Embed Size (px)
Citation preview

mRNA Trans-Splicing in the Human Parasitic Cestode Echinococcus
multilocularis
Klaus Brehm*, Katharina Jensen, and Matthias Frosch
Institut für Hygiene und Mikrobiologie, Universität Würzburg, Josef-Schneider-Straße 2,
97080 Würzburg, Germany
* To whom correspondence should be addressed: Tel: +931-201-3936; Fax: +931-201-
3445; Email: [email protected]
Running title:
Trans-splicing in the cestode Echinococcus multilocularis §
Copyright 2000 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on September 5, 2000 as Manuscript M006091200 by guest on June 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

SUMMARY
An identical 36 nucleotide exon was identified at the 5’ termini of different mRNAs from the cestode
Echinococcus multilocularis. We provide evidence that this exon constitutes a new spliced leader (SL) which is
obviously spliced in trans to echinococcal pre-mRNAs, donated by a non-polyadenylated, trimethylguanosine-
capped SL-RNA of 104 nucleotides. Sequence comparisons indicated that cestode and trematode SLs are likely
to be derived from a common ancestor gene. No conservation was, however, observed concerning the spectrum
of mRNAs which is trans-spliced in cestodes and trematodes, indicating that trans-splicing of a particular
flatworm mRNA is not correlated with the function of the encoded protein. We also show that the echinococcal
gene elp, encoding a member of the ERM protein family, is expressed via two alternative transcripts, spliced
either in cis or in trans at an identical splice acceptor site. This was accompanied by the formation of different
elp primary transcripts, harboring a complete or a truncated upstream intron, which supports the hypothesis that
alternative cis/trans-splicing depends on the presence or absence of an upstream splice donor site. A putative
SL gene was also identified on chromosomal DNA of Echinococcus granulosus, indicating widespread
utilization of trans-splicing in the genus.
INTRODUCTION
2
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Trans-splicing is a mechanism of mRNA processing which involves the fusion of exons from independent
primary transcripts to form a mature mRNA (for recent reviews, see Refs. 1-5). In the most common form of
trans-splicing, called SL1 trans-splicing, a small „mini-exon“ (or SL) is added to the 5’ ends of pre-mRNA
molecules eventually forming the 5’ terminal exon of the mature mRNA. In all cases investigated so far, the SL
donor molecule is a small, non-polyadenylated nuclear RNA (the SL-RNA) with structural properties similar to
the snRNAs which function in „conventional“ cis-splicing (1-5). SL trans-splicing was first described in
kinetoplastid protozoans where all mature mRNAs contain an identical leader sequence at the 5’ end (6). Among
metazoans, SL trans splicing is known for several parasitic and non-parasitic nematodes (1-6) and, in the
phylum Platyhelminthes, for parasitic trematodes (7-9) and free-living turbellarians (9). In contrast to
kinetoplastids, where trans-splicing is the predominant form of splicing, transcripts of metazoans are generally
spliced both in cis, usually at introns within the coding region, and in trans, at the 5’ end (1-5). A further
difference is that in metazoans not all mRNAs are trans-spliced. In C. elegans, for example, around 60 % of all
transcripts contain one leader, called SL1, and about 10-15 % contain an alternative mini-exon, called SL2 (1-
5). Besides SL1 and SL2, three additional SLs were identified in C. elegans which are, however, less frequently
found at mRNA 5’ ends (10).
Although significant data has been obtained on the biochemistry of the trans-splicing mechanism itself (1-6),
the biological in vivo functions of SLs are less well understood. At least one cellular mechanism which employs
trans-splicing is the processing of polycistronic messages into individual coding units as demonstrated for
trypanosomes and C. elegans (1-5). A role of the SL in resolving polycistronic messages has also been
suggested for the trematode S. mansoni (11). However, only a fraction of around 25 % of all C. elegans genes
are expressed as operons and among the monocistronically transcribed mRNAs, some are trans-spliced and
some are not (3, 4). This strongly implies additional functions of SLs and it has been proposed that they may
participate in the regulation of translation, in controlling mRNA stability, or in directed transport of transcripts
within the cell (1, 4, 12). So far, no specific sequence motifs critical for SL function have been identified since
SLs of different phylogenetic groups are highly divergent at the level of primary sequence. A particular role for
SL function in helminths could, however, have the TMG-cap of trans-spliced transcripts which derives from
the donor RNA and which distinguishes these mRNAs from non-trans-spliced transcripts containing a MMG-
cap (13-15).
3
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

After the description of SLs in parasitic nematodes and trematodes, some, as yet unsuccessful, attempts have
been made to also identify trans-splicing in the third group of medically important helminths, the cestodes (7,
9). In this study we have investigated gene expression mechanisms in the cestode Echinococcus multilocularis,
the so-called fox-tapeworm. The larval stage of E. multilocularis is the causative agent of alveolar
echinococcosis, which is considered to be the most lethal helminthic infection in humans (16). As in the case of
the closely related dog-tapeworm E. granulosus (16), the causative agent of cystic echinococcosis, no
information on gene regulatory mechanisms is available for E. multilocularis and only a few genes have been
characterized for both species. We now present evidence that the echinococcal gene elp, encoding a member of
the ERM-family of proteins (17, 18), is expressed via two alternatively spliced transcripts of which one
contains an echinococcal spliced leader at its 5’ end. We have identified several echinococcal mRNAs which are
processed by trans-splicing and provide a detailed characterization of the SL encoding gene and the SL-RNA.
4
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTAL PROCEDURES
Organisms- Origin and characteristics of the natural E. multilocularis isolates H-95 as well as E. granulosus
larvae from an Italian bovine hydatid cyst have been described before (19, 20). E. multilocularis isolate K188
was obtained as infective eggs from the intestine of a fox from the region of Carinthia, Austria, and further
propagated in mongolian jirds as described before (19).
Parasite cultivation and nucleic acid isolation- E. multilocularis larvae were routinely kept in mongolian jirds
(Meriones unguiculatus) as described (19). In vitro cultivation of metacestodes was performed according to (19)
and protoscoleces were isolated as described in (21). Chromosomal DNA and total RNA was isolated from in
vivo cultivated parasite material according to previously described protocols (22). For RNA isolation from in
vitro cultivated metacestodes and protoscoleces the RNeasy-KIT (Quiagen, Hilden, Germany) was used
according to the manufacturers instructions.
Blot analyses and cloning procedures- For Northern blot experiments, RNA samples were separated on 2 %
formaldehyde agarose gels or on 8 % polyacrylamide/7 M urea gels, transferred, and hybridized with 32P-
labeled oligonucleotide probes as described previously (7). Cloning of PCR products and PCR generated
cDNAs was performed by TA-cloning employing the TOPO-TA cloning KIT (Invitrogen) according to the
manufacturers instructions.
5’- and 3’-RACE- 5’-RACE was performed using oligo-dG-tailing of ss-cDNA and subsequent PCR-
amplification essentially as described before (17). For 3’-RACE of the SL-RNA, total RNA was first poly-A-
tailed using polyA-polymerase, reverse transcribed, and then amplified using an internal primer for the SL (5’-
TGC AGT TTT GTA TGG TGA GT-3’) and a primer for the polyA-tail according to a previously described
protocol (23). The SL RNA 3’ end was subsequently determined by sequencing of obtained PCR products.
Preparation of polyA+ RNA and TMG-immunoprecipitation- For the isolation of polyA+ and polyA- RNA
fractions the Oligotex mRNA purification -KIT (Quiagen, Hilden, Germany) was used according to the
manufacturers instructions. After repeated application of total RNA to oligo-dT columns, both the polyA+ and
the polyA- fractions were collected, precipitated and subjected to gel electrophoresis as described above. Anti
TMG immunoprecipitation was performed as described before (24) using the monoclonal anti-TMG antibody
K121 (Calbiochem, Schwalbach, Germany).
PCR and RT-PCR amplification- For standard PCR amplifications, the Eurogentec (Seraign, Belgium)
5
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

GoldStar Taq-system was used. For the amplification of long products the Advantage 2 PCR Kit (Clontech,
Palo Alto, CA, USA) was employed according to the manufacturers instructions. In RT-PCR reactions, mRNA
was reverse transcribed using Omniscript reverse transcriptase (Quiagen, Hilden, Germany) and subsequently
either PCR amplified as described above or using the One Step PCR Kit (Quiagen, Hilden, Germany). Detection
of pre-mRNA was performed using the sensitive Titan One Step RT-PCR System of Boehringer (Mannheim,
Germany). PCR primers were purchased from ARK Scientific (Darmstadt, Germany). Sequences of
oligonucleotides described in the text were: SL-5 (5’-AAG GAC CGA TTA ACG GTG-3’) and SL-3 (5’-
ACC TTG CAG TTT TGT ATG-3’) for cloning the SL repeat; E10-6 (5’-ATC GTA CTG TTC CTT GCT G-
3’), E10-24 (5’-GAT GAT GCT CCG CAT AAT CCA-3’), E10-19 (5’-CGT TGG CAG TAT TCT CAC
CG-3’), E10-31 (5’-TTC ACT TCT CGT GAA CAG TAG-3’), and E10-29 (5’-CAG TAG CCT CCT TGT
TTG AG-3’) for mapping the pre-mRNA start point in the 5’-intron.
cDNA library construction- 1 µg of total E. multilocularis RNA was reverse transcribed as described above
using primer A-37 (5-GGC CGC ATG CCG ACT AGT ACT17-3) directed to the mRNA polyA-tail. The
resulting ss-cDNA was PCR amplified with primer A-20 (5-GGC CGC ATG CCG ACT AGT AC-3) and
primer SL-5PR (5’-CAC CGT TAA TCG GTC CTT AC-3’) specific for the echinococcal SL using the
Advantage 2 PCR Kit (Clontech, Palo Alto, Ca, USA) and employing 20 PCR cycles (1 min. at 94 °C, 1 min. at
57 °C, 5 min. at 68 °C) to avoid over-representation of small PCR products. The products were subsequently
cloned as described above.
Competitive PCR- Competitive PCR experiments were performed in adaptation to previously described
methods (25, 26). For measuring amounts of conventionally and trans-spliced elp-transcripts, one single
competitor was constructed as follows: by overlap extension (27) of PCR generated elp chromosomal and
cDNA fragments, a 584 bp fragment was constructed which contained the primers EC5 (5’-GGG CTC CCG
TTG ATT GCA GT-3’; see cDNA nt 11-30 in Ref. 17), specific for exon I, and E10-6 (5’-ATC GTA CTG
TTC CTT GCT GG-3’), spanning the exon IV/exon V boundary (17), at its ends. Due to the presence of elp
intron II (but absence of the 5’ intron and intron I), this fragment was 74 bp longer than the conventionally
spliced mRNA fragment between EC5 and E10-6. By further overlap extension, oligonucleotide ET5 (5’-TCC
TTA CCT TGC AGT TTT GT-3’), specific for the SL, was fused to the EC5 binding site of the above
fragment. The resulting fragment for ET5 and E10-6 was 94 bp longer than the corresponding fragment for the
6
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

trans-spliced elp mRNA. Serial dilutions of the resulting fragment were then used in competitive PCR with
constant amounts of cDNA produced from RNA isolated from metacestodes and protoscoleces applying 30
cycles of 1 min., 94 °C, 1 min., 57 °C, 1 min., 72 °C and using the primer combinations EC5 x E10-6 for elp
transcript A as well as ET5 x E10-6 for transcript B. As a control for total elp mRNA, the competitor was used
for PCR experiments with primer E10-6 and E10-15 (5’-AAT AAG GTC AGG GTG ACT AC-3’; see cDNA
nt 69-89 in Ref. 17), specific for elp exon III. Equimolarity of cDNA and competitor was determined by 2 %
agarose gel electrophoresis and ethidium bromide staining.
7
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

RESULTS
Identification of alternatively spliced elp transcripts - We have previously characterized the chromosomal elp-
locus and investigated elp transcription by 5’-RACE (17). One transcript, herein termed transcript A, was
identified which derived from a fusion of the non-translated 32 nt exon I to the translational start-codon
containing exon II thus revealing the presence of a 386 nt intron in the 5’ non-translated region (a so-called 5’-
intron) (Fig. 1). In the present study, we have analyzed further 5’-RACE clones and identified a second
transcript, B, in which exon I was precisely replaced by a so far unknown 36 nt sequence (“spliced leader” in
Fig. 1). This alternative exon showed no homology to exon I and was not present within the elp locus or in
chromosomal regions of around 2 kb down- or upstream of the coding region. Sequence analysis also revealed
that transcripts A and B were identical from exon II to the polyA addition site and that, indeed, the only
difference was the alternative exon at the 5’ end. Southern hybridization (28) and PCR analysis of E.
multilocularis chromosomal DNA furthermore indicated that elp is present as a single copy gene per haploid
genome and that the isolates investigated in this study were homozygous for the locus (data not shown). Hence,
transcripts A and B are likely to be derived from the same allele.
< Figure 1 suggested >
We were interested in the relative abundance of both transcripts and conducted competitive PCR experiments
using upstream primers specific for exon I or the alternative exon and downstream primers specific for the exon
IV/V boundary of elp. We consistently measured amounts of transcript A and transcript B in a relation of
around 1:2 using parasite material isolated from Meriones unguiculatus as an intermediate host (not shown).
Since this parasite material was composed of two different developmental stages of E. multilocularis, the
metacestode and the protoscolex, we investigated whether stage specifical differences in elp expression could
lead to the alternative transcripts. However, in protoscoleces manually picked from parasite material as well as
in metacestode vesicles after in vitro cultivation, a constant relation of about 1:2 between transcript A and B was
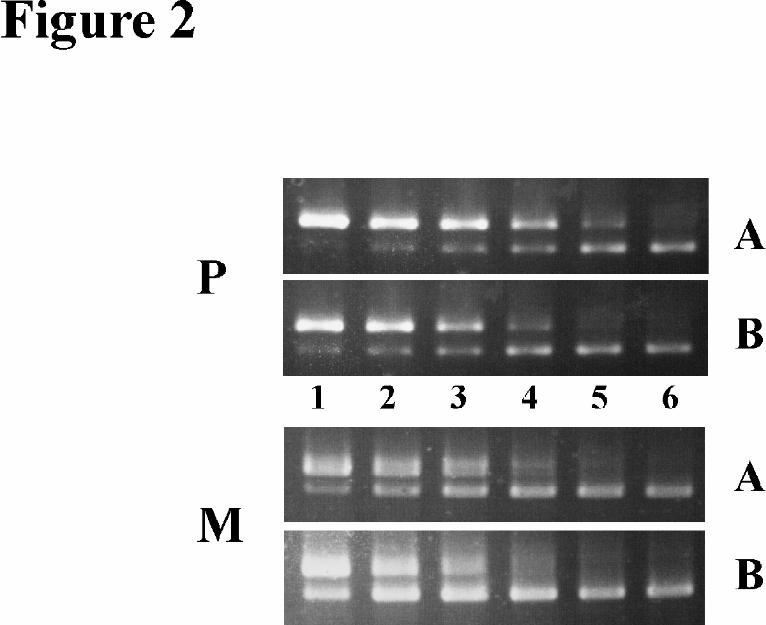
observed (Fig. 2).
< Figure 2 suggested >
8
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Identification of an E. multilocularis SL-RNA- Based on the data outlined above we assumed that exon I was
fused to exon II by „conventional“ cis-splicing and that the alternative 36 nt exon derived from trans-splicing
involving an echinococcal SL-RNA. This hypothesis was tested by Northern blot experiments using antisense
oligonucleotides specific for exon I (probe A) and the 36 nt exon (probe B) as probes. As expected, probe A
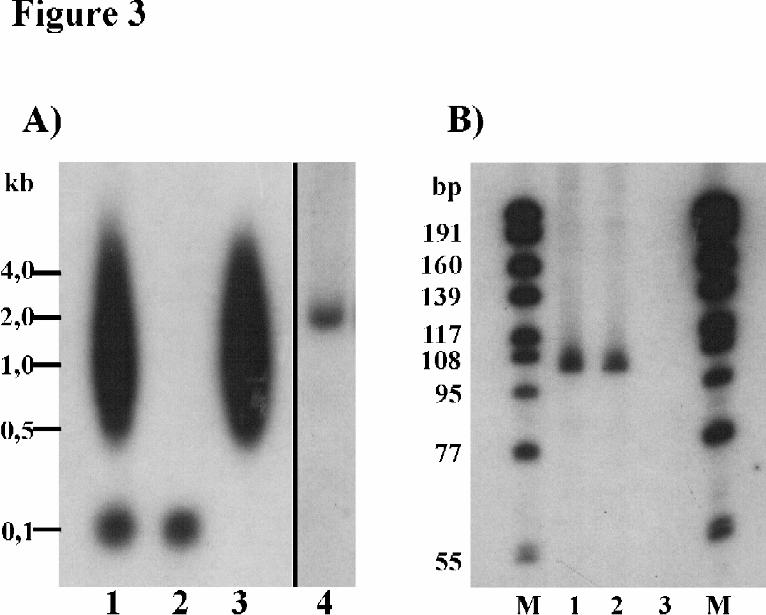
identified a single band of ~ 2,1 kb (Fig. 3A) which corresponded to the previously determined size of the elp
transcript (20). Probe B, on the other hand, produced a hybridization smear for polyadenylated transcripts
ranging between 500 nt and several kb (Fig. 3A). In addition, probe B produced a strong hybridization signal for
a non-polyadenylated RNA of ~ 100 nt (Fig. 3A/B) which was present in the TMG-precipitable RNA fraction
but absent in the non-TMG fraction (Fig. 3B). These considerable similarities to trans-splicing elements of
nematodes and trematodes (see above) led us to conclude that the 36 nt alternative exon of elp constitutes a SL
from E. multilocularis which was donated to a set of different transcripts by a SL-RNA of around 100 nt. It
should be noted that results similar to that described above were also obtained when RNA from protoscolex or
metacestode was analyzed (not shown). Hence, at least for these two developmental stages, no stage specific
patterns of SL expression or trans-splicing were observable.
< Figure 3 suggested >
To test whether the TMG-cap deriving from the donor RNA was still present on trans-spliced echinococcal
transcripts, we performed elp-specific RT-PCRs on mRNA of the TMG-precipitated fraction. In these
experiments, we could clearly identify the trans-spliced transcript B of elp whereas no amplification products
were obtained for the conventionally spliced transcript A (not shown). Hence, conventionally and trans-spliced
elp transcripts could be clearly distinguished by the absence or presence, respectively, of a 5’ TMG-cap.
Cloning and characterization of the SL-RNA encoding gene- In all organisms investigated so far, the SL-RNA
encoding genes are present on DNA-fragments which are directly repeated more than 100-fold on the
chromosome (6-8). Assuming that this is also the case for E. multilocularis, we chose a PCR approach to clone
the SL encoding gene, emsl, using two primers („SL-5“, „SL-3“) which were specific for the SL but which
were oriented in opposite directions. After PCR amplification from chromosomal E. multilocularis DNA as a
template, a 1513 bp fragment was obtained, cloned in E. coli and entirely sequenced. As expected, the 36 nt SL
sequence was immediately followed by a consensus splice donor sequence (5’-GTGCGT-3’) (Fig. 4A). We
9
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

then used the sequence information of the SL downstream region to (i) verify the SL 5’ end present on the 5’-
RACE product by primer extension (not shown), and (ii) precisely map the SL-RNA 3’ end by 3’-RACE (see
experimental procedures) to a position 104 bp downstream of the SL 5’ end. A computer-generated, energy
minimized RNA secondary structure for the SL-RNA is shown in Fig. 4B. All conserved features of other SL-
RNAs (4, 6, 8, 9), including three stem-loop structures, a putative Sm-binding sequence between stem-loop 2
and 3, and the typical location of the SL within the first stem-loop are present.
Sequence analysis of the 1513 bp fragment harboring emsl did not indicate the presence of putative protein-
coding regions or of other snRNA-encoding genes. Furthermore, unlike as for nematode SL repeats, no
sequences homologous to 5S rRNAs were identified in the E. multilocularis sequence.
Southern blot analysis revealed that the 1,5 kb fragment was, indeed, directly repeated on the E. multilocularis
chromosome (data not shown). To measure the copy number we chose a competitive PCR approach using as a
reference the elp - locus which was known to be present as a single copy gene (28). On chromosomal DNA of
different natural and clinical E. multilocularis isolates we constantly measured a relation of around 20 : 1
between emsl and elp (data not shown), revealing a SL-gene copy number significantly lower than that
previously estimated for trematodes (100-200; Refs. 7, 8) and nematodes (100; Ref. 6).
Characterization of the SL repeat and the SL gene, egsl, from E. granulosus- To investigate the possibility of
trans-splicing in the dog-tapeworm, we performed PCR experiments using several primers specific for the E.
multilocularis SL repeat and chromosomal E. granulosus DNA as a template (not shown). These studies led to
the characterization of a 1555 bp repeated element which shared 87 % homology with the E. multilocularis
repeat. Two nucleotide substitutions were observed for the SL-RNA of which one was located in the SL
sequence (Fig. 4A). A sequence comparison of the echinococcal SL genes and the previously characterized SL-
RNAs of the parasitic flatworms S. mansoni and F. hepatica is depicted in fig. 4A. Apart from some conserved
regions, particularly around the splice donor site, we could merely detect an overall homology of around 30 %
between the cestode and trematode SLs although higher degrees of conservation (50-55 %) were measurable for
the SLs. No significant homology was observable between echinococcal SL-RNAs or SLs and those of
nematode or protozoan origin.
< Figure 4 suggested >
10
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Characterization of additional trans-spliced mRNAs- To verify that the hybridization smear for polyadenylated
RNAs shown in Fig. 3 did indeed represent a set of mRNAs which harbor the SL at the 5’ end, a cDNA-library
specific for trans-spliced transcripts was constructed. After reverse transcription of E. multilocularis mRNA and
PCR amplification of the ss-cDNA with primers specific for the polyA-tail and for the 5’ end of the SL (see
also experimental procedures), a smear of products between 500 bp and ~ 4 kb was obtained (not shown). The
PCR products were cloned in E. coli and the recombinant plasmids of 40 randomly chosen colonies were
sequenced. In all cases, the 18 bp of the SL-specific amplification primer were followed by the remaining 18 bp
of the SL identified for elp, indicating that the cDNAs derived from trans-spliced mRNAs and that in all an
identical SL was present. For 3 of the cDNAs (Em4231, Em1342, Em2222) we also determined the
chromosomal situation and found consensus splice acceptor sequences immediately upstream of the mRNA SL
addition site (not shown). ORFs could be identified in the inserts of 39 plasmids and the deduced amino acid
sequences of 17 revealed significant database matches with already known proteins. Six were homologous to
proteins with unknown functions. The remaining 11 are listed in table 1. Concerning the putative biological
function of the encoded proteins, no common patterns were identified although 3 of the factors, corresponding
to clones Em4231, Em3332, and Em4121, are potentially involved in splicing processes. The remaining factors
are potentially involved in cellular processes as different as, for instance, glycolysis, pyrimidine metabolism, the
electron transport chain, or translation initiation (see Table 1).
< Table 1 suggested >
With its 3’ end the echinococcal SL provides trans-spliced mRNAs with an AUG codon which, according to the
first-AUG-rule (32), constitutes a potential ORF translation initiation start point. However, when analyzing the
5’ ends of the above mentioned trans-spliced echinococcal mRNAs we found the SL-AUG in 7 of 12 cases out
of frame to the actual start-AUG as predicted by sequence comparison (Table 1). In 3 cases, including elp, the
SL-AUG was present in the same reading frame as the predicted start-AUG. For elp this could possibly result
in the production of two different proteins from the 5’ cis- and trans-spliced transcripts (see Fig. 1). N-terminal
sequencing of immunoprecipitated Elp protein, however, revealed a uniform N-terminus identical to the
sequence deduced from the 5’ cis-spliced transcript A (not shown). We therefore conclude that in both elp
transcripts the same AUG start-codon is used and that the SL-AUG of transcript B does not serve for
11
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

translation initiation. For 2 transcripts (Em1433, Em1431), on the other hand, sequence comparisons indicated
that the SL-AUG codon most probably serves as the actual translation start. In both cases, translation from
alternative downstream AUGs would result in products which lack a considerable part of structures that are
highly conserved among the homologous members of the respective protein families.
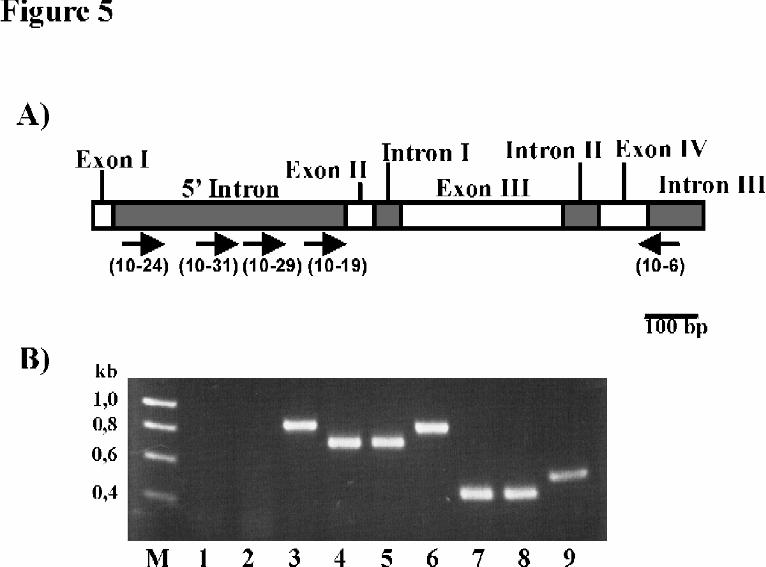
Identification of alternative elp primary transcripts. In a final set of experiments we were interested whether
alternative cis/trans-splicing at the splice-acceptor site upstream of elp exon II did occur at the same pre-
mRNA or whether two different primary transcripts were involved. By primer extension experiments, only the
completely spliced 5’ ends of transcripts A and B could be identified (data not shown) presumably because the
concentration of pre-mRNA in the echinococcal RNA preparations was below detection level. We therefore
chose RT-PCR using one downstream primer spanning the exon IV/exon V boundary of the elp coding region
(primer „10-6“) and two different upstream primers which were located close to the 5’ intron’s splice-donor
(„10-24“) and splice-acceptor sites („10-19“), respectively. Interestingly, with the combination 10-6 x 10-19,
a PCR product was obtained which, as determined by DNA-sequencing, was completely spliced for the introns
I, II and III of elp but still retained the 3’ part of the 5’ intron (see Fig. 5). The combination 10-6 x 10-24, on
the other hand, did not reveal a PCR product, even with elevated amounts of RNA. The likeliest explanation for
these results is the presence of an additional transcriptional start point within the 5’ intron which leads to the
expression of a second elp primary transcript that lacks the 5’ intron splice donor site. This indicates that cis-
splicing can most probably precede trans-splicing in E. multilocularis, similar to the situation previously
described for the nematode Onchocerca volvulus (33). Using further primer combinations, we localized the
alternative starting point to a region approximately between 256 and 244 nt upstream of the 5’ intron splice
acceptor site since the combination 10-6 x 10-29 still revealed a completely spliced fragment for the introns I -
III while the combination 10-6 x 10-31 gave no product (Fig. 5, Fig. 1).
< Figure 5 suggested >
Since trans-splicing can be associated with the expression of polycistronic transcripts, we investigated similar
aspects concerning elp. However, neither Northern blot hybridization experiments, nor RT-PCR amplification
using primers specific for elp and chromosomal upstream or downstream regions led to any indication of
polycistronic pre-mRNAs even when elevated amounts of RNA were applied (not shown). In addition, 800 bp
12
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

upstream of the elp 5’ intron we identified a putative ORF encoding a protein with 86 % amino acid sequence
identity to nuclear proteins of Drosophila melanogaster and Homo sapiens. This reading frame was, however,
transcribed in the opposite direction than elp (not shown). Up to 2 kb downstream of elp we could not identify
any additional reading frames. Taken together, these data indicate that elp is most probably expressed
monocistronically.
13
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

DISCUSSION
Several evidences, provided in this study, are clearly indicative of SL trans-splicing in E. multilocularis: (i) an
identical 36 nt exon was found at the 5’ ends of different echinococcal mRNAs at a position where, in the
respective chromosomal loci, a consensus splice acceptor site was located; (ii) the 36 nt exon was not part of
these chromosomal loci but was encoded by a gene, emsl, located elsewhere on the chromosome on a directly
repeated 1,5 kb fragment; (iii) the 36 nt exon formed part of a 104 nt, non-polyadenylated, TMG-capped RNA
with structural characteristics of snRNAs, where it was immediately followed by a splice donor consensus site.
These characteristics are highly reminiscent of SLs and SL-RNAs from kinetoplastid, nematode and trematode
origin (1-6). We conclude that the 36 nt exon is a SL from E. multilocularis and that the 104 nt RNA is the
corresponding SL-RNA. A gene highly homologous to emsl, egsl, was identified on chromosomal DNA of E.
granulosus, suggesting that mRNA trans-splicing is also used for gene expression in the dog-tapeworm.
By the identification of SLs in the genus Echinococcus we have, for the first time, provided evidence for trans-
splicing in tapeworms. Earlier studies on the distribution of trans-splicing in the phylum Platyhelminthes led to
the identification of SLs in several trematode species (7, 8, 11) and in the polyclad turbellarian Stylochus zebra
(11), representing two of the three major flatworm groups (34). The failure to detect similar SLs in cestodes
raised questions on a possible polyphyletic origin of flatworms, the loss of trans-splicing in flatworm-
subgroups, or the presence of evolutionary divergent SL-RNAs (11). Our results now indicate that trans-
splicing is, indeed, a widespread feature of the phylum, similar to the situation described for kinetoplastids and
nematodes (1-6). Furthermore, due to the observed sequence homologies in their primary sequences we suggest
that all flatworm SLs identified so far derive from a common ancestor gene.
Similarities and differences of the echinococcal trans-splicing elements to that of trematodes, nematodes, and
protozoa correlate with the phylogenetic position of cestodes. The Echinococcus SL-RNA displays certain
homologies to that of trematodes, but none to those of nematodes or kinetoplastids. The SL encoding genes of
all flatworms are located on chromosomal repeats which, albeit variable in length and copy number, do not
contain the 5S rRNA gene, a characteristic of SL repeats from nematodes (1), euglena (35), and some
trypanosomes (36). Furthermore, and typical for metazoans, not all echinococcal mRNAs are trans-spliced.
Analysis of elp transcripts revealed that trans-spliced and non trans-spliced mRNAs can even derive from one
single gene locus and, recently, we have characterized β-tubulin encoding transcripts of E. multilocularis (21)
which are exclusively expressed without spliced leader. Based on a preliminary analysis of 150 randomly
14
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

chosen, full-length cDNAs (data not shown) we estimate that ~ 25 % of all E. multilocularis mRNAs acquire
the SL, a figure close to what has been suggested for trematodes (37).
Two structural features, the 5’ TMG cap which is conserved among all SLs, and the 3’ AUG codon which is
specifically conserved in flatworm SLs, were also present in the echinococcal SLs. For C. elegans it has been
shown that the TMG cap is still retained on the mature mRNAs after trans-splicing (14, 15) and our data on the
alternatively spliced elp-transcripts suggest that this is also the case for flatworms. Using an Ascaris in vitro
system, evidence for a TMG-cap involvement in translation initiation has been obtained (12) and, recently,
several isoforms of the translation initiation factor 4E (eIF4E) were identified in C. elegans which can
differentiate between TMG and MMG containing transcripts (38). It will be worthwhile to investigate whether
alternative eIF4E expression also occurs in flatworms and whether this feature could be involved in translational
control mechanisms of trans-spliced mRNAs. Concerning the 3’ AUG codon and its possible role as a
translational start point, the situation in E. multilocularis closely parallels that in F. heptica (37). In 2 of 12
analyzed transcripts this AUG codon obviously provided the translation start point, whereas at least in 8 a
similar role could be excluded. Like in the case of the TMG cap, further experiments are required to investigate
whether the occasional provision of a translation start codon is the only function of the 3’ AUG.
A remarkable difference between cestodes and trematodes was the variability in genes which are expressed via
trans-splicing. Although the echinococcal UbCRBP and FBPA proteins identified in this study have clear
homologs in F. hepatica and S. mansoni (11, 31), the respective mRNAs are trans-spliced in E. multilocularis
but not in trematodes. Further examples are the enolase encoding mRNA which is trans-spliced in trematodes
but lacks SL sequences in 4 different cestode species (11) and a S. japonicum factor homologous to E.
multilocularis Elp which is obviously expressed independently of trans-splicing (39). Hence, differences in the
pattern of trans-spliced factors, previously observed between trematodes and nematodes (4), can also be
observed within the same phylum and concerning SLs of the same phylogenetic origin. This strongly argues
against a general correlation between trans-splicing of a mRNA and the biological function of the encoded
protein.
The mode of regulation of the E. multilocularis elp gene is highly remarkable. We have identified two
transcripts which (i) apparently derive from alternative cis/trans splicing at an identical splice acceptor site, (ii)
only differ in a small 5’ exon and in the presence or absence of a TMG-cap, and (iii) obviously encode an
identical protein. Although alternative cis/trans-splicing at internal introns was observed before in S. mansoni
15
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

(7, 40), this was always accompanied by the formation of full length and truncated forms of the encoded protein
and, therefore, probably associated with regulatory aspects acting at the protein level. The production of an
identical protein by alternative cis/trans-spliced transcripts is, to our knowledge, a novel finding and raises
several questions regarding its transcriptional regulation and biological significance. By use of reporter gene
constructs in C. elegans, Conrad et al. (41, 42) previously showed that introduction or removal of a splice donor
site upstream of a given splice acceptor site can alter the mode of splicing of resulting mRNAs from cis to trans.
This led to the hypothesis that solely the presence or absence of a 5’ splice site upstream of a 3’ splice site, and
not the sequence context around the splice acceptor, determines the mode of splicing. Our data on the presence
of two alternative elp primary transcripts, one containing the full length 5’ intron, the other a truncated version,
are in perfect agreement with this hypothesis. It is, therefore, reasonable to assume that the 5’ cis-spliced elp
transcript derives from the longer primary transcript whereas the 5’ trans-spliced elp mRNA is determined by
the truncated pre-mRNA. Why should E. multilocularis express Elp via transcripts spliced either conventionally
or in trans? On the one hand, this could be a simple consequence of developmentally regulated switching
between both elp promoters, leading to different pre-mRNAs for metacestode and protoscolex, or differential
expression of elp as part of monocistronic and polycistronic messages, accompanied by resolving polycistrons
via trans-splicing. Due to the constitutive expression pattern of conventional and trans-spliced elp mRNAs in
two developmental stages and the probable expression of elp as a monocistron we consider these scenarios
unlikely. The constant relation between the conventionally and trans-spliced elp transcripts in developmental
stages as different as metacestode and protoscolex indicates that both transcripts are most probably expressed at
the same time within the same cell. It remains to be established whether this expression pattern is related with
particular, as yet unknown functions or control mechanisms of the gene product in the echinococcal cell.
Currently, experiments are performed in our laboratory to measure differential stability, translational efficiency,
and intracellular location of the alternative elp transcripts. We predict that a closer analysis of the unusual
splicing mode of elp mRNAs will lead to valuable insights concerning the biological significance of SL trans-
splicing.
The discovery of trans-splicing in protozoans, nematodes and trematodes has greatly facilitated the PCR based
construction of full length cDNA libraries from these organisms (5, 37, 43-46). Particularly in the case of
parasitic helminths, of which usually only limited amounts of biological material are available, the presence of
identical leaders on mRNA subsets proved of high value for the identification of novel genes (5, 37, 46, 47).
16
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Based on our results, similar investigations are now possible for the two important human parasitic cestodes E.
multilocularis and E. granulosus, and possibly for other cestodes such as Taenia solium or T. saginata. The
conserved utilization of trans-splicing in all three major groups of parasitic helminths, nematodes, trematodes,
and cestodes, offers a potential target for the development of chemotherapeutic agents against a variety of
parasitic diseases.
Acknowledgements-We thank Kerstin Kronthaler for excellent technical assistance and Kerstin Hubert for
many helpful suggestions.
17
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

REFERENCES
1. Nilsen, T. W. (1993) Annu. Rev. Microbiol. 47, 413-440
2. Nilsen, T. W. (1995) Mol. Biochem. Parasitol. 73, 1-6
3. Blumenthal, T. (1995) Trends Genet. 11, 132-136
4. Davis, R. E. (1996) Parasitol. Today 12, 33-40
5. Blaxter, M., and Liu, L. (1996) Int. J. Parasitol. 26, 1025-1033
6. Donelson, J. E., and Zeng, W. (1990) Parasitol. Today 6, 327-334
7. Rajkovic, A., Davis, R. E., Simonsen, J. N., and Rottman, F. M. (1990) Proc. Natl. Acad. Sci. USA 87, 8879-8883
8. Davis, R. E., Singh, H., Botka, C., Hardwick, C., Ashraf el Meanawy, M., and Villanueva, J. (1994) J. Biol. Chem. 269, 20026-20030
9. Davis, R. E. (1997) Mol. Biochem. Parasitol. 87, 29-48
10. Ross, L. H., Freedman, J. H., and Rubin, C. S. (1995) J. Biol. Chem. 270, 22066-22075
11. Davis, R. E., and Hodgson, S. (1997) Mol. Biochem. Parasitol. 89, 25-39
12. Maroney, P. A., Denker, J. A., Darzynkiewicz, E., Laneve, R., and Nilsen, T. W. (1995) RNA 1, 714-723
13. Thomas, J. D., Conrad, R. C., and Blumenthal, T. (1988) Cell 54, 533-539
14. Liou, R. F., and Blumenthal, T. (1990) Mol. Cell. Biol. 10, 1764-1768
15. Van Doren, K., and Hirsh, D. (1990) Mol. Cell. Biol. 10, 1796-1772
16. Rausch, R. L. (1995) in Echinococcus and hydatid disease (eds. Thompson, R. C. A., Lymbery, A. J.) pp. 89-134, CAB International, Wallingford, UK
17. Brehm, K., Jensen, K., Frosch, P., and Frosch, M. (1999) Mol. Biochem. Parasitol. 100, 147-152
18. Hubert, K., Cordero, E., Frosch, M., and Solomon, F. (1999) Cell Motil. Cytoskel. 42, 178-188
19. Jura, H., Bader, A., Hartmann, M., Maschek, H. J., and Frosch, M. (1996) Infect. Immun. 64, 3484-3490
20. Frosch, P., Mühlschlegel, F., Sygulla, L., Hartmann, M., and Frosch, M. (1994) Parasitol. Res. 80, 703-705
21. Brehm, K., Kronthaler, K., Jura, H., and Frosch, M. (2000) Mol. Biochem. Parasitol. 107, 297-302
22. McManus, D. P., Knight, M., and Simpson, J. G. (1985) Mol. Biochem. Parasitol. 16, 251-266
23. Adams, D. S., Noonan, D., Burn, T. C., and Skinner, H. B. (1987) Gene 54, 93-103
24. Adams, D. S., Herrera, R. J., Luhrmann, R., and Lizardi, P. M. (1985) Biochemistry 24, 117-125
25. Diviacco, S., Norio, P, Zentilin, L., Menzo, S., Clementi, M., Biamonti, G., Riva, S., Falaschi, A., and Giacca, M. (1992) Gene 122, 313-320
26. Jayagopala Reddy, N. R., Wilkie, B. N., Mallard, B. A. (1996) BioTechniques 21, 868-875
18
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

27. Horton, R. M., Hunt, H. D., Ho, S. N., Pullen, J. K. and Pease, L. R. (1989) Gene 77, 61-68
28. Hemmings, L., McManus, D. P. (1991) Mol. Biochem. Parasitol. 44, 53-62
29. Milligan, J. F., Groebe, D. R., Witherell, G. W., and Uhlenbeck, O. C. (1987) Nucl. Acids Res. 15, 8783-8798
30. Mathews, D. H., Sabina, J., Zuker, M., and Turner, D. H. (1999) J. Mol. Biol. 288, 911-940
31. El-Dabaa, E., Mei, H., El-Sayed, A., Karim, A. M., Eldeskoy, H. M., Fahim, F. A., LoVerde, P. T., and Saber, M. A. (1998) J. Parasitol. 84, 954-960
32. Kozak, M. (1991) J. Cell Biol. 115, 887-903
33. Shiwaku, K., and Donelson, J. E. (1995) Biochem. Biophys. Res. Comm. 211, 49-53
34. Carranza, S., Baguna, J., and Riutort, M. (1997) Mol. Biol. Evol. 14, 485-497
35. Keller, M., Tessier, L. H., Chan, R. L., Weil, J. H. and Imbault, P. (1992) Nucl. Acids Res. 20, 1711-1715
36. Aksoy, S., Shay, G. L., Villanueva, M. S., Bears, C. B., and Richards, F. F. (1992) Gene 113, 239-243
37. Davis, R. E., Hardwick, C., Tavernier, P., Hodgson, S., and Singh H. (1995) J. Biol. Chem. 270, 21813-21819
38. Keiper, B. D., Lamphear, B. J., Deshpande, A. M., Jankowska-Anyshka, M., Aamodt, E. J., Blumenthal, T., and Rhoads, R. E. (2000) J. Biol. Chem. 275, 10590-10596
39. Kurtis, J. D., Ramirez, B. L., Wiest, P. M., Dong, K. L., El-Meanawy, A., Petzke, M. M., Johnson, J. H., Edmison, J., Maier, R. A., and Olds, G. R. (1997) Infect. Immun. 65, 344-347
40. Hamdan, F. F., and Ribeiro, P. (1998) Parasitol. Res. 84, 839-842
41. Conrad, R., Thomas, J., Spieth, J., and Blumenthal, T. (1991) Mol. Cell. Biol. 11, 1921-1926
42. Conrad, R., Liou, R. F., and Blumenthal, T. (1993) EMBO J. 12, 1249-1255
43. Martin, S. A., Thompson, F. J., and Devaney, E. (1995) Mol. Biochem. Parasitol. 70, 241-245
44. El-Sayed, N. M. A., Alarcon, C. M., Beck, J. C., Sheffield, V. C., and Donelson, J. E. (1995) Mol. Biochem. Parasitol. 73, 75-90
45. Levick, M. P., Blackwell, J. M., Connor, V., Coulson, R. M. R., Miles, A., Smith, H. E., Wan, K. L., and Ajioka, J. W. (1996) Mol. Biochem. Parasitol. 76, 345-348
46. Blaxter, M. L., Raghavan, N., Ghosh, I., Guiliano, D., Lu, W., Williams, S. A., Slatko, B., and Scott, A. L. (1996) Mol. Biochem. Parasitol. 77, 77-93
47. Henkle-Duhrsen, K., Tawe, W., Warnecke, C., and Walter, R. D. (1995) Biochem. J. 308, 441-446
19
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

FOOTNOTES:
§ This work was supported by grant Fr689/9-2 from the Deutsche Forschungsgemeinschaft (to M.F.). The costs
of publication of this article were defrayed in part by the payment of page charges. This article must therefore be
hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EBI Data Bank with
accession number(s) AJ29365 – AJ29377
1 The abbreviations used are: SL, spliced leader; SL-RNA, spliced leader RNA; snRNA,
small nuclear RNA; TMG, 2,2,7-trimethylguanosine; MMG, 7-methylguanosine; ERM,
ezrin/radixin/moesin; RACE, rapid amplification of cDNA ends; RT, reverse transcription;
PCR, polymerase chain reaction; ss, single stranded; nt, nucleotide(s); bp, basepair(s); kb,
kilobase(s); UbCRBP, ubiquinol-cytochrome C-reductase binding protein; FBPA, fructose-
bisphosphate-aldolase.
20
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

LEGENDS
Table 1. Trans-spliced mRNAs of E. multilocularis. A cDNA library for trans-spliced
mRNAs of isolate H95 was constructed as indicated in experimental procedures. The inserts
of 11 recombinant clones which yielded significant BLAST scores were entirely sequenced
and are listed. Indicated are for each clone: the cDNA size (in bp; excluding the polyA-tail
and the 36 nt SL sequence), the length of the predicted ORF (in amino acids), the homology
(in % identical amino acids) of the deduced amino acid sequence to the best matching amino
acid sequence in databases, the predicted identification according to homology, and the
accession numbers of best matching proteins together with their origin. Further shown are the
5’ terminal nt sequences of the cDNAs, potential protein coding regions (bold letters) and the
first amino acids of predicted ORFs (below the nt sequence). The SL addition site is marked
by a slash (/) with the SL 3’ ATG indicated to the left. In clones Em1433 and Em 1431, the 3’
AUG of the SL is predicted to serve as translation start. In clones Em2342 and Em2313, the
SL AUG is in frame with the putative translation start whereas in all others the SL AUG is
out of frame to the predicted ORF.
Fig. 1. Identification of alternative elp transcripts. The chromosomal situation around the 5’
intron of the E. multilocularis elp locus (from Ref. 17) together with the 5’ terminal
sequences of conventionally (transcript A) and trans-spliced (transcript B) elp mRNAs as
21
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

determined by 5’ RACE is shown. Exonic sequences are represented by capitals, intronic and
non-transcribed regions by small letters. Size and location of elp exon I, exon II, and the 5’
intron are indicated by lines and small arrows (<__>) above the nt sequence. Exon-intron
boundaries are shown as double-slashes (//) and the 5’ intron splice donor and splice acceptor
regions are marked above the intron sequence. Sequence and location of the oligonucleotides
10-31 and 10-29 used for mapping the alternative transcriptional start point within the 5’
intron are shown as arrows (---à) below the nt sequence. Alternative 5’ exons of transcript A (exon I)
and B (spliced leader) are underlined and the transition-point to exon II is marked by a dash (-). The deduced
amino acid sequence of the Elp protein N-terminus, which has been confirmed by direct protein sequencing of
immunoprecipitated Elp, is shown below the nt sequence in upper case letters.
Fig. 2. Competitive PCR experiments on alternative elp transcripts in E. multilocularis metacestodes and
protoscoleces. Upon in vitro cultivation of E. multilocularis larvae (19), 50 protoscoleces (P) and 30
metacestode vesicles (mean diameter 1 mm) (M) were manually picked, total RNA was isolated, reverse
transcribed using an oligo-dT primer and the resulting cDNA was diluted to 50 µl. Per PCR reaction, 1 µl of the
diluted cDNA was mixed in a 30 µl final volume with 1x106 (lane1), 5x105 (lane 2), 2,5x105 (lane3), 1,2x105
(lane4), 6x104 (lane 5), and 3x104 (lane 6) molecules of the competitor which was constructed as described in
experimental procedures. For competitive PCR the primer combinations 10-6 x EC5, specific for the exon I-
containing transcript A (A) and 10-6 x ET5, specific for the SL-containing transcript B (B) were added and 30
cycles were applied. 10 µl of each reaction were separated on a 2 % agarose gel and stained with ethidium
bromide. In each lane, the upper band corresponded to the competitor, the lower to the cDNA. Equimolarity
between competitor and cDNA was reached at ~ 1.2x105 molecules for transcript A and ~ 2.5x105 molecules
for transcript B in the protoscolex fraction. For the metacestode fraction, the respective values were ~ 2.5x105
for transcript A and ~ 5x105 for transcript B. Thus, a constant relation of about 1:2 between transcripts A and B
was measurable in both developmental stages.
22
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Fig. 3. Northern blot analysis of transcripts containing the SL. A) Analysis of polyA+ and polyA- fractions. E.
multilocularis was cultivated in Meriones unguiculatus and parasite RNA was isolated. 10 µg total RNA (lanes
1, 4) and equivalent portions of polyA- (lane 2) and polyA+ RNA (lane 3) were separated on a 2 %
formaldehyde gel, blotted, and hybridized with 32P-labeled oligonucleotide probes specific for elp exon I
(probe A; lane 4) or the SL (probe B; lanes 1, 2, 3). Molecular size marker bands are indicated on the left. Probe
A hybridized to a single 2,1 kb fragment. Probe B strongly hybridized to a non-polyadenylated RNA of ~100 nt
and to a smear of polyadenylated transcripts between 500 nt and > 4 kb. B) Analysis of anti-TMG
immunoprecipitated RNA. 5 µg of total RNA (lane 1) and equivalent portions of RNA in the pellet (lane 2) or
the supernatant (lane 3) after immunopreciptation with the anti-TMG cap specific antibody K121 were
separated on a 8 % polyacrylamide/7 M urea gel, transferred, and probed with a 32P-labeled, anti-SL specific
oligonucleotide (probe B). Size markers at the left and right were produced from differently sized PCR products
containing the SL sequence after in vitro transcription using T7 polymerase (29). Probe B strongly hybridized to
a RNA of ~100 nt which was present in the anti-TMG precipitated fraction but absent in the non-TMG
fraction.
Fig. 4. Sequence comparison and structure of echinococcal SL RNAs. A) Comparison of flatworm SL RNA
gene sequences. Aligned are nt sequences from E. multilocularis (Em) and E. granulosus (Eg), determined in
this study, and those from F. hepatica (Fh) and S. mansoni (Sm) taken from (11). Nucleotides identical to that of
the E. multilocularis SL RNA are marked by dots (.) and dashes (-) represent gaps inserted to maximize the
alignment. Further indicated are the length of the SL genes (in bp) and homologies to the E. multilocularis SL
gene for the SL region alone and for the full length SL RNA. B) Possible secondary structure of the E.
multilocularis SL RNA. Indicated are: the splice site by an arrow; the SL in bold letters; the
trimethylguanosine-cap (TMG) by an asterisk. A putative Sm binding site with homology to previously
presented flatworm Sm binding sequences (11) is underlined. The RNA was folded using an algorithm of
Mathews et al. (30) with the constraint that the putative Sm binding sequence is single stranded.
Fig. 5. Mapping of a transcriptional start point within the elp 5’ intron. A) Schematic of the chromosomal
situation around the elp 5’ intron indicating size and location of exons (white boxes) and introns (striped boxes).
23
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Oligonucleotide primers used for mapping the transcriptional start point are indicated by small arrows. B) PCR
and RT-PCR analysis. 2 µg total RNA of the E. multilocularis isolates H95 (lanes 1, 4, 7) and K188 (lanes 2, 5,
8) were RT-PCR amplified in a 30 µl volume employing a sensitive one step RT-PCR system (Titan,
Boehringer, Mannheim, Germany) and using the primer combinations 10-31 x 10-6 (lanes 1, 2), 10-29 x 10-6
(lanes 4, 5), and 10-19 x 10-6 (lanes 7, 8). 10 µl of each reaction were separated on a 2 % agarose gel and
stained with ethidium bromide. In addition, all PCR products were directly sequenced. As controls, the same
primer combinations were used for PCR amplification of chromosomal DNA of isolate H95 (lane 3: 10-31 x
10-6; lane 6: 10-29 x 10-6; lane 9: 10-19 x 10-6). Products completely spliced for introns I, II, and III but still
containing parts of the 5’ intron up to the 10-29 primer binding site were obtained for both isolates (lanes 4, 5,
7, 8). No products were obtained for 10-31 x 10-6 although this primer combination gave a clear positive signal
on chromosomal DNA.
24
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Figure 1
Chromosome <___________Exon I ______________><_donor 5 ’ Intron ___________________________________àgttgcttGCCCGTGGTTGGGCTCCCGTTGATTGCAGTTT//gtgcgt..<171bp>..ttcacttctcgtgaacagtagcctccttgtttgag. Primer „ 10-31“ --------------------à Primer „10-29“ -----------------à
à__5 ’ Intron acceptor _><____________Exon II __________________..<230bp>..cttatattttag//ACTAAAACCATGTTGAAGAGGAGTAAGAA........ M L K R S K N........
Transcript A 5’-GCCCGUGGUUGGGCUCCCGUUGAUUGCAGUUU-ACUAAAACCAUGUUGAAGAGGAGUAAGAA... Exon I M L K R S K N
Transcript B 5’-CACCGUUAAUCGGUCCUUACCUUGCAGUUUUGUAUG-ACUAAAACCAUGUUGAAGAGGAGUAAGAA... spliced leader M L K R S K N
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from

Figure 4
A). . . . .Em CACCGTTAATCGGTCCTTACCTTGCAGTTT--TGTATGGTGAGTATCGAT 48Eg ..........................A........... ............ 48Fh A...T.-..CG.T..TC.G..C..T.TA..AG..C... ..A..A....T. 48Sm A.....-C.CG.T.TTAC-T....TGA...GT..C... ..A..A.C..TC 48
. . . . .Em GCAGC-TGAGGCTGTGCCTACGAGCTGAC---CCAGTATTTGGCTGGTCC 94Eg .......C.......................................... 94Fh .G.C.A.CG.T.CAAA..C.TT.TT..G.TAG..TCC...CT..A.AGG. 98Sm .-.C.AA..AT.GAA.TT--TTCTT..G.AGC..TAC.CAC----..GG. 91
hom. to E. mult.- . length: SL SL-snRNAEm TTCGAGGGCC 104 100% 100%Eg .......... 104 97% 98%Fh .AA..AT-CC 107 55% 34%Sm .A 93 50% 30%
B)
G U U C-G G-C G-C A-U A C G-C U G G U-A U-A C-G C-G C-G G-C U-G U U U A-U G-CU UGUA GGUGAG AUCGAUGC-GACCCAGTATTTGGCUGG-C |||| |||||| ||||||U ACGU CCAUUC UGGCUA G U C A U U GCCAC-*TMG-cap
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from
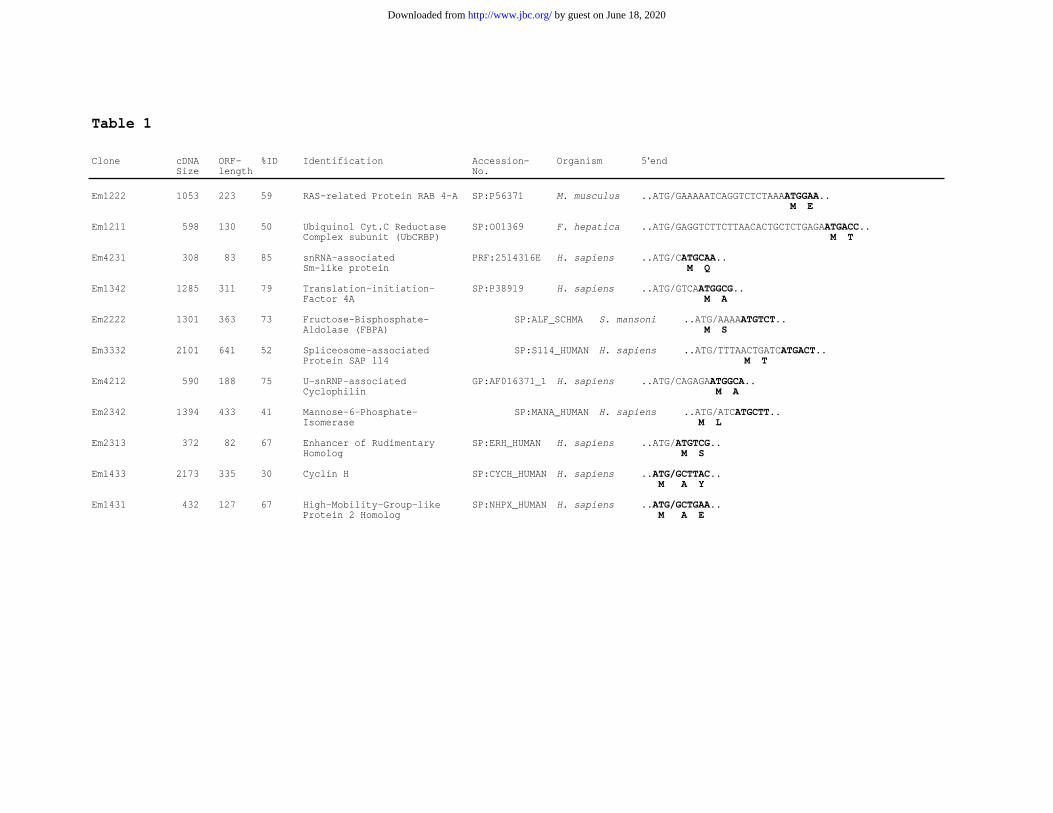
Table 1
Clone cDNA ORF- %ID Identification Accession- Organism 5‘endSize length No.
Em1222 1053 223 59 RAS-related Protein RAB 4-A SP:P56371 M. musculus ..ATG/GAAAAATCAGGTCTCTAAAATGGAA.. M E
Em1211 598 130 50 Ubiquinol Cyt.C Reductase SP:O01369 F. hepatica ..ATG/GAGGTCTTCTTAACACTGCTCTGAGAATGACC..Complex subunit (UbCRBP) M T
Em4231 308 83 85 snRNA-associated PRF:2514316E H. sapiens ..ATG/CATGCAA..Sm-like protein M Q
Em1342 1285 311 79 Translation-initiation- SP:P38919 H. sapiens ..ATG/GTCAATGGCG..Factor 4A M A
Em2222 1301 363 73 Fructose-Bisphosphate- SP:ALF_SCHMA S. mansoni ..ATG/AAAAATGTCT..Aldolase (FBPA) M S
Em3332 2101 641 52 Spliceosome-associated SP:S114_HUMAN H. sapiens ..ATG/TTTAACTGATCATGACT..Protein SAP 114 M T
Em4212 590 188 75 U-snRNP-associated GP:AF016371_1 H. sapiens ..ATG/CAGAGAATGGCA..Cyclophilin M A
Em2342 1394 433 41 Mannose-6-Phosphate- SP:MANA_HUMANH. sapiens ..ATG/ATCATGCTT..Isomerase M L
Em2313 372 82 67 Enhancer of Rudimentary SP:ERH_HUMAN H. sapiens ..ATG/ATGTCG..Homolog M S
Em1433 2173 335 30 Cyclin H SP:CYCH_HUMAN H. sapiens ..ATG/GCTTAC.. M A Y
Em1431 432 127 67 High-Mobility-Group-like SP:NHPX_HUMAN H. sapiens ..ATG/GCTGAA..Protein 2 Homolog M A E
by guest on June 18, 2020http://www.jbc.org/Downloaded from

Klaus Brehm, Katharina Jensen and Matthias FroschmRNA trans-splicing in the human parasitic cestode Echinococcus multilocularis
published online September 5, 2000J. Biol. Chem.
10.1074/jbc.M006091200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 18, 2020http://w
ww
.jbc.org/D
ownloaded from