Embed Size (px)
Citation preview
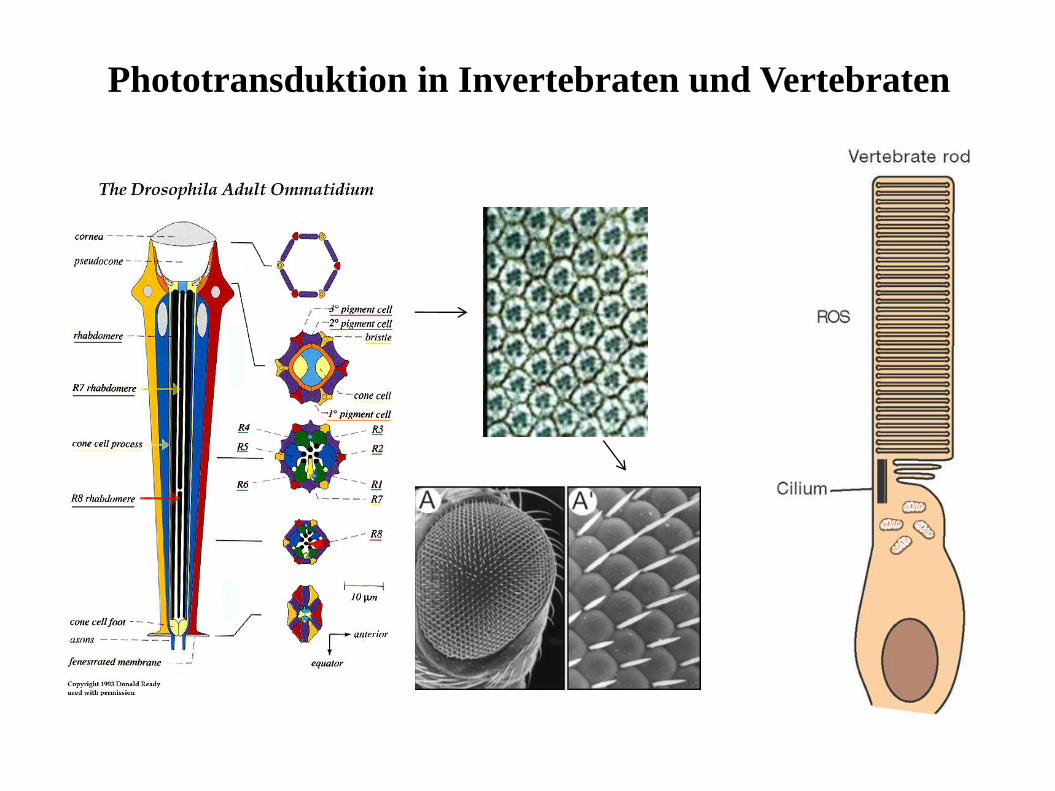
Phototransduktion in Invertebraten und Vertebraten

Einige wichtige Merkmale der Phototransduktionbei Invertebraten und Vertebraten
•Das Lichtsignal (Photon) wird von einem Rhodopsin Molekül
absorbiert und letztendlich in ein elektrisches Signal umgewandelt, das
von der Photorezeptorzelle gebildet wird, und über Interneurone an
das Gehirn weitergeleitet wird.
•Die Photorezeptoren sind in der Lage einzelne Photonen war
zunehmen
•Photorezeptoren können adaptieren—d.h. bei andauernder
Lichtreizung wird die Verstärkung der Signale reduziert

Phototransduction in Drosophila: Phototaxis and das Elektroretinogram
(ERG) als Mittel um die beteiligten Gene zu identifizieren.

Isolierung einer Vielzahl nicht-postiv-phototaktischer Drosophila Mutanten
.....gefolgt von der Analyse der Photorezeptorströme im Auge nach Lichtreizung
(Elektroretinogramm, oder ERG)

Was ist ein Elektroretinogramm (ERG)?
• Drosophila Rezeptorzellen depolarisieren auf Lichtreize,
wodurch Aktionspotentiale ausgelöst werden können.
•Das ERG misst die synchrone Antwort vieler Rezeptorzellen,
die als Antwort auf einen Lichtreiz gemeinsam depolarisieren
•Das ERG wird extrazellulär, mit einer Elektrode direkt unter
der Augenoberfläche gemessen

Das ERG set-up
Stark lab
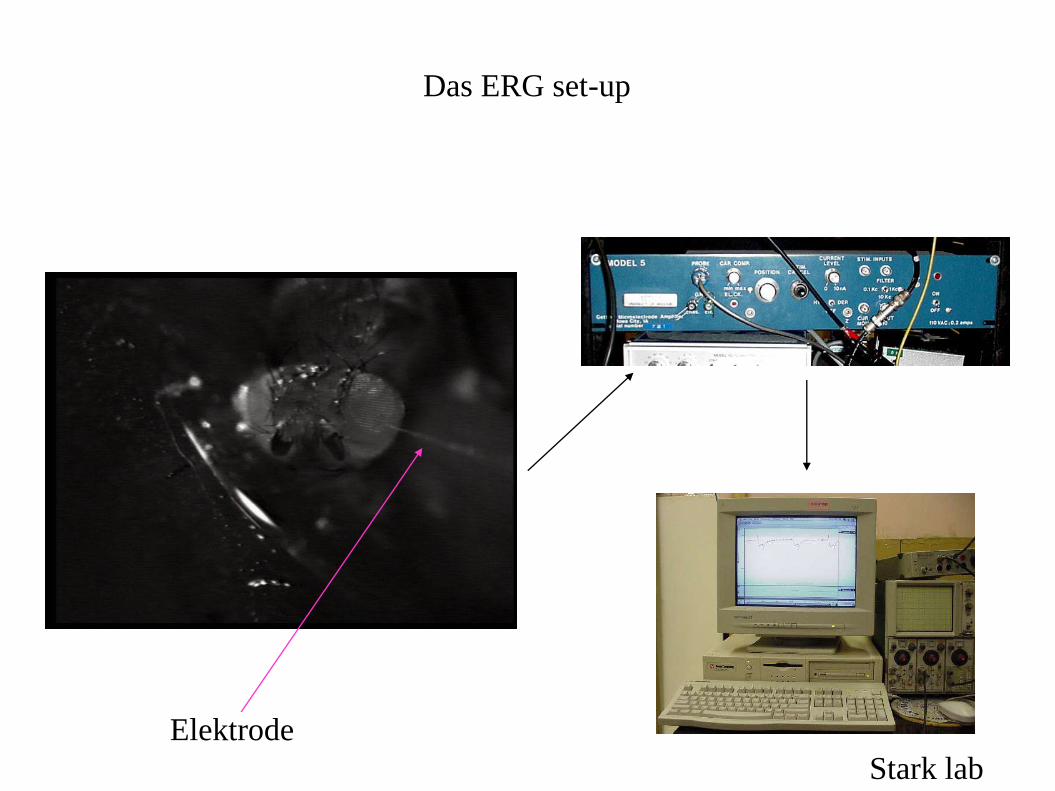
Elektrode
Stark lab
Das ERG set-up
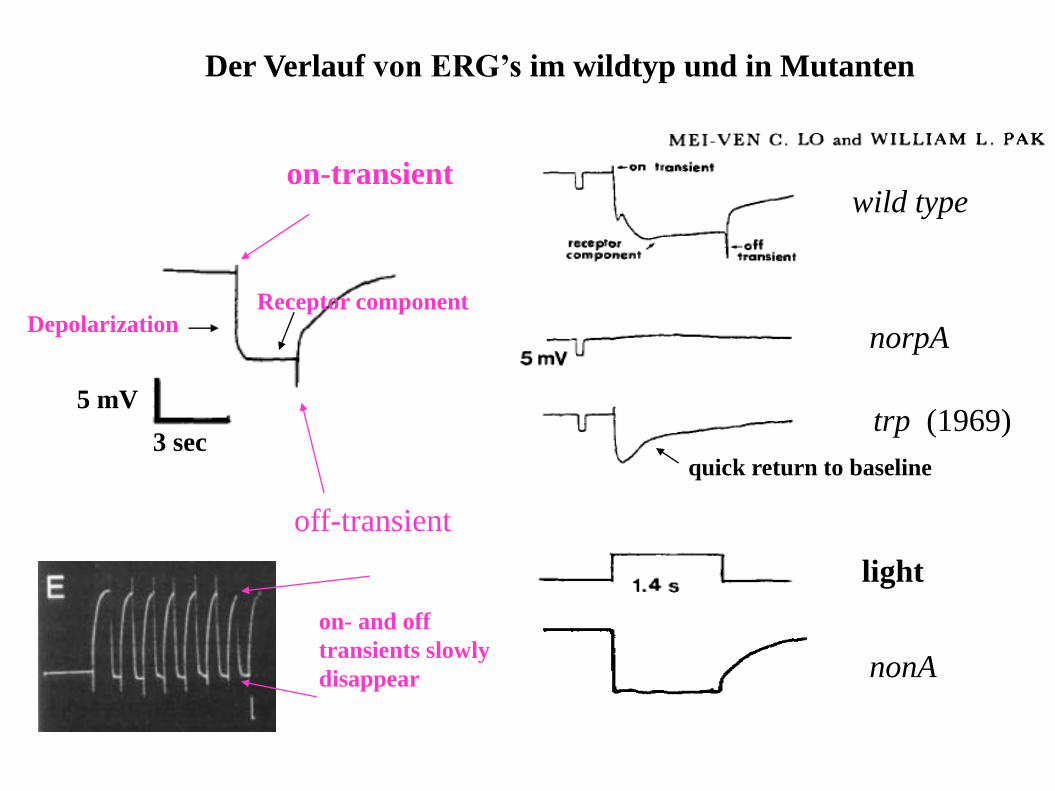
Der Verlauf von ERG’s im wildtyp und in Mutanten
3 sec
5 mV
on-transient
off-transient
Depolarization
on- and off
transients slowly
disappear
wild type
norpA
trp (1969)
light
nonA
quick return to baseline
Receptor component
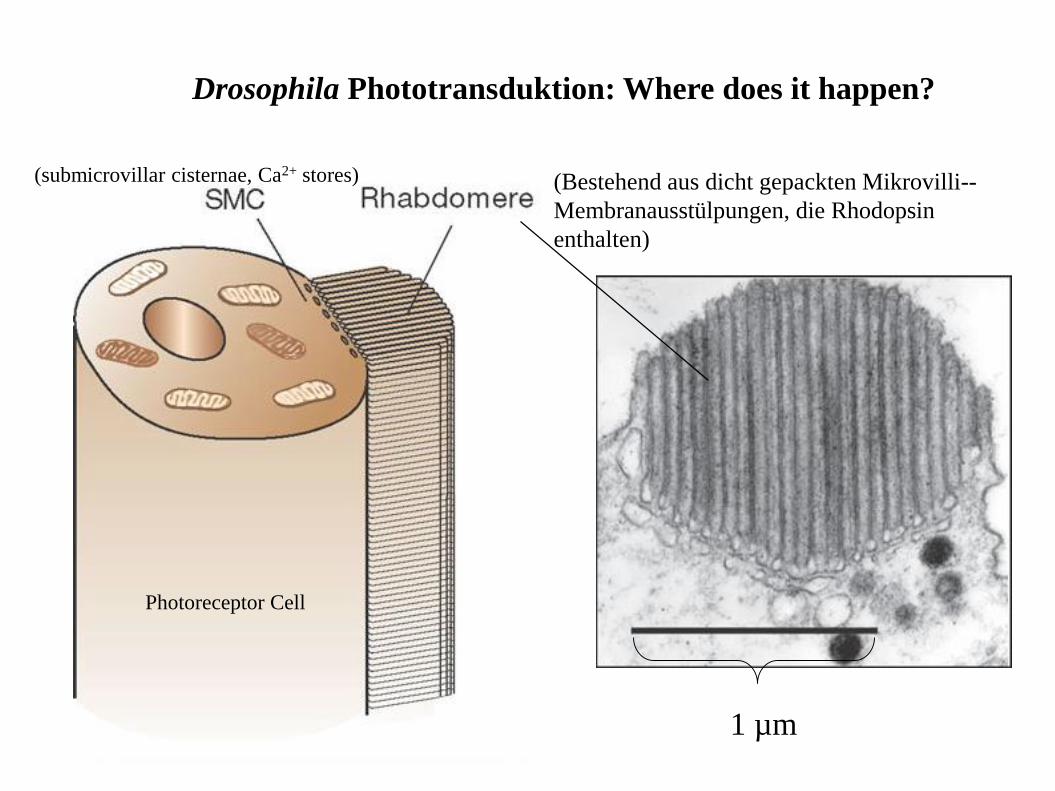
1 µm
(submicrovillar cisternae, Ca2+ stores) (Bestehend aus dicht gepackten Mikrovilli--
Membranausstülpungen, die Rhodopsin
enthalten)
Drosophila Phototransduktion: Where does it happen?
Photoreceptor Cell
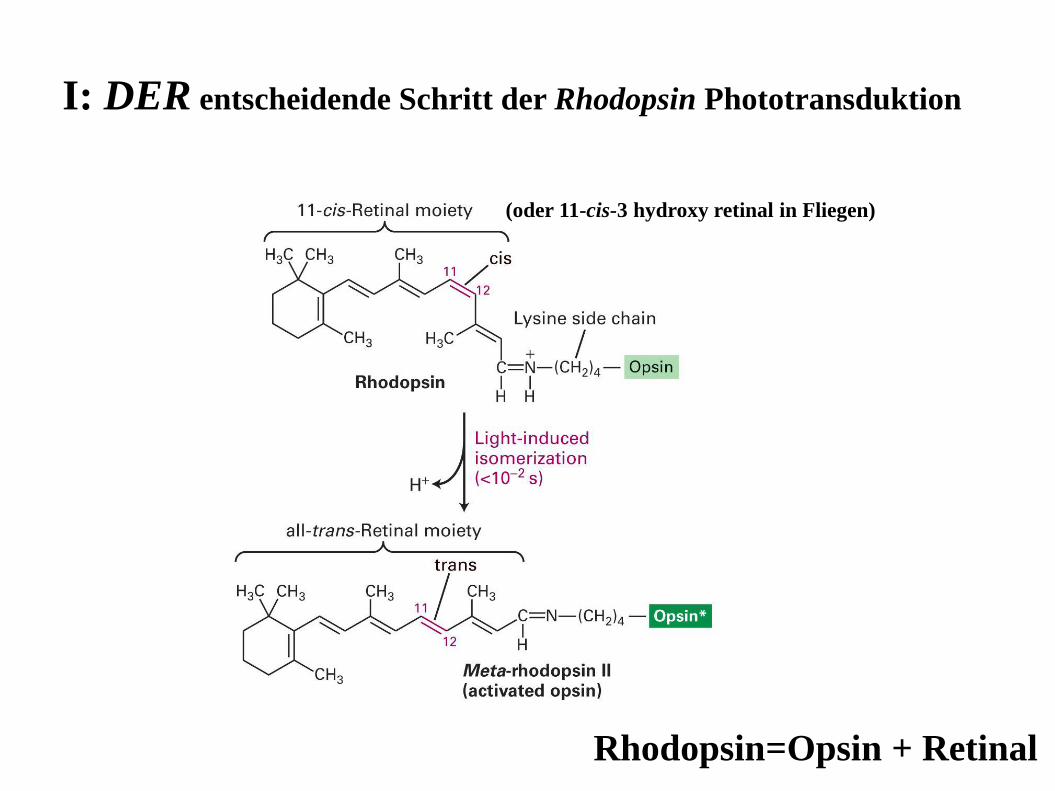
I: DER entscheidende Schritt der Rhodopsin Phototransduktion
(oder 11-cis-3 hydroxy retinal in Fliegen)
Rhodopsin=Opsin + Retinal

II: Rhodopsin* ist ein 7TM Rezeptor
‘serpentine’, oder 7TM Rezeptoren
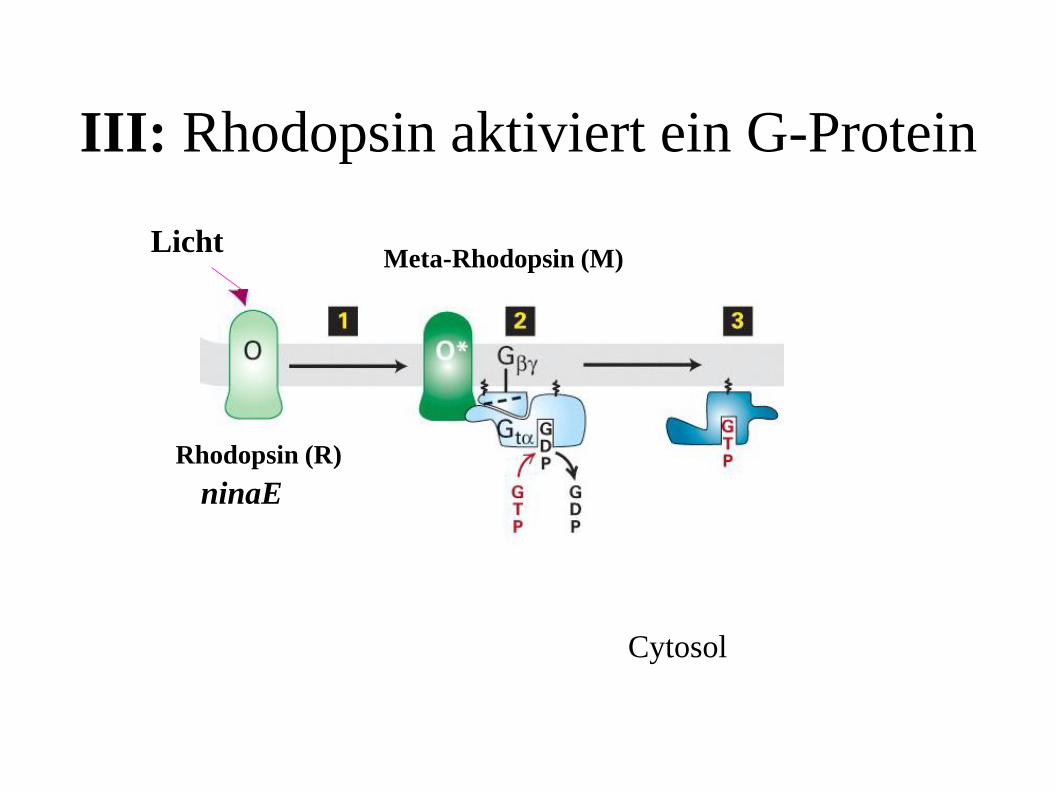
III: Rhodopsin aktiviert ein G-Protein
Licht
Rhodopsin (R)
Meta-Rhodopsin (M)
Cytosol
ninaE
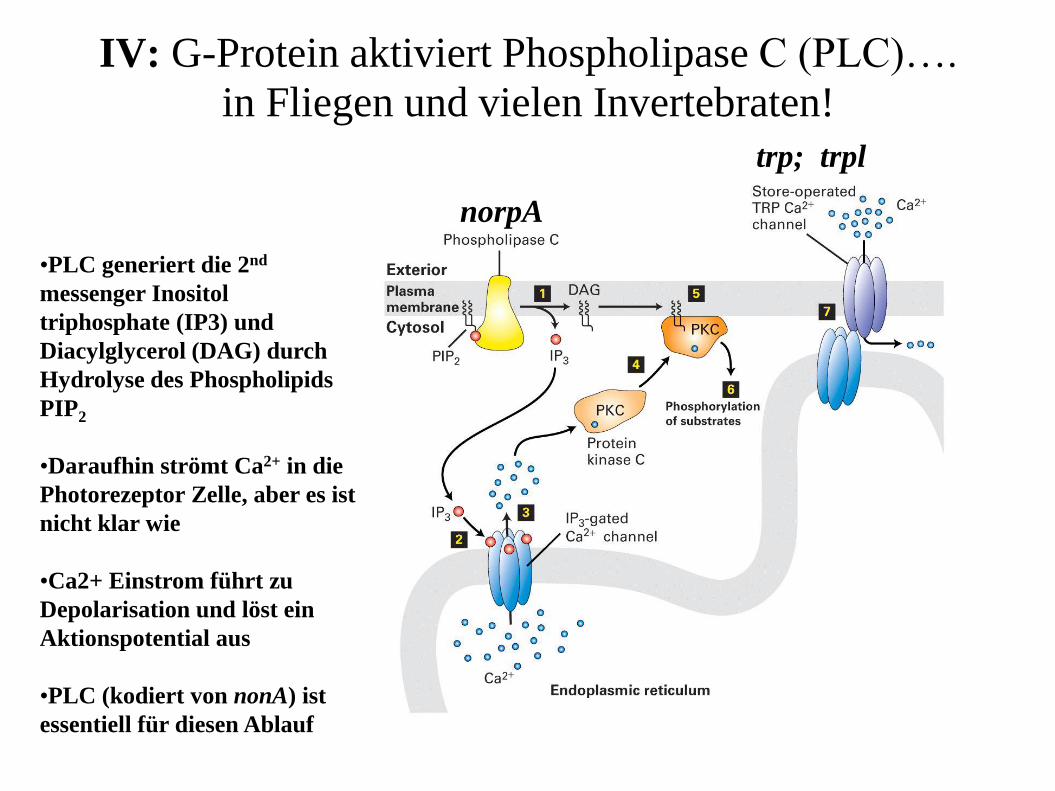
IV: G-Protein aktiviert Phospholipase C (PLC)….
in Fliegen und vielen Invertebraten!
•PLC generiert die 2nd
messenger Inositol
triphosphate (IP3) und
Diacylglycerol (DAG) durch
Hydrolyse des Phospholipids
PIP2
•Daraufhin strömt Ca2+ in die
Photorezeptor Zelle, aber es ist
nicht klar wie
•Ca2+ Einstrom führt zu
Depolarisation und löst ein
Aktionspotential aus
•PLC (kodiert von nonA) ist
essentiell für diesen Ablauf
norpA
trp; trpl

Das Ergebnis: Calcium Einstrom in eine Drosophila Photorezeptor Zelle
C. Zuker
Response to a 33 millisecond light-flash

Aber was genau bewirkt die Öffnung der
trp/trpl Kanäle???
•PLC ist essentiell
•PLC generiert IP3 (insositol triphosphat) und DAG (diacylglycerol)
aus PIP2 (phosphatidylinositol biphosphate )
•Sind IP3 und/oder DAG wichtig für die Öffnung der Kanäle?

Experimente mit IP3
Modell 1: IP3-induzierte Ca2+
Release aus internen Stores aktiviert
Trp durch einen ‘store operated’
Mechanismus
ABER:
•Experimentelle Freisetzung von caged
Ca2+ in Dunkel-adaptierten Zellen ruft
keine Aktivierung hervor
•IP3 Rezeptor Mutanten haben keinen
Effekt auf die normale
Phototransduction
Modell kommt nicht in Frage
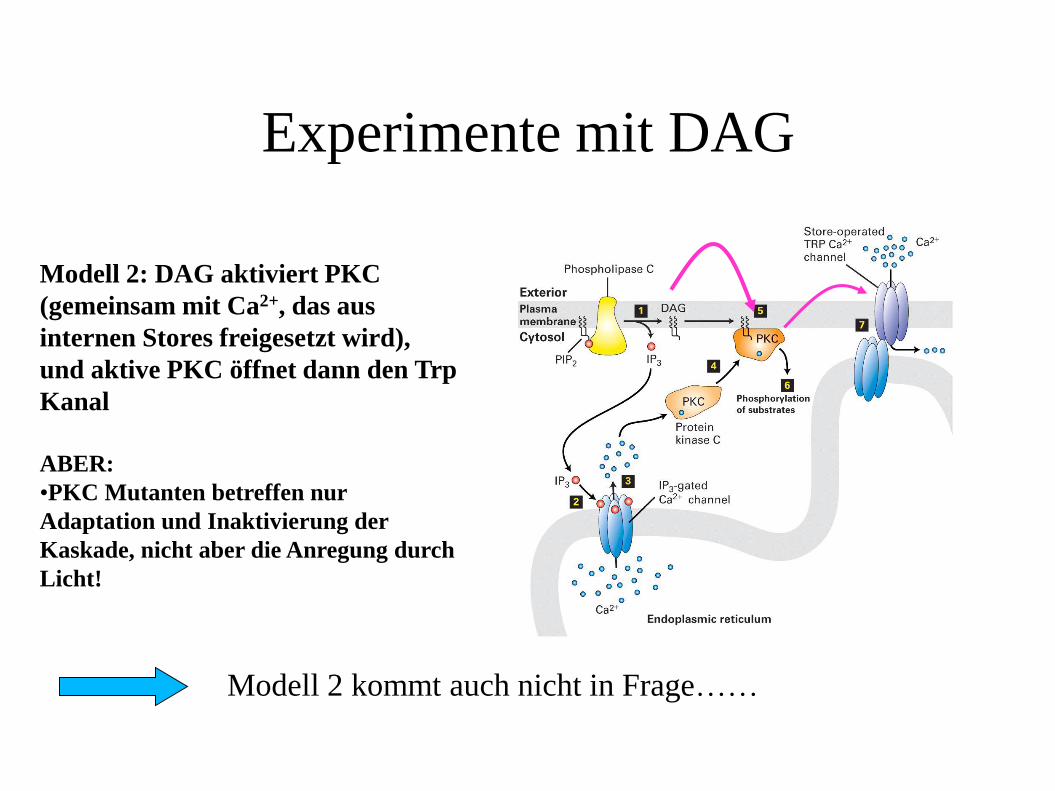
Experimente mit DAG
Modell 2: DAG aktiviert PKC
(gemeinsam mit Ca2+, das aus
internen Stores freigesetzt wird),
und aktive PKC öffnet dann den Trp
Kanal
ABER:
•PKC Mutanten betreffen nur
Adaptation und Inaktivierung der
Kaskade, nicht aber die Anregung durch
Licht!
Modell 2 kommt auch nicht in Frage……

Experimente mit DAG
DAG
DAG-LipasePoly unsaturated fatty acids (PUFAs)
z.B. arachidonic acid
TrpTrpl
DAG turnover ist wichtig für die Terminierung der Kaskade. Trp Kanäle sind
konstitutiv geöffnet in rdgA Mutanten, was zu Degradation der
Photorezeptorzellen führt (retinal degeneration)
DAG-Kinase
Phosphatid Säure
(Vorläufer von PIP2)
rdgA
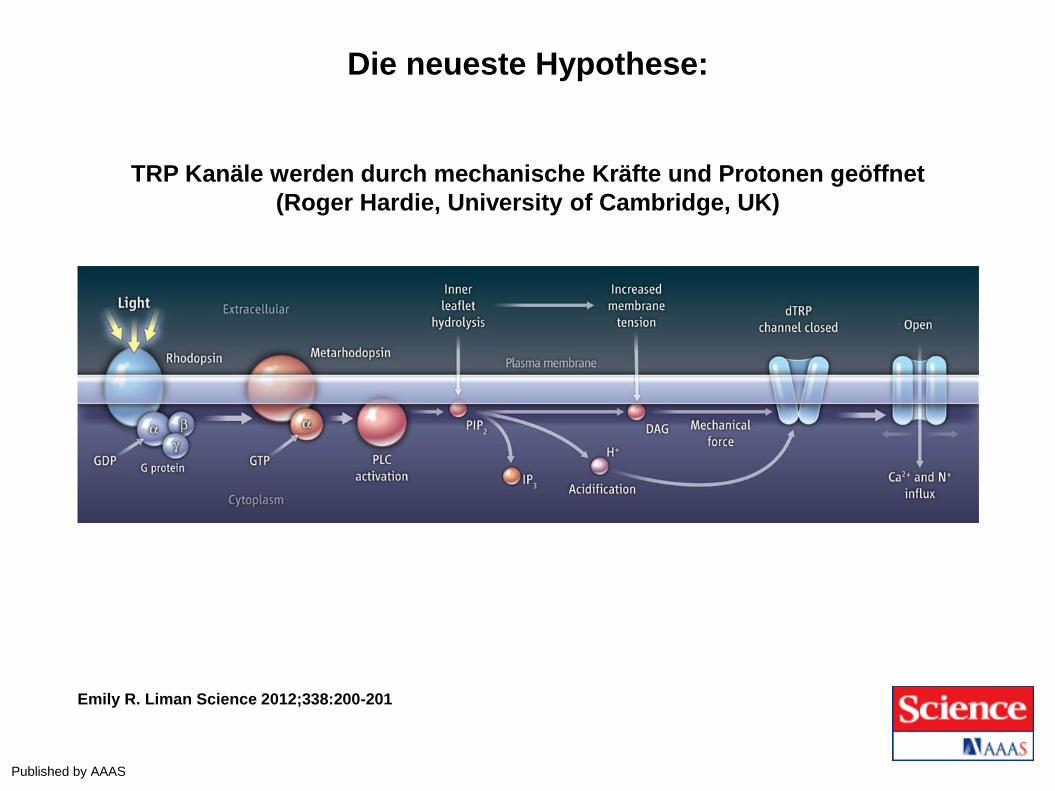
Die neueste Hypothese:
TRP Kanäle werden durch mechanische Kräfte und Protonen geöffnet
(Roger Hardie, University of Cambridge, UK)
Emily R. Liman Science 2012;338:200-201
Published by AAAS

Adaptationsmechanismen:
•Die normale Anpassung des Auges an sich ändernde
Lichtintensitäten.
•Die Reduzierung der ‘firing Frequenz’ eines Neurons,
insbesondere eines Rezeptorneurons, unter Bedingungen
konstanter Stimulierung
Adaptation

Licht-induzierte Translokation in den Rhabomeren
Trpl und Gqα lokalisieren in
Rhabdomeren dunkel-adaptierter
Fliegen, aber nach 1-hr
Lichtexposition translozieren sie
in den Zellkörper
Das Myosin III Protein (kodiert
vom ninaC Gen) ist für diese
Translokation essentiell
Wie konnte das gezeigt werden??
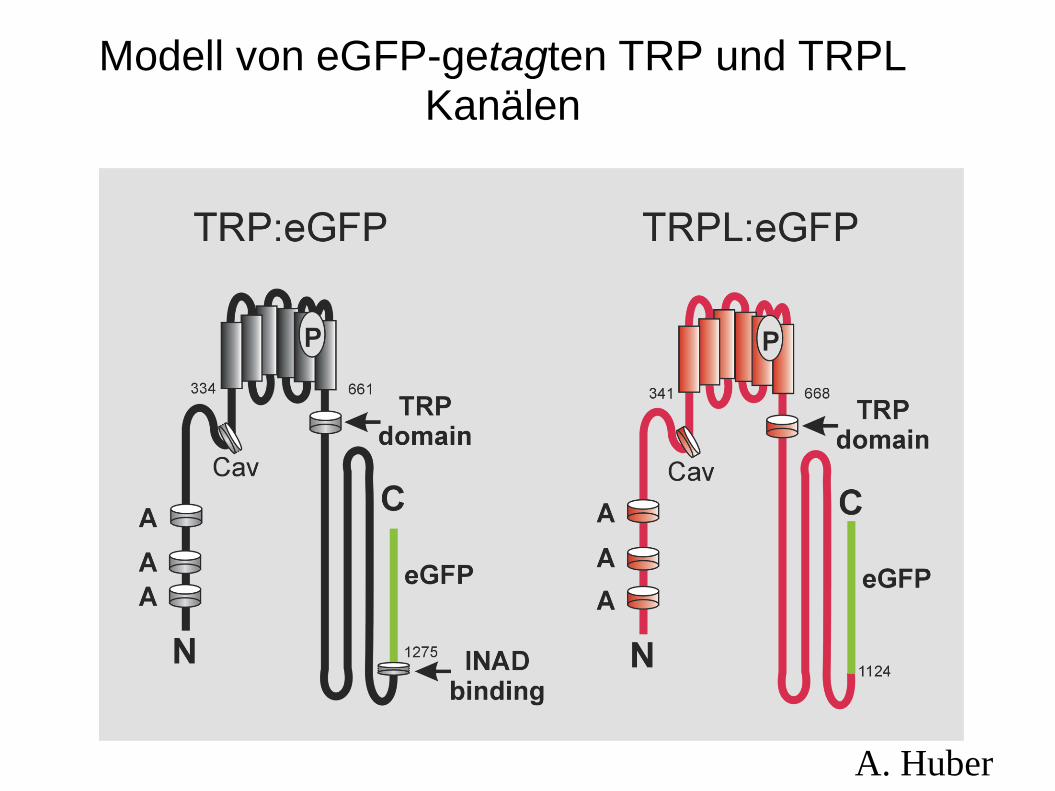
Modell von eGFP-getagten TRP und TRPL Kanälen
A. Huber
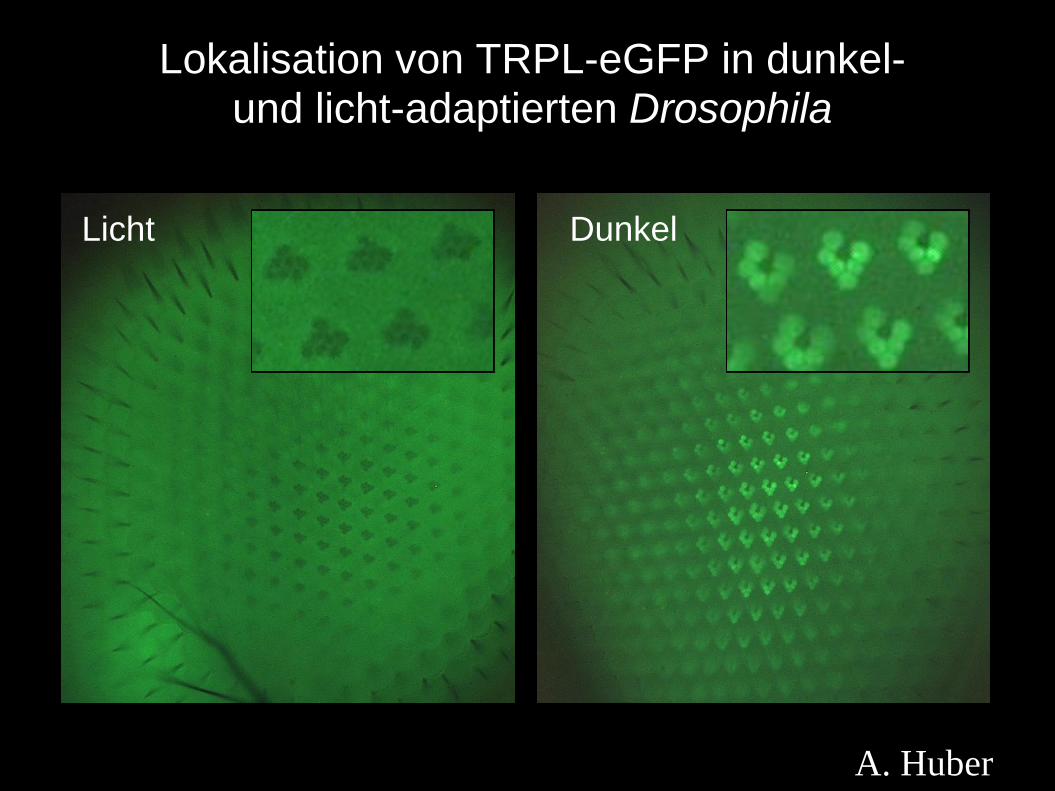
Lokalisation von TRPL-eGFP in dunkel-und licht-adaptierten Drosophila
Licht Dunkel
A. Huber
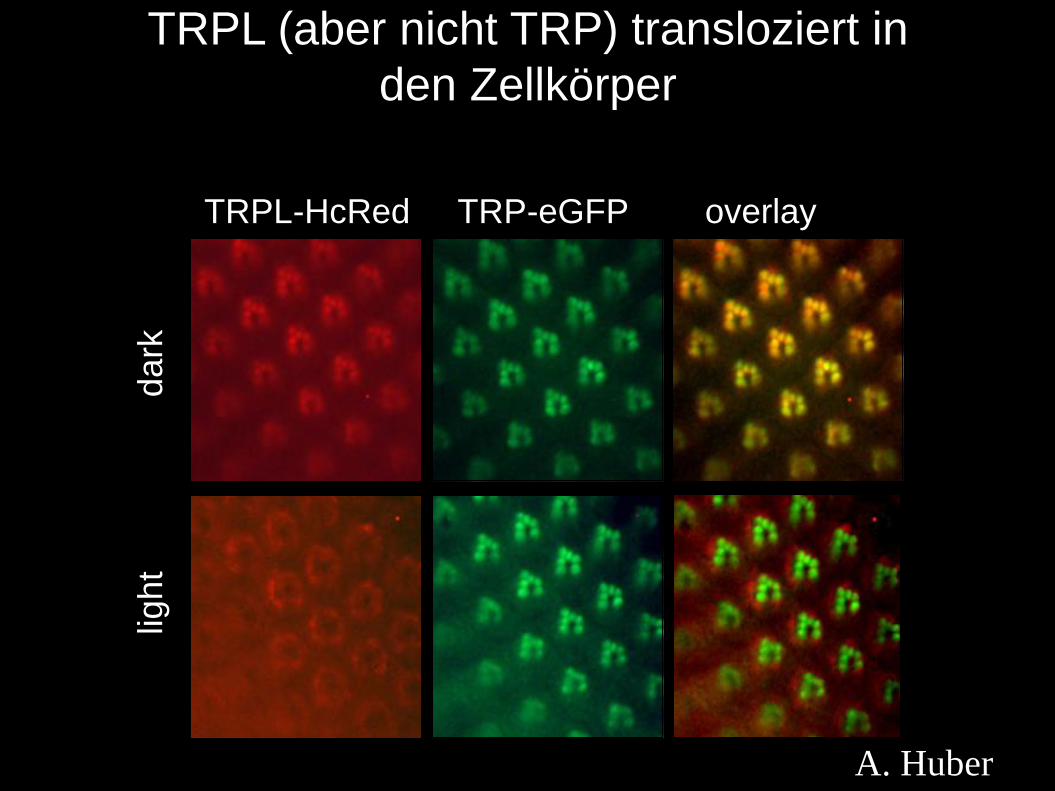
dark
light
TRPL (aber nicht TRP) transloziert in
den Zellkörper
TRPL-HcRed TRP-eGFP overlay
A. Huber

Der Signalplex: Proteine bilden mit Hilfe des INAD
Proteins ein Cluster
•Trp, PLC, und PKC, benötigen
INAD zur Lokalisation in den
Rhabdomeren
•Trpl, Rhodopsin, NinaC, und
CaM kinase lokalisieren ohne
INAD in den Rhabdomeren
•Ermöglicht (im Prinzip) Licht-
regulierte Lokalisation dieser
Proteine
Signalplex trägt zur Schnelligkeit der Phototransduktionskaskade bei
(schnellste bekannte G protein-gekoppelte Signalskaskade überhaupt!)
Rhabdomere membrane

Ca2+
CytosolicExtracellular
R Gq
InsP3
DAG PUFAPIP2
Ca2+
Ca2+
TRPSMC
TRPL
Na+
Zusammenfassung: Drosophila Phototransduktionskaskade
A. Huber
DAG
Lipase
PLUS: Membran Kontraktion und Azidifizierung (H+) der TRP Kanäle

Warum muß die Phototransduktionskaskade
wieder inaktiviert werden??
•Um die maximale zeitliche Auflösung der Antwort auf
Lichtreize zu gewährleisten
•Konstitutive Aktivierung führt zur Degeneration der
Photorezeptoren (z.B., rdgA Mutanten)

I: Meta-Rhodopsin* wird durch Bindung an Arrestin inaktiviert
After C. Zuker
All-trans Retinal bleibt an
Rhodopsin gebunden und
photo-isomerisiert zu 11-cis-3
hydroxy-retinal (durch länger
welliges Licht verglichen mit
dem induzierenden
Lichtstimulus).
Die Funktion der Rhodopsin
Phosphorylierung ist nicht
bekannt, aber RK Mutanten
beeinflussen ERG Amplitude
PUFA’s
DAG-Lipase
Trp/Trpl
Calcium Einstrom/Depolarization
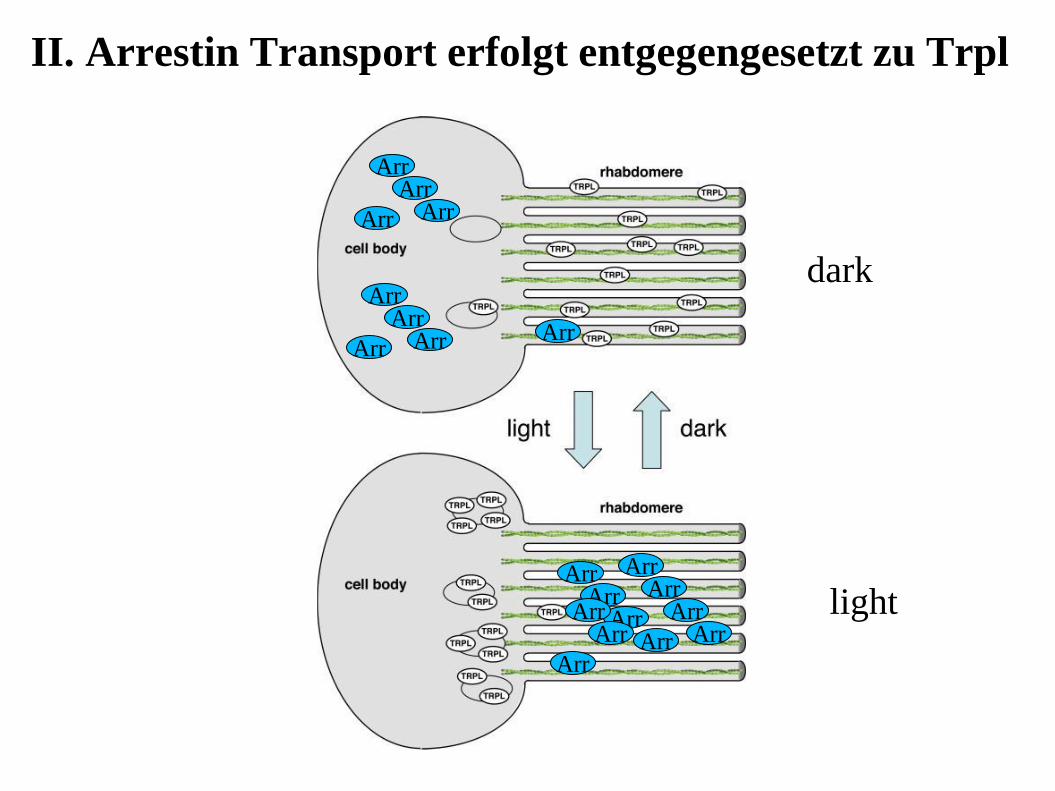
II. Arrestin Transport erfolgt entgegengesetzt zu Trpl
ArrArr
ArrArr
ArrArr
ArrArr
Arr
ArrArr
Arr
Arr
ArrArr
Arr
Arr
Arr
ArrArr
light
dark
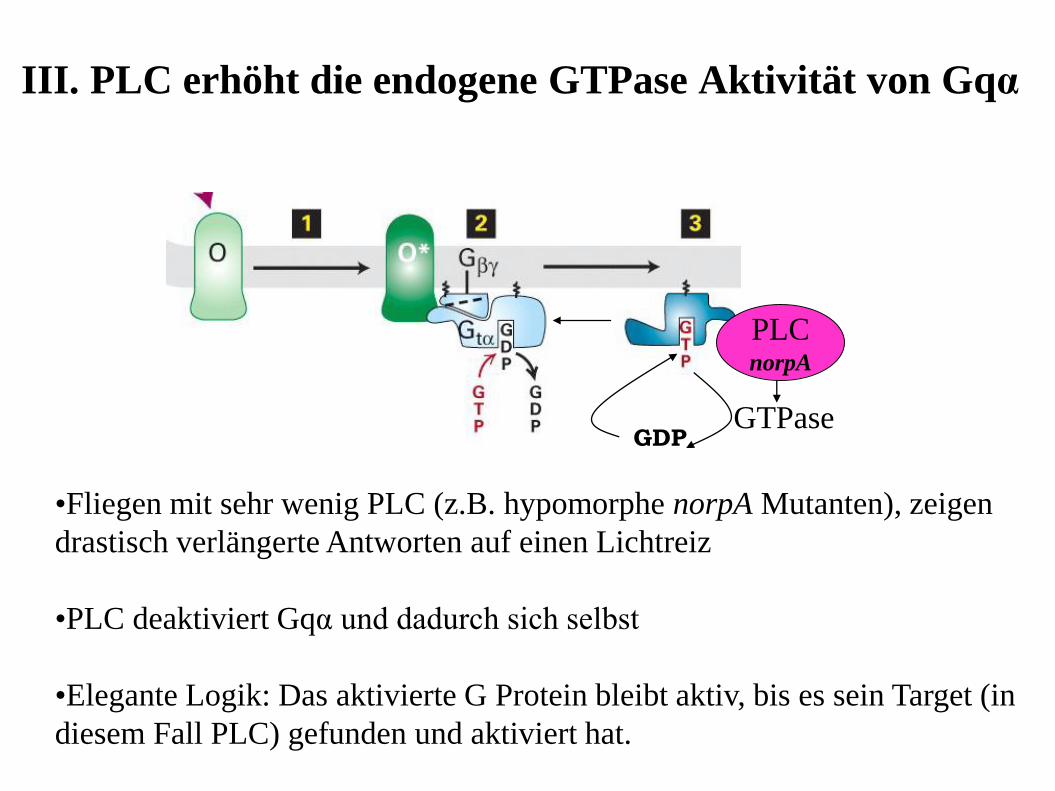
III. PLC erhöht die endogene GTPase Aktivität von Gqα
PLCnorpA
GDPGTPase
•Fliegen mit sehr wenig PLC (z.B. hypomorphe norpA Mutanten), zeigen
drastisch verlängerte Antworten auf einen Lichtreiz
•PLC deaktiviert Gqα und dadurch sich selbst
•Elegante Logik: Das aktivierte G Protein bleibt aktiv, bis es sein Target (in
diesem Fall PLC) gefunden und aktiviert hat.

IV: Calcium wir aktiv aus der Zelle transportiert
durch den Na+/Ca2+ transporter CalX
Extrazellulär
Cytosol
C. Montell
•Durch reduzierte trp Funktion induzierte Retinale Degeneration kann
durch gleichzeitige Reduktion der CalX Function ‘gerettet’ werden
•Durch konstitutive trp Funktion (Kanäle immer offen) induzierte Retinale
Degeneration kann durch Überexpression von CalX gerettet werden
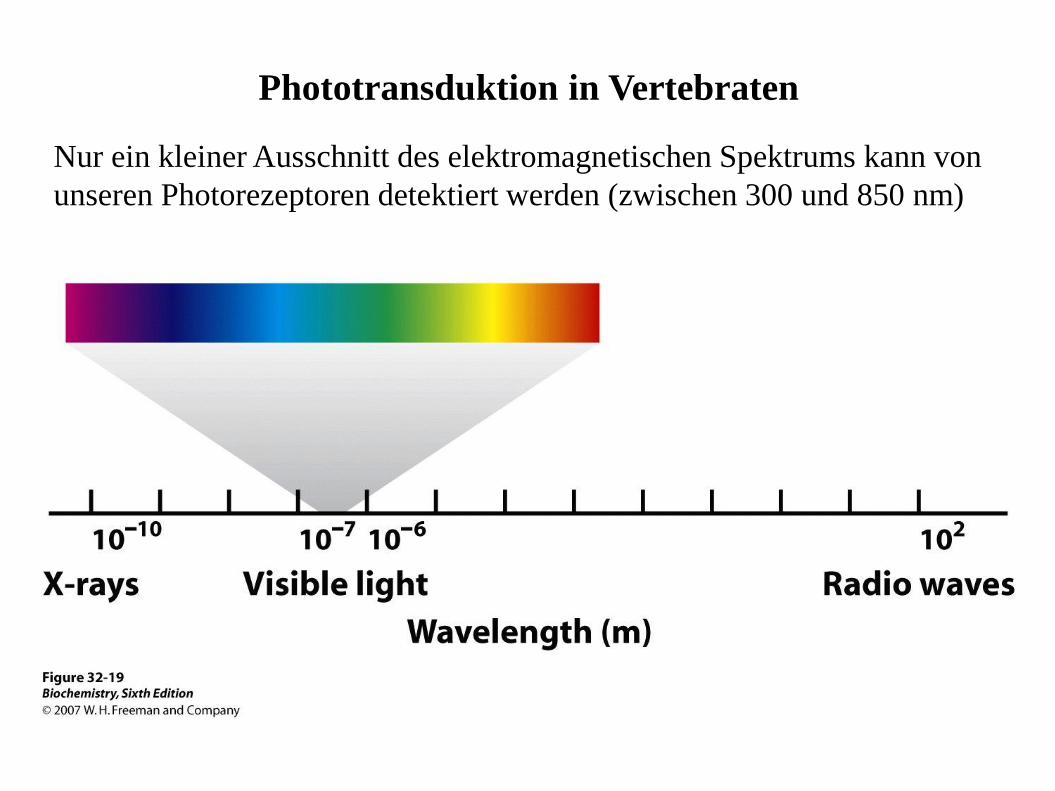
Phototransduktion in Vertebraten
Nur ein kleiner Ausschnitt des elektromagnetischen Spektrums kann von
unseren Photorezeptoren detektiert werden (zwischen 300 und 850 nm)

Phototransduktion in Vertebraten: Einige wichtige
Fakten
•Zapfen (cones): Farbsehen und helles Licht
•Stäbchen (rods): Schwarz/Weiss und Dämmerungslicht
•Die menschliche Retina enthält 6 Millionen Zapfen und 125 Millionen
Stäbchen
•Eine Stäbchenzelle kann ein EINZELNES Photon wahrnehmen, und das
Gehirn benötigt nur < 10 Antworten einer Zelle um sie als Lichtblitz
einzuordnen

Phototransduktion in Vertebraten: Where does it happen?
Stäbchenzellen

Photorezeptoren
Stäbchen Zapfen
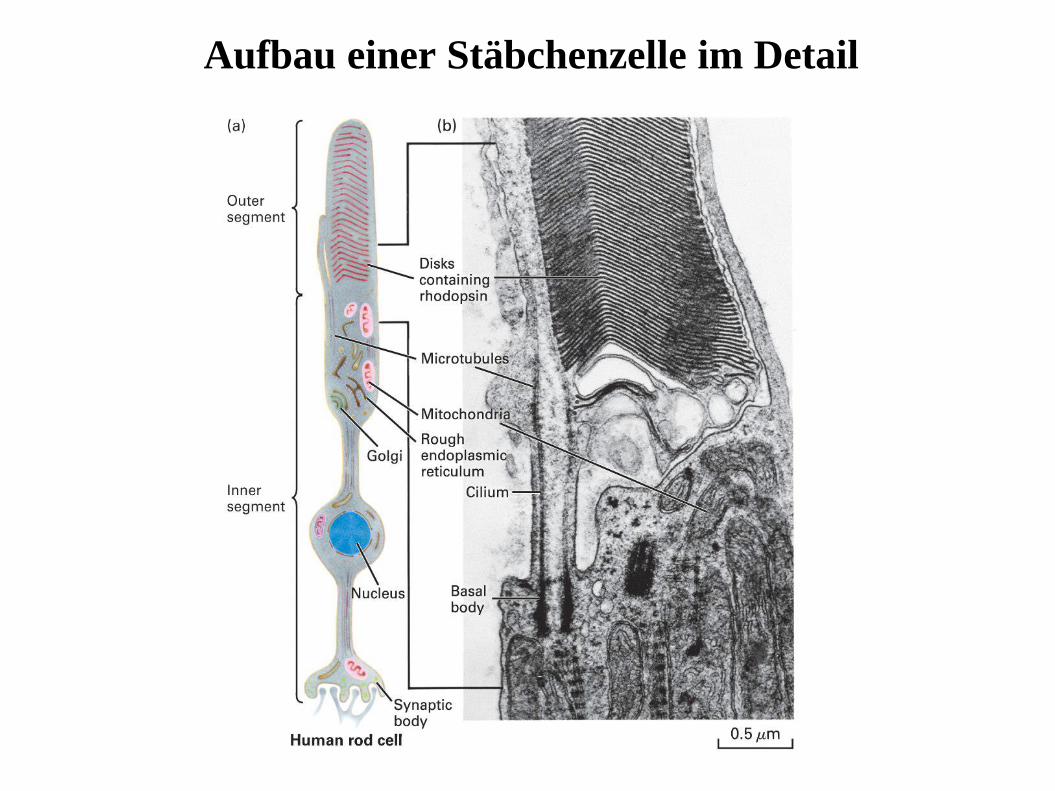
Aufbau einer Stäbchenzelle im Detail

Die Phototransduktionskaskade in Vertebraten
•Die Phototransduktionskaskade von Invertebraten wurde in der
Hauptsache durch Genetik entschlüsselt (z.B., norpA, trp, trpl
Mutanten).
•Die Phototransduktionskaskade der Vertebraten wurde in der
Hauptsache biochemisch entschlüsselt.
•Rinderopsin war das erste Protein der 7TM-Rezeptor Familie, und die
Struktur wurde im Jahr 2000 gelöst.

I: Der wichtigste Schritt bei der Rhodopsin Photorezeption……ist sehr ähnlich bei Vertebraten und Invertebraten
Die Lichtenergie eines Photons wird in atomare Bewegung umgesetzt (5 Ǻ
Drehung des N Atoms, das Retinal und Protein über eine Schiff’sche Base
verbindet)
Licht
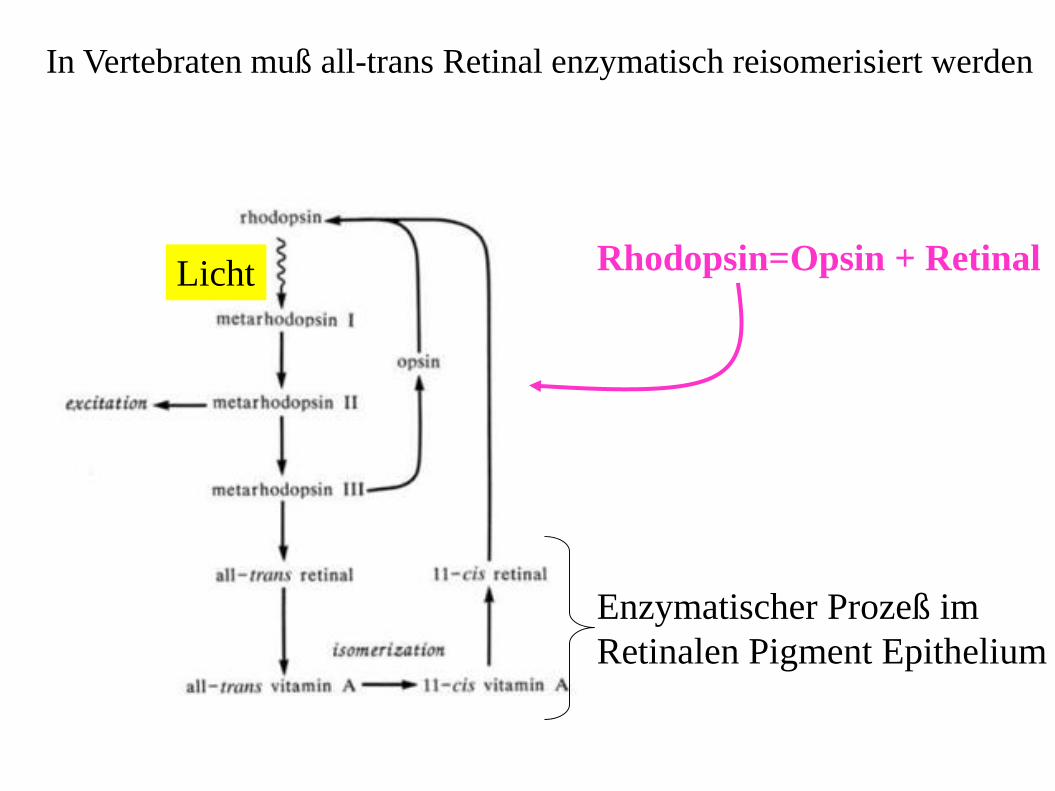
In Vertebraten muß all-trans Retinal enzymatisch reisomerisiert werden
Enzymatischer Prozeß im
Retinalen Pigment Epithelium
Rhodopsin=Opsin + Retinal
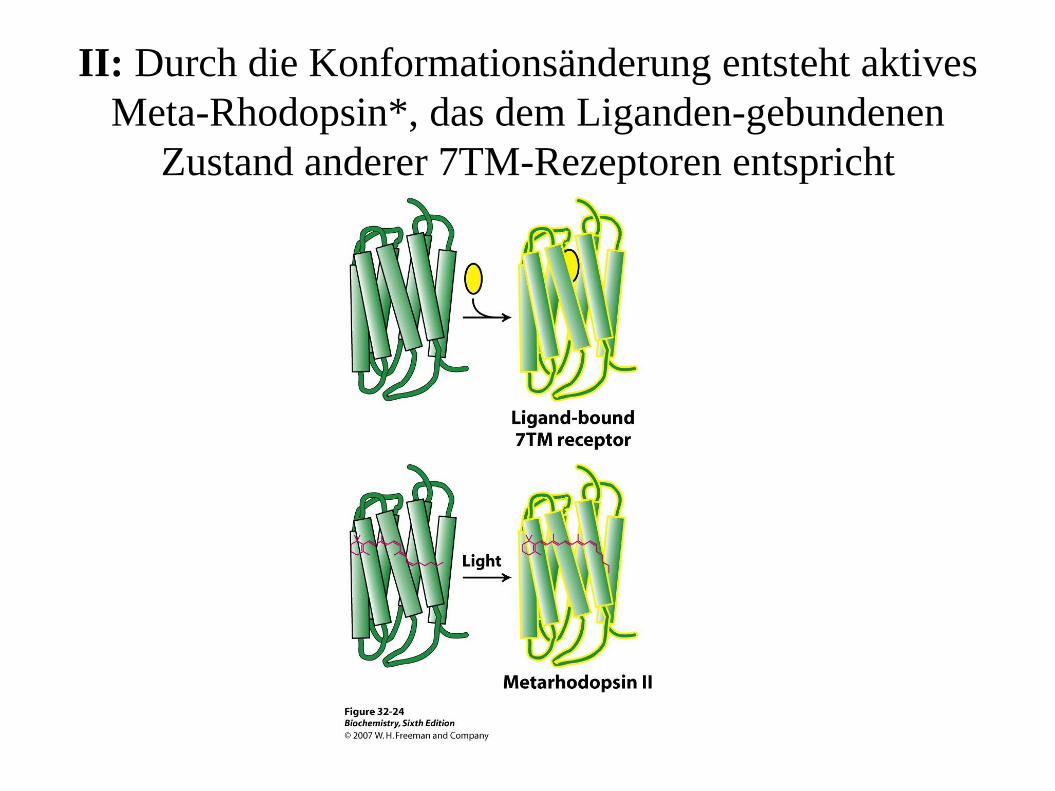
II: Durch die Konformationsänderung entsteht aktives
Meta-Rhodopsin*, das dem Liganden-gebundenen
Zustand anderer 7TM-Rezeptoren entspricht

III. Meta-Rhodopsin* aktiviert das G-protein
Transducin
….bisher alles wie in Fliegen
Transducin
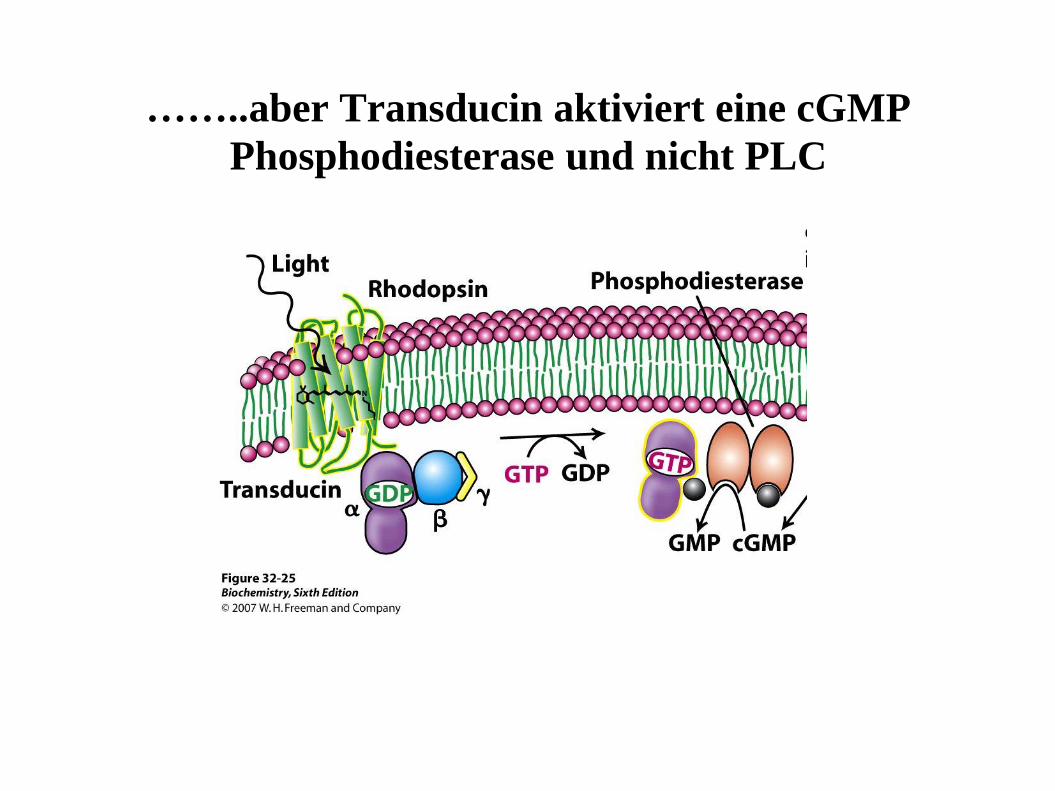
……..aber Transducin aktiviert eine cGMP
Phosphodiesterase und nicht PLC
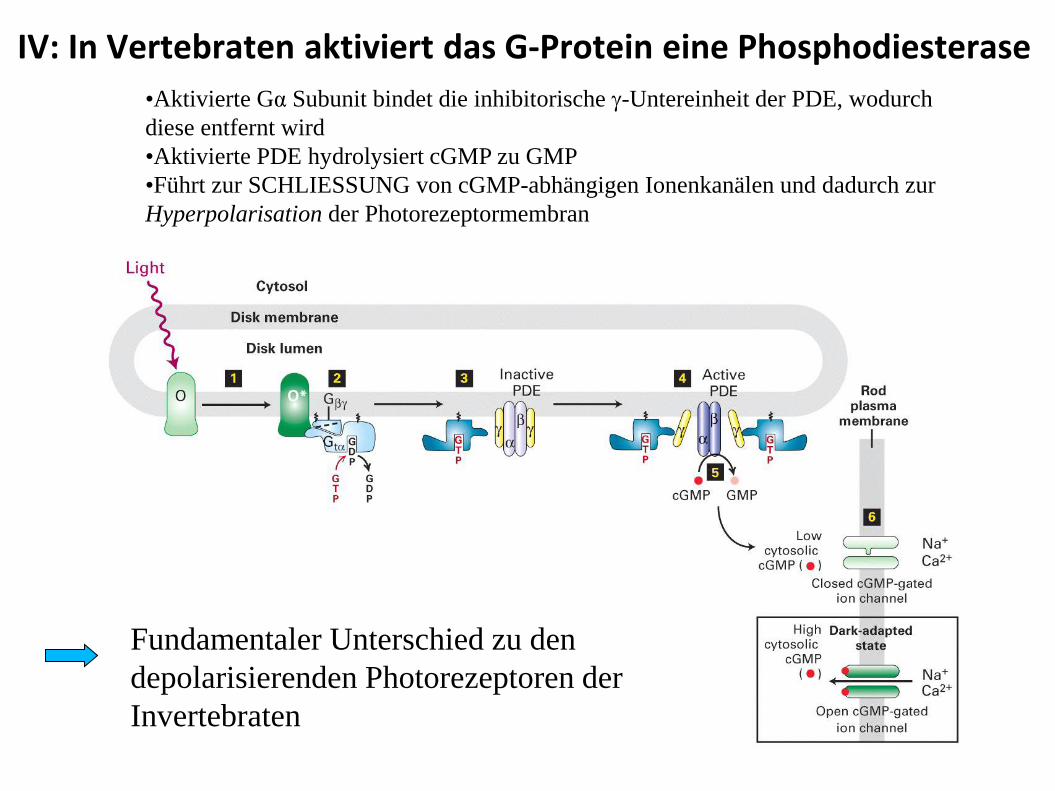
IV: In Vertebraten aktiviert das G-Protein eine Phosphodiesterase
•Aktivierte Gα Subunit bindet die inhibitorische -Untereinheit der PDE, wodurch
diese entfernt wird
•Aktivierte PDE hydrolysiert cGMP zu GMP
•Führt zur SCHLIESSUNG von cGMP-abhängigen Ionenkanälen und dadurch zur
Hyperpolarisation der Photorezeptormembran
Fundamentaler Unterschied zu den
depolarisierenden Photorezeptoren der
Invertebraten

Dunkelstrom
Na+ InfluxK+ Efflux
Dunkelheit
Kein Na+ Influx
K+ EffluxLicht
Depolarisierung
Hyperpolarisierung

Wie kann Hyperpolarisierung zur Weiterleitung des
elektrischen Signals an das Gehirn führen?
Bipolar Zellen werden aktiviert, wenn weniger (in diesem Falle
inhibierendes) Glutamat von den Stäbchen und Zapfen
ausgeschüttet wird

Signalverstärkung
•In Vertebraten aktiviert ein Rhodopsin Molekül bis zu 800 G-Proteine
•Ein PDE Molekül katalysiert die Hydrolyse von bis zu 6 cGMP
Molekülen
•In der Summe führt die Aktivierung von einem Rhodopsin Molekül zur
Schließung von ~200 Ionenkanälen (entspricht einer Membranpotential
Änderung von 1 mV)
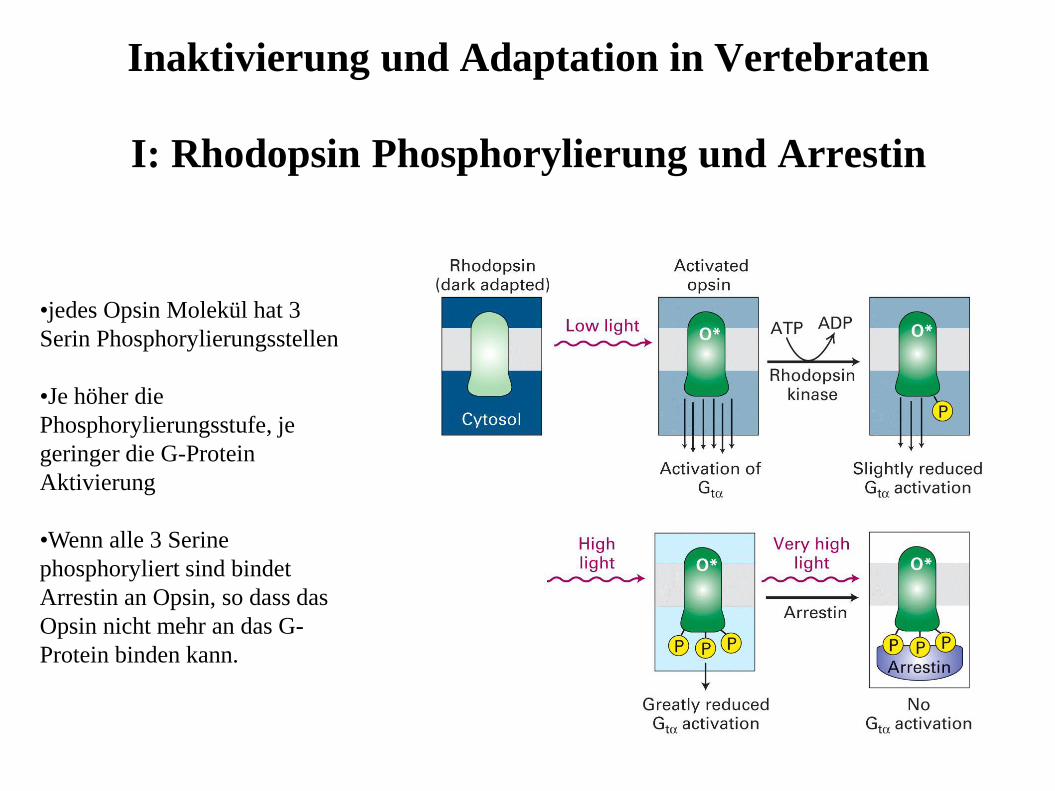
Inaktivierung und Adaptation in Vertebraten
I: Rhodopsin Phosphorylierung und Arrestin
•jedes Opsin Molekül hat 3
Serin Phosphorylierungsstellen
•Je höher die
Phosphorylierungsstufe, je
geringer die G-Protein
Aktivierung
•Wenn alle 3 Serine
phosphoryliert sind bindet
Arrestin an Opsin, so dass das
Opsin nicht mehr an das G-
Protein binden kann.

Inaktivierung und Adaptation in Vertebraten
II: Translokation von Phototransduktion Molekülen
•Retinas Dunkel-adaptierter
Ratten vor und nach kurzer
Lichtexposition gefärbt mit
anti -Gα and Gβ Antikörpern
•Nur im outer segment (OS)
kann das G-protein mit
Rhodopsin interagieren
•Wie in Fliegen transloziert
Arrestin in die
entgegengesetzte Richtung

III. PDE erhöht die intrinsische GTPase Aktivität von Gα
PDE
GTPaseGDP
Gα-GDP re-assoziiert mit Gβγ
Ähnlich wie PLC in Fliegen!

IV: Um die cGMP-abhängigen Kanäle wieder zu
öffnen, muss die cGMP Konzentration erhöht werden
GTP
[cGMP]guanylate cyclase
Ca2+
Ca2+
Schließung der cGMP Kanäle nach Lichtexposition
verringert die [Ca2+]i, wodurch Guanylat Cyclase
aktiviert wird
open
[cGMP] closed

Einige Hauptunterschiede zwischen Vertebraten
und Invertebraten Phototransduktion
Vertebraten Invertebraten
G-protein aktiviert PDE G-Protein aktiviert PLC
c-GMP abhängige
Ionenkanäle schließen;
Photorezeptorzellen
hyperpolarisieren
Trp, Trpl Kanäle öffnen;
Photorezeptorzellen
depolarisieren
all-trans Retinal dissoziiert
vom aktiven Opsin und muss
enzymatisch reisomerisiert
werden. Trägt zu langsamer
Dunkeladaptation bei.
all-trans Retinal bleibt Opsin-
gebunden und wird schnell
photoisomerisiert. Trägt zu
schneller Dunkeladaptation
bei

Farbensehen
Stäbchen exprimieren Rhodopsin Zapfen exprimieren 3 Farbopsine

Für die unterschiedlichen spektralen Eigenschaften der ‘grün’ und ‘rot’
Opsine sind nur einige wenige Aminosäuren verantwortlich
identische Reste
unterschiedliche Reste
unterschiedlich und verantwortlich
für Absorptionsspektrum

Farbblindheit—ein genetischer Defekt
•Die Gene für rote und grüne Opsine liegen benachbart auf dem X-Chromosom
•Homologe Rekombination zwischen den 2 Genen führt zur Farbblindheit

Wie häufig ist Farbblindheit? 1/12 Männern (8%) und 1/200 Frauen (0.5%) sind
rot/grün farbenblind.
Defekte in blauen Zapfenzellen (Typ 3: Tritanopes) sind sehr selten (Blaues Opsin liegt auf Chromosome 7)
Rot-Grün Blindheit ist sehr häufig (bei Männern). In
einem Raum mit 25 Menschen, sollte sich im
Durchschnitt eine farbenblinde Person befinden….
Typ 1 (Protanopes) und Typ 2
(Deuteranopes):
Defekte in roten und grünen
Zapfenzellen
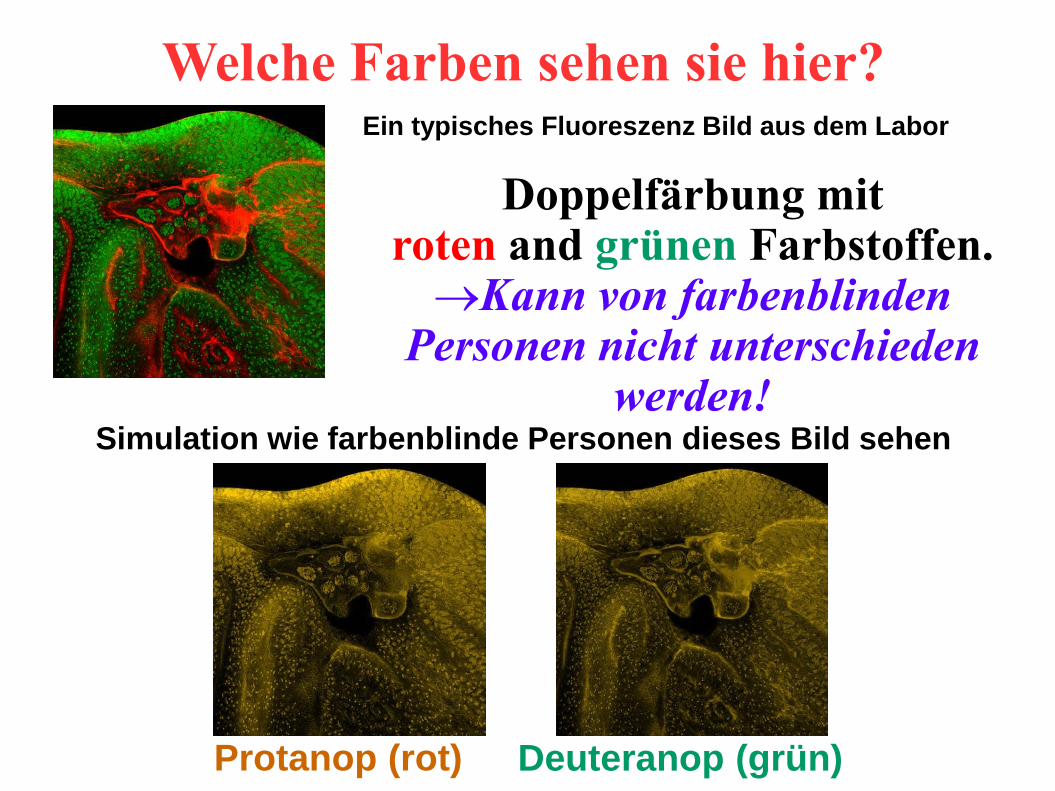
Doppelfärbung mit roten and grünen Farbstoffen.Kann von farbenblinden
Personen nicht unterschieden werden!
Protanop (rot) Deuteranop (grün)
Welche Farben sehen sie hier?
Simulation wie farbenblinde Personen dieses Bild sehen
Ein typisches Fluoreszenz Bild aus dem Labor

Ein kleiner Exkurs über nicht-visuelle Photorezeptoren

Schlaf-Wach Rhythmus eines französichen Höhlenforschers (Michel Siffre, 1962)
t = 24,8 h
Circadiane Rhythmen in Menschen
Tageszeit

“Bunker Experimente”(418 Experimente mit 447 Freiwilligen
[Studenten] zwischen 1964 - 1989)
Erling-Andechs
Jürgen Aschoff (1913 – 1998)
Rütger Wever(1923 – 2005)

Schlaf-Wachrhythmen im Bunker
Kontakt mit der
Außenwelt?
Organismen besitzen eine innere Uhr, die mit der Umwelt synchronisiert werden kann
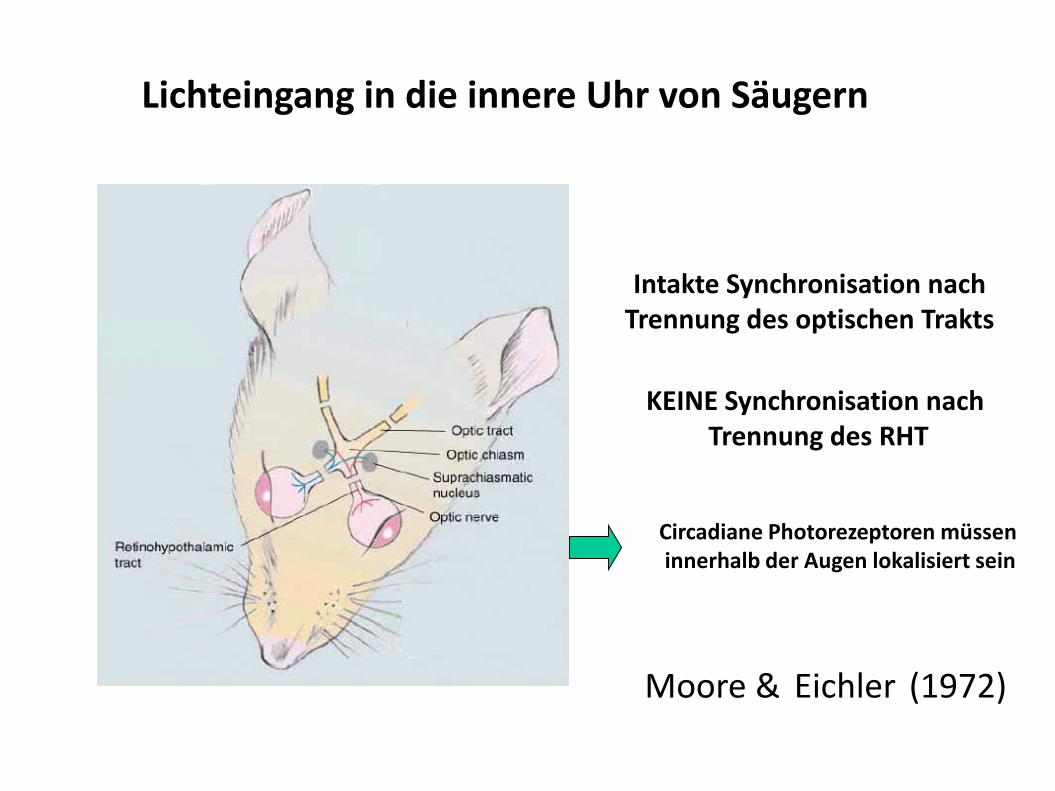
Moore & Eichler (1972)
Intakte Synchronisation nach Trennung des optischen Trakts
KEINE Synchronisation nach Trennung des RHT
Circadiane Photorezeptoren müssen innerhalb der Augen lokalisiert sein
Lichteingang in die innere Uhr von Säugern

Roelof Hut
Wie kann man testen, ob der Lichteingang in die innere Uhr funktioniert?
Laufaktivität
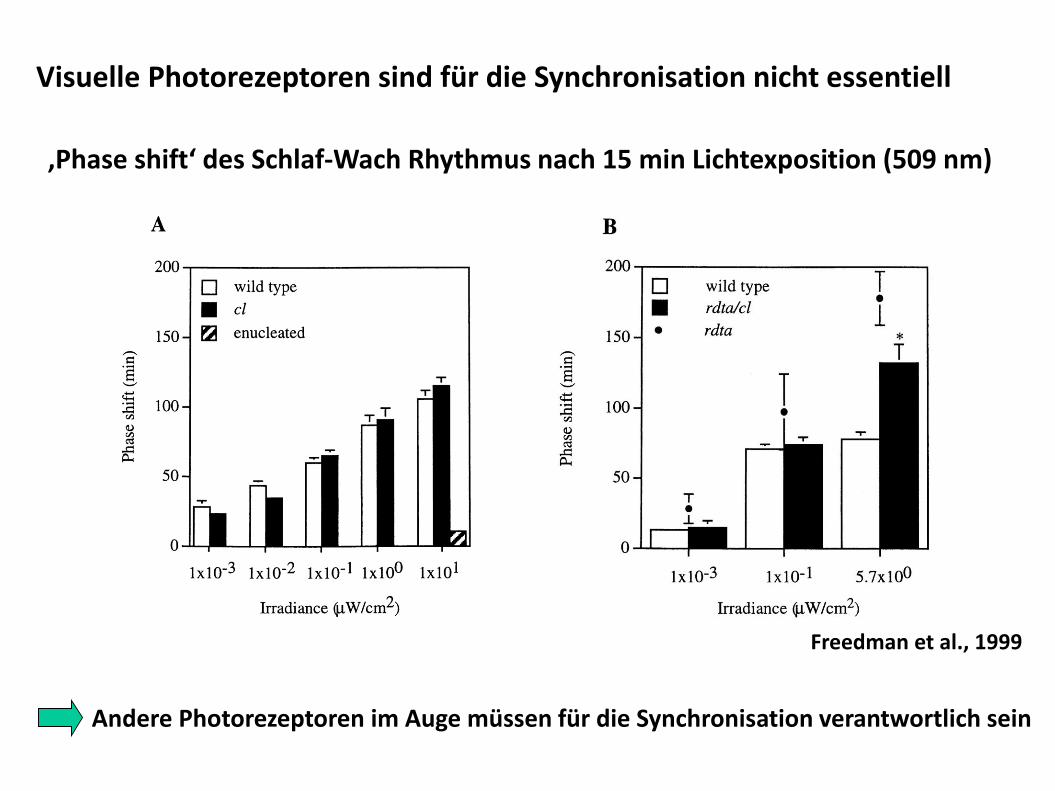
Visuelle Photorezeptoren sind für die Synchronisation nicht essentiell
‚Phase shift‘ des Schlaf-Wach Rhythmus nach 15 min Lichtexposition (509 nm)
Andere Photorezeptoren im Auge müssen für die Synchronisation verantwortlich sein
Freedman et al., 1999

Ein Subset (1-2%) der RGC exprimieren Melanopsin
1-2% Melanopsin positiv
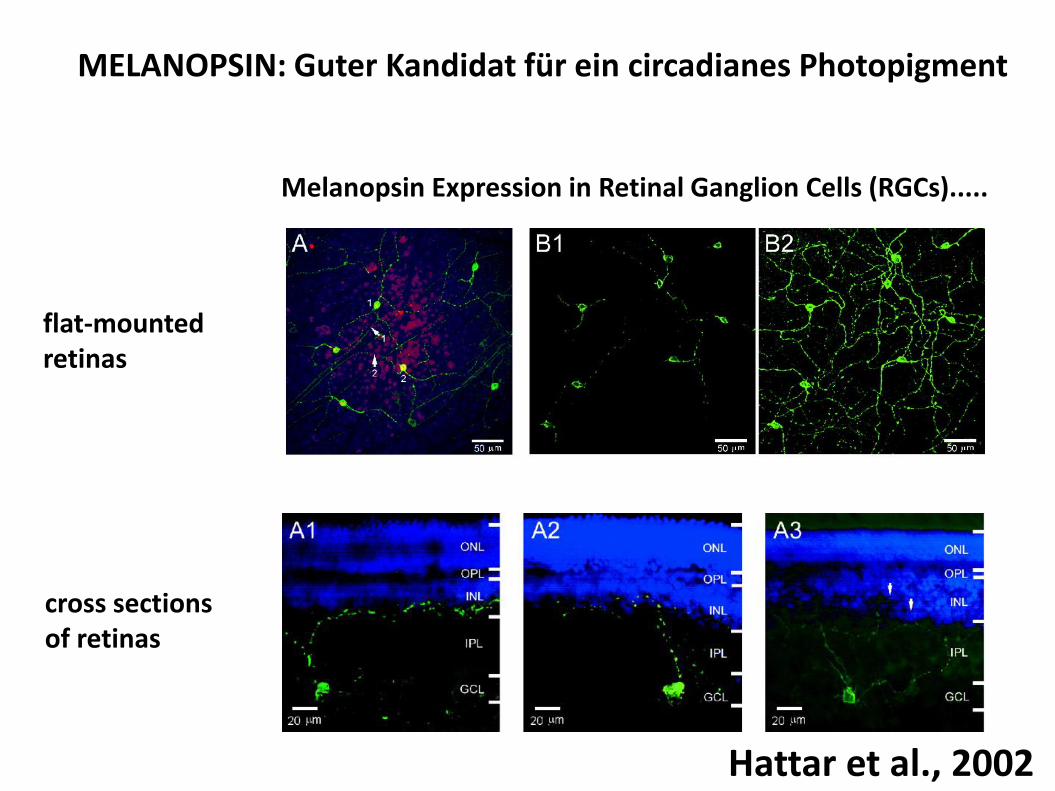
MELANOPSIN: Guter Kandidat für ein circadianes Photopigment
Melanopsin Expression in Retinal Ganglion Cells (RGCs).....
cross sectionsof retinas
flat-mounted retinas
Hattar et al., 2002

...Melanopsin RGCs projizieren zum SCN
lacZ Färbung der RHT Projektionenin den SCN
tau-lacZ Reporter inseriert im Melanopsin Gen
Hattar et al., 2002
RGC cell
Science; 295(5557):1065-70.
MELANOPSIN: Guter Kandidat für ein circadianes Photopigment

... RGCs sind intrinsisch photosensitiv (unabhängig von Stäbchen und Zapfen)
Spektrale Sensitivität passt auf ein 484nm Opsin
Einzelne RGC depolarisieren nach Lichtexposition
Berson et al., 2002
Science; 295(5557):1070-3.
MELANOPSIN: Guter Kandidat für ein circadianes Photopigment

Melanopsin, aber auch Stäbchen und Zapfen tragen zur Licht-Synchronisation bei!
Panda et al., 2003
100 lux
100 lux100 lux
100 lux
800 lux
Science; 301(5632):525-7
Wildtyp Maus Kein Melanopsin in RGCs
Keine Stäbchen und ZapfenWeder noch

Melanopsin ist enger verwandt mit Rhodopsinen der Invertebraten als mit den visuellen Opsinen der Vertebraten
Langer cytoplasmatischer C-terminus: existiert nicht in den visuellen Opsinen der Vertebraten
Cytoplasmatische Loops vermitteln G-Protein Bindung: ähnlich zu Fliegen Rhodopsinen
Nayak et al., 2007
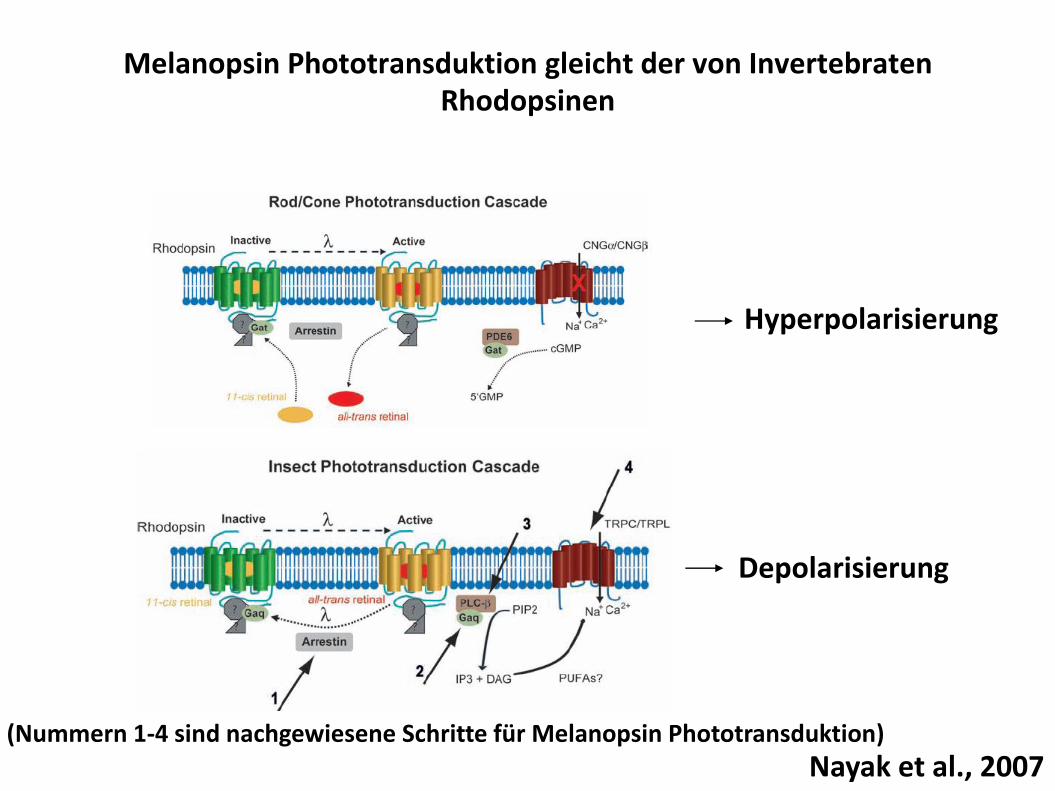
Melanopsin Phototransduktion gleicht der von Invertebraten Rhodopsinen
Hyperpolarisierung
Depolarisierung
Nayak et al., 2007(Nummern 1-4 sind nachgewiesene Schritte für Melanopsin Phototransduktion)

Eigenschaften von Melanopsin und RGCs
•11-cis Retinal Photo re-isomerisiert (……..wie in Fliegen)
•RGC depolarisieren nach Lichtreizung (…..wie in Fliegen)
•Melanopsin wird nicht in spezialisierten Membranstrukturen (z.B., ‘discs’) exprimiert, sondern gleichmäßig verteilt in der RGC Plasmamembran von Soma, Axon und Dendrit
•Ablation der Melanopsin RGC führt zum totalen Verlust der Synchronisation. D.h. die zur Synchronisation beitragenden Stäbchen und Zapfen sind mit diesen RGC verschaltet.





























