Embed Size (px)
Citation preview
Molekulare Genetik der Eukaryoten
Vorlesung Nr. 441: Biochemie I für Mediziner Josef Jiricny
4. Auflage, August 2002

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten
Aug-02 J. Jiricny
Inhaltsverzeichnis
1. Organisation des Eukaryotischen-Genoms.......................................................................................1
1.1 DNA-Methylierung......................................................................................................................1
2. Der Zellzyklus .....................................................................................................................................2
3. DNA-Synthese: Replikation...............................................................................................................3
3.1 Telomerase..................................................................................................................................4
4. RNA-Synthese: Transkription ...........................................................................................................4
4.1 Spleissen......................................................................................................................................5 4.1.1 Hemmstoffe von Replikation und/oder Transkription .................................................................................................6
5. Topoisomerasen ..................................................................................................................................6
5.1 Hemmstoffe der Topoisomerasen...............................................................................................7
6. Synthese (Translation) von Proteinen...............................................................................................7
6.1 Hemmstoffe der Proteinsynthese ...............................................................................................9
7. Genumordnungen.............................................................................................................................10
7.1 Retroviren.................................................................................................................................11 7.1.1 Inhibitoren der DNA-Synthese des HIV Virus............................................................................................................12
8. Mutagenese und DNA Reparatur ....................................................................................................13
8.1 Schädenumkehrung ...................................................................................................................13
8.2 Basenexcisionsreparatur (BER) ...............................................................................................14
8.3 Nukleotidexcisionsreparatur.....................................................................................................14
8.4 Fehlpaarungsreparatur ..............................................................................................................15
8.5 Strangbruchsreparatur ........................................................................................................................15

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 1
Aug-02 J. Jiricny
1. Organisation des Eukaryotischen-Genoms
Die DNA bildet mit den basischen Histonen geordnete Strukturen, die Nukleosomen. Ein Nukleosom Core besteht aus etwa 146 Basenpaaren, die um je zwei H2A, H2B, H3 und H4 Histonmoleküle so herum gewunden sind, dass 13/4 Windungen einer linksgängigen Superhelix entstehen. Einzelne Nukleosomen sind über kurze DNA-Abschnitte perlschnurartig miteinander verbunden. Die einzelnen Chromatinfäden liegen in den 46 Chromosomen in stark kondensierter Form vor: Totale Länge der DNA = 2 m, totale Länge aller 46 Chromosomen (in Metaphase) = 200 µm. Chromatinkondensierung ist abhängig auch vom DNA- Methylierungszustand und der Histonmodifikation (Acetylierung, Methylierung, ADP-Ribosylierung, Phosphorylierung). Methylierte DNA (siehe unten) interagiert stärker mit Histon H1. Nukleosome die H1 enthalten besitzen ungefähr 166 Basenpaare DNA. H1 wird unmittelbar vor der Mitose phosphoryliert und nach der Mitose wieder dephosphoryliert, was vermuten lässt, dass diese kovalente Modifikation seine Fähigkeit steuert, die DNA zu verdichten.
Acetylierung an Lysinresten von Histonen (z. B. H4) kann ihre Affinität für DNA beeinflussen. Das Zusammenspiel zwischen Histon-Acetylasen und –Deacetylasen, DNA Methylasen und Histon-Kinasen ist insgesamt für die Dichte der Chromatinverpackung verantwortlich. Die am stärksten komprimierte DNA findet man in Sper-mienköpfen, wo die Histone durch Protamine ersetzt sind. Die Protamine sind argininreiche Proteine, welche durch Bindung an die DNA hochgradig α-helikal werden.
Weniger als 1% der Gesamt -DNA ist in den Mitochondrien lokalisiert als kleine zirkuläre DNA. Mitochondriale Proteine werden teils durch nukleäre, teils durch mitochondriale DNA kodiert. Menschliche DNA enthält pro diploide Zelle ca. 6.6 x 109 Basenpaare. Dies würde für ca. 3 x 106 Gene reichen. Tat-sächlich rechnet man aber mit nur ca. 35’000 Genen. Dies hätte eine viel kürzere DNA zur Folge, aber stark repetitive DNA und Introns (nicht kodierende DNA-Abschnitte zwischen den Exons) sind u.a. für die jetzige Länge der menschlichen DNA verantwortlich.
1.1 DNA-Methylierung
Das C-5 des Cytosins in CG–Sequenzen der Säuger-DNA (in Pflanzen auch in CNG) wird durch die DNA-Methyltransferase modifiziert. Die Methylgruppe wird von S-Adenosylmethionin übertragen. Ungefähr 90% der CG–
Sequenzen in menschlicher DNA sind methyliert. Im allgemeinen sind aktive Gene weniger stark methyliert als inaktive, vor allem in Promotoren. Methylierte DNA wird von Histon H1 und anderen Proteinen (MBD, methylated DNA binding proteins) stärker gebunden. Dadurch wird sie eher in Heterochromatin verpackt.
Viele "Haushaltgene" haben CG-reiche Promotoren, die so genannten "CG-Inseln", die in der Regel immer unmethyliert bleiben.
N
N
NH2
HO
CH3
5-Methylcytosin

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 2
Aug-02 J. Jiricny
2. Der Zellzyklus
Voraussetzung für jede Zellteilung (Mitose) ist die Replikation der DNA, um identische Kopien für jede Tochterzelle bereitzustellen. Die Replikation erfolgt in der synthetischen oder S-Phase während der Interphase (zwischen zwei Mitosen). Ein ganzer Zellzyklus (G1 - S - G2 - M) dauert bei kontinuierlich sich teilenden Zellen (z.B. Darmmucosa, Hautepithel, Erythrozyten-Vorstufe) ca. 16 - 20 Std. Andere Zellarten teilen sich nur auf ein Signal hin (z.B. Lymphozyten), selten (Leber- und Nierenzellen) oder sozusagen nie (Nerven- und Muskelzellen). In diesen Fällen ist die G1-Phase von ca. 5 Std. auf Tage oder Wochen verlängert. Eine derart lange G1-Phase wird auch als G0 bezeichnet. S-, G2- und M-Phase dauern ca. 7, 3 und 2 Std. Der Eintritt in die Mitose wird von einer Protein-Kinase (cdc2-Kinase) kontrolliert. Sie wird durch Cyclin B aktiviert, ein im Zellzyklus synthetisiertes und abgebautes Protein.
Die katalytische Aktivität der cdc2-Kinase hängt auch von ihrem Phosphorylierungszustand ab, der durch Signale von Wakstumsfaktoren kontrolliert wird.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 3
Aug-02 J. Jiricny
3. DNA-Synthese: Replikation
In höheren Eukaryoten sind mehr als zehn DNA-Polymerasen charakterisiert worden. Polymerase-α, -δ und -ε werden bei der DNA-Replikation und -Reparatur gebraucht. Polymerase-β ist für die Basenexcisions Reparatur verantwortlich, während Polymerase-γ für die Replikation des mitochondriellen Genoms sorgt. Die anderen sind in „By-pass“ von DNA-Schäden involviert.
Die eukaryotische DNA wird semikonservativ und semidiskontinuierlich von mehreren tausend Startpunkten aus repliziert. Der Leitstrang wird von der DNA-Polymerase-δ synthetisiert, deren Prozessivität ist durch die Komplexbildung mit PCNA (Homolog der β-Untereinheit der bakteriellen Polymerase III) erhöht. Der Folgestrang wird diskontinuierlich von DNA-Polymerase-α synthetisiert.
Eukaryotisches Enzym Bakterielles Homolog Polymerase-δ 3‘-5‘ Exonuclease
Pol III (α-Untereinheit) Pol III (ε-Untereinheit)
Polymerase-α Primase (2) 3‘5‘ Exonuclease
Pol III (α-Untereinheit) DNA-Primase Pol III (ε-Untereinheit)
Polymerase-ε Pol I Replikations-Faktor C (RFC) (5) Pol III (γδ-Untereinheiten) Replikations-Protein A (RPA) (3) Ssb DNA Helikase DNA B-Protein PCNA (Homotrimer) Pol III (β-Untereinheit) Topoisomerase I, II DNA-Gyrase DNA-Ligase I DNA-Ligase
Die Länge der Okazaki-Fragmente bei Eukaryoten liegt bei 100-200 Nukleotiden, was darauf hindeutet, dass jeweils ein Nukleosom pro Okazaki-Fragment entfernt wird. Alte Histone verbleiben bei der DNA-Doppelhelix, die den Leitstrang enthält, während sich an der Doppelhelix mit dem Folgestrang neue Histone zusammenlagern.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 4
Aug-02 J. Jiricny
3.1 Telomerase
Telomere, die Enden der Chromosomen, werden von einer Reversen Transkriptase repliziert, die eine RNA-Matrize enthält. Die Telomerase synthetisiert Tandemwieder-holungen von G-reichen Hexanukleotiden (AGGGTT in menschlicher DNA), um die Synthese des Folgestrangs zu vollenden. Weil somatische Zellen keine, oder nur wenig Telomeraseaktivität haben, wird die Telomerenlänge mit jeder Zellteilung kürzer. Daraus ist die Hypothese entstanden, dass die Telomeraseaktivität die Ge schwin-digkeit des Alterungsprozesses festlegt. Krebszellen zeigen eine erhöhte Telomeraseaktivität und haben die Tendenz „immortal“ zu werden. Die Telomerase könnte deswegen ein Ziel der Tumortherapie werden.
4. RNA-Synthese: Transkription
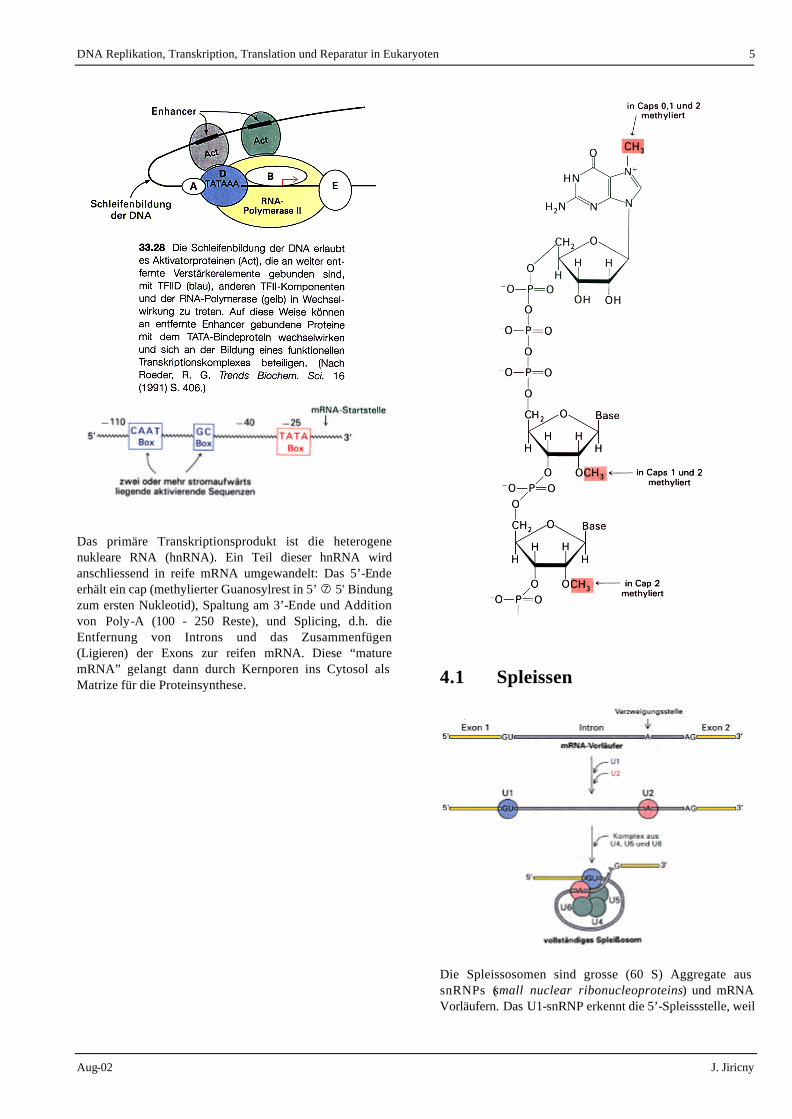
In Eukaryoten sind drei RNA-Polymerasen bekannt. Die RNA-Polymerase I sorgt für die Synthese der ribosomalen RNA, RNA Polymerase II für die mRNA Synthese und Polymerase III ist für tRNA-Synthese zuständig. Anders als bei der Replikation der DNA werden bei der Transkription immer nur Teile der DNA transkribiert. Für die Initiation der Transkription werden besondere Er-kennungssequenzen und auf diesen Erkennungssequenzen bindende Proteine (Transkriptions- faktoren) gebraucht: der Promoter (Ort der initialen Bindung der RNA Polymerase) und ev. regulatorische Regionen (Enhancer). Ein Promoter besteht meist aus einer TATA-Box (ca. 30 Basen upstream von der Initiationsstelle) und aus einer GC-Region, oft mit einer CAAT-Box, 50 - 100 Basen upstream. Enhancer können up- oder downstream liegen.
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 5
Aug-02 J. Jiricny
Das primäre Transkriptionsprodukt ist die heterogene nukleare RNA (hnRNA). Ein Teil dieser hnRNA wird anschliessend in reife mRNA umgewandelt: Das 5’-Ende erhält ein cap (methylierter Guanosylrest in 5’ � 5' Bindung zum ersten Nukleotid), Spaltung am 3’-Ende und Addition von Poly-A (100 - 250 Reste), und Splicing, d.h. die Entfernung von Introns und das Zusammenfügen (Ligieren) der Exons zur reifen mRNA. Diese “mature mRNA” gelangt dann durch Kernporen ins Cytosol als Matrize für die Proteinsynthese.
4.1 Spleissen
Die Spleissosomen sind grosse (60 S) Aggregate aus snRNPs (small nuclear ribonucleoproteins) und mRNA Vorläufern. Das U1-snRNP erkennt die 5’-Spleissstelle, weil

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 6
Aug-02 J. Jiricny
die U1-snRNA eine Sequenz erkennt, die komplementär zu diese Spleissstelle ist. Nun bindet das U2-snRNP die Verzweigungsstelle in dem Intron. Die Assoziation von U1 und U2 bringt die 5’- und 3’-Enden des Introns zusammen, was U1 in die Lage versetzt, auch mit der 3’-Spleissstelle eine Basenpaarung einzugehen. Zu diesem Komplex stösst jetzt eine bereits gebildete Anordnung aus U4, 5 und 6. Die U2 und U6-snRNPs bilden höchstwahrscheinlich das katalytische Zentrum des Spleissosoms.
4.1.1 Hemmstoffe von Replikation und/oder Transkription
Hemmstoffe von Replikation und/oder Transkription bei Eukaryoten können toxisch wirken (Aflatoxin, α-Amanitin) oder als Cystostatika zur Krebsbekämpfung eingesetzt werden (Cytosin-Arabinosid, Actinomycin); als Antibiotika können Stoffe verwendet werden, welche selektiv nur die Replikation oder Transkription von Prokaryoten hemmen (Rifampicin). Die Wirksamkeit Acyclovirs bei Herpes simplex Infektionen ist dadurch gegeben, dass diese Substanz erst durch die virale Thymidin-Kinase phosphoryliert wird. Acyclovirtriphosphat ist nur Substrat für die virale, nicht aber für die menschliche DNA Polymerase. Deswegen wird es ausschliesslich in die virale DNA eingebaut.
ACYCLOVIR
5. Topoisomerasen
Die meisten natürlich vorkommenden DNA-Moleküle sind negativ superspiralisiert. Die negative Superspiralisierung bereitet die DNA auf Prozesse vor, die eine Trennung der beiden Stränge erfordern. Dazu gehören die Replikation, die Rekombination und die Transkription. Die Verwin-dungszahl der DNA ist aber ein dynamisches Phänomen. Prozesse, die Proteinbewegungen auf der DNA-Matrize erfordern, ändern kontinuierlich den Superspirali-sierungstand vor und hinter sich.
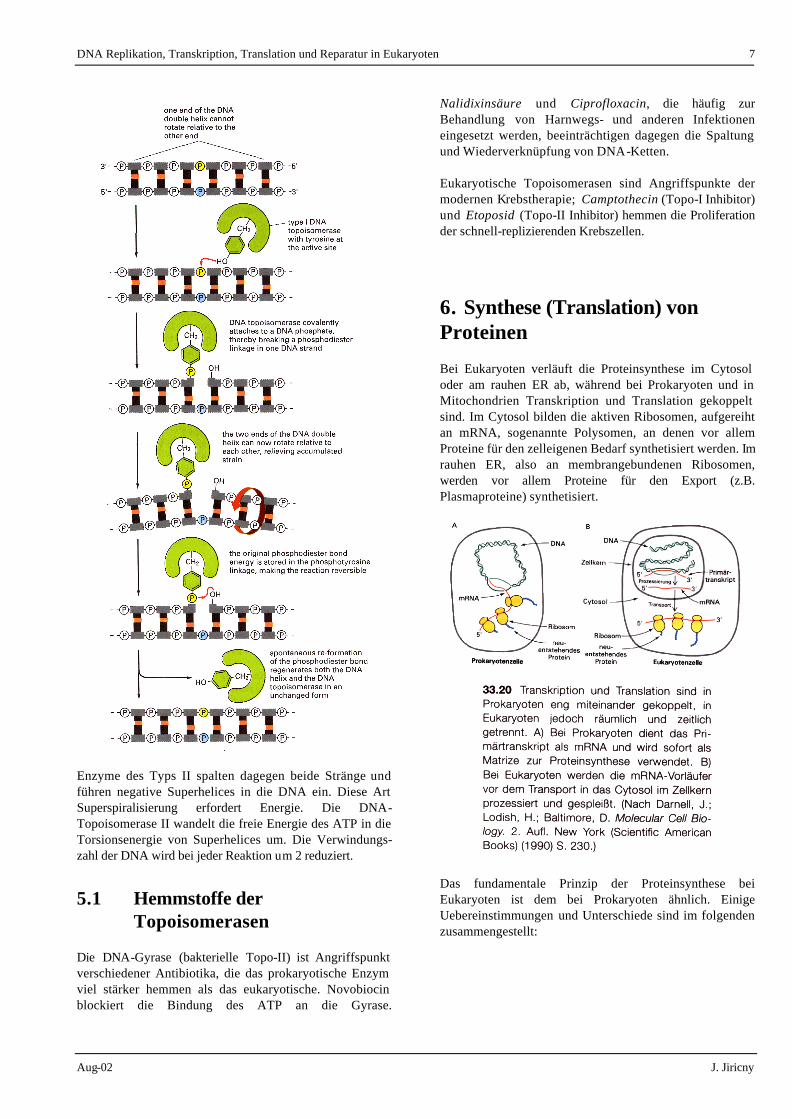
Topoisomerasen verändern die Verwindungszahl der DNA, indem sie einen dreistufigen Prozess katalysieren: 1) Spaltung eines oder beider Stränge der DNA 2) Durchtritt eines DNA-Abschnitts durch die
entstandene Lücke 3) die Wiederverknüpfung der DNA-Bruchstelle. Die Typ-I- Topoisomerasen spalten nur einen DNA-Strang. Die Reaktion der Entspannung einer negativ-superspiralisierten DNA ist thermodynamisch gesehen mit Energiegewinn verbunden. Die Verwindungszahl erhöht sich bei jedem Katalysendurchgang um 1 (Relaxation).
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 7
Aug-02 J. Jiricny
Enzyme des Typs II spalten dagegen beide Stränge und führen negative Superhelices in die DNA ein. Diese Art Superspiralisierung erfordert Energie. Die DNA-Topoisomerase II wandelt die freie Energie des ATP in die Torsionsenergie von Superhelices um. Die Verwindungs-zahl der DNA wird bei jeder Reaktion um 2 reduziert.
5.1 Hemmstoffe der Topoisomerasen
Die DNA-Gyrase (bakterielle Topo-II) ist Angriffspunkt verschiedener Antibiotika, die das prokaryotische Enzym viel stärker hemmen als das eukaryotische. Novobiocin blockiert die Bindung des ATP an die Gyrase.
Nalidixinsäure und Ciprofloxacin, die häufig zur Behandlung von Harnwegs- und anderen Infektionen eingesetzt werden, beeinträchtigen dagegen die Spaltung und Wiederverknüpfung von DNA-Ketten. Eukaryotische Topoisomerasen sind Angriffspunkte der modernen Krebstherapie; Camptothecin (Topo-I Inhibitor) und Etoposid (Topo-II Inhibitor) hemmen die Proliferation der schnell-replizierenden Krebszellen.
6. Synthese (Translation) von Proteinen
Bei Eukaryoten verläuft die Proteinsynthese im Cytosol oder am rauhen ER ab, während bei Prokaryoten und in Mitochondrien Transkription und Translation gekoppelt sind. Im Cytosol bilden die aktiven Ribosomen, aufgereiht an mRNA, sogenannte Polysomen, an denen vor allem Proteine für den zelleigenen Bedarf synthetisiert werden. Im rauhen ER, also an membrangebundenen Ribosomen, werden vor allem Proteine für den Export (z.B. Plasmaproteine) synthetisiert.
Das fundamentale Prinzip der Proteinsynthese bei Eukaryoten ist dem bei Prokaryoten ähnlich. Einige Uebereinstimmungen und Unterschiede sind im folgenden zusammengestellt:
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 8
Aug-02 J. Jiricny
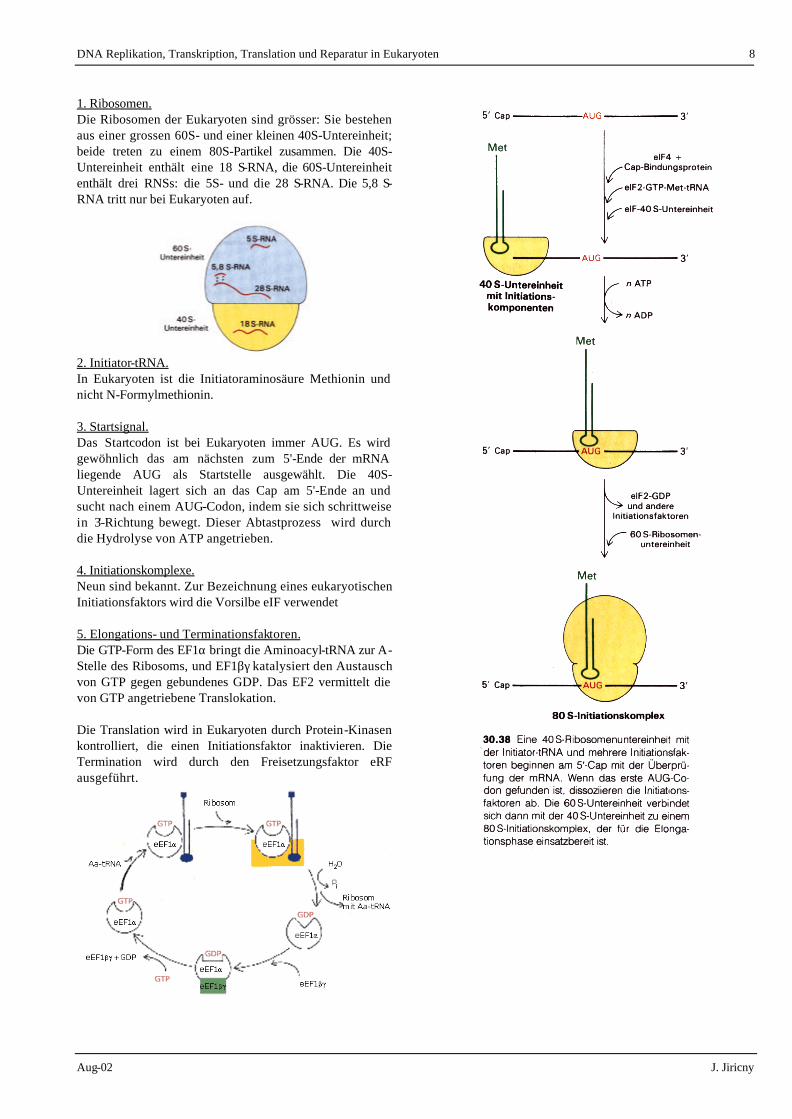
1. Ribosomen. Die Ribosomen der Eukaryoten sind grösser: Sie bestehen aus einer grossen 60S- und einer kleinen 40S-Untereinheit; beide treten zu einem 80S-Partikel zusammen. Die 40S-Untereinheit enthält eine 18 S-RNA, die 60S-Untereinheit enthält drei RNSs: die 5S- und die 28 S-RNA. Die 5,8 S-RNA tritt nur bei Eukaryoten auf.
2. Initiator-tRNA. In Eukaryoten ist die Initiatoraminosäure Methionin und nicht N-Formylmethionin. 3. Startsignal. Das Startcodon ist bei Eukaryoten immer AUG. Es wird gewöhnlich das am nächsten zum 5'-Ende der mRNA liegende AUG als Startstelle ausgewählt. Die 40S-Untereinheit lagert sich an das Cap am 5'-Ende an und sucht nach einem AUG-Codon, indem sie sich schrittweise in 3'-Richtung bewegt. Dieser Abtastprozess wird durch die Hydrolyse von ATP angetrieben. 4. Initiationskomplexe. Neun sind bekannt. Zur Bezeichnung eines eukaryotischen Initiationsfaktors wird die Vorsilbe eIF verwendet 5. Elongations- und Terminationsfaktoren. Die GTP-Form des EF1α bringt die Aminoacyl-tRNA zur A-Stelle des Ribosoms, und EF1βγ katalysiert den Austausch von GTP gegen gebundenes GDP. Das EF2 vermittelt die von GTP angetriebene Translokation. Die Translation wird in Eukaryoten durch Protein-Kinasen kontrolliert, die einen Initiationsfaktor inaktivieren. Die Termination wird durch den Freisetzungsfaktor eRF ausgeführt.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 9
Aug-02 J. Jiricny
6.1 Hemmstoffe der Proteinsynthese
Die Unterschiede zwischen den Proteinsynthesemaschinen der Pro- und Eukaryoten ermöglichen den Einsatz von Inhibitoren der Proteinsynthese von Prokaryoten als Antibiotika:
Antibiotikum Wirkung Streptomycin und andere Aminoglykoside
hemmen die Initiation und verursachen Fehlablesungen der mRNA (bei Prokaryoten)
Tetracyclin lagert sich an die 30S-Untereinheit an und hemmt die Bindung der Aminoacyl-tRNAs (bei Prokaryoten)
Chloramphenicol hemmt die Peptidyltransferaseaktivität der 50S-Ribosomenuntereinheit (bei Prokaryoten)
Cycloheximid hemmt die Peptidyltransferaseaktivität der 60S- Ribosomenuntereinheit (bei Eukaryoten)
Erythromycin lagert sich an die 50S-Untereinheit an und hemmt die Translokation (bei Prokaryoten)
Puromycin verursacht vorzeitigen Kettenabbruch, weil es als Analogon der Aminoacyl-tRNA wirkt (bei Prokaryoten und Eukaryoten)
Inhibitoren der Eukaryoten-Proteinsynthese sind gefährliche Toxine. Diphtherie-Toxin z.B. blockiert die Pro-teinsynthese durch ADP-Ribosylierung von EF-2, was die Translokation verunmöglicht.
Röntgenstruktur der 30S-Untereinheit der E. coli Ribosoms
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 10
Aug-02 J. Jiricny
7. Genumordnungen
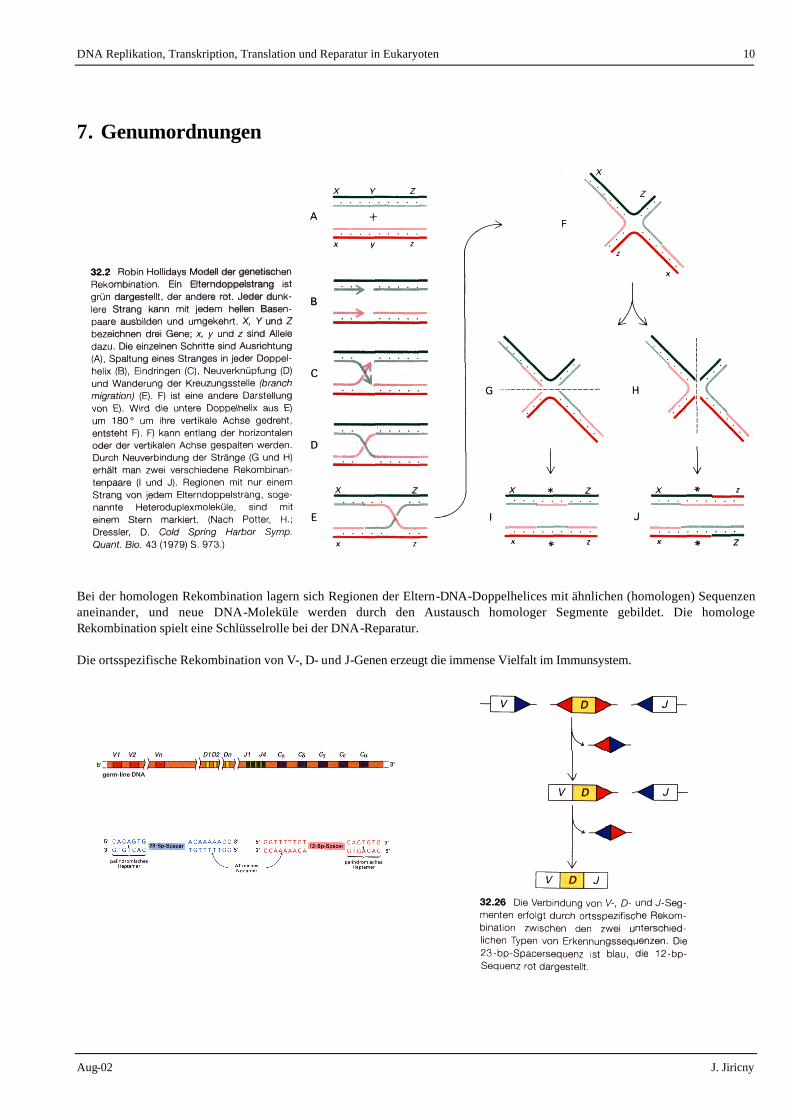
Bei der homologen Rekombination lagern sich Regionen der Eltern-DNA-Doppelhelices mit ähnlichen (homologen) Sequenzen aneinander, und neue DNA-Moleküle werden durch den Austausch homologer Segmente gebildet. Die homologe Rekombination spielt eine Schlüsselrolle bei der DNA-Reparatur. Die ortsspezifische Rekombination von V-, D- und J-Genen erzeugt die immense Vielfalt im Immunsystem.
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 11
Aug-02 J. Jiricny
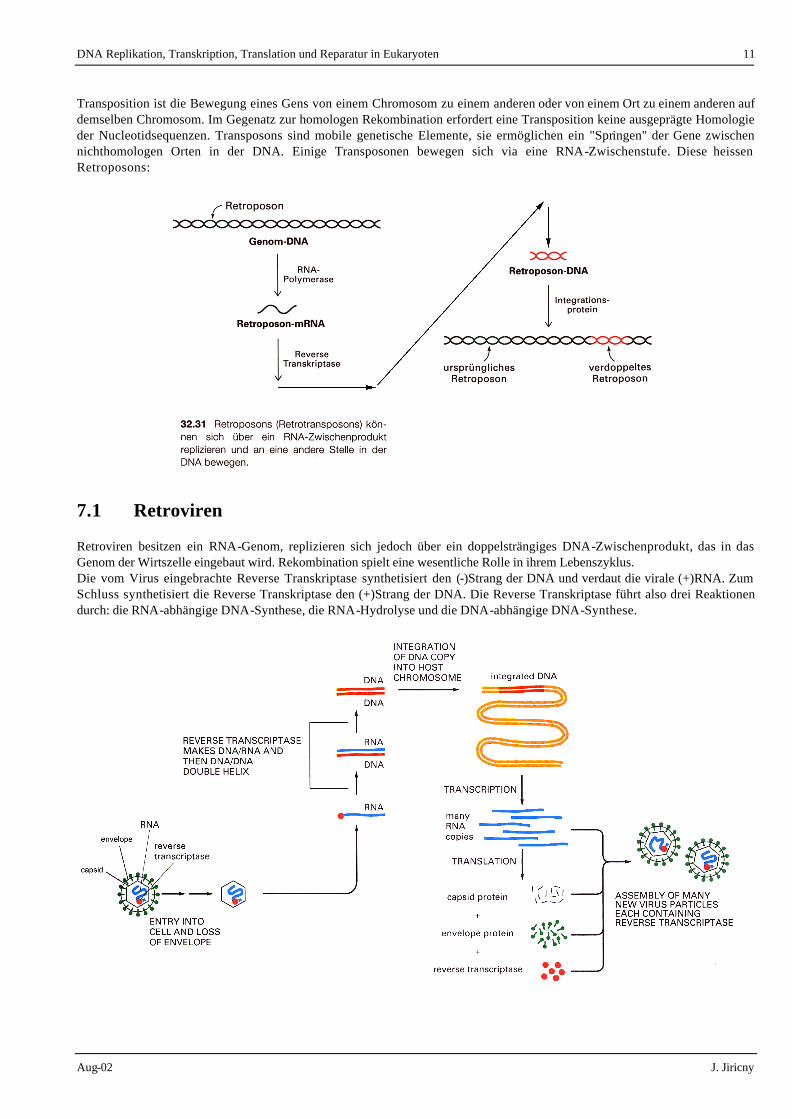
Transposition ist die Bewegung eines Gens von einem Chromosom zu einem anderen oder von einem Ort zu einem anderen auf demselben Chromosom. Im Gegenatz zur homologen Rekombination erfordert eine Transposition keine ausgeprägte Homologie der Nucleotidsequenzen. Transposons sind mobile genetische Elemente, sie ermöglichen ein "Springen" der Gene zwischen nichthomologen Orten in der DNA. Einige Transposonen bewegen sich via eine RNA-Zwischenstufe. Diese heissen Retroposons:
7.1 Retroviren
Retroviren besitzen ein RNA-Genom, replizieren sich jedoch über ein doppelsträngiges DNA-Zwischenprodukt, das in das Genom der Wirtszelle eingebaut wird. Rekombination spielt eine wesentliche Rolle in ihrem Lebenszyklus. Die vom Virus eingebrachte Reverse Transkriptase synthetisiert den (-)Strang der DNA und verdaut die virale (+)RNA. Zum Schluss synthetisiert die Reverse Transkriptase den (+)Strang der DNA. Die Reverse Transkriptase führt also drei Reaktionen durch: die RNA-abhängige DNA-Synthese, die RNA-Hydrolyse und die DNA-abhängige DNA-Synthese.
DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 12
Aug-02 J. Jiricny
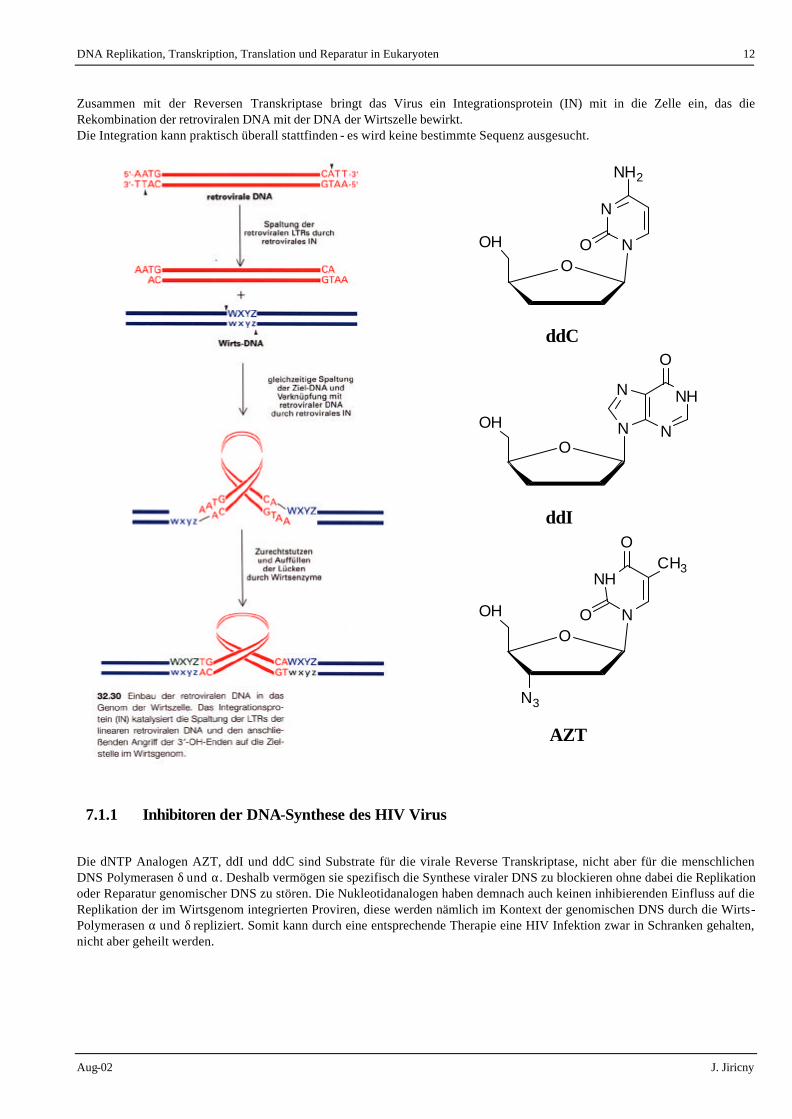
Zusammen mit der Reversen Transkriptase bringt das Virus ein Integrationsprotein (IN) mit in die Zelle ein, das die Rekombination der retroviralen DNA mit der DNA der Wirtszelle bewirkt. Die Integration kann praktisch überall stattfinden - es wird keine bestimmte Sequenz ausgesucht.
O
OH N
NHN
N
O
ddI
N
N
NH2
OO
OH
ddC
N
NH
O
OO
OH
AZT
N3
CH3
7.1.1 Inhibitoren der DNA-Synthese des HIV Virus
Die dNTP Analogen AZT, ddI und ddC sind Substrate für die virale Reverse Transkriptase, nicht aber für die menschlichen DNS Polymerasen δ und α. Deshalb vermögen sie spezifisch die Synthese viraler DNS zu blockieren ohne dabei die Replikation oder Reparatur genomischer DNS zu stören. Die Nukleotidanalogen haben demnach auch keinen inhibierenden Einfluss auf die Replikation der im Wirtsgenom integrierten Proviren, diese werden nämlich im Kontext der genomischen DNS durch die Wirts-Polymerasen α und δ repliziert. Somit kann durch eine entsprechende Therapie eine HIV Infektion zwar in Schranken gehalten, nicht aber geheilt werden.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 13
Aug-02 J. Jiricny
8. Mutagenese und DNA Reparatur
Mutationen werden durch verschiedenartige Veränderungen der Basensequenz in der DNA verursacht. Verschiedene Mutationstypen sind bekannt: 1) die Substitution eines Basenpaares durch ein anderes, 2) die Deletion eines oder mehrerer Basenpaare und 3) die Insertion (oder Addition) eines oder mehrerer Basenpaare. Die Substitution eines Basenpaares durch ein anderes ist der verbreitetste Mutationstyp. Eine Transition ist der Ersatz eines Purins durch ein anderes oder eines Pyrimidins durch ein anderes. Dagegen ist eine Transversion der Ersatz eines Purins durch ein Pyrimidin oder umgekehrt.
Einige chemische Mutagene wirken sehr spezifisch (z.B. 5-Bromuracil und 2-Aminopurin, die AT→GC Transitionen verursachen). Andere Mutagene wirken durch chemische Modifikation der DNA-Basen (z.B. salpetrige Säure und Hydroxylamin, die durch Desaminierung von Cytosin und Adenin, Transitionen von CG→AT und AT→GC verursachen). Flache aromatische Moleküle (Benzopyrene, Akridine) interkalieren in die DNA. Sie führen dadurch zur Insertion oder Deletion von einem oder mehreren Basenpaaren. Einwirkung von ultraviolettem Licht auf DNA bildet Pyrimidindimere, die auch mutagen wirken können.
DNA-Polymerasen machen Fehlpaarungen. Manche DNA-Modifikationen blockieren die Replikations- und Transkriptionsprozesse. Mutationen in Genen können zu Zelltransformation und Krebs führen. Durch unreparierte Schäden können Mutationen entstehen, die nach
Zellteilung an die nächste Generation weitergegeben werden. Die Änderung der Genstruktur durch Mutation kann sich in Verlust oder Änderungen der Aktivität eines Proteins, seiner Regulierbarkeit, seiner Fähigkeit spezifische Bindungen einzugehen (z.B. mit Cofaktoren, Substraten) oder seiner Stabilität auswirken. Beispiele für Störungen im Katabolismus sind: Galaktosämie, Phenylketonurie, Glucose-6-P-Mangel, Glykogenspeicher-krankheiten. Beispiel für Störungen im Anabolismus: Porphyrien. Beispiele für Störungen komplexer Funktionen: Hämoglobinopathien, Hämophilie, Xeroderma Pigmentosum, HNPCC. Alle DNA-Schäden und Modifikationen müssen deswegen effizient repariert werden. DNA-Schäden können repariert werden, da die genetische Information in beiden Strängen der Doppelhelix gespeichert ist, so dass die in einem Strang verlorengegangene Information aus der im anderen Strang ersetzt werden kann.
8.1 Schadenumkehrung
Ein UV-Pyrimidindimer kann photochemisch gespalten werden und zurück zu zwei Thymidinen rekonvertiert werden. Dies wird mit Licht und DNA-Photolyase katalysiert. 6-O-Methylguanin, ein Produkt der Chemotherapie, wird durch den Transfer der Methylgruppe auf das “Kamikaze”-Enzym, DNA-Alkyltransferase, zu Guanin konvertiert. DNA-Ligasen reparieren einfache (3’-OH, 5’-Phosphat) Strangbrüche.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 14
Aug-02 J. Jiricny
8.2 Basenexcisionsreparatur (BER)
Beschädigte oder modifizierte Basen werden generell durch Basenexcision entfernt. Z.B. Cytosin in der DNA desaminiert zu einem merklichen Prozentsatz auch spontan zu Uracil. Die Desaminierung des Cytosins ist potentiell mutagen, da sich während der Replikation Uracil mit Adenin paart. Somit wird eines der Tochtermoleküle ein AU-Basenpaar anstelle des ursprünglichen GC-Basenpaares haben.
Ein Reparatursystem beugt dieser Mutation vor: das Uracil wird als fremde Base in der DNA erkannt und entfernt.
8.3 Nukleotidexcisionsreparatur
Excision eines Thymindimeres, das durch Einwirkung von ultraviolettem Licht auf DNA gebildet wird:
Xeroderma pigmentosum entsteht durch einen Defekt in der menschlichen UV-Excinuclease. Die betroffenen Gene sind XPA-XPG. Das XPV (Variant) Protein ist nicht beteiligt bei der Reparatur. Das Gen kodiert für eine Polymerase, die Pyrimidindimere toleriert und übergehen kann (by-pass polymerase), ohne durch die Lesion blockiert zu werden.

DNA Replikation, Transkription, Translation und Reparatur in Eukaryoten 15
Aug-02 J. Jiricny
8.4 Fehlpaarungsreparatur
Bei der Reparatur der Fehlpaarungen ist die Identifizierung des Matrizen- und des Tochterstranges erforderlich. Betrachten wir ein GT-Basenpaar in einem neusynthetisierten DNA-Molekül. Wie unterscheidet die Reparaturmaschine zwischen dem authentischen Eltern-strang und dem fehlerhaften Tochterstrang? Die Mar-kierungen zur Identifizierung der Stränge bestehen in E. coli aus Methylgruppen an Adeninresten in GATC-Sequenzen.
In Eukaryoten ist kein MutH Homolog gefunden worden. MutS existiert als Heterodimer von hMSH2 und hMSH6, MutL als Heterodimer von hMLH1 und hPMS2. Der erbliche nichtpolypöse kolorektale Krebs (HNPCC, hereditary nonpolyposis colorectal cancer) ist die Folge einer defekten DNA-Fehlpaarungsreparatur. Die am häufigsten mutierten Gene in HNPCC Familien sind das mutS homolog hMSH2 und mutL homolog hMLH1.
8.5 Strangbruchsreparatur
Strangbrüche werden entweder durch homologe Rekombination (A) oder durch "non-homologous end-joining (NHEJ)" (B) repariert. Während der homologen Rekombination wird das Schwesterchromatid als Matrize verwendet. Somit geht keine genetische Information verloren. Während des "non-homologous end-joining" werden die beiden Enden zuerst exonukleolytisch abgebaut und dann religiert. Dieser Prozess führt zum Verlust von genetischer Information. A
B





























