprechenden Taxa wieder.
39
MATERIAL UND METHODEN
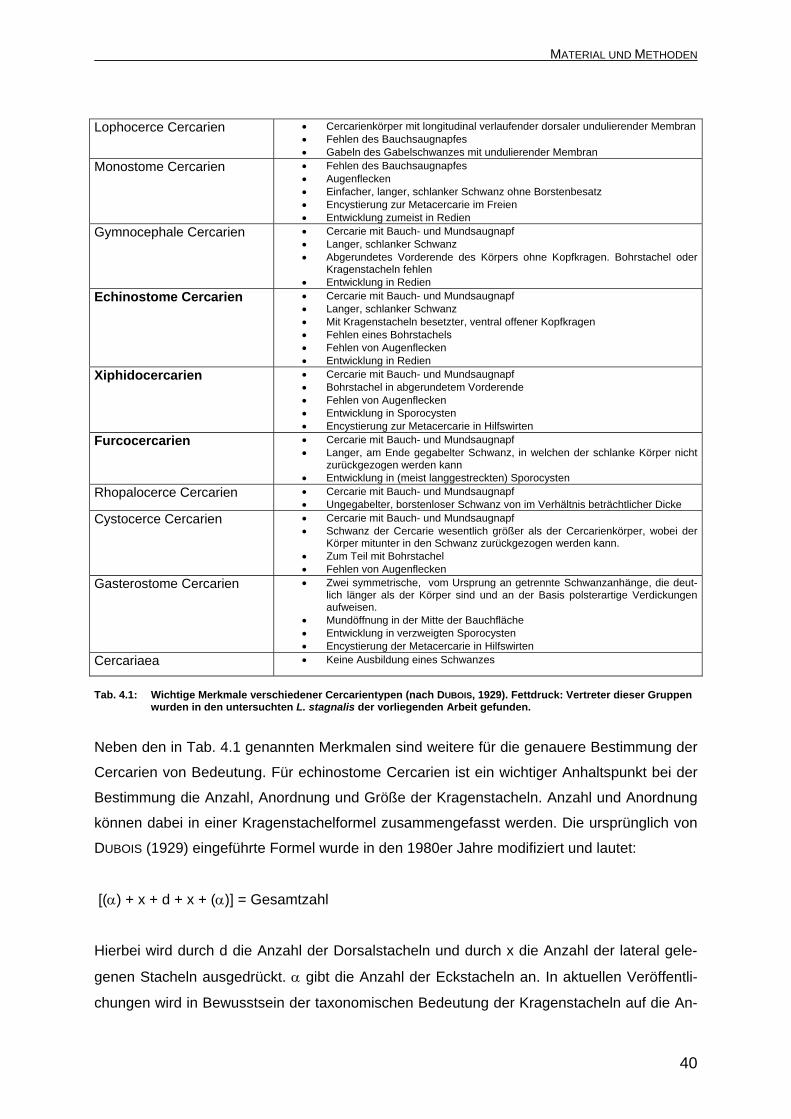
Lophocerce Cercarien • Cercarienkörper mit longitudinal verlaufender dorsaler undulierender Membran • Fehlen des Bauchsaugnapfes • Gabeln des Gabelschwanzes mit undulierender Membran
Monostome Cercarien • Fehlen des Bauchsaugnapfes • Augenflecken • Einfacher, langer, schlanker Schwanz ohne Borstenbesatz • Encystierung zur Metacercarie im Freien • Entwicklung zumeist in Redien
Gymnocephale Cercarien • Cercarie mit Bauch- und Mundsaugnapf • Langer, schlanker Schwanz • Abgerundetes Vorderende des Körpers ohne Kopfkragen. Bohrstachel oder
Kragenstacheln fehlen • Entwicklung in Redien
Echinostome Cercarien • Cercarie mit Bauch- und Mundsaugnapf • Langer, schlanker Schwanz • Mit Kragenstacheln besetzter, ventral offener Kopfkragen • Fehlen eines Bohrstachels • Fehlen von Augenflecken • Entwicklung in Redien
Xiphidocercarien • Cercarie mit Bauch- und Mundsaugnapf • Bohrstachel in abgerundetem Vorderende • Fehlen von Augenflecken • Entwicklung in Sporocysten • Encystierung zur Metacercarie in Hilfswirten
Furcocercarien • Cercarie mit Bauch- und Mundsaugnapf • Langer, am Ende gegabelter Schwanz, in welchen der schlanke Körper nicht
zurückgezogen werden kann • Entwicklung in (meist langgestreckten) Sporocysten
Rhopalocerce Cercarien • Cercarie mit Bauch- und Mundsaugnapf • Ungegabelter, borstenloser Schwanz von im Verhältnis beträchtlicher Dicke
Cystocerce Cercarien • Cercarie mit Bauch- und Mundsaugnapf • Schwanz der Cercarie wesentlich größer als der Cercarienkörper, wobei der
Körper mitunter in den Schwanz zurückgezogen werden kann. • Zum Teil mit Bohrstachel • Fehlen von Augenflecken
Gasterostome Cercarien • Zwei symmetrische, vom Ursprung an getrennte Schwanzanhänge, die deut-lich länger als der Körper sind und an der Basis polsterartige Verdickungen aufweisen.
• Mundöffnung in der Mitte der Bauchfläche • Entwicklung in verzweigten Sporocysten • Encystierung der Metacercarie in Hilfswirten
Cercariaea • Keine Ausbildung eines Schwanzes
Tab. 4.1: Wichtige Merkmale verschiedener Cercarientypen (nach DUBOIS, 1929). Fettdruck: Vertreter dieser Gruppen
wurden in den untersuchten L. stagnalis der vorliegenden Arbeit gefunden.
Neben den in Tab. 4.1 genannten Merkmalen sind weitere für die genauere Bestimmung der
Cercarien von Bedeutung. Für echinostome Cercarien ist ein wichtiger Anhaltspunkt bei der
Bestimmung die Anzahl, Anordnung und Größe der Kragenstacheln. Anzahl und Anordnung
können dabei in einer Kragenstachelformel zusammengefasst werden. Die ursprünglich von
DUBOIS (1929) eingeführte Formel wurde in den 1980er Jahre modifiziert und lautet:
[(α) + x + d + x + (α)] = Gesamtzahl
Hierbei wird durch d die Anzahl der Dorsalstacheln und durch x die Anzahl der lateral gele-
genen Stacheln ausgedrückt. α gibt die Anzahl der Eckstacheln an. In aktuellen Veröffentli-
chungen wird in Bewusstsein der taxonomischen Bedeutung der Kragenstacheln auf die An-
40

MATERIAL UND METHODEN
gabe einer Kragenstachelformel, zugunsten ausführlicher Beschreibungen mit gleichzeitiger
Angabe der Größe der Stacheln, zumeist verzichtet.
Als weiteres für verschiedene Cercariengruppen taxonomisch wertvolles Merkmal wird die
Struktur des Exkretionssystems angesehen (ODENING, 1962, 1971). Anzahl und Anordnung
der Protonephridien lassen sich dabei in einer Protonephridialformel ausdrücken (FAUST,
1932).
So bedeutet beispielsweise die Protonephridialformel 2{[1+1+1]+[3+(1)]} = 14 für T. o-cellata, dass drei einzelne Protonephridien unabhängig voneinander in den vorderen Ex-kretionsgang ableiten [1+1+1]. Die Exkretionsgänge drei weiterer Terminalzellen laufen zunächst zusammen, um dann gemeinsam in den hinteren Exkretionsgang abzuleiten [3]. Zwei zusätzliche im Schwanzstamm gelegene Protonephridien entsorgen ihre Exkrete ebenfalls in dieses Gefäß (2). Im Schwanzstamm gelegene Protonephridien werden durch Klammern gekennzeichnet. Da das Exkretionssystem paarig angelegt ist ergibt sich die Summe von 14 Flammenzellen. Über Art und Struktur der Exkretionsblase, in die sich die Exkretionsgänge entleeren, macht die Protonephridialformel keine Aussage.
Weitere wichtige bei der Bestimmung herangezogene Merkmale können die Anordnung und
Beschaffenheit von Penetrationsdrüsen und ihren Ausführgängen sowie das Vorkommen
von Flossensäumen am Cercarienschwanz, sowie die Form und Größe von Stiletten sein.
Die Bestimmung von Trematodenspezies anhand der Cercarien ist aus verschiedenen
Gründen problematisch und führt nur in wenigen Fällen bis zur Art.
Zunächst fehlen brauchbare Bestimmungsschlüssel für Cercarien. Mit den existierenden
Schlüsseln sind z.T. nur grobe Einordnungen möglich und zumeist beziehen sie sich nur auf
bestimmte Regionen, zum Teil nur auf bestimmte Gewässer (DUBOIS, 1929, KHAN, 1961,
KHAN, 1962, ZAJICEK, 1963, MEYER, 1964, NASIR & ERASMUS, 1964, REIMER, 1971, BOCK,
1980), sodass nach erfolgter grober Bestimmung jeweils die Originalliteratur der Beschrei-
bung der vermuteten Arten herangezogen werden muss. Da die modernen computergestütz-
ten Systeme der Literatursuche zumeist Publikationen vor den 1980er Jahren nicht oder nur
teilweise erfassen, viele der Erstbeschreibungen aber deutlich früher veröffentlicht wurden,
sind Beschreibungen von Trematodenlarven häufig nicht auffindbar. Dies gilt besonders für
frühe Publikationen aus dem ehemaligen Ostblock, wo zum Teil umfassende Forschung zu
Trematoden betrieben wurde.
Häufig sind Cercarien nah verwandter Arten (z.B. innerhalb einer Gattung) nahezu identisch
aufgebaut. So sind beispielsweise die Cercarien der Gattung Trichobilharzia aufgrund ihrer
morphologisch-anatomischen Merkmale keiner Art zuzuordnen (SZIDAT, 1942, DÖNGES,
1965, BLAIR & ISLAM, 1983, MÜLLER & KIMMIG, 1994, HORAK et al., 1998). Ein wichtiger An-
41

MATERIAL UND METHODEN
haltspunkt bei genauerer Zuordnung identischer Cercarien können die Zwischenwirtschne-
cken sein, da Trematoden gegenüber dem ersten Zwischenwirt häufig eine ausgeprägte
Wirtsspezifität aufweisen. Eine verlässliche Bestimmung von Trematoden kann nach MÜLLER
& KIMMIG (1994) jedoch nur eine komplette Aufdeckung des Entwicklungszyklus unter Be-
rücksichtigung aller Wirte und Entwicklungsstadien liefern, da auch die Adulti nah verwandter
Arten mitunter morphologisch identisch sind (KANEV et al., 1998). In der Praxis ist eine sol-
che Forderung aufgrund der Komplexität der Lebenszyklen der Digenea nur in den seltens-
ten Fällen realisierbar.
Die Bestimmung der Cercarien wird weiterhin erschwert durch die lange Tradition der Cerca-
rienbestimmung (s.o.). In den Anfängen bestand keine einheitliche Methode der Beschrei-
bung, weshalb für die Bestimmung der Cercarien unterschiedliche Merkmale herangezogen
wurden. Wichtigstes Bestimmungsmerkmal waren, neben der Angabe auffälliger Besonder-
heiten wie Stilette, Kragenstacheln, Saugnäpfe etc., die Körperabmessungen, welche heut-
zutage aufgrund der hohen Variabilität und der hohen Verformbarkeit des Trematodenkör-
pers nur von zweitrangiger Bedeutung sind. Auch die unterschiedlichen Methoden der Prä-
paration führten zu abweichenden Ergebnissen bei der Betrachtung morphologischer Merk-
male. Eine Zuordnung der damaligen Beschreibungen zu heute gefundenen Arten ist daher
nur in Einzelfällen möglich. In der Folge resultierten aus der uneinheitlichen Beschreibung
der Cercarien eine Vielzahl von Doppelt- und Mehrfachbeschreibungen, die eine Bestim-
mung heute zusätzlich erschweren, zumal gleiches für die Adulti gilt. So sind nach KANEV
(1994) allein für Echinostoma revolutum vier Synonyme (s.u.) und 47 falsche Zuordnungen in
wissenschaftlichen Arbeiten (auch jüngeren Datums) veröffentlicht.
42

MATERIAL UND METHODEN
4.3.3 Vergleich verschiedener Methoden der Erfassung von Infektions- raten: Cercarienschlupftest, Sektion, nPCR
Der Vergleich der Methoden zur Erfassung von Infektionsraten wurde in zwei aufeinander
folgenden Untersuchungen durchgeführt.
4.3.3.1 Vergleich der Eignung von Cercarienschlupftest und Sektion bei Ermittlung von Infektionsraten von L. stagnalis in Tümpel J2
Da die Folgen der nachhaltigen Entnahme von Schnecken aus dem Gewässer nicht abzu-
schätzen waren und eine dadurch bedingte Veränderung der Schneckenpopulation und der
Zusammensetzung der Trematodenfauna wahrscheinlich war, wurde die Untersuchung nicht
an Weiher J1, sondern wie beschrieben an Tümpel J2 durchgeführt (vgl. Kap. 4.1).
Zunächst wurden im dem Weiher J1 benachbarten Tümpel J2 für die Jahre 2001 und 2002
Schnecken, wie in Kap. 4.3.1 beschrieben, gesammelt und zunächst mit dem Cerca-
rienschlupftest untersucht. Nachfolgend wurde dann eine Sektion der Schnecken durchge-
führt und nicht patente Infektionen erfasst. Dabei wird davon ausgegangen, dass patente
Infektionen immer auch durch eine Sektion nachgewiesen werden können. Für alle der in
L. stagnalis gefundenen Trematoden trifft diese Voraussetzung zu, da sie den Bereich der
Gonaden und der Mitteldarmdrüse besiedeln und dabei das Platzangebot komplett aus-
schöpfen. Entsprechend lassen sich patente Infektionen bei einer Sektion einfach nachwei-
sen.
4.3.3.2 Vergleich der Eignung von Cercarienschlupftest, Sektion und nPCR bei Ermittlung von Infektionsraten am Beispiel T. ocellata – infizierter L. stagnalis aus Weiher J1
Diese Untersuchung wurde zum Ende der Beprobung des Tümpels J1 im September 2003
durchgeführt, da sich in Tümpel J2 der Bestand an L. stagnalis nach dem Winter 2002 / 2003
zum Untersuchungszeitpunkt noch nicht erholt hatte. Die bei der letzten Probennahme ge-
sammelten 261 Schnecken wurden zunächst mit Hilfe des Cercarienschlupftest auf Trema-
todenbefall getestet. Von diesen erwiesen sich 99 Schnecken als infiziert. Da davon ausge-
gangen werden kann, dass diese Infektionen auch per Sektion entdeckt worden wären, wur-
den diese 99 Schnecken wieder in das Gewässer ausgesetzt. Die verbleibenden 162 Schne-
cken wurden bis zur weiteren Verarbeitung bei -20°C eingefroren. Nach Auftauen wurden
sie per Sektion auf Trematodenbefall untersucht und auftretende Infektionen protokolliert.
Nach Entfernen des Magen-Darm-Traktes wurden die Gewebeproben dann einzeln in Auf-
bewahrungsbehälter gegeben und bis zur Untersuchung durch die Trichobilharzia – nPCR
43

MATERIAL UND METHODEN
(s. Kap. 4.3.6) eingefroren. Die zur Präparation genutzten Utensilien wurden dabei jeweils
nur einmal benutzt, um Kontaminationen zwischen den Proben zu vermeiden. Anschließend
wurden sie, wie in Kap. 4.3.6.5 beschrieben, dekontaminiert und erneut verwendet. Die DNA-
Extraktion wurde wie in Kap. 4.3.4.3 beschrieben durchgeführt. Die nPCR folgte der Vorge-
hensweise, wie in Kap. 4.3.6.2 dargestellt. Zur Verifikation T. ocellata – positiver Proben
wurde anschließend eine Restriktionsanalyse der Amplifikate der 2. PCR-Runde durchge-
führt. Es sei darauf hingewiesen, dass die Untersuchung der Proben mit der Trichobilharzia –
nPCR selbstverständlich nur Infektionen von T. ocellata nachweisen konnte. Für die anderen
in L. stagnalis diagnostizierten Trematoden existiert bis jetzt kein entsprechendes Diagnose-
verfahren.
4.3.4 Nukleinsäureextraktion 4.3.4.1 Nukleinsäureexktraktion aus Cercarien
1. Proben aus Ethanol oder Formol werden zweimal mit 500 µl Digestionspuffer gewaschen. Fri-sche Proben finden direkt Verwendung.
2. Probe mit 100 µl Digestionspuffer versetzen und homogenisieren.
3. Proteinase K (200 µl/ml) zugeben und 60 min bei 37°C inkubieren.
4. Mit 100 µl Na-Acetat-Lsg. versetzen und 15 min auf Eis inkubieren.
5. 200 µl Probenmenge in PhaseLockGel-Reaktionsgefäß 1,5 ml geben, mit 200 µl Roti®-Phenol versetzen und sanft schütteln. Anschließend 10 min bei 14000 rpm zentrifugie-ren.
6. Obere Phase in ein 1,5 ml Reaktionsgefäß pipettieren.
7. Mit 200 µl Chloroform/Isoamylalkohol 24:1 versetzen und sanft schütteln. 10 min bei 14000 rpm (FA-45-30-11) zentrifugieren.
8. Obere Phase in ein 1,5 ml Reaktionsgefäß pipettieren.
9. Probe mit doppelter Menge 98% EtOH (-20°C) versetzen und Prezipitation bei – 20 °C über Nacht.
10. Probe 15 min bei 4°C und 14000 rpm (FA-45-30-11) zentrifugieren und Überstand entfernen.
11. Probe mit 500 µl 70% EtOH (-20°C) für 10 min in Gefrierbox (-20°C) inkubieren.
12. Probe für 15 min bei 4°C mit 14000 rpm (FA-45-30-11) zentrifugieren und EtOH vollständig mit Einwegspritzen entfernen. Anschließend 7 min bei 60°C inkubieren.
13. Mit 100 µl TE-Puffer für 10 min bei 50°C im Wasserbad inkubieren und anschließend bei -20°C einfrieren.
44

MATERIAL UND METHODEN
4.3.4.2 Nukleinsäureexktraktion aus Plankton Die Nukleinsäureextraktion aus Plankton folgt einem veränderten Protokoll nach WINNEPEN-
NINCKX et al. (1993). Das Verfahren basiert dabei auf der Fähigkeit von N-Cetyl-N,N,N-
trimethyl-ammoniumbromid (CTAB) bei hohen Salzkonzentrationen bevorzugt Proteine und
Polysaccharide im Vergleich zu DNA zu binden.
1. Die Planktonprobe wurde in ein 15 ml Falcon-Gefäß überführt und 10 min bei 800 rpm im Aus-
schwenkrotor (A-4-44) abzentrifugiert.
2. Verwerfen des Überstands und versetzen der Probe mit 1,5 ml erhitztem CTAB-Puffer (60°C).
3. Auffüllen der Probe mit CTAB-Puffer (60°C) auf 3 ml und Zusatz von 5 mAU/ml Proteinase K.
4. Erhitzen der Probe für 60 min bei 60°C.
5. Zentrifugation für 10 min bei 8000 rpm im Festwinkelrotor (F34-6-38).
6. 0,9 ml Überstand mit 0,9 ml 3M Na-Acetat-Lösung in ein 2 ml Reaktionsgefäß pipettieren und 10 min auf Eis inkubieren.
7. 650 µl der Probe mit 450 µl Roti®-Phenol in ein 2 ml PhaseLockGel-Reaktionsgefäß geben und sanft schütteln.
8. Zentrifugation für 10 min bei 14000 rpm im Festwinkelrotor (FA-45-30-11).
9. Überstand in ein 2 ml Reaktionsgefäß pipettieren und mit doppelter Menge Chloro-form/Isoamylalkohol 24:1 versetzen.
10. Zentrifugation für 10 min bei 14000 rpm im Festwinkelrotor (FA-45-30-11).
11. Obere Phase in ein 1,5 ml Reaktionsgefäß pipettieren und mit doppelter Menge EtOH 98% versetzen.
12. Inkubation der Probe über Nacht bei – 20°C.
13. Zentrifugation der Probe für 15 min bei 4°C und 14000 rpm (FA-45-30-11).
14. Überstand vom Pellet abnehmen, Probe mit 500 µl EtOH 70% versetzen und 10 min bei – 20°C in der Gefrierbox inkubieren.
15. Zentrifugation der Probe für 15 min bei 4°C und 14000 rpm (FA-45-30-11).
16. EtOH vollständig mit Einwegspritze entfernen und Trocknung der DNA für 7 min bei 60°C.
17. Probe mit 100 µl TE-Puffer und 50 U Ribonuklease A für 15 min bei 37°C inkubieren und an-schließend bis zur Verwendung bei – 20°C lagern.
45

MATERIAL UND METHODEN
4.3.4.3 Nukleinsäureexktraktion aus Schneckengewebe
Die Nukleinsäureextraktion aus Schneckengewebe folgt einem veränderten Protokoll
nach WINNEPENNINCKX et al. (1993) (vgl. Kap. 4.3.4.2).
1. Entfernen des Schneckengehäuses.
2. Schnecken mit einer Gehäusehöhe < 2 cm wurden im Ganzen weiterverarbeitet. Größere Schnecken wurden seziert und Magen und Darmtrakt extrahiert, da in ihnen PCR-hemmende Stoffe vorhanden sein können (WINNEPENNINCKX et al., 1993).
3. Übergießen der Probe mit flüssigem Stickstoff und Zerreiben mit Mörser und Pistill.
4. Zugabe von 3 ml auf 60°C erhitztem CTAB-Puffer und Überführung in 15 ml Falcon-Gefäße
5. Auffüllen der Gefäße mit CTAB-Puffer (60°C) auf 6 ml und Zusatz von 5 mAU/ml Proteinase K
6. Inkubation der Proben für 60 min bei 60°C
7. Zentrifugation für 10 min bei 8000 rpm im Festwinkelrotor (F34-6-38).
8. 0,9 ml Überstand mit 0,9 ml 3M Na-Acetat-Lösung in ein 2 ml Reaktionsgefäß pipettieren und 10 min auf Eis inkubieren.
9. 650 µl der Probe mit 450 µl Roti®-Phenol in ein 2 ml PhaseLockGel-Reaktionsgefäß geben und sanft schütteln.
10. Zentrifugation für 10 min bei 14000 rpm im Festwinkelrotor (FA-45-30-11).
11. Überstand in ein 2 ml Reaktionsgefäß pipettieren und mit doppelter Menge Chloro-form/Isoamylalkohol 24:1 versetzen.
12. Zentrifugation für 10 min bei 14000 rpm im Festwinkelrotor (FA-45-30-11).
13. Obere Phase in ein 1,5 ml Reaktionsgefäß pipettieren und mit doppelter Menge EtOH 98% versetzen.
14. Inkubation der Probe über Nacht bei – 20°C.
15. Zentrifugation der Probe für 15 min bei 4°C und 14000 rpm (FA-45-30-11).
16. Überstand vom Pellet abnehmen, Probe mit 500 µl EtOH 70% versetzen und 10 min bei – 20°C in der Gefrierbox inkubieren.
17. Zentrifugation der Probe für 15 min bei 4°C und 14000 rpm (FA-45-30-11).
18. EtOH vollständig mit Einwegspritze entfernen und Trocknung der DNA für 7 min bei 60°C.
19. Probe mit 100 µl TE-Puffer und 50 U Ribonuklease A für 15 min bei 37°C inkubieren und an-schließend bis zur Verwendung bei – 20°C lagern.
46

MATERIAL UND METHODEN
4.3.5 Sequenzierung der 18s rDNA von Trichobilharzia ocellata Um eine auf der 18s rDNA beruhende spezifische nPCR für T. ocellata entwickeln zu kön-
nen, wurde zunächst die 18s rDNA des Parasiten sequenziert, da zu Beginn der Arbeit noch
keine entsprechende Sequenz veröffentlich war (vgl. Kap 5.1.1).
4.3.5.1 Ausgangsmaterial und Aufreinigung
Als Ausgangsmaterial dienten im Untersuchungstümpel gesammelte L. stagnalis. Mittels
Cercarienschlupftest (s. Kap. 4.3.2) wurden mit T. ocellata infizierte Schnecken identifiziert
und die aus ihnen gewonnene Cercarien zunächst aufgereinigt. Dazu wurde die stark aus-
geprägte positive Phototaxis von T. ocellata (NEUHAUS, 1952) in der Weise ausgenutzt, dass
die per Cercarienschlupftest gewonnenen Cercarien zunächst im Lichtpunkt einer Schwa-
nenhalslampe aufkonzentriert und anschließend in ein 15 ml Falcongefäß pipettiert wurden.
Dieses Gefäß wurde mit sterilisiertem H2O dest. aufgefüllt und anschließend am Boden mit
der Schwanenhalslampe bestrahlt. Aufgrund ihrer positiv phototaktischen Bewegung sam-
melten sich die Cercarien am Gefäßboden. Der Überstand wurde abgenommen, und das
Gefäß wurde mit frischem sterilisiertem H2O dest. aufgefüllt. Diese Reinigung wurde insge-
samt dreimal durchgeführt. Anschließend wurden die Cercarien in ein 1,5 ml Reaktionsgefäß
überführt und dreimal mit EtOH 70% gewaschen. Die Cercarien wurden dabei jeweils durch
Zentrifugation aufkonzentriert.
4.3.5.2 DNA – Sequenzierung Als Ausgangsmaterial für die Sequenzierung dienten PCR-Produkte, die aus DNA-Extrakten
von aufgereinigten Cercarien von T. ocellata (s. Kap. 4.3.5.1) gewonnen wurden. Entspre-
chend der Vorgehensweise von HANELT et al. (1997) wurde die Nukleotidsequenz mittels
„walking in“ ermittelt. Dazu wurde innerhalb der mit Hilfe von 18s rDNA-Universalprimern
amplifizierten Nukleotidsequenzen und mittels des Primer-Suchprogramms Primer 3 (White-
head Institute for Biomedical Research, Cambridge) neue geeignete Primer gesucht und
diese als Startpunkt für eine weitergehende Amplifikation bis zur Überlappung der beiden
DNA-Stränge genutzt.
Die Polymerase-Kettenreaktionen wurden nach den in Kap. 4.3.6.2 aufgeführten PCR-
Protokollen mit den in Kap. 4.3.3.1 angegebenen Primern durchgeführt. Die erste mit der
T. ocellata – DNA durchgeführte Polymerase-Kettenreaktion erfolgte dabei mit den von der
IIT GmbH zur Verfügung gestellten 18s rDNA-Universalprimer 18s-x1 und 18s-y1 (MEDLIN et
47

MATERIAL UND METHODEN
al., 1988). Die Aufreinigung und Sequenzierung der erhaltenen PCR-Produkte wurde vom
hausinternen Sequenzierservice der IIT GmbH durchgeführt.
Zur Erstellung der DNA-Sequenz wurden ausschließlich die Basen genutzt, deren Identifizie-
rung durch das Sequenzierungsprogramm als „verlässlich“ eingestuft wurde. Auf die durch
eine vollständige Sequenzierung beider DNA-Stränge verminderte Fehlerwahrscheinlichkeit
wurde verzichtet, da die Sequenz nicht für phylogenetische Zwecke interpretiert werden soll-
te und eine falsche Sequenz durch eine nicht funktionierende PCR angezeigt worden wäre.
Mittels des Programms CLUSTAL W 1.8 (European Bioinformatics Institute, EBI) wurde ein
Alignment der erhaltenen DNA-Fragmente vorgenommen und die einzelnen Sequenzen zu
einer Gesamtsequenz verknüpft.
4.3.6 Polymerasekettenreaktion (PCR)
4.3.6.1 „Nested“ – PCR zum spezifischen Nachweis der Gattung Tricho-bilharzia
Für die vorliegende Arbeit wurde eine „Nested“-PCR (nPCR) zum spezifischen Nachweis der
Gattung Trichobilharzia in Anlehnung an eine von HANELT et al. (1997) entwickelte nPCR
zum spezifischen Nachweis von Schistosoma mansoni etabliert. Im Vergleich zu anderen
molekularbiologischen Methoden des Nachweises digeneischer Parasiten (HAMBURGER et
al., 1991, SERMSWAN et al., 1991, SHUBKIN et al., 1992, KAPLAN et al., 1995, HERTEL et al.,
2002, HERTEL et al., 2004) vermeidet eine nPCR–basierende Methode die zeitaufwändigen
und arbeitsintensiven Protokolle einer RT-PCR oder Hybridisierung und ist zumeist deutlich
sensitiver als diese Methode oder eine konventionelle PCR. Die Zielregion der vorliegenden
nPCR war die für die SSU 6 der Ribosomen codierende 18s rDNA des Parasiten. Grund für
diese Wahl war einerseits, dass eine Vielzahl von entsprechenden nPCR bereits entwickelt
wurden, von denen an dieser Stelle exemplarisch nur die Arbeit von HANELT et al. (1997)
angeführt werden soll, da sie eine nPCR für den phylogenetisch nahe stehenden Tremato-
den S. mansoni etablieren konnten. Andererseits wurde diese DNA-Sequenz gewählt, da sie
in mehreren Kopien im Genom vorliegt und Universalprimer zur Sequenzierung der Region
bekannt sind. Weiterhin bietet die 18s rDNA hinreichend Variabilität um eine spezifische Er-
kennung der Gattung Trichobilharzia zu ermöglichen und gestattet damit eine Abgrenzung
von verwandten Trematoden, die keine Dermatitis hervorrufen.
6 small subunit
48

MATERIAL UND METHODEN
Bei der nPCR werden zwei Primerpaare benötigt, die in zwei aufeinander folgenden PCR-
Runden zum Einsatz kommen. Dabei dient das Amplikon der vorangehenden PCR als
Template für die zweite PCR. Die Primer dieser zweiten PCR hybridisieren dabei innerhalb
des Abschnittes des ersten Primerpaares. Die Durchführung dieser zweiten („nested“) PCR
führt dabei zu einer Steigerung der Sensitivität und Spezifität. Gegenüber der Methodik einer
konventionellen PCR und Hybridisierung ist die Sensitivität dabei zumeist deutlich erhöht
(HAMBURGER et al., 1991, SERMSWAN et al., 1991, HANELT et al., 1997, HERTEL, et al., 2002,
HERTEL, et al., 2004)(vgl. Kap 5.1.3).
Die Auswahl Trichobilharzia – spezifischer Primer Um geeignete Primer zu finden, wurde zunächst ein Alignment der 18s rDNA von Schistos-
oma mansoni (HANELT et al., 1997) und der durch Sequenzierung gewonnenen 18s rDNA
von T. ocellata (s. Kap. 5.1.1 und Anh., S. 142) mittels des Programms CLUSTAL W 1.8
(EBI) vorgenommen. S. mansoni war bei Durchführung der Arbeiten der phylogenetisch
nächststehende Organismus mit bekannter Sequenz der 18s rDNA. Die Primer wurden in-
nerhalb der Nukleotidsequenz so gewählt, dass Bereiche mit möglichst großem Unterschied
zur Sequenz von S. mansoni gesucht wurden. Gleichzeitig wurden sie in der Art und Weise
gewählt, dass entsprechend der Methode der nPCR ausreichend lange Amplifikate entste-
hen. Weiterhin wurde die Suche der Primer mit dem Programm Primer 3 (Whitehead Institute
for Biomedical Research, Cambridge) dahingehend spezifiziert, Primer zu wählen, die deut-
lich verschiedene Annealing-Temperaturen aufwiesen. Da bei der nPCR stets eine Probe der
ersten PCR-Runde als Template genutzt wird, können in geringer Konzentration Primer die-
ses Ansatzes übertragen werden. Unterschiedliche Annealingtemperaturen zwischen den
beiden Primerpaaren minimieren dabei die Gefahr unerwünschter PCR-Produkte.
4.3.6.2 PCR-Protokolle und Reaktionsansätze
PCR-Protokolle der Sequenzierung der 18s rDNA von Trichobilharzia ocellata Die von der IIT GmbH zur Verfügung gestellten Universalprimer 18s-x1 und 18s-y1 weisen
eine sehr niedrige Schmelztemperatur von 55°C auf. Da die Sequenzierung nach Information
des IIT bei Schmelztemperaturen um die 60°C mit höherer Genauigkeit läuft, wurden die
folgenden Primer entsprechend dieser Temperatur gewählt. Die verwendeten PCR-
Protokolle sind Tab. 4.2 und 4.3 zu entnehmen.
49

MATERIAL UND METHODEN
T
T
ab. 4.2: Schematische Darstellung des PCR-Protokolls für die Primer 18s-x1 und 18s-y1
50
Initi
ale
Den
atur
ieru
ng
Den
atur
ieru
ng
Ann
ealin
g
DN
A-S
ynth
ese
Fina
le
DN
A-S
ynth
ese
Temperatur 94°C 94°C 55°C 72°C 72°C 4°C
Zeit in sec 240 60 30 60 420 HOLD
29 Zyklen; pro Zyklus verlängert sich die DNA-
Synthese um 4 sec
ab. 4.3: Schematische Darstellung des PCR-Protokolls für die Primer to18s-x2,
to18s-x3, to18s-y2 und to18s-y3
Die Reaktionsansätze (50 µl) der zur Sequenzierung genutzten PCR-Produkte setzten sich wie folgt zusammen: Reagenzien Eingesetztes Volumen
DNA – Isolat 2 µl
10x Taq Reakt.-puffer enth. 15 mM Mg++ 5 µl
dNTP-Mix 10 mM 6 µl
Primer 1 10 pmol/µl 2 µl
Primer 2 10 pmol/µl 2 µl
MgCl2-Lsg. 25 mM 2 µl
Taq-Polymerase 5 U / µl 0,5 µl
H2O 30,5 µl
Initi
ale
Den
atur
ieru
ng
DN
A-S
ynth
ese
DN
A-S
ynth
ese
Den
atur
ieru
ng
Fina
le
Ann
ealin
g
Temperatur 94°C 94°C 60°C 72°C 72°C 4°C
Zeit in sec 240 60 30 60 420 HOLD
29 Zyklen; pro Zyklus verlängert sich die DNA-
Synthese um 4 sec
MATERIAL UND METHODEN
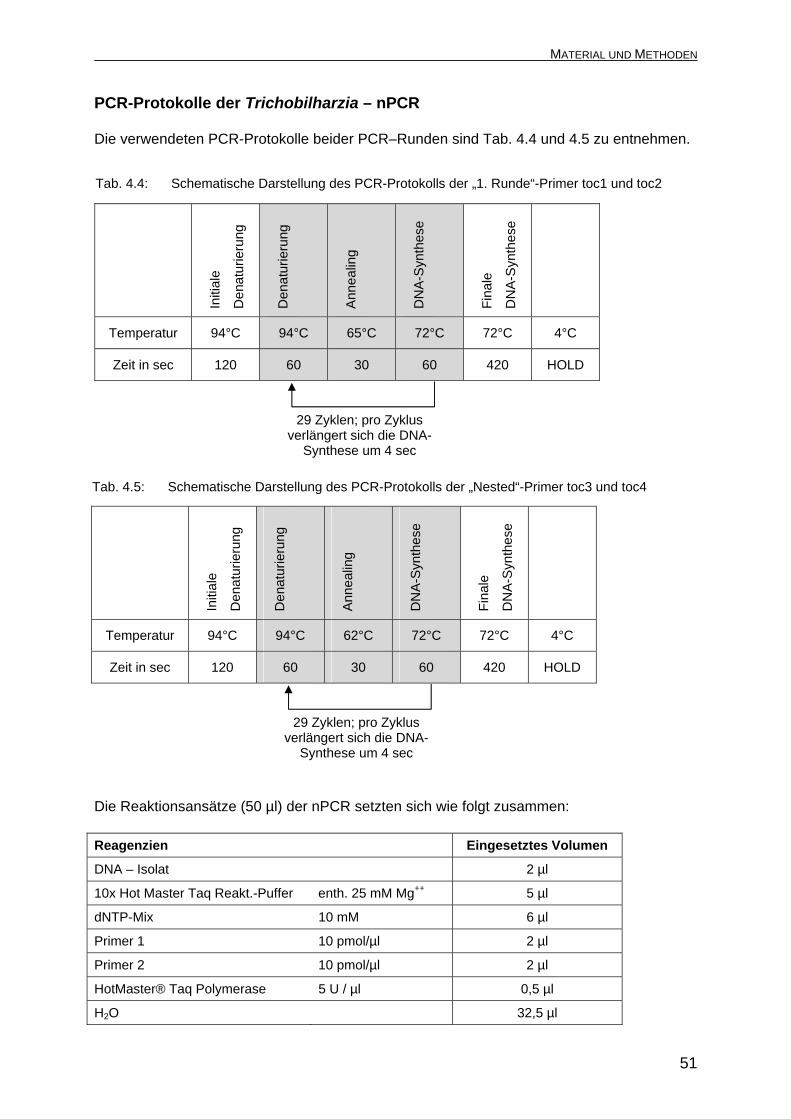
PCR-Protokolle der Trichobilharzia – nPCR Die verwendeten PCR-Protokolle beider PCR–Runden sind Tab. 4.4 und 4.5 zu entnehmen.
Tab. 4.4: Schematische Darstellung des PCR-Protokolls der „1. Runde“-Primer toc1 und toc2 Die Reaktionsansätze (50 µl) der nPCR setzten sich wie folgt zusammen: Reagenzien Eingesetztes Volumen
DNA – Isolat 2 µl
10x Hot Master Taq Reakt.-Puffer enth. 25 mM Mg++ 5 µl
dNTP-Mix 10 mM 6 µl
Primer 1 10 pmol/µl 2 µl
Primer 2 10 pmol/µl 2 µl
HotMaster® Taq Polymerase 5 U / µl 0,5 µl
H2O 32,5 µl
Initi
ale
Den
atur
ieru
ng
DN
A-S
ynth
ese
DN
A-S
ynth
ese
Den
atur
ieru
ng
Fina
le
Ann
ealin
g
Temperatur 94°C 94°C 65°C 72°C 72°C 4°C
Zeit in sec 120 60 30 60 420 HOLD
29 Zyklen; pro Zyklus verlängert sich die DNA-
Synthese um 4 sec
Tab. 4.5: Schematische Darstellung des PCR-Protokolls der „Nested“-Primer toc3 und toc4
Initi
ale
Den
atur
ieru
ng
DN
A-S
ynth
ese
DN
A-S
ynth
ese
Den
atur
ieru
ng
Fina
le
Ann
ealin
g
Temperatur 94°C 94°C 62°C 72°C 72°C 4°C
Zeit in sec 120 60 30 60 420 HOLD
29 Zyklen; pro Zyklus verlängert sich die DNA-
Synthese um 4 sec
51

MATERIAL UND METHODEN
4.3.6.3 Spezifitätsanalyse der Trichobilharzia – nPCR Die Spezifität der entwickelten nPCR wurde getestet, indem Template DNA verschiedener
Trematoden, L. stagnalis – Gewebe und Plankton (bestehend aus versch. Arten von Grünal-
gen, Kieselalgen, Cladoceren, Insektenlarven) eingesetzt wurde. Dabei wurden alle im un-
tersuchten Gewässer angetroffenen (auch aus anderen Wirtschnecken stammende) Trema-
todenlarven getestet. Zusätzlich zu diesen aber noch die phylogenetisch nahe stehenden
Arten Trichobilharzia franki, Schistosoma mansoni und Bilharziella polonica (FARLEY, 1971,
SNYDER et al., 2001, LOCKYER et al., 2003). Die DNA aller in der Spezifitätsanalyse geteste-
ten Proben wurde, wie in Kap. 4.3.4.1 beschrieben, gewonnen. Die Planktonprobe wurde
aus einem Weiher (J5) entnommen, in dem kein Befall der Schnecken mit Trichobilharzia-
Arten festgestellt werden konnte und der sich ca. 1 km nördlich des Untersuchungsgebiets
an der Jölle befindet.
Da für diesen Test nur relativ kleine Probenmengen nötig waren, ist auch die DNA der
Schneckengewebe- und der Planktonprobe nach diesem Protokoll isoliert worden. Tab. 4.5
ist eine Auflistung der getesteten DNA-Proben und ihre Herkunft zu entnehmen.
Tab. 4.6: Herkunft der DNA-Isolate des Spezifitätstests
DNA-Probe Wirtstier Herkunft
Trichobilharzia ocellata Cercarien L. stagnalis Weiher J1
Trichobilharzia franki Cercarien R. auricularia Weiher Mühlbachtal
Bilharziella polonica Adulti A. platyrhynchos Wildfang, Extertal
Schistosoma mansoni Cercarien B. glabrata Laborstamm
Diplostomum sp. Cercarien L. stagnalis Weiher J1
Echinostoma revolutum Cercarien L. stagnalis Weiher J1
Hypoderaeum conoidum Cercarien L. stagnalis Weiher J1
Pseudoechinoparyphium echinatum Cercarien L. stagnalis Weiher J1
Opistoglyphe ranae Cercarien L. stagnalis Weiher J1
Plagiorchis sp. Cercarien L. stagnalis Weiher J1
Astiotrema sp. Cercarien P. corneus Weiher J1
Opisthodiscus nigrivasis Cercarien A. vortex Weiher J1
Haematoloechus sp. Cercarien A. vortex Weiher J1
Lymnaea stagnalis Weiher J1
Plankton Weiher J5
52

MATERIAL UND METHODEN
4.3.6.4 Bestimmung der Nachweisgrenze der Trichobilharzia – nPCR Zur Ermittlung der Nachweisgrenze der entwickelten nPCR wurden verschiedene Tests
durchgeführt.
Zunächst wurden dabei die Nachweisgrenzen der verwendeten Primerpaare toc1/toc2 und
toc3/toc4 in der Weise ermittelt, dass Polymerase-Kettenreaktionen mit fallenden Konzentra-
tionen von Template-DNA in Anwesenheit konstant hoher Menge (100 ng) von Fremd-DNA
(L. stagnalis) durchgeführt wurden. Die Ausgangskonzentration der verwendeten DNA wurde
dabei fotometrisch bestimmt. Die eingesetzten DNA-Konzentrationen wurden durch eine
Verdünnungsreihe hergestellt und betrugen für T. ocellata-DNA 10 ng bis 100 ag in Zehntel
Abstufung.
Ein weiterer Sensitivitätstest betraf die Methodik der Planktonanalyse. Dazu wurden Plank-
tonproben aus einem nicht mit T. ocellata infizierten Gewässer (J5) mit einzelnen Cercarien
von T. ocellata bestückt und anschließend die DNA aus den Proben nach dem in Kap.
4.3.4.2 angegebenen Verfahren isoliert. Jeweils 0,75 g Plankton wurde dazu in 15 ml Fal-
congefäße gegeben, mit 50, 10, 5 oder einer Cercarie von T. ocellata versetzt und auf 15 ml
mit H2O aufgefüllt. Nachfolgend wurde die nPCR nach den in Kap. 4.3.6.2 aufgeführten Pro-
tokollen durchgeführt.
Entsprechend wurde auch die Methodik der DNA-Extraktion aus Gewebe von L. stagnalis
und der Nachweis T. ocellata-infizierter Schnecken getestet. Da sich die in der Schnecke
vorkommende Entwicklungsstufe des Trematoden, die Sporocyste, nicht rein isolieren lässt
und Miracidien (das für die Schnecke infektiöse Stadium) des Parasiten nicht zur Verfügung
standen, wurden den Proben auch hier zum Testen der Sensitivität Cercarien zugesetzt. Da-
bei ist die Cercarie nach NEUHAUS (1952) ungefähr 4,5-fach größer als die kleinste in der
Schnecke vorkommende Form des Parasiten, das Miracidium7. Zum DNA-Gehalt dieser bei-
den Parasitenstadien gibt es keine Erkenntnisse. Für den Sensitivitätstest wurden parasiten-
freie im Labor gehaltene adulte Nachzuchten von L. stagnalis mit einer Gehäusehöhe von
3,8 cm benutzt8. Dies entspricht nach Entfernung des Schneckengehäuses und Extraktion
des Magendarmtraktes einem Frischgewicht von ca. 2,0 g. Die Proben wurden nach Entfer-
nen des Schneckengehäuses mit 50, 25, 10, 5, 3 oder einer Cercarie versetzt und, wie in
Kap. 4.3.4.3 beschrieben, verarbeitet. Die isolierte DNA wurde anschließend per nPCR, wie
oben beschrieben, getestet. 7 Das Miracidium wandelt sich unmittelbar nach Infektion der Schnecke zur Muttersporocyste, aus der die folgende Sporocystengeneration und schließlich Cercarien hervorgehen. 8 Dies entspricht der maximalen bei uns in Laborhaltung für L. stagnalis erreichten Gehäusehöhe. Freilandschnecken sind teilweise deutlich größer.
53

MATERIAL UND METHODEN
4.3.6.5 Prävention vor falsch-positiven Ergebnissen Da zum Nachweis von T. ocellata eine hoch sensitive n PCR genutzt wurde, musste beson-
deres Augenmerk auf die Vermeidung von Kontaminationen gelegt werden. Dabei wurde
zunächst auf die strikte räumliche Trennung der PCR-Vorgänge Wert gelegt. Die Aufberei-
tung der Proben und die DNA-Extraktion sowie die Amplifikation im Thermocycler fand dabei
an separaten Tischen statt, die PCR-Ansätze wurden unter der Impfbank angesetzt. Die Ge-
lelektrophorese sowie die Färbung der Gele und ihre Dokumentation wurden in einem sepa-
raten Raum durchgeführt. Insbesondere die DNA-Extraktion aus zum Teil mit T. ocellata infi-
ziertem Schneckengewebe erzwang zusätzliche Verfahren zur Reinigung und Dekontamina-
tion der bei der Präparation eingesetzten Materialen wie z.B. Mörser, Pistill, Sezierbesteck.
Die eingesetzten Geräte wurden dazu zunächst gründlich bei 70°C in der Geschirrspülma-
schine gewaschen, anschließend im Autoklaven hitzesterilisiert und nachfolgend unter einem
Sterisol®-Intensivstrahler Typ NN 15/44 der Fa. Heraeus Nobelight mit UV-C-Licht bei einer
Leistung von 1900 µW/cm2 für 15 min dekontaminiert. Nachfolgende Kontrollen dieser so
behandelten Geräte mittels nPCR konnten in keinem Fall Rest-DNA von T. ocellata nach-
weisen.
4.3.6.6 Nachweis und Analyse von PCR-Produkten Agarosegelelektrophorese Zum Nachweis der PCR-Produkte wurde ein 1,7%iges Agarosegel verwendet. Jeweils 8 µl
PCR-Probe wurden mit 2 µl des Ladepuffers gemischt und in die Gel-taschen pipettiert. Der
gebrauchsfertige Laufpuffer wurde aus 200ml TAE Puffer 50x (Merck AG) und 10 l H2O auf
Vorrat angesetzt. Zum Abschätzen der DNA-Fragmentgrößen wurde ein DNA-
Längenstandard (GeneRulerTM 100 bp DNA Ladder Plus, MBI Fermentas) mitgeführt. Die
Auftrennung der DNA-Fragmente erfolgte bei einer Spannung von 75V in ca. 105 min. Dabei
erfolgte das Abschalten der Spannungsquelle, wenn die Trennung der beiden Farbstoffe des
Ladepuffers ausreichend war. Das Gel wurde für 3 min in Ethidiumbromidlösung gefärbt und
anschließend ca. 45 min im TAE-Pufferbad entfärbt. Die DNA wurde mit Hilfe eines Transil-
luminators (Herolab) bei 312 nm sichtbar gemacht und das Gel ausgewertet. Die Längen der
PCR-Produkte wurden durch Vergleich mit dem DNA-Längenstandard ermittelt.
Die verwendeten Puffer, Chemikalien und Geräte sind in Kap. 4.2.4 bis 4.2.6 detailliert auf-
gelistet.
54
MATERIAL UND METHODEN
Analytische Restriktionsspaltung von „2. Runde“-PCR-Produkten Für die aus der nPCR erhaltenen PCR-Produkte, die einer Länge von 816 bp entsprachen,
wurde mittels einer Restriktionsanalyse eine Verifikation durchgeführt. Bei der Restriktions-
analyse werden die PCR-Produkte mit Hilfe sequenzspezifischer Endonukleasen gespalten.
Die Länge der entstehenden Spaltungsprodukte ist aufgrund der bekannten Nukleotidse-
quenz des 816 bp – Amplifikats vorhersagbar. Stimmen die Längen der entstanden Frag-
mente mit der Voraussage überein, so gilt die Identität des Fragments als verifiziert.
Eine geeignete Restriktionsendonuklease wurde mit Hilfe des im Internet zugänglichen Pro-
gramms CUTTER (www.justbio.com) ermittelt. Die Restriktionsanalyse wurde mit dem En-
zym BamH I (MBI Fermentas) durchgeführt, welches das 816 bp große DNA-Amplifikat in
zwei Fragmente mit den Längen 186 bp und 630 bp spaltet. Die Durchführung erfolgte unter
Verwendung des vom Hersteller mitgelieferten Restriktionspuffers.
Reaktionsansatz:
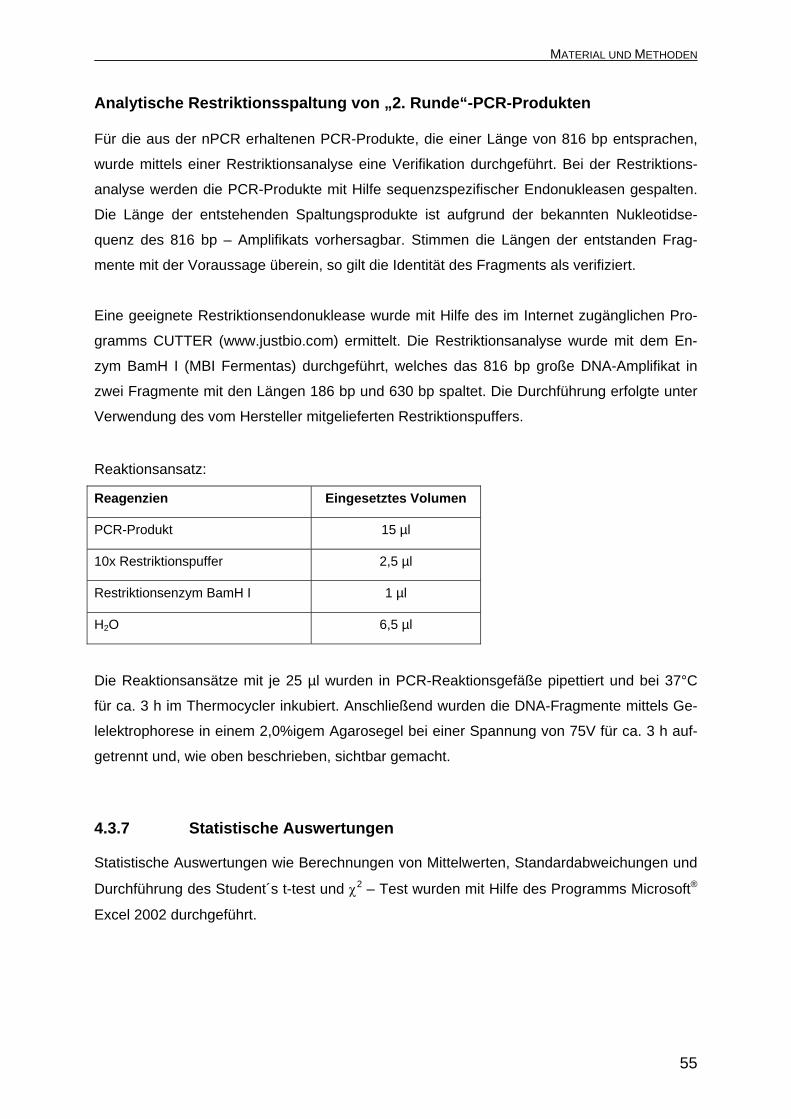
Reagenzien Eingesetztes Volumen
PCR-Produkt 15 µl
10x Restriktionspuffer 2,5 µl
Restriktionsenzym BamH I 1 µl
H2O 6,5 µl
Die Reaktionsansätze mit je 25 µl wurden in PCR-Reaktionsgefäße pipettiert und bei 37°C
für ca. 3 h im Thermocycler inkubiert. Anschließend wurden die DNA-Fragmente mittels Ge-
lelektrophorese in einem 2,0%igem Agarosegel bei einer Spannung von 75V für ca. 3 h auf-
getrennt und, wie oben beschrieben, sichtbar gemacht.
4.3.7 Statistische Auswertungen Statistische Auswertungen wie Berechnungen von Mittelwerten, Standardabweichungen und
Durchführung des Student´s t-test und χ2 – Test wurden mit Hilfe des Programms Microsoft®
Excel 2002 durchgeführt.
55





























