Embed Size (px)
Citation preview

Zeitschrift fiJr
Z Parasitenkd (1982)67:147-153 Parasitenkunde Parasitology Research
�9 Springer-Verlag 1982
Ultrastructure of Gamonts and Gametes and Fertilization of Sarcocystis sp. from the Roe Deer (Capreolus capreolus) in Dogs
R. Entzeroth Abteilung fiir Protozoologie, Zoologisches Institut, der Universit/it, Poppelsdorfer Schlo6, 5300 Bonn 1, Federal Republic of Germany
Abstract. G am on t s o f Sarcocystis sp. f rom the roe deer were examined in the intestine of dogs 10 h after inoculation. Early macrogamont s were limited by a three-membranous pellicle, and situated in a paras i tophorous vacuole. Female sexual stages during fertilization, the macrogametes , were limited by five membranes, and microgametes were observed in the paras i tophorous vacuole. The outer membranes of the microgamete and macrogamete fuse, and the nucleoplasm of the microgamete enters the cytoplasm of the macroga- mete. N o wall-forming bodies were observed in macrogamont s and macroga- metes.
Introduction
The biology of Sarcocystis species is characterized by an obl igatory two-host life cycle with multiplication (merogonies) occurr ing in the intermediate host (herbivore) and g a m o g o n y in the carnivorous final host (Rommel et al. 1972; Heydorn 1977; Mehlhorn and H eydo rn 1978). G a m o g o n y takes place within cells of the lamina propr ia of the intestine of the final host. Ultrastructural studies on g a m o g o n y stages have been done in tissue cultures (Vetterling 1973; Mehlhorn and H eydo rn 1979; Becket et al. 1979) and in the final host (Zaman and Colley 1975; Schfiffler 1979; Sheffield e ta l . 1980; Entzeroth 1980). In the present study early g a m o g o n y stages and fertilization of Sarcocystis sp. of the roe deer were studied in the intestine of dogs.
Materials and Methods
Muscle tissue of diaphragm and esophagus containing mature cysts of Sarcocystis sp. were taken from roe deer collected in the area of Bonn, Federal Republic of Germany, and repeatedly fed to an adult dog. Another coccidia-free puppy (6 weeks old) was inoculated by stomach tube with a suspension of Sarcocystis cyst merozoites that had been isolated from 200 g diaphragm and esophagus muscle of roe deer by trypsin digestion according to the method of Erber (1977). The first dog, which was repeatedly inoculated 19 days (to collect sporocysts), 24 h, 53 h before, was killed 10 h after the last inoculation. The second dog was killed 10 h after a single inoculation
0044-3255/82/0067/0147/$01.40

I48 R. Entzeroth
with Sarcocystis merozoites. Segments of duodenum, jejunum, and ileum were fixed in 2.5% glutaral- dehyde in cacodylate buffer, dehydrated in ethanol, and embedded in Spurr low viscosity medium. Ultrathin sections were poststained with lead citrate and examined with a Zeiss EM 9 $2 electron microscope.
Results
The intestines of both dogs were heavily infected with Sarcocystis sp. In the first dog sporulated sporocysts from the first inoculation as well as developing gamonts from the last inoculation were present. After entering epithelial and submucosa cells of the small intestine, merozoites lose several organdies usually seen in motile stages of coccidia (rhoptries, micronemes, and sub-pellicular microtubules), and round up. At 10 h postinoculation (p.i.) merozoites trans- forming into gamonts were situated in a parasitophorous vacuole limited by a membrane of the host cell, which was reinforced by an osmiophilic layer up to 50 nm thick and formed pocket-like enlargements (Fig. 1). In some cases, the parasite with the membrane limiting the parasitophorous vacuole is displaced from its original position within the host cell and appears to be extracellular between the basal membrane of the epithelial layer and the lamina propria. Young gamonts at this time measured 5-7 gm in diameter. They were character- ized by a prominent nucleus with numerous chromatin plaques (Fig. 1, arrows), and well-developed endoplasmic reticulum and mitochondria. Remnants of mi- cronemes were present. During further development the macrogamonts increased in size (7-10 Ixm) and their cytoplasm and nucleus became paler (Fig. 2). Mito- chondria and lamellae of the endoplasmic reticulum, but no storage granules or wall-forming bodies, were present at this stage of development. In several cases and in both dogs, microgametes could be observed in the parasitophorous vacuole of macrogametes (Figs. 2 and 3). The microgametes measured 2-3 ~tm in length and 0.5-0.8 gm in width. They were bound by a unit membrane and contained a mitochondrion with tubular cristae and granular osmiophilic, chromatin-like material (Fig. 3, OZ). Typical flagella could not be found; how- ever, single microtubules, or groups of three and four, were present inside the microgamete and in the parasitophorous vacuole; those microtubules within the parasitophorous vacuole were surrounded by a membrane (Fig. 3). Union of the gametes occurred by a narrow channel through which there was cytoplas- mic continuity. At the periphery of the channel, at its narrowest point, the membranes of both gametes were fused, forming a continuous entity. At the point of fusion the gamete membranes formed an osmiophilic ring-like structure (OS), which appeared wedge-like in cross-section (Fig. 3). Through this ring the osmiophilic chromatin-like material (OZ), representing the microgamete nucleoplasm, entered the cytoplasm of the macrogamete.
Of interest is the membranous limitation of the macrogamete, which consists of five membranes (M1-M5). The outer membrane of the parasite (M1) is the plasmalemma bordering the gamont adjacent to the parasitophorous vacuole. Underneath are two membranes (M2 and M3), 10-30 nm apart, with granular material inbetween. Two additional membranes (M4+M5) underneath appear in the form of a membrane complex with interruptions. At this stage of develop- ment the nucleus of the macrogamete was situated at the opposite side of
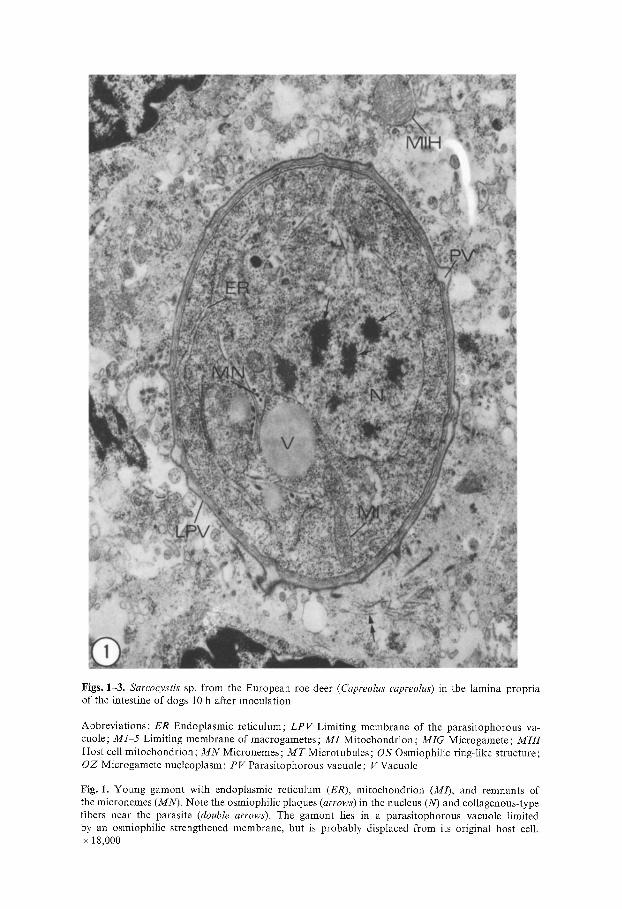
Figs. 1-3. Sarcocystis sp. from the European roe deer (Capreolus capreolus) in the lamina propria of the intestine of dogs 10 h after inoculation
Abbreviations: ER Endoplasmic reticulum; LPV Limiting membrane of the parasitophorous va- cuole; M1 5 Limiting membrane of macrogametes; MI Mitochondrion; MIG Microgamete ; MIH Host cell mitochondrion ; M N Micronemes; M T Microtubules; OS Osmiophilic ring-like structure; OZ Microgamete nucleoplasm; PV Parasitophorous vacuole; V Vacuole
Fig. 1. Young gamont with endoplasmic reticulum (ER), mitochondrion (M/), and remnants of the micronemes (MN). Note the osmiophilic plaques (arrows) in the nucleus (N) and coIlagenous-type fibers near the parasite (double arrows). The gamont lies in a parasitophorous vacuole limited by an osmiophilic strengthened membrane, but is probably displaced from its original host cell. x 18,000
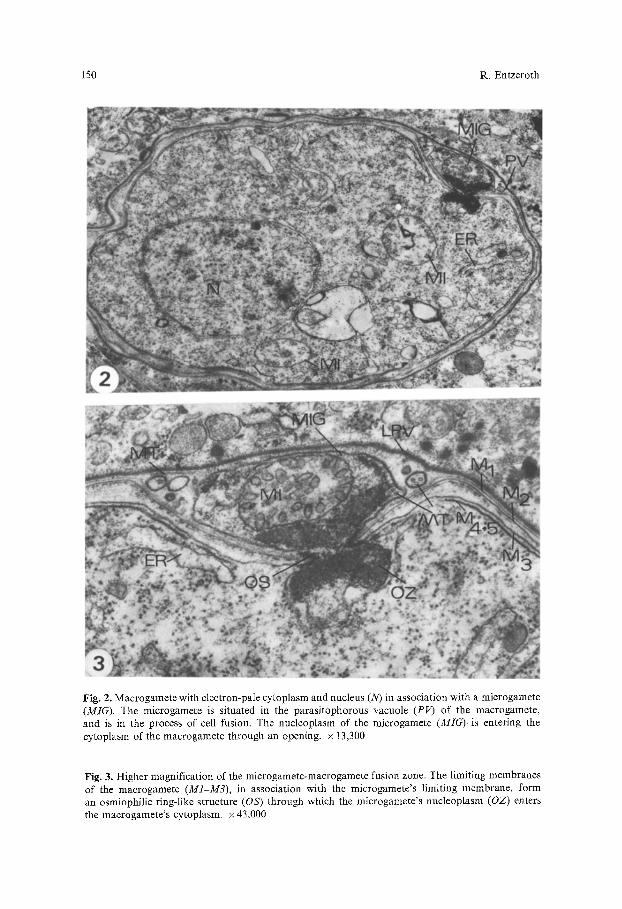
150 R. Entzeroth
Fig. 2. Macrogamete with electron-pale cytoplasm and nucleus (N) in association with a microgamete (MIG). The microgamete is situated in the parasitophorous vacuole (PV) of the macrogamete, and is in the process of cell fusion. The nucleoplasm of the microgamete (MIG)is entering the cytoplasm of the macrogamete through an opening, x 13,300
Fig. 3. Higher magnification of the microgamete-macrogamete fusion zone. The limiting membranes of the macrogamete (M1-M3), in association with the microgamete's limiting membrane, form an osmiophilic ring-like structure (OS) through which the microgamete's nucleoplasm (OZ) enters the macrogamete's cytoplasm, x 43,000

Ultrastructure of Gamonts and Fertilization of Sarcocystis sp. 151
the microgamete's entrance (Fig. 2). Thus, fusion of the microgamete nucleo- plasm with that of the macrogamete was not observed.
Discussion
Immediately after entering intestinal cells of dogs, cyst merozoites of Sarcocystis sp. from the roe deer develop into gamonts. Fertilization occurs between 10 and 12 h p.i. This rapid development is common in Sarcocystis species and has been observed in Sarcocystis of sheep in cats (Mehlhorn and Scholtyseck 1974), Sarcocystis of rats in the python (Zaman and Colley 1975), Sarcocystis of mice in cats (Ruiz and Frenkel 1976), and also in in vitro studies of Sarcocystis sp. (Vetterling et al. 1973; Mehlhorn and Heydorn 1978; Becker et al. 1979). In the present study the development of the gamonts occurred in a parasitophor- ous vacuole limited by a membrane thickened by osmiophilic material. The same observation has been made in gamogony studies of other Sarcocystis species (Zaman and Colley 1975; Mehlhorn and Heydorn 1978; Becker et al. 1979). Occasionally, gamonts were observed intercellularly surrounded by the limiting membrane of the parasitophorous vacuole, probably dislocated from the original host cell. Similar observations have been made in Sarcocystis bovi- canis (Sheffield and Fayer 1980).
Of interest is the fact that Sarcocystis macrogametes and oocysts are sur- rounded by a number of membranes. In Sarcocystis of grackles seven membranes have been reported (Vetterling et al. 1973), in S. suihominis (Mehlhorn and Heydorn 1978) and Sarcocystis sp. (Becker et al. 1979) six membranes and an additional outer layer were seen. In the present study, macrogametes prior to and during fertilization were surrounded by five membranes, thus showing a configuration described in Eimeria species (Scholtyseck and Mehlhorn 1971 ; Chobotar et al. 1980).
Also there have been different observations concerning wall-forming bodies in Sarcocystis gamonts. While no wall-forming bodies have been seen in the present study, in S. suihominis (Mehlhorn and Heydorn 1978) both types of wall-forming bodies were reported to be present. In Sarcocystis of grackles (Vetterling et al. 1973) granules resembling WF 1 were seen, whereas in S. bovicanis (Sheffield and Fayer 1978) no wall-forming bodies were found. Further studies are necessary to clarify whether there are actually wall-forming bodies involved in the formation of the relatively thin oocyst wall in Sarcocystis species.
Microgametes but no microgamonts were found in the present study. In light microscopic studies of S. fusiformis (Fayer 1972) and of S. hemionilatrantis (Speer et al. 1980) no microgamonts were seen either. From tissue cultures of Sarcocystis sp. of grackles (Fayer 1972), it is known that microgamonts accounted for only 18% of all gamonts formed. In S. suihominis (Mehlhorn and Heydorn 1979) microgamonts were relatively small (10 gin) and produced 20 30 microgametes.
In the present study microgametes that had already entered the parasitophor- ous vacuole or were in the act of fertilization lacked typical flagella. The same observation has been made in S. bovicanis microgametes (Sheffield and Fayer 1980) and S. suicanis (Schfiffler 1979).

152 R. Entzeroth
P r e c e d i n g fe r t i l i za t ion the p l a s m a l e m m a o f the m i c r o g a m e t e and m a c r o g a -
m e t e fuse a n d o n l y the n u c l e o p l a s m o f the m i c r o g a m e t e appea r s to en te r the
m a c r o g a m e t e c y t o p l a s m . These obse rva t ions , h o w e v e r , d i f fe r f r o m those in Ei- meria species w h e r e the w h o l e m i c r o g a m e t e seems to be i n c o r p o r a t e d in to the m a c r o g a m e t e ( H a m m o n d and S c h o l t y s e c k 1970; M a d d e n and Ve t t e r l i ng 1977).
F u s i o n o f the m i c r o - and m a c r o g a m e t e p l a s m a l e m m a is a p p a r e n t l y c o m m o n
a m o n g the A p i c o m p l e x a , as it has a lso been o b s e r v e d in three o t h e r genera ,
Parahaemoproteus velans (Desse r 1972), Haemoproteus columbae (Ga l lucc i 1974),
and Plasmodium yoelii nigeriensis (S inden et al. 1976).
Acknowledgements. The author wishes to thank Prof. Erich Scholtyseck for his guidance during this study and Prof. Bill Chobotar for reviewing the manuscript. This work was supported by a Feodor Lynen stipend of the Alexander von Humboldt Foundation.
References
Becker B, Mehlhorn H, Heydorn AO (1979) Light and electron microscopic studies on gamogony and sporogony of five Sarcocystis species in vivo and in tissue cultures. Zbl Bakt Hyg I Abt Orig A 244 : 394 404
Chobotar B, S~naud J, Ernst JV, Scholtyseck E (1980) Ultrastructure of macrogametogenesis and formation of the oocyst wall of Eimeria papillata in Mus musculus. Protistologica 16 : 115-124
Desser SS (1972) Gametocyte maturation, exflagellation and fertilization in Parahaemoproteus (= Haemoproteus velans (Coatney & Roudabush) (Haemosporidia : Haemoproteidae) : an ultrastruc- tural study. J Protozool 19:287296
Entzeroth R (1980) Licht- und elektronenmikroskopische Untersuchungen zum Lebenszyklus yon Sarcocystis sp. (Sporozoa, Coccidia) im Reh (Capreolus capreolus) und Hund (Canis familiaris). Dissertation Universitfit Bonn
Erber M (1977) M6glichkeiten des Nachweises und der Differenzierung von zwei Sarcocystis-Arten des Schweines. Beri Mfinch Tierfirztl Wochenschr 90:480482
Fayer R (1972) Gametogony of Sarcocystis sp. in ceil culture. Science 175:65-67 Gallucci BB (1974) Fine structure of Haemoproteus columbae Kruse during macrogametogenesis
and fertilization. J Protozool 21:254-263 Hammond D M, Scholtyseck E (1970) Observations concerning the process of fertilization in Eimeria
boris. Naturwissenschaften 8 : 399~400 Heydorn AO (1977) Beitrfige zum Lebenszyklus der Sarkosporidien. IX. Entwicklungszyklus von
Sarcocystis suihominis n. spec. Berl M/inch Tier/irztl Wochenscbr 90:218224 Madden PA, Vetterling JM (1977) Scanning electron microscopy of Eimeria tenella microgametogen-
esis and fertilization. J Parasitol 63:607-610 Mehlhorn H, Heydorn AO (1978) The sarcosporidia (Protozoa, Sporozoa): life cycle and fine
structure. Adv Parasitol :43-93 Mehlhorn H, Heydorn AO (1979) Electron microscopical study on gamogony of Sarcocystis suiho-
minis in human tissue cultures. Z Parasitenkd 58:97-113 Mehlhorn H, Scholtyseck E (1974) Light and electron microscope studies on stages of Sarcocystis
tenella in the intestine of cats. Z Parasitenkd 43:251-270 Rommel M, Heydorn AO, Gruber F (1972) Beitrfige zum Lebenszyklus der Sarkosporidien. I.
Die Sporozyste von S. tenella in den F/izes der Katze. Berl M/inch Tier/irztl Wochenschr 85:101 I05
Ruiz A, Frenkel JK (1976) Recognition of cyclic transmission of Sarcocystis muris by cats. J Infect Dis 133:409 418
Sch/iffler M (1979) Licht- und elektronenmikroskopische Untersuchungen zur Gamogonie und Sporogonie yon Sarcocystis suicanis im D/inndarm experimentell infizierter Hunde. Dissertation Universitfit M/inchen

Ultrastructure of Gamonts and Fertilization of Sarcocystis sp. 153
Scholtyseck E, Mehlhorn H, Hammond DM (1971) Fine structure of macrogametes and oocysts of coccidia and related organisms. Z Parasitenkd 37:143
Sheffield HG, Fayer R (i978) Electron microscopy of in situ development of Sarcocystis cruzi oocysts. ICOPA IV, Warsaw
Sheffield HG, Fayer R (t980) Fertilization in the coccidia: fusion of Sarcocystis bovicani~' gametes. Proc Helminthol Soc Wash 47:118 121
Sinden RE, Canning EU, Spain B (1976) Gametogenesis and fertilization in Plasmodiurn yoelii nigeriensis." a transmission electron microscope study. Proc R Soc Lond [Biol] 193:55 76
Speer CA, Pond DB, Ernst JV (1980) Development of Sarcocystis hemionilatrantis Hudkins and Kistner, 1977 in the small intestine of coyotes. Proc Helminthol Soc Wash 47:i06-113
Vetterling JM, Pacheco ND, Fayer R (1973) Fine structure of gametogony and oocyst formation in Sarcocystis sp. in cell culture. J Protozool 20:28-48
Zaman V, Colley FC (1975) Light and electron microscopic observations of the life cycle of Sarcocys- tis orientalis sp. n. in the rat (Rattus norvegicus) and the Malaysian reticulated python (Python reticulatus). Z Parasitenkd 47 : 169-185
Received October i5, 198I





























