Embed Size (px)
Citation preview

Osteolyse beim Multiplen Myelom: Untersuchungen am Zellmodell
Markus Bergmann

ii

iii
Osteolyse beim Multiplen Myelom: Untersuchungen am Zellmodell
Von der Medizinischen Fakultät der Rheinisch-Westfälischen Technischen Hochschule Aachen
zur Erlangung des akademischen Grades eines Doktors der Medizin genehmigte Dissertation
vorgelegt von
Markus Bergmann
aus
Dorsten
Berichter: Herr Privatdozent Dr. med. Johann Lorenzen Herr Universitätsprofessor Dr. rer. nat. Wilhelm Jahnen-Dechent Tag der mündlichen Prüfung: 11.Mai 2005 Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfügbar

iv
Die Durchführung der Sequenzierung der Amplifikate bei der Primer-Etablierung wurde freundlicherweise von Frau S. Reuschenberg und Frau I. Losen übernommen. Alle anderen dieser Arbeit zugrunde liegenden Experimente sind nach entsprechender Anleitung durch Herrn Prof. Dr. med. Lorenzen und Mitglieder seiner Forschungsgruppe von mir selbst ausgeführt worden.

v
Danksagung Mein Dank gilt an erster Stelle Herrn Universitätsprofessor Dr. med. J. Lorenzen für die Überlassung des Dissertationsthemas und die intensive Betreuung dieser Arbeit. Das Engagement und die Unterstützung, die er der Arbeitsgruppe und gerade auch den Doktoranden entgegengebracht hat, sind sicher nicht selbstverständlich. Weiterhin bedanke ich mich bei Frau E. Stellbrink für die geduldige Einweisung in die Arbeitstechniken der Zellkultur, der PCR und der Immunfluoreszenzfärbung. Viele weitere Anregungen und nützliche Hinweise habe ich von Herrn Dr. Y. Chen und den anderen Doktoranden der Arbeitsgruppe erhalten. Ein besonderer Dank gilt Frau S. Reuschenberg, die mit wertvollen Tipps und Unterstützung viel zur Arbeitsatmosphäre und zur Motivation beigetragen hat. Besonders möchte ich meiner Partnerin Petra danken, die viel Geduld und Verständnis während des Entstehungsprozesses dieser Arbeit aufbrachte und weit mehr für mich getan hat als zeitaufwendige Korrekturen sprachlicher und inhaltlicher Aspekte. Und natürlich bedanke ich mich bei meinen Eltern ohne deren Unterstützung mein Studium und diese Dissertation niemals möglich gewesen wären.
Juni 2004

vi
Inhalt
Abkürzungen viii
1 Einleitung 1 1.1 Das multiple Myelom 1
1.1.1 Historischer Rückblick 1 1.1.2 Pathologie und Verlauf 2
1.2 Molekularbiologische Erkenntnisse zur Ätiologie 3 1.2.1 B-Zell Entwicklung 3 1.2.2 Erkenntnisse über genetische Ursachen 4
1.3 Interaktionen der Myelomzellen mit dem Mikromilieu des Knochenmarks 5 1.3.1 Knochenstoffwechsel 5 1.3.2 Das OPG/RANK/RANKL System 7 1.3.3 Syndecan 1 10 1.3.4 Einbruch der Myelomzellen in das System 11 1.3.5 Die Rolle von IL-6 13 1.3.6 Die Rolle von IL-15 15
1.4 Fragestellung 16
2 Material und Methoden 17 2.1 Zellkultur 17
2.1.1 Zelllinien 17 2.1.2 Kultivierung 17 2.1.3 Stimulation mit Zytokinen 17 2.1.4 Kokulturen 17 2.1.5 Behandlung mit konditionierten Medien 18
2.2 RNA-Präparation 18 2.3 Semiquantitative Realtime RT-PCR 19
2.3.1 Reverse Transkription 19 2.3.2 Primerdesign und Etablierung 19 2.3.3 Semiquantitative PCR mit dem LightCycler 21 2.3.4 Auswertung der Messdaten mit der LightCycler Data Analysis-Software 23
2.4 Immun-Fuoreszenz-Färbung 26
3 Ergebnisse 28 3.1 Ergebnisse der Primer-Etablierung 28
3.1.1 Beta-Actin 28 3.1.2 Syndecan 1 (SDC1) 30

vii
3.1.3 Osteoprotegerin (OPG) 32 3.1.4 Receptor Activator of NF-κB Ligand (RANKL) 34
3.2 Syndecan 1: Expression des Proteins bei U-266 36 3.3 Syndecan 1: Expression der mRNA bei IM-9 und U-266 37
3.3.1 Vergleich der Nativ-Expression 37 3.3.2 mRNA-Expression bei IM-9 unter Einfluss von IL-6, IL-15 und IL-15AK 37 3.3.3 mRNA-Expression bei U-266 unter Einfluss von IL-6, IL-15 und IL-15AK 38 3.3.4 mRNA-Expression in Kokultur mit Fibroblasten 40 3.3.5 mRNA-Expression unter Einfluss von konditioniertem Medium 40
3.4 Osteoprotegerin: Expression der mRNA bei Fibroblasten 41 3.4.1 mRNA-Expression in Kokultur mit Myelomzellen 41 3.4.2 mRNA-Expression unter Einfluß von konditioniertem Medium 42
3.5 Receptor Activator of NF-κB Ligand 43
4 Diskussion 44 4.1 Untersuchungen zur Expression von OPG 44
4.1.1 Veränderung der OPG-Expression in Kokultur 44 4.1.2 Veränderung der OPG-Expression unter Einfluss des durch Myelomzellen
konditionierten Mediums 45 4.2 Untersuchungen zur Expression von RANKL 47 4.3 Untersuchungen zur Expression von SDC1 49
4.3.1 U-266 bietet polarisiertes Verteilungsmuster von SDC1 50 4.3.2 SDC1-mRNA-Expression ist bei U-266 siebenmal stärker als bei IM-9 50 4.3.3 IL-6, IL-15 oder IL-15AK bewirken keine Veränderung der SDC1-mRNA-
Expression und auch bei Behandlung mit konditioniertem Medium bleibt die SDC1-mRNA-Expression konstant 51
4.3.4 SDC1-mRNA-Expression in Kokultur von U-266 und Fibroblasten 53 4.4 Diskussion der Methode 53
4.4.1 Das Problem Beta-Actin als Housekeeping-Gen 54 4.4.2 Betrachtung der Reproduzierbarkeit 55
4.5 Ausblick auf praktische Anwendung 56 Zusammenfassung 58 Summary 59
5 Literatur 60
Lebenslauf 73

viii
Abkürzungen AS Aminosäure(n) bp Basenpaare CD cluster of differentiation cDNA complemantary DNA CFU-GM granulocyte/macrophage colony forming unit CP crossing point DAPI 4,6-Diaminidin-2-phenylindol-dihydrochlorid DDH death domain homologous DMEM Dulbecco´s modified Eagle´s Medium DNA desoxyribonucleic acid dNTP Desoxynucleotidtriphosphat fas fibroblast associated FITC Fluoresceinisothiocyanat F-Primer Forward-Primer, 5´-Primer GAG Glycosaminoglycan HGF hepatocyte growth factor HSPG Heparansulfatproteoglycan IL Interleukin Jak Janus-Kinase kD Kilodalton LCDA LightCycler Data Analysis MAPK mitogen activated protein kinase M-CSF macrophage colony stimulating factor MGUS monoclonal gammopathy of unknown origin mRNA messenger RNA NCBI National Institute for Biotechnology Information OAF Osteoklasten aktivierender Faktor ODAR osteoclast differentiation and activation receptor →RANK ODF osteoclast differentiation factor →RANKL OPG Osteoprotegerin OPG-L Osteoprotegerin Ligand →RANKL PBS phosphate based saline PCR polymerase chain reaction PG Prostaglandin PKB Proteinkinase B PTH Parathyroidales Hormon RANK receptor activator of nuclear factor κB RANKL receptor activator of nuclear factor κB ligand

ix
RNA ribonucleic acid R-Primer Reverse-Primer, 3´-Primer RT Reverse Transkriptase SDC1 Syndecan 1 SEM standard error of the mean sRANKL soluble RANKL STAT signal transducer and activator of transcription Taq thermus aquatus TNF Tumornekrosefaktor TRANCE TNF-related activation-induced cytokine →RANKL tRNA transfer RNA


1
1 Einleitung
1.1 Das multiple Myelom Das multiple Myelom (Plasmozytom, M. Kahler) ist der häufigste generalisierte Knochen-tumor und geht als B-Zell-Neoplasie von bösartig transformierten Plasmazellen des Knochenmarks aus. Er kann in einem Knochen solitär oder multipel entstehen und als Rarität auch extramedullär auftreten [112]. Es kommt zur Zerstörung des Knochens, Ver-drängung der normalen Blutbildung und Produktion monoklonaler Immunglobuline. Die Inzidenz beträgt in den meisten westlichen Industrienationen etwa 3/100.000. Unterschiede zeigen sich in einem deutlichen Nord-Südgefälle in Europa (Schweden > Rumänien) und in verschiedener Rassenzugehörigkeit (Chinesen < Kaukasier < Afroamerikaner). Betroffen sind überwiegend Männer im 6. und 7. Lebensjahrzehnt (♂:♀ = 3:2). Das mediane Patientenalter bei Diagnosestellung liegt in Mitteleuropa bei 69 Jahren [87, 93].
1.1.1 Historischer Rückblick Im Jahre 1850 beschrieb der Londoner Arzt William MacIntyre den Fall des 44 jährigen Mr. McBean, der an einer von ihm als „mollities et fragilitas ossium“ bezeichneten Krankheit litt [90]. Der Patient klagte über wechselnd heftige Knochenschmerzen, zunehmende Abgeschlagenheit, Gewichtsverlust und Infektanfälligkeit. Im Urin dieses Mannes konnte MacIntyre´s Kollege Henry Bence Jones eine „eigenartige albuminöse Substanz“ - die später nach ihm benannten Proteine - sowie Kalziumphosphat- und Kalziumoxalatkristalle nachweisen [9]. Trotz aller Bemühungen seiner Ärzte starb Mr. McBean Anfang des Jahres 1846. Die makroskopische pathologische Untersuchung zeigte Rippen, Sternum sowie große Teile der Wirbelsäule weich und zerbrechlich und von einer gelatinösen Masse infiltriert [90]. Erst die exakte mikroanatomische Auswertung dieser Befunde an zwei Wirbelkörpern und einer Rippe durch John Dalrymple eröffnete durch den Fund eines bestimmten spezifischen Zellbildes die Möglichkeit der Abgrenzung zu anderen Formen der „mollities ossium“ [34]. In einer Veröffentlichung des Jahres 1873 bezeichnete der Pathologe von Rustizky die Knochentumoren eines unter ähnlichen Symptomen verstorbenen und von ihm autopsierten Mannes erstmalig als multiple Myelome, erkannte jedoch noch nicht die Malignität des Krankheitsbildes [117]. Größere Aufmerksamkeit lenkte Otto Kahler mit seiner Publikation 1889 in der Prager medicinischen Wochenschrift auf das Krankheitsbild [67]. Der Zusammenhang mit Plasmazellen wurde erst 1900 von Wright hergestellt [157].

2
1.1.2 Pathologie und Verlauf Die zu großen Teilen schon von den Erstbeschreibern angeführten Symptome Paraprotein-ämie und -urie, Knochenmarksinfiltration durch Plasmazellen und Osteolysen zählen heute zu den Diagnosekriterien des Plasmozytoms. Die Paraproteinämie entsteht durch die monoklonale Wucherung Immunglobulin-produ-zierender Zellen. Die einheitliche Wanderungsgeschwindigkeit der jeweils identischen Paraproteine in der Serumelektrophorese manifestiert sich als hoher, schmalbasiger Peak, genannt M-Gradient, vorzugsweise im Gammaglobulinbereich, seltener auch im Beta- und Alpha-2-Bereich [87]. Mittels Immunelektrophorese lässt sich der Paraproteintyp näher klassifizieren, wobei die Häufigkeitsverteilung mit 54 % IgG, 25 % IgA und nur 1 % IgD oder IgE [56] in etwa dem normalen Vorkommen der Immunglobuline entspricht. Bei etwa 20 % handelt es sich ausschließlich um nierengängige Leichtketten (κ und λ) den Bence-Jones-Proteinen. In diesen Fällen kommt es zur Paraproteinurie, nicht jedoch zum Auftreten eines M-Gradienten. Die neoplastisch wuchernden Plasmazellen beschränken sich meist auf das Knochenmark, wo sie bevorzugt nodulär zusammenliegen. Das Vorliegen von mehr als 15% Plasmazellen im Knochenmarksausstrich oder Knochenmarksbiopsat gilt als Kriterium für die Diagnose eines Plasmozytoms [105]. Es sind jedoch gerade beim Vorliegen gut differenzierter Zellen benigne monoklonale Gammopathien und reaktive Plasmozytosen durch andere Parameter abzugrenzen. Prognostisch günstig ist ein reifer Zelltyp oder Marschalko-Typ (nach Marschalkos erster genauer Beschreibung der Plasmazelle [92]) sowie ein nichtnoduläres Infiltrationsmuster. Noduläres Befallsmuster und ein blastärer Zelltyp sind assoziiert mit einer schlechten Prognose [105]. Einen weiteren pathognomonischen Befund für das Multiple Myelom stellen die lytischen Skelettdestruktionen dar. Charakteristischerweise handelt es sich um scharfrandig ausge-stanzte Knochendefekte, die keine randständige Knochenneubildung aufweisen. Betroffen sind in erster Linie Brust- und Lendenwirbelsäule, Rippen, Becken, Sakrum und Schädel. Weiterhin können proximale Femora, Klavikeln Schultern und Sternum befallen sein, während distale Anteile der langen Röhrenknochen und kleine Hand- und Fußknochen meist verschont bleiben. Osteolysen der Rippen und Wirbelkörper führen häufig zu pathologischen Frakturen, während Befall der Schädelkalotte sich typischerweise als so genannter „Schrotschussschädel“ zeigt. Der weitaus größte Teil der Patienten (um 65 %) leidet zum Zeitpunkt der Diagnosestellung sowohl unter Osteolysen als auch unter einer allgemeinen Osteopenie, die auf Mikroosteolysen zurückgeführt wird [87, 88, 105]. Neben den drei Kardinalsymptomen kommt es in vielen Fällen zum Immundefektsyndrom (mit 70 % häufigste Todesursache) [87], Hyperviskositätssyndrom [8, 57], Hyperkalzämie-syndrom (terminal 31 %) [78, 87, 88], zur Nephropathie (als zweithäufigste Todesursache

3
und terminal bei über 50 % nachweisbar) [87, 88], Anämie (50 %) [87] und zur Amyloidose (2-7 %) [78, 120]. Die Klassifikation erfolgt anhand der allgemein anerkannten Stadieneinteilung nach Salmon und Durie von 1975 [41]. Als zusätzliche Parameter sind β2-Microglobulin (als Ausdruck der Krankheitsaktivität) und der Plasmazell-Labeling-Index LI% (als Wachs-tumsratenindikator) etabliert [52]. Weitere Parameter sind CRP (korreliert meist mit IL-6) [7], Serumkreatinin und Alter des Patienten.
1.2 Molekularbiologische Erkenntnisse zur Ätiologie Obwohl das multiple Myelom in seiner klinischen Präsentation bereits sehr gut untersucht ist, sind viele Fragen zu seiner Entstehung und seinem Fortschreiten Gegenstand aktueller Forschung. Es scheint inzwischen Einigkeit darin zu bestehen, dass sowohl Veränderungen der Myelomzellen selbst als auch ihre veränderte Interaktion mit dem Knochenmarks-mikromilieu zu Genese und Progression beitragen [10].
1.2.1 B-Zell Entwicklung Die normale Entwicklung der B-Lymphozyten beginnt im Knochenmark. Frühe Pro-B-Zellen haften über ihre CD44-Rezeptoren an Hyaluronat der Knochenmarksstromazellen. Die Bindung des stromalen Stammzellfaktors SCF an ihren c-kit-Rezeptor führt zur Proliferation und zur Ausbildung von IL-7-Rezeptoren. Die Zellen werden zu Prä-B-Lymphozyten und sind in ihrem Wachstum abhängig vom IL-7 der Knochenmarks-stromazellen. Funktionelles Rearrangement der Gene für die schweren (Chr. 14) und leichten Ketten (κ Chr. 2 und λ Chr. 22) der Immunglobuline und strenge Selektion der selbsttoleranten Varianten während dieser Phase erzeugen eine unreife B-Zelle, die funktionsfähiges IgM auf ihrer Oberfläche trägt. Diese verlässt das Knochenmark als naive B-Zelle um bei der primären Immunantwort ihre Differenzierung im sekundären lymphatischen Gewebe fortzusetzen. Produktiver Antigenkontakt führt dabei zur Bildung von Lymphoblasten, die sich anfangs über Plasmablasten zu kurzlebigen Plasmazellen ausbilden und eine Lebensdauer von wenigen Tagen haben. Ein Klassenwechsel von IgM zu IgG ist hierbei möglich. Bei weiterem Antigenkontakt oder in einer sekundären Immun-antwort wandern die Lymphoblasten in die Keimzentren der Lymphknoten, wo die somatische Hypermutation der V-Regionen ihrer IgH- und IgL-Gene und die Selektion der hochaffinen Mutanten stattfindet. Diese differenzieren nach Rückkehr ins Knochenmark zu langlebigen Plasmazellen aus, die mindestens 3 Wochen lang Immunglobuline bilden können [49, 53, 86, 163].

4
1.2.2 Erkenntnisse über genetische Ursachen Um die Vielfalt an Antikörpern zu gewährleisten, sind also mehrere Veränderungen der embryonalen DNA der B-Zelle notwendig, was eine dementsprechend große Gefahr für Fehler darstellt. So konnten in den letzten Jahren von mehreren Forschungsgruppen chromosomale und genetische Abnormitäten bei nahezu allen B-Zell-Neoplasien nach-gewiesen werden. Diese umfassen Deletionen oder Amplifikationen ganzer Chromosomen oder subchromosomaler Abschnitte und, im besonderen, Translokationen. Der Immun-globulin-Genlokus erweist sich dabei als Translokationspartner mit hoher Promiskuität [154]. Entsprechend der Tatsache, dass Myelomzellen phänotypisch reifen B-Zellen gleichen und dass Myelome non-IgM-Immunglobuline exprimieren, betreffen die meisten dort gefundenen Translokationen die S-Regionen der IgH-Loci [76]. So scheint es möglich und sogar logisch, dass das onkogene Ereignis nicht mit der normalen B-Zell-Entwicklung interferiert und erst bei der langlebigen Plasmazelle zutage tritt [53]. Die Translokation t(4;14)(p16;q32) ist bei 25% aller Myelomfälle nachweisbar und führt zur Überexpression von fibroblast growth factor receptor 3 (FGFR3), einem Tyrosin-kinase-Rezeptor, der bei IL-6-abhängigen B-Zellen Apoptosen inhibiert. Es sind Muta-tionen beschrieben, die durch Daueraktivierung zu Stromazell-unabhängigem Wachstum führen. Weiterhin wird MMSET in die Regionen starker Enhancer gebracht [29, 30, 154]. Eine weitere Translokation t(6;14)(p25;q32) ist bei 20 % aller Fälle nachweisbar. Betroffen ist das Gen von interferon regulatory factor 4 (IRF4) einem wichtigen Transkriptionsfaktor in der Entwicklung der Plasmazellen [94]. 25 % der Myelome sind für die Translokation t(14;16)(q32;q23) positiv, betreffend den Transkriptionsfaktor c-MAF. Die Rolle von c-MAF selbst ist noch nicht ganz klar, andere Faktoren seiner Genfamilie sind jedoch in viele zelluläre Prozesse verwickelt, wie Proliferation, Differenzierung und Ansprechbarkeit auf IL-6 [154]. Die Translokation t(11;14)(q13;q32) führt zur Überexpression von cyclin D1, einem Zellzyklus-Cyclin, dass die G1-Phase kontrolliert. In den meisten Fällen von Mantelzell-lymphomen ist das cyclin D1-Gen, ursprünglich als bcl-1 bekannt, in ein IgH J-Segment transloziert, in 25 % der Fälle von multiplem Myelom jedoch stattdessen in die IgH S-Region [28, 154]. Weitere Translokationen betreffen die Gene für MUM2/MUM3 (t(1;14)(q21;q32)) und c-myc (t(8;14)(q24;q32)) [53, 154]. Ebenso wichtig für die Progression der Krankheit scheinen im weiteren Verlauf Mutationen zu sein, die zu unabhängigem Wachstum führen (z. B. ras im IL-6-Signalweg), oder die die Apoptose verhindern (p53 und bcl-2) [53]. Insgesamt wurden in der letzten Zeit vor allem Modelle entworfen, die Entstehung und Progression des multiplen Myeloms als einen mehrschrittigen Transformationsprozess - etwa über MGUS und intramedulläres Myelom zum extramedullären oder multiplen Myelom - beschreiben [5, 53].

5
1.3 Interaktionen der Myelomzellen mit dem Mikromilieu des Knochenmarks
Die wie ausgestanzt wirkenden osteolytischen Skelettveränderungen beim Plasmozytom sind so charakteristisch, dass sie von Paläopathologen selbst an antiken Knochenfunden von bereits 3300 v. Chr. nachgewiesen werden können [137]. Sie werden durch normale Osteoklasten verursacht, die auf eine Vielzahl von Stimuli reagieren, ausgehend von den Myelomzellen selbst oder von weiteren Zellen des Knochenmarkmikromilieus. Zusätzlich fehlt Knochenneuformierung, die normalerweise an Stellen vorausgegangener Knochen-resorption vorhanden ist. Myelomzellen scheinen also durch lokal produzierte Mediatoren einerseits für extensiv gesteigerte Osteoklastenaktivität, andererseits für stark verminderte Knochenneubildung verantwortlich zu sein [23].
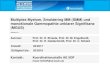
1.3.1 Knochenstoffwechsel Die zwei Komponenten des ständigen Knochenumbaus (Remodeling) - Resorption und Neubildung - sind eng miteinander verwoben. Sie finden immer am gleichen Ort statt, Resorption geht immer der Knochenneuformung voraus, und die Menge an alter, in der Resorptionsphase entfernter Knochensubstanz wird normalerweise in der darauf folgenden Neubildungsphase ersetzt. Diese Prozesse verlangen eine fein abgestimmte Koordination von Osteoblasten und Osteoklasten über all ihre Entwicklungsphasen hinweg [50]. Osteoblasten stammen wie Knochenmarksstromazellen von undifferenzierten Mesenchym-zellen des Knochenmarkes ab, die sich zu Präosteoblasten oder Knochenstammzellen entwickeln. Aus ihnen gehen Osteoblasten und später Osteozyten hervor. Osteoblasten bilden die organische Knochenmatrix, an die anorganische Hartsubstanz angelagert wird. Osteoklasten sind mehrkernige Riesenzellen, die von der Monozyten/Makrophagen-Linie des Knochenmarks abstammen [86]. Zellen aus Kolonie-formenden Einheiten der Granulo-zyten/Makrophagen-Linie (CFU-GM) entwickeln sich unter Einfluss von M-CSF und RANKL (s.u.) der Stromazellen zu unreifen, mehrkernigen Präosteoklasten. Einmal zur Knochenfläche vorgestoßen, differenzieren sie sich durch Aktivierung mit IL-1 und wiederum RANKL zu reifen Osteoklasten, die fähig sind, Knochen abzubauen. Terminiert wird die Knochenresorption durch Apoptoseeinleitung mit TGF-β, therapeutisch auch mit Bisphosphonaten (Abb. 1) [114]. Osteoklasten resorbieren Knochen, indem sie Proteasen abgeben, die die Matrix verdauen, und Säure, die das Knochenmineral herauslöst. Wichtige Proteasen sind MMP-9, eine Matrix-Metalloprotease und Kathepsin K, eine Cystein-Protease. Die abgegebenen Protonen werden durch Carboanhydrase II gebildet und über eine H+-ATPase ähnlich der der Nierentubulusepithelien transportiert (Abb. 2) [114].
6
Die Koordination der Knochen-resorption erfolgt über eine Vielzahl von Hormonen und Zytokinen. In den frühen 90er Jahren stellte sich dabei heraus, dass die meisten dieser Mediatoren, beispielsweise PTH, Vit-D3, IL-1β, TNF, IL-6 oder PGE2, nicht über Rezeptoren der Osteoklasten wirken, sondern über Osteo-blasten und Knochenmarks-stromazellen, die ihrerseits Einfluss auf die Osteoklasten
Abb. 1: Osteoklasten-Entwicklung (mod. n. [70, 114]). Aus Vorläuferzellen entwickeln sich unter Wirkung von M-CSF auf den c-Fms Rezeptor Präosteoklasten, die fusionieren und nach Aktivierung Knochenresorbieren können. Gesteuert wird diese Entwicklung durch die Expression von RANKL durchStromazellen, das auf den Rezeptor RANK der Osteoklasten wirkt. OPG wirkt als Bremse diesesMechanismus, indem es aktives RANKL bindet und so neutralisiert. Weitere Erläuterungen im Text. (OCL=Osteoklasten)
Abb. 2: Knochenresorption durch Osteoklasten (n.[114]). Erläute-rungen im Text.
MMPsCathepsin K
H+
HCO3-Cl-
CA II H+
Cl-
Aktivierung Apoptose
Fusion
unreifer OCL
CFU-GM
reifer OCL
M-CSF
RANKL
sRANKL
TGFβ
Proliferation Differenzierung
Präosteoblast/Stromazelle
c-Fms
RANK
OPG

7
nehmen. Die Ausnahme dieser generellen Regel zeigt Kalzitonin, das direkt über einen Osteoklastenrezeptor wirkt [1]. Der Osteoklasten-aktivierende Faktor der Osteoblasten schien zunächst membrangebunden zu sein [146] und wurde osteoclast differetiation factor (ODF) genannt. Nagai et al. konnten im Jahre 2000 die Existenz einer löslichen Form von ODF nachweisen. Diese wurde von Plattenepithelkarzinomzelllinien, deren Ursprungs-gewebe starke Hyperkalzämien bei den betroffenen Patienten ausgelöst hatten, sezerniert [99]. ODF ist inzwischen in RANKL umbenannt [1].
1.3.2 Das OPG/RANK/RANKL System Während der letzten fünf Jahre konnte ein essentielles Zytokinsystem für die Osteoklasten-Biologie charakterisiert werden. Es besteht aus einem Liganden, receptor activator of NF- κB ligand (RANKL) [80], seinem zellulären Rezeptor (RANK) [65] und einem löslichen Attrappenrezeptor, Osteoprotegerin (OPG) [131]. Während RANKL mehrere Aspekte der Osteoklastenfunktion über RANK stimuliert und so die Knochenresorption steigert, blok-kiert OPG die Bioaktivität von RANKL und verhindert Knochenresorption (Abb. 1) [129].
Osteoprotegerin 1997 forschten Simonet et al. an mit dem TNF-Rezeptor zusammenhängenden Molekülen, die eine potentielle therapeutische Bedeutung haben könnten. Eine der untersuchten cDNAs kodierte dabei für ein dem TNF-Rezeptor ähnliches Protein, das allerdings keine hydrophobe Transmembrandomäne enthielt. Dieser Klon hätte sich nicht bedeutend von anderen interessierenden cDNAs unterschieden, hätten nicht transgene Mäuse, die dieses Gen überexprimierten, einen bemerkenswerten Phänotyp gehabt, denn sie wiesen eine deutliche Osteopetrose auf. Weitere Analysen zeigten, dass dies Folge eines enormen Abfalls der Osteoklasten war, was darauf hinwies, dass OPG eine Schlüsselrolle in der Osteoklastogenese zukam [131]. Fast zeitgleich entdeckten Tsuda et al. bei der Suche nach Stimulatoren und Inhibitoren der Osteoklastogenese einen von embryonalen Fibroblasten gebildeten Inhibitor, den sie OCIF nannten [145]. Wenig später war klar, dass sie das gleiche Molekül wie Simonet et al. identifiziert hatten [161]. OCIF und andere Namen für das gleiche Molekül (TR-1, FDCR-1, TNFRSF-11B) wurden zugunsten der Bezeichnung OPG aufgegeben [1]. Das initiale Protein besteht aus 401 AS, von denen 380 AS nach Abspaltung eines Signalpeptids das reife Protein bilden. N-terminal befinden sich vier cysteinreiche Domänen (D1-D4), während die C-terminale Region zwei zu den „Death-Domains“ des TNF-Rezeptors p55 homologe (DDH) Regionen (D5,D6), eine Heparin-bindende Domäne (D7) und einen Cysteinrest zur Bildung von Homodimeren enthält (Abb. 3) [131, 159, 161]. Die DDH-Regionen sowie die Bildung von Homodimeren scheinen keine Bedeutung für die Wirkung zu haben [159]. Die Heparin-bindende Domäne vermittelt nicht nur die

8
experimentelle Bindung an Heparin, sondern auch die Bindung an das Heparansulfat-proteoglycan Syndecan 1 [15].
OPG-mRNA konnte in vielen Geweben nachgewiesen werden, darunter Lunge, Herz, Niere, Leber, Magen, Darm, Hirn und Rückenmark, Schilddrüse und Knochen [131, 161]. Die Bedeutung von OPG in anderen Geweben ist Gegenstand aktueller Forschung. So entwickelten beispielsweise OPG--Knockout-Mäuse neben starker Osteoporose auch profunde Verkalkungen der großen Arterien und deutliche Intima- und Mediaprolifera-tionen [20].
Receptor Activator of NF-κB Ligand Kurz nach der Entdeckung von OPG identifizierten beide Gruppen auch den passenden Liganden, den sie vorerst OPG-L/ODF nannten [80, 162]. Es stellte sich bald heraus, dass dieser Ligand identisch war mit zwei bereits bekannten Mitgliedern der TNF-Ligand Familie, mit TRANCE und RANKL [3, 156]. OPG-L, ODF, TRANCE sowie SOFA und TNFSF-11A sind inzwischen der Bezeichnung RANKL gewichen [1]. Die membranständige Form von RANKL ist ein 317 AS langes Peptid, das sich zu einem Typ II Transmembranprotein faltet. Es besitzt kein Signalpeptid und N-terminal liegt die zytoplasmatische Domäne. C-terminal befindet sich die extrazelluläre Domäne, die die
Abb. 3: Strukturen von OPG, RANK und RANKL vom N- zum C-Terminus (n. [61, 138]. Weitere Erläuterungen im Text. (EC=extrazellulärer Anteil, IC=intrazellulärer Anteil, TM=trans-membraner Anteil, SP=Signalpeptid, HB=Heparin-bindende Region, Cys400=Cysteinrest zur Bildung von Homodimeren)
1 401
Cys400
SP Cys-reiche Domänen DDH-Regionen HB
22 198 N C
OPG
Cys-reiche Domänen
1
SP
TM
616 29 213 234
EC IC
N C
RANK
TM
3171
Domäne homolog zur TNF Liganden Familie
EC IC
70 148 49 N C
RANKL

9
aktive Liganden-Region mit 10 β-Faltblatt-bildenden Sequenzen enthält (Abb. 3). Diese Struktur ist bei allen Mitgliedern der TNF Liganden Familie anzutreffen [61]. Über Ursprung der löslichen Form sRANKL besteht gewisse Uneinigkeit. Nakashima et al. wiesen die Protein-Expression zweier unterschiedlich großer Formen von RANKL bei murinen Stromazelllinien mit einem Antikörper gegen die C-terminale Sequenz von RANKL nach. Die Expression der kürzeren Form ließ sich durch den Metallopro-teaseninhibitor KB-R8301 hemmen, während die membranständige Form daraufhin über-exprimiert wurde. Die N-terminale Sequenzierung der löslichen Form ließ den Schluß zu, dass die kürzere Variante durch posttranslationale Spaltung vor dem Phenylalanin an Position 139 durch eine Metalloprotease zustande kommt [100]. Im Gegensatz dazu beschrieben Nagai et al. kurz zuvor die Entdeckung einer cDNA, die potentiell für ein RANKL-homologes Protein kodierte, dem allerdings die ersten 73 AS mit der intra-zellulären und der transmembranen Domäne fehlen würden. Dieses Protein konnte mit Antikörpern gegen RANKL in durch Plattenepithelkarzinomzelllinen konditioniertem Medium nachgewiesen werden. Auch die Osteoklastogenese-induzierende Aktivität konnte gezeigt werden. Das Fehlen eines Signalpeptids bei beiden Formen weist darauf hin, dass sowohl RANKL als auch sRANKL nicht den normalen Sekretionspfad über endo-plasmatisches Retikulum und Golgiapparat gehen [99]. RANKL-mRNA wird in hohem Maße in Knochen und Knochenmark und in Lymph-gewebe (Lymphknoten, Thymus, Milz, fetaler Leber und Peyer-Plaques) exprimiert [3, 80, 156, 162]. RANKL ist wichtiger Regulator der Osteoklastogenese, der Lymphozyten-entwicklung und der Organogenese der Lymphknoten. RANKL-negative Knockout-Mäuse entwickelten durch völliges Fehlen von Osteoklasten starke Osteopetrose und Defekte in der frühen T- und B-Zell-Entwicklung. Ihnen fehlte außerdem die Entwicklung von Lymphknoten bei normaler Milzentwicklung [73].
Receptor Activator of NF-κB Mit der Entdeckung, dass der Ligand für OPG identisch war mit TRANCE/RANKL, war das fehlende Teil des Puzzles so gut wie gefunden. Denn schon 1997 hatten Anderson et al. RANK als einen stimulierende Einflüsse übertragenden Rezeptor auf T-Zellen und dendritischen Zellen identifiziert [3]. Hsu et al. zeigten, dass RANK-mRNA von osteo-klastischen Vorläufern und von reifen Osteoklasten stark exprimiert wurde und nannten den Rezeptor deshalb ODAR (osteoclast activation and differentiation receptor) [65]. Im Allgemeinen wurde jedoch die Bezeichnung RANK übernommen [1]. Das humane RANK ist ein 616 AS langes Protein mit einem 28 AS langen Signalpeptid, einer N-terminalen extrazellulären Domäne, die aus vier cysteinreichen Regionen besteht, einer kurzen Transmembrandomäne von 21 AS und einer langen C-teminalen cytoplas-matischen Domäne (Abb. 3) [3].

10
Vor allem Zellen der Monozyten/Makrophagen-Linie exprimieren RANK, darunter Präosteoklasten, T- und B-Zellen, dendritische Zellen und Fibroblasten [3, 65]. Knockout-Mäuse, denen RANK oder eine Komponente seines Signalweges, beispielsweise TRAF-6 oder NF-κB, fehlte, entwickelten Osteopetrose aufgrund der Abwesenheit von Osteoklasten. Außerdem fehlten RANK-Knockout-Mäusen die peripheren Lymphknoten und ihre B- und T-Zell-Reifung war defekt [81].
1.3.3 Syndecan 1 Syndecan 1 (CD138) ist Mitglied einer Familie von 4 Heparansulfatproteoglycanen (HSPG) bei Säugetieren. HSPG sind Proteine, die kovalent an sulfatierte Glycosaminoglycan-Ketten (GAG) gebunden sind. Strukturell mit dem gerinnungs-hemmenden Heparin verwandt, bestehen diese Ketten aus sich abwechselnden Einheiten von Glucuronsäure und N-Acetylglucosamin [11, 84]. Das Kernprotein von Syndecan 1 besteht aus 310 AS. Der C-terminalen zytoplas-matischen Domäne folgt eine einzelne Trans-membrandomäne und eine N-terminale Ekto-domäne. Letztere besteht aus dem Abschnitt, an dem die GAGs gebunden sind, und einer Region, die, sicher zumindest bei Syndecan 4, wichtig für Zelladhäsion ist (Abb. 4) [109]. Interessan-terweise kann die gesamte Ektodomäne des Syn-decan 1 von Myelomzellen in vivo als auch in vitro nach extrazellulär abgegeben werden [39, 111], was dazu führt, dass es eine membranstän-dige und eine lösliche Form gibt. Der Mechanis-mus hierfür ist noch nicht hinreichend erklärt, vieles spricht aber dafür, dass die Ektodo-mäne nachträglich durch eine Metalloprotease vom non-Matrix-Typ abgespalten wird [63]. HSPG finden sich in zahlreichen Geweben als extrazelluläre Matrixkomponenten und membrangebundene Proteine. Es wird allgemein angenommen, dass sie bei wichtigen biologischen Prozessen als Gerüststrukturen andere Proteinen durch nichtkovalente Bindung an ihren GAG-Ketten aufnehmen und so auf die Zelladhäsion, Zellmigration und auch auf die Wirkung von Wachstumsfaktoren und Zytokinen Einfluss nehmen. SDC1 selbst wird auf vielen Epithelien exprimiert, wo es zur Zelladhäsion und epithelialen
Abb. 4: Struktur von SDC1 (n. [109]). (EC=extrazellulärer Anteil, IC=intrazellul-ärer Anteil, TM=Transmembrandomäne).
Typspezifisch
Heparan Sulfat Bindungs-Stellen
Zellbindende Domäne (SDC4)
TM
EC
IC
C
N
310
306
286
276

11
Morphogenese beiträgt [11]. Im Immunsystem wird es vor allem von Zellen der B-Zell-Reihe, so von Prä-B-Zellen und Plasmazellen, und einigen ihrer Lymphome, darunter B-CLL, lymphoplasmazytoides Lymphom und Plasmozytom, exprimiert [121]. Im Jahre 2000 zeigten Børset et al., dass die Expression von Syndecan 1 auf humanen Myelomzelllinien nicht immer gleichmäßig, sondern teilweise polarisiert erfolgt [15]. Dabei war der Anteil der polarisierten Zellen, die auf der einen Seite Lamellopodien, auf der anderen Seite so genannte Uropodien ausbildeten, bei unterschiedlichen Zelllinien verschieden groß. Außer von SDC1 war auch die Expression anderer Adhäsionsmoleküle, wie ICAM-1 (CD54) und CD44 klar auf diese Uropodien ausgerichtet. Es fanden sich Hinweise, dass die Uropodien eine wichtige Rolle in der aktiven Migration, der Zell-Matrix-Adhäsion und der Zell-Zell-Kommunikation spielen. In Bezug auf letzteres war SDC1 vor allem an Zellkontaktstellen zu finden. Auch wurde gezeigt, dass SDC1 in der Lage ist, OPG und HGF (hepatocyte growth factor) an den Uropodien zu binden. Die Wirkung von HGF auf andere Zellen kann durch lösliches SDC1 abgeschwächt werden [123], während die membranständige Form den Einfluss auf die Myelomzellen selbst verstärkt [37]. Das von Myelomzellen stark exprimierte SDC1 greift so in wichtige Prozesse bei der Krankheitsprogression und Knochendestruktion ein.
1.3.4 Einbruch der Myelomzellen in das System Die Fähigkeit humaner Myelomzellen an endotheliale Zellen im Knochenmarksmikro-milieu zu binden, lässt darauf schließen, dass die Interaktion dieser beiden Zelltypen von entscheidender Bedeutung für den als „Homing“ bezeichneten Prozess der Tumorzell-migration in das Knochenmark sein könnte. Dazu exprimieren Myelomzellen einen hoch-affinen Rezeptor, der an Laminin-1, Hauptkomponente der Basalmembran, bindet. Direkter Kontakt zu endothelialen Zellen verstärkt dessen Expression, und die Bindung von Laminin-1 stimuliert die Migration von Myelomzellen weiter [149]. Außerdem ex-primieren Knochenmarksstromazellen die Chemokine MCP-1, -2 und -3, die über den Chemokinrezeptor CCR2 auf Myelomzellen deren Migrationsfähigkeit nochmals herauf-setzen [19]. Die Adhäsion der malignen Zellen an das lokale Knochenmarksgewebe hat eine verstärkte Expression und Abgabe von für die Tumorzellen überlebenswichtigen Zytokinen zur Folge, die damit einen wichtigen Stimulus für die Ansammlung der Tumor-zellen im Knochenmark darstellt. TNFα erhöht dabei über Aktivierung von NF-κB die Expression von Adhäsionsmolekülen, die die Myelomzellen zur Bindung an die Knochen-marksstromazellen brauchen [115]. Die Bindung von β1-Integrin der Tumorzellen an Fibronectin der Stromazellen und die extrazelluläre Matrix scheint für die Myelomzellen zu entscheidenden Überlebensvorteilen zu führen, die zu verstärktem Tumorwachstum

12
führen. Diese Überlebensvorteile beinhalten unter anderem Resistenz gegenüber Chemo-therapeutika, Dysregulation der fas-induzierten Apoptose und Verstärkung der IL-6 Freisetzung [115]. Weitere das Wachstum unterstützende Mechanismen scheinen die Abgabe von neovaskularisierenden Faktoren wie VEGFs, FGF und Angiopoetin 1 sein. Gerade die mit dem Myelom assoziierte Osteolyse ist Thema vieler aktueller Arbeiten. Für den Knochenabbau verantwortliche Osteoklasten sammeln sich in der Nähe von Plasmazellnestern. Deren Stimulation scheint demnach lokal stattzufinden. Zellkulturen humaner Myelomzellen produzieren eine ganze Reihe Osteoklasten-aktivierender Faktoren (OAFs), darunter TNFβ, IL-1β, IL-6, PTHrP, HGF, TNFα und Metalloproteasen wie MMP9, MMP2 oder MMP1. Diese scheinen aber nicht allein für die Knochenzerstörung in vivo verantwortlich zu sein, denn keiner dieser Faktoren ist bei einer Mehrheit der Patienten in hohen Konzentrationen nachweisbar oder korreliert mit deren Krankheits-aktivität [23]. Nach neuesten Erkenntnissen werden die stimulierenden Effekte über das RANK/RANKL-System übertragen. Die Herkunft von RANKL dabei ist Gegenstand aktueller Auseinandersetzungen. Sezer et al. und andere wiesen die direkte Expression von RANKL auf Myelomzellen nach [55, 127], andere Gruppen wie Giuliani et al. bestätigen diese Ergebnisse jedoch nicht [45]. Sie halten nach wie vor die Induktion der Expression von RANKL durch Myelomzellen in anderen Zellen des Mikromilieus für entscheidend. OPG stellt als neutralisierender Atrappenrezeptor den natürlichen Gegenspieler von RANKL dar. Terpos et al. zeigten erst kürzlich die Korrelation des Verhältnisses von sRANKL zu OPG zur Überlebenswahrscheinlichkeit und zum Ausmaß der Osteolysen [140]. OPG wird als Heparin-bindendes Protein von dem membranständigen HSPG Syndecan 1 der Myelomzellen gebunden [15], was die RANKL/OPG-Ratio zusätzlich in Richtung Osteolyse verschiebt. Interessanterweise scheinen gerade die durch ein Überangebot an RANKL und einen Mangel an OPG rekrutierten Osteoklasten einen weiteren bedeutenden Überlebensfaktor für die Tumorzellen darzustellen. Yaccoby et al. behandelten Mäuse des SCID-hu Myelommodells, ein Modell, bei dem immunodefizienten Mäusen humane Myelomzellen und humane Knochenmarksstromazellen injiziert werden, mit Inhibitoren der Osteoklastenaktivität, darunter die Bisphosphonate Pamidronat und Zoledronat und der spezifische RANKL-Inhibitor RANK-Fc. Bei Mäusen mit intramedullärem Myelom wurde daraufhin nicht nur die Osteoklastenaktivität gehemmt, sondern auch das Tumorwachtum deutlich eingeschränkt. Bei Mäusen mit extramedullärem Myelom blieb das Tumor-wachstum unbeeinflusst [158]. Die erhöhte osteoklastische Aktivität führt beim Myelom des Knochenmarks zu einer Freisetzung von IL-6, TGFβ, FGFs und IGFs. Diese Zytokine stimulieren direkt oder indirekt weiteres Tumorwachstum und schließen so einen Teufelskreis, in dem Myelomzellen die Knochenresorption stimulieren und Knochen-resorption zu gesteigertem Tumorwachtum führt (Abb. 5) [144].

13
1.3.5 Die Rolle von IL-6 Hirano et al. gelang es 1986 erstmals die cDNA des bis dahin BSF2 (B-Zell-stimulierender-Faktor 2) genannten Zytokins zu klonieren [58]. 1987 konnten van Damme et al. zeigen, dass der kurz zuvor beschriebene, von Makrophagen gebildete und für Überleben und Proliferation von Plasmozytomzellen benötigte Faktor [103] identisch ist mit BSF2 [148]. Bald darauf wurde dieser Faktor IL-6 genannt. Das 26 kD schwere humane IL-6 Protein besteht aus 184 AS und ist der wichtigste Wachstumsfaktor für das multiple Myelom beim Menschen [71]. Zahlreiche Erkenntnisse unterstreichen die Rolle von IL-6 bei dieser Krankheit. So wurde gezeigt, dass die Serum-spiegel von IL-6 und IL-6R bei Myelompatienten umgekehrt mit ihrer Überlebensrate korrelieren [79, 89], dass Therapie mit monoklonalen Anti-IL-6-Antikörpern die Krank-heitsmanifestationen vorübergehend aufhalten kann [6] und dass die Proliferation von Myelomzellen in vitro durch Antikörper gegen IL-6 oder seinen zellulären Rezeptor gehemmt wird [135, 153].
Abb. 5: Knochendestruktion beim Multiplen Myelom (mod. n. [144]). Über lokale Mechanismen wie Zellkontakt oder Freisetzung von OAFs stimulieren Tumorzellen die Knochenmarksstromazellen zur Produktion von RANKL und sRANKL. Der natürliche Gegenspieler von RANKL, der Atrappenrezeptor OPG, wird durch Bindung an SDC1 auf den Myelomzellen inaktiviert, was die Wirkung von RANKL zusätzlich verstärkt. OCL-Vorläufer differenzieren unter RANKL zu reifen OCL, deren Aktivität wiederum Zytokine freisetzt, die das Tumorwachstum vorantreiben. (OAF=Osteoklasten aktivierende Faktoren, OCL=Osteo-klasten)
RANKL
sRANKL
RANK
OPG
SDC1 OAFs
TGFβ FGF1&2 IGF-I&II IL-6
Myelomzellen
OCL-Vorläufer
reife OCL
Stromazellen

14
Die zelluläre Herkunft von IL-6 beim Plasmozytom wird kontrovers diskutiert. Einige Autoren zeigen eine Produktion des Zytokins durch die Myelomzellen selbst [68], während andere auf die parakrine Produktion durch Knochenmarksstromazellen hinweisen [72, 141]. IL-6 ist für Myelomzellen nicht nur ein proliferativer, sondern auch ein antiapoptotischer Faktor, denn es verhindert spontane [82], durch Dexamethason induzierte [54] und fas-induzierte [27] Apoptosen. Diese zwei Effekte, der proliferative und der antiapoptotische, basieren dabei auf unterschiedlichen Signalwegen [134]. Interaktion des IL-6 mit der α-Kette des Rezeptors (gp80) induziert die Formierung multimerer Komplexe, die aus zwei α-Ketten, zwei β-Ketten (gp130) und zwei IL-6-Molekülen bestehen. Durch Aktivierung von Tyrosinkinasen (JAKs und src-Kinasen) werden einerseits Signalwege via STAT1 und STAT3 eingeschaltet, andererseits der ras/raf/MAPK-Pfad aktiviert [53]. Aktivierung des ras/raf/MAPK-Signalweges induziert Zellwachstum, während der STAT3-Weg anti-apoptotische Effekte über Aktivierung des Bcl-xL-Gens ausübt [26, 43]. Erst 1999 zeigten Catlett-Falcone et al. die konstitutionelle Aktivierung des STAT3-Weges bei den meisten ihrer von Myelompatienten isolierten Zellen sowie bei der auch in dieser Arbeit verwende-ten Zelllinie U-266. Letztere trägt eine autokrine IL-6-Schleife und ist abhängig von der eigenen IL-6-Produktion. Im Kontrast dazu zeigte die von IL-6 unabhängige Myelomzell-linie RPMI 8226, die eine Mutation des ras-Signalweges aufweist, folgerichtig keine STAT-Aktivierung, jedoch entsprechend Aktivierung anderer Signalwege. Der Aktivie-rungsgrad des STAT-Weges bei U-266 war durch die autokrine IL-6-Schleife bei weitem nicht ausgereizt. So konnte die STAT3-DNA-Bindung durch Zugabe von exogenem IL-6 noch stark erhöht werden [26]. Die lymphoblastoide Zelllinie IM-9 scheint keinen autokrinen IL-6-Loop zu zeigen [153], weist jedoch die Expression des IL-6-Rezeptors auf (unveröffentlichte Daten unserer Arbeitsgruppe). Die Auswirkungen des Zytokins auf den Knochenstoffwechsel sind weit weniger klar beschrieben. IL-6 stimuliert die Formierung humaner Osteoklasten in vitro [77] und in vivo [14], und es verstärkt die Wirkungen anderer osteoklastogenetischer Faktoren wie PTH und PTHrP [36]. IL-6 beeinflusst auch die Effekte anderer inflammatorischer Zyto-kine, wie IL-1 und TNFα, auf Osteoklasten-Formierung [38]. Die Korrelation zwischen hohen IL-6-Serumspiegeln und dem Vorhandensein von Osteolysen jedoch wird nach wie vor kontrovers diskutiert. Sati et al. fanden keinen Zusammenhang zwischen IL-6-Expression und den klinischen Erscheinungen des multiplen Myeloms wie Osteolysen oder Hyperkalzämie [119]. So spielt IL-6 für Myelomzellen mit Sicherheit eine bedeutende Rolle als wachstums-fördernder und antiapoptotischer Faktor, seine Funktion als Osteoklasten-aktivierender Faktor und die Mechanismen seiner dahingehenden Wirkung bleiben aber weiter unklar.

15
1.3.6 Die Rolle von IL-15 Aufgrund seiner Eigenschaft, die Proliferation der IL-2 abhängigen T-Zelllinie CTLL-2 in Anwesenheit von neutralisierenden Anti-IL-2-Antikörpern zu stimulieren, wurde IL-15 1994 von zwei unabhängigen Forschungsgruppen identifiziert [22, 51]. Durch seine Fähigkeit, den wachstumsstimulierenden Effekt von IL-2 auf T-Zellen nachzuahmen, wurde der funktionelle und biochemische Zusammenhang dieser beiden Zytokine schnell untersucht. Es zeigte sich, dass ihre Rezeptoren die selben βγ-Untereinheiten benutzen, und dass diese Gemeinsamkeit zu einem gleichen Effekt in vitro geführt hatte, dass ihre Funktion in vivo jedoch, bestimmt durch die Verschiedenheit ihrer α-Ketten und deren ungleiche Verteilung, eine unterschiedliche ist [42]. Das 14 bis 15 kD schwere, 114 AS lange, reife IL-15-Protein wird von 4 Exons des IL-15-Gens in Bande q31 auf Chromosom 4 kodiert [51, 75]. Es enthält zwei Disulfidbrücken, und zwar zwischen Cys42 und Cys88 und zwischen Cys 35 und Cys 85. Als Tertiärstruktur wird eine 4α-Helix Struktur angenommen [107], sehr ähnlich der des IL-2. Weil IL-2 und IL-15 Signalkomponenten teilen (IL-2/15RβγC), weist vieles heute darauf hin, dass die Interaktion von IL-15 mit seinem Rezeptorkomplex in mehreren Zellen zu einer Signalkas-kade führt, die ähnlich, wenn nicht gar identisch der von IL-2 ist [139]. Diese umfasst die Aktivierung des Janus-Kinase (Jak)/signal-transducer-and-activator-of-transcription (STAT)-Pfades [66]. Die β-Einheit des Rezeptors führt bei IL-15-Bindung zur Phosphory-lierung von STAT3 über Jak1, die γC-Einheit zu der von STAT5 via Jak3 [83, 95]. Weitere Signalwege umfassen Induktion von bcl-2 und Stimulation des ras/raf/MAPK-Weges. Der letztere wie auch der über Jak1/STAT3 werden auch von IL-6 wahrgenommen. IL-15 wird von vielen Zelltypen exprimiert, darunter Monozyten/Makrophagen [25, 40] und Knochenmarksstromazellen [97]. Aktuelle Ergebnisse unterstreichen vor allem die Bedeutung von IL-15 bei Proliferation, Reifung und Überleben von Natural-Killer(NK)-Zellen [24, 42] sowie bei Wachstum und Stimulation von T-Zellen [51]. Des Weiteren konnten proliferative und differenzierende Effekte auf B-Zellen nachgewiesen werden [4]. Bulfone Paus et al. konnten zeigen, dass IL-15 Anti-CD3-, Anti-fas-, Anti-IgM- oder Dexamethason-induzierte Apoptosen bei aktivierten T-Zellen und B-Zellen verhindern kann [21]. Im Jahr 2000 wiesen Tinhofer et al. die Existenz einer autokrinen IL-15-Schleife bei Myelomzellen nach [142], ähnlich der von Kawano et al. schon 1988 vermuteten Schleife für IL-6 [68]. Sie zeigten, dass die Expression aller IL-15-Rezeptor-Untereinheiten in sämtlichen ihrer 6 Myelomzelllinien und in Plasmazellen aller ihrer 14 Patienten mit multiplem Myelom vorhanden war. Insbesondere konnte der Nachweis der IL-15-spezifischen α-Untereinheit gelingen, und die Expression derselben spezifisch für IL-2 ausgeschlossen werden. Sie wiesen auch IL-15-mRNA in allen 6 Zelllinien nach und konnten zeigen, dass Unterbrechung der postulierten autokrinen Zyklen für IL-15 oder IL-6 die Zahl an spontanen Apoptosen erhöhte, sowie die Abhängigkeit der Zellen für das

16
jeweils andere Zytokin verstärkte. Zugesetztes IL-15 reduzierte die Zahl spontaner Apop-tosen und schützte, wie teilweise bereits erwähnt, die Zellen zu großen Teilen vor fas-in-duzierter Apoptose und der zytotoxischen Wirkung von Vincristin und Doxorubicin. Diese Daten ergänzen so die Liste der für Myelomzellen wachstums- und überlebensfördernden Zytokine um das IL-15 und zeigen, dass IL-15 in autokriner Weise aktiv sein kann, ein Grund, in dieser Arbeit die Auswirkung der Unterbrechung oder Verstärkung der auto-krinen Schleife auf die Expression wichtiger Merkmale der Myelomzellen zu untersuchen.
1.4 Fragestellung Der Befall des Knochens beim multiplen Myelom, der zu starken Schmerzen und Spontan-frakturen führt, ist sicherlich einer der augenscheinlichsten Aspekte der Krankheit. Erst in den letzten Jahren gelang es, die biochemischen Grundlagen der Osteolysen in Grundzügen zu verstehen. Es eröffnete sich der Blick auf ein Netzwerk aus zahlreichen gegenseitigen Einflussnahmen zwischen Zellen des Knochens, Stromazellen und Myelomzellen. Ziel dieser Arbeit ist es, eine Methode zur Untersuchung einiger Elemente dieses Netzwerkes zu etablieren und mit Hilfe dieser Methode weiteren Fragen auf den Grund zu gehen. Als vereinfachtes Zellmodell sollen dabei Zellen einer humanen Myelomzelllinie und, als direkte Verwandte der Knochenmarksstromazellen, primäre Fibroblasten dienen. Im Speziellen wird weiter untersucht: 1. Lässt sich das polarisierte Verteilungsmuster von SDC1 auf den Myelomzellen
bestätigen? 2. Wie verändert sich die Expression von SDC1 bei Myelomzellen als Inhibitor von OPG
unter folgenden Bedingungen: • Stimulation mit IL-6 • Verstärkung oder Unterbrechung der autokrinen Schleife mit IL-15 oder Antikörpern
gegen IL-15 • Kultivierung in Kokultur mit Fibroblasten • Kultivierung in durch Fibroblasten konditioniertem Medium
3. Welche Veränderungen zeigt die Expression von OPG durch Fibroblasten in Kokultur
mit Myelomzellen oder unter dem Einfluss von durch Myelomzellen konditioniertem Medium?
4. Lässt sich der Osteoklasten-aktivierende Faktor RANKL auf einer der Zelllinien
nachweisen?

17
2 Material und Methoden
2.1 Zellkultur
2.1.1 Zelllinien Die Myelomzelllinie U-266 [102] sowie die lymphoblastoide Zelllinie IM-9 [147] stammen aus der Deutschen Sammlung von Mikroorganismen und Zellkulturen (DSMZ, Braunschweig, BRD). Bei den Fibroblasten handelt es sich um humane primäre dermale Fibroblasten, isoliert von einem Präputium-Resektat. Diese Zellen wurden uns freundlicherweise von Frau PD Dr. Anja Bosserhoff überlassen.
2.1.2 Kultivierung Die Zelllinie IM-9 wurde in RPMI 1640 Medium (PAA, Cölbe, BRD), die Zelllinie U-266 und die Fibroblasten in DMEM Medium (PAA, Cölbe, BRD) kultiviert. Zusätzlich enthielt das Medium jeweils 10% fetales Kälberserum (PAA, Cölbe, BRD) sowie 10 U/ml Penicillin und 10 µg/ml Streptomycin (Biochrom, Berlin, BRD). Die Kulturen wuchsen in Falcon-Flaschen (Becton-Dickinson, Le Pont de Claix, Frankreich) à 25 bzw. 75 ml in einem Heraeus-Brutschrank bei 37 °C in einer mit Wasserdampf gesättigten Atmosphäre und bei 5 % CO2 .
2.1.3 Stimulation mit Zytokinen Zur Stimulation eingesetzte Zytokine waren rekombinantes humanes Interleukin 6 (Upstate, Lake Placid, USA) in einer Konzentration von 0,24 ng/ml, rekombinantes humanes Interleukin 15 (R&D, Wiesbaden, BRD) in einer Konzentration von 6 ng/ml und murine Antikörper gegen humanes IL-15 (R&D, Wiesbaden, BRD) in einer Konzentration von 600 ng/ml. Die Konzentrationen entsprechen jeweils der dreifachen ED50 bzw. ND50 (50 %-Effektivdosis bzw. 50 %-Neutralisationsdosis).
2.1.4 Kokulturen Dazu wurden Fibroblasten und Zellen der Zelllinie U-266 jeweils in Medium suspendiert, ausgezählt und bis auf gleiche Konzentration verdünnt. In je 5 ml DMEM-Medium in Kulturflaschen à 25 ml zu gleichen Anteilen zusammengebracht, wuchsen sie unter normalen Kulturbedingungen weiter. Dem Vergleich dienten zwei weitere Kulturflaschen mit Fibroblasten sowie mit Zellen der Zelllinie U-266 gleicher Ausgangszellzahl. Aufgrund fehlender Trennbarkeit der Zellen, sowie unterschiedlicher Wachstumsge-schwindigkeiten, hatten diese Experimente nur wegweisenden Charakter.

18
2.1.5 Behandlung mit konditionierten Medien Jeweils 30 ml Medium (DMEM) wurden in mit U-266 bzw. Fibroblasten voll bewachsenen 75-ml-Kulturflaschen für 24 bzw. 48 h konditioniert. Zentrifugieren (400 xg, 5 min) und Dekantieren trennte das Medium von den Zellen. Anschließend wurde es mit Minisart Spritzenfiltern (Sartorius, Göttingen, BRD) mit einer Porengröße von 0,2 µm steril filtriert und bis zum weiteren Gebrauch bei –20 ºC eingefroren. Die Behandlung der in ihrer Expression zu untersuchenden Zellen mit dem so konditionierten Medium erfolgte unter normalen Kulturbedingungen in Flaschen à 25 ml. Zur Gewinnung der unbehandelten Zellen (Vergleichswert) wurden gleiche Zellen mit frischem Medium für 24 h weiter kultiviert.
2.2 RNA-Präparation Die Aufreinigung der zellulären Gesamt-RNA erfolgte mit dem RNeasy Mini Kit der Firma Qiagen (Qiagen, Hilden, BRD). Eine spezielle Silicat-Gel-Membran bindet dabei unter hohen Salzkonzentrationen RNA, die länger als 200 bp ist. Zelluläre Proteine, DNA, kürzere RNA (z.B. tRNA) und sonstige im Lysat enthaltenen Bestandteile werden mit Waschpuffern eluiert, bis nur noch die RNA in der Membran gebunden ist und mit Wasser herausgelöst werden kann. Die Membran sitzt in einer kleinen Säule mit austauschbarem Auffanggefäß; Das Lysat sowie die Waschpuffer werden mittels Zentrifugation hindurch befördert. Die interessierenden Zellen wurden dazu in Medium suspendiert, adhärente Zellen trypsiniert und dann suspendiert. Die Zellen wurden gezählt und durch Zentrifugation vom Medium getrennt. Es folgte eine Resuspension in PBS (Gibco Life Technologies, Karlsruhe, BRD) um Mediumreste auszuwaschen. Erneutes Zentrifugieren pelletierte die Zellen, und der Überstand wurde verworfen. Je nach Zellmenge wurde das Pellet in 350 oder 600 µl Lysepuffer RLT + β-Mercaptoethanol (M6250 Sigma, Deisenhohen, BRD; 14,5 M, 10 µl auf 1 ml Lysepuffer) aufgenommen und homogenisiert. Der Lysepuffer RLT enthält stark denaturierendes Guanidin-Isothiocyanat (GITC), durch welches zelluläre RNasen sofort inaktiviert werden. Nun wurden 350 oder 600 µl Ethanol 70%ig zugesetzt und das Lysat auf die mitgelieferte Säule überführt. Zentrifugieren bei 8000 xg für 15 sec beförderte den Lysepuffer in das Auffanggefäß, die RNA blieb in der Membran haften. Es folgte ein Waschgang mit 70 µl Waschpuffer RW1, nach dem Zentrifugieren (8000 xg, 15 sec) wurde Auffanggefäß und Eluat verworfen. War das Auffanggefäß erneuert, konnten zwei weitere Waschgänge mit je 500 µl Waschpuffer RPE folgen. Danach wurde mit 10000 xg für 2 min zentrifugiert um die Membran zu trocknen, denn in den Puffern enthaltener Alhohol hätte mit nachfolgenden Reaktionen interferiert. Nun lösten 50 µl RNase-freies Wasser die RNA aus der Membran und wurden in ein 1,5 ml Eppendorff-

19
Gefäß zentrifugiert. 1 µl DNase, RNase-frei (Roche, Mannheim, BRD), zugesetzt und bei 37 ºC für 1 h inkubiert, zerstörte eventuell übrig gebliebene DNA. Anschließend wurde der Gehalt an RNA sowie ihr Reinheitsgrad photometrisch (Ultrospec 3000, Pharmacia Biotech, Cambridge, England) bestimmt. Die Lagerung der RNA-Lösung erfolgte anschließend bei –80 ºC.
2.3 Semiquantitative Realtime RT-PCR
2.3.1 Reverse Transkription Da die Taq-Polymerase in der Polymerasekettenreaktion zur Amplifikation DNA braucht, mußte die Einzelstrang-RNA mittels einer reversen Transkriptase (RT) in cDNA umgeschrieben werden. Dieser Schritt wurde unabhängig von der eigentlichen PCR durchgeführt, es handelte sich also um eine Two-Step RT-PCR. Zum Einsatz kam das AMV-Kit der Firma Roche (Roche, Mannheim, BRD). Die reverse Transkriptase in diesem Kit stammt vom Avian Myeloblastosis Virus und hat ein Temperaturoptimum von 42 ºC, wobei sie effizienter arbeitet als andere RTs. Als Primer wurde der enthaltene Random Primer p(dN)6 ausgewählt. Dieser transkribiert zwar nicht nur die interessierende mRNA, jedoch besteht auch bei längeren RNAs eine statistisch gleichmäßige Umschreibung (i. Ggs. zum Oligo p(dT)-Primer). Die RNA wurde bis zu einem Gehalt von 1 µg in 8,2 µl Wasser verdünnt, bei 65 ºC für 15 min denaturiert und für 5 min auf Eis gesetzt. Der während dieser Zeit pipettierte Master-Mix enthielt pro Probe 2 µl 10x Reaktionspuffer, 4 µl 25 mM MgCl2, 2 µl Desoxynucleotid Mix, 2 µl Random Primer, 1 µl RNase-Inhibitor und 0,8 µl der AMV RT. Das Primer-Annealing erfolgte bei 25 ºC für 10 min, die cDNA-Synthese bei 42 ºC für 60 min. Nach abgeschlossener Synthese wurde das Enzym durch fünfminütiges Erhitzen auf 99 ºC inaktiviert und die Probe wurde auf Eis gelegt. Die Lagerung der cDNA erfolgte bei –20 ºC.
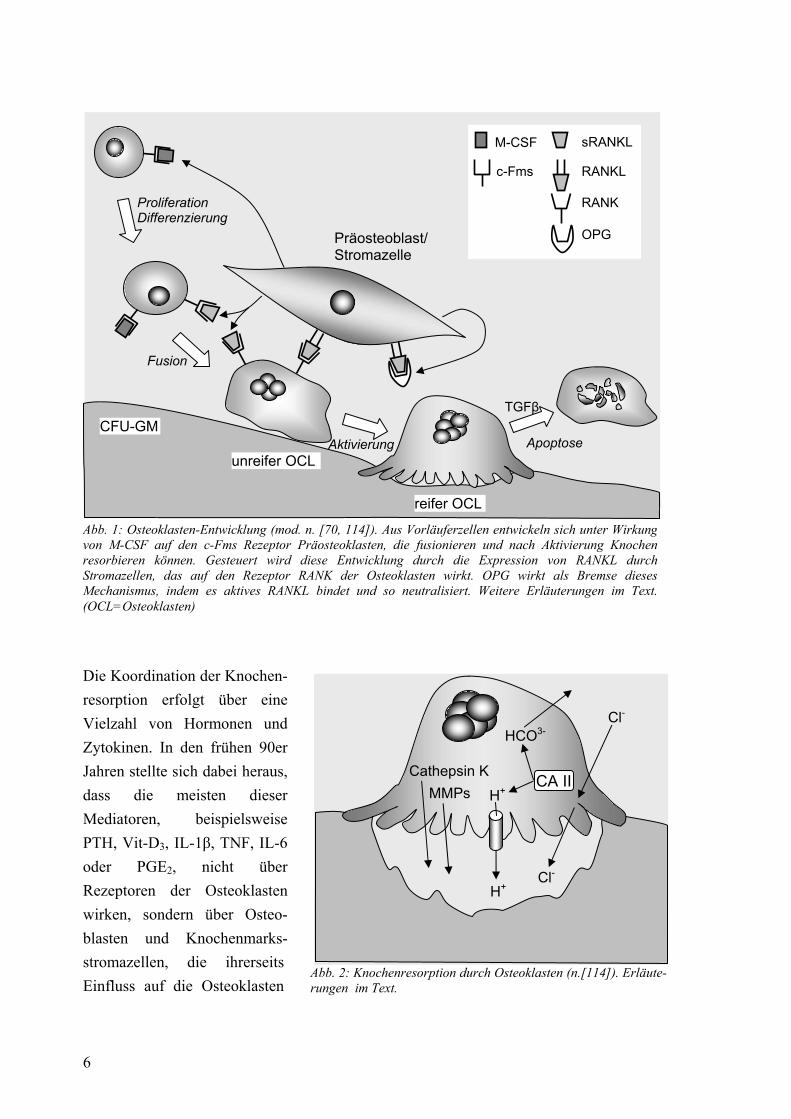
2.3.2 Primerdesign und Etablierung Die Oligonukleotid-Sequenzen der verwendeten Primer wurden anhand der Sequenzen der mRNAs aus der Gen-Datenbank des National Center for Biotechnology Information (NCBI, NIH, Bethesda, USA) ausgewählt. Entscheidend war GC-Gehalt (50-60 %), Länge (18-24-mer) und Schmelztemperatur. Die spätere Annealing-Temperatur mußte in etwa zu der der β-Actin-Primer passen, da diese als Quantifizierungsstandard verwendet werden sollten. Außerdem sollte das amplifizierte Fragment eine Länge von etwa 100 bis 300 bp haben. Mit der Primersynthese wurde die Firma MWG-Biotech (Ebersberg, BRD) beauftragt. Eine Auflistung der Primer mit ihren Sequenzen zeigt Tab. 1.
20
Sequenz Fragment-Länge Referenz
F 5´-CTA CGT CGC CCT GGA CTT CGA GC-3´ β-Actin R 5´-GAT GGA GCC GCC GAT CCA CAC GG-3´ 385 bp NM_001101
F 5´-ATC CGA CTG CTT TGG ACC TAA ATG-3´ SDC1 R 5´-CCA TAG ATT AGG GAA GCA AGA TGG G-3´ 297 bp NM_002997
F 5´-GCA GTG TCT TTG GTC TCC TG-3´ OPG R 5´-AAC CTG AAG AAT GCC TCC TC-3´ 130 bp HSU94332
F 5´-GCC AGT GGG AGA TGT TAG AC-3´ RANKL R 5´-GCA CAT GAC CAG GGA CCA AC-3´ 154 bp NM_003701 NM_033012
Tab. 1: Sequenzen der Oligonukleotidprimer (F=Forward-, R=Reverse-Primer), Fragmentlängen und Referenzen nach NCBI Daran schloß sich die Etablierung der Primerpärchen an. OPG- und RANKL-Primer wurden an Zellmaterial aus einem Lymphknoten mit pseudolymphomartiger Parenchym-veränderung etabliert. Dieser war uns freundlicherweise von Herrn Prof. Dr. Büttner zur Verfügung gestellt worden. SDC1-Primer konnten an Zellen der Zelllinie U-266 etabliert werden. Die Primer für β-Actin waren bereits im Vorfeld durch unsere Arbeitsgruppe etabliert worden. Hier sollen nur die für die Methodik wichtigen Daten genannt sein, genauere Beschreibungen finden sich im Ergebnisteil. Die Spezifität und Effizienz einer PCR hängt im wesentlichen von zwei Faktoren ab, einerseits von der Magnesiumchlorid-Konzentration, denn Mg2+ ist Kofaktor der Taq-DNA-Polymerase und MgCl2 bildet mit den dNTPs lösliche Komplexe, die die Polymerase erkennt. Andererseits ist die Annealing-Temperatur, d. h. die Temperatur, bei der die Primer spezifisch an das DNA-Template binden, effizienzbestimmend. So mußten also durch mehrere Testläufe optimale Bedingungen gefunden werden. Die daraufhin verwendeten Temperaturen und Konzentrationen zeigt Tab. 2.
Annealing- Temperatur
MgCl2- Konzentration Effizienz
β-Actin 58-65 °C 3 mM 1,94 SDC1 59 °C 4 mM 1,93 OPG 58 °C 2 mM 1,80 RANKL 61 °C 2 mM 1,85
Anhand der Standardkurve konnte über die LightCycler® Data-Analysis-Sofware die Effizienz bestimmt werden (Kap. 2.3.4). Zusätzlich wurde zwecks Überprüfung auf unspezifische Nebenprodukte eine Gelelektrophorese der PCR-Produkte gemacht.
Tab. 2: Optimale PCR-Bedingungen der verwendeten Primer und experimentelllermittelte Effizienz

21
Den letztendlichen Beweis für die Spezifität der Primer lieferte die Sequenzierung des Amplifikats. Zur Aufreinigung des PCR-Produktes verwendeten wir Sephacryl S-300 HR Micro Spin Columns (Amersham Pharmacia Biotech, Little Chalont, England). Ein in den Säulen enthaltenes Austauscherharz hielt überschüssige und die Sequenzierung störende Primer beim Hindurchzentrifugieren (735 xg, 2 min) zurück. Die Sequenzierung erfolgte mit einer Modifizierung der Kettenabbruchmethode nach Frederick Sanger [118] an einem ABI Prism 310 Sequenzer (Applied Biosystems, Weiterstadt, BRD). Der Ansatz für das Thermocycling enthielt 4 µl Terminator Ready Reaction Mix (DNA-Polymerase, Puffer, Desoxynucleotide und Didesoxynucleotide, basenspezifisch fluoreszenzfarbstoffmarkiert), 4 µl Template mit einem DNA-Gehalt von 30-90 ng, 1 µl Primer (10 pmol/µl) und 11 µl Aqua dest.. Um nun eine ausreichende Menge an Ketten herzustellen, die jeweils bei Kopplung an ein basenspezifisch farbstoffmarkiertes Didesoxynucleotid abbrechen, wurde ein GeneAmp 9600 Thermocycler (Perkin-Elmer Corp., Norwalk, USA) mit 25 Zyklen verwendet. Ein Zyklus bestand aus der Denaturierungsphase bei 96 °C für 10 sec, der Annealingphase für 5 sec bei primerspezifischer Annealingtemperatur und der Extensions-phase bei 60 °C für 4 min. Durch Fällung mit 10 µl 3 M Natriumazetat (pH 4,6) und 250 µl reinem Alkohol in 80 µl Wasser, anschließende Zentrifugation und nochmaliges Waschen mit 250 µl 70%igem Alkohol konnte die DNA pelletiert und dann getrocknet werden. Die Vorbereitung auf den Detektionsprozeß im Sequenzer umfaßte das Wiederauflösen der DNA in 25 µl TSR und die Denaturierung in Einzelstränge bei 90 °C für 5 min. Im Sequenzer wurde die DNA in einer Spur des Sequenziergels aufgetrennt und mit einem Laserphotometer detektiert.
2.3.3 Semiquantitative PCR mit dem LightCycler Das LightCycler-System (Roche, Mannheim, BRD) verbindet die Prozesse der Amplifikation und Detektion eines PCR-Produktes durch die Kombination eines Thermocyclers mit einem Fluorimeter (Abb. 6). Die Verwendung fluoreszeierender Farbstoffe ermöglicht es, die Amplifikatmenge nach jedem einzelnen Zyklus fluorimetrisch zu bestimmen. Die Daten aus der exponentiellen Phase dieser „kinetischen“ PCR werden unter Verwendung entsprechender Standards (absolute oder relative) zur Quantifizierung herangezogen [155]. Bei den zur Detektion verwendeten Fluorochromen handelt es sich entweder um Farbstoffe, die an kurze korrespondierende Oligonukleotid-Pärchen gekoppelt sind und spezifisch an bestimmte DNA-Abschnitte binden (Hybridisierungssonden) oder um Substanzen, die sich ähnlich wie Ethidiumbromid an doppelsträngige DNA anlagern (SYBR Green I). Wir entschieden uns für den Farbstoff SYBR Green I, dessen Spezifität im Wesentlichen von der Spezifität der Primer abhängt [96]. Außerdem verwendeten wir

22
die FastStart Taq DNA-Polymerase. Diese ist durch Blockierung mit hitzelabilen Gruppen bei Raumtemperatur inaktiv und wird erst unmittelbar vor der PCR durch Erhitzen aktiviert (sog. „Hot-Start“-PCR), was im Endeffekt weniger Nebenprodukte durch unspezifische Primerbindung bedeutet. So wird Spezifität und Sensitivität der PCR signifikant verbessert [13, 31, 69].
Zur Verwendung kam so das LightCycler-FastStart DNA Master SYBR Green I – Kit der Firma Roche. Im „ready-to-use“-Reaktions-Mix waren Polymerase, Reaktionspuffer, Desoxynukleotid-Mix und SYBR Green-Farbstoff enthalten. Die Beladung der Reaktions-kapillaren erfolgte im Kühlblock mit je 1 µl Forward-Primer (10 pmol/µl), 1 µl Reverse-Primer (10 pmol/µl), 2 µl Reaktions-Mix, MgCl2-Lösung bis zur für die jeweiligen Primer optimalen Konzentration, sowie Wasser, aufgefüllt bis zur Gesamtmenge von 19 µl. Vom Produkt der reversen Transkription wurde 1 µl als Template zugegeben. Ein Lauf setzte sich zusammen aus einem Denaturierungsprogramm, das gleichzeitig der Aktivierung der Taq-Polymerase diente, einem Amplifikationsprogramm mit entsprechend häufiger Wiederholung, einem Schmelzkurvenprogramm und einer Abkühlphase. Der Ablauf der Programme und die Zeitpunkte der Messungen der Fluoreszenz konnten mit der LightCycler-Run-Software programmiert werden.
Abb. 6: Arbeitsprinzip des LightCyclers (n. [113]): Kombination aus Thermocycler und Fluorimeter. Die Pro-ben befinden sich in Glaskapillaren, die durch ein Luftheiz-/ kühl-System den PCR-Zyklen unterzogen werden. Ein Karussell als Halterung sorgt dafür, dass die Proben nach jedem einzelnen Zyklus über die Fuori-metereinheit bewegt werden können, um den Gehalt an Produkt zu bestimmen.

23
In der Denaturierungsphase erfolgte Erhitzen auf 95 °C für 10 min. Die FastStart Taq-Polymerase wurde aktiviert und die DNA lag danach in Einzelsträngen vor. Die Amplifikationszyklen bestanden aus drei Segmenten, wiederum aus einer Denaturierungsphase (95 °C, 5 sec), die nach jedem Zyklus die Doppelstränge wieder in Einzelstränge trennte, einer Annealingphase (5 sec bei primerspezifischer Annealingtem-peratur), in der die Primer an die Eizelstränge banden, und einer Extensionsphase (72 °C, 16-30 sec), in der die Stränge kopiert wurden. Danach erfolgte eine Fluoreszenz-Messung um die Menge an dsDNA zu bestimmen. Im Normalfall wurden 40-45 Zyklen gefahren. Im Schmelzkurvenprogramm wurden alle Doppelstränge nochmals denaturiert (95 °C) dann auf 50 °C abgekühlt und langsam (0,1 °/sec) wieder auf 95 °C erhitzt. Dabei wurde kontinuierlich die Fluoreszenz gemessen. Wenn die Schmelztemperatur eines Amplifikationsprodukts erreicht war, gab es dementsprechend einen Abfall der Fluoreszenz, der in der negativen ersten Ableitung der Fluoreszenz (F) nach der Temperatur (T) (-d(F)/dT) als Peak sichtbar wurde. Die Schmelzkurve diente vor allem dem Nachweis der Spezifität der Amplifikation. Letzter Bestandteil eines Laufs war die Abkühlung der Proben auf angenehme 40 °C. Häufig wurden zu Kontrollzwecken die Proben anschließend in 1,5 ml Eppendorf-Gefäße zentrifugiert, mit 5 µl Loading-Buffer (Blaumarker) versetzt und gelelektrophoretisch analysiert. Die hierzu verwendeten 2%igen Agarose-Gele bestanden für 50 ml Gel aus 1 g Agarosepulver (Sigma, Deisenhofen, BRD), 5 ml TBE-Puffer, 45 ml Wasser und 5 µl 0,1%igem Ethidiumbromid. Nach dem Erkalten wurden jeweils 15 µl Probe in die Fächer des Gels gegeben. Eine 100 bp DNA-Ladder (Gibco Life Technologies, Karlsruhe, BRD) diente der Fragmentgrößenorientierung. Die Auftrennung erfolgte mit 110 V über etwa 35 min auf Gelträgern der Firma Serva (Heidelberg, BRD). Der Dokumentation diente eine digitale Fotodokumentationsanlage (MWG-Biotech, Ebersberg, BRD) mit UV-Tisch.
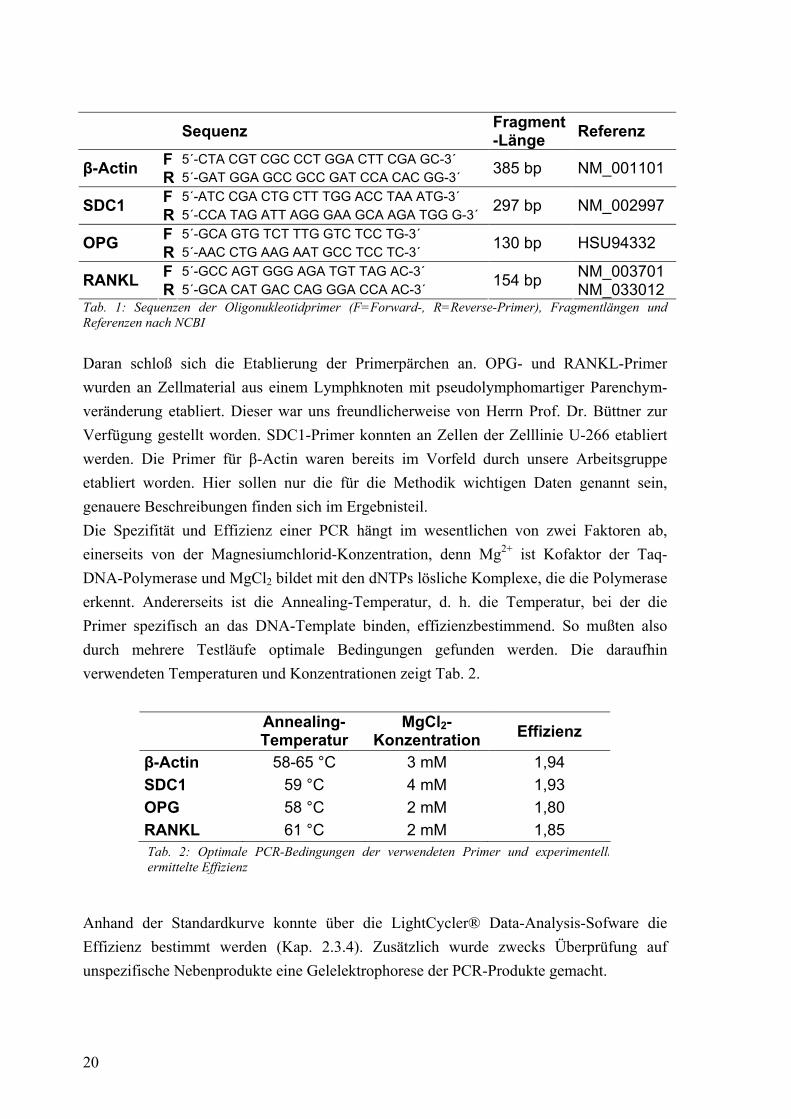
2.3.4 Auswertung der Messdaten mit der LightCycler Data Analysis-Software Zur Verwendung kam in den meisten Fällen die Analyse-Software in der Version 3.5, einige der ersten Datensätze wurden mit der Version 3.0 analysiert. Die Software berechnet Konzentrationen unbekannter Proben, indem sie deren Fluoreszenzen mit denen verschieden verdünnter Standards vergleicht. Für diese Analyse werden die Daten aus der exponentiellen (log-linearen) Phase der Amplifikation heran-gezogen. Durch setzen eines Noise-Bands kann Hintergrundfluoreszenz von der Analyse ausgeschlossen werden. Wird die Fluoreszenz logarithmisch gegen die Zyklusanzahl aufgetragen, so lässt sich am log-linearen Anteil der Fluoreszenzkurven der einzelnen Proben eine Gerade bestimmen, die eine Parallele zur x-Achse (sog. Crossing-Line) in je einem bestimmten Punkt schneidet. Dieser für jede Probe spezifische Schnittpunkt wird als
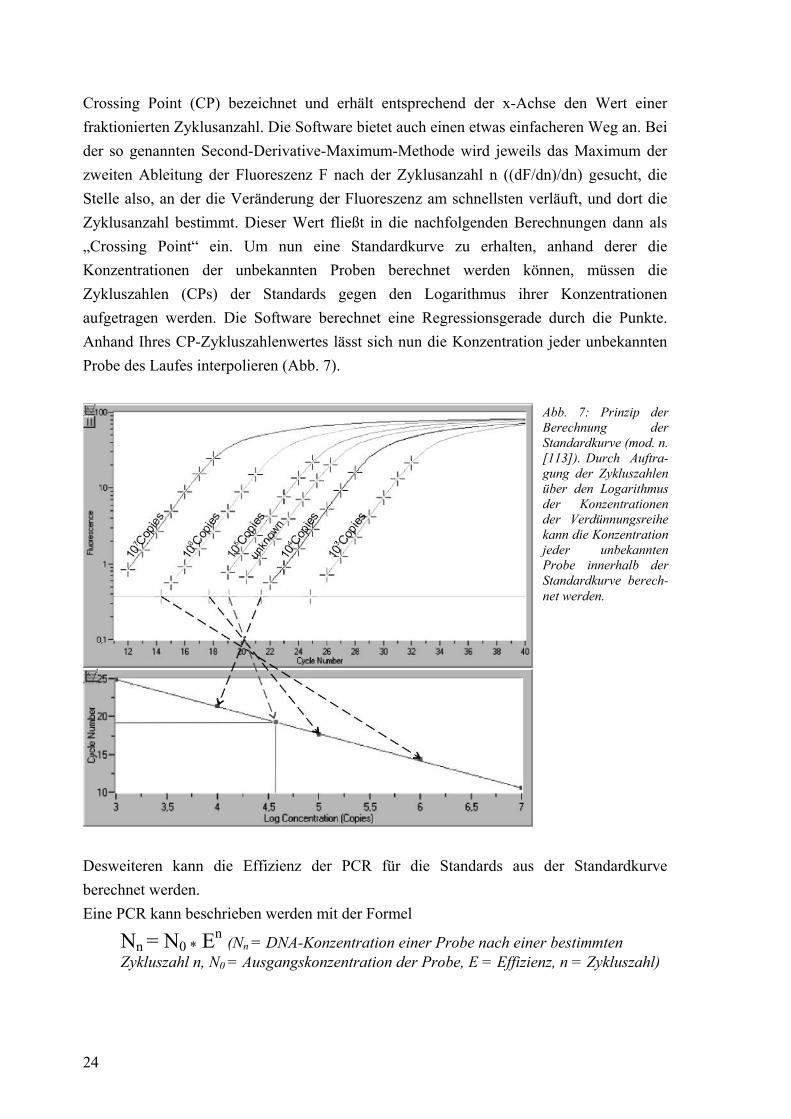
24
Crossing Point (CP) bezeichnet und erhält entsprechend der x-Achse den Wert einer fraktionierten Zyklusanzahl. Die Software bietet auch einen etwas einfacheren Weg an. Bei der so genannten Second-Derivative-Maximum-Methode wird jeweils das Maximum der zweiten Ableitung der Fluoreszenz F nach der Zyklusanzahl n ((dF/dn)/dn) gesucht, die Stelle also, an der die Veränderung der Fluoreszenz am schnellsten verläuft, und dort die Zyklusanzahl bestimmt. Dieser Wert fließt in die nachfolgenden Berechnungen dann als „Crossing Point“ ein. Um nun eine Standardkurve zu erhalten, anhand derer die Konzentrationen der unbekannten Proben berechnet werden können, müssen die Zykluszahlen (CPs) der Standards gegen den Logarithmus ihrer Konzentrationen aufgetragen werden. Die Software berechnet eine Regressionsgerade durch die Punkte. Anhand Ihres CP-Zykluszahlenwertes lässt sich nun die Konzentration jeder unbekannten Probe des Laufes interpolieren (Abb. 7).
Desweiteren kann die Effizienz der PCR für die Standards aus der Standardkurve berechnet werden. Eine PCR kann beschrieben werden mit der Formel
Nn = N0 * En (Nn = DNA-Konzentration einer Probe nach einer bestimmten Zykluszahl n, N0 = Ausgangskonzentration der Probe, E = Effizienz, n = Zykluszahl)
Abb. 7: Prinzip der Berechnung der Standardkurve (mod. n. [113]). Durch Auftra-gung der Zykluszahlen über den Logarithmus der Konzentrationen der Verdünnungsreihe kann die Konzentration jeder unbekannten Probe innerhalb der Standardkurve berech-net werden.

25
Bei oben genannter Auftragung der Zyklusanzahl gegen den Logarithmus der Konzentration ist demnach die Steigung der Geraden
S = -1/log E (S = Steigung, E = Effizienz) und somit die Effizienz E
E = 10 –1/S Fast alle Proben wurden mehrfach mit unterschiedlichen Ansätzen gemessen. Beispielhaft zeigt Tab. 3 die Beladung des LightCyclers für einen Lauf. Pos. Probe Standard Primer
1 unbehandelt 1,0 β-Actin 2 unbehandelt (Wdh. Pos.1) 1,0 β-Actin 3 unbehandelt (Wdh. Pos.1) 1,0 β-Actin 4 unbehandelt 0,1 β-Actin 5 unbehandelt 0,01 β-Actin 6 unbehandelt 0,001 β-Actin 7 behandelt - β-Actin 8 behandelt (Wdh. Pos.7) - β-Actin 9 behandelt (Wdh. Pos.7) - β-Actin 10 negativ (H2O) - β-Actin 11 unbehandelt - SDC1 12 behandelt - SDC1 13 negativ (H2O) - SDC1 14 unbehandelt (Wdh. Pos.11) - SDC1 15 behandelt (Wdh. Pos.12) - SDC1 16 negativ (H2O) - SDC1 17 unbehandelt (Wdh. Pos.11) - SDC1 18 behandelt (Wdh. Pos.12) - SDC1 19 negativ (H2O) - SDC1
Die Amplifikation von β-Actin als Housekeeping-Gen diente einerseits als relativer Standard, andererseits zur Normalisierung der Messwerte bei Vergleichen zweier unterschiedlicher Proben (im Normalfall unbehandelte Zellen und behandelte Zellen). Meist wurden die Messwerte der eine bestimmte Zeit behandelten Zellen (xi) zu den Werten zum Zeitpunkt Null (x0) in Relation gesetzt. Als Normalisierungsfaktor diente dann das Verhältnis von β-Actin-Wert bei unbehandelten (b0) zu dem bei behandelten Zellen (bi). Demnach berechnete sich die Abweichung (A) vom Nullwert:
A = xi/x0 * b0/bi
Tab. 3: Beladung des LightCyclers am Beispiel einer Messung der Ver-änderung der Expression von Syndecan 1 (SDC1); Mess-werte für Proben 1,2 und 3 ergeben b0, für 7,8 und 9 entsprechend bi. Die Proben 11, 14 und 17 dienen der Berechnung von x0, 12, 15 und 18 der von xi; Die Proben 10, 13, 16 und 19 dienen als Negative der Kontrolle von Kontamination

26
Diese Handhabung erfolgte bei den Messungen zur Expression von SDC1 an Myelomzelllinien (3.3.2, 3.3.3). Bei den Untersuchungen zur Gen-Expression nach Behandlung mit konditionierten Medien (3.3.5, 3.4.2), an den Kokulturen (3.3.4, 3.4.1) sowie zur Nativ-Expression von SDC1 (3.3.1) wurden die Messwerte jeweils nur auf die Expression von β-Actin als Housekeeping-Gen bezogen. Diese ist dort demnach als 1,0 definiert. So erhält man relative, untereinander vergleichbare Werte. Die relative Konzentration (Crel) berechnete sich dann:
Crel = xi/bi Da alle Faktoren auf gemittelten Werten beruhen, erfolgte die Berechnung des Gesamtfehlers nach dem Gauß´schen Gesetz der Fehlerfortpflanzung. Es wird im Folgenden, soweit nicht anders bezeichnet, immer der mittlere Fehler des Mittelwertes (SEM) angegeben. Bei der Herstellung der graphischen Darstellungen der Messwerte kam das Programm StatView® in der Version 5.0 der Firma SAS (SAS, Cary, USA) zum Einsatz.
2.4 Immun-Fuoreszenz-Färbung Die Fluoreszenz-Färbung diente der Demonstration der Syndecan 1 Expression auf der Zelloberfläche von Zellen der Linie U-266. Hierzu wurden Lab-Tek Chamber Slide Objektträger (Nunc Inc., Naperville, USA) mit 2 Kulturkammern verwendet. Die Zellen der Linie U-266 wuchsen 24 h auf den Objektträgern fest. Danach wurde das Medium abgesaugt, wurden die Zellen mit PBS-Puffer gewaschen, die Kunststoffkammerwände entfernt und die Zellen luftgetrocknet. Später erfolgte eine Rehydrierung der Zellen mit PBS, dann eine Überschichtung mit den Antikörpern gegen Syndecan 1. Es handelte sich hierbei um monoklonale murine Anti-human CD138 Antikörper des häufig verwendeten Klons B-B4 (Serotec, Oxford, Großbritannien), dessen Spezifität in der Literatur bereits gut untersucht ist [44]. Diese wurden 1:30 verdünnt mit 1 % BSA (bovines Serumalbumin) als Blocking-Buffer in PBS-Puffer, auf die Objektträger gegeben und 1 h bei 37 °C inkubiert. Nach sorgfältigem Spülen der Objektträger mit PBS-Puffer wurden die Zellen mit 3%iger Formaldehydlösung (Merck, Darmstadt, BRD) 5 min fixiert. Nach erneutem Waschen erfolgte die Überschichtung mit FITC (=Fluoresceinisothiocyanat) markierten Anti-Mouse-Sekundärantikörpern (DAKO, Glostrup, Denmark) für 30 min bei Raumtemperatur zur Markierung des gebundenen Anti-CD138. Dann schloß sich nach wiederholtem Waschen mit PBS die DAPI-Färbung an. Dazu wurden die Objektträger für 1 min in DAPI-Färbelösung (4,6-Diaminidin-2-phenylindol-dihydrochlorid (Boehringer, Mannheim, BRD)) mit einer Konzentration von 1 µg/ml gebracht. Diese letzte Färbung diente der Darstellung der DNA innerhalb der Zelle. Vor dem Eindecken wurden 5 µl Antifade Eindeckmedium (Appligene-Oncor/Q-Biogene, Heidelberg, BRD) auf die Zellen

27
gegeben. Die monochromatische Anregung der Fluoreszenzfarbstoffe erfolgte an einem ORBIT-System der Firma Improvision (Heidelberg, BRD), basierend auf einem Zeiss Axiovert S 100 Mikroskop (Carl Zeiss, Göttingen, BRD). Die Bilder wurden mit einer Hamamatsu Digitalkamera (Hamamatsu Photonics, Herrsching, BRD) aufgezeichnet und an das auf einer Apple Macintosh G4 Recheneinheit (Apple Computer, Cupertino, USA) basierende Bildverarbeitungssystem Open Lab® 2.2.5 übertragen.

28
3 Ergebnisse
3.1 Ergebnisse der Primer-Etablierung
3.1.1 Beta-Actin Die Primer für die mRNA von β-Actin waren bereits durch unsere Arbeitsgruppe auf anderen PCR-Geräten etabliert worden (Abb. 8). Es wurde überprüft, ob die ermittelten Bedingungen auch für die PCR mit dem LightCycler gelten. 1 cgcgtccgcc ccgcgagcac agagcctcgc ctttgccgat ccgccgcccg tccacacccg 61 ccgccagctc accatggatg atgatatcgc cgcgctcgtc gtcgacaacg gctccggcat 121 gtgcaaggcc ggcttcgcgg gcgacgatgc cccccgggcc gtcttcccct ccatcgtggg 181 gcgccccagg caccagggcg tgatggtggg catgggtcag aaggattcct atgtgggcga 241 cgaggcccag agcaagagag gcatcctcac cctgaagtac cccatcgagc acggcatcgt 301 caccaactgg gacgacatgg agaaaatctg gcaccacacc ttctacaatg agctgcgtgt 361 ggctcccgag gagcaccccg tgctgctgac cgaggccccc ctgaacccca aggccaaccg 421 cgagaagatg acccagatca tgtttgagac cttcaacacc ccagccatgt acgttgctat 481 ccaggctgtg ctatccctgt acgcctctgg ccgtaccact ggcatcgtga tggactccgg 541 tgacggggtc acccacactg tgcccatcta cgaggggtat gccctccccc atgccatcct 601 gcgtctggac ctggctggcc gggacctgac tgactacctc atgaagatcc tcaccgagcg 661 cggctacagc ttcaccacca cggccgagcg ggaaatcgtg cgtgacatta aggagaagct 721 gtgctacgtc gccctggact tcgagcaaga gatggccacg gctgcttcca gctcctccct 781 ggagaagagc tacgagctgc ctgacggcca ggtcatcacc attggcaatg agcggttccg 841 ctgccctgag gcactcttcc agccttcctt cctgggcatg gagtcctgtg gcatccacga 901 aactaccttc aactccatca tgaagtgtga cgtggacatc cgcaaagacc tgtacgccaa 961 cacagtgctg tctggcggca ccaccatgta ccctggcatt gccgacagga tgcagaagga 1021 gatcactgcc ctggcaccca gcacaatgaa gatcaagatc attgctcctc ctgagcgcaa 1081 gtactccgtg tggatcggcg gctccatcct ggcctcgctg tccaccttcc agcagatgtg 1141 gatcagcaag caggagtatg acgagtccgg cccctccatc gtccaccgca aatgcttcta 1201 ggcggactat gacttagttg cgttacaccc tttcttgaca aaacctaact tgcgcagaaa 1261 acaagatgag attggcatgg ctttatttgt tttttttgtt ttgttttggt tttttttttt 1321 tttttggctt gactcaggat ttaaaaactg gaacggtgaa ggtgacagca gtcggttgga 1381 gcgagcatcc cccaaagttc (...) 1793
Abb. 8: Sequenz der mRNA von β-Actin. Translatierter Bereich in fett, Positionen der Primer kursiv und grau hinterlegt. Basen ab Position 1400 sind nicht weiter aufgeführt.
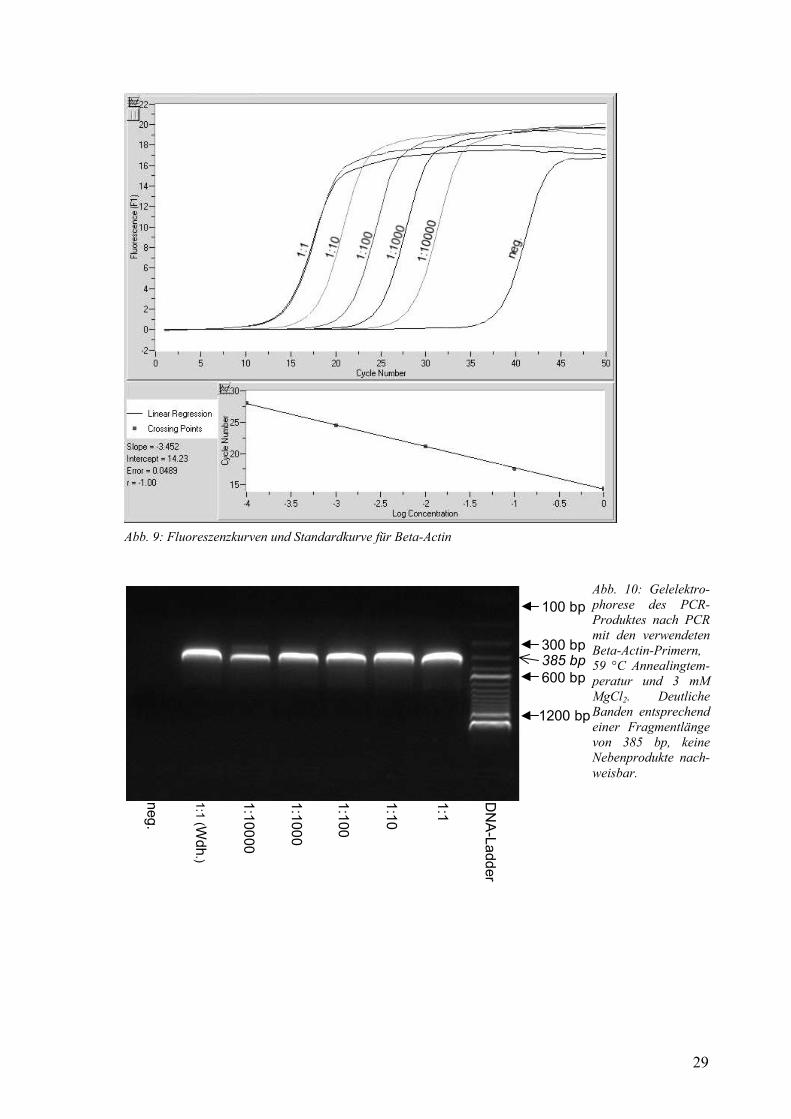
Die Verdünnungsreihe bis 1:10000 lieferte bei 59 °C und 3 mM MgCl2 eine sehr genaue Standardkurve (Abb. 9). In der Gelelektrophorese zeigte sich ein einziges Produkt mit den erwarteten 385 bp Länge (Abb. 10). Die Reaktion hatte eine Effizienz von 1,94. Reaktionen bei 58 °C und 61 °C lieferten fast identische Ergebnisse. Die verwendeten Primer eignen sich unter den in den nachfolgenden Experimenten angewendeten Bedingungen zum Nachweis und zur Quantifizierung von β-Actin.
29
Abb. 9: Fluoreszenzkurven und Standardkurve für Beta-Actin
Abb. 10: Gelelektro-phorese des PCR-Produktes nach PCR mit den verwendeten Beta-Actin-Primern, 59 °C Annealingtem-peratur und 3 mM MgCl2. Deutliche Banden entsprechend einer Fragmentlänge von 385 bp, keine Nebenprodukte nach-weisbar.
1:10000
neg.
1:1000
DN
A-Ladder
1:100
1:1 (Wdh .)
1:10
1:1
100 bp
300 bp
600 bp
1200 bp
385 bp

30
3.1.2 Syndecan 1 (SDC1) Ursprung der SDC1-mRNA ist die Region 2p24.1 auf Chromosom 2. Ihre Länge beträgt 3148 Basen, von denen die Basen 249-1181 zum 310 AS langen Produkt translatiert werden. Der verwendete F-Primer entspricht den Basen 1507-1530, der R-Primer ist komplementär zu den Basen 1778-1803. Das entsprechend amplifizierte Fragment hat demnach eine Länge von 297 bp (Abb. 11). Alle bisher beschriebenen Variationen der mRNA-Sequenz liegen außerhalb des amplifizierten Fragments. 1 gtgggaaggg cctgtgggtt tattataagg cggagctcgg cgggagaggt gcgggccgaa 61 tccgagccga gcggagagga atccggcagt agagagcgga ctccagccgg cggaccctgc 121 agccctcgcc tgggacagcg gcgcgctggg caggcgccca agagagcatc gagcagcgga 181 acccgcgaag ccggcccgca gccgcgaccc gcgcagcctg ccgctctccc gccgccggtc 241 cgggcagcat gaggcgcgcg gcgctctggc tctggctgtg cgcgctggcg ctgagcctgc 301 agccggccct gccgcaaatt gtggctacta atttgccccc tgaagatcaa gatggctctg 361 gggatgactc tgacaacttc tccggctcag gtgcaggtgc tttgcaagat atcaccttgt 421 cacagcagac cccctccact tggaaggaca cgcagctcct gacggctatt cccacgtctc 481 cagaacccac cggcctggag gctacagctg cctccacctc caccctgccg gctggagagg 541 ggcccaagga gggagaggct gtagtcctgc cagaagtgga gcctggcctc accgcccggg 601 agcaggaggc caccccccga cccagggaga ccacacagct cccgaccact catcaggcct 661 caacgaccac agccaccacg gcccaggagc ccgccacctc ccacccccac agggacatgc 721 agcctggcca ccatgagacc tcaacccctg caggacccag ccaagctgac cttcacactc 781 cccacacaga ggatggaggt ccttctgcca ccgagagggc tgctgaggat ggagcctcca 841 gtcagctccc agcagcagag ggctctgggg agcaggactt cacctttgaa acctcggggg 901 agaatacggc tgtagtggcc gtggagcctg accgccggaa ccagtcccca gtggatcagg 961 gggccacggg ggcctcacag ggcctcctgg acaggaaaga ggtgctggga ggggtcattg 1021 ccggaggcct cgtggggctc atctttgctg tgtgcctggt gggtttcatg ctgtaccgca 1081 tgaagaagaa ggacgaaggc agctactcct tggaggagcc gaaacaagcc aacggcgggg 1141 cctaccagaa gcccaccaaa caggaggaat tctatgcctg acgcgggagc catgcgcccc 1201 ctccgccctg ccactcacta ggcccccact tgcctcttcc ttgaagaact gcaggccctg 1261 gcctcccctg ccaccaggcc acctccccag cattccagcc cctctggtcg ctcctgccca 1321 cggagtcgtg gggtgtgctg ggagctccac tctgcttctc tgacttctgc ctggagactt 1381 agggcaccag gggtttctcg cataggacct ttccaccaca gccagcacct ggcatcgcac 1441 cattctgact cggtttctcc aaactgaagc agcctctccc caggtccagc tctggagggg 1501 agggggatcc gactgctttg gacctaaatg gcctcatgtg gctggaagat cctgcgggtg 1561 gggcttgggg ctcacacacc tgtagcactt actggtagga ccaagcatct tgggggggtg 1621 gccgctgagt ggcaggggac aggagtccac tttgtttcgt ggggaggtct aatctagata 1681 tcgacttgtt tttgcacatg tttcctctag ttctttgttc atagcccagt agaccttgtt 1741 acttctgagg taagttaagt aagttgattc ggtatccccc catcttgctt ccctaatcta 1801 tggtcgggag acagcatcag ggttaagaag actttttttt ttttttttaa actaggagaa 1861 ccaaatctgg aagccaaaat gtaggcttag tttgtgtgtt (...) 3148
Abb. 11: Sequenz der mRNA von Syndecan 1. Translatierter Bereich in fett, Positionen der Primer kursiv und grau hinterlegt. Basen ab Position 1901 sind nicht weiter aufgeführt.
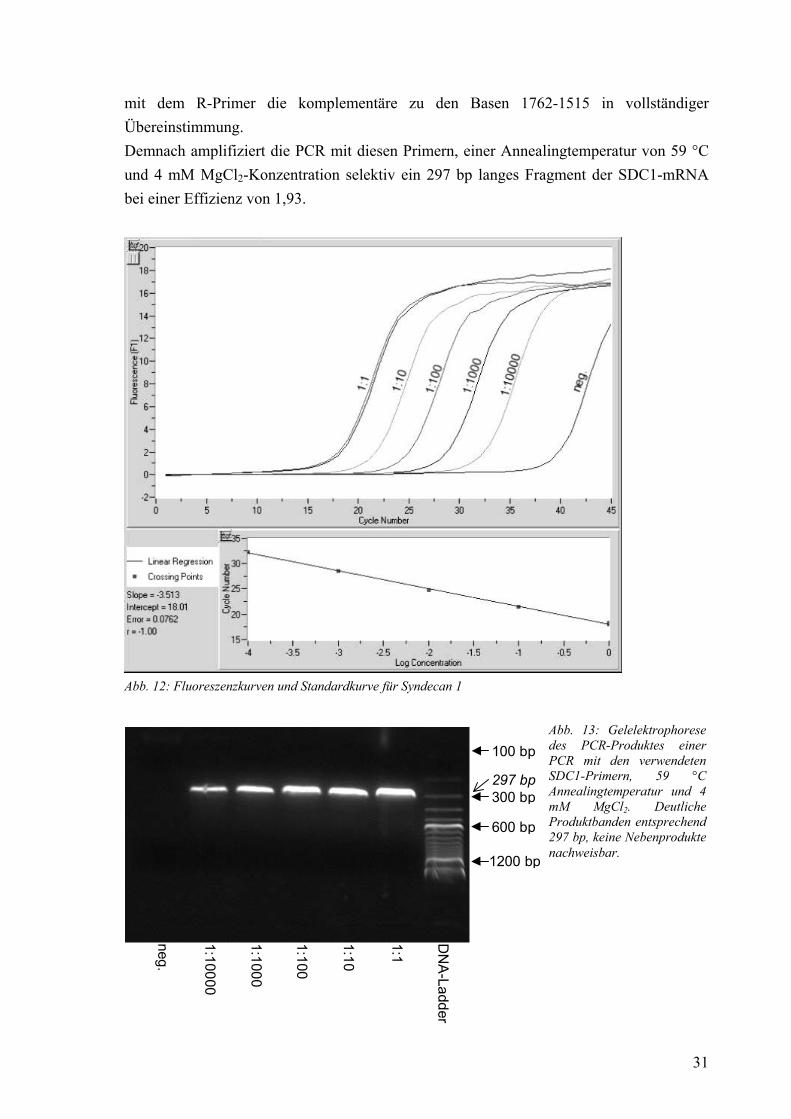
Aus der PCR ergab sich eine fehlerarme Standardkurve über einen Verdünnungsbereich bis 1:10000 (Abb. 12). Die anschließende Gelelektrophorese zeigte ein Produkt im Längenbereich um 300 bp sowie keine Nebenprodukte (Abb. 13). Die Sequenzierung des Produkts lieferte mit dem F-Primer die Sequenz der Basen 1538-1793 der SDC1-mRNA,
31
mit dem R-Primer die komplementäre zu den Basen 1762-1515 in vollständiger Übereinstimmung. Demnach amplifiziert die PCR mit diesen Primern, einer Annealingtemperatur von 59 °C und 4 mM MgCl2-Konzentration selektiv ein 297 bp langes Fragment der SDC1-mRNA bei einer Effizienz von 1,93.
Abb. 12: Fluoreszenzkurven und Standardkurve für Syndecan 1
100 bp
300 bp
600 bp
1200 bp
297 bp
1:10000
neg.
1:1000
DN
A-Ladder
1:100
1:10
1:1
Abb. 13: Gelelektrophorese des PCR-Produktes einer PCR mit den verwendeten SDC1-Primern, 59 °C Annealingtemperatur und 4 mM MgCl2. Deutliche Produktbanden entsprechend 297 bp, keine Nebenprodukte nachweisbar.

32
3.1.3 Osteoprotegerin (OPG) Die bisher beschriebene Sequenz der OPG-mRNA mit einer Länge von 1356 Basen umfasst vor allem den translatierten Bereich von Position 95-1300, der ein 401 AS langes Proteinprodukt liefert. Die Ursprungssequenz ist in der Region q23-24 auf Chromosom 8 zu finden. Der verwendete F-Primer entspricht den Basen 591- 610, der R-Primer ist komplementär zu den Basen 701-720. Die Länge des amplifizierten Fragment beträgt demnach 130 bp (Abb. 14). Variationen der mRNA-Sequenz sind bisher nicht berichtet worden. 1 gtatatataa cgtgatgagc gtacgggtgc ggagacgcac cggagcgctc gcccagccgc 61 cgyctccaag cccctgaggt ttccggggac cacaatgaac aagttgctgt gctgcgcgct 121 cgtgtttctg gacatctcca ttaagtggac cacccaggaa acgtttcctc caaagtacct 181 tcattatgac gaagaaacct ctcatcagct gttgtgtgac aaatgtcctc ctggtaccta 241 cctaaaacaa cactgtacag caaagtggaa gaccgtgtgc gccccttgcc ctgaccacta 301 ctacacagac agctggcaca ccagtgacga gtgtctatac tgcagccccg tgtgcaagga 361 gctgcagtac gtcaagcagg agtgcaatcg cacccacaac cgcgtgtgcg aatgcaagga 421 agggcgctac cttgagatag agttctgctt gaaacatagg agctgccctc ctggatttgg 481 agtggtgcaa gctggaaccc cagagcgaaa tacagtttgc aaaagatgtc cagatgggtt 541 cttctcaaat gagacgtcat ctaaagcacc ctgtagaaaa cacacaaatt gcagtgtctt 601 tggtctcctg ctaactcaga aaggaaatgc aacacacgac aacatatgtt ccggaaacag 661 tgaatcaact caaaaatgtg gaatagatgt taccctgtgt gaggaggcat tcttcaggtt 721 tgctgttcct acaaagttta cgcctaactg gcttagtgtc ttggtagaca atttgcctgg 781 caccaaagta aacgcagaga gtgtagagag gataaaacgg caacacagct cacaagaaca 841 gactttccag ctgctgaagt tatggaaaca tcaaaacaaa gcccaagata tagtcaagaa 901 gatcatccaa gatattgacc tctgtgaaaa cagcgtgcag cggcacattg gacatgctaa 961 cctcaccttc gagcagcttc gtagcttgat ggaaagctta ccgggaaaga aagtgggagc 1021 agaagacatt gaaaaaacaa taaaggcatg caaacccagt gaccagatcc tgaagctgct 1081 cagtttgtgg cgaataaaaa atggcgacca agacaccttg aagggcctaa tgcacgcact 1141 aaagcactca aagacgtacc actttcccaa aactgtcact cagagtctaa agaagaccat 1201 caggttcctt cacagcttca caatgtacaa attgtatcag aagttatttt tagaaatgat 1261 aggtaaccag gtccaatcag taaaaataag ctgcttataa ctggaaatgg ccattgagct 1321 gtttcctcac aattggcgag atcccatgga tgataa
Abb. 14: Sequenz der mRNA von Osteoprotegerin. Translatierter Bereich in fett, Positionen der Primer kursiv und grau hinterlegt.
Bei 58 °C Annealingtemperatur und 2 mM MgCl2 ergab sich unter Verwendung dieser Primer eine Standardkurve mit niedrigen Fehlerwerten über einen Verdünnungsbereich bis 1:10000 (Abb. 15). Die Gelelektrophorese des PCR-Produktes zeigte ein Fragment von etwa 130 bp bei fehlenden Nebenprodukten (Abb. 16). Die Sequenzierung des Amplifikats ergab mit dem F-Primer die Sequenz der mRNA der Basen 626-720 und mit dem R-Primer die komplementäre Sequenz zu den Basen 678-591. Unter den oben genannten PCR-Bedingungen bieten diese Primer also die Möglichkeit, OPG-mRNA selektiv nachzuweisen und zu quantifizieren. Die Effizienz ist mit 1,80 akzeptabel.
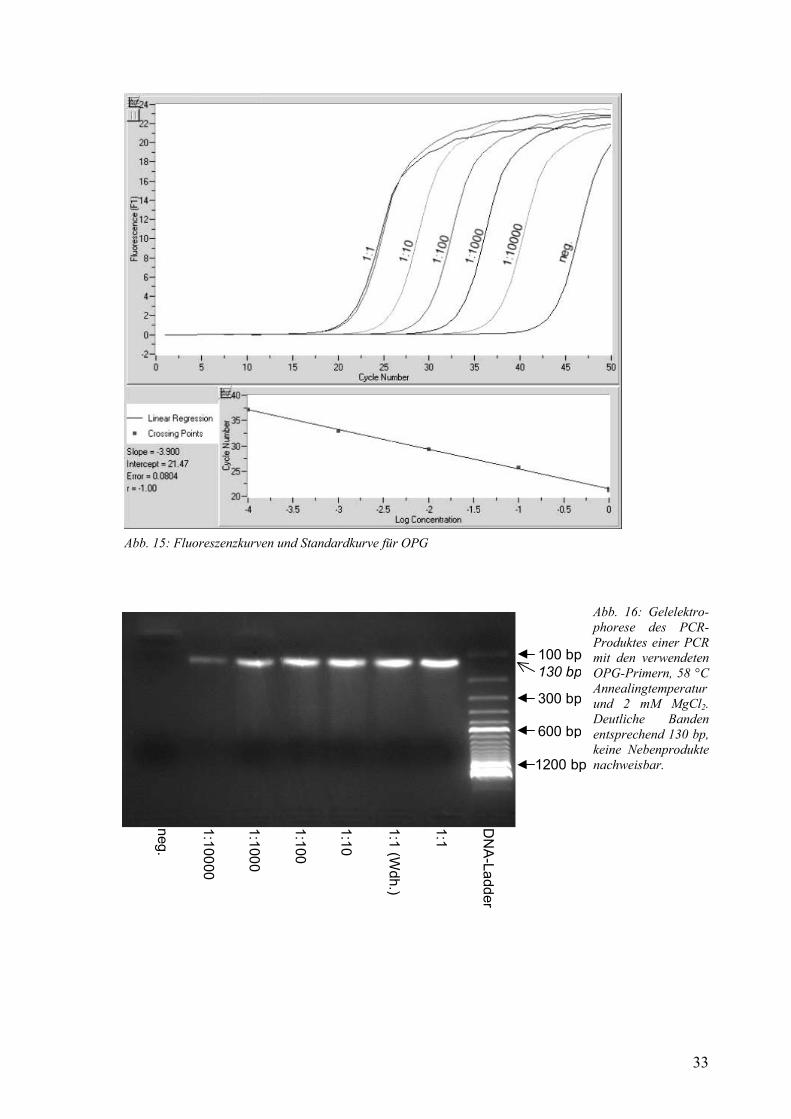
33
Abb. 15: Fluoreszenzkurven und Standardkurve für OPG ---Abb. „GrafikETABOPGgel“---
1:1000
neg.
1:100
DN
A-Ladder
1:10
1:10000
1:1 (Wdh.)
1:1
100 bp
300 bp
600 bp
1200 bp
130 bp
Abb. 16: Gelelektro-phorese des PCR-Produktes einer PCR mit den verwendeten OPG-Primern, 58 °C Annealingtemperatur und 2 mM MgCl2. Deutliche Banden entsprechend 130 bp, keine Nebenprodukte nachweisbar.

34
3.1.4 Receptor Activator of NF-κB Ligand (RANKL) Von RANKL sind die Sequenzen zweier Splicevarianten beschrieben. Variante 1 liefert ein 317 AS langes Protein-Produkt durch Translation der Basen 157-1110. Bei Variante 2 ist der erste Teil der Sequenz bis zur Position 371 durch eine andere ersetzt, sodass das Triplet an Position 376-78 zum Startcodon wird. Das resultierende Translationsprodukt ist entsprechend um 73 AS verkürzt. Der verwendete F-Primer entspricht den Basen 1378-1397, der R-Primer ist komplementär zu den Basen 1431-1412. Das Fragment hat eine Länge von 154 Basen und liegt in der gemeinsamen Sequenz beider Varianten (Abb. 17). Bekannte Variationen der Sequenz liegen nicht innerhalb der Primersequenzen. 1 ctcgacccac gcgtccgcgc gccccaggag ccaaagccgg gctccaagtc ggcgccccac 61 gtcgaggctc cgccgcagcc tccggagttg gccgcagaca agaaggggag ggagcgggag 121 agggaggaga gctccgaagc gagagggccg agcgccatgc gccgcgccag cagagactac 181 accaagtacc tgcgtggctc ggaggagatg ggcggcggcc ccggagcccc gcacgagggc 241 cccctgcacg ccccgccgcc gcctgcgccg caccagcccc ccgccgcctc ccgctccatg 301 ttcgtggccc tcctggggct ggggctgggc caggttgtct gcagcgtcgc cctgttcttc 361 tatttcagag cgcagatgga tcctaataga atatcagaag atggcactca ctgcatttat 421 agaattttga gactccatga aaatgcagat tttcaagaca caactctgga gagtcaagat 481 acaaaattaa tacctgattc atgtaggaga attaaacagg cctttcaagg agctgtgcaa 541 aaggaattac aacatatcgt tggatcacag cacatcagag cagagaaagc gatggtggat 601 ggctcatggt tagatctggc caagaggagc aagcttgaag ctcagccttt tgctcatctc 661 actattaatg ccaccgacat cccatctggt tcccataaag tgagtctgtc ctcttggtac 721 catgatcggg gttgggccaa gatctccaac atgactttta gcaatggaaa actaatagtt 781 aatcaggatg gcttttatta cctgtatgcc aacatttgct ttcgacatca tgaaacttca 841 ggagacctag ctacagagta tcttcaacta atggtgtacg tcactaaaac cagcatcaaa 901 atcccaagtt ctcataccct gatgaaagga ggaagcacca agtattggtc agggaattct 961 gaattccatt tttattccat aaacgttggt ggatttttta agttacggtc tggagaggaa 1021 atcagcatcg aggtctccaa cccctcctta ctggatccgg atcaggatgc aacatacttt 1081 ggggctttta aagttcgaga tatagattga gccccagttt ttggagtgtt atgtatttcc 1141 tggatgtttg gaaacatttt ttaaaacaag ccaagaaaga tgtatatagg tgtgtgagac 1201 tactaagagg catggcccca acggtacacg actcagtatc catgctcttg accttgtaga 1261 gaacacgcgt atttacagcc agtgggagat gttagactca tggtgtgtta cacaatggtt 1321 tttaaatttt gtaatgaatt cctagaatta aaccagattg gagcaattac gggttgacct 1381 tatgagaaac tgcatgtggg ctatgggagg ggttggtccc tggtcatgtg ccccttcgca 1441 gctgaagtgg agagggtgtc atctagcgca attgaaggat catctgaagg ggcaaattct 1501 tttgaattgt (...) 2226
Abb. 17: Sequenz der mRNA von RANKL. Translatierter Bereich in fett, Positionen der Primer in kursiv und grau hinterlegt. Basen ab Position 1511 sind nicht weiter aufgeführt.
Bei einer Annealingtemperatur von 61 °C, einer MgCl2-Konzentration von 2 mM und der Verwendung der oben genannten Primer gelang die Erstellung einer Standardkurve bis zu einer Verdünnung von 1:1000 (Abb. 18). Die Gelelektrophorese zeigte ein Produkt von etwa 150 bp (Abb. 19). Die Sequenzierung lieferte mit dem F-Primer die Sequenz der mRNA der Basen 1317-1431, mit dem R-Primer die komplementäre der Basen 1394-1284. Mit den hier verwendeten Primern lässt sich also RANKL-mRNA selektiv nachweisen und quantifizieren. Die Effizienz dieser Reaktion liegt bei 1,85. Der Versuch, ein
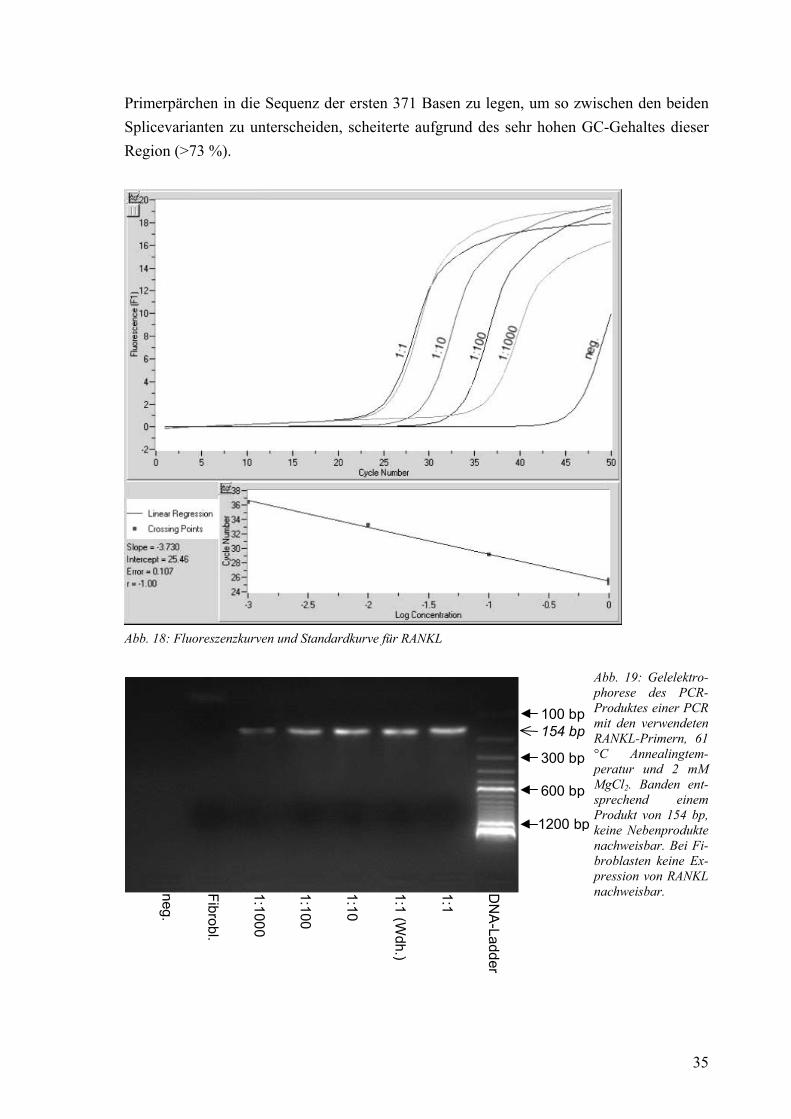
35
Primerpärchen in die Sequenz der ersten 371 Basen zu legen, um so zwischen den beiden Splicevarianten zu unterscheiden, scheiterte aufgrund des sehr hohen GC-Gehaltes dieser Region (>73 %).
Abb. 18: Fluoreszenzkurven und Standardkurve für RANKL
100 bp
300 bp
600 bp
1200 bp
154 bp
1:1000
neg.
1:100
DN
A-Ladder
1:10
Fibrobl.
1:1 (Wdh.)
1:1
Abb. 19: Gelelektro-phorese des PCR-Produktes einer PCR mit den verwendeten RANKL-Primern, 61 °C Annealingtem-peratur und 2 mMMgCl2. Banden ent-sprechend einem Produkt von 154 bp, keine Nebenprodukte nachweisbar. Bei Fi-broblasten keine Ex-pression von RANKL nachweisbar.
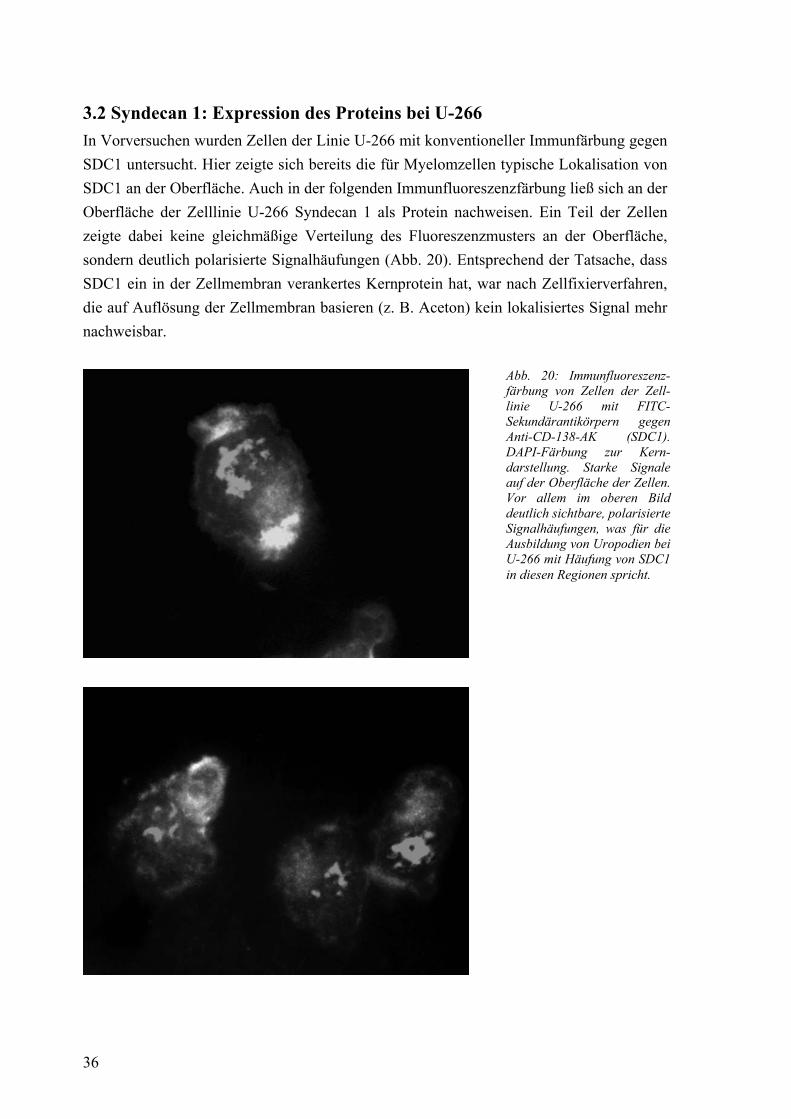
36
3.2 Syndecan 1: Expression des Proteins bei U-266 In Vorversuchen wurden Zellen der Linie U-266 mit konventioneller Immunfärbung gegen SDC1 untersucht. Hier zeigte sich bereits die für Myelomzellen typische Lokalisation von SDC1 an der Oberfläche. Auch in der folgenden Immunfluoreszenzfärbung ließ sich an der Oberfläche der Zelllinie U-266 Syndecan 1 als Protein nachweisen. Ein Teil der Zellen zeigte dabei keine gleichmäßige Verteilung des Fluoreszenzmusters an der Oberfläche, sondern deutlich polarisierte Signalhäufungen (Abb. 20). Entsprechend der Tatsache, dass SDC1 ein in der Zellmembran verankertes Kernprotein hat, war nach Zellfixierverfahren, die auf Auflösung der Zellmembran basieren (z. B. Aceton) kein lokalisiertes Signal mehr nachweisbar.
Abb. 20: Immunfluoreszenz-färbung von Zellen der Zell-linie U-266 mit FITC-Sekundärantikörpern gegen Anti-CD-138-AK (SDC1). DAPI-Färbung zur Kern-darstellung. Starke Signale auf der Oberfläche der Zellen. Vor allem im oberen Bild deutlich sichtbare, polarisierte Signalhäufungen, was für die Ausbildung von Uropodien bei U-266 mit Häufung von SDC1 in diesen Regionen spricht.
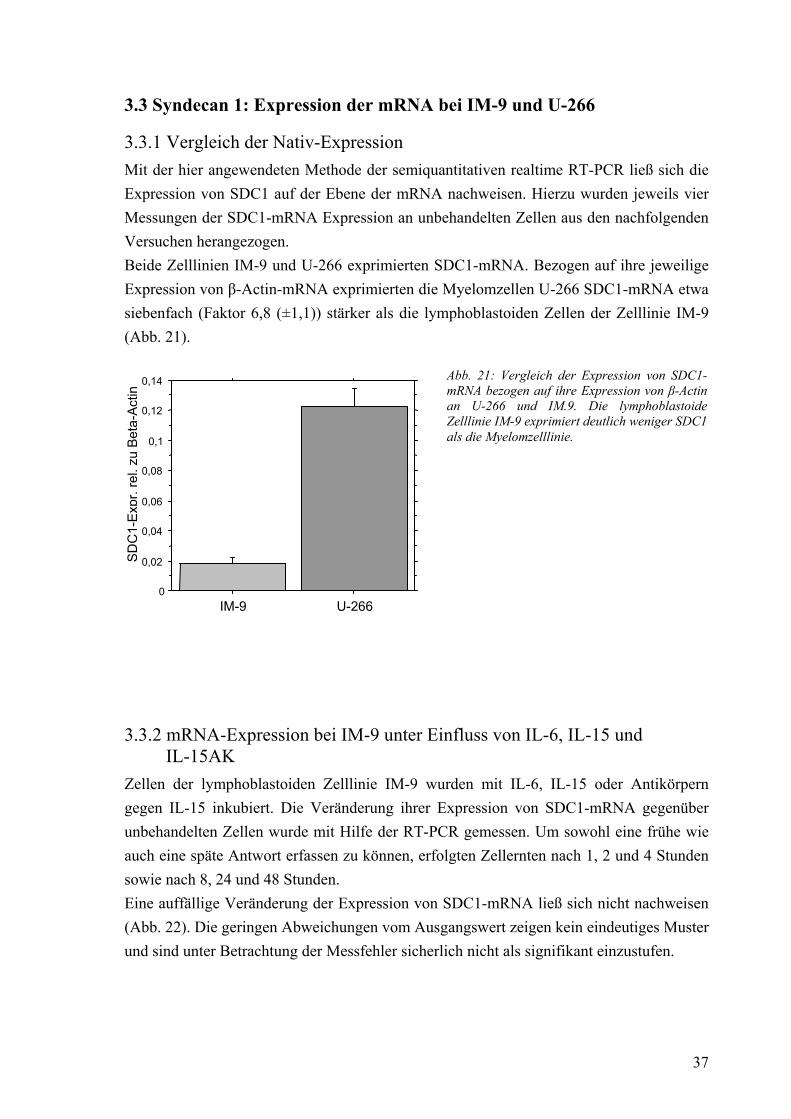
37
3.3 Syndecan 1: Expression der mRNA bei IM-9 und U-266
3.3.1 Vergleich der Nativ-Expression Mit der hier angewendeten Methode der semiquantitativen realtime RT-PCR ließ sich die Expression von SDC1 auf der Ebene der mRNA nachweisen. Hierzu wurden jeweils vier Messungen der SDC1-mRNA Expression an unbehandelten Zellen aus den nachfolgenden Versuchen herangezogen. Beide Zelllinien IM-9 und U-266 exprimierten SDC1-mRNA. Bezogen auf ihre jeweilige Expression von β-Actin-mRNA exprimierten die Myelomzellen U-266 SDC1-mRNA etwa siebenfach (Faktor 6,8 (±1,1)) stärker als die lymphoblastoiden Zellen der Zelllinie IM-9 (Abb. 21).
3.3.2 mRNA-Expression bei IM-9 unter Einfluss von IL-6, IL-15 und IL-15AK
Zellen der lymphoblastoiden Zelllinie IM-9 wurden mit IL-6, IL-15 oder Antikörpern gegen IL-15 inkubiert. Die Veränderung ihrer Expression von SDC1-mRNA gegenüber unbehandelten Zellen wurde mit Hilfe der RT-PCR gemessen. Um sowohl eine frühe wie auch eine späte Antwort erfassen zu können, erfolgten Zellernten nach 1, 2 und 4 Stunden sowie nach 8, 24 und 48 Stunden. Eine auffällige Veränderung der Expression von SDC1-mRNA ließ sich nicht nachweisen (Abb. 22). Die geringen Abweichungen vom Ausgangswert zeigen kein eindeutiges Muster und sind unter Betrachtung der Messfehler sicherlich nicht als signifikant einzustufen.
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
IM-9 U-266
SD
C1-
Exp
r.re
l.zu
Beta
-Act
in
Abb. 21: Vergleich der Expression von SDC1-mRNA bezogen auf ihre Expression von β-Actin an U-266 und IM.9. Die lymphoblastoide Zelllinie IM-9 exprimiert deutlich weniger SDC1 als die Myelomzelllinie.
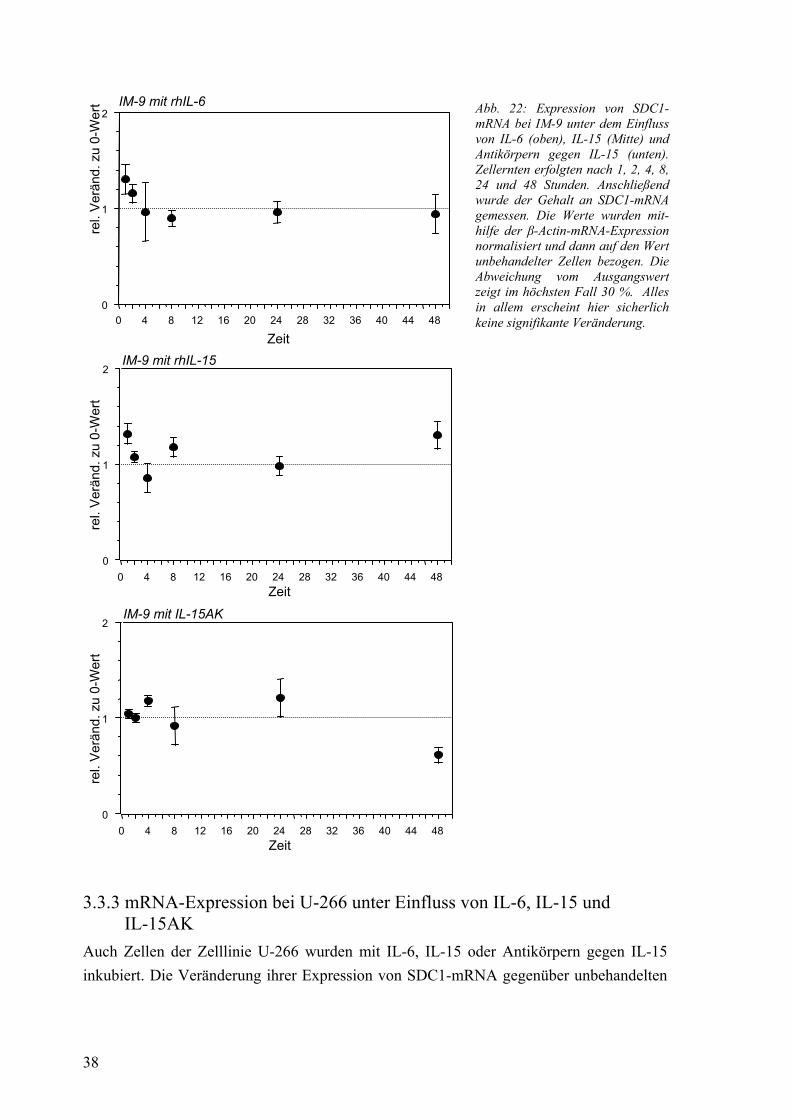
38
3.3.3 mRNA-Expression bei U-266 unter Einfluss von IL-6, IL-15 und IL-15AK
Auch Zellen der Zelllinie U-266 wurden mit IL-6, IL-15 oder Antikörpern gegen IL-15 inkubiert. Die Veränderung ihrer Expression von SDC1-mRNA gegenüber unbehandelten
0
1
2
0 4 8 12 16 20 24 28 32 36 40 44 48
Zeit
IM-9 mit rhIL-6 re
l. V
erän
d. z
u 0-
Wer
t
0
1
2
rel.
Ver
änd.
zu
0-W
ert
0 4 8 12 16 20 24 28 32 36 40 44 48Zeit
IM-9 mit rhIL-15
0
1
2
rel.
Ver
änd.
zu
0-W
ert
0 4 8 12 16 20 24 28 32 36 40 44 48Zeit
IM-9 mit IL-15AK
Abb. 22: Expression von SDC1-mRNA bei IM-9 unter dem Einfluss von IL-6 (oben), IL-15 (Mitte) und Antikörpern gegen IL-15 (unten). Zellernten erfolgten nach 1, 2, 4, 8, 24 und 48 Stunden. Anschließend wurde der Gehalt an SDC1-mRNA gemessen. Die Werte wurden mit-hilfe der β-Actin-mRNA-Expression normalisiert und dann auf den Wert unbehandelter Zellen bezogen. Die Abweichung vom Ausgangswert zeigt im höchsten Fall 30 %. Alles in allem erscheint hier sicherlich keine signifikante Veränderung.
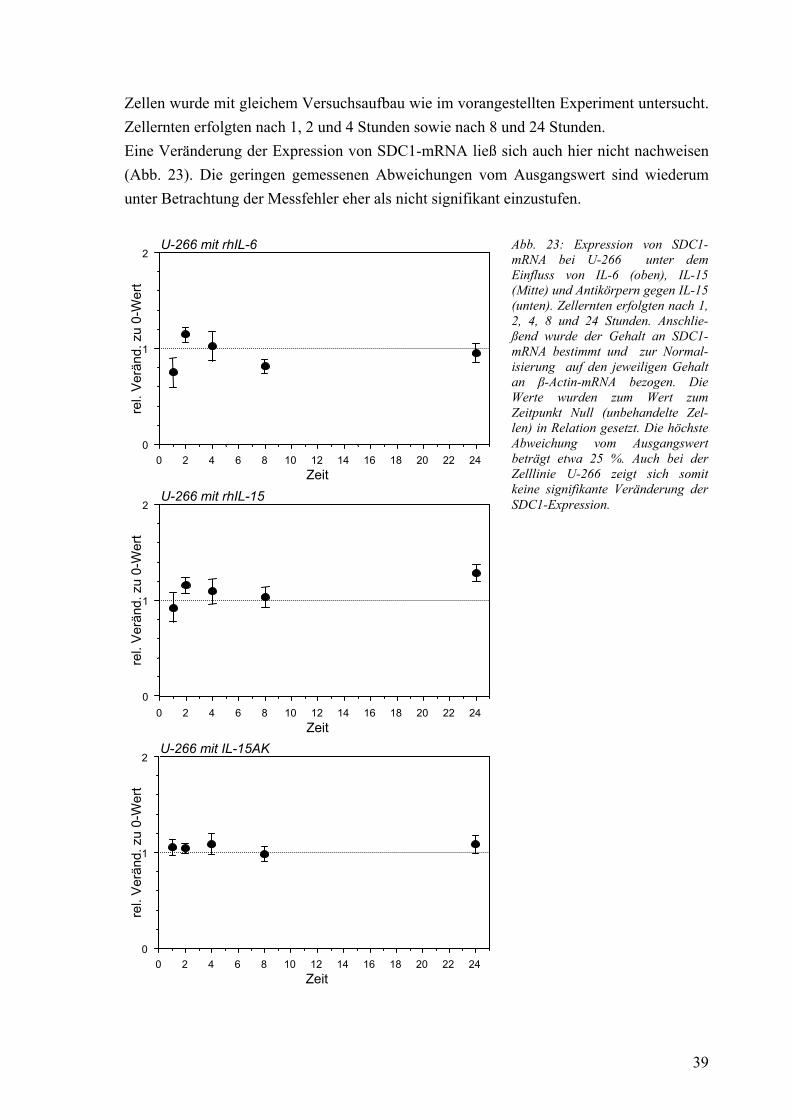
39
Zellen wurde mit gleichem Versuchsaufbau wie im vorangestellten Experiment untersucht. Zellernten erfolgten nach 1, 2 und 4 Stunden sowie nach 8 und 24 Stunden. Eine Veränderung der Expression von SDC1-mRNA ließ sich auch hier nicht nachweisen (Abb. 23). Die geringen gemessenen Abweichungen vom Ausgangswert sind wiederum unter Betrachtung der Messfehler eher als nicht signifikant einzustufen.
0
1
2
rel.
Ver
änd.
zu
0-W
ert
0 2 4 6 8 10 12 14 16 18 20 22 24Zeit
U-266 mit rhIL-6
0
1
2
rel.
Ver
änd.
zu
0-W
ert
0 2 4 6 8 10 12 14 16 18 20 22 24Zeit
U-266 mit rhIL-15
0
1
2
rel.
Ver
änd.
zu
0-W
ert
0 2 4 6 8 10 12 14 16 18 20 22 24Zeit
U-266 mit IL-15AK
Abb. 23: Expression von SDC1-mRNA bei U-266 unter dem Einfluss von IL-6 (oben), IL-15 (Mitte) und Antikörpern gegen IL-15 (unten). Zellernten erfolgten nach 1, 2, 4, 8 und 24 Stunden. Anschlie-ßend wurde der Gehalt an SDC1-mRNA bestimmt und zur Normal-isierung auf den jeweiligen Gehalt an β-Actin-mRNA bezogen. Die Werte wurden zum Wert zum Zeitpunkt Null (unbehandelte Zel-len) in Relation gesetzt. Die höchste Abweichung vom Ausgangswert beträgt etwa 25 %. Auch bei der Zelllinie U-266 zeigt sich somit keine signifikante Veränderung der SDC1-Expression.
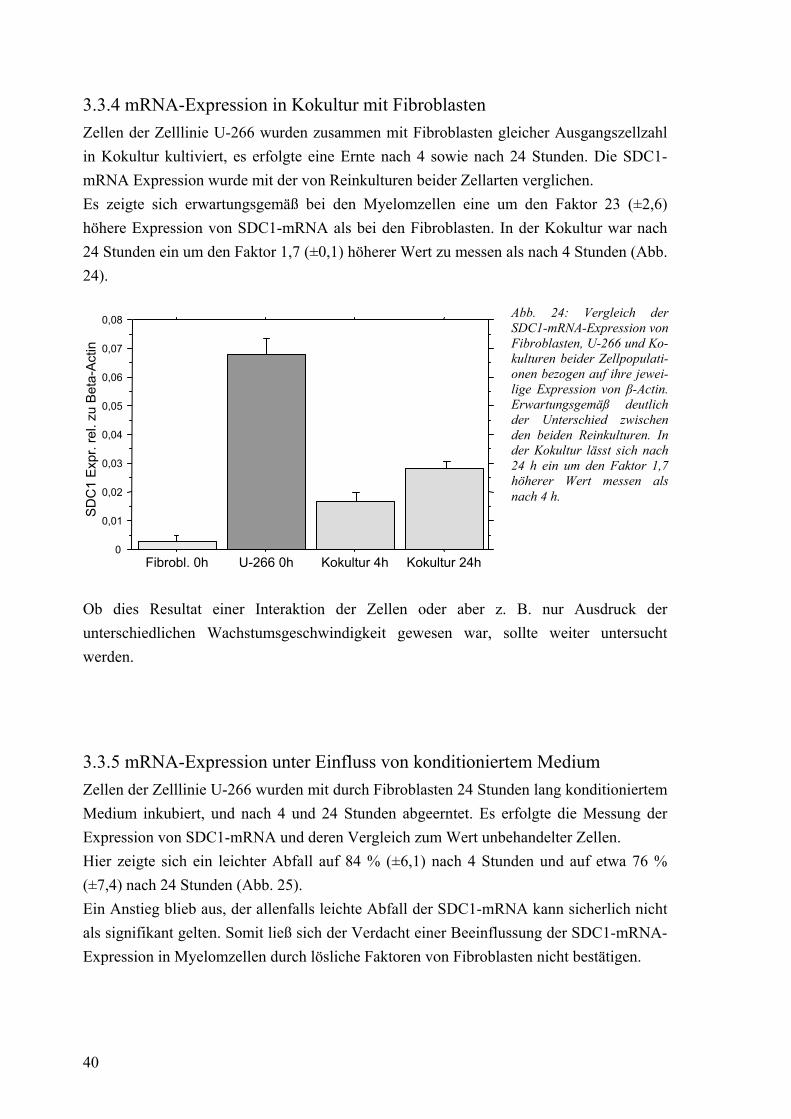
40
3.3.4 mRNA-Expression in Kokultur mit Fibroblasten Zellen der Zelllinie U-266 wurden zusammen mit Fibroblasten gleicher Ausgangszellzahl in Kokultur kultiviert, es erfolgte eine Ernte nach 4 sowie nach 24 Stunden. Die SDC1-mRNA Expression wurde mit der von Reinkulturen beider Zellarten verglichen. Es zeigte sich erwartungsgemäß bei den Myelomzellen eine um den Faktor 23 (±2,6) höhere Expression von SDC1-mRNA als bei den Fibroblasten. In der Kokultur war nach 24 Stunden ein um den Faktor 1,7 (±0,1) höherer Wert zu messen als nach 4 Stunden (Abb. 24).
Ob dies Resultat einer Interaktion der Zellen oder aber z. B. nur Ausdruck der unterschiedlichen Wachstumsgeschwindigkeit gewesen war, sollte weiter untersucht werden.
3.3.5 mRNA-Expression unter Einfluss von konditioniertem Medium Zellen der Zelllinie U-266 wurden mit durch Fibroblasten 24 Stunden lang konditioniertem Medium inkubiert, und nach 4 und 24 Stunden abgeerntet. Es erfolgte die Messung der Expression von SDC1-mRNA und deren Vergleich zum Wert unbehandelter Zellen. Hier zeigte sich ein leichter Abfall auf 84 % (±6,1) nach 4 Stunden und auf etwa 76 % (±7,4) nach 24 Stunden (Abb. 25). Ein Anstieg blieb aus, der allenfalls leichte Abfall der SDC1-mRNA kann sicherlich nicht als signifikant gelten. Somit ließ sich der Verdacht einer Beeinflussung der SDC1-mRNA-Expression in Myelomzellen durch lösliche Faktoren von Fibroblasten nicht bestätigen.
SD
C1
Expr
. rel
. zu
Bet
a-Ac
tin
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
Fibrobl. 0h Kokultur 24hKokultur 4hU-266 0h
Abb. 24: Vergleich der SDC1-mRNA-Expression von Fibroblasten, U-266 und Ko-kulturen beider Zellpopulati-onen bezogen auf ihre jewei-lige Expression von β-Actin. Erwartungsgemäß deutlich der Unterschied zwischen den beiden Reinkulturen. In der Kokultur lässt sich nach 24 h ein um den Faktor 1,7 höherer Wert messen als nach 4 h.
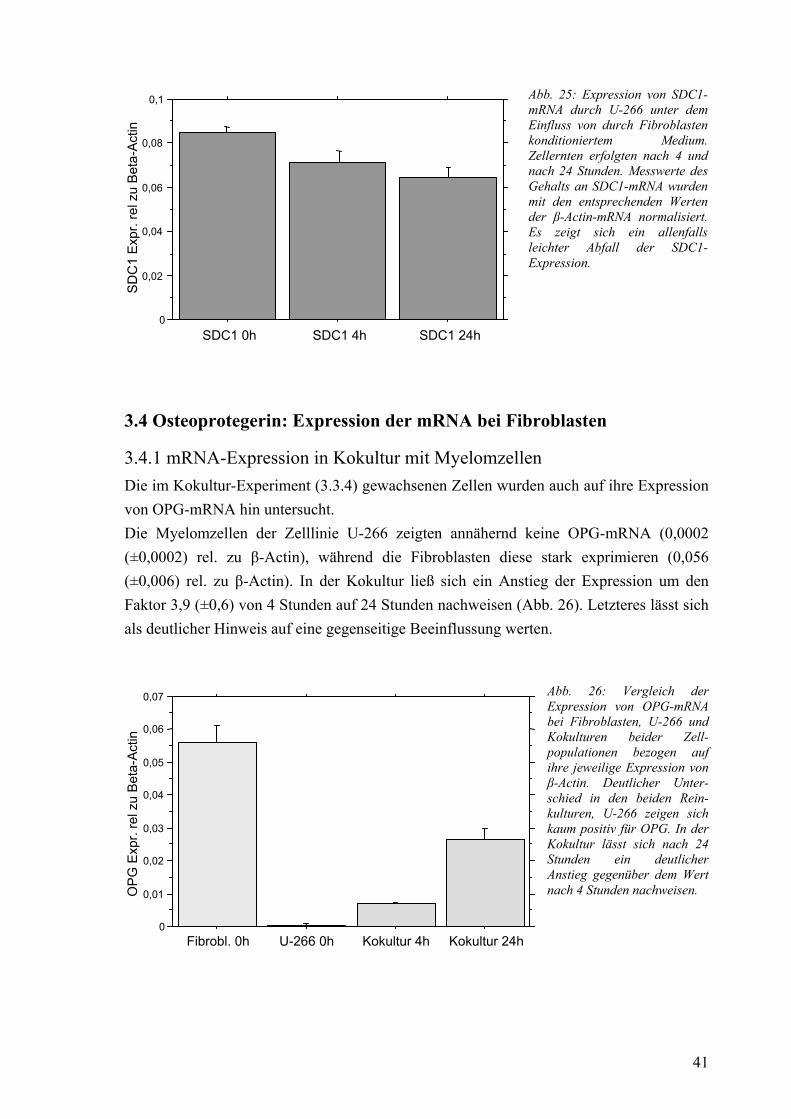
41
3.4 Osteoprotegerin: Expression der mRNA bei Fibroblasten
3.4.1 mRNA-Expression in Kokultur mit Myelomzellen Die im Kokultur-Experiment (3.3.4) gewachsenen Zellen wurden auch auf ihre Expression von OPG-mRNA hin untersucht. Die Myelomzellen der Zelllinie U-266 zeigten annähernd keine OPG-mRNA (0,0002 (±0,0002) rel. zu β-Actin), während die Fibroblasten diese stark exprimieren (0,056 (±0,006) rel. zu β-Actin). In der Kokultur ließ sich ein Anstieg der Expression um den Faktor 3,9 (±0,6) von 4 Stunden auf 24 Stunden nachweisen (Abb. 26). Letzteres lässt sich als deutlicher Hinweis auf eine gegenseitige Beeinflussung werten.
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
OP
G E
xpr.
rel z
u B
eta-
Actin
Fibrobl. 0h Kokultur 24hKokultur 4hU-266 0h
0
0,02
0,04
0,06
0,08
0,1
SDC1 0h SDC1 4h SDC1 24h
SD
C1
Expr
. rel
zu
Bet
a-Ac
tin
Abb. 26: Vergleich der Expression von OPG-mRNA bei Fibroblasten, U-266 und Kokulturen beider Zell-populationen bezogen auf ihre jeweilige Expression von β-Actin. Deutlicher Unter-schied in den beiden Rein-kulturen, U-266 zeigen sich kaum positiv für OPG. In der Kokultur lässt sich nach 24 Stunden ein deutlicher Anstieg gegenüber dem Wert nach 4 Stunden nachweisen.
Abb. 25: Expression von SDC1-mRNA durch U-266 unter dem Einfluss von durch Fibroblasten konditioniertem Medium. Zellernten erfolgten nach 4 und nach 24 Stunden. Messwerte des Gehalts an SDC1-mRNA wurden mit den entsprechenden Werten der β-Actin-mRNA normalisiert. Es zeigt sich ein allenfalls leichter Abfall der SDC1-Expression.
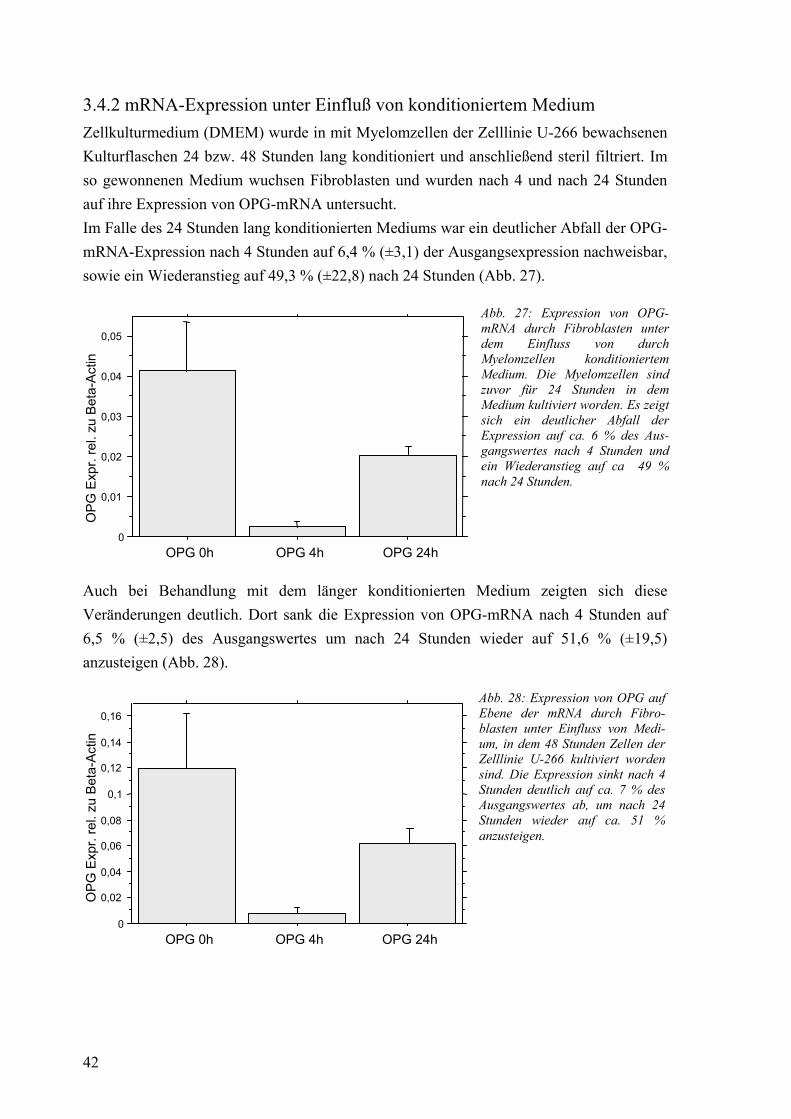
42
3.4.2 mRNA-Expression unter Einfluß von konditioniertem Medium Zellkulturmedium (DMEM) wurde in mit Myelomzellen der Zelllinie U-266 bewachsenen Kulturflaschen 24 bzw. 48 Stunden lang konditioniert und anschließend steril filtriert. Im so gewonnenen Medium wuchsen Fibroblasten und wurden nach 4 und nach 24 Stunden auf ihre Expression von OPG-mRNA untersucht. Im Falle des 24 Stunden lang konditionierten Mediums war ein deutlicher Abfall der OPG-mRNA-Expression nach 4 Stunden auf 6,4 % (±3,1) der Ausgangsexpression nachweisbar, sowie ein Wiederanstieg auf 49,3 % (±22,8) nach 24 Stunden (Abb. 27).
Auch bei Behandlung mit dem länger konditionierten Medium zeigten sich diese Veränderungen deutlich. Dort sank die Expression von OPG-mRNA nach 4 Stunden auf 6,5 % (±2,5) des Ausgangswertes um nach 24 Stunden wieder auf 51,6 % (±19,5) anzusteigen (Abb. 28).
0
0,01
0,02
0,03
0,04
0,05
OPG 0h OPG 4h OPG 24h
OP
G E
xpr.
rel.
zu B
eta-
Act
in
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
0,16
OPG 0h OPG 4h OPG 24h
OP
G E
xpr.
rel.
zu B
eta-
Act
in
Abb. 27: Expression von OPG-mRNA durch Fibroblasten unter dem Einfluss von durch Myelomzellen konditioniertem Medium. Die Myelomzellen sind zuvor für 24 Stunden in dem Medium kultiviert worden. Es zeigt sich ein deutlicher Abfall der Expression auf ca. 6 % des Aus-gangswertes nach 4 Stunden und ein Wiederanstieg auf ca 49 % nach 24 Stunden.
Abb. 28: Expression von OPG auf Ebene der mRNA durch Fibro-blasten unter Einfluss von Medi-um, in dem 48 Stunden Zellen der Zelllinie U-266 kultiviert worden sind. Die Expression sinkt nach 4 Stunden deutlich auf ca. 7 % des Ausgangswertes ab, um nach 24 Stunden wieder auf ca. 51 % anzusteigen.

43
Hier ließ sich also zeigen, dass die unter 3.4.1 beschriebene Wirkung von Myelomzellen auf die Expression von OPG-mRNA durch Fibroblasten tatsächlich vorhanden ist, dass sie offensichtlich durch einen humoralen Faktor erfolgt, und dass der Effekt dieses Faktors nur für kurze Zeit anhält. Außerdem machte es keinen Unterschied, ob die Myelomzellen nun 24 oder 48 Stunden in dem Medium kultiviert worden waren.
3.5 Receptor Activator of NF-κB Ligand Weder U-266 noch die verwendeten Fibroblasten exprimierten RANKL stark genug um es mit der hier angewandten Methode zuverlässig zu quantifizieren. Sämtliche Messwerte für RANKL lagen weit außerhalb der Standardkurven, das Verhältnis von RANKL zu β-Actin bei den Fibroblasten betrug in diesen Messungen 1:3,2*105, bei U-266 sogar mehr als 1:106, was Crossing Points von 38,5 beziehungsweise 38,7 entspricht. Dementsprechend sind die Ergebnisse der Kokulturexperimente zu RANKL wenig aussagekräftig. Auch in der Gelelektrophorese ließ sich für die PCR der Fibroblastenprobe kein Produkt nachweisen (Abb. 19, Kap. 3.1.4). Dennoch steht mit der hier etablierten Methode sicherlich ein geeignetes Instrument zur zukünftigen Untersuchung der Expression von RANKL auf Ebene der mRNA zur Verfügung.

44
4 Diskussion Als wesentliche Ergebnisse der vorliegenden Untersuchungen sind festzuhalten: 1. Die Expression von OPG-mRNA in humanen Fibroblasten wird durch Kokultur mit
Myelomzellen und durch die Kultivierung in durch Myelomzellen konditioniertem Medium signifikant supprimiert.
2. Die in diesem Zellmodell verwendeten Zellen sind negativ für die Expression von RANKL-mRNA.
3. Das polarisierte Verteilungsmuster des SDC1-Proteins auf Myelomzellen wird bestätigt. Die Expression der mRNA von SDC1 in Myelomzellen ist nicht beeinfluss-bar durch IL-6, IL-15 oder Antikörper gegen IL-15. Auch durch Fibroblasten konditioniertes Medium verändert die Expression nicht.
4.1 Untersuchungen zur Expression von OPG Kurz nach Abschluss der Experimente zur vorliegenden Arbeit erschienen zahlreiche Veröffentlichungen, die den Erkenntnisstand über das OPG/RANKL-System beim multiplen Myelom entscheidend erweitern und unsere Ergebnisse in einigen wichtigen Punkten bestätigen. Weitere Teilaspekte sind bisher nicht veröffentlicht oder wider-sprechen Resultaten anderer Gruppen.
4.1.1 Veränderung der OPG-Expression in Kokultur Die Untersuchung an den Kokulturen aus Zellen der Zelllinie U-266 und Fibroblasten präsentierte deutliche Hinweise auf eine Beeinflussung der Expression von OPG-mRNA in Fibroblasten. Der Wert nach vier Stunden entsprach nur etwa 12,2 % (±1,6) des Wertes der Fibroblasten allein, sollte er doch bei konstanter Expression 50 % betragen haben, denn U-266 und Fibroblasten wurden zu genau gleichen Teilen zusammengeführt, und die Myelomzellen sind für OPG-mRNA quasi negativ. Nach 24 Stunden war die Expression der OPG-mRNA in der Kokultur wieder um fast das vierfache gegenüber dem Wert nach vier Stunden angestiegen, was in etwa 50 % des Wertes der Fibroblasten allein entsprach. Ein direkter Ausgangswert der Kokulturen wurde in diesem Experiment nicht bestimmt, und so kann der anfänglich gemessene Abfall unter Umständen durch Zählungenauigkeiten bei der Zusammenführung der Zellen beeinflusst sein. Dies gilt jedoch nicht für den messbaren Wiederanstieg der Expression, dafür sei als einzige systematische Fehlerquelle die eventuell unterschiedliche Wachstumsgeschwindigkeit der beiden Zelllinien erwähnt. Die untersuchten Fibroblasten zeigten sich für OPG-mRNA deutlich positiv, die Zellen der Zelllinie U-266 im Gegensatz dazu annähernd negativ. Dies entspricht weitestgehend den Resultaten anderer Gruppen. So waren bei der Entdeckung des Osteoprotegerins durch

45
Tsuda et al. 1997 Fibroblasten die Quelle für das damals noch OCIF genannte OPG. Das von ihnen isolierte Protein hemmte die durch drei unterschiedliche Signalwege (Vit. D3, PTH, IL-11) stimulierte Osteoklastogenese [145]. Giuliani et al. konnten bei zehn humanen Myelomzelllinien, darunter die hier verwendete Zelllinie U-266, mittels RT-PCR keine Expression von OPG-mRNA feststellen. Durch ELISA war kein OPG-Protein in durch die Zellen konditioniertem Medium nachweisbar. Die von ihnen untersuchten Präosteoblasten und Knochenmarksstromazellen erwiesen sich demgegenüber als OPG-positiv [45]. Auch in den Experimenten von Shipman et Croucher waren alle drei verwendeten Myelom-zelllinien negativ für OPG, während Osteoblasten und Knochenmarksstromazellen deutlich positiv waren [130]. Ähnliche Untersuchungen an Kokulturen führten Pearse et al. Ende 2001 durch [104]. Sie wiesen bei Kokulturen von Myelomzellen mit Osteosarkomzellen den Rückgang der OPG-Expression nach. Zusätzlich konnte der die OPG-Expression stimulierende Effekt von TGF-1β bei den Osteosarkomzellen durch die Kokultur unterdrückt werden. Genauere quantitative Aussagen wurden nicht gemacht. Giuliani et al. untersuchten wenig später ein Zellmodell aus Knochenmarksstromazellen und den Myelomzelllinien XG-1 und XG-6 [45]. Auch sie stellten einen deutlichen Abfall der Osteoprotegerin-mRNA fest, und zwar auf 36 % nach 12 Stunden und auf 54 % nach 24 Stunden bei XG-6. Bei Kokultur mit XG-1 betrug die Konzentration nach 24 Stunden 63 % und nach 48 Stunden 48 %. Auch bei frisch von Patienten isolierten Myelomzellen ließ sich dieser Effekt nachweisen. Der hier beschriebene Wiederanstieg der Konzentration in der Zeit von 12 auf 24 Stunden bei Kokultur mit XG-1 könnte durchaus der von uns festgestellten Dynamik von 4 auf 24 Stunden entsprechen. Nach Meinung der Gruppe um Giuliani ist der die OPG-Expression unterdrückende Effekt abhängig von einem direkten Zell-zu-Zell Kontakt zwischen Knochenmarksstromazellen und Myelomzellen. Bei Untersuchungen an Kokulturen, bei denen die beiden Zellpopulationen durch zellundurchlässige Membranen getrennt waren, zeigten sich weiter oben beschriebene Effekte nicht. Auch die Behandlung der Stromazellen mit durch Myelomzellen konditioniertem Medium war bei Giuliani et al. ohne Auswirkung auf die OPG-Expression [45]. Gerade letzterem widersprechen jedoch unsere weiteren Ergebnisse (4.1.2).
4.1.2 Veränderung der OPG-Expression unter Einfluss des durch Myelomzellen konditionierten Mediums
Eine Suppression der Expression von OPG-mRNA unter Einfluss des konditionierten Mediums ließ sich deutlich nachweisen, und zwar unabhängig davon, ob das Medium 24 oder 48 Stunden konditioniert worden war. Dieser Effekt muss demnach von einem humoralen Faktor, abgegeben von den Myelomzellen, vermittelt worden sein. So ist nicht unbedingt ein direkter Zellkontakt erforderlich, damit Faktoren der Myelomzellen, die zu

46
einem Abfall der OPG-Expression in fibroblastischen Zellen führen, wirksam werden. Außerdem lässt sich der Wiederanstieg nach 24 Stunden auf etwa 50 % des Ausgangs-wertes als Zeichen einer Gewöhnung der Zellen werten, beispielsweise im Sinne einer „Down-Regulation“ der vermittelnden Rezeptoren. Diese Ergebnisse widersprechen also den Resultaten von Guiliani et al., die unter Einfluss von konditioniertem Medium keine Veränderung der OPG-Expression feststellten [45]. Jedoch erfassten sie mit einem Messzeitpunkt nach erst 24 Stunden keine besonders frühen Effekte, anders als in unserer Untersuchung nach bereits 4 Stunden. Der von uns festgestellte Wiederanstieg der Expression auf etwa 50 % des Ausgangswertes nach 24 Stunden zeigt die Problematik eines zu spät gewählten Messzeitpunktes im Falle eines Gewöhnungseffektes der Zellen. In unseren Untersuchungen wurde bisher lediglich die Aktivität eines humoralen Faktors gezeigt, und so stellt sich die Frage nach der Natur dieses Faktors. Bisher sind erst wenige Negativregulatoren der OPG-Expression bekannt, Stimulatoren dagegen reichlich (Tab. 4).
↑ ↓ IL-1α [16, 59, 150, 151] Glukokortikoide [16, 60, 108, 150] IL-1β [59] PGE2 [16, 17] TNF-α [16, 18] Forskolin FSK [16] TNF-β [18] Phorbol-12,13-Dibutyrat [16] Vit. D3 [59] BMP-2 [59] Östrogen [62] TGF-β1 [98] IL-18 [91]
Tab. 4: Stimulierende (↑) und hemmende (↓) Faktoren, die die OPG-Expression beeinflussen So konnte mit IL-1α und -1β, mit TNF-α und -β, mit Vit. D3, Östrogen und TGF-β1 die OPG-Expression durch osteoblastenartige Zellen gesteigert werden. Phorbol-12,13-Dibutyrat, ein Aktivator des Proteinkinase C Signalweges, und BMP-2 (Bone Morpho-genetic Protein-2) hatten ebenfalls diesen Effekt. Von Makiishi-Shimobayashi et al. ist auch für IL-18 ein stimulierender Einfluss nachgewiesen, dessen Signalweg wahrschein-lich über das MAPK-System verläuft [91]. Hemmende Einflüsse konnten bisher nur für Glukokortikoide und für Prostaglandin E2 nachgewiesen werden. Bei PGE2 könnte dieser Einfluss über die Proteinkinase A erfolgen, da Forskolin, ein starker Aktivator dieses Signalweges, ebenfalls zu einer Reduzierung der OPG-Expression führt. Die negative Beeinflussung der OPG-Expression und damit der Sekretion beim Plasmozytom scheint nicht nur lokal nachzuweisen zu sein. Seidel et al. zeigten 2001 in

47
einer größer angelegten Studie, dass die OPG-Spiegel im Serum von Patienten mit multiplem Myelom signifikant niedriger sind als bei Gesunden [124]. Der Grad dieser Erniedrigung war assoziiert mit dem Grad der radiografisch beurteilten Skelettdestruktion. Weiterhin waren die OPG-Spiegel assoziiert mit dem WHO-Performance-Status und Markern der Knochenneubildung. Lipton et al. und Standal et al. bestätigten 2002 die niedrigeren OPG-Spiegel bei Myelompatienten [85, 136]. Bei der Untersuchung von Patienten mit anderen malignen Erkrankungen waren interessanterweise unter Individuen mit metastasierten soliden Tumoren eher höhere Serumspiegel nachweisbar als bei gesunden Kontrollpersonen, jedoch zeigten jene Patienten mit Knochenmetastasen normale Serumspiegel. Keiner der Patienten wies aber derart hohe OPG-Spiegel auf, wie sie zur Hemmung der Osteoklastogenese nach den von Tsuda et al. [145] durchgeführten Experimenten nötig wären. Daraus ergibt sich eine rationale Basis für OPG als eine Möglichkeit zur Behandlung der Knochendestruktion beim multiplen Myelom. Im Jahr 2000 beschrieben Honore et al. die Behandlung mit OPG als eine Möglichkeit, die durch maligne Zellen ausgelösten Knochenaffektionen zu vermindern [64]. An einem murinen Modell mit Knochensarkomen zeigten sie die Verringerung von Knochendestruktion, Knochenschmerz und schmerzbedingtem, zur Chronifizierung führendem, neuro-chemischem Umbau des Rückenmarks nach der Gabe von OPG. Der von Croucher et al. 2001 verwendete Mäuseklon hatte spontan ein multiples Myelom entwickelt [33]. Auch an diesem Modell ließ sich die Knochendestruktion durch Gabe von OPG hemmen. Standal et al. gehen in der Deutung ihrer Ergebnisse noch einen Schritt weiter. Wie in Kapitel 4.3 näher erläutert, zeigten sie, dass Myelomzellen die Fähigkeit besitzen, OPG zu binden, aufzunehmen und abzubauen, und dass dabei HSPGs wie SDC1 als Bindungsstellen eine zentrale Rolle übernehmen. Die Bindungsstelle des OPG-Moleküls für SDC1 erwies sich dabei nicht als entscheidend für die RANKL-Bindung [136]. So ist als eine mögliche Zukunftsvision sicherlich die Behandlung von Knochenaffektionen bei Myelompatienten mit rekombinantem OPG, dem die Heparin bindende Domäne fehlt, in Erwägung zu ziehen.
4.2 Untersuchungen zur Expression von RANKL Die Etablierung der RANKL-Primer erfolgte an Lymphknotengewebe, von dem eine zuverlässige Expression von RANKL hinreichend bekannt ist [3, 80, 156, 162]. Ergebnisse der Etablierung zeigt Kapitel 3.1.4. Beim Vergleich der Nativ-Expression der mRNA von RANKL in Fibroblasten und Myelomzellen der Zelllinie U-266 waren die Messwerte der Expression in Fibroblasten dreifach höher, jedoch müssen diese Ergebnisse aufgrund der Normalisierung mit der Expression von β-Actin kritisch betrachtet werden. Das Verhältnis von RANKL zu β-Actin bei den Fibroblasten beträgt in diesen Messungen 1:3,2*105, bei

48
U-266 sogar mehr als 1:106, was Crossing Points von 38,5 beziehungsweise 38,7 entspricht. So wird ersichtlich, dass bei diesen Experimenten mit Verdünnungsreihen bis 1:104 die Messwerte weit unterhalb der Standardkurve liegen und deshalb wenig Aussagekraft besitzen. Ähnlich niedrige Werte können bei der hier angewandten Methode allein durch unspezifische Nebenprodukte entstehen [110]. So ist es nicht verwunderlich, dass in der gelelektrophoretischen Auftrennung weder bei Fibroblasten (Kap. 3.1.4, Abb. 19) noch bei U-266 (ohne Abb.) ein spezifisches Produkt nachweisbar war. Ein Variationskoeffizient der Messwerte von 40,8 % bei U-266 und sogar 141 % bei den Fibroblasten bestätigt die Ungenauigkeit bei derart wenigen, wenn überhaupt vorhandenen, Ausgangskopien. Die von uns verwendeten Zellen müssen demnach als negativ oder zumindest nicht signifikant positiv für die Expression von RANKL-mRNA gelten. Es besteht inzwischen kein Zweifel mehr, dass im Knochenmark von Myelompatienten nicht nur die Konzentration von OPG erniedrigt, sondern auch die von RANKL erhöht ist. Über die Quelle von RANKL herrscht jedoch bislang noch keine Einigkeit. So berichten verschiedene Autoren über RANKL-Expression durch primäre Myelomzellen von erkrankten Patienten [55, 127, 128] oder durch humane oder murine Myelomzelllinien [2, 33]. Heider et al. arbeiteten mit Antikörpern gegen RANKL, mit denen sie auf den Myelomzellen selbst die Expression des RANKL-Proteins nachwiesen [128]. In erst kürzlich veröffentlichten Ergebnissen beschreiben sie, dass diese Expression mit dem Ausmaß an osteolytischer Aktivität bei den Patienten korreliert [55]. Auf der anderen Seite ordnen Roux et al. die Expression von RANKL klar den Stromazellen zu. Auch sie verwendeten Antikörper zum Nachweis der Proteinexpression, bei ihren Experimenten bei Knochenmarksbiopsien von Myelompatienten und MGUS-Patienten [116]. Pearse et al., die vornehmlich mit PCR arbeiteten, berichten, dass Myelomzellen zur Knochen-destruktion führen, indem sie die Expression von OPG in Knochenmarksstromazellen vermindern und die von RANKL in den selben steigern. Eine Expression von RANKL auf der Ebene der mRNA durch die Myelomzellen selbst konnten sie nicht feststellen [104]. Auch Giuliani et al. demonstrierten den Einfluss von Myelomzellen auf Knochen-marksstromazellen in Kokulturexperimenten. Zusätzlich konnte auch hier für keine der verwendeten Myelomzelllinien, darunter auch die von uns verwendete Myelomzelllinie U-266, die Expression von RANKL-mRNA nachgewiesen werden [45]. Demnach ordnet sich unser Ergebnis in Bezug auf U-266 in letztere Gruppe ein. In Bezug auf Fibroblasten widerspricht unser Ergebnis anderen Autoren. Nach Giuliani et al., Roux et al. und Pearse et al. könnte bei Fibroblasten eine Expression von RANKL erwartet werden [45, 104, 116], da Fibroblasten nahe Verwandte der Knochenmarksstromazellen sind. Zusätzlich wiesen Quinn et al. die Expression von RANKL-mRNA auch bei dermalen Fibroblasten selbst nach [108].

49
Eine weitere, bisher unerwähnte Rolle bei der Expression von RANKL scheinen T-Lymphozyten zu haben. Giuliani et al. konnten unter dem Einfluss vom Myelomzellen eine erhöhte Expression von RANKL-mRNA in aktivierten T-Lymphozyten feststellen. Außerdem wiesen sie im Knochenmark von Myelompatienten erhöhte Konzentrationen von IL-7 nach. Ihrer Vermutung nach geben Myelomzellen unter Einfluß von IL-6 dieses IL-7 ab, stimulieren damit T-Zellen zur RANKL-Expression und liefern so einen weiteren Anstoß zur Knochendestruktion [47]. Die Wechselwirkungen der verschiedenen Zellen im Knochenmarksmikromilieu scheinen also verzweigter zu sein, als anfänglich erwartet, und die Frage nach der Herkunft von RANKL in Osteolyseherden beim Multiplen Myelom entwickelte sich zur Jahreswende 2003/04 zur Kernfrage wissenschaftlicher Auseinandersetzungen [46].
4.3 Untersuchungen zur Expression von SDC1 Wie eingangs erwähnt, sequestriert SDC1 das Zytokin HGF. HGF scheint über die Induktion der IL-11-Produktion durch Osteoblasten die Knochendestruktion voranzu-treiben [143] und über die Stimulation des PI3-Kinase/PKB- und ras/raf/MAPK-Signalweges in Myelomzellen Überleben respektive Proliferation der Tumorzellen zu begünstigen [37]. Dabei wirkt SDC1 als Korezeptor verstärkend mit. Seidel et al. zeigten das sich die IL-11 induzierende Wirkung von HGF auf Saos-2 Zellen durch Zugabe von SDC1 auf fast 200 % steigern ließ. Allerdings verwendeten sie die lösliche Form des SDC1 [123]. Derksen et al. untersuchten in Myelomzellen direkt den durch HGF stimulierten Signalweg über Met, die Rezeptortyrosinkinase des HGF. Sie wiesen eine deutlich reduzierte Tyrosinphosphorylierung von Met nach, wenn die Heparansulfatketten des membranständigen SDC1 spezifisch enzymatisch entfernt worden waren. Außerdem wurde dadurch die Aktivierung von Downstream-Effektormolekülen des Met/HGF-Signalweges stark gehemmt, darunter die aktive PKB und die MAP Kinasen Erk1 und Erk2, indikativ für antiapoptotische und proliferative Effekte von HGF. Ein von diesen Autoren entworfenes Modell der Wechselwirkung von SDC1 mit dem HGF/Met-Komplex beschreibt eine „ménage a trois“ zwischen den drei Komponenten und kann sicherlich als ein Modell für die Wirkungsweise von SDC1 gelten [37]. Die Bindung von OPG an das SDC1 auf Myelomzellen hat demgegenüber die Internalisierung und den Abbau des OPGs zur Folge, was das in dieser Arbeit vornehmlich untersuchte OPG/RANK/RANKL-System in Richtung Knochendestruktion verschiebt. Standal et al. konnten in ihren Experimenten mit Fluoreszenzfarbstoff-markiertem OPG an lebenden Zellen der Myelomzelllinie CAG dieses Phänomen der Internalisierung und Degradierung nachweisen. Ihrer Meinung nach wirkt das im Überfluß vorhandene OPG als Puffer für Veränderungen in der RANKL-Expression. Ein Verlust dieser Pufferkapazität durch verstärkten Abbau oder verminderte Produktion des OPG könnte die Knochen-

50
homöostase schon für kleinste Änderungen der RANKL-Expression sehr empfindlich machen [136]. Daher war es für uns interessant, die Lokalisation und die Expression von SDC1 in unserem Zellmodell genauer zu analysieren. Auch der Einfluss zweier für das Myelomzellwachstum wichtiger Zytokine auf die Expression sollte untersucht werden.
4.3.1 U-266 bietet polarisiertes Verteilungsmuster von SDC1 Das im Jahre 2000 von Børset et al. beschriebene Verteilungsmuster von SDC1 auf Myelomzellen ließ sich bei der Zelllinie U-266 bestätigen [15]. Der Anteil der Zellen mit polarisiertem Verteilungsmuster bei U-266 wurde dort mit 13 % angegeben. Trotzdem in unseren Untersuchungen keine größeren Zellzahlen ausgezählt wurden, entspricht dies in etwa dem von uns abgeschätzten Verhältnis von 1:10. Børset et al. konnten zeigen, dass, wenn Uropodien ausgebildet sind, SDC1 spezifisch dort zu finden ist, während bei den anderen Zellen eine gleichmäßige Verteilung auf der Oberfläche stattfindet. Weiterhin korrelierte die Fähigkeit der Myelomzellen zur aktiven Migration mit dem Anteil des polarisierten Phänotyps. Die Uropodien scheinen an Zell-Zell-Kontakten der Myelom-zellen untereinander sowie zu anderen Zellen entscheidend beteiligt zu sein, und das dort vorhandene SDC1 sequestriert Heparin bindende Moleküle wie HGF oder OPG [15]. Weitere Untersuchungen der gleichen Gruppe brachten die überraschende Erkenntnis, dass die Verschiebung des SDC1 auf der Zelloberfläche zu den Uropodien entscheidend von den Heparansulfatketten abzuhängen scheint, und zwar spezifisch von denen des Syndecans. So führte die Zugabe von Heparin oder einer Heparitinase zu polarisierten Myelomzellen oder lymphoblastoiden Zellen zu einer raschen gleichmäßigen, also depolarisierten Verteilung des SDC1 auf der Zelloberfläche. Desweiteren fanden sich andere HSPGs wie Glypican-1 oder Betaglycan nie an den Uropodien konzentriert, was dafür spricht, dass in den Heparansulfatketten des SDC1 vorhandene, einzigartige Strukturmotive für die spezifische Umlagerung verantwortlich sind [160].
4.3.2 SDC1-mRNA-Expression ist bei U-266 siebenmal stärker als bei IM-9 Obwohl sich die malignen Plasmazellen leicht durch ihre Expression von CD38 und SDC1 identifizieren lassen, ist ihre Aufreinigung und ihre in vitro Vermehrung immer noch schwierig. Das führte zur Etablierung von Myelomzelllinien, die heute immer noch essentiell sind für die Erforschung der Biologie des Multiplen Myeloms. 1977 zeigte Nillson, dass die Etablierung kontinuierlicher Zelllinien aus dem Knochenmark oder dem peripheren Blut von Myelompatienten zu zwei völlig unterschiedlichen Typen von Zelllinien geführt hatte. Zum einen handelt es sich um lymphoblastoide Zelllinien, die durch Immortalisierung durch Epstein-Barr-Virus aus nichtmalignen B-Zellen entstehen, zum anderen um echte Myelomzelllinien, die die Immortalität von Tumorzellen zeigen und

51
das gleiche Ig-Gen-Rearrangement wie die frischen Tumorzellen besitzen [101]. Aufgrund ihrer verschiedenen Eigenschaften dürfen diese beiden Typen von Zelllinien keinesfalls undifferenziert in Studien zum Multiplen Myelom verwendet werden. In Untersuchungen des European Myeloma Resarch Network zur Identifikation geeigneter Unter-scheidungsparameter fanden sich für lymphoblastoide Zellinien CD19, CD20, Oberflächen-Ig und die Adhäsionsmoleküle CD11a und CD49e, für echte Myelomzell-linien CD38, SDC1 und CD28. Für die Kennzeichen des jeweils anderen Typs waren die Zelllinien weitgehend negativ. Für SDC1 waren aber auch die lymphoblastoiden Zellen leicht positiv [106]. In unseren Untersuchungen exprimierte die Myelomzelllinie U-266 die mRNA von SDC1 etwa siebenmal stärker als die lymphoblastoide Zelllinie IM-9. Quantitative Ergebnisse hierzu finden sich in der Literatur zwar nicht, im semiquantitativen Vergleich lässt sich dieses Ergebnis jedoch gut einordnen. Bei Pellat-Deceunynk et al. ist die lymphoblastoide Zelllinie IM-9 für SDC1 nur leicht positiv (-/+), die Myelimzelllinie U-266 dem gegenüber deutlich positiv (+++) [106]. Gooding et al. beschreiben die lymphoblastoiden Zelllinien ARH-77 und HS-Sultan beide als SDC1-negativ (Expression kleiner 10 %), während sich auch hier die Zelllinie U-266 deutlich positiv (Expression 50-90 %) darstellt [48]. In letzterer Untersuchung konnten auch die kennzeichnenden Parameter CD19 und CD20 für die lymphoblastoiden Zellen bestätigt werden. Spezifisch für die echten Myelomzelllinien waren hier SDC1 und CD56 [48].
4.3.3 IL-6, IL-15 oder IL-15AK bewirken keine Veränderung der SDC1-mRNA-Expression und auch bei Behandlung mit konditioniertem Medium bleibt die SDC1-mRNA-Expression konstant
Veränderungen der Expression von SDC1 unter Einfluss von IL-6, IL-15 oder Antikörpern gegen IL-15 konnten auf der Ebene der mRNA bei keiner der beiden Zelllinien festgestellt werden. Auch bei der Behandlung mit durch Fibroblasten konditioniertem Medium konnte keine signifikante Veränderung der SDC1-mRNA-Expression nachgewiesen werden. Die Ergebnisse zu der Wirkung von IL-6 auf die mRNA-Expression des SCD1 entsprechen den Aussagen von Sneed et al., die den Effekt dieses Zytokins an murinen B-Lymphozyten untersucht hatten. Sie beschreiben zwar einen deutlichen Rückgang der SDC1-Protein-Expression unter IL-6 auf 40 % bei 10 U/ml, was etwa den von uns eingesetzten 0,24 ng/ml entspricht, und auf 20 % bei 1000 U/ml, räumten aber ein, dass die Expression der SDC1-mRNA unverändert bliebe. Es sei davon auszugehen, dass die Reduktion der Expression durch IL-6 durch posttranskriptionale Mechanismen bewirkt werde [132]. Auch bei den von Day et al. untersuchten Darmepithelzellen war es wie bei den in der vorliegenden Arbeit untersuchten Myelomzellen durch IL-6 zu keiner signifikanten Änderung der SDC1-mRNA-Expression gekommen. Es fand lediglich eine kaum

52
merkliche Steigerung der Sekretion von löslichem Syndecan in das Medium statt [35]. Sebestyén et al. beschreiben die Wirkung von IL-6 auf die Expression von SDC1 bei anderen Zellarten. Unter IL-6-Einfluß zeigte sich hier ein Rückgang der SDC1-Expression bei Rattenhepatozyten, Hepatomzellen und humanen Hautfibroblasten, eine verstärkte Expression hingegen bei Rattenleberfibroblasten [122]. Über die Wirkung des erst 1994 identifizierten IL-15 auf die SDC1-Expression bei Myelomzellen oder anderen Zellen ist in der eingängigen Literatur bisher nichts beschrieben, bekam es doch erst im Jahr 2000 durch die Entdeckung einer autokrinen Schleife durch Tinhofer et al. einen Bezug zum Multiplen Myelom [142]. Auch über Experimente mit durch Fibroblasten konditioniertem Medium ist bisher nicht berichtet worden. Die Wirkungen anderer Zytokine wie TNFα, IL-1β, IFNγ und TGFβ1 auf verschiedene Zelltypen sind bei Sebestyén et al. ebenfalls beschrieben (Tab. 5) [122]. Weiterhin berichten Day et al. über einen Abfall der SDC1-Expression in den Epithelzellen unter TNFα und IL-1β. Außerdem wirkte sich die Kokultur mit PBMCs (peripheral blood mononuclear cells) negativ auf die Expression aus [35]. In einer Arbeit von Kovalszky et al. wird die Expression von SDC1 in von Kupffer´schen Sternzellen durch Endotoxin und IFNγ induziert [74] (Tab. 5).
↓ ↑ TNFα Ep [35] Hom [122]
TNFα RHep [122]
IL-1β Ep [35] RHep, Hom [122]
IL-1β RLFib [122]
IL-6 BLym [132] RHep, Hom, HFib [122]
IL-6 RLFib [122]
IFNγ Hom [122]
IFNγ, Endotoxin vKuSZ [74]
PBMCs Ep [35]
TGFβ1 RHep, Hom, RLFib [122]
Die dargestellten Daten deuten darauf hin, dass der regulatorische Effekt auf ein spezifisches HSPG in nicht unerheblichem Maße vom Zelltyp abhängig ist. Die regulatorische Region des SDC1-Gens enthält Konsensussequenzen für zahlreiche
Tab. 5: Die SDC1-Expression stimulierende (↑) und hemmende (↓) Faktoren, untersucht an Darmepithelzellen (Ep), Hepatomzelllinien (Hom), Rattenhepato-zyten (RHep), Rattenleberfibroblasten (RLFib), humanen Hautfibroblasten (HFib), murinen B-Lymphozyten (BLym) und von Kupffer´schen Sternzellen (vKuSZ)

53
Transkriptionsfaktorbindungsstellen wie Antennapedien, MyoD(E-Box), NF-κB, Sp1(GC-Box), C/EPBs bindene Sequenzen und CAAT- und TATA-ähnliche Strukturen [152]. So dürfte der Effekt verschiedener Zytokine in verschiedenen Zellen von der aktuellen Aktivierung bestimmter Transkriptionfaktoren abhängen. All das spricht gegen die Klassifizierung bestimmter Zytokine als stimulierend oder inhibierend in Bezug auf die Regulation von HSPGs. Bei den hier verwendeten Zelllinien ließ sich für IL-6 und IL-15 keine Beeinflussung der SDC1-mRNA-Expression nachweisen. Demnach scheint diese unabhängig von den autokrinen Schleifen des IL-6 [68] und IL-15 [142] zu sein.
4.3.4 SDC1-mRNA-Expression in Kokultur von U-266 und Fibroblasten Es konnte gezeigt werden, dass U-266 die SDC1-mRNA etwa um den Faktor 23 stärker exprimieren als die von uns verwendeten Fibroblasten. Weiterhin war in der Kokultur nach 24 Stunden ein um den Faktor 1,7 höherer Wert der SDC1-mRNA-Expression als nach 4 Stunden zu messen. Ersteres sind Werte aus Reinkulturen, zur Vergleichbarkeit normalisiert auf die Expression von β-Actin. Fehlerquelle ist hier also eine möglicherweise von Natur aus unterschiedliche Expression von β-Actin in den beiden unterschiedlichen Zelltypen. Bei den Kokulturen stand ein geeignetes Trennverfahren, das den Ansprüchen der PCR genügt, nicht zur Verfügung. Es wurden deshalb, wie auch bei Giuliani in den Experimenten zur OPG-Expression [45], Gesamtexpressionen bestimmt. Ist dabei nur eine der beiden Zellarten für die zu untersuchende mRNA positiv, so ist, neben Meß- und Mischungenauigkeiten, als einzige Fehlerquelle die eventuell unterschiedliche Wachstums-geschwindigkeit anzusehen. Diese würde durch die Normalisierung auf die Expression von β-Actin zu Verzerrungen führen. Auch die Möglichkeit, dass die eine Zellart erst die Expression einer mRNA bei der anderen induziert darf hier nicht unerwähnt bleiben. Sind beide Zellarten für die zu untersuchende mRNA positiv, können dementsprechend Veränderungen nicht mehr eindeutig einer Zellart zugeordnet werden. Ein Anstieg der Expression von SDC1-mRNA im Zeitraum zwischen 4 und 24 Stunden um den Faktor 1,7 kann also sowohl den Fibroblasten als auch den Myelomzellen zuzuordnen sein. Angesichts des geringen Effekts und der Uneindeutigkeit dieses Ergebnisses, sollte die Aussagekraft dieses Experiments also nicht überschätzt werden.
4.4 Diskussion der Methode Die hier vorliegende Arbeit stellt eine geeignete Methode zur semiquantitativen Untersuchung einiger an der Knochendestruktion beim Multiplen Myelom beteiligter Faktoren vor. Im speziellen wurde eine Methode der semiquantitativen RT-PCR für Syndecan 1 (SDC1, CD138), Osteoprotegerin (OPG) und Receptor Activator of NF-κB Ligand (RANKL) etabliert. Weitere Untersuchungen mit dieser Methode wurden an einem

54
Zellmodell aus primären Fibroblasten und Myelomzellen durchgeführt. Im Folgenden soll nicht der gesamte Arbeitsprozess hinterfragt werden, sondern auf einige kritische Punkte eingegangen werden. Einerseits gibt es in letzter Zeit kritische Stimmen zum Thema β-Actin als Housekeeping-Gen, andererseits interressiert bei einer Methode die Reproduzierbarkeit der Ergebnisse.
4.4.1 Das Problem Beta-Actin als Housekeeping-Gen Um mit der Methode der RT-PCR Genexpressionen quantifizieren zu können, werden interne Standards gebraucht. Die im Gegensatz zu absoluten Standards auf so genannten Housekeeping-Genen basierenden relativen Standards verbannen die RT-PCR dabei auf ein nur semiquantitatives Level. Trotzdem bietet dieser recht einfache Ansatz mit endogenen internen Standards einige praktische Vorteile, wenn eine absolute Quantifizierung nicht erforderlich ist. Da das Verhältnis von Standard- zu Zieltranskripten in einer RNA-Probe konsistent bleibt, ist beispielsweise die Menge an Ausgangs-RNA nicht entscheidend, die Probe wird rechnerisch normalisiert. Auch ist es nicht erforderlich, die Proben mit einer immer genau gleichen Menge an Kontrollsubstanz zu versehen. Werden Housekeeping-Gene als endogene interne Standards verwendet, ist nur das Primer-Set nötig, dessen Reaktionsbedingungen zu denen des Primer-Sets der Zielsequenzen passt. Der Gebrauch solcher Gene als interne Standards erfolgt unter der Prämisse, dass sie eine konstante basale Expression zeigen, die nicht reguliert wird und unabhängig vom Zellzyklus ist. Einer der am weitesten verbreiteten internen Standards in der Molekularbiologie ist β-Actin. In den letzten Jahren zeigten sich jedoch Hinweise, dass einige der bisher verwendeten Housekeeping-Gene gewisse Grundforderungen an interne Standards nicht erfüllen. Einige kritische Arbeiten zu β-Actin als Housekeeping-Gen sollen an dieser Stelle nicht unerwähnt bleiben. So zeigten Selvey et al. eine dosisabhängige Hemmung der β-Actin-mRNA-Expression in murinen Fibroblasten durch die Behandlung mit Matrigel, einer Extrazellularsubstanz aus Rattentumoren. Sie überprüften die Ergebnisse einer RT-PCR, in der die Expression einer Metalloprotease unter Einfluß von Matrigel erfaßt werden sollte, durch anschließende elektrophoretische Auftrennung der PCR-Produkte und laserphotometrische Quanti-fizierung. Bei reiner Betrachtung der Ergebnisse der PCR wurde durch einen Abfall der β-Actin-Expression ein Anstieg der Expression der Metalloprotease vorgetäuscht. Laserphotometrisch blieb die Expression letzterer konstant. In einem weiteren Versuch unter Verwendung von 18S rRNA als internem Standard konnte diese Konstanz bestätigt werden [125]. Für β-Actin wurde von Cornelius et al. schon 1988 ein Abfall der mRNA um 50 % nachgewiesen, wenn Präadipozyten der Zelllinie 3T3-L1 zu Adipozyten

55
ausdifferenzieren [32]. Auch Serazin-Leroy et al. bemerkten den niedrigeren Gehalt an β-Actin-mRNA in Adipozyten im Vergleich zu Präadipozyten [126]. Diese kritischen Stimmen können und sollen sicherlich nicht zahllose Ergebnisse in Frage stellen, viele nicht unbedeutende der hier erwähnten Arbeiten basieren auf der Verwendung von β-Actin als internem Standard [45, 104, 122], dennoch ist die Suche nach dem idealen internen Standard nicht beendet. 18S rRNA scheint in einigen Punkten den Forderungen an einen idealen internen Standard ein Stück näher zu kommen und ein konsistentes Maß für den Gehalt an Gesamt-RNA in einer Probe während des gesamten Zellzyklus zu sein [125].
4.4.2 Betrachtung der Reproduzierbarkeit Die Reproduzierbarkeit einer quantitativen PCR hängt von vielfältigen Faktoren ab, beginnend mit der Präparation der Proben bis zur Amplifikation und Detektion. Weitere Einflüsse sind beispielsweise die statistische Verteilung (Poisson´sches Gesetz) bei der Arbeit mit sehr niedrigen Kopienzahlen, was dann starken Einfluß auf Präparation und Amplifikation hat. Präzision des Pipettiervorgangs, vor allem kritisch bei kleinen Volumina, Effizienz der PCR und das Instrumentarium für die Detektion sind weitere kritische Determinanten für die Reproduzierbarkeit. In einer Arbeit von Betzl et al., veröffentlicht im firmeneigenen Journal „Biochemica“ der Firma Roche, wird auf letzteres genauer eingegangen. In einem ersten experimentellen Ansatz wurde die Sequenz der vorher in ein Plasmid geklonten humanen TNFα-cDNA als Zielsequenz verwendet. Die Differenzierung der Ausgangskonzentrationen von 5000, 10000 und 25000 gelang in einem PCR-System mit einem 131 bp langem Amplifikat mit einem Variations-koeffizienten von 4,3 %, 6,6 % und 7,2 %. In einem zweiten System mit einer Amplifikatlänge von 385 bp wurden Variationskoeffizienten von 13,2 %, 8,9 % und 7,9 % gemessen. SYBR Green I und Hybridisierungssonden lieferten sehr ähnliche Reproduzier-barkeiten. In einem zweiten Ansatz wurde anhand des 131 bp-Systems die Repro-duzierbarkeit mit Hybridisierungssonden von 10 bis 10000 Ausgangskopien bestimmt. Bei nur 10 Ausgangskopien lag der Variationskoeffizient bei 41,2 % [12]. Bei sehr wenigen Ausgangskopien empfiehlt sich die Korrektur der Daten anhand der Schmelzkurve. Diese Methode ist Bestandteil der LightCycler Data Analysis Software und subtrahiert die Fluoreszenzanteile unzpezifischer Nebenprodukte vor allem bei weniger als 1000 Ausgangskopien [110]. Die von uns durchgeführten Experimente an U-266 mit der Zielsequenz des SDC1 (3.3.3) hatten für β-Actin einen durchschnittlichen Variationskoeffizienten von 6,6 %, für SDC1 8,5 %. Bei den gleichen Untersuchungen an der Zelllinie IM-9 (3.3.2) ergab sich für β-Actin ein durchschnittlicher Variationskoeffizient von 7,6 % und für SDC1 13,9 %. Bei
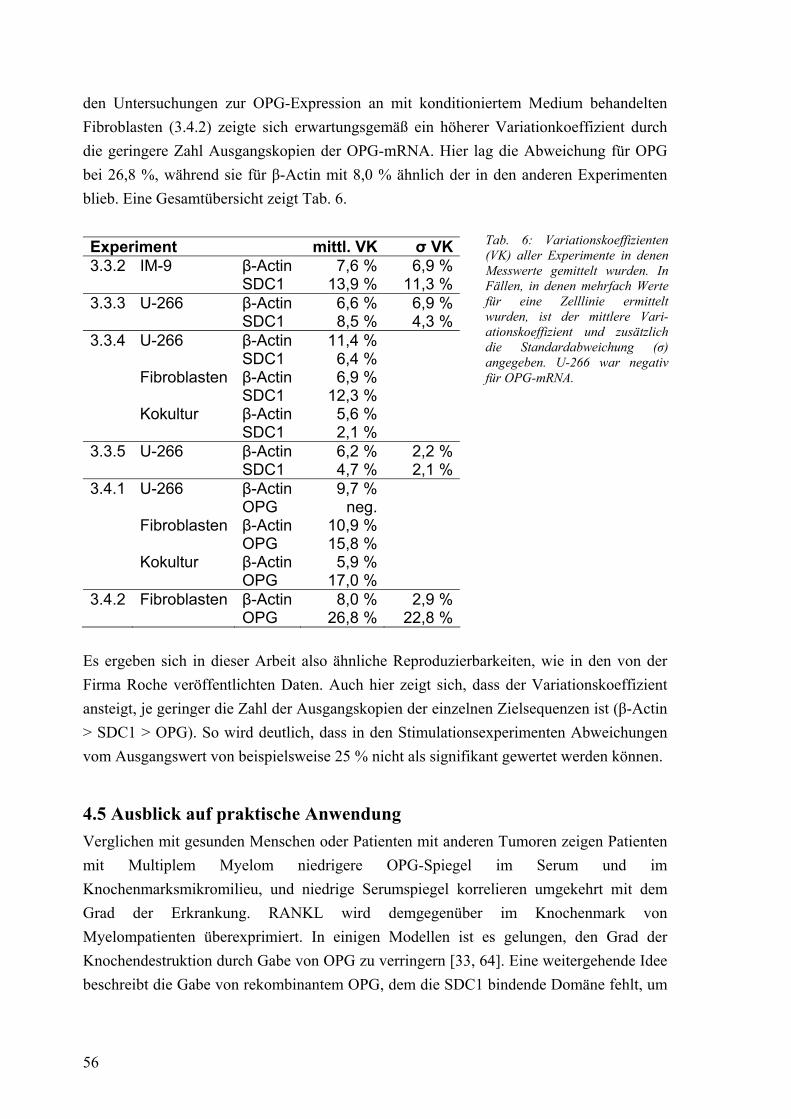
56
den Untersuchungen zur OPG-Expression an mit konditioniertem Medium behandelten Fibroblasten (3.4.2) zeigte sich erwartungsgemäß ein höherer Variationkoeffizient durch die geringere Zahl Ausgangskopien der OPG-mRNA. Hier lag die Abweichung für OPG bei 26,8 %, während sie für β-Actin mit 8,0 % ähnlich der in den anderen Experimenten blieb. Eine Gesamtübersicht zeigt Tab. 6. Experiment mittl. VK σ VK3.3.2 IM-9 β-Actin 7,6 % 6,9 % SDC1 13,9 % 11,3 %3.3.3 U-266 β-Actin 6,6 % 6,9 % SDC1 8,5 % 4,3 %3.3.4 U-266 β-Actin 11,4 % SDC1 6,4 % Fibroblasten β-Actin 6,9 % SDC1 12,3 % Kokultur β-Actin 5,6 % SDC1 2,1 %3.3.5 U-266 β-Actin 6,2 % 2,2 % SDC1 4,7 % 2,1 %3.4.1 U-266 β-Actin 9,7 % OPG neg. Fibroblasten β-Actin 10,9 % OPG 15,8 % Kokultur β-Actin 5,9 % OPG 17,0 %3.4.2 Fibroblasten β-Actin 8,0 % 2,9 % OPG 26,8 % 22,8 %
Es ergeben sich in dieser Arbeit also ähnliche Reproduzierbarkeiten, wie in den von der Firma Roche veröffentlichten Daten. Auch hier zeigt sich, dass der Variationskoeffizient ansteigt, je geringer die Zahl der Ausgangskopien der einzelnen Zielsequenzen ist (β-Actin > SDC1 > OPG). So wird deutlich, dass in den Stimulationsexperimenten Abweichungen vom Ausgangswert von beispielsweise 25 % nicht als signifikant gewertet werden können.
4.5 Ausblick auf praktische Anwendung Verglichen mit gesunden Menschen oder Patienten mit anderen Tumoren zeigen Patienten mit Multiplem Myelom niedrigere OPG-Spiegel im Serum und im Knochenmarksmikromilieu, und niedrige Serumspiegel korrelieren umgekehrt mit dem Grad der Erkrankung. RANKL wird demgegenüber im Knochenmark von Myelompatienten überexprimiert. In einigen Modellen ist es gelungen, den Grad der Knochendestruktion durch Gabe von OPG zu verringern [33, 64]. Eine weitergehende Idee beschreibt die Gabe von rekombinantem OPG, dem die SDC1 bindende Domäne fehlt, um
Tab. 6: Variationskoeffizienten (VK) aller Experimente in denen Messwerte gemittelt wurden. In Fällen, in denen mehrfach Werte für eine Zelllinie ermittelt wurden, ist der mittlere Vari-ationskoeffizient und zusätzlich die Standardabweichung (σ) angegeben. U-266 war negativ für OPG-mRNA.

57
den Abbau des OPG durch Myelomzellen zu verringern [136]. Durch RANKL-Blockade kann so die Osteoklastogenese und Osteolyse wirksam gehemmt werden. Der blockierende Effekt von OPG könnte dabei auch durch OPG- oder RANK-Fusionsproteine nachgeahmt werden, wie Versuche mit der extrazellulären Domäne von RANK, fusioniert mit der Fc-Domäne menschlichen Immunglobulins, zeigen [133]. Eine andere Möglichkeit böte die Blockade von RANK, des Rezeptors auf Osteoklasten. All diese Ansätze haben zur Zeit noch experimentellen Charakter oder Studiencharakter. So bleibt abzuwarten, was die Zukunft durch nach biochemischem Verständnis konstruierte Therapeutika an Beeinflussung des Krankheitsverlaufes und vor allem an Verbesserung der Lebensqualität für die betroffenen Patienten bringt.

58
Zusammenfassung Das multiple Myelom ist charakterisiert duch osteolytischen Befall der Knochen, der zu starken Schmerzen und Spontanfrakturen führt. Erst in den letzten Jahren begann man, die Biochemie dieser Osteolysen in Grundzügen zu verstehen. Ein für die Balance zwischen Osteogenese und Osteolyse wichtiges System besteht aus einem Rezeptor, receptor activator of NF-κB (RANK), seinem Ligand (RANKL) und dem löslichen Atrappenrezeptor Osteoprote-gerin (OPG). Die Bindung von RANKL an RANK führt zu Osteoklastogenese und Osteolyse, wohingegen die Bindung an OPG die Wirkung von RANKL inaktiviert. Myelomzellen scheinen das Knochenmarksmikromilieu in einer Weise zu stören, dass dieses Gleichgewicht in Richtung Osteolyse abgleitet. Einerseits kann Syndecan 1 (SDC1), ein transmembranes Heparansulfatproteoglycan auf Myelomzellen, OPG sequestrieren und zu Internalisierung und Abbau von OPG in Myelomzellen führen. Andererseits wird angenommen, dass Myelom-zellen durch die Beeinflussung der Knochenmarksstromazellen, der Hauptquelle für OPG und RANKL im Knochenmark, ein Ungleichgewicht im OPG/RANKL System induzieren. Diese Arbeit beschreibt eine suffiziente Methode zur Untersuchung der Expression von OPG, RANKL und SDC1 auf der Ebene der mRNA, basierend auf der semiquantitativen Realtime RT-PCR. Unter Anwendung dieses Verfahrens wird gezeigt, dass die Behandlung von humanen Vorhautfibroblasten mit durch Myelomzellen konditioniertem Medium und Kokultur von Fibroblasten mit Myelomzellen zu einem signifikanten Abfall der Expression von OPG-mRNA in den Fibroblasten führt. Dies zeigt, dass Myelomzellen Einfluss nehmen auf die OPG-Expression in Fibroblasten, hier als Modell für Zellen des Knochenmarksstromas, dass sie das OPG/RANKL-System empfindlich stören und so die Knochendestruktion vorantreiben. Im Gegensatz zur Meinung anderer Autoren geschieht dies über einen löslichen Faktor, wie durch den experimentellen Ansatz mit konditioniertem Medium demonstriert wird. Weiterhin sind U-266 und Fibroblasten nativ und in Kokultur negativ für RANKL. Die Expression der SDC1-mRNA in den Myelomzelllinien U-266 und IM-9 wird nicht beeinflusst durch die Verstärkung der autokrinen IL-6-Schleife oder die Verstärkung oder Unterbrechung der autokrinen IL-15-Schleife. Des Weiteren verändert auch die Behandlung von U-266 mit durch Fibroblasten konditioniertem Medium oder die Kokultur von Myelomzellen und Fibroblasten die SDC1-mRNA-Expression nicht signifikant. Die polarisierte Expression des SDC1-Proteins auf der Oberfläche von U-266 kann durch Immunfluoreszenzfärbung bestätigt werden. Zytokine und Zellkontakt in der Interaktion zwischen Myelomzellen und mesenchymalen Zellen lassen im Mikromilieu von Myelominfiltraten das Gleichgewicht des Knochens in Richtung Osteolyse abgleiten. Dafür scheint hauptsächlich eine Störung des OPG/RANKL-Systems verantwortlich zu sein. Die Expression von SDC1 durch Myelomzellen wird durch mesenchymale Zellen nicht verändert und ist unabhängig von der autokrinen Schleife von IL-6 oder IL-15.

59
Summary Myeloma is characterized by bone disease leading to severe bone pain and pathologic fractures. Only over the last few years the biochemistry of osteolysis in multiple myeloma has become clearer. A basic cytokine network responsible for balancing ostgenesis and osteolysis has been described consisting of a receptor called receptor activator of NF-κB (RANK), its ligand (RANKL) and the soluble decoy receptor osteoprotegerin (OPG). Binding of RANKL to RANK leads to osteoclastogenesis and osteolysis whereas binding to OPG inactivates RANKL. Myeloma cells seem to disturb bone marrow microenvironment in a manner tipping the balance towards osteolysis. On one hand syndecan 1 (SDC1), a transmembrane heparan sulfate proteoglycan on myeloma cells, is able to sequester OPG and to mediate internalization and degradation of OPG in myeloma cells. On the other hand, myeloma cells are thought to imbalance the OPG/RANKL system by influencing bone marrow stromal cells, the major source of OPG and RANKL in bone marrow microenvironment. This thesis presents a sufficient procedure for investigating expression of OPG, RANKL and SDC1 on the mRNA level based on semiquantitative realtime RT-PCR. Using this method we show that treatment of human foreskin fibroblasts with myeloma cell contitioned media and fibroblast/myeloma cell coculture both lead to a significant decrease in expression of OPG mRNA in fibroblasts. These data indicate that myeloma cells influence OPG expression in fibroblasts as a model for bone marrow stromal cells and, thus, disrupt RANKL/OPG cytokine axis to trigger bone destruction. In contrast to the published literature, this happens through a soluble factor as demonstrated by the conditioned media experiments. Moreover fibroblasts and U-266 both are negative for RANKL even if grown in coculture. The levels of SDC1 mRNA in myeloma cell lines U-266 and IM-9 are not affected by enhancing the autocrine IL-6 loop or by enhancing or interrupting the autocrine IL-15 loop. Furthermore treatment of U-266 with fibroblast conditioned media or myeloma cell/fibroblast coculture does not alter expression of SDC1 mRNA significantly. A polarized expression of SDC1 protein on U-266 cell surface can be confirmed by immune fluorescence staining. Cytokine and contactmediated interactions between myeloma cells and mesenchymal cells shift the balance towards osteolysis within the microenvironment of myeloma infiltrates. Disruption of the OPG/RANKL cytokine axis seems to be of major importance. Expression of SDC1 by myeloma cells is not affected by mesenchymal cells and is independent of IL-6 or IL-15 autocrine loop.

60
5 Literatur
1. Proposed standard nomenclature for new tumor necrosis factor family members involved in the regulation of bone resorption. The American Society for Bone and Mineral Research President's Committee on Nomenclature. J Bone Miner Res, 15(12): p. 2293-6, 2000
2. Altamirano, C. V., Neeser, J. A., and Manyak, S., Malignant multiple myeloma cells expressing RANKL induce the formation of TRAP positive multinucleated cells (abstract). Blood, 98: p. 637a, 2001
3. Anderson, D. M., Maraskovsky, E., Billingsley, W. L., Dougall, W. C., Tometsko, M. E., Roux, E. R., Teepe, M. C., DuBose, R. F., Cosman, D., and Galibert, L., A homologue of the TNF receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature, 390(6656): p. 175-9, 1997
4. Armitage, R. J., Macduff, B. M., Eisenman, J., Paxton, R., and Grabstein, K. H., IL-15 has stimulatory activity for the induction of B cell proliferation and differentiation. J Immunol, 154(2): p. 483-90, 1995
5. Asosingh, K., De Raeve, H., Van Riet, I., Van Camp, B., and Vanderkerken, K., Multiple myeloma tumor progression in the 5T2MM murine model is a multistage and dynamic process of differentiation, proliferation, invasion, and apoptosis. Blood, 101(8): p. 3136-41, 2003
6. Bataille, R., Barlogie, B., Lu, Z. Y., Rossi, J. F., Lavabre-Bertrand, T., Beck, T., Wijdenes, J., Brochier, J., and Klein, B., Biologic effects of anti-interleukin-6 murine monoclonal antibody in advanced multiple myeloma. Blood, 86(2): p. 685-91, 1995
7. Bataille, R., Boccadoro, M., Klein, B., Durie, B., and Pileri, A., C-reactive protein and beta-2 microglobulin produce a simple and powerful myeloma staging system. Blood, 80(3): p. 733-7, 1992
8. Bataille, R., Durie, B. G., Sany, J., and Salmon, S. E., Myeloma bone marrow acid phosphatase staining. A correlative study of 38 patients. Blood, 55: p. 802-5, 1980
9. Bence Jones, H. I., On a new substance occuring in the urine of a patient with mollities ossium. Philos Trans R Soc (London), 138: p. 55-62, 1848
10. Berenson, J. R., Etiology of multiple myeloma: what's new. Semin Oncol, 26(5 Suppl 13): p. 2-9, 1999
11. Bernfield, M., Gotte, M., Park, P. W., Reizes, O., Fitzgerald, M. L., Lincecum, J., and Zako, M., Functions of cell surface heparan sulfate proteoglycans. Annu Rev Biochem, 68: p. 729-77, 1999
12. Betzl, G., Seller, M., Eichner, C., Kalbe, A., Geyer, M., Kleider, W., and van Miltenburg, R., Reproducibility of PCR on the LightCycler System. Biochemica, (1): p. 22-6, 2000
13. Birch, D. E., Simplified hot start PCR. Nature, 381(6581): p. 445-6., 1996
14. Black, K., Garrett, I. R., and Mundy, G. R., Chinese hamster ovarian cells transfected with the murine interleukin-6 gene cause hypercalcemia as well as cachexia, leukocytosis and thrombocytosis in tumor-bearing nude mice. Endocrinology, 128(5): p. 2657-9, 1991

61
15. Borset, M., Hjertner, O., Yaccoby, S., Epstein, J., and Sanderson, R. D., Syndecan-1 is targeted to the uropods of polarized myeloma cells where it promotes adhesion and sequesters heparin-binding proteins. Blood, 96(7): p. 2528-36, 2000
16. Brandstrom, H., Bjorkman, T., and Ljunggren, O., Regulation of osteoprotegerin secretion from primary cultures of human bone marrow stromal cells. Biochem Biophys Res Commun, 280(3): p. 831-5, 2001
17. Brandstrom, H., Jonsson, K. B., Ohlsson, C., Vidal, O., Ljunghall, S., and Ljunggren, O., Regulation of osteoprotegerin mRNA levels by prostaglandin E2 in human bone marrow stroma cells. Biochem Biophys Res Commun, 247(2): p. 338-41, 1998
18. Brandstrom, H., Jonsson, K. B., Vidal, O., Ljunghall, S., Ohlsson, C., and Ljunggren, O., Tumor necrosis factor-alpha and -beta upregulate the levels of osteoprotegerin mRNA in human osteosarcoma MG-63 cells. Biochem Biophys Res Commun, 248(3): p. 454-7, 1998
19. Broek, I. V., Asosingh, K., Vanderkerken, K., Straetmans, N., Van Camp, B., and Van Riet, I., Chemokine receptor CCR2 is expressed by human multiple myeloma cells and mediates migration to bone marrow stromal cell-produced monocyte chemotactic proteins MCP-1, -2 and -3. Br J Cancer, 88(6): p. 855-62, 2003
20. Bucay, N., Sarosi, I., Dunstan, C. R., Morony, S., Tarpley, J., Capparelli, C., Scully, S., Tan, H. L., Xu, W., Lacey, D. L., et al., osteoprotegerin-deficient mice develop early onset osteoporosis and arterial calcification. Genes Dev, 12(9): p. 1260-8, 1998
21. Bulfone-Paus, S., Ungureanu, D., Pohl, T., Lindner, G., Paus, R., Ruckert, R., Krause, H., and Kunzendorf, U., Interleukin-15 protects from lethal apoptosis in vivo. Nat Med, 3(10): p. 1124-8, 1997
22. Burton, J. D., Bamford, R. N., Peters, C., Grant, A. J., Kurys, G., Goldman, C. K., Brennan, J., Roessler, E., and Waldmann, T. A., A lymphokine, provisionally designated interleukin T and produced by a human adult T-cell leukemia line, stimulates T-cell proliferation and the induction of lymphokine-activated killer cells. Proc Natl Acad Sci U S A, 91(11): p. 4935-9, 1994
23. Callander, N. S. and Roodman, G. D., Myeloma bone disease. Semin Hematol, 38(3): p. 276-85, 2001
24. Carson, W. E., Fehniger, T. A., Haldar, S., Eckhert, K., Lindemann, M. J., Lai, C. F., Croce, C. M., Baumann, H., and Caligiuri, M. A., A potential role for interleukin-15 in the regulation of human natural killer cell survival. J Clin Invest, 99(5): p. 937-43, 1997
25. Carson, W. E., Ross, M. E., Baiocchi, R. A., Marien, M. J., Boiani, N., Grabstein, K., and Caligiuri, M. A., Endogenous production of interleukin 15 by activated human monocytes is critical for optimal production of interferon-gamma by natural killer cells in vitro. J Clin Invest, 96(6): p. 2578-82, 1995
26. Catlett-Falcone, R., Landowski, T. H., Oshiro, M. M., Turkson, J., Levitzki, A., Savino, R., Ciliberto, G., Moscinski, L., Fernandez-Luna, J. L., Nunez, G., et al., Constitutive activation of Stat3 signaling confers resistance to apoptosis in human U266 myeloma cells. Immunity, 10(1): p. 105-15, 1999
27. Chauhan, D., Kharbanda, S., Ogata, A., Urashima, M., Teoh, G., Robertson, M., Kufe, D. W., and Anderson, K. C., Interleukin-6 inhibits Fas-induced apoptosis and

62
stress-activated protein kinase activation in multiple myeloma cells. Blood, 89(1): p. 227-34, 1997
28. Chesi, M., Bergsagel, P. L., Brents, L. A., Smith, C. M., Gerhard, D. S., and Kuehl, W. M., Dysregulation of cyclin D1 by translocation into an IgH gamma switch region in two multiple myeloma cell lines. Blood, 88(2): p. 674-81, 1996
29. Chesi, M., Nardini, E., Brents, L. A., Schrock, E., Ried, T., Kuehl, W. M., and Bergsagel, P. L., Frequent translocation t(4;14)(p16.3;q32.3) in multiple myeloma is associated with increased expression and activating mutations of fibroblast growth factor receptor 3. Nat Genet, 16(3): p. 260-4, 1997
30. Chesi, M., Nardini, E., Lim, R. S., Smith, K. D., Kuehl, W. M., and Bergsagel, P. L., The t(4;14) translocation in myeloma dysregulates both FGFR3 and a novel gene, MMSET, resulting in IgH/MMSET hybrid transcripts. Blood, 92(9): p. 3025-34, 1998
31. Chou, Q., Russell, M., Birch, D. E., Raymond, J., and Bloch, W., Prevention of pre-PCR mis-priming and primer dimerization improves low- copy-number amplifications. Nucleic Acids Res, 20(7): p. 1717-23., 1992
32. Cornelius, P., Enerback, S., Bjursell, G., Olivecrona, T., and Pekala, P. H., Regulation of lipoprotein lipase mRNA content in 3T3-L1 cells by tumour necrosis factor. Biochem J, 249(3): p. 765-9, 1988
33. Croucher, P. I., Shipman, C. M., Lippitt, J., Perry, M., Asosingh, K., Hijzen, A., Brabbs, A. C., van Beek, E. J., Holen, I., Skerry, T. M., et al., Osteoprotegerin inhibits the development of osteolytic bone disease in multiple myeloma. Blood, 98(13): p. 3534-40, 2001
34. Dalrymple, J., On the microscopical character of mollities ossium. Dublin Quart J Med Sci, 2: p. 85-95, 1846
35. Day, R. M., Mitchell, T. J., Knight, S. C., and Forbes, A., Regulation of epithelial syndecan-1 expression by inflammatory cytokines. Cytokine, 21(5): p. 224-33, 2003
36. de la Mata, J., Uy, H. L., Guise, T. A., Story, B., Boyce, B. F., Mundy, G. R., and Roodman, G. D., Interleukin-6 enhances hypercalcemia and bone resorption mediated by parathyroid hormone-related protein in vivo. J Clin Invest, 95(6): p. 2846-52, 1995
37. Derksen, P. W., Keehnen, R. M., Evers, L. M., van Oers, M. H., Spaargaren, M., and Pals, S. T., Cell surface proteoglycan syndecan-1 mediates hepatocyte growth factor binding and promotes Met signaling in multiple myeloma. Blood, 99(4): p. 1405-10, 2002
38. Devlin, R. D., Reddy, S. V., Savino, R., Ciliberto, G., and Roodman, G. D., IL-6 mediates the effects of IL-1 or TNF, but not PTHrP or 1,25(OH)2D3, on osteoclast-like cell formation in normal human bone marrow cultures. J Bone Miner Res, 13(3): p. 393-9, 1998
39. Dhodapkar, M. V., Abe, E., Theus, A., Lacy, M., Langford, J. K., Barlogie, B., and Sanderson, R. D., Syndecan-1 is a multifunctional regulator of myeloma pathobiology: control of tumor cell survival, growth, and bone cell differentiation. Blood, 91(8): p. 2679-88, 1998

63
40. Doherty, T. M., Seder, R. A., and Sher, A., Induction and regulation of IL-15 expression in murine macrophages. J Immunol, 156(2): p. 735-41, 1996
41. Durie, B. G. and Salmon, S. E., A clinical staging system for multiple myeloma. Cancer, 36: p. 842-54, 1975
42. Fehniger, T. A. and Caligiuri, M. A., Interleukin 15: biology and relevance to human disease. Blood, 97(1): p. 14-32, 2001
43. Fukada, T., Hibi, M., Yamanaka, Y., Takahashi-Tezuka, M., Fujitani, Y., Yamaguchi, T., Nakajima, K., and Hirano, T., Two signals are necessary for cell proliferation induced by a cytokine receptor gp130: involvement of STAT3 in anti-apoptosis. Immunity, 5(5): p. 449-60, 1996
44. Gattei, V., Godeas, C., Degan, M., Rossi, F. M., Aldinucci, D., and Pinto, A., Characterization of anti-CD138 monoclonal antibodies as tools for investigating the molecular polymorphism of syndecan-1 in human lymphoma cells. Br J Haematol, 104(1): p. 152-62, 1999
45. Giuliani, N., Bataille, R., Mancini, C., Lazzaretti, M., and Barille, S., Myeloma cells induce imbalance in the osteoprotegerin/osteoprotegerin ligand system in the human bone marrow environment. Blood, 98(13): p. 3527-33, 2001
46. Giuliani, N., Colla, S., Rizzoli, V., Barille-Nion, S., and Bataille, R., Do Human Myeloma Cells Directly Produce the Receptor Activator of Nuclear Factor kappaB Ligand (RANKL) or Induce RANKL in the Bone Marrow Microenvironment? Cancer Res, 64(2): p. 772-3, 2004
47. Giuliani, N., Colla, S., Sala, R., Moroni, M., Lazzaretti, M., La Monica, S., Bonomini, S., Hojden, M., Sammarelli, G., Barille, S., et al., Human myeloma cells stimulate the receptor activator of nuclear factor-kappa B ligand (RANKL) in T lymphocytes: a potential role in multiple myeloma bone disease. Blood, 100(13): p. 4615-21, 2002
48. Gooding, R. P., Bybee, A., Cooke, F., Little, A., Marsh, S. G., Coelho, E., Gupta, D., Samson, D., and Apperley, J. F., Phenotypic and molecular analysis of six human cell lines derived from patients with plasma cell dyscrasia. Br J Haematol, 106(3): p. 669-81, 1999
49. Goossens, T., Klein, U., and Kuppers, R., Frequent occurrence of deletions and duplications during somatic hypermutation: implications for oncogene translocations and heavy chain disease. Proc Natl Acad Sci U S A, 95(5): p. 2463-8, 1998
50. Gori, F., Hofbauer, L. C., Dunstan, C. R., Spelsberg, T. C., Khosla, S., and Riggs, B. L., The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology, 141(12): p. 4768-76, 2000
51. Grabstein, K. H., Eisenman, J., Shanebeck, K., Rauch, C., Srinivasan, S., Fung, V., Beers, C., Richardson, J., Schoenborn, M. A., Ahdieh, M., et al., Cloning of a T cell growth factor that interacts with the beta chain of the interleukin-2 receptor. Science, 264(5161): p. 965-8, 1994
52. Greipp, P. R., Katzmann, J. A., O´Fallon, W. M., and Kyle, R. A., Value of B2-microglobulin level and plasma cell labeling indices as prognostic factors in patients with newly diagnosed myeloma. Blood, 72: p. 219-23, 1988

64
53. Hallek, M., Bergsagel, P. L., and Anderson, K. C., Multiple myeloma: increasing evidence for a multistep transformation process. Blood, 91(1): p. 3-21, 1998
54. Hardin, J., MacLeod, S., Grigorieva, I., Chang, R., Barlogie, B., Xiao, H., and Epstein, J., Interleukin-6 prevents dexamethasone-induced myeloma cell death. Blood, 84(9): p. 3063-70, 1994
55. Heider, U., Langelotz, C., Jakob, C., Zavrski, I., Fleissner, C., Eucker, J., Possinger, K., Hofbauer, L. C., and Sezer, O., Expression of receptor activator of nuclear factor kappaB ligand on bone marrow plasma cells correlates with osteolytic bone disease in patients with multiple myeloma. Clin Cancer Res, 9(4): p. 1436-40, 2003
56. Herold, G., Innere Medizin - Eine Vorlesungsorientierte Darstellung. Ausgabe 2000. Gerd Herold, Köln, 2000
57. Heyburn, P. J., Child, J. A., and Peacock, M., Relative importance of renal failure and increased bone resorption in the hypercalcaemia of myelomatosis. J Clin Pathol, 34: p. 54-7, 1981
58. Hirano, T., Yasukawa, K., Harada, H., Taga, T., Watanabe, Y., Matsuda, T., Kashiwamura, S., Nakajima, K., Koyama, K., Iwamatsu, A., et al., Complementary DNA for a novel human interleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature, 324(6092): p. 73-6, 1986
59. Hofbauer, L. C., Dunstan, C. R., Spelsberg, T. C., Riggs, B. L., and Khosla, S., Osteoprotegerin production by human osteoblast lineage cells is stimulated by vitamin D, bone morphogenetic protein-2, and cytokines. Biochem Biophys Res Commun, 250(3): p. 776-81, 1998
60. Hofbauer, L. C., Gori, F., Riggs, B. L., Lacey, D. L., Dunstan, C. R., Spelsberg, T. C., and Khosla, S., Stimulation of osteoprotegerin ligand and inhibition of osteoprotegerin production by glucocorticoids in human osteoblastic lineage cells: potential paracrine mechanisms of glucocorticoid-induced osteoporosis. Endocrinology, 140(10): p. 4382-9, 1999
61. Hofbauer, L. C., Khosla, S., Dunstan, C. R., Lacey, D. L., Boyle, W. J., and Riggs, B. L., The roles of osteoprotegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J Bone Miner Res, 15(1): p. 2-12, 2000
62. Hofbauer, L. C., Khosla, S., Dunstan, C. R., Lacey, D. L., Spelsberg, T. C., and Riggs, B. L., Estrogen stimulates gene expression and protein production of osteoprotegerin in human osteoblastic cells. Endocrinology, 140(9): p. 4367-70, 1999
63. Holen, I., Drury, N. L., Hargreaves, P. G., and Croucher, P. I., Evidence of a role for a non-matrix-type metalloproteinase activity in the shedding of syndecan-1 from human myeloma cells. Br J Haematol, 114(2): p. 414-21, 2001
64. Honore, P., Luger, N. M., Sabino, M. A., Schwei, M. J., Rogers, S. D., Mach, D. B., O'Keefe P, F., Ramnaraine, M. L., Clohisy, D. R., and Mantyh, P. W., Osteoprotegerin blocks bone cancer-induced skeletal destruction, skeletal pain and pain-related neurochemical reorganization of the spinal cord. Nat Med, 6(5): p. 521-8, 2000
65. Hsu, H., Lacey, D. L., Dunstan, C. R., Solovyev, I., Colombero, A., Timms, E., Tan, H. L., Elliott, G., Kelley, M. J., Sarosi, I., et al., Tumor necrosis factor receptor

65
family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc Natl Acad Sci U S A, 96(7): p. 3540-5, 1999
66. Ihle, J. N., Witthuhn, B. A., Quelle, F. W., Yamamoto, K., and Silvennoinen, O., Signaling through the hematopoietic cytokine receptors. Annu Rev Immunol, 13: p. 369-98, 1995
67. Kahler, O., Zur Symptomatologie des multiplen Myeloms. Beobachtung von Albuminosurie. Prag med Wschr, 14: p. 45-9, 1889
68. Kawano, M., Hirano, T., Matsuda, T., Taga, T., Horii, Y., Iwato, K., Asaoku, H., Tang, B., Tanabe, O., Tanaka, H., et al., Autocrine generation and requirement of BSF-2/IL-6 for human multiple myelomas. Nature, 332(6159): p. 83-5, 1988
69. Kellogg, D. E., Rybalkin, I., Chen, S., Mukhamedova, N., Vlasik, T., Siebert, P. D., and Chenchik, A., TaqStart Antibody: "hot start" PCR facilitated by a neutralizing monoclonal antibody directed against Taq DNA polymerase. Biotechniques, 16(6): p. 1134-7., 1994
70. Khosla, S., Minireview: the OPG/RANKL/RANK system. Endocrinology, 142(12): p. 5050-5, 2001
71. Kishimoto, T., The biology of interleukin-6. Blood, 74(1): p. 1-10, 1989
72. Klein, B., Zhang, X. G., Jourdan, M., Content, J., Houssiau, F., Aarden, L., Piechaczyk, M., and Bataille, R., Paracrine rather than autocrine regulation of myeloma-cell growth and differentiation by interleukin-6. Blood, 73(2): p. 517-26, 1989
73. Kong, Y. Y., Yoshida, H., Sarosi, I., Tan, H. L., Timms, E., Capparelli, C., Morony, S., Oliveira-dos-Santos, A. J., Van, G., Itie, A., et al., OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature, 397(6717): p. 315-23, 1999
74. Kovalszky, H., Gallai, M., Armbrust, T., and Ramadori, G., Syndecan-1 gene expression in isolated rat liver cells (hepatocytes, Kupffer cells, endothelial and Ito cells). Biochem Biophys Res Commun, 204(2): p. 944-9, 1994
75. Krause, H., Jandrig, B., Wernicke, C., Bulfone-Paus, S., Pohl, T., and Diamantstein, T., Genomic structure and chromosomal localization of the human interleukin 15 gene (IL-15). Cytokine, 8(9): p. 667-74, 1996
76. Kuipers, J., Vaandrager, J. W., Weghuis, D. O., Pearson, P. L., Scheres, J., Lokhorst, H. M., Clevers, H., and Bast, B. J., Fluorescence in situ hybridization analysis shows the frequent occurrence of 14q32.3 rearrangements with involvement of immunoglobulin switch regions in myeloma cell lines. Cancer Genet Cytogenet, 109(2): p. 99-107, 1999
77. Kurihara, N., Bertolini, D., Suda, T., Akiyama, Y., and Roodman, G. D., IL-6 stimulates osteoclast-like multinucleated cell formation in long term human marrow cultures by inducing IL-1 release. J Immunol, 144(11): p. 4226-30, 1990
78. Kyle, R. A., Multiple Myeloma. Review of 869 cases. Mayo Clin Proc, 50: p. 29-40, 1975

66
79. Kyrtsonis, M. C., Dedoussis, G., Zervas, C., Perifanis, V., Baxevanis, C., Stamatelou, M., and Maniatis, A., Soluble interleukin-6 receptor (sIL-6R), a new prognostic factor in multiple myeloma. Br J Haematol, 93(2): p. 398-400, 1996
80. Lacey, D. L., Timms, E., Tan, H. L., Kelley, M. J., Dunstan, C. R., Burgess, T., Elliott, R., Colombero, A., Elliott, G., Scully, S., et al., Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell, 93(2): p. 165-76, 1998
81. Li, J., Sarosi, I., Yan, X. Q., Morony, S., Capparelli, C., Tan, H. L., McCabe, S., Elliott, R., Scully, S., Van, G., et al., RANK is the intrinsic hematopoietic cell surface receptor that controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc Natl Acad Sci U S A, 97(4): p. 1566-71, 2000
82. Lichtenstein, A., Tu, Y., Fady, C., Vescio, R., and Berenson, J., Interleukin-6 inhibits apoptosis of malignant plasma cells. Cell Immunol, 162(2): p. 248-55, 1995
83. Lin, J. X., Migone, T. S., Tsang, M., Friedmann, M., Weatherbee, J. A., Zhou, L., Yamauchi, A., Bloom, E. T., Mietz, J., John, S., et al., The role of shared receptor motifs and common Stat proteins in the generation of cytokine pleiotropy and redundancy by IL-2, IL-4, IL-7, IL-13, and IL-15. Immunity, 2(4): p. 331-9, 1995
84. Lindahl, U., Kusche-Gullberg, M., and Kjellen, L., Regulated diversity of heparan sulfate. J Biol Chem, 273(39): p. 24979-82, 1998
85. Lipton, A., Ali, S. M., Leitzel, K., Chinchilli, V., Witters, L., Engle, L., Holloway, D., Bekker, P., and Dunstan, C. R., Serum osteoprotegerin levels in healthy controls and cancer patients. Clin Cancer Res, 8(7): p. 2306-10, 2002
86. Löffler, G. and Petrides, P. E., Biochemie und Pathobiochemie. 5th ed. Springer-Verlag, Berlin, Heidelberg, New York, Barcelona, Hongkong, London, Mailand, Paris, Santa Clara, Singapur, Tokio, 1997
87. Ludwig, H., Multiples Myelom. 1st ed. Springer-Verlag, Berlin, Heidelberg, New York, 1982
88. Ludwig, H., Die Klinik des multiplen Myeloms. Onkologie, 9(4): p. 202-8, 1986
89. Ludwig, H., Nachbaur, D. M., Fritz, E., Krainer, M., and Huber, H., Interleukin-6 is a prognostic factor in multiple myeloma. Blood, 77(12): p. 2794-5, 1991
90. MacIntyre, W., Case of mollities and fragilitas ossium, accompanied with urine strongly charged with animal matter. Med Chir Trans (London), 33: p. 211-32, 1850
91. Makiishi-Shimobayashi, C., Tsujimura, T., Iwasaki, T., Yamada, N., Sugihara, A., Okamura, H., Hayashi, S., and Terada, N., Interleukin-18 up-regulates osteoprotegerin expression in stromal/osteoblastic cells. Biochem Biophys Res Commun, 281(2): p. 361-6, 2001
92. Marschalko, T. V., Über die sogenannten Plasmazellen: Ein Beitrag zur Kenntnis der Herkunft der entzündlichen Infiltrationszellen. Arch Dermatol Syphiol, 30: p. 1-52, 1895
93. Mittag, M. Y., Die historische Entwicklung der Erkennung, Erforschung und Behandlung des Plasmozytoms. 1st ed. Schattauer, Stuttgart, New York, 1992
94. Mittrucker, H. W., Matsuyama, T., Grossman, A., Kundig, T. M., Potter, J., Shahinian, A., Wakeham, A., Patterson, B., Ohashi, P. S., and Mak, T. W.,

67
Requirement for the transcription factor LSIRF/IRF4 for mature B and T lymphocyte function. Science, 275(5299): p. 540-3, 1997
95. Miyazaki, T., Kawahara, A., Fujii, H., Nakagawa, Y., Minami, Y., Liu, Z. J., Oishi, I., Silvennoinen, O., Witthuhn, B. A., Ihle, J. N., et al., Functional activation of Jak1 and Jak3 by selective association with IL-2 receptor subunits. Science, 266(5187): p. 1045-7, 1994
96. Morrison, T. B., Weis, J. J., and Wittwer, C. T., Quantification of low-copy transcripts by continuous SYBR Green I monitoring during amplification. Biotechniques, 24(6): p. 954-8, 960, 962., 1998
97. Mrozek, E., Anderson, P., and Caligiuri, M. A., Role of interleukin-15 in the development of human CD56+ natural killer cells from CD34+ hematopoietic progenitor cells. Blood, 87(7): p. 2632-40, 1996
98. Murakami, T., Yamamoto, M., Ono, K., Nishikawa, M., Nagata, N., Motoyoshi, K., and Akatsu, T., Transforming growth factor-beta1 increases mRNA levels of osteoclastogenesis inhibitory factor in osteoblastic/stromal cells and inhibits the survival of murine osteoclast-like cells. Biochem Biophys Res Commun, 252(3): p. 747-52, 1998
99. Nagai, M., Kyakumoto, S., and Sato, N., Cancer cells responsible for humoral hypercalcemia express mRNA encoding a secreted form of ODF/TRANCE that induces osteoclast formation. Biochem Biophys Res Commun, 269(2): p. 532-6, 2000
100. Nakashima, T., Kobayashi, Y., Yamasaki, S., Kawakami, A., Eguchi, K., Sasaki, H., and Sakai, H., Protein expression and functional difference of membrane-bound and soluble receptor activator of NF-kappaB ligand: modulation of the expression by osteotropic factors and cytokines. Biochem Biophys Res Commun, 275(3): p. 768-75, 2000
101. Nilsson, K., Established cell lines as tools in the study of human lymphoma and myeloma cell characteristics. Haematol Blood Transfus, 20: p. 253-64, 1977
102. Nilsson, K., Bennich, H., Johansson, S. G., and Ponten, J., Established immunoglobulin producing myeloma (IgE) and lymphoblastoid (IgG) cell lines from an IgE myeloma patient. Clin Exp Immunol, 7(4): p. 477-89., 1970
103. Nordan, R. P. and Potter, M., A macrophage-derived factor required by plasmacytomas for survival and proliferation in vitro. Science, 233(4763): p. 566-9, 1986
104. Pearse, R. N., Sordillo, E. M., Yaccoby, S., Wong, B. R., Liau, D. F., Colman, N., Michaeli, J., Epstein, J., and Choi, Y., Multiple myeloma disrupts the TRANCE/ osteoprotegerin cytokine axis to trigger bone destruction and promote tumor progression. Proc Natl Acad Sci U S A, 98(20): p. 11581-6, 2001
105. Peest, D., Multiples Myelom - in Hämatologie/Internistische Onkologie, in Thiemes Innere Medizin - TIM, 1st ed., p. 925-31, Georg Thieme Verlag: Stuttgart, New York, 1999
106. Pellat-Deceunynk, C., Amiot, M., Bataille, R., Van Riet, I., Van Camp, B., Omede, P., and Boccadoro, M., Human myeloma cell lines as a tool for studying the biology of multiple myeloma: a reappraisal 18 years after. Blood, 86(10): p. 4001-2, 1995

68
107. Pettit, D. K., Bonnert, T. P., Eisenman, J., Srinivasan, S., Paxton, R., Beers, C., Lynch, D., Miller, B., Yost, J., Grabstein, K. H., et al., Structure-function studies of interleukin 15 using site-specific mutagenesis, polyethylene glycol conjugation, and homology modeling. J Biol Chem, 272(4): p. 2312-8, 1997
108. Quinn, J. M., Horwood, N. J., Elliott, J., Gillespie, M. T., and Martin, T. J., Fibroblastic stromal cells express receptor activator of NF-kappa B ligand and support osteoclast differentiation. J Bone Miner Res, 15(8): p. 1459-66, 2000
109. Rapraeger, A. C., Syndecan-regulated receptor signaling. J Cell Biol, 149(5): p. 995-8, 2000
110. Rasmussen, R., Morrison, T., Herrmann, M., and Wittwer, C., Quantitative PCR by continuous fluorescence monitoring of a double strand DNA specific binding dye. Biochemica, (2): p. 8-11, 1998
111. Ridley, R. C., Xiao, H., Hata, H., Woodliff, J., Epstein, J., and Sanderson, R. D., Expression of syndecan regulates human myeloma plasma cell adhesion to type I collagen. Blood, 81(3): p. 767-74, 1993
112. Riede, U.-N. and Schaefer, H.-E., Allgemeine und spezielle Pathologie. 4th ed. Georg Thieme Verlag, Stuttgart, New York, 1995
113. Roche, LightCycler Operator´s Manual. Version 3.5. Roche Molecular Biochemicals, Mannheim, 2000
114. Roodman, G. D., Biology of osteoclast activation in cancer. J Clin Oncol, 19(15): p. 3562-71, 2001
115. Roodman, G. D., Role of the bone marrow microenvironment in multiple myeloma. J Bone Miner Res, 17(11): p. 1921-5, 2002
116. Roux, S., Meignin, V., Quillard, J., Meduri, G., Guiochon-Mantel, A., Fermand, J. P., Milgrom, E., and Mariette, X., RANK (receptor activator of nuclear factor-kappaB) and RANKL expression in multiple myeloma. Br J Haematol, 117(1): p. 86-92, 2002
117. Rustizky, J. v., Multiples Myelom. Deutsche Zeitschr f Chir, 3: p. 162-72, 1873
118. Sanger, F. and Coulson, A. R., A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J Mol Biol, 94(3): p. 441-8., 1975
119. Sati, H. I., Greaves, M., Apperley, J. F., Russell, R. G., and Croucher, P. I., Expression of interleukin-1beta and tumour necrosis factor-alpha in plasma cells from patients with multiple myeloma. Br J Haematol, 104(2): p. 350-7, 1999
120. Schubert, G. E., Die Plasmocytomniere. I. Häufigkeit pathologisch-anatomischer Veränderungen. Klin Wschr, 52: p. 763-70, 1974
121. Sebestyen, A., Berczi, L., Mihalik, R., Paku, S., Matolcsy, A., and Kopper, L., Syndecan-1 (CD138) expression in human non-Hodgkin lymphomas. Br J Haematol, 104(2): p. 412-9, 1999
122. Sebestyen, A., Gallai, M., Knittel, T., Ambrust, T., Ramadori, G., and Kovalszky, I., Cytokine regulation of syndecan expression in cells of liver origin. Cytokine, 12(10): p. 1557-60, 2000

69
123. Seidel, C., Borset, M., Hjertner, O., Cao, D., Abildgaard, N., Hjorth-Hansen, H., Sanderson, R. D., Waage, A., and Sundan, A., High levels of soluble syndecan-1 in myeloma-derived bone marrow: modulation of hepatocyte growth factor activity. Blood, 96(9): p. 3139-46, 2000
124. Seidel, C., Hjertner, O., Abildgaard, N., Heickendorff, L., Hjorth, M., Westin, J., Nielsen, J. L., Hjorth-Hansen, H., Waage, A., Sundan, A., et al., Serum osteoprotegerin levels are reduced in patients with multiple myeloma with lytic bone disease. Blood, 98(7): p. 2269-71, 2001
125. Selvey, S., Thompson, E. W., Matthaei, K., Lea, R. A., Irving, M. G., and Griffiths, L. R., Beta-actin--an unsuitable internal control for RT-PCR. Mol Cell Probes, 15(5): p. 307-11, 2001
126. Serazin-Leroy, V., Denis-Henriot, D., Morot, M., de Mazancourt, P., and Giudicelli, Y., Semi-quantitative RT-PCR for comparison of mRNAs in cells with different amounts of housekeeping gene transcripts. Mol Cell Probes, 12(5): p. 283-91, 1998
127. Sezer, O., Heider, U., Jakob, C., Eucker, J., and Possinger, K., Human bone marrow myeloma cells express RANKL. J Clin Oncol, 20(1): p. 353-4, 2002
128. Sezer, O., Heider, U., Jakob, C., Zavrski, I., Eucker, J., Possinger, K., Sers, C., and Krenn, V., Immunocytochemistry reveals RANKL expression of myeloma cells. Blood, 99(12): p. 4646-7; author reply 4647, 2002
129. Sezer, O., Heider, U., Zavrski, I., Kuhne, C. A., and Hofbauer, L. C., RANK ligand and osteoprotegerin in myeloma bone disease. Blood, 101(6): p. 2094-8, 2003
130. Shipman, C. M. and Croucher, P. I., Osteoprotegerin is a soluble decoy receptor for tumor necrosis factor-related apoptosis-inducing ligand/Apo2 ligand and can function as a paracrine survival factor for human myeloma cells. Cancer Res, 63(5): p. 912-6, 2003
131. Simonet, W. S., Lacey, D. L., Dunstan, C. R., Kelley, M., Chang, M. S., Luthy, R., Nguyen, H. Q., Wooden, S., Bennett, L., Boone, T., et al., Osteoprotegerin: a novel secreted protein involved in the regulation of bone density. Cell, 89(2): p. 309-19, 1997
132. Sneed, T. B., Stanley, D. J., Young, L. A., and Sanderson, R. D., Interleukin-6 regulates expression of the syndecan-1 proteoglycan on B lymphoid cells. Cell Immunol, 153(2): p. 456-67, 1994
133. Sordillo, E. M. and Pearse, R. N., RANK-Fc: a therapeutic antagonist for RANK-L in myeloma. Cancer, 97(3 Suppl): p. 802-12, 2003
134. Spets, H., Jernberg-Wiklund, H., Sambade, C., Soderberg, O., and Nilsson, K., The effects on growth and survival of IL-6 can be dissociated in the U-266-1970/U-266-1984 and HL407E/HL407L human multiple myeloma cell lines. Br J Haematol, 98(1): p. 126-33, 1997
135. Sporeno, E., Savino, R., Ciapponi, L., Paonessa, G., Cabibbo, A., Lahm, A., Pulkki, K., Sun, R. X., Toniatti, C., Klein, B., et al., Human interleukin-6 receptor super-antagonists with high potency and wide spectrum on multiple myeloma cells. Blood, 87(11): p. 4510-9, 1996

70
136. Standal, T., Seidel, C., Hjertner, O., Plesner, T., Sanderson, R. D., Waage, A., Borset, M., and Sundan, A., Osteoprotegerin is bound, internalized, and degraded by multiple myeloma cells. Blood, 100(8): p. 3002-7, 2002
137. Steinbock, T., Paleopathological diagnosis and interpretation. Verlag Thomas, Illinois, 1976
138. Takahashi, N., Udagawa, N., and Suda, T., A new member of tumor necrosis factor ligand family, ODF/OPGL/TRANCE/RANKL, regulates osteoclast differentiation and function. Biochem Biophys Res Commun, 256(3): p. 449-55, 1999
139. Taniguchi, T., Miyazaki, T., Minami, Y., Kawahara, A., Fujii, H., Nakagawa, Y., Hatakeyama, M., and Liu, Z. J., IL-2 signaling involves recruitment and activation of multiple protein tyrosine kinases by the IL-2 receptor. Ann N Y Acad Sci, 766: p. 235-44, 1995
140. Terpos, E., Szydlo, R., Apperley, J. F., Hatjiharissi, E., Politou, M., Meletis, J., Viniou, N., Yataganas, X., Goldman, J. M., and Rahemtulla, A., Soluble receptor activator of nuclear factor kappaB ligand-osteoprotegerin ratio predicts survival in multiple myeloma: proposal for a novel prognostic index. Blood, 102(3): p. 1064-9, 2003
141. Thomas, X., Anglaret, B., Magaud, J. P., Epstein, J., and Archimbaud, E., Interdependence between cytokines and cell adhesion molecules to induce interleukin-6 production by stromal cells in myeloma. Leuk Lymphoma, 32(1-2): p. 107-19, 1998
142. Tinhofer, I., Marschitz, I., Henn, T., Egle, A., and Greil, R., Expression of functional interleukin-15 receptor and autocrine production of interleukin-15 as mechanisms of tumor propagation in multiple myeloma. Blood, 95(2): p. 610-8, 2000
143. Tomoyasu, A., Goto, M., Fujise, N., Mochizuki, S., Yasuda, H., Morinaga, T., Tsuda, E., and Higashio, K., Characterization of monomeric and homodimeric forms of osteoclastogenesis inhibitory factor. Biochem Biophys Res Commun, 245(2): p. 382-7, 1998
144. Tricot, G., New insights into role of microenvironment in multiple myeloma. Lancet, 355(9200): p. 248-50, 2000
145. Tsuda, E., Goto, M., Mochizuki, S., Yano, K., Kobayashi, F., Morinaga, T., and Higashio, K., Isolation of a novel cytokine from human fibroblasts that specifically inhibits osteoclastogenesis. Biochem Biophys Res Commun, 234(1): p. 137-42, 1997
146. Udagawa, N., Takahashi, N., Akatsu, T., Sasaki, T., Yamaguchi, A., Kodama, H., Martin, T. J., and Suda, T., The bone marrow-derived stromal cell lines MC3T3-G2/PA6 and ST2 support osteoclast-like cell differentiation in cocultures with mouse spleen cells. Endocrinology, 125(4): p. 1805-13, 1989
147. van Boxel, J. A. and Buell, D. N., IgD on cell membranes of human lymphoid cell lines with multiple immunoglobulin classes. Nature, 251(5474): p. 443-4., 1974
148. Van Damme, J., Opdenakker, G., Simpson, R. J., Rubira, M. R., Cayphas, S., Vink, A., Billiau, A., and Van Snick, J., Identification of the human 26-kD protein, interferon beta 2 (IFN-beta 2), as a B cell hybridoma/plasmacytoma growth factor induced by interleukin 1 and tumor necrosis factor. J Exp Med, 165(3): p. 914-9, 1987

71
149. Vande Broek, I., Vanderkerken, K., De Greef, C., Asosingh, K., Straetmans, N., Van Camp, B., and Van Riet, I., Laminin-1-induced migration of multiple myeloma cells involves the high-affinity 67 kD laminin receptor. Br J Cancer, 85(9): p. 1387-95, 2001
150. Vidal, N. O., Brandstrom, H., Jonsson, K. B., and Ohlsson, C., Osteoprotegerin mRNA is expressed in primary human osteoblast-like cells: down-regulation by glucocorticoids. J Endocrinol, 159(1): p. 191-5, 1998
151. Vidal, O. N., Sjogren, K., Eriksson, B. I., Ljunggren, O., and Ohlsson, C., Osteoprotegerin mRNA is increased by interleukin-1 alpha in the human osteosarcoma cell line MG-63 and in human osteoblast-like cells. Biochem Biophys Res Commun, 248(3): p. 696-700, 1998
152. Vihinen, T., Auvinen, P., Alanen-Kurki, L., and Jalkanen, M., Structural organization and genomic sequence of mouse syndecan-1 gene. J Biol Chem, 268(23): p. 17261-9, 1993
153. Villunger, A., Egle, A., Kos, M., Hittmair, A., Maly, K., and Greil, R., Constituents of autocrine IL-6 loops in myeloma cell lines and their targeting for suppression of neoplastic growth by antibody strategies. Int J Cancer, 65(4): p. 498-505, 1996
154. Willis, T. G. and Dyer, M. J. S., The role of immunoglobulin translocations in the pathogenesis of B-cell malignancies. Blood, 96(3): p. 808-22, 2000
155. Wittwer, C. T., Ririe, K. M., Andrew, R. V., David, D. A., Gundry, R. A., and Balis, U. J., The LightCycler: a microvolume multisample fluorimeter with rapid temperature control. Biotechniques, 22(1): p. 176-81., 1997
156. Wong, B. R., Rho, J., Arron, J., Robinson, E., Orlinick, J., Chao, M., Kalachikov, S., Cayani, E., Bartlett, F. S., 3rd, Frankel, W. N., et al., TRANCE is a novel ligand of the tumor necrosis factor receptor family that activates c-Jun N-terminal kinase in T cells. J Biol Chem, 272(40): p. 25190-4, 1997
157. Wright, J. H., A case of multiple Myeloma. John Hopkins Hosp Rep, 9: p. 359-66, 1900
158. Yaccoby, S., Pearse, R. N., Johnson, C. L., Barlogie, B., Choi, Y., and Epstein, J., Myeloma interacts with the bone marrow microenvironment to induce osteoclastogenesis and is dependent on osteoclast activity. Br J Haematol, 116(2): p. 278-90, 2002
159. Yamaguchi, K., Kinosaki, M., Goto, M., Kobayashi, F., Tsuda, E., Morinaga, T., and Higashio, K., Characterization of structural domains of human osteoclastogenesis inhibitory factor. J Biol Chem, 273(9): p. 5117-23, 1998
160. Yang, Y., Borset, M., Langford, J. K., and Sanderson, R. D., Heparan sulfate regulates targeting of syndecan-1 to a functional domain on the cell surface. J Biol Chem, 278(15): p. 12888-93, 2003
161. Yasuda, H., Shima, N., Nakagawa, N., Mochizuki, S. I., Yano, K., Fujise, N., Sato, Y., Goto, M., Yamaguchi, K., Kuriyama, M., et al., Identity of osteoclastogenesis inhibitory factor (OCIF) and osteoprotegerin (OPG): a mechanism by which OPG/OCIF inhibits osteoclastogenesis in vitro. Endocrinology, 139(3): p. 1329-37, 1998

72
162. Yasuda, H., Shima, N., Nakagawa, N., Yamaguchi, K., Kinosaki, M., Mochizuki, S., Tomoyasu, A., Yano, K., Goto, M., Murakami, A., et al., Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci U S A, 95(7): p. 3597-602, 1998
163. Zhan, F., Tian, E., Bumm, K., Smith, R., Barlogie, B., and Shaughnessy, J., Jr., Gene expression profiling of human plasma cell differentiation and classification of multiple myeloma based on similarities to distinct stages of late-stage B-cell development. Blood, 101(3): p. 1128-40, 2003

73
Lebenslauf Markus Bergmann, geboren am 21. April 1975 in Dorsten Schulbildung: Dr. Adenauer Grundschule Geseke 1981-1985 Gymnasium Antonianum Geseke 1985-1994, Abitur 1994 Zivildienst: Krankenpflege in der Abteilung für Innere Medizin, Kreiskrankenhaus Sögel 09/1994-12/1995 Praktikum: Eisengießerei Schonlau, Geseke 12/1995-04/1996 u.a. Instandhaltungsschlosserei, Elektrowerkstatt, Gießereibetrieb
Studium: Humanmedizin an der RWTH Aachen, Immatrikulation 10/1996 Examina: Ärztliche Vorprüfung 08/1998 Erster Abschnitt der Ärztlichen Prüfung 08/1999 USMLE Step 1 11/1999 Zweiter Abschnitt der Ärztlichen Prüfung 03/2002 Dritter Abschnitt der Ärztlichen Prüfung 05/2003
Praktische Tätigkeiten: Famulaturen: Chirurgie, Innere Medizin, Allgemeinmedizin, Pathologie, Psychiatrie,
Anästhesie Praktisches Jahr: 22.04.2002-23.03.2003 Medizinisches Zentrum Kreis Aachen und Royal Devon & Exeter Hospital, Exeter, GB
Wahlfach Neurologie Weiterbildung: seit 15.09.2003 zunächst als AIP, seit 01.10.2004 als Assistenzarzt
Abteilung für Chirurgie, Johanniter-Krankenhaus Bonn
Königswinter, den 14. Mai 2005





















![Ganzkörper- MRT beim multiplen Myelom: Vergleich ... · klinischen und pathomorphologischen Symptomen eines multiplen Myeloms zum ersten Mal Proteine im Urin [10]. Erst 117 Jahre](https://img.pdfslide.org/doc/110x75/605fae9a8404eb0e4d38fff5/ganzkrper-mrt-beim-multiplen-myelom-vergleich-klinischen-und-pathomorphologischen.jpg)