Embed Size (px)
Citation preview

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 1
Bewegungslehre und Methodik dargestellt am Beispiel
des Gerätturnens
Prof. Dr. Klaus Wiemann © 2013
5 Sensomotorische Grundlagen..............................................................................3
5.1. Grundzüge der Struktur des Nervensystems und seiner motorischen Funktionen......3 5.2 Motorische Aufgaben des Rückenmarkes: Dehnungs- und Spannungsreflexe..........8
5.2.1 Die Dehnungsrezeptoren des Muskels............................................................8 5.2.2 Der Dehnungsreflex und die Längenregelung des Muskels..........................10 5.2.3 Reflex der antagonistischen Hemmung.......................................................14 5.2.4 Spannungsreflex und Spannungsregelung des Muskels.............................16 5.2.5 Beuge- und Streckreflexe............................................................................17

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 2
5.3 Stützmotorik..............................................................................................................21
5.3.1 Regelung des aufrechten Standes..............................................................24 5.3.2 Orientierung im Raum.................................................................................31 5.3.3 Orientierungs- und Tonusreflexe beim Menschen.......................................33
5.4 Zielmotorik ................................................................................................................43 5.4.1 Motivation und Handlungsbereitschaft.......................................................48 5.4.2 Strategische Planung ................................................................................49 5.4.3 Bildung des Bewegungsentwurfes.............................................................51 5.4.4 Bewegungsprogrammierung......................................................................63 5.4.5 Start und Realisation der Bewegung..........................................................65 5.4.6 Bewegungsrückmeldung und Bewegungskorrektur...................................69
5.5. Biologische Grundlagen des motorischen Lernens.................................................81
5.5.1 Neuronale Ketten, Kreise und Netze..........................................................81 5.5.2 Konditionierung als Basis motorischer Lernvorgänge.............................. 85

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 3
5 Sensomotorische Grundlagen
Wie in kaum einer anderen Sportart stellt Gerätturnen mit seinen vielgestaltigen Änderungen der Körperpositionen und der Drehbewegungen des ganzen Körpers um alle nur denkbaren Körperachsen spezifische Anforderungen an die sensomotorische Leistungsfähigkeit. Dies betrifft insbesondere denjenigen Bereich der Motorik, der die Regelung der Körperposition sowie die Sicherung der Orientierung im Raum bei Drehbeschleunigungen zur Aufgabe hat. Daraus erwachsen entsprechende Anforderungen an methodische Maßnahmen und Trainingsprinzipien.
Sensomotorische Leistungen sind an die Funktion des zentralen Nervensystems (ZNS) gebunden, so dass es geboten erscheint, die wichtigsten Grundlagen in der Funktion des Nervensystems hier aufzuzeigen. Dabei wird – mit Ausnahme einiger grundlegender Funktionen – das Hauptaugenmerk auf diejenigen Aspekte beschränkt bleiben, die für das Gerätturnen, die Realisation turnerischer Elemente sowie für entsprechende methodische Maßnahmen von besonderer Bedeutung erscheinen. Natürlich kann an dieser Stelle keine ausführliche Einführung in die Anatomie und die Physiologie des ZNS dargestellt werden. Statt dessen soll versucht werden, nur die für das Verständnis der anstehenden Fragen notwendigsten Strukturen des ZNS anzubieten und die Zusammenhänge in einer Weise darzustellen, dass sie mit den gebotenen Grundkenntnissen zu verstehen sind. Für Leser, die sich - aufbauend auf solchen Grundkenntnissen – weiterbilden wollen, sei R.F. SCHMIDT: Medizinische Physiologie des Menschen, München 1983, empfohlen. Für naturwissenschaftlich geschulte Leser stehen eine Reihe von Lehrbüchern zur Verfügung, von denen hier beispielhaft SCHMIDT / THEWS / LANG: Physiologie des Menschen, Berlin 2000 und BIRBAUMER / SCHMIDT: Biologische Psychologie, Berlin 2003 genannt werden sollen.
5.1. Grundzüge der Struktur des Nervensystems und seiner motorischen Funktionen
Im Laufe der Evolution der Nervensysteme entstand aus einem einfach strukturierten Nervenrohr durch schrittweise Überlagerung höher organisierter, jeweils übergeordneter Schichten das menschliche Zentralnervensystem (Abb. 5.1) als ein hierarchisch strukturiertes Gebilde.
Gemäß dieser Entwicklungsschritte lassen sich 4 Schichten (von unten nach oben) voneinander trennen:
1. das Rückenmark, 2. der Hirnstamm mit dem Kleinhirn, 3. das entwicklungsgeschichtlich „alte“ Großhirn, 4. das entwicklungsgeschichtlich „neue“ Großhirn (genauer: neue Großhirnrinde).

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 4
Abb. 5.1: Halbschematische Darstellung der stammesgeschichtlichen Entwicklung des zentralen Nervensystems der Wirbeltiere. 1 Vorderhirn;
2 Mittelhirn; 3 Hinterhirn; 4 Rückenmark; 5 Zentralkanal; 6a „altes“ Großhirn; 6b „neues“ Großhirn; 7 Zwischenhirn; 8 Brückenhirn; 9 verlängertes Rückenmark; 10 Kleinhirn; 11 Längsspalt zwischen den Großhirnhemisphären; 12 Balken. 13 und 14 erste und zweite Gehirnkammer; 15 dritte Gehirnkammer; 16 vierte Gehirnkammer. Hirnstamm: 2+(7+) 8+9; Limbisches System: Teile von 6a und 7.
Während dieser Entwicklung erhielten die jeweils neuen (jüngeren, übergeordneten) Schichten bezüglich der Auslösung und Steuerung der Motorik jeweils neue Aufgaben, während die alten „untergeordneten“ Schichten einige Aufgaben teilweise oder ganz an übergeordnete Schichten abgaben, andere Aufgaben aber in größtmöglicher Eigenkompetenz beibehielten. Das auf diese Weise entstandene System funktioniert nach dem sogenannten Subsidiaritätsprinzip:
Das folgende Schema soll die Schichtenstruktur des zentralen Nervensystems verdeutlichen und ihre vorwiegenden Aufgaben erläutern:

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 5
Abb. 5.2: Schematische Darstellung der Schichten des zentralen Nervensystems und ihrer Aufgaben im Rahmen der Motorik
Das Zentralnervensystem ist ein Organ aus vielen Milliarden Nervenzellen mit ihren Nervenfasern, wobei die Nervenzellen und ihre Nervenfasern nicht gleichmäßig über das gesamte Nervensystem verteilt sind, sondern die Nervenzellen (genauer: ihre Zellkörper) sind als „graue“ Substanz von der „weißen“ Substanz, die ausschließlich aus den Nervenfasern der in der grauen Substanz liegenden Nervenzell-Körper besteht, getrennt.
Die hierarchisch angeordneten Schichten des menschlichen zentralen Nervensystems arbeiten bei der Steuerung der Bewegungen nach dem Subsidiaritätsprinzip: Für jede Schicht so viel Selbständigkeit wie möglich! Für jede übergeordnete Schicht so wenig helfende Ergänzung für die unteren Schichten wie notwendig!
Hirnstamm und „altes“ Kleinhirn
neue Großhirnrinde und „neues“ Kleinhirn
„altes“ Großhirn
Rückenmark einfache Bewegungsmuster, Muskelreflexe
Zielmotorik: Erwerbsmotorik
Zielmotorik: Erbmotorik
Orientierung im Raum, Stützmotorik

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 6
Abb. 5.3: Schematische Darstellung der Verteilung von grauer und weißer Substanz im Zentralnervensystem sowie derjenigen Hirnstrukturen, die im Folgenden zur Erklärung der Sensomotorik Verwendung finden. punktierte Bereiche: graue Substanz
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 7
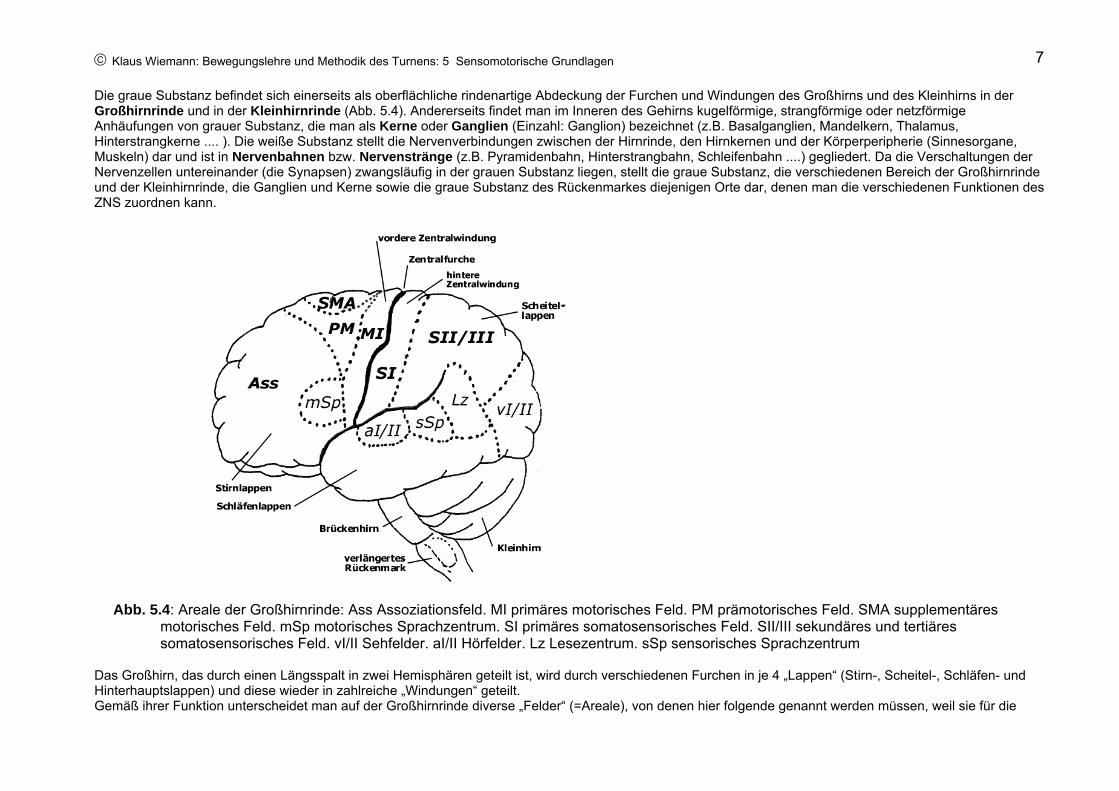
Die graue Substanz befindet sich einerseits als oberflächliche rindenartige Abdeckung der Furchen und Windungen des Großhirns und des Kleinhirns in der Großhirnrinde und in der Kleinhirnrinde (Abb. 5.4). Andererseits findet man im Inneren des Gehirns kugelförmige, strangförmige oder netzförmige Anhäufungen von grauer Substanz, die man als Kerne oder Ganglien (Einzahl: Ganglion) bezeichnet (z.B. Basalganglien, Mandelkern, Thalamus, Hinterstrangkerne .... ). Die weiße Substanz stellt die Nervenverbindungen zwischen der Hirnrinde, den Hirnkernen und der Körperperipherie (Sinnesorgane, Muskeln) dar und ist in Nervenbahnen bzw. Nervenstränge (z.B. Pyramidenbahn, Hinterstrangbahn, Schleifenbahn ....) gegliedert. Da die Verschaltungen der Nervenzellen untereinander (die Synapsen) zwangsläufig in der grauen Substanz liegen, stellt die graue Substanz, die verschiedenen Bereich der Großhirnrinde und der Kleinhirnrinde, die Ganglien und Kerne sowie die graue Substanz des Rückenmarkes diejenigen Orte dar, denen man die verschiedenen Funktionen des ZNS zuordnen kann.
Abb. 5.4: Areale der Großhirnrinde: Ass Assoziationsfeld. MI primäres motorisches Feld. PM prämotorisches Feld. SMA supplementäres
motorisches Feld. mSp motorisches Sprachzentrum. SI primäres somatosensorisches Feld. SII/III sekundäres und tertiäres somatosensorisches Feld. vI/II Sehfelder. aI/II Hörfelder. Lz Lesezentrum. sSp sensorisches Sprachzentrum
Das Großhirn, das durch einen Längsspalt in zwei Hemisphären geteilt ist, wird durch verschiedenen Furchen in je 4 „Lappen“ (Stirn-, Scheitel-, Schläfen- und Hinterhauptslappen) und diese wieder in zahlreiche „Windungen“ geteilt. Gemäß ihrer Funktion unterscheidet man auf der Großhirnrinde diverse „Felder“ (=Areale), von denen hier folgende genannt werden müssen, weil sie für die

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 8
Sensomotorik von besonderer bedeutung sind: 1. Primäre sensorische Felder (vI, aI, SI): Funktion: Bewusstwerden visueller, auditiver und somatosensorischer Empfindungen. Auf der primärsensorischen Rinde (SI) ist der gesamte Körper des Menschen „präsent“, jedoch nicht in der realen Proportion, sondern die einzelnen Körperabschnitte beanspruchen einen Platz gemäß der Wichtigkeit ihrer sensorischen Leistungen: sensorischer Homunkulus (Abb. 5.3, links oben). 2. Sekundäre und tertiäre sensorische Felder vII, aII, SII/III): Funktion: Verknüpfen visueller, auditiver und somatosensorischer Meldungen mit gespeicherten Informationen zur Bildung der Wahrnehmung (einschließlich Erkennen). 3. Supplementäres motorisches Feld (SMA): Funktion: Bilden von Bewegungsentwürfen bzw. Bewegungsvorstellungen. Höchste Instanz zur Kombination der Meldungen verschiedenster sensorischer Systeme zur Bewegungswahrnehmung. 4.Assoziationsfeld (Ass): Funktion: logisches Denken; Entwerfen motorischer Handlungsfolgen; strategische Entscheidungen. 5. Primäres motorisches Feld (MI; Ursprungsgebiet der Pyramidenbahn): Funktion: schnelle Innervation der Muskulatur, Bewegungsexekution, geringfügige Kurskorrektur der laufenden Bewegung. Entsprechend den Bedingungen bei SI lässt sich hier ein motorischer Homunkulus festlegen (Abb. 5.3, oben rechts).Zusätzlich sind zu nennen: 6. Lese- und Schreibzentrum (Lz): Funktion: Erkennen geschriebener Wörter; das Sinnverständnis wird jedoch erst in Kombination mit dem sensorischen Sprachzentrum (sSp) gewährleistet. 7. Sensorisches Sprachzentrum (sSp): Funktion: Sinnverständnis gehörter und gelesener Wörter und Sätze, Sprechen sinnvoller Wort- und Satzzusammenhänge. 8. Motorisches Sprachzentrum (mSp): Funktion: Sprechen und Schreiben längerer Wortkombinationen und vollständiger, grammatikalisch richtiger Sätze.
5.2 Motorische Aufgaben des Rückenmarkes: Dehnungs- und Spannungsreflexe
Die "unterste" Koordinationsinstanz für die Motorik bildet das Rückenmark. Seine koordinative Leistung liegt in der Produktion einiger recht einfacher, aber höchst bedeutungsvoller Reflexe und Reaktionen, ohne die es dem Organismus unmöglich wäre, sinnvolle und zielgerichtete Bewegungen zu realisieren. Dazu gehören die Dehnungs- und Spannungsreflexe, sowie die Beuge- und Streckreflexe.
5.2.1 Die Dehnungsrezeptoren des Muskels
Die Sensoren für die Längenkontrolle des Muskels sind die Muskelspindeln. Diese stellen wenige Millimeter lange sensorische Organe dar, die parallel an den Faserhüllen der die Kontraktionsarbeit verrichtenden Muskelfasern (den Arbeitsmuskelfasern)
Die wichtigsten motorischen Aufgaben des Rückenmarkes sind: - Dehnungsreflexe und Längenkontrollsystem des Muskels, - Spannungsreflexe sowie - Beuge- und Streckreflexe.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 9
angeheftet sind (Abb. 5.5). Die Muskelspindeln enthalten in einer Kapsel kleine längsgerichtete Muskelfasern, die Spindelfasern (intrafusale Fasern), die zwei kontraktionsfähige Pole und ein dehnfähiges Mittelstück besitzen. Durch die Art der Anheftung an der Faserhülle der Arbeitsmuskelfasern ist gewährleistet, dass bei einer Dehnung des Muskels die Muskelspindelfasern, speziell ihre Mittelstücke, mit gedehnt werden bzw. bei einer konzentrischen Kontraktion des Muskels sich entdehnen können. Um das Mittelstück (die Äquatorialregion) einer jeden Muskelspindelfaser sind spiralförmige Rezeptoren gewickelt, die auf Dehnung reagieren und die Meldung über den Dehnungszustand des Muskels auf dem Wege über sensorische Nerven (Ia- und II-Fasern) dem ZNS zuführen. Die kontraktionsfähigen Pole der Muskelspindelfasern erhalten auf dem Weg über motorischen Nervenfasern (γ-Fasern) Kontraktionsbefehle vom ZNS. Auf diese Weise kann durch eine Kontraktion der beiden Spindelpole das Mittelstück der Muskelspindelfaser auch bei einem entdehnten bzw. durch Kontraktion verkürzten Muskel in einem funktionsfähigen Dehnungszustand gehalten werden.
Abb. 5.5: Schematische Darstellung des Aufbaues und der nervösen Versorgung der Muskelspindel. 1 Hülle der Arbeitsmuskelfaser. 2 Kapsel der Muskelspindel. 3 kontraktile Pole der Kernsack-Faser der Muskelspindel. 4 kontraktile Pole der Kernketten-Fasern der Muskelspindel. 5 sensible Äquatorregion der Kernsack-Faser. 6 sensible Äquatorregion der Kernketten-Faser. 7 schnelle afferente Nervenfaser (Ia-Faser). 8 afferente Nervenfasern (II-Fasern) der Kernketten-Fasern. 9 Dehnungsrezeptor der Ia-Nervenfaser (Primärrezeptor). 10 und 11 Dehnungsrezeptoren der II-Nervenfasern (Sekundärrezeptoren). 12 motorische Nervenfasern (γ-Fasern). 13 motorische Endplatten der γ-Fasern. 14 motorischen Endnetze der γ-Fasern.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 10
Die Ia-Rezeptoren der Kernsack-Fasern reagieren vorwiegend bei schnellen Längenänderungen während die Rezeptoren der Kernketten-Fasern fast ausschließlich den Ist-Zustand der Muskellänge messen. Das bedeutet, dass die Kernsack-Fasern vorwiegend dynamische, die Kernketten-Fasern statische Informationen übermitteln. Während die dynamischen Informationen speziell für die im Folgenden zu besprechenden Dehnungsreflexe von Bedeutung sind, liefern die statischen Informationen die Basis für das Längenkontrollsystem des Muskels ( Kap. 5.2.4).
5.2.2 Der Dehnungsreflex und die Längenregelung des Muskels
Wird ein Muskel entgegen der vom Gehirn vorgesehenen Länge, die über eine längere oder kürzere Zeit beizubehalten ist, durch eine äußere Wirkung gedehnt, antwortet der Muskel mit einer schnellen unwillkürlichen Reaktion, um die beabsichtigte Länge wiederherzustellen. Diese Reaktion des Muskels ist der Dehnungsreflex (auch: Eigenreflex), der allein in der Verantwortung des Rückenmarkes steht.
Die motorische Bedeutung des Dehnungsreflexes, den jeder in Form des Kniesehnenreflexes kennt, soll an folgendem Beispiel erläutert werde:
Allgemein ist ein Reflex eine automatische Reaktion eines Muskels bzw. einer Muskelgruppe auf einen Reiz ohne Einfluss des bewussten Wollens. Dabei ist der Reflexbogen die Verbindung einer Sinneszelle (eines „Rezeptors“, eines Sensoren) oder eines Sinnesorgans auf dem Wege über das zentrale Nervensystem mit einem Erfolgsorgan (hier: Muskel oder Muskelgruppe).
Eine Aufgabe der Muskelspindeln ist das ständige Überwachen der Länge (des Dehnungszustandes) des gesamten Muskels, um a) äußere Störgrößen rechtzeitig zu entdecken und b) dem ZNS eine permanente Information über die augenblickliche Muskellänge (den Dehnungsgrad) zu liefern.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 11
Will eine Turnerin am Boden die Bewegungsfolge Radwende-Salto rückwärts turnen, muss sie während des Bodenkontaktes der Füße zwischen Radwende und Salto durch die Wadenmuskeln einen kurzen, aber starken Absprung-Kraftstoß erzeugen. Nimmt man an, dass die Turnerin während der Bewegungsfolge eine Horizontalgeschwindigkeit (v) von 5 m/s besitzt und ihr Körperschwerpunkt während des Bodenkontaktes einen Weg von 0,5 m zurücklegt, bleibt ihr für die Erzeugung des Absprungkraftstoßes eine Zeitspanne von 0,1 s.
Abb. 5.6: Beispiel eines prellenden Absprunges mit kurzer Bodenkontaktzeit
Diese kurze Zeit reicht nicht aus, mit dem Beginn des Bodenkontaktes eine Willkürkontraktion der Sprungmuskeln zu entwerfen und erfolgreich durchzuführen; dazu würde das Nervensystem einen deutlich längeren Zeitraum benötigen (Kap. 5.4; Abb. 5.41). Statt dessen muss die Turnerin schon beim Auftreffen der Füße auf den Boden die Sprungmuskeln, insbesondere die Wadenmuskulatur maximal kontrahiert haben, so dass der Körper durch einen elastischen Stoß (= durch die Elastizität der angespannten Wadenmuskeln) vom Boden abprellt (s. Kap. 3.11). In der Zeit des Bodenkontaktes zeigt somit die kontrahierte Wadenmuskulatur eine kurze durch den Bodenwiderstand erzeugte Dehnung und eine entsprechende elastische Entdehnung (s. Abb. 3.83). Etwa 0,03 s (= 30 Millisekunden, ms) nach dem Auftreffen auf den Boden ist die Dehngeschwindigkeit stark genug, um die Entstehung eines

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 12
Dehnungsreflexes in der Wadenmuskulatur auszulösen, der unter günstigen Bedingungen den elastischen Stoß durch eine reflektorische Kontraktion unterstützen kann.
In den Wadenmuskeln reagieren die Muskelspindeln auf die Dehnung und senden über die zugehörigen sensorische Nervenfasern eine kurze Serie von Nervenimpulsen zum Rückenmark. Hier werden auf dem Wege über Nervenkontakte (Synapsen) motorische Nervenzellen angeregt, eine kurze Serie von Impulsen zurück zur Wadenmuskulatur zu schicken (Abb. 5.7), die daraufhin (etwa 30 ms nach Dehnung der Rezeptoren) durch eine zusätzliche Kontraktion den Absprungkraftstoß verstärkt.
Abb. 5.7: Schematische Darstellung des Dehnungsreflexes. Pfeile: Richtung der Nervenleitung
Da in diesem Reflexbogen die sensorische Nervenzelle jeweils nur mittels einer Synapse direkt mit der motorischen Nervenzelle verbunden ist, spricht man hier auch vom monosynaptischen Dehnungsreflex. Aufgrund einer Leitungsgeschwindigkeit der beteiligten Nervenfasern von rund 100 m/s benötigt der Reflex im obigen Beispiel eine Laufzeit von rund 25 ms, so dass etwa 60 ms nach Aufsetzen der Füße auf den Boden (rd. 30 ms Dehnungszeit + rd. 30 ms Reflexzeit) die reflektorische Kontraktion in der Wadenmuskulatur beginnen und die Absprungaktion rechtzeitig im Laufe der Entdehnungsphase unterstützen kann. Da im Leistungsturnen die Bewegungsgeschwindigkeiten häufig wesentlich höher als im vorliegenden Beispiel und die Kontaktzeiten entsprechend kürzer sind, ist zu vermuten, dass hier der Dehnungsreflex zur Unterstützung prellender Absprünge „zu spät“ kommt. D.h., der Turner ist beim Absprung allein auf die Elastizität der Kontrahierten Muskelfasern angewiesen.
Querschnitt durch das Rückenmark
Muskelspindel
Wadenmuskel
sensorische Nervenfaser
motorische Nervenfaser
Synapse
motorische Endplatte
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 13
Selbstverständlich ist bei diesem und in Abb. 5.7 dargestellten Geschehen nicht nur eine Muskelspindel, eine sensorische und eine motorische Nervenfaser beteiligt, wie man aus Abb. 5.7 (sowie Abb. 5.9 und Abb. 5.11 u.a.) aufgrund der vereinfachten Darstellung schließen könnte. Da die Wadenmuskulatur aus mehreren 100 motorischen Einheiten besteht ( eine motorische Einheit enthält ein Motoneuron mit einer motorischen Nervenfaser und mehreren 100 Muskelfasern, die von dem Motoneuron innerviert werden), werden entsprechend viele Muskelspindeln und sensorische Nervenfasern benötigt.
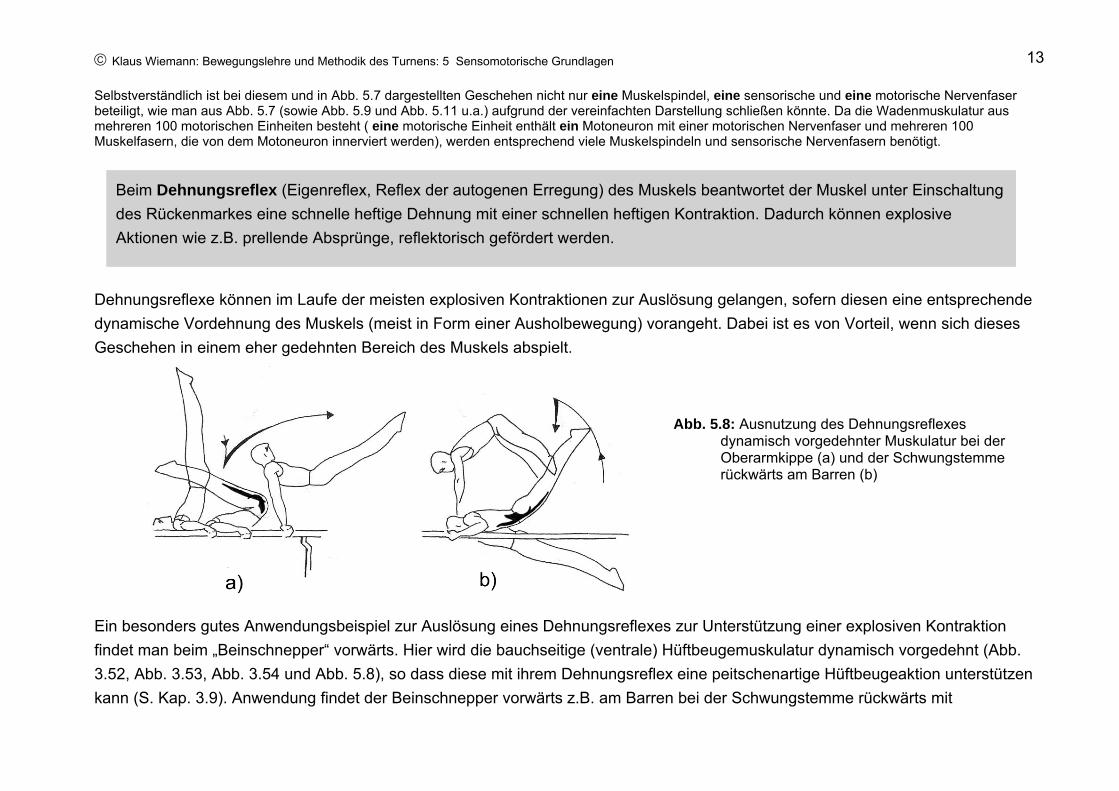
Dehnungsreflexe können im Laufe der meisten explosiven Kontraktionen zur Auslösung gelangen, sofern diesen eine entsprechende dynamische Vordehnung des Muskels (meist in Form einer Ausholbewegung) vorangeht. Dabei ist es von Vorteil, wenn sich dieses Geschehen in einem eher gedehnten Bereich des Muskels abspielt.
Ein besonders gutes Anwendungsbeispiel zur Auslösung eines Dehnungsreflexes zur Unterstützung einer explosiven Kontraktion findet man beim „Beinschnepper“ vorwärts. Hier wird die bauchseitige (ventrale) Hüftbeugemuskulatur dynamisch vorgedehnt (Abb. 3.52, Abb. 3.53, Abb. 3.54 und Abb. 5.8), so dass diese mit ihrem Dehnungsreflex eine peitschenartige Hüftbeugeaktion unterstützen kann (S. Kap. 3.9). Anwendung findet der Beinschnepper vorwärts z.B. am Barren bei der Schwungstemme rückwärts mit
Beim Dehnungsreflex (Eigenreflex, Reflex der autogenen Erregung) des Muskels beantwortet der Muskel unter Einschaltung des Rückenmarkes eine schnelle heftige Dehnung mit einer schnellen heftigen Kontraktion. Dadurch können explosive Aktionen wie z.B. prellende Absprünge, reflektorisch gefördert werden.
Abb. 5.8: Ausnutzung des Dehnungsreflexes dynamisch vorgedehnter Muskulatur bei der Oberarmkippe (a) und der Schwungstemme rückwärts am Barren (b)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 14
Übergrätschen der Beine (Abb. 5.8), am Reck bei der Stemme rückwärts mit Bücke, dem Jägersalto gebückt, dem Doppelsato rückwärts gehockt über die Stange (Kovacs, Abb. 3.47) u.a.
Beim „Beinschnepper“ rückwärts kann dieses Prinzip jedoch nicht ausgenutzt werden. Das ausholende Hüftbeugen führt nicht zu einer genügend tiefen Hüftbeugestellung, in der die Hüftstreckmuskeln einen hinreichend starken Dehnungsreiz erfahren würden. Gleiches scheint auch für weitere Hüftstreckbewegungen zu gelten. Einzig bei der Oberarmkippe am Barren könnte eine tiefe Oberarmkipplage – sofern sie dynamisch eingenommen wird – die Hüftstreckmuskeln genügen stark vordehnen, um eine kräftige Kippstreckung reflektorisch zu unterstützen (Abb. 5.8).
5.2.3 Reflex der antagonistischen Hemmung
Auch der Reflex der antagonistischen Hemmung ist ein Dehnungsreflex. Seine wesentliche Aufgabe liegt darin, dafür Sorge zu tragen, dass die in einem Muskel durch einen Dehnungsreflex erzeugte Aktion durch seinen Gegenspieler (den Antagonisten) nicht gestört wird. Dies wird dadurch bewerkstelligt, dass von den sensorischen Nervenfasern des gedehnten Muskels (Agonist) im Rückenmark Fasern abzweigen, welche ihrerseits hemmende Nervenzellen aktivieren. Diese wiederum verhindern durch ihre hemmende Wirkung, dass die motorischen Nervenzellen des Gegenspielers in der Phase, in der der Agonist die reflektorische Kontraktion durchführt, den Gegenspieler zu einer Kontraktion anregen und dadurch die Aktion des Agonisten behindern könnten (Abb. 5.9)
Im vorliegenden Beispiel muss der vordere Schienbeinmuskel als Gegenspieler der Wadenmuskulatur angesehen werden.
Will man im Laufe turnerischer Fertigkeiten die Dynamik von Hüftaktionen durch Ausnutzung des Dehnungsreflexes steigern, muss der Aktion eine dynamische Ausholbewegung vorangehen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 15
Während die besprochnen Dehnungsreflexe durch die dynamischen Rezeptoren innerhalb der Muskelspindeln ausgelöst werden, wird das Längenkontrollsystem durch die statischen Rezeptoren der Muskelspindeln bestimmt. Sie sind der Ausgangspunkt für einen Servomechanismus, das Längenkontrollsystem der Muskulatur. Ähnlich wie man eine gewünschte Raumtemperatur am Thermostaten des Heizkörpers einstellen kann, wird die bei einer jeden Haltung notwendige Länge eines Muskels von den motorischen Instanzen des Gehirns durch die Einstellung der Empfindlichkeit der Muskelspindelrezeptoren vorbestimmt. Auf dem Wege über die Dehnungsreflexe wird nun dafür gesorgt, dass die Muskellänge beibehalten und damit die gewünschte Haltung oder Stellung (eine Standwaage, ein Schwebestütz oder einfach nur ein aufrechter Stand) eingehalten und eine durch äußere Störgrößen bedingt Abweichung möglichst unverzüglich beseitigt wird.
[5.4] Beim Reflex der antagonistischen Hemmung (Reflex der reziproken Hemmung) wird unter Einschaltung des Rückenmarkes bei einer schnellen heftigen Dehnung eines Muskels, die zu einem Dehnungsreflex dieses Muskels führt, der Gegenspieler kurzfristig gehemmt. Dadurch kann reflektorisch sichergestellt werden, dass explosive Aktionen wie z.B. prellende Absprünge, nicht durch die gleichzeitige Kontraktion der Gegenspieler gestört werden.
Abb. 5.9: Schematische Darstellung des Reflexes der antagonistischen Hemmung

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 16
Auch im Laufe einer jeden Bewegung ist auf die Wirkung der Dehnungsreflexe nicht zu verzichten. Hier verändern die motorischen Hirninstanzen die Empfindlichkeit der Dehnungsrezeptoren der Muskelspindeln fortlaufend, so dass auf dem Wege der Dehnungsreflexe die Muskellänge diesen Veränderungen fortlaufend folgt und dadurch die gewünschte Bewegung garantiert wird: Folge-Servomechanismus.
5.2.4 Spannungsreflex und Spannungsregelung des Muskels
Tritt in einem Muskel, der sich gerade in einem Kontraktionszustand befindet, durch äußere Einflüsse kurzfristig eine hohe Spannung auf, kann unter Mithilfe des Rückenmarkes die Kontraktion für kurze Zeit (wenige Millisekunden) abgeschwächt oder gar völlig unterbrochen werden. Dadurch werden Muskel und Sehne kurzfristig entlastet und vor einer Überlastung und einer damit möglicherweise verbundenen Schädigung geschützt.
In dem oben angeführten Beispiel könnte die Turnerin beispielsweise beim Absprung zum Salto mit den Füßen auf einen unerwartet unelastischen Boden treffen. Da sich die Wadenmuskulatur schon kurz vor dem Auftreffen der Fußballen auf den Boden in einem hohen Kontraktionszustand befindet, könnte sich beim Aufsetzen der Füße in der Wadenmuskulatur kurzfristig eine zu hohe Spannung entwickeln. In diesem Falle reagieren in den Wadenmuskeln die Sehnenspindeln (Golgi-Sehnenorgane), Sensoren bzw. Rezeptoren am Übergang der Muskelfasern zur Sehne, und senden über sensorische Nervenfasern eine kurze Serie von Nervenimpulsen zum Rückenmark. Hier werden auf dem Wege über Nervenkontakte (Synapsen) hemmende Nervenzellen angeregt, die mit den motorischen Nervenzellen der Wadenmuskulatur Kontakt haben (s. auch Abb. 5.9). Auf diese Weise werden die motorischen Nervenzellen der Wadenmuskeln kurzfristig gehemmt, wodurch sich die Kontraktion der Wadenmuskeln abschwächt oder für wenige Millisekunden sogar völlig unterbrochen wird. Dies reicht in den meisten Fällen aus, die Sehne vor einer Schädigung zu bewahren. Die Golgi- Sehnenorgane werden auch als Rezeptoren eines Spannungsregelsystems angesehen, das zusammen mit dem Servomechanismus der Längenregelung eine funktionelle Einheit bildet. Wenn man bedenkt, dass die von den motorischen Zentren des Gehirns an die Muskulatur geschickten Kontraktionsbefehle zur Ausführung einer Bewegung im Grunde nichts anderes bedeuten , als in bestimmten Zeitabschnitten bestimmte Muskellängen und Längenänderungen gegen äußere Kräfte einzustellen, wird deutlich, dass die Spannungsregelung eine bedeutungsvolle Ergänzung in der Absicherung der Bewegungsabläufe gegenüber äußeren oder inneren Störgrößen (z.B. auch schwindender Kraft bei Ermüdung) darstellt.
Auch innerhalb der Spannungskontrolle der Muskulatur findet man einen Einfluss auf den Gegenspieler (Reflex der reziproken bzw. antagonistischen Erregung): Von den sensorischen Fasern, die vom Agonisten zum Rückenmark ziehen, zweigen im Rückenmark Fasern ab, die erregende Nervenzellen aktivieren. Letztere üben auf die motorischen Nervenzellen des Gegenspielers (Antagonisten) einen erregenden Einfluss aus, so dass der Gegenspieler kontrahiert. Man findet hier somit eine doppelte Sicherung: Bei einem unvorhergesehenen Spannungsanstieg im Agonisten wird nicht nur dieser kurzfristig deaktiviert und dadurch die

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 17
Spannung im Agonisten gemildert, sondern durch die zusätzliche Kontraktion des Antagonisten wird die auf den Agonisten wirkende äußere Spannung zusätzlich reduziert.
5.2.5 Beuge- und Streckreflexe
Ähnliche Schutzfunktionen wie der Spannungsreflex übernehmen die Beuge- und Streckreflexe, die auch über das Rückenmark - also vorwiegend ohne Beteiligung des Gehirns – ablaufen. Tritt beispielsweise ein Turner beim Anlauf zu einer akrobatischen Folge auf eine Unebenheit, etwa den Mattenrand, könnte die Gefahr des Fußumknickens mit nachfolgenden Verletzungen auftreten (in Abb. 5.10 beispielsweise mit dem linken Bein). In diesem Fall wird ein Reflexgeschehen ausgelöst, das dazu dient, den (linken) Fuß aus dem Gefahrenbereich zu entfernen:
Rezeptoren in der Haut, im Bindegewebe oder direkt an den Gelenken, die durch die besondere Situation gereizt werden, schicken eine entsprechende Meldung zum Rückenmark (Abb. 5.10 a). Von hier werden die Meldungen einerseits zum Gehirn weitergeleitet, andererseits aber direkt im Rückenmark auf dem Wege über mehrere erregende und hemmende Schaltzellen zu den motorischen Nervenzellen des betroffenen (linken) Beines weitergeleitet, und zwar in der Weise, dass die Streckmuskeln, die gerade aktiv sind , um die Stützphase des Laufschrittes auszuführen, gehemmt und somit daran gehindert werden, diese Aufgabe zu Ende zu führen (Abb. 5.11). Gleichzeitig werden die motorischen Nervenzellen der Beugemuskeln des gleichen Beines aktiviert, damit diese das Bein im Hüft- und Kniegelenk beugen und so aus der Gefahrenzone entfernen (Abb. 5.10 b).
Da bei diesem Reflexgeschehen die Informationen oft aus mehreren Rezeptorsystemen stammen, im Rückenmark über mehrere Schalt-Nervenzellen laufen und sogar in andere Körpersegmente auf- oder absteigen müssen (Abb. 5.11), da zudem oft noch unterschiedliche Muskeln zu hemmen bzw. zu aktivieren sind, ist die Zeit vom Reiz bis zum Beginn der Reaktion (die Reflexzeit) mit
[Beim Spannungsreflex (Reflex der autogenen Hemmung) des Muskels beantwortet der Muskel unter Einschaltung des Rückenmarkes einen schnellen heftigen Spannungsanstieg mit einer schnellen kurzfristigen Unterbrechung der Kontraktion. Dadurch wird ein Schutz des Muskels vor Verletzungen gewährleistet.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 18
40 ms bis 60 ms deutlich länger als z.B. der Dehnungsreflex – aber immer noch kurz genug, um in den meisten Fällen eine Verletzung zu verhindern.
Bei diesem Reflexgeschehen, das sowohl die Beuge- als auch die Streckmuskeln desselben Beines tangiert, steht funktional die Beugeaktion im Vordergrund, deshalb die Bezeichnung Beugereflex. Da der Beugereflex die Gliedmaße auf derselben Körperseite beeinflusst, auf der auch der auslösende Reiz wirkt, lautet die vollständige Bezeichnung: gleichseitiger (bzw. ipsilateraler) Beugereflex. Da er im Gegensatz zum Dehnungsreflex (= monosynaptischer Reflex) einerseits und dem Reflex der antagonistischen Hemmung und den Spannungsreflexen (= bisynaptische Reflexe) über mehr als zwei Synapsen laufen muss, spricht man auch von polysynaptischen Reflexen.
Zusätzlich zum Beugen des Beines kann die Reaktion auch noch von einem unterstützenden Beugen des Rumpfes und der Arme und einem Neigen des Kopfes (Beugen der Halswirbelsäule) begleitet sein. Eine turnerisch korrekte Bewegungsweise wird dadurch deutlich unterbrochen.
ab)
Abb. 5.10: Darstellung des Informationsflusses beim Beugereflex (linkes Bein) und beim gekreuzten Streckreflex (rechtes Bein)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 19
Abb. 5.11: Schematische Darstellung der Nervenverschaltung zum Beugereflex (rechts) und zum gekreuzten Streckreflex (links)
Beim gleichseitigen Beugereflex (Flexorreflex) wird durch Reizung von Haut- Schmerz- oder Gelenkrezeptoren die Beugemuskeln einer Gliedmaße aktiviert und die Streckmuskeln derselben Gliedmaße gehemmt und auf diese Weise die Gliedmaße gebeugt und somit eine evtl. gerade ablaufende Streckaktionen unterbrochen, um die Gliedmaße aus einem Gefahrenbereich zu entfernen.
sensorischeNervenfasern sensorische
Nervenfasern
sensorische Nervenfasern innerhalb des Rückenmarkes
Streckmuskel (gehemmt)
motorische N f
erregende und hemmende (schwarz) Schaltzellen
Streckmuske Beugemuskel
Beugemuskel (gehemmt)
Querschnitte durch das Rückenmark
motorische Nervenzellen
motorische Nervenfasern

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 20
Sinnvoller Weise ist dieser Beugereflex mit einem Reflex gekoppelt, der vorwiegend zur Sicherung oder Wiederherstellung der aufrechten Körperposition beiträgt, dem gekreuzten Streckreflex. Die (in unserem Beispiel auf der linken Körperseite) einlaufenden sensorischen Meldungen werden im Rückenmark direkt auch auf die andere Körperseite (hier: die rechte) übertragen und erzeugen dort entgegengesetzte Reaktionen: die Beugemuskeln werden gehemmt und die Streckmuskeln aktiviert (Abb. 5.11, linke Hälfte der Abbildung) und somit eine gerade ablaufende Beugeaktion des (rechten Beines) abgebrochen und statt dessen eine Streckaktion in Gang gesetzt (Abb. 5.10 b). Auf diese Weise wird sichergestellt, dass der Körper mit Auslösung des Beugereflexes nicht in sich zusammenbricht, sondern das Gegenbein schnell auf den Boden setzt.
Im Übrigen funktionieren der ipsilaterale Beugereflex und der gekreuzte (kontralaterale) Streckreflex bedeutungsgemäß auch in den oberen Extremitäten. In biologischen Lehrbüchern wird als Beispiel für diesen Funktionszusammenhang häufig das Greifen auf eine heiße Herdplatte verwendet, es fällt jedoch nicht schwer, entsprechende Beispiele beim Gerätturnen , z.B. bei den verschiedensten Stützaktionen, zu finden.
Die Meldungen, die aus den genannten Rezeptoren zum Rückenmark gesendet werden, um die besprochenen Reflexe in Gang zu setzen, werden – wie schon angedeutet – auch dem Gehirn zugeleitet. In dem Augenblick, in dem diese Informationen im Gehirn ankommen und zu einer bewussten Wahrnehmung verarbeitet werden, sind die Reflexe jedoch schon in Gang gesetzt. Es würde drei mal so viel Zeit wie beim Beugereflex vergehen (mindestens 150 ms), wenn der Körper darauf warten müsste, bis das Gehirn die Meldung analysiert und entschieden hätte, wie zu reagieren wäre. Eine entsprechende Verletzung wäre nicht mehr zu vermeiden. Trotzdem hat der Betroffene den Eindruck, die Geschehnisse wären wie folgt abgelaufen: Wahrnehmen des Schmerzes – Durchführung der Schutzreaktion.
Beim gekreuzten Streckreflex wird - parallel zum Beugereflex in der einen Gliedmaße - die Streckmuskeln der Gliedmaße der anderen Körperseite aktiviert und die Beugemuskeln gehemmt und auf diese Weise durch ein schnelles Strecken dieser Gliedmaße ein Beibehalten der Körperstellung abgesichert.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 21
5.3 Stützmotorik
Unter dem Begriff der Stützmotorik wird eine Gruppe von Bewegungsphänomenen zusammengefasst, die zwei wesentliche Aufgaben innerhalb der Motorik erfüllen:
Für die Durchführung der Stützmotorik ist vorwiegend die zweite, dem Rückenmark übergeordnete Schicht des Zentralenervensystems zuständig, das Stammhirn und Teile des Kleinhirns (des stammesgeschichtlich „alten“ Kleinhirns, Abb. 5.2). Die Rezeptoren, die Ausgangspunkt der Körperstellreflexe darstellen, sind die Muskelspindeln der Augenmuskulatur, die Rezeptoren des Gleichgewichtssinnes (des Labyrinthes), die Muskel- und Sehnenspindeln der Hals- und Rückenmuskulatur sowie die Gelenkrezeptoren vorwiegend der Halswirbelsäule sowie letztendlich die Hautrezeptoren und die Rezeptoren der Tiefensensibilität der Gliedmaßen. Die Reflexbögen verlaufen teils über den Hirnstamm, teils über das Rückenmark (Abb. 5.12). Als Erfolgsorgan kommt die gesamte Skelettmuskulatur, einschließlich der Augenmuskeln in Betracht.
Die Stützmotorik a) garantiert, dass der Organismus seine aufrechte Körperhaltung beibehält oder - nach einer Störung - wieder einnimmt, und b) gewährleistet, dass der Körper dem sich mit Hilfe der Fernsinne in der Umwelt orientierenden Kopf folgt.
Großhirn
Augenmuskeln
Labyrinth Halswirbelsäule
Hirnstamm + Kleinhirn
Rückenmark
Rückenwirbelsäule
Abb. 5.12: Verdeutlichung des kettenartigen Verlaufes der Reflexbögen der Stützmotorik

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 22
Da verschiedene Mechanismen der Stützmotorik für die strukturgerechte Ausführung vieler turnerischer Fertigkeiten sowie für die Planung lehrmethodischer Maßnahmen von Bedeutung sind, müssen sie hier ausführlicher besprochen werden.
Die Mechanismen der Stützmotorik funktionieren bei Mensch und Tier in gleicher Weise. Sie lassen sich jedoch beim Tier deutlicher beobachten, weil beim erwachsenen, gesunden Menschen der größte Teil der Mechanismen der Stützmotorik durch übergeordnete Regelungen überformt ist. Aus diesem Grunde soll die Beschreibung stützmotorischer Äußerungen jeweils mit Beispielen begonnen werden, die aus Untersuchungen des Verhaltens von Vierfüßern gewonnen wurden. In der sportmethodischen Literatur werden diese Mechanismen gelegentlich angesprochen und mit dem Schlagwort Halstonusreflexe gekennzeichnet. Diese Bezeichnung wird der Komplexität der Stützmotorik jedoch nicht gerecht und bezieht sich nur auf einen beschränkten Ausschnitt aus dem gesamten Geschehen. Statt dessen müssen hier vier Funktionen angesprochen werden, die sich teils wechselseitig beeinflussen (Abb. 5.13).
Auge
Augenmuskeln
Netzhaut (Bildverschiebung)
Dehnungsrezeptoren der Augenmuskeln
Großhirn (zielmotorisches Blicken)
Äußere Störeinflüsse
Halsmuskeln / Dehnungsrezeptoren
Labyrinth
Rumpfmuskeln / Dehnungsrezeptoren
Extremitäten
Tast- und Druck-rezeptoren
Abb. 5.13: Wirkungsmechanismen innerhalb der Stützmotorik: Dicke Pfeile: zielmotorischer Eingriff. Gestrichelte Pfeile: optische Orientierungsreflexe. Punktierte Pfeile: Labyrinth-Stellreflexe. Dünne Pfeile: taktil ausgelöste Stellreaktionen. Grau unterlegt: Tonusreflexe
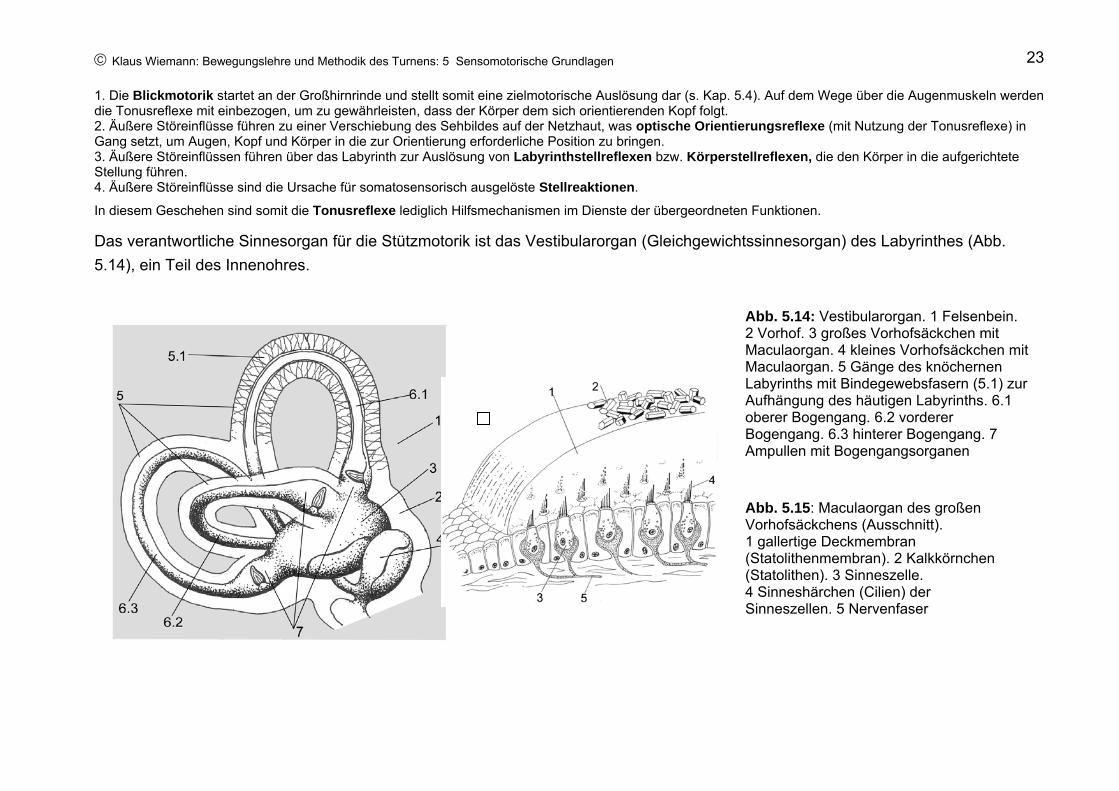
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 23
1. Die Blickmotorik startet an der Großhirnrinde und stellt somit eine zielmotorische Auslösung dar (s. Kap. 5.4). Auf dem Wege über die Augenmuskeln werden die Tonusreflexe mit einbezogen, um zu gewährleisten, dass der Körper dem sich orientierenden Kopf folgt. 2. Äußere Störeinflüsse führen zu einer Verschiebung des Sehbildes auf der Netzhaut, was optische Orientierungsreflexe (mit Nutzung der Tonusreflexe) in Gang setzt, um Augen, Kopf und Körper in die zur Orientierung erforderliche Position zu bringen. 3. Äußere Störeinflüssen führen über das Labyrinth zur Auslösung von Labyrinthstellreflexen bzw. Körperstellreflexen, die den Körper in die aufgerichtete Stellung führen. 4. Äußere Störeinflüsse sind die Ursache für somatosensorisch ausgelöste Stellreaktionen.
In diesem Geschehen sind somit die Tonusreflexe lediglich Hilfsmechanismen im Dienste der übergeordneten Funktionen.
Das verantwortliche Sinnesorgan für die Stützmotorik ist das Vestibularorgan (Gleichgewichtssinnesorgan) des Labyrinthes (Abb. 5.14), ein Teil des Innenohres.
Abb. 5.14: Vestibularorgan. 1 Felsenbein. 2 Vorhof. 3 großes Vorhofsäckchen mit Maculaorgan. 4 kleines Vorhofsäckchen mit Maculaorgan. 5 Gänge des knöchernen Labyrinths mit Bindegewebsfasern (5.1) zur Aufhängung des häutigen Labyrinths. 6.1 oberer Bogengang. 6.2 vorderer Bogengang. 6.3 hinterer Bogengang. 7 Ampullen mit Bogengangsorganen
Abb. 5.15: Maculaorgan des großen Vorhofsäckchens (Ausschnitt). 1 gallertige Deckmembran (Statolithenmembran). 2 Kalkkörnchen (Statolithen). 3 Sinneszelle. 4 Sinneshärchen (Cilien) der Sinneszellen. 5 Nervenfaser

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 24
Das Vestibularorgan besitzt zwei nahezu senkrecht zueinander angeordnete Sinnesfelder (Maculaorgane, Gleichgewichtssinnesorgane) zur Registrierung von Linearbeschleunigungen (somit auch Erdbeschleunigung) und drei Bogengangsorgane zur Registrierung von Drehbeschleunigungen, die sich in je einer Ampulle an der Basis von drei jeweils senkrecht aufeinander stehenden Bogengängen befinden (Abb. 5.14 und Abb. 5.16 b).
5.3.1 Regelung des aufrechten Standes
Steht ein vierfüßiges Säugetier in mittlerer Winkelstellung der Gliedmaßengelenke der Gliedmaßen mit Blick geradeaus nach vorn auf einer Unterlage und kippt diese Unterlage um einen bestimmten Winkel nach vorn, werden charakteristische, kettenartig ablaufende Reaktionen zu beobachten sein (Abb. 5.17):
b)a)
Die Gleichgewichtssinnesorgane (= die Maculaorgane) registrieren die Lage des Kopfes in Bezug zur Erdbeschleunigung und senden bei Abweichung von der Normalstellung Informationen an den Hirnstamm, der daraufhin die aufrechte Körperhaltung reflektorisch wiederherstellt.
Abb. 5.16: a: Mechanik der Maculaorgane (1 - 5: s. Abb. 5.15). F und F’: Horizontalbeschleunigung. Fg: Erdbeschleunigung mit tangentialer (Ft) und normaler Komponente (Fn). b: Mechanik des Bogengangsorgan. 1 Wand des häutigen Bogengangs. 2 Ampulle. 3 Sinnesleiste mit Sinneszellen. 4 Cupula. Ausgezogener Pfeil: Drehung des Kopfes und des Bogenganges. Gestrichelte Pfeile: Bewegung der Endolymphe in Bezug zum Bogengang.
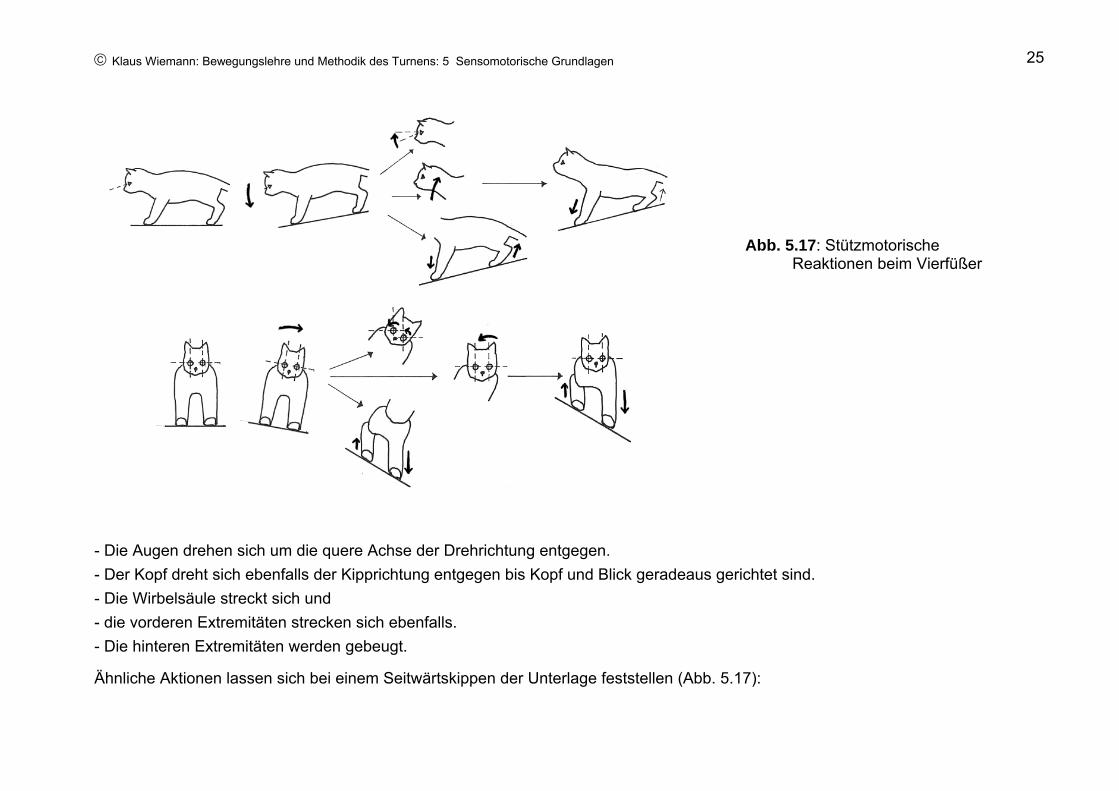
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 25
- Die Augen drehen sich um die quere Achse der Drehrichtung entgegen. - Der Kopf dreht sich ebenfalls der Kipprichtung entgegen bis Kopf und Blick geradeaus gerichtet sind. - Die Wirbelsäule streckt sich und - die vorderen Extremitäten strecken sich ebenfalls. - Die hinteren Extremitäten werden gebeugt.
Ähnliche Aktionen lassen sich bei einem Seitwärtskippen der Unterlage feststellen (Abb. 5.17):
Abb. 5.17: Stützmotorische Reaktionen beim Vierfüßer

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 26
- Die Augen drehen sich um die Tiefenachse der Drehrichtung entgegen. - Der Kopf richtet sich in die Senkrechte auf. - Die Extremitäten der "Innenseite" werden gebeugt, während die Extremitäten der "Außenseite" gestreckt werden.
Ausgangspunkt für diese Regulationen der Körperstellung sind einerseits die Gleichgewichtsorgane (das Labyrinth), die reflexartig bewirken, dass sich die Augen der von außen erzeugten Körper- und Kopfdrehung entgegendrehen, um ein auf der Netzhaut aufrecht stehendes Abbild der Umwelt beizubehalten. Durch die damit verbundene Änderung in der Dehnungssituation der Augenmuskeln lösen die Dehnungsrezeptoren der Augenmuskeln eine entsprechende Mitbewegung des Kopfes durch Kontraktion der Halsmuskeln aus. Dies wiederum führt zu einer Reizung von Dehnungsrezeptoren in den Halsmuskeln und Gelenkrezeptoren der Halswirbelsäule, was zur Auslösung nachfolgender Reflexbögen (Abb. 5.12) und somit zur Mitbewegung der Rumpfwirbelsäule und u.U. zu unterstützenden Aktionen der Gliedmaßen beiträgt (Abb. 5.17). Auch von den Tastrezeptoren der Gliedmaßen, die eine Veränderung der Druckverteilung an den Kontaktstellen mit der Unterlage registrieren, werden Reflexbögen zu den Gliedmaßenmuskeln zur Durchführung entsprechender Ausgleichsbewegungen geschaltet.
Beim Menschen kommt den Körperstellreflexen aufgrund der aufrechten Haltung eine noch größere Bedeutung zur Regulation des aufrechten Standes im Schwerefeld der Erde zu als beim Vierfüßer, da das Verhältnis der Standfläche zur Schwerpunkthöhe deutlich kleiner und somit das Gleichgewicht entsprechend labiler ist. Allerdings zeigen die Reflexe zur Regelung zumindest von Augen-, Kopf- und Rumpfhaltung annähernd die gleiche Wirkung wie beim Vierfüßer: Bei einem Seitneigen des gesamten Körpers kommt es durch einen Einfluss der Gleichgewichtssinnesorgane auf die Augenmuskulatur zuerst zu einem Gegenrotieren der Augen um die Sagittalachse, danach durch die Verbindung der Augenmuskelrezeptoren mit den Halsmuskeln zu einem Aufrichten des Kopfes. Parallel dazu werden – ausgehend von den Gleichgewichtssinnesorganen - durch weitere kettenartigen Reflexbögen und entsprechende Spannungsänderungen in der Beinmuskulatur, die u.U. einen oder mehrere Seitschritte ausführen können, Aktionen
Bei der stützmotorischen Regulation der aufrechten Körperstellung werden Stördrehungen des Körpers um die Längs- oder Querachse sukzessive mit Gegendrehungen erst der Augen, dann des Kopfes, Halses und des Rumpfes mit Unterstützung der Gliedmaßen beantwortet.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 27
zur Aufrechterhaltung der senkrechten Körperposition ausgelöst. Zusätzlich beteiligen sich die Arme an der Durchführung von Ausgleichsbewegungen. Auch wenn sich beim Menschen der Gesichtssinn, der Gleichgewichtssinn und der Tastsinn (Durckverteilung an der Fußsohle) gemeinsam wechselseitig unterstützend an der Gleichgewichtsregulation beteiligen, muss jedoch festgestellt werden:
Letzteres kann jeder leicht an einem Selbstversuch feststellen, indem er Balancieraufgaben mit geschlossenen Augen ausführt.
In der Normallage des Kopfes zeigen die Maculazeptoren eine mittlere Ruheentladung (Abb. 5.18a). Bei Seitneigung des Kopfes steigt die Entladungsrate der Maculasinneszellen der einen Körperseite, auf der Gegenseite nimmt sie ab. Das bedingt auf der „Innenseite“ eine Erhöhung der Aktivität der Streckmuskeln des Beines und eine Abnahme der Aktivität der Beugemuskeln des Beines, auf der Gegenseite eine Aktivierung der Beugemuskeln und eine Hemmung der Strecker, so dass der Körper in Normalstellung gelangt (Abb. 5.18b). Parallel dazu bedingt die unterschiedliche Entladungsrate ein Gegenrollen der Augen (Abb. 5.18c), so dass ein aufrechtstehendes Bild gewährleistet wird, sowie eine Aktivierung der Halsmuskeln der „Außenseite“ und eine Hemmung der Halsmuskeln der Gegenseite, so dass der Kopf (Abb. 5.18d) und anschließend der Gesamtkörper die Normalstellung zurückerlangt.
Wird der Kopf in horizontaler Richtung linear (physikalisch richtig: translatorisch) beschleunigt (Abb. 5.16 a, F), zeigen die Statolithen die Tendenz, unter der Wirkung der Trägheit am Ort zu verharren. Es kommt zu einer Verschiebung zwischen Sinnesfeld und Deckmembran, was ein Abbiegen der Sinneshaare und
Bei der Kontrolle der aufrechten Körperstellung des Menschen spielt der optische Sinn eine herausragende Rolle. Bei Beeinträchtigung einer optischen Orientierung ist die Balancierleistung deutlich verschlechtert.
Abb. 5.18: Wirkung der Maculaorgane
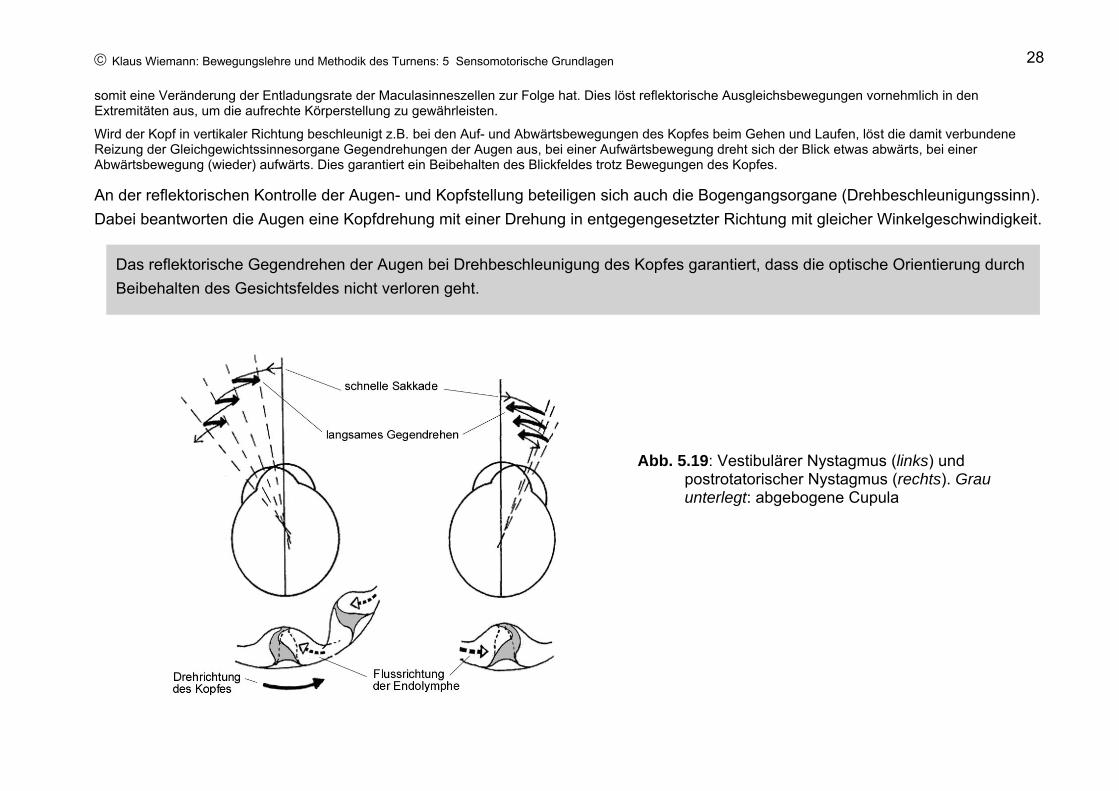
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 28
somit eine Veränderung der Entladungsrate der Maculasinneszellen zur Folge hat. Dies löst reflektorische Ausgleichsbewegungen vornehmlich in den Extremitäten aus, um die aufrechte Körperstellung zu gewährleisten.
Wird der Kopf in vertikaler Richtung beschleunigt z.B. bei den Auf- und Abwärtsbewegungen des Kopfes beim Gehen und Laufen, löst die damit verbundene Reizung der Gleichgewichtssinnesorgane Gegendrehungen der Augen aus, bei einer Aufwärtsbewegung dreht sich der Blick etwas abwärts, bei einer Abwärtsbewegung (wieder) aufwärts. Dies garantiert ein Beibehalten des Blickfeldes trotz Bewegungen des Kopfes.
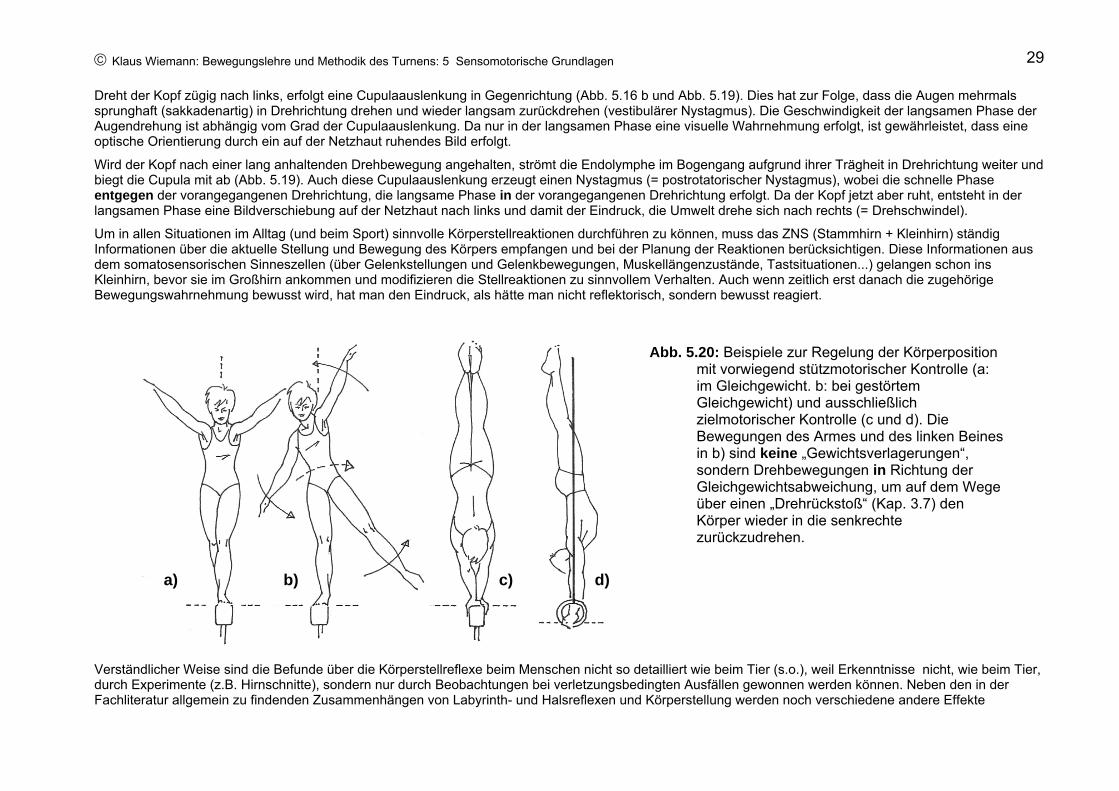
An der reflektorischen Kontrolle der Augen- und Kopfstellung beteiligen sich auch die Bogengangsorgane (Drehbeschleunigungssinn). Dabei beantworten die Augen eine Kopfdrehung mit einer Drehung in entgegengesetzter Richtung mit gleicher Winkelgeschwindigkeit.
Das reflektorische Gegendrehen der Augen bei Drehbeschleunigung des Kopfes garantiert, dass die optische Orientierung durch Beibehalten des Gesichtsfeldes nicht verloren geht.
Abb. 5.19: Vestibulärer Nystagmus (links) und postrotatorischer Nystagmus (rechts). Grau unterlegt: abgebogene Cupula
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 29
Dreht der Kopf zügig nach links, erfolgt eine Cupulaauslenkung in Gegenrichtung (Abb. 5.16 b und Abb. 5.19). Dies hat zur Folge, dass die Augen mehrmals sprunghaft (sakkadenartig) in Drehrichtung drehen und wieder langsam zurückdrehen (vestibulärer Nystagmus). Die Geschwindigkeit der langsamen Phase der Augendrehung ist abhängig vom Grad der Cupulaauslenkung. Da nur in der langsamen Phase eine visuelle Wahrnehmung erfolgt, ist gewährleistet, dass eine optische Orientierung durch ein auf der Netzhaut ruhendes Bild erfolgt.
Wird der Kopf nach einer lang anhaltenden Drehbewegung angehalten, strömt die Endolymphe im Bogengang aufgrund ihrer Trägheit in Drehrichtung weiter und biegt die Cupula mit ab (Abb. 5.19). Auch diese Cupulaauslenkung erzeugt einen Nystagmus (= postrotatorischer Nystagmus), wobei die schnelle Phase entgegen der vorangegangenen Drehrichtung, die langsame Phase in der vorangegangenen Drehrichtung erfolgt. Da der Kopf jetzt aber ruht, entsteht in der langsamen Phase eine Bildverschiebung auf der Netzhaut nach links und damit der Eindruck, die Umwelt drehe sich nach rechts (= Drehschwindel).
Um in allen Situationen im Alltag (und beim Sport) sinnvolle Körperstellreaktionen durchführen zu können, muss das ZNS (Stammhirn + Kleinhirn) ständig Informationen über die aktuelle Stellung und Bewegung des Körpers empfangen und bei der Planung der Reaktionen berücksichtigen. Diese Informationen aus dem somatosensorischen Sinneszellen (über Gelenkstellungen und Gelenkbewegungen, Muskellängenzustände, Tastsituationen...) gelangen schon ins Kleinhirn, bevor sie im Großhirn ankommen und modifizieren die Stellreaktionen zu sinnvollem Verhalten. Auch wenn zeitlich erst danach die zugehörige Bewegungswahrnehmung bewusst wird, hat man den Eindruck, als hätte man nicht reflektorisch, sondern bewusst reagiert.
Verständlicher Weise sind die Befunde über die Körperstellreflexe beim Menschen nicht so detailliert wie beim Tier (s.o.), weil Erkenntnisse nicht, wie beim Tier, durch Experimente (z.B. Hirnschnitte), sondern nur durch Beobachtungen bei verletzungsbedingten Ausfällen gewonnen werden können. Neben den in der Fachliteratur allgemein zu findenden Zusammenhängen von Labyrinth- und Halsreflexen und Körperstellung werden noch verschiedene andere Effekte
a) b) c) d)
Abb. 5.20: Beispiele zur Regelung der Körperposition mit vorwiegend stützmotorischer Kontrolle (a: im Gleichgewicht. b: bei gestörtem Gleichgewicht) und ausschließlich zielmotorischer Kontrolle (c und d). Die Bewegungen des Armes und des linken Beines in b) sind keine „Gewichtsverlagerungen“, sondern Drehbewegungen in Richtung der Gleichgewichtsabweichung, um auf dem Wege über einen „Drehrückstoß“ (Kap. 3.7) den Körper wieder in die senkrechte zurückzudrehen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 30
beschrieben. So soll beispielsweise die Halsstellung eine Bestimmungsgröße für die Intensität von Dehnungsreflexen, für die Kraft von Willkürkontraktionen der Handbeuger und für die Schnelligkeit von Armstreck- und Armbeugebewegungen sein. Ebenso wird eine führende reflektorische Beeinflussung der Hals-, Rumpf- und Gliedmaßenmuskeln durch die Augenstellung vermutet, da die visuelle Kontrolle bei Balancieraufgaben leistungsfähiger ist als die vestibuläre oder taktile Kontrolle. Die Führung des visuellen Systems bei der Regelung der Körperhaltung ist sogar derart ausschließlich, dass bei experimenteller visueller Täuschung von Versuchspersonen das vestibuläre und das somatosensorische System nicht korrigierend eingreifen und die Versuchspersonen das Gleichgewicht verlieren können. In diesem Falle kann erst nach einer Übungsphase die Balancierleistung wieder etwas verbessert werden, indem sich die Personen vorwiegend durch Tastinformationen aus den Fußsohlen über ihre Körperstellung orientieren. Allerdings sind in diesem Fall die Balancierleistungen geringer. (GEORGE 1972; MÜLLER 1974; HAYES 1975; LEE/LISHMAN 1975; WILLIAMS/DUBOIS 1980).
Im Gerätturnen spielt die Regulierung der Körperstellung in Bezug zum Schwerefeld der Erde insbesondere beim aufrechten Balancieren auf dem Schwebebalken sowie beim Einnehmen eines sicheren Standes nach einem Abgang eine besondere Rolle (Abb. 5.20, a und b). Sobald sich der Turner jedoch in einen Handstand begibt, also in eine Position, die in der stammesgeschichtlichen Entwicklung als Teil der Alltagsmotorik „nicht vorgesehen“ ist, kann er sich beim Ausbalancieren nicht mehr auf die Körperstellreflexe verlassen (Abb. 5.20, c und d).
Das bedeutet: der Turner muss ein sicheres Balancieren im Handstand durch ausgiebiges Üben erwerben, indem er sich auf die Druckempfindungen zwischen Hand und Stützfläche konzentriert und auf Gewichtsverlagerungen entsprechend reagiert. Mit der Stützmotorik sind auch Schutzreflexe verbunden, die in der Regel den Zweck verfolgen, in Gefahrensituationen, die durch eine massive Störung des Gleichgewichtes entstehen oder intuitiv als solche empfunden werden, den Körper, insbesondere die empfindlichere Körpervorderseite, vor Verletzungen zu bewahren. Dazu werden in der Regel der gesamte Körper, aber auch Arme und Beine gebeugt, der Kopf auf die Brust genommen und die Arme und Beine vor den Rumpf gebracht. Zusätzlich erhalten die Muskeln einen hohen Tonus, um vor allem die Gliedmaßen in gebeugter Stellung zu versteifen. Vor allem dann, wenn turnerische Elemente neu erlernt werden und ihr Ablauf noch nicht gefestigt ist, können solche Schutzreaktionen – oft aus belanglosen Anlässen - den Lernprozess massiv stören. Es gilt deshalb, derartige Situationen in der Unterrichts- und Trainingspraxis konsequent zu vermeiden und beim Übenden möglichst zu keiner Zeit ein Gefühl der Unsicherheit aufkommen zu lassen.
Unsicherheiten können auch dann auftreten, wenn der Turnende ungewohnte Drehbeschleunigungen erfährt. Ausgangspunkt sind dabei in der Regel die Bogengangsorgane (Drehbeschleunigungssinn), die als Folge reflektorische Gegendrehungen der Augen bewirken. Zeigen letztere hohe Drehgeschwindigkeiten verbunden mit Blicksprüngen (Sakkaden, optokinetischer Nystagmus), geht die optische Orientierung verloren. Der Turner „weiß nicht mehr, wo er ist“. Durch langes intensives Üben kann (und muss) sich der Turner an diese Drehbeschleunigungen gewöhnen, bis es ihm gelingt, auch bei hohen Drehbeschleunigungen
Die Körperstellreflexe sind zum Ausbalancieren eines Handstandes völlig ungeeignet. Aufgrund der ungewöhnlichen Position des Kopfes kommen sie nicht zur Auslösung bzw. müssen willkürlich unterdrückt werden. Balancieren im Handstand muss also erlernt werden!

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 31
die Bewegungen seines Körpers bewusst zu überwachen. Wichtig ist dabei zu beachten, dass diese Gewöhnungen nach längeren Perioden ohne Übung wieder verloren gehen können. Dieser Verlust kann eine Quelle von Unfällen sein. Wenn beispielsweise ein ehemals versierter Turner nach Jahren ohne Turnpraxis glaubt, eine früher automatisch beherrschte Turnübung spontan vorführen zu können, kann die Übung auf Grund der verschwundenen Anpassung an die Drehbeschleunigung völlig misslingen – auch bei wiederholten Versuchen und auch, wenn keine sonstigen körperlichen Defizite (z.B. im Kraftbereich) zu erkennen sind.
Die Auswirkungen der Überreizung der Bogengangsorgane können bei völlig Bewegungsunerfahrenen noch massiver sein. Dies lässt sich besonders bei Schülern beobachten, z.B. wenn von ihnen verlangt wird, mehrere Rollen vorwärts hintereinander zu turnen. Hier kann neben Drehschwindel sogar körperliches Unwohlsein, u,U. sogar mit Brechreiz verbunden, auftreten. Dies ist dadurch erklärbar, dass das Vestibularorgan mit den vegetativen Zentren des Zwischenhirns und somit mit dem vegetativen Nervensystem in Wechselbeziehung steht.
Die Reflexe der Rückenmarksmotorik funktionieren nach dem „Alles-oder-nichts-Prinzip“, d.h., wenn am Rezeptor eine bestimmte Reizschwelle überschritten ist, wird die Reaktion in der vorgegebenen Stärke realisiert. Dies gilt für die Reflexe der Stützmotorik nicht. Diese stehen unter der Kontrolle des Großhirns, können somit durch willkürliche Einflüsse unterdrückt, abgeschwächt, ausgenützt oder umgeformt werden.
5.3.2 Orientierung im Raum
Will ein vierfüßig lebendes Tier zum Zweck der optischen Orientierung z.B. aus einer gebeugten Ausgangsstellung einen hochgelegenen Punkt visuell fixieren, strecken sich im Zuge der Orientierungsreaktion nach dem Aufwärtsdrehen der Augen und des Kopfes zuerst die Hals- , dann erst die Brust- und schließlich die Lendenwirbelsäule und die vorderen Extremitäten, während die hinteren Extremitäten gebeugt werden (Abb. 5.21). Sinngemäß beugt sich der Rumpf bei einer Orientierung nach unten sukzessive abwärts. Neben diesen „symmetrischen“ Reaktionen gibt es sinngemäße „asymmetrische“ Reaktionen bei einer Orientierung zur Seite oder bei Drehungen des Kopfes. Ausgangspunkt dieser Reflexketten ist in der Regel eine Orientierungsreaktion: Bewegt sich das Abbild eines optisch zu fixierenden Objektes aus der Sehgrube, dem Ort des schärfsten Sehens, heraus auf periphere Bereiche der Netzhaut, wird ein Orientierungsreflex ausgelöst, der die Augen dem Sehobjekt folgen lässt, um es wieder in der Sehgrube abbilden zu können. Durch damit verbundenen Dehnungsänderung der Augenmuskeln werden nachfolgend die oben beschriebenen Reflexketten in Gang gesetzt. Gleiches geschieht natürlich auch, wenn die Augen zwecks optischer Orientierung willkürlich in eine beabsichtigte
Wird die natürliche Körperposition massiv gestört, können die Körperstellreflexe allein die Regulation der Körperstellung nicht bewerkstelligen. Orientierungs- und Tonusreflexe (s.u.) müssen helfend eingreifen.
Bei Verlust der Orientierung, z.B. im Falle ungewohnter Drehbeschleunigungen, dominieren Schutzreflexe alle Körperstell- und Orientierungsreaktionen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 32
Richtung gedreht werden. Das Ziel ist ein optisches Orientieren mit einem möglichst niedrigen Anstrengungsgrad in den verschiedenen Körpersegmenten. Die mit einer willkürlichen Orientierung verbundene Kopfbewegung könnte nun durch das Gleichgewichtsorgan Körperstellreflexe auslösen, um den Körper wieder in die Ausgangslage zurückzuführen. Damit wäre der Organismus praktisch in der Ausgangsposition gefangen und könnte diese nicht verlassen. Um dies zu vermeiden, sind die Orientierungsreaktionen den Labyrinthreflexen übergeordnet, was bedeutet, dass die Labyrinthreflexe während der Orientierungsaktionen und Orientierungsreaktionen neuronal gehemmt sind.
Für all diese Reflexerscheinungen zur Körperstellung und -haltung gelten folgende bedeutende Bedingungen:
1. Entsprechend ihrer Aufgabe laufen die Tonusreflexe von vorn nach hinten (genauer: in cranio-caudaler Richtung = vom Kopf bis zur Schwanzwirbelsäule) über den Körper, gleichgültig, ob die Reflexbögen direkt in der Augenmuskulatur oder etwa erst durch
Um eine möglichst mühelose optische Orientierung zu garantieren, lösen Augendrehungen (Blickwendungen) Reflexketten aus, die den Körper nach und nach dem sich orientierenden Kopf folgen lassen (Tonusreflexe). Dabei ist die Auslösung von Körperstellreflexen weitgehend unterbunden bzw. an die Zwecke der Blickbewegungen angepasst.
Abb. 5.21: Orientierungsaktion mit Halsreflexen beim Vierfüßer

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 33
willkürliches Bewegen des Kopfes in der Halsmuskulatur ausgelöst werden. In keinem Fall aber laufen die Reflexketten in umgekehrter Richtung. So hat z.B. eine willkürliche Spannungsänderung der Rückenmuskulatur im Brust- und Lendenbereich keine reflektorische Wirkung auf die Halsmuskulatur.
2. Zur Auslösung der einzelnen Reflexbögen ist eine bestimmte Reizstärke nötig. Eine Augendrehung hat nur dann eine Mitbewegung des Kopfes zur Folge, wenn eine bestimmte Reizschwelle für die Rezeptoren der Augenmuskulatur überschritten wird. Gleiches gilt auch für die übrigen Reflexbögen der Tonus- und Körperstellreflexe. Das bedeutet: je stärker der auslösende Reiz ist, um so weiter läuft die Auslösung der Reflexe nach hinten (in cranio-caudaler Richtung) über den Körper.
3. Die einzelnen Reflexbögen der Körperstellreflexe sind nicht „unbedingter“ Natur, wie etwa die Dehnungsreflexe, die nach dem „Alles-oder-nichts-Prinzip“ funktionieren, sondern können (und müssen) durch willkürliche Impulse unterdrückt oder abgeändert werden.
5.3.3 Orientierungs- und Tonusreflexe beim Menschen
Beim Menschen unterliegen die in Tierversuchen eingehend erforschten Körperstellreflexe den gleichen Wirkungsprinzipien wie beim Vierfüßer. Naturgemäß erfahren sie jedoch eine - wenn auch geringe - Abänderung durch die Tatsache, dass der Mensch den Körper auf den ursprünglich hinteren Extremitäten aufgerichtet hält, aber:
Auch beim Menschen - haben die Orientierungs- und Tonusreflexe die Aufgabe, den Körper dem sich hauptsächlich durch den Gesichtssinn orientierenden Kopf folgen zu lassen, - wirken die Reflexe nur in cranio-caudaler Richtung - also von oben nach unten bzw. von der Halswirbelsäule bis zur Lendenwirbelsäule, - ist die Stärke des auslösenden Reizes für den Grad der Reflexausbreitung caudalwärts (steißwärts) von Bedeutung, - können (und müssen - je nach Bewegungsziel) die Orientierungs- und Tonusreflexe durch bewusste Impulse beeinflusst werden.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 34
Im folgenden soll nun die Bedeutung der Körperstellreflexe des Menschen für Bewegungen, die außerhalb des naturgegebenen Bewegungsvolumens (Kap. 4.1) liegen, nämlich für Gerätübungen, abgegrenzt werden. Bei allen Turnübungen handelt es sich um Bewegungen, bei denen durch vorherbestimmte Bewegungsakte ein festgelegtes Bewegungsziel erreicht werden soll. Im Vordergrund steht in jedem Fall die geforderte Körperhaltung und Bewegungsform ohne Rücksicht auf das natürliche Orientierungsbedürfnis in den verschiedenen Körperlagen, die während der Übung eingenommen werden müssen. So ist die Orientierung bei den meisten Gerätübungen auf den „Gewebssinn“ (Somatosensibilität) beschränkt, während der optische Sinn zum großen Teil den bewegungsführenden Einfluss einbüßt. Deutlich wird diese Tatsache bei ungeübten Turnern, die, sobald ihnen durch eine ungewohnte Körperstellung die Möglichkeit zur optisch regulierten Beibehaltung des Gesichtsfeldes genommen wird, die „Orientierung verlieren“.
Um zu zeigen, welchen Bedingungen die Orientierungs- und Tonusreflexe bei den verschiedenen turnerischen Bewegungen unterliegen, werden die turnerischen Bewegungen in mehrere Gruppen zusammengefasst.
In einer ersten Gruppe finden wir Gerätübungen, bei denen die gesamte Kette der Orientierungs- und Tonusreflexe in ihrer natürlichen Form von der Augenmuskulatur caudalwärts zur Wirkung kommt. In diese Gruppe gehört z.B.: Handstandüberschlag rückwärts (Abb. 5.22, links) und Salto rückwärts gestreckt am Boden, alle Vorwärtsrollen (Abb. 5.22, rechts) und die Übungen, die eine Vorwärtsdrehung um die quere Schwerpunktachse zeigen. Diese Übungen beginnen mit einer orientierenden Augendrehung in Bewegungsrichtung, was nacheinander ein Folgen des Kopfes und der Hals- und Rumpfwirbelsäule nach sich zieht (Abb. 5.22), beim Hangstandüberschlag rückwärts eine zunehmende Überstreckung, bei der Rolle vorwärts eine entsprechende Beugung. Außerdem
Abb. 5.22: Handstandüberschlag rückw. und Rolle vorw. als Beispiele für Übungen, in deren Verlauf die gesamte Kette der Orientierungs- und Tonusreflexe zur Auslösung gelangen. OP: optischer Orientierungspunkt

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 35
werden teilweise die Arme in der für die Tonusreflexe charakteristischen Weise beeinflusst. Für diese Gruppe von Übungen muss die unter Punkt 2. in Kap. 5.3.2 angeführte Bedingung methodisch angewendet werden: Je stärker der auslösende Reiz, desto intensiver die Reflexwirkung in caudaler Richtung. Muss nämlich eine Übung unter starker Extremstellung der Wirbelsäule im Sinne einer Dorsalflexion (Überstreckung bis zur Hohlkreuzhaltung) oder Ventralflexion (Einrollen des Rumpfes und Abwinkeln im Hüftgelenk) ausgeführt werden, sollte diese Stellung durch einleitendes, möglichst intensives Bewegen des Kopfes dorsal- oder ventralwärts ausgelöst werden – am zweckmäßigsten vorbereitet durch eine entsprechende optische Orientierung in die gewünschte Richtung u.U. durch Verwendung eines Orientierungspunkts.
In einer zweiten Gruppe von Turnelementen kommen die Tonusreflexe zwar auch zur Anwendung, dabei muss jedoch das natürliche Bedürfnis zur optischen Orientierung unterdrückt werden, so bei allen mit gestrecktem Körper geturnten Vorwärtsüberschlägen. Beim Handstandüberschlag vorwärts am Boden versucht der Anfänger, durch ein Einrollen des Kopfes in Bewegungsrichtung möglichst schnell ein senkrecht stehendes Bild zu gewinnen, um sich über die Lage des Körpers im Raum orientieren zu können (Abb. 5.23, oben). Diese reflektorische Bewegung muss sowohl der Wirkung der Augenmuskelreflexe, als auch der Labyrinthreflexe zugeschrieben werden. Durch das Einrollen des Kopfes wird aufgrund der Tonusreflexe die für den Handstandüberschlag verlangte Bogenspannung (Dorsalflexion) des Rumpfes aufgehoben.
Dieser Orientierungsreflex wird am zweckmäßigsten unterbunden, wenn dem Übenden ein neuer Orientierungspunkt gegeben wird, der in der Stützphase und der anschließenden Flugphase so lange wie möglich anzuschauen ist (Abb. 5.23, unten). Auf diese Weise wird das Beibehalten der korrekte Körperhaltung durch die Auslösung der gesamten Kette der Orientierungs- und Tonusreflexe unterstützt, beginnend beim natürlichen Anfangsglied der Kette, der orientierenden Augendrehung (gleichsam auch das „Führungsglied“), bis hin zur Streckung des Hüftgelenkes. Diese Methode scheint erfolgreicher als ein Einstieg an irgend einer anderen Stelle der Kette, beispielsweise der Halswirbelsäule, etwa durch die Forderung, den “Kopf in den Nacken zu nehmen!“. Ähnliches gilt für die Kopf- und Nackenüberschläge (s. jedoch unten, vierte Gruppe) und im entgegengesetzten Sinne für die Rolle rückwärts am Boden und für den gehockten Salto rückwärts.
Gerade bei der letztgenannten Übung nimmt auch manch fortgeschrittener Turner noch den Kopf auf Grund eines Orientierungsbedürfnisses in den Nacken. Methodiker fordern diese Bewegung sogar, obwohl sie aufgrund der ausgelösten Tonusreflexe eine schlechte Hockphase zur Folge hat; denn es besteht vielfach die irrige Auffassung, man könne durch ein

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 36
Rückführen des Kopfes nach dem Absprung die Rückwärtsdrehung einleiten oder vergrößern (s. Kap. 3.6 und Abb. 3.41). Bei exakter Ausführung des Salto rückwärts wird spätestens direkt nach Verlassen des Bodens der Kopf auf die Brust genommen, um die gesamte Ventralflexion des Körpers reflektorisch zu unterstützen.
Zur optischen Orientierung beim Salto rückwärts gehockt muss festgestellt werden, dass zwar ein Orientierungsbedürfnis besteht wie in allen ungewohnten Körperlagen, dass aber die Möglichkeit einer durch optische Orientierung ausgelösten Bewegungskorrektur nicht gegeben ist. Die Zeit, die dem Turner zur Verfügung steht, reicht bei weitem nicht aus, die Lage im Raum optisch zu analysieren, eine Korrekturbewegung zu planen und zu realisieren (Kap. 5.4; Abb. 5.41).
In einer dritten Gruppe von turnerischen Fertigkeiten kann die Haltung des Kopfes auch die Stellung der Arme bzw. den Tonus der Armmuskeln beeinflussen. Beim Bauchwellaufschwung am Reck werden durch eine Ventralflexion der Halswirbelsäule nicht nur die ventrale Bauchmuskulatur, sondern auch die Beuger und Retraktoren der Arme durch die Tonusreflexe zu einer Klimmbewegung angesteuert (Abb. 5.24 b).
Abb. 5.23: Handstandüberschlag vorwärts. oben: turntechnisch schlechte Ausführung durch Auslösung von Orientierungs- und Tonusreflexen. unten: korrekte Ausführung durch Verwendung eines optischen Orientierungspunktes (OP)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 37
Sobald jedoch - etwa durch eine falsche Bewegungsvorstellung oder eine unzutreffende methodische Maßnahme - der Kopf in den Nacken genommen wird, entsteht - vor allem beim Anfänger - ein reflektorischer Einfluss auf die Armmuskeln, der zu einer Streckbewegung der Arme führen kann und die Übung misslingen lässt (Abb. 5.24 c).
Dieser Einfluss der Kopfhaltung auf die Spannung der Armmuskulatur scheint auch bei der Schwungstemme rückwärts am Barren ausgenutzt zu werden. Bei den Schwungstemmen rückwärts soll die Abschwungphase mit leicht gewinkeltem Hüftgelenk geturnt werden, während zu Beginn der Aufschwungphase die Beine nach hinten zur Hohlkreuzhaltung des Rumpfes geschwungen werden müssen (Abb. 5.25). Hier lässt sich beobachten, dass manche Turner nicht den Kopf in den Nacken nehmen, wie es die
a)
b)
c)
Abb. 5.24: Turntechnisch korrekter Salto rückwärts (a), Bauchwellaufschwung mit reflektorischer Unterstützung der Klimmbewegung der Arme (b) und mit fehlerhaftem Strecken der Arme (c)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 38
Überstreckung der Wirbelsäule eigentlich voraussetzt, sondern ihn gegen die Brust bewegen (ventral beugen). Möglicherweise wird auf diese Weise die Beuge- bzw. Zugbewegung der Arme reflektorisch unterstützt.
Außer den bisher besprochenen Bewegungen gibt es noch eine vierte Gruppe bewegungsverwandter Übungen, bei denen nach einer mehr oder weniger starken Hüftbeugung der Körper zügig oder schlagartig gestreckt wird: die Kippen und Felgen. Die Streckung beginnt bei diesen Übungen aus sehr ähnlichen Ausgangspositionen, nämlich aus einer Kipplage, einem Sturzhang oder Sturzhangschwung, bei denen der Rücken zur Erde hinweist, während sich die Beine oberhalb des Rumpfes befinden (Abb. 5.26). Von hier aus kann die Streckung einerseits nach vorn oben, andererseits mehr oder weniger senkrecht nach oben oder gar nach hinten oben führen. All diese Kipp- und Felgbewegungen beginnen, gleichgültig in welcher Richtung sie erfolgen, mit einer Streckung im Hüftgelenk. Erst im Laufe der weiteren Streckbewegung löst sich – bei einigen Übungen - auch die durch die gesamte Körperbeugung bedingte Ventralflexion der Wirbelsäule auf. Je nach der Richtung der Übung wird die Streckbewegung noch bei einer schwachen Beugestellung des Körpers gebremst (Abb. 5.26 a, b), oder sie führt zur völligen Streckung des Körpers, wobei Rumpf und Beine annähernd mit der Längsachse zusammenfallen (Abb. 5.26 c). In Ausnahmefällen endet die Bewegung sogar in einer leicht überstreckten Körperhaltung (Abb. 5.26 d). Da aber bei all diesen Übungen die Streckbewegung im Hüftgelenk beginnen muss und sich in cranialer Richtung fortsetzt, kommen die Tonusreflexe und somit auch die Halsreflexe, die nur in caudaler Richtung arbeiten, bei der Auslösung und Durchführung dieses Bewegungsaktes nicht zur Wirkung. So wird im allgemeinen bei der
Abb. 5.25: Schwungstemme rückwärts am Barren (nach Filmanalyse)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 39
Ausführung der Kipp- und Felgbewegungen der Kopf sowohl zu Beginn als auch im Laufe der Streckung entweder auf der Brust oder „locker“ in Verlängerung der Wirbelsäule gehalten (Abb. 5.26).
Eine Ausnahme bilden der Nackenüberschlag und die Rolle rückwärts am Boden in den Handstand (Abb. 5.27). Hier kann ein rechtzeitiges Drücken des Kopfes in den Nacken zwei Effekte erreichen: 1. Durch ein kräftiges Drücken des Hinterkopfes gegen die Unterlage kann der Körper aufgrund des sehr günstigen Kraftmomentes unterstützend aufwärtsbeschleunigt werden – ein Effekt, auf den man insbesondere bei Schülern und Turnanfängern nicht verzichten sollte. 2. Da durch diese Aktion die Halswirbelsäule gestreckt wird, mag sie auf dem Wege über Tonusreflexe die anschließende Körperstreckung unterstützen können, obwohl diese schon durch die Willküraktion „Hüftstreckbewegung“ eingeleitet wurde.
a) b)
c)
d)
Abb. 5.26: Turnerische Fertigkeiten, in deren Verlauf die Körperstreckbewegung im Hüftgelenk beginnt. a) Oberarmkippe am Barren. b) Unterschwung („Schwabenkippe“) am Barren. c) Felgaufschwung am Reck. d) „Schleudern“ an den Ringen

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 40
Selbst bei manchen Übungen, die eine Streckbewegung bis zum völlig gestreckten Körper zeigen, beobachtet man, dass der Kopf in Ventralhaltung verharrt (Abb. 5.28 rechts). Wird bei einzelnen Übungen der Kopf doch in den Nacken genommen, sollte diese Bewegung von geringer Intensität sein, damit keine reflektorisch bedingten Fehlbewegungen entstehen. Außerdem lässt die Tatsache, dass die Dorsalflexion des Halses nicht zu Beginn, sondern erst im Laufe der Streckbewegung einsetzt, erkennen, dass damit die Tonusreflexe nicht betätigt werden.
a)
b)
Abb. 5.27: Nackenüberschlag (a) und Rolle rückw. in den Handstand (b)
Abb. 5.28: Felge am Barren (nach Filmanalyse)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 41
Der Sinn der Streckung der Halswirbelsäule (= „Hochnehmen“ des Kopfes) ist entweder eine optische Orientierung über die Lage des Körpers im Raum (nur bei Anfängern notwendig), oder ein Richten des Blickes auf das Gerät zwecks optischer Kontrolle des Griffwechsel (zum großen Teil auch nur bei Anfängern notwendig), oder ein abschließender Handstand verlangt eine leichte Dorsalhaltung des Kopfes. In diesen Fällen sollte man sich im Hinblick auf methodische Anweisungen davor hüten, die Kopfbewegung für die Auslösung der Felg- und Kippbewegungen verantwortlich zu machen.
Auch bei einer noch zu besprechenden fünften Gruppe von Übungen, den freien Überschlägen rückwärts an Reck, Barren und Ringen, beginnt die Streckung im Hüftgelenk. Allerdings können hier zwei Ausführungsarten beobachtet werden. Bei der ersten werden nach einem Anwinkeln des Hüftgelenks zu Beginn der Aufschwungphase die Hüfte und die Rumpfwirbelsäule heftig gestreckt - ohne Ausnutzung der Tonusreflexe (Abb. 5.29a). Der Kopf wird verhältnismäßig lange ventralwärts gehalten und erst in der letzten Phase der Übung, nach Vollendung der Körperstreckung, zwecks Orientierung schnell dorsalwärts bewegt.
Bei der zweiten Ausführungsart wird der Kopf schon zu Beginn der Aufschwungphase in den Nacken genommen. Offensichtlich sollen hier (wahrscheinlich eher willentlich geplant als intuitiv) die Tonusreflexe zur Überstreckung des Körpers ausgelöst werden, obwohl im
a)
b)
Abb. 5.29: Überschlag rückwärts am Barren mit spätem (a) und frühem (b) Einsatz der Dorsalflexion der Halswirbelsäule (nach Filmanalyse)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 42
Hüftgelenk die Beugebewegung noch nicht abgeschlossen ist (Abb. 5.29b). Inwieweit dadurch die Effektivität der Hüftbeugebewegung, die eigentlich zur Verstärkung des Schwunges dient, negativ beeinflusst wird, kann nur vermutet werden. Im Laufe des weiteren Aufschwunges mag sich aufgrund der Tonusreflexe die Streckbewegung der Halswirbelsäule über die Brust- und Lendenwirbelsäule auch auf das Hüftgelenk übertragen. Welche der beiden Ausführungsarten die von biologischer und mechanischer Sicht zweckmäßigere ist, muss offen bleiben.
Aus der biologischen Betrachtung der Körperstell-, Orientierungs- und Tonusreflexe resultieren Grundsätze, die bei der Planung methodischer Maßnahmen zum Erwerb turnerischer Fertigkeiten zu beachten sind:
Bei der Anwendung der Körperstell-, Orientierungs- und Tonusreflexe für methodische Überlegungen sind die „Halsreflexe“ bzw. „Halstonusreflexe“ nicht isoliert zu betrachten, da sie nur ein Glied in der Reflexkette zur Körperstellung- und Körperhaltung sind!
Die Orientierungs- und Tonusreflexe sind am sichersten am natürlichen Ausgangspunkt der Reflexkette, nämlich beim Augenstellreflex, und weniger zweckmäßig an einem Zwischenglied zu beeinflussen. Folglich: Optische Orientierungspunkte anbieten!
Das natürliche Orientierungsbedürfnis dort ausnutzen, wo es die geforderte Bewegungsstruktur unterstützt und dort durch Zusatzaufgaben umlenken, wo es zu fehlerhaften Bewegungsabläufen führen könnte!
Die Auslösung von Orientierungsreflexe und Schutzreflexen, die den Bewegungsablauf verfälschen könnten, durch solche methodische Maßnahmen vermeiden, die zu einem Gefühl der körperlichen Sicherheit führen!

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 43
5.4 Zielmotorik
Nahezu alle Bewegungen, die der Mensch im Alltag, bei der Arbeit und im Sport produziert, werden der Zielmotorik zugeordnet. Bei der Auswahl des Begriffes „Zielmotorik“ war offensichtlich maßgeblich, dass man den Bewegungen in der Regel ein Ziel zuordnen kann, das willentlich (absichtlich, geplant...) angesteuert werden soll. Die innere Koordination der Zielmotorik obliegt der höchsten Schicht des Zentralnervensystems, dem Großhirn (Kap. 5.1), wobei die Zielmotorik auf der Stützmotorik und der Rückenmarksmotorik aufbaut und ohne sie nicht gelingen kann. Allerdings ist das Merkmal des Beabsichtigten, Zielgerichteten oder Willkürlichen nicht ausschließliches Merkmal von Zielmotorik; denn je nach Situation können Bewegungsabläufe, die zur Zielmotorik zu rechnen sind, auch unwillkürlich, unbeabsichtigt, ungeplant, gleichsam reflexartig oder automatisch auftreten, wie folgende Beispiele verdeutlichen sollen:
Ein Schüler läuft an, um eine Hocke über den längs gestellten Kasten auszuführen. Natürlich sind die Laufbewegungen beabsichtigt und verfolgen ein äußeres Ziel. Sie werden somit vom Großhirn, in dessen Zuständigkeitsbereich alle Willkürbewegungen fallen, in Gang gesetzt. Sie sind der Zielmotorik zuzuordnen. Gleichwohl beinhalten sie stützmotorische Anteile, die sicherstellen, dass der Anlauf in der aufrechten Körperhaltung zu Ende geführt wird. Vollendet nun der Schüler im Moment des Absprunges die Hocke nicht, „verweigert“ er den Sprung, weicht statt dessen dem Kasten aus und läuft seitlich an ihm vorbei, gehört diese Ausweichbewegung nach wie vor zur Zielmotorik, obwohl der Schüler diese nicht absichtlich, sondern eher unwillkürlich vollzogen hat und obwohl sie mit einem hohen Anteil von Schutzreflexen versehen ist oder gar auf der Ebene der Schutzreflexe ausgelöst wurde.
Noch deutlicher wird dies an folgendem Beispiel: Ein Turner beabsichtigt, am Reck zwei Vorschwünge mit Kontergrätsche („Tkatchev“) unmittelbar hintereinander auszuführen. Nach der erfolgreich absolvierten ersten Kontergrätsche „verweigert“ er jedoch die zweite und turnt unwillkürlich einen Riesenumschwung rückwärts (Abb. 5.30). Hier kann niemand behaupten, es handele sich um einen Schutzreflex aus dem Bereich der Stützmotorik; denn ein „angeborener“ Schutzreflex „Riesenumschwung“ ist im natürlichen (stammesgeschichtlich begründbaren) Bewegungsvolumen des Menschen nicht vorgesehen (Kap. 4.2). Aus diesen Beispielen wird deutlich, dass es kaum möglich ist, Aktionen der Zielmotorik und Stützmotorik durch äußere Merkmale voneinander zu trennen. Selbst die Differenzierung in beabsichtigte bzw. unwillkürliche Auslösung hilft nicht. Dieses Problem spiegelt sich auch in der Wahl des Begriffes Zielmotorik. Es fällt schwer, einer unwillkürlichen „Ausweichbewegung“ ein Handlungsziel zu unterstellen. Auf ähnliche Schwierigkeiten stoßen Ersatzbegriffe, die vorgeschlagen wurden, wie z.B. Willkürmotorik oder Handlungsmotorik. Auch verbietet es sich, hier nach Erbmotorik und Erwerbmotorik zu differenzieren; denn auch die erbmotorischen Aktionen des Menschen wie Gehen, Laufen, Greifen usw. werden vom Großhirn koordiniert.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 44
In Abb. 5.31 a sind bei der Turnerin auf dem Schwebebalken das graziöse Anheben des rechten Beines und das gezierte Spreizen der Finger eindeutig Aktionen der Zielmotorik. Sie werden entweder absichtlich geplant und im Ablauf bewusst kontrolliert (Anheben des Beines) oder treten nach langjähriger Gewohnheit unwillkürlich auf. Ausgleichsbewegungen des Fußes und eventuelle Balancierbewegungen der Arme müssen jedoch der Stützmotorik zugerechnet werden. Im fiktiven Beispiel des über den Kasten springenden Schülers (Abb. 5.31 b und c), das schon in Kap. 3, Abb. 3.40, zur Erläuterung des „Drehrückstoßes“ diente, müssten ein unwillkürliches „Einrollen“ des Kopfes und ein ebenso unwillkürliches Vorwärtskreisen der Arme (Abb. 5.31 b) den Körperstellreflexen und den Schutzreflexen zugeordnet werden, sind somit keine zielmotorische Aktionen. Auch wenn der Schüler es nach der Landung so empfunden hätte, können diese Aktionen nicht absichtlich geplant sein. Diese Zuordnung erfolgt aber lediglich aufgrund
Alle turnerischen Fertigkeiten unterliegen den Bedingungen der Zielmotorik, enthalten jedoch stets mehr oder weniger ausgedehnte Komponenten der Stützmotorik (Kap. 5.3) und erfordern als unverzichtbare Basis die Rückenmarksmotorik (Kap. 5.2).
Abb. 5.30: Vorschwung mit Kontergrätsche
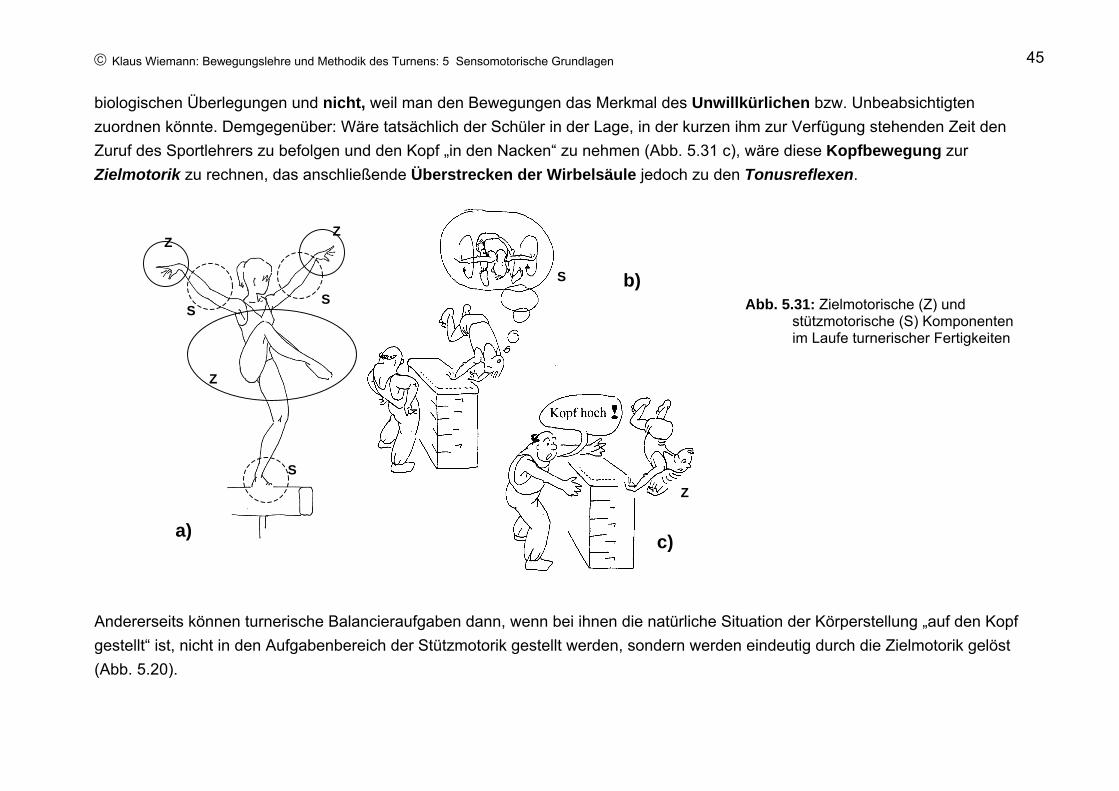
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 45
biologischen Überlegungen und nicht, weil man den Bewegungen das Merkmal des Unwillkürlichen bzw. Unbeabsichtigten zuordnen könnte. Demgegenüber: Wäre tatsächlich der Schüler in der Lage, in der kurzen ihm zur Verfügung stehenden Zeit den Zuruf des Sportlehrers zu befolgen und den Kopf „in den Nacken“ zu nehmen (Abb. 5.31 c), wäre diese Kopfbewegung zur Zielmotorik zu rechnen, das anschließende Überstrecken der Wirbelsäule jedoch zu den Tonusreflexen.
Andererseits können turnerische Balancieraufgaben dann, wenn bei ihnen die natürliche Situation der Körperstellung „auf den Kopf gestellt“ ist, nicht in den Aufgabenbereich der Stützmotorik gestellt werden, sondern werden eindeutig durch die Zielmotorik gelöst (Abb. 5.20).
a)
SS
S
S
ZZ
Z
Z
b)
c)
Abb. 5.31: Zielmotorische (Z) und stützmotorische (S) Komponenten im Laufe turnerischer Fertigkeiten

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 46
Abb. 5.32: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen
Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv). Grau unterlegt: bewusstseinspflichtige Funktionen -(verändert nach POPPER/ECCLES 1982). Grau unterlegt: bewusstseinspflichtige Funktionen.
zentrale Speicher Limbisches
System Motivation
Assoziations-rinde
strategische Planung
SMA Bewegungsentwurf
Primärmot. Rinde
Bewegungs-start
Neukleinhirn Programmierung Erwerbmotorik
Altkleinhirn
stato-vestib. Regulation
sensorischer Thalamus
motorischer Thalamus
Motoneurone Innervation
Muskeln Realisation
Hirnstamm
Großhirn
Rücken-mark
Basalganglien
Programmierung Erbmotorik
sensorische Rinde
Bewegungs- wahrnehmung
S I
Hinterstang- kerne
Sinnesorgane Rezeption
muskuläre und propriozeptive
Rezeptoren
mechanische Umwelt
Instruktionen
soziale Umwelt

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 47
Der gesamte Prozess soll anhand des Beispieles von Abb. 5.33, der Bewegungsfolge Anlauf – Radwende – Salto rückwärts, erläutert werden. Das vollständige Wirkungsgefüge zur Auslösung, Steuerung und Regelung eines zielmotorischen Ablaufes ist in Abb. 5.32 dargestellt.
Prinzipiell zeigen die gesamten Hirnprozess zur Erzeugung einer turnerischen Fertigkeit einen 6-stufigen Ablauf. Diese sechs Stufen, die man entsprechenden unterschiedlichen Regionen des Großhirns zuordnen muss (Abb. 5.32 und Abb. 5.41), sind: - Motivation und Handlungsbereitschaft (limbisches System), - strategische Planung (Assoziationsrinde des Großhirns), - Bildung des Bewegungsentwurf, (supplementäres motorisches Areal, SMA) - Programmierung (subkortikale Funktionsgeneratoren: Basalganglien, Neukleinhirn) - Start und Bewegungsausführung (primärmotorischen Rinde des Großhirns und Rückenmark) - Bewegungsrückmeldung und Bewegungskorrektur (Rückenmark aufsteigend bis Assoziationsrinde).
Bei der grafischen Darstellung solch komplexer Systeme wird naturgemäß auf reduktionistische Modelle zurückgegriffen Mit steigender Reduktion ist stets die Gefahr zunehmender Vereinfachung und Verfälschung verbunden. Dabei steht das Ausmaß der Reduktion in der Verantwortung des Verfassers und seiner Auffassung darüber, welches Maß an Verfälschung vertretbar ist. Im Endeffekt ließe sich das Modell zur Koordination der Zielmotorik auf einfache Regelkreise und Steuerketten reduzieren (s. Meinel/Schnabel 1989). Hier soll jedoch das in Abb. 5.32 dargestellte Flussdiagramm verwendet werden, weil es größere Entsprechungen zur Hirnfunktion bietet. Zur Verbesserung der Übersichtlichkeit wird dieses Diagramm schrittweise auf die einzelnen Funktionsabschnitte im Laufe der Produktion einer sportmotorischen Fertigkeit reduziert (Abb. 5.34 bis Abb. 5.49).
Abb. 5.33: Verbindung turnerischer Fertigkeiten am Boden

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 48
5.4.1 Motivation und Handlungsbereitschaft
Ein Sportler, der sich in der Turnhalle befindet und sich anschickt, eine sportliche Aktivität aufzunehmen, etwa die Ausführung der Bewegungskombination Anlauf – Radwende – Salto rückwärts, führt dieses Vorhaben auf eine von ihm geleistete Willensentscheidung zurück. Diese Phase zur Bildung der Handlungsbereitschaft bzw. Motivation stellt die Voraussetzung dafür dar, dass der Sportler überhaupt aktiv ist, dass er jetzt turnen und speziell die beabsichtigte Bewegungskombination ausführen will und nicht irgend etwas anderes zu tun beabsichtigt.
Die neurophysiologischen Vorgänge, denen diese Handlungsantriebe und Willensentscheidungen zugrunde liegen, sind nicht - wie früher angenommen wurde – allein an die Funktion der Großhirnrinde, speziell der primärmotorischen Rinde der vorderen Zentralwindung, gekoppelt, sondern werden in "tieferen" archaischen Hirnschichten verursacht, nämlich im Limbischen System (s. Abb. 5.1 und Abb. 5.41), dem stammesgeschichtlich ältesten Teil des Großhirns. Es umgreift eng
Limbisches System
Motivation
Assoziations-rinde
strategische Planung
Großhirn
Instruktionen
soziale Umwelt
Abb. 5.34: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Phase: Motivation und Handlungsbereitschaft. Grau unterlegt: bewusstseinspflichtige Hirnfunktionen

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 49
das Zwischenhirn und nimmt mit dessen Zentren, die die emotionale Situation des Menschen färben, Verbindungen auf. Das Limbische System wird in der englischsprachigen Fachliteratur auch als "emotional brain" bezeichnet, was darauf hindeuten soll, dass dieser Hirnabschnitt die Motivation zum Tun des Menschen bestimmt und für den emotionalen Hintergrund des Handelns verantwortlich ist.
Die Phase der Motivation (Abb. 5.34) kann von unterschiedlicher zeitlicher Ausdehnung sein und unterliegt vielfältigen äußeren Faktoren (Umgebungssituation, Licht, Geräusche, Gerüche), körperlichen Befindlichkeiten (Müdigkeit, Schmerz, Hunger, Durst, Müdigkeit) und emotionalen Zuständen (Freude, Ärger, Aversion, Zuneigung, Angst) und im Lehr- und Lernprozess den Instruktionen und Maßnahmen der Lehrperson (Abb. 5.34). Sie muss jedoch mindestens 0,5 s vor Bewegungsbeginn zu einem Ergebnis, einer Entscheidung, geführt haben (Abb. 5.41). Da die Motivation von großem Einfluss auf jeglichen Lern- und Trainingserfolg ist, sollte ihm stets gebührende Beachtung zukommen. Normaler Weise ist sie schon gegeben, wenn der Turner sich in die Turnhalle begibt. Bei Schülern ist ansonsten die Motivation auch direkt vor Erledigung der Aufgabe zu steigern, indem auf das erstrebenswerte Kunststück hingewiesen, Bewegungs- und Erfolgserlebnisse angeboten und generell für eine positive Stimmung gesorgt wird. Dabei sind Furcht vor Misserfolgen und Frustrationen und Angst vor Verletzungen zu vermeiden.
5.4.2 Strategische Planung
Die Phase der Handlungsbereitschaft leitet in der Regel in eine Phase über, in der der Sportler darüber entscheidet, in welcher Weise er sein Vorhaben in die Tat umsetzen will (Abb. 5.34), er plant die ersten und sehr häufig auch noch später folgenden Tätigkeiten voraus. Diese Phase der strategischen Planung gründet sich weniger auf emotionale Befindlichkeiten, sondern mehr auf logische und taktische Überlegungen, bei denen Wissen und Erfahrungen aus früheren Situationen eine entscheidende Rolle spielen, im vorliegenden Beispiel (Abb. 5.399 etwa über Länge, Richtung und Geschwindigkeit des Anlaufes, Lage des Handaufsatzpunktes, Vermeidung von Bewegungsfehlern und vieles mehr. Damit fällt diese Phase der Vorbereitung einer anstehenden motorischen
Die Handlungsbereitschaft stellt die Grundvoraussetzung für die Ausführung turnerischer Fertigkeiten dar. Sie ist Ausdruck der Funktion des Limbischen Systems im Großhirn.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 50
Aufgabe in den Zuständigkeitsbereich derjenigen Hirnbereiche, an die logisches Denken und planende Überlegungen gekoppelt sind, der Großhirnrinde, speziell im Bereich des Stirnlappens, des sogenannten Assoziationsfeldes. Allerdings muss auch hier mit einem starken Einfluss des Limbischen Systems, des "emotional brain", gerechnet werden; denn die angestellten Überlegungen sind ständig zu bewerten, zu verwerfen und neu zu bilden, bis die als richtig vermutete Lösung gefunden ist. Es scheint demnach in dieser Phase ein ständiges "Wechselgespräch" zwischen der Assoziationsrinde (Zentrum der logischen Verknüpfungen) und dem limbischen System (Evaluationszentrum) stattzufinden.
Die Phasen der Entscheidungsleistung und der Planung müssen in der Regel jeder äußeren zielbestimmten Bewegung vorangehen. Sie zeigen jedoch in unterschiedlichen Situationen unterschiedliche zeitliche Ausdehnungen und unterschiedliche Gewichtungen. Vor der Ausführung angeborener Tätigkeiten wie Gehen und Laufen können sie ganz fehlen, da die zugehörigen Bewegungsabläufe vorprogrammiert sind, notfalls werden nur äußere Ziele wie Richtung und Geschwindigkeit geplant. Bei der Aufgabe, schwierige erlernte Bewegungsabläufe wie im Gerätturnen mental vorzubereiten, kommt der Phase der strategischen Planung größere
Limbisches System
Motivation
Assoziations-rinde
strategische Planung
SMA Bewegungsentwurf
Großhirn
Instruktionen
soziale Umwelt
zentrale Speicher
Abb. 5.35: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Phase: Strategische Planung und Bildung des Bewegungsentwurfes. Grau unterlegt: bewusstseinspflichtige Funktionen

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 51
Bedeutung zu, in Abhängigkeit vom Trainingszustand des Einzelnen. Der wenig geübte Turner wird sich vor der Ausführung schwieriger Übungen ausführlicher überlegen müssen, wie er vorgehen will, welche Einzelaktionen er besser in anderer Weise durchführen muss, welche Ratschläge des Trainers zu berücksichtigen sind, während der hoch trainierte Wettkämpfer keine strategische Planung durchzuführen braucht und sofort zur nächsten Stufe in der Bewegungsvorbereitung übergehen kann.
Die Abgrenzung der Phase der strategischen Planung ist nicht klar von der nächsten Phase, der Bildung des Bewegungsentwurfes, abzugrenzen. In der Regel mündet sie in diese Phase ein, um direkt zur Programmierung und zum Bewegungsstart überzuführen. Aus diesem Grunde wird erst im Laufe der Besprechung der nächsten Phase, der Bildung des Bewegungsentwurfes, auf die turnpraktische Bedeutung eingegangen
5.4.3 Bildung des Bewegungsentwurfes
Erst nach der Phase der bewegungsstrategischen Planung beginnt der Sportler, die Absichten in Bewegungen umzusetzen. Diese 3. Stufe der Zielmotorik ist sowohl für die Bewegungslehre, als auch für die Trainingslehre und Methodik von besonderer Bedeutung. Aus diesem Grunde muss sie an dieser Stelle (und später im Kapitel zur Bewegungsvorstellung und zum motorischen Lernen, Kap. 6.2) ausführlicher behandelt werden.
Diejenigen Hirnabschnitte des ZNS des Menschen, die die Programmierung der Bewegung organisieren, also ein sinnvolles Muster von Kontraktionsbefehlen für die Muskulatur erstellen (Basalganglien und Kleinhirn), arbeiten nicht von sich aus, sondern benötigen einen exakten Auftrag, den Bewegungsentwurf. Um einzusehen, was man sich unter dem Bewegungsentwurf vorzustellen hat, muss etwas weiter ausgeholt werden:
Die strategische Planung vor der Ausführung turnerischer Fertigkeiten, eine Aufgabe vorwiegend des Assoziationsfeldes der Großhirnrinde, stellt vor allem für Turner, deren Abläufe noch nicht automatisiert sind, eine bedeutsame Phase in der sensomotorischen Vorbereitung jedes Bewegungsablaufes dar.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 52
Zuerst muss man sich darüber klar werden, welche Informationen das Großhirn (genauer: die Großhirnrinderinde) zur Verfügung hat darüber, wie z.B. eine Radwende „aussieht“. Das Hirn kann sich nicht außerhalb des Körpers stellen und den Körper beim Turnen der Radwende zusehen. Auch wenn es Informationen aus dem visuellen System über die visuelle Struktur einer Radwende erhielte, könnte es zunächst mit diesen Kenntnissen wenig anfangen, den Bewegungsentwurf für eine Radwende zu bilden. Statt dessen muss es sich auf diejenigen Informationen stützen, die im Hirn (in Form der Bewegungsrückmeldung, der sensorischen Konsequenzen, response feedback) dann einlaufen, wenn der zum Hirn gehörende Körper gerade eine Radwende ausführt. Diese Informationen stammen vorwiegend aus dem Gewebs- und Muskelsinn (Somatosensibilität), dem Tastsinn (Oberflächensensibilität) und entsprechenden Komponenten aus dem optischen Sinn. Sie melden dem Gehirn, wie sich die Radwende „anfühlt“. Dieser Komplex an Informationen wird mit den Koordinationsprogrammen zur Radwende neuronal verknüpft und gespeichert und dient als Material zur Bildung des Bewegungsentwurfes (Abb. 5.35).
Wollte man das neuronale Geschehen, dass den Auslöseauftrag der SMA an die Koordinationsinstanzen darstellt, als eine Art Depesche in Worte fassen, müsste er in etwa wie folgt lauten: "Bitte eine solche Bewegung programmieren und durch die Muskulatur durchführen lassen, die eine derartige sensorische Rückmeldung liefert, wie ich (das SMA) sie gerade durch Erinnerung vorweg realisiert habe und in der Anlage als Muster mitschicke!" Da die Assoziationszentren zwar neuronale Verbindungen zu den Koordinationszentren besitzen, letztere aber ihrerseits keine Nervenfasern zu den Assoziationszentren schicken, da also eine unidirektionale Verbindung vorliegt, "wissen" die Assoziationszentren nicht, welche Programme in den Koordinationszentren zur Verfügung stehen. Die einzigen Informationen, die in den Assoziationszentren über die dem Organismus möglichen Körperbewegungen vorliegen, sind die mit dem Ablauf der einzelnen Bewegungen verbundenen charakteristischen Kombinationen sensorischer Rückmeldungen.
Somit ist der Bewegungsentwurf (physiologisch) mit einer Rekonstitution der Bewegungsrückmeldung von der zugehörigen motorischen Fertigkeit gleichzusetzen. Er wird vorwiegend von einem Teil der prämotorischen Großhirnrinde, dem „supplementären motorischen Areal“ (SMA), gebildet. Psychologisch stellt er somit eine Rekapitulation der Bewegungswahrnehmung (image of response feedback, die Bewegungsvorstellung) dar (s. Kap. 6.2).

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 53
Bei Hirnstrommessungen bzw. bei Bestimmung der Durchblutungsrate der Großhirnrinde hat sich gezeigt, dass das supplementäre motorische Areal (SMA) der Großhirnrinde gleichermaßen bei der Bildung der Bewegungswahrnehmung und der Bildung des Bewegungsentwurfes gegenüber anderen Bereichen der Großhirnrinde aktiviert ist (s. auch WIEMANN 1989), was darauf hindeutet, dass das SMA an beiden Phänomenen beteiligt ist. Die auf die Bildung des Bewegungsentwurfes zurückzuführende Aktivität (das Bereitschaftspotential) lässt sich u.U. schon 400 ms vor dem Bewegungsstart (und früher) beobachten, wird jedoch erst 150 ms vor Bewegungsbeginn signifikant und muss spätestens 120-100 ms vor Beginn der Bewegung endgültig aufgetreten sein. Die Aktivitäten zeigen bezüglich der topographischen Ausdehnung und hinsichtlich der Kontralateralität Ähnlichkeiten mit denjenigen elektrischen Phänomenen, die 90 bis 160 ms nach Beginn der Bewegung im EEG auftreten und das Einlaufen der Bewegungsrückmeldung bzw. die Bildung der Bewegungswahrnehmung anzeigen. Der Bewegungsentwurf (bzw. die zentrale antizipatorische Repräsentation der sensorischen Konsequenzen einer Bewegung oder die Bewegungsvorstellung) muss demnach ähnliche oder gleiche sensorische Qualitäten berühren wie die Bewegungswahrnehmung, wenn davon ausgegangen wird, dass beide Phänomene in denselben Hirnstationen realisiert werden. (KORNHUBER / DEECKE, 1965; Mc ADAM, 1974; LASSEN u.a., 1980; ROLAND u.a., 1980; SHIBASAKI u.a., 1980; WIESENDANGER, 1981; BRUNIA u.a., 1985; DEECKE, 1985).
Für Bewegungen der Alltagsmotorik (genauer: Erbmotorik) wie Gehen, Laufen, Springen.... brauchen natürlich keine Bewegungsentwürfe gebildet zu werden. Ihre Programme sind in den Basalganglien fest verankert und es bedarf nur eines kurzen Auslösers, etwa der Idee, von A nach B gehen zu wollen, um die Koordinationsprogramme zu aktivieren und die Bewegungen zu starten.
Der Übergang der vorangegangenen Phase (Phase der strategischen Planung) zur Phase der Bildung des Bewegungsentwurfes kann nicht nur fließend erfolgend, sondern auch gekennzeichnet sein durch ein häufiges „Wechselgespräch“ zwischen denjenigen Großhirnarealen, die für die beiden Funktionen verantwortlich sind, nämlich dem Assoziationsfeld und dem supplementären motorischen Areal (SMA), und das besonders dann, wenn nach zusätzlichen Informationen zur Bildung des Bewegungsentwurfes gesucht wird. Es kann angenommen werden, dass diese „Unschärfe“ mit zunehmender Übung abnimmt, der „Lernanfänger“ mag sich bei der Bildung des Bewegungsentwurfes noch mehrfach durch „Rückfragen“ strategischer Art absichern, den Könner zeichnet aus, dass er „auf Anhieb“ den richtigen Bewegungsentwurf parat hat. Gemäß Befunden aus der Wahrnehmungspsychologie (SCHALTENBRAND 1969, KEIDEL 1970) kann gefolgert werden, dass das SMA in einer kürzeste Zeitspanne von rd. 140 ms, dem sogenannten physiologischen Augenblick, nur einen Inhalt „bearbeiten“, also nur einen Bewegungsentwurf bilden kann. Erst nach Erledigung dieser Aufgabe, somit frühestens nach Ablauf dieser Zeitspanne, ist das SMA in der Lage, mit der Bildung eines neuen Bewegungsentwurfes zu beginnen.
Für die Bildung eines Bewegungsentwurfes benötigt das SMA eine Zeitspanne von mindestens 140 ms, der Zeitspanne des „physiologischen Augenblicks“. Erst danach kann das SMA die Bildung eines nächsten Bewegungsentwurfes in Angriff nehmen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 54
Da für die Programmierung und den Bewegungsstart mindestens 120 ms benötigt wird, gilt zusätzlich Folgendes:
Die angegebenen Zeitspannen sind Durchschnittswerte und werden in der Literatur unterschiedlich diskutiert, je nach eher biologisch-physiologischem oder eher psychologischem Ausgangspunkt (z.B. 63 ms; WEISS). Bemerkenswert ist in jedem Fall, dass mit der Bildung des Bewegungsentwurfes mindestens 260 ms vor Bewegungsstart begonnen werden muss.
Aus den obigen drei Feststellungen entstehen auch (bzw. insbesondere) beim Gerätturnen entscheidende, sich wechselseitig beeinflussende Konsequenzen sowohl auf die Bewegungspraxis als auch auf die Trainingspraxis und Methodik:
a) Zeitbedarf für die Bildung des Bewegungsentwurfes und Fertigkeitsniveau
Es scheint offensichtlich, dass der Zeitbedarf für das Entwerfen einer turnerischen Fertigkeit im SMA sowohl von der Komplexität als auch vom Schwierigkeitsgrad der Fertigkeit abhängig ist. Ein wenig geübter Turner (bzw. sein SMA) wird für die Bildung des Bewegungsentwurfes zu einer Rolle rückwärts in den Handstand deutlich mehr Zeit benötigen als für eine gehockte Rolle rückwärts. Das lässt sich schon am Verhalten des Turners erkennen, der sich vor Ausführung der Rolle rückwärts in den Handstand u.U. längere Zeit „konzentriert“ als vor der gehockten Rolle rückwärts, die er nahezu „spontan“ realisieren kann.
Innerhalb einer Zeitspanne von mindestens 140 ms kann das SMA sich nur mit einem Bewegungsentwurf befassen. Parallel dazu ist das SMA nicht in der Lage, eine weitere Aufgabe zu erfüllen, etwa die Bildung eines zusätzlichen Bewegungsentwurfes oder die Bearbeitung einlaufender Meldungen, etwa zur bewussten Überwachung der gerade ablaufenden Bewegung.
Die Bildung des Bewegungsentwurfes muss spätestens 120 ms vor Bewegungsstart abgeschlossen sein, d.h. spätestens 120 ms vor Bewegungsstart muss der Auftrag zur Bewegungsprogrammierung vom SMA an die Koordinationsinstanzen abgeschickt werden.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 55
b) Simultanität von Ablaufkontrolle und Bildung eines Bewegungsentwurfes
Da das SMA nicht in der Lage ist, zwei oder mehrere Aufgaben zeitlich parallel zu erledigen, kann während des Ablaufes einer Fertigkeit mit hohem Schwierigkeitsgrad, die einer permanenten Ablaufkontrolle bedarf, nicht gleichzeitig ein Bewegungsentwurf für eine folgende Fertigkeit gebildet werden, d.h., dass erst nach Ablauf der ersten Fertigkeit das SMA die nächste Fertigkeit entwerfen kann, wozu eine Mindestzeit von 140 ms (bei schwierigen Bewegungsabläufen entsprechend länger) notwendig ist. Das bedeutet für die Bewegungskombination von Abb. 5.33: Der ungeübte Turner muss zwischen den Elementen Pausen zur Bildung des nächsten Bewegungsentwurfes schieben, die unter Umständen nur einige Zehntel Sekunden betragen, den Bewegungsfluss aber deutlich stören. Diese Pausen werden mit zunehmender Übung kürzer. Nur der geübte Turner bewegt sich fließend ohne Unterbrechungen (s. unter a).
c) Verkürzung der Zeitspanne zur Bildung eines Bewegungsentwurfes
Somit erscheint es sinnvoll den Zeitbedarf zur Bildung des Bewegungsentwurfes bis auf den Mindestbedarf von 140 ms zu verkürzen, was nur durch entsprechendes Training, d.h. durch Festigung der Bewegungsprogramme möglich ist. Ziel ist es, die Komplexität des Bewegungsentwurfes zunehmend zu reduzieren, so dass auf eine kurzes, im SMA gebildetes „Signal“ die Programmierung des zugehörigen Bewegungsablaufes gestartet werden kann.
d) Automatsierung der Bewegungsabläufe
Die Festigung der Bewegungsprogramme bedeutet gleichzeitig, dass nach dem Start der Programmierung und der Exekution die Bewegung ohne weitere Kontrolle durch das SMA, also „automatisch“, ablaufen kann und das SMA „frei“ ist für weitere Aufgaben, z.B. für die Bildung eines Bewegungsentwurfes für eine sich anschießende Bewegung.
Die Punkte a) bis d) sind anhand praktischer Beispiele zu erläutern: Da sich die einzelnen Punkte wechselseitig bedingen, werden sie nicht in der vorn genannten Reihenfolge besprochen.
1. Will ein (wenig geübter) Turner die Bewegungsfolge Radwende – Rolle rückwärts gehockt turnen, hat er anfangs – in der Ausgangsstellung - genügend Zeit, den Bewegungsentwurf für die Radwende zu bilden und benötigt dann im Übergang von der

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 56
ersten zur zweiten Übung nur wenig Zeit für den Bewegungsentwurf zur (in der Ausführung leichteren) Rolle rückwärts gehockt. Folglich wird ihm die Bewegungsfolge flüssig und ohne nennenswerte Pause zwischen beiden Übungen gelingen. Anders ist es, wenn er an die Radwende einen Handstandüberschlag rückwärts anschließen will. Ist dieser wenig geübt, muss der Turner nach der Radwende eine Pause zur Bildung des Bewegungsentwurfes des Flick-Flack einlegen. Tut er das nicht, läuft er Gefahr, dass er für die zweite Übung nur einen unzureichenden Bewegungsentwurf bildet: die Übung kann misslingen.
Ist für einen Turner die Kontergrätsche am Reck (Abb. 5.30) eine anspruchsvolle Übung, d.h., muss er diese durch einen ausführlichen Bewegungsentwurf vorbereiten, tut er gut daran, vor dem Vorschwung, der die Kontergrätsche einleitet, einen einfachen Riesenumschwung rückwärts zu turnen, damit er ungestört und rechtzeitig den Bewegungsentwurf für die Kontergrätsche bilden kann – und zwar während des Ablaufes des einleitenden Riesenumschwunges, den er sicher beherrscht, d.h., der „automatisch“, ohne bewusste Kontrolle, gemäß den gespeicherten Koordinationsprogrammen, abläuft. In der Regel wird der Turner an die Kontergrätsche einen zweiten Riesenumschwung anschließen, der in seiner Vorbereitung im SMA keinen großen zeitlichen Aufwand verlangt. An eine Kontergrätsche direkt eine zweite anzufügen (Abb. 5.30) setzt voraus, dass die erste Kontergrätsche derart gut beherrscht wird, dass schon während ihres Ablaufes der Bewegungsentwurf für die zweite Kontergrätsche gebildet werden kann und dafür auch noch genügend Zeit zur Verfügung steht. Geht man davon aus, dass etwa mit dem Zugreifen nach der ersten Kontergrätsche die Aktionen für den folgenden Vorschwung beginnen müssen, muss rd. 120 ms vor dem Zugreifen die Bildung des Bewegungsentwurfes beendet sein, da mit dem Zugreifen feststehen muss, ob eine zweite Kontergrätsche oder ein einfacher Riesenumschwung angeschlossen werden soll; denn in beiden Fällen unterscheiden sich die Aktionen in charakteristischer Weise. Das bedeutet aber auch, dass spätestens 260 ms vor dem Zugreifen mit der Bildung des Bewegungsentwurfes begonnen werden muss – in der Regel aber noch deutlich früher. Nimmt man weiterhin an, dass der freie Flug eine Steighöhe von 1 m zeigt, dauert der freie Flug etwa 0,6s. Folglich muss die Bildung des Bewegungsentwurfes für die zweite Kontergrätsche im günstigsten Fall etwa im Gipfelpunkt der Flugkurve der ersten Kontergrätsche begonnen werden.
Das gilt in gleichem Maße auch für die Bewegungskombination Abb. 5.33.
Mehrere turnerische Fertigkeiten lassen sich nur dann fließend hintereinander turnen, wenn die Bildung des Bewegungsentwurfes für die an zweiter oder dritter Stelle in der Bewegungsfolge stehende Einzelfertigkeit keine zu große Zeitspanne in Anspruch nehmen muss.
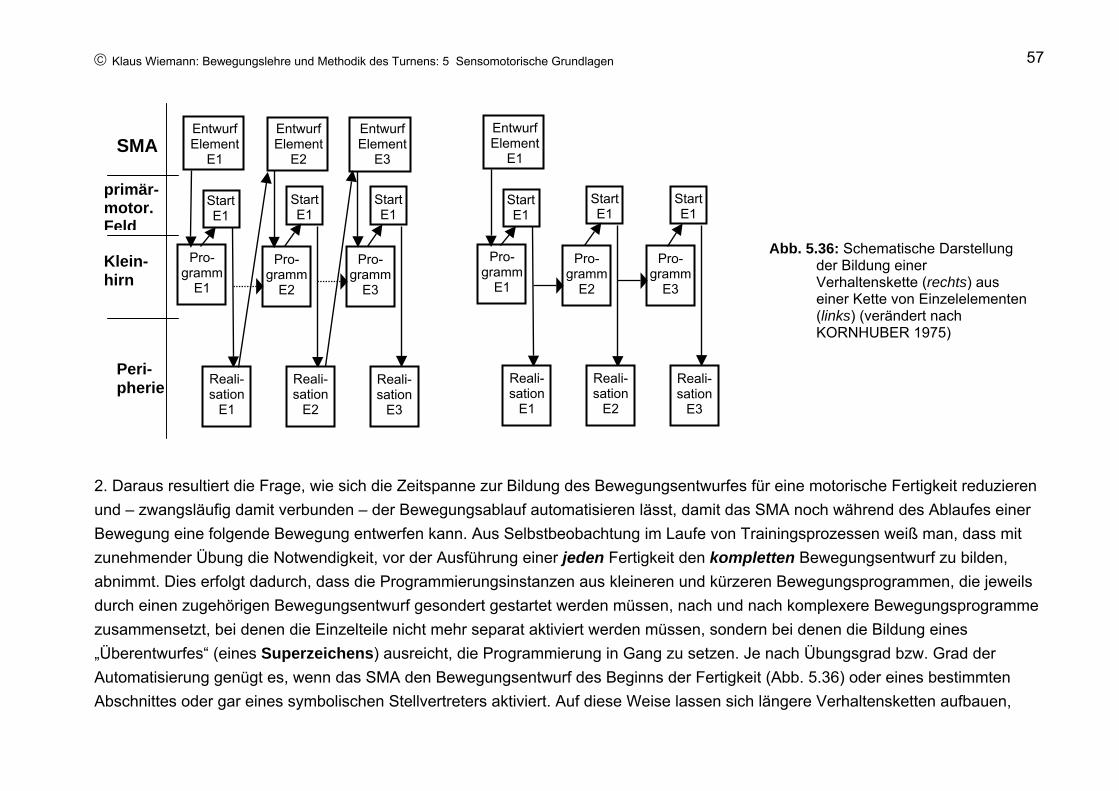
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 57
2. Daraus resultiert die Frage, wie sich die Zeitspanne zur Bildung des Bewegungsentwurfes für eine motorische Fertigkeit reduzieren und – zwangsläufig damit verbunden – der Bewegungsablauf automatisieren lässt, damit das SMA noch während des Ablaufes einer Bewegung eine folgende Bewegung entwerfen kann. Aus Selbstbeobachtung im Laufe von Trainingsprozessen weiß man, dass mit zunehmender Übung die Notwendigkeit, vor der Ausführung einer jeden Fertigkeit den kompletten Bewegungsentwurf zu bilden, abnimmt. Dies erfolgt dadurch, dass die Programmierungsinstanzen aus kleineren und kürzeren Bewegungsprogrammen, die jeweils durch einen zugehörigen Bewegungsentwurf gesondert gestartet werden müssen, nach und nach komplexere Bewegungsprogramme zusammensetzt, bei denen die Einzelteile nicht mehr separat aktiviert werden müssen, sondern bei denen die Bildung eines „Überentwurfes“ (eines Superzeichens) ausreicht, die Programmierung in Gang zu setzen. Je nach Übungsgrad bzw. Grad der Automatisierung genügt es, wenn das SMA den Bewegungsentwurf des Beginns der Fertigkeit (Abb. 5.36) oder eines bestimmten Abschnittes oder gar eines symbolischen Stellvertreters aktiviert. Auf diese Weise lassen sich längere Verhaltensketten aufbauen,
Entwurf Element
E1
Entwurf Element
E2
Entwurf Element
E3
Start E1
Start E1
Start E1
Pro-gramm
E1
Pro-gramm
E3
Pro-gramm
E2
Reali-sation
E3
Reali-sation
E2
Reali-sation
E1
Entwurf Element
E1
Start E1
Start E1
Start E1
Pro-gramm
E1
Pro-gramm
E3
Pro-gramm
E2
Reali-sation
E3
Reali-sation
E2
Reali-sation
E1
Peri-pherie
primär-motor. Feld
Klein-hirn
SMA
Abb. 5.36: Schematische Darstellung der Bildung einer Verhaltenskette (rechts) aus einer Kette von Einzelelementen (links) (verändert nach KORNHUBER 1975)

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 58
wobei der Zeitbedarf zur Bildung des Bewegungsentwurfes der einzelnen Elemente der Bewegungsfolge zunehmend geringer wird oder – bei Bewegungsautomatisierung - völlig verschwinden kann.
Die angesprochenen Probleme gelten natürlich nicht für angeborenen (genetisch vorprogrammierte) Bewegungsgrundmuster wie Gehen, Laufen, Springen, Greifen, Klimmen, Klettern, Werfen u.a.m. sondern nur für komplexe vom Menschen ausgedachte Bewegungskunstformen wie es die meisten Gerätübungen darstellen. Da das Gehirn des Menschen überfordert wäre, müsste es für jede aus mehreren motorischen Elementen bestehende Verhaltenskette ein eigenes Programm bilden und abspeichern, bleibt nur die Möglichkeit anzunehmen, dass die unterschiedlichen Verhaltensketten aus einer beschränkten Anzahl von Elementen, für die jeweils ein Programm zur Verfügung steht, zusammengesetzt werden, vergleichbar mit der Bildung einer Vielzahl von Wörtern aus nur wenigen Buchstaben. Wie dieses Geschehen aus physiologischer Sicht funktioniert, ist noch relativ unbekannt. Tatsache scheint nur zu sein, dass das Neukleinhirn (Neocerebellum) dabei eine entscheidende Rolle spielt. Gemäß einem Modell von KORNHUBER (1975) könnte die rechtzeitige Programmierung der einzelnen Elemente der Verhaltensketten durch eine interne Rückmeldung gewährleiste werden: Startet von der primären motorischen Rinde der Exekutionsbefehl zur Realisierung des ersten Verhaltenselementes zu den Muskeln, werden noch innerhalb des Gehirns mittels Nervenbahnen, die von den die Kontraktionsbefehle übermittelnden Nerven abzweigen, die Programmierungsinstanzen (hier: Kleinhirn) über den erfolgten Start informiert und sie zur Programmierung des nächst folgenden Elementes angeregt, so dass die Programmierung und die Bildung des Exekutionsbefehls für das folgende Verhaltenselement abgeschlossen sind, bevor noch die Realisation des voraufgehenden Elementes beendet ist (Abb. 5.36).
3. Was für die Ausführung zweier hintereinander geturnter Fertigkeit gilt, sollte auch für das Turnen von längeren Übungsfolgen oder kompletter Pflicht- oder Kürübungen gelten. Da in jedem Fall die Bildung des Bewegungsentwurfes (oder eines symbolischen Abschnittes desselben) einige 100 ms in Anspruch nimmt und spätestens 120 ms vor Bewegungsbeginn abgeschlossen sein muss, ist es beim Turnen von Übungsfolgen erforderlich, dass die Bildung der Bewegungsentwürfe bzw. der Ketten von Bewegungsentwürfen zeitversetzt dem aktuellen Geschehen voranläuft, d.h., der Bewegungsentwurf für die nachfolgende Fertigkeit ist schon zu einem Zeitpunkt zu bilden, in dem die vorangehende Bewegung noch nicht abgeschlossen ist (s.o.!). Bei der Übungsverbindung Anlauf – Radwende – Handstandüberschlag rückwärts - Salto rückwärts – Grätschwinkelsprung (Abb. 5.37) ist der Bewegungsentwurf für den Salto rückwärts etwa in der Stützphase des Handstandüberschlages rückwärts zu bilden. Erst dadurch ist gewährleistet, dass die
Durch ständiges Üben lässt sich der Bewegungsentwurf, den das SMA der Großhirnrinde vor Bewegungsstart zu bilden hat, in seiner Komplexität reduzieren, so dass sich der für die Bildung des Bewegungsentwurfes notwendige Zeitbedarf bis auf ein Minimum (140ms) einschränken lässt.
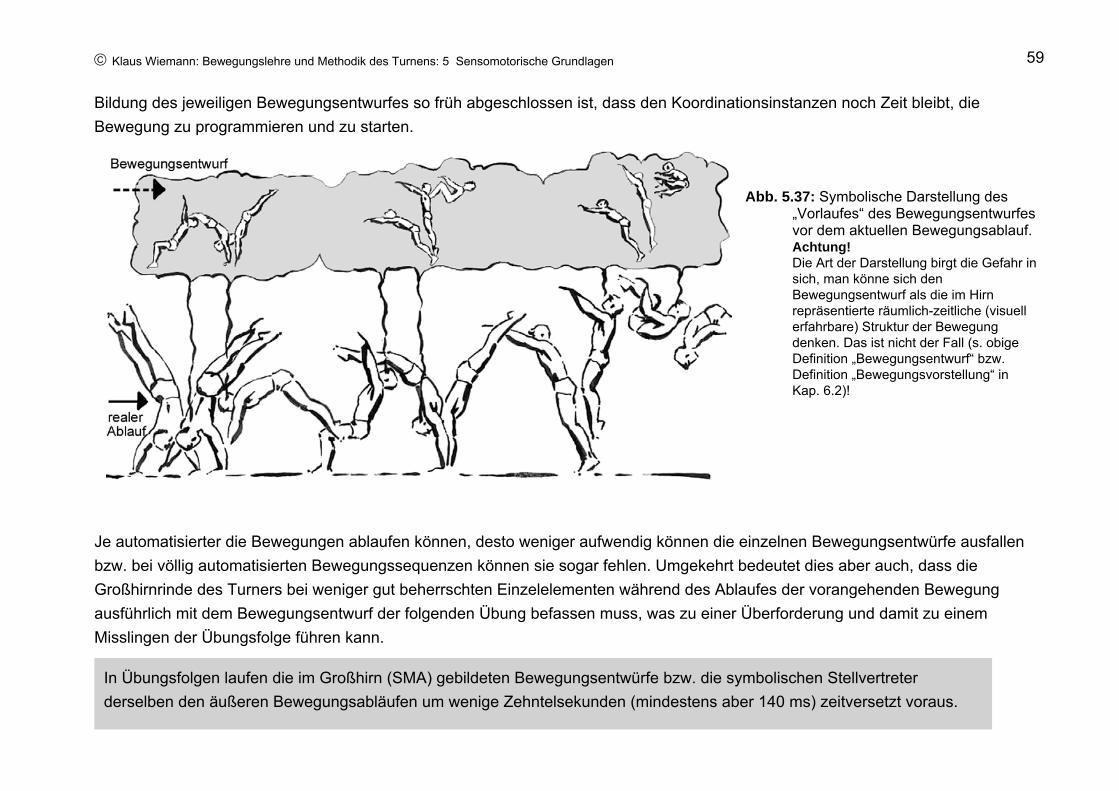
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 59
Bildung des jeweiligen Bewegungsentwurfes so früh abgeschlossen ist, dass den Koordinationsinstanzen noch Zeit bleibt, die Bewegung zu programmieren und zu starten.
Je automatisierter die Bewegungen ablaufen können, desto weniger aufwendig können die einzelnen Bewegungsentwürfe ausfallen bzw. bei völlig automatisierten Bewegungssequenzen können sie sogar fehlen. Umgekehrt bedeutet dies aber auch, dass die Großhirnrinde des Turners bei weniger gut beherrschten Einzelelementen während des Ablaufes der vorangehenden Bewegung ausführlich mit dem Bewegungsentwurf der folgenden Übung befassen muss, was zu einer Überforderung und damit zu einem Misslingen der Übungsfolge führen kann.
In Übungsfolgen laufen die im Großhirn (SMA) gebildeten Bewegungsentwürfe bzw. die symbolischen Stellvertreter derselben den äußeren Bewegungsabläufen um wenige Zehntelsekunden (mindestens aber 140 ms) zeitversetzt voraus.
Abb. 5.37: Symbolische Darstellung des „Vorlaufes“ des Bewegungsentwurfes vor dem aktuellen Bewegungsablauf. Achtung! Die Art der Darstellung birgt die Gefahr in sich, man könne sich den Bewegungsentwurf als die im Hirn repräsentierte räumlich-zeitliche (visuell erfahrbare) Struktur der Bewegung denken. Das ist nicht der Fall (s. obige Definition „Bewegungsentwurf“ bzw. Definition „Bewegungsvorstellung“ in Kap. 6.2)!

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 60
Für die Turnpraxis folgt daraus: Will ein Turner in einer Übungsverbindung ein für ihn schwieriges Element ausführen, sollte er direkt vor diesem Element ein leichteres Element ausführen, damit er ungestört - während des Ablaufes des leichteren Elementes - den Bewegungsentwurf für das schwierige Element bilden kann, und er sollte nach dem schwierigen Element ein leichtes Element einplanen, da er während des Ablaufes des schwierigen Elementes keine Gelegenheit hat, einen komplexen Bewegungsentwurf für ein folgendes Element zu bilden.
4) Letztendlich entspricht auch der Aufbau einer einzelnen sportmotorischen Fertigkeit aus mehreren Einzelaktionen, die teilweise als motorische Grundmuster angesehen werden können, einer Überführung von einzeln produzierten Bewegungsentwürfen zu den
Abb. 5.38: Beispiel von simultan ablaufenden Körperaktionen im Laufe der Stützkehre vorwärts am Barren. Die horizontalen Balken verdeutlichen die annähernd zeitliche Ausdehnung der folgenden Aktionen: a) Vorschwingen und geschlossen und gestreckt Halten der Beine. b) Verdrehen (Verschrauben) des Rumpfes. c) Abdrücken und Umgreifen des Außenarmes. d) Umsetzen des Innenarmes

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 61
Teilaktionen zu einem komplexen Bewegungsentwurf der Fertigkeit. Im Laufe der Stützkehre vorwärts am Barren sind unter anderem die in Abb. 5.38 dargestellten Einzelaktionen zu realisieren. Soll diese Fertigkeit erstmalig ausgeführt werden, wäre die motorische Hirnrinde (das SMA) überfordert, müsste sie die Bewegungsentwürfe aller zur Ausführung notwendigen Aktionen einzeln in der korrekten Abfolge erstellen. Um die Ausführung trotzdem zu ermöglichen, müssen Bedingungen geschaffen werden, die es dem Hirn gestatten, auf die Bildung des einen oder anderen Bewegungsentwurfes zu verzichten. Dies erfolgt durch Lernhilfen, die die Ausführung (und damit auch die neuronale Planung) der einen oder anderen Aktion erübrigen, so dass sich das SMA vorerst auf die Erstellung der Bewegungsentwürfe für die bedeutendsten Aktionen beschränken kann (Abb. 5.39a).
Hat sich durch entsprechendes Üben aus den Teilaktionen eine Verhaltenskette konsolidiert, können die Lernhilfen reduziert werden: Das SMA benötigt geringere Zeitspannen zur Bildung der Bewegungsentwürfe und kann im Laufe der Bewegung – Schritt für Schritt - früher umschalten auf die Planung kommender Aktionen, etwa ein Schwingen im Stütz (Abb. 5.39b) oder einen Vorschwung im Stütz mit Ablegen auf die Oberarme zwecks Ausführung einer Oberarmkippe (Abb. 5.39c) oder auf ein direkt nach dem Stützfassen folgendes Senken aus dem Stütz in den Sturzhang zwecks Ausführung eine Felge (Abb. 5.39d). Da die Stützkehre am Barren kein angeborenes Grundmuster darstellt und somit in den Programmierungsinstanzen kein entsprechendes Grundprogramm „Stützkehre“ vorliegt, muss im Laufe eines Lernprozesses aus den Programmen einfachster Grundmuster (Stützen, Greifen ...) und schon gelernter turnerischer Fertigkeiten (Schwingen im Stütz, Körperdrehung um die Längsachse mit gestreckt-geschlossenen Beinen...) das Programm für die Verhaltenskette „Stützkehre“ zusammengesetzt werden.
In der methodischen Planung (Kap. 7) zu turnerischen Fertigkeiten ist es zentrales Anliegen, methodische Reihen oder Lernprogramme zu entwickeln, die es dem Hirn ermöglichen, aus den Bewegungsentwürfen von Einzelaktionen den Bewegungsentwurf einer turnerischen Fertigkeit zu erstellen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 62
Abb. 5.39: Stufen im Fertigkeitsniveau der Stützkehre am Barren. Grau ausgefüllte Bewegungsphase: Das Gehirn ist mit der Kontrolle der ablaufenden Bewegung (=Stützkehre) beschäftigt und kann keinen Bewegungsentwurf für kommende Aktionen bilden. Erst in der weiß ausgefüllten Bewegungsphase ist das Gehirn „frei“ zur Bildung neuer Bewegungsentwürfe.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 63
5.4.4 Bewegungsprogrammierung
Sobald der Bewegungsentwurf gebildet ist und als Programmierungsauftrag an die unter der Großhirnrinde gelegenen (= subkortikalen) Programmierungsinstanzen geschickt worden ist, haben diejenigen Hirnstationen, an deren Funktion bewusste Entscheidungen hinsichtlich der Bewegungsplanung gekoppelt sind, ihren Einfluss auf das weitere Geschehen abgegeben (Abb. 5.40, rechts, Abb. 5.41). Erst mit dem Einlaufen der Bewegungsrückmeldung, also frühestens 90-100 ms (in der Regel aber erst nach 140-200 ms) nach Beginn des äußeren Bewegungsablaufes wird der Organismus bzw. sein Bewusstsein, erfahren, ob die Bewegung wie beabsichtigt vollzogen worden ist.
Als subkortikale Generatoren der Bewegungsprogramme kommen einerseits die Basalganglien (an der Basis des Großhirns gelegene Schaltstationen; Abb. 5.3), andererseits das Kleinhirn in Betracht. Beide arbeiten „parallel“ an der Programmierung, wobei die Basalganglien die Grundprogramme der geplanten Aktionen (Abb. 5.40, rechts), insbesondere den erbmotorischen Anteil, also diejenigen Programme zur Koordination der Bewegungsgrundmuster liefern, die als angeboren bezeichnet werden müssen.
Dem Kleinhirn obliegen gleich mehrere Aufgaben. Da dem Kleinhirn (insbesondere dem stammesgeschichtlich älteren Teil) die gesamte Information über die augenblickliche körperliche Situation zur Verfügung steht, z.B. über die momentane Lage des Körpers im Raum, über die Gelenkstellungen, über die gerade ablaufenden muskulären Aktionen und über taktile Gegebenheiten, fällt ihm die Aufgabe zu, die vorherrschende innere und äußere Situation des Körpers bei der Programmierung zu berücksichtigen. Wenn z.B. der Bewegungsentwurf „Rolle vorwärts am Boden“ gebildet wurde, ist bei der Programmierung von Bedeutung, ob die Rolle vorwärts aus dem Hockstand startet oder etwa aus einem Handstand geturnt werden soll. Bei der Erstellung der Kontraktionsbefehle für die Muskulatur verlangt die unterschiedliche Ausgangsposition entsprechend unterschiedliche motorische Kommandos.
Nach Abschluss des Bewegungsentwurfes, sobald die Innervationskaskade von dem SMA in tiefere Hirnschichten abgetaucht ist, kann das weitere Geschehen, die Programmierung und der Bewegungsstart, willentlich nicht mehr geändert werden. Die Bewegung wird so ablaufen, wie es im Bewegungsentwurf vorgesehen ist.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 64
Limbisches System
Assoziations-rinde
strategische Planung
SMA Bewegungsentwurf
Primärmot. Rinde
Bewegungs-start
Neukleinhirn Programmierung Erwerbmotorik
Altkleinhirn
stato-vestib. Regulation
sensorischer Thalamus
motorischer Thalamus
Hirnstamm
Großhirn
Basalganglien
Programmierung Erbmotorik
zentrale Speicher
Abb. 5.40: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Bewegungsprogrammierung. Grau unterlegt: bewusstseinspflichtige Funktionen

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 65
Dieses Beispiel ist natürlich nicht in jedem Fall zutreffend. Ein wenig erfahrener Turner wird schon bei der Bildung des Bewegungsentwurfes die unterschiedlichen Ausgangspositionen berücksichtigen, so dass er bei der Rolle vorwärts aus dem Hockstand einen anderen Bewegungsentwurf produziert als bei der Rolle vorwärts aus dem Handstand. Das bedarf jedoch einer erhöhten Aufmerksamkeit und einer längeren Planungszeit. Je mehr aber ein Turner seine Bewegungen automatisiert hat und je symbolhafter seine Bewegungsentwürfe ausfallen, desto weitgehender übernehmen die subkortikalen Funktionsgeneratoren (insbesondere das Neukleinhirn) die Differenzierung gemäß den vorherrschenden äußeren Bedingungen.
Dem Kleinhirn kommt eine zweite Aufgabe zu, insbesondere dem stammesgeschichtlich jungen Teil der Kleinhirnhemisphären (dem „Neukleinhirn“, Neocerebellum): Es muss als Hauptinitiator der Programme für die schnelle erlernte Zielmotorik gelten. Damit muss das Neukleinhirn als das motorische Lernhirn des Menschen gewertet werden.
Die motorischen Funktionsgeneratoren (Basalganglien und Kleinhirn) benötigen – beginnend mit dem Absenden des Bewegungsentwurfes vom SMA 40 – 60 ms für die Programmierung der Bewegung (Abb. 5.41), wobei – grob vereinfacht – die Basalganglien bestimmen, welche Muskeln in welcher Reihenfolge tätig werden sollen und das Kleinhirn festlegt, in welcher Intensität und zeitlichen Ausdehnung dies geschehen soll.
5.4.5 Start und Realisation der Bewegung
Die von den Basalganglien und dem Kleinhirn entworfenen Programme werden auf dem Weg über den Thalamus dem primärmotorischen Areal der Großhirnrinde (MI, s. Abb. 5.3, und Abb. 5.41) zur Verfügung gestellt, das auf dem Wege über die schnellleitende Pyramidenbahn (Abb. 5.3) und die α-Motoneuronen die Muskelfaser zur Kontraktion anregt und somit die Bewegung startet. Neben den Pyramidenbahnen versorgen noch weitere motorische Nervenbahnen die Muskulatur. So laufen z.B. aus den Basalganglien langsamer leitende Fasern (ohne Umweg über die Großhirnrinde) direkt zu den Motoneuronen im Rückenmark. Ebenso werden letztere von Fasern innerviert, die über Umschaltstationen im Hirnstamm aus dem Kleinhirn stammen. Während letztere vorwiegend für die Stützmotorik (s. Kap. 5.3) verantwortlich sind, liefern erstere
Gemäß Auftrag des SMA (also: gemäß Bewegungsentwurf) werden a) von den Basalganglien und b) vom Neocerebellum die Programme der beabsichtigten Bewegung erstellt.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 66
einen Beitrag zur Zielmotorik, indem sie z.B. die γ-Motoneurone und somit die Muskelspindeln (s. Abb. 5.5) „vorinnervieren“, d.h. auf die durch die Pyramidenbahnen zu erwartenden Bewegungsaufträge vorbereiten
Das primärmotorische Areal (MI) der Großhirnrinde wird zusammen mit den schnellleitenden Pyramidenbahnen zum pyramidal-motorischen System zusammengefasst. Da dieses System in dieser Ausprägung nur beim Menschen vorkommt, wurde früher dieses System für die Planung und Ausführung der bewussten menschlichen Willkürmotorik verantwortlich gemacht und dem extrapyramidalen System, den außerhalb des Pyramidensystems gelegenen motorischen Strukturen des ZNS, die die unwillkürlichen Bewegungen steuern sollten, gegenübergestellt. Seit mehr als 20 Jahren, insbesondere seit den Forschungen von ECCLES (z.B. 1975, 1982), KORNHUBER (1975) und anderen weiß man, dass diese Trennung unberechtigt ist, dass beide Systeme zusammen funktionieren und das Pyramidensystem lediglich ein „Hilfsorgan“ zum schnellen Start der Zielmotorik darstellt – ohne Möglichkeit der bewussten Kontrolle. Denn auch „unwillkürlich“ ausgelösten zielmotorischen Aktionen, also auch bei unwillkürlich durchgeführten turnerischen Fertigkeiten funktionieren die diversen Hirnstationen nach dem hier dargestellten Schema (Abb. 5.32, Abb. 5.41 ), wobei die Bildung des Bewegungsentwurfes derart „automatisch“ erfolgt, dass das Geschehen nicht ins Bewusstsein vordringt. Allerdings findet man in der sportwissenschaftlichen Literatur auch heute noch die funktionelle Trennung der beiden Systeme, was zu entscheidenden Missverständnissen in der angewandten Trainingslehre und Methodik führen kann.
Innerhalb des gesamten Prozesses der Planung und Durchführung einer zielmotorischen Aktion stellt aus trainingstheoretischer und lehrmethodischer Sicht die Bildung des Bewegungsentwurfes „die“ entscheidende Phase dar, weil sie einerseits letztendlich über die Form der geplanten Bewegung entscheidet und andererseits der zeitlich letzte bewusstseinsfähige Prozess vor Beginn der Bewegung darstellt. Ebenso ist von Bedeutung, dass der Informationsfluss von dem SMA bis zur Muskulatur (nahezu) rückwirkungsfrei, also unidirektional abläuft.
Die primärmotorische Großhirnrinde startet (ohne Beteiligung des Bewusstseins) die geplante Bewegung rd. 30 ms vor Bewegungsbeginn und schickt die Bewegungsbefehle über die Pyramidenbahn zu den α-Motoneuronen des Rückenmarkes und von dort weiter zur Muskulatur.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 67
Primärmot. Rinde
Bewegungs-start
Motoneurone Innervation
Muskeln Realisation
Hirnstam
Großhirn
Rücken-k
mechanische Umwelt
Limbisches System Motivation
Assoziationsrinde strategische
SMA Bewegungsentwurf
Neukleinhirn
Program-mierung Erwerb-
Basal-ganglien Program-mierung
Erbmotorik
Thalamus
Primärmot. Rinde Bewegungsstart
Motoneurone Innervation
Muskeln Realisation
Abb. 5.41: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv). links: Bewegungsstart. rechts: unidirektionaler Informationsfluss vom Abschluss der Bildung des Bewegungsentwurfes bis zur Realisation (verändert nach POPPER/ECCLES 1982). Grau unterlegt: bewusstseinspflichtige Funktionen.
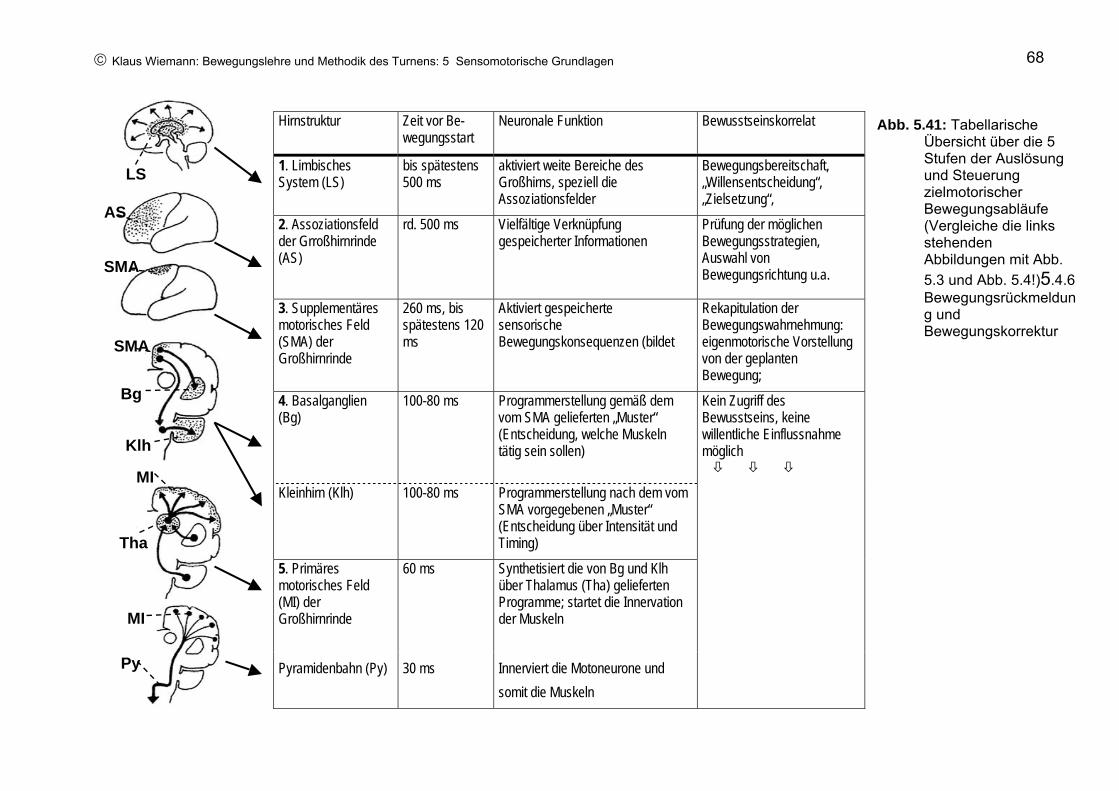
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 68
Hirnstruktur Zeit vor Be-wegungsstart
Neuronale Funktion Bewusstseinskorrelat
1. Limbisches System (LS)
bis spätestens 500 ms
aktiviert weite Bereiche des Großhirns, speziell die Assoziationsfelder
Bewegungsbereitschaft, „Willensentscheidung“, „Zielsetzung“,
2. Assoziationsfeld der Grroßhirnrinde (AS)
rd. 500 ms Vielfältige Verknüpfung gespeicherter Informationen
Prüfung der möglichen Bewegungsstrategien, Auswahl von Bewegungsrichtung u.a.
3. Supplementäres motorisches Feld (SMA) der Großhirnrinde
260 ms, bis spätestens 120 ms
Aktiviert gespeicherte sensorische Bewegungskonsequenzen (bildet
Rekapitulation der Bewegungswahrnehmung: eigenmotorische Vorstellung von der geplanten Bewegung;
4. Basalganglien (Bg)
100-80 ms Programmerstellung gemäß dem vom SMA gelieferten „Muster“ (Entscheidung, welche Muskeln tätig sein sollen)
Kein Zugriff des Bewusstseins, keine willentliche Einflussnahme möglich
Kleinhirn (Klh)
100-80 ms Programmerstellung nach dem vom SMA vorgegebenen „Muster“ (Entscheidung über Intensität und Timing)
5. Primäres motorisches Feld (MI) der Großhirnrinde
60 ms Synthetisiert die von Bg und Klh über Thalamus (Tha) gelieferten Programme; startet die Innervation der Muskeln
Pyramidenbahn (Py) 30 ms Innerviert die Motoneurone und somit die Muskeln
Abb. 5.41: Tabellarische Übersicht über die 5 Stufen der Auslösung und Steuerung zielmotorischer Bewegungsabläufe (Vergleiche die links stehenden Abbildungen mit Abb. 5.3 und Abb. 5.4!)5.4.6 Bewegungsrückmeldung und Bewegungskorrektur
LS
AS
SMA
SMA
Bg
Klh
Py
Tha
MI
MI

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 69
5.4.6 Bewegungsrückmeldung und Bewegungskorrektur
Sobald eine motorische Aktion durch die Funktion der Muskulatur gestartet ist, wird die biomechanische Situation des sich bewegenden Körpers sowie die mechanische Umwelt in unterschiedlicher Form verändert, was auf den Körper des sich Bewegenden zurückwirkt (Abb. 5.45). Diese Rückwirkungen werden von den diversen sensorischen Systemen registriert und dem Zentralnervensystem zur Auswertung zur Verfügung gestellt.
Als entscheidendes Kriterium für die Bewegungsrückmeldung und Bewegungskorrektur gilt die Zeit, die einerseits die Ausführung der Bewegung in Anspruch nimmt und die andererseits die Rückmeldung von den Sinneszellen (Rezeptoren) bis zu den verarbeitenden Instanzen des ZNS benötigt (Abb. 5.46). Aus diesem Grunde sind bei der Besprechung der Bewegungsrückmeldung und der zugehörigen Bewegungskorrektur prinzipiell zwei Bewegungstypen zu unterscheiden, nämlich a) schnelle „ballistische“ Bewegungen, die mit relativ hohen Beschleunigungen einhergehen, durch explosive Muskelkontraktionen gekennzeichnet sind und in der Regel nach 100 ms – 200 ms abgeschlossen sind, und b) langsame („rampenförmige“) Nachführbewegungen, Halteaktionen und rhythmische Bewegungswiederholungen, deren Ablaufzeiten sich über mehrere Sekunden und Minuten erstrecken können. In der physiologischen Literatur werden den explosiven ballistischen Aktionen die langsamen „rampenförmigen“ Aktionen gegenübergestellt. Da die Verwendung dieses Begriffes eher eine physiologische Rechtfertigung besitzt, umgangssprachlich aber missverständlich sein kann, soll im Folgenden der Begriff „Nachführaktionen“ verwendet werden, mit der Maßgabe, hier Hebe-, Absenk- und Halteaktionen mit einzubeziehen.
Aus sensomotorischer Sicht lassen sich alle turnerischen Fertigkeiten in folgende Gruppen von Bewegungstypen gliedern:
Ballistische Bewegungen (Aktionen) sind impulsive Schlag-, Wurf-, Stoß-, Schwung- oder Sprungaktionen. Sie kennzeichnen sich durch eine kurze (80-200 ms dauernde) heftige (= explosiv-ballistische) Kontraktion eines Muskels bzw. durch simultan ablaufende, allenfalls geringfügig zeitversetzte, Kontraktionen mehrerer Muskeln, die zu einer entsprechen explosive Beschleunigung (oder Abbremsung) eines Körperteils oder des ganzen Körpers führen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 70
1) Ballistische Aktionen: Diese können neben der ballistischen (Muskel-) Aktion eine oder wenige vorangehende vorbereitende ballistische Aktionen enthalten und können in einer weiteren ballistischen Aktion, meistens aber in Nachführ- oder Halteaktionen ausklingen. Beispiel: Der Salto vorwärts gliedert sich in drei ballistische Abschnitte, den Absprung, die Einrollbewegung der Flugphase und die Landung. Der Anlauf zum Salto muss zum folgenden Bewegungstyp (2) gerechnet werden. Die ballistische Aktion des Aufschwingens in den Handstand klingt im Handstand (Abb. 5.42), einer Halteaktion, aus.
2) Zyklisch sich wiederholende ballistische Aktionen: Beispiele: Alle Anläufe zu Sprungbewegungen, aber auch Schwünge im Stütz oder Hang, wobei sich einzelne ballistische Aktion (Laufschritte, Vor- und Rückschwünge) periodisch wiederholen. Die einzelne ballistische Aktion zeigt dabei in jedem Fall die zugehörigen dynamischen (= explosiv) und zeitlichen (= 80-200 ms) Charakteristika, die Gesamtaktion kann jedoch mehrere Sekunden (Anlauf zum Sprung) oder einige Stunden (Marathonlauf) in Anspruch nehmen.
3) Vorwiegend „geführte“ Aktionen: Beispiel: Geführte Aktionen treten gelegentlich bei Übergängen von einer ballistische Aktion zur nächsten auf. Nach einem ballistischen Vorschwung im Oberarmhang kann der Körper zur Gestaltung einer weiträumigen Bewegung durch eine Kerzenposition bis in die gewinkelte Kipplage geführt werden, wo sich dann der ballistische Kippstoß anschließt.
4) Aktionen des Hebens und Senkens: Beispiele: Dies sind vielfach „Kraftelemente“ wie Heben in den Handstand oder Senken aus dem Sturzhang an den Ringen in den Seitspannstütz. Aber auch im Laufe komplexer Aktionen können solche „Nachführaktionen“ ballistische Aktionen vorbereiten oder miteinander verbinden (Abb. 5.42).
Abb. 5.42: Beispiel einer Folge turnerischer Fertigkeiten, bestehend aus einer Kombination von ballistischen Aktionen (s. schwarze Balken: Abdruckaktion zum Aufschwingen in den Handstand; schnelles Anhocken der Beine) und verschiedenen Halte-, Absenk- und Hebeaktionen (Senken zum Aufsetzen der Hände auf den Boden, Halten des Körpers im Handstand, Aufrichten in den Stand)
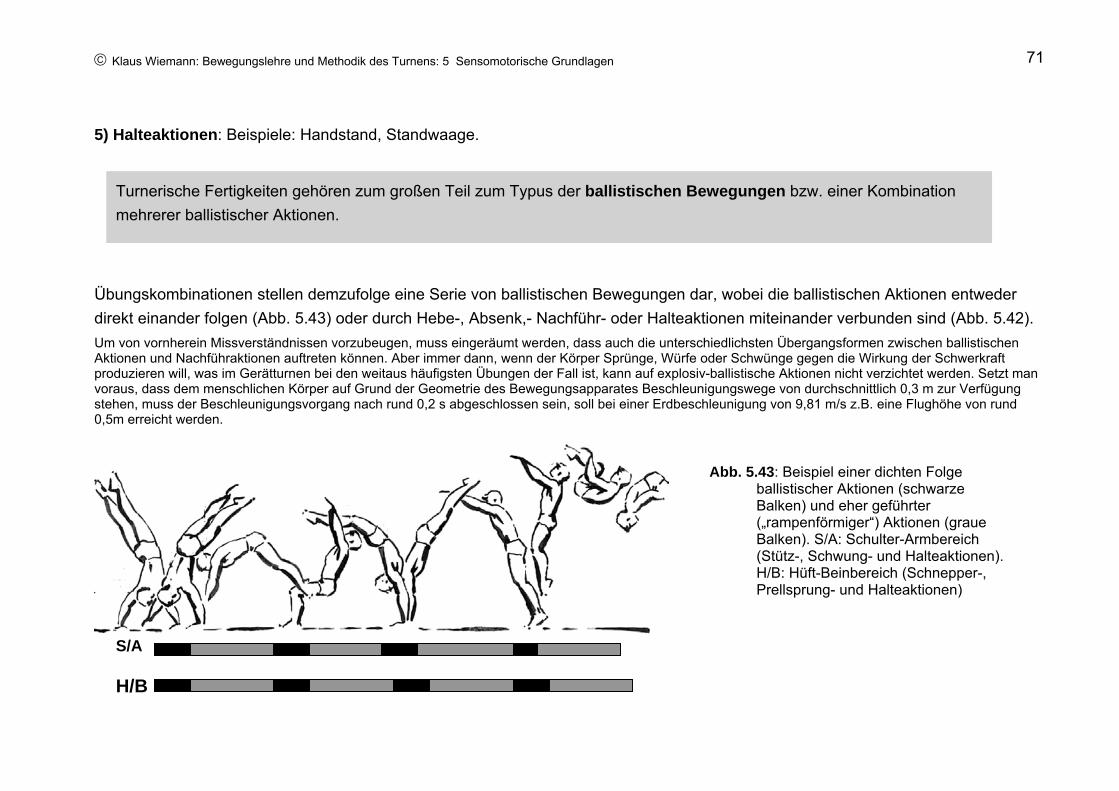
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 71
5) Halteaktionen: Beispiele: Handstand, Standwaage.
Übungskombinationen stellen demzufolge eine Serie von ballistischen Bewegungen dar, wobei die ballistischen Aktionen entweder direkt einander folgen (Abb. 5.43) oder durch Hebe-, Absenk,- Nachführ- oder Halteaktionen miteinander verbunden sind (Abb. 5.42). Um von vornherein Missverständnissen vorzubeugen, muss eingeräumt werden, dass auch die unterschiedlichsten Übergangsformen zwischen ballistischen Aktionen und Nachführaktionen auftreten können. Aber immer dann, wenn der Körper Sprünge, Würfe oder Schwünge gegen die Wirkung der Schwerkraft produzieren will, was im Gerätturnen bei den weitaus häufigsten Übungen der Fall ist, kann auf explosiv-ballistische Aktionen nicht verzichtet werden. Setzt man voraus, dass dem menschlichen Körper auf Grund der Geometrie des Bewegungsapparates Beschleunigungswege von durchschnittlich 0,3 m zur Verfügung stehen, muss der Beschleunigungsvorgang nach rund 0,2 s abgeschlossen sein, soll bei einer Erdbeschleunigung von 9,81 m/s z.B. eine Flughöhe von rund 0,5m erreicht werden.
Turnerische Fertigkeiten gehören zum großen Teil zum Typus der ballistischen Bewegungen bzw. einer Kombination mehrerer ballistischer Aktionen.
S/A
H/B
Abb. 5.43: Beispiel einer dichten Folge ballistischer Aktionen (schwarze Balken) und eher geführter („rampenförmiger“) Aktionen (graue Balken). S/A: Schulter-Armbereich (Stütz-, Schwung- und Halteaktionen). H/B: Hüft-Beinbereich (Schnepper-, Prellsprung- und Halteaktionen)
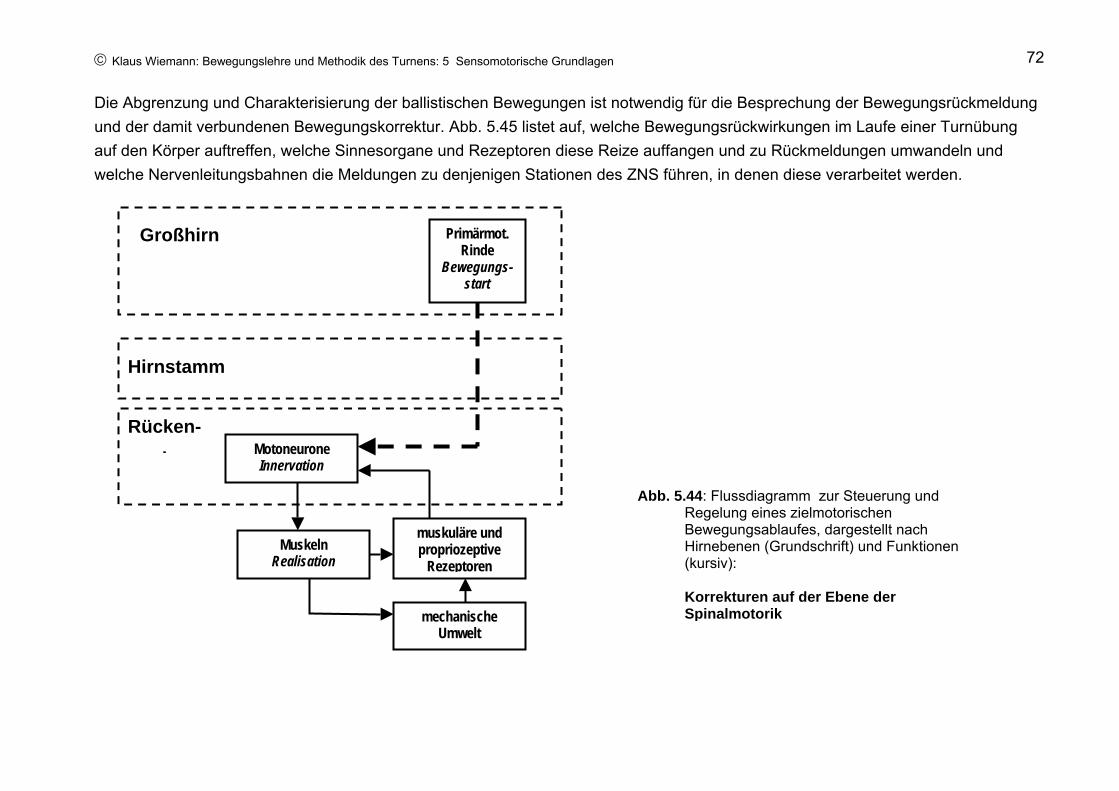
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 72
Die Abgrenzung und Charakterisierung der ballistischen Bewegungen ist notwendig für die Besprechung der Bewegungsrückmeldung und der damit verbundenen Bewegungskorrektur. Abb. 5.45 listet auf, welche Bewegungsrückwirkungen im Laufe einer Turnübung auf den Körper auftreffen, welche Sinnesorgane und Rezeptoren diese Reize auffangen und zu Rückmeldungen umwandeln und welche Nervenleitungsbahnen die Meldungen zu denjenigen Stationen des ZNS führen, in denen diese verarbeitet werden.
Motoneurone Innervation
Muskeln Realisation
Rücken-k
muskuläre und propriozeptive
Rezeptoren
mechanische Umwelt
Primärmot. Rinde
Bewegungs-start
Hirnstamm
Großhirn
Abb. 5.44: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Korrekturen auf der Ebene der Spinalmotorik
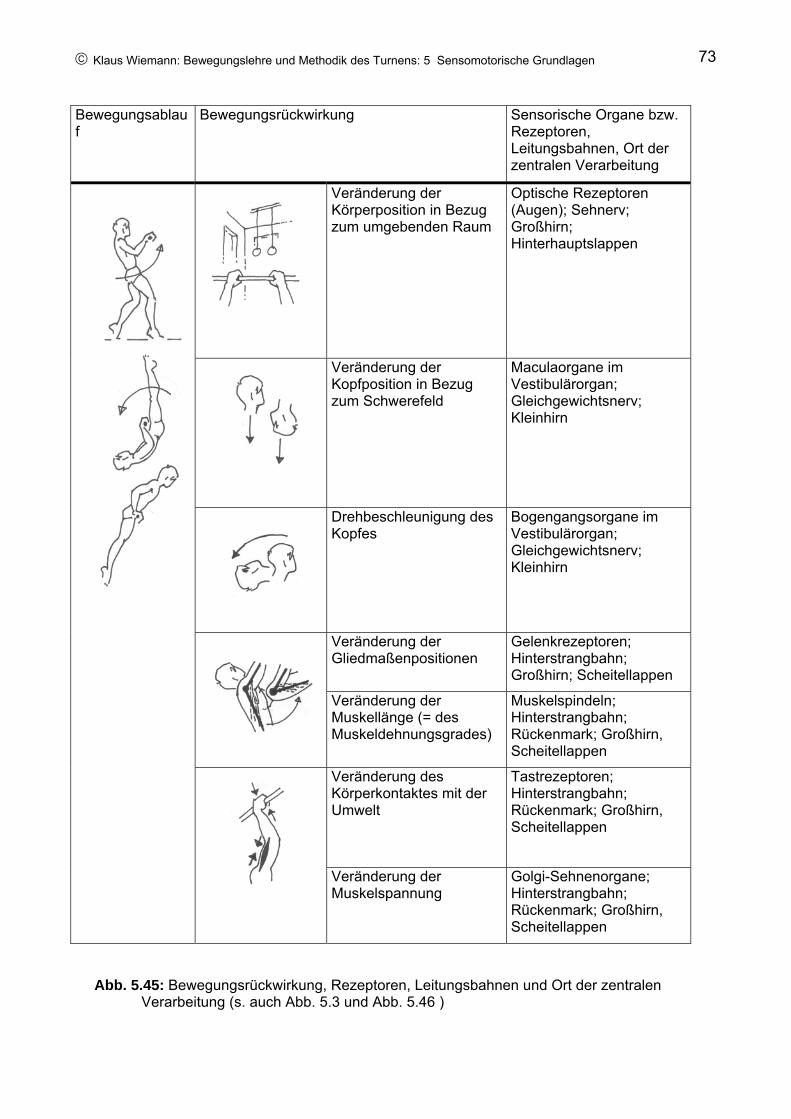
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 73
Bewegungsablauf
Bewegungsrückwirkung Sensorische Organe bzw. Rezeptoren, Leitungsbahnen, Ort der zentralen Verarbeitung
Veränderung der Körperposition in Bezug zum umgebenden Raum
Optische Rezeptoren (Augen); Sehnerv; Großhirn; Hinterhauptslappen
Veränderung der Kopfposition in Bezug zum Schwerefeld
Maculaorgane im Vestibulärorgan; Gleichgewichtsnerv; Kleinhirn
Drehbeschleunigung des Kopfes
Bogengangsorgane im Vestibulärorgan; Gleichgewichtsnerv; Kleinhirn
Veränderung der Gliedmaßenpositionen
Gelenkrezeptoren; Hinterstrangbahn; Großhirn; Scheitellappen
Veränderung der Muskellänge (= des Muskeldehnungsgrades)
Muskelspindeln; Hinterstrangbahn; Rückenmark; Großhirn, Scheitellappen
Veränderung des Körperkontaktes mit der Umwelt
Tastrezeptoren; Hinterstrangbahn; Rückenmark; Großhirn, Scheitellappen
Veränderung der Muskelspannung
Golgi-Sehnenorgane; Hinterstrangbahn; Rückenmark; Großhirn, Scheitellappen
Abb. 5.45: Bewegungsrückwirkung, Rezeptoren, Leitungsbahnen und Ort der zentralen Verarbeitung (s. auch Abb. 5.3 und Abb. 5.46 )

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 74
Struktur neuronale Funktion und Zeitbedarf
begleitende bewusste
Phänomene Propriozeptoren und Tastrezeptoren (P+T), Rückenmark (Rm), Vestibular apparat (Va), Augen, Hirnstamm
Aufnahme und Weiterleitung der bewegungsbedingten Reize, 20 - 60 ms nach Bewegungsbeginn; reflektorische Korrekturen
Hinterstrangbahn; Hinterstrangkerne (Hsk); Schleifenbahn; Thalamus (Tha)
Leitung der Rückmeldungen zum Thalamus; Selektion der Meldungen und Kontrastschärfung; ab 30 ms nach Bewegungsbeginn
Thalamus (Tha); primäres somato-sensorisches Feld (SI); primäres motorisches Feld (MI)
Leitung direkt zu MI oder über SI zu MI (50 ms nach Bewegungsbeginn): grobe Kurzkorrektur länger anhaltender Bewegungen (>100 ms: „geführter“ Bewegungen) möglich
Empfindungen sensorischer Natur (jedoch kein bewusster Zugriff zur Bewegungskorrektur)
Hinterstrangkerne (Hsk); Kleinhirn (Klh); Thalamus; primäres motorisches Feld (MI)
Korrektur von Intensität und Timing langsamer Bewegungen oder zyklischer Aktionen (nicht früher als 100 ms nach Bewegungsbeginn); Korrektur und Speicherung der Programme
Sensorische Felder; (SI, SII/III supplementäres motorisches Feld (SMA)
Verarbeitung, Verknüpfung der Meldungen und Rückmeldungen, Verrechnung mit gespeicherten Informationen; Vergleich mit Bewegungsentwurf; Bewegungskorrektur, falls Bewegung noch nicht abgeschlossen ist; ab 150 ms nach Bewegungsstart
Wahrnehmen der Bewegung; bei schnellen, kurz dauernden Aktionen nur Wahrnehmung des Bewegungsresultats; Vergleich mit der Bewegungsvorstellung (Bewegungsabsicht); bewusste Korrektur
Assoziationsfelder (ASS); Limbisches System (LS)
Bewertung und Speicherung der Meldungen
Emotionale Bewertung der Bewegung und des Resultats
Abb. 5.46: Tabellarische Übersicht über die Stufen der Bewegungsrückmeldung und deren zentraler Verarbeitung
SI
MI
MI SI
Tha
Tha
Klh
Tha
Hsk
Hsk
ASS
LS
SMA SII/III SI
P+T Va
Rm

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 75
Abb. 5.46 verdeutlicht schematisch den Verlauf der Nervenleitungsbahnen zu den verarbeitenden Hirnstationen und gibt vor allem den Bedarf an Zeit wieder, der für die diversen Nerven- und Hirnfunktionen anfällt und der für die einzelnen Korrekturmaßnahmen von Bedeutung sind. Diese Korrekturmaßnahmen sollen in folgende Stufen eingeteilt werden:
1) reflektorische Korrekturen auf dem Niveau des Rückenmarkes, 2) stützmotorische Korrekturen auf der Kleinhirnebene, 3) willkürliche, durch das Großhirn gesteuerte Korrekturen und 4) Korrektur des Bewegungsentwurfes.
Zu 1) reflektorische Korrekturen: Frühestens 30 ms nach Bewegungsbeginn können die Dehnungsreflexe und Spannungsreflexe (s. Kap. 5.21) korrigierend in den Bewegungsablauf eingreifen, indem sie explosiv-ballistische Kontraktionen wie z.B. ein prellendes Abspringen oder ein Abstützen mit den Händen unterstützen (Abb. 5.44). Ob diese Unterstützung gelingt, hängt allerdings von verschiedenen Faktoren ab, die in Kap. 5.21 diskutiert wurden.
Anders verhält sich die dehnungsbedingte Regelung von länger dauernden Nachführaktionen. Hier kann der Organismus mit Hilfe des Längen-Kontrollmechanismus durch ständiges Überwachen der Muskellänge Abweichungen vom Sollwert, der supraspinal vorgegeben wird, beseitigen. Als Rezeptoren für diese Abweichungen kommen die Muskelspindeln in Frage (Kap. 5.21 und 5.22). Die Gelenkrezeptoren, die als Messfühler für die Gelenkstellung und Gelenkbewegung dienen, haben keinen Einfluss auf die reflektorische Kontrolle von Halte- und Nachführbewegungen.
Zur reflektorischen Korrektur explosiv-ballistischer Aktionen können auch die Spannungsreflexe (Kap. 5.23 und 5.24) beitragen, indem sie in einem Zeitrahmen von 30-60 ms nach Bewegungsbeginn zu hohe Muskelspannung reduzieren. Sie liefern damit eher einen Schutz des Organismus vor Überlastungen, als eine Bewegungskorrektur. Zusätzlich kann angenommen werden, dass auf dem Wege über den Spannungs-Servomechanismus Muskelspannungen geregelt werden, indem bei einem unvorhergesehenen Spannungsanstieg im Agonisten nicht nur dieser kurzfristig deaktiviert und dadurch die Spannung im Agonisten gemildert, sondern durch die zusätzliche Kontraktion des Antagonisten die auf den Agonisten wirkende äußere Spannung zusätzlich reduziert wird.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 76
Primärmot. Rinde
Bewegungs-start
Neukleinhirn Programmierung Erwerbmotorik
Altkleinhirn
stato-vestib. Regulation
sensorischer Thalamus
motorischer Thalamus
Motoneurone Innervation
Muskeln Realisation
Hirnstam
Großhirn
Rücken-
sensorische Rinde
Bewegungs- wahrnehmun
g
Hinterstang- kerne
SinnesorganeRezeption
muskuläre und propriozeptive
Rezeptoren
mechanische Umwelt
Abb. 5.47: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Korrekturen auf der Ebene der Stützmotorik. Grau unterlegt: bewusstseinspflichtige Funktionen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 77
Als letzte Möglichkeit der reflektorischen Bewegungskorrektur auf dem Niveau des Rückenmarkes müssen die Beuge- und Streckreflexe dienen (Kap. 5.25). Diese dienen jedoch vornehmlich der Sicherung der aufrechten Körperposition beim Stehen, Gehen und Laufen.
Zu 2) stützmotorische Korrekturen (Abb. 5.48): Da die Rückmeldungen zur stützmotorischen Kontrolle und Korrektur von motorischen Aktionen aus verschiedenen Körperbereichen und Sinnesorganen (Tast- und Gelenkrezeptoren, Augenmuskelrezeptoren, Gleichgewichts- und Drehbeschleunigungssinnesorgan) stammen, über unterschiedliche Nervenbahnen und diversen Schaltzentralen (Hinterstrangkerne, s. Abb. 5.46, Kleinhirnkerne) zum Kleinhirn gesendet und dort frühesten 100 ms nach Bewegungsbeginn verarbeitet werden, ist es verständlich, dass die äußeren Reaktionen erst nach mehreren 100 ms wirksam werden. Damit wird deutlich, dass sie im Rahmen ballistischer Aktionen keine bedeutende Rolle spielen können, sondern ihre vorrangige Bedeutung in der Kontrolle von Haltungen, speziell Körperhaltungen, im Laufe von länger dauernden Nachführbewegungen besitzen. Im Rahmen ballistischen Aktionen muss die Körperhaltung schon vor Beginn der Bewegung im Zuge der Bildung des Bewegungsentwurfes eingeplant sein.
Da die Stützmotorik sowie ihre Bedeutung für das Gerätturnen in Kap. 5.3 ausführlich behandelt wurde, braucht an dieser Stelle nicht weiter darauf eingegangen zu werden.
Zu 3) willkürliche, durch das Großhirn gesteuerte Korrekturen:
Voraussetzung für eine willkürliche Korrektur von Bewegungen ist das Aufsteigen der Rückmeldungen bis zur Großhirnrinde, mindestens zu den primären sensorischen Zentren (SI) und weiter zu den sekundären und tertiären sensorischer Zentren (SII/III) und zum somatosensorischen Areal (SMA; s. Abb. 5.46). Berücksichtigt man einerseits, dass vom Beginn der Bewegungsrückwirkungen bis zur Verarbeitung der Rückmeldung an der Hirnrinde mindestens 150 ms vergehen, andererseits zur Bildung eines neuen Bewegungsentwurfes zur willkürlichen Korrektur der Bewegung sowie zur Programmierung und zum Start der Bewegung noch mal mehr als 120 ms benötigt werden, wird verständlich, dass sich ballistische Bewegung mit einer Ablaufdauer von wenigen 100 ms willkürlich nicht korrigieren lassen; denn die motorischen Befehle zur Durchführung der Bewegungskorrektur würden frühestens 270 ms nach Bewegungsstart bei den Muskeln ankommen, nach einer Zeitspanne also, nach der explosiv-ballistische Aktionen häufig schon abgeschlossen sind.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 78
Auch wenn der Turner zu Beginn der ballistischen Aktion erkennt, dass sie nicht korrekt geplant war oder nicht in der vorgesehenen Form abzulaufen droht, wird er in der Regel an deren Ablauf selbst nichts mehr ändern können. Allenfalls kann er in der Lage sein, darauf im Hinblick auf die anschließenden Aktionen zu reagieren. Allerdings müssen dazu einige Bedingungen berücksichtigt werden, die an die Funktionsmöglichkeiten der motorischen Hirnrinde, speziell des SMA (s. Kap. 5.4.3), gekoppelt sind, z.B.:
-- Wie rechtzeitig wird der Großhirnrinde die Abweichung mitgeteilt?
-- Mit welcher Funktion sind die motorischen Areal der Großhirnrinde beim Einlaufen der Fehlermeldung beschäftigt?
-- Wie viel Zeit wird beansprucht, eine motorische Aktion zum Abfangen der Störung zu entwerfen?
-- Steht evtl. ein „vorgefertigter“ Bewegungsentwurf zum Abfangen der Störung parat?
Wie sich die Reaktion durch „vorgefertigte“ Bewegungsentwürfe beeinflussen lässt, kann man an der Beobachtung erkennen, dass erfahrene Sprinter ihre Reaktionszeit unter 100 ms drücken können. Dies scheint nur möglich, weil in der Startposition das ZNS keine besonderen Kontrollfunktionen zu erfüllen hat und sich ganz auf das Bereithalten des Bewegungsentwurfes „Start“ konzentrieren kann.
Diese und ähnliche Fragen sind jedoch erst zufriedenstellend zu beantworten, wenn (wie im folgenden Kap. 6 vorgesehen) die Ebene der reinen Hirnfunktionen verlassen wird und die Wahrnehmungs- und Vorstellungsprozesse, die an die Funktion der Großhirnrinde gekoppelt sind, also die korrelierenden Phänomene des Bewusstseins, mit in die Betrachtung einbezogen werden. Danach werden auch weitergehende Fragestellungen zu beantworten sein, wie z.B.:
-- Woran liegt es bzw. kann es gelegen haben, wenn die Bewegung nicht so abläuft, wie im Bewegungsentwurf vorgesehen?
-- Woran es liegt, dass manchmal der Bewegungsentwurf falsch, der Übende aber - nach einer mehr oder weniger langen Übungsperiode - doch zur korrekten Ausführung gelangt?
-- Ändert sich mit fortschreitendes Übung auch die Form und Komplexität des Bewegungsentwurfes?
Der Ablauf turnerischer Fertigkeiten vom Typus explosiv-ballistischer Bewegungen, der eine Ablaufzeitspanne von weniger als 300 ms benötigt, lässt sich willkürlich nicht korrigieren. Die Bewegung läuft in derjenigen Form ab, in der sie entworfen wurde. Jeder willkürliche Korrekturversuch „käme zu spät“.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 79
Limbisches System
Motivation
Assoziations-rinde
strategische Planung
SMA Bewegungsentwurf
Neukleinhirn Programmierung Erwerbmotorik
Muskeln Realisation
Hirnstamm
Großhirn
sensorische Rinde
Bewegungs- wahrnehmung
S I
Bewegungsablauf
zentrale Speicher
Sinnesorgane Rezeption
sensorischer Thalamus
motorischer Thalamus
Abb. 5.48: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Korrektur des Bewegungsentwurfes durch sensorische Rückmeldungen. Grau unterlegt: bewusstseinspflichtige Funktionen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 80
Zu 4) Korrektur des Bewegungsentwurfes
Erst nach der Besprechung des Themas „Bewegungswahrnehmung und Bewegungsvorstellung“ sind die wichtigsten Grundlagen zur Bearbeitung der lehrmethodischen Maßnahmen gelegt.
Limbisches System
Assoziations-rinde
strategische Planung
SMA Bewegungsentwurf
Neukleinhirn Programmierung Erwerbmotorik
Muskeln Realisation
Hirnstamm
Großhirn
Bewegungsablauf
Instruktionen
soziale Umwelt
zentrale Speicher
Abb. 5.49: Flussdiagramm zur Steuerung und Regelung eines zielmotorischen Bewegungsablaufes, dargestellt nach Hirnebenen (Grundschrift) und Funktionen (kursiv): Korrektur des Bewegungsentwurfes durch Lehrmaßnahmen. Grau unterlegt: bewusstseinspflichtige Funktionen

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 81
Eine erste Korrektur des Bewegungsentwurfes erfolgt seitens des Übenden durch den Vergleich des Bewegungsentwurfes mit den einlaufenden Rückmeldungen. Ergibt sich hier eine weitgehende Entsprechung, kann die Ausführung der Bewegung als gelungen gewertet werden. Sind jedoch Abweichungen festzustellen, muss dies für den Übende Anlass sein, seinen Bewegungsentwurf zu korrigieren (Abb. 5.48). Im Hinblick auf eine Regelung tierischer Bewegungsabläufe mit einem theoretisch geforderten Istwert-Sollwert-Vergleich wurde von HOLST / MITTELSTAEDT (1950) das Reafferenzprinzip entwickelt. Dieses hypothetisierte die Bildung einer Kopie der Efferenz, also einer Kopie der Gesamtheit der Innervationsmuster an die Muskulatur, und deren Speicherung in niederen Hirnabschnitten. Mit dieser Efferenzkopie sollte die nach der Realisation einlaufende Reafferenz, also die Gesamtheit der sensorischen Rückmeldungen, verglichen und Abweichungen zur Nachbesserung der Efferenz benutzt werden – ohne Beteiligung der höchsten Hirninstanzen. Dieses Reafferenzprinzip wurde auch auf die Steuerung der menschlichen Motorik angewendet und auch heute noch werden gelegentlich Begriffe wie Efferenzkopie und Reafferenz in der sportwissenschaftlichen Bewegungslehre benutzt. Spätestens seit den Forschungen von Eccles (1987). Kornhuber (1975) und anderen ist deutlich, dass für die Bildung einer solchen Efferenzkopie für zielmotorische Abläufe allenfalls die primärmotorische Großhirnrinde in Frage käme, die jedoch nur in sehr beschränktem Maße auf dem Weg über das primäre somatosensoriche Feld (SI) in eine Bewegungskorrektur eingreifen könnte (Abb. 5.47). Somit bleibt für die vorliegende Fragestellung nur, eine durch das bewusstseingesteuerte Korrektur der Bewegungsentwürfe (= Bewegungsvorstellung, Kap. 6.2) anzunehmen.
Mit der Korrektur des Bewegungsentwurfes (Abb. 5.49) durch Lehrmaßnahmen wird die Ebene der sensormotorischen Erklärungen verlassen und auf das Kapitel der Grundlagen der methodischen Maßnahmen (Kap. 7) verwiesen.
5.5. Biologische Grundlagen des motorischen Lernens
5.5.1 Neuronale Ketten, Kreise und Netze
Lernen allgemein und motorisches Lernen im Besonderen vollzieht sich im Zentralnervensystem (ZNS). Dieses besteht aus einer Ansammlung von vielen Milliarden Nerverzellen (Neuronen; Abb. 5.50a), den Funktionseinheiten des ZNS, die in (neuronalen) Ketten, Kreisen oder Netzen (Abb. 5.50 b, c und d) miteinander verknüpft sind. Jede einzelne Nervenzelle besteht in der Regel aus einem Zellleib mit mehreren kürzeren verzweigten Fortsätzen (Dendriten) und meist einem längeren Fortsatz (Neurit, Axon), der am Ende eine Aufzweigung besitzt. An jedem Ende dieser Axonaufzweigungen befindet sich je ein Endknöpfchen, das mit der Membran einer nachfolgenden Nervenzelle (am Zellleib oder an den Dendriten) Kontakt aufnimmt und die sogenannte Synapse bildet. Die Synapsen sind diejenigen Stellen im Verbund der Nervenzellen, an denen die Information von einer Nervenzelle an die nächste weitergegeben

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 82
wird. Dies geschieht jedoch stets nur in einer Richtung, nämlich vom synaptischen Endknöpfchen hin zur Membran der nachgeschalteten Nervenzelle. Auf die Erregungsbildung von Nervenzellen, auf die Entstehung fortgeleiteter Aktionspotentiale sowie auf die synaptische Übertragung von Informationen kann hier nicht im Einzelnen eingegangen werden. Es wird auf die physiologische Literatur verwiesen.
Dendrit
Zellkörper mitZellkern
Endverzweigung mit Endknöpfchen
Neurit
Synapse a)
d)
c)
b)
Abb. 5.50: Grundstruktur einer Nervenzelle (a). Neuronale Kette (b). Neuronaler Kreis (c). Neuronales Netz (d). Pfeile: Richtung der Informationsübertragung.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 83
Im ZNS gibt es sowohl erregende Nervenzellen, die die Weitergabe von Informationen an den Synapsen fördern („bahnen“, facilisieren), als auch hemmende Nervenzellen, die die Weitergabe der Informationen drosseln oder völlig unterdrücken (inhibitieren). Durch das Zusammenspiel erregender und hemmender Nervenzellen wird gewährleistet, dass ausgewählte Informationen gezielt weitergeleitet werden, und verhindert, dass sich ein Nervenimpuls ungehindert im gesamten Nervensystem ausbreitet.
Diese Veränderungen, die sowohl erregende als auch hemmende Synapsen betreffen, können sein: - Freisetzung von Nervenüberträgerstoff (Neurotransmitter in den synaptischen Spalt, - erhöhte Freisetzung von Nervenüberträgerstoff (Neurotransmitter) in den synaptischen Spalt, - verlängerte Verweildauer des Neurotransmitters im synaptischen Spalt, - Einbau von Proteinen in die Membranen der Synapse zur Förderung des Ionentransportes und der Transmitterfreisetzung, - Vermehrung der Anzahl der synaptischen Kontakte. (z.B. SINZ, 1977; KANDEL u.a. 1996).
Sobald eine über längere Zeit bestehen bleibende Bahnung hergestellt ist, spricht man von der Bildung einer neuen zentralnervösen Verknüpfung, die als physische Grundlage von Lernvorgängen angesehen werden muss. Eine irreversible Verknüpfung (Konsolidierung) kann sich im Extremfall schon bilden, wenn nur eine einzige Impulsserie eine Neuronenkette durchläuft (z. B. im Fall der Prägung), in der Regel wird jedoch erst eine mehr oder weniger häufige Wiederholung der gleichen Impulsserie zu einer bleibenden Verknüpfung führen, wobei sich eine Verknüpfung um so leichter und schneller herstellt, je intensiver Synapsen emotionaler Systeme (limbisches System) mitaktiviert sind. Letzteres ist aber gerade immer auch dann der Fall, wenn die übermittelte Information einen hohen Neuigkeitswert besitzt (KUGLER 1981, KANDEL u.a. 1996).
Jedes Mal, wenn an einer Synapse eine Information an die nachgeschaltete Nervenzelle weitergegeben wird, vollzieht sich an der Synapse eine kurzfristige (wenige Millisekunden bis Sekunden dauernde), mittelfristige (mehrere Stunden oder Tage anhaltende) oder bleibende Veränderung. Die Wahrscheinlichkeit einer solchen Veränderung steigt bei Wiederholung des Vorganges unter gleichen Bedingungen.

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 84
Diese Veränderungen sind die Grundlage für die Veränderungen des Verhaltens allgemein sowie des Lernens und des motorischen Lernens im Besonderen. Ein bedeutender Einfluss kommt in diesem Zusammenhang den Motivationssystemen des ZNS, insbesondere dem Limbischen System, zu. Diese können durch spezielle Nervenbahnen und durch synaptische Kontaktaufnahme (Abb. 5.51) mit den Nerven der sensorischen und motorischen Systeme die Synapsenfunktion derart beeinflussen, dass die Informationsübertragung verstärkt (gebahnt) oder gehemmt wird und die bleibenden Veränderungen an den Synapsen beschleunigt und vertieft werden. Letzteres ist aber gerade immer auch dann der Fall, wenn die übermittelte Information einen hohen Neuigkeitswert besitzt.
Durch das Eingreifen der Motivationssysteme in die Nervenübertragung werden der Einbau bleibender Veränderungen an den Synapsen und somit das Lernen allgemein und das motorische Lernen im Besonderen beschleunigt.
Zms
Zzv Zzv
EKzv
EKms Abb. 5.51: Beispiel einer
Einflussnahme einer Nervenzelle eines Motivationssystems (Zms) auf die Informationsübertragung zwischen Nervenzellen einer zentralen Verschaltung (Zzv). links: Im Überblick. Rechts: Ausschnitt der Synapsen; Ekms Endknöpfchen der motivationalen Nervenzelle; Ekzv Endknöpfchen der zentralen Verschaltung; Zzv Nervenzelle der zentralen Verschaltung. Schwarze Pfeile: Richtung der Informationsübermittlung
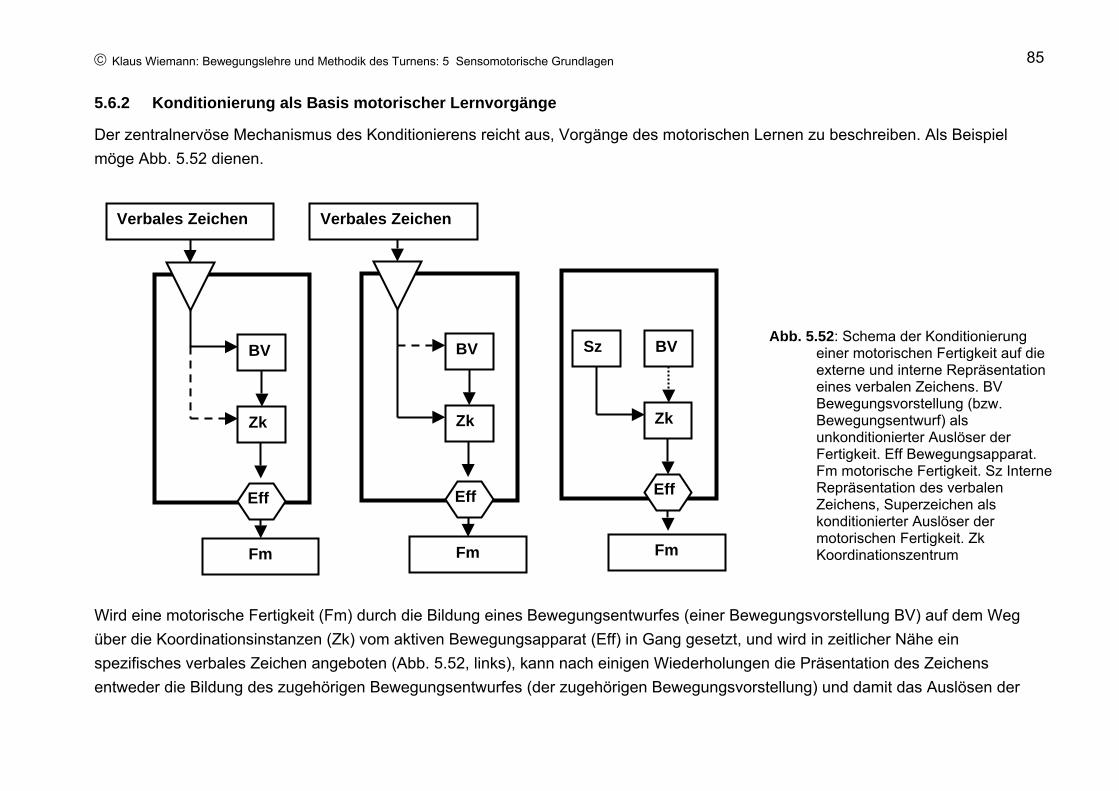
© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 85
5.6.2 Konditionierung als Basis motorischer Lernvorgänge
Der zentralnervöse Mechanismus des Konditionierens reicht aus, Vorgänge des motorischen Lernen zu beschreiben. Als Beispiel möge Abb. 5.52 dienen.
Wird eine motorische Fertigkeit (Fm) durch die Bildung eines Bewegungsentwurfes (einer Bewegungsvorstellung BV) auf dem Weg über die Koordinationsinstanzen (Zk) vom aktiven Bewegungsapparat (Eff) in Gang gesetzt, und wird in zeitlicher Nähe ein spezifisches verbales Zeichen angeboten (Abb. 5.52, links), kann nach einigen Wiederholungen die Präsentation des Zeichens entweder die Bildung des zugehörigen Bewegungsentwurfes (der zugehörigen Bewegungsvorstellung) und damit das Auslösen der
BV
Fm
Verbales Zeichen
Zk
Eff
BV
Zk
Eff
Fm
Verbales Zeichen
BV
Zk
Eff
Fm
Sz Abb. 5.52: Schema der Konditionierung einer motorischen Fertigkeit auf die externe und interne Repräsentation eines verbalen Zeichens. BV Bewegungsvorstellung (bzw. Bewegungsentwurf) als unkonditionierter Auslöser der Fertigkeit. Eff Bewegungsapparat. Fm motorische Fertigkeit. Sz Interne Repräsentation des verbalen Zeichens, Superzeichen als konditionierter Auslöser der motorischen Fertigkeit. Zk Koordinationszentrum

© Klaus Wiemann: Bewegungslehre und Methodik des Turnens: 5 Sensomotorische Grundlagen 86
Bewegung anregen oder aber direkt die Koordination der Bewegung anstoßen (Abb. 5.52, mitte). Das bedeutet, dass die Ausführung der Fertigkeit auf das verbale Zeichen konditioniert wird. Wird das Zeichen zentral gespeichert, kann es als Superzeichen (Sz) bei interner Repräsentation sowohl die Koordination der Bewegung in Gang setzen, gleichsam als Ersatz für den Bewegungsentwurf (die Bewegungsvorstellung; Abb. 5.52, rechts). Ebenso ist es möglich, dass die Interne Repräsentation des Superzeichens die Bildung der Bewegungsvorstellung auslöst, ohne die zugehörige Bewegung extern zu realisieren.
Das Schema von Abb. 5.52, rechts könnte auch ein vereinfachtes Modell der Beobachtung darstellen, dass es mit fortschreitender Übung zunehmend überflüssig wird, den kompletten Bewegungsentwurf (die kompletten Bewegungsvorstellung) zur angestrebten Fertigkeit zu bilden und es ausreicht, nur einen repräsentativen Teil (meistens die ersten Abschnitte) der gesamten Bewegungsvorstellung oder gar einen symbolischen Stellvertreter intern zu repräsentieren. In ähnlicher Weise kann die Koordination der geplanten Fertigkeit auf die Bewegungsrückmeldung vom Ablauf einer vorangehenden Fertigkeit konditioniert werden. Auf diese Weise ist das Erlernen längerer Verhaltensketten denkbar (s. auch Kap. 5.4.3 und Kap. 6.2).